
Pyrococcus abyssi sp. nov., a new hyperthermophilic archaeon isolated from a deep-sea hydrothermal...
12
Arch Microbiol (1993) 160:338-349 Archives of Micrnbiology 9 Springer Verlag 1993 Pyrococcus abyssi sp. nov., a new hyperthermophilic archaeon isolated from a deep-sea hydrothermal vent Ga~l Erauso ~, Anna-Louise Reysenbach z, Anne Godfroy 3, Jean-Roch Meunier 4, Byron Crump s, Frfidfiric Partensky ~, John A. Baross 5, Viggo Marteinsson 1, Georges Barbier 3, Norman R. Pace 2, Daniel Prieur 1 CNRS UPR 4601, Station Biologique, F-29680 Roscoff, France 2 Department of Biology, Indiana University, Bloomington, IN 47405, USA 3 IFREMER Centre de Brest: Environment Profond, F-29280 Plouzan~, France 4 Institut Pasteur, Unit6 des ent6robact6ries, 28 rue du docteur Roux, F-75724 Paris C6dex, France 5 School of Oceanography, University of Washington, Seattle, WA 98195, USA Received: 22 April 1993/Accepted: 6 July 1993 Abstract. A novel, hyperthermophilic, anaerobic, sulfur- metabolizing archaeon was isolated from a fluid sample from recently discovered hydrothermal vents in the North Fiji basin (SW Pacific), at 2000 m depth. The new organism, strain GE5, is a gram-negative, highly motile coccus. It grows between 67 ~ and 102 ~ under atmo- spheric pressure, with an optimum at 96 ~ (doubling time 33 rain). The upper growth temperature is extended by at least 3 ~ when cells are cultivated under in situ hydrostatic pressures (20 MPa). Strain GE5 is an obligate heterotroph, fermenting peptides, or mixtures of amino acids to acetate, isovalerate, isobutyrate, propionate, H2 and CO 2. Hydrogen inhibits growth unless sulfur is present. In the presence of sulfur, HzS is then produced. Phylogenetic analyses of the 16 S rRNA sequence of strain GE5 places the new isolate within the Thermococcales. By its high growth temperature and physiological features the new isolate ressembles Pyrococcus sp. However it differs by a 7%mol upper G + C-content and shows low level of DNA similarity with the two previously described species. Based on these differences the description of strain GE5 as a new species Pyrococcus abyssi (CNCM 1-1302) is proposed. Key words: Archaea - Hyperthermophile - Hydrostatic pressure - Pyrococcus - Deep-sea vent As a result of the hydrostatic pressure at deep-sea vents, geothermally heated seawater remains liquid at tem- peratures up to 400 ~ (Spiess et al. 1980; Delaney et al. 1984). When the hot fluid enriched in polymetal sulfides and gasses, mixes with cold (2 ~ seawater, minerals precipitate and form the so-called "chimneys'" or "smokers" from which the hot fluids are emitted. Many novel hyperthermophilic archaea (Woese et al. 1990) have been isolated and described from such envi- ronments. They represent different metabolic types, pri- mary producers such as methanogens and mixotrophic Correspondence to" G. Erauso sulfate reducers, and consumers represented by he- terotrophic sulfur-metabolizers. These new isolates in- clude members of the genera Methanococcus (Jones et al. 1983; Zhao etal. 1988; Burggraf etal. 1990a), Me- thanopyrus (Huber etal. 1989; Kurr et al. 1991), Ar- chaeoglobus (Stetter 1988; Burggraf etal. 1990b) Sta- phylothermus (Fiala et al. 1986), Desulfurococcus (Zillig et al. 1982; Jannasch et al. 1988) and Pyrodictium (Stetter etal. 1983; Pley etal. 1991). With the exception of Methanopyrus, these genera are not unique to deep-sea hydrothermal vents, but have been isolated from shallow marine or terrestrial environments too. This report describes a novel hyperthermophilic, pep- tide-fermenting archaeon, Pyrococcus abyssi, sp. nov., isolated from a newly discovered deep-sea vent with a temperature up to 296 ~ at 2000 m depth in the North Fiji Basin. Strain GE5 was selected as a type strain among 5 isolates (Godfroy et al. unpublished results). According to the 16 S rRNA sequences and DNA-DNA hybridiza- tions data the 5 isolates belong to the same species (J.-R. Meunier et al. unpublished data). The taxonomic position of the new organism, and the effects of elevated hydrostatic pressures on growth are discussed. Materials and methods Reference strains Thermococcus celer (DSM 2476), Thermococcus litoralis (DSM 5474), Thermocoecus stetteri (DSM 5262), Pyrococcus furiosus (DSM 3638), Pyrococcus woesei (DSM 3773), and Thermotoga maritima (DSM 3109) were obtained from the Deutsche Sammlung von Mikroorganismen (DSM) Braunschweig-St6ckheim, Ger- many. Culture conditions Standard medium referred to as "YPS" was used to cultivate references strains and new isolates. The mineral base used for the preparation this media consisted of the following (per liter): (A) NaC1 23.4 g, MgC12 9 6 H20 10.8 g, Na2SO 4 4 g, KC1 0.7 g, Nail CO3 0.2 g, CaC12 92H20 0.2 g, KBr 0.09 g, SrC12 ' 6H20 0.025 g, H3BO 3 0.025 g, NaF 0.003 g; (B) 10 ml of trace element solution (Balch et al. 1979); (C) 1 ml Na2WO 4 10 mM and 1 ml FeC13
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Pyrococcus abyssi sp. nov., a new hyperthermophilic archaeon isolated from a deep-sea hydrothermal...

Arch Microbiol (1993) 160:338-349 Archives of
Micrnbiology �9 Springer Verlag 1993
Pyrococcus abyssi sp. nov., a new hyperthermophilic archaeon isolated from a deep-sea hydrothermal vent
Ga~l Erauso ~, Anna-Louise Reysenbach z, Anne Godfroy 3, Jean-Roch Meunier 4, Byron Crump s, Frfidfiric Partensky ~, John A. Baross 5, Viggo Marteinsson 1, Georges Barbier 3, Norman R. Pace 2, Daniel Prieur
1 CNRS UPR 4601, Station Biologique, F-29680 Roscoff, France 2 Department of Biology, Indiana University, Bloomington, IN 47405, USA 3 IFREMER Centre de Brest: Environment Profond, F-29280 Plouzan~, France 4 Institut Pasteur, Unit6 des ent6robact6ries, 28 rue du docteur Roux, F-75724 Paris C6dex, France 5 School of Oceanography, University of Washington, Seattle, WA 98195, USA
Received: 22 April 1993/Accepted: 6 July 1993
Abstract. A novel, hyperthermophilic, anaerobic, sulfur- metabolizing archaeon was isolated from a fluid sample from recently discovered hydrothermal vents in the North Fiji basin (SW Pacific), at 2000 m depth. The new organism, strain GE5, is a gram-negative, highly motile coccus. It grows between 67 ~ and 102 ~ under atmo- spheric pressure, with an optimum at 96 ~ (doubling time 33 rain). The upper growth temperature is extended by at least 3 ~ when cells are cultivated under in situ hydrostatic pressures (20 MPa). Strain GE5 is an obligate heterotroph, fermenting peptides, or mixtures of amino acids to acetate, isovalerate, isobutyrate, propionate, H2 and CO 2. Hydrogen inhibits growth unless sulfur is present. In the presence of sulfur, HzS is then produced. Phylogenetic analyses of the 16 S rRNA sequence of strain GE5 places the new isolate within the Thermococcales. By its high growth temperature and physiological features the new isolate ressembles Pyrococcus sp. However it differs by a 7%mol upper G + C-content and shows low level of DNA similarity with the two previously described species. Based on these differences the description of strain GE5 as a new species Pyrococcus abyssi (CNCM 1-1302) is proposed.
Key words: Archaea - Hyperthermophile - Hydrostatic pressure - Pyrococcus - Deep-sea vent
As a result of the hydrostatic pressure at deep-sea vents, geothermally heated seawater remains liquid at tem- peratures up to 400 ~ (Spiess et al. 1980; Delaney et al. 1984). When the hot fluid enriched in polymetal sulfides and gasses, mixes with cold (2 ~ seawater, minerals precipitate and form the so-called "chimneys'" or "smokers" from which the hot fluids are emitted. Many novel hyperthermophilic archaea (Woese et al. 1990) have been isolated and described from such envi- ronments. They represent different metabolic types, pri- mary producers such as methanogens and mixotrophic
Correspondence to" G. Erauso
sulfate reducers, and consumers represented by he- terotrophic sulfur-metabolizers. These new isolates in- clude members of the genera Methanococcus (Jones et al. 1983; Zhao etal. 1988; Burggraf etal. 1990a), Me- thanopyrus (Huber etal. 1989; Kurr et al. 1991), Ar- chaeoglobus (Stetter 1988; Burggraf etal. 1990b) Sta- phylothermus (Fiala et al. 1986), Desulfurococcus (Zillig et al. 1982; Jannasch et al. 1988) and Pyrodictium (Stetter etal. 1983; Pley etal. 1991). With the exception of Methanopyrus, these genera are not unique to deep-sea hydrothermal vents, but have been isolated from shallow marine or terrestrial environments too.
This report describes a novel hyperthermophilic, pep- tide-fermenting archaeon, Pyrococcus abyssi, sp. nov., isolated from a newly discovered deep-sea vent with a temperature up to 296 ~ at 2000 m depth in the North Fiji Basin. Strain GE5 was selected as a type strain among 5 isolates (Godfroy et al. unpublished results). According to the 16 S rRNA sequences and DNA-DNA hybridiza- tions data the 5 isolates belong to the same species ( J . -R. Meunier et al. unpublished data). The taxonomic position of the new organism, and the effects of elevated hydrostatic pressures on growth are discussed.
Materials and methods
Reference strains
Thermococcus celer (DSM 2476), Thermococcus litoralis (DSM 5474), Thermocoecus stetteri (DSM 5262), Pyrococcus furiosus (DSM 3638), Pyrococcus woesei (DSM 3773), and Thermotoga maritima (DSM 3109) were obtained from the Deutsche Sammlung von Mikroorganismen (DSM) Braunschweig-St6ckheim, Ger- many.
Culture conditions
Standard medium referred to as "YPS" was used to cultivate references strains and new isolates. The mineral base used for the preparation this media consisted of the following (per liter): (A) NaC1 23.4 g, MgC12 �9 6 H20 10.8 g, Na2SO 4 4 g, KC1 0.7 g, N a i l CO3 0.2 g, CaC12 �9 2 H 2 0 0.2 g, KBr 0.09 g, SrC12 ' 6 H 2 0 0.025 g, H3BO 3 0.025 g, NaF 0.003 g; (B) 10 ml of trace element solution (Balch et al. 1979); (C) 1 ml Na2WO 4 10 mM and 1 ml FeC13

339
25 raM; (D) 5 ml KH2PO 4 7% (w/v) and 5 ml of NH4C1 14% (w/v). B and C were filter sterilized and added to cooled autoclaved A + D. The YPS medium was obtained by adding Bactoyeast extract 0.1% (w/v), Bactopeptone 0.4% (w/v) (Difco), and sulfur (0.5%, steam sterilized at 100 ~ 1 h, on three successive days) to the mineral base. Resazurin (0.1 nag/l) was used as a redox indicator. Unless otherwise specified, the medium was buffered at pH 6.8 with PIPES (10 raM). Medium was dispensed into sterile vials and air was evacuated and replaced by Nz by alternately applying a vacuum and saturating with gas (Balch and Wolfe 1976). Final anaerobiosis was achieved by adding neutral sterile N a 2 S ' 9 H 2 0 to a final concentration of 0.025%. Isolate GE5 was plated on YPS medium solidified by Gelrite (Kelco Div. of Merck and Co., San Daego, Calif., USA) 14 g/1. Plates were incubated in an anaerobic jar at 95 ~ (gas phase Nz/H2/CO 2 90:5: 5; 100 kPa). Sulfur-free reduced medium "YP" was prepared by omitting elemental sulfur and Na2S and by the adding titanium (Ti III) citrate to 1 mM final concentra- tion as a reductant (Zehnder and Wuhrmann 1976). Influence of the gas phase on growth was determined by using the following headspaces at an initaal pressure of 200 kPa: COa, H2/CO2 (80 : 20), N 2 / C O 2 (80 : 20), N 2 / H 2 (80 : 20).
Storage
Cultures in exponential growth phase were stored at 4 ~ after the gas phase had been briefly flushed with N ; in order to remove the H2S produced during growth. These cultures could serve as an moculum for at least 1 year. For long term storage, pure cultures were stored anaerobically at - 80 ~ in culture medium containing 20% (w/v) glycerol.
Cell counts
Bacterial growth was measured either by microscopy or by flow cytometry. For microscopy, samples of cultures were fixed with 0.5% (w/v) neutralized glutaraldehyde. Appropriate dilutions (ma- ximum 10 -3) were made using the mineral base filtered onto a 0.2 gm filter cartridge (Mediacap, Microgon, Calif., USA). Then cells were stained with 0.01% acridine orange (Hobbie et al. 1977), filtered onto 0.2 l~m polycarbonate membrane filters (Nuclepore. Pleasanton, Calif., USA) and counted using an Olympus (Tokyo, Japan) BH-2 epifluorescent microscope.
For flow cytometry, bacterial samples were fixed with 1% (w/v) glutaraldehyde for 10 rain at ambient temperature, then either stored at 4 ~ in the dark or frozen in liquid nitrogen (Vaulot et al. 1989) and stored at - 8 0 ~ until analysis. After thawing, cell DNA was stained for at least 15rain at room temperature with DAPI (4',6-diamino-phenylindole, Sigma Chemicals, St. Louis, Mo., USA) at a final concentration of 2 ~tg/ml. Cells samples were diluted 10 or 100 times and counted twice using an EPICS 541 flow cytometer. UV excltatlon light was provided by a 6-W argon laser (Coherent, Palo Alto, Cahf., USA) at 500 mW. Scattered hght was measured at forward (FALS) and right (RALS) angles. DAPI-DNA blue fluorescence was measured through a 485 nm band-pass filter (Glen Spectra, Sanmore, UK). All three signals were recorded in integral mode on 3-decade logarithmac scales and bacteria were counted on a DAPI-DNA fluorescence histogram after gating on a FALS vs RALS cytogram. A good correlation was found between microscopic and flow cytometric counts. Growth rates were calculated using hnear regression anlysis from 3 to 5 points along the logarithmic part of the resulting growth curves.
Determination of growth parameters
Cultures were grown in 100-ml serum bottles containing 40 ml of medium, sealed with black butyl rubber stoppers (Bellco, Vineland, NJ, USA). The Nz gas phase was passed through a cartridge of cold catalyst to remove any traces of oxygen. An initial gas pressure of 200 kPa was applied to the head space to prevent boiling of the medium at temperature above 100 ~ and was renewed after each
sampling. Cultures were incubated in a rotary shaker (200 rpm) water bath (Bioblock, Illkirch, France) filled wath dimethyl silicone oil (Prolabo, Paris. France). Temperature was monitored with an electronic thermometer (Cole-Parmer, Chicago, Ill., USA) and precision thermocouples (type K) directly inserted into the culture vial through the stopper. No significative differences were observed between the temperature m the oil bath and the actual temperature m the culture. The pH and salinity growth curves were determined at 90 ~ For determination of specific growth rates at different pH values, the following chemicals (all from Sigma) were used to buffer the YPS medium, each at 10mM: pH 3.5 to 4.5, no buffer; pH 5.0 to 6.0, MES; pH 6.5 and 7.0, PIPES, pH 7.5 to 8.5, HEPES; pH 9.0, no buffer. Specific growth rates at different salt concentrations were determined using a 4 x solution (A) diluted appropriately in YPS medium.
Pressure experiments
Growth experiments under hydrostatic pressures were conducted at 20 MPa (101.325 kPa = 1 atm) and a range of temperature between 75 and 112 ~ growth at 40 MPa was also tested at 100 and 105 ~ Pressure experiments were run as described by Reysen- bach and Deming 1991. In an anaerobic chamber (Forma Scientific, Marietta, Ohio, USA), 200 ml of YP medium were inoculated with a 1% inoculum culture in exponential phase (ca. 1 x 108 cells/ml) grown at 95 ~ After mixing, 10 ml of inoculated medium were dispensed, in sterile, gas tight, glass syringes (Becton Dickson, Mountain Vies, Calif., USA) with cut barrel and plug, containing 0.1 g of sulfur. Gas phase was expelled prior to tightening the seal on each syringe. Duphcate syringes were brought to the desired pressure and temperature in preheated, stainless steel pressure vessels by using tap water as hydraulic fluid as described by Deming and Baross 1986. Growth curves were generated by decompressing one vessel (containing two syringes) per time point, according to standard procedures developed for the determination of bacterial growth rates under hydrostatic pressures (Yayanos et al. 1982; Deming et al. 1984; Yayanos 1986). Low pressure controls were run in duplicate for each temperature tested. In these cases YPS medium was inoculated and dispensed as described before, except that Hungate test tubes were used instead of syringes. Control tubes were pressurized at 200 kPa of N2 to prevent boiling at temperature above 100 ~ and incubated in a silicon oil bath; growth curves were drawn using 2 tubes per time point.
Determination of growth requirements
GE5 was grown in 27 ml Hungate tubes contaimng 10ml of medium. Tubes were inoculated with an exponentially growing culture at a final concentration of 5 x 105 cells/ml and incubated at 90 ~ Individual carbon sources were added to the mineral base medium supplemented with sulfur (0.5%) and vitamins 10 ml/1 of a filter sterilized solution (Balch et al. 1979). A mixture of the 20 amino acids each at 0.1 mM, and an amino acids mixture referred as "5 + amino acids" described by Pledger and Baross (1991) were also tested. To check for possable growth by fermentation or by other energy yielding reactions, these tests were run with and without sulfur added, and with various gas mixtures as described above. Autotrophic growth was also tested in the mineral base medium and in filtered natural sea water at pH 6.8, supplemented with vitamins (10 ml/t) and sulfur (0.5% w/v) under a H2/COz (80:20) gas phase. Selected nitrogenous compounds were tested for suitabili- ty as nitrogen sources, using ammonium-free mineral medium with 40 mM pyruvate as the carbon and energy source, and with 0.05% of yeast extract to stimulate growth. Nitrogenous compounds were added at 20 mM final concentration except for complex organics which were added at 0.2% (w/v). Electron acceptors were tested in YPS medium without Na/SO4 and sulfur, under the same conditions as described for carbon sources. A control in which sulfur 0.5% (w/v) was added with the putative electron acceptor was also used to check for possible growth inhibition.

340
Antibiotic sensitivity
The sensitivity to antibiotics was tested at 80 ~ in YPS medium at a concentration of 150 ~tg/ml. Controls were performed with an antibiotic sensitive eubacterlum, Thermotoa maritima, cultivated under the same conditions to demonstrate the efficiency of the antibiotic at the temperature of the test.
Determination of oxygen sensitivity
Duplicate cultures of GE5 were grown at 95 ~ in 120-ml serum bottles containing 40 ml YPS medium. When cultures were in the late logarithmic phase of growth (about 1 x 108 cells/ml), one culture was progressively cooled to 4 ~ while the other was maintained at 95 ~ Both cultures were then flushed vigorously with air (10 rain) until resazurin turned pink. The cultures were incubated at 4 ~ and at 95 ~ respectively. The number of viable cells was determined by serial dilutions.
Analysis of metabolic products
Samples for H2S determination were stabilized as ZnS by combining the samples with an excess of ZnSO4 in 1 N NaOH (Ingvorsen and J6rgensen 1979). Sulfide determination was carried out according to Cline (1969). Volatile fatty acids were analysed from acidified culture samples (I M HC1 final concentration) on a Carlo Erba (Rodano, Italy) 5300 gas chromatograph and a flame ionisation detector equipped with a column Carbovax (Supelco Inc., Bellefonte, USA) 58 CB (air pressure 120 kPa, carrier gas H a 40 kPa, com- bustion gas H2 80 kPa, oven temperature 80 to 200 ~ detector temperature 120 ~ injector temperature 150 ~ H 2 and COz were determined on a Carbosieve 100/200 packed column (Supelco) by a thermal conductivity detector.
Electron microscopy
Cells were harvested in early and in late log phase. The pellets were resuspended and fixed for 1 h in a ASW (buffered with PIPES 20 mM pH 7.0) containing 3% (w/v) glutaraldehyde, and post fixed for 2 h with 1% (w/v) OsO~ in the same buffer. After dehydration with increasing ethanol concentration, ceils were embedded in Spurr's resin, thin sectionned, contrasted with 1% (w/v) uranyl acetate and 1% (w/v) lead citrate (EM grade, Bio-Rad S.A., Ivry, France) and examined in a Phillips (Eindhoven, The Netherlands)
Fig. 1. Phase contrast micgrographs of strain Ge5 in mid-log phase from an agitated hquid culture. Bar represents 1 gm
model EM201 transmission electron mmroscope. For negative staining, 20 gl of bacterial suspension fixed with 2% (w/v) glutar- aldehyde in ASW was dropped on formvar/carbon coated grids (400 mesh) and stained with 1% (w/v) uranyl acetate.
Lipid analysis
Cells for hpid analysis were grown in YPS medmm at 95 ~ Lyophilized ceil material was extracted with the single-phase chloroform/methanol/water system of Bigh and Dyer (1959). The lipid-extracted residue was recovered from the aqueous phase by centrifugation. Residue consisted of ceil wall material and any mineral solids which are not soluble in chloroforme or water. The total hpid extract was fractionated into neutral lipid, glycolipid, and polar lipid fractions by silicic acid column chromatography (White et al. 1979). The glycolipid, polar lipid and residue were treated with a strong acid methanolysis to cleave the ether lipids from phosphate or saccharide head groups or from the cell wall (Hedrick et al. 1991). The liberated ether-alcohols were quantified by super- critacal fluid chromatography (SFC) (Hedrick et al. 1991). Identifica- tion was performed by comparing SFC retention times with ether lipids isolated from archaea with known lipid profiles.
Isolation of DNA
Genomic DNAs were extracted and purified by CsCl equihbrium gradient centrifugation previously described (Charbonnier et al. 1992). DNA purity was tested spectrophomemcally.
DNA base composition
The G + C content of the DNA was determined from the melting point in 0.1 • SSC according to Marmur and Doty (1962) using Escheriehia coli DNA (57% G + C, Sigma) as a standard. The direct nucleotide analysis was performed by HPLC after digesUon of the DNA with nuclease P1 as desribed by Mesbah et al. (1989), using calf thymus DNA (42% G + C; Sigma) and phage ;t (48.9% DNA; Boehringer Biochemica (Mannheim, Germany)) as standards.
16 S rRNA gene sequence analysis
16 S rDNA was amplified by the polymerase chain reaction (Saiki et al 1988) as described by Reysenbach et al. (1992). PCR products were cloned into the phagemide vectors Bluescript KS + and KS- (Stratagene, San Diego, Calif., USA) using standard methods (Sambrook et al. 1989) and sequenced by the dideoxy chain termina- tion method (Sanger et al. 1977). Two clones from two independant amplification reactions were sequenced by double-stranded se- quencing. Both strands of each clones were sequenced. The sequence was aligned on the basis of the secondary structure with a represen- tative collection of 16 S rRNAs. Phylogenetic trees were constructed using a least-squares distance-matrix method (Olsen et al. 1992). The Genbank accession number for strain GE5 is 19921.
D NA-D NA hybridizations
Hybridizations were performed in solution by the nuclease S1 method as outlined by Johnson (1985a). Genomic DNAs (0.5 p.g/ml) in TE buffer were sonicated to give uniform length-distributaon (0.4 to 1.6 kbp as estimated by electrophoresis in 1.2% agarose). The preparanons were then denatured by boiling for 10 rain and, after cooling in ice, centrifuged at 12000 • g for 15 rain to remove any particulate material. About 200 ng of each DNA preparation were used as templates for the synthesis of [c~-32P] dATP-labelled DNA (Feinberg and Vogelstein 1983) using a Boehringer-primed DNA labeling kit. Unincorporated nucleotides were removed by passing the DNA over a Sephadex (Pharmaeia- France, St. Quentin, Yvelines, France) G-50 spun column (Sam- brook et al. 1989). Random priming may produce large quantities of "snap-back" DNA, and the labelled DNA was denaturated by

boiling for 5 rain and then aItowed to incubate at 60 ~ for 30 mm to allow snap-back renaturation (Wermann et al. 1990). The DNA was run over a hydroxyapatite column (Bio-Rad) to remove the snap-back DNA, which constitutes 20-30% of the labelled DNA (Grimont et al. 1980). The eluted DNA was then concentrated using n-butanol, and desalted by passing over a Sephadex G-50 spun column. Approximatively 5 n g of each labelled DNA ("tracer") were
341
hybridized with 10 btg of each unlabelled ("driver") DNA. hybridiza- tions were carried out in 0.8 M NaC1, 1 mM HEPES, pH 7 (final volume of 50 gl) in 1.5 ml microtubes with O-ringed screw-caps (Sarsted, Nfimbrecht, Germany). A few drops of mineral oil were added to delay evaporation. Tubes were incubated for 60 h. The temperature of hybridization chosen (65 ~ was in the optimal range: Tm- 25 ~ to Tm- 30 ~ (Johnson 1985b; Ivanova et al. 1988) in the hybridization buffer. After nuclease St digesnon, the radioactivity of the trichloroacetic precipitable fraction was collec- ted on GF/F glass filters (Whatman, Maidstone, UK) and counted m a Hewlett-Packard, Cupertino, Calif., USA 1500 Tri-Carb liquid scintillation counter. The degree of homology for each pair was calculated (Grimont et al. 1980) by determining the ratio between the input counts in the nuclease-treated (duplicates) and the nuclease-untreated (duplicatges) samples. Results were then normal- lzed to the homologous reaction, after the percentage of S 1-resistant material of the control tubes ("tracer" DNA with 10 ~tg of sheared native salmon sperm DNA) was substracted.
Fig. 2a-e. Ultrathin section of strain GE5. a Shape and size variability; b double S-layers; c paracrystalline structure in the cytoplasm; d dividing cell: "diploform", and clumps of globular particles; e striated bundles of filaments. Bars, a, d: 1 gin; b, c, e. 0.1 um
Results
Collection of samples
In 1989, during the French-Japanese cruise "Starmer", new hydrothermal vents were discovered by the sub- mersible "Nauti le" at 2000 m depth in the Nor th Fiji Basin (173 5'E; 1 59'S). The macrofauna associated with the vents appeared different f rom those known in the eastern Pacific or the Atlantic, and mainly consisted of gastropods (Jollivet etal . 1989; Debruy6res etal . in press). The active chimneys were composed almost exclu- sively of anhydrite (Grimaud et al. 1991). Venting hot fluids (maximum temperature recorded: 296 ~ were sampled with titanium syringes. Pieces of active hydro- thermal chimneys and rock debris were collected by the submersible and brought to the surface in an insulated box. Aboard the mother ship R. V. "Nadi r" solid samples were immediately transferred into 100 ml serum bottles filled with 50 ml degassed sterile ASW, capped with black butyl rubber stoppers and aluminium seals, flushed with N 2 and then vigorously shaken. Anaerobiosis was achie- ved by adding 1 ml of a 2.5% (w/v) solution of NazS. 9 H 2 0 ; resazurin 0.001% was used as redox indicator.
Enrichments and isolation
Samples (5 ml) of rock suspensions and o fhydro the rmal fluids were transferred into 50-ml serum bottles contai- ning 20ml of SME medium (Stetter et al. 1983) supplenqented with 0.1% yeast extract and 0.4% peptone under N 2 as the gas phase (200 kPa). Cultures were incubated in an oven at 85 ~ and 95 ~ After 2 to 5 days, numerous enrichment cultures were turbid, consis- ting of a mixed populat ion of rods and cocci. Only the coccoid cells could be successfully subculture& Cultures were stored at 4 - 5 ~
Since better growth occurred in this medium, YPS medium was selected for further investigations one a part of culture collection. Twenty isolates were purified by 5 series of serial dilutions (1 : 10) in YPS medium and by streaking on "Gelri te" plates. After 3 to 4 days of incubation at 90 ~ small (about 1 m m in diameter) ochre colonies formed for several isolates. One colony served

342
as inoculum for a liquid culture of the pure strain. During the purification step, isolate GE5 (obtained from a fluid sample having an estimated temperature of 296 ~ showed the fastest growth (up to dilution 10 .8 on overnight incubation in serial dilutions) and was studied in detail.
Morphology and ultrastructure
Light microscope observation of viable cells showed highly motile (only when slide the was heated at 80-90 ~ regular to slightly irregular cocci of 0.8 to 2 gm in width (Fig. 1) with a polar tuft of flagella (not shown). In exponentially growing cultures, they were usually elongated and often variably constricted. In static cultures, GE5 forms aggregates of up to 100 cells, often attached to sulfur particles. Atypical forms in size and shape, especially "giant cells" sometimes more than 5 ~tm in diameter and with low phase contrast, occurred under specific conditions that limit growth, such as temperature, salinity, or pH limits of the growth range, and nutrient limitation (Fig. 2a). Gram staining reaction was negative. Thin sections showed a bilayered cell envelope which tightly fits to the cytoplasmic membrane (Fig. 2b). The two layers, each about 15 nm wide, were separated by an electron dense layer about 5 nm wide. Cells divided most likely by constriction, as indicated in Fig. 2d. However, budding cells have also been observed in growing cultures (not shown). A paracrystalline structure 70 nm in diameter was sometimes observed in the cyto- plasm (Fig. 2c). Dense clumps of small spherical particles (Fig. 2d), and sometimes bundles of striated fibers are present (Fig. 2e). These two peculiar structures have been also observed in thin sections of Pyrococcus woesei (Zillig et al. 1987).
Effect of pH and salt contrention
Strain GE5 grew within a pH range of pH 4.0 to 8.5 with an optimum around pH 6.8 (Fig. 3a). GE5 was found to require salt for growth. No growth occurred below 0.7% or above 5% NaC1, and optimum was 3% NaC1 (Fig. 3b).
Growth temperature and influence of hydrostatic pressures
Under 200 kPa of N2, strain GE5 grew between 67 ~ and 102 ~ with an optimum at 96 ~ (doubling time 33 min; Fig. 3 c). Shaking (200 rpm) during incubation of cultures resulted in a 1.2-1.6 times increase of growth rates, but the final cell yields were unaffected. Attempts to cultivate strain GE5 at temperatures above 102 ~ invariably failed, either in shaking or quiescent conditions. Specific growth rates determined at different pressures and a range of temperatures are presented in Table 1. It appeared that elevated hydrostatic pressures increased the maximal growth temperature of strain GE5 by at least 3 ~ i.e. from 102 ~ (determined under about 2 atm of N2) to 105 ~ The optimal growth temperature was also shifted upwards in a similar range (from 96 to 100 ~ Significant growth was observed at 105 ~ under 20 and 40 MPa
(about 1 h doubling time and final cell densities above 108 cells/ml). In an attempt to determine the actual temperature limit for growth, we also tested GE5 for growth at 107, 108, 109, 111 and 112 ~ under in situ hydrostatic pressure (20 MPa). Weak growth was repea-
1.4
1.2
1
0.8
0.6
0.4
0.2
0
a o
/
4 5 6 7 8 pH
1.4
1.2
1
0.8
0.6 ~o
0.4
r 0.2
b
1 2 3 4 5 6
NaCt concentration (%)
1.4.
1.2 C
1 t
0.8
0.6
0.4
0.20 I~_
60 70 80 90 100 110 Temperature (~
Fig. 3a-c . Temperature, pH, and NaC1 optima for growth of isolate GE5 cultivated on YPS medium with agitation (200 rpm). Specific growth rates (calculated as described in 'Material and methods' as a function of pH a, sahnlty b, and temperature c

Table 1. Effect of temperature and hydro- static pressure on the specific growth rate (#) of strain GE5 and ratios of the specific growth rates determined at high hydrostatic pressures and low gas pressure (0.2 MPa N2)
343
Temperature Specific growth rate" Ix (h-1) at Ratio Ratio (~ different pressures (in MPa) #20MPa/gO.2MPa fi4OMPa/]~O,2MPa
0.2 20 40
75 0.10 _+ 0.03 0.23 nd 2.30 nd 80 0.25 _+ 0.04 0.32 nd 1.28 nd 85 0.41 _+ 0.03 0.43 nd 1.04 nd 90 0.56 +_ 0.10 0.63 + 0.06 nd 1.12 nd 95 0.82 _+ 0.05 0.87 + 0.07 nd 1.06 nd
100 0.71 • 0.04 0.90 ___ 0.09 0.98 _+ 0.08 1.27 1.38 105 ng 0 74 +_ 0.11 0.70 108 ng go go
a vatues of specific growth rates are means _+ SD for 2 separate experiments when available. nd, not determined; ng, no growth; go, growth observed by epiftuorescence microscopy, growth rate not determined
tedly observed at 107-108 ~ The presence of numerous dividing cells suggested that the cultures were in the exponential growth phase. No growth was detected at 109 ~ and higher temperatures tested.
Nutritional requirements
Strain GE5 appears to be an obligate heterotroph. No growth was detected in mineral media amended with vitamins and with a H2 /CO 2 (80:20) headspace, with or without elemental sulfur. Maximum growth rates and cell yields were achieved in the presence of S ~ on a combination of yeast extract (0.l% w/v) with either peptone, tryptone, or casein hydrolysate (each at 0.4% w/v). Under optimal conditions (pH 6.8, 96 ~ with these combinations, the final cell density was over 7 x 108 cells/ml after 12 h.
Good growth was also observed when yeast extract, peptone, tryptone, casein hydrolysate, beef extract, brain heart infusion and casamino acids, when tested in- dividually. No growth was detected on carbohydrates, alcohols, organic acids, and individual amino acids, with or without sulfur, nor did they stimulate growth in presence of 0.1% (w/v) yeast extract (Table 2). No effect was observed with different gases (N2, Nz/H2, H2/C02, CO2). High growth yields (up to 5 x l0 s cells/ml) were observed on a mixture of the 20 natural amino-acids at 0.1 mM each and on the "5+ amino-acids mixture" described by Pledger and Baross (1991) if a vitamin solution (Balch et al. 1979) wad added to the medium. The initial cultures in the amino-acids mixtures, resulted in a very long lag phase (5 to 11 h), with a generation time of 4 to 7 h. However, after several transfers in these "amino acids media" lag phase and generation time were both reduced to about 2-3 h.
Growth on 40 mM pyruvate was poor (1 x 107 cells/ml final density), but was stimulated 10- to 20-fold in presence of 0.05% yeast extract. This concentration of yeast extract in the absence of pyruvate was found to support only poor (5 x 106 cells/m1, final density) growth. This combination of pyruvate and yeast extract was utilized in the ammonium-free medium used for testing different nitrogen sources. The results (Table 2) indicated that strain GE5 was able to grow equally well on different
protein hydrolysates, amino acid mixtures, glutamate or ammonium as nitrogen sources. Tryptone stimulated growth. NO 2 was found to inhibit growth.
Influence of sulfur compounds
Addition of elemental sulfur stimulated growth (10% increase of the final cell yield at 90 ~ and about 60% decrease in the doubling time) but was not absolutely required. When sulfur was present in the medium, the gas phase did not influence growth. However, in a sulfur-free medium, with a H2 /CO 2 (80:20, 200 kPa) headspace, no growth was observed (Table 2), thus indicating a strict sulfur-dependence for growth in pre- sence of H 2.
Of the alternate electron acceptors tested, strain GE5 used cystine and polysulfides (Table 2). Growth of GE5
1 0 c"
1 0 ~
I o -j
1 0 E
0
I I I I ( I
2 4 6 8 10 12
Time (h )
8
7
6
5
co
E 3
2
1
0
14
Fig. 4. Growth and sulfide production of strain GE5, cultivated at 90 ~ with agitation (200 rpm). (A) growth without S~ (*) growth with S~ (e) H2S production in YPS medium; (A) production in uninoculated control
344
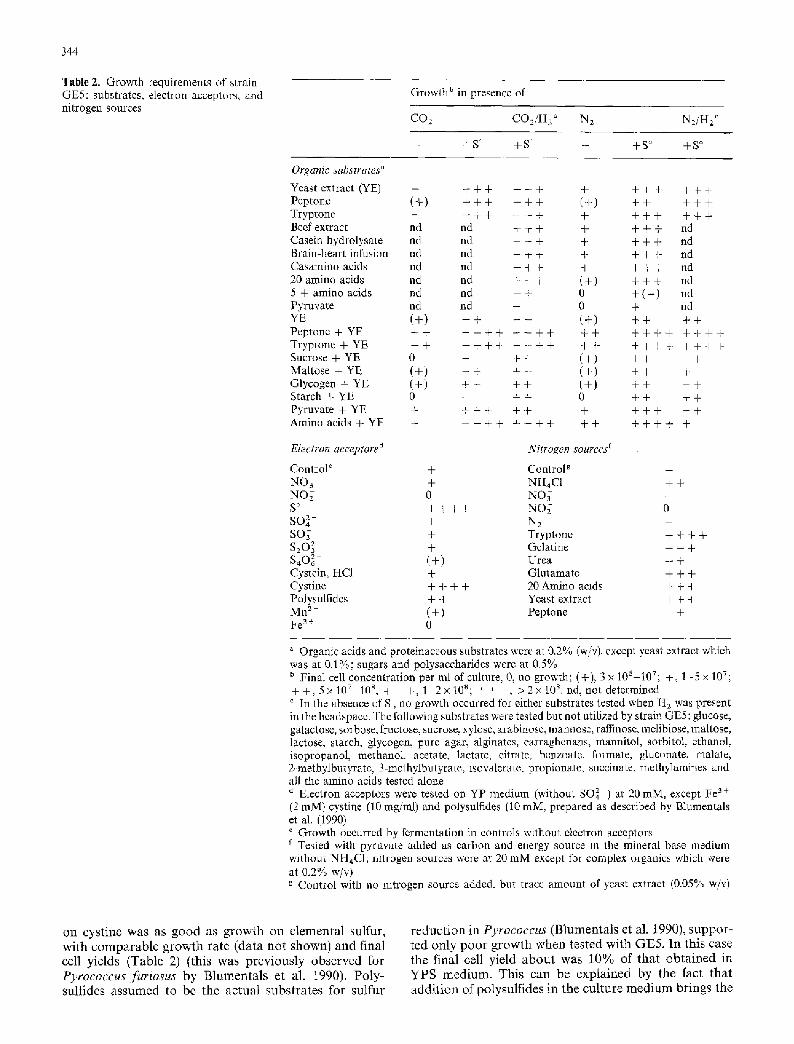
Table 2. Growth requirements of strain GE5: substrates, electron acceptors, and nitrogen sources
Growth b in presence of
CO2 CO2/H2 ~ N 2 N 2 / H 2 c
+S ' +S ' - +S ~ +S ~
Organic substrates a
Yeast extract (YE) + + + + + + + + + + + + + + Peptone (+) + + + + + + (+) + + + + + Tryptone + + + + + + + + + + + + + + Beef extract nd nd + + + + + + + nd Casein hydrolysate nd nd + + + + + + + nd Brain-heart infusion nd nd + + + + + + + nd Casamino acids nd nd + + + + + + + nd 20 amino acids nd nd + + + (+) + + + nd 5 + amino acids nd nd + + 0 + (+) nd Pyruvate nd nd + 0 + nd YE (+) + + + + (+) + + + + P e p t o n e + Y E + + + + + + + + + + + + + + + + + + + + T r y p t o n e + Y E + + + + + + + + + + + + + + + + + + + + Sucrose + YE 0 + + + + (+) + + + + Maltose + YE (+) + + + + (+) + + + Glycogen + YE (+) + + + + (+) + + + + Starch + YE 0 + + + 0 + + + + P y r u v a t e + Y E + + + + + + + + + + + + A m i n o a c i d s + Y E + + + + + + + + + + + + + + + +
Electron acceptors a Nitrogen sources r
ControP + Control g + NO~ + NH~C1 + + NO;- 0 N O j + S ~ + + + + NO~- 0 SO 2- + N2 + SO~ + Tryptone + + + + $20~ + Gelatine + + + S4062- (+) Urea + + Cystein, HC1 + Glutamate + + + Cystine + + + + 20 Amino acids + + + Polysulfides + + Yeast extract + + + M n 2 + ( + ) Peptone + + Fe 3 + 0
Organic acids and proteinaceous substrates were at 0.2% (w/v), except yeast extract which was at 0.1%; sugars and polysaccharides were at 0.5% b Final cell concentration per ml of culture, 0, no growth; (+), 3 x 106-10v; + , 1-5 x 10v; + + , 5 x 1 0 7 - 1 0 8 , + + + , 1 2 x l 0 S ; + + + , > 2 x l 0 s .nd, not determined c In the absence of S, no growth occurred for either substrates tested when H2 was present in the headspace. The following substrates were tested but not utilized by strain GE5: glucose, galactose, sorbose, fructose, sucrose, xylose, arabinose, mannose, raffinose, melibiose, maltose, lactose, starch, glycogen, pure agar, alginates, carraghenans, mannitol, sorbitol, ethanol, isopropanol, methanol, acetate, lactate, citrate, benzoate, formate, gluconate, malate, 2-methylbutyrate, 3-methylbutyrate, lsovalerate, propionate, succinate, methylamines and all the amino acids tested alone a Electron acceptors were tested on YP medium (without SO4 z-) at 20 raM, except Fe 3 + (2 mM) cystine (10 mg/ml) and polysulfides (10 raM, prepared as described by Blumentals et al. (1990) e Growth occurred by fermentation in controls without electron acceptors f Tested with pyruvate added as carbon and energy source in the mineral base medium without NH4C1; nitrogen sources were at 20 mM except for complex organics which were at 0.2% w/v) g Control with no nitrogen source added, but trace amount of yeast extract (0.05% w/v)
on cys t ine was as g o o d as g r o w t h on e l e m e n t a l sulfur, w i t h c o m p a r a b l e g r o w t h r a t e (da ta n o t shown) a n d f inal cell y ie lds (Tab le 2) (this was p r e v i o u s l y o b s e r v e d for Pyrococcus furiosus by B l u m e n t a l s et al. 1990). Po ly - sul f ides a s s u m e d to be the a c t u a l subs t r a t e s fo r su l fu r
r e d u c t i o n in Pyrococcus ( B l u m e n t a l s et al. 1990), s u p p o r - t ed o n l y p o o r g r o w t h w h e n tes ted w i t h G E 5 . I n this case t he f inal cell y ie ld a b o u t was 1 0 % of t h a t o b t a i n e d in Y P S m e d i u m . T h i s c a n be e x p l a i n e d by the fact t ha t a d d i t i o n of po lysu l f ides in the cu l tu re m e d i u m b r ings the

pH to 8.5-9.0, which is very far from the optimal pH for growth.
Growth on elemental sulfur or cystine was accompa- nied by exponential H2S production which paralleled growth (Fig. 4). H2S concentration reached up to 13 btmol/ml when cells entered in the stationary phase. Growth in sulfur containing media produced an un- pleasant smell characteristic of mercaptans. This has also been reported for cultures of Desulfurococcus (Zillig et al. 1982; Jannasch et al. 1988), Thermococcus (Zillig et al. 1983), Pyrococcus (Fiala and Stetter 1986; Jannasch et al. 1992) and Hyperthermus (Zillig et al. 1990).
Sulfide toxicity was investigated at 90 ~ using YPS medium reduced with neutralized Na2S " 9 H 2 0 at diffe- rent concentrations (the control was reduced with tita- nium citrate or ascorbic acid). Specific growth rates and final cell densities showed no significant differences be- tween cultures reduced with ascorbate (0.4% w/v), tita- nium citrate, or 0.5 mM Na2S. However, the final cell densities for cultures reduced with 25 mM NazS were 10% of those reduced with 0.5 mM Na2S (Fig. 5). No growth was detected with 40 mM Na2S.
Fermentation products
The same fermentation patterns were found in cultures grown with or without sulfur (Table 3). The relative yields of products were roughly similar except that Hz replaced H2S as the main reduced product in cultures grown on sulfur-free medium.
Antibiotic sensitivity
GE5 was totally insensitive to the antibiotics chlor- amphenicol, vancomycin, penicillin, streptomycin, and rifampicin at a concentration of 150 rtg/ml.
Oxygen sensitivity
GE5 did not grow when traces of oxygen were present in the culture medium, as indicated by the pink color of the resazurin (Eh above - 70 mV). The cells remained
1 0 0
--" 8 0 ~
60
40 ~ X \
20 \ (,.,5 ~ ' e.,,
0 1'0 2 0 3 0 4 0 5 0 NaeS (mM)
Fig. 5. H2S inhibition of growth of strain GE5. Cell yields after 24 h of incubation in the presence of various Na2S concentrations presented as percentage of the Na2S-free control (stock solutions of NazS were titrated to pH 7.5 before ad&tion to the culture tubes). (o) Static culture, (�9 agitated culture (200 rpmt
345
Table 3. Fermentation products of strata GE5 in the presence and in the absence of S ~
Medium/Fermentation products" (p, mol/ml) YPS YP
Acetate 9.5 1.5 Isovalerate 1.6 0.6 Isobutyrate 1.0 0.2 Propionate 0.1 0.2 COz 5.2 1.54 H2N 9.0 0 H2 0 1.6
All values are average of data obtained from cultures grown 20 h at 90 ~ in triplicate, under N2 gas phase; uninoculated medium m duplicate was used as control. Average final cell concentrations in YPS and YP media were respectively: 4 x 108 and 7 x 107 cells/ml
viable for about 15 min after exponentially growing cultures were exposed to oxygen by aerating at 95 ~ After 30 min, only i % of the cells were still viable and there were no survivors after 8 h. When the culture was exposed to oxygen after cooling to 4 ~ the titer of viable cells decreased exponentially in the first 12 h with a half life of about 10 h (Fig. 6). The fraction of oxygen-resistant cells remained around 102 cells/ml for at least three weeks, but no cells survived after one month under these conditions.
DNA base composition
The G + C content of the DNA of strain GE5 was determined by the melting point method (Marmur and Doty 1962), and by direct analysis (Mesbah et al. 1989) to be 44.3% and 45.2% respectively.
LipM composition
The isolate GE5 contained both the typical archaeal phytanyl glycerol diethers and dibiphytanyl tetraethers, and 85% of its lipids were in the tetraether form. No significant amounts of fatty acids were detected. Over 80% of the ether lipids of GE5 were residue tetraether with 73.5% of the total as acyclic, and 9.5% as bipent- acyclic C4o biphytanyl chains.
1 I ,~ 'o . . . . . . . . . 0 10 20 30 40 50 60 70 80 90 100
Twne (h )
Fig. 6. Oxygen sensitivity of ceils of strain GE5 at different tem- peratures. (O) Viable cells during exposure to oxygen at 95 ~ (at) Viable cells number during exposure to oxygen at 4 ~

346
ermococcu$ celer
GE5
nosp~rdlum hungatei ethanothrix soehngemt
alobaeterium volcanit alococcus morrhuae
~ Methanobacterium thermoautotrophicum I t......_ Methanothermusfervidus L Methanococcus vanmehi
Thermofilum pendens Sulfolobus solfatartcus
- - [ ' Sulfolobus shibatae Desulfurococcus rnobtlis
Pyrodtcttum occultum Escherichta col•
0 0.05 0A0 0.15 0 2 0.25
Evoluttonary Dmtance (fixed mutations per nucleotide)
Fig. 7. Phylogenetic relationships based on 16 S rRNA sequences. The GenBank accession numbers for the organisms used in this analysis are as follows: Sulfolobus solfataricus, X03235; Thermo- coccus celer, M21529 I; Desulfurococcus mobil• M36474; Me- thanobacterium thermoautotrophicum, X66789 X66791; Halococ- cus morrhuae, $39858; Methanococcus vannielii, M36507; Methano- thermus f ervidus, M59145; Methanothrix soehng enii, X16932; Pyro- dictium occultum, M21087; Halobact erium volcanii, L00922; Sulf olo- bus shibatae, M32504; Methanospirillum hungatei, M60880; Ther- roof• pendens, X14835; Escherichia col• V00348. For references to the original publications of 16S rRNA sequences see Olsen et al. (1992)
16 S rRNA sequence analysis
A total of 1423 nucleotides of the 16 S rRNA sequence of strain GE5 were determined, corresponding to about 96% of the entire molecule. The inferred secondery structure of GE5 showed no idiosyncrasies in the expec- ted structure and conserved features and therefore there is no obvious evidence for errors introduced during the amplification process. The sequence was aligned with other known archaeal 16 S rRNA sequences and com- pared in a phylogenetic tree (Fig. 7) based on distance- matrix methods (Olsen 1987). The resulting tree was tested using bootstrap analysis (Felsenstein 1985). Strain GE5 was specifically associated with Thermococcus celer in 100% of the 50 bootstrap trees tested.
DNA : DNA hybridizations
DNA-DNA hybridizations (Table 4), revealed only a weak similarity between the new isolate and Pyrococcus
species. No significant similarity could be detected in this way with any of the Thermococcus species tested. Inter- estingly, in the light of DNA-DNA hybridization (Table 4), P.furiosus (Fiala and Stetter 1986) and P. woesei (Zillig et al. 1987) previously described as distinct species on the basis of the differences in morphological and metabolic properties, formed an unambiguously coherent species. Indeed, the observed similarity (about 85%) is well above the 70% level believed to indicate affiliation of two organisms to the same species (Johnson 1985b; Wayne et al. 1987).
Discussion
The novel marine hyperthermophilic strain GE5 belongs to the phylogenetic domain ofArchaea (Woese et al. 1990) on the basis of its resistance to antibiotics (Hilpert et al. 1981), the presence of isopranyl ether lipids, and its 16 S rRNA sequence. The latter criterion clearly shows that strain GE5 belongs to the order Thermococcales (Fig. 7), which is so far represented by only genera: Thermococcus (Zillig et al. 1983) and Pyrococcus (Fiala and Stetter 1986). This classification is consistent with the mor- phological and physiological features of strain GE5 that are characteristic of this order. Due to its optimal and maximal growth temperature, and its very short doubling time (less than 35 rain) under optimal growth conditions, strain GE5 ressembles Pyrococcus furiosus (Fiala and Stetter 1986) and Pyrococcus woesei (Zillig et al. 1987), the two representative species of the Pyrococcus genus. However, unlike these previously described species, strain GE5 does not utilize carbohydrates but shows very good growth on a mixture of amino acids as sole carbon and energy source. This ability has been so far reported only for the archaeal, hyperthermophilic, chemoorganotroph strain ES4 (Pledger and Baross 1991). Like most of hyperthermophilic archae, the major complex lipids (85% of total lipids) present in GE5 ( and related strains) are derivatives of glycerol dialkyl glycerol tetraether (GDGT). This composition sharply differs from that reported for Thermococcus celer (De Rosa et al. 1987) and P. woesei (Lanzotti et al. 1989) which contain almost exclusively glycerol diether core lipid. Since lipid com- position of other representatives of Thermococcus and Pyrococcus genera is lacking it is difficult to know if this difference in the tetraether/diether ratio has or not a
Table 4. DNA-DNA hybridizations between strain GE5 and references species of the Thermococcales
Tracer Driver DNA DNA
GE5 Thermo- Thermo- Thermo- Pyrococcus Pyrococcus coccus coccus coccus furiosus woesei celer litoralis stetteri
GE5 100_+ 1 5 a ! 1 8 • 0 19_+ 3 13_+2 T. celer 3 • I 100 + 1 5 +_ 1 1 2 ! 2 nd b 0 T. litoralis 1 3 _ 3 7_+ 3 1 0 0 • 5 • 1 7 • 8 • 3 T. stetterii nd 10 +_ 2 6 + 1 100 • 4 0 0 P.furiosus 22 • 5 6 _ 1 4 • 1 2 • 1 100 • 2 87 • 3 P. woesei 18 • 1 0 9 • 2 nd 83 • 5 100 • 3
a) Normalized percent of Homology; values are mean __ SD b) not determined

347
taxonomic value (Trincone et al. 1992). Also the lipid com- position may vary with the culture conditions as observ- ed for strain ES-4 (D. B. Hedrick, R. J. Pledger, D. C. Whi- te, and J. A. Baross, unpublished results). Therefore in examining this hypothesis the lipid composition of Pyro- coccus, Thermococcus and the new isolate should be analysed after cultivation under the same conditions in order to avoid culturing artefacts.
The G + C content of strain GE5 (45%) is si- gnificantly different from that of Thermococcus and Pyrococcus genera, originally distinguished mainly with respect to their G § C content (i.e. 56% and 38% respectively; Fiala and Stetter 1986). In DNA/DNA hybridization experiments, strain GE5 shows only 21% homology with P. furiosus and therefore represents most likely a new species of Pyrococcus. Since the DNA homologies of the new isolate with Thermococcus species are insignificant (Schleifer and Stackebrandt 1983; Fox et al. 1992) the new isolate is not a Thermococcus. This is in line with the 16S rRNA sequence comparison of GE5 with the corresponding sequence of T. celer which shows only 95.4% of identity. This degree of similarity is typical of intergeneric relationships in the Thermo- coccales (A. L. Reysenbach, in preparation). As other representatives of the Thermococcales, strain GE5 is seen to be slowly evolving (Achenbach-Richter et al. 1988).
Taking in account both ressembling and discrimina- tive features the new isolate represents a new species of Pyrococcus, named Pyrococcus abyssi by reference to its deep-sea biotope.
Effects of high pressures on bacterial growth and metabolism have been recently investigated with several deep-sea hyperthermophilic archaea (Nelson et al. 1991; Reysenbach et al. 1991 ; Nelson et al. 1992; Jannasch et al. 1992). Although the natural habitat of these strains is characterized by high hydrostatic pressures (ranging between 200 and 360 atm) they were obtained from enrichment cultures at atmospheric pressure, as was GE5. All these strains exhibited a marked tolerance to elevated hydrostatic pressures at their optimal growth tempera- ture, and some of them had a slight barophilic response to increases in pressure. A trend towards barophily at pressures greater than encountered in situ was demon- strated for strain AL2 whose specific growth rate at 440 atm was significantly higher than at low-pressure (Reysenbach et al. 1991). Similar results were obtained with strain ES4 (Pledger and Baross 1991) for which increasing the hydrostatic pressure from 35 to 500 arm resulted in a 60% increase in growth rate (Nelson et al. 1992). A slight barophilic response was also observed at in situ pressure for strains SY and MAR-7C (Jannasch et al. 1992). Although detailed growth studies have not yet been carried out. The preliminary results presented here indicate that strain GE5 has a tendency towards barophily at the upper temperature for growth. This is consistent with the observation that the optimal and maximal growth temperatures of strain GE5 are shifted upwards, by at least 3 ~ under high hydrostatic pressu- res. To our knowledge this is the first reported case, where pressure extents the upper temperature of a thermophilic organism. Miller et al. 1988, working with
the deep-sea thermophilic methanogen Methanococcus jannaschii, demonstrated that hyperbaric He pressure of up to 750 arm stimulated growth and extended the high-temperature limit for methanogenesis by several degrees (from 90 to 98 ~ although no growth occurred above 90 ~ A shift of the temperature growth range has thus only been described for the mesophilic, ob- ligately barophilic, bacterium, Colwellia hadaliensis (De- ruing et al. 1988).
Although strain GE5 was isolated from a geogra- phically distant area (North Fiji Basin) it has numerous features in common with strains ES4 and AL2 isolated on the Juan de Fuca Ridge (Pledger and Baross 1991; Reysenbach and Deming, 1991 respectively). These ar- chaeal strains are characterized by their heterotrophic mode of nutrition, their hyperthermophily (growing up to 108-110~ and as discussed above, their trend towards barophily. Additional similarities between GE5 and ES4 include their unique ability to utilize amino acids as sole carbon and energy sources and their ether lipids composition. This similarity was further confirmed by dot blot DNA-DNA hybridizations (data not shown), where under optimal conditions (T m - 25 ~ both strains AL2 and ES4 yielded a weak cross hybridization signal (detected by autoradiography) with GE5 but not with other references species of the Thermococcales order. Based on the 16 S rRNA sequences these Pyrococcus-like isolates group together phylogenetically with P. abyssi (A. L. Reysenbach et al., unpublished results).
Description of Pyrococcus abyssi
Pyrococcus abyssi sp. nov. Erauso, Reysenbach, Pace, and Prieur. ab.y'ss.i Gr. neutr.n, abyssos bottomless, immense depth, living within the ocean depths.
Cells are gram-negative, slightly irregular cocci, varia- ble in size (usually 0.8 to 2 ~tm) and shape depends upon growth conditions, type strain (GE5) is highly motile by means of a polar tuft of flagella. Cell envelope consists of a double- S-layer. Cell division occurs by constriction. Obligate anaerobe. Grows optimally at 0.2-0.3% NaC1, and pH around 7.0. At atmospheric pressure, growth occurs between 67 ~ and 102 ~ (Optimum 96 ~ corre- sponding to 33 min doubling time). Facultatively baro- philic: elevated hydrostatic pressures significantly extend upper growth temperature and stimulates growth. Habi- tat: anaerobic hot sites of deepsea hydrothermal systems. Barotolerant to facultatively barophilic.
Obligate chemoorganotroph, fermenting proteolysis products (e.g. tryptones and gelatin), and mixtures of amino acids and pyruvate in presence of vitamins in H2, CO2, and volatil fatty acids. H z produced inhibits growth, but this can be alleviated by addition of S ~ cystine, or polysulfides. In this case, H2S and mercaptans are pro- duced.
Membrane lipids are composed mainly of tetraether lipids with 0 to 4 cyclopentane rings in the isoprenoid cha'ns. G + C-content is 45%. 16S rRNA sequence comparisons affiliate P. abyssi with the thermococcalean archaea. GenBank accession number for the 16 S rRNA

348
sequence is 19921. Isolated from hot fluid from an active chimney in the North Fiji Basin (SW Pacific).
Type strain is P. abyssi, GE5, CNCM 1-1302, Collec- tion Nationale de Culture de Microorganismes, Institut Pasteur, Paris, France.
Acknowledgements. David B. Hedrick and Jean Guezennec are gratefully acknowledged for providing lipid analysis and interpreta- tion of lipids results. Franqoise Gaill is gratefully acknowledged for electron microscopy. We thank Bernard Ollivier for gas analysis. The technical assistance of Christophe Richard. St6phane L'harmdon, G6rard Ragu6nes and Anne-Lise Tr6dan is greately appreciated. We thank Nadia Benbouzid-Rollet and Jody Deming for helpfull discussion and critical reading of the manuscript. We wish to thank our anonymous referees for their pertinent comments. Thanks are also due to Daniel Desbruy+res and Suguru Ohta, cochief scientists of the Starmer cruise organized by Ifremer and Jamstec, for inviting two of us to participate to the "Starmer" cruise in the North Fiji Basin aboard the R/V "Nadir" and dive aboard the submersible "Nautile". This work was supported by CNRS, Ifremer, and grants of the SN Elf Aquitaine to G.E A.L.R. and N.R.P. were supported by Department of Energy grant DE-FG02- 92ER20088.
References
Achenbach-Richter L, Gupta R, Zillig W, Woese CR (1988) Roo- ting the archaebacterial tree: the pivotal role of Thermococeus celer in archaebacterial evolution. Syst Appl Microblol 10: 231-240
BalchW, Wolfe RS (1976) new approach to the cultivation of methanogenic bacteria: 2-mercaptoethaanesulfonic acid (HS- CoM)-dependent growth of Methanobacterium ruminanuum in a pressurized atmosphere. Appl Environ Microbiol 32:781-791
Balch WE, Fox GE, Magrum LS, Woese CR, Wolfe RS (1979) Methanogens: reevaluation of a unique biological group. Mi- crobml Rev 43:260-296
Bligh EG, Dyer WJ (1959) A rapid method for lipid extraction and purification. Can J Microbiol 35:911-917
Blumentals II, ItohM, OlsonGJ, Kelly RM (1990) Role of polysulfides in reduction of elemental sulfur by the hyperthermo- philic archaebacterium Pyrococcus furiosus. Appl Environ Mi- crobiol, 56:1255-1262
BurggrafS, Jannasch HW, Nicolaus B, Stetter CO (1990a) Ar- chaeoglobus profundus sp. nov., represents a new species within the sulfate-reducing archaebacteria. Syst Appl Microbiol 13: 24-28
BurggrafS, Fricke H., Neuner AM, Kristjansson J, Rouvier P, Mandelco L, Woese CR. Stetter KO (1990b) Methanocoeeus igneus sp. nov., a novel hyperthermophihc methanogen from a shallow submarine hydrothermal system. Syst Appl Microbiol 13:263 269
Charbonmer F, Erauso G, Barbeyron T, Prieur D, Forterre P (1992) Evidence that a plasmid from a hyperthermophilic archaebacterium is relaxed at physiological temperatures. J Bacteriol 174:6103-6108
Cline JD (1969) Spectrophotometric determination of hydrogen sulfide in natural waters. Limnol Oceanogr 14:454-458
De Rosa M, Gambacorta A, Trincone A, Basso A, Zillig W, Holz I (1987) Lipids of the Thermococeus celer, a sulfur-reducing archaebacterium: structure and biosynthesis. Syst Appl Mi- crobiol 9 :1 -5
Delaney JR, McduffRE, Lupton JE (1984) Hydrothermal fluid temperatures of 400 ~ on the endeavour segment, northern Juan de Fuca. Trans Am Geophys Union 65:973
Deming JW, Baross JA (1986) Solid medium for culturing black smoker bacteria at temperatures to 120 ~ Appl Environ Microbiol 51:238-243
Deming JW, Hada H, Colwell RR, Luehrsen, Fox GE (1984) The ribonucleotide sequence of 5S rRNA from two strains of deep-sea barophilic bacteria. J Gen Microbiol 130: 1911 - 1920
Deming JW, Somers LK, Straube WL, Swartz DG, Macdonell MT (1988) Isolation of an obligately barophilic bacterium and description of a new genus, Coh4,ellia gen. nov. Syst Appl Microbiol 10:152-160
Desbruy6res D, Alayse AM, Otha S (1993) Deep-sea hydrothermal communities in the Southwestern Pacific back-arc basins (the North-Flji and Lau Basins): composition, microdistribution and food web. Marine Geol (in press)
Feinberg AP, Vogelstein B (1983) A technique for radiolabeling DNA restriction endonuclease fragments to high specific ac- tivity. Anal Biochem 132:6-13
Felsenstein J (1985) Confidence limits on phylogenies: an approach using bootstrap. Evolution 39:783 79I
Fiala G, Stetter KO (1986) Pyrococcusfuriosus sp. nov. represents a new genus of marine heterotrophic archaebacteria growing optimally at 100 ~ Arch Microbiol 145:56-61
Fiala G, Stetter KO, Jannasch HW, Langworthy TA, Madon J (1986) Staphylothermus marinus sp. nov. represents a novel genus of extremeiy thermophilic submarine heterotrophic ar- chaebacteria growing up to 98 ~ Syst Appl Microbiol 8: i06-113
Fox GE, Wisotzkey JD, Jurtshuk P (1992) How close is close: 16 S rRNA sequence identity may not be sufficient to guarantee species identity. Int J Syst Bacteriol 42: 166-170
Grimaud D, Ish~bashi JI, Lagabriele Y, Auzende JM, Urabe T (1991) Chemistry of hydrothermal fluids from the 17 ~ active site on the North Eiji basin ridge (SW Pacific). Chem Geol 93: 209-218
Grimont PAD, PopoffMY, Gnmont E, Coynault C, Lemelin M (1980) Reproducibility and correlation study of three deoxyribo- nucleic acid hybridization procedures. Curr Mlcrobiol 4: 325-330
Hedrick DB, Guckert JB, White DC (1991) Archaebacterial ether lipid diversity analyzed by supercritical fluid chromatography: integration with a bacterial lipid protocol. J Lipid Res 32: 659-666
Hilpert R, Winter J, Hammes W, Kandler O (1981) The sensitivity of archaebacterla to antibiotics. Zentralbl Bakt Hyg I Abt Orig C2:11-20
Hobbie JE, Dahney RJ, Jasper S (1977) Use of nucleopore filters for counting bacteria by fluorescence microscopy. Appl Environ Microbiol 33 : 1225-1228
Huber R, Kurr M, Jannasch HW, Stetter KO (1989) A novel group of abyssal methanogenic archaebacteria (Methanopyrus) gro- wing at 110 ~ Nature 342:833-834
IngvorsenK, JorgensenBB (1979) Combined measurement of oxygen and sulfide in water samples. Limnol Oceanogr 24: 390 393
Ivanova TL, Turova TP, Antonov AS (1988) DNA-DNA hybridi- zation studies on some purple non sulfur bacteria. Syst Appl Microbiol 10:259-263
Jannasch HW, Wirsen CO, Molyneaux S J, Langworthy TA (1988) Extremely thermophilic fermentative archaebacteria of the genus DesuIfurococcus from deep-sea hydrothermal vents. Appl Envi- ron Microbiol 54:1203-1209
Jannasch HW. Wlrsen CO, Molyneaux S J, Langworthy TA (1992) Comparative physiological studies of hyperthermophilic archae isolated from deep-sea hot vents with emphasis on Pyrococcus strain GB-D. Appl Envaron Microbiol 58:3472-3481
Johnson JL (1985a) DNA reassociation and RNA hybridisatlon of bacterial nuclei acids. Methods Microbiol 18:33-74
Johnson JL (1985b) Nucleic acid in bacterial classification. In: Krieg NR. Holt JG (eds) Bergey's manual of systematic bac- teriology, vol I. Williams & Wilkins, Baltimore
Jollivet D, Hashimoto J, Auzende JM (1989) Premieres observa- tions de communaut6s animales associ6es ~t l'hydrothermalisme arri+re arc du bassin Nord-Fidjien. CR Acad Sci Paris 309: 301-308

349
Jones WJ, Leigh JA, Mayer F, Woese CR, Wolfe RS (1983) Me- thanococcus jannaschii sp. nov., an extremely thermophihc me- thanogen from a submarine hydrothermal vent. Arch Microbiol 136:254-261
Kurr M, Huber R, Konig H, Jannasch HW, Fricke H, Trlncone A, Kristjansson JK, Stetter KO (1991) Methanopyrus kandleri, gen. and sp. nov. represents a novel group of hyperthermophilic methanogens, growing at 110 ~ Arch Microbiol 156:239-247
Lanzotti V, Trincone A, Nlcolaus B, ZiIlig W, De Rosa M, Gamba- corta A (1989) Complex lipids of Pyrococcus and AN1, thermo- phdic members of the archaebacteria belonging to the Thermo- coccales. Biochim Biophys Acta 1004:44-48
Marmur J, Doty P (1962) Determination of the base composition of deoxyribonucleic acid from its thermal denaturation tem- perature. J Mol Biol 5:109-118
Mesbah M, Premachandran U, Whitman WB (1989) Precise mea- surement of the G + C content of dideoxyribonucleic acid by high performance liquid chromatography. Int J Syst BacterioI 39:159-167
Miller JF, Shah NN, Nelson CN, Ludlow JM, Clark DS (1988) Pressure and temperature effects on growth and methane production of the extreme thermophile Methanococcus janna- schii. Appl Environ Microbiol 54:3029 3042
Nelson CM, Shuppenhauer MR, Clark D S (1991) Effect of hyperba- ric pressure on a deep-sea archaebacterium in stainless steel and glass-lined vessels. Appl Environ Microbiol 57:3576 3580
Nelson CM, Shuppenhauer MR, Clark DS (1992) High pressure, high temperature bioreactor for comparing effects of hyperbaric and hydrostatic pressure on bacterial growth. Appl Environ Microbiol 58:1789 1793
Olsen GJ (1987) The earliest phylogenetic branchings: comparing rRNA-based evolutionary trees inferred with various techmques. Cold Spring Harbor Symp Quant Biol 52:825 838
Otsen G J, Overback R, Larsen N, Marsh TL, Mc Canghey M J, Macinkenas MA, Kuan WM, Macke T J, Xmg Y, Woese CR (1992} The ribosomal database project. Nucleic Acids Res 20: 2199 2200
Pledger R J, Baross JA (1991) Preliminary description and nutritio- nal characterizatmn of a chemoorganotrophic archaebactermm growing of up to 110 ~ isolated from a submarine hydrothermal vent environment. J Gen Microbml 137:203-211
Pley U0 Schipka J, Gambacorta A0 Jannasch HW, Fricke H, Ra- chel R, Stetter KO (1991) Pyrodictizon abyssi sp. nov. represents a novel heterotrophic marine archaeaI hyperthermophile gro- wing at 110 ~ Syst Appl Microbiol 14:245-253
Reysenbach AL, Deming JW (1991) Effects of hydrostatic pressure on growth of hyperthermophilic archaebacteria from the Juan de Fuca Ridge. Appl Environ Microbiol 57:1271-1274
Reysenbach AL, Giver L J, Wickham GS, Pace NR (1992) Differen- tial amphfication f rRNA genes by polymerase chain reaction. Appl Environ Microbiol 58:3417-3418
Saikl RK, Gelfand DH, Stoffel S J, Scharf S, Higuchi R, Horn GT, Mullis HB, Erlich HA (1988) Primer-directed enzymatic am- plification of DNA with a thermostable DNA polymerase. Science 230:1350-1354
Sambrook J, Fritch EF, Maniatis T (1989) Molecular cloning: a laboratory manual, 2nd ed. Cold Spring Harbor Laboratory Press, Cold Spring Habor, NY
Sanger F, Nicklen S, Coulson AR (1977) DNA sequencing with chain terminating inhibltors. Proc Natl Acad Scl USA 74: 5463 5483
SchleiferKH, StuckebrandtE (1983) Molecular systematics of prokaryotes. Ann Rev Microbiol 37:143-187
Spiess FN, Macdonald KV, Atwater Y, Ballard R, Carranza A, Cordoba D, Cox C, Diaz Garcia VM, Francheteau J, Guer- rero J, Hawkins J, Haymon R, Hessler R, Juteau T, Kastner M, Larson R, Luyendyk B, Macdougall JD, Miller S, Normark W, Orcutt J. Rangm C (1980) Wast Pacific Rise: hot springs and geophysical experiments. Science 207: 1421 - 1433
Stetter KO (1988) Archaeoglobusfervidus gem nov., sp. nov. :a new taxon of extremely thermophilie archaebacteria. Syst Appl Microbiol 10: t72-i73
Stetter KO, Konig H, Stackebrandt E (1983) Pyrodicttum gen. nov., a new genus of submarine disc-shaped sulphur reducing ar- chaebacteria growing optimaly at 105 ~ Syst Appl Microblol 4:535-551
Trincone A, Nicolaus B, Palmieri G, De Rosa M, Huber R, Stet- ter KO, Gambacorta A (1992) Distribution of complex and core lipids within new hyperthermophilic members of the Archaea Domain. Syst Appl Microbiol 15:11-17
Vanlot D, Courties C, Partensky F (1989) A simple method to preserve oceanic phytoplankton for flow cytomemc analyses. Cytomctry 10:629-635
Wayne LG, Brenner D J, Colwell RR, Grimont PAD, Kandler O, Krichevsky MI, Moore LH, Moore WEC, Murray HE, Stak- kebrandt E, Starr MP, Truper HG (1987) Report of the ad hoc committee on reconciliation of the approaches to bacterial systematics. Int Syst Bacteriol 37:463-464
Wermann SD, Springer MS, BrittenRJ (1990) Nucleic acids I' DNA-DNA hybridization. In: Hillis DM, Moritz C (eds) Mole- cular systematic. Sinauer Associates, Massachusetts, USA, pp 204-245
White DC, Bobbie RJ, King JD, Nickels JS, Amoe P (1979) Lipid analysis of sediments for microbial biomass and community structure. In: Litch CD, Seyfried PL (eds) Methodology for biomass determinations and microbial activities in sediments. ASTM, Washington, pp 87 103
Woese CR, Kandler O, Wheelis ML (1990) Towards a natural system of organisms: proposal for the domains Archaea. Bac- iena, and Eucarya. Proc Natl Acad Sci USA 87:4576-4579
Yayanos AA (1986) Evolution and ecological implications of the properties of deep-sea barophilic bacteria. Proc Natl Acad Sea USA 83:9542-9546
Yayanos AA, Dietz AS, Boxtel R van (1982) Dependence of repro- duction rate on pressure as a hallmark of deep-sea bacteria. Appl Environ Microbiol 44:1356-1361
Zehnder AJB, Wuhrmann K (1976) Titanium(III)-citrate as a non- toxic oxidation-reduction buffering system for the culture of obligate anaerobes. Science 194:1 t65- t i66
Zhao H, Wood AG, Widdel F, Bryant MP (1988) An extremely thermophilic Methanococcus from a deep sea hydrothermal vent and its plasmid. Arch Macrobiol 150:178 183
Zillig W, Stetter KO, Prangishwilli D, Schafer W, Wunderl S, Jane- kovm D, Holz I, Palm P (1982) Desu~lrococcaeae, the second family of extremely thermophilic, anaerobic sulfur-respiring Therrnoproteales. Zentralbi Bakt Hyg I Abt Orig 3:304-311
Zillig W, Gierl A, Schreiber S, Wunderl S, Janekovic D, Stetter KO, Klenk HP (1983) The archaebacterium Thermophilum pendens represents a novel genus of the thermophilic, anaerobic sulfur respiring Thermoproteales. Syst Appl Microbiol 4:79-87
Zillig W, Holz I, Klenk HP, Trent J, Wunderl S, Janekovic D, Imsel E, Haas B (1987) Pyrococcus woesei, sp. nov., an ultra- thermophilic marine archaebacterlum, representing a novel order, Thermococcales. Syst Appl Microbiol 9:62-70
Zilhg W, Holz I, Janekovic D, Klenk HP, Imsel E, Trent J, Wun- derl S, Forjaz VH, Coutinho R, Ferreira T (1990) Hyperthermus butylicus, a hyperthermophilic sulfur redncing archaebacterium that ferments peptides. J Bacterioi 172:3959-3965




















