
Enzymes Involved in Defense Functions of Hemocytes of Mussel Mytilus galloprovincialis
10
Enzymes Involved in Defense Functions of Hemocytes of Mussel Mytilus galloprovincialis Marı ´a J. Carballal,* Carmen Lo ´pez,* Carlos Azevedo,** and Antonio Villalba* *Centro de Investigacio ´ns Marin ˜as, Xunta de Galicia, P.O. Box 208, E-36600 Vilagarcı´a de Arousa, Spain; and **Department of Cell Biology, Institute of Biomedical Sciences, University of Oporto, 4000 Porto, Portugal Received June 27, 1996; accepted February 28, 1997 Mytilus galloprovincialis hemocytes contain phos- phatases, esterases, proteases, and glucosidases as revealed by a semiquantitative colorimetric method. The activity levels of some enzymes changed through- out the months of the experiment without a seasonal pattern. No differences in the enzyme levels were found between the two groups of mussels of different age (cultivated 11 and 19 months on ropes hung from the same raft). Light microscopy study of hemocyte enzymes showed that hyalinocytes and granulocytes had reaction of acid phosphatase, b-glucuronidase, a-naphthyl butyrate esterase, naphthol AS-D chloroac- etate esterase, and a-naphthyl acetate esterase. Pheno- loxidase and peroxidase activities were also detected in monolayers of hemocytes. Ultrastructural study revealed that the granules of hemocytes contained acid phosphatase, b-glucuronidase, nonspecific ester- ases, peroxidase, and NADH oxidase. Products of b-glucuronidase and peroxidase reaction were also localized in the plasma membrane. Lysozyme activity level was higher in hemocytes than in serum. Compari- son between the two groups of different age showed a higher activity level of lysozyme in younger mussels. r 1997 Academic Press KEY WORDS: Mytilus galloprovincialis; hemocytes; cel- lular defense; hydrolytic enzymes; phenoloxidase; per- oxidase; NADH oxidase. INTRODUCTION Two mechanisms for cell killing have been described for mollusc hemocytes during phagocytosis: the release of lysosomal enzymes and other lysins (Cheng, 1981) and the respiratory burst which involves the release of reactive oxygen metabolites (Nakamura et al., 1985; Dikkeboom et al., 1988; Bache `re et al., 1991; Pipe, 1992). In order to understand the interaction pathogen– hemocyte it is necessary to have a better knowledge of these cytotoxic mechanisms. There are numerous publi- cations of bivalve hemocyte enzymes (Cheng, 1981; Huffman and Tripp, 1982; Mohandas et al., 1985; Chu and La Peyre, 1989; Auffret, 1989; Pipe, 1990; Pipe et al., 1993; Holden et al., 1994; Coles and Pipe, 1994). However, studies on changes of hemocyte enzymes in normal and pathological conditions are limited. The aim of the present study was to determine the contents of enzymes involved in defense functions of Mytilus galloprovincialis hemocytes and their varia- tions according to season and age. The location of enzymes within the hemocyte and the differences in enzyme contents among hemocyte types were also studied. MATERIALS AND METHODS Cultured adult specimens of M. galloprovincialis were sampled from the Rı ´a de Arosa (Galicia, NW Spain). Hemolymph samples were collected from the posterior adductor muscle of the mussels using sterile syringes containing different solutions depending on the enzyme assays. API ZYM test A colorimetric semiquantitative method, API ZYM (BioMe ´rieux), was used to estimate activity levels of hydrolytic enzymes in mussel hemocytes. Hemolymph from five mussels was directly withdrawn 1:3 in Alsev- er’s solution (Bache `re et al., 1988) and pooled. A sample of 2.5 3 10 6 hemocytes was then spun (750g, 10 min) to eliminate the serum. Cells were resuspended in 1.4 ml distilled water, and 65 μl of this suspension was deposited in each well of a plate containing the sub- strates for each enzyme reaction. After 4 hr of incuba- tion at 37°C, the enzyme activity was revealed using the kit reagents. The intensity of reaction was evalu- ated according to the kit color scale. This test was used to estimate the seasonal variation of hemocyte enzymes. For this purpose, monthly samples (12 months) of five commercial mussels (7–10 cm) were collected from the same raft. This test was also applied to study the enzyme content of hemocytes from two groups of five mussels of different ages (after JOURNAL OF INVERTEBRATE PATHOLOGY 70, 96–105 (1997) ARTICLE NO. IN974670 96 0022-2011/97 $25.00 Copyright r 1997 by Academic Press All rights of reproduction in any form reserved.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Enzymes Involved in Defense Functions of Hemocytes of Mussel Mytilus galloprovincialis

Enzymes Involved in Defense Functions of Hemocytesof Mussel Mytilus galloprovincialis
Marıa J. Carballal,* Carmen Lopez,* Carlos Azevedo,** and Antonio Villalba**Centro de Investigacions Marinas, Xunta de Galicia, P.O. Box 208, E-36600 Vilagarcıa de Arousa, Spain; and **Department of Cell Biology,
Institute of Biomedical Sciences, University of Oporto, 4000 Porto, Portugal
Received June 27, 1996; accepted February 28, 1997
Mytilus galloprovincialis hemocytes contain phos-phatases, esterases, proteases, and glucosidases asrevealed by a semiquantitative colorimetric method.The activity levels of some enzymes changed through-out the months of the experiment without a seasonalpattern. No differences in the enzyme levels werefound between the two groups of mussels of differentage (cultivated 11 and 19 months on ropes hung fromthe same raft). Light microscopy study of hemocyteenzymes showed that hyalinocytes and granulocyteshad reaction of acid phosphatase, b-glucuronidase,a-naphthyl butyrate esterase, naphtholAS-D chloroac-etate esterase, anda-naphthyl acetate esterase. Pheno-loxidase and peroxidase activities were also detectedin monolayers of hemocytes. Ultrastructural studyrevealed that the granules of hemocytes containedacid phosphatase, b-glucuronidase, nonspecific ester-ases, peroxidase, and NADH oxidase. Products ofb-glucuronidase and peroxidase reaction were alsolocalized in the plasma membrane. Lysozyme activitylevel was higher in hemocytes than in serum. Compari-son between the two groups of different age showed ahigher activity level of lysozyme in younger mussels.r 1997Academic Press
KEYWORDS:Mytilus galloprovincialis;hemocytes; cel-lular defense; hydrolytic enzymes; phenoloxidase; per-oxidase; NADH oxidase.
INTRODUCTION
Two mechanisms for cell killing have been describedfor mollusc hemocytes during phagocytosis: the releaseof lysosomal enzymes and other lysins (Cheng, 1981)and the respiratory burst which involves the release ofreactive oxygen metabolites (Nakamura et al., 1985;Dikkeboom et al., 1988; Bachere et al., 1991; Pipe,1992). In order to understand the interaction pathogen–hemocyte it is necessary to have a better knowledge ofthese cytotoxicmechanisms. There are numerous publi-cations of bivalve hemocyte enzymes (Cheng, 1981;Huffman and Tripp, 1982; Mohandas et al., 1985; Chu
and La Peyre, 1989; Auffret, 1989; Pipe, 1990; Pipeet al., 1993; Holden et al., 1994; Coles and Pipe, 1994).However, studies on changes of hemocyte enzymes innormal and pathological conditions are limited.The aim of the present study was to determine the
contents of enzymes involved in defense functions ofMytilus galloprovincialis hemocytes and their varia-tions according to season and age. The location ofenzymes within the hemocyte and the differences inenzyme contents among hemocyte types were alsostudied.
MATERIALS AND METHODS
Cultured adult specimens of M. galloprovincialiswere sampled from the Rıa de Arosa (Galicia, NWSpain). Hemolymph samples were collected from theposterior adductor muscle of the mussels using sterilesyringes containing different solutions depending onthe enzyme assays.
API ZYM test
A colorimetric semiquantitative method, API ZYM(BioMerieux), was used to estimate activity levels ofhydrolytic enzymes in mussel hemocytes. Hemolymphfrom five mussels was directly withdrawn 1:3 in Alsev-er’s solution (Bachere et al., 1988) and pooled. A sampleof 2.5 3 106 hemocytes was then spun (750g, 10 min) toeliminate the serum. Cells were resuspended in 1.4 mldistilled water, and 65 µl of this suspension wasdeposited in each well of a plate containing the sub-strates for each enzyme reaction. After 4 hr of incuba-tion at 37°C, the enzyme activity was revealed usingthe kit reagents. The intensity of reaction was evalu-ated according to the kit color scale.This test was used to estimate the seasonal variation
of hemocyte enzymes. For this purpose, monthlysamples (12 months) of five commercial mussels (7–10cm) were collected from the same raft. This test wasalso applied to study the enzyme content of hemocytesfrom two groups of five mussels of different ages (after
JOURNAL OF INVERTEBRATE PATHOLOGY 70, 96–105 (1997)ARTICLE NO. IN974670
960022-2011/97 $25.00Copyright r 1997 by Academic PressAll rights of reproduction in any form reserved.

11 and 19 months of culture, respectively, hung fromthe same raft).
Cytochemical Techniques
Light Microscopy
Hemolymph from three commercial mussels wasextracted in Alsever’s solution and pooled, on differentdates. Cytospins were prepared with 3 3 104 cells. Thehemocyte monolayers were fixed in 2.5% glutaralde-hyde in 0.1 M Pipes buffer at pH 5 7.4 for 5 min andthen incubated in specific media for each enzyme. Inthe assays of naphtholAS-D chloroacetate esterase anda-naphthyl acetate esterase, hemolymph was extracted1:4 in filtered seawater (0.22 µm) because Alsever’ssolution contains EDTA which interferes with enzymereaction (Sigma Diagnostics Procedure No. 91). For thephenoloxidase assay, hemolymph was directly with-drawn 1:1 in fixative (2.5% glutaraldehyde in 0.1 M PBSbuffer at pH 7.2).The activity of a-naphthyl acetate esterase, naphthol
AS-D chloroacetate esterase, a-naphthyl butyrate ester-ase, and b-glucuronidase was demonstrated by SigmaStaining Techniques, Procedure Nos. 181 B and 91 foresterases and Procedure No. 181 C for b-glucuronidase.For this last enzyme, three incubation times weretested: 90 min, 5 hr, and 18 hr. Controls involvedincubation in a medium without substrate.Acid phosphatase activity was revealed with Gomo-
ri’s technique (Borgers et al., 1991). The medium waspreincubated overnight prior to use. Hemocytes werethen incubated for 1 and 5 hr in this solution, stainedwith 1% ammonium sulfide in distilled water, andfinally counterstained with methylene blue. Controlsconsisted of hemocytes incubated in a medium withoutsubstrate and in a medium containing the inhibitorFNa (0.01 M).Diaminobenzidine (DAB) method (Deimann et al.,
1991) was followed to demonstrate peroxidase activity.The incubation medium consisted of 50 ml of 0.1 M
Pipes buffer (pH 6.5) containing 25 mg of DAB and 0.5ml of 1% H2O2. Hemocytes were incubated in thismedium for 10 and 45 min at room temperature indarkness. The reaction was amplified by incubation ofcells in copper nitrate for 2 min. Hemocytes were thencounterstained with light green. Controls were carriedout by incubation in media without DAB or addition ofthe inhibitors 10 mM CNK or 10 mM NaN3.The phenoloxidase assay was performed by incubat-
ing hemocytes in 0.1 M PBS (pH 7.2) containing 1mg/mlL-DOPA(Sigma) (Holden et al., 1994). Monolayers werefixed, washed in buffer, and then incubated in L-DOPAmedium for 120 min at 30°C. Samples for control wereincubated in medium without substrate.The number of reactive hemocytes was evaluated in
20 microscopic fields at 31000 in all cytochemicalassays.
Electron Microscopy
Hemolymph from 10 or more mussels was directlyextracted in fixative solutions or in buffer and pooled.
Acid Phosphatase
Hemolymphwas collected 1:1 in fixative (5% glutaral-dehyde in 0.1 M Pipes buffer, pH 7.2, 7% sucrose). Cellswere allowed to fix for 30 min and then rinsed in Pipesbuffer. Hemocytes were washed in 0.05 M acetate buffer(pH 5, 7% sucrose) and incubated for 30 min in Gomori-chloride medium at 37° (Lobo da Cunha and Azevedo,1990). Hemocytes were then washed in Pipes bufferwith sucrose (30 min) and postfixed in 1% osmiumtetroxide in Pipes buffer containing 1.5% potassiumferrocyanide for 1 hr. After dehydration, cells wereembedded in 1.5% agar (40°C) and in Epon. Controlswere carried out by adding the inhibitor 10 mM NaF tothe incubation medium.
Nonspecific Esterases
The fixative was 5% glutaraldehyde in 0.1 M cacodyl-ate buffer (pH 7.4) containing 7% sucrose. After 1 hr offixation, cells were washed twice in cacodylate bufferwithout sucrose and containing 20% dimethylsulfoxide.Hemocytes were then placed in incubation medium(Hanker, 1972) for 45 min at 4°C. At the end of theincubation process, cells were washed in cold distilledwater. The reaction was amplified by incubation of cells(20 min at 4°C) in 10 ml of acetate buffer (0.05 M, pH5.6) containing 20 mg of 3-38diaminobenzidine. Afterbeing washed in distilled water, cells were postfixed in1% osmium tetroxide and embedded in agar and inEpon. Control involved incubation of hemocytes inmedium without substrate.
Peroxidase
The fixative contained less glutaraldehyde (2.5%)than the fixative used for esterases, since a higherquantity inhibits the enzyme reaction (Deimann et al.,1991). Cells were left to fix for 1 hr and then werewashed in 0.1 M Pipes buffer (pH 7.2) with sucrose. Theenzyme activity was demonstrated using the methoddescribed for light microscopy. After 45 min of incuba-tion in darkness, hemocytes were washed in incubationbuffer (1 hr) and then rinsed in 0.1 M Pipes buffer.Finally, cells were embedded in agar and in Epon.Controls involved incubation in medium without DABand inmedium containing the inhibitors 0.01 M 3-amino-1,2,4-triazole or 0.01 M CNK.
97ENZYMES OF MUSSEL HEMOCYTES

b-Glucuronidase
Hemolymph was extracted in the same fixative as inperoxidase assay. After 1 hr of fixation, hemocytes werewashed twice in 0.1 M Pipes buffer (pH 7.2, 7% sucrose)and then incubated in the samemedium as that used todemonstrate this enzyme activity in light microscopy.The incubation was carried out at 37°C for 5 hr and 45min. Finally, the samples were washed in buffer, post-fixed, and embedded in agar and Epon. Controls in-volved incubation of hemocytes in medium withoutsubstrate.
NADH Oxidase
Cells were fixed after incubation. Hemolymph wasextracted 1:4 in 0.1 M Tris–Maleato buffer (pH 7.5, 4°C)containing 20% sucrose. Hemocytes were preincubated(10 min, 37°) in Tris–Maleato buffer containing thecatalase inhibitor 3-amino-1,2,4-triazole (0.1 M).Samples were placed in incubation medium (Briggs etal., 1986) at 37°C for 20 min. After incubation, cellswere washed in Tris buffer and then in 0.1 M Pipes (pH7.2, 4°C) with sucrose. The fixation was then carriedout in 2.5% glutaraldehyde in Pipes buffer (1 hr at 4°C).Finally, hemocytes were postfixed and embedded inagar and Epon. Samples for control were incubated inmedium without substrate.For all techniques, ultrathin sections were observed
without counterstaining through a JEOL 100 CXIItransmission electron microscope.
Lysozyme
Lysozyme activity was studied in both sera andhemocytes of two groups of mussels which have beencultivated 11 and 19 months, respectively, hung fromthe same raft. For these assays, hemolymph from threemussels of each group was extracted and pooled. Threehundred microliters of hemolymph from each groupwas diluted 1:3 in antiaggregant Alsever’s solution todetermine the number of hemocytes. The hemolymphsamples were centrifuged at 3000g for 30 min (4°C) toseparate cells and serum. The cells were subsequentlyresuspended in 2500 µl of 0.1 M phosphate buffer (pH6.4) and then sonicated 20 sec at 60 W.The enzyme activity was determined spectrophoto-
metrically according to the method of Shugar (1952)and Chu and La Peyre (1989). Serum (0.3 ml) in bufferwas added to 1.7 ml of the Micrococcus lysodeikticussuspension, and the decrease in absorbance was re-corded at 450 nm every minute using a Beckman DU 70spectrophotometer. Sonicated cells in buffer underwentthe same process. The assays were performed at roomtemperature for 4min. Themeasurements were tripled.Hen egg white lysozyme (Sigma) served as the stan-dard. Lysozyme activity was measured as a concentra-
tion of hen egg white lysozyme equivalent in micro-grams per milliliter.
Statistical Analysis
Differences in lysozyme levels between sera andplasma of the two groups of mussel of different ageswere analyzed by two-way analysis of variance, usingMINITAB Statistical Software. The factors of theANOVA were lysozyme location (serum or cells) andmussel age.
RESULTS
Api Zym Test
Mytilus galloprovincialis hemocytes showed activityof phosphatases, esterases, proteases, and glucosi-dases. Only two glucosidases (a-galactosidase and b-glu-cosidase) and two proteases (trypsin and a-chymotryp-sin) were not detected. The levels of activity of someenzymes changed through the year although no sea-sonal pattern of variation was observed (Fig. 1).The results of the comparison of hemocyte enzyme
contents between two groups of mussels of differentages are shown in Table 1. Both groups had similarenzyme levels, except for N-acetyl-b glucosaminidasewhich was higher in younger mussels, and a-glucosi-dase which was higher in older mussels.
Enzyme Cytochemistry
Cytospins
a-Naphthyl butyrate esterase activity was detectedin both hemocyte types of the mussel (Fig. 2a) (Carbal-lal et al., submitted). The 98% of hemocytes had astrong reaction for this enzyme. Hyalinocytes andgranulocytes also showed a strong reaction for a-naph-thyl acetate esterase (100% of the hemocytes) andnaphtholAS-D chloroacetate esterase (95% of the hemo-cytes) (Fig. 2b).
b-Glucuronidase activity was detected in hyalino-cytes and granulocytes (Fig. 2c). Only a few hemocytesshowed weak reaction after 90 min of incubation. Thepercentage of reacting hemocytes increased with theincrement of incubation time. Reaction deposits wereobserved in 55% of cells after 18 hr of incubation.Both hemocyte types contained reaction deposits
from acid phosphatase activity (Fig. 2d). A weak reac-tion was detected after 1 hr of incubation. Only 16% ofhemocytes showed reaction after 5 hr of incubation.Peroxidase activity was detected after 10 min of
incubation. A very strong reaction was observed in the100% of hemocytes after 45 min of incubation (Fig. 2e).The reaction was completely inhibited by NaN3 andonly partially by CNK. Slides incubated in mediumwithout DAB did not show reaction.
98 CARBALLAL ET AL.
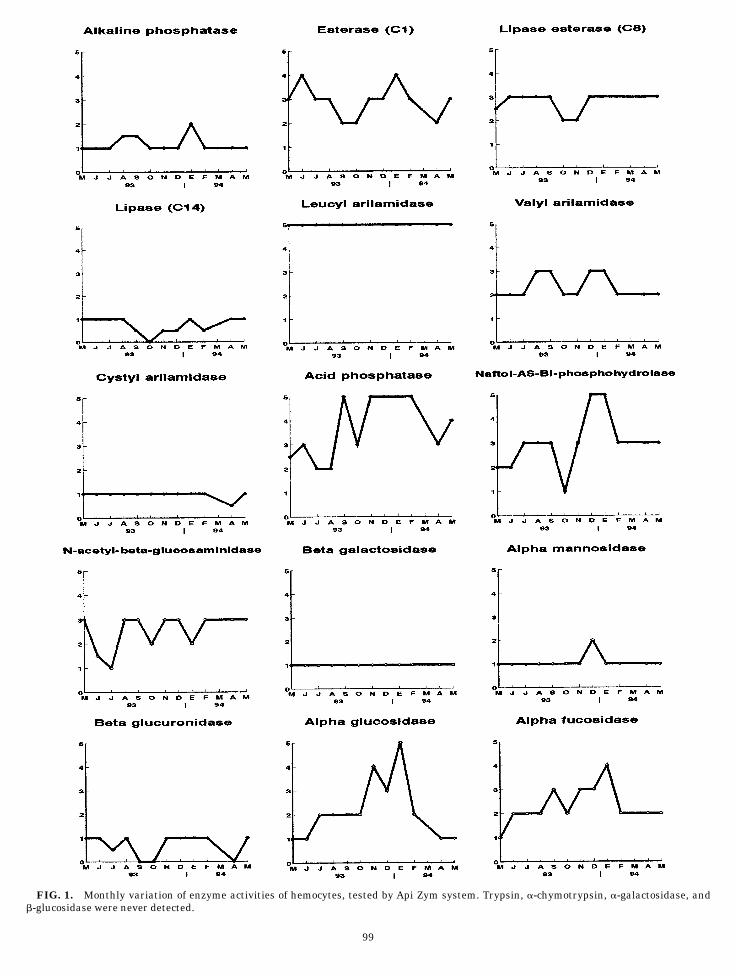
FIG. 1. Monthly variation of enzyme activities of hemocytes, tested by Api Zym system. Trypsin, a-chymotrypsin, a-galactosidase, andb-glucosidase were never detected.
99

The percentage of hemocytes containing reactionproducts from phenoloxidase activity was 73% (Fig. 2f ).The strong reaction was observed in cells of differentsizes but it was impossible to distinguish differenthemocyte types.Control slides showed no reaction deposits from any
of the enzymes.
Ultrastructural Localization of Enzymes
Acid phosphatase. The deposits of lead phosphate,which result from acid phosphatase activity, were local-ized inside cytoplasmic granules (Fig. 3a). The reactionwas observed in granulocytes containing only smallgranules (diameter ,0.5 µm) and in granulocytes con-
TABLE 1EnzymeActivity Levels of Two Groups of Mussels
of Different Ages Tested by Api Zym System
AP E EL L LA VA CA F NF G AG NG M AF
P1 1 3 3 1 5 2 1 3 2 1 1 3 1 2P2 1 3 3 1 5 2 1 3 2 1 2 2 1 2
Note. P1, mussels cultivated 11 months; P2, mussels cultivated 19months. AP, alkaline phosphatase; E, esterase (C1); LE, lipaseesterase (C8); L, lipase (C14); LA, leucyl arilamidase; VA, valylarilamidase; CA, cystyl arilamidase; P, acid phosphatase; NP, naph-thol AS-BI phosphohydrolase; G, b-galactosidase; AG, a-glucosidase;NG, N-acetyl-b-glucosaminidase; M, a-mannosidase; AF, a-fucosi-dase. The values represent the number of nanomoles of substratehydrolyzed in enzyme reaction. 0, 0 nmol; 1, 5 nmol; 2, 10 nmol; 3, 20nmol; 4, 30 nmol; 5, 40 nmol.
FIG. 2. Lightmicrographs of cytospins ofMytilus galloprovincialis hemocytes showing deposits of reaction (arrows) of several enzymes. (a)a-naphthyl butyrate esterase activity. (b) a-naphthyl acetate esterase and naphthol AS-D chloroacetate esterase activities. (c) b-Glucuroni-dase activity. (d) Acid phosphatase activity. (e) Peroxidase activity. (f ) Phenoloxidase activity.
100 CARBALLAL ET AL.
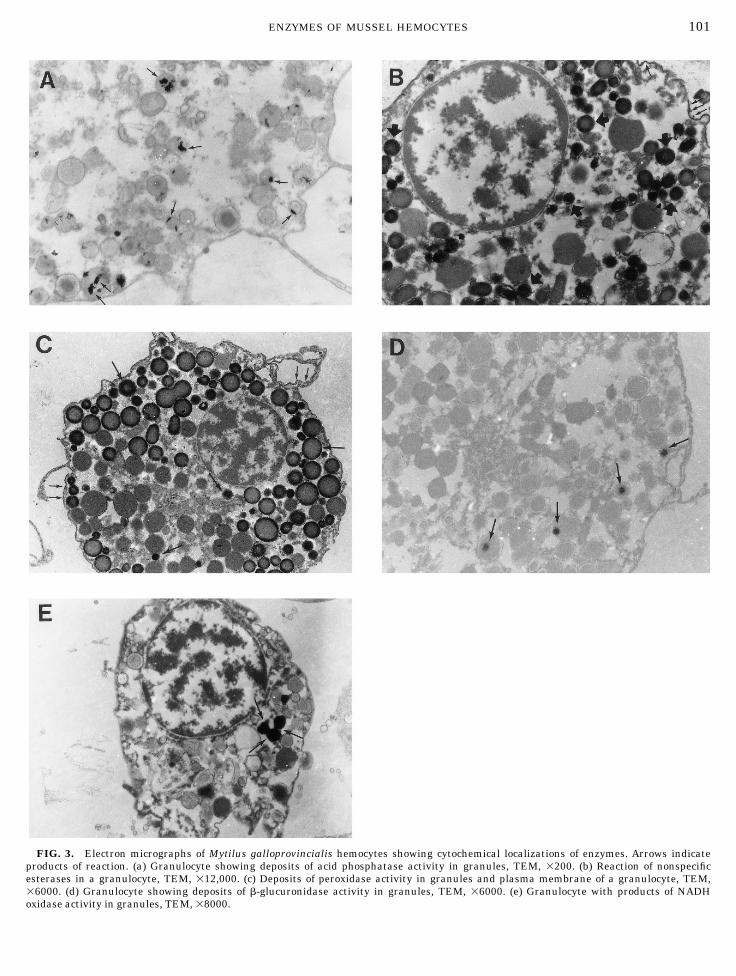
FIG. 3. Electron micrographs of Mytilus galloprovincialis hemocytes showing cytochemical localizations of enzymes. Arrows indicateproducts of reaction. (a) Granulocyte showing deposits of acid phosphatase activity in granules, TEM, 3200. (b) Reaction of nonspecificesterases in a granulocyte, TEM, 312,000. (c) Deposits of peroxidase activity in granules and plasma membrane of a granulocyte, TEM,36000. (d) Granulocyte showing deposits of b-glucuronidase activity in granules, TEM, 36000. (e) Granulocyte with products of NADHoxidase activity in granules, TEM, 38000.
101ENZYMES OF MUSSEL HEMOCYTES

taining larger granules (diameter . 0.5 µm). Few gran-ules of each hemocyte demonstrated enzyme activityand in many cases only a part of the granule containedreaction product. The hyalinocytes showed a lightenzyme activity in some of their scarce granules. Hemo-cytes incubated in medium with the inhibitor FlNa didnot have any reaction deposits.
Nonspecific esterases. The precipitates of the reac-tion were formed within the granules and in theirmembranes (Fig. 3b). Reaction deposits were also seenin plasma membranes. The reaction was observed onlyin granules whose diameters were shorter than 0.5 µmand was detected in granulocytes containing only smallgranules and in granulocytes containing larger gran-ules. The control did not show enzyme activity.
Peroxidase. The enzyme activity was observed ingranules of granulocytes. Reaction products were seenin the periphery and center of the small granules and inthe periphery of large granules (Fig. 3c). Plasma mem-brane also showed deposits of reaction. Neither thegranulocytes containing only small granules nor thehyalinocytes showed reaction. The enzyme activity wasnot detected in hemocytes incubated in medium with-out substrate. The reaction was completely inhibited byadding CNK to the medium. However, the addition ofthe catalase inhibitor 3-amino-1,2,4 triazole only slightlyinhibited the reaction.
b-Glucuronidase. Very few hemocytes showed en-zyme activity. The reaction was seen only in granulo-cytes. The deposits of reaction were localized in gran-ules whose diameter was longer than 0.3 µm (Fig. 3d).Deposits of reaction were not observed in granulocytescontaining mainly large granules. The hemocytes incu-bated in medium without substrate did not showenzyme activity.
NADH oxidase. NADH oxidase activity was presentin a few granulocytes. Deposits of reaction were seen tofill completely some cytoplasmic granules (Fig. 3e).Hyalinocytes and granulocytes containing only smallgranules did not show reaction. The enzyme activitywas not seen in cells incubated in medium withoutsubstrate.
Lysozyme. Higher levels of lysozyme activity weredetected in cells than in serum in both groups ofmussels of different ages. Lysozyme levels were higherin the group of younger mussels (Table 2). Results ofANOVA demonstrated that differences due to lysozymelocation and mussel age were significant. Interactionbetween both factors was also significant, since differ-ences between serum and cells were more pronouncedin the group of younger mussels (Table 3).
DISCUSSION
The occurrence of various hydrolytic enzymes inhemocytes of M. galloprovincialis was confirmed bycytochemical techniques. The study of monthly varia-tion of the enzymes demonstrated that the activitylevels of some of them varied throughout the year,although without a seasonal pattern.Several authors have distinguished hemocyte types
according to their enzyme content (Moore and Lowe,1977; Bayne et al., 1979; Pipe, 1990). However, inmonolayers of M. galloprovincialis hemocytes, bothhyalinocytes and granulocytes showed reaction to thesame enzymes, although the reaction was usuallylighter in hyalinocytes. It was impossible to distinguishamong granulocyte classes using these techniques, buta-naphthyl acetate esterase activity was detected in100% of hemocytes indicating that all classes of granu-locytes contain this enzyme. The deposits of phenoloxi-dase activity were observed in hemocytes of differentsizes but hyalinocytes and granulocytes could not beidentified.The ultrastructural study revealed that the granules
of M. galloprovincialis hemocytes contained acid phos-phatase, thus indicating that they are either lysosomesor some form of lysosomes (De Duve et al., 1955). Thedeposits of reaction were observed in hyalinocytes andall classes of granulocytes. The granules of hemocytesalso contain the lysosomal enzymes b-glucuronidaseand nonspecific esterase. b-Glucuronidase is consid-ered as a nonspecific humoral defense molecule, since it
TABLE 2LysozymeActivity in Serum and Hemocytes
ofM. galloprovincialis
Hemocytes Serum
Mussels cultivated 11 months 2.7268 6 0.0497 0.6272 6 0.791Musssels cultivated 19 months 0.5940 6 0.0257 0.3597 6 0.0316
Note. The enzyme activity (means 6 SD) is expressed in micro-grams/milliliter hemolymph.Although the samples of two groups hada different number of hemocytes, the values of enzyme activity werefitted and correspond to the same number of hemocytes.
TABLE 3Analysis of Variance of Lysozyme Levels of Two Groups
of Mussels of Different Ages a
Source DF SS MS F P
Locationb 1 5.0234 5.0234 979.36 0.000Age 1 4.7357 4.7357 923.28 0.000Locationb Age 1 2.0823 2.0823 405.95 0.000Error 8 0.0410 0.0051Total 11 11.8825
a Cultured 11 and 19 months, hung from same raft.b Hemocytes or serum.
102 CARBALLAL ET AL.

can hydrolyze acid mucopolysaccharides, which areconstituents of bacterial cells and teguments of hel-minths (Cheng, 1976). The ultrastructural study showedthat only the granulocytes containing granules whosediameter was higher than 0.3 µm had deposits fromreaction. Similar results were found by Pipe (1990) inM. edulis hemocytes. Nevertheless, the reaction wasobserved in granulocytes and hyalinocytes of M. gallo-provincialis in the light microscopy study.Nonspecific esterase activity was observed only in a
few granulocytes at the ultrastructural level. However,the reaction was very strong in the light microscopystudy and could be seen in both hyalinocytes andgranulocytes. The only bivalve species in which theultrastructural location of these enzymes was deter-mined were the oysters Ostrea edulis and Crassostreagigas (Chagot, 1989). Both oyster species showed depos-its from reaction in vesicles of hyalinocytes but not ingranulocytes. In the present study, the reaction wasseen in granules and also in the plasma membrane.Esterases of vertebrate blood cells have ultrastructurallocations similar to those of mussels (Borgers andVerheyan, 1985).In addition to hydrolytic enzymes,M. galloprovincia-
lis hemocytes also contain the enzymes phenoloxidase,peroxidase, and NADH oxidase. Phenoloxidase is in-volved in the prophenoloxidase system which is adefense mechanism of crustaceans and arthropods (Jo-hansson and Soderhall, 1989, 1992). Melanin, a well-known bactericidal pigment, is synthesized throughthis mechanism. Phenoloxidase activity was observedonly in hemocytes of another bivalve,M. edulis (Holdenet al., 1994; Coles and Pipe, 1994). Although the preciserole of phenoloxidase in Mytilus is unknown (Coles etal., 1994), it is possible that mussels have this cellulardefense system. In addition, melanin was found in thedeposits characterizing brown ring disease of clamswhich was interpreted as a defense reaction against theetiological agent of this disease, Vibrio P1 (Paillardet al., 1994).The deposits of peroxidase reaction were located in
granules and plasmamembrane ofM. galloprovincialisgranulocytes. Peroxidase reaction occurred in the pres-ence of the catalase inhibitor and did not in thepresence of the peroxidase inhibitor CNK, thus demon-strating that the reaction is catalyzed by peroxidase. Inaddition, the incubation of hemocytes was carried outat pH 6.5 which stimulated this enzyme activity(Deimann et al., 1991). Although the reaction wasobserved in hyalinocytes and all classes of granulocytesunder light microscopy, these results should be inter-preted carefully since the reaction was not completelyinhibited by CNK and interference by other enzymesmight have happened.Ultrastructural localization of NADH oxidase activ-
ity was studied only in hemocytes of the bivalves
Crassostrea gigas and Ostrea edulis (Chagot, 1989).The deposits of reaction were seen in the plasmamembranes of both species. In the present study, thereaction was observed in granules but not in theplasma membrane. In human leukocytes and phago-cytic cells of mammals, this reaction was also producedin plasma membrane, phagosomes, and vesicles (Borg-ers and Verheyen, 1985). This enzyme and also peroxi-dase participate in the oxidative metabolism, a cyto-toxic mechanism coupled to phagocytosis in vertebratesand some bivalves (Nakamura et al., 1985; Bachere etal., 1991; Le Gall et al., 1991; Friedl et al., 1992;Anderson et al., 1992; Pipe, 1992; Noel et al., 1993). Theoccurrence of the oxidative metabolism has also beenfound in M. galloprovincialis hemocytes (Carballalet al., in prep.).The cytochemical study of hemocytes revealed that
not all granules within the same cell were reactive.This can be explained by deficiencies in techniquesused, or as stated by Yoshino and Cheng (1976), thegranules are chemically heterogeneous or there is anonsynchronized chemical cycle which occurs in gran-ules. Also, enzyme reaction was not observed in allhemocytes which suggests that hemocytes are in adifferent state of enzyme synthesis or there are differ-ences among hemocyte subpopulations.Lysozyme is a lysosomal enzyme with an important
defense role, since it can hydrolyze components ofbacterial walls and also participate in digestion, be-cause bacteria are a part of the diet of many marinemolluscs (Cheng, 1983). Lysozyme activity has beendetected in the hemolymph of many molluscs (McDadeand Tripp, 1967; Feng and Canzonier, 1970; Cheng andRodrick, 1974; Cheng et al., 1978; Pickwell and Stein-ert, 1984; Chu and La Peyre, 1989, 1993; Pipe, 1990)and also in serum of M. galloprovincialis (Santaremet al., 1994). The results of our study revealed thatlysozyme activity was higher in hemocytes than inserum of this mussel species. Cheng et al. (1974, 1975)found similar results in Biomphalaria glabrata andMercenaria mercenaria. However, the levels of lyso-zyme activity were higher in serum than in hemocytesin both Crassostrea virginica and Mya arenaria (Ro-drick and Cheng, 1974). These differences in resultsmay have been caused by the different physiological orpathological stage of animals. According to our results,determination of lysozyme levels would be more accu-rate from hemocytes or total hemolymph than fromserum.There are few studies of enzyme levels in mollusc
hemocytes of different ages. Cheng et al. (1978) de-tected that the aminopeptidase level in serum of B.glabrata increased with age, and Dikkeboom et al.(1984) found that hemocytes of young snails Lymnaeastagnalis had lower peroxidase levels than adults. Theresults of the present study demonstrate that the
103ENZYMES OF MUSSEL HEMOCYTES

enzyme levels of hemocytes of two adult groups M.galloprovincialis do not change significantly with ageexcept for lysozyme which shows a higher level in theyounger mussels. Several authors have found seasonalvariation of lysozyme levels and associate this varia-tion with changes in the gonad cycle (Feng and Canzo-nier, 1970; Chu and La Peyre, 1989; Santarem et al.,1994). In our case, both younger and older mussels hadvery probably ripe gonad, since were collected in March(Villalba, 1995).
ACKNOWLEDGMENTS
We are grateful to Ms. Laura Corral, Ms. Elena Penas, Ms. MarıaIsabel Melendez, Ms. Veronica Ferro, andMs. Eulalia Fernandez whoprovided technical assistance. We thank Ms. Milagros Madrinan andthe crew of the ‘‘Gaivotina’’ for sampling the mussels and Mr. LuisLosada who supplied the mussels.
REFERENCES
Anderson, R., Oliver, L. M., and Brubacher, L. L. 1992. Superoxideanion generation by Crassostrea virginica hemocytes as measuredby nitroblue tetrazolium reduction. J. Invertebr. Pathol. 59, 303–307.
Auffret, M. 1989. Comparative study of the hemocytes of two oysterspecies: The european flat oyster, Ostrea edulis, Linnaeus, 1750and the Pacific Oyster, Crassostrea gigas (Thunberg, 1793). J.Shellfish Res. 2, 367–373.
Bachere, E., Chagot, D., and Grizel, H. 1988. Separation of Cras-sostrea gigas hemocytes by density gradient centrifugation andcounterflow centrifugal elutriation. Dev. Comp. Immunol. 14,261–268.
Bachere, E., Hervio, D., and Mialhe, E. 1991. Luminol-dependentchemiluminescence by hemocytes of two marine bivalves, Ostreaedulis and Crassostrea gigas. Dis. Aquat. Org. 11, 173–180.
Bayne, C. J., Moore, M. N., Carefoot, T. H., and Thompson, R. J. 1979.Hemolymph functions inMytilus californianus: The cytochemistryof hemocytes and their responses to foreign implants and hemo-lymph factors in phagocytosis. J. Invertebr. Pathol. 34, 1–20.
Borgers, M., and Verheyen, A. 1985. Enzyme cytochemistry. Int. Rev.Cytol. 95, 163–227.
Borgers, M., Firth, J. A., Stoward, P. J., and Verheyen, A. 1991.Histochemical methods for phosphatases. In ‘‘Histochemistry’’ (Sto-ward and Pearse, Eds.), Vol. 3, 4th ed., pp. 591–606. ChurchillLivingstone, NewYork.
Briggs, R. T., Robinson, J. M., Karnovsky, M. L., and Karnovsky, M. J.1986. Superoxide production by polymorphonuclear leucocytes. Acytochemical approach.Histochemistry 84, 371–378.
Chagot, D. 1989. ‘‘Caracterisation morphologique et fonctionelle deshemocytes d’Ostrea edulis et de Crassostrea gigas, mollusquesbivalves. Etude in vitro de leurs interactions avec le protozoaireBonamia ostreae (Ascetospora).’’ These de l’Ecole Pratique desHautes Etudes, Montpellier.
Cheng, T. C., and Rodrick, G. E. 1974. Identification and characteriza-tion of lysozyme from the hemolymph of the softshelled clam,Mya arenaria. Biol. Bull. 147, 311–320.
Cheng, T. C., Rodrick, G. E., Foley, D. A., and Koehler, S. A. 1975.Release of lysozyme from hemolymph cells of Mercenaria merce-naria during phagocytosis. J. Invertebr. Pathol. 25, 261–265.
Cheng, T. C. 1976. Beta-glucuronidase in the serum and hemolymphcells of Mercenaria mercenaria and Crassostrea virginica (Mol-lusca: Pelecypoda). J. Invertebr. Pathol. 27, 125–128.
Cheng, T. C., Guida, V. G., and Gerhart, P. L. 1978. Aminopeptidaseand lysozyme activity levels and serum protein concentrations inBiomphalaria glabrata (Mollusca) challenged with bacteria. J.Invertebr. Pathol. 32, 204–210.
Cheng, T. C. 1981. Bivalves. In ‘‘Invertebrate Blood Cells, Vol. 2’’(N. A. Ratcliffe and A. F. Rowley, Eds.), pp. 231–300. AcademicPress, London.
Cheng, T. C. 1983. The role of lysosomes in molluscan inflamation.Am. Zool. 23, 129–144.
Chu, F. L. E., and La Peyre, J. F. 1989. Effect of environmental factorsand parasitism on hemolymph lysozyme and protein of Americanoysters (Crassostrea virginica). J. Invertebr. Pathol. 54, 224–232.
Chu, F. L. E., and La Peyre, J. F. 1993. Perkinsusmarinus susceptibil-ity and defense-related activities in eastern oysters Crassostreavirginica: Temperature effects.Dis. Aquat. Org. 16, 223–234.
Coles, J. A., and Pipe, R. K. 1994. Phenoloxidase activity in thehaemolymph and haemocytes of the marine musselMytilus edulis.Fish Shellfish Immunol. 4, 337–352.
De Duve, C. B., Pressman, B. C., Gianetto, R., Waltiaux, R., andApplemans, F. 1955. Tissue fractionation studies 6. Intracellulardistribution patterns of enzymes in rat-liver tissue. Biochem. J. 60,604–617.
Deimann,W.,Angermuller, S., Stoward, P. J., and Fahimi, H. D. 1991.Peroxidases. In ‘‘Histochemistry’’ (Stoward and Pearse, Eds.), Vol.3, 4th ed., pp. 135–159 and 573–576. Churchill Livingstone, NewYork.
Dikkeboom, R., van der Knaap, W. P. W., Meuleman, E. A., andSminia, T. 1984. Differences between blood cells of juvenile andadult specimens of the pond snail Lymnaea stagnalis. Cell TissueRes. 238, 43–47.
Dikkeboom, R., van der Knaap, W. P. W., van der Bovenkamp, W.,Tijnagel, J. M. G. H., and Bayne, C. J. 1988. The production of toxicoxygen metabolites by hemocytes of different snail species. Dev.Comp. Immunol. 12, 509–520.
Feng, S. Y., and Canzonier, W. J. 1970. Humoral responses in theAmerican oyster (Crassostrea virginica) infected withBuchephalussp. andMinchinia nelsoni. Am. Fish. Soc. Spec. Publ. 5, 479–510.
Friedl, F. E., and Alvarez, M. R. 1992. Oxidant production byhemocytes of eastern oyster, Crassostrea virginica (Gmelin). Aqua-culture 107, 125–129.
Hanker, J. S., Yates, P. E., Clapp, D. H., and Anderson, W. A. 1972.Newmethods for the demonstration of lysosomal hydrolases by theformation of osmium black.Histochemie 30, 201.
Holden, J. A., Pipe, R. K., Quaglia, A., and Ciani, G. 1994. Blood cellsof the arcid clam Scarpharca inaequivalvis. J. Mar. Biol. Ass. U. K.74, 287–299.
Huffmann, J. E., and Tripp, M. R. 1982. Cell types and hydrolyticenzymes of soft shell clam (Mya arenaria) hemocytes. J. Invertebr.Pathol. 40, 68–74.
Johansson, M. W., and Soderhall, K. 1989. Cellular immunity incrustaceans and the proPo system. Parasitol. Today 6, 171–176.
Johansson, M. W., and Soderhall, K. 1992. Cellular defense and celladhesion in crustaceans. Anim. Biol. 1, 97–107.
Le Gall, G., Bachere, E., and Mialhe, E. 1991. Chemiluminescenceanalysis of the activity of Pecten maximus hemocytes stimulatedwith zymosan and host-specific Rickettsiales-like organisms. Dis.Aquat. Org. 11, 181–186.
Lobo da Cunha, A., and Azevedo, C. 1990. Enzyme cytochemistry ofthe alveolar sacs and golgian-like cisternae in the ciliate Icthy-ophthirius multifiliis. J. Protozool. 37(3), 206–211.
McDade, J. E., and Tripp, M. R. 1967. Lysozyme in the hemolymph ofthe oyster, Crassostrea virginica. J. Inver. Pathol. 9, 531–535.
104 CARBALLAL ET AL.

Mohandas, A., Cheng, T. C., and Cheng, L. B. 1985. Mechanism oflysosomal release from Mercenaria mercenaria granulocytes: ascanning electron microscope study. J. Invert. Pathol. 46, 189–197.
Moore, M. N., and Lowe, D. M. 1977. The cytology and cytochemistryof the hemocytes ofMytilus edulis and their responses to experimen-tally injected particles. J. Invert. Pathol. 29, 18–30.
Nakamura, M., Mori, K., Inooka, S., and Nomura, T. 1985. In vitroproduction of hydrogen peroxide by the amoebocytes of the scallopPatinopecten yessoensis (Jay).Dev. Comp. Immunol. 9, 407–417.
Noel, D., Bachere, E., and Mialhe, E. 1993. Phagocytosis associatedchemiluminescence of hemocytes inMytilus edulis (Bivalvia). Dev.Comp. Immunol. 17, 483–493.
Paillard, C., Maes, P., and Oubella, R. 1994. Brown ring disease inclams. Annu. Rev. Fish Dis. 4, 219–240.
Pickwell, G. V., and Steinert, S. A. 1984. Serum biochemical andcellular responses to experimentally cupric ion challenge in mus-sels.Mar. Environ. Res. 14, 245–265.
Pipe, R. K. 1990. Hydrolytic enzymes associated with the granularhaemocytes of the marine musselMytilus edulis. Histochem. J. 22,595–603.
Pipe, R. K. 1992. Generation of reactive oxygen metabolites by thehaemocytes of the musselMytilus edulis. Dev. Comp. Immunol. 16,111–122.
Pipe, R. K., Porte, C., and Livingstone, D. R. 1993. Antioxidantenzymes associated with the blood cells and haemolymph of themusselMytilus edulis. Fish Shellfish Immunol. 3, 221–223.
Rodrick, G. E., and Cheng, T. C. 1974. Kinetic properties of lysozymefrom the hemolymph of Crassostrea virginica. J. Invertebr. Pathol.24, 41–48.
Santarem, M. M., Robledo, J. A. F., and Figueras, A. 1994. Seasonalchanges in hemocytes and serum defense factors in the blue musselMytilus galloprovincialis. Dis. Aquat. Org. 18, 217–222.
Shugar, D. 1952. Measurement of lysozyme activity and the ultravio-let inactivation of lysozyme. Biochim. Biophys. Acta 8, 302–308.
Villalba, A. 1995. Gametogenic cycle of cultured mussel, Mytilusgalloprovincialis, in the bays of Galicia (N. W. Spain). Aquaculture130, 269–277.
Yoshino, T. P., and Cheng, T. C. 1976. Fine structural localization ofacid phosphatase in granulocytes of the pelecypod Mercenariamercenaria. Trans. Amer. Micros. Soc. 95, 215–220.
105ENZYMES OF MUSSEL HEMOCYTES




















