
Comparative genomics and systems biology of ... - CORE
469
Comparative genomics and systems biology of environmental stress responses relevant to fungal virulence A thesis presented for the degree of Doctor of Philosophy of the University of London and the Diploma of Imperial College by Andrew N. McDonagh Department of Microbiology Imperial College Armstrong Road, London SW7 2AZ OCTOBER 2010
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of Comparative genomics and systems biology of ... - CORE

Comparative genomics and systems biology of environmental stress responses relevant to fungal
virulence
A thesis presented for the degree of
Doctor of Philosophy of the University of London
and the
Diploma of Imperial College
by
Andrew N. McDonagh
Department of Microbiology
Imperial College
Armstrong Road, London SW7 2AZ
OCTOBER 2010

2
I certify that this thesis, and the research to which it refers, are the product of my own work, and that any ideas or quotations from the work of other people, published or otherwise, are fully acknowledged in accordance with the standard referencing practices of the discipline.
Signed: r

3
For Dad (1945 - 2007).
For my son Fin. Stick to the arts subjects kid.
And for Mrs Chippy, who pioneered the way.

4
Abstract
The ability to sense and adapt to environmental stress is a major factor for fungi such as Candida glabrata and Aspergillus fumigatus to colonize a host niche and therefore for patho-genesis. Herein, I assess the role of the putative C. glabrata stress activated protein kinase signalling architecture in hyperosmotic stress adaptation using gene disruption, and gene ex-pression microarrays. I report that, apart from the induction of glycerol biosynthetic genes, the transcriptional response of wild-type C. glabrata subjected to hyperosmotic stress is dissimilar to S. cerevisiae (< 20% concordance) despite sharing approximately 90% of their genetic repertoire. Transcription profiling of mutants disrupted in signalling genes (HOG 1 and STE11), revealed a significant component (approximately 30%) of transcription that was independent of the extant Shol signalling. In parallel, a library of null mutant strains disrupted in 186 C. glabrata putative transcription factor ORFs was developed and screened for sensitivity to the anti-fungal compound fluconazole, identifying novel determinants of fluconazole sensitivity (including a regulator of aromatic acid biosynthesis and several me-diator complex subunits). In a separate study, to avoid using in vitro approximations of host niche stressors, the transcriptional response of A. fumigatus cells during early stage infection in a murine model in invasive aspergillosis was investigated. This revealed an A. fumigatus gene expression programme governing metabolic and physiological adaptation, which may allow the organism to prosper within the mammalian niche. The transcrip-tional response to nutritional cues was also studied in C. glabrata and mapped to molecular interaction networks inferred by homology with S. cerevisiae. Transcription profiling of C. glabrata exposed to serum (10% v/v) revealed upregulation of amino acid uptake and catabolism and repression of carbon catabolic pathways in favour of high mannose glycan biosynthesis.

5
Acknowledgments
There are many people to thank for their scientific and emotional support during this chal-lenging endeavour. Accordingly, I would like to thank my excellent mentor, and friend, Ken Haynes for his unfaltering support. I would also like to thank Elaine Bignell and Janet Quinn for giving me the opportunity to analyze some interesting transcription data and to Michael Stumpf for informative and entertaining discussions in bioinformatics and computa-tional biology. I am indebted to members of the Butler group at University College Dublin for schooling me in the practical aspects of microarray technology, notably Anne-Marie Jennings. I thank Geraldine Butler for her generous gift of Candida glabrata microarrays. I would also like to thank Darius Armstrong-James, Helen Findon, Hsueh-Lui Ho and Lucy Holcombe for their guidance in the early stages of my PhD, and Piers Ingram and fellow CisBIC bunker detainees for technical support in the latter stages. I extend special thanks to Tatiana Munera and Emily Cook at Imperial College for expert technical assistance and patience. I am thankful for the Medical Research Council doctoral training award that supported this research. Finally: thanks to Ernest, Frank, Thomas, Sir James Caird and Dudley Docker for inspiration.

6
Table of contents
Abstract 4
1 Introduction 23 1.1 The increasing prevalence of opportunistic mycoses 23 1.2 The medical burden of invasive candidiasis and invasive pulmonary aspergillosis 24
1.2.1 Candida spp. and candidiasis 24 1.2.2 Aspergillus fumigatus and aspergillosis 28
1.3 Current treatment options for systemic mycoses: the need for better anti- fungal drugs 28
1.4 Identifying therapeutic targets: strategies for identifying virulence determi- nants in Candida spp. and A. fumigatus 31
1.5 A brief review of virulence factors important for Candida and Aspergillus mediated pathogenesis 34
1.6 Sensing and adapting to environmental stress responses 43 1.6.1 Oxidative stress response 43 1.6.2 Ambient pH adaptation 44 1.6.3 Hyperosmotic stress adaptation 44 1.6.4 Thermotolerance 45 1.6.5 The general stress response of S. cerevisiae 45
1.7 Identifying therapeutic targets: potential for a systems approach to fungal virulence studies 47
1.8 Specific aims addressed in this thesis 53
2 Materials and Methods 55 2.1 Experimental Methods 55
2.1.1 Fungal strains and culture media 55 2.1.2 Bacterial strains and culture media 56 2.1.3 Assays to identify altered growth phenotypes of mutant C. glabrata
and S.cerevisiae strains 56 2.1.3.1 Solid agar assays of C. glabrata and S.cerevisiae strain growth 56 2.1.3.2 Miniaturized optical density assays of mutant Candida glabrata
strain growth 57 2.1.4 Oligonucleotide primer synthesis 57 2.1.5 Polymerase Chain Reaction (PCR) 58

7
2.1.6 Ribonucleic acid (RNA) extraction and quantification 2.1.7 Linear DNA and PCR extraction and purification 2.1.8 Agarose gel electrophoresis 2.1.9 Yeast DNA extraction 2.1.10 Transformation of yeast cells with electroporation 2.1.11 Western blots
2.1.11.1 Design of probes 2.1.11.2 Microarray slides
2.1.12 Reverse transcription and fluorescent labelling 2.1.12.1 Hybridization, washing and scanning 2.1.12.2 Image acquisition and data analysis
2.1.13 Statistical analysis of microarray data 2.2 Computational Methods
2.2.1 Computation of network statistics 2.2.2 k-clique percolation and community finding
3 The Hogip and Stellp dependent transcriptional effects of hyperosmotic stress in Candida glabrata 63 3.1 Chapter Overview 63 3.2 Specific Methods 70
3.2.1 Construction of a C. glabrata hogl null mutant 70 3.2.1.1 Western blot analysis 70 3.2.1.2 Genome-wide transcript profiling 71 3.2.1.3 Downstream bioinformatic analyses 71
3.3 Results 72
3.3.1 C. glabrata, hyperosmotic stress dependent, Hog1p phosphorylation is abolished in a stell background
72
3.3.2 The transcriptional response of wild-type C. glabrata cells exposed to transient hyperosmotic stress
74
3.3.3 Comparison of S. cerevisiae, C. albicans and C. glabrata transcrip- tional responses to hyperosmotic stress
85
3.3.4 The effect of 1-10G1 or STE11 deletion on hyperosmotic stress in- duced transcriptional responses in C. glabrata
88
3.3.5 The hogl and stel 1 specific transcriptional responses to hyperosmotic shock: evidence for transcriptional cross-talk between putative C glabrata MAPK cascades 92
3.3.6 Identification of a hypothetical hyperosmotic stress regulon in C glabrata
94 3.3.7 Identification of hyperosmotic stress sensitive C. glabrata transcrip-
tion factor null mutant strains 101 3.4 Discussion 120 3.5 Chapter Summary 133
of cDNA of cDNA microarrays
58 58 59 59 59 59 60 60 60 61 61 61 62 62 62

8
4 Sub-telomere directed gene expression during initiation of invasive as- pergillosis 134 4.1 Chapter Overview 134 4.2 Specific Methods 137
4.2.1 A. fumigatus strains and growth conditions 137 4.2.2 Murine Infections 138 4.2.3 RNA extraction and amplification 139 4.2.4 Estimation of fidelity of transcript abundance following mRNA am-
plification 139 4.2.5 Microarray hybridisations 140
4.3 Results 140 4.3.1 A. fumigatus RNA extraction and mRNA amplification 140 4.3.2 Impact of mRNA amplification on preservation of transcript ratios 141 4.3.3 A. fumigatus transcript profile during initiation of mammalian infection144 4.3.4 Lineage specificity and locational analyses of differentially expressed
genes 150 4.3.5 Clustering of induced genes 157 4.3.6 Comparative analyses of murine adaptation- and in vitro gene expres-
sion datasets 158 4.3.7 Nutrient limitation is a relevant physiological cue during A. fumigatus
initiation of murine infection 160 4.3.8 The LaeA regulon is represented among genes preferentially expressed
during intitation of murine infection 162 4.4 Discussion 163 4.5 Chapter Summary 171
5 Functional genomics of fluconazole sensitivity in Candida glabrata using a library of transcription factor knock-outs: a pilot study 172 5.1 Chapter Overview 172 5.2 Specific Methods 175
5.2.1 Construction of the CGTFKOLib library of transcription factor null mutants 175
5.2.2 Liquid culture assay to determine strain growth dynamics 177 5.2.3 Fitting growth dynamics data to a logistic model of population growth178 5.2.4 Fitting truncated growth dynamics data to an exponential model of
population growth 178 5.2.5 Fitting the inhibitory effect of fluconazole at a single time point in a
linear model 179 5.3 Unsupervised learning of strain clusters based on growth parameter estimates 179 5.4 Results 179 5.5 Discussion 182 5.6 Chapter Summary 192
6 Conclusions and future work 193 6.1 Future work 193

9
A Chapter 1 Supplementary Data 199
B Media Recipes 205
C Strain Genotypes 206
D Chapter 3 Supplementary Data 207
E Chapter 4 Supplementary Data 274 E.1 Dataset S1 reference 306 E.2 Dataset S2 reference 306 E.3 Dataset S3 reference 306 E.4 Dataset S4 reference 306
F Chapter 5 Supplementary Data 307
G Transcription profiling of C. glabrata serum stress responses 330 G.1 Chapter overview 330 G.2 Specific methods 331 G.3 Transcriptional response of C. glabrata cells exposed to 10% Foetal Calf
Serum (FCS) 332
References 469

10
List of Figures
1.1 Maximum likelihood phylogeny of 153 universally distributed fungal genes . 26
2.1 Overview of k-clique percolation algorithm 62
3.1 Architecture of the HOG pathway in S. cerevisiae 64 3.2 Architecture of MAPK cascades of the model organism S. cerevisiae, incuding
the HOG osmoadaptation cascade 65 3.3 The C. glabrata HOG pathway is activated by a single MAPKKK, Ste11p 73 3.4 Schematic of experimental design for transcriptional profiling of hyperosmotic
stress responses 76 3.5 Single channel normalization methods preserve slide intensity distributions
for cross-slide comparison and construction of synthetic log ratios of tran- script abundance 77
3.6 Heatmap featuring column and row based cluster dendrograms of genome- wide transcriptional response to hyperosmotic stress in wild type, hogl and ste / / cells 78
3.7 Heatmap subset of genome featuring column and row based cluster dendro-grams of loge (Cy5T') - log2 (Cy3TM ) for transcripts induced by hyperosmotic stress in wild-type cells, compared with the equivalent values in hyperosmotic stressed hogl and sten cells 79
3.8 A transcript encoding glycerol dehydrogenase, an enzyme involved in glyc-erophospholipid metabolism is induced in wild-type C. glabrata cells exposed to 1 M sorbitol for 15 minutes 80
3.9 Three transcripts involved in glycerolipid metabolism are induced in wild- type C. glabrata cells exposed to 1 M sorbitol for 15 minutes 81
3.10 Three transcripts encoding enzymes involved in the ergosterol biosynthesis pathway, are repressed in wild-type C. glabrata cells exposed to 1 M sorbitol for 15 minutes 84
3.11 Venn diagram of transient hyperosmotic stress and basally regulated genes in wild-type, hogl and sten cells. 90
3.12 Architecture of MAPK cascades of the model organism S. cerevisiae, incuding the HOG osmoadaptation cascade annotated with evidence of transcriptional cross-talk in conserved C. glabrata transcripts 103
3.13 Overview of the hypothetical hyperosmotic stress regulon 104

11
3.14 Distributions of betweenness centrality and degree in the hypothetical os- mostress regulon network (HORN) 105
3.15 HORN transcription factor activity versus node connectivity 106 3.16 HORN subnetwork comprising putative hyperosmotic stress regulators Skolp,
Skn7p, Msn2p, Msn4p and their immediate k = 1 neighbours 107 3.17 HORN subnetwork comprising highly active regulators and their immediate
k = 1 neighbours 108 3.18 HORN subnetwork comprising hub regulators and their immediate k = 1
neighbours 109 3.19 HORN subnetwork comprising low activity and low connectivity regulators
and their immediate k = 1 neighbours 110 3.20 HORN subnetwork comprising the pair of transcriptional regulators account-
ing for the largest proportion of the regulated set and their k = 1 neighbours 111 3.21 HORN subnetwork comprising candidate GPD2 regulators and their k = 1
neighbours 112 3.22 HORN subnetwork comprising candidate GCY1 regulators and their k = 1
neighbours 113 3.23 HORN subnetwork comprising candidate ALD6 regulators and their k = 1
neighbours 114 3.24 HORN subnetwork comprising direct targets of Cin5p 115 3.25 HORN subnetwork comprising direct targets of Stel2p 116 3.26 HORN subnetwork comprising the minimal connections between 12 ortholo-
gous S. cerevisiae genes required for growth in hyperosmotic stress 117 3.27 Four C. glabrata mutants, each lacking a single putative transcription factor
are hypersensitive to 0.5 M KC1 in a liquid growth medium assay 118 3.28 Three C. glabrata mutants, each lacking a single putative transcription factor
are hypersensitive to 0.5 M KC1 in a liquid growth medium assay 119 3.29 A working model of C. glabrata ATCC 2001 hyperosmotic stress adaptation 122 3.30 Alternate hypotheses for hyperosmotic stress signal transduction through the
Shol branch of C. glabrata ATCC 2001 130 3.31 Four C. glabrata transcripts, orthologous to S. cerevisiae genes encoding core
and regulatory subunits of the 26S proteasome are induced in cells exposed to 1 M sorbitol for 15 minutes 131
3.32 Two C. glabrata transcripts, orthologous to S. cerevisiae genes encoding members of the adjacent cell integrity pathway are induced in cells exposed to 1 IVI sorbitol for 15 minutes 132
4.1 Comparative time-course of A. fumigatus Af293 germination and hyphal de- velopment in the murine lung, and laboratory culture 142
4.2 Correlation of log2 ratios resulting from comparative transcriptional analysis of the laboratory cultured A. fumigatus cell populations To and T60 under varying mRNA amplification protocols. 143
4.3 A genome-wide transcriptional snapshot of A. fumigatus Af293 during intia- tion of murine infection
151

12
4.4 Distribution of lineage specific and telomere-proximal genes among differen- tially expressed host adaptation dataset 152
4.5 Overlap between murine adaptation and in vitro stress datasets 153 4.6 Comparative analysis of A. fumigatus gene expression datsets. 154 4.7 Characterisation of A. fumigatus growth on YPD, murine lung tissue and
minimal medium supplemented with ammonium or hydroxyproline 155 4.8 Expression of LaeA-regulated genes during initiation of murine infection 156
5.1 Molecular approach adopted for targeted disruption of C. glabrata transcrip- tion factor ORFS (schematic). 176
5.2 Logistic growth curves of a) wild-type strain ATCC 2001and b) the library parental strain his3 in YEPD and YEPD + fluconazole (5-10 pg-m1-1) . . . 183
5.3 Fluconazole dependent distribution of midpoint standard errors and number of inestimable growth midpoints 184
5.4 A liquid growth phenotype assay identifies two categories of fluconazole- sensitive null mutant strains 185
5.5 Growth curves of strains belonging to a cluster (C1) characterized by wild-type like growth in YEPD but sensitivity to YEPD + fluconazole (5-100 ug•ml-1) 186
5.6 Growth curves of strains belonging to a cluster (C5) characterized by slow growth in YEPD and YEPD + fluconazole (5-100 pg•ml-1) 187
5.7 Growth curves of strains belonging to a cluster (C3) characterized by wild- type like growth in YEPD and YEPD + fluconazole (5-100 mg•ml-1) . 188
A.1 Licensing timeline for antifungal drugs with systemic bioavailability 200 A.2 Species specific incidence rates of invasive candidiasis 201 A.3 Common antifungal drug molecules and their mechanism of action 202 A.4 Schematic summary of Candida albicans virulence factors 203 A.5 Proportion of conserved virulence factors in the Ascomycota 204
D.1 Solid agar phenotype assay screen of CGTFKOlib plate 1 on hypertonic agar (1 M Sorbitol) 262
D.2 Solid agar phenotype assay screen of CGTFKOlib plate 2 on hypertonic agar (1 M Sorbitol) 263
D.3 Calculated dN dS rates for Saccharomyces paradoxes, Saccharomyces bayanus, Saccharomyces mikatae and C. glabrata 264
E.1 Schematic representation of hybridisations performed in the amplification control study 275
E.2 Comparative analysis of A. fumigatus gene expression datasets 276 E.3 Concordance between estimates of relative abundance from triplicate qPCR
measurements and microarray experiments for selected A. fumigatus ORFs. 277 E.4 Box and whisker plots of Cy5TM and Cyr intensities for slides hybridized with
amplified and non-amplified RNA 278 E.5 Genes rejected in the amplification control experiment transposed onto cor-
relation plots 279

13
F.1 Assessment of pipette channel cross contamination on a robotic platform . . 328 F.2 Example growth curves for four strains following incubation and sampling on
a robotic platform 329
G.1 Exploratory data analysis plots for C. glabrata environmental stress microar- ray hybridizations 337
G.2 MA plots for each hybridization performed in the C. glabrata environmental stress study 338
G.3 Volcano plots of differentially expressed transcripts in the C. glabrata envi- ronmental stress study 338
G.4 Genes differentially transcribed following exposure to 10% FCS (1 hour) mapped to the orthologous global metabolic map of S. cerevisiae 339
G.5 Genes differentially transcribed following exposure to 10% FCS (1 hour) mapped to the orthologous phenylalanine, tyrosine and trytophan biosyn- thetic pathway of S. cerevisiae 340
G.6 Genes differentially transcribed following exposure to 10% FCS (1 hour) mapped to the orthologous valine, leucine and isoleucine biosynthetic path- way of S. cerevisiae 341
G.7 Genes differentially transcribed following exposure to 10% FCS (1 hour) mapped to the orthologous valine, leucine and isoleucine degradation path- way of S. cerevisiae 342
G.8 Genes differentially transcribed following exposure to 10% FCS (1 hour) mapped to the orthologous arginine and proline metabolic pathway of S cerevisiae 343
G.9 Genes differentially transcribed following exposure to 10% FCS (1 hour) mapped to the orthologous alanine, aspartate and glutamate metabolic path- way of S. cerevisiae 344
G.10 Genes differentially transcribed following exposure to 10% FCS (1 hour) mapped to the orthologous lysine degradation pathway of S. cerevisiae . . . 345
G.11 Genes differentially transcribed following exposure to 10% FCS (1 hour) mapped to the orthologous glycolysis pathway of S. cerevisiae 346
G.12 Genes differentially transcribed following exposure to 10% FCS (1 hour) mapped to the orthologous pyruvate metabolism pathway of S. cerevisiae . 347
G.13 Genes differentially transcribed following exposure to 10% FCS (1 hour) mapped to the orthologous oxidative phosphorylation pathway of S. cere- visiae 348
G.14 Genes differentially transcribed following exposure to 10% FCS (1 hour) mapped to the orthologous biotin biosynthesis pathway of S. cerevisiae . . 349
G.15 Genes differentially transcribed following exposure to 10% FCS (1 hour) mapped to the orthologous pantothenoate and CoA biosynthesis pathway of S. cerevisiae 350
G.16 Genes differentially transcribed following exposure to 10% FCS (1 hour) mapped to the orthologous unsaturated fatty acid biosynthesis pathway of S. cerevisiae 351

14
G.17 Genes differentially transcribed following exposure to 10% FCS (1 hour) mapped to the orthologous starch and sucrose metabolism pathway of S cerevisiae 352
G.18 Genes differentially transcribed following exposure to 10% FCS (1 hour) mapped to the orthologous high mannose type N-glycan biosynthesis path- way of S. cerevisiae 353
G.19 Genes differentially transcribed following exposure to 10% FCS (1 hour) mapped to the orthologous o-mannosyl glycan biosynthesis pathway of S cerevisiae 354
G.20 Genes differentially transcribed following exposure to 10% FCS (1 hour) mapped to the orthologous v-snare vesicular trafficking pathway of S. cerevisiae355
G.21 Genes differentially transcribed following exposure to 10% FCS (1 hour) mapped to the orthologous meiosis pathway of S. cerevisiae 356
G.22 Genes differentially transcribed following exposure to 10% FCS (1 hour) mapped to the orthologous cell cycle pathway of S. cerevisiae 357
G.23 Genes differentially transcribed following exposure to 10% FCS (1 hour) mapped to the orthologous MAPK signalling pathways of S. cerevisiae . . . 358

15
List of Tables
3.1 Conservation of induced transcripts between publicly available, hyperosmotic stress affected, transcript profiles in S. cerevisiae, C. albicans and this study 88
4.1 Over-represented Gene Ontology terms among differentially expressed genes 146 4.2 Functional classification of selected genes with altered transcript abundance,
relative to laboratory culture, in the murine lung 147 4.3 Distribution of induced and repressed genes among lineage specific cohorts
and chromosomal locations 150 4.4 Comparative analysis of genetic distribution among genes having differen-
tial transcript abundances in microarray analyses using A. fumigates RNA following exposure to in vitro or murine-adaptive stress, or laeA gene deletion.159
A.1 Factors involved in the development of opportunistic mycoses 199
B.1 Recipe for Synthetic Complete culture medium (1 L) 205 B.2 Recipe for Yeast Extract Peptone Dextrose culture medium (1 L) 205
C.1 Fungal strains used in this study 206
D.1 Oligonucleotides used in the three step PCR strategy for HOG1 disruption in C. glabrata 207
D.2 Biological effects encoded by linear model contrasts 208 D.3 Candida glabrata transcripts induced in wild-type cells following exposure to
YEPD + 1M sorbitol for 15 minutes 209 D.4 Candida glabrata transcripts repressed in wild-type cells following transient
exposure to 1M sorbitol for 15 minutes 224 D.5 Profile of transcripts involved in metabolism in wild-type, hog1 and stell cells228 D.6 Profile of transcripts involved in protein degradation in wild-type, hog1 and
ste11 cells 230 D.7 Profile of transcripts involved in protein trafficking in wild-type, hog1 and
stell cells 231 D.8 Profile of transcripts involved in transcriptional regulation in wild-type, hog1
and ste11 cells 233 D.9 Profile of induced transcripts conserved between S. cerevisiae (Gasch et al.,
and C. glabrata after exposure to hyperosmotic stress 234

16
D.10 Profile of induced transcripts conserved between S. cerevisiae (O'Rourke & Herskowitz 1M sorbitol) and C. glabrata after exposure to hyperosmotic stress236
D.11 Profile of induced transcripts conserved between S. cerevisiae (Rep et al.,) and C. glabrata after exposure to hyperosmotic stress 237
D.12 Profile of induced transcripts conserved between S. cerevisiae (O'Rourke & Herskowitz 0.5M KC1) and C. glabrata after exposure to hyperosmotic stress 239
D.13 C. glabrata hyperosmotic stress repressed transcripts that depend on a func- tional putative SHOI signalling branch 241
D.14 C. glabrata hyperosmotic stress induced transcripts that are independent of HOG1 and STE11 245
D.15 HOG1-dependent hyperosmotic stress induced C. glabrata transcripts . . 250 D.16 STE/ / -dependent hyperosmotic stress induced C. glabrata transcripts . . 252 D.17 C. glabrata hyperosmotic stress repressed transcripts that depend on a func-
tional putative SHOI signalling branch 256 D.18 HOG1-dependent hyperosmotic stress repressed C. glabrata transcripts . 258 D.19 STE/ / -dependent hyperosmotic stress repressed C. glabrata transcripts . 259 D.20 S. cerevisiae transcripts whose hyperosmotic stress dependent induction is
affected by HOG1 deletion. Data from Rep et al., 260 D.21 Betweenness centrality measures and degree of tube stations in the London
Underground network 261 D.22 C. glabrata ORFs with dNIdS > 1 265 D.23 Profile of induced transcripts conserved between S. cerevisiae stell (O'Rourke
& Herskowitz 0.5M KC1 sorbitol) and C. glabrata after exposure to hyperos- motic stress 272
E.1 RNA Samples and associated amplification factors 280 E.2 Filtering per array in the amplification study 281 E.3 Over-represented Gene Ontology (GO) terms amongst genes having increased
transcript abundance. 282 E.4 Over-represented Gene Ontology (GO) terms amongst genes having repressed
transcript abundance. 287 E.5 A. fumigatus genes having increased transcript abundance, relative to labo-
ratory culture, in the murine lung and S. cerevisiae orthologs regulated by rapamycin-mediated TOR kinase inhibition 292
E.6 A. fumigatus genes having decreased transcript abundance, relative to labo-ratory culture, in the murine lung and S. cerevisiae orthologs regulated by rapamycin-mediated TOR kinase inhibition 294
E.7 A. fumigatus specific genes having elevated transcript abundance in the murine lung, relative to laboratory culture. 298
E.8 A. fumigatus specific genes having repressed transcript abundance in the murine lung, relative to laboratory culture. 299
E.9 Affc-specific specific genes having elevated transcript abundance in the murine lung, relative to laboratory culture 300
E.10 Affc-specific specific genes having repressed transcript abundance in the murine lung, relative to laboratory culture 301

17
E.11 Nitrogen starvation gene clusters 303 E.12 Oligonucleotides used in qPCR 305
F.1 Summary of null mutant CGTFKOLib strain growth relative to his3 parental strain in YEPD and YEPD + 5 - 10 pg m1-1 fluconazole.
F.2 Summary of null mutant CGTFKOLib strains showing insufficent growth in YEPD (0-11.5 hours) for nonlinear regression based estimation of midpoint and upper asymptote
F.3 Summary of null mutant CGTFKOLib strains showing insufficent growth in YEPD + 5pgm1-1 (0-11.5 hours) for nonlinear regression based estimation of midpoint and upper asymptote
F.4 Summary of null mutant CGTFKOLib strains showing insufficent growth in YEPD + 10,u,gm1-1 (0-11.5 hours) for nonlinear regression based estimation of midpoint and upper asymptote
F.5 Summary of hierarchical clustering membership with associated median growth rates in YEPD ± fluconazole (5 - 10pg m1-1)
F.6 Effect of timepoint and growth condition on Z factor F.7 Fitted parameter values for a linear model describing optical density at 11.5
hours as a function of strain genotype, growth condition and the interaction between growth condition and genotype. Plate 1 of 2
F.8 Fitted parameter values for a linear model describing optical density at 11.5 hours as a function of strain genotype, growth condition and the interaction between growth condition and genotype. Plate 2 of 2
G.1 Candida glabrata transcripts induced in ATCC 2001 following exposure to Synthetic Complete + 10% (v/v) foetal calf serum (FCS)
G.2 Candida glabrata transcripts repressed in ATCC 2001 following exposure to Synthetic Complete + 10% (v/v) foetal calf serum (FCS)
G.3 Over-represented GO component terms for C. glabrata transcripts induced by 10% FCS (w/v) 432
G.4 Over-represented GO process terms for C. glabrata transcripts induced by 10% FCS (w/v) 432
G.5 Over-represented GO function terms for C. glabrata transcripts induced by 10% FCS 433
G.6 Over-represented GO component terms for C. glabrata transcripts repressed by 10% FCS (w/v) 433
G.7 Over-represented GO process terms for C. glabrata transcripts repressed by 10% FCS (w/v) 433
G.8 Over-represented GO function terms for C. glabrata transcripts repressed by 10% FCS 434
307
311
311
313
319 319
320
324
359
39'7

18
Nomenclature
lc Micro - 10-6
Microgram
Ohm
°C Degree celsius
AA Absorbance at A
aH Activity of hydrogen ions
L. Chirality identifier: laevorotatory
as amino acid
ABC ATP binding cassette
ACM Aspergillus complete medium
AMM Aspergillus minimal medium
ATCC American type culture collection
ATP Adenosine triphosphate
BLAST Basic local alignment search tool
bp Base pairs
cDNA Complementary DNA
CFU Colony forming units
CHiP Chromatin Immunoprecipitation
COGEME Consortium for the Functional Genomics of Microbial Eukaryotes

19
CWPs Cell wall proteins
Cy3 Cyanine 3. Fluorescent label incorporated in deoxyribonucleosides.
Cy5 Cyanine 5. Fluorescent label incorporated in deoxyribonucleosides.
dATP Deoxyadenosine triphosphate
dCTP Deoxycytidine triphosphate
dGTP Deoxyguanosine triphosphate
DMSO Dimethyl sulphoxide
DNA Deoxyribonucleic acid
dTT Dithiothreitol
dTTP Deoxythymidin triphosphate
EASE Expression Analysis Systematic Explorer
EDTA Ethylenediaminetetraacetic acid
ER Endoplasmic reticulum
ESR Environmental stress response
F Farad
FCS Foetal calf serum
g Gram
GDP Guanosine diphosphate
GO Gene ontology
GPR Genepix results
h Hour
H202 Hydrogen peroxide
JCVI The J Craig Venter Institute of Genomic Research
KAN Kanomycin

20
kb Kilobase
KEGG Kyoto Encyclopaedia of Genes and Genomes
kg Kilogram
kV Kilo volt
L litre
LB Lurian Bertinin
log2 Binary logarithm. For 16 bit images, the theoretical range of microarray intensities
is 216. Binary logarithm transformation puts these values in the range r < 16 > 0
Lowess Locally weighted scatterplot smoothing
M Molar
m Milli - 10-3
M13 Bacteriophage M13.
MAPK Mitogen activated protein kinase. See MAPKKK
MAPKK Mitogen activated protein kinase kinase. See MAPKKK
MAPKKK Mitogen activated protein kinase kinase kinase. Hierarchical nature of this class of protein's activity is reflected in the nomenclature. MAPKKK phosphorylate
MAPKK which in turn phosphorylate MAPK
mg Milligram
min Minutes
MLT Murine lung tissue
mM Millimolar
mRNA Messenger RNA
Mw Molecular weight
n Nano - 10-9
NADPH Nicotinamidr adenine dinucleotide phosphate
NO Nitric oxide or nitrogen monoxide

21
Null mutation A mutation that results in the complete loss of function of the encoded gene product
ODA Optical density at wavelength A
ORF Open reading frame. Stretch of DNA that could potentially be translated into a polypeptide or RNA
PAMP Pathogen associated molecular pattern
PCR Polymerase chain reaction
pH The decimal logarithm of the hydrogen ion activity in aqueous solution. A measure of acidity and alkalinity of a solution -logioax
PLOS Public library of science
R The R statistical computing framework
Regulon Collection of genes under regulation by the same regulatory protein
RIA Reductive iron assimilation
RNA Ribonucleic acid
ROS Reactive oxygen species
RPM Revolutions per minute
RT Reverse transcription
s Second
SAP Secreted aspartic proteases or secreted aspartyl proteases
SAPK Stress activated protein kinase
SDS Sodium dodecyl sulphate
SGD Saccharomyces genome database
SH3 SRC homology domain 3
STM Signature tagged mutagenesis
t-BOOH tertiary butyl hydroperoxide or ,1-1, dimethylethyl hydroperoxide
T-DNA Transferred DNA of tumour inducing plasmid

22
T7 Bacteriophage T7
Tet Tetracycline
TF Transcription factor
TIGR The Institute of Genomic Research
TLR Toll-like receptor
TOR Target of rapamycin
UTR Untranslated region
UV Ultraviolet
UV Volts
v/v Volume / volume. Percentage solution expressing volume of the concentrated solu- tion as a proportion of dissolving solvent (in excess)
v/v Weight / volume. Percentage solution expressing mass of the solute as a proportion of dissolving solvent
WT Wild-type
YEPD Yeast extract peptone dextrose
YKO Yeast knock-out
YPD YEPD

23
Chapter 1
Introduction
1.1 The increasing prevalence of opportunistic mycoses
The increasing incidence of life-threatening mycoses emerged as a public health concern in the last three decades of the twentieth century (456; 156; 259; 479; 411). It steadfastly remains so, with the rapidly widening spectrum of opportunistic agents outpacing the emer-gence of effective, novel therapies (Supplementary Figure A.1 and A.2). This increase has been attributed, in part, to a shift in western demography and medical practise. The use of broad-spectrum antibiotics, cytotoxic chemotherapies and transplantation in combination with an increasingly aging, immunocompromised population elevates the risk for both com-mon and uncommon opportunistic mycoses (507; 541; 724). The majority of these mycoses come from the Candida (72.8 million cases annually), Cryptococcus (65.5 cases annually) and Aspergillus (12.4 million cases annually) species (542), ubiquitous organisms (107; 394) that can colonize a range of mammalian anatomical sites. Unlike primary pathogens, where disease incidence is influenced by the communicability of the organism, Candida and As-
pergillus - the pathogenic fungi studied in this thesis - are ubiquitous, and transmission of the organism to a susceptible individual is not a major factor in disease propagation. More important, is the degree and heterogeneity of immune dysfunction required for these otherwise commensal organisms (202; 704) to evade the immune system, proliferate and affect human health (Supplementary Table A.1).

1.2 The medical burden of invasive candidiasis and invasive pulmonary aspergillosis 24
1.2 The medical burden of invasive candidiasis and invasive pulmonary asp ergillosis
1.2.1 Candida spp. and candidiasis
Candida is a heterogeneous and taxonomically challenging genus of 154 species belonging to the Ascomycetes class, which accounts for, approximately, 50% of all known fungal species and 80% of primary and opportunistic-pathogen species (258). The majority of Candida
spp. are not associated with human morbidity or mortality and exist as commensals of a healthy oral, gut, skin, and vaginal flora as well as saprophytes in other non-mammalian niches'. Increasingly, in some at-risk groups, emerging Candida spp. can switch from be-nign commensals to well adapted and prolific pathogens. Systems for fungal taxonomy2 are
vital for accurate diagnosis of aetiological agents of candidiasis and aspergillosis, early and accurate diagnosis of which can improve clinical outcomes (720; 7). Candida are morpho-logically and sexually diverse, ranging from meiospores (714; 107) to obligate mitospores, with some organisms able to produce colonies containing budding yeasts, pseudohyphae and true hyphae (reviewed in (253; 92; 348)) - features that have been used as distinguishing characteristics in fungal systematics. Fungal taxonomists have traditionally used multiple classifying dimensions3, often without consensus, to classify species. Microscopically iden-tified morphological differences have perhaps been the most commonly adopted, but these techniques do do not accurately depict evolutionary relationships with a common ancestor. These traditional systems of Ascoinycete classification have been criticized for their reliance on an insufficient number of distinguishing morphological characteristics to resolve higher taxonomic relationships and have recently been augmented with methods based on molec-ular sequence evolution, which is becoming increasingly available in the post-genomic era of fungal biology.
In the post-genomic era (reviewed in (457)), fungal research is benefiting from the ac-crual of molecular sequence data (reviewed in (455)) and phylogenetic inference methods (198; 148; 763). Between 1990 and 2003, 560 fungal research papers reporting phylogenies were published, of which approximately 84% were derived using the ribosomal DNA (rDNA) (119). The 18S rDNA gene is frequently chosen for phylogenetic inference because it is ubiq-uitous, vertically transferred and has slowly evolving sites. Nevertheless, phylogenies based
'Such as on the surface of fruits in the soil and in leaf litter and the guts of insects that feed on it (497). C. glabrata can be a major cause of food spoilage. Paradoxically, it has also been used as a competitive agent in the prevention of Botrytis rot in strawberries (186)
2comprehensively reviewed in (258).
3These include morphological, physiological and biochemical techniques (194), diagnostic use of secondary metabolite profiles (527; 215), cell wall composition and protein composition (258)

Chapter 1. Introduction 25
on a single-gene do not necessarily reflect the ancestry of the organisms possessing them and can be affected by lateral gene transfer4. This is particularly applicable to protein coding genes - phylogenies based on genes involved with vital physiological or adaptive processes do not always correlate with rDNA derived trees, but they do have the capacity to resolve deep taxonomic relationships. In an attempt to combine the beneficial attributes of both phylogenetic approaches, researchers have sought to incorporate all available phylogenetic data to form consensus trees.
Recently, Fitzpatrick et al., constructed a fungal phylogeny based on 42 complete genomes derived from a supertree and combined gene analysis (205). This method used 4,805 orthologous genes families and the resulting gene-family phylogenies as input (Figure 1.1) for two different consensus tree inference methods: matrix representation with parsi-mony (MRP) and the average consensus method (AV). The consensus phylogenies provide supporting evidence for the classification of phyla, sub-phyla and orders and also shows ev-idence that the Saccharomycotina is a monophyletic Glade containing organisms that either translate CTG either as serine or as leucine. This Glade contains two sub-groups, one con-taining the fully sexual species Candida lusitaniae, Candida guilliermondii, Debaryomyces hansenii and the second group containing Candida albicans, Candida dubliniensis, Candida tropicalis and Lodderomyces elongisporus. This CTG Glade is distinct from species that have undergone a genome duplication event - which notably contains Candida glabrata, in phylogenetic terms a closer relative of Saccharomyces spp. than Candida spp.
Genome duplication events are believed to be important evolutionary processes in a variety of eukaryotes, where they play a role in the evolution of new molecular functions (515; 6; 654). The sequencing of Saccharomyces cerevisiae genome (246) has provided new insight into the evolutionary processes that have shaped its genome size and content. Shortly after the S. cerevisiae sequence was published, Wolfe and Shields (752) proposed that the species is a degenerate tetraploid resulting from whole genome duplication after its divergence from Kluyveromyces lactis based on the number and orientation of these conserved blocks, further evidence being provided by analyzing paired regions between the Kluyveromyces waltii and S. cerevisiae genomes (352). With the rapid accumulation of other hemiascomycete genome sequences (246)5 Dujon and co-workers (182) investigated the distribution of tandem gene duplications and blocks of ancestral duplications between S. cerevisiae, C. glabrata, K. lactis, D. hansenii and Y. lipolytica. They demonstrated that duplicate blocks exist in each of the four investigated species, but their number decreases with the evolutionary distance from S. cerevisiae and are highly conserved between S.
4this is common in prokaryotes
5For details of sequencing efforts consult references in (182)

1.2 The medical burden of invasive candidiasis and invasive pulmonary aspergillosis 26
Zygomycota
Basidiom cota flAri/upo tallSdis kirialims r l 111"
02.71/000(1'41A ne0,0.11 rn Grlt
Pirairermtmete el;ry mnpvirnr Hymenamycetes TOD Cop, tent riiirmiA
SchimThruharum,vrm pnrthr
16')
Sordarlomycetes„,...16
t v•
Trilluntermer rrrsrl 0CLit.varru on m grormincorr
rIrTiTrintf71 ITnicilliVideS
N riA,e4: "V run. Amine CIMAGI
rikihnpOrei Me/WM
C kiernmirim gfnincurr Pezizornycotina
Ascomycota
Stagonospora etothirum liistopia eiVilfhl Win
tu:) Uncinrarrpri frrsii 1001.—CaTirlioideN irrmitis ktpergillus aitlufwi AverOlktine*atus. ice
80 AAffe.twitki 1:ry,-fir 63 Aspergiihrs farms
Yorrricia lirolytfrct
TOO
71; t toy
Etirotionycetes
160 ta:„..4*C""f}tdryPt Leotiomycetes Sete rutini mit. rot foram
Candi Al XIII 11 rakl
70
Saccharomycotina
GTG
WG0as
5t Sfweivroviyers mairkle
04 f IITJ1171rr t (*Mirth;
StirrhILIVITIrri r re 111 icra• 10
'7 Sarcharn,wyrrs pgradorKv 95
SIN.ThaMinyet'S htlyallfiS
'01"..54.ri)111/7:/ttp:V.V
rliattlfi.yrrA.I'liY ;;IL;;":7(7" 10
athyd grmAypii Kit Tvirorykys farris Corditht heti torthre Candid(' gnilliermonchi
081 priN fri*T.VITyrrS Linn rnii Caadi &it pqraimllasis
trevideeth.i 1
ice
ice 4.N yirwrts
dflithriir#1.41'
4.1
Figure 1.1: Maximum likelihood phylogeny of 153 universally distributed fungal genes. The concatenated alignment contains 42 taxa and exactly 38,000 amino acid posi-tions. Taken from Fitpatrick et al (205). At the bottom of the tree, the Saccharomycotina family consist of two clades encompassing Candida species that translate CTG as serine in-stead of leucine. This Glade can be further divided into species that are meiosporic. The second major Glade within the Saccharomycotina contains species who have undergone a whole genome duplication (WGD).

Chapter 1. Introduction 27
cerevisiae and C. glabrata but not between other species. Although the number and degree of conservation of these blocks is higher between S. cerevisiae and C. glabrata, they are structurally different, the latter, on average, being smaller (27 - 89 kb versus 42 - 243 kb in S. cerevisiae), a feature of reductive evolution in C. glabrata. These inferences from molecular sequence data imply that C. glabrata is phylogenetically distant from its other pathogenic relations, and is more closely related to organisms that have undergone a whole genome duplication and are not normally associated with disease (189). This evolutionary proximity of C. glabrata and Saccharomyces sensu strict() species presents the model organism S. cerevisiae, with its comprehensive molecular tools (55; 634; 595; 559; 749; 239; 173), as useful resource for the study of conserved biological processes of relevance to pathogenesis. Candida species can colonize a range of anatomical sites in immunocompetent hosts (causing genitourinary, oropharyngeal , vulvovaginal and cutaneous infections (107)), where evasion of the immune system and proliferation of the organism can lead to medical complaints with varying degrees of morbidity and mortality. Genitourinary (40) and superficial infections such as vulvovaginal and oropharyngeal candidiasis (28; 29) are common, a leading cause of GP visits and prescription expense, but not life-threatening (645). Although superficial Candida infections are trivial in terms of mortality, impairment of epithelial barriers at these anatomical sites can facilitate hematogeneous dissemination of the organism, a condition called invasive candidiasis (IC).
The medical burden of such invasive Candida infections has been evaluated using analy-ses of hospital discharge information (745; 542; 736) and population surveillance initiatives (751). Estimates vary substantially for the number of invasive candidiasis infections in the US (ranging from 7,000 (736) to 63,000 per annum (542)), but crude mortality is more stably estimated around 40%. Taken together, these results have been used to estimate the annual deaths associated with nosocomial candidiasis in the US to be in the range of 2,800 to 25,300. A long-term study of nosocomial fungal infections in the U.S.A6 reported that the Candida species were the most prevalent, accounting for 78.3% of cases (214). Although C. albicans remains the principal infective species, recent epidemiological evidence indicates an increased prevalence of the non-albicans species (496; 64)7. In a seven year study of 24,179 nosocomial bloodstream infections (BSI), conducted in the USA, Wisplinghoff et al., ranked Candida species as having the fourth highest BSI incidence rate, behind Staphylococcus au-reus and Enterococcus species (751). To date, more than 17 aetiological agents of human IC have been identified, but 90% of invasive infections are due to only five species: C. albicans,
6This study, of fungal and non-fungal infections included surveillance cultures from 115 hospitals between 1980 - 1990. The data are collected by the National Nosocomial Infections Surveillance system
7Concomitant advances in classification may have also increased detection rates, suggesting that the overall epidemiology might not have changed so significantly. See Haynes (278) and references therein

1.3 Current treatment options for systemic mycoses: the need for better anti-fungal drugs 28
C. glabrata, C. tropicalis and C. krusei. C. albicans is the most common of clinical Candida isolates in BSIs (66% of all Candida species). Surveillance studies have shown that the rates are decreasing with simultaneous increases in incidence rates of other Candida spp (Supplementary Figure A.2). Of these emergent species, C. glabrata has particular medical significance because it is associated with high mortality (748; 212) and increased anti-fungal resistance (543).
1.2.2 Aspergillus fumigatus and aspergillosis
Invasive aspergillosis describes several diseases (aspergilloma, allergic bronchopulmonary aspergillosis and invasive aspergillosis) that can affect several anatomical sites (lungs, liver, brain and kidney) whose aetiological agents belong to the genus Aspergillus. Of the ap-proximately 185 Aspergillus species identified to date, approximately 11% (20 species) are recognised as human pathogens. Prevalent species include Aspergillus fumigatus, Aspergillus nidulans, Aspergillus flavus, Aspergillus clavatus, and Aspergillus niger (394). Invasive pul-monary aspergillosis (IPA) is the most life-threatening of these diseases, with an associated mortality of 70-90 % (168) and is principally caused by A. fumigatus (394). Although the spores of this organism are among the least prevalent found in air samples (616), they are disproportionately linked to the majority of air-borne fungal infections (616). The size of A. fumigatus spores (2-3 ii1VI diameter) not only facilitates their sustained dispersion (394), but also permits entry into the pulmonary alveoli of the mammalian lung (576), whereupon the level of host immunocompetence (95; 169; 168; 583; 371), and the organism's ability to adapt to environmental stress, determines its fate, and conversely, the morbidity of the host (394; 773). Invasive pulomonary aspergillosis (IPA) has become a leading cause of death among haematology patients. The average incidence of IPA is estimated to be 5 to 25% in patients with acute leukemia, 5 to 10% after allogeneic bone marrow transplant, and 0.5 to 5.0% after cytotoxic treatment of blood diseases or autologous bone marrow transplant and solid-organ transplantation. IPA which follows solid-organ transplantation is most common in heart-lung transplant patients (19 to 26%) (reviewed in (394)). The emergence of such pernicious, life-threatening mycoses necessitates effective therapies. In the following section, treatment options and their limitations are briefly reviewed.
1.3 Current treatment options for systemic mycoses: the need for better anti-fungal drugs
In the absence of clinically available vaccines (311), the primary therapeutic option for the treatment of mycoses is the use of small molecule anti-fungals such as the polyenes, allylamines and azoles, the majority of which owe their activity to the physical disruption

Chapter 1. Introduction 29
of ergosterol or its biosynthesis (mechanisms of action are reviewed in (238)). Ergosterol is required for optimal functioning of membrane-bound enzymes and transporters (706; 155), pheromone signalling (332) and is a key determinant of plasma membrane structure (2). The most established drugs used in the treatment of invasive mycoses belong to the polyene class of anti-fungals. Members of this class, such as nystatin and amphotericin B, have been available for several decades, and amphotericin B was the standard therapy for candidemia for many years (668). By binding directly to cell membrane sterols the larger polyenes, such as amphotericin B are thought to form pores lined by the inward facing polyene hydroxyl residues (160; 294). These pores alter membrane permeablility and lead to leakage of important cytoplasmic components including intracellular potassium and ultimately cell death (354; 355). Although amphotericin B has been the gold standard therapy for invasive fungal infections (660), a narrow therapeutic index (238) and poor efficacy (53) necessitate other clinical options.
Despite the existence of many licensed anti-fungal molecules, less than half of them have a route of administration that is suitable for treatment of invasive mycoses8. The azole class of anti-fungals (273; 613; 599; 101; 503) was developed in order to improve upon the poor bioavailability of contemporary anti-fungals and until recently had been used ex-tensively in the treatment and prophylaxis of invasive candidiasis (124) and aspergillosis (187). The azoles (e.g. fluconazole, itraconazole and posaconazole) disrupt the biosyn-thesis of ergosterol by inhibiting catalytic enzymes involved in the step-wise conversion of lanosterol to ergosterol (Supplementary Figure A.3). The resulting altered flux in this path-way results in the depletion of ergosterol and the concomitant accumulation of 14a-methyl sterols (lanosterol, 4,14-dimethylzymosterol and 24-methylenedihydrolanosterol) whose C-4 methyl groups contribute to the altered structural properties of the azole effected fungal cell membrane. The primary target of the first azoles is encoded by ERG11 (also known as Cyp51 in Aspergillus), whose protein product lanosterol 14-a-demethylase co-catalyzes cytochrome P-450-dependent 14a-demethylation of lanosterol (288), although more recent triazole derivatives9 partially owe their activity to inhibition of cytochrome P-450 depen-dent 14a-sterol demethylase (encoded by ERG24 ) (604). The widespread use of the effica-ceous fluconazole has been tempered by the emergence of drug resistance in Aspergillus spp (298; 120; 250) and Candida spp. (543).
Although compensatory mechanisms exist to scavenge exogenous sterols for example in C. glabrata (492), azole resistance is generally caused by diminishing the concentration of the active compound at the active site of the target enzyme. This can be achieved in several
8Data obtained from http://www.accessdata.fda.gov/scripts/cder/drugsatfda/
8such as fluconazole, itraconazole and voriconazole

1.3 Current treatment options for systemic mycoses: the need for better anti-fungal drugs 30
ways, including mutations in the target enzyme effecting drug but not natural substrate binding (712; 741), overproduction of the target enzyme (711), biophysical changes in the membrane that impede intracellular azole accumulation (437), and active efflux mechanisms (reviewed in (110)). The molecular basis of such azole efflux mechanisms has been well characterized in S. cerevisiae, C. albicans and C. glabrata. There are two main classes of efflux pumps that act to translocate anti-fungal compounds across the cell membrane: i) the ABC (ATP-binding cassette (335)) transporters and ii) the MFS (major facilitator superfamily (600)) pumps. ABC transporters are primary transporters, using the energy derived from ATP hydrolysis to pump azoles out of the cell and include the pleiotropic drug resistance (PDR) group of transporters such as Pdr5p identified in S. cerevisiae (46; 69; 306) and orthologues in C. albicans, C. glabrata (697; 719; 606) and A. fumigatus. An elegant study by Thakur et al probed the mechanism of Pdrlp-mediated transcriptional control of efflux pumps and demonstrated that direct binding of anti-fungal molecules to Pdrlp causes this transcription factor to activate transcription of efflux pumps via an interaction with the mediator complex subunit Ca111p (687) in S. cerevisiae and C. glabrata. MFS transporters are secondary transporters that utilize proton-motive force across the plasma membrane to actively translocate azoles out of the cell and include the C. albicans Mdrlp, Flu1p and A. fumigatus Mdr3p proteins (110). The importance of efflux mechanisms in Aspergillus spp. is less clear. Much of the research performed in A. nidulans has identified candidate efflux transporters (21; 165; 23), however few clinical isolates of A. fumigatus overexpress efflux pumps, and there is evidence to suggest that target enzyme alteration is a major mechanism in addition to other uncharacterized mechanisms (154; 442; 493).
Clinicians have sought to quickly address the problem of emerging drug resistance by investigating the efficacy of combination therapy with existing drugs. Sanati at al (605) investigated the effects of amphotericin B and fluconazole monotherapy and combination therapy on mortality and tissue fungal density in neutropenic mice. They demonstrated that the survival of neutropenic mice with disseminated candidiasis was prolonged when animals were treated with a combination of amphotericin B and fluconazole. This seemingly strong in vitro evidence in favour of combination therapy belies the clinical, pharmacoeco-nomic and regulatory drawbacks. Clinical trials of combination therapy are limited and often empirical, and concerns remain over the putative mechanism and the increased po-tential for adverse pharmacokinetic interactions (370; 406; 713; 256). In their attempts to exploit the unmet medical need for efficacious, cost-effective anti-fungal therapies, pharma-ceutical companies have had some success with a new generation of molecules such as the triazoles (posaconazole, isavuconazole, ravuconazole and albaconazole) and echinocandins (caspofungin, micafungin and anidulafungin) (199; 244). Voriconazole is now the primary treatment of most forms of invasive aspergillosis. A clinical trial comprising 277 patients

Chapter 1. Introduction 31
with invasive pulmonary aspergillosis demonstrated that a successful outcome was observed in 52% of patients treated with voriconazole in comparison to 32% of patients who received amphotericin B (283). Caspofungin is better tolerated and as effective as amphotericin B (651) and voriconazole was as effective as amphotericin B followed by fluconazole in a large clinical trial that included 370 patients with systemic candidiasis (378). Mortality rates were comparable in the groups that received voriconazole or fluconazole with amphotericin B (36% versus 42% respectively), however voriconazole led to much better treatment out-comes for patients infected with C. tropicalis. These are small advances in clinical outcomes, however, significant cross-resistance to next generation triazoles has also been reported af-ter prolonged exposure to fluconazole and itriconazole (523). Although triazoles are an effective option for new cases, patients with persistent candidemia and prior second or first generation azole exposure may still have few effective treatment options. Clearly, this poses a major challenge for clinicians and drug discovery companies alike: how can such increas-ingly prevalent, drug-resistant, pernicious mycoses be managed? Focusing on such narrow range of targets restricts the chemical space that can be explored in designing potent, selec-tive and safe drug molecules. Indeed, a recent chemical genomics (287) study in which 316 unique compound treatment conditions spanning established anti-fugal molecules, natural products and chemical probes and showed that 97% of the knockout strains were suscepti-ble to at least one drug condition. This systematic investigation indicates that there is a largely unexplored chemical space available for exploitation. In this clinical context, biolo-gists aim to understand the molecular basis of virulence. In doing so they hope to identify and exploit new anti-fungal targets - aspects of fungal genetics that are important for fungal pathogenesis.
1.4 Identifying therapeutic targets: strategies for identifying virulence determinants in Candida spp. and A. fumigatus
Molecular mycologists have employed a variety of molecular and genetic techniques to iden-tify criticial determinants of virulence, including gene deletion strategies, gene expression strategies, and the use of systematic disruption libraries in the model organism S. cerevisiae to characterize processes required for virulence. Before the advent of genomics, classical for-ward genetic approaches' were the cornerstone of research in sexually reproductive species such as A. nidulans and S. cerevisiae and have been used extensively to formulate ge-netic maps of such diverse cellular processes as nitrogen-metabolite and carbon-catabolite
10These involve identifying a chemically or physically induced mutation that confers a phenotype of inter-est, followed by characterization of a wild-type copy of the mutated gene, e.g using functional complemen-tation.

1.4 Identifying therapeutic targets: strategies for identifying virulence determinants in Candida spp. and A. fumigatus 32
repression (34), conidial development and mitosis (481) Unfortunately, these classical techniques are not yet accessible to map the virulence phe-
notype or related phenotypes in Candida spp and A. fumigatus. Most Candida species are
diploid and devoid of a sexual cycle. This limits the frequency at which storable recessive mutations are obtained and prevents segregation of relevant genotypes of interest. Even with these limitations, chemical mutagenesis techniques were adopted in early A. fumiga-
tus studies (365; 372) but were largely abandoned in ensuing years. However, the recent exciting discovery of a sexual cycle in A. fumigatus (513) may yet re-open this avenue of
virulence research in A. fumigatus. In the absence of suitable forward genetics approaches, functional complementation studies (5) have been used in C. albicans, C. glabrata and A.
fumigatus. This technique works by cloning a gene of interest, or a library of genes into a heterologous host to complement a defined phenotype. It has been used to identify molec-ular determinants of epithelial adhesion (231; 220), multi-drug resistance (207; 8; 677) and morphogenesis (663), in C. albicans; cell wall maintenance (734; 490; 500) in C. glabrata
and calcium homeostasis (650) in A. fumigatus. Functional complementation experiments make an assumption that the transforming gene is expressed in the heterologous host in the same manner to its native locus in the donor strain, and that it has the same function in the host genetic context. However, fundamental differences relating to codon usage (608; 91), and to background-specific functions have been reported (677).
Reverse genetics approaches approaches have also been adopted to associate loci with phenotypes. Unlike classical forward genetics studies, reverse genetics methods expose a strain, typically deficient in a known protein-coding chromosomal locus, to a selection of in vitro assays designed to evaluate the contribution of that locus to a specific biological process. Typically, this leads to a loss of function in in vitro assays or in in vivo models of invasive mycosis, however some studies have revealed enhanced function in the null mutant strain. For example deletion of the C. glabrata ACE2 locus results in hyper-virulence
in a murine model of candidiasis (341), and deletion of CAS1, a C. neoformans putative
membrane protein rendered cells more virulent (324). The development of high copy number plasmids (545), robust transformation protocols (247; 582; 171) and multiple selectable markers (582; 628), facilitated gene disruption techniques (208; 746; 483; 750) in C. albicans, elucidating the function of genes involved in e.g. invasive growth and virulence (728), salt tolerance (647) and gluconase function (251). Similarly, transformation procedures (519) and single gene disruption techniques using antibiotic resistance markers (447; 561) or auxotrophic markers (166) have been developed in A. fumigatus, elucidating the function of genes involved in virulence related processes such as calcium tolerance (649), multi-drug resistance (460), cation transport (423), iron assimilation (618) and alkaline adaptation
(66).

Chapter 1. Introduction 33
Although some progress has been made in functional genomics of pathogenic fungi, the laborious nature of targeted gene disruption methods has limited its scope and impact in C. albicans, A. fumigatus and C. glabrata. To date there are no genome-wide knockout collections for these organisms and this still represents a significant technical challenge. A genome-wide knockout collection is of particular value because it enables unbiased pheno-typic screens, in contrast to small scale targeted single gene deletions which are usually chosen on the basis of homologues in a related species. In their excellent review, Scherens and Goffeau (614) summarized the value of this unbiased approach by focusing on the S. cerevisiae knockout library (749). At the time of their publication, they identified 33 pub-lished studies which used this collection to screen approximately 100 conditions, identifying 5000 new phenotypes including over 100 new genes resulting in a defective bipolar budding pattern (498) and 71 essential genes that were previously thought to be dispensable (257). Massively parallel random mutagenesis techniques were developed to maximize genome cov-erage with limited resources (595; 282; 98; 116; 135; 356). Variations on this approach have been adopted to successfully identify virulence determinants in A. fumigatus (94) and C. glabrata (342; 341; 139; 141). Despite these positive findings, the propensity for mobile elements to integrate in non-coding regions and limited transformation efficiency means that exhaustive coding region coverage has not yet been achieved and systematic unbiased screens consequently remain elusive.
In contrast, collaborative targeted disruption programs, although labour intensive, pro-vide much greater coverage of the target genome. This has been performed on an industrial scale in S. cerevisiae (749; 239), and there are near comprehensive libraries of null mu-tants in this organism. The collection has also been informative in dissecting the molecular basis of fundamental biological processes such as meiosis and sporulation (498), chromoso-mal segregation (445), cytoskeletal organization, cell-wall biosynthesis, microtubule-based chromosome segregation and DNA metabolism (692; 693), glycogen accumulation (747), accumulation of mutations (307), response to abiotic chemical stresses induced by a va-riety of drug molecules including anti-fungal agents and anti-cancer drugs (25; 26; 240) and has also made a large contribution to our understanding of conserved signal transduc-tion pathways pertinent to Candida spp. (484; 63; 739; 310) and A. fumigatus virulence, although a more complete insight into the function of pathogen specific genes necessitates gene disruption in the appropriate species (61), moreover conserved genes can have opposite functions in related species, for example ace2 strains of S. cerevisiae and C. albicans are avirulent, whereas the C. glabrata ace2 strain is hyper-virulent in an immunocompromised mouse model (430).Gene disruption studies and heterologous expression studies mentioned above have also been allied with techniques that monitor the expression of genes within the host or in conditions that mimic environmental cues experienced within the host niche. In

1.5 A brief review of virulence factors important for Candida and Aspergillus mediated pathogenesis 34
non-fungal pathogens, where virulence may be associated with a single expressed genetic element, transcriptional profiling techniques compare the mRNA abundance of each gene in one or more conditions pertinent to pathogenesis.
This technique allows researchers to identify genes whose altered expression may be part of an adaptive response to environmental stress signals. Before gene expression microarrays were widely adopted, this was monitored in pathogenic organism with reporter gene assays, differential subtraction (68), RNA fingerprinting (133) or in vitro expression technology (IVET)
1.5 A brief review of virulence factors important for Candida and As-pergillus mediated pathogenesis
The techniques described in the previous section have elucidated important processes and molecules involved in pathogenesis caused by Candida spp and Aspergillus spp.; the follow-ing section is a non-exhaustive overview of some of those critical determinants of patho-genesis. Unlike some primary enteric pathogens, whose pathogenic properties are tightly linked to a very small set of molecules (e.g. type 3 secretion proteins of Salmonella spp. (671)), virulence determinants (VDs) of opportunistic fungal pathogens are more diffuse, and pathogenesis is determined by the overall fitness of the fungus within the host. Although some researchers favour the strict definition of a virulence determinant as a microbial en-coded entity that damages the host, a more subtle definition which presents virulence as a continuum in which host immunocompetence and pathogen fitness has been suggested by Casadevall and Pirofski (112). Much research has focused on the molecular and genetic nature of processes that enable fungal pathogens to adapt to the host niche. Although Can-dida spp and A. fumigatus primarily colonize different anatomical sites, the host imposes some common constraints on these opportunistic pathogens: they must adhere to mucosal layers of the host and sense and adapt to the environmental and nutritional stresses of their mammalian niche. Colonization of the host niche requires adherence to a substrate; a process that has been studied in depth in C. albicans and to a lesser extent in C. glabrata and A. fumigatus. Radioligand binding assays were used in conjunction with human buccal (HBECs) or vaginal cells (HVECs) to identify spontaneous, cerulenin-resistant strains of C. albicans with impaired adherence (108; 289) and to demonstrate a correlation between the degree of innate adherence of Candida species and their virulence (358). Techniques were later developed to identify the molecular determinants of adhesion, initially in S. cerevisiae, where heterologous gain of function in non adherent strains (231; 220) demonstrated the genetic basis of adherence, and subsequently in C. albicans (208). The molecular basis of this process has largely been dissected by Lois Hoyer (304) and colleagues.

Chapter 1. Introduction 35
They initially described the similarity of ALS1 gene product to the S. cerevisiae a—agglutinin, a protein that is involved in adhesion of a mating types to a ligand on the complementary a type cells during mating (413). The agglutinin-like sequence ALS gene family consists of at least nine genes (302; 231; 340; 303; 300) that share a conserved (55% - 90% sequence iden-tity) 5' domain, one or more putatively glycosylated central motifs of 108bp and a variable 3' domain. Immunohistochemical studies have shown that the gene products of ALS genes are localized to the cell surface in vitro and in vivo due to the amino and carboxy-terminal hydrophobic sequences that act as a secretory signal sequence and glycosylphosphatidyli-nositol (GPI) anchor addition site respectively (303; 301; 346). The amino terminal domain has been the focus of other structural (76; 301), although not crystallographic, and func-tional studies, with some groups predicting that it assumes an immunoglobin fold motif (76; 129). The abundant glycosylation observed in the carboxy-terminal and central motifs is likely to confer an extended conformation of this domain, whose exposed hydrophobicity may explain the adhesive nature of the molecule to epithelial cells.
Possibly as a consequence of its epidemiological importance, and the earlier availability of genetic tools for systematic investigations of virulence, the Candida literature offers many more molecular studies of virulence attributes in C. albicans than other species. Although commonly used as point of reference for discussion of the molecular basis of virulence in C. glabrata and other non-albicans species, this is not always useful. For example, although relatively well defined in C. albicans, adhesins required for virulence in models of candidiasis are not conserved in most other aetiological agents of invasive candidiasis. Inspection of the Candida genome database" (CGD) data shows that two genes of the ALS family that are required for virulence in mouse models of candidiasis (ALS1 (340) and ALS/ (770)) are not conserved in any of the major aetiological agents of IC apart from C. tropicalis (Sup-plementary Figure A.5). Indeed adhesion is mediated by a separate family in C. glabrata (140; 117; 1; 141), the EPA adhesins. Most of the genes belonging to this family are located in subtelomeric regions (1), which is an important determinant of their regulation. The importance of the chromosomal location was demonstrated by inserting DNA cassettes be-tween the telomere and EPA6 and EPA7 in order to vary their telomeric proximity (117). Large insertions, which increase the distance between the telomere and the EPA genes by approximately 18 kb resulted in hyperadherance, mediated by the loss of transcriptional silencing associated with their close proximity to telomeric regions. Transcriptional silenc-ing is a function of the SIR-family (silent information regulator SIR2p, SIR3p, SIR4p) modulated heterochromatic state of these chromosomal regions, which serves to restrict transcription in those areas. Studies of subtelomeric silencing in S. cerevisiae showed that
lhttp://www.candidagenome.org

1.5 A brief review of virulence factors important for Candida and Aspergillus mediated pathogenesis 36
a complex of Sir2p/Sir3p/Sir4p is recruited to the telomere by interaction with two telom-ere associated proteins Rap1p and Hfdlp (465; 700). Sir2p is an NAD+ dependent histone deacetylase that has been implicated as a major regulator of urinary tract infection, where nicotinamide limiting conditions in the bladder is associated with derepression of EPA1, EPA6, EPA 7 and increased adherence in wild-type cells (180; 390; 682).
The cell wall of C. albicans harbours other important adhesins that are not conserved in C. glabrata such as the /3-glucan linked hypha cell wall specific protein Hwp1p (656) which encodes an outer surface mannoprotein. The proline and glutamine rich amino ter-minus of this protein has structural similarity to substrates for mammalian transglutaminase (TGases) involved in crosslinking epithelial cell wall proteins, and it was postulated that this protein contributes to the proliferation of C. albicans in keratinized epithelial cells by interacting with these TGases responsible for maintaining an essential host epithelial bar-rier. To test this hypothesis, Staab et al generated a recombinant, truncated Hwp1p and showed a fourfold elevation in covalent incorporation of a radiolabelled TGase2 substrate relative to a negative control. Furthermore, homozygous null mutants of HWP1 showed significantly decreased (23% relative to wild type) stable adhesion to BECs and less viru-lence than parental or single-gene deleted strains in a hematogenously disseminated murine model (655), although this was later re-evaluated by the authors themselves (673) and an-other prominent group in the field (626). They have questioned the veracity of the survival phenotype data and have demonstrated that the expression of the reconstituted URA3 gene, used as the selection marker in the disruption strategy may actually account for the filamentation and survival phenotypes observed. Although not identified as virulence factor in the CGD repository, further literature review of adhesins reveals that the C. albicans cell wall has integrin-like molecules that bind several extracellular matrix ligands (fibronectin, collagen I and IV and laminin) as well as specific monoclonal antibodies raised against the a-chain of integrin proteins.
A putative integrin-like molecule Int 1p, which has low sequence identity to the human integrin protein at the level of the ligand binding domain, has been described in C. albi-cans. This transmembrane protein has an external EF-hand domain that is consistent with a cation binding function, and a cytoplasmic talin interacting domain that may interact with the actin cytoskeleton. Molecular studies of INT1 gene disruptions reveal impaired adhesion to epithelia, attenuated virulence in mice and inhibition of hypha formation (224) as wells as abrogation of super-antigen like effects on human T-lymphocytes (174). Het-erologous gene transformation studies of non adherent S. cerevisiae strains showed that INT1 confers increased adherence and induced filamentous growth. The adhesins of A. fumigatus are poorly understood. A. fumigatus conidia also express proteins that permit specific binding to proteins expressed on host mucosal layers (24)(78; 79; 90; 241). To date,

Chapter 1. Introduction 37
most studies of A. fumigates adhesion have mainly focused on the conidial outer cell wall, and its largely proteinaceous rodlet layer in particular (59). This thin layer of regularly arranged rodlets is thought to confer buoyancy and aid dispersion in air currents, and is also where hydrophobins, a family of hydrophobic proteins (528), are located. Another important process determining the virulence of C. albicans is its ability to grow in differ-
ent morphological states (e.g. blastospores, pseudohyphae and hyphae) (86). C. glabrata can form pseudohyphae, but not true hyphae in response to nitrogen starvation (150) and germ-like tubes in certain colony forms. Pseudohyphae are not true septate hyphae and are distinguished from "true hyphae" by their method of growth, relative frailty and lack of cytoplasmic connection between the cells. C. albicans forms true hyphae in response to environmental cues associated with host colonization (92; 667; 760).
For example, simultaneous changes in ambient temperature and pH, both of which are departures from the optimal growth conditions (471; 438), induce filamentation. Further-more, many of the culture media used to induce hyphal growth in vitro impose some degree of nitrogen starvation. For example, serum, a powerful inducer of germ-tube formation is composed mainly of proteins that are an inaccessible nutrient source until they are hy-drolysed. N-acetylglucosamine, a poor source of carbon and nitrogen will induce hyphae alone, which can be reversed by supplementing the media with amino acids. These in vitro environmental cues reflect starvation or depletion of easily accessible nutrients. Moreover, this polymorphic behaviour is shared with more distant fungal relatives whose mycelial networks penetrate woodland floors where they can influence the control of nitrogen and phosphorus supply, scavenging and sequestering nutrients from the soil and leaf litter that they decompose (213). It has been suggested that in the mammalian host, different mor-phological states confer advantages at specific stages of pathogenesis. For example hyphae can penetrate endothelial and epithelial cell layers during invasion whereas the unicellular yeast form can disseminate more easily through the bloodstream (253; 593). Studies in S.
cerevisiae and C. albicans have identified several genes in distinct pathways that influence filamentous growth. Like in the studies of adhesion, S. cerevisiae was used extensively in
the early characterization of C. albicans morphogenetic regulation and focused on the re-lationship between filamentation and mating, the coherence of which is dependent on the transcription activator Ste12p and the mitogen activated protein kinases that regulate its activation (136; 414; 397; 469; 416). C. glabrata orthologues belonging to this putative pathway have also been investigated (105; 103; 104). Strains disrupted in the orthologous C. glabrata STEW and STE1 1 loci were shown to be attenuated for virulence in an im-munocompromised murine model of candidiasis. Ste2Op and Stellp are both required for adaptation to hyperosmotic stress, and Ste2Op is required for a fully functional cell integrity pathway whereas Ste11p is largely dispensable (103; 104).

1.5 A brief review of virulence factors important for Candida and Aspergillus mediated pathogenesis 38
The C. albicans homologues of STE12, CPH1 (415; 87) complements both the mating and pseudohyphal defects of ste12 S. cerevisiae strains. Functional homologues of S. cere-visiae STE genes have now been identified in C. albicans, and null mutations at each of these loci cause analagous hyphal defects in the cphl null mutant, providing evidence that these genes lie on the same signalling pathway. Further epistasis analyses have placed CST20, HST7 and CPH1 genes in positions equivalent to their homologues in S. cerevisiae and sug-gest a conserved architecture between the two species. Deletion of a different transcriptional activator in the Ras-protein kinase A pathway, EFGJ, which belongs to a family of basic helix-loop-helix transcription factors, results in decreased filamentous growth and attentu-ated virulence (529). Epistasis analysis has suggested that EFG1 and CPH1 lie on distinct, parallel pathways that activate hyphal development. Culture conditions that induce hypha formation also induce the expression of SAP4 and SAP6 genes encoding secreted aspartyl proteases. There is evidence to suggest that co-ordinated regulatory pathways exist for hy-pha production and proteinase secretion, since deletion of a transcriptional activator of Sap expression, TEC1 abrogates SAP4 and SAP6 expression and prevents hypha formation (620). Moreover, deletion of EFG1 results in impaired hypha formation and a reduction in SAP4 and SAP6 expression. Strains lacking regulators of morphogenetic shift have been used in studies of transcription during the yeast-hyphal transition (491) and provide fur-ther evidence for a co-ordinated regulation between hypha formation and SAP expression. C. albicans can undergo phenotypic switching in response to environmental stress and is implicated in the control of the sexual cycle and in virulence traits such as Sap secretion and hyphal formation. This phenomenon was also reported in C. glabrata by Lachke and colleagues, who showed rapid cell morphology changes, associated with altered expression of the metallothionein gene MTII and the haemolytic HLP in response to copper stress (384; 385). Increased high frequency phenotypic switching, identified in invasive, infectious C. albicans strains (336) suggests a role for this mechanism in virulence. Dimorphism, was also detected amongst the different copper-stress induced phenotypes after 2-3 days of growth. The proportions of budding and pseudohyphal cells was similar amongst four core phenotypes, but one phenotype displayed a far greater proportion of pseudohyphae. Previous work indicated that C. glabrata did not form pseudohyphae, and this assumption was used as a discriminating characteristic for its identification (45), but there are later reports of pseudohyphal-like projections under nitrogen starvation (150).
Hydrolytic enzyme secretion is a feature of many pathogenic organisms including bac-teria (203; 204), protozoa (454) and pathogenic yeasts (514). Although extracellular pro-teinases of saprophytic fungi such as A. niger or Neurospora crassa are secreted primarily to provide nutrients for the cell, pathogenic fungi appear to have adapted this biochem-ical property to fulfill a number of specialized functions during the infective process in

Chapter 1. Introduction 39
addition to the simple role of digesting molecules for nutrient acquisition, for example in adhesion (237). The 10 Secreted aspartyl proteases (Saps) of C. albicans are preproenzymes processed in the secretory pathway (211), and play an important and complicated role in virulence (597)12 . Between them, the Sap enzymes have a broad substrate specificity where hydrolysis of peptide linkages can provide access to vital nitrogen sources, but can also deactivate important host molecules. These include mucosal layer protective proteins such as mucin and IgA and host defence molecules such as lactoferrin, lactoperoxidase, cathep-sin D complement (115; 4; 158). Saps are expressed in vivo during leukocyte engulfment and in murine models of disseminated candidiasis and gastrointestinal persistance (491). In a model of vaginal candidiasis, Schaller et al showed that Saps may contribute to the induction of an inflammatory host immune response; sap mutants, lacking either SAP1 or SAPS caused reduced tissue damage and had a significantly reduced potential to stimulate cytokine expression. In contrast, the vaginopathic and cytokine-inducing potential of sap4 and sap6 mutants was similar to that of the wild-type strain (612).
Saps are also importantly associated with other virulence processes. Sap production is correlated with adherence - strongly proteolytic C. albicans strains adhere more strongly to HBECs (237) and show a higher degree of tissue colonization. Inhibition of Saps with proteinase inhibitors such as pepstatin A inhibit germ tube dependent invagination of the vascular epithelium (362). The mechanism by which Saps confer adherence is not fully understood, but two different hypotheses exist; the first hypothesis is that Saps have ad-hesive properties themselves by acting as ligands for surface moieties of host cells. The second hypothesis, perhaps more consistent with their known functions is that Sap depen-dent hydrolysis could the modify the structure and increase adherence of host molecules (474). A correlation between virulence and proteolytic activity has been demonstrated for A. fumgatus clinical isolates and in experimental models of infection (365; 373; 585), however a specific role for proteases in pathogenesis is unclear. The major secreted pro-tease of A. fumigates is a serine alkaline protease (Alpp), belonging to the subtilisin family ((217; 326; 365; 475; 573)), however a metalloprotease (Mepp) and a pepsin family aspartic protease (Pepp) have also been identified in this species ((398; 574; 574)). A mutant strain lacking the Alpp gene showed abrogated elastin but not collagen hydrolysis, however this mutant strain was no less virulent than an isogenic Alpp secreting strain in an experimental model of aspergillosis ((475; 640; 680; 681)). Similarly, cloning and disruption of Mepp (325; 636) in a A* background results in total abrogation of extracellular proteolytic ac-tivity at neutral pH in vitro is still virulent in immunosuppressed mice in a manner similar to the wild-type cells (325). Although proteolytic activity is correlated with virulence it
12comprehensively reviewed in (491)

1.5 A brief review of virulence factors important for Candida and Aspergillus mediated pathogenesis 40
may not be causative relationship. Indeed, histopathological studies have shown that in the lung tissue of humans or experimental animals, the collagen and elastin matrix is tra-versed without any apparent proteolytic degradation associated with the invading fungus (170; 325)). Furthermore, a monocolonal antibody that inhibits A. fumigatus elastase did not reduce the virulence of the fungus in immunocompromised mice (219; 218). Secreted proteases are important for nutrient acquistion in Candida spp. and A. fumigatus, however acquistion of nutrients other than carbon and nitrogen is also of importance in the host, for example siderophore biosynthesis is required for A. fumigatus iron acquisition and virulence in a murine model of aspergillosis (618). Recently, the zafA zinc-responsive transcriptional activator was shown to regulate the expression of genes required for growth in zinc depleted media in vitro (478). Moreno et al., demonstrated that the A. fumigatus zafA gene com-plements the ZAP1 transcription factor gene of S. cerevisiae, is induced in zinc deficient medium and is repressed by the addition of zinc. Deletion of zafA impairs the germination and maximal growth of A. fumgatus in zinc-limiting media and attentuates the virulence phenotype (478).
Fungi that produce fruiting bodies such as A. fumigatus, demonstrate co-ordinate expres-sion of biologically active, toxic and immunosuppresive secondary metabolites and spore-related products during development (109). Indeed, A. fumigatus is equipped with a di-verse arsenal of toxins ranging from specific inhibitors of the 28S ribosome (344; 345; 389) through natural steroid antibiotics (632; 470; 99), haemolytic proteins (184; 221; 377), afla-toxins (537; 510) and small molecular weight toxins such as gliotoxin (GT). Gliotoxin is a mycotoxin that belongs to the group of epipolythiodioxopiperazines, which damages ep-ithelial cells (67), is cytotoxic to mouse macrophages (731) impedes the function of ciliary cells (18), and inhibits the activiation of NADPH oxidase of polymorphonuclear leucocytes (701). This inhibitory action on a major killing mechanism employed by major effectors of cell-mediated defence, macrophages and neutrophils (611; 236), means that gliotoxin has been implicated in virulence (486; 526). Although the role of gliotoxin in virulence remains unclear (74; 145; 379; 652; 670), there has been renewed interest in the role of secondary metabolites in Aspergillus infection since the discovery of a global transcriptional regulator of secondary metabolite biosynthesis in this organism, laeA (73). laeA deletion does not affect growth or sporulation in media in vitro, but it does result in reduced virulence in mice. This impairment in the ability to cause infection can be correlated with a loss of detectable gliotoxin as well as mis-regulation of gene expression within 13 gene clusters (539). Siderophore-mediated iron uptake and storage, also partially under laeA control (539) is indispensable for A. fumigatus virulence (618; 619) and adequate nutrient acquisi-tion within the host niche is linked to pathogenesis of invasive disease being necessary to support sufficient growth to promote infection (281; 375; 374).

Chapter 1. Introduction 41
Biofilm formation is one of the major factors contributing to the virulence of Candida species (621). Candida biofilms are surface-attached microbial communities surrounded my a matrix of extracellular polymers and are widespread amongst medical devices involved with the systemic circulation (567)13 , where a breached epithelial layer poses a major risk for tissue invasion and invasive candidiasis. Biofilms often display phenotypic features that are inconsistent with those of their planktonic constituents. Candida biofilms were 30 - 2000 times as resistant as planktonic cells to various anti-fungal agents, including amphotereicin B, fluconazole, itraconazole and ketoconazole (276). The first, critical, step of adhesion14 is initially mediated by hydrophobic and electrostatic interactions but is subsequently stabi-lized by the expression of specific adhesion molecules. Iraqui et al (316) used an insertional mutant library of C. glabrata to isolate mutant strains with an increased ability to form biofilms. They showed that silencing mutations of C. glabrata, which displayed Yak1p de-pendent transcriptional induction of EPA6 and EPA 7 also showed dramatically induced in vitro biofilm formation. Candida species differ in their ability to form biofilms, with C. glabrata generally showing poor growth and scant blastosphore distribution in comparison to the more proilific biofilm forming species C. albicans, C. krusei and C. dublininensis. The YWP1 gene of Candida albicans is predicted to be a 533 amino acid polypeptide with an N-terminal. A homozygous ywpl/ywpl null mutant, created by recombination of both YWP1 alleles in C. albicans exhibited increased adhesiveness and biofilm formation sug-gesting that it may promote dispersal of yeast forms of C. albicans (254). This protein is absent from C. gabrata. A. fumigatus also forms biofilms
The discovery of melanin production in C. neoformans was a major breakthrough in un-derstanding the basis of virulence in that species and in other pathogens such as Wangiella dermatitidis (177) and Mycobacterium leprae (738). Fungal melanins are produced by at least two different pathways (738; 234). In C. neoformans, melanin synthesis is catalyzed by lactase, a reaction that requires exogeous substrate in comparison to other fungi that pro-duce melanin from dihydroxynapthalene by the pentaketide pathway (738). When grown in the presence of certain o-diphenolic compounds, such as 3,4-dihydroxyphenylalanine (L-dopa), C. neoformans cells turn a dark colour, due to the presence of melanin (113). Trans-mission electron micrographs and electron spin resonance show that this pigment has a high electron density and is located in the cell wall. Classic genetic and gene disruption studies have demonstrated that melanin producing C. neoformans strains are more virulent than their respective albino mutants (383; 382; 586; 602). Further evidence of the importance of
13such as shunts, stents, implants, endotracheal tubes, pacemakers and venous catheters
14A biofilm typically develops over four sequential steps: i) adhesion to a usually flat and hydrophobic sub-strate, ii) discrete colony formation, iii) secretion of extracellular polymeric substances and iv) dissemination of progeny biofilms

1.5 A brief review of virulence factors important for Candida and Aspergillus mediated pathogenesis 42
melanization in vivo came from studies were a hyperpigmented C. neoformans strain was
associated with reduced production of the pro-inflammatory tumor necrosis factor a and also lymphoproliferation in infected mice (308); furthermore, binding peptides which react with melanized C. neoformans cell walls were used in immunohistochemical studies to show
that infected tissues contained melanized C. neoformans cell walls (506). Moreover, the iso-lation of anti-melanin antibodies following infection of mice with C. neoformans indicated an immune response to this pigment during infection (506). Direct biochemical evidence for in vivo biosynthesis of melanin was provided by the isolation of melanin particles from the tissues of infected rodents (594). Melanin confers several physiological advantages that partially contribute to C. neoformans virulence, most notably a resistance to oxidative stress (725; 546; 321; 322; 188)), amphotericin B (726) and microbiocidal peptides (179). Melanized cells are are also less susceptible to ingestion by macrophages and killing by alveolar macrophages (418).
A. fumigatus strains lacking polyketide synthase and scyatolone dehydratase (encoded
by ALB1 and ARP1 respectively (696; 698), and pigmentless wild-type strains (394) are less pathogenic than pigmented wild-type strain. Pigmented, or "white" conidia bind to complement more effectively than do wild-type cells and are also more permeable to anti-fungal drugs. A recent study in which 0 ace2 mutants were shown to produce an orange pigment (185). The relevance of these findings in virulence is limited however, since the wild-type strains did not produce the pigment and the increased null strain virulence phenotype could be a result of the morphoogical changes associated with the disruption of a pleiotropic gene such as A CE2; furthermore, although "white" conidia are more virulent, all observed wild-type strains have green conidia with hyaline hyphae (394). C. albicans also produced
melanin in vitro and during infection of immunocompetent mice (482) although there are no studies to date on its mechanistic role in stress protection or virulence. There are also no reports of melanin production by C. glabrata, although studies of phenotypic switching
in C. glabrata (385; 384) identify core phenotypes that are light brown, dark brown or very dark brown this is assumed to be due to the graded levels of cupric and cuprous copper
ions in the cells. C. glabrata does however produce indole alkaloid pigments (pityriacitrin, malasseziaindole, pityrianhydride and) when tryptophan is the sole source of nitrogen in the culture medium (450). Pityriacitrin, first isolated in Paracoccis spp. are protective
against UV radiation, and pityarubins A, B and C are all capable of inhibiting the human neutrophil respiratory burst. Although the role of these pigments in C. glabrata remains
undetermined, Mayser et al., speculate that these pigments may be protective in vivo.

Chapter 1. Introduction 43
1.6 Sensing and adapting to environmental stress responses
Sessile microorganisms must be able to adapt to a wide range of physicochemical conditions. Such innate robustness requires homeostatic mechanisms (538; 368; 554; 50; 190; 315; 480; 261; 89; 292) that sense these fluctuations and direct adaptive processes which either mod-ulate the signal or obviate the effects of the perturbation. Understanding the molecular and genetic basis of these homeostatic mechanisms is of fundamental importance in biology. With a mechanistic insight, we can try to fix them when they are broken, or conversely try to break them when we want to sabotage this cellular robustness. Accordingly environmen-tal stress research transcends the field of mycology (627; 555; 260). However, the model fungal research communities have powerful molecular toolboxes at their disposal, and sig-nificant advances have been made in our understanding of environmental stress adaptation in S. cerevisiae and Schizosaccharomyces porn be. In particular, comprehensive genome-wide transcription surveys (230; 118) have significantly advanced our understanding of the core environmental stress response in S. cerevisiae, and specific stress responses pertinent to the host niche of a fungal pathogen. In the context of fungal virulence studies we are primarily interested in the specific environmental stresses of the host niche, and how fungi adapt to them. Although there is some debate over the definition of an opportunistic fungal viru-lence factor (511; 278), one of the uncontentious molecular Koch's postulates (195) is the requirement for virulence determinant expression within the host. So far, genome-wide transcription profiling in the host has been inaccessible due to technical limitations in re-covering sufficient fungal material for microarray experiments, however gene expression has been monitored within the host on a gene-family basis (22; 376). While these technical limitations remain, host environmental stress has been approximated in vitro (e.g. oxida-tive stress, thermotolerance, exposure to human polymorphonuclear leukocytes and human whole blood (191; 192; 210)), with the majority of genome-wide transcription profiling pe-formed in C. albicans and A. fumigatus. At the start of this thesis, the environmental stress
response of C. glabrata had not been characterized either by transcriptome surveys or in-vestigations of signal transduction pathways. A non-exhaustive discussion of environmental important environmental stress response follows.
1.6.1 Oxidative stress response
Several C. albicans studies have focused on important effector (611; 236; 366) cells of host immunity (210; 596) and the fungicidal molecules they release (192; 191; 305; 718). Can-
dida spp. and A. fumigatus are predominantly engulfed by macrophages and neutrophils, whose reactive oxygen species (ROS) burst is a fungicidal mechanism (421; 466). Envi-ronmental stress response studies in S. cerevisiae using hydrogen peroxide, menadione and

1.6 Sensing and adapting to environmental stress responses 44
diamide as sources of exogenous reactive oxygen species (230; 118) revealed co-ordinated expression of genes encoding oxidative stress protection proteins such as the thioredoxin TRX1 and glutaredoxin TTRI and the glutathione biosynthetic enzyme. These are largely conserved in C. albicans cells exposed to hydrogen peroxide (191) and after exposure to ploymorphonuclear leukocytes. Comparison of transcript profiles generated after exposure of C. albicans cells to 11202 and polymorponuclear leukocytes reveals conserved upregu-lation of transcripts encoding oxidative stress protection proteins: CTA1, TTRI, GLR1, TRX1, GTT, BMR, YHB2. Oxidative stress protection genes such as glutaredoxin GRX1 (125), catalase CTA1 (758), and cell surface superoxide dismutases SOD! and SOD5 (216) are also required for virulence in animal models of candidiasis.
1.6.2 Ambient pH adaptation
In addition to the specific stress imposed by the host immune system, the fungus must adapt to physicochemical constraints of the host such as the ambient pH, temperature and energy source availability. Candida spp. and A. fumigatus can adapt to a wide ambient pH range, an attribute that in part enables these fungi to survive in a diversity of habitats, ranging from compost and fruit surfaces where they are saprophytes to a wide range of anatomical sites in immunocompetent host where they are commensals. Significant insight into the molecular basis of ambient pH adaptation has come from studies of the signal transduction components and PacC transcription factor of A. nidulans (reviewed in (536; 33)), and orthologous Rim101 pathway counterparts in C. albicans (reviewed in (157)) and S. cerevisiae. Ambient pH adaptation is required for virulence in A. nidulans (66) and C. albicans but the role of Rim101/pacC in C. glabrata or A. fumigatus remains undetermined.
1.6.3 Hyperosmotic stress adaptation
Hyperosmotic stress adaptation pathways (89; 292) are particularly well characterized in S. cerevisiae and S. pombe. Although the assertion that hyperosmotic stress is imposed on pathogenic fungi in the host environment is not entirely convincing, a recent observation that Hog1p MAPK function has diverged from related model organisms and is required for virulence in C. albicans (14; 126; 192) and C. neoformans (42) makes this pathway especially interesting in the broader context of fungal virulence. The pathway architecture was deduced in S. cerevisiae with a series of genetic screens and consists of at least two signalling branches, defined by the putative osmo-sensors at the top of each cascade: the two component like Slnl phosphorelay branch (553; 575), and the mucin-like transmembrane receptor Shol branch (551; 152). A MAPKKK architecture lying at the core of each (551; 552), relays the upstream signal to the MAPKK Pbs2p (89) and ultimately the MAPK,

Chapter 1. Introduction 45
Hog1p (89). Osmoadaptation proceeds in part (737) by the Nmd5p (201) mediated nuclear translocation of phosphorylated Hog1p mediating the induction of glycerol biosynthetic enzymes (11; 505) and the closure of aquaglyceroporins (678; 428), resulting in the net accumulation of glycerol and decreased water potential of the cell. The role of orthlogous A. fumigatus osmosensing proteins in virulence is largely undetermined, however two genes encoding putative osmoregulatory proteins (mpkA and SHOT ) were shown to have severe growth defects in osmotic stress conditions but were not required for virulence (709; 710; 429).
1.6.4 Thermotolerance
Thermotolerance plays an important role in virulence gene expression in pathogenic bacte-ria, including Escherichia coli, Salmonella spp., Shigella spp., and Yersinia spp., where the heat shock response is an important determinant of pathogenesis, and this has long been suggested to be a key determinant of virulence in A. fumigatus, which unlike its mesophilic relatives can grow in temperatures ranging from 20°C - 60°C.To investigate the metabolic adaptation of A. fumigatus at such elevated temperatures, Nierman et al., (499) performed transcript profiling of A. fumigatus cultures at 30°C (tropical soil temperature), 37°C (mam-malian body temperature) and 48°C (compost temperature) and identified a cluster of 135 genes that were specifically induced at mammalian body temperature, including genes en-coding putative heat shock proteins and oxidative stress protection proteins. Heat shock proteins are thought to act as chaperones to maintain correct protein folding, however their role in virulence may be more complicated (521; 520), particularly in C. albicans where heat shock proteins are induced during the hyphal transition
1.6.5 The general stress response of S. cerevisiae
The concept of a co-ordinated, general stress response or environmental stress response (ESR) emerged after the effects of normally lethal doses of one stress were sublethal after pre-treatment with a sub-lethal dose of a different stress (467; 468). Furthermore, genes that were specifically thought to protect the cell against heat shock were shown to be induced under a variety of independent stressors. Analysis of the upstream promoter region of these genes identified a conserved sequence element that became known as the stress response element (STRE), which was later shown to be the consensus sequence binding site for the zinc-finger transcription factors, Msn2p and Msn4p (615; 615; 81). The ESR was originally described using microarray expression analysis in S. cerevisiae by the independent groups Gasch et al(230) and Causton et a/(118). In combination these groups investigated the transcriptional responses to a range of environmental cues including heat shock (230), ex-

1.6 Sensing and adapting to environmental stress responses 46
tremes of pH (118), oxidative stress (hydrogen peroxide (118) or menadione induced (230)), hyper-osmotic stress (NaC1 (118) or sorbitol (230; 118) induced), hypo-osmotic shock (230), amino acid starvation (230), nitrogen depletion (230), and sub-optimal carbon sources (230). Gasch et al identified a set of approximately 900 genes, whose transcript abundance was elevated or decreased across almost all of the environmental stress conditions. The set of stress decreased transcripts (600) and the set of stress-increased transcripts ('300), were remarkably similar in their average fold-change and kinetics, being almost equal and opposite across the time-course; transcripts that are co-expressed often behave with sim-ilar kinetics and this was taken as evidence that these genes were in part controlled by a common transcriptional regulation process. A remarkable feature of the ESR data is its large proportion of un-annotated transcripts (approximately 50% in the repressed set and approximately 45% in the induced set). Although common transcriptional patterns may be functionally related (716), a significant portion of the ESR remains poorly characterized. Induced ESR transcript functions spanned diverse processes, such as oxidative stress protec-tion, autophagy, protein degradation, carbohydrate metabolism and respiration; repressed ESR transcripts were predominantly involved in protein synthesis.
Since the identification of the general stress response, the model in which two tran-scription factors Msn2p and Msn2p are global regulators (615; 446) of the co-ordinated response has been refined. Although the majority (251, 88%) (230) of induced ESR genes contain the consensus binding site (STRE element CCCCT) for the "general" transcription factors Msn2p and Msn4p, they are not necessary for expression of all genes in the ESR. Indeed, several transcription factors regulate specific stresses within the broader context of the ESR. For example, Hot1p and Msnlp are required for the expression of hyperos-motic stress genes of the ESR (581). Further regulatory complexity was discovered by condition specific behaviour of transcription factor null strains in S. cerevesiae. A study (581) of msnl, msn2, msn4 and hotl transcription factor knockout strains subjected to osmotic stress and heat shock revealed that the gene encoding glycerol-3-phosphate dehy-drogenase GPD1 is predominantly dependent on Hot1p in response to osmotic shock, but was dependent on Msn2p and Msn4p in response to heat shock (581). Similar observa-tions supporting the notion of stress specific transcription factor behaviour come from heat shock studies. The heat shock proteins Hsp26p and Hsp104p are induced after a heatshock in an Hsflp dependent manner, however their induction in response to carbon starvation, osmotic shock and oxidative stress is dependent on Msn2p and Msn4p (19; 695). To fully understand the complex nature of these regulons, comprehensive time-course transcription profiling and phenotypic analysis of transcription factor null mutants and double mutants will be necessary. There is also evidence for epigenetic control of gene expression in re-sponse to stress ((641; 732; 756; 226; 579; 337)), however the relationship between these

Chapter 1. Introduction 47
regulatory factors, stress specific transcription factors and other pathways that regulate the ESR (e.g. the TOR (58) and PKA (227; 252; 638) pathways) is unclear. It has been proposed that this co-ordinated transcription programme is part of a conserved protective response to protect critical features of cellular physiology in budding yeast (229). How-ever, initial transcriptional studies in C. albicans did not reveal a general stress response (191) (in oxidative, hyperosmotic stress and a 23 - 27°C temperature shift) nor was any cross protection observed when cells pre-treated with a mild heat shock were subsequently exposed to oxidative stress. In a contradictory study, Smith et al., (639) did demonstrate HOG/ -dependent cross-protection to 0.3M NaC1 after pre-treatment with a sublethal dose of H202. A subsequent systematic comparison of general stress response data sets of C. albicans, S. pombe and S. cerevisiae (192) did reveal a general stress response with almost 50-fold fewer genes than in S. cerevisiae. While there is evidence for a conserved ESR in the model fungi S. cervisiae and S.pombe (229), the evidence for such a comprehensive ESR in C. albicans is unconvincing. At the start of this study, the general stress response of C. glabrata had not been investigated, and the relevance of the ESR in opportunistic fungal pathogens is still unclear.
In summary, gene deletion and transcription profiling methods have been used to iden-tify the critical components for growth, and to identify a set of genes that are adaptively regulated in response to stress conditions pertinent to the host. Eukaryotic pathogens such as the opportunistic fungi co-ordinate many processes in order to survive in the host. Fo-cusing on single gene deletion strategies alone may not uncover the complexity of these processes. For example, the virulence of A. fumigatus is increasingly regarded as a com-plex multi-factorial attribute (36), underpinned by different networks of genes that have evolved to support the organism in a variety of environments. A critical question remains: why is C. glabrata a pathogen when its close phylogenetic neighbour S. cerevisiae is rarely so? Similarly, what makes A. fumigatus such a prolific pathogen in relation to its close relatives? By understanding the co-ordination of these complex processes, rather than the role of individual genes, we may be able to better understand the molecular basis for their survival in an immunocompromised host. In order to do so, genomic data may need to be interpreted in the context of pathways and regulatory processes, a procedure that requires methods for system-wide data integration and analysis.
1.7 Identifying therapeutic targets: potential for a systems approach to fungal virulence studies
A systems approach to studying fungal virulence may overcome these limitations in the future. Although it is not a novel concept (635; 735), the confluence of post-genomic data

1.7 Identifying therapeutic targets: potential for a systems approach to fungal virulence studies 48
(424), advances in the understanding of complex networks (47; 48; 733) and a burgeoning research community (293) make it a tractable proposition in biology. The motivation of systems biology is to develop mathematical and computational techniques to predict the behaviour of a complex system (359; 648; 313; 272) which would otherwise be limited by a reductionist approach and has been the focus of much speculation and skepticism with regard to the potential value in understanding multigenic human diseases. Although the field is immature, there are signs that this approach might be of real value in the fight against complex multigenic diseases (617; 130; 610). Despite being regarded as the antithe-sis of the reductionist approach, systems biology is necessarily reductionist at this time, currently focusing on either subsets of complex pathways within a cell (755; 151; 128), or a specific organ (501) within an organ system. The distinction is not that systems biology is anti-reductionist, rather that it is integrative and simply less reductionist. By integrating information about the parts of the system, how they interact, and the kinetics of these interactions it is possible to identify emergent system-level properties - behaviours that are not explicable in terms of the components of the system alone. For example, swarming intelligence (530) is an emergent property of systems of unintelligent agents that collec-tively demonstrate intelligence (740). Individual ants are behaviorally simple insects with limited memory; collectively, however, they manage to solve complicated problems (114) such as searching for and retrieving food and the feeding of larvae by worker ants. When mycelial fungi sense carbon and nitrogen depletion, they develop hyphal projections which radiate from their established substrate towards richer nutrient sinks, eventually forming an aggregated mycelial network composed of other individual fungal cells. These mycelia aggregate behind the growing margin and fuse to form high capacity nutrient transport channels (570; 102). Although each cell within the aggregate each has individual mech-anisms for nutrient acquisition, recent studies in Phanerochaete velutina revealed highly specialized components of the mycelial aggregate that preferentially transport nitrogen to carbon rich areas of the network in an oscillatory fashion (213).
Emergent properties have also been discovered in cell signalling pathways of the cell. Bhalla and Iyenger (65) reconstructed in silico biochemical pathways using experimentally determined parameters and connected these pathways to create more complex networks composed of either two (epidermal growth factor - EGF - pathways) or four (glutamate synaptic transmission in hippocampal CAl neurons) of these discrete pathways, and demon-strated that the interconnected network possessed several properties distinct from the indi-vidual discrete pathways. For example, when two epidermal growth factor pathways (EGF mediated stimulation of MAPK 1 and MAPK2, EGF mediated activation of phospholipase C-'y) were connected with a feedback loop, the resulting system was bi-stable. Such a bi-stable system showed thresholding behaviour whereby only certain doses and exposure times

Chapter 1. Introduction 49
elicited sustained activation of MAPK or phospholipase C--y. In addition to this switching behaviour, bi-stable systems also demonstrated a memory property (759), whereby path-way activation can occur after the stimulus is removed. This is of particular importance to the EGF pathway, sustained activation of which is a hypothetical therapeutic strategy for cholangiocarcinoma (699; 765). bi-stable switches also direct the population level synthesis of antibiotics in Streptomyces coelicolor (the bytryolcatone regulon (458)), haematopoietic stem cell fate (393) and have a regulatory role in the cell cycle (547). Although Bhalla and Iyenger connected pathways into large networks composed of up to approximately 15 reaction members, it should be noted that the bistability can also occur in a simple reaction scheme composed of two stochastically expressed genes (30; 533).
The two best examples of integrated dynamic models in yeast to date have focused on hyperosmotic stress adaptation (361; 569) and cell cycle control (128). The Klipp-Hohmann integrative model of the response of yeast to osmotic shock (361) was constructed and pa-rameterized to fit experimental measurements of Hog1p activation, glycerol accumulation and glycerol biosynthetic enzyme activation in S. cerevisiae. Simulations from their pa-rameterized model closely matched experimental data and also provided new insight into the regulatory mechanisms of pathway activation. For example, although the Slnlp-Ypdlp phosphorelay is similar to the two-component signalling cascades of bacteria, the incorpo-ration of a second phosphorelay step provides a switch like behaviour that requires supra-threshold input to cause an effect, and a delayed activity analagous to the findings of Balla and Iyenger in their EGF pathway simulation (65). Another important prediction of the model was that sufficient glycerol accumulation can be achieved with basal glycerol pro-duction and rapid Fps1p closure (the aquaglyceroporin through which glycerol leaves the cell), which was supported by later evidence from Westfall et al., who demonstrated S. cere-visiae cells could still proliferate if Hog1p was prevented from entering the nucleus where it directs the transcription of glycerol biosynthetic enzymes (737). They also demonstrated that phosphatases do not need to be induced above a threshold in order to elicit feedback, moreover subsequent pathway activation would not be possible if phosphatase levels are induced greater than two-fold in their model. Chen et al., (128) used literature searches to create a wiring diagram of components involved in the cell cycle and represented the mech-anism as a series of algebraic and nonlinear differential equations. Using experimentally determined and fitted parameters, their model accurately predicts the phenotypic conse-quences of a panel of approximately 100 existing null mutant strains, and more importantly predicted the phenotypic effects of novel mutants. Another key feature of using integrated dynamical models is to identify inconsistencies with simulations and the consensus opinion of mechanisms. This interplay between simulation and experimentation serves to either improve parameterization or description of the model, or more fundamentally to formulate

1.7 Identifying therapeutic targets: potential for a systems approach to fungal virulence studies 50
hypotheses that form the basis of future experimentation. Although dynamic models are most desirable, the required physiological parameters are often immeasurable or unmea-sured (262). Consequently, contrary to the strict definitions of systems biology posited by some eminent researchers, other modelling techniques that may ordinarily be described as integrative genomics have been incorporated into the systems biology lexicon.
One such research field is that of network theory and its specific application to molec-ular interaction networks of model organisms (reviewed in (665). Network theory is a relatively new branch of statistical physics that extended the pioneering work of social scientists (463) and ecologists, and has advanced our understanding of how complex sys-tems are organized, how they may evolve, and how their topology is linked to system behaviour. The recent interest in network theory was sparked by a seminal paper published by Barabasi and Albert (47; 48) who overturned the tacit assumption that nodes in real networks were randomly wired together and best described by the random graphs of Eras and Renyi (193). By studying diverse "real" network data obtained for power grids, movie-collaborations and the world wide web they observed that the degree distribution of these graphs followed a power law and retained the small world property (733). These so-called scale-free networks characteristically have a majority of poorly connected nodes (i.e much lower than the average number of connections in the network), and a minority of highly connected (i.e. much greater than average number of connections in the network) nodes that are often termed hubs. Ensuing technological advances furnished the S. cerevisiae (705; 318; 232; 233; 693; 692), Caenorhabditis elegans (407) and Drosophila melanogaster (243) research communities with large-scale interaction data, whose topological character-istics have also been investigated and shown to be scale-free networks (328). In addition, metabolic networks whose nodes may represent metabolites, reactions or enzymes and whose edges connecting them represent mass flow or catalytic regulation are also scale-free (329). Scale free networks have particular topological properties that determine aspects of system function. For example, topology dependent robustness of scale-free networks has been stud-ied using vertex attack methods. In the context of scale-free networks, robustness refers to the persistence of network structural properties after sequential, random removal of nodes. Albert et al., (10) demonstrated that networks display an unexpected degree of robustness to random vertex removal, however, scale-free networks are not robust if high degree nodes are removed. Although it is clear that dynamical behaviour also governs system robustness, these in silico attack tolerance simulations have also been interpreted in the light of gene knockout data in S. cerevisiae. Genome-wide surveys of essentiality in S. cerevisiae indicate that approximately 10% of known ORFs with less than 5 connections are essential, whereas over 60% of proteins having 15 or more links are essential, i.e. targeted attack of hubs has a catastrophic effect on network robustness (328).

Chapter 1. Introduction 51
In addition to power law degree distributions, scale free networks have high average clus-tering coefficients (9), a measure of local connectedness in the network. The high average clustering coefficients of metabolic and protein-protein interactions reveal that they consist of subgraphs more highly connected to each other than the network as a whole, and that these subgraphs are connected to each other by hubs of high degree (764; 49). These bio-logical "modules" consist of highly interconnected and functionally related proteins (522), a property that has led some researchers to propose the function of uncharacterized pro-teins by their co-location in such modules (405; 625). Interaction networks have also been used as a scaffold for the integration of genome-wide data sets using visualization tools such as cytoscape (624; 137) and statistical methods to identify subgraphs of the network enriched with differentially expressed genes (314) and network motifs with putative regu-latory function (629). Transcriptome and deletome data have been integrated to identify cellular pathways activated in response to alkaline (31) and transition metal stresses in S. cerevisiae (333; 274). Haugen et al., (274) mapped arsenic III specific transcription profil-ing and gene deletion data to metabolic and protein interaction networks. This approach identified significant subgraphs of the protein network that were enriched with differentially expressed genes, and proteins central to that network with known regulatory function were proposed to be regulators of the arsenic III stress response (e.g. Msn2p, Msn4p, Yap1p, Hsflp). Furthermore, significant metabolic subnetworks were also identified by integrating gene deletion data with the global yeast metabolic network. In doing so they showed that genes conferring sensitivity to arsenic III belong to the shikimate and serine/threonine/ glutamate pathways lie upstream of the genes that are transcriptionally regulated. The development of the synthetic genetic array technology in the Boone and Andrews laborato-ries (693; 692) enabled a systematic global screen for synthetic global genetic interactions, which has recently been extended to approximately 75% of the genome in the DryGIN project (142). Using this technology they identified complex regulatory connections that only emerge in this unbiased genome-scale network approach. For example they identified a select subset of genes involved in cell polarity functions that appear to be regulated by the elongator and tRNA modification pathway; also they predict that the similar regula-tion of microtubule and cytoskeletal organization genes underlies a human disorder, familial dysautomia. The positive correlation between genetic interaction degree and the chemical genetic degree published by Giaever et al., (287) also revealed that hubs in the chemical genetic network are predictive of the hubs of the genetic interaction network. These data sets were integrated to propose a mechanism of action for an uncharacterized compound called erodoxin. Network analysis showed that the drug sensitivity profile of this drug most resembled the genetic interaction profile of ERO1 which encodes an enzyme participating in disulfide bond formation into its oxidized state.

1.7 Identifying therapeutic targets: potential for a systems approach to fungal virulence studies 52
The use of S. cerevisiae as a model organism has been described above, and has princi-pally focused on orthologous expression profiles and deletion phenotypes to elucidate diverse cellular processes. As S. cerevisiae enters an integrative systems based era, it is important to assess the degree to which a model organism can be used as a framework for a systems approach in a related organism that lacks fully described interaction data and functional genomics screens. In this thesis, parallel gene deletion assays and transcriptional profil-ing were conducted and integrated with a protein-protein interaction network inferred by othology to S. cerevisiae. This was performed in an attempt to identify significant subnet-works and to formulate hypotheses on the nature of environmental stress adaptation in C. glabrata. Specific aims are listed below.

Chapter 1. Introduction 53
1.8 Specific aims addressed in this thesis
1. Recent studies in the non-pathogenic models yeasts and C. albicans have provided evidence that signalling pathways, transcription factors and transcriptional responses to virulence related environment cues have significantly diverged in C. albicans. The environmental stress response of a yeast with higher innate drug resistance and an increasing cause of invasive candidiasis, C. glabrata is largely unexplored at the tran-scriptional level. Furthermore, to date, no studies have been conducted to investigate the contribution of stress signalling pathway components to these transcriptional re-sponses. In Chapter 3 I aimed to:
(a) Identify whether HOG1 is required for growth in hyperosmotic stress conditions by constructing a hogl strain and assessing its growth in hyperosmotic stress medium relative to rich culture medium.
(b) Assess how Hog1p functions under hyperosmotic stress conditions with respect to its phosphorylation kinetics and determine if any observed phosphorylation depends on upstream MAPKKK architecture conserved with related fungi.
(c) Further assess any divergence in fungal hyperosmotic stress adaptation by assay-ing the transcript profile of wild-type C. glabrata cells subjected to hyperosmotic stress and comparing it with published data for S. cerevisiae and C. albicans.
(d) Assess how divergent HOG1-dependent transcriptional responses are by further comparing the transcript profile of hogl cells subjected to hyperosmotic stress, with equivalent data in S. cerevisiae and C. albicans.
(e) Identify a candidate regulatory network by mapping regulated genes to a tran-scriptional regulatory network inferred from S. cerevisiae and use this to identify candidate regulators of hyperosmotic stress induced transcription.
2. Investigating the transcriptional responses to environmental cues in vitro is an ap-proximation of the host-niche, and is useful in Candida spp. where fungal material is virtually impossible to recover post-infection. In contrast, an immunocompromised mouse model of invasive aspergillosis does permit the re-capture of germlings in the early stages of infection. In Chapter 4, I report on joint work with Natalie Federova and Elaine Bignell. In this section we aimed to:
(a) Optimize the time-course for infection of mice immunocompetent mice with A. fumigates Af293 such that fungal germlings are simultaneously easy to recover and also in growth phase of relevance to infection.

1.8 Specific aims addressed in this thesis 54
(b) Employ bioinformatic and molecular strategies for global profiling of A. fumigatus gene expression in germlings rescued directly from the murine lung, a tool which will empower the analysis of virulence in this pathogen
(c) Identify fungal attributes preferentially employed during adaptation to the host niche, and thus contributing to the virulence of A. fumigatus
(d) Compare the transcriptomes of developmentally matched A. fumigatus isolates following laboratory culture or initiation of infection in the neutropenic murine lung.
(e) Compare host-niche regulated genes with data generated in vitro to identify environmental cues relevant to the host niche
(f) Assess the lineage specificity of genes preferentially expressed in the host-niche, which might provide evolutionary clues to the unique success of A. fumigatus as a pathogen in comparison to closely related Aspergilli.
3. The groundbreaking sequencing and annotation of the S. cerevisiae genome some thir-teen years ago (246) facilitated a new era of yeast research in which reverse genetics approaches became the catalyst for a burgeoning supply of genome-wide phenotypic data. Although the S. cerevisiae knockout libraries have been informative in elucidat-ing the molecular nature of conserved processes, a remaining challenge is to establish a comprehensive, genome-wide C. glabrata deletion mutants akin to that developed by the Saccharomyces genome deletion project. In chapter 5 I aimed to:
(a) Report on initial progress in creating a prototypic targeted disruption library in C. glabrata
(b) Provide a quantitative measure of C. glabrata fitness when exposed to fluconazole with a view to comparing equivalent null strains (lacking a single orthologous gene) in S. cerevisiae

55
Chapter 2
Materials and Methods
2.1 Experimental Methods
All chemicals used in this study were purchased from Sigma (UK) or Fisher Scientific (UK), unless otherwise stated. All culture media constituents were purchased from Oxoid Ltd (Hampshire, UK), BDH (Poole, UK) or Formedium (Norwich, UK). Unless otherwise
stated, all buffers and solutions were prepared according to standard recipes (603).
2.1.1 Fungal strains and culture media
Saccharomyces cerevisiae and Candida glabrata were routinely cultured in synthetic com-
plete (SC, Supplementary Table B.1) or yeast extract peptone dextrose (YEPD, Supple-mentary Table B.2) broths and equivalent solid media. Solid or liquid S. cerevisiae and C. glabrata cultures were incubated in either a static or an orbital shaking (180 rpm) incubator
(New Brunswick Scientific, USA) at 30°C or 37°C respectively.
All S. cerevisiae strains used in this study come from the Yeast Knockout (YKO) Mata
collection (749) (Open Bio Systems, USA). These strains, derived from BY4741, were cre-ated by homologous recombination with a kanamycin (KanMX) antibiotic resistance cas-sette. Accordingly, cultures of these strains were supplemented with geneticin G418 (200
µg/mL) to maintain selection of the correct genetic background. A. fumigates were routinely cultured at 37°C in supplemented Aspergillus Minimal
Media (AMM) (NaNO3, 6.0 g/L; KC1, 0.52g/L; KH2PO4, 1.52 g/L; MgSO4 . 7H20, 0.52
g/L; and 1 mL of trace element solution per litre) or Aspergillus Complete Media (ACM) (2.5% glucose (w/v), 0.5% yeast extract (w/v), 1.5% agar (w/v)) according to Pontecorvo
et al. (548) containing 1% (w/v) glucose as carbon source and 5 mM ammonium ± tartrate as nitrogen source. Non-standard Aspergillus culture media are explained in the relevant
results section (Chapter 4).

2.1 Experimental Methods 56
2.1.2 Bacterial strains and culture media
XL-10 Escherichia coli competent cells were cultured in Luria Bertani (LB) medium, 2% (w/v) GibcoBRL LB broth base (Life Technologies, UK). Liquid cultures were routinely incubated at 37°C, 150 rpm. Solid medium was produced by adding 2% (w/v) agar prior to autoclaving. When required, ampicillin was added to a final concentration of 100 pg/mL) after autoclaving and once the media had cooled to below 55°C.
2.1.3 Assays to identify altered growth phenotypes of mutant C. glabrata and S. cerevisiae strains
2.1.3.1 Solid agar assays of C. glabrata and S.cerevisiae strain growth
YKO library strains, individually stored as glycerol stocks in 96 well microtitre plates, were thawed close to a Bunsen flame and agitated in situ on a bench-top orbital plate-shaker (10 min, 10 rpm).
Stocks were aspirated from library plates using a multi-channel pipette and dispensed into new, sterile 96 well microtitre plates containing YEPD (final density 1 x 106 cells / ml) and incubated until the cells reached stationary phase (16 hours, 30°C, 180 rpm). In parallel, an equivalent cell density 5 mL culture of BY4741, the library parental strain, was created in a separate Sterilin universal tube.
Cells were harvested from sub-cultured microtitre plates with a centrifuge equipped with plate-adpators (Wolf Labs, UK, 3000 rpm, 5 min). The supernatant was discarded by aspiration with a multi-channel pipette, leaving cell pellets of YKO library strains, which were then washed twice with 100 pl sterile ddH2O using the harvesting steps described above.
Cell density was estimated for 5-7 wells using a microscope and a haemocytometer (Neubauer, USA). The average cell density was taken for these wells and used to calculate a global dilution factor for daughter plates containing wells seeded at a density of 106 cells / mL. The parental strain, at the same cell density, was added to an empty well on this plate.
Cells were then transferred from these working stock plates to pre-poured YEPD or SC solid agar (+ environmental stress) in 50 mL Petri dishes using a 96-pronged multi-replicator tool.
Inoculated plates were covered with lids and allowed to dry for one hour on the bench before inversion and incubation in a static incubator (30°C, 48-72 hours).
Colony growth of null mutant strains was qualitatively scored (either no growth or sen-sitive) in relation to the co-inoculated parental strain in YEPD and YEPD ± environmental stress.

Chapter 2. Materials and Methods 57
A conditional growth phenotype was defined for strains showing growth equivalent to the parental strain in YEPD but impaired growth in agar supplemented with an environmental stress on which the parental strain growth was unaffected.
2.1.3.2 Miniaturized optical density assays of mutant Candida glabrata strain growth
Glycerol stocks of each library strain (either the C. glabrata transcription factor knockout library CGTFKOLib or the equivalent null strains in the YKO library), collectively main-tained in two 96-well microtitre plates at -80°C, were sampled (5 itl) using a multi-channel pipette and dispensed into corresponding sterile, round-bottomed 96-well plate containing 150 it of YEPD broth. Library strains were sub-cultured to stationary phase for 18 hours in an orbital shaker (30°C, 180 RPM) before commencing the growth curve assay.
Sub-cultured cells were harvested (3000rpm, 5 mins) in a centrifuge rotor equipped with plate adaptors (Wolf Laboratories, UK) and washed three times with sterile, distilled water. Washed cells from each library plate were transferred (final OD600=0.1) into microtitre plates containing YEPD ± environmental stress. Plates were sealed with a breathable membrane and incubated for 12 hours. Optical density (0D600) measurements were bi-hourly recorded on a Multiskan Ascent absorbance plate-reader (Thermo Scientific, U.S.A).
Time course optical density (0D600) data, used as a measure of cell population dynamics, were fitted to a three parameter (slope, midpoint and upper asymptote) logistic function of growth using non-linear regression (SSLogis function) in the R statistical computing framework (562). The logistic function of growth adopted in the analysis is shown in equation 2.1:
f (x, (b, d, e)) = (2.1) 1 + exp{ b (log(x) — log(e))}
In this model, logistic growth is a function of time x, the maximum cell density d (upper asymtote), the midpoint of the growth e and the slope around this midpoint b. Parameters d, e and b are quantitative phenotypic traits of each strain and are estimated from samples of cell density at x = 0 — 11.5 hours.
Strains showing significant, environmental stress specific growth defects were identified in null strains either by a) a complete lack of logistic growth in the presence of environmen-tal stress but not YEPD (hypersensitive) or b) significantly greater midpoint (p < 0.05) estimates than wild-type strains in fluconazole but not YEPD (sensitive).
2.1.4 Oligonucleotide primer synthesis
All primers used in this study were purchased from Sigma-Genosys (Dorset, UK) and are listed in the relevant Appendices. See relevant chapters featuring molecular work.

2.1 Experimental Methods 58
2.1.5 Polymerase Chain Reaction (PCR)
Colony PCR cycling and reaction parameters, used for verifying the identity of yeast knock-out (YKO) strains, were adapted from published protocols (601). Genomic DNA template was obtained by adding a prior high-temperature lysis step (100°C, 15 minutes) to single colony picks of the knockout strain in the presence of oligonucleotide primers and reaction buffer; the reaction was initialized by addition of 0.5U of BioTaq polymerase (BioLine, UK). All reactions were performed in a thermocycler (Thermofisher, UK), in a total volume of 50 1d. YKO strain genotypes were validated using previously described primers, designed specifically for each null mutant (consult the original paper, (749) and the accompanying websitel). PCR products were inspected using agarose gel electrophoresis and a 1kb ladder (New England Biolabs, USA).
2.1.6 Ribonucleic acid (RNA) extraction and quantification
S. cerevisiae and C. glabrata cells (0D600 = 0.4 — 0.8) were harvested with a benchtop centrifuge (3000 rpm, 5 min) and washed twice in sterile distilled water. Fungal total RNA was extracted by mechanical disruption of cells (FastPrep and acid washed glass beads, BI0101, US) followed by purification with RNEasy mini-kit columns and reagents, incorporating an in situ DNase digestion step (Qiagen, UK). Quantification of samples (0 —3.7 pg/mL) was performed by measuring A260 on a nanodrop ND-1000 spectrophotometer (Labtech, US). A2601280 and A260/230 ratios were used as indicators of protein and salt contamination respectively. Samples were retained if these ratios where in the 1.8 — 2.5 and 2.0 — 2.5 ranges respectively.
RNA sample quality was assessed with the bioanalyzer nano series microfluidics platform (Agilent, UK); quanitification of 26S and 18S peaks was performed using the Eukaryotic RNA assay protocol within the Agilent Expert software (version B.02.02). Samples showing distinguishable 26S and 18S bands on the synthetic electropherogram, and 26S:18S peak height ratios > 1.7 were retained for use as templates in reverse transcription and fluorophore labelling reactions.
2.1.7 Linear DNA and PCR extraction and purification
Linear DNA and PCR fragments were routinely purified from 1% (w/v) agarose gels or from solution using a QlAquickTM (QIAGen LTD, UK) column according to the manufacturer's protocol.
Ihttp://www-sequence.stanford.edu/group/yeast_deletion_project/protocols.html

Chapter 2. Materials and Methods 59
2.1.8 Agarose gel electrophoresis
Samples of DNA were routinely separated on 1% (w/v) agarose made with TBE (Tris-Borate: 4 mM EDTA, 8.9 mM TriZMATm base), containing a 1:10000 dilution of SYBR Safe DNA gel stain (Invitrogen, USA). Gels were inspected under a Gel Dock transilluminator (Alpha Innotech, USA).
2.1.9 Yeast DNA extraction
DNA extractions were performed using the MasterPure Yeast DNA Purification Kit (Epi-centre Biotechnologies, Madison, USA) according to manufacturer's instructions.
2.1.10 Transformation of yeast cells with electroporation
Yeast cells were transformed using a modification of the electroporation protocol (39). Briefly, 2.5 mL of a saturated overnight culture was transferred into 50 mL pre-warmed YEPD and incubated for a further 4 h at 37°C (C. glabrata) or 30°C (S. cerevisiae), 180 rpm. Cells were harvested by centrifugation, washed in sterile water and resuspended in 8 mL dd H20, 1 mL 10 x TE (10 mM Tris-HC1 (pH 8), 1 EDTA (pH 8)) and 1 mL LiAc (1 M) and incubated at 30°C for 30 mM at 130 rpm. Then 250 µL dithiotheitol DTT (1 M) was added and incubated for a further 60 min at 30°C at 130 rpm, 40 mL of dd H2O added and the cells harvested by centrifugation.
All subsequent procedures were performed at 4°C. The cells were resuspended in 25 mL ice-cold dd H20, harvested by centrifugation, washed in 5 mL sorbitol (1 M) and resuspended in 550 ,uL sorbitol (1 M). The competent cells (45 ,u,L per reaction) were transferred to pre-chilled electroporation cuvettes and 100 ng DNA added, mixed and left at 4°C for 10 mM. The cuvettes were pulsed at 1.5 kV, 25 pF, 200 ft The cells were recovered by gentle mixing with 950 it L YEPD and incubated for 4 h at 30°C and plated onto YEPD plates containing 200 pg/mL nourseothicin and incubated for 2-4 days at 30°C or 37°C
2.1.11 Western blots
To prepare samples for the phosphorylation assay, C. glabrata ATCC 2001 cells were cul-tured in 100 mL of YEPD (37°C, 180 rpm) to OD600= 0.55, at which time 25 mL samples were collected and diluted either in 25 mL YEPD or YEPD supplemented with sorbitol (1 M final concentration) to provide the hyperosmotic stress. Cultures were incubated for a further 15 mM prior to harvesting by centrifugation (2000 rpm, 2 min, 4°C). The super-natant was discarded, pellets were retained and snap-frozen in liquid nitrogen before storage at -80°C. Phosphorylation assays were performed as described previously in Candida albi-

2.1 Experimental Methods 60
cans using an anti-phospho-p38 MAPK (Thr180/Tyr182) antibody (9211S, New England Biolabs, USA).
2.1.11.1 Design of probes
S. cerevisiae microarray oligonucleotides were developed by Agilent using oligonucleotides designed in (134).
C. glabratra microarray oligonucleotides used in the Agilent platform were designed by Butler laboratory at University College Dublin, Eire and produced by custom production spotting 5908 69- or 70-mer oligonucleotides synthesized replicated in quadruplicate for each of the 4x 44k arrays on a slide. A. fumigatus microarray oligonucleotides were developed by TIGR, now the JCVI (499).
2.1.11.2 Microarray slides
The Agilent 4x 44k microarray platform was adopted for environmental stress studies in C. glabrata (AMADID 015761) and S. cerevisiae .
COGEME consortium microarrays were used for the hyperosmotic stress transcript profiling of ATCC 2001 and stell and hogl strains (Chapter 3)
All A. fumigatus transcription profiling experiments used Af293 DNA amplicon mi-croarrays (499).
2.1.12 Reverse transcription and fluorescent labelling of cDNA
For environmental stress experiments in S. cerevisiae and C. glabrata reverse transcription and fluorophore labelling was performed with the ChipShotTM direct labelling and clean-up system (Promega, UK) using a thermocycler for all incubation steps. Priming of mRNA within the total RNA population was achieved by incubating RNA template with oligo dT and random primers (65°C, 10 minutes) before rapid chilling to allow primer annealing (ice, 5 minutes) and subsequent addition of reverse transcription reaction components to the primed mix (final volume 40 pl).
Final reaction concentrations were as follows: total RNA template (1.25 pg p1-1), ran-dom primers (75 pg p1-1), oligo dT primer (50 pg p1-1), ChipShotTM reaction buffer (1 x), MgC12 (12.5 mM), dNTP mix (12.5 mM), Cy3Thor Cy5TM-dCTP (25 pg p1-1) and ChipShot'M reverse transcriptase. Reaction mixtures were briefly mixed with a vortex, spun in a micro-centrifuge (13000 rpm, 1 minute) and pre-incubated in a dark room (20-25°C, 30 minutes) before transfer to the thermocycler (42°C, 2 hours). Thereafter, cDNA synthesis reactions were terminated by incubation of samples with RNaseH (1 pl, 37°C, 15 minutes). The re-action was neutralized by adding 4 pl of sodium acetate (5 M). Fluorophore labelled cDNA

Chapter 2. Materials and Methods 61
samples were added to ChipShotTM binding buffer and transferred to ChipShotTM columns for a cycle of three washes (500 pi, 80% (w/v) ethanol, 13000 rpm, 1 minute). Samples were eluted in 600 of elution buffer and quantified using the microarray procedure on the nanodrop ND-1000. Acceptable sample yield ranges for Cy3TM and Cy5TM labelled cDNA were 1.2 - 2.4 pg and 1.0 - 2.4 pg respectively; associated acceptable yields of incorporated dye were 100 - 225 and 50 - 160 pmol respectively.
2.1.12.1 Hybridization, washing and scanning of cDNA microarrays
2.1.12.2 Image acquisition and data analysis
For C. glabrata and S. cerevisiae (both agilent and COGEME arrays), intensities of hy-bridized probes were oriented and measured with a GenePix 4000b semiconfocal microarray scanner and GenePix version 6.0 software (Axon Instruments, USA). Scanning parameters were adjusted for the high density spotting format of the agilent slide. Pixel size was set to 5,um with 3 lines used to average the spot intensity. The photomultiplier tube (PMT) voltage settings for each channel (532 nm and 635 nm) were adjusted for each scan to give an overall intensity ratio across each slide of 1, a background intensity at 532 nm of approximately 100, and a maximal intensity close to saturation (2 x 1016) for a 16-bit TIFF image. The GenePix proprietary spotfinding algorithm was used in combination with the "gal" meta-data file to map intensity values to oligonucleotides on the array that represent ORF in either the S. cerevisiae or C. glabrata genome. Image files were ex-ported as GenePix results files (GPR) which were used in subsequent statistical analyses. Hybridised A. fumigatus slides were scanned using the Axon GenePix 4000B microarray scanner and the TIFF images generated were oriented and analyzed using TIGR Spotfinder (http : //www t igr org/ sof twar e /) to identify poor quality spots.
2.1.13 Statistical analysis of microarray data
All data processing and statistical analyses were performed using the R statistical frame-work (562) for scientific computing, bioconductor (643) and the limma package (644) for linear modelling of microarray data. Detailed descriptions of exploratory and design specific analysis can be found in the relevant chapters. Briefly, poor quality spots were filtered as previously described and systematic spatial effects were normalized within arrays using the print-tip lowess method and quantile between array normalization encoded in (644).
Least squares regression was used to fit the average log2 intensity ratios, residuals and the p values, corrected for multiple testing using empirical Bayes methods (644) or Benjamini-Hochberg correction. Non standard pre-processing steps, such as single channel normal-ization or estimation of amplification effects are detailed in the relevant chapters (Chapter

k-Clique Community Discovery Algorithm
5 1
°
1 1 0 0
1 1 0 0
O 0 0 0
O 0 0 0
0 0 0
5
1 0
O 0
O 0
O 0
1 0
a 0 0
5 3 2 1 3 1
4 2 1 1 1
2 2 3 2 1 2
Z 3 0 1
I 1, 0 1 2
1 1 2 1 2 4
clique clique overlap matrix •
(b) Outcome of 4 -clique percolation (a) Input network
2.2 Computational Methods 62
3 and 4). Least squares regression and linear modelling for microarrays are described in detail (642; 643; 644)
2.2 Computational Methods
2.2.1 Computation of network statistics
Graphical representation of networks and their statistical computation was performed using the nodebox and networkx toolboxes developed for python (263; 717). Cytoscape was also used for graphical representation of networks (624; 137).
2.2.2 k-clique percolation and community finding
The majority of community finding algorithms detect standard partitions of a network. The problem of graph partitioning concerns dividing the nodes into discrete groups of predefined size, such that the number of edges lying between them is minimal. Most of these methods prohibit multiple community membership for a single node, however in real graphs, single nodes are often shared between communities, and the problem of detecting overlapping communities has recently been addressed by Palla and colleagues (522) (Figure 2.1).
Figure 2.1: Overview of k-clique percolation algorithm adapted from (522)

63
Chapter 3
The Hogip and Stellp dependent transcriptional effects of hyperosmotic stress in Candida glabrata
3.1 Chapter Overview
Yeasts, devoid of any behavioural homeostatic facility, have evolved co-ordinated signalling pathways to adapt to the fluctuating physicochemical nature of their habitat. Several sens-ing and adaptive mechanisms have been discovered to date ((538; 368; 554; 50; 190; 315; 480; 261), however, the high osmolarity glycerol response (HOG) pathway (89; 292), and its role in the adaptation of S. cerevisiae to hyperosmotic stress in particular, is one of the best characterized. This model pathway forms the basis for studying hyperosmotic stress adaptation in related pathogenic fungi where homologous mitogen activated protein kinases coupled to stress adaptation are referred to as stress activated protein kinase (SAPK) cas-cades (192). A hyperosmotic stress is imparted on a cell when the external water potential of the culture medium is less than the sum of the water potential of the soluble component of the cell and the turgor pressure imparted by the mechanical structures bounding the cell, leading to a net outflow of water from the cell until equilibrium is reached (443).
The HOG pathway modulates the rate at which equilibrium is reached by inducing cellular changes leading to the accumulation of intracellular glycerol (12; 392), a compat-ible solute that serves to decrease intracellular water potential and therefore the rate of water outflow. This is achieved by increasing the metabolic flux through glycerol biosyn-thetic pathways, and also by closing aquaglyceroporins (678; 428; 473) located in the cell

Hyperobrpotic Signal
A #= no C. glabrata ortholog
= truncated in C.giabrata
PTP2 PTP3
Glycerate-3-phosphate D-glycerate
10
ALD4. ALD5, ALD6
04
B
3.1 Chapter Overview 64
membrane that permit glycerol efflux. This exemplar hyperosmotic adaptation pathway
architecture was deduced with a series of genetic screens and consists of at least two func-tionally redundant signalling branches, defined by the putative osmo-sensors at the top of
each cascade: the two component like Slnl phosphorelay branch (553; 575), and the mucin-like transmembrane receptor Shol branch (551; 152). A MAPKKK architecture lying at the core of each (551; 552), relays the upstream signal to the MAPKK Pbs2p (89) and ultimately the MAPK, Hog1p (89) - a major effector of hyperosmotic stress adaptation in
S. cerevisiae (Figure 3.1).
DAK1, DAK2
GRP
l'OD-glyceraldehyde
•,.0 Glycerol 0.4 04
Glycerone phosphate Glycerine
HOF72, RriN ' II &Hi
GPD1, Gun' 0
II.Nn-glycerot-3-phosphate
Adapted from Kegg pathway. Kaneisha laboratories (http://www.genome.jp/kegg/).
Figure 3.1: Architecture of the HOG pathway in S. cerevisiae. See text for descrip-tions.

• . • Several Factors
(71'
?
0 O!00
0 0 MAPKKK
0 O MAPKK O MAPKK
Chapter 3. The Hog1p and Stellp dependent transcriptional effects of hyperosmotic stress in Candida glabrata 65
Mating Morphological Cell Wall Osmoregulation Cell Wall Sporulation Response Switch Integrity Construction Response
Figure 3.2: Architecture of MAPK cascades of S. cerevisiae, incuding the HOG osmoadaptation cascade. Adapted from (292). Orange circles denote putative pro-teins embedded in the plasma membrane, connected to the upstream G-protein (Stel8p, Ste4p, Ras2p, Rholp, Cdc24p, Cdc42p, Gpalp, Rom2p) or phosphorelay (Ypdlp, Ssklp) control components (black circles). Kinases (either p21 activated kinases Ste2Op or serine/ threonine kinases Pkclp, Spslp), depicted by grey circles, lie upstream of the core MAP kinase architecure (IVIAPKKK (Ste11p, Ssk2p, Ssk22p, Bcklp) MAPKK (Ste7p, Pbs2p, Mkklp, Mkk2p) —p MAPK (Fus3p, Ksslp, Hoglp, Slt2p, Smklp)) whose activation re-sults in recruitment of transcription factors (Stel2p, Rlmlp) which mediate transcriptional changes in response to the detected signals. MAPKKK —÷ MAPK components are depicted as blue circles colour-graded in relation to their hierarchy; the darkest hue representing a MAPKKK and the lightest blue representing a transcription factor activated by the ap-propriate signal cascade. Coherence of osmoadaptation signalling is maintained by the Hoglp MAPK, which prevents cross-talk with the mating response pathway (516). Genes disrupted in this study are depicted by shadowed circles with red boundaries. Unknown inputs, regulators or transcriptional events are denoted by a question mark.

3.1 Chapter Overview 66
The HOG pathway comprises at least two signalling branches that transduce a signal detected in the plasma membrane by putative osmosensors (Figure 3.1 A orange circles) to the upstream G-protein related proteins (Cdc24p, Cdc42p) or phosphorelay (Ypdlp, Ssklp) control components (Figure 3.1 A black circles). These upstream regulatory components transduce the signal to a MAPKKK (Figure 3.1 A dark blue circles , Ssk2, Ste11) —> MAPKK (sky blue circles, Pbs2p) —> MAPK (Figure 3.1 A cyan circles, Hog1p) signalling cascade, in the case of the Slnl branch this is via p21-activated serine-threonine kinase (Ste20p, Figure 3.1 A grey circle). The Slnl branch (435; 531), similar to the canonical two-component-like signalling cascades found in prokaryotes and other fungi (reviewed in (662)), functions as a negative regulator of the HOG pathway and comprises the Sln1p histidine sensor-kinase, the Ypdlp phosphorelay protein (553) and the Ssklp response regulator (553). The autophosphorylation of Sln1p is modulated by external water potential, with the greatest level of phosphorylation occurring in hypotonic culture media (553; 552). Switching cells to a hypertonic environment (denoted by the flat arrow head and negative symbol in Figure 3.1 A) results in a relative dephosphorylation of Sln1p and Ypdlp, and ultimately the Ssklp response regulator. In its dephosphorylated form, Ssklp acts as a positive regulator of Ssk2p/Ssk22p activity, and also the downstream MAPKKs (Pbs2p) and MAPK (Hog1p) (435).
In the second branch, the Sholp transmembrane protein contains an intracellular Src homology 3 (SH3) domain, which binds to a proline-rich region of the Pbs2p MAPKK (435) which is activated in response to hypertonicity (denoted by the arrow head and positive symbol in Figure 3.1 A). Sho1p recruits Pbs2p to the cell membrane following osmotic stress, and both Sholp and Pbs2p bind to the MAPKKK Sten') (684). Sten') is acti-vated by phosphorylation which requires the Rho-like GTPase Cdc42p, the p21-activated serine/threonine protein kinase Ste20p, and the adaptor protein Ste5Op (715; 564) (scaf-fold denoted by grey oval shape encompassing these proteins in Figure 3.1 A). A third osmosensing branch requires the Msb2p protein, which has a single membrane-spanning do-main and a large predicted extracellular domain; this sensing branch is partially redundant with Sholp (517). Once activated, the phosphorylated Hog1p translocates to the nucleus in an Nmd5p mediated fashion (201), where it promotes adaptive transcriptional changes. Hog1p controls the transcriptional response to hyperosmotic stress by activating members of distinct transcription factor families that in turn drive transcriptional responses to hyper-osmotic stress in S. cerevisiae: Skolp (557; 558), Hot1p (13; 578), Msnlp, Msn2p/Msn4p (581), Sgdlp (410) and Gen4p (532) (not shown). Nuclear Hog1p mediates the transcrip-tion of key transcripts highlighted in this figure including glycerol-3-phosphate dehydro-genase (GPD1,GPP2) (reaction catalyzed shown in Figure 3.1 B), and phosphotyrosine phosphatases (Figure 3.1 green circles, PTP2, PTP3) which provide negative feedback by

Chapter 3. The Hog1p and Stellp dependent transcriptional effects of hyperosmotic stress in Candida glabrata 67
dephosphorylating Hog1p (449; 757; 448). Type two phosphotyrosine proteases (Figure 3.1 green circles PTC1, PTC2) also dephosphorylate Hog1p (729; 767). During the course of this work, Gregori et al., showed that the C. glabrata ATCC 2001 strain contains a prema-ture stop codon in the SSK2 gene that leads to a dysfunctional Slnl signalling branch (255) - denoted by the * (Figure 3.1 A) adjacent to the Ssk2p component of the Shol branch. C. glabrata also lacks an Ssk22 homolog (denoted by the # in Figure 3.1 A) - a functionally redundant paralogue of the Ssk2p MAPKKK.
The HOG pathway and adjacent MAPK signalling cascades contain common functional elements both in the upper parts of the regulatory hierarchy, and in the downstream mod-ules that regulate the phosphorylation signal on MAPK proteins (Figure 3.2). Hoglp's position within the signalling cascades means that it not only plays an important role in signal transduction for hyperosmotic stress adaptation, but also maintains the coherence of this signal, namely the correct relay to the downstream effectors of osmotic stress adap-tation and not to the morphogenesis, cell integrity or mating pathways (516). O'Rourke and Herskowitz demonstrated that a hogl mutant subjected to an osmotic upshift aber-rantly activates the pheromone response pathway as evidenced by activation of a FUS1::lacZ reporter, morphological changes, and mating in ste4 and ste5 mutants. This cross-talk re-quired the osmosensor Sho1p, as well as Ste20p, Ste50p, the pheromone response MAPK cascade (Stellp, Ste7p, and Fus3p or Ksslp), and Stel2p but not Ste4p or the MAPK scaf-fold protein, Ste5p(516). More recently, Shock et al., demonstrated that the Hog1p MAPK interrupts signal transduction between the Ksslp MAPK and the Tecip transcription fac-tor to maintain pathway specificity between the osmoadaptation, mating and filamentous growth pathways (633). The transcriptional responses to hyperosmotic stress have been studied extensively in S. cerevisiae, both in broader investigations of the environmental stress response (ESR) (230; 118), and also in more focused studies of hyperosmotic stress imparted by high salinity (550) or high exogenous concentrations of an impermeable solute such as sorbitol (581; 230).
These pathways have taken on new importance in the context of related, pathogenic fungi; deletion of MAPKs in some human pathogens significantly attenuates virulence (14; 42), whilst such pathways are dispensable for virulence in a number of plant pathogens. For example, in C. albicans it has been demonstrated that the Hog1p MAPK is activated in response to diverse stimuli, many of which are likely to be encountered in the host, such as: oxidative stress (15), osmotic stress, the morphogenic agent serum, and the quorum-sensing molecule farnesol (639). The SAPK network in this pathogen has diverged from corresponding networks in the non-pathogenic model yeasts S. cerevisiae and Schizosac-charomyces pombe, which suggests that specialised, niche-specific, Hoglp-mediated stress responses have evolved in this pathogenic fungus (639). A mathematical modelling approach

3.1 Chapter Overview 68
has been used in conjunction with single-cell measures of GFP-promoter fusion strains (452) to investigate how signal specificity is maintained in MAPK/SAPK signalling cascades. Mc-Clean et al., showed that the hyperosmotic stress and pheromone response pathways which share a common MAPKKK (Ste11p) achieve specificity by mutual inhibition rather than insulation and that this mutual inhibition must occur downstream of the components com-mon to both pathways; Zou et al., later used a modelling approach to provide evidence that Pbs2p and Hog1p are essential for maintenance of this specificity (774). SAPKs are critical for survival of some opportunistic fungi in the host, as C. albicans cells lacking HOG1 display significantly attenuated virulence in a mouse model of systemic candidiasis (14). Similarly, a recent study in C. neoformans revealed that the Hog1p SAPK is essential for virulence in the highly pathogenic serotype A Glade, and, significantly, uncovered clear differences in the mechanisms underlying Hog1p regulation in highly virulent and less viru-lent serotypes (42). Collectively, these studies demonstrate: (i) that stress responses are an essential virulence attribute of human pathogenic fungi and (ii) the importance of SAPK pathways in these stress responses.
The role of C. glabrata SAPK pathways has been partially investigated in our labora-tory by Ana-Maria Calcagno-Pizarelli, who identified C. glabrata homologues encoding the S. cerevisiae upstream p21- activated serine/threonine kinase Ste2Op (103), the MAPKKK Stel1p (104) and the transcription factor Stel2p (105). Cloning, disruption and functional complementation studies were used to investigate the role of Ste2Op in hyperosmotic stress adaptation, filamentation and mating, cell wall integrity and in a mouse model of candidia-sis. S. cerevisiae ura3/ura3 ste20/ste20 cells were transformed with the pAMC86 plasmid bearing the C. glabrata STE20 gene and a URA3 marker, with subsequent selection of uracil prototrophs. These colonies were selected and tested in mating and filamentation defect assays which revealed that C. glabrata STE20 complements the mating and filamen-tation defects of S. cerevisiae ste20 strains. Although C. glabrata Ste2Op is required for growth on solid hypertonic growth medium (containing either 1M NaC1 or 1.5 M sorbitol) and agar supplemented with calcofluor white (lmg/ml), it is not required for nitrogen star-vation induced filamentation. Moreover, in a mouse model of candidiasis, ste20 strains are partially attenuated for virulence in comparison to reconstituted ste20:STE20 strains (103). Similar cloning, complementation and disruption studies were also performed for the C. glabrata STE11 gene encoding a MAPKKK: the mating and filamentation defects of a S. cerevisiae strain are complemented following transformation of a plasmid bearing the C. glabrata STEN gene. Cell wall integrity and hyperosmotic stress adaptation as-says were also carried out for the C. glabrata stel 1 strain and revealed that this gene is required for adaptation to hyperosmotic stress but is largely dispensable for maintenance of cell wall integrity (104). Survival analysis conducted in the murine model of candidiasis

Chapter 3. The Hogip and Stellp dependent transcriptional effects of hyperosmotic stress in Candida glabrata 69
demonstrated that sta./ mutants are also partially attenuated for virulence in comparison to the re-constituted stell:STE11 strain (104). These results suggested that although the components of SAPK pathways are conserved, they play different roles in the biology of each species. In this section we aim to expand our investigation into the divergence of this pathway by studying the role of an homologous MAPK (putative SAPK) in the re-lated pathogenic fungus C. glabrata. I present an investigation into the putative role of the homologous C. glabrata Hog1p as a regulator of hyperosmotic stress adaptation, although there are plans to assess the role of Hog1p in a variety of environmental stress conditions. In collaboration with Janet Quinn (University of Newcastle), we had the following aims:
1 Identify whether HOG1 is required for growth in hyperosmotic stress conditions by constructing a hog1 strain and assessing its growth in hyperosmotic stress medium relative to rich culture medium.
2. Assess how Hog1p functions under hyperosmotic stress conditions with respect to its phosphorylation kinetics and determine if any observed phosphorylation depends on upstream MAPKKK architecture conserved with related fungi.
3. Further assess any divergence in fungal hyperosmotic stress adaptation by assaying the transcript profile of wild-type C. glabrata cells subjected to hyperosmotic stress and comparing it with published data for S. cerevisiae and C. albicans.
4. Assess how divergent HOG1-dependent transcriptional responses are by further com-paring the transcript profile of hog1 cells subjected to hyperosmotic stress, with equiv-alent data in S. cerevisiae and C. albicans.
5. Identify a hypothetical hyperosmotic stress regulation network by mapping regulated transcripts to a hypothetical osmostress regulation network inferred from S. cerevisiae consensus transcription factor binding sites. This hypothetical network can be queried to identify potential regulators of the hyperosmotic stress response in C. glabrata.
It is important to stress that my involvement in this work has only been in the technical analysis and biological discussion of the gene expression microarray data in context of the initial phenotype discoveries. I am very grateful to Janet Quinn for involving me in this project. Unless otherwise stated, all figures in this section have been generated from my analysis.

3.2 Specific Methods 70
3.2 Specific Methods
3.2.1 Construction of a C. glabrata hogl null mutant
BLAST (16) searches of the C. glabrata genome (630) identied an open reading frame, CAGLOM11748g, encoding a protein with 85% amino acid identity to S. cerevisiae Hog1p. A three-step overlapping PCR strategy was devised by our collaborator, Janet Quinn (Uni-versity of Newcastle), to generate a HOG1 disruption cassette based on this nucleotide sequence, for subsequent lithium acetate based transformation of a C. glabrata histidine auxotrophic strain. A brief description of the construction method follows:
In the first step, two fragments, an 870 bp 5' fragment (-942 to -73) and a 512 bp 3' fragment spanning coding sequence, stop codon and 3' untranslated region (+1224 to +1735) were amplified with M13 overhangs from C. glabrata ATCC 2001 genomic DNA template by high fidelity PCR using primer pairs HOG15-5 + HOG15-3 and HOG13-5 + HOG13-3 respectively (Supplementary Table D.1). Secondly, primer pair HOG1-HIS5 + HOG1-HIS3 was used to amplify a HISS marker gene template that had been previously cloned into the M13 site of vector pTW25 (734), yielding the HISS gene with M13 overhangs and HOG1 5' (-73 to -94) and 3' (+1224 -+1243) sequences. Finally, the HOG1 disruption cassette was constructed using the products from the above two reactions. The disruption cassette was amplified by PCR using the 5' and 3' HOGI fragments from the first step which overlap the amplified HISS template from the second step at the 5' and 3' ends. The resulting fragment was used to transform C. glabrata AHT6 (734), and restoration of histidine prototrophy was used to select putative hog1 colonies. Putative hog1 mutants were tested for homologous integration and gene replacement by PCR, using the primers HOG13ex (external to the disruption cassette) and HIS3INTF (internal HIS3). The selected hog1 strain used in this study was given the laboratory name JG7.Two strains disrupted in the STE11 and STENO loci respectively were created by Ana Maria-Calcagno Pizarelli (106; 103; 104). Full details of these strains and JG7 can be found in the supplementary data and references therein (Supplementary Table C.1).
3.2.1.1 Western blot analysis
Western blot analysis was performed on whole cell extracts from wild-type, ste11 and ste20 C. glabrata cells following treatment with 1 M sorbitol. Blots were probed with an anti-p38 MAPK antibody (New England Biolabs 9211S), which recognises the phosphorylated, active form of C. glabrata Hoglp (phospho-Hoglp). Total levels of Hog1p were determined by stripping and reprobing the blot with an anti-Hoglp antibody (Y-215 Santa Cruz). Full details are described in the materials and methods section.

Chapter 3. The Hog1p and Ste11p dependent transcriptional effects of hyperosmotic stress in Candida glabrata
3.2.1.2 Genome-wide transcript profiling
Transcript profiling was performed in biological triplicate using C. glabrata strains and CO-GEME consortium arrays: ATCC 2001 (wild type), stell (AI\43, (104)) and hog' (JG7, this study). Hybridizations were also performed using a dye-swap to account for potential dye-bias in the experiments. Strains were cultured in YEPD at 37°C, 180 rpm to OD600 = 0.55 and harvested after 15 minutes of mock stress treatment with YEPD or hyperos-motic stress treatment with 1 1VI sorbitol. Hyperosmotic stress was induced by adding 60 ml of pre-warmed YEPD + 2.4 M sorbitol to 50 ml of OD600 = 0.55 cell suspension to give a final concentration of 1 M sorbitol and an OD600 = 0.23. Cells were collected by centrifugation (3000 rpm, 5 minutes), snap-frozen in liquid nitrogen and stored at -80°C before isolating total RNA. Total RNA was prepared by shearing frozen cell pellets with glass beads and a Mini-Beadbeater-8 (Biospec Products, Bartlesville, USA) and extraction with Trizol Reagent (GibcoBRL), as described previously (275). Hybridization and scan-ning was performed as described in the materials and methods section. Relative abundance of transcripts is quantified by log2 intensity ratios (also known as M values) of intensity measured in the CySTM channel (red) and the Cy3T' channel (green), values for which are dependent on each hybridization comparison. Average intensity A values are quantified as (0.5 x log2 (Cy5Tm .) + log2(Cy3m.)) and are used in conjunction with M values in quality control procedures to assess how log2 ratios vary as a function of spot brightness. All sta-tistical analysis was performed in the R statistical computing framework (562) using the limma package (643) for linear modelling of microarray data and single channel normaliza-tion procedures. Hierarchical clustering was performed using the hclust function, with the default settings of complete agglomeration and Euclidean measures of similarity:
For two n—dimensional points, P = (pi , p2, • • • ,pri) and Q = (gi,g2 ,• • • , g„) then Eu-clidean distance is
D _ Om. qi ) 2 + (192 (22 ) 2 ± (prt qn ) 2 (3.1)
Heatmaps of cluster dendrograms were created using the heatmap.2 function provided in the gplots package.
3.2.1.3 Downstream b ioinform at ic analyses
Over-representation of gene ontology terms (35) was investigated using the Saccharomyces Genome Database' hosted GO term finder (83), using a C. glabrata gene ontology inferred by homology to S. cerevisiae. Over-represented GO Process, GO Function and GO Corn-
lhttp://www.yeastgenome.org/
71

3.3 Results 72
ponent terms were identified in relation to this inferred whole-genome background set. The null distribution was approximated by a hypergeometric distribution and statistically sig-nificant over-represented terms were defined as having a Fisher's exact p value of < 0.05. Annotated metabolic pathways were created by permission of the Kyoto Encyclopaedia of Genes and Genomes (KEGG) (343). The colour pathway tool was used for the C. glabrata reference species. At this point, it is appropriate to note the following analysis of transcript regulation is encumbered with two caveats. First, transcription profiles are often discussed in lieu of protein levels, but gene expression profiles themselves describe only a part of the adaptive responses to stress. Turnover of mRNA, post-translational modification of pro-teins and patterns of localization can all significantly affect the exact nature of a cellular response. Nevertheless, these profiles provide useful clues to how the cell may orchestrate a response to an environmental stress. Second, unlike the collegial and prolific S. cerevisiae research community, the poorer C. glabrata cousins have very few giants' shoulders upon which to stand - much of our knowledge of C. glabrata is inferred by evolutionary com-putational annotation. With this in mind, homologous S. cerevisiae gene names are used, and unless otherwise stated, it should be assumed that appropriate S. cerevisiae comple-mentation studies have not been performed. Furthermore, functional assignment is based on descriptions cited in the SGD (131) database for each transcript discussed in these re-sults and is used throughout the discussion and results section. Supplementary data tables comprise C. glabrata ORF names alongside their S. cerevisiae homologous standard and canonical names.
3.3 Results
3.3.1 C. glabrata, hyperosmotic stress dependent, Hogip phosphorylation is abol-ished in a ste// background
Colony growth assays and western-blotting of Hog1p phosphorylation indicate that Hog1p is phosphorylated upon exposure to hyperosmotic stress, and that is required for growth in high osmolarity medium (Figure 3.3). All three mutant strains (hogl, stel 1 and stab') showed equivalent growth to the wild-type on YEPD agar, but impaired growth in solid medium supplemented with 1 M sorbitol. The hogl and stel 1 stains showed equivalent growth defects in 1 M sorbitol medium over the range of cell titres used in this study, whereas the ste20 was not as sensitive (some growth at highest cell titre - 104 cells).
A two-stage Western blotting analysis of the Hog1p putative SAPK, using an anti-p38 antibody to detect phosphorylation specific epitopes, and an S. cerevisiae anti-Hoglp revealed a low level of Hog1p phosphorylation in rich culture medium, that is rapidly up-regulated and sustained for twenty minutes upon application of exogenous sorbitol (1 M)
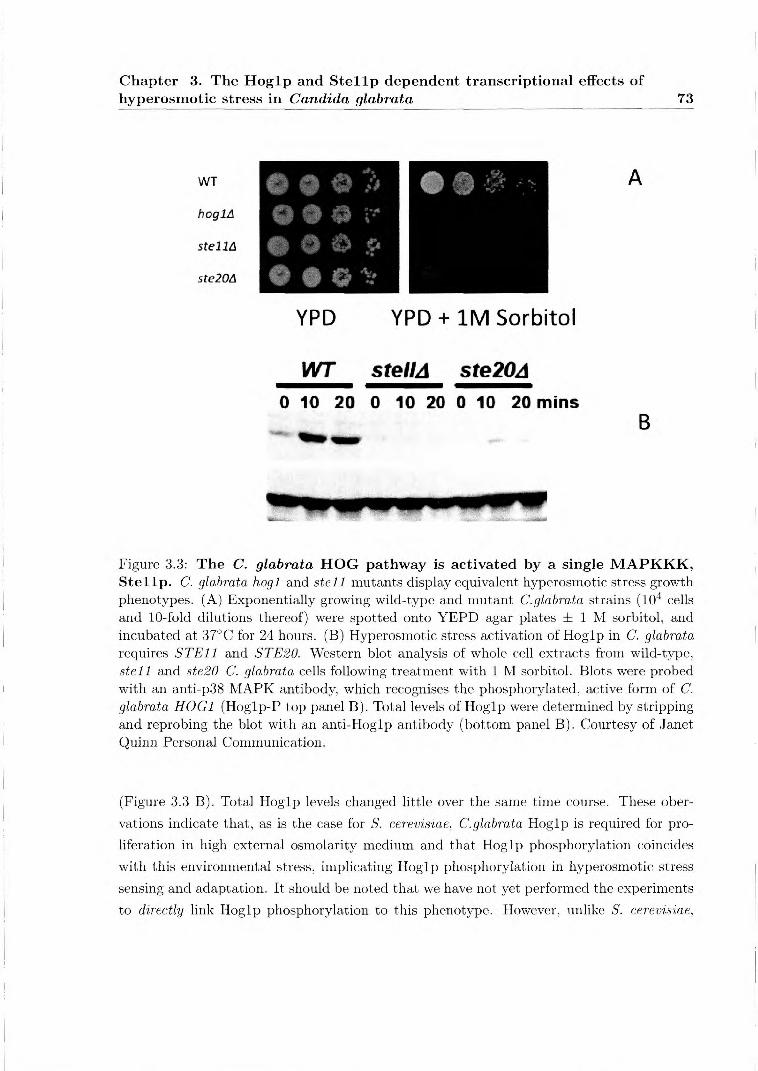
Chapter 3. The Hog1p and Stellp dependent transcriptional effects of hyperosmotic stress in Candida glabrata 73
WI
hogld
ste118
ste20d
0 ze
*COOP • • ►t•
A
YPD
YPD + 1M Sorbitol
WT stelld ste2O4 0 10 20 0 10 20 0 10 20 mins
B Ism
Figure 3.3: The C. glabrata HOG pathway is activated by a single MAPKKK, Stellp. C. glabrata hogl and stell mutants display equivalent hyperosmotic stress growth phenotypes. (A) Exponentially growing wild-type and mutant C.glabrata strains (104 cells and 10-fold dilutions thereof) were spotted onto YEPD agar plates ± 1 M sorbitol, and incubated at 37°C for 24 hours. (B) Hyperosmotic stress activation of Hoglp in C. glabrata requires STEll and STE20. Western blot analysis of whole cell extracts from wild-type, step and ste20 C. glabrata cells following treatment with 1 M sorbitol. Blots were probed with an anti-p38 MAPK antibody, which recognises the phosphorylated, active form of C. glabrata HOG1 (Hog1p-P top panel B). Total levels of Hoglp were determined by stripping and reprobing the blot with an anti-Hoglp antibody (bottom panel B). Courtesy of Janet Quinn Personal Communication.
(Figure 3.3 B). Total Hoglp levels changed little over the same time course. These ober-
vations indicate that, as is the case for S. cerevisiae. C.glabrata Hoglp is required for pro-
liferation in high external osmolarity medium and that Hoglp phosphorylation coincides
with this environmental stress, implicating Hoglp phosphorylation in hyperosmotic stress
sensing and adaptation. It should be noted that we have not yet performed the experiments
to directly link Hoglp phosphorylation to this phenotype. However, unlike S. cerevisiae,

3.3 Results 74
where input signals from both osmosensing branches must be disrupted to block hyperos-motic stress induced Hogip phosphorylation (435), this can be achieved by deletion of a single MAPKKK belonging to the predicted Shol branch upstream of Cg-Hoglp suggesting that C. glabrata signals only through the Shol signalling branch. These results were later demonstrated independently by the Kuchler laboratory (255). We next investigated the transcriptional effects of hyperosmotic stress on C. glabrata, and the roles of Hog1p and Ste1 1p in regulating these transcriptional responses.
3.3.2 The transcriptional response of wild-type C. glabrata cells exposed to transient hyperosmotic stress
Hybridizations were designed (Figure 3.4) specifically to compare the wild-type against hogl and stell strains in rich culture medium and in rich culture medium supplemented with a source of exogenous hyperosmotic stress (1 M sorbitol).
To increase the value of this design, I also wanted to investigate the wild-type hyperos-motic stress affected transcriptome of C. glabrata so that it could be compared with pub-lished data in S. cerevisiae, even though conventional analysis of the initial design precluded this. To overcome this limitation, I treated the two-colour gene expression microarray data as a single-colour experiment where intensity values are measured for one channel only and relative abundances (log ratios) are constructed across channels. This is unusual for the analysis of two-colour chips but is common place in other platforms such as on Affymetrix chips. Since the specific hybridizations compare wild-type in one channel (Cy3TM) and a null strain in the other channel (Cy5TM) with and without hyperosmotic stress, it is theoreti-cally possible to obtain the wild-type hyperosmotic stress response by comparing the single channels of wild-type cells before and after stress (i.e. Cy3TM versus Cyr), and therefore construct a log ratio of intensities spanning hybridizations. This poses particular problems in removing systematic sources of noise to reveal the biological effects of interest. To this end, a standard lowess normalization technique (762), accounting for within-slide bias, was complemented with specific techniques that ensure comparable distributions of Cy5TM and Cy3TM intensities between slides, thus facilitating meaningful construction of log ratios across hybridizations. This was performed with an adapted quantile normalization by Yang and Thorne (690) (Figure 3.5). This method extends the idea of normalizing for equivalent medians or quartiles of the single-channels by requiring every quantile across channels be equivalent, and thus forcing each channel to share a common distribution.
The distribution is estimated by averaging across channels for each quantile. After normalizing within arrays using quantile normalization (Figure 3.5 a) and M values between slides using quantile normalization, intensity values were normalized with "A"-quantile

Chapter 3. The Hogip and Stellp dependent transcriptional effects of hyperosmotic stress in Candida glabrata 75
normalization to make intensity values comparable across slides and suitable for downstream comparison (Figure 3.5 c and d) in both the red and green channels (Figure 3.5 e and f). In this study, the genome-wide transcriptional response (Figure 3.6 and 3.7) of wild-type C. glabrata was constrained to a small proportion (< 5%) of the total genetic repertoire, in a similar manner to published data in S. cerevisiae (Supplementary Data Tables D.3 and D.4) .
This analysis reveals 169 induced and 39 repressed transcripts in the wild-type C. glabrata response to transient hyperosmotic stress. Functional classification of these tran-scripts was performed by inspection of the SGD annotation descriptions and by mapping differentially transcribed genes to annotated KEGG pathways (343) and a gene ontology (GO) (35) inferred by S. cerevisiae sequence homology. Collectively, these downstream analyses reveal the transcriptional regulation of several important biological processes, that are perhaps unsurprisingly categorized under the GO process terms: "Response to chemical stimulus" and "Response to abiotic stimulus". More specifically, induced transcripts are involved in: metabolic processes (14%), vesicular trafficking (9%), transcriptional regula-tion (8%), cell wall remodelling (7%), protein degradation and autophagy (7%), phosphate metabolism (5%), transport of inorganic ions and organic molecules (3%), the protein un-folding response (3%), oxidative stress and DNA damage repair (3%) in addition to a number of uncharacterized proteins (10%). Importantly, key glycerol biosynthetic enzymes are induced (Figure 3.8 and 3.9). The NAD-dependent glycerol 3-phosphate dehydrogenase GPD2 transcript levels are induced, however the array design does not include a specific probe for the isoforms GPD1 or GPD3 and so the transcriptional activity of these paralo-gous genes is indeterminable in this study. The NADP(+) coupled glycerol dehydrogenase GCY1, is proposed to be involved in an alternative pathway for glycerol production is also induced after hyperosmotic stress (122; 121; 504; 143) (Supplementary Data Table D.5).

3.3 Results 76
1
1 Sorbitol
0_0001
stelld
2
hogld CG2001
3
} Sorbitol
0,2L 0
stelld
4
CG2C01
hogld
stelld
} Sorbitol
1 rSorbitol
hogid
mend
CG2001
hogld
} -Sorbitol
CG2001
0,2001
} +Sorbitol
CG2001
,G2001
Figure 3.4: Gene expression microarray hybridization scheme adopted to inves-tigate the HOG1 and STE11 dependency of hyperosmotic stress induced tran-criptional changes in C. glabrata. Cultures of stell, hogl and their parental strain - ATCC 2001 were incubated (YEPD, 37°C, 3.5 hours, 180 rpm) to mid-logarithmic phase (0D600=0.4) and either transiently exposed to hyperosmotic stress (1 M Sorbitol, 15 min) or left untreated. Bi-directional arrows denote dye-swapping of cDNA samples labelled with Cy5
TM or Cy3TM dCTP (black and white rectangles respectively). Biological effects are
numbered 1 through 9. Effects 1 - 4 are measured directly in the hybridization, effects 5 - 9 are estimated indirectly by fitting loge intensity values in a linear model followed by extraction of the relevant effect by application of a contrast matrix.

111111 I I I I I 1
Chapter 3. The Hogip and Stellp dependent transcriptional effects of hyperosmotic stress in Candida glabrata 77
1111111111111111 111111
1 3 5 7 9 11 13 15 17 19 21 23
Slide Number
(a)
1 3 5 7 9 11 13 15 17 19 21 23
Slide Number
(b)
111,1111111111111111
1 3 5 7 9 11 13 15 17 19 21 23
Slide Number
(c)
RG densities
1 1 1 1 1 1
3 5 7 9 11 13 15 17 19 21
Slide Number
(d)
RGdensiMs
-10
0 5
5 -s
10
Intensity Venally
(e)
(f)
Figure 3.5: Single channel normalization methods preserve slide intensity distri-butions for cross-slide comparison and construction of synthetic log ratios of transcript abundance. The effects of normalization procedures on background corrected intensity distributions prior to single-channel linear modelling analysis. Box and whisker plots of M values (log2 (Cy5T9 ) - log2(Cy3TN )) are shown for each slide after lowess normal-ization (a) and lowess normalization + quantile normalization (b). Similarly, equivalent plots of A values (0.5 x log2 (CyC") log2 (Cy3-m )) are shown for each slide after lowess normalization (c) and lowess normalization + "A"-quantile normalization (d). Coloured boxes denote the biological effect (see Supplementary Data Table D.2) measured in the hybridization. Black boxes = STE.0 - WT.U, red boxes = HOG.0 - WT.U, blue boxes = STE.S - WT.S and cyan boxes = HOG.S - WT.S. The pattern of boxes repeats six times, accounting for three dye-swapped biological replicates of each effect. Density plots of red

3.3 Results 78
C o
0 (0
-4 0 4 Value
0 0
Figure 3.6: Heatmap featuring column and row based cluster dendrograms of genome-wide log ratios (log2 (Cy5TM ) - log2 (Cy3TM )) for hyperosmotic stressed wild-type (WT.S - WT) cells compared with stell (STE.S - STE.U) and hogl cells (HOG.S HOG.U) (defined in Supplementary Data Table D.2). The heat-map depicts the magnitude of intensity ratios for each gene on a hue scale ranging from dark blue (repressed) to bright yellow (induced), as indicated by the colour key histogram of log ratio in the top left inset box. Cluster dendrograms, based on the hierarchical clustering of Euclidean distances, indicate the degree of similarity in expression profiles between genes (rows) and between experimental conditions (columns). Vertical dashed lines represent the median loge (Cy5TM ) - log2 (Cy3TM ). Transcripts induced by hyperosmotic stress in wild-type cells form the basis for a "zoomed" view in the related Figure 3.7

0 2 4 6 Value
STE. S
-STE
.0
Chapter 3. The Hog1p and Stellp dependent transcriptional effects of hyperosmotic stress in Candida glabrata 79
Figure 3.7: Heatmap featuring column and row based cluster dendrograms of log2 (Cy5TM ) - log9 (Cy3TM ) for transcripts induced by hyperosmotic stress in wild-type cells, compared with the equivalent values in hyperosmotic stressed hogl and stel1 cells. The heat-map depicts the magnitude of intensity ratios for each gene in the subset on a hue scale ranging from dark blue (repressed) to bright yellow (induced) as indicated by the colour key histogram of log ratio in the top left inset box. Cluster den-drograrns, based on the hierarchical clustering of Euclidean distances, indicate the degree of similarity in expression profiles between genes (rows) and between experimental condi-tions (columns). Vertical dashed lines represent the median log2 (Cy5 - log2 (Cy3TM ) This heatniap is a "zoomed view" of the entire genome and is restricted to the subset of the genome that was significantly induced in the wild-type strain after hyperosmotic stress and compares the expression profile with the equivalent conditions in the hogl and ste11 strains. In this subset, the colour key histogram median is much higher than the genome-wide median of 0, because it is skewed by focusing on the comparison of transcripts induced by hyperosmotic stress in wild-type cells. Panel A denotes HOG1-dependent transcripts with lower log ratios in hyperosmotic stress treated hogl cells than wild type or stell cells. Panel B denotes STE/ / -dependent transcripts with lower log ratios in ste11 cells than wild-type or hogl cells. Panel D denotes transcripts with higher log ratio in hogl cells than stell or wild-type cells subjected to transient hyperosmotic stress. Contrasts are defined in Supplementary Data Table D.2

0.
0
7.4 to; f---0
*, Q°' t .4 I IA g 0
9 i 1 .,'?. F., E
,-, ' g 0 .; g 7-,
O -0. 1 LA
2 L'' 1 0, 1 1 a n .;,• ..... ,g Na t., U1 0 $ii, § r. P.A ...LT.... l`6 1
r2
1.0
3.3 Results 80
Figure 3.8: A transcript encoding the enzyme glycerol dehydrogenase (IUBMB Enzyme code EC 1.1.1.6), involved in glycerophospholipid metabolism is in-duced in wild-type C. glabrata cells exposed to 1 M sorbitol for 15 minutes. Differentially transcribed transcripts were identified by gene expression microarray analysis and mapped to C. glabrata pathways of metabolism in the KEGG database. Red boxes de-note enzymes whose transcripts were more abundant after exposure to hyperosmotic stress imposed by addition of 1 M sorbitol to rich culture medium for 15 minutes.

Chapter 3. The Hog1p and Stellp dependent transcriptional effects of hyperosmotic stress in Candida glabrata 81
Figure 3.9: Three transcripts encoding the enzymes aldehyde dehydroge-nase, alcohol dehydrogenase and glycerol-1-phosphatase (IUBMB Enzyme code EC 1.2.1.3, EC 1.1.1.2 and 3.1.3.21 respectively), involved in glycerolipid metabolism are induced in wild-type C. glabrata cells exposed to 1 M sorbitol for 15 minutes. Differentially transcribed transcripts were identified by gene expression microarray analysis and mapped to C. glabrata pathways of metabolism in the KEGG database. Red boxes denote enzymes whose transcripts were more abundant after exposure to hyperosmotic stress imposed by addition of 1 M sorbitol to rich culture medium for 15 minutes.

3.3 Results 82
Transcriptional changes that may lead to the accumulation of alternative compatible solutes (e.g. induction of biosynthetic enzymes or transporters) adopted in other organisms such as inositol (664), arabitol, mannitol (319) and trehalose (459; 297; 296; 432) are not evident. Moreover, inositol monophosphatase INM2, which is involved in the biosynthesis of inositol, is repressed under hyperosmotic stress conditions. Taken together, these tran-script profiles provide indirect evidence for an increased metabolic flux through the glycerol biosynthetic pathway after hyperosmotic stress, similar to that reported previously S. cere-visiae (12; 392). Another key feature of the induced transcripts is the involvement of their gene products in energy production pathways such as glycolysis (pyruvate kinase CDC/9), the TCA cycle (pyruvate carboxylase PYC1, succinyl Co-A ligase LSC2, mitochondrial di-carboxylate carrier DIC1, pantothenate synthase PANG, riboflavin kinase FMN1), pentose phosphate pathway regulation (fatty acid dehydrogenase HFD1) and oxidative phospho-rylation (cytochrome b5 reductase MCR1). Transcript levels are also elevated for genes encoding proteins involved in the oxidative stress response (methionine-S-sulphoxide reduc-tase MXR1, monothiol glutaredoxin GRX7, NADPH-dependent aldo-keto reductase YPR1, and the cytosolic alcohol dehydrogenase ALD6). Transcripts annotated to protein degra-dation processes are induced in the wild-type strain after exposure to 1 M sorbitol for 15 minutes (Supplementary Data Table D.6). Within this set of transcripts there are those en-coding key proteins in ubiquitin conjugation (ubiquitin UBI4, the genes encoding ubiquitin protein ligase HRD1 and UFD2 a ubiquitin chain assembly factor and RODI a membrane protein that binds the ubiquitin ligase). These genes are co-induced with regulatory and catalytic subunits of the 26S proteasome, which degrades ubiquitinated substrates (RPN11, a metalloprotease subunit of the 19S regulatory particle; RPT3 an ATPase of the 19S reg-ulatory particle; RPN6, an essential, non-ATPase regulatory subunit of the 26S proteasome lid; PRES a2 subunit of the 20S proteasome, RPN3, an essential, non-ATPase regulatory subunit of the 26S proteasome lid), suggesting that hyperosmotic stress leads to an enhanced (relative to rich culture medium) turnover of proteins in the stressed wild-type.
Some transcripts related to protein trafficking are also induced by hyperosmotic stress (Supplementary Data Table D.7); gene products of which are involved in sorting proteins in the ER and Golgi (SEC8, SNX4) apparatus or in endocytosis and uptake of arginine (BTN2). Furthermore, gene products of several transcripts induced by hyperosmotic stress are located in the vesicles (VMA5, DAP2, PRB1, VPS31 and MVP/ ). Ubiquitination mediated by the induced ubiquitin-protein ligase HRD1 is also required for endoplasmic reticulum-associated degradation of misfolded proteins. Several other transcripts belonging to the heat-shock protein family, which are also involved in protecting misfolded proteins, are also induced in response to hyperosmotic stress (HSP78, HSP104, HRD1, KAR2 and APJ1)

Chapter 3. The Hogip and Stellp dependent transcriptional effects of hyperosmotic stress in Candida glabrata 83
Several transcriptional regulators are also induced, but none of the known downstream targets of S. cerevisiae Hog1p (Supplementary Data Table D.8) were shown to be induced in this study. However, Hog1p and Stellp dependent induction of the epigenetic modulator of transcription SIR2 - was demonstrated, as was a component of the Rpd3 histone deacetylase complex: PH023. Three basic leucine zipper transcription factor genes - FZF1, CIN5 and YAP1, and two zinc finger containing transcription factor genes SUT1 and SET5 were also induced. Hyperosmotic stress causes a largely Hog1p and Stel 1p independent induction of transcripts encoding polyamine (PUT4 ), glucose (HXT3) and multidrug transporter genes (YOR1) as well as the P-type ATPase sodium pump gene (ENA1), involved in Na+ and Li+ efflux to allow salt tolerance. Although no transcriptional changes were observed for the ac-tual signalling components of the HOG pathway, hyperosmotic stress did induce transcripts involved in modulation and feedback control of Hoglp phosphorylation state (PTC1). Other significantly induced phosphatase transcripts included the genes encoding both calcineurin subunits ( CMP2 and CNB1) which are required for activation of another stress response pathway mediated by the transcription factor Crzlp. Repressed transcripts largely belong to different biological processes (ribosome biogenesis, mRNA processing, inositol biosyn-thesis, GPI anchor biosynthesis, sterol synthesis, DNA repair/replication and other as yet uncharacterized functions), although some of those biological processes mentioned above are also represented in the repressed transcripts (cell wall maintenance and transcriptional regulation, endocytosis, ion transport). Transcripts of three enzymes involved in ergosterol biosynthesis are repressed upon exposure to hyperosmotic stress: squalene mono-oxygenase ERG1, C-5 sterol desaturase ERG3 and C-4 methyl sterol oxidase ERG25 (Figure 3.10). The hyperosmotic stress induced repression of both ERG1 and ERG2 is abolished in hog1 and sten cells, suggesting that the sorbitol dependent repression relies on a functional putative Shol signalling branch. In summary, transcription profiling of C. glabrata, and the summary of biological themes by inference from S. cerevisiae identified a largely divergent hyperosmotic stress response to that observed in S. cerevisiae. Although the induction of glycerol biosynthesis genes was observed, these analyses suggest that enhanced cellular traf-ficking, probably in delivery of remodelled cell wall components to the cell wall and targeted turnover of plasma membrane occurs in parallel with proteasomal degradation.

Non-merelonate pathway Texpenoid biosynthesis )
4
11.17.121 2-Phospho-4-(cridire 5,diphospho)-2-C-methyl-D-erythrhol
V aline, leonine axsl isoleuctne degradation Mevelonerte pathway
• K
epoxide Tocopherol
• O r 2.5.1i iHro 1:4147„Ytel- Omaprenyl- &phosphate
5.439.8
24-Methylene-cycloarte.nol Cycloartenol 0---I Din
(S)-Sguelene-23-epaxide 5.4.99.7
24,25-D ihydro- 0.4_1 13,74_1"• Lanmerci lenosteml
ECM
Cycloeucalenol
EMI 4 ..4-D imethyl cholesta-
1424-rtrienal
4-M ethylzymosterol-carboxylate
Obutosifaliol 'GYPSUM'
5814-Sterol
Sterol biosynthesis 4A-Dimethyl- 04_1 14-D emethyl- cholesta-13-en-313-o1 1 .3.1.72
Phytosterol Gelato:1ml 1.14.13.131-11.0 Calcitriol
BIOSYNTHESIS OF STEROIDS
1-1.15droxy-2-methyl- 2-C-Methyl-D-erythiltal 2-butenyi 4-diphosphate 2,4-cycloalphosphate
• MEM • =El • EZIECI •
pyrvvate (CInrolysis ) 1-Deozy-D-rylulose • 5-phosphate
E. • GLIMI • BM • - 2-C-M ethyl-D-erythntol 4-phosphate
4-(Cytidine 5,diphospho)-2-C-methyl-D-eryluitol
D-Glweraldehyde 3-phosphate
• MOM • EMI
25.120
Coenzyme Q
I 2.5.1.1
ct-Tocapherol (Vitamin E)
2.5.1.1
Geranyl-PP 2.5.1.10] Presgu.elarte-PP
ZIIIrpeat Mg.,= Heptaprenyl-diphosphate
Sguelere
4.1.1.331-04—I 2/.4.2 1-04-12.7.1.36I-0*—L1 1.154 1›.( Synthesis end deegradatiOn rievalonerte-5PF Meralonate-5P Marelonate 3-Hydroxy- of ketone bodies
3-rnethylglutaryl-CaA 1.0 Vitamin K Redacted Vitamin K
5.332 1-1.• ISOperderyl-PP
Farresyl-PP
I I
VO ( Carmen= biosynthesis )
t.---I 1.1.4.1 I
2-Methy1-6-phytylguinol
(Li le
2,3-D imethy1-5-phytylgutnol Y /El 1-1.0-1 G-TMTI-Ile b--.1 13.1.- Irtie Phytyl-PP
I all-trans- I 5-Tocopherol 13-Tocopiverca 1 Geranylgeranyl-PP I I , -Po( Porphyrin and chlorophyll metabolism )
itamln K cyck and Vitamin E biosynthesis
Rubber D iinethylell;11;
3-Ket0-4-Methyl- 9 zymosrexol
I inzml
9 * 24-Methylene- • 24-Ethy lidene-
4-M ethylsymosterol EZI
I 5.3.35 I
C?--I ERG6 1-0.9
rol Fecostero l
I 05021 *
lophenol
Zymose
EEGS • Episteml
piliop
7-A
henol
5 nrenesteral
* 0 CI:talent-7 ,24- diem-313-M =row 9 5-Dehydroep1ster01 0 5-D ehyiroavenastend
I DW7 I DTI5 I
♦ 7-0 °hydro- 7,24(28)-Ergoste- * 51 MP
24 -Methylene- + O desmosterol 9 trierca 5, cholesterol 9 Isafocamerol
I E1ZG5 I I DWI, I I I DWI I
0 Desmoslthol 57 2224(28)-Ergosta- + S Sitosterol
II C,..iemol letreenal 0 Campesterol
I I CY1.710A1 II I ao41 I II + I
S I's- - - *IC Bile acid biosynthesis) 1; Ergosterol v Blassirec•Wzi3)
000005000501
II' - - - -rtit.(Steroid hormone metabolism)
biosynthesis 0
Vitamin D2
• 4 e-Methylfecosteml
•
EEO Lathosteml
7-D ertrydracholesurol
tv
Cholesta-8-en-3D-01
V hamin D3
00100 214109 (c) Kartehisa Latoratones
3.3 Results 84
Figure 3.10: Three transcripts encoding enzymes involved in the ergosterol biosynthesis pathway, are repressed in wild-type C. glabrata cells exposed to 1 M sorbitol for 15 minutes. Repressed transcripts were identified by gene expres-sion microarray analysis and mapped to C. glabrata pathways of metabolism in the KEGG database. Red boxes denote enzymes whose transcripts where less abundant after exposure to hyperosmotic stress imposed by addition of 1 M sorbitol to rich culture medium for 15 minutes.

Chapter 3. The Hog1p and Stellp dependent transcriptional effects of hyperosmotic stress in Candida glabrata 85
3.3.3 Comparison of S. cerevisiae, C. albicans and C. glabrata transcriptional re-sponses to hyperosmotic stress
Having found phenotypic evidence of a different hyperosmotic stress sensing architecture, we hypothesized that the core transcriptional responses of hyperosmotic stress would be conserved, assuming that a) phosphorylation of Hog1p was necessary and sufficient for osmoadaptation and b) the regulatory components downstream of Hog1p are functionally conserved in C. glabrata. To assess the conservation of transcriptional changes induced by hyperosmotic stress between related species, I performed a simple comparison of induced and repressed transcription profiles between C. glabrata, S. cerevisiae and C. albicans. Several studies have investigated the affect of an osmotic upshift on the transcriptional behaviour of S. cerevisiae, using a range of solutes and exposure times. In an early study, Rep et al., (581) applied hyperosmotic stress by exposing S. cerevisiae cells to 0.7M NaC1, while more recently O'Rourke and Herskowitz (518) investigated the effects of both 0.5 M KC1 and 1 M sorbitol in wild-type and hogl strains (Supplementary Data Table 3.1). In a broader investigation into the environmental stress response of S. cerevisiae, Gasch et al., (230) applied hyperosmotic stress with 1 M sorbitol and recorded the transcriptional changes over a time-course.
To assess the degree of functional conservation between each available data set, I iden-tified homologous genes of C. glabrata and S. cerevisiae that showed conserved2 transcrip-tional responses between the two species after exposure to hyperosmotic stress. Comparison of induced C. glabrata transcripts identified in this study with the S. cerevisiae equivalents identified by Gasch et al., revealed little functional conservation between the two species. Curiously, even though the vast majority (93.5%) of hyperosmotic stress affected transcripts have a S. cerevisiae homologue, less than ten percent of the induced (Supplementary Data Table D.9) (7.10%) and repressed (5.12%) transcripts are similarly regulated in S. cerevisiae cells exposed to virtually identical hyperosmotic stress treatment (1 M sorbitol, 15 minutes). This discordance was also shown when data from this study were compared finding to a separate study, by O'Rourke and Herskowitz (518), who also used 1 M sorbitol as the source of hyperosmotic stress (Supplementary Data Table D.10), providing greater support for a different transcriptional response of C. glabrata to hyperosmotic stress. 3
The dissimilarity of transcriptional responses between C. glabrata and S. cerevisiae also
tin terms of the relative abundance. Conserved induction requires homologues to be more abundant after hyperosmotic stress treatment, whereas conserved repression requires homologues to be less abundant after hyperosmotic stress treatment
3The set induced of C. glabrata transcripts was also compared to Gasch and colleagues' 1 M sorbitol affected transcriptome at all available time-points, to see if a similar profile existed but with a delayed onset, however there was no concordance at all with the later time points.

3.3 Results 86
extends to studies that have used permeable solutes such as NaC1 and KC1. In the study by Rep et al., where 0.7M NaC1 was used as the hyperosmotic stress treatment, only 11.32% of hyperosmotic stress induced transcripts are functionally conserved with the equivalent tran-scripts identified in this study (Supplementary Data Table D.11). The sets of functionally conserved, repressed transcripts were similarly discordant: only three repressed transcripts are conserved, including: FUR1, a gene encoding a uracil phosphoribosyltransferase in-volved in the pyrimidine salvage pathway, and two genes encoding enzymes involved in the ergosterol biosynthesis pathway (ERGS encoding a putative C-5 sterol desaturase and ERG25 encoding C-4 methyl sterol oxidase). Furthermore, O'Rourke and Herskowitz (518) extended their microarray experiments to incorporate a treatment with 0.5 M KC1 as well as sorbitol discussed above. Again, transcriptional changes caused by 0.5 M KC1 treatment showed little similarity with those caused by 1 M sorbitol in C. glabrata (Supplementary Data Table D.12). Hyperosmotic stress induced transcriptional changes were also compared between a wild-type C. albicans data set (192) and this study. Enjalbert et al., identified 95 induced transcripts in a wild-type C. albicans strain subjected to an osmotic shift imposed by 0.3M NaC1 for 10 minutes. As is the case for S. cerevisae, there is very little similarity between the transcriptional response of C. albicans reported by Enjalbert et al., and the findings in this study.
Following the same protocol adopted for the comparison with S. cerevisiae data above, only 8 induced transcripts were shared in the C. albicans and C. glabrata hyperosmotic stress data sets. Functionally conserved transcripts were annotated as (S. cerevisiae/ C. albicans): NADPH-dependent medium chain alcohol dehydrogenase ADH6/113F20/04 ; pheromone-regulated protein PRM1011PF13607; mitochondrial NADH-cytochrome b5 reductase MCR1 and DNA damage-responsive protein DDR48. Search criteria were also relaxed to identify induced genes that belong to similar gene families or where annotated to different paralo-gous genes. For example, both data sets show elevated transcript levels for different isoforms of glycerol 3-phosphate dehydrogenase. The annotations of isoforms are slightly different between studies: GPD2 was found to be homologous to YDL022w in the C. albicans study but is annotated as YOL059W on microarrays used in this study. Similarly, hallmarks of HOG pathway activation such a putative C. albicans hexose transporter, HXT3 and a sodium transporter ENA22, each having different S. cerevisiae homologues to those in C. glabrata are also induced.
Nevertheless, even under these relaxed criteria, there are few similarities between the hyperosmotic stress affected transcriptome shown in this study and that of Enjalbert et al. A meta-analysis of microarray data from several independent studies, spanning different laboratories and sources of transcriptional stress, is problematic. Uncontrolled systematic differences between studies may equally well explain the observed discordance, casting some

Chapter 3. The Hogip and Stellp dependent transcriptional effects of hyperosmotic stress in Candida glabrata 87
doubt over whether the proposed divergence is really a biological phenomenon. Indeed, the highest degree of similarity between transcript profiles is observed in experiments performed in the same study with different sources of hyperosmotic stress (1 M sorbitol and 0.5 M KC1 in O'Rourke and Herskowitz), where perhaps systematic bias is best controlled be-tween data sets. Notwithstanding these technical differences, the proportion of functionally conserved transcripts is greater within S. cerevisiae data sets utilizing different sources of hyperosmotic stress than between S. cerevisiae and C. glabrata data sets using the same source of hyperosmotic stress (Table 3.1), which suggests that the transcriptional response to hyperosmotic stress may be due to fundamental differences in the biology of the species.

3.3 Results 88
Table 3.1: Conservation of induced transcripts between publicly available, hy-perosmotic stress affected, transcript profiles in S. cerevisiae, C. albicans and this study. Figures shown are percentage of induced transcripts functionally conserved between each study.
1 M Sorbitol* 0.7M NaC1 0.5 M KC1I 1 M Sorbitol' 0.3M NaCF 1 NI Sorbitol 1 M Sorbitoll 61.46 74.14 53.44 12.01 12.50 0.7M NaClt 22.00 - 39.60 21.90 4.20 9.40 0.5 M KC1I 33.52 48.35 - 40.11 1.10 10.44 1 M Sorbitol* 51.90 63.29 92.41 - 6.33 12.66 0.3M NaCl° 17.02 21.28 21.28 10.64 - 4.26 1 M Sorbitol 7.55 11.32 11.95 6.29 1.26 -
* 1 M Sorbitol, S. cerevisiae, Gasch et al., (230); t 0.7M NaC1, S. cerevisiae, Rep et al., (581); t 0.5 M KC1, S. cerevisiae, O'Rourke & Herskowitz (518); 1 M Sorbitol KC1, S. cerevisiae, O'Rourke
& Herskowitz (518); ° 0.3M NaC1, C. albicans, Enjalbert et al., (192); m 1 1\4 Sorbitol, C. glabrata,
This study.
3.3.4 The effect of HOG1 or STEll deletion on hyperosmotic stress induced tran-scriptional responses in C. glabrata
Transcript profiling of hog1 and stell under hyperosmotic stress conditions serves three purposes. First, Hog1p dependent hyperosmotic stress responses can be determined by identifying transcripts whose level is altered as a consequence of HOG1 deletion. Second, the analagous consequences of STE11 deletion can be determined, and compared with the above to identify which transcripts are affected by an individual gene deletion, either deletion, or both deletions. The dependency of transcript profiles can be interpreted as gene specific (i.e. wild-type response altered as a consequence of HOG1 deletion only (Figure 3.7 panel A), or STET1 deletion only (Figure 3.7 panel B)), branch specific (i.e. wild-type response altered by either HOG1 or STE11 deletion) or branch independent (wild-type response unaffected by HOG1 or STE11 deletion). I define a signalling branch as a hierarchical input of phosphorylation signal to Hog1p, in this sense Ste11p phosphorylates Pbs2p which in turn phosphorylates Hog1p; for a working hypothesis I assume that these proteins act only as a relay and that phosphorylation of Hog1p is sufficient to cause the downstream transcriptional events. Third, by observing transcriptional changes specific to stressed hogl
or stell strains, it is possible to identify points of hyperosmotic stress induced cross-talk between signalling pathways. In this section, I report the effects of HOGI or STE11 deletion on wild-type transcriptional response to hyperosmotic stress by comparing overlapping and disjoint transcript sets (Figure 3.11). Dealing with hyperosmotic stress induced transcripts

Chapter 3. The Hog1p and Stellp dependent transcriptional effects of hyperosmotic stress in Candida glabrata 89
first (Figure 3.11 b): Sixty two transcripts (36.70% of the induced set) show hyperosmotic stress dependent
induction that is abolished in either ste11 or hogl backgrounds. These transcripts (Supple-mentary Data Table D.13) are dependent on a functional putative signalling branch rather than on either of the two specific components investigated in this study (HOG1 or STE11). Notably, Shol branch dependent transcripts include those encoding key glycerol biosyn-thetic enzymes GPD2 and GCY2 and other metabolic process (alkaline ceramidase YPC1, /3 subunit of succinyl-CoA ligase LSC2, carbamoyl phosphate synthetase CPA1, serine hy-drolase FSH1, a pyruvate carboxylase isoenzyme PYC1, MCR1, malonyl-CoA:ACP trans-ferase MCT1 and mitochondrial dicarboxylate carrier involved in the TCA cycle D/C/ ). Some transcripts involved in the oxidative stress response are also dependent on the putative Shol branch (aldehyde dehydrogenase ALD6, glutaredoxin GRX7, methionine-S-sulfoxide reductase MXR1 and MBR1), although a key regulator of this response YAP1 depends specifically on STE11.
A small proportion of induced transcripts involved in protein degradation are Shol branch dependent (transcripts encode a subunit of a proteasomal metalloprotease RPN11, ubiquitin U1/3/4, mitochondrial inner membrane m-AAA protease YTA12), however these transcripts are largely uninvolved with the proteasome. Transcripts involved in vesicular fusion, transport and sorting (SEC18, VMA5, SWF1, GRH1, PKH2 and MVP/ ) are also induced by hyperosmotic stress. Similarly, there is small proportion of transcripts involved in cell wall maintenance that are generally dependent on the Shol branch (chitin synthase CHS1, a1,3-mannosyltransferase ALG3 and a cell wall beta-glucan assembly glycoprotein KRE9)
A functional Shol branch is also required for hyperosmotic stress dependent induction of transcripts involved with phosphate metabolism, whose protein products act either as direct regulators of MAPK phosphorylation status (e.g. transcripts encoding phosphotyrosine-specific protein phosphatase PTP3 and type 2C protein phosphatase PTC/ ) or in other metabolic processes such as glycogen metabolism (e.g. the regulatory subunit of protein phosphatase Glc7p GIP2) and sphingolipid metabolism, e.g. putative regulatory subunit of Nemlp-Spo7p phosphatase holoenzyme SPOT). A general disruption of Sho1 branch also affects the hyperosmotic stress dependent induction of several transcripts that share little or no sequence identity with S. cerevisiae (CAGLOE04554g) or are homologous to uncharacterized ORFS (YPR127W, YJL017W and YDR379W).
A surprisingly large proportion of transcripts are unaffected by deletion of HOG1 or STE11 (50, 29.58%) (Supplementary Data Table D.14), suggesting that a significant com-ponent of the transcriptional regulation is independent of the SHO1 signalling branch in C. glabrata. Protein products of these transcripts are involved in several biological pro-

3.3 Results 90
(a) hyperosmotic repressed (b) hyperosmotic induced
HOG .0-WT.0 STE.0-WT.0
2 0 0
5051
(c) Basal repressed (d) Basal induced
Figure 3.11: Venn diagrams of transient hyperosmotic stress repressed (a) and induced (b) genes in wild-type (WT.S - WT.U), hogl (HOG.S - HOG.U) and stell (STE.S - STE.U) cells (defined in Supplementary Data Table D.2). Induced and repressed genes were defined as having log2(Cy5TM) - log2(Cy3TM) >= 2.0 and <= -2.0 respectively and Benjamini-Hochberg corrected p values < 0.05. Basally repressed (c) and induced (d) genes are compared in hogl and stell cells grown in rich culture medium (HOG.0 - WT.U) and (STE.0 - WT.U) respectively. For description of contrasts: STE.0 - WT.0 etc see Supplementary Data Table D.2.

Chapter 3. The Hog1p and Stellp dependent transcriptional effects of hyperosmotic stress in Candida glabrata 91
cess categories including stress protection such as the heat-shock proteins (HSP78, HSP42, HSP104 and APJ1), as well as transporters of inorganic ions (ENA1) and organic molecules (HXTS, YOR1, TPO4) and regulators of glucose availability (SKS1, NRG2 and RGS2). The most induced transcript, BTN2 is also induced in a hogl or ste11 background along with other transcripts encoding proteins involved with vesicular trafficking ( VPS/2 and
SBE22). Transcripts belonging to the cell integrity pathway (the putative cell integrity receptor MID2, and the calcineurin subunit encoding CMP2 and CPA2) are also indepen-
dent of HOG1 and STE11 deletion. Twenty one transcripts (12.42% of the induced set) are specifically HOG1-dependent. These transcripts (Supplementary Data Table D.15) are induced in both wild-type and ste11, but not hogl cells and largely encode proteins in-volved with vesicular sorting and transport (HSE1, YPT52) or protein degradation either in the vesicle (PRB/ ) or in the 26S proteasome (RPT3, PRE8, HRD1). However, there are several poorly characterized induced transcripts that are specifically HOG1-dependent (AIM7, AIMS, YMR253C, YLR392C).
Thirty six transcripts (21.30% of the induced set) are specifically STE11-dependent. These transcripts (Supplementary Data Table D.16) are induced in both wild-type and hogl cells, but not stell cells and are largely uncharacterized (YIL055C, YLR414C, YLR194C, YOL057W, YN:305C, YGR205W, YDL190C, RCN2 and YJL171C), although several tran-scripts are involved in cell wall maintenance (KTR6, GSC2, SLT2, MTL1 and MN/V4), transcription regulation (SUT1, TFG1, YAP1, BDF1, SRB6) and vesicular sorting (APM1, SNX4 and KAR2).The repressed transcripts show less dependency on a specific component of the SHO1 signalling branch than induced transcripts (Figure 3.11 a). Twenty nine tran-scripts (74.30% of the repressed set) show hyperosmotic stress dependent repression that is abolished in both ste11 and hogl backgrounds (Table D.17). HOG1 and STE11 inde-pendent repressed transcripts include two that encode key ergosterol biosynthesis enzymes (squalene monooxygnease ERG1 and ERG25 C4 methyl sterol oxidase). Transcripts are also involved in mRNA processing (HSH49, ALB1 and UTPIS) transcriptional regulation (POG1, MOT1 and TRA1).
Seven transcripts (18.00% of the repressed set) are specifically HOG1-dependent. These transcripts (Supplementary Data Table D.18) are repressed in both wild-type and ste11, but not hogl cells and encode proteins involved in axial bud-site selection Bud4p and iron acquisition Ftrlp respectively.Two transcripts (5.20% of the repressed set) are specifically Stel1p-dependent. These transcripts (Supplementary Data Table D.19) are repressed in both wild-type and hogl cells, but not stell cells and includes the gene encoding the C-5 sterol desaturase ERGS of the ergosterol biosynthetic pathway. In contrast to the hyperosmotic stress induced transcripts, only a single transcript (CAGLOI04026g/PBP4 ) (2.50% of the repressed set) is unaffected by deletion of HOG1 or STE11. Transcript

3.3 Results 92
repression therefore relies almost entirely on the Shol branch, in contrast to the induced genes which has a larger Sho1 branch independent component.
A small number of transcripts are basally regulated by HOG1 or STE11. These tran-scripts were identified by comparing wild-type and hogl or ste11 cells grown in YEPD with-out sorbitol. Two transcripts, encoding a mitochondrial protein CAGLOG07931g/MRPS/2 and a putative glucokinase, CAGLOF00605g/ GLK1, are more abundant in hogl cells than wild-type cells which indicates that HOG1 is required for basal repression of these genes. Three transcripts are less abundant in hogl cells than wild-type cells and therefore require HOG1 for basal expression in rich culture medium. These transcripts, encode a putative hexose transporter CAGLOA01804g/HXT3, a lectin-like protein CAGL0005665g/FL 0/0 and an uncharacterized ORF CAGLOA02277g/YCL049c.
The effects of HOG1 deletion on hyperosmotic stress induced transcriptional changes has also been investigated in S. cerevisiae (581). Publicly available data from the study by Rep et al., were mapped to the C. glabrata annotation (Supplementary Data Table D.20) and compared with the HOG/ -dependent, STE11-dependent and SHO/ -branch dependent transcripts identified in this study. No homologous, HOG1-dependent transcripts were functionally conserved between this study and that of Rep et al.,. This is largely due to differences in the wild-type transcriptional responses; twenty one (75%) HOG1-dependent transcripts identifed by Rep et al., are not induced in C. glabrata cells exposed to hyperos-motic stress. In this case, a relaxation of search criteria to include HOG/ -dependent mem-bers of conserved gene families revealed some degree of conservation. The HOG1-dependent induction of mitochondrial aldehyde dehydrogenase ALD4 in stressed S. cerevisiae cells is not observed in stressed C. glabrata cells, however a Shot branch-dependent, related cytoso-lic aldehyde dehydrogenease homologue ALD6 is evident. Similarly, uncharacterized genes belonging to the AIM (Altered Inheritance rate of Mitochondria) family are Shol branch dependent (AIM9) or STE11 dependent in this study, and HOG1 dependent in S. cere-visiae. Surprisingly, four Hoglp-dependent, stress induced S. cerevisiae transcripts (ENA1, HSP104, PRM10, PHM8) were found to be induced by hyperosmotic stress in C. glabrata, but independent of HOG1 and STE11 deletion, suggesting altered regulatory control over two genes encoding genes expressed in the response to pheromone (PR/1410 and PHM8).
3.3.5 The hogl and stel 1 specific transcriptional responses to hyperosmotic shock: evidence for transcriptional cross-talk between putative C. glabrata MAPK cascades
Null mutant strains, disrupted in either the HOG1 or STE11 loci, provide an insight into how C. glabrata maintains coherence in hyperosmotic stress signalling. Hyperosmotic stress

Chapter 3. The Hog1p and Stellp dependent transcriptional effects of hyperosmotic stress in Candida glabrata 93
induced transcriptional responses, specific to each null mutant strain were investigated in the context of the orthologous MAPKKK MAPK architecture of S. cerevisiae. Remarkably, several components of the adjacent signalling cascades, spanning four levels of the hierarchy are only induced in the hogl strain under hyperosmotic stress (Figure 3.12).
Remarkably, several components of these signalling cascades, spanning four levels of the hierarchy, are aberrantly induced or repressed as a consequence of HOG1 deletion (Figure 3.12). The hogl specific osmostress-dependent induction of the gene encoding the tran-scription factor STE12 is intriguing because it regulates several downstream processes in S. cerevisiae, including the cell wall integrity pathway, the morphological switching pathway and the mating response pathway, and it's many functional guises, two of which - mor-phological switching and mating - are observed rarely or have never been observed in C. glabrata, make the interpretation of these findings problematic. Given that C. glabrata is currently thought to be an obligate haploid (754; 487), devoid of any observed sexual cycle, it is perplexing to not only see aberrant induction of a downstream transcriptional acti-vator of (amongst others) the mating response pathway, but to see an apparent feedback to a pheromone signalling sensor gene (STE2 (100)) in the plasma membrane, and regula-tory components upstream of the MAPKKK MAPK architecture. Furthermore, Far7p a protein involved in G1 cell cycle arrest in response to pheromone (353) is also aberrantly induced in hogl cells exposed to hyperosmotic stress in addition to other genes involved with the cell cycle progression (NISI encodes a protein localized to the bud neck at G2/M phase (320), WTM1 encodes a transcriptional modulator involved in regulation of meiosis (535), CLN3 a G1 cyclin involved in cell cycle progression and SPS1 a putative protein serine/threonine kinase expressed at the end of meiosis and localized to the prospore mem-brane of S. cerevisiae). S. cerevisiae Stel2p also activates genes involved in pseudohyphal/ invasive growth pathways of S. cerevisiae. Although, described as a third osmosensor in S. cerevisiae, Msb2p was also identified as an important regulator of filamentous growth by promoting differential activation of Ksslp - the MAPK for the filamentous growth pathway. Domain deletion studies show that Msb2 (153)plays an inhibitory role in mucin dependent MAPK pathway activation. If transcriptional induction results in increased amount of func-tional Msb2p in the plasma membrane and a greater inhibitory signal, then hogl cells will suppress the filamentous growth pathway when exposed to hyperosmotic stress.
In addition to incoherent signalling through, cross-talk is observed in another cascade in which Stel2p is not a downstream effector - the cell wall construction pathway. Although two genes, encoding important components of this pathway: the transmembrane spanning MUM and the MAPK, SLT2 are induced in stressed wild-type cells, the downstream tran-scriptional regulator RLM1 is only induced in stressed hogl cells, suggesting that Hog1p is required for suppression of this transcription factor in stressed wild-type cells. Other down-

3.3 Results 94
stream transcripts, relating to activation of this pathway are also only induced in stressed hog1 cells including cell-wall mannoprotein CWP1, chitin trans-glycosidase CRH1, DIE2 and CAX4. In addition to STE12, several other transcriptional regulators are specifically in-duced in hog1 cells exposed to hyperosmotic stress, including TAF3, VHR1, YAP7, SOK2, WTM1 and GCR1. These findings suggest that, like in S. cerevisiae MAPK pathways, Hogip is required to maintain coherent signalling.
3.3.6 Identification of a hypothetical hyperosmotic stress regulon in C. glabrata
In order to estimate the regulatory processes controlling the gene expression patterns ob-served after exposing C. glabrata to hyperosmotic stress, I used the YEASTRACT plat-form (686; 477) to construct a hypothetical osmostress regulon network (HORN) by infer-ring regulatory interactions from the model organism S. cerevisiae. Specifically, induced and repressed transcripts (identified using methods described above) were pooled to cre-ate a "regulated" set whose transcriptional control we sought to understand. For each C. glabrata gene in the regulated set, an S. cerevisiae orthologue was identified (using the IN-PARANOID determined orthologues available at Genolevures (630)); YEASTRACT was then used to identify all known conserved transcriptional regulators of this osmotic stress regulated gene. Assuming that these interactions are conserved between the two species, the regulatory interactions are represented by a directed edge from the transcription factor to the regulatory target. When performed for all genes, this process creates an orthologous network of transcriptional regulators and their interactions, with target genes having "in" edges from one or more transcription factors and transcription factors having "out" edges to one or more targets. Although the assumption of conserved regulatory interactions is quite strong, the analysis was performed only to generate hypotheses about regulation that can be experimentally verified or refuted in later studies.
I introduce some pertinent definitions prior to presenting the network analyses: a simple graph is a collection of items called nodes or vertices and their relations, called links or edges4 . I represent a simple graph as g (V, 5) where V = {vi, v2, , vM} is the set of M nodes and E = {ei, e2, , eEl is the set of E edges. Two nodes of a graph are adjacent if there is an edge between them. The entire graph can be represented by an adjacency matrix G E RM"71, containing entries of the form G (i, j), representing an edge between two nodes vi and 'Undirected graphs exclusively contain edges with no orientation and therefore have symmetric adjacency matrices. In contrast, directed graphs - also known as digraphs - have directed edges or arcs, a = (x, y) from the tail x to the head y and can have asymmetric adjacency matrices. These graphs can be used to depict influence
4for clarity in this section, we will consistently define graphs with nodes connected by edges

Chapter 3. The Hog1p and Stellp dependent transcriptional effects of hyperosmotic stress in Candida glabrata 95
between variables, for example in a Bayesian network or to indicate physical constraints in real paths (over Konigsburg bridges for example). The degree of a node in a network is simply the number of edges it has to other nodes; betweenness centrality is a measure of a node's importance in a network's structure. Let o-st = ats denote the number of shortest paths from s E V to t E V where mss = 1. Let crst]( ,) denote the number of shortest paths from s to t that some v E V lies on, then a commonly used betweenness centrality CB (v) measure is:
C B (V) E sty (3.2) Est V
and and is computed using an implementation of the algorithm developed by Brandes in networkx and python (263; 717). In essence, high centrality scores indicate that a node can reach others on relatively short paths, or that that the node lies on a high proportion of shortest paths connecting other nodes - i.e. a node of high traffic. Drawing comparisons with more familiar networks, such as the London Underground rail system, Baker Street is an example of a station that has high betweenness centrality (Supplementary Data Table D.21); if a passenger were to travel between each pair of stations in the network - taking the shortest route possible then Baker Street lies on a higher proportion of those shortest routes than any other stations.
The resulting HORN consists of 282 nodes and 863 edges in two connected components (overview in Figure 3.13), the largest of which comprises > 99% of the nodes and edges (280 nodes and 862 edges), the smallest of which is simply a directed edge from the origin replication complex Sumlp to the induced PFS2 gene, an integral subunit of the pre-mRNA cleavage and polyadenylation factor (CPF) complex (an overview of this regulon is shown in Figure 3.13). The largest connected component comprises transcription factors (large nodes in Figure 3.13 that are both transcriptionally repressed (coloured blue in Figure 3.13), induced (coloured yellow in Figure 3.13) and unchanged (coloured grey in Figure 3.13) by hyperosmotic stress; target genes are represented by smaller rectangles. The majority of nodes in the HORN are targets (167 nodes, 59%) in comparison to transcription factors (115 nodes, 41%), however the HORN does capture the majority of nodes that are annotated as transcription regulators in YEASTRACT (115 out of 183, 63%), which suggests that the structure might not be specific to the hyperosmotic stress regulated genes. To address this question of specificity, although not performed here, future work will require a resampling analysis to assess if the appearance of these transcription factors is specifically dependent
5Although perhaps oxymoronic, a rational terrorist might select this tube station as a potential target to maximize disruption with a single terrorist strike

3.3 Results 96
on the choice of input data (i.e. the hyperosmotic stress signal) rather than the size of the regulated gene set used to construct the HORN. Although the specificity of the presented HORN may be in question, I investigated the phenotypic enrichment of the HORN by taking 10,000 random 282 node samples from the genome and calculating the number of nodes that were annotated with hyperosmotic stress growth decreased, abnormal or absent in SGD. The permuted null distribution had an average of < 5 sensitive phenotypes, whereas the observed number of hyperosmotic stress sensitive nodes in the HORN (12 denoted by red vertical line in INSET B of Figure 3.13) was significantly greater than expected in a random sample. This evidence suggests that the HORN controls the expression of genes of significant number of genes that are required for the adaptation to hyperosmotic stress than would be expected by chance selection of genes from the genome. On a cautionary note, this assertion is based on a rather weak inductive argument (because phenotypic enrichment is also calculated on the basis of homology with S. cerevisiae).
While the HORN approach described above makes strong assumptions about the con-servation of transcription factor binding sites several approaches have been developed to identify statistically significant motifs within nucleotide sequences. I adopted one such approach, a Gibbs motif recursive sampling strategy, to identify candidate transcription factor binding motifs in the upstream promoter regions (-1000 to -1 bp before start codon, downloaded from Genolevures database (630)) of the regulated set described above. This sampling strategy used the Eukaryotic default settings and took 72 hours to complete on a Macbook Pro laptop computer. The resulting position weight matrices were then used to create a sequence logo with the graphical seqLogo toolkit in the R statistical framework (Inset of Figure 3.13). This process identifies in a 16 nucleotide motif (GAGATAAACCCGCTCT),
containing two sub-motifs (GAGAT and GCTCT) with higher information content, separated by a ] portion (AAACCC). The entire motif was used to search YEASTRACT transcription factor binding site motifs, allowing 2 substitutions and aligned with S. cerevisiae consensus sequences of Gsmlp, Yrrlp, Yaplp, Xbplp, Uga3p, Tec1p, Stp2p, Stplp, Stb5p, Rtglp, Rtg3p, Rgtlp, Raplp, Pho4p, Nrg1p, Msn2p, Msn4p, Mot3p, Mcmlp, Haclp, Gln3p, Ace2p, Swi5p, Aft1p, Aship, Bas1p, Cat8p and Sip4p.
There is also some information in the set of transcription factors that are not incorpo-rated into the HORN. For example, GO term searches of the unrepresented transcriptional regulators factors reveals processes that may not be regulated during hyperosmotic stress in-cluding (not exhaustive list): protein deubiquitination and urmylation (OTU1, ELP6), neg-ative regulation of transcription by glucose (M/G2), nitrogen utilization ( UGA3), positive regulation of RNA polymerase II by pheromones (KAN), positive regulation of gluconeo-genesis (S1134 and CATS), purine base biosynthesis (BAS1), cellular zinc ion homeostasis (ZAP1) and reciprocal meiotic recombination (BAS/ ). One obvious limitation of this ap-

Chapter 3. The Hog1p and Stellp dependent transcriptional effects of hyperosmotic stress in Candida glabrata 97'
proach is the omission of transcripts lacking S. cerevisiae orthologues, i.e the C. glabrata specific genes that may confer a different hyperosmotic stress phenotype to the model or-ganism. Although using a model organism can be useful in studying conserved regulatory processes, there is a stark requirement for experimenal tools and statistical inference meth-ods to elucidate the nature of these regulatory process directly in the pathogenic fungi. Such a top-down approach to modelling is still in it infancy, but might yet be truly informative in understanding the molecular basis of environmental stress adaptation in non-model or-ganisms. Moreover, this approach is severely limited by the lack of dynamic transcription measurements.
In order to assess how the HORN structure is associated with hyperosmotic stress de-pendent transcription, I adopted two separate approaches to investigate the hypothetical network organization. The first method involved computing simple summary statistics of network structure (node degree and node betweenness centrality - Figure 3.14) and assess-ing the relationship between these measures and gene expression. The HORN contains very many nodes that are sparsely connected (i.e. with degree below the mean degree depicted by the red line in the degree histogram), and very few nodes that have many connections (bars to the far right of the distribution). For the sake of discussion, I defined a hub as a node with degree k > 20, and subsequently identified 13 hubs in the HORN: Sok2p (k = 45), Rap1p (k = 45), Fhllp (k = 45), Teel') (k = 34), Stel2p (k = 32), Uthlp (k = 27), Cin5p (k = 27), Roxlp, Phdlp (k = 25), ENA1 (k = 21) and Ino4p (k = 20). Interestingly, all of these hubs apart from ENA1 are transcription regulators; ENA1 is annotated as a P-type ATPase sodium pump, involved in sodium and lithium efflux, which is apparently under the regulatory influence of many transcription factors.
The connectivity of HORN nodes was then investigated in the context of the observed gene expression. I defined a simple expression of regulator activity, Ant and Ar,.. This mea-sure is simply the number of target genes that the regulator induces or represses respectively, expressed as a proportion of the total number of target genes k. These expressions pro-vide a measure of activity for each of the HORN transcriptional regulators and can be used to rank transcription factors implicated in hyperosmotic stress adaptation, with the top ranking regulators having the highest proportion of their targets regulated. Notably, transcription factors ranked in this manner do not necessarily regulate the largest number of targets in the regulated set. The HORN can also be classified in terms of An, and k, in order to identify groups of functionally related transcriptional regulators. A k-means clustering method with 4 centroids was adopted to do so (Figure 3.15), and revealed four distinct groups. The first group (red points and red shaded text box Figure 3.15) consists of non-hubs (k < 20) with the highest target activity Art (> 80%). Remarkably this highly active group contains four transcription factors that have been implicated in S. cerevisiae

3.3 Results 98
hyperosmotic stress regulation: 1VIsn2p, Msn4p, Skn7p, Sko1p in addition to Stel2p which has a role in the mating, cell wall maintenance and morphological switch pathways, which suggests that they may also be active in hyperosmotic stress regulation in C. glabrata. The subnetwork of these transcription factors and their immediate neighbour nodes is shown in Figure 3.16. The second group (green points and green shaded text box Figure 3.15) consists of HORN hubs (k > 20) with moderate target activity Ari (> 80%); these tran-scriptional regulators have the highest connectivity (Roxlp, Rap1p, Mcm1p, Ino4p, Cin5p, Tec1p, Sockp, Fhllp, Phdlp and Flo8p) within the HORN but activate or repress a lower proportion of their putative targets (Figure 3.18). Although these hubs do not activate or repress every target, they do regulate larger and distinct proportions of the overall reg-ulated set than those identified in the red group e.g. Sok2p (20%), Rap1p (18%), Fhllp (16%), Teclp (16%), Stel2p (15%); moreover when all pairings of transcription factors were considered, the combination of two transcription factors regulating the largest proportion (51 genes, 30%) of the regulated set was found to be Sok2p-Rap1p (Figure 3.20).
Although present in the HORN, IVIsn2p and Msn4p are not themselves transcription-ally induced and have only a few direct targets which include the induced genes HSP78, ROD1, BTN2, HSP104, LST8, CMP2, IPT1, APJ1, GSC2, PIH1 and HSP42, GO term analysis of which reveals that this set is enriched in the protein unfolding response genes (corrected p value < 0.05) and also in the membrane fraction (corrected p value < 0.05) of the GO component ontology terms(IPT1, LST8 and GSC2). Other direct targets include the putative transcriptional regulators Roxlp and Yap1p. Motif searching (implemented in cytoscape (624; 137) NetMatch based on the Alon motif finding algorithm (629; 464)) reveals that Msn4p and Msn2p both have feedforward loop structures (a potential regula-tory motif discussed in (629; 464)) with YAP1 and DDR48. By identifying the subgraph of Msn2p, Msn4p, Skn7p and Sko1p with the k = 1 neighbours it was also possible to identify other transcription factors that are local to this subnetwork and may co-operate with these putative hyperosmotic stress regulators to drive the observed transcriptional changes. For example, Skn7p appears to direct the transcription of NRG2 which encodes a transcription factor that mediates glucose repression and negatively regulates filamentous growth, and also appears to direct the transcription of ROX1 and YAP1 in co-operation with Msn2p and Msn4p. The induced direct targets of Skn7p that are not transcription factors in-clude genes encoding a polyamine transporter ( TP04), chitin synthase I ( CHS/ ) and a type 2C protein phosphatase (PTC/ ). Surprisingly, SUR7, a gene required for growth in hyperosmotic stress conditions in S. cerevisiae is repressed in this study, apparently due to the influence of Skn7p. Transcriptional regulators recruited by Skolp, Skn7p, Msn2p and Msn4p are themselves regulators of the oxidative stress response (Yaplp and Roxlp).
In order to predict hypothetical regulators of glycerol biosynthesis, I searched the HORN

Chapter 3. The Hog1p and Stellp dependent transcriptional effects of hyperosmotic stress in Candida glabrata 99
for all transcriptional regulators with an edge incident upon induced transcripts having a putative role in glycerol biosynthesis (GPD2 - Figure 3.21, GCY1 - Figure 3.22 and ALD6 - Figure 3.23), all of which are Shol branch-dependent (as depicted by the hexagonal node shape in each figure). Although several transcription factors had edges to GPD2 (Tec1p, Hcm1p, Ino2p, Rap1p, Ino4p, Cin5p), only one (Cin5p) was induced after exposure to hy-perosmotic stress, and was induced independently of the extant signalling branch. This transcriptional regulator also has an edge to ALD6 (Figure 3.24) and several other tran-scripts, the majority of which are Shol branch dependent. Cin5p seems to be an obvious candidate regulator of glycerol biosynthesis, although by an inductive argument one might expect Cin5p transcription to also be Shol branch dependent if there is a direct regulatory role. However, this interpretation is confounded by the "hidden" activation profile of this protein. For example, it is conceivable that the transcriptional induction is not equivalent to activation, and some other Shol branch dependent activation event occurs that may account for the Shol branch dependent profile of its targets. Perhaps more important, in acknowledgement of the nature of complex networks, is the potential co-operative role with other transcriptional regulators and their targets (e.g. ALD6 has several candidate regulators: Pdr3p, Tec1p, Gcn4p, Flo8p, Pdrlp, Stel2p, Stb5p) in the network that are not tractable with the simple analyses adopted here, but may warrant investigation with approaches such as boolean networks. However, the observed Sho1 branch dependent in-duction of these glycerol biosynthetic enzyme does suggest that glycerol production under hyperosmotic stress conditions is regulated by the extant SAPK architecture in C. glabrata. Recent work conducted in our laboratory by Emily Cook (Personal Communication) shows that glycerol accumulation is independent of Hog1p in C. glabrata but Hog1p is required for growth in these conditions. Taken together with our findings of Shol branch dependent induction of glyerol biosynthetic enzymes, it appears that induction of these enzymes is not required for proliferation under hyperosmotic stress conditions, and there may exist other Hog1p dependent mechanisms that are required for proliferation in hyperosmotic stress. Glycerol accumulation is also the net effect of glycerol turnover and export, and the regu-lation of C. glabrata Fps1p may be an important determinant of glycerol accumulation in hyperosmotic stress conditions. One speculation is that Hog1p is involved in the regulation of Fps1p closure or endocytosis to reduce glycerol efflux in C. glabrata which is under pos-itive selection in comparison to sensu stricto species (Supplementary Data Table D.22 and Supplementary Figure D.3).
I used the same HORN approach to make predictions regarding the nature of C. glabrata stel2 null strain phenotypes observed in previous work conducted in our laboratory (Figure 3.25). A total of five (SKS1, UTH1, DDR48, BDF1 and SUT1) induced Stel2p targets are required for growth in S. cerevisiae cells exposed to hyperosmotic stress (depicted by red

3.3 Results 100
borders of nodes in Figure 3.25). The sensitivity of C. glabrata ste12 strains to sorbitol (1.5 M) may possibly be due to the regulatory effects of Stel2p on these transcripts however, these transcripts are also apparently regulated by other transcription factors in the HORN so future experimentation will be required to assess the regulatory influence on these genes whose S. cerevisiae required for growth. Moreover analysis methods that may take into account the complexity of the interactions (such as boolean networks (753; 743; 37; 708; 167) or hidden variable dynamical modelling (51; 52)) may be more complex approaches that are still easy to implement, providing time series experiments are conducted.
In general, there is a strong positive correlation (r = 0.89 between k and CB(v) such that hubs are also highly central to the HORN. However, there are some hubs that deviate from this correlation with some having higher centrality (above the dotted red line in Figure 3.14 C: Ino4p, ENA1, UTH1, Rap1p and Sok2p) and some that have lower than average centrality (below the dotted red line in Figure 3.14 C: Cin5p, Flo8p, Roxip, Fhllp, Tec1p, Stel2p and Phdlp). The more highly connected set consists exclusively of transcriptional regulators, whereas the more highly central set consists of transcriptional regulators and two target genes ENA1 and UTR1 that are both required for wild-type growth in S. cerevisiae cells subjected to hyperosmotic stress. In this study, the transcription of these highly central transcripts was independent of Hog1p and Ste11p and potentially controlled by several regulators.
In a second approach, I used a k-clique percolation search strategy to identify communi-ties in the HORN based purely on network topology, the aim being to find how transcription factors are organized in terms of their topological relationship to each other. In this sense, while the investigation described above deals with local network structure, the following approach addresses the topological structure at a higher level of organization. While there is no accepted mathematical definition of a "community", one interpretation comes from an arguably intuitive understanding of social community structure. A community therefore consists of entities that are more closely related to each other than to other members of the entire society (encyclopaedically reviewed by Porter (549), Newman (495) and also by Fortunato (209)). In terms of graph structure, a community finding algorithm seeks sub-networks that contain a greater proportion of edges between each other than to the wider network, and provides an unbiased strategy to identify modularity in the network based on topology alone. Most algorithms find partitions of the graph and therefore do not allow overlapping communities, which confines nodes to a single context. By analogy, standard partition methods may identify a community of British National Party (BNP) voters, but would not also permit simultaneous membership with a West Ham United Football Club (WHUFC) supporters community, whereas k-clique percolation may uncover membership of the BNP, WHUFC and Fathers 4 Justice. This strategy identified three overlapping com-

Chapter 3. The Hog1p and Ste11p dependent transcriptional effects of hyperosmotic stress in Candida glabrata 101
munities (Figure 3.13) that regulate 50% of the targets in the HORN. The largest identified community (community 2) has 15 members including several putative transcription factors (Phdlp, Teclp, Yap6p, Rim101p, Skolp, Pdrlp, Cin5p and Yap7p) and overlaps with a smaller community (community 3) that contains the putative transcription factors Roxlp, Msn2p and Skn7p. The points of overlap are not generally transcriptional regulators (apart form Skn7p): overlap b (Figure 3.13) for example consists of DDR48 and YPRO15C which appear to be under the complex regulation of two communities of transcriptional regulators.
In summary these approaches have assessed the relationship between local network struc-ture (degree and centrality measures) and hyperosmotic stress dependent transcription and the hierarchical organisation of these nodes (k-clique percolation). The local network statis-tics have been used to classify transcriptional regulators in terms of their activity and connectivity. I identified one class of transcription factors with high activity and low con-nectivity that contained several putative regulators of hyperosmotic stress adaptation in S. cerevisiae. In comparison to the HORN hubs, these regulators are incident on a smaller proportion of the whole regulated set. Hubs with moderate activity (e.g Mcmlp, Rap1p, Roxlp, Ino4p, Cin5p, Tec1p. Sok2p, Fhllp, PhDlp and Flo8p) are likely to be important regulators of hyperosmotic stress response in C. glabrata, Moreover, k-clique percolation identified communities of transcription regulators that show similarity in function. For example community 3 consists of transcriptional regulators (Roxlp, Hap1p, Msn2p and Skn7p) that regulate gene expression in response to hypoxia and oxidative stress whereas community 2 consists of transcriptional regulators involved in filamentation or morpholog-ical changes (Phdlp,Stb6p, Teclp and Sok2p). Although the observations are based on hypothetical network structure, the value of this approach is in the contextualization of gene expression in the network structure, and as such is merely a visualization tool rather than a true systems biology approach. Moreover, by only focusing on transcription data, to say this work is an integrative genomics approach might also be stretching the point. How-ever, such contextualization does permit hypothesis development for future experimental work that might not be immediately obvious from looking at gene lists alone.
3.3.7 Identification of hyperosmotic stress sensitive C. glabrata transcription factor null mutant strains
We screened 186 null mutants, with disrupted putative transcription factor ORFs to identify transcription factors that are required for growth in hyperosmotic stress (1 M sorbitol) using liquid growth and solid agar assays (Figures D.1 and D.2, performed by Lauren Ames). We reasoned that, if the downstream targets of Hog1p are conserved and required for growth in hyperosmotic stress then strains deficient in these orthologous genes would show impaired

3.3 Results 102
growth in 1 M sorbitol and 0.5 M KC1 which are similarly hypertonic (Emily Cook, Per-sonal Communication). The so-called C. glabrata Transcription Factor Knockout Library (CGTFKOLib) employed here comprises 186 strains (created by Biao Ma), disrupted in ORFs encode transcription regulators in S. cerevisiae. Growth was measured using solid agar screens (performed by Lauren Ames, Figures D.1 and D.2) assays (see materials and methods section).
The strains showing the highest degree of growth impairment were: pho4 (CAGLOD05170g) a bHLH transcription factor involved in the transcriptional control of the phosphatase reg-ulon (512); not3 (CAGLOL01001g) a subunit of the CCR4-NOT complex involved in tran-scription initiation, elongation and in mRNA degradation (417); srb8 (CAGLOE00627g) a subunit of the RNA polymerase II mediator involved in glucose repression (44); msn4 (CAGLOM13189g) transcriptional activator related to Msn2p; activated in stress conditions in both S. cerevisiae and C. glabrata (589; 615). The gain (CAGLOF00803g) subunit of the RNA polymerase II mediator complex; associates with core polymerase subunits to form the RNA polymerase II holoenzyme; affects transcription by acting as target of acti-vators and repressors(472); hstl (CAGL0005357g) a NAD(+)-dependent histone deacety-lase required for ORC-dependent silencing and mitotic repression; yox1 (CAGLOG07249g),
taf.14 (CAGLOIVI02739g) a major component in transcriptional regulation complexes TFIID, TFIIF, IN080, SWI/SNF, and NuA3 (694; 269; 400; 268; 489; 540; 399); gcr2 (CAGLOG05467g) a transcriptional activator of genes involved in glycolysis; and arg81 (CAGLOH06875g) a zinc-finger transcription factor of the Zn(2)-Cys(6) binuclear cluster domain type, involved in the regulation of arginine-responsive genes.

Chapter 3. The Hog1p and Stellp dependent transcriptional effects of hyperosmotic stress in Candida glabrata 103
• • • Ste -12 Stel2 Several Factors Rim, ?
N......) Ste-12
Mating Morphological Cell Wall Osmoregulation Cell Wall Sporulation Response Switch Integrity Construction Response
Figure 3.12: Architecture of MAPK cascades of S. cerevisiae, incuding the HOG osmoadaptation cascade annotated with evidence of transcriptional cross-talk in conserved C. glabrata transcripts. Adapted from (292). Orange circles denote putative osmosensors embedded in the plasma membrane, connected to the upstream G-protein (Stel8p, Ste4p, Ras2p, Rho1p, Cdc24p, Cdc42p, Gpalp, Rom2p) or phosphorelay (Ypdlp, Ssklp) control components (black circles). Kinases (either p21 activated kinases Ste2Op or serine/threonine kinases Pkclp, Spslp), depicted by grey circles, lie upstream of the core MAP kinase architecure (MAPKKK (Stel 1p, Ssk2p, Ssk22p, Bcklp) —> MAPKK (Ste7p, Pbs2p, Mkklp, 1V1kk2p) —> MAPK (Fus3p, Ksslp, Hoglp, Slt2p, Smklp)) whose activation results in recruitment of transcription factors (Stel2p, R1m1p) which mediate transcriptional changes in response to the detected signals. Coherence of osmoadaptation signalling is maintained by the Hoglp MAPK, which prevents cross-talk with the mating response pathway (516). Genes disrupted in this study are depicted by shadowed circles with red boundaries. Unknown inputs, regulators or transcriptional events are denoted by a question mark. An aberrant transcriptional response, observed only in hog 1 cells, manifests itself as a hyperosmotic stress dependent transcriptional induction (yellow circle boundaries) or repression (blue circle boundary) in connected signalling branches that respond to a specific, non-hyperosmotic signalling input in wild-type cells.

B INSET
community 3: R0X1, HAP1, MSH2, ERGZS SOO
overlay a: UTRZ POG1
community 1: H5P104, RAPT, PUT2, GTS1
community 2: PH01, STR6, COC19, TEC], YAPR, MGA1, SOKZ YHM1, R1M101, FSH1, AGOZ SK01, PORI, OHS, YAP?, PITS, Aux. HSH49, HX71 PH023, KRA 71C
overlap b: DOR48,YPRO15C
INSET A
3.3 Results 104
Figure 3.13: Overview of the hypothetical hyperosmotic stress regulon. See text for full descriptions (Subsection 3.3.6). A sequence logo of a statistically significant mo-tif (GAGATAAACCCGCTCT), discovered by Gibbs sampling of the upstream (-1000 to -1 bp) regions of C. glabrata genes, is displayed in the bottom left corner of the image. The mo-tif was discovered using the default Eukaryotic settings. This results in a 16 nucleotide motif (INSET A), containing two sub-motifs (GAGATAAACCCGCTCT and GCTCT) with higher information content, separated by a low scoring portion. This motif was used to search YEASTRACT transcription factor binding site motifs, allowing 2 substitutions and aligned S. cerevisiae consensus sequences of Gsmlp, Yrrlp, Yaplp, Xbplp, Uga3p, Tecip, Stp2p, Stplp, Stb5p, Rtglp, Rtg3p, Rgtlp, Raplp, Pho4p, Nrglp,Msn2p, Msn4p, Mot3p, Mcmlp, Haclp, Gln3p, Ace2p, Swi5p, Aft1p, Ashlp, Bas1p, Cat8p and Sip4p. Phenotypic enrich-ment of the HORN was investigated by taking 10,000 random 282 node samples from the genome and calculating the number of nodes that were annotated with hyperosmotic stress growth decreased, abnormal or absent in SGD. The permuted null distribution had an average of 1.38 (range = 0 - 10) sensitive phenotypes, whereas the observed number of hyperosmotic stress sensitive nodes in the HORN (12) was significantly greater than ex-pected in a random sample (INSET B). This evidence suggests that the HORN controls the expression of genes of significant number of genes that are required for the adaptation to hyperosmotic stress than would be expected by chance selection of genes from the genome.

/ • /
• •./ •
• / •
•
Chapter 3. The Hog1p and Ste11p dependent transcriptional effects of hyperosmotic stress in Candida glabrata 105
Histogram of Degree in HORN Histogram of Centrality in HORN
O
LL
0
10
20
30 40 0.00 0.02 0.04 0.06 0.08 0.10 0.12
(a)
(b)
HORN Degree v Centrality
0.00 0.02 0.04 0.06 0.06 0.10 0.12
(c)
•
Figure 3.14: Distributions of degree (A) and betweenness centrality (B) and their correlation (C) in the hypothetical osmostress regulon network (HORN). The degree k of a node in a network is simply the number of edges it has to other nodes; betweenness centrality C B (V) (abbreviated as C in figure (B)) is a measure of a node's traffic in the network's structure (Equation 3.2). Vertical red lines depict the mean k (6.12, A) and CB(v) (0.008, B) observed in the HORN. In general there is a strong positive correlation (r = 0.89) between k and CB(v) (C) such that hubs are also highly central to the HORN. However, there are some hubs that have higher than average centrality (red dots, Ino4p, ENA1, UTH1, Rap1p and Sok2p) and some that have lower than average centrality (blue dots, Cin5p, Flo8p, Roxlp, Fhllp, Teclp, Stel2p and Phdlp). The hub threshold in C is depited by the horizontal dashed line at k = 20

MCM1, RAPS. ROX2, 1N04, CANS, TEC1, 5OK2, FHLI, PNDI, FLOG Proportion Upregulated v Degree
• • •
Hub Threshold I
• • • • • •
••• • • •
• • • •• • • • S. • • • • • • *
• • •
•
••• 4150
•
•
•
3,3 Results 106
OAF1,, MET32. SUMS, STP1, MIG1, NDT80, PDR8, ARG80, 0171, GCRI UPC2, Ph102, DOTE, STP2, GIF3 1XR1, RGT1, MET31, MAL33 AZF1, YRM1, TYE1, GAT1, TOSS, DAL81, MOTS, HSF1, mum M641, tION2 PDRI, ACE2, NISPE YOX1 XBP1, Ent GLA13, oz. STBS, HOME YAPT, GCR2,
coo
DPC2, DALSO, GA73, RGM1, META, ME CUPS, RFX1, HAP4, F2F1, NRG2 51J11 0 10 20 30 40
k
Figure 3.15: HORN transcription factor activity versus node connectivity. A simple expression of regulator activity Ant was defined as the number of target genes that the regulator induces expressed as a proportion of the total degree k for each node. Regulator activity (y axis, Ant ) is plotted versus node degree (x axis, k) and classified into four groups (red, blue, green and grey) on the basis of this profile using k-means clustering (4 centroids using the Hartigan-Wong method in the R statisical computing framework). Transcriptional regulators in the red class have the highest Ant (> 80%) and an associated low k (less than the mean value of 6). Transcriptional regulators in the green class are hubs (k > 20, threshold depicted by vertical red line) with the highest degree k > 20 and moderate Art , while those in the grey class are non-hubs with low Art . Blue class transcriptional regulators have moderate Art and k. Remarkably, this method co-classifies several transcription factors that are implicated as regulators of hyperosmotic stress adaptation with low k and high Art (Msn2p, Msn4p, Skolp, Skn7p)

U
Chapter 3. The Hog1p and Ste11p dependent transcriptional effects of hyperosmotic stress in Candida glabrata 107
Figure 3.16: HORN subnetwork comprising putative hyperosmotic stress reg-ulators Sko1p, Skn7p, Msn2p, Msn4p and their immediate k = 1 neighbours. Large rectangular nodes depict transcriptional regulators, whose directed arrows represent a regulatory influence to smaller rectangle target nodes. Yellow nodes represent genes whose transcription is induced after exposure to hyperosmotic stress, whereas blue nodes represent genes whose transcription is repressed under the same conditions. Nodes with red borders represent genes that are required for wild-type like growth in S. cerevisiae cells exposed to hyperosmotic stress. The shape of the node depicts the Hog1p, Stel 1p or Sho1-branch dependence of expression. Ellipses represent Hoglp-specific regulation, triangles represent Stel 1p-specific regulation and octagons represent Shol-branch dependent expression. Rect-angular nodes whose expression is altered independently of Hog1p or Ste11p after exposure to hyperosmotic stress are represented as rounded rectangles.

3.3 Results 108
Figure 3.17: HORN subnetwork (red group in Figure 3.15) comprising highly active regulators and their immediate k = 1 neighbours. Large rectangular nodes depict transcriptional regulators, whose directed arrows represent a regulatory influence to smaller rectangle target nodes. Yellow nodes represent genes whose transcription is induced after exposure to hyperosmotic stress, whereas blue nodes represent genes whose transcrip-tion is repressed under the same conditions. Nodes with red borders represent genes that are required for wild-type like growth in S. cerevisiae cells exposed to hyperosmotic stress. The shape of the node depicts the Hog1p, Stellp or Shol-branch dependence of expression. Ellipses represent Hog1p-specific regulation, triangles represent Stellp-specific regulation and octagons represent Shol-branch dependent expression. Rectangular nodes whose ex-pression is altered independently of Hog1p or Stellp after exposure to hyperosmotic stress are represented as rounded rectangles.

Chapter 3. The Hog1p and Ste11p dependent transcriptional effects of hyperosmotic stress in Candida glabrata 109
Figure 3.18: HORN subnetwork (green group in Figure 3.15) comprising hub regulators and their immediate k = 1 neighbours. Large rectangular nodes depict transcriptional regulators, whose directed arrows represent a regulatory influence to smaller rectangle target nodes. Yellow nodes represent genes whose transcription is induced after exposure to hyperosmotic stress, whereas blue nodes represent genes whose transcription is repressed under the same conditions. Nodes with red borders represent genes that are required for wild-type like growth in S. cerevisiae cells exposed to hyperosmotic stress. The shape of the node depicts the Hog1p, Ste11p or Shol-branch dependence of expression. Ellipses represent Hog1p-specific regulation, triangles represent Ste11p-specific regulation and octagons represent Shol-branch dependent expression. Rectangular nodes whose ex-pression is altered independently of Hog1p or Stel1p after exposure to hyperosmotic stress are represented as rounded rectangles.

GAT3
3.3 Results 110
Figure 3.19: HORN subnetwork (grey group in Figure 3.15) comprising low ac-tivity and low connectivity regulators and their immediate k = 1 neighbours. Large rectangular nodes depict transcriptional regulators, whose directed arrows represent a regulatory influence to smaller rectangle target nodes. Yellow nodes represent genes whose transcription is induced after exposure to hyperosmotic stress, whereas blue nodes represent genes whose transcription is repressed under the same conditions. Nodes with red borders represent genes that are required for wild-type like growth in S. cerevisiae cells exposed to hyperosmotic stress. The shape of the node depicts the Hog1p, Ste1 1p or Shol-branch dependence of expression. Ellipses represent Hog1p-specific regulation, triangles represent Stellp-specific regulation and octagons represent Sho1-branch dependent expression. Rect-angular nodes whose expression is altered independently of Hog1p or Stellp after exposure to hyperosmotic stress are represented as rounded rectangles.

...s161./
Chapter 3. The Hog1p and Stellp dependent transcriptional effects of hyperosmotic stress in Candida glabrata 111
Figure 3.20: HORN subnetwork comprising the pair of transcriptional regulators accounting for the largest proportion of the regulated set and their k = 1 neigh-bours. Large rectangular nodes depict transcriptional regulators, whose directed arrows represent a regulatory influence to smaller rectangle target nodes. Yellow nodes represent genes whose transcription is induced after exposure to hyperosmotic stress, whereas blue nodes represent genes whose transcription is repressed under the same conditions. Nodes with red borders represent genes that are required for wild-type like growth in S. cerevisiae cells exposed to hyperosmotic stress. The shape of the node depicts the Hoglp, Stellp or Shol-branch dependence of expression. Ellipses represent Hoglp-specific regulation, tri-angles represent Stellp-specific regulation and octagons represent Shol-branch dependent expression. Rectangular nodes whose expression is altered independently of Hoglp or Stellp after exposure to hyperosmotic stress are represented as rounded rectangles.

I
3.3 Results 112
Figure 3.21: HORN subnetwork comprising candidate GPD2 regulators and their k = 1 neighbours. Large rectangular nodes depict transcriptional regulators, whose di-rected arrows represent a regulatory influence to smaller rectangle target nodes. Yellow nodes represent genes whose transcription is induced after exposure to hyperosmotic stress, whereas blue nodes represent genes whose transcription is repressed under the same condi-tions. Nodes with red borders represent genes that are required for wild-type like growth in S. cerevisiae cells exposed to hyperosmotic stress. The shape of the node depicts the Hoglp, Stellp or Shol-branch dependence of expression. Ellipses represent Hoglp-specific regula-tion, triangles represent Stellp-specific regulation and octagons represent Shol-branch de-pendent expression. Rectangular nodes whose expression is altered independently of Hoglp or Stellp after exposure to hyperosmotic stress are represented as rounded rectangles.

Chapter 3. The Hog1p and Stellp dependent transcriptional effects of hyperosmotic stress in Candida glabrata 113
(
014
Figure 3.22: HORN subnetwork comprising candidate G CY 1 regulators and their k = 1 neighbours. Large rectangular nodes depict transcriptional regulators, whose di-rected arrows represent a regulatory influence to smaller rectangle target nodes. Yellow nodes represent genes whose transcription is induced after exposure to hyperosmotic stress, whereas blue nodes represent genes whose transcription is repressed under the same condi-tions. Nodes with red borders represent genes that are required for wild-type like growth in S. cerevisiae cells exposed to hyperosmotic stress. The shape of the node depicts the Hoglp, Stellp or Shol-branch dependence of expression. Ellipses represent Hoglp-specific regula-tion, triangles represent Stellp-specific regulation and octagons represent Shol-branch de-pendent expression. Rectangular nodes whose expression is altered independently of Hoglp or Stellp after exposure to hyperosmotic stress are represented as rounded rectangles.

3.3 Results 114
Figure 3.23: HORN subnetwork comprising candidate ALD6 regulators and their k = 1 neighbours. Large rectangular nodes depict transcriptional regulators, whose di-rected arrows represent a regulatory influence to smaller rectangle target nodes. Yellow nodes represent genes whose transcription is induced after exposure to hyperosmotic stress, whereas blue nodes represent genes whose transcription is repressed under the same condi-tions. Nodes with red borders represent genes that are required for wild-type like growth in S. cerevisiae cells exposed to hyperosmotic stress. The shape of the node depicts the Hog1p, Stelip or Shol-branch dependence of expression. Ellipses represent Hog1p-specific regula-tion, triangles represent Ste11p-specific regulation and octagons represent Shot-branch de-pendent expression. Rectangular nodes whose expression is altered independently of Hogip or Steilp after exposure to hyperosmotic stress are represented as rounded rectangles.

-,414102) •
Chapter 3. The Hog1p and Stellp dependent transcriptional effects of hyperosmotic stress in Candida glabrata 115
Figure 3.24: HORN subnetwork comprising direct targets of Cin5p. Large rectan-gular nodes depict transcriptional regulators, whose directed arrows represent a regulatory influence to smaller rectangle target nodes. Yellow nodes represent genes whose transcrip-tion is induced after exposure to hyperosmotic stress, whereas blue nodes represent genes whose transcription is repressed under the same conditions. Nodes with red borders rep-resent genes that are required for wild-type like growth in S. cerevisiae cells exposed to hyperosmotic stress. The shape of the node depicts the Hog1p, Stellp or Shot-branch dependence of expression. Ellipses represent Hog1p-specific regulation, triangles represent Ste11p-specific regulation and octagons represent Shol-branch dependent expression. Rect-angular nodes whose expression is altered independently of Hog1p or Stellp after exposure to hyperosmotic stress are represented as rounded rectangles.

3.3 Results 116
Figure 3.25: HORN subnetwork comprising direct targets of Stel2p. Large rectan-gular nodes depict transcriptional regulators, whose directed arrows represent a regulatory influence to smaller rectangle target nodes. Yellow nodes represent genes whose transcrip-tion is induced after exposure to hyperosmotic stress, whereas blue nodes represent genes whose transcription is repressed under the same conditions. Nodes with red borders rep-resent genes that are required for wild-type like growth in S. cerevisiae cells exposed to hyperosmotic stress. The shape of the node depicts the Hog1p, Stelip or Shol-branch dependence of expression. Ellipses represent Hoglp-specific regulation, triangles represent Ste11p-specific regulation and octagons represent Shol-branch dependent expression. Rect-angular nodes whose expression is altered independently of Hog1p or Stel.l.p after exposure to hyperosmotic stress are represented as rounded rectangles.

ie up c-T3 a. 77./ROX1\
CMS pm (FLcz) 'UAL 1;;10 (AM 1,GrA1
CPA2r (cPA) '!.isr
11;;;i) , PANG
PCLI
ST12 RTAL t,:)2
-FA
"SI it Atn:06 1 V1'.k
NOM
Chapter 3. The Hog1p and Stellp dependent transcriptional effects of hyperosmotic stress in Candida glabrata 117
Figure 3.26: HORN subnetwork comprising the minimal connections between or-thologous S. cerevisiae genes required for growth in hyperosmotic stress. Large rectangular nodes depict transcriptional regulators, whose directed arrows represent a reg-ulatory influence to smaller rectangle target nodes. Yellow nodes represent genes whose transcription is induced after exposure to hyperosmotic stress, whereas blue nodes repre-sent genes whose transcription is repressed under the same conditions. Nodes with red borders represent genes that are required for wild-type like growth in S. cerevisiae cells exposed to hyperosmotic stress. The shape of the node depicts the Hoglp, Stellp or Shol-branch dependence of expression. Ellipses represent Hoglp-specific regulation, triangles represent Stellp-specific regulation and octagons represent Shol-branch dependent expres-sion. Rectangular nodes whose expression is altered independently of Hogip or Stellp after exposure to hyperosmotic stress are represented as rounded rectangles.

B 9.0 —
8.5 —
■ wt YPD ■ parental YPD ■ Null strains YPD
8.0 —
7.5 —
7.0 —
6.5 —
6.0 —
5.5 —
Tim
e to
Mid
poin
t (hr
s)
1
3.3 Results 118
9.0 —
Tim
e to
Mid
poin
t (hr
s)
8.5 —
8.0
7.5 —
7.0
6.5 —
6.0 —
5.5 —
■ wt + 0.5M KCI ❑ parental + 0.5M KCI ❑ Null strains + 0.5M KCI
Gene name inferred by SC homology
Gene name inferred by SC homology
Figure 3.27: Four C. glabrata mutants, each lacking a single, different putative transcription factor are hypersensitive to 0.5 M KC1 in a liquid growth medium assay. A library of 186 C. glabrata null mutants (plate one shown here), each disrupted in one putative transcription factor ORF (based on sequence homology with S. cerevisiae) was incubated in an orbital shaker to stationary phase (18 hours, 37°C, 180 RPM) in YEPD and then sub-cultured in clear well micro-titre plates containing YEPD with (A) or without (B) 0.5 M KC1. Bi-hourly recorded optical density (0D600) measurements, used in place of cell counts, were fitted to a three parameter logistic function of growth using non-linear regression function SSLogistic from the R statistical computing framework. The slowest growing strains in 0.5 M KC1 were identified as hyperosmotic stress sensitive strains: (hstl, srb8, msnl, ga11.1, gisl)

Chapter 3. The Hog1p and Stellp dependent transcriptional effects of hyperosmotic stress in Candida glabrata 119
9.0 —
■ wt + 0.5M KCI ■ parental + 0.5M KCI ■ Null strains + 0.5M KOI
8.5 —
8.0
Tim
e to
Mid
poin
t (hr
s)
7.5
7.0
6.5
6.0 —
5.5 —
8
Gene name inferred by SC homology
B
■ wt YPD ■ parental YPD ■ Null strains YPD
9.0 —
8.5 —
8.0 —
Tim
e to
Mid
poin
t (hr
s)
7.5 —
7.0 —
6.5 —
6.0 —
5.5 —
1j5g3 .! ,AgiAf,:cia. 2 1! zt F, 2 2 ts 31° DA! ;
Gene name inferred by SC homology
Figure 3.28: Three C. glabrata mutants, each lacking a single, different putative transcription factor are hypersensitive to 0.5 M KC1 in a liquid growth medium assay. A library of 186 C. glabrata null mutants (plate two shown here), each disrupted in one putative transcription factor ORF (based on sequence homology with S. cerevisiae) was incubated in an orbital shaker to stationary phase (18 hours, 37°C, 180 RPM) in YEPD and then sub-cultured in clear well micro-titre plates containing YEPD with (A) or without (B) 0.5 M KC1. Bi-hourly recorded optical density (0D600) measurements, used in place of cell counts, were fitted to a three parameter logistic function of growth using non-linear regression function SSLogistic from the R statistical computing framework. The slowest growing strains in 0.5 M KC1 were identified as hyperosmotic stress sensitive strains (rgtl, tafl4, rnsn4).

3.4 Discussion 120
3.4 Discussion
Stress activated protein kinases are important molecular signalling molecules involved in environmental sensing and adaptation, a critical requirement for host colonization. In this study, we used the consensus HOG pathway architecture, taken from an array of genetic interaction and comparative genomics studies in S. cerevisiae (comprehensively reviewed in (292)), as a conserved signalling pathway for understanding Hog1p function in C. glabrata. Using hogl and stel 1 null mutants in tandem with transcription profiling, we report a difference between the genetic architecture of the HOG pathway in C. glabrata ATCC 2001 and that of the model organism S. cerevisiae (Figure 3.3). The latter has two signalling branches that must both be disrupted to prevent phosphorylation of Hog1p. In contrast, like C. albicans, C. glabrata ATCC 2001 relies on only one (Ste11p) MAPKKK for Hog1p phosphorylation and osmoadaptation after an osmotic upshift (126), but unlike C. albicans this MAPKKK (Ste1 1p belongs to the putative Shol signalling branch and not the SLN1 signalling branch. Initially, it was appealing to speculate on these differences in an evo-lutionary context; however during the course of this work, a study of C. glabrata ATCC 2001 by Gregori et al., (255) revealed the same sole dependency on the Shol branch for osmoadaptation, which was not evident in two other C. glabrata strains (BG2 and Cg2633) and therefore is not generalizable to the species.
In initial gene deletion studies, adopting a non-prototrophic selection method suitable for all three strains, Gregori and colleagues demonstrated that disruption of PBS2 led to an osmosensitive phenotype in all three strains, whereas deletion of SHO1 led to an osmosen-sitive phenotype only in the ATCC 2001 background. Crucially, this observation arises from strain-dependent differences in a single Slnl branch component: SSK2. Of the three strains, the ATCC 2001 strain uniquely possesses a SSK2 allele (ssk2-1 ) with a premature stop-codon upstream of the phosphotransfer domain, which blocks Sln1p branch dependent phosphorylation of Pbs2p. As such, Gregori and colleagues relay a cautionary message in in-terpreting phenotypic results of gene deletion studies: "all mutations or deletions generated in the ATCC 2001-derived genetic backgrounds are in fact double mutants also carrying a Cgssk2-1 allele. Thus, any stress-related phenotypes and molecular cross talk between different signaling pathways may be caused by synthetic genetic interactions due to the presence of the mutated Cgssk2-1 allele and the lack of the second gene". However, this can be said of many C. glabrata null mutants that have been created with mutagenic trans-formation protocols, since there exists no facility to back cross C. glabrata strains to obviate the mutagenic nature of direct gene disruption. Moreover, this genetic quirk provides an opportunity to investigate the Shol branch contribution to hyperosmotic stress adaptation without deletion of relay components upstream of Pbs2p in the Slnl branch.

Chapter 3. The Hog1p and Ste11p dependent transcriptional effects of hyperosmotic stress in Candida glabrata 121
Reconstituting ATCC 2001 with a full-length SSK2 ORF from another C. glabrata strain such as BG2 would ensure that the upstream architecture of S. cerevisiae and this C. glabrata strain are collectively orthologous (however this has been particularly difficult to achieve at the chromosomal locus, Christa Gregori Personal Communication). Transcrip-tional differences could then be explained in terms of how each component relays osmostress signals, or in terms of mechanisms downstream of Hog1p rather than differences in SAPK architecture per se. In the absence of such a reconstituted strain, it was possible to compare the transcriptional response of a S. cerevisiae sskl strain treated with 0.5 M KC1 to our "wild-type" (ssk2-1) ATCC 2001 strain (518). Although this comparison is not ideal, it per-mits comparison of C. glabrata and S. cerevisiae strains lacking a functional Slnl branch. Although the proportion of functionally conserved transcripts is slightly higher between these strains (20.10%) (Supplementary Data Table D.23) than in the comparison of ATCC 2001 strain to the wild-type S. cerevisiae strain, there are still prominent hyperosmotic transcription signatures in C. glabrata ATCC 2001 that are absent in S. cerevisiae ssk1. Therefore, even with a more similar SAPK architecture, we could still identify hyperosmotic stress induced transcriptional differences between two species, spanning several biological processes.
Although the comparisons with S. cerevisiae data are likely to be confounded by the lack of direct experimental comparison, my analyses recovered only a small conserved re-sponse (< 15%) between C. glabrata and S. cerevisiae (Supplementary Data Tables D.9, D.10, D.11 and D.12), encompassing genes encoding glycerol biosynthetic enzymes ( GPD1/ 2, GCY1, ALD6 (230)), DNA damage response proteins (DDR48 and RCN2 (230; 518)), a heat shock protein (HSP42 (230; 518)), a v-snare binding protein that facilitates spe-cific protein retrieval from the late endosome (BTN2 (230; 518)) and a proline permease (PUT4 (518)). The regulation of some (37%) transcripts is consistent with the S. cerevisiae consensus HOG pathway model (GPD1/2, GCY1, ALD6, PTC1, PTP3 are Shol branch dependent), moreover this analysis identifies a novel Sho1 branch dependency for some transcripts induced under hyperosmotic stress (ALG3,UBI4, RPN11 and CPA1). Impor-tantly, a large component (30%) of the hyperosmotic stress induced transcriptional changes occurs independently of the extant Shol branch (summarized in Figure 3.29), and includes transcripts whose hyperosmotic stress induction is also conserved with S. cerevisiae (BTN2, P HM8 and PRM10 and DDR48), a proteasomal lid subunit (RP N6) and a transcriptional regulator of the oxidative stress response (CIN5). Indeed, the BTN2 transcript showed the highest fold-induction in response to hyperosmotic shock in C. glabrata ATCC 2001 cells. This gene encodes a v-snare that mediates late endosome-Golgi sorting (339), and modulates L-arginine uptake through localization of Canlp. It is not clear if modulation of L-arginine levels is part of an adaptive response to hyperosmotic stress in S. cerevisiae or
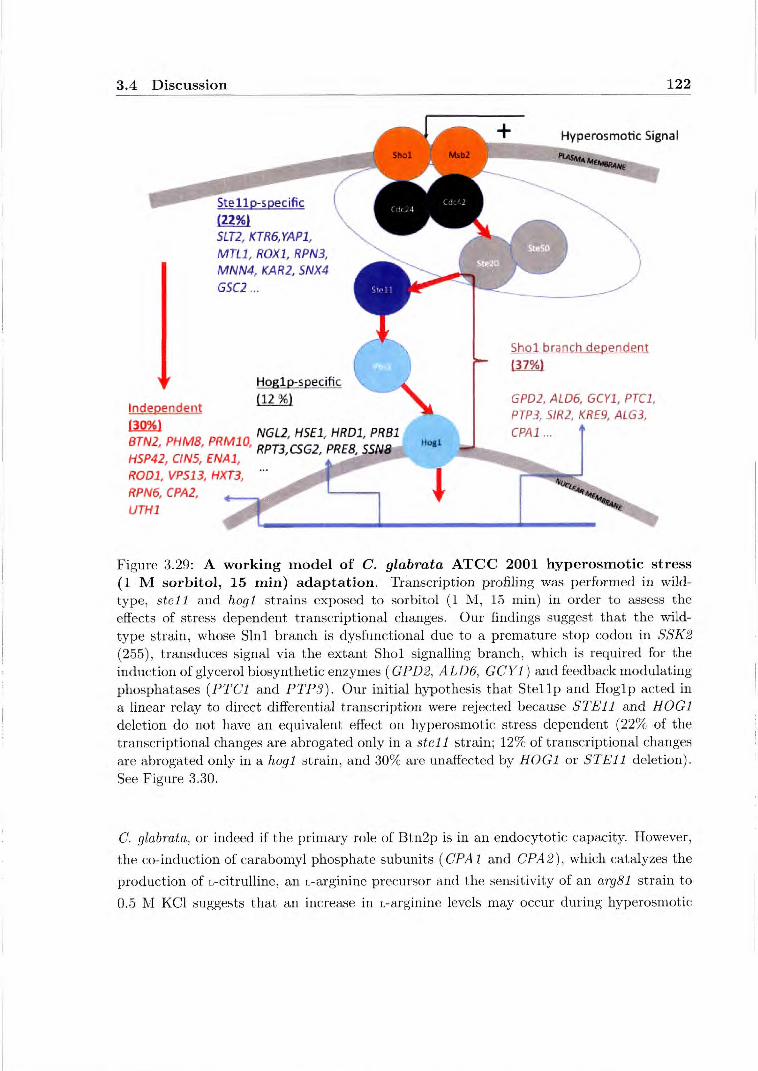
Hyperosmotic Signal
'PWAISt4tmeRA
1
Independent
130%1 BTN2, PHM8, PRM1 HSP42, CIN5, ENA1, ROD1, VP513, HXT3, RPN6, CPA2, UTH1
Hog1p-specific (12 %)
NGL2, HSE1, HRD1, PRB1 RPT3,CSG2, PRES, SSN8
Stellp-specific (22%1 SLT2, KTR6,YAP1, MTL1, ROX1, RPN3, MNN4, KAR2, SNX4 GSC2
Shol branch dependent 07%1
GPV2, ALD6, GCY1, PTC1, PTP3, S1R2, KRE9, ALG3, CPA1 ...
3,4 Discussion 122
Figure 3.29: A working model of C. glabrata ATCC 2001 hyperosmotic stress (1 M sorbitol, 15 min) adaptation. Transcription profiling was performed in wild-type, ste11 and hog1 strains exposed to sorbitol (1 M, 15 min) in order to assess the effects of stress dependent transcriptional changes. Our findings suggest that the wild-type strain, whose Slnl branch is dysfunctional due to a premature stop codon in SSK2 (255), transduces signal via the extant Shol signalling branch, which is required for the induction of glycerol biosynthetic enzymes ( GPD2, ALD6, GCY1) and feedback modulating phosphatases (PTC1 and PTP3). Our initial hypothesis that Stellp and Hog1p acted in a linear relay to direct differential transcription were rejected because STE11 and HOG1 deletion do not have an equivalent effect on hyperosmotic stress dependent (22% of the transcriptional changes are abrogated only in a sten strain; 12% of transcriptional changes are abrogated only in a hog1 strain, and 30% are unaffected by HOG1 or STE11 deletion). See Figure 3.30.
C. glabrata, or indeed if the primary role of Btn2p is in an endocytotic capacity. However,
the co-induction of carabomyl phosphate subunits (CPA1 and CPA2), which catalyzes the
production of L-citrulline, an L-arginine precursor and the sensitivity of an arg81 strain to
0.5 M KC1 suggests that an increase in L-arginine levels may occur during hyperosmotic

Chapter 3. The Hogip and Ste11p dependent transcriptional effects of hyperosmotic stress in Candida glabrata 123
stress adaptation. The inequivalent effects of HOG1 and STE11 deletion on hyperosmotic stress induced
transcription suggest that Stellp does not act as a simple relay to Hogip (via Pbs2p) to elicit transcriptional changes (Figures 3.29 and 3.30). The identification of a Stellp specific hyperosmotic transcriptional response (Figure 3.30 a) implies that 22% of the tran-scriptional changes are mediated by Stellp, independently of Hog1p, possibly by recruiting additional downstream signalling proteins that regulate transcriptional events. An alter-nate hypothesis is that Stellp is required to phosphorylate Pbs2p, which in turn has other non-Hoglp substrates that regulate the transcriptional hyperosmotic stress response. The striking, hyperosmotic stress dependent, transcriptional activation of components of the ad-jacent cell wall remodelling pathway (Figure 3.32) which was previously undescribed in C. glabrata and S. cerevisiae may be caused by cross-talk at the level of Stellp (as evidenced by Stellp specific transcription of SLT2, KTR6, GSC2 and MNN4 ). A key difference between this study and transcript profiling studies of hyperosmotic stress in S. cerevisiae, is this induction of genes whose products affect cell wall structural integrity, both at the regulatory and effector level. For example, the rapid induction of genes encoding enzymes involved in the biosynthesis of the carbohydrate components of the cell wall (chitin syn-thase I CHS1, KRE9 - required for /3-1,6 glucan assembly, GSC2 - the catalytic subunit of 0-1,6 glucan synthase) indicate that C. glabrata mounts a significant re-modelling of the cell wall carbohydrate content in response to sorbitol treatment. Moreover, genes involved in glycoprotein mannosylation (KRE2, KTR6, MNN4 and GUK/ ) and N-linked glycosy-lation (ALG3) are also induced in the same time-frame. The simultaneous repression of ergosterol biosynthetic enzymes (ERG1, ERGS and ERG25) and induction of chitin syn-thase I ( CHS1) are likely to have effects on the physical properties of the plasma membrane (decreased fluidity) and cell wall (increased tensile strength), which I hypothesize may in-crease the turgor pressure of the cell. However, the cell wall remodelling is likely to be more complicated. An alternate speculation is that pre-existing cross-linked glucan and chitin moieties are replaced with new components that do not become cross-linked, as suggested by the concomitant repression of UTR2, a cell wall protein that functions as a chitin trans-glycosidase in the cross-linking of chitin to 13(1-6)-glucan. Indeed, reduced cross-linking of chitin, as artificially imposed by calcofluor white staining in growing cells, may results in a (counter-intuitive) reduction in cell wall strength even when chitin levels are elevated (565). Furthermore, it is also important to consider other physical changes that may occur as a result of this proposed cell wall remodelling. For example how does cell wall permeability change with respect to its cell wall composition and does this instead confer some osmoad-aptive advantage? De Nobel and colleagues showed that N-linked mannoproteins (whose gene expression is induced in this study) play a major role in limiting cell wall porosity in

3.4 Discussion 124
S. cerevisiae (161). The upregulated N-linked mannosylation may alter the permeability of the cell to limit water efflux during osmotic stress.
In addition to an Stellp specific component, a Hog1 p specific hyperosmotic transcrip-tional response was identified (Figure 3.30 b), which implies that the hyperosmotic stress signal is partially transduced independently of Stellp. The majority of these Hoglp-specific, hyperosmotic stress induced transcripts are involved in mediating proteolysis, either in the vacuole or in the proteasome, although the regulation of transcripts involved in proteolysis was not exclusively Hog1p dependent. We identified C. glabrata-specific, Hogip and Stellp dependent induction of transcripts involved in the ubiquitination of proteins ( UB/-4, HRD1, UFD2). The covalent attachment of ubiquitin (encoded by UB/4 ) to a lysine residue of a substrate protein is a versatile regulatory post-translational modification that can de-termine the fate of a protein depending on the degree of conjugation. This process may involve conjugation of a single ubiquitin (monoubiquitination), several ubiquitin molecules (multiubiquitination) or the attachment of ubiquitin chains (polyubiquitination) to a sub-strate protein via a three step process involving the sequential action of activating (El), conjugating and ligase enzymes (E3) (HRD1 ), which can recruit proteins in a variety of cellular processes such as proteasomal and endoplasmic reticulum associated degradation, signal transduction, membrane trafficking, DNA repair, chromatin remodelling and peroxi-some biogenesis (175; 264; 285) that are important in environmental stress adaptation. The consequences of ubiquitination are diverse, and it is possible that one or more of the above cellular processes is recruited in response to hyperosmotic stress in C. glabrata. However, the co-induction of genes encoding subunits of the proteasomal (Figure 3.31) degradation ma-chinery (RPN11, RPT3, PRES and RPN3) provides supporting evidence for an osmostress induced, HOG1 and STE// -dependent induction of genes having protein degradation func-tion. The functional roles of 19S proteasome subunits remain poorly understood, however Rpn3p is required for controlling cyclin levels in Gi /S transition (43; 338). Recent stud-ies by Posas et al., showed that Hoglp mediates cell cycle S-phase delay in response to hyperosmotic stress in S. cerevisiae (761); in the light of these findings and this study, future work may involve studying the role of the proteasome in controlling cyclin levels and cell cycle progression under hyperosmotic stress in C. glabrata. The subunit (Rpn3, RpnS, Rpn6, Rpn7, Rpn8, Rpn9, Rpnl 1 and Rpn12) stoichiometry of the 19S proteasome is typically invariant, however mature proteasomes are also directly modified in response to environmental stress (reviewed in (245)). The differential expression of genes encoding these subunits observed in this study may be an artifact of transcript profiling noise, however it is possible that this leads to an altered stochiometry of these regulatory subunits and an altered regulatory function. Indeed, RPN6 - induced by hyperosmotic stress in a Hoglp and Stellp independent manner, encodes a non-ATPase regulatory subunit that binds to im-

Chapter 3. The Hog1p and Stel1p dependent transcriptional effects of hyperosmotic stress in Candida glabrata 125
portant components of adjacent MAPK signalling cascades of S. cerevisiae (Ksslp, Tec1p, Ste12p, Slt2p, Bcklp and Ste11p) and to other proteins induced in hyperosmotic stress conditions (Hxt6p, Grx3p, Pho84p - this study) (291). An intriguing possibility that Shol branch independent expression of Rpn6p may regulate core proteasomal degradation of MAPK components to maintain coherent hyperosmotic stress signalling warrants further investigation.
Why might the HOG pathway regulate protein degradation in hyperosmotic stress con-ditions? One explanation is that Stelip and Hogip control the proteasome in its function as a housekeeper - preventing the intracellular accumulation of aberrantly folded, functionally impaired and potentially toxic proteins that arise as a result of hyperosmotic stress (248). Evidence supporting this role includes the co-induction of heat-shock proteins encoded by HSP78, HSP104 and APJ1. Although the expression of heat-shock protein folding chaper-ones is known to be induced in response to heat-denatured proteins (674), they can also be expressed in a variety of other environmental stress conditions (230; 118) including osmotic stress. The proteasome has also been shown to act as a regulatory feedback mechanism in the preferential degradation of unphosphorylated Ssklp (609). Assuming C. glabrata ATCC 2001 cells maintain an active signalling branch (Ypdlp, Sln1p) upstream of the truncated Ssklp lacking a phosphoreceiver domain, then hyperosmotic stress would lead to an ac-cumulation of unphosphorylated Ssklp relative to cells with a complete and active Slnl branch exposed to hyperosmotic stress, which may require modulation by this degradative mechanism. Placed in the context of the physical changes that must occur in the plasma membrane and in the cell wall during hyperosmotic stress adaptation, this likely involves targeted endocytosis of proteins that would normally increase either the permeability of the cell to water or glycerol, or alter the composition of the cell wall in order to modulate tur-gor pressure. A second, explanation of Hog1p dependent control of the proteasome is that it may provide a more subtle, stress-dependent mechanism for targeted turnover of active proteins, which augments the transcriptional repression and induction employed to alter the protein content of the cell in response to hyperosmotic stress. Such a targeted turnover of active proteins, may also be achieved by altering the cellular location of proteins rather than by their degradation. For example, the co-induction of ROD1 (also known as ART4), an arrestin-related protein that targets specific plasma membrane proteins for endocytic downregulation by serving as adaptors for Rsp5 (409), suggests that one consequence of the induced ubiquitin machinery genes may involve targeting membrane protein cargoes for endocytosis (409). Such cargo specific endocytosis, mediated by arrestin-related proteins is believed to be important in stress induced plasma membrane remodelling of S. cerevisiae, particularly in alkaline stress adaptation (284; 631).
I also identified an hyperosmotic stress dependent induction of transcripts that are

3.4 Discussion 126
normally associated with oxidative stress protection in S. cerevisiae. Monothiol glutaredoxin (encoded by GRX7) and peptide methionine sulphoxide reductase (encoded by MXR1) are induced in response to hyperosmotic stress by a mechanism that requires Hog1p or Ste11p, as is the putative transcriptional regulator of the oxidative stress response ( YAPI). Maintaining the redox state of cysteine thiol groups - which govern protein tertiary structure and co-factor binding - is critically important for correct maintenance of protein function. Without intervention, reactive oxygen species can disrupt this optimal redox state leading to misfolded and functionally impaired proteins. Glutaredoxins typically act as glutathione-dependent disulphide oxidoreductases, reducing superoxide anion or hydroperoxide in place of proteins' reactive sulphydryl groups to prevent the formation of mixed disulphides in proteins (425), however GRX7 is not involved in the the general maintenance of disulphide bonds, but is required for reduction of cysteine residues in certain substrate proteins of the secretory pathway that require reduced thiol residues for their functionality (462).
Methionine residues and iron-sulphur clusters are similarly vulnerable to oxidation by cellular reactive oxygen species, and the methionine sulphoxide reductases, such as that encoded by MXR1 reduce oxidized methionines and protect iron-sulphur clusters from re-active oxygen species in an analagous fashion to the glutaredoxin mediated protection of cysteine-sulphydryl groups. The induction of oxidative stress response genes by (ostensi-bly) independent stressors is well documented in the S. cerevisiae (but not in C. albicans (191)) literature (230; 118), in what has become known as the general stress response, and latterly the environmental stress response (ESR). The cross-protection against difference stresses, evident from the fact that cells exposed to a mild dose of one stress become resis-tant to a normally lethal dose of another stress (467; 742), has been interpreted as a general mechanism of cellular protection that is induced when cells are exposed to stressful stimuli. However, a fundamental assumption underpinning this interpretation is that the "indepen-dent" stress conditions used have no common effect, or that during the adaptive response the cell itself does not produce stressful by-products in the course of finding an optimal solution to the immediate cellular threat. In this study, the induction of oxidative stress response genes by hyperosmotic stress treatment, is a likely consequence of the metabolic demands of ATP-dependent proteasomal degradation, glycerol production and remodelling of the cell wall. Indeed, induction of transcripts involved in glycolysis, the TCA cycle and oxidative phosphorylation would support this notion - oxidative phosphorylation, in partic-ular, is known to produce reactive oxygen species (235; 676; 525; 403; 80). Furthermore, the production of reactive oxygen species is an inevitable consequence of protein modification in the endoplasmic reticulum (440).

Chapter 3. The Hog1p and Stellp dependent transcriptional effects of hyperosmotic stress in Candida glabrata 127
In his entertaining review paper, entitled "can a biologist fix a radio?" (396)6 Lezebnik asks how would molecular biologists and electrical engineers go about fixing a complex system that they know nothing about? Which methodologies typify the separate disciplines and what are the relative merits of each approach? More importantly: who would you bet on to fix the radio? In short, he advocates the approach of the engineer, based on a quantitative approach to reverse engineering the radio's wiring diagram. In systems biology, we have a considerably harder problem: our systems are noisy, model parts are not yet fully characterized7 and most model parameters are immeasurable with current technologies. Our knowledge of how model components interact is also still embryonic, but technological advances have furnished the community with some interaction data: methods exist to identify physical interactions between proteins (705; 318) and genetic interactions in single and double null mutant strains of S. cerevisiae (404; 693; 692). However, these data also have well documented caveats (568; 41; 661; 162; 363; 265). For example, the protein-protein interactome of S. cerevisiae is incomplete (271) both in terms of the number of known nodes that can participate in the network, and also the incompleteness of edges that exist in the nodes that are present. The ensuing subnetworks are also biased towards proteins from certain cellular environments, towards more ancient, conserved proteins and towards highly expressed proteins (721). This has important implications in assertions made about the properties of the unobserved "complete" network. Most studies of networks have only looked at the small subsets of the true network. Only if the degree distributions of the network randomly sampled subnets belong to the same family of probability distributions is it possible to extrapolate from sub-network properties to the full network from which the subgraph is taken. Although some theoretical graphs indeed satisfy these criteria, scale-free degree distributions, which caused such a stir a decade ago, do not satisfy this condition (666). Clearly problems are abound, even in a model organism.
The picture is even bleaker in non-model organisms, where the parts-list is largely in-ferred by computational annotation, and interaction data are virtually non-existent. In a progressive step, a project has recently commenced in our laboratory (Hsueh-Lui Ho, Per-sonal Communication) to obtain large-scale protein protein interaction data in C. glabrata. However, in the absence of these richer data sets, I have adopted a rather crude method to propose a candidate regulatory circuit for hyperosmotic stress in C. glabrata. The HORN was created by searching for transcription factor associations with the differentially tran-
6This biologist has never fixed a radio. However with an aging mother in-law, I have become proficient in explaining the intricacies of a VCR
7genome snapshot http://www.yeastgenome.org/cache/genomeSnapshot.html reveals 968 uncharacter-ized ORFs in september 2009, representing approximately 15% of predicted gene sequences. Moreover, a peptidocentric view underestimates the complexity of the parts list

3.4 Discussion 128
scribed orthologous C. glabrata genes which relies on a strong assumption that transcrip-
tional regulators share conserved regulatory behaviour in C. glabrata and S. cerevisiae.
Borneman et al., (77) used chromatin immunoprecipitation and gene expression microar-ray (CHIP-chip) analysis to demonstrate the divergence of Tec1p and Stel2p binding sites between members of the Saccharomyces sensu stricto species, with species specific Stel2p and Tec1p binding and activation profiles varying greater than gene sequences between these species. Such differences in gene regulation are likely to be a major cause in species divergence and should be investigated in C. glabrata which is even more distantly related, and potentially harbours greater TF binding site divergence. CHIP-chip technologies are likely to be superceded by higher resolution techniques in the near future, and any such in-vestigations of transcription factor binding site divergence should be developed with higher throughput, more cost efficient methods such as the multiplexed CHIP-seq protocol recently adopted in S. cerevisiae (401).
Mapping transcriptional status to the HORN does not fully describe the activation status of the incorporated transcriptional regulators, which may be affected by e.g. phos-phorylation (228), redox state (646) and cellular localization (658). A more detailed un-derstanding of hyperosmotic stress regulation may therefore require further experimental work to determine the phosphorylation status of potential targets using high throughput phosphoproteomic methods (71; 72; 683) or mathematical methods for their in silico pre-diction. Hidden variable dynamical modelling (HVDM) has been used with some success to predict downstream targets of p53 (52), and has since been improved upon by the same group (51) to remove the requirement of prior knowledge of some targets to train the model. In their revamped genome-wide transcriptional modelling (GWTM) approach, Barenco et al., directly explained 70% of the 200 most significantly upregulated genes in the DNA damage response of a mammalian cell line, but required time course measurements of gene expression and estimates of transcript degradation rate that have not yet been performed for hyperosmotic stress adaptation in C. glabrata. Despite these major caveats, some hy-potheses arise from the analysis of local network topology and community structure. Simple analysis of target activity and node degree suggest that there are transcription regulators other than the putative S. cerevisiae Hog1p targets (1\ilsn2p/Msn4p, Sko1p and Skn7p) in-volved in hyperosmotic stress regulation in C. glabrata. HORN hubs with moderate activity (i.e. approximately 80% of direct targets showing differential expression) such as Mcm1p, Rap1p, Roxlp, Ino4p, Cin5p, Sok2p, Fhllp, Phdlp and Flo8p may also play a role in governing the transcriptional response to hyperosmotic stress in C. glabrata. Furthermore, transcription factors are likely to be co-ordinated and redundant in their function as solid agar and liquid culture assays of single gene knockout CGTFKOlib strains did not identify any hypersensitive (i.e. completely abrogated growth) strains.

Chapter 3. The Hog1p and Ste11p dependent transcriptional effects of hyperosmotic stress in Candida glabrata 129
In the community structure analysis, three HORN communities were discovered by k—clique percolation (522), however the majority of nodes (85%, 242 nodes) were not at-tributed to any particular community. Accordingly, an incomplete and inaccurate picture of HORN organization may have been presented. The method of k—clique percolation is based on the concept of a k—clique, which is a complete subgraph of k connected with all k(k — 1)/2 possible links and relies on the observation that communities seem to consist of several smaller cliques that share many nodes with other cliques of the same commu-nity. A k—clique community is then defined by the union of adjacent k—cliques, which share k — 1 nodes and is particularly sensitive to the choice of k, with k = 2 effectively reducing to link percolation (k = 2 signifies an edge rather than any complicated subgraph) and larger values of k imposing more stringent requirement for communities (e.g cliques with large k values are likely to be multi-subunit complexes). The use of such cliques as a basis for community detection does enable the detection of overlapping communities, with nodes existing in multiple contexts, however this comes at the cost of detection sensitivity. Is this a worthwhile cost? Overlapping community detection appears to be of significant interest in the social sciences, where community membership might be used to infer po-litical (412) and, more controversially (and as yet unpublished), sexual preferences (331), in addition to consumer appetites (590). We would also like to make analagous inferences in computational biology; for example can a protein community be used to infer biological function? In this study, there seems to be little scope for such an approach, simply because of the small number of detected communities. However, DDR48 and the uncharacterized YPRO15C genes are the point of overlap for two functionally diverse HORN communities. Community 2 (Figure 3.13) contains transcriptional regulators involved in filamentation (Phdlp, Tec1p, Sok2 (242; 727; 387)), ambient pH adaptation (RimlOip (388)), hyperos-motic stress adaptation (Skolp (580)), multi-drug resistance (Pdrlp (441)) and oxidative stress adaptation (Cin5p (461; 222; 646) and Yap7p (200)). Community 3 is more homo-geneous and contains transcriptional regulators involved in hypoxic gene reguation and the oxidative stress response (Skn7p Roxlp, Hap1p and Msn2p (563; 771; 118; 380; 381)). The combined regulatory context of DDR48 and the uncharacterized YPRO15C indicates that they might be regulated during filamentation, alkaline adaptation, hypoxia and azole efflux. There is some literature to support these predictions: DDR48 is regulated in C. albicans during exposure to ketoconazole (419), hypoxia (622), hyperosmotic stress (191), ambient pH adaptation (62) and is also essential for filamentation (176). Although YPRO15C is less well characterized, the C. albicans orthologue of YPRO15C NRG1 lorf19.7150 is a negative regulator of filamentation (88). The community context identified here suggests that the gene product may also play a role in stress responses in a similar manner to Ddr48p.

(a)
Specific Hogip Requirement Hyperosmotic Signal
PLASMA M[~A9AAYL
Hogip-specific 112 961
NGL2, HSE1, NRD1, PRS1 RPT3,CSG2, PRES, SSN8
(b)
Specific Stellp Requirement
•
Stel le-specific 122%1
KT116,YAP1, 4."*.
• • ***** * MTLI, ROX1, RPN3, 2
MNN4, KAR2, SNX4 GSC2 ..
Hyperosmotic Signal
'ftW
3.4 Discussion 130
Figure 3.30: Alternate hypotheses for hyperosmotic stress signal transduction through the Sho1 branch of C. glabrata ATCC 2001. The identification of a Stellp specific hyperosmotic transcriptional response (a) implies that 22% of the transcriptional changes are mediated by Stellp independently from Hogip, possibly by recruiting addi-tional downstream signalling proteins (*1) that regulate the induction of genes such as (incomplete list) SLT2, KTR6, ROX1, RPN3 and MNN4. Another alternate explanation is that Stellp is required to phosphorylate Pbs2p (*2), which in turn has other non-Hogip substrates that are involved in the hyperosmotic stress response. The identification of a Hog1p specific hyperosmotic transcriptional response (b) implies that the hyperosmotic stress signal is partially transduced independently of Ste11p, leading to the transcriptional regulation of (incomplete list) NGL2, HSEJ, HRDI, RPT3, PRES. This suggests that sig-nal may be transduced via upstream components that are not yet characterized (?1), and /or directly to Pbs2p (?2)

Chapter 3. The Hog1p and Stel1p dependent transcriptional effects of hyperosmotic stress in Candida glabrata 131
Figure 3.31: Four C. glabrata transcripts, orthologous to S. cerevisiae genes en-coding core and regulatory subunits of the 26S proteasome are induced in cells exposed to 1 M sorbitol for 15 minutes. Differentially transcribed transcripts were identified by gene expression microarray analysis and mapped to C. glabrata pathways of metabolism and cell signalling in the KEGG database. Red boxes denote subunits whose transcripts were more abundant after exposure to hyperosmotic stress imposed by addition of 1 M sorbitol to rich culture medium for 15 minutes. Four transcripts encode regula-tory subunits of the 19S proteasome lid (RPNS, RPN6, RPN11) or base (RPT3) and one transcript encodes a subunit of the proteasome core a subunit (PRE8)

MAPIC
MAPK SIGNALING PATHWAY
MAPIOCK INAPECK
04011 1 1.9709 Kenebsa Laboratories
3.4 Discussion 132
Figure 3.32: Two C. glabrata transcripts, orthologous to S. cerevisiae genes encoding members of the adjacent cell integrity pathway are induced in cells exposed to 1 M sorbitol for 15 minutes. Differentially transcribed transcripts were identified by gene expression microarray analysis and mapped to C. glabrata pathways of metabolism and cell signalling in the KEGG database. Red boxes denote subunits whose transcripts were more abundant after exposure to hyperosmotic stress imposed by addition of 1 M sorbitol to rich culture medium for 15 minutes.

Chapter 3. The Hog1p and Ste11p dependent transcriptional effects of hyperosmotic stress in Candida glabrata 133
3.5 Chapter Summary
1. Disruption of a MAPKKK (STE11 ) or MAPK (HOG/ ) in the putative SHO1 sig-nalling branch is sufficient to prevent sorbitol stress (1 M) induced Hog1p phospho-rylation and proliferation in a strain derived from C. glabrata ATCC 2001. This observation was independently reported during the course of the study (255), and is not a universal feature of all C. glabrata strains.
2. Transcription profiling of ATCC 2001 exposed to sorbitol (1 M) for 15 minutes iden-tified a transcriptome that is dissimilar (< 15%) to relevant studies in S. cerevisiae and C. albicans. The transcript profile of S. cerevisiae sskl strain, a background more closely related to ATCC 2001, subjected to hyperosmotic stress is also poorly conserved (< 21%). Our hypothesis that Hoglp phosphorylation would be sufficient to cause a highly conserved transcriptional response to hyperosmotic stress between S. cerevisiae and C. glabrata was rejected.
3. Using an annotation inferred from S. cerevisiae, several biological themes emerged in the set of induced and repressed transcripts. In keeping with the current model of osmoadaptation, key glycerol biosynthetic enzymes were induced, although not all were Hoglp dependent. Other biological processes represented in the set of induced transcripts include, ubiquitin mediated proteolysis, cell wall re-modelling and response to oxidative stress.
4. Transcription profiling of putative C. glabrata HOG pathway mutants allowed us to investigate the architectural dependency of the hyperosmotic stress transcript pro-files. A total of 30% of transcripts induced on exposure to hyperosmotic stress were independent of Hoglp or Stellp; 37% of induced transcripts depended on Stellp or Hoglp; 13% of induced transcripts were specifically dependent on Hoglp and 20% were specifically dependent on Stellp.
5. The observation that STE11 deletion and HOG1 deletion do not have the same effects on hyperosmotic stress induced gene expression is inconsistent with a model in which Stellp and Hoglp function together only as a simple relay of signal status.

134
Chapter 4
Sub-telomere directed gene expression during initiation of invasive aspergillosis
4.1 Chapter Overview
Aspergillus fumigatus is a common mould whose small asexual spores (2-3 tiM diameter) enable prolonged suspension in the normal airborne flora (394). The size of these conidia not only facilitates their sustained dispersion, but also permits entry into the pulmonary alveoli of the mammalian lung (576), whereupon the level of host immunocompetence determines their fate, and conversely, the morbidity of the host (394; 773).The host niche imposes common constraints on opportunistic pathogens such as A. fumigatus and some Candida spp; to successfully exploit an immunocompromised host, these organisms must be able to withstand purgative mechanical forces in order to establish themselves on a substrate (164; 1), sense the local environment, exploit the available nutrients (584) and evade host defence mechanisms (657) that function to expunge the foreign organism. Such commonality is reflected in the study of A. fumigatus virulence, with much research focused on the study of host-pathogen adhesion (e.g. the effect of hydrophobins), the role of secreted proteolytic enzymes such as aspartic proteases, (572; 573) serine alkaline proteases (e.g. Alpp (365; 444; 485; 571) the metalloprotease (Mepp) (217; 327; 365; 476; 653)), dipeptidylpeptidases (56; 57) and enzymes involved in the dissipation of reactive oxygen species (266; 279). However, in keeping with their phylogenetic differences, virulence studies in A. fumigatus have also diverged, notably in an effort to understand the role of pigments (391; 323), allergens (364; 386; 566) and toxins in virulence. Fungi that produce fruiting bodies such as A. fumigatus, demonstrate co-ordinated expression of biologically active, toxic and immunosuppresive

Chapter 4. Sub-telomere directed gene expression during initiation of invasive aspergillosis 135
secondary metabolites and spore-related products during development (109). Indeed, A.
fumigatus is equipped with a diverse arsenal of toxins ranging from specific inhibitors of the 28S ribosome (344; 345; 389) through natural steroid antibiotics (632; 470; 99), haemolytic proteins (184; 221; 377), aflatoxins (537; 510) and small molecular weight toxins such as gliotoxin (GT).
Gliotoxin is a mycotoxin that belongs to the group of epipolythiodioxopiperazines, which damages epithelial cells (67), is cytotoxic to mouse macrophages (731) impedes the function of ciliary cells (18), and inhibits the activiation of NADPH oxidase of polymorphonuclear leucocytes (701). This inhibitory action on a major killing mechanism employed by major effectors of cell-mediated defence, macrophages and neutrophils (611; 236), means that gliotoxin has been implicated in virulence (486; 526). Although the role of gliotoxin in virulence remains unclear (74; 145; 379; 652; 670), there has been renewed interest in the role of secondary metabolites in Aspergillus infection since the discovery of a global transcriptional regulator of secondary metabolite biosynthesis in this organism, LaeA (73). LaeA deletion does not affect growth or sporulation in media in vitro, but it does result in reduced virulence in mice. This impairment in the ability to cause infection can be correlated with a loss of detectable gliotoxin as well as mis-regulation of gene expression within 13 gene clusters (539). Siderophore-mediated iron uptake and storage, also partially under LaeA control (539) is indispensable for A. fumigatus virulence (618; 619) and adequate nutrient acquisition within the host niche is linked to pathogenesis of invasive disease being necessary to support sufficient growth to promote infection (281; 375; 374).While single gene deletion strategies have been informative in the study of Saccharomyces cerevisiae biology (749; 239), and to some extent in the elucidation of virulence traits in C. albicans and C. glabrata (341), there is little evidence that single gene knockout methods alone are informative in study of A. fumigatus virulence (reviewed in (394)) because virulence is multifactorial (36). In the absence of practical methods for disrupting multiple loci, transcriptional profiling studies can be informative. In this scenario, it is possible to identify genes that are adaptively transcribed or repressed in response to a perturbation in the culture environment. Indeed this has been adopted in a variety of in vitro environmental cues (alkaline stress, calcium stress, Omar Loss PhD thesis (423)). Moreover, even before microarray technologies were widely adopted in post-genomic fungal research, groups studying bacterial infection foresaw the potential of profiling the transcriptional changes within the host environment (637; 22). In vivo expression technology (IVET) was originally conceived upon the premise that most virulence genes are transcriptionally induced at one or more times during infection, and although certain host environmental parameters can be mimicked in vitro to induce a subset
of virulence genes, the full repertoire is only expressed in vivo. The renewed focus of attention on an important transcriptional regulator of secondary

4.1 Chapter Overview 136
metabolism, and the paucity of data obtained from single gene deletion methods turned our attention to profiling the transcriptional changes that occur during the early stages of A. fumigatus infection in an immunocompromised murine lung, and to compare the findings with existing in vitro data that can be viewed as components of the overall host environment. The feasibility of performing microarray analyses on limited sample material has been tested by a number of researchers employing linear amplification of mRNA, an approach which has recently proven successful in murine candidiasis (688), though a truly global transcriptional signature has yet to be reported for any fungal pathogen initiating mammalian invasive disease. The Eberwine method of mRNA amplification (183) involves reverse transcription of mRNA with an oligo dT primer bearing a T7 RNA polymerase promoter site, to direct in vitro transcription of antisense RNA after double stranded cDNA synthesis and is favoured for linear mRNA amplification from limited quantities of starting material. It provides the basis for the methodology employed in our study, and in the majority of reported instances where mRNA amplification has been applied to samples destined for microarray analysis (509). To identify fungal attributes preferentially employed during adaptation to the host niche, and thus contributing to the virulence of the saprophytic fungus A. fumigatus, we compared the transcriptomes of developmentally matched A. fumigatus isolates following laboratory culture or initiation of infection in the neutropenic murine lung. In this section, I report the employment of a methodology for global profiling of A. fumigatus gene expression in germlings rescued directly from the murine lung, a tool which will empower the analysis of virulence in this pathogen.
Our methodology employs simple molecular manipulations which, combined with cus-tom bioinformatic scripts based on the latest annotations of the A. fumigatus Af293 genome, mark a significant advance in understanding orchestration of fungal gene expression in vivo. Our analyses identify iron limitation, alkaline stress and nutrient deprivation as relevant host-imposed stresses during early-stage A. fumigatus infection and reveal a biased distri-bution of host-adaptation genes (relative to laboratory culture) in subtelomeric regions of chromosomes. Finally we assess lineage specificity of functions favoured during initiation of infection with other members of the genus Aspergillus.
This work was a collaborative effort, across several departments and universities. To place my work in context, and to maintain a narrative flow, findings from other co-authors are included, and appropriately cited. My contribution, as a joint first author to the manuscript - apart from a major contribution to the writing, was in the analysis of gene expression microarray data and in assessing the fidelity of log ratios following linear ampli-fication. This work was published in PLoS Pathogens (453).

Chapter 4. Sub-telomere directed gene expression during initiation of invasive aspergillosis 137
4.2 Specific Methods
4.2.1 A. fumigatus strains and growth conditions
In vitro isolates of A. fumigatus Af293 used in murine infection experiments were cultured in liquid YEPD medium, in an orbital shaking incubator (37°C, 150 rpm). For oxidative stress treatment, Aspergillus complete medium (CM) (144) was inoculated with 107 Af293 conidia and cultured in an orbital shaking incubator (37°C, 16 hours, 150 rpm). The resulting hyphal culture was transferred to CM containing 17 mM H202 and aliquots were harvested for RNA isolation either instantaneously (To) or after 1 hour of H202 exposure (T60). To impose acid stress, the same fungal burden (107 conidia) was incubated in liquid CM (37°C, 6 hours, 150 rpm) prior to sampling of two batches: a) a batch required for pH adjustment (pH 3.0) using HC1 and b) an unadjusted batch used as a reference. Aliquots of each were taken at the initiation of the subculture (To), and after 1 hour (T60), and culture material was used for RNA isolation and microarray expression analysis. Culture pH was verfied at the initiation and termination of the sub-culturing period. To impose alkaline stress, A. fumigatus germlings were cultured in an orbital shaking incubator for 16 hours in liquid AMM containing 100 mM glycolic acid pH 5.0, 10 ml / 1 vitamin solution (526), 5 mM ammonium tartrate and 1% w/v glucose. Germlings were filtered using Miracloth and shifted to similar, pre-warmed medium containing 100 mM Tris-HC1 pH 8.0. After 1 hour of incubation at 37°C, mycelia were washed with cold AMM (pH 5.0) and RNA extraction was performed immediately. For anaerobic stress treatment, hyphae (107 conidia) were statically cultured in 100 mm Petri dishes containing 20 ml of CM (16 hours at 37°C). Three plates were placed in anaerobic chambers and anaerobic conditions were established using 2 palladium catalysts (GasPak Plus, Becton Dickinson). A control plate was placed in a chamber without palladium catalysts and all plates were returned to a 37°C incubator for the duration of the experiment. Plates placed in anaerobic chambers were removed after 1 hour, the mycelium rapidly harvested on Miracloth, rinsed with ice-cold water and rapidly frozen in liquid nitrogen. The RNA was extracted and labelled cDNA was used in competitive hybridization with the equivalent material derived from the 16 hour control RNA sample. For neutrophil exposure assays, 107 germlings were initially cultured in 20 ml of RPMI 1640 (Sigma) with L-glutamine, 25 mM HEPES, and 5% (v/v) fetal bovine serum for 7 hours at 37°C in 100 mm Petri dishes, prior to addition of 107 human neutrophils for 1 hour, after which time cells were harvested for RNA isolation.For iron limitation treatment, A. fumigatus isolate ATCC46645 was grown for 15 hours at 37°C in Aspergillus minimal medium with depleted iron (AMM -Fe, iron depleted conditions) according to Pontecorvo (548) containing 1% (w/v) glucose as carbon source, 20 mM L-glutamine as a nitrogen source. After this time, 10 itM FeSO4 was added to the medium and germlings

4.2 Specific Methods 138
were harvested after a further 1 hour. Growth curve analyses were performed in triplicate using liquid YEPD, or AMM containing 1% glucose and either 5 mM ammonium tartrate or 5 mM hyroxyproline as nitrogen source. Cultures were inoculated to a final concentration of 5x 106 spores/ml and incubated under aerobic conditions at 37°C with shaking at 150 rpm. At selected timepoints mycelia were harvested on Miracloth, encased in Whatmann paper and dried at 37°C for 48 hours before weighing. Fungal growth is presented as the accumulation of biomass as a function of time. To prepare MLT medium we harvested uninfected murine lung tissue and homogenised the tissue in 100 ul of sterile distilled water per lung. The homogenate was added 70/30 v/v to a 2% water/oxoid No.1 agar mix and allowed to set prior to inoculation with A. fumigatus spores.
4.2.2 Murine Infections
Groups of 24 outbred male mice (strain CD1, 18-22 g, Harlan Ortech) were housed in indi-vidually vented cages and allowed free access to food and water. Infection was performed on day Do of the experiment and preceding steps are named in relation to this step e.g D_1 is one day prior to infection etc. Mice received cyclophosphamide (150 mg•kg-1), ENDOX-ANA, Asta Medica) by intraperitoneal injection on days D_3 and D_1 of the experiment. A single dose of hydrocortisone acetate (112.5 mg / kg), HYDROCORTISTAB, Sovereign Medical) was administered subcutaneously on day D_1. All mice received tetracycline hy-drochloride 1 mg / L and ciprofloxacin 64 mg / L in drinking water as prophylaxis against bacterial infection. Aspergillus spores for inoculations were grown on solid ACM, contain-ing 5 mM ammonium (-0-tartrate and 1% (w/v) Oxoid Agar number 3 for 5 days prior to infection. Conidia were freshly harvested using sterile saline (Baxter Healthcare Ltd. Eng-land) and filtered through MIRACLOTH (Calbiochem). Conidial suspensions were spun for 5 minutes at 3000g, washed twice with sterile saline, counted using a haemocytometer and re-suspended to a concentration of 2.5 x101° CFU / ml. Viable counts from administered inoculations were determined following serial dilution by plating on Aspergillus complete medium and growth at 37°C. Mice were anaesthetized by halothane inhalation and infected by intranasal instillation of 108 conidia in 40 ,u1 of saline solution. Groups of infected mice were culled and processed collectively during a 2 hour window corresponding to a time point 12-14 hours post-infection. Bronchoalveolar lavage was performed immediately after culling using three 0.5 ml aliquots of pre-warmed sterile saline. BALFs were snap frozen immediately following harvest using liquid nitrogen.

Chapter 4. Sub-telomere directed gene expression during initiation of invasive aspergillosis 139
4.2.3 RNA extraction and amplification
Prior to RNA extraction snap-frozen BALF samples were harvested by centrifugation, washed with 1 ml of ice-cold sterile water to lyse contaminating host cells and pooled. Following a further cycle of snap freezing pelleted pooled product was homogenized using a pestle and mortar and liquid nitrogen. RNA extraction was performed immediately using the filamentous fungi protocol of the RNeasy mini kit (Qiagen). RNA concentration was quantified on a Nanodrop spectrometer. RNA from reference samples was prepared from snap-frozen homogenized germlings or young mycelium.
4.2.4 Estimation of fidelity of transcript abundance following mRNA amplification
The corrected, multiple t—testing methodology of Nygaard (509) formed the basis of our investigation. However, for computational ease it was modified to use the limma linear modelling framework for the R statistical environment (562) (Supplementary Data Figure E.1).Spots were filtered as previously described (509) and systematic spatial effects were normalized within arrays using the print-tip lowess method (limma package for R (643)); no between-array normalization was performed in order to preserve the amplification-protocol variation existent between arrays. Least squares regression was used to fit the average log ratio, and error term of each transcript in each of the amplification protocols. The resulting coefficient vectors contained the mean relative abundance of each transcript (To v T60) in one of the three amplification protocols: aRNAri (amplified one round), aRNAr2 (ampli-fied two rounds) or totRNA (total RNA, unamplified). The amplification protocol effects and their associated Benjamini-Hochberg (290) corrected p-values (contrasts, contrast2 and contrast3 in Supplementarty Figure E.1) were extracted by fitting a contrast matrix to the original linear model. We defined the null hypothesis Ho as no difference in log ratios due to the amplification protocol, and an alternative hypothesis H1 as loss of log ratio fidelity due to the amplification protocol. Spots were partitioned into the three sets based on the Benjamini-Hochberg (290) adjusted p-values associated with each of the fitted contrasts. The rejected, undetermined and conservative sets were populated by genes whose adjusted p-values were 0<p<0.01, 0.01<p<0.1 and 0.1<p<1 respectively. This approach rejected 2325 (8.49%) of the spots in the aRNAi - totRNA comparison, i.e. the genes whose log ratios are significantly altered as a result of one round of mRNA amplification. The anal-ogous rejected sets for the aRNA2 - totRNA and aRNAi - aRNA2 comparisons contained 2604 (9.52%) and 73 (0.27%) rejected spots respectively. Comparison of the rejection sets showed that 80.69% of the spots in the aRNAi- totRNA rejection set were also present in the aRNA2- totRNA rejection set. This finding, in addition to the observed small number of genes in the aRNAi - aRNA2 rejection set, suggests that most of the amplification specific

4.3 Results 140
noise is introduced in the first round.
4.2.5 Microarray hybridisations
All experiments used Af293 DNA amplicon microarrays (499). Labelling reactions with RNA and hybridisations were conducted as described in the TIGR standard operating proceduresl. Independent verification of selected log2 ratios was achieved using quantita-tive RT-PCR (See Supplementary Figure E.3 and Supplementary Table E.12). Hybridised slides were scanned using the Axon GenePix 4000B microarray scanner and the TIFF images generated were analyzed using TIGR Spotfinder2 to identify poor quality spots. mRNA underwent two rounds of linear amplification by T7 promoter-directed transcrip-tion. Data processing and analysis was performed in the R statistical framework, using the limma, multcomp and fdrtool packages3. The intensity data retained after filtering (91.5% of the total spots) were normalized within (print-tip lowess) and between (scale) arrays. Least squares regression (1mFit) was then used to estimate a vector of regression coefficients (representing the mean loge (Cy5/Cy3) across the 5 biological replicates for the in vivo effect), the associated residuals and unadjusted p-values. Differentially regulated transcripts (Benjamini-Hochberg (290) corrected p-value < 0.05) were further categorized by the magnitude and direction of the fitted log ratios. Up-regulated and down-regulated transcripts were defined as having log2 (Cy5/Cy3) greater than an arbitrary threshold of plus, or minus, two respectively. Accuracy of microarray data was independently verified by quantitative RT-PCR on selected transcripts (See Figure E.3). Oligonucleotides used for this analysis are detailed in Table E.12). In vitro stress experiments were performed as previously described (499).
4.3 Results
4.3.1 A. fumigates RNA extraction and mRNA amplification
Microarray analyses are constrained by the availability of sufficient RNA for fluorophore labelling and hybridisation. One strategy to overcome the requirement for large quantities of material is global amplification of the sample. This approach has been employed for a number of reported microarray expression analyses (509) with favourable results. We chose to analyse an early time point of infection to facilitate separation of fungal cells from
ihttp://atarrays.tignorg
2http://www.tignorg/software/
3http://www.bioconductor.org

Chapter 4. Sub-telomere directed gene expression during initiation of invasive aspergillosis 141
those of the host with the advantage of studying a stage at which germinating hyphae have sensed, and are adapting to, their host. Early time points of A. fumigatus infection rep-resent a vulnerable phase of morphogenesis in vivo since epithelial invasion and formation of mycelial mass have yet to occur. They also mark a point at which diagnoses capable of distinguishing infection from carriage of fungal spores would be most desirable, and antifun-gal therapy most effective. We firstly characterised the time course of hyphal development in the sequenced clinical isolate Af293 by histopathological examination of infected neu-tropenic murine lung tissues (Figure 4.1). Lung sections collected and formalin-fixed at 4, 6, 8 and 12 hours post-infection contained numerous A. fumigatus spores in close associa-tion with murine epithelium in the bronchioles and alveoli (Figure 4.1 a). At 12 - 14 hours post-infection > 80% of A. fumigatus conidia had undergone germination and primary hy-phal production. We therefore performed all BAL extractions for downstream analyses on concurrently infected neutropenic mice within a two hour window of infectious growth cor-responding to 12 - 14 hours post-infection. At this time point recovery of germlings in BAL fluid was routinely achievable in the order of 103 germlings per lavaged lung (Figure 4.1 b).
4.3.2 Impact of mRNA amplification on preservation of transcript ratios
A suitable global mRNA amplification protocol should provide sufficient material for fluo-rophore cDNA labelling reactions whilst preserving the samples' original relative transcript abundance. To estimate amplification-related error (and distinguish such error from system-atic error inherent to microarray methodologies) a mock experiment was devised by Elaine Bignell to quantify the proportion of transcripts having significantly aberrant log2 ratios as a result of RNA amplification. This was achieved by indirectly comparing cDNA sam-ples from different amplification protocols in a statistical linear model and fitting relevant contrast matrices (see Materials and Methods and Figure E.1). For the mock experiment total RNA (totRNA) was isolated from two A. fumigatus cell populations To and T60, and subjected to either one (aRNAri) or two (aRNA,2) rounds of mRNA amplification prior to cDNA fluorophore labelling and microarray hybridisation (Figure 4.2). To evaluate whether ratios are preserved between amplification protocols, we adopted the approach of Nygaard et. at., (509) who used corrected gene-wise t-tests to identify differences in the mean log intensity ratios between aRNA2 and total RNA populations. In preparing the data for mul-tiple t—testing, we excluded spots that were flagged by the the TIGR spotfinder software, or where the intensity was lower than twice the background intensity in either the Cy5 or Cy3 channel. The proportion of excluded spots was used as an indication of the hybridis-ation quality of the slide as a whole. Slides hybridised with aRNA had fewer spots (13.49 - 18.22%) removed by the filtering process than did slides hybridised with cDNA (44.56 -48.95%), see Table E.2. This apparent amplification-related improvement in hybridisation

12 - 14 hours 12 hours
4 hours
8 hours
6 hours
12 hours
A
B C
4.3 Results 142
quality has been previously reported (508). In our study, this can be attributed to a greater
signal to noise ratio, and more specifically to increased foreground intensity on these slides
(Supplementary Figure E.4). We identified 8.49% of retained spots showing evidence of am-plification protocol dependent differences in the log2 ratios. Thus our estimated measures of
confidence from the above quality-control (QC) exercises fall within a range conducive for deriving biologically useful information, based upon reports of analytical studies published
to date (508) where Pearson correlation coefficients range from 0.75 to 0.99 (Figure 4.2)
and rejected gene sets approximate 10% of spots included in QC analysis.
Figure 4.1: Comparative time-course of A. fumigatus Af293 germination and hy-phal development in the murine lung, and laboratory culture. (A) Time-course of Af293 germination and hyphal development in the neutropenic murine lung. (B) Mi-croscopic appearance of Af293 germlings recovered from a typical single murine BALF, (harvested at 12-14 hours post-infection). (C) Microscopy of developmentally matched lab-oratory cultured Af293 germlings, following liquid culture for 12 hours in YEPD at 37°C. Infection/harvesting procedures and imaging performed by Elaine Bignell.

•i0
10 .10
0
011001
• 1 0
Ci
0
M RI2
Chapter 4. Sub-telomere directed gene expression during initiation of invasive aspergillosis 143
M Unamp
Figure 4.2: Correlation of log2 ratios resulting from comparative transcriptional analysis of the laboratory cultured A. fumigatus cell populations To and To under varying mRNA amplification protocols. Correlation of technically replicated log2 ratios between competitive hybridizations using single (aRNA1), double (aRNA2) and non-amplified (totRNA) RNA samples. (A and B) Correlation between log2 ratios obtained using highest correlations between slides using cDNA derived from amplified RNA (aRNA1 v aRNA2 r=0.74-0.8). Cross-protocol pairings (C) revealed highest correlations between slides using cDNA derived from amplified RNA (aRNA1 v aRNA2 r = 0.88-0.01). Surprisingly technical replicates of slides derived using cDNA derived from total RNA (totRNA r = 0.8) were comparable to cross-protocol pairings. Rejected sets are transposed onto correlation plots in Figure E.5

4.3 Results 144
4.3.3 A. fumigatus transcript profile during initiation of mammalian infection
A common reference design was adopted for the microarray experiment from which data were analysed and processed as described in the Materials and Methods section. We per-formed the infection experiment 5 times in total generating pooled (n = 24) BALFs from five independent Af293 infections (samples A to E, Table E.1). These samples were co-hybridised with cDNA derived from similarly amplified mRNA prepared from developmentally matched laboratory cultured Af293 germlings (Materials and Methods and Table E.1). Fluorescent signals from 9075 out of a possible 9516 represented ORFs were detectable from these hybridisation analyses (Supplementary Dataset E.1 and Array Expresso). Of 2180 genes (22.6% of the whole genome) having a fold-change in log intensity ratio of 2 or greater, 1281 were up-regulated and 897 were down-regulated. The entire expression dataset is graphi-cally represented in Figure 4.3 which plots log2 ratios against chromosomal locus. Initial interpretations of the dataset were performed by the Expression Analysis Systematic Ex-plorer (295) (EASE) to infer function by homology to Saccharomyces cerevisiae and identify over-represented Gene Ontology (GO) terms (35) among differentially expressed genes. The results of these analyses are partially listed in Table 4.1 (full listings in Supplementary Ta-ble E.3 and E.4). Distinct trends among favoured cellular processes are evident from these analyses which reveal a marked utilization by the host-adapting A. fumigatus cell, of genes encoding proteins involved in transport of metal ion, cation, carbohydrate and siderophore iron. Given the importance of iron acquisition for microbial pathogenesis in general, and the absolute requirement for siderophore biosynthesis during murine A. fumigatus patho-genesis in this model of infection (618; 619) we expected transcripts from genes involved in iron mobilization and transport to be differentially abundant in this analysis, relative to iron-rich laboratory culture media. Accordingly we could identify a minimum of eleven siderophore biosynthesis/transport genes as important during growth in the murine lung (Table 4.2) including two ferric-chelate reductases (Afu1g17270 and Afu6g13750).
Thirteen amino acid permease genes were more abundantly represented during host-adaptation than growth in YEPD (Table 4.2) including 4 4-aminobutyrate GABA (Afu8g01450, Afu7g0040, Afu5g14660 and Afu5g00710), and three proline permeases (Afu2g11220, Afu8g02200 and Afu7g01090) as well as the general amino acid permease, Gapl (Afu7g04290). Nine genes annotated as maltose permeases or transporters in the current Af293 annotation were also more abundantly represented during initiation of murine infection (Table 4.2). Extracellular proteases have been implicated as virulence factors in invasive aspergillosis, as well as antigens causing inflammatory irregularity during allergic A. fumigatus disease (394). Our analysis identified increased abundance of transcripts from the elastinolytic
4http://www.ebi.ac.uk/microarray-as/aer/#?ae-main0AccessionnumberE-TABM-327

Chapter 4. Sub-telomere directed gene expression during initiation of invasive aspergillosis 145
metalloprotease (Mep) (Afu8g07080), an aorsin-like serine protease (Afu6g6g10250) and three dipeptidylpeptidases (Afu4g09320, Afu2g09030 and Afu3g07850). Thus transcription of this subset of A fumigatus proteases is significantly higher in the murine lung relative to rich laboratory culture. Functional categories of ergosterol biosynthesis, haem biosynthe-sis and aerobic respiration were significant among genes underrepresented during infection, relative to laboratory culture (Table 4.1) as well as multiple functional categories represent-ing ribosome biogenesis and assembly, and protein biosynthesis and processing. This may reflect the poor nutritional value of murine lung relative to YEPD and/or reduced growth (due to any number of stresses) during host-adaptation compared to broth culture. This trend is evidenced on multiple levels within our dataset, comprising repression of genes directing ribosomal protein synthesis, rRNA synthesis, RNA polymerase I and II activity, translation initiation and elongation, tRNA processing and synthesis, intracellular traffick-ing, secretion and vesicular trafficking (Table 4.1 and Supplementary Table E.3 and E.4). While such metabolic dampening is often observed in microbial systems under stress, the observed A. fumigatus regulatory signature mimics that of rapamycin-mediated TOR ki-nase (111) inhibition and typifies fungal starvation. The S. cerevisiae TOR proteins TOR1p and TOR2p are phosphatidylinositol kinase homologues, first identified as the targets of the immunophilin-immunosuppressant complex FKBP-rapamycin (280), combined deletion of which causes yeast cells to arrest growth, undergo a reduction in protein synthesis, accu-mulate the storage carbohydrate glycogen and acquire thermotolerance. Comparison of our dataset to that obtained following rapamycin-induced TOR inhibition in S. cerevisiae (111) reveals extensive overlap in induced (n = 35) and repressed (n = 90) homologous genes be-tween the two datasets (Tables E.5 and E.6). Thus a clear TOR repression-like starvation signature, relative to laboratory culture, is observable during early-stage infection, which may derive from the relative nutritional status of the tested conditions and/or slower growth within the context of our experiment. The indicated cellular down-turn in metabolism ob-served is strongly countered by up-regulation of genes encoding functions associated with amino acid and carbohydrate catabolism (Tables 4.1 and E.3 and E.4). These analyses were performed by myself and Elaine Bignell.

4.3 Results 146
Table 4.1: Over-represented Gene Ontology terms among differentially expressed genes. Table lists selected biological processes significantly over-represented among dif-ferentially expressed genes, with respect to their occurrence in the A. fumigatus Af293
genome. To identify over-represented Gene Ontology terms, loci having significantly dif-ferent expression were analyzed by the Expression Analysis Systematic Explorer (EASE)
(PMID:14519205), which is implemented in MEV within the TIGR TM4 microarray data
analysis suite. Numbers of genes in the indicated Gene Ontology categories were subjected to statistical analysis by EASE to identify categories overrepresented compared with the whole genome data set. Only categories with Fisher's exact (FE) test probabilities < 0.05
were included. Induced genes are represented above the double horizontal line. Repressed genes are represented are represented below the double horizontal line. Full results of the
analysis can be found in Table E.3 and E.4
GO ID GO Term List hits List size Pop hits Pop size FE
GO:0006810 transport 133 448 761 4219 1.08 x 10-10
GO:0008643 carbohydrate transport 19 448 40 4219 1.08 x 10-19
GO:0009063 amino acid catabolism 14 448 35 4219 3.41X10-9
GO:0044242 cellular lipid catabolism 10 448 19 4219 5.37x10-6
GO:0044270 nitrogen compound catabolism 15 448 42 4219 6.30x10-6
GO:0030001 metal ion transport 19 448 74 4219 1.28X10-5
GO:0015892 siderophore-iron transport 5 448 8 4219 1.76x10-4
GO:0006812 cation transport 21 448 94 4219 5.63x10-4
GO:0019541 propionate metabolism 3 448 4 4219 6.45x10-4
GO:0006830 high-affinity zinc ion transport 2 448 2 4219 4.38x10-2
GO:0006631 fatty acid metabolism 13 448 62 4219 1.17 x 10-2
GO:0019538 protein metabolism 174 561 781 4219 7.35x10-15
GO:0042254 ribosome biogenesis and assembly 51 561 161 4219 5.56 X 10-19
GO:0044237 cellular metabolism 366 561 2251 4219 6.51 x 10-19
GO:0007046 ribosome biogenesis 46 561 140 4219 1.08 X 10-9
GO:0016072 rRNA metabolism 41 561 118 4219 1.37X10-9
GO:0006364 rRNA processing 40 561 115 421.9 2.12x10-9
GO:0044238 primary metabolism 334 561 2028 4219 3.30 x 10-9
GO:0006457 protein folding 23 561 50 4219 1.48x10-8
GO:0006412 protein biosynthesis 72 561 302 4219 1.63x10-7
GO:0006783 heme biosynthesis 6 561 13 4219 4.03x10-3
GO:0009060 aerobic respiration 13 561 48 4219 8.02 x 10-3
GO:0006696 ergosterol biosynthesis 11 561 38 4219 8.45X10-3

Chapter 4. Sub-telomere directed gene expression during initiation of invasive aspergillosis 147
Table 4.2: Functional classification of selected genes with altered transcript abun-dance, relative to laboratory culture, in the murine lung.
GO ID GO Term List hits FE Ergosterol and Heme Biosynthesis
-2.07 Afu1g05720 c-14 sterol reductase -2.14 Afu1g03950 cytochrome P450 sterol C-22 desaturase -2.18 Afu8g07210 hydroxymethylglutaryl-CoA synthase -2.30 Afu5g02450 farnesyl-pyrophosphate synthetase -2.35 Afu7g03740 14-alpha sterol demethylase Cyp51I3 -2.90 Afulg07140 c-24(28) sterol reductase -2.92 Afulg03150 c-14 sterol reductase -2.97 Afu2g00320 sterol delta 5 -3.38 Afu6g05140 sterol delta 5 -3.42 Afu5g14350 c-24(28) sterol reductase -3.93 Afu4g06890 14-alpha sterol demethylase Cyp51A -4.40 Afu4g09190 S-adenosyl-methionine-sterol-C- methyltransferase -4.30 Afulg07480 coproporphyrinogen III oxidase -2.44 Afu5g06270 5-aminolevulinic acid synthase -2.13 Afu5g07750 ferrochelatase precursor -3.02 Afu6g07670 cytochrome c oxidase assembly protein cox15
Iron acquisition 2.39 Afu7g06060 siderochrome-iron transporter (Sitl) 2.55 Afu7g04730 siderochrome-iron transporter 3.97 Afu6g13750 ferric-chelate reductase 6.11 Afu3g03440 MFS family siderophore transporter 3.40 Afu4g14640 low affinity iron transporter 2.64 Afu3g03650 sidG 6.45 Afu3g03640 siderochrome-iron transporter (MirB) 6.14 Afu3g03420 sidD 6.45 Afu3g03400 siderophore biosynthesis acetylase Acel (sidF) 3.41 Afu3g03390 siderophore biosynthesis lipase/esterase 4.85 Afu3g03350 nonribosomal peptide synthase (sidE) 4.01 Afu3g01360 siderochrome-iron transporter 5.29 Afulg17270 ferric-chelate reductase (Fret) 2.66 Afulg17200 nonribosomal peptide synthase (sidC) 2.61 Afu8g01310 metalloreductase (FRE1)
Nitrate assimilation -4.05 Afu1g12840 nitrite reductase -0.30 Afu1g12850 nitrate transporter (nitrate. permease) -2.33 Afu1g12830 nitrate reductase NiaD 0.47 Afu5g10420 nitrate reductase 7.06 Afu1g17470 high affinity nitrate transporter NrtB 0.56 Afu6g13230 Nit protein 2
Carbohydrate transport 5.70 Afu7g06390 maltose permease 5.88 Afu7g05190 maltose permeate 4.69 Afu6g11920 maltose permease 2.72 Afu5g00500 maltose permeate 4.75 Afu3g01700 maltose permeate 2.92 Afu2g10910 maltose permease 2.61 Afu1g03280 maltose permease 3.20 Afu3g03380 maltose 0-acetyltransferase 2.56 Afu8g07070 maltase 4.68 Afu7g06380 maltase 3.48 Afu4g00150 MFS maltose transporter 2.24 Afu8g07240 MFS maltose permease 3.08 Afu6g01860 MFS lactose permeate 2.08 Afu1g17310 MFS lactose permease 4.35 Afu3g01670 MFS hexose transporter 4.69 Afu2g08120 MFS monosaccharide transporter (IIxt8) 3.31 Afu5g14540 MFS monosaccharide transporter
continued on next page

4.3 Results 148
continued from previous page GO ID GO Term List hits FE
4.86 Afu4g00800 MFS monosaccharide transporter 5.06 Afu7g00780 MFS monocarboxylate transporter 2.77 Afu3g03320 MFS monocarboxylate transporter 3.04 Afu3g03240 MFS monocarboxylate transporter 2.81 Afu6g03060 monosaccharide transporter 3.47 Afu5g01160 monosaccharide transporter 3.77 Afu4g13080 monosaccharide transporter 2.81 Afu7g05830 MFS sugar transporter 6.97 Afu6g14500 MFS sugar transporter 2.05 Afu5g06720 MFS sugar transporter 4.47 Afulg11050 MFS sugar transporter 2.42 Afu3g12010 high-affinity hexose transporter 2.89 Afu3g00430 high-affinity glucose transporter
Nitrate assimilation -4.05 Afu1g12840 nitrite reductase -0.30 Afulg12850 nitrate transporter (nitrate permease) -2.33 Afu1g12830 nitrate reductase NiaD 0.47 Afu5g10420 nitrate reductase 7.06 Afu1g17470 high affinity nitrate transporter NrtB 0.56 Afu6g13230 Nit protein 2
Secreted Proteins 7.40 Afu5g14190 beta-glucanase 5.90 Afulg17510 lipase/esterase 5.64 Afu2g09380 cutinase 5.50 Afu8g07090 extracellular proline-serine rich protein 5.07 Afu2g05150 cell wall galactomannoprotein Mpg 5.01 Afu5g00540 extracellular signaling protein FacC 4.97 Afu7g01180 extracellular lipase 4.93 Afulg16250 alpha-glucosidase B 4.70 Afu3g14030 alkaline phosphatase 4.68 Afu2g00490 glycosyl hydrolase 4.68 Afu7g06380 maltase 4.66 Afu8g01050 lipase/esterase 4.63 Afu3g14910 extracellular signalling protein (factor C) 4.63 Afu8g01130 alpha-galactosidase C 4.58 Afu4g01070 acid phosphatase 4.54 Afu7g05610 glucanase 4.32 Afu4g00870 antigenic cell wall galactomannoprotein 4.07 Afu6g02980 extracellular exo-polygalacturonase 3.88 Afu8g04710 xylosidase 2.18 Afu3g07850 dipeptidyl aminopeptidase Stel3 3.50 Afu2g09030 secreted dipeptidyl peptidase 3.82 Afu6g11500 dipeptidase
Antigens 5.64 Afu4g09320 antigenic dipeptidyl-peptidase Dpp4 4.32 Afu4g00870 antigenic cell wall galactomannoprotein 7.13 Afu4g09580 major allergen Asp F2 6.44 Afu8g07080 elastinolytic metalloproteinase Mep 3.22 Afu6g10250 alkaline serine protease AorO
Carbohydrate and/or protein glycosylation 0 2.54 Afu8g02020 glycosyl transferase 6.10 Afu4g14070 glycosyl transferase 2.23 Afu5g00670 glycosyl hydrolase family 35 2.27 Afu6g11910 glycosyl hydrolase family 3 3.18 Afu4g00390 glycosyl hydrolase 2.07 Afu2g03270 glycosyl hydrolase 4.68 Afu2g00490 glycosyl hydrolase 2.14 Afu2g03120 cell wall glucanase (Utr2) 3.08 Afu8g05610 cell wall glucanase (Scw11) 5.07 Afu2g05150 cell wall galactomannoprotein Mp2
Carbohydrate catabolism 2.83 Afu6g14490 beta-glucosidase
continued on next page

Chapter 4. Sub-telomere directed gene expression during initiation of invasive aspergillosis 149
continued from previous page GO ID GO Term List hits FE
2.11 Afu3g00230 beta-glucosidase 7.40 Afu5g14190 beta-glucanase 3.17 Afu5g14550 beta-galactosidase 2.18 Afulg14170 beta-galactosidase 7.16 Afu7g06140 beta-D-g]ucoside glucohydro]ase 3.59 Afu6g08700 beta glucosidase 3.25 Afulg16700 beta galactosidase 4.93 Afu1g16250 alpha-glucosidase B 3.83 Afu4g10150 alpha-glucosidase 4.63 Afu8g01130 alpha-galactosidase C 6.51 Afulg01200 alpha-galactosidase 2.87 Afu8g07300 alpha/beta hydrolase 4.46 Afu8g00570 alpha/beta hydrolase 2.44 Afu8g00530 alpha/beta hydrolase 2.04 Afu7g00830 alpha/beta hydrolase 5.21 Afu3g01280 alpha/beta hydrolase
Amino acid transport 4.72 Afu7g04290 amino acid permease (Gapl) 4.05 Afu2g08800 amino acid permease (Dips)
amino acid permease 3.73 Afu8g06090 amino acid permease
2.96 Afu2g10560 amino acid permease 3.27 Afulg09120 amino acid permease 2.96 Afu8g02200 proline permease 5.71 Afu7g01090 proline permease 3.38 Afu2g11220 proline permease 4.19 Afu8g01450 GABA permease 2.39 Afu7g00440 GABA permease 2.91 Afu5g14660 GABA permease 6.28 Afu5g00710 GABA permease 6.73 Affilg14700 allantoate transporter
Metal ion transport/homeostasis 4.97 Afu5g09360 calcineurin A 5.71 Afu7g01030 Calcium-transporting ATPase 1 (PMC1) 3.30 Afu3g10690 calcium-translocating P-type ATPase(PMCA-type) 2.25 Afu3g08540 Ca2+ binding modulator protein (A1g2) 2.77 Afu6g00470 plasma membrane zinc ion transporter 4.26 Afu5g03550 plasma membrane H(+)ATPase 3.26 Afu1g02480 plasma membrane ATPase 4.13 Afulg01550 high affinity zinc ion transporter 3.10 Afu8g01890 Na+/H-I- exchanger family protein 3.32 Afu7g04570 Na/K ATPase alpha 1 subunit
Oxidative stress resistance 4.27 Afu1g14550 Mn superoxide dismutase MnSOD -0.71 Afu4g11580 Mn superoxide dismutase (SodB) 2.34 Afu8g01670 bifunctional catalase-peroxidase Cat2

4.3 Results 150
Table 4.3: Distribution of induced and repressed genes among lineage specific cohorts and chromosomal locations. x2 analysis (with one degree of freedom) was used to test the distribution of induced and repressed genes with respect to lineage specificity and sub-genomic locations, based upon the null hypothesis that equal numbers of induced and repressed genes occur in each cohort. Asterisked gene populations are listed in Tables E.7, E.8, E.9, E.10. Table entries above the horizontal line denote lineage statistics; table entries below the horizontal line denote chromosome location statistics. Key to abbreviations: n = number of genes; n de = number differentially expressed; Induced obs = observed number of genes induced; Repressed obs = observed number of genes repressed; Induced E = expected number of induced genes; Repressed E = expected number of repressed genes.
rt. nd, Induced obs Down obs Induced E Repressed E X2 Aspergillus core 5095 965 414 551 483 483 19.45 < 0.0001 Affc-core 6907 1468 791 677 734 734 8.86 0.0029
Affc-specific 180 64* 54 10 32 32 30.25 < 0.0001 Mu-specific 428 135* 77 58 68 68 2.68 0.102
0-300 kb 1326 470 367 103 235 235 148.289 < 0.0001
301-600 kb 1504 365 229 136 182.5 182.5 23.696 < 0.0001
>600 5779 1345 686 659 672.5 672.5 0.5 0.4795
4.3.4 Lineage specificity and locational analyses of differentially expressed genes
From an evolutionary perspective, relative proportions of genes being over- and underrepre-sented in the analysis differed significantly within lineage-specific gene cohorts (Figure 4.4 A and Tables E.7, E.8, E.9, E.10). Genes having increased transcript abundance during infection are significantly enriched (p<0.0001 by chi-square analysis, see Tables E.7, E.8, E.9, E.10) among differentially expressed genes (n = 64) unique to the A. fumigatus lin-eage. Thus 93.6% of A. fumigatus genes having orthologues restricted to two very closely related, but differentially virulent, species Neosartorya fischeri (anamorph of Aspergillus fis-cherianus) (Genbank sequenced genome identifier AAKE00000000) and Aspergillus clavatus (Genbank sequenced genome identifier AAKD00000000) are more abundantly represented during the initiation of infection (Supplementary Data Tables E.7, E.8, E.9, E.10). In contrast only 8% of genes having orthologues in all six Aspergillus species sequenced to
date (i.e. the Aspergillus core genome) are more abundantly represented under these con-ditions (n = 5095). This invariable core genome encodes many functions associated with information processing, central metabolism and cell growth, retention of which is most likely to be essential for cellular survival (197). Narrowing the phylogenetic sampling to include only A. fumigatus and its relatives N. fischeri and A. clavatus distinguishes several subtelomeric genomic islands upon which phenotypic variation between species, including differing pathogenicity, might depend (197). Accordingly we found differentially expressed genes to be unevenly distributed amongst A. fumigatus chromosomes (Figure 4.4 B and

.J M Cr Lin
F
CO
ii S
a
r
• or:
7=4
Chapter 4. Sub-telornere directed gene expression during initiation of invasive aspergillosis 151
Figure 4.3: A genome-wide transcriptional snapshot of A. fumigates A1293 dur-ing intiation of murine infection. Red and green horizontal lines correspond to individ-ual up- and down-regulated genes, respectively. Thin light gray horizontal lines indicate the positions of all other genes. (SM) and (aspcore) are density graphs of secondary metabo-lite and Aspergillus-core genes, respectively, expressed as a percentage of the total bases contained per gene type, per non-overlapping 2 kb of chromosomal sequence. Induced and repressed gene clusters, are depicted by red and green rectangles, respectively, below each chromosome. A complete listing of genes housed in these co-regulated clusters can be found in Table S6. Light blue/gray horizontal bars represent putative centromeres and the pink horizontal bar in chromosome 4 represents a region of ribosomal DNA. Figure produced by Jonathon Crabtree. Higher resolution figure available at http: //ukpmc .ac. uk/articlerender.cgi?artid=16630688mendertype=figure&id=ppat-1000154-g003

4.3 Results 152
Aspergillos core Affc-core Affc-specific
Afu-specific A n=5095 n - 6907 n = 180 n = 428 •
> 600 kb
301 - 600 kb
0 — 300 kb
(Distal)
(Intermediate)
(subtelomeric)
B n = 6494 n = 1634 n = 1509
Increased transcript abundance
II Decreased transcript abundance
Figure 4.4: Distribution of lineage specific and telomere-proximal genes among differentially expressed host adaptation dataset. (A) Lineage specificity of A. fu-migatus genes having altered transcript abundances, relative to laboratory culture, in the murine lung. The Aspergillus-core (Asp-core) set contains A. fumigatus Af293 proteins that have orthologues in A. clavatus (AAKD00000000), N. fischeri (AAKE00000000), As-pergillus terreus NIH2624 (AAJNO1000000), Aspergillus oryzae RIB40 (431), A. nidulans FGSC A4 (223) and Aspergillus niger CBS 513.55 (534). The Affc-core set were defined as A. fumigatus Af293 proteins that have orthologues in N. fischeri and A. clavatus. The Affc-unique set is a sub-set of Affc-core proteins that do not have orthologues in A. terreus, A. oryzae, A. nidulans or A. niger. Asterisks indicate gene sets which are listed in Table E.7, Table E.8, Table E.9 and Table E.8. Underlined values significantly deviate from the null hypothesis that an equal number of induced and repressed genes will occur in each cohort, as estimated by Chi-square analysis (Tables E.7, E.8, E.9, E.10). (B) Chromoso-mal distribution of A. fumigatus genes having altered transcript abundances, relative to laboratory culture, in the murine lung. Distances from telomeres (kb) are noted above pie charts. Asterisked gene sets are listed in Supplementary Table Table E.7, Table E.8, Table E.9 and Table E.8. Underlined values significantly deviate from the null hypothesis that an equal number of induced and repressed genes will occur in each cohort, as estimated by Chi-square analysis (Tables E.7, E.8, E.9, E.10).

7 0 0 11
136 13 89 1026
465
Nitrogen (626)
4
Nouse (1262)
104
1 0
Chapter 4. Sub-telomere directed gene expression during initiation of invasive aspergillosis 153
Alkaline (211)
Figure 4.5: Overlap between murine adaptation and in vitro stress datasets. Venn diagrammatic representation of overlap between murine adaptation dataset and those of nitrogen starvation, iron starvation and alkaline shift. Genes are listed in Dataset E.3. Figure produced by Elaine Bignell.

I
4.3 Results 154
. I. 0 0.0 1.0
.0.40011/0 CO < nptttr.****** IssA loll'/••r«...• IS., pfti•savo-
-11.1,1.01020 .001 min4 Of 4.•••.e. p .*• • Pals0001010 CI tronstraptist ****** Rosh
..... y1•0001 , erg spi....e• -P.C,..1.0111. C. h.,. p • At014111160 p
nn. • P. • • -1,11• ....... 11.0.011000 pte•sph..1 ,11....• 01.0,
1101.1.001,0 Pvp•oksts•.1 p ..... C1..em.. .. .. p•ststsvo-
-P.tiol.01006 sist ..... CUBA, - P.0•6111 SOO•*.1 MO •wh!•0$201 •14,101.1,4 • 'MIDS./MONO. elver! 4strartfor as. relative 040/44/10 AaphatooLyaan "Atts2,1114,0 plan. isnekbo.so• Loy sies.” sine \IC 1111, pritistivo• .1111.4,101/10 sdposhleir 141111' 0iNs401140 .014kattas• GIs? A64100,0 .Atialt341.40 tate ttoolt p Mtp1014sp42, t ..... At01,0,2:0 O•phospho1.c1•,•2-Iti.s• I A(.9140/10 4' ID. atm• ...live satire foal, p .114•44.11/40 111 .. .... .41 • '1\1'06,11110 t isrllsep p , p 11/00014$0 ••.4 hr.thetseal p '1fir41.09110 ellests.me e p Ws Cepa pacts.... .M•74454,0 TN alopeatent ematosoductos• potats.e. -10404401410 aleatal 4,0.14rager.ase rat A14104.41•0 • oniart.4 47.0044" al protest 111...1,032110 rv.1 h47oths1seal p 1e.a0/01110 CO 1.01/feirciot Ika A00101))00 n11.wJ 111141tipl,N4 vita. 0140 Atuls1104110 cor.sairosi hrp•stlieSse01 •Ilfatt11140 Ill frail, fcerac•eh.lass pecntsve '4(01032f:0 ..ph.ssylis. 4l.pW0ph1, pa...044 • 1halil1110 ,,,..reed typotItati 1101 riototn .04•0243/00 ... E•etn.l ornsChane eases. p P.,0011.1, Il(0f102110 ..).se red Afss1013110 ..Afr494113110 Cs s ram, p.y.4.s.ne• -kta,c440441 .....• ehroweisse isti. pslibtivo. Affulp1.191141 se Cr 4,•,1,4th A(01e01930 4411•10.r.disup p.t•se. lade )144400) )40con...m.4 Wats ta col .14.441411100 00.*. I see A720-13.11. -Ate4041140 st.lertettaat•I art I eaa., , pa) atlaa• •Afal4111141 ler.r.ctear..fatei-aszAteak Its.. 11(000)420 avast...al
40 1 /SO alpta-1, I- gllat r... a, mutat as ...... -0!.144142 sa.4.eoptawe 01•11,01,t1.1 .4)1 1 ase &eat , pa AC•144/3430 ADC Issesp•es. 1.1T Aess140/410ateli ..... (malt. ..... it 1111•2/01440 1-..11.es - oat ...... 'Maly) )440 00: cilarepho.s• asset tests... pulals.- 'A6001411 Alf sults:A., Soars, keta)03640 1gf3 ..lerochsome •gen !ramparter eel
Figure 4.6: Comparative analysis of A. fumigatus gene expression datsets.. A Cross experimental comparison of A. fumigatus gene expression aligning log2 ratios ob-tained during host adaptation (mice); exposure to neutrophils (neut), increased expression in parental strain versus AlaeA mutant, acid shift (acid), iron starvation (iron), oxygen depletion (anaer) and oxidative stress (H202) for various genes. The colour bar indicates the range of log2 expression ratios; grey bars indicate genes from which signals were un-detectable for technical reasons. Experimental conditions are described in Materials and Methods. LaeA dataset is taken from Perrin et. al. (539). Comparative analyses were implemented in TM46. Figure produced by Natalie Federova.

A
Rad
ial g
row
th (
mm
) 40
35
30
25 -
20 -
15-
10-
YPO
MLT
5
0
WA
Day 1 Day 2
Day3
Day 4 Day 5 Day 6 Day 7
B 0.80
0.70
0.69
E 0.50
GAO
0.30
50 150 100
- -a- • - YPD —t MM+NH4
•—MM+Hydroxyproline
Chapter 4. Sub-telomere directed gene expression during initiation of invasive aspergillosis 155
Time
Figure 4.7: Characterisation of A. fumigates growth, relative to YEPD. (A) Com-parative analysis of Af293 radial growth on YEPD and synthetic murine lung tissue medium (114LT). Triplicated, spot-inoculated plates containing single 100 spore inocula were incu-bated at 37°C. (B) Growth curve analysis of Af293, performed in triplicate using liquid YEPD, or AMM containing 1% glucose and either 5 mM ammonium tartrate or 5 mM hy-roxyproline as nitrogen source. Cultures were inoculated to a final concentration of 5 x 106 spores/ml and incubated under aerobic conditions at 37°C with shaking at 150 rpm. At selected timepoints mycelia were harvested on Miracloth, encased in Whatmann paper and dried at 37°C for 48 hours before weighing. Experiments and figures produced by Timothy Cairns

23 genes in secondary metabolism clusters
Subtelomeric (1509)
23 subtelomeric
4.3 Results 156
Up in vivo Up in wt versus A/aeA
(1266)
(415)
Up in vivo Up in WT versus laeA (1282) (415)
Secondary metabolism clusters (209 [133 subtelomeric])
Figure 4.8: Expression of LaeA-regulated genes during initiation of murine infec-tion. Venn diagram representation of overlap between genes repressed in AlaeA (539) and those having increased transcript abundance during murine infection, according to propor-tions having subtelomeric locations, and secondary metabolism functionality (on the basis of annotation).

Chapter 4. Sub-telomere directed gene expression during initiation of invasive aspergillosis 157
Supplementarty Data Tables E.7, E.8, E.9, E.10). Induced genes form a significantly in-creased proportion of differentially regulated functions in intermediate (p < 0.001) and subtelomeric (p < 0.001) regions of the chromosomes (Figure 4.4 B and Supplementary Data Tables E.7, E.8, E.9, E.10). While only 16% of the predicted A. fumigatus gene reper-toire is housed within 300 kb of telomeres (classed as the subtelomeric gene repertoire in our analyses), 29% of transcripts having increased abundance, relative to laboratory cul-ture, in the murine lung are located to such subtelomeric areas, compared to just 11% of down-regulated transcripts. Moreover, 28% of the entire subtelomeric gene repertoire is represented in the induced dataset compared to only 8% of subtelomeric genes represented among down-regulated functions (Supplementary Data Tables E.7, E.8, E.9, E.10). These analayses were performed by Natalie Federova.
4.3.5 Clustering of induced genes
Regarding our A. fumigatus gene expression dataset as a function of chromosomal locus (Figure 4.3) we discovered that many induced A. fumigatus genes are found in contigu-ous clusters. To investigate this further a custom script, developed by Jonathon Crabtree, was used to automate cluster identification. This identified numerous genomic loci within which co-ordinate regulation of a minimum of 5 closely neighbouring genes can be observed (Figure 4.3 and Table S6). Co-ordinated expression of physically clustered genes is a promi-nent feature of the induced, but not repressed, gene set and we observe a large proportion (40%) of up-regulated physically clustered genes to reside within 300 kb of chromosome ends (Figure 4.3 and Table 4.4). The clusters are comprised of up to 34 co-ordinately expressed genes and include loci directing biosynthesis of siderophores (cluster 33) and two known sec-ondary metabolites, pseurotin and gliotoxin (clusters 72 and 60, respectively). The gliotoxin biosynthetic cluster is not subtelomerically located, being 700 kb from the telomere (as annotated by Perrin et. al. (539)). The pseurotin biosynthetic cluster, however, (as anno-tated by Maiya et. al. (439)) is contained within the fumitremorgen cluster (Afu8g00100-Afu8g00720) at 100 kb from the telomere (539). Pseurotin (439; 367; 277) is a neuritogenic, nematicidal quinone and gliotoxin an immunotoxin which supports A. fumigatus virulence in some murine models of invasive pulmonary aspergillosis (74; 145; 379; 652; 670). We observed four other postulated, but uncharacterised, secondary metabolite gene clusters in-duced during early stage A. fumigatus infection, including a large proportion of genes on the left arm of chromosome 8, predicted to encode a fumitremorgen biosynthesis supercluster (75). Thus, it would seem that selective expression of a subset of secondary metabolite loci facilitates initiation of mammalian infection. While gliotoxin biosynthesis is dispensable for virulence in some murine models, our analysis demonstrates that this host environment is nonetheless conducive to immunotoxin production, and further insights on virulence mech-

4.3 Results 158
anisms relevant to neutropenia and/or corticosteroid therapy await comparative analyses of fungal gene expression in each of these strikingly different host settings, an analysis which is currently underway in our laboratory. A fully annotated table of clusters, indicating between-species synteny within cluster loci, and all accession numbers, can be viewed in Supplementary Dataset E.2.
4.3.6 Comparative analyses of murine adaptation- and in vitro gene expression datasets
The complexity of the transcriptional signature derived from host adaptation analyses is likely to originate from convergence of multiple environmental cues, coupled with metabolic and morphologic effects. Although functional categorisation of differentially expressed func-tions (Tables 4.1, E.3 and E.4) could identify metabolic and physiological trends during initiation of infection, environment-related signatures were less easy to discern. To ascer-tain physiologically relevant features of the host environment we compared the transcrip-tome of host-adapting germlings to those of in vitro A. fumigatus cultures exposed to iron limitation, nutrient limitation, alkaline stress, acid stress, neutrophils, oxidative stress or anaerobic stress. The resulting transcriptomic responses varied in magnitude (Table 4.4) but assignment of cut-off log2 ratio values of +2 and -2 across all of the analyses enabled us to distinguish several important aspects of the A. fumigatus host-adaptive response (Fig-ure 4.5). The transcriptional signatures of paramount importance among those examined were alkaline adaptation, iron deprivation and nutrient starvation, which are remarkably prominent in the infection dataset. Using the aforementioned log2 cut-offs, 24 of 43 iron-regulated genes were identified as differentially expressed during host adaptation, of these 18 were more abundantly represented (Figure 4.5 and Supplementary Dataset E.3), and 6 less abundantly represented.

Chapter 4. Sub-telomere directed gene expression during initiation of invasive aspergillosis 159
Table 4.4: Comparative analysis of genetic distribution among genes having dif-ferential transcript abundances in microarray analyses using A. fumigatus RNA following exposure to in vitro or murine-adaptive stress, or laeA gene deletion. Gene density in telomere-proximal (0 - 300 kb from telomeres), intermediate (300 - 600 kb from telomeres) and telomere-distal (>600 kb from telomeres) regions of the A. fumiga-tus genome are indicated (Genome stats); (no sec mets) indicates omission of secondary
metabolite biosynthetic genes (identified as detailed in the Materials and Methods section) from the tested dataset.
Experimental condition Total genes in dataset Proximal % Intermediate % Distal % A. fumigates-specific % Genome stats 9632 1509 16 1634 17 6494 67 2201 23
Clustered in vivo up 658 227 34 129 20 302 46 218 33 AlaeA up 415 102 32 59 14 254 61 122 29 in vivo up 1282 367 29 229 18 686 54 374 30
Nitrogen starvation up 626 180 29 115 18 331 53 33 33 In vivo up (no sec mets) 1196 327 27 223 19 646 54 355 30
Neutrophils 60 min up 312 83 27 81 26 148 47 80 26 Alkaline adaptation up 211 53 25 39 18 113 54 6 6
AlaeA up (no sec mets) 318 70 22 58 18 190 60 50 16 Acid stress up 83 18 22 21 25 44 53 43 52
Iron limitation up 28 4 14 4 15 20 71 8 30 AlaeA down 528 93 18 99 19 336 64 108 20
Oxidative stress up 54 9 17 9 17 36 67 22 54 In vivo down 898 103 11 136 15 659 73 126 14
Relaxing the cut-off criterion to encompass all differentially regulated genes in the host
adaptation dataset allowed complete capture of the iron regulon (Figure 4.6). Among these genes are the siderophore biosynthetic genes sidA (Afu2g07680), sidD (Afu3g03420) and sidC (Afu1g17200), the latter two discerning biosynthesis of both intra- and extracel-
lular siderophores during infection as substantiated by previous findings (619), and four siderochrome/siderophore transporter proteins Afu7g06060, Afu7g04730, Afu3g03440 and Afu3g03640. Preferential gene expression following a 60 minute shift from acid to alka-line medium could also be strongly correlated with that observed during infection (Figure
4.5 and Supplementary Dataset E.3). Alkaline adaptive capability, previously found to be essential for A. nidulans virulence in neutropenic mice (66), is likely to be important for growth of A. fumigatus spores at physiological pH. Accordingly we identified 102 genes pref-erentially expressed during both murine infection and in vitro alkaline adaptation (Figure 4.5 and Supplementary Dataset E.3). Among them are 36 genes having unknown function, two sodium ATPases (Afu6g03690 and Afu4g09440), the plasma membrane zinc ion trans-
porter (Afu6g00470) and an alkaline phosphatase (Afu3g14030). Interestingly, we found no concordance between iron starvation and alkaline adaptation (Figure 4.5). Since acidifica-tion of the macrophage phagolysosome is an essential step in ROS-mediated A. fumigatus killing we also assessed the transcriptome of Af293 germlings upon shift from pH 7 to pH 3, using a rich medium and hydrochloric acid. No concordance between the resulting dataset and that of host adaptation was evident, indeed (despite the magnitude of the

4.3 Results 160
murine infection dataset) most functions upregulated in response to an in vitro acid shift were less abundantly represented in our infection analyses (n = 18, Supplementary Figure E.2) thus we conclude that acid stress, at least within the context tested in this analysis (which can only approximate conditions encountered in the host) is not relevant during host adaptation. This agrees with our observation that alkaline adaptation is a physiologi-cal cue of primary importance in this murine model of infection (Figure 4.5). An essential component of phagocyte defence against A. fumigatus conidia and hyphae is the NADPH oxidase-mediated respiratory burst which generates reactive oxygen species (ROS) required for fungal killing. To assess the A. fumigatus transcriptional response to oxidative stress, conidia were grown in rich medium at 37°C prior to shift into similar medium containing 17 mM hydrogen peroxide. Comparison of the resulting dataset to that obtained from host-adapting germlings revealed some concordance, this time revealing a subset of genes having decreased transcript abundance both in vitro and during murine infection (Figure E.2).
A common theme among this group of genes is ergosterol and haem biosynthesis, evidenced by common behaviour of both 14-alpha sterol demethylase Cyp51A-encoding genes (Afu4g06890 and Afu7g03740) and a coproporphyrinogen III oxidase homologue (Afu1g07480). Oxygen depletion was achieved by transfer of A. fumigatus hyphae, fol-lowing 16 hour growth in rich medium, to anaerobic chambers containing two palladium catalysts. We were unable to correlate gene expression under these anaerobic stress con-ditions with gene expression during infection (Figure 4.6). Finally, we assessed germlings grown in 20 ml of RPMI1640 with L-glutamine, 25 mM HEPES and 5% fetal bovine serum for 7 hours at 37°C following exposure to human neutrophils at a multiplicity of infection of 1:1 for 60 minutes, the reference sample for this analysis being germlings incubated in the absence of neutrophils. Of 57 A. fumigatus genes upregulated in response to neutrophil ex-posure in vitro, 18 (60%) were also more abundantly represented during initiation of murine infection (Figures 4.6 and E.2). Interestingly these included the two major known A. fu-migatus antioxidant enzymes, Mn superoxide dismutase (Afu1g14550) and the bifunctional catalase-peroxidase Cat2 (Afu8g01670). Whether representation of these transcipts among those differentially expressed in both murine and laboratory culture is indeed neutrophil-specific remains to be determined.
4.3.7 Nutrient limitation is a relevant physiological cue during A. fumigatus initia-tion of murine infection
To investigate the relevance of nutrient starvation during host adaptation we made sev-eral growth and gene expression analyses. Hypothesising that YEPD is nutritionally more robust than murine lung tissue we compared radial growth of A. fumigatus, in triplicate

Chapter 4. Sub-telomere directed gene expression during initiation of invasive asp ergillosis 161
from an inoculum of 100 spores grown at an agar/air interface on Petri dishes, on YEPD and on a synthesized murine lung tissue medium (MLT) composed of homogenised murine lung tissue (80%) and water/agar (20%), overlaid upon a water/agar baseplate. Growth of A. fumigatus was completely unsupported by water/agar base with no evidence of conidial germination after 7 days. Growth on YEPD produced conidiating colonies averaging 66 mm (n = 3) in diameter after 7 days at 37°C, whereas MLT as growth medium supported significantly less growth, reaching a maximum colony diameter of 44 mm (Figure 4.7A). Any concern that reduced radial growth observed on MLT medium originates from iron deprivation can be allayed by reference to radial growth analysis of two independent wild type A. fumigatus isolates, CEA10 and ATTC46645 (618), where equivalent growth is ob-served in the presence and absence of iron, and on blood agar medium. Thus, from this solid growth analysis, we conclude that MLT supports slower A. fumigatus colony growth than YEPD under laboratory conditions. Our MLT analysis could not support the volume of liquid culture required to perform growth curve analyses, moreover, the viscosity of the medium would have hindered dry weight measurements. To support our conclusions on nutritive status of the host environment, within the context of the experimentation per-formed during our infection analyses, we assessed log2 ratios obtained from competitive microarray hybridisation, using doubly amplified A. fumigatus mRNA extracted from ni-trogen starved germlings, and the same YEPD reference sample used for the initial host adaptation analysis (samples F and G, respectively, Supplementary Table E.1). Nitrogen starvation was exerted in shaken liquid culture using minimal medium, with hydroxyproline as nitrogen source, for a period of five hours. Hydroxyproline is a rational candidate nitrogen source during initiation of mammalian pulmonary infection, being a widely used surrogate marker of lung injury whose concentration in bronchoalveolar lavage fluid permits quantita-tive assessment of collagen breakdown (3; 451). Transcript levels of the proline permeases Afu2g11220, Afu8g02200 and Afu7g01090 suggest induction of proline uptake during initia-tion of infection, and laboratory culture on solid medium confirms that hydroxyproline can support aconidial filamentous growth of A. fumigatus on minimal medium in the presence of a repressing carbon source such as glucose (data not shown). To confirm the inferiority of hydroxyproline as a nitrogen source (relative to YEPD) we performed dry weight growth curve analyses (Figure 4.7B) including a widely used Aspergillus minimal medium (MM) for comparison. Nitrogen starvation rendered 1047 genes subject to differential expression, relative to the doubly amplified YEPD reference. Several notable features of the resultant dataset (Dataset E.4) support our conclusion that, relative to YEPD laboratory culture, initiation of murine infection occurs under nutrient stress. As with our analyses of host adaptation, over-represented Gene Ontology (GO) terms among differentially expressed genes identified ribosome biogenesis and assembly, and protein biosynthesis and processing

4.3 Results 162
as the most significantly down-regulated functional categories (Table E). We also identified significant over-representation of cell cycle-related functions among
genes preferentially expressed during growth in hydroxyproline, including cell cycle regula-tion, mitosis, nuclear migration, chromosome segregation and karyogamy (Table E). This is a particularly satisfying finding since, supported by our growth curve analyses (Figure 4.7 B), a clear distinction between A. fumigatus growth phase in the compared media is discern-able. This was not a feature of the murine versus YEPD comparison which, importantly, lessens the probability that differences associated with cell cycle stage or growth rate pre-side over environmental cues within the host adaptation dataset. Direct comparisons of the murine and nitrogen starvation datasets identified an overlap of 280 differentially expressed genes common to both, of these 24 genes are also common to the TOR kinase nutrient limitation geneset, and are indicated in Supplementary Data Tables E.5 and E.6. 156 genes preferentially expressed under nitrogen starvation conditions (relative to YEPD laboratory culture) were similarly favoured during host adaptation (Supplementary Dataset E.4).
Assessing locational bias among the hydroxyproline dataset, 29% of the geneset was found to reside subtelomerically (Table 4.4) with 180 out of 634 preferentially expressed genes housed within 300 kb of telomeres. This is comparable to the level of subtelomeric gene expression identified during initiation of murine infection (Table 4.4) and indicates that, relative to growth in a nutrient-rich laboratory culture, adaptation to growth in a nu-tritionally challenging environment prompts expression of a subtelomeric gene repertoire. A number of physically linked coregulated genes came to prominence in the hydroxyproline starvation dataset (Supplementary Data Table E.11). 16 clusters conforming to the previ-ously applied cluster algorithm were identified, 8 of which were subtelomeric and 3 of which encompass genes in secondary metabolite loci, as defined by bioinformatic analyses. Of the three latter loci one cluster (Afu6g03390 - Afu6g03490) subject to regulation by the LaeA methyltransferase (539), the product of which is currently unknown, is also expressed dur-ing murine infection. Beyond this, correlation between clustered gene regulation in murine and nitrogen starved growth was modest.
4.3.8 The LaeA regulon is represented among genes preferentially expressed during intitation of murine infection
Given the predominance of clustered and subtelomeric gene loci among host adaptation genes, we compared our dataset to that produced by Perrin et. al. (539) during study of laeA gene deletion. An A. fumigatus AlaeA (Afu1g14660) mutant, which has decreased virulence in both neutropenic and hydrocortisone treated mice (350; 669), demonstrates significantly lower expression of genes in 13 secondary metabolite biosynthetic clusters in-

Chapter 4. Sub-telomere directed gene expression during initiation of invasive aspergillosis 163
eluding that of gliotoxin. LaeA was found to influence expression of a subset of lineage and species-specific genes therefore we tested the overlap between functions down-regulated in the absence of LaeA and functions having greater transcript abundance during murine infec-tion, hoping to decipher a link between genes under LaeA control, and those active during initiation of murine infection. Out of 415 genes down-regulated in the absence of LaeA, we identified 99 genes having increased abundance during initiation of murine infection (Figure 4.8). Functional categorisation of shared genes revealed that 40% (n = 40) were involved in secondary metabolite biosynthesis, among these we could identify three complete secondary metabolite clusters, those directing gliotoxin and pseurotin biosynthesis as well as the ge-netic locus mentioned above (Afu6g03390) whose biosynthetic product is unknown. Perrin et. al. also found 54% of the LaeA-regulated gene clusters showing differential expression under laboratory conditions were located within 300 kb of the telomeres. We therefore ex-tended our analysis still further, determining the proportions of subtelomeric and secondary metabolism cluster genes shared between the two datasets. This identified 49 and 40 genes, having subtelomeric locations and secondary metabolite biosynthetic functions respectively (Figure 4.8).
Finally we analysed all of the in vitro and in vivo datasets comparatively to assess the occurrence of subtelomeric bias among differentially expressed genes, aiming to deter-mine whether the induction of genes at the telomeric extremities of chromosomes was a standard feature of adaptation to environmental alterations rather than a host adaptation phenomenon (Table 4.4). The analysis identified murine adaptation, LaeA deletion, neu-trophil exposure and nitrogen starvation as the conditions most enriching for transcript abundance among subtelomeric genes, where 29%, 32%, 27% and 29% of the respective cohorts were located. Interestingly, removal of secondary metabolism genes from the anal-ysis dramatically reduced the LaeA regulated subtelomeric gene cohort to 22% while only minimally impacting representation of murine adaptation genes (Table 4.4). Taken together these analyses indicate that a significant component of the LaeA regulon, comprised mainly of secondary metabolism genes, is represented among transcripts more abundant during in-fection. Furthermore the subtelomeric bias observed among differentially expressed murine adaptation genes extends beyond secondary metabolite biosynthesis and does not appear to be a general feature of adaptation to environmental change.
4.4 Discussion
We present a methodology for A. fumigates transcript profiling during initiation of murine infection and a comparative analysis of global transcriptional programming, in laboratory culture and the mammalian lung. We were able to optimize a technical and statistical

4.4 Discussion 164
framework sufficiently robust to reproducibly quantify relative transcript abundances using minute samplings of A. fumigatus germlings. We chose to co-hybridize mock samples from the same amplification protocol (thereby determining the true biological effect in a bench-mark sample, in this case total RNA) and then, by comparison to cohybridisations using unamplified samples, indirectly estimate the systematic effect of the amplification in our analysis. The t—testing framework (509; 508) then allowed us to identify genes where the log ratios are significantly different due to one factor i.e. the amplification protocol. This en-abled an estimation of the proportion of genes showing amplification-dependent bias which we carried into the murine experiment, where comparison of unamplified versus amplified material is impossible. Clearly, we assume that this estimate can be reliably applied to sim-ilar experiments, in doing so we also assume that the amplification process depends only on the protocol adopted, and not the underlying gene expression dynamics. A further technical consideration in planning experiments comparing murine and laboratory samples is differ-ential treatments. Given the nature of our analysis, differential treatment of the germling samples was necessary to avoid osmotic shock in either instance. Water lavage would impose an osmotic shift on germlings rescued from the lung and saline treatment would osmotically shock laboratory cultured germlings. Thus the technical limitations imposed by such com-parative analysis must be accepted, however, in terms of maximally preserving transcript abundances within the context of the experiment, we believe our treatments of the samples to be appropriate.
To identify factors governing adaptation to the host niche we compared laboratory and murine lung samples aiming to find fungal attributes preferentially employed during in-fection. Therefore our findings document genes having increased transcript abundance in the A. fumigatus sample of murine origin, relative to that derived from laboratory cul-tured fungus. We performed our comparative analysis using doubly amplified mRNA from developmentally matched A. fumigatus germlings following laboratory culture, or growth in neutropenic murine lungs. In total we identified 2164 genes having altered transcript abundance. Functional analysis of the dataset flagged certain putatively relevant physio-logical cues which we then pursued by additional in vitro analyses. Careful interpretation of the dataset with respect to specific nutrient acquisition mechanisms, and within the con-text of the comparison performed here, can lend powerful insight into accessible nutrients in the mammalian niche. Nitrate assimilation (which is strongly inducible by nitrate in the absence of preferred nitrogen sources ammonium and glutamine) (17) is not preferen-tially employed in the murine host as evidenced by co-ordinate down-regulation of crnA (Afu1g12850), niaD (Afu12830) and niiA (Afu1g12840). This may be due to a) equivalent nutritional status of YEPD and murine lung or b) to the absence of nitrate as a nitrogen source during initiation of infection, or both. We were able to confirm, by radial growth

Chapter 4. Sub-telomere directed gene expression during initiation of invasive aspergillosis 165
analyses in vitro using YEPD and a synthetic lung tissue medium IVILT, that YEPD is nu-tritionally superior to MLT based upon the rate of radial growth supported (Figure 4.7A). Coupled with the observation that the nitrogen metabolite repression gene product AreA is required for full virulence (281) a likely explanation for slowed growth and repression of nitrogen assimilation, is the utilisation of alternative non-preferred nitrogen sources during establishment of disease, such as amino acids.
Strong support for this conclusion is provided by high level induction of the areA-dependent nitrogen-scavenging enzyme L-amino acid oxidase LaoA (Afu7g06810) which enables Aspergilli to catabolise a broad spectrum of amino acids in nitrogen starvation conditions (436). Notably a by-product of such catabolism is ammonium. However the likelihood that sufficient ammonium is produced by these reactions to prevent starvation is diminished by the observed general starvation response in our dataset. Catabolism of amino acids during initiation of infection is also evidenced by induction of the methyl citrate synthase enzyme (Afu6g03590) which acts to detoxify the intermediates of propionyl-coA generating carbon sources (436), such as cysteine, isoleucine and methionine. An essen-tial role for methylcitrate synthase in murine aspergillosis has recently been demonstrated (312) thereby demonstrating the value of our approach in generating physiological infor-mation on virulence mechanisms within the context of murine infection. Strong themes among genes having lowered transcript abundance during murine infection are ribosome biogenesis and assembly, and protein bisoynthesis and folding (Table 4.1). Such signatures are commonly observed among microbes under stress, and in this instance might indicate a slowing of growth in the murine lung, relative to laboratory culture. We reasoned that a mechanism possibly linking such transcriptional profiles to nutrient starvation is TOR mediated ribosomal gene regulation, which tightly couples protein synthesis and cellular growth to availability of nutrients and physiological status to balance the opposing forces of protein synthesis and degradation. This is pivotal for cellular fate determination in many organisms (270; 172) directing cells towards either proliferation (through the cell cycle) or vegetative growth (increase in size). Comparison of our dataset to that generated following rapamycin-mediated TOR kinase inhibition in S. cerevisiae revealed a very marked overlap (n = 125) in differentially regulated homologous genes (Supplementary Data Tables E.5 and E.6). This is in keeping with defined roles for TOR kinase function in S. cerevisiae, which includes the regulation of transcription in response to nutrients (111). A strong correlation between developmental programming and microbial secondary metabolite biosynthesis has been well-documented (351) and while plasticity of nitrogen metabolism demonstrably sup-ports A. fumigatus virulence (375) the role of the single, and likely essential, A. fumigatus TOR kinase homologue, TorA, (Afu2g10270) in this process remains untested. Intriguingly, however, a link between TOR kinase function and secondary metabolite production, par-

4.4 Discussion 166
tially through AreA, has recently been established in the rice pathogen Fusarium fujikuroi (685) where, in addition to the target genes in common with yeast and other eukaryotes, the AreA-regulated giberellin and bikaverin biosyntheis genes are also under the control of TOR. This raises the possibility of a relatively limited investment in secondary metabolism, within the context of our analysis, at the tested timepoint of murine infection.
Genes expressed during nitrogen starvation are also expressed during virulence in Mag-neporthe grisea (181), where two wide-range regulators of nitrogen catabolism genes, NPR1 and NPR2 (395) are required for virulence. Contrary to our analysis nitrate and nitrite reductase activities were found to be relevant for M. grisea during rice infection (181), how-ever, a number of amino acid transporting proteins predominated among induced functions during both in vitro nitrogen starvation and infection.
Common to that analysis, and ours, was increased abundance of proline oxidase (Afu3g02300) and proline permease (Afu2g11220, Afu7g10190, Afu8g02200) encoding genes, which, in the context of our experiment might have special relevance given the high hydroxyproline of collagen tissue, and the notable proline requirement of an attenuated A. fumigatus deletion mutant lacking the Ras-related protein RhbA (524). To test the effect of nitrogen limitation on gene expression, in the context of the host adaptation study performed, we returned to the YEPD reference sample employed for the initial analysis (sample F, Supplementary DataTable E.1), this time performing competitive hybridisations with doubly-amplified RNA obtained from a nitrogen-starved laboratory culture (sample G, Supplementary Data Table E.1). Given the predominance of hydroxyproline among collagen amino acids content, and its release into bronchoalveolar lavage fluid upon tissue injury (3) we used hydroxypro-line as sole source of nitrogen for the starvation experiment. Hydroxyproline supported markedly slowed growth of A. fumigatus in liquid culture (Figure 4.7 B) relative to YEPD. We identified a significant overlap between the two datasets amounting to 280 genes.
Functional anlaysis of the resulting datasets revealed that, in keeping with the murine adaptation dataset, ribosome biogenesis and protein biosynthesis were markedly down reg-ulated, moreover, functions associated with mitosis and cell cycle were more abundantly represented among the categories favoured under nitrogen starvation, relative to rich labo-ratory growth. Given the differences between the tested media in terms of ability to support A. fumigatus growth (Figure 4.7 B) it is pleasing to see that such a theme was not apparent from the murine adaptation dataset where the degree of nutrient limitation is unlikely to be as severe as that imposed in the nitrogen starvation conditions we used here. Interestingly 29% of the genes preferentially expressed during nitrogen starvation were subtelomerically located, and a degree of clustered gene expression was observable. Thus induction of the subtelomeric gene repertoire becomes important during nutrient deprivation, a trend which was not observable in response to any of the other in vitro stresses, other than neutrophil

Chapter 4. Sub-telomere directed gene expression during initiation of invasive aspergillosis 167
exposure the physiological relevance of which requires further investigation. The uptake and catabolism of other amino acids released by proteolytic digestion of
murine lung parenchyma might provide a source of nitrogen during A. fumigatus infection, which is supported in our analyses by increased transcript abundance of various secreted proteases (Table 4.2). Extracellular proteases are implicated as virulence factors in invasive aspergillosis (394). An elastolytic protease, produced when A. fumigatus is cultured on the insoluble matrix from bovine lung is also produced during spore germination in infected lungs of neutropenic mice, as judged by immunogold cytochemical localization (365), and mutants unable to produce this protease were deficient for virulence in the murine model. An elastinolytic metalloprotease, characterized by the same group was similarly visualized during murine lung infection (444). Many species of human pathogenic fungi secrete pro-teases in vitro or during infection. Full virulence associated with A. fumigatus protease mutants (394; 325) is presumed to be due to redundancy among the many enzymes pro-duced by this organism which may degrade the lung parenchyma to release utilisable carbon and nitrogen during infection. A directed analysis of the role of such proteins in virulence might now be possible on the strength of our data. Time course analyses of A. fumigatus growth during murine infection will reveal stage-specific gene expression in the absence of confounding sample treatments. These analyses are underway in our laboratories.
Fungal oxygen-sensing mechanisms have been linked to cell membrane sterol levels in S. pombe and C. neoformans where homologues of the mammalian Sterol Regulatory Element Binding Protein (SREBP) transcription factors, in complex with SREBP cleavage-activating protein (SCAP) partners, undergo cellular translocation (from the endoplasmic reticulum to the golgi) prior to proteolytic SREBP activation (309), (123). The C. neoformans SREBP homologue, Sre1p, plays an important role in low oxygen adaptation and infection (123). Biosynthesis of sterols and unsaturated fatty acids is an aerobic process in S. cerevisiae and in C. neoformans, the hypoxia-mimicking agent cobalt chloride and oxygen limitation target sterol biosynthetic gene expression. This was intriguing to us given the broadly observed reduction in transcript abundance among ergosterol biosynthetic genes, relative to laboratory culture (Table 4.2).
Our initial hypothesis attributed this effect to oxygen limitation in the murine lung environment since airway obstruction, intraalveolar exudates and inflammation, or damage to alveolar capillaries (all observed in our murine modelling of pulmonary aspergillosis) pose a significant barrier to proper oxygenation in human lungs (703). Oxygen depriva-tion in mammals leads to a transcriptional induction of genes for adaptation to hypoxia (84) and efforts to characterise the transcription profile of murine immune responses to A. fumigatus infection indicate that hypoxia is relevant in the neutropenic murine lung at 24 hours post-infection (32). Co-hybridisation of murine lung cDNAs, derived from in-

4.4 Discussion 168
fected immunocompetent and immunocompromised mice with a murine immunology array set identified upregulation of murine ARNT (log2 ratio = 2.09) the obligate heterodimeric binding partner for the hypoxia induced factor HIF-1a (84). However, on comparing the A. fumigatus murine adaptation gene expression signature to that observed following exposure to anaerobic stress in vitro (Figure 4.5A) no support for this hypothesis could be gleaned. Rather, repression of the two A. fumigatus 14-alpha sterol demethylase genes (Afu4g06890 and Afu03740), representing a critical step in ergosterol biosynthesis, was observed following hydrogen peroxide-mediated oxygen stress. Therefore, it would seem that this component of the H202-mediated oxidative stress response is relevant in vivo.
This was found to be distinct from antioxidative action of the A. fumigatus Mn super-oxide dismutase (Afu1g14550) and the bifunctional catalase-peroxidase Cat2 (Afu8g01670), both of which were more abundant following murine lung (Table 4.2) or neutrophil expo-sure (Figures 4.6 and E.2), suggesting multiple modes of oxidative stress encountered dur-ing murine infection. Interestingly H202 gradients are detectable across the S. cerevisiae plasma membrane upon H202 exposure, suggesting a mechanism other than diffusion for H202 entry into cells. This, coupled with the observation that S. cerevisiae mutants erg30 and erg60, having increased ergosterol biosynthesis, show increased permeability to H202 might suggest down-regulation of ergosterol biosynthesis as a protective response against oxidative stress (85). All of the previously characterised components of the A. fumigatus siderophore biosynthetic pathway (618; 619) were more abundantly represented at tran-script level during murine infection. Comparison of the murine dataset with that generated during in vitro iron limitation confirmed the importance of this environmental deficit dur-ing murine adaptation (Figure 4.6). With respect to extracellular iron mobilization fungal siderophores bind ferric iron with a high affinity, delivering the ferric chelate to specific receptors at the cell surface for translocation into the cytoplasm. Existing evidence sup-ports an essential role for this uptake mechanism during infection (618; 619). Reductive iron assimilation (RIA) is dispensable for murine virulence (618) but may assist in iron acquisition during infection since RIA inhibition in the absence of extracellular siderophore biosynthesis prevents growth in vitro (619). This hypothesis is supported by our finding that numerous RIA components are induced during initiation of infection (Table 4.2) in addition to siderophore biosynthetic genes. We were also able to correlate differential gene expression following in vitro alkaline shift to the host adaptation transcriptome (Figure 4.5). This was to be expected given the broad requirement for fungal pH adaptation dur-ing mammalian pathogenesis (536) but is nonetheless pleasing to observe particularly given that the in vitro acid stress transcriptome showed an opposite trend (Supplementary Fig-ure E.2) and since pleiotropic activity of the virulence-directing family of PacC/Rim101 pH sensing transcription factors (536) does not permit virulence defects to be solely, or thus

Chapter 4. Sub-telomere directed gene expression during initiation of invasive aspergillosis 169
far absolutely, correlated with pH growth phenotypes. Fungal secondary metabolite biosynthesis is mediated at the level of transcription, from
clusters of physically linked, co-ordinately regulated genes and is profoundly affected by en-vironmental factors such as pH and nutrient availability (351). For the most part, defined cues prompting gene expression from such clusters remain unknown, as do the functions of the molecules produced by them in the natural environment, but popular theory regards mi-crobial secondary metabolites as chemical determinants of selective advantage (591). Noting that clusters of physically linked, co-ordinately regulated genes were a prominent feature of our dataset we applied a custom script to systematically identify them. To allow for noise in the microarray analysis, which can lead to exclusion of some spots in the final analysis, we allowed for gaps of up to a maximum of 4 non-up- or down- regulated genes permissable per cluster. Gap size variation influenced the architecture of clusters detectable in the dataset, further widening the extremities of pre-existing clusters rather than creat-ing new ones (a complete breakdown of the cluster anlaysis is available as Supplementary Dataset E.2). Thus a gap size of zero identified 14 upregulated gene clusters and a gap size of one identified 38. The maximum gap size applied (gap size = 4) identified 65 groups of physically linked genes having increased transcript abundance during murine infection (Figure 4.3). A large proportion (34%) of up-regulated, physically clustered, genes was found to reside within 300 kb of chromosome ends (Figure 4.3 and Table 4.4). The clusters are comprised of up to 34 co-ordinately expressed genes and include loci directing biosyn-thesis of siderophores (cluster 33) and two known secondary metabolites, pseurotin and gliotoxin (clusters 70 and 59, respectively). The importance of fungal gene clusters in viru-lence has recently been characterised in the fungal plant pathogen, Ustilago maydis where co-induction of physically linked, secreted protein-encoding genes is seen during infection (299).
We were unable to find evidence of clustered secreted protein genes from our analyses of A. fumigatus (data not shown) however, from an evolutionary perspective, clusters 8, 40, 53, 68 (down-regulated), 69 and 70 (Table S6) deviate significantly from the least viru-lent sequenced species considered here, N. fischeri, and therefore merit further analysis as they are linked to a specific, virulent Glade. N. fischeri is extremely rarely identified as a human pathogen (132; 420; 672), while prolonged exposure to A. clavatus spores can cause extrinsic allergic alveolitis known as malt worker's lung (70). From a functional perspective some relevant deductions regarding the clusters identified by our analyses might be possible using present genome annotations. However, closer scrutinization of clusters is required to address the likelihood that A. fumigatus genes located in them encode functions required for virulence. We anticipate that the inference of shared functionality among neighbouring co-regulated genes in our dataset will empower the functional annotation of the A. fumiga-

4.4 Discussion 170
tus genome; moreover, it significantly raises the profile of such gene-regulatory paradigms within the context of fungal pathogenicity. Our observations of biased localization among genes overrepresented during growth in the murine lung (Figures 4.3 and 4.4, Tables E.7, E.8, E.9, E.10) prompted comparison of our dataset to that obtained by Perrin et. al. (539) who identified genes under control of the global seconday metabolism regulator, LaeA. Sig-nificant overlap is observable, notably among secondary metabolism genes, most specifically between genes in the gliotoxin and pseurotin biosynthetic gene clusters.
Hypovirulence of a AlaeA deletion mutant cannot be explained solely on the basis of a lack of gliotoxin biosynthesis, and no investigation of the role of other secondary metabolite clusters, with respect to virulence, has been undertaken in A. fumigatus, therefore at this time it is not possible to reach any conclusions on the contribution made to virulence by the molecules whose synthesis is directed by these loci. With regard to our comparison of functions under LaeA control and those important during murine infection, differing ex-perimental conditions (Perrin study (539) performed at 25°C in liquid shaking culture and glucose minimal medium (548) for 60 hours) and methodologies pose a contentious issue. A truly illuminating analysis of LaeA activity during murine pathogenesis might be forth-coming from time course analyses in neutropenic mice, which is currently being attempted (Timothy Cairns, Bignell Lab). DNA sequences in subtelomeric regions undergo ectopic recombination at a much higher rate than expected for homologous recombination (196) allowing the expansion and diversification of gene families located at chromosome ends. For some organisms gene expression is governed by subtelomeric localisation, for example two A. nidulans secondary metabolism gene clusters, directing penicillin and sterigmatocystin biosynthesis, are activated following deletion of the hdaA histone deacetylase gene [65]. In other organisms subtelomeres provide an ideal setting for genes involved in antigenic varia-tion, such as in the parasites Plasmodium falciparum, and Trypanosoma brucei (54) and in cytoadhesion, such as the EPA family of C. glabrata adhesins (1). Thus the importance of sub-telomeric chromosomal regions within the context of eukaryotic pathogenesis is gaining significance in this post-genomic era of microbial studies.
These analyses of transcript abundance, drawn from minute quantities of fungal material convey a programme of A. fumigatus cellular regulation directly from the site of pulmonary infection. From a fungal physiological perspective the mammalian host restricts iron, and likely various nutrients, as well as exerting multiple degrees of oxidative stress. Regarded as a function of the genomic landscape the observed transcriptional changes reveal genome organisation and subtelomeric diversity as effectors of the remarkable versatility of A. fu-migatus with respect to the niches it successfully inhabits, one of which is the neutropenic human lung.

Chapter 4. Sub-telomere directed gene expression during initiation of invasive aspergillosis 171
4.5 Chapter Summary
1. Recent genome sequencing of several Aspergillus species provides an exceptional op-portunity to analyse the transcription of potential fungal virulence attributes within a genomic and evolutionary context.
2. To identify genes preferentially expressed during adaptation to the mammalian host niche, we generated multiple gene expression profiles from minute samplings of A. fumigatus germlings during initiation of murine infection
3. They reveal a highly co-ordinated A. fumigatus gene expression programme, governing metabolic and physiological adaptation, which allows the organism to prosper within the mammalian niche. As functions of phylogenetic conservation and genetic locus, 28% and 30%, respectively, of the A. fumigatus subtelomeric and lineage-specific gene repertoires are induced relative to laboratory culture, and physically clustered genes including loci directing pseurotin, gliotoxin and siderophore biosyntheses are a prominent feature
4. Locationally biased A. fumigatus gene expression is not prompted by in vitro iron limitation, acid, alkaline, anaerobic or oxidative stress. However, subtelomeric gene expression is favoured following ex vivo neutrophil exposure and in comparative anal-yses of richly and poorly nourished laboratory cultured germlings.
5. We found remarkable concordance between the A. fumigatus host-adaptation tran-scriptome and those resulting from in vitro iron depletion, alkaline shift, nitrogen starvation and loss of the methyltransferase LaeA
6. This first transcriptional snapshot of a fungal genome during initiation of mammalian infection provides the global perspective required to direct much-needed diagnostic and therapeutic strategies and reveals genome organisation and subtelomeric diversity as potential driving forces in the evolution of pathogenicity in the genus Aspergillus.

172
Chapter 5
Functional genomics of fluconazole sensitivity in Candida glabrata using a library of transcription factor knock-outs: a pilot study
5.1 Chapter Overview
The groundbreaking sequencing and annotation of the S. cerevisiae genome some thirteen years ago (246) ushered in a new era of yeast research in which reverse genetics approaches became the catalyst for a burgeoning supply of genome-wide phenotypic data. Unlike for-ward genetics, reverse genetics exposes a strain, typically deficient in a known protein-coding chromosomal locus, to a selection of in vitro assays designed to evaluate the contribution of that locus to a specific biological process. With an abstract notion of systematically removing each functional gene, and a research community newly furnished with sequence data, various implementations emerged. These were reliant either on the promiscuity of mobile genetic elements in target genomes, or the collaborative spirit of yeast researchers and their collective purpose to disrupt open reading frames in a targeted fashion.
The earliest comprehensive deployment of mobile genetic elements, was undertaken by Ross-Macdonald et al. (595) who used a mTn3xHA/lacZ mini transposon to determine dis-ruption phenotypes for nearly 8,000 strains in 20 different growth conditions. This technique provided supporting evidence for some established gene functions, but more importantly elu-cidated the function of previously uncharacterized ORFs involved in sporulation and vege-tative growth. The collection of mutants was also used in separate studies to identify genes required for growth in high-salt medium (159), survival of hypo-osmotic shock and growth

Chapter 5. Functional genomics of fluconazole sensitivity in Candida glabrata using a library of transcription factor knock-outs: a pilot study 173
at 15°C in addition to nitrogen starvation, invasive or filamentous growth and sporulation (675).Although transposons are widely adopted for random mutagenesis in yeast, other approaches have also been used, including mutagenesis by T-DNA from Agrobacterium tumefaciens (98), linearized plasmid DNA (135) and the use of non-homologous insertion of DNA cassettes (356). There are some benefits to using a random mutagenesis approach (e.g. promoter regions can be disrupted, gain-of-function phenotypes can be observed), however the propensity for mobile elements to integrate in non-coding regions means that exhaustive coding region coverage has not yet been achieved; on the other hand, targeted disruption techniques, although labour intensive, provide much greater coverage of the tar-get genome. The best example of this co-ordinated disruption effort is the library of deletion mutants provided by Winzeler et al (749) and Giaever et a/.,(239).
This library comprises over 20,000 haploid and diploid knockout strains corresponding to deletions of each of 5,916 genes (including 1,159 essential genes) and one haploid strain of each mating type for every non-essential gene (4,757 genes). It has been used extensively to uncover the role of genes in a range of environmental stresses including: extremes of salinity (730; 239), hyperosmotic stress (239), alternative carbon sources (659; 239), alkaline stress (239), temperature shock (239), oxidative stress (239) and DNA damaging agents (60; 225). The collection has also been informative in dissecting the molecular basis of fundamental biological processes such as meiosis and sporulation (498), chromosomal segregation (445), cytoskeletal organization, cell-wall biosynthesis, microtubule-based chromosome segregation and DNA metabolism (692; 693), glycogen accumulation (747), accumulation of mutations (307), response to abiotic chemical stresses induced by a variety of drug molecules including anti-fungal agents and anti-cancer drugs (25; 26; 240). A valuable feature of this library is the incorporation of unique identifiers within the disruption cassette sequence. This enables parallel screening of pooled mutants in a range of environmental stress conditions, in an analogous manner to the signature tagged mutagenesis (STM) approach developed by the Holden laboratory (282). This technology relies on hybridization to microarrays, rather than filter membranes, spotted with complementary barcode sequences and allows data to be analyzed with statistical measures and unsupervised learning techniques for classification and functional prediction of genes (93).
As S. cerevisiae research enters a new integrative phase (reviewed in (293)), it does so from a solid platform of reductionist biology based, in part, on the use of disruption libraries described above. Meanwhile, Candida researchers lag somewhat behind, lacking the equiva-lent comprehensive disruption libraries. Existing S. cerevisiae disruption libraries have been useful in characterizing orthologous C. abicans pathways (63; 739), but do not provide any insight into the function of genes specific to Candida species that may explain differences in virulence phenotype between species. Candida researchers have tried to follow suit. In-

5.1 Chapter Overview 174
deed, mutagenesis protocols have been successfully adopted in C. albicans and C. glabrata. The Haynes laboratory adopted a signature tagged mutagenesis approach to identify a C. glabrata mutant, aces, with a cell separation defect, that is hypervirulent in a murine model of candidiasis aces (341); Cormack and Falkow (140; 139) developed a random mutagen-esis approach based on transformation of non homologous linearized DNA to identify a novel family of genes (EPA) mediating adhesion of C. glabrata to human epithelial cells. A Tn7 transposon based mutatgenesis protocol developed by (116) has been used to screen for altered sensitivity to fluconazole, (347) however coverage was limited to approximately 25% of the genome, with 75% of disruptions occuring in coding sequences. Therefore, a remaining challenge is the establishment of a comprehensive, genome-wide collection of C. alb jeans and C. glabrata deletion mutants akin to that developed by the Saccharomyces genome deletion project. In this section I report on initial progress in creating a prototypic targeted disruption library in C. glabrata. The overall aim is to provide genome-wide cov-erage, however for the initial pilot study, a collection of putative functionally related ORFs was chosen. In the first instance, we chose to disrupt ORFs encoding putative regulators of transcription, because the laboratory's earlier work with ACES suggested that transcrip-tion factors play an important role in modulating virulence. Moreover, the altered function of transcriptional circuitry in evolutionary timescales has emerged as an important source of diversity between organisms that share similar transcriptional components (reviewed in (702) and (592)). With an orthologous S. cerevisiae transcription factor knockout library already available, we hope that a quantitative growth assay will form the first part of a study of transcriptional rewiring between non pathogenic fungus and related opportunis-tic pathogen. Creation of this library will facilitate comparison of S. cerevisiae and C. glabrata transcription factor regulons in gene expression microarray experiments, and also phenotypic traits of null mutants exposed to a variety of environmental stress conditions.
The aims of this pilot study were to provide a quantitative measure of fitness in envi-ronmental stress conditions so that S. cerevisiae and C. glabrata phenotypic traits can be compared.

Chapter 5. Functional genomics of fluconazole sensitivity in Candida glabrata using a library of transcription factor knock-outs: a pilot study 175
This work was presented at the following conference: Third FEBS Advanced Lecture CourseHuman Fungal Pathogens:
Molecular Mechanisms of Host-Pathogen Interactions and Virulence May 2 - 8, 2009
Biao Ma, Andrew McDonagh, Emily Cook, Melanie Puttnam and Ken Haynes Functional genomics of fluconazole sensitivity in Candida glabrata using a library of
transcription factor knock-outs.
5.2 Specific Methods
5.2.1 Construction of the CGTFKOLib library of transcription factor null mutants
Through the study of sequence data on the Saccharomyces Genome Database 1 and liter-ature searches, Biao Ma - a post-doctoral colleague in the Haynes laboratory - identified 208 non-essential S. cerevisiae genes encoding known or putative regulators of transcrip-tion, encompassing the many categories of genes that can regulate transcription, including: DNA-binding TFs, subunits of large protein complexes that regulate gene transcription and also chromatin re-modelling factors that play important accessory roles in transcrip-tion regulation. The C. glabrata homologues of these 208 genes were identified by BLAST (16) search on the Genolevures website2 (630). For ten S. cerevisiae genes, two C. glabrata paralogues can be identified. Hence, in total, 218 C. glabrata genes were identified as dis-ruption candidates for inclusion in a gene knock-out collection known as the CGTFKOLib (Candida glabrata Transcription Factor Knockout Library).
Based on published methods for constructing gene knock-outs on a large scale in C. albicans (502), and methods previously developed in the Haynes and Cormack laboratories, a highly efficient knockout protocol was developed by Biao Ma over the course of a year. The method (Figure 5.1) is summarized below. Approximately 500 bp of the 5' and the 3' un-translated regions (UTRs) of each target gene in addition to a 1.4 kb selectable NATR cassette, which confers resistance to nourseothricin, are individually amplified by PCR and purified by gel extraction. Several features are also integrated in the amplified 5' and 3' UTRs through the primers `-1-NATR' and `4-1-NAT-F'. For the amplified 5' UTR, two 18 bp universal priming sequences, Ul and U2, flanking a unique 20 bp sequence tag (Up tag) are incorporated at its 3' end. For the amplified 3' UTR, two 18 bp universal priming sequences, D2 and D1, flanking another unique 20 bp sequence tag (Down tag) are incorporated at its 5 end. The 'Up tag' and the 'Down tag' enable the future identification
lhttp://www.yeastgenome.org
2http://www.genolevures.org/

FAT J2 NAT* eass to FRTV2i
Fusion PCR -500-F
5 UTR TIT Ulf FRr . tiAr cassette
DefeLian CafffIrflc;
oatiri tau
!FIRTIO4 , 01. 3' UTR
V5OWn is
+1-NAT-F
UTR I-500 bp) •
4500-ft
-500-F
[ UTR (-500 bp(
oc ur -1-111AT-R
5.2 Specific Methods 176
Tranclormeng to C gta•hrazir
ORF ot tar get gene 3' UTR +600-R
iartrt.R
4004 5' UTR
Up vity NAT -3`-F c'-wmn I 1 .t!.. 1 VirrErr; NATR cassette IFRTlC2 jr]
NAT-5'4 Correct Otiottoo 01 target grog!
01‘ 3' UTR .600-R
Figure 5.1: Molecular approach adopted for targeted disruption of C. glabrata transcription factor ORFS (schematic). The procedure consists of three stages: 1) The creation of three fusion components. This involves the amplification of approximately 500 bp of the 5' (-500-F, -1-NATR primer pair) and the 3' (1-NAT-F, +500-R primer pair) un-translated regions (UTRs) of each target gene in addition to a 1.4 kb nourseothricin (628) resistance cassette (NATR) (Top panel). Amplified upstream and downstream UTRs comprise 151-U2 and D2-D1 tags respectively - universal amplification sites flanking molec-ular barcodes that uniquely identify the disrupted strain. 2) Fusion. U2 and D2 sequences incorporated in the amplified 5' and 3' UTRs, respectively, overlap with the 5' and 3' end of the amplified NATR cassette. Thus, the 5' UTR, the NATR cassette, and the 3' UTR of the target gene can be joined together through both overlaps by a fusion PCR to yield a deletion construct. 3) Transformation and screening. Each deletion construct is individu-ally transformed into a C. glabrata his3 strain by electroporation and disrupted strains are identified by screening 20 to 60 transformants by PCR (-600-F, -int-F and -int-R. + 600-R primer pairs for amplification of targeted ORF - present if not disrupted; -600-F, NAT-5'-R. NAT-3'-F, +600-R for amplification of disruption cassette at the chromosomal locus).
400-F 5' UTR

Chapter 5. Functional genomics of fluconazole sensitivity in Candida glabrata using a library of transcription factor knock-outs: a pilot study 177
of each gene knock-out through a tag-array-based method, and the Ul, U2 sites and the D2, D1 sites can be used to amplify all the Up tags and Down tags, respectively. The `Up tag' and 'Down tag sequences were chosen to match the homologous identifiers used in generating the complete S. cerevisiae gene knock-out strain collection, allowing existing parallel knockout screening tools to be re-deployed in C. glabrata. Moreover, the U2 and D2 sequences incorporated in the amplified 5' and 3' UTRs, respectively, overlap with the 5' and 3' end of the amplified NATR cassette. Thus, the 5' UTR, the NATR cassette, and the 3` UTR of the target gene can be joined together through both overlaps by a fusion PCR to yield a deletion construct. Each deletion construct is individually transformed into a C. glabrata his3 strain (Schwarzmiiller unpublished data (691)) (a derivative of C. glabrata ATCC 2001) by electroporation. Subsequently, 20 to 60 transformants are screened by PCR for the desired gene knock-out through PCR. The PCR screening confirms the correct integration of the deletion construct in the target gene locus and the loss of the open reading frame of the target gene. Finally, individual frozen stocks are made of single colonies derived from two independent transformants that contain the correct knock-out of each target gene. For the 218 candidate genes, 1,960 DNA primers were designed for generating deletion constructs as well as for confirming correct gene knock-outs. Using the protocol described above, gene knock-out strains for 186 genes have been generated, with a success rate of 85%.
5.2.2 Liquid culture assay to determine strain growth dynamics
Glycerol stocks of each library strain, collectively maintained in two 96-well microtitre plates at -80°C, were sampled (5 pl) using a multi-channel pipette and dispensed into correspond-ing sterile, round-bottomed 96-well plate containing 150 µl of YEPD broth. Library strains were sub-cultured to stationary phase for 18 hours in an orbital shaker (30°C, 180 RPM) before commencing the growth curve assay. Sub-cultured cells were harvested (3000rpm, 5 mins) in a centrifuge rotor equipped with plate adaptors (Wolf Laboratories, UK) and washed three times with sterile, distilled water. Washed cells from each library plate were transferred (final OD600=0.1) into microtitre plates containing YEPD, YEPD + 5 µg-m1-1 fluconazole, YEPD + 10 iug.m1-1, YEPD + 30 µg•ml-1 and YEPD + 100 µg•ml-1 (fi-nal concentration). Plates were sealed with a breathable membrane and incubated for 12 hours. Optical density (0D600) measurements were bi-hourly recorded on a Multiskan Ascent absorbance plate-reader (Thermo Scientific, U.S.A). Assays were performed in bio-logical quadruplicate unless otherwise stated.

5.2 Specific Methods 178
5.2.3 Fitting growth dynamics data to a logistic model of population growth
Time course optical density (0D600) data, used as a measure of cell population dynamics, were fitted to a three parameter (slope, midpoint and upper asymptote) logistic function of growth using non-linear regression (SSLogis function) in the R statisitical computing framework. The logistic function of growth adopted in the analysis is shown in equation 5.1:
f (x, (b, d, e)) = (5.1) 1 + exp{ b (log(x) — log(e))}
In this model, logistic growth is a function of time x, the maximum cell density d (upper asymtote), the midpoint of the growth e and the slope around this midpoint b. Parameters d, e and b are quantitative phenotypic traits of each strain and are estimated from samples of cell density at x = 0 —11.5 hours. Strains showing significant, fluconazole specific growth defects were identified in null strains either by a) a complete lack of logistic growth in the presence of fluconazole but not YEPD (hypersensitive) or b) significantly greater midpoint (p < 0.05) estimates than wild-type strains in fluconazole but not YEPD (sensitive). Missing midpoint estimates, resulting from failed non-linear regression were imputed and set to a value outside the measured range (15 hours).
5.2.4 Fitting truncated growth dynamics data to an exponential model of population growth
Optical density data (0 - 8 hours), were considered for estimating the growth rate of strains in an exponential model of growth, in an attempt to disregard the systematic problems of estimating a logistic fit to data that poorly defined the stationary phase of growth. In this approach, optical density y is described by a simple model of exponential growth:
y = a + bert (5.2)
Where a is the initial population at time t = 0 and r is the growth rate of the strain. Model parameters were estimated using the non-linear least squares regression function in the R statistical framework. Starting values for estimated parameters were fixed for each strain: a = 0.04, r = 0.2, and 1000 iterations were allowed for convergence. 95% Confidence intervals were simulated from the standard errors of the estimated parameters using the confint function provided by the multcomp package in the R statistical framework. Sensitive strains were identified if the upper confidence interval of r for the mutant strain was less than the lower confidence of interval of r for the parental strain in media supplemented with fluconazole. Furthermore, these strains must show no difference in growth in YEPD

Chapter 5. Functional genomics of fluconazole sensitivity in Candida glabrata using a library of transcription factor knock-outs: a pilot study 179
medium lacking fluconazole.
5.2.5 Fitting the inhibitory effect of fluconazole at a single time point in a linear model
In a departure from the above non-linear regression modelling approaches, the optical den-sity y at a single time point is modelled as a linear relationship between the strain genotype 131, the environmental condition 02 and an interaction term between the two, which repre-sents the fluconazole-specific sensitivity of each strain.
y =131X+02X+ /302X +E (5.3)
Least squares regression estimates of these terms and the residuals E was performed using the lm function of the R framework for statistical computing.
The Z factor statistic, commonly used in high throughput screening assays as a quality metric (769) was used as a rational basis for choosing the single time point that maximized the signal to noise ratio. The Z factor is defined as:
=7 1 3 x (6p + &n)
I uip —fin l
(5.4)
Where i7p is the estimated sample standard deviation for the positive control samples (i.e. the parental strain growth), ern is the estimated sample standard deviation for the negative control samples (i.e. blank wells), A p is the mean of the positive control samples and ri p is the mean of the negative control samples.
5.3 Unsupervised learning of strain clusters based on growth parameter estimates
Hierarchical clustering was performed with the hclust function in the R statistical framework using a Euclidean distance metric between midpoint estimates and average linking. Where insufficient growth occurred for logistic regression based estimation of parameters, growth of each null strain was expressed as a percentage of the parental strain growth, his3 in each growth condition.
5.4 Results
My aim was to develop an assay that could identify null mutant strains showing either hypersensitivity or sensitivity to fluconazole, and where possible, to provide estimates of growth rate, so that strains could be classified by unsupervised learning techniques such

5.4 Results 180
as hierarchical clustering. This would form the basis for quantitative comparison of S. cerevisiac and C. glabrata environmental stress regulation by putative transcription factors using a functional genomics approach, instead of gene expression microarrays. The standard phenotypic growth assay adopted in the Haynes lab relies on a discrete scoring (growth, no growth or sensitive) of colonies spotted onto solid agar plates, and thus provides only a very coarse classification of strains that is uniform across each environmental stress condition. For this newer assay to be valuable to the overall aim, it must not only be possible to easily identify each of the categories described above, but also to provide a measure of sensitivity to provide finer grained categorization. This relies on reproducible estimates of the growth midpoints for each strain, but most importantly the control strain, with which the null mutant strains are compared.
Hypersensitive strains, classified as "no growth" strains in the existing method, were easily identified even in the untreated YEPD culture medium. Ten null mutant strains (rpd3, gln3, pop2, ssn6, nggl, snf6, gcn5, rtfl, asfl and spt3) grew so slowly in relation to the parental his3 strain (range = 11.64% - 74.42% of his3 growth at 11.5 hours) that logistic regression estimates of growth parameters were not achieved with the SSlogis function (Table F.2). For the sake of discussion, these strains were defined as "growth defect" strains. Were logistical fits were impossible, curves are omitted from all plots.
In contrast, both the library parental strain, his3, and the wild-type ATCC 2001 strain showed logistic growth in YEPD that was sensitive to fluconazole in a concentration-dependent manner (Figure 5.2). Disappointingly, variable midpoint estimates of these reference strains prevented the identification of "sensitive" null mutant strains with the hypothesis testing methods adopted. The estimate of ATCC 2001 growth midpoint in YEPD, showed the least error of the two reference strains (8.44 hours, SEM = 0.37 hours), corresponding to a coefficient of variance of 7.58%.
Estimation of 95% confidence intervals, based on assumption of normally distributed midpoints, places the midpoint in the range of 7.72 - 9.16 hours; addition of the lowest concentration of fluconazole (5pg-m1-1) to the culture medium shifts the upper confidence interval of the midpoint beyond the detectable range (8.134 - 14.46 hours). Thus, although 81 strains have midpoint estimates greater than that of the ATCC 2001 strain, error in the estimates of reference and null strains prevents the rejection of the null hypothesis i.e. no difference in growth rates as a consequence of gene deletion.
The same is true if the his3 strain is used as the reference strain - a more suitable control because all null mutant strains in the CGTFKOLib were derived from this background. The estimate of the his3 growth midpoint in YEPD, was greater than ATCC 2001 (9.39 hours, SEM = 1.33 hours), corresponding to a coefficient of variance of 24.49%,and shows a more poorly defined upper asymptote than that of the ATCC 2001 strain (Figure 5.2), indicating

Chapter 5. Functional genomics of fluconazole sensitivity in Candida glabrata using a library of transcription factor knock-outs: a pilot study 181
that its growth is further away from the stationary phase at the maximum sampling time-point. Similar estimates of 95% confidence intervals, place the midpoint in the range of 8.13 -14.46 hours; addition of the lowest concentration of fluconazole (5jzg•m1-1) to the culture medium also shifts the midpoint beyond the detectable range (3.33 - 19.39 hours). Errors associated with midpoint estimates increased with fluconazole concentration (Figure 5.3), such that only data from the 5µg•ml-1 and 10pg•m1-1 fluconazole treatment were retained in the final analysis.
Twenty four strains were identified as hypersensitive in YEPD supplemented with 5teg•ml-1 fluconazole. These stains showed no obvious increased in optical density be-tween 0 - 8 hours. This includes 14 strains that showed no difference in growth to reference strains in YEPD (), in addition to those 10 strains previously defined as having growth defects in YEPD (Table F.4). Of the 14 strains that showed fluconazole-specific growth inhibition, two strains carried disruption cassettes in ORFs encoding known multi-drug resistance effectors, PDR1 and MIG1.
Other strains that were hypersensitive to fluconazole (5,ug-m1-1) are disrupted in ORFs whose S. cervisiae orthologues are involved in histone modification (hat2, ada2, rphl and hosl), chromatin modification (taf14), whereas others are transcriptional activators of genes modulated by levels of purines (basl), aromatic amino-acids (aro80), or the levels of haem and oxygen (hap1). Some fluconazole sensitive strains are disrupted in ORFs that encode proteins belonging to multi-subunit regulators of RNA polymerase II (ssn6) such as the SAGA complex (spt8) and the mediator complex (ssn2)
Hierarchical clustering of midpoint estimates was used to classify strains based on their growth in untreated YEPD, and YEPD supplemented with fluconazole (5-10pg•ml-1). Where midpoint estimates were missing, due to insufficient growth, data were imputed and set to an arbitrary value beyond the detectable range (15 hours), thus artificially plac-ing hypersensitive strains more closely together than may actually be the case if the range of timepoints was increased. Nevertheless, hierarchical clustering of midpoints identified seven categories of null mutant strains, two of such categories are shown in Figure 5.4.
Although the initial aim was to fully characterize the logistic growth of C. glabrata strains, growth curves (Figure 5.2, 5.5 and 5.7) show that stationary phase is not reached in every strain with the experimental method adopted. To achieve improved logistic fits, the experimental system can be adjusted to increase the range of sampling times or to increase the incubation temperature so that growth midpoints are decreased. However, comparison of phenotypic traits at this elevated temperature may need to be complemented with comparison at 30°C in order to identify temperature dependent growth sensitivities in S. cerevisiae.
To be practicable, a longer time-course assay requires automation, the initial results of

5.5 Discussion 182
which have shown promise, in terms of the reproducibility of data, the lack of contamination (Figure F.1) and the definition of the stationary phase of growth (Figure F.2). With a robotic platform, the assay can run overnight without supervision and be extended to 20 hours. In initial experiments, only three strains out of 96 failed to reach stationary phase after incubation in YEPD, for 20 hours (37°C, 300 RPM).
Moreover, it was possible to identify 32 strains that saturated at significantly lower optical density (< 0D600 9.04 - 10.05, p < 0.05) in comparison to the parental strain, and to identify 16 sensitive strains that showed logistic growth but with a greater midpoint than the parental strain (> 4.12 — 5.22 hours, p < 0.05). Unfortunately, this system has not yet been deployed in a screen of fluconazole sensitivity because it is awaiting repair.
Two alternate approaches, which disregard the requirement for a logistic model and make use of the existing data, are a) to estimate the fluconazole dependent inhibition of growth at a single time point, or b) to use the range of time points that determine the exponential phase of growth and therefore do not require a defined stationary phase. In a), the inhibitory concentration of fluconazole and the choice of time point must be considered carefully. In this method, only the inhibitory effects of YEPD supplemented with 5 pg•ml-1 fluconazole were considered as the assay showed the least systematic variability at this concentration (Figure 5.3). Furthermore, a time-point that maximizes the signal to noise ratio, was chosen.
In this instance, the Z factor assay quality score (see methods above) was maximized at 11.5 hours (Table F.6). This method identified eight strains with significant (p < 0.05), flu-conazole (5 pg-m1-1) specific growth sensitivity. Four strains were identified as fluconazole sensitive in both the linear regression and logistic fitting method (pdrl, srb8, hap/ -like and rph1). The linear regression method also classified strains differently: two were classified as having growth defects in the logistic method, but fluconazole sensitivity in the linear regression method (nggl, gcn5) (Table F.7 and Table F.8).
5.5 Discussion
In this pilot study, I aimed to develop a quantitative assay that could identify null mu-tant strains whose growth is significantly more susceptible to environmental stress than the parental strain. In this instance we used fluconazole sensitivity as an exemplar con-dition. Although well established methods already exist for assessing microbial sensitivity to antimicrobial compounds (544; 494), we intend this screen to be applicable to a wider number of environmental stress conditions, and aim to quantify differences in growth rate, and population size at stationary phase.
In recent years, quantitative phenotype scores have been adopted proved advantageous.

CG2001
• YEPD 6 YEPD 5ug/m1 Flu o YEPD 1Oug/m1 Flu
2 4 6 8
Time (hours)
(a) ATCC 2001
10
CG2001 his3—
• YEPD o YEPD 6ug/m1 Flu o YEPD 10ugiml Flu
O
2 4 6
Time (hours)
(b) his3
10
Chapter 5. Functional genomics of fluconazole sensitivity in Candida glabrata using a library of transcription factor knock-outs: a pilot study 183
Figure 5.2: Logistic growth curves of a) wild-type strain ATCC 2001 and b) the library parental strain his3 in YEPD and YEPD fluconazole (5-10 µg-m1-1). Panels show optical density measurement (0D600= 0 - 1.5, as a function of time = 0 - 11.5 hours) for both strains, each depicting growth in YEPD (closed blue circles), YEPD + 5 pg-m1-1 fluconazole (open red triangles) and YEPD + 10 µg-m1-1 fluconazole (open black squares). Data points derived from growth assays in higher concentrations of fluconazole are omitted for clarity. ATCC 2001 showed logistic growth in YEPD (midpoint estimate =-8.44 hours, SEM = 0.37 hours) that was inhibited by the addition of 5 µg•ml-1 fluconazole (midpoint estimate = 11.29, SEM = 1.61 hours). Growth was sufficiently impaired in YEPD + fluconazole (10-100 µg•ml-1) to prevent logistic regression estimates of growth parameters. At these higher concentrations of fluconazole (YEPD + 10 iug•m1-1, YEPD + 50 µg-ml-1 and YEPD + 100 pg-m1-1), growth was 41.03%; SEM = 5.00%, 34.02%; SEM =-6.58%, 30.80%; SEM = 13.21% of that observed in YEPD without fluconazole respectively. The his3 parental strain also showed logistic growth in YEPD (midpoint estimate = 9.39 hours, SEM = 1.33 hours) and YEPD + 5 pg-m1-1 (midpoint estimate = 11.32, SEM = 4.62 hours), and a pattern or failed logistic regression fits in YEPD + fluconazole (10-100 ,ug•m1-1). Growth of his3 at these higher concentations of fluconazole (YEPD + 10 tig-m1-1,YEPD + 50 pg•ml-1 and YEPD + 100 pg-m1-1) was 62.46%; SEM = 14.65%, 35.21%; SEM = 9.38% and 34.68%; SEM = 10.13 respectively.

0 0
0
0 0 0
0 o 0
5.5 Discussion 184
0 5 10 50 1000 Fluconazole concentration (µg/m1)
(a)
Number of strains
0 5 10 50 100 Fluconazole concentration (µ,g/m1)
(b)
Figure 5.3: Fluconazole dependent distribution of midpoint standard errors and number of inestimable growth midpoints. (a) Box and whisker plots depict increasing median standard error as a function of increasing fluconazole concentration. Median stan-dard errors of midpoint estimates ranged from 4.68 (5pg•ml-lml fluconazole) - 76 hours (100pg•ml-lml fluconazole). (b) The number of strains showing insufficient growth for estimation of midpoint also increased with fluconazole concentration, reaching a peak at 10µg•m1-1 fluconazole.
600
200-
a
r
100
80
60
40
20
0

yeD
!Ot4171 FN
IT C 0
O
•TT
O O
Chapter 5. Functional genotnics of fluconazole sensitivity in Candida glabrata using a library of transcription factor knock-outs: a pilot study 185
Figure 5.4: A liquid growth phenotype assay identifies two clusters of fluconazole-sensitive null mutant strains. A library comprising wild-type and parental auxotrophic strains and 186 C. glabrata null mutants, each disrupted in one putative transcription factor ORF (based on sequence homology with S. cerevisiae) was incubated overnight in an orbital shaker to stationary phase (18 hours, 30°C, 180 RPM, in YEPD broth) and then sub-cultured (18 hours, 30°C, 180 RPM, in YEPD broth) in micro-titre plates containing YEPD broth ± fluconazole (5 - lOug /ml). Optical density (0D600) measurements were bi-hourly recorded on a Multiskan Ascent absorbance plate-reader (Thermo Scientific, U.S.A), and data were fitted to a three parameter (slope, midpoint and upper asymptote) logistic function of growth using non-linear regression in the R statisitical computing framework. Hierarchical clustering of midpoint estimates was used (average linkage, Euclidean distance) to classify strains based on their growth in untreated YEPD, and YEPD supplemented with fluconazole (5-10 pg•ml-1). Where midpoint estimates were missing, due to insufficient growth, data were imputed and set to an arbitrary value beyond the detectable range (15 hours). Two clusters, Cl and C5, showing fluconazole sensitivity and general growth defects respectively are depicted. Midpoint estimates are depicted in a hue scale rangeing from red (low) to white (highh)

4
T.;•,JJ. • A
y I
(j) hose
5.5 Discussion 186
AMP 1131.1,
„.—
(a) pdr 1
(b) mig 1
(c) aro80 (d) ashl
MR1-11110
• la
(m) sfpl (n) srb8
(o) ssn2 (p) ssn3
(e) cti6
(i) hosl
sFP
(f) hap/-like (g) hap. (h) hate
(1) rphl.
„..„-j KM. I0.111 Raj
1.1041 4089
1
Figure 5.5: Growth curves of strains belonging to a cluster (C1) characterized by wild-type like growth in YEPD but sensitivity to YEPD fluconazole (5-10 pg•ml-1. Panels show optical density measurement (0D600= 0 - 1.5, as a function of time = 0 - 11.5 hours) for twelve null mutant strains, each depicting growth in YEPD (closed blue circles), YEPD + 5 fig•m1-1 (open red triangles) and YEPD + 10 tzg-m1-1 (open black squares). Data points derived from growth assays in higher concentrations of fluconazole are omitted for clarity.

(c) nggl (d) pop2
D06.1.1616,0
(a) gcn5 (b) mksl
(h) snf5
ID
(g) rtg2 (e) rpd3 (f) rtfl
11••• i•••441
• I
EICH11 011410 11113011 1•0162
411.0111
. . .
66•• 64..1
repo g:=1L,
a 1
51.6
• •66o
(i) snf6 (j) sPt3 (k) spt8 (I) ssn6
:
• A m ;
, 4 I
Chapter 5. Functional genomics of fluconazole sensitivity in Candida glabrata using a library of transcription factor knock-outs: a pilot study 187
Figure 5.6: Growth curves of strains belonging to a cluster (C5) characterized by slow growth in YEPD and YEPD fluconazole (5-10 pg-m1-1). Panels show optical density measurement (0D600= 0 - 1.5, as a function of time = 0 - 11.5 hours) for twelve null mutant strains, each depicting growth in YEPD (closed blue circles), YEPD 5 pg-m1-1 (open red triangles) and YEPD 1- 10 itg-m1-1 (open black squares). Data points derived from growth assays in higher concentrations of fluconazole are omitted for clarity.

(a) crzl (b) cst6 (c) hon.! (d) met31
Prnonaoai 7.4 ,••,••1
1
KU,
1
(f) ph023
(j) spt2
(g) pip2
• • •
(k) sutl (1) tupi
(e) msn4
10
(i) snf4
(h) sir4
1
BO
5.5 Discussion 188
Figure 5.7: Growth curves of strains belonging to a cluster (C3) characterized by wild-type like growth in YEPD and YEPD fluconazole (5-10 pg•ml -1 ). Panels show optical density measurement (0D600= 0 - 1.5, as a function of time = 0 - 11.5 hours) for twelve null mutant strains, each depicting growth in YEPD (closed blue circles), YEPD + 5 µg•m1-1 (open red triangles) and YEPD + 10 pg•ml-1 (open black squares). Data points derived from growth assays in higher concentrations of fluconazole are omitted for clarity.

Chapter 5. Functional genomics of fluconazole sensitivity in Candida glabrata using a library of transcription factor knock-outs: a pilot study 189
Synthetic genetic array scoring has facilitated correlation maps between proteins that al-low cross species comparison (178) as well as guilt by association functional predictions of uncharacterized S. cerevisiae proteins (178). The methods developed in the Andrews lab have relied on image analysis of colony size, although the SGA scoring algorithm and software analysis has yet to be published, this may be something that we re-visit in the future. Furthermore, although the strains have been barcoded, and are amenable to fitness profiling using the microarray method described previously (239); we proceeded with the growth curve assay on economic grounds and, more importantly, because strains do not need to be pooled before the assay.
Optical density measurements are used in lieu of cell counts, although currently there exists no calibration curve relating cell number to the optical density on our absorbance measurement instrument, this is an important technical requirement for future screening. Furthermore, by using optical density in place of cell counts I have made an assumption that differences in optical density between strains exposed to stress over a timecourse are due only to population size. However, it is conceivable that null mutations can confer differences in size and shape, which can affect optical density measurements. Although I expect cell morphological differences to be a small contributing factor to differences in optical density measurements, future work should include a comparison of the morphological characteristics of the null mutant strains in normal culture medium and culture medium exposed to fluconazole.
The manual methods initially adopted here suffered from variability and a poorly defined stationary phase in most strains. The initial development of a robotic screening platform has addressed these two problems, and allows us to perform high throughput screening. The reliability of midpoint estimates of 96 parental strain growth curves conducted on this platform is good (with a midpoint coefficient of variance of 5.23%). We anticipate this platform to be the basis for a large scale test of null mutant strains of C. glabrata and strains deleted in orthologous S. cerevisiae genes in a large number of environmental stress conditions.
Extending the screen to incorporate different environmental stress conditions is a vital step in dissecting the functional role of transcription factor ORFs in environmental stress adaptation, at least two of which are likely to be pleiotropic (e.g. Msn2p and Msn4p regulate the expression of approximately 200 genes in response to several stresses, including heat shock, osmotic shock, oxidative stress, low pH, glucose starvation, sorbic acid and high ethanol concentrations (446; 230; 118).
In this pilot study, fluconazole, a specific inhibitor of lanosterol C14-demethylase (766), acts to knock down the activity of a single gene; as a result, sensitive or hypersensitive null mutant strains effectively reveal synthetic sick or synthetic lethal interactions with

5.5 Discussion 190
disrupted ergosterol biosynthesis. Such synthetic interactions may occur in the ergosterol biosynthetic pathway itself ((20; 623)), in a dependent process (e.g. calcium signalling (347; 707), respiration (369), iron uptake (556)), or in downstream processes that are affected by ergosterol levels in the plasma membrane (e.g tryptophan permease targeting to the plasma membrane (706)). The dissection of this phenotype is further complicated by the type of genes profiled in this analysis. Null mutants like those profiled in this pilot study -transcription factors - are likely to be regulators of one or more of those pathways, rather than effectors in the pathway.
Further work should assess the growth of these null mutants in a range of antibiotic agents, including related azole compounds (e.g. itraconazole, posaconazole, ketoconazole and voriconazole) as well as antibiotics that do not target the ergosterol biosynthetic path-way (e.g. cyclohexamide, dexamethosone, hygromyin, amphotericin B). This will help to identify which transcription factors are specifically involved in azole sensitivity. Further-more, fitness profiling in conditions that identify respiratory deficient mutants will be nec-essary. Typically, respiratory mutants have the petite phenotype (96; 163), and therefore spotted colony phenotypic screens on 3% yeast peptone glycerol agar (in addition to the liquid screens described) should be performed; other sole carbon sources such as ethanol, acetate and 2-deoxyglucose should be considered (357; 127; 408). Null mutants with an iron auxotrophy can be identified either with dropout synthetic medium or by adding chelators bathophenanthrolene disulfonic acid and ferrozine to the YEPD medium (556).
Apart from adding the suggested environmental stress conditions to the phenotypic screen, the function of each of these transcription factors may be further elucidated by whole genome experimental approaches. It will be beneficial to compare the transcriptional changes that occur after exposure to fluconazole between parental and null mutant strains using gene expression microarrays. These experiments will help to define the sets of genes that are induced and repressed in response to fluconazole exposure, and how they depend on each transcription factor (e.g. (719)). Chromatin immunoprecipitation in tandem with gene expression microarray profiling (CHiP-chip analysis) may also complement the gene expres-sion microarray approaches described above (577; 27). Although null mutant libraries are informative tools in functional genomics, they only identify a subset of regulatory influences, that can be complemented by overexpression libraries (334; 82).
In this screen, I identified two clusters of strains that were defined by either an impaired growth profile in YEPD (C5) or YEPD supplemented with fluconazole 5 - 10 iug•ml-1 flu-conazole (C1). Several fluconazole sensitive C. glabrata strains identified in this study were disrupted in ORFs whose S. cerevisiae orthologues encode components of the C. glabrata RNA polymerase II mediator complex (CAGLOE00627g/SRB8, CAGLOJ10472g/SSN2 and CAGLOM06875g/SSN8), a requirement for activator-dependent stimulation of RNA poly-

Chapter 5. Functional genomics of fluconazole sensitivity in Candida glabrata using a library of transcription factor knock-outs: a pilot study 191
merase II (349; 206) and Tupl-Ssn6 co-mediated transcriptional repression (641; 768). A specific co-operative role for a mediator complex subunit in directing xenobiotic resis-
tance gene expression was recently discovered in S. cerevisiae and C. glabrata (687). After studying orthologous multi-drug resistance pathways in mammalian cells, where the preg-nane X receptor upregulates ABC transporters after binding structurally diverse xenobiotics (360; 744), Thakur et al., demonstrated that the S. cerevisiae and C. glabrata multidrug resistance transcription factor Pdrlp binds to a related antifungal - ketoconazole - and that deletion of the mediator subunit Gall 1p results in hypersensitivity to this compound.
In their screen of S.cerevisiae mutant strains, Thakur et al., also profiled other medi-ator subunit mutants for ketoconazole hypersensitivity (ssn2, srb8, sr1)2, cse2, ssn8, pgdl, medl, sohl, sin4 and srb5) but did not identify any additional hypersensitive strains. In this study, a C. glabrata srb8 strain was identified as fluconazole sensitive. Although C.
glabrata srb8 mutants were not screened for fluconazole sensitivity in (687), a separate study identified three other hypersensitive strains disrupted in ORFs encoding mediator subunits, including SRB8 (347). In addition, two other mediator subunit mutants from the CGTFKOlib profiled here(ssn2 and ssn8), were also fluconazole sensitive, suggesting a different role for these mediator subunits in fluconazole versus ketoconazole susceptibility, or intriguingly between C. glabrata and S. cerevisiae.
Zinc cluster proteins (434) and basic leucine zipper proteins have a more established mechanistic role in the regulation of multi-drug resistance gene expression (? ). Therefore, it was not surprising to identify several fluconazole sensitive strains that were disrupted in ORFs encoding zinc finger domain transcription factors. The role of Pdrlp in regulat-ing the expression of multi-drug transporters, which confer fluconazole resistance, is well known in both S. cerevisiae (433) and C. glabrata (719). Other well established fluconazole dependent processes were fluconazole sensitive. Ergosterol synthesis in S. cerevisiae is an energy expensive, aerobic process (772) requiring haem (249; 38; 422) and molecular oxy-gen (422; 598). Perhaps unsurprisingly, this study identified fluconazole sensitive mutant strains lacking ORFs whose S. cerevisiae orthologs have regulatory roles in these processes (CAGLOK05841g/hap/ (286; 679), CAGL0001485g/cti6 (560; 149) and CAGLOA01628g I mig.1 (427), reviewed in (607)),In contrast, there are no previous reports of fluconazole sensitivity in an aro80 strain in C. glabrata (CAGLOF03025g), S. cerevisiae or C. albicans. Although a role for a transcriptional regulator of aromatic amino acid catabolism in flucona-zole sensitivity is intriguing, it remains unclear why aro80 strains are more susceptible to fluconazole than the parental his3 strain. In S. cerevisiae, Aro8Op positively regulates the transcription of the aromatic transferase II Aro9p which catalyzes the first step of trypto-phan, phenylalanine and tyrosine (317) catabolism. Previous work in S. cerevisiae reveals a link between depleted ergosterol levels and the improper targeting of the high affinity

5.6 Chapter Summary 192
tryptophan permease in the plasma membrane (706). Thus, a plausible explanation for the fluconazole sensitivity observed in the aro80 strain might be that reduced ergosterol levels in the cell membrane lead to impaired tryptophan uptake, which might otherwise be rescued by an intact aromatic acid catabolic pathway, governed by Aro80p. This can be investigated by supplementing the YEPD + fluconazole test condition with a tenfold excess of tryptophan, or by over expressing the genes encoding effectors downstream of Aro80p, ARO9 and AR010, in a aro80 background.
5.6 Chapter Summary
This preliminary work has provided the basis for the development of an automated plat-form for functional genomics studies in C. glabrata although significant optimization is still required. A medium throughput knockout methodology has been adopted to differentiate between wild-type and null mutant growth rates in medium supplemented with fluconazole, although this can be extended to other environmental cues. This study has identified some candidate genes that may play a role in fluconazole sensitivity that have not previously been reported in the S. cerevisiae literature.

193
Chapter 6
Conclusions and future work
6.1 Future work
The following points for further research are applicable to chapter 3:
1. In chapter 3 we constructed a Candida glabrata hog1 null mutant and performed parallel phosphorylation, growth and transcription profiling assays in cells that were either mock treated or exposed to hyperosmotic stress (sorbitol, 15 minutes). The hogl mutant showed simultaneous growth and phosphorylation defects in relation to the parental strain, and wild-type equivalent growth and Hog1p phosphorylation in rich culture medium (YEPD).
2. Although this implicated Hog1p phosphorylation in hyperosmotic stress adaptation, no direct link was demonstrated. The evidence in support of Hog1p phosphorylation mediated hyperosmotic stress adaptation would be greatly enhanced by preventing phosphorylation of conserved target residues (Thr174 and Tyr176) using site directed mutagenesis, or by overexpression of the specific Hog1p phosphatase PTC1. To be consistent with our hypothesis, we would expect such an expressed and mutated Hog1p allele to confer normal growth in rich culture medium, and conversely, abrogated phosphorylation and growth in culture medium supplemented with sorbitol (1 M).
3. Transformation protocols adopted in gene disruption can be mutagenic and introduce secondary site mutations that confound the interpretation of phenotypic results. In order to confirm that the hyperosmotic stress induced growth defect is indeed spe-cific to HOG1 deletion, phosphorylation, growth and transcription profiling assays should be performed with the reconstituted strain. If the observed phenotypic and transcription traits are specific to HOG1 deletion, a reconstituted strain would res-cue wild-type behaviour in the above assays. Heterologous complentation assays of

6.1 Future work 194
S. cerevisiae hogl strain would also provide evidence that the gene has a similar biochemical profile when expressed in a heterologous host.
4. Apart from the secondary mutations arising from mutagenic transformation proce-dures, gene deletion methods can lead to multiple (non-targeted) sites of cassette integration. It will be prudent to a) confirm the genotype of the HOG1 locus null by using a PCR based method (upstream HOG1 and internal HOG1 primer pair versus upstream HOG1 and internal HISS pair and b) perform a Southern analysis using a HISS probe directed to genomic DNA extracted from the ATCC 2001, AHT6 and JG7 to confirm a single site of integration in the JG7 strain.
5. Gene expression microarray profiling identified a radically different osmostress tran-scriptome to published data for Candida albicans and S. cerevisiae (references in chapter 3), however comparing gene expression data sets across laboratories and mi-croarray platforms does not adequately control for different sources of systematic noise. In order to make more convincing comparisons with related species, the tran-scription profiling should be performed in parallel with equivalent S. cerevisiae and C. albicans strains. A particularly good example of this approach was recently published by Lelandais et al., (402) who simultaneously performed transription profiling of S. cerevisiae and C. glabrata after benomyl exposure.
6. During the course of the study, a truncated SSK2 allele was identifed as the cause of the functionally impaired Slnl branch of C. glabrata (references in chapter 3), and this was not evident in two other common C. glabrata strains. Prior to our transcription profiling we hypothesized that Hog1p phosphorylation would be sufficient to recover a S. cerevisiae-like transcription profile if the downstream transcriptional modules are dependent on Hog1p and conserved in C. glabrata, We rejected this hypothesis on the evidence of the observed transcript profiles, however the fact that the specific architecture of the ATCC 2001 strain is not generalizable to the whole species, raises the question of whether these transcript profiles are an aberration, peculiar to the ATCC 2001 strain background. The study would be strengthened by profiling other mutants derived from other C. glabrata strain backgrounds such as BG2, that have both signalling branches in tact or by reconstituion of a functional SSK2 gene either episomally or at the chromosomal locus. HOG1-dependent transcripts should be the same in the ATCC 2001 and BG2 backgrounds if this depends only on Hoglp phosphorylation rather than a specific background.
7. Assertions regarding the transcriptional activity of important transcripts and their STEII and HOG1 dependency should be independently verified with non-microarray

Chapter 6. Conclusions and future work 195
based methods that ideally use the same RNA samples, e.g qFCR or Northern blot analysis.
8. Apart from the technical verification steps above, there are some potentially interest-ing biological phenomena that should be investigated. Several transcriptional regula-tors are induced wild-type ATCC 2001 cells exposed to 1 M sorbitol. CIN5 is expressed in hogl and sten cells exposed to 1 M sorbitol. Inspection of CHiP-chip data for S. cerevisiae reveals that Cin5p binds upstream of GPD1, a key glycerol biosynthetic enzyme. Assuming that C. glabrata Cin5p binds to the orthologous GPD2 promoter region, there may exist a HOG1-independent regulator of GPD2 transcription and glycerol biosynthesis. Electrophoretic mobility shift assays should be designed to as-sess if C. glabrata Cin5p binds to the upstream GPD2 promoter region. A cin5 null mutant should also be constructed. GPD2 transcript levels should be measured anal-ysis in ATCC 2001 and cin5 cell extracts grown in rich culture medium ± sorbitol (1 M)
9. To further investigate the role of CIN5 in glycerol production, internal glycerol ac-cumulation assays should be performed in cin5 and gpd2 cells grown in rich culture medium ± sorbitol (1 M). Since internal glycerol accumulation is a net effect of glyc-erol biosynthesis and efflux, the study may have higher resolution if the efflux system is impaired. Accordingly these experiments may need to be performed in an fpsl background or in strains that have a constitutively closed Fps1p aquaglyceroporin.
10. The analysis of the STE11 and HOG1 dependency of the transcriptional response highlighted four major sets of genes: a) a set of genes that were dependent on the extant Shot signalling branch, b) a set of genes that were not dependent on HOG1 or STE11, c) a set of genes that were specifically dependent on STE11 and d) a set of genes that were specifically dependent on HOG1. Analysis of over-represented GO terms showed that the Shol branch dependent genes predominantly encode genes in-volved in metabolism, including glycerol biosynthetic genes. Those genes that were independent of HOG1 and STE11 contained calcineurin signalling components sug-gesting that the calcineurin pathway is not under the control of Hog1p or Ste11p. The STE11 dependent genes were predominantly associated with cell wall remodelling and oxidative stress protection whereas the HOG1 dependent genes predominantly encoded genes involved in proteolysis.
11. The induction of genes involved in cell wall remodelling, both at the level of signalling and catalysis suggests that changes in the cell wall may be part of the adaptive response to hyperosmotic stress in C. glabrata. Notably, genes encoding signalling

6.1 Future work 196
components of the calcineurin and protein kinase C cell integrity pathways are co-induced with their downstream targets (see data in Chapter 3). To individually test whether transcriptional induction of the PKC cell integrity signalling genes SLT2, MID2 and BCK1 is required for proliferation in hyperosmotic stress null mutants of these strains should be constructed and tested for growth on agar supplemented with 1 M sorbitol or 1 M Na Cl. Assuming that deletion of a single component of the PKC cell integrity pathway is sufficient to disrupt its function, one would expect all of these strains to be sensitive to the above supplementations if this pathway is required for the hyperosmotic stress adapation in C. glabrata. A similar experiment can be conducted by chemical epistasis if such knockout strains prove hard to obtain. The protein kinase C pathway can be inhibited by supplementation of the culture medium with caffeine (1- 10 mM) (267). If the protein kinase C pathway is required for osmoadaption then wild-type C. glabrata cells grown in 1 M sorbitol and caffeine (1- 10 mM) will have more impaired growth than strains cultured in sorbitol alone or caffeine alone. Physical evidence of cell wall remodelling can also be investigated by assaying chitin levels qualitatively and quantitatively using calcofluor white staining and (722; 488)
The following points for further research are applicable to chapter 4:
1. In chapter 4 we generated multiple gene expression profiles from minute samplings of A. fumigatus germlings during initiation of murine infection in order to identify genes preferentially expressed during adaptation to the mammalian host niche. This analy-sis revealed a highly co-ordinated A. fumigatus gene expression programme, governing metabolic and physiological adaptation, which allows the organism to prosper within the mammalian niche. As functions of phylogenetic conservation and genetic locus, 28% and 30%, respectively, of the A. fumigatus subtelomeric and lineage-specific gene repertoires are induced relative to laboratory culture, and physically clustered genes including loci directing pseurotin, gliotoxin and siderophore biosyntheses are a promi-nent feature. Locationally biased A. fumigatus gene expression is not prompted by in vitro iron limitation, acid, alkaline, anaerobic or oxidative stress. However, sub-telomeric gene expression is favoured following ex vivo neutrophil exposure and in comparative analyses of richly and poorly nourished laboratory cultured germlings. We found remarkable concordance between the A. fumigatus host-adaptation tran-scriptome and those resulting from in vitro iron depletion, alkaline shift, nitrogen starvation and loss of the methyltransferase laeA, a putative global regulator of sec-ondary metabolite production.

Chapter 6. Conclusions and future work 197
2. The regulation of such secondary metabolite production is unclear, however envi-ronmental signals such as pH changes and carbon availability may alter production of metabolites through the actions of proteins such as PacC (146). Although tran-scriptional regulation of secondary metabolites by cis-acting transcriptional regulators which simultaneously activate or repressall the genes of the cluster has been described (74), chromatin regulation is also thought to play a significant role. Deletion of the laeA gene has been shown to result in the loss of numerous secondary metabolites (75; 539)), and transcriptional comparisons between in vitro cultures of wild-type and
laeA strains using microarrays revealed 13 out of 22 secondary metabolite clusters were down regulated in the AlaeA mutant. In these clusters, regulation by laeA is limited to genes within the cluster and not to surrounding genes (539). Additionally, the laeA protein contains motifs which are similar to histone methlytransferases, sug-gesting laeA regulation via chromatin remodelling. Interestingly, the AlaeA strain shows attenuation in murine models of virulence, with no significant difference in in vitro growth rates (73). It is presumed that the attenuation of virulence is in some way related to the decrease in multiple toxin production.
3. A clearer mechanistic understanding of the putative epigenetic control of transcription and secondary metabolite production may elucidate a major mechanism of A. fumi-gates adaptation within the host and also identify future candidates for therapeutic intervention. Future work should focus on the functional role of LaeA in histone mod-ification and its mediatory role in transcription within the host. The first step will be to evaluate those LaeA dependent secondary metabolism clusters by comparing the virulence and transcription profiles of AlaeA strain and wild-type strains in vivo. In addition, the role of secondary metabolite production in virulence can be more specifically addressed by creating mutants with deficiencies in secondary metabolite production. For example ApsoA and AftmA lack a hybrid NRPS/PKS gene and a NRPS gene respectively. The virulence of these strains should be investigated in both cyclophosphamide and hydrocortisone treated models of murine infection.
4. Whilst our initial investigations are a snapshot of the fungal response to the host, relative to a nutrient rich in vitro culture, a more detailed time-series investigation of transcription in wild-type and A.laeA strains, coupled with NMR and LCMS analy-sis of secondary metabolite production will determine whether secondary metabolite synthesis is stage-specific and laeA-dependent.
The following points for further research are applicable to chapter 5:
1. In chapter 5, we showed that targeted disruptions of C. glabrata open reading frames

6.1 Future work 198
can be achieved systematically. We adopted these methods to disrupt orthologues of S. cerevisiae transcription factors in C. glabrata. We used these genetic tools for a functional genomics study of fluconazole susceptibilty in order to identify novel mediators of resistance mechanisms.
2. Sensitive strains should now be profiled in clinically acceptable sensitivity assays such as the NCLSS (138) or EUCAST (588) microbroth methods. Furthermore, several mechanisms involved in fluconazole sensitivity have been previously described (see (97)). Sensitive strains should therefore be profiled for respiratory deficiency by growth on yeast peptone glycerol agar or by cytometric analysis of rhoadmine 123 accumulation in the mitochondria; in addition the ergosterol content of strains should also be measured to rule out any defects in ergosterol biosynthesis that may be com-pounded by fluconazole treatment.
3. Gene expression microarray experiments of sensitive transcription factor null strains can be performed to define the regulon of these genes with and without exposure to fluconazole to further define the processes that are controlled by these genes, which may identify novel pathways involved in fluconazole resistance.

Factor Fungal Pathogens Mucosal and cutaneous barrier disruption
Neutrophil dysfunction
Candida spp. Aspergillus spp
Candida spp. Aspergillus spp.
Trichosporon
Defects in cell-mediated immunity Cryptococcus endemic mycoses
Metabolic disorders Candida spp. Zygomycetes
Exposures Aspergilllus spp
endemic mycoses
Extremes of age (< 1 > 70 years) Candida spp
199
Appendix A
Chapter 1 Supplementary Data
Table A.1: Factors involved in the development of opportunistic mycoses. Par-tial list of pre-disposition categories involved in the development of opportunistic mycoses. Taken from (542)

Ketoc663zoie
Itracanazole
Posacannale
Yori tale
Flucoataanie
1450 1972 1976 1983 15146 1990 15051 6
Appendix A. Chapter 1 Supplementary Data 200
KEY: Anti-metabolite Atoles
2006
Figure A.1: Licensing timeline for antifungal drugs with systemic bioavailability. Data taken from Drugs@FDA http://www.accessdata.fda.gov/scripts/cder/drugsatfda/

A B
2003 1998 1999 2000 2001 2002 2003
year
1998 1999 2000 2001 2002
year
C
ro
.0 a
2000 2001 2002 2003
Appendix A. Chapter 1 Supplementary Data 201
year
Figure A.2: Species specific incidence rates of invasive candidiasis. Aetiological agents of invasive candidiasis (IC) and their associated incidence rates. Incidence data taken from (542) indicate Candida albicans is the primary aetiologial agent of invasive candidiasis (IC) (A), its prevalence decreased between 1998 and 2003, with concomitant increases in non-albicans species Candida glabrata, Candida tropicalis, Candida parapsilop-sis and Candida krusei (B). The shaded panel in (B) denotes the period in which new aetiological agents rapidly emerged as causes of IC (C).
5 f,
0 0 - •9
ro a
.,,
2
o
— C. albicans - - C.glabrata — C.tropiCalis - - C.parepsilopsis — C.krusei
— C. inconspicua - C.norvengsis
— C.dubliniensis Clipolytica
— C.zelanoides - - C.pelliculose

Amphotericin B VON, 0
N CI
0 OH rTh,
CI 0
Itraconazole
CI
Appendix A. Chapter 1 Supplementary Data 202
off
Ketoconazole
N
F
Fluconazole
---1 Hydroxy-methyl-glutaryl (HMG)-CoA)
1,1vIG-CoA Reductase
Al Iylamines
Mevalonic acid
Squalene
I Squalene epoxidase IERG/)
Lanosterol
A"— saturase (ERG3) Azoles ---
eplethyl-lanosterol cp1,-,d
L1-1-t eductase (ERG24)
Fecosterol
- .sorne rase (ERG21
Polyenes_
Episterol
Ergosterol
Figure A.3: Common antifungal drug molecules and their mechanism of action. Structure of commonly used antifungals (A) and schematic represenation of the mechanism of action of selected classes of antifungal drugs affecting the ergosterol biosynthesis pathway (B). Biochemical precursors are depicted as blue rectangles; the enzymes catalyzing reaction steps are shown to their right with encoding genes in parentheses. Antifungal drug classes are reported in grey outlined rectangles to the left of the relevant biosynthetic step, or in the case of polyenes directly toward the molecular target. Adapted from Lupetti et al (426)
C14a demethylase (ERG11)
14a-methyl-3,6-diol Amo ro Hine

Blofilm Formation
Fit. rrent ati on
51, PEP1-7, /PC CST20
T Issue Invasion
R.4052,1E11
1,
Endosomal Transport
Cell Wall integrity
CHS2,CHS.i.1114. G5C1,440041
COMPit475 JradEMVPi
CCCI. CTIY3 FRE10,F1,112 GC/44,6,01 I801,11ArG1, PEP7A4P1-8 Str1.114,1,
SAP1-8
Adhesiori
71,5t 11 PAS1
Al A. VPSJ
CiAL I
Appendix A. Chapter 1 Supplementary Data 203
Figure A.4: Schematic summary of Candida albicans virulence factors stored in the CGD database. Gene names bounded by a white box are required for the associated function, which is denoted by the caption in the grey box, but are not necessary or virulence. Gene names bounded by the red boxes are required for virulence. Data sourced from CGD.

Appendix A. Chapter 1 Supplementary Data 204
N
Y.lipolytica 111.111.1111111111111.1.1.111111.1111111.11
C. g I ab rata 1111111111.111111111111.111.11.111111.111
S.cereyistae
S.kluyveri
A.gossypii
K.lactis 1111111111111111111111111111111•111111111111111111111111111111111111•111
C.lusitanae
C.guilliermondii
D.hansenii
C. parapsilopsis
C.tropicalis 1111111111111111111111111111111111111111 -11111111111.1111111111111111111.11111111111111111111111111111
0 10 20 30 40 50 60 70 80 90
Proportion of conserved virulence genes (%)
en Ascomycota l'arrmi.ra lrlrnlmiia
Candido pirotmitta
Sacchairomycohtna
W OD S Chan' Air '4•1 CM lent!
Sur Y: baron iy4 CA AlikarCle
100 SYa OYA111141.1
ril-ht4r7Nrity.reN rroinvir3 141. 1.11 1117.1T 4' tAtlyitill,11A
Aro (741r. ce.y iiiinur LI: rill irsq.preJni s Iva {
Ita.5-1'ffir(.1711Y'et kiatyvero
‘Way,;.1 gym rai
,5. rit,,vemovy4-ew ?at' A
CC4Flifi4*/ iitraraniar C'artdiriro ,gr4diarrniornlii
100
GTG 00
Dri'xu-viArr5rrr hiamer4 Candider itklritiMike. AA
uruliilaa trrviralit
7C:10 etlablatir bbirli rri 100 Ca 'Oak OHM lAITA
Figure A.5: Proportion of virulence factor (VF) reciprocal best BLAST (587; 16) hits in the Saccharomycotina as a function of evolutionary distance from Candida albicans. Species located in the same subgroup as C. albicans (orange bars), that translate the colon CTG as serine, show a higher proportion of conserved, published VFs than whole genome duplication (WGD) species (green bars).

205
Appendix B
Media Recipes
Table B.1: Recipe for Synthetic Complete culture medium (1 L). Components above the double solid line are added to 800 ml of distilled water and dissolved with the aid of a magnetic stirrer. Solutions are made up to 1 L and agar is added if solid medium is required. Culture media are sterilized by heating at 121°C for 15 minutes.
Constituent Mass (g) Final Concentration YNB - amino acids, - ammonium sulphate
6.78 0.68 % (w/v)
Complete Supplement Medium 0.798 0.0798 % (w/v) d-Glucose 20 2 % (w/v) Agar 20 2 % (w/v)
Table B.2: Recipe for Yeast Extract Peptone Dextrose culture medium (1 L). Components above the double solid line are added to 800 ml of distilled water and dissolved with the aid of a magnetic stirrer. Solutions are made up to 1 L and agar is added if solid medium is required. Culture media are sterilized by heating at 121°C for 15 minutes.
Constituent Mass (g) Final Concentration Bacteriological peptone 20 2 % (w/v) Yeast extract 10 1 % (w/v) d-Glucose 20 2 % (w/v) Agar 20 2 % (w/v)

206
Appendix C
Strain Genotypes
Table C.1: Fungal strains used in this study. The genotype describes the strain back-
ground used and the gene disrupted separated by a ",". Reconstitution is represented by
genes between square braces. Plasmid details are in the accompanying plasmid table.
Species Strain Name Genotype Origin
S. CCT,I4Siae BY4741 MATa Ahis3Aleu2Amet15Aura3 EuroScarf C. glabrata C. glabrata C. glabrata C. glabrata C. glabrata C. glabrata
ATCC 2001 2001AHT6
AM3 AM22 AM25 JG7
Type strain Aura3::pUC18, Atrp/ Aura3,his3::URA3
2001AHT6, ste11,HIS3 2001AHT6, .ste20,HIS3
AM22, pAMC85 [STE20, TRP1] 2001AHT6, hog1,111S3
American Type Culture Collection (734)
(106; 104) (106; 103) (106; 103) This study

207
Appendix D
Chapter 3 Supplementary Data
Table D.1: Oligonucleotides used in the three step PCR strategy for HOG1 disruption in C. glabrata
Oligonucleotide Sequence 5'-3' HOG15-5 GTATCCATACTTGCGTTTTGG HOG15-3 gtcatagctgttt cctgTGTATAGTGTAGTATATCCAAG HOG13-5 tacaacgt cgtgactgggCAGCTCGAATTCTGCAACAA HOG13-3 TTTTGGAGGACTATTTCCGATT
HOG1-HIS5 CTTGGATATACTACACTATACAcaggaaacagctatgac HOG1-HIS3 TTGTTGCAGAATTCGAGCTGcccagt cacgacgttgt a HOG13-ex CGAAGGAGCAACAAAAGAGG HIS3INTF TGGGCAAGGAATATGCTAGTGG CgHOG F1 GCGCGGATCCGAACTGATTAAGATGTTCAGAGACGAGG CgHOG R1 GCGCGGATCCTGAAATATTCGCTAAGGGTATGTAG

Appendix D. Chapter 3 Supplementary Data 208
Table D.2: Biological effects encoded by linear model contrasts
Figure 3.4 ID
Effect Name Biological Effect
1 STE.0 - WT.0 Untreated stell cells versus untreated ATCC 2001 cells
2 HOG.0 - WT.0 Untreated hog1 cells versus untreated ATCC 2001 cells
3 STE.S - WT.S Hyper-osmotic stress treated stell cells versus untreated ATCC 2001 cells
4 HOG.S - WT.S Hyper-osmotic stress treated stell cells versus untreated ATCC 2001 cells
5 WT.S - WT.0 Hyper-osmotic stress treated ATCC 2001 cells versus untreated ATCC 2001 cells
6 HOG.S - HOG.0 Hyper-osmotic stress treated hog1 cells versus untreated hog1 cells
7 STE.S - STE.0 Hyper-osmotic stress treated ste11 cells versus untreated stell cells
8 HOG.0 - STE.0 Untreated hog1 cells versus untreated stell cells
9 HOG.S - STE.S Hyper-osmotic stress treated hog1 cells versus Hyper-osmotic stress treated stell cells

Appendix D. Chapter 3 Supplementary Data 209
Table D.3: Candida glabrata transcripts induced in wild-type cells following ex- posure to YEPD 1M sorbitol for 15 minutes. Induced transcripts are defined as having log2(Cy5) - log2(Cy3) > 2.0 and Benjamini-Hochberg corrected p-values < 0.05. Estimated log-ratios correspond to the WT.S - WT.0 effect described in Table D.2
CG ORF SC ORF gene name Description WT.S - WT.0 CAGLOI10010g YGR142W BTNS v-SNARE binding protein that fa-
cilitates specific protein retrieval from a late endosome to the Golgi; modulates arginine uptake, possi- ble role in mediating pH homeosta-sis between the vacuole and plasma membrane H(+)-ATPase
5.55
CAGL0CO2321g YER037W PHM8 Protein of unknown function, ex- pression is induced by low phos- phate levels and by inactivation of
5.41
Pho85p CAGL0G04433g YJL108C PRM10 Pheromone-regulated protein, pre-
to have 5tpreadnsmembrane dictedt segments; induced by treatment with 8-methoxypsoralen and UVA irradiation Description not
map 5.14
CAGLOE04554g - - 4.96 CAGLOM09713g YMR152W YIMI Protein of unknown function; null
mutant displays sensitivity to DNA damaging agents; the authentic, non-tagged protein is detected in highly purified mitochondria in high-throughput studies
4.9
CAGLOK08338g YOR385W - Description not mapped 4.57 CAGLOA02816g YDR368W YPRI NADPH-dependent aldo-keto
reductase, reduces multi- ple substrates including 2- methylbutyraldehyde and D,L- glyceraldehyde, expression is induced by osmotic and oxidative stress; functionally redundant with other aldo-keto reductases
4.43
CAGLOM06963g YNR034W SOLI Protein with a possible role in tRNA export; shows similarity to 6-phosphogluconolactonase non-catalytic domains but does not ex- hibit this enzymatic activity; ho- mologous to Sol2p, Sol3p, and
4.31
Sol4p CAGLOJ10846g YHR071W PCL5 Cyclin, interacts with and phos-
phorylated by Pho85p cyclin- dependent kinase (Cdk), induced by Gcn4p at level of transcrip- tion, specifically required for
4.09
Gcn4p degradation, may be sensor of cellular protein biosynthetic capacity
CAGL0105390g YPL026C SKSI Putative serine/threonine protein kinase; involved in the adaptation to low concentrations of glucose in-dependent of the SNF3 regulated pathway
3.97
continued on next page

Appendix D. Chapter 3 Supplementary Data 210
continued from previous page CG ORF SC ORF gene name Description WT.S - WT.0
CAGLOM08822g YDR258C HSP78 Oligomeric mitochondria] matrix chaperone that cooperates with
3.93
Ssclp in mitochondrial thermotol-erance after heat shock; able to pre-vent the aggregation of misfolded proteins as well as resolubilize pro-tein aggregates
CAGLOH08151g - - Description not mapped 3.86 CAGLOE00803g YDR171W HSP42 Small heat shock protein (sHSP)
with chaperone activity; forms barrel-shaped oligomers that sup-press unfolded protein aggregation; involved in cytoskeleton reorganiza-tion after heat shock
3.82
CAGLOL05016g YKL072W STB6 Protein that binds Sin3p in a two- hybrid assay
3.64
CAGLOG03883g YLL026W HSP/04 Heat shock protein that cooper- ates with Ydjlp (Hsp40) and Ssalp
3.61
(Hsp70) to refold and reactivate previously denatured, aggregated proteins; responsive to stresses in-cluding: heat, ethanol, and sodium arsenite; involved in [PSI-I-] propa-gation
CAGLOH06523g YIL145C PANG Pantothenate synthase, also known as pantoate-beta-alanine ligase, re-quired for pantothenic acid biosyn-thesis, deletion causes pantothenic acid auxotrophy, homologous to E. coli panC
3.6
CAGLOF08745g YLR327C TAIA10 Protein of unknown function that associates with ribosomes
3.53
CAGLOH08173g YOR028C CIN5 Basic leucine zipper (bZIP) tran- scription factor of the yAP-1 fam- ily, mediates pleiotropic drug re- sistance and salt tolerance; nude-arly localized under oxidative stress and sequestered in the cytoplasm by
3.49
Lot6p under reducing conditions CAGLOG01540g YNL036W NCE103 Carbonic anhydrase; poorly tran-
scribed under aerobic conditions and at an undetectable level under anaerobic conditions; involved in non-classical protein export path-way
3.47
CAGLOM12474g YIL055C - Description not mapped 3.46 CAGLOM07381g YPL053C KTR6 Probable mannosylphosphate
transferase involved in the syn- thesis of core oligosaccharides in protein glycosylation pathway; member of the KRE2/MNT1 mannosyltransferase family
3.45
CAGLOF09119g YHR207C SET5 Zinc-finger protein of unknown function, contains one canonical and two unusual fingers in unusual arrangements; deletion enhances replication of positive-strand RNA virus
3.38
CAGLOK00715g YGR213C RTA1 Protein involved in 7- aminocholesterol resistance; has seven potential membrane-spanning regions; expression is induced un-der both low-heme and low-oxygen conditions
3.3
continued on next page

Appendix D. Chapter 3 Supplementary Data 211
continued from previous page CG ORF SC ORF gene name Description WT.S - WT.0
CAGLOM12320g
CAGLOG00858g
YAL053W
YLR332W
FLC2
MID2
Putative FAD transporter; required for uptake of FAD into endoplas-mic reticulum; involved in cell wall maintenance 0-glycosylated plasma membrane protein that acts as a sensor for cell wall integrity signaling and activates the pathway; interacts with Rom2p, a guanine nucleotide exchange factor for Rho1p, and with cell integrity pathway protein
3.28
3.23
Zeolp CAGL0G00242g YGR281W YORI Plasma membrane ATP-binding
cassette (ABC) transporter, rnul- tidrug transporter mediates export of many different organic anions including oligomycin; similar to hu-man cystic fibrosis transmembrane receptor (CFTR)
3.21
CAGL0102684g YMR285C NGL2 Protein involved in 5.85 rRNA pro- cessing; Ccr4p-like RNase required for correct 3'-end formation of 5.8S rRNA at site E; similar to Ngllp and Ngl3p
3.2
CAGLOK02761g YHLOO2W HSE1 Subunit of the endosomal Vps27p- 3.2 H se cnInert requiredm for sorting of ubiquitinated membrane proteins into intralumenal vesicles prior to vacuolar degradation, as well as for
Description
1p app ed
recycling of Golgi proteins and for-mation of lumens] membranes
CAGLOL09251g - - 3.17 CAGL00O2233g YER042W AfXR / Peptide methionine sulfoxide reduc-
tase, reverses the oxidation of me-thionine residues; involved in oxida-tive damage repair, providing resis-tance to oxidative stress and regu-lation of lifespan
3.14
CAGLOJ00539g YHR030C SLT2 Serine/threonine MAP kinase in- volved in regulating the mainte- nance of cell wall integrity and pro-gression through the cell cycle; reg-ulated by the PKC1-mediated sig-naling pathway
3.13
CAGLOK00539g YPL259C APM1 Mul-like medium subunit of the clathrin-associated protein complex
3.1
(AP-1); binds clathrin; involved in clathrin-dependent Golgi protein sorting
CAGLOD03916g YHR034C P1111 Protein of unresolved function; may function in protein folding and/or rRNA processing, interacts with a chaperone (Hsp82p), two chromatin remodeling factors (Rvblp, Rvb2p) and two rRNA processing factors
3.1
(Rrp43p, Nop58p) CAGLOM12034g YAL038W CDC19 Pyruvate kinase, functions as a ho-
motetramer in glycolysis to convert phosphoenolpyruvate to pyruvate, the input for aerobic (TCA cycle) or anaerobic (glucose fermentation) respiration
3.09
continued on next page

Appendix D. Chapter 3 Supplementary Data 212
continued from previous page CG ORF SC ORF gene name Description WT.S - WT.0
CAGLOH08844g YMR173W DDR48 DNA damage-responsive protein, expression is increased in response to heat-shock stress or treatments that produce DNA lesions; contains multiple repeats of the amino acid sequence NNNDSYGS
3.06
CAGL0104246g YGL162W S UT/ Transcription factor of the 3.04 Zn[II]2Cys6 family involved in sterol uptake; involved in induction of hypoxic gene expression
CAGLOD05082g YLL039C U/34 Ubiquitin, becomes conjugated to proteins, marking them for selec- tive degradation via the ubiquitin-
2.99
26S proteasome system; essential for the cellular stress response; en-coded as a polyubiquitin precursor comprised of 5 head-to-tail repeats
CAGLOH04631g YML007W YAPI Basic leucine zipper (bZIP) tran- scription factor required for oxida- tive stress tolerance; activated by
2.98
11202 through the multistep forma-tion of disulfide bonds and transit from the cytoplasm to the nucleus; mediates resistance to cadmium
CAGLOD05368g YBR253W SRB6 Subunit of the RNA polymerase II mediator complex; associates with core polymerase subunits to form the RNA polymerase II holoen- zyme; essential for transcriptional regulation
2.97
CAGLOF08107g YGR244C LSC2 Beta subunit of succinyl-CoA lig- ase, which is a mitochondrial en- zyme of the TCA cycle that catalyzes the nucleotide-dependent conversion of succinyl-CoA to suc-cinate
2.96
CAGLOB04257g YBRO31W RPL4A N-terminally acetylated protein component (60S) ribosomal subunit, nearly identical to Rpl4Bp and has similarity to
2.96
E. coli L4 and rat L4 ribosomal proteins Description not
o mtahpepedlarge
CAGLOE03025g - - 2.94 CAGLOM02255g YPL 152W RRD2 Activator of the phosphoty-
rosyl phosphatase activity of 2.92
PP2A,peptidyl-prolyl cis/trans- isomerase; regulates G1 phase progression, the osmoresponse, microtubule dynamics; subunit of the Tap42p-Pph2lp-Rrd2p complex
CAGLOJ01595g YPRO15C - Description not mapped 2.91 CAGLOJ00385g YHR049W FSHI Putative serine hydrolase that lo-
calizes to both the nucleus and cy- toplasm; sequence is similar to S. cerevisiae Fsh2p and Fsh3p and the human candidate tumor suppressor
2.89
OVCA2 CAGLOE05632g YOR348C PUT4 Proline permease, required for high-
affinity transport of proline; also transports the toxic proline ana-log azetidine-2-carboxylate (AzC);
2.85
PUT4 transcription is repressed in ammonia-grown cells
continued on next page

Appendix D. Chapter 3 Supplementary Data 213
continued from previous page CG ORF SC ORF gene name Description WT.S - WT.0
CAGLOG04763g YOR107W RGS2 Negative regulator of glucose- induced cAMP signaling; directly activates the GTPase activity of the heterotrimeric G protein alpha subunit Gpa2p
2.84
CAGLOJ00253g YGRO23W MTL1 Protein with both structural and functional similarity to Mid2p, which is a plasma membrane sensor required for cell integrity signaling during pheromone-induced morpho-genesis; suppresses rgdl null muta-tions
2.83
CAGLOJ03256g YER075C PTP3 Phosphotyrosine-specific protein phosphatase involved in the in- activation of mitogen-activated protein kinase (MAPK) during osmolarity sensing; dephospory- lates Hog1p MAPK and regulates its localization; localized to the cytoplasm
2.83
CAGLOL05434g YKR042W UTH/ Mitochondrial outer membrane and cell wall localized SUN family mem-ber required for mitochondrial au-tophagy; involved in the oxidative stress response, life span during starvation, mitochondrial biogene-sis, and cell death
2.83
CAGL0005137g YOL059W GPD2 NAD-dependent glycerol 3- phosphate dehydrogenase, homolog of Gpdlp, expression is controlled by an oxygen-independent signal-ing pathway required to regulate metabolism under anoxic con- ditions; located in cytosol and mitochondria
2.81
CAGLOG07601g YBR287W - Description not mapped 2.79 CAGLOA02046g YOR018W ROD1 Membrane protein that binds the
ubiquitin ligase Rsp5p via its 2 PY motifs; overexpression confers re- sistance to the GST substrate o- dinitrobenzene,zinc, and calcium; proposed to regulate the endocyto-sis of plasma membrane proteins
2.79
CAGLOM01628g YDR389W SAC7 GTPase activating protein (GAP) for Rho1p, involved in signaling to the actin cytoskeleton, null muta-tions suppress tor2 mutations and temperature sensitive mutations in actin; potential Cdc28p substrate
2.77
CAGLOH09812g YDROO6C SOK1 Protein whose overexpression sup- presses the growth defect of mu-tants lacking protein kinase A ac-tivity; involved in cAMP-mediated signaling; localized to the nucleus; similar to the mouse testis-specific protein PBS13
2.77
CAGLOF09163g YER182W FMP10 Putative protein of unknown func- tion; the authentic, non-tagged pro- tein is detected in highly purified mitochondria in high-throughput studies
2.74
continued on next page

Appendix D. Chapter 3 Supplementary Data 214
continued from previous page CG ORF SC ORF gene name Description WT.S - WT.0
CAGLOG01166g YLR348C Did Mitochondria] dicarboxylate carrier, integral membrane pro- tein, catalyzes a dicarboxylate- phosphate exchange across the inner mitochondrial membrane, transports cytoplasmic dicar- boxylates into the mitochondria] matrix
2.7
CAGLOH07557g YGL254W FZF1 Transcription factor involved in sul- fite metabolism, sole identified reg-ulatory target is SSU1, overexpres-sion suppresses sulfite-sensitivity of many unrelated mutants due to hy-peractivation of SSU1, contains five zinc fingers
2.68
CAGLOF06941g YGL062W PYC1 Pyruvate carboxylase isoform, cyto- plasmic enzyme that converts pyru-vate to oxaloacetate; highly similar to isoform Pyc2p but differentially regulated; mutations in the human homolog are associated with lactic acidosis
2.66
CAGLOH02519g YMR253C - Description not mapped 2.65 CAGLOL07480g YBRO66C NRG2 Transcriptional repressor that me-
diates glucose repression and nega-tively regulates filamentous growth; has similarity to Nrglp
2.64
CAGLOM11066g YGR109C CLB6 B-type cyclin involved in DNA replication during S phase; acti- vates Cdc28p to promote initiation of DNA synthesis; functions in for- mation of mitotic spindles along with C1b3p and Clb4p; most abun-dant during late GI
2.64
CAGL0003201g YLL040C VPS13 Protein of unknown function; het- erooligomeric or homooligomeric complex; peripherally associated with membranes; homologous to human COH1; involved in sporula-tion, vacuolar protein sorting and protein-Golgi retention
2.63
CAGLOL00803g YER054C CIP2 Putative regulatory subunit of the protein phosphatase Glc7p, in- volved in glycogen metabolism; contains a conserved motif (GVNK motif) that is also found in Gaclp, Pig1p, and Pig2p
2.62
CAGL0,11.0296g YNL077W APJ1 Putative chaperone of the HSP40 2.62 (DNAJ) family; overexpression interferes with propagation of the [Psi+) prion; the authentic, non-tagged protein is detected in highly purified mitochondria in high-throughput studies
CAGLOK12034g YDR040C ENA 1 P-type ATPase sodium pump, in- volved in Nab- and Li+ efflux to al-low salt tolerance
2.61
CAGLOM01782g YBRO80C SEC18 ATPase required for the release of 2.59 Secl7p during the 'priming' step in homotypic vacuole fusion and for ER to Golgi transport; homolog of the mammalian NSF
continued on next page

Appendix D. Chapter 3 Supplementary Data 215
continued from previous page CG ORF SC ORF gene name Description WT.S - WT.0
CAGLOJ01353g YMR089C YTA.12 Component, with Afg3p, of the mitochondrial inner membrane m-
2.57
AAA protease that mediates degra-dation of misfolded or unassembled proteins and is also required for cor-rect assembly of mitochondrial en-zyme complexes
CAGLOI01100g YOR120W GCY1 Putative NADP(-1-) coupled glyc- erol dehydrogenase, proposed to be involved in an alternative pathway for glycerol catabolism; member of the aldo-keto reductase (AKR) fam-ily
2.56
CAGLOI07007g YGL219C MDM3.4 Mitochondrial outer membrane pro- tein required for normal mito- chondrial morphology and inher- itance; localizes to dots on mi- tochondrial surface near mtDNA nucleoids; ubiquitination target of
2.55
Mdm30p; interacts genetically with MDM31 and MDM32
CAGLOB01991g YDR126W S WF/ Palmitoyltransferase that acts on the SNAREs Snclp, Syn8p, Tlglp and likely on all SNAREs; mem-ber of a family of putative palmi-toyltransferases containing an Asp-
2.55
His-His-Cys-cysteine rich (DHHC- CRD) domain; may have a role in vacuole fusion
CAGLOF07095g YGL122C NAB2 Nuclear polyadenylated RNA- binding protein required for nuclear mRNA export and poly(A) tail length control; binds nuclear pore protein MIplp; autoregulates mRNA levels; related to human hnRNPs; nuclear localization sequence binds Kap104p
2.54
CAGLOH05137g YPL061W ALD6 Cytosolic aldehyde dehydrogenase, activated by Mg2+ and utilizes
2.53
NADP4- as the preferred coenzyme; required for conversion of acetalde- hyde to acetate; constitutively ex- pressed; locates to the mitochon- drial outer surface upon oxidative stress
CAGLOE00649g YCR079W PTC6 Mitochondrial protein phosphatase of type 2C with similarity to mam- malian PP1Ks; involved in mi- tophagy; null mutant is sensitive to rapamycin and has decreased phos-phorylation of the Pdal subunit of pyruvate dehydrogenase
2.53
CAGLOM06347g YBR183W YPC1 Alkaline ceramidase that also has reverse (CoA-independent) ce- ramide synthase activity, catalyzes both breakdown and synthesis of phytoceramide; overexpression confers fumonisin B1 resistance
2.52
CAGLOL09152g YFROO4W BPN11 Metalloprotease subunit of the 19S regulatory particle of the 26S pro-teasome lid; couples the deubiqui-tination and degradation of protea-some substrates
2.52
continued on next page

Appendix D. Chapter 3 Supplementary Data 216
continued from previous page CG ORF SC ORF gene name Description WT.S - WT.0
CAGLOA02024g
CAGLOJ11176g CAGLOJ04466g CAGLOH03729g
YOL025W
YNL176C YLR4I4C YNLOO6W
LAG2
- -
LST8
Protein involved in determination of longevity; LAG2 gene is preferen-tially expressed in young cells; over-expression extends the mean and maximum life span of cells Description not mapped Description not mapped Protein required for the transport of amino acid permease Gap1p from the Golgi to the cell surface; com-ponent of the TOR signaling path- way; associates with both Tor1p and Tor2p; contains a WD-repeat
2.52
2.51 2.5
2.48
CAGLOG07689g YKR014C YPT52 GTPase, similar to Ypt5lp and 2.48 Ypt53p and to mammalian rab5; re-quired for vacuolar protein sorting and endocytosis
CAGLOL04708g YGR111W - Description not mapped 2.48
CAGLOA04081g YLR194C - Description not mapped 2.48
CAGLOL00605g YKL190W CNB1 Calcineurin B; the regula- tory subunit of calcineurin, a
2.46
Cal-l-/calmodulin-regulated type 2B protein phosphatase which regulates Crzlp (a stress-response transcription factor), the other calcineurin subunit is encoded by CNA1 and/or CMP1
CAGLOL10186g YOR052C - Description not mapped 2.45
CAGLOL11572g YHR103W SBE'22 Protein involved in the transport of cell wall components from the
2.43
Golgi to the cell surface; similar in structure and functionally redun-dant with Sbe2p; involved in bud growth
CAGLOE06424g YKL150W MCR1 Mitochondrial NADH-cytochrome b5 reductase, involved in ergosterol biosynthesis
2.42
CAGLOK00891g YGR205W - Description not mapped 2.42
CAGLOJ03322g YER080W AIMS Putative protein of unknown func- tion; the authentic, non-tagged pro- tein is detected in highly purified mitochondria in high-throughput studies
2.4
CAGLOM10263g YCLOO2C - Description not mapped 2.39
CAGLOJ09108g YDL190C UFD2 Ubiquitin chain assembly fac- tor (E4) that cooperates with a ubiquitin-activating enzyme (El), a ubiquitin-conjugating enzyme
2.39
(E2), and a ubiquitin protein ligase (E3) to conjugate ubiquitin to substrates; also functions as an E3
CAGLOD05434g YPRO65W ROX1 Heme-dependent repressor of hy- poxic genes; contains an HMG do-main that is responsible for DNA bending activity
2.38
CAGLOL06028g YGL181W GTS1 Protein that localizes to the nu- cleus and is involved in transcrip- tion regulation; also localizes to actin patches and plays a role in en-docytosis; N-terminus contains an
2.38
ARF-GAP domain and C-terminus contains a Gln-rich domain
continued on next page

Appendix D. Chapter 3 Supplementary Data 217
continued from previous page CG ORF SC ORF gene name Description WT.S - WT.0
CAGLOM14047g
CAGL0004499g
YMR318C
YDLOO6W
ADM;
PTC1
NADPH-dependent medium chain alcohol dehydrogenase with broad substrate specificity; member of the cinnamyl family of alcohol dehydro-genases; may be involved in fusel al-cohol synthesis or in aldehyde toler-ance Type 2C protein phosphatase (PP2C); inactivates the osmosens-ing MAPK cascade by dephospho- rylating Hog1p; mutation delays mitochondria] inheritance; dele- tion reveals defects in precursor tRNA splicing, sporulation and cell separation
2.36
2.35
CAGLOJ04158g YOR220W RCN2 Protein of unknown function; green fluorescent protein (GFP)-fusion protein localizes to the cytoplasm and is induced in response to the
2.35
DNA-damaging agent MMS; phos-phorylated in response to alpha fac-tor
CAGL0104554g YBRO14C GRX7 Cis-golgi localized monothiol glutaredoxin; more similar in activ-ity to dithiol than other monothiol glutaredoxins; involved in the oxidative stress response; does not bind metal ions; functional overlap with GRX6
2.34
CAGLOGD5313g YDR072C IPTI Inositolphosphotransferase, in- volved in synthesis of mannose-
2.34
(inositol-P)2-ceramide (M(IP)2C), the most abundant sphingolipid;, can mutate to resistance to the antifungals syringomycin E and DmAMP1 and to K. lactis zymocin
CAGLOD04752g YPR127W - Description not mapped 2.33 CAGLOB00836g YCL033C - Description not mapped 2.33
CAGLOE02299g YOL013C HRD1 Ubiquitin-protein ligase required for endoplasmic reticulum- associated degradation (ERAD) of misfolded proteins; genetically linked to the unfolded protein response (IJPR); regulated through association with Hrd3p; contains an H2 ring finger
2.31
CAGLOG06886g Y.11.017W - Description not mapped 2.3 CAGLOK03509g YMR110C 11FD1 Putative fatty aldehyde dehydro-
genase, located in the mitochon- dria) outer membrane and also in lipid particles; has similarity to hu-man fatty aldehyde dehydrogenase
2.3
(FALDH) which is implicated in Sjogren-Larsson syndrome
CAGL0G05049g YD11.063W AIM7 Putative protein of unknown func- tion; green fluorescent protein
2.28
(GFP)-fusion protein localizes to the cytoplasm and nucleus; YDR063W is not an essential gene
CAGLOA02321g YDR345C HXT3 Low affinity glucose transporter of the major facilitator superfamily, expression is induced in low or high glucose conditions
2.28
continued on next page

Appendix D. Chapter 3 Supplementary Data 218
continued from previous page CG ORF SC ORF gene name Description WT.S - WT.IJ
CAGLOA03058g YDR379W RGA2 GTPase-act ivating protein for the polarity-establishment protein
2.27
Cdc42p; implicated in control of septin organization, pheromone response, and haploid invasive growth; regulated by Pho85p and Cdc28p
CAGL0000363g YJL174W KRE9 Glycoprotein involved in cell wall beta-glucan assembly; null muta- tion leads to severe growth defects, aberrant multibudded morphology, and mating defects
2.27
CAGLOL07238g YDL097C RPN6 Essential, non-ATPase regulatory subunit of the 26S proteasome lid required for the assembly and activ-ity of the 26S proteasome; the hu-man homolog (S9 protein) partially rescues Rpn6p depletion
2.25
CAGLOI04774g YBRO36C CSG2 Endoplasmic reticulum membrane protein, required for mannosylation of inositolphosphorylceramide and for growth at high calcium concen-trations
2.25
CAGLOJ11506g YNL192W CHSI Chitin synthase I, requires activa- tion from zymogenic form in or- der to catalyze the transfer of
2.25
N-acetylglucosamine (G1cNAc) to chitin; required for repairing the chitin septum during cytokinesis; transcription activated by mating factor
CAGLOG04873g YLR371W ROM2 GDP/GTP exchange protein (GEP) for Rho1p and Rho2p; mutations are synthetically lethal with muta-tions in roml, which also encodes a
2.25
GEP CAGLOM06545g YKL124W SSH4 Specificity factor required for 2.24
Rsp5p-dependent ubiquitination and sorting of cargo proteins at the multivesicular body; identified as a high-copy suppressor of a SHR3 deletion, increasing steady-state levels of amino acid permeases
CAGLOH07007g YML092C PRE8 Alpha 2 subunit of the 20S protea- some
2.23
CAGLOA04587g YBL082C ALG3 Dolichol-P-Man dependent alpha(1- 2.23 3) mannosyltransferase, involved in the synthesis of dolichol-linked oligosaccharide donor for N-linked glycosylation of proteins
CAGL0004433g YDL003W MCDI Essential subunit of the cohesin complex required for sister chro-matid cohesion in mitosis and meio-sis; apoptosis induces cleavage and translocation of a C-terminal frag- ment to mitochondria; expression peaks in S phase
2.22
CAGLOM11242g YMR226C TMA29 NADP(+)-dependent dehydro- genase; acts on serine, L-allo- threonine, and other 3-hydroxy acids; green fluorescent protein fusion protein localizes to the cytoplasm and nucleus; may in- teract with ribosomes, based on co-purification experiments
2.21
continued on next page

Appendix D. Chapter 3 Supplementary Data 219
continued from previous page CG ORF SC ORF gene name Description WT.S - WT.0
CAGLOM09108g YJR091C JSNI Member of the Puf family of 2.21 RNA-binding proteins, interacts with mRNAs encoding membrane-associated proteins; overexpression suppresses a tub2-150 mutation and causes increased sensitivity to beno-myl in wild-type cells
CAGL0005093g YOL057W - Description not mapped 2.21 CAGLOM03839g YNL305C - Description not mapped 2.2 CAGL0108239g YER021W RPN3 Essential, non-ATPase regulatory
subunit of the 26S proteasome lid, similar to the p58 subunit of the hu-man 26S proteasome; temperature-sensitive alleles cause metaphase ar-rest, suggesting a role for the pro-teasome in cell cycle control
2.2
CAGLOK12892g YFLO47W RGD2 GTPase-activating protein 2.2 (RhoGAP) for Cdc42p and Rho5p
CAGLOM04081g YNL317W PFS2 Integral subunit of the pre-mRNA cleavage and polyadenylation factor
2.2
(CPF) complex; plays an essential role in mRNA 3'-end formation by bridging different processing factors and thereby promoting the assem-bly of the processing complex
CAGLOJ04136g YOR221C MC T/ Predicted malonyl-CoA:ACF trans- ferase, putative component of a type-II mitochondrial fatty acid synthase that produces intermedi-ates for phospholipid remodeling
2.19
CAGLOK11022g YDR236C FMNI Riboflavin kinase, phosphory- lates riboflavin to form riboflavin monophosphate (FMN), which is a necessary cofactor for many enzymes; localizes to microsomes and to the mitochondrial inner membrane
2.18
CAGLOD01474g YBR108W AIMS Protein interacting with Rvs167p; 2.17 YBR108W is not an essential gene
CAGLOG08382g YDR195W REF2 RNA-binding protein involved in the cleavage step of mRNA 3'-end formation prior to polyadenylation, and in snoRNA maturation; part of holo-CPF subcomplex APT, which associates with 3'-ends of snoRNA-and mRNA-encoding genes
2.17
CAGLOF07513g YKL093W MBRJ Protein involved in mitochondrial functions and stress response; over- expression suppresses growth de- fects of hap2, hap3, and hap4 mu-tants
2.16
CAGLOL05126g YKL080W VMA5 Subunit C of the eight-subunit 2.15 V1 peripheral membrane domain of vacuolar H-k-ATPase (V-ATPase), an electrogenic proton pump found throughout the endomembrane sys- tem; required for the V1 domain to assemble onto the vacuolar mem-brane
continued on next page

Appendix D. Chapter 3 Supplementary Data 220
continued from previous page CG ORF SC ORF gene name Description WT.S - WT.0
CAGLOM06875g YNL025C SSN8 Cyclin-like component of the RNA polymerase II holoenzyme, involved in phosphorylation of the RNA polymerase II C-terminal domain; involved in glucose repression and telomere maintenance
2.15
CAGLOI07513g YOL 100W PKH2 Serine/threonine protein kinase in- volved in sphingolipid-mediated sig-naling pathway that controls en- docytosis; activates Ypklp and
2.15
Ykr2p, components of signaling cas- cade required for maintenance of cell wall integrity; redundant with Pkhlp
CAGLOH09130g YKL201C MNN4 Putative positive regulator of mannosylphosphate transferase
2.14
(Mnn6p), involved in manno- sylphosphorylation of N-linked oligosaccharides; expression in- creases in late-logarithmic and stationary growth phases
CAGLOM08206g YJL171C - Description not mapped 2.14 CAGLOI00418g YGL055W OLEI Delta(9) fatty acid desaturase, re-
quired for monounsaturated fatty acid synthesis and for normal dis-tribution of mitochondria
2.13
CAGLOD03322g YLR023C IZH3 Membrane protein involved in zinc metabolism, member of the four-protein IZH family, expression in- duced by zinc deficiency; deletion reduces sensitivity to elevated zinc and shortens lag phase, overezpres-sion reduces Zap1p activity
2.12
CAGLOJ00583g YHR028C DAPS Dipeptidyl aminopeptidase, synthe- sized as a glycosylated precursor; localizes to the vacuolar membrane; similar to Stel3p
2.12
CAGLOL10912g YOR273C TPO4 Polyamine transport protein, rec- ognizes spermine, putrescine, and spermidine; localizes to the plasma membrane; member of the major fa-cilitator superfamily
2.12
CAGLOF00583g YDR517W GRH1 Acetylated, cis-golgi localized pro- tein involved in ER to Golgi trans-port; homolog of human GRASP65; forms a complex with the coiled- coil protein Bug1p; mutants are compromised for the fusion of ER-derived vesicles with Golgi mem-branes
2.12
CAGLOD02948g YJL034W KAR2 ATPase involved in protein import into the ER, also acts as a chap-erone to mediate protein folding in the ER and may play a role in ER export of soluble proteins; regulates the unfolded protein response via interaction with Ire1p
2.12
CAGLOK05995g YLR26OW LCB5 Minor sphingoid long-chain base ki- ease, paralog of Lcb4p responsible for few percent of the total activ-ity, possibly involved in synthesis of long-chain base phosphates, which function as signaling molecules
2.12
continued on next page

Appendix D. Chapter 3 Supplementary Data 221
continued from previous page CG ORF SC ORF gene name Description WT.S - WT.0
CAGLOK01287g YGRO74W SMD I Core Sm protein Sm Dl; part of heteroheptameric complex (with
2.11
Smblp, Smd2p, Smd3p, Smeip, Smx3p, and Smx2p) that is part of the spliceosomal Ul, U2, U4, and U5 snRNPs; homolog of human Sm D1
CAGLOA03806g YOR298W MUMS Protein of unknown function in- volved in the organization of the outer spore wall layers; has similar-ity to the tafazzins superfamily of acyltransferases
2.11
CAGL00O2673g YLR392C - Description not mapped 2.11 CAGL00O2717g YAL009W SPO T Putative regulatory subunit of 2.1
Nemlp-Spo7p phosphatase holoen-zyme, regulates nuclear growth by controlling phospholipid biosyn- thesis, required for normal nuclear envelope morphology, premeiotic replication, and sporulation
CAGLOJ01001g YJL036W SNX4 Sorting nexin, involved in retrieval of late-Golgi SNAREs from post-
2.1
Golgi endosomes to the trans- Golgi network and in cytoplasm to vacuole transport; contains a PX phosphoinositide-binding do- main; forms complexes with Snx4lp and with Atg2Op
CAGLOL09889g YKL025C PA N3 Essential subunit of the Pan2p- 2.09 Pan3p poly(A)-ribonuclease com- plex, which acts to control poly(A) tail length and regulate the stoi-chiometry and activity of postrepli-cation repair complexes
CAGLOK09526g
.
YDR394W riPT3 One of six ATPases of the 19S reg- ulatory particle of the 26S protea-some involved in the degradation of ubiquitinated substrates; substrate of N-acetyltransferase B
2.09
CAGLOB03619g YEL060C P 1-03 1 Vacuolar proteinase B (yscB), a ser- ine protease of the subtilisin fam-ily; involved in protein degradation in the vacuole and required for full protein degradation during sporula-tion
2.09
CAGLOJ09328g YDL127W PCL2 G1 cyclin, associates with Pho85p cyclin-dependent kinase (Cdk) to contribute to entry into the mitotic cell cycle, essential for cell morpho-genesis; localizes to sites of polar-ized cell growth
2.09
CAGLOG01034g YGRO32W GSC2 Catalytic subunit of 1,3-beta- glucan synthase, involved in formation of the inner layer of the spore wall; activity positively regulated by Rholp and negatively by Smklp; has similarity to an alternate catalytic subunit, Fkslp
2.08
(Gsclp) continued on next page

Appendix D. Chapter 3 Supplementary Data 222
continued from previous page CG ORF SC ORF gene name Description WT.S - WT.0
CAGLOD05478g YGR186W TFG1 TFIIF (Transcription Factor II) largest subunit; involved in both transcription initiation and elonga-tion of RNA polymerase II; homol-ogous to human RAP74
2.08
CA GLOJ11704g YMR004W MVPI Protein required for sorting pro- teins to the vacuole; overproduction of Mvplp suppresses several domi-nant VPS1 mutations; Mvplp and
2.08
Vpslp act in concert to promote membrane traffic to the vacuole
CAGL0G06556g YNL097C PH023 Probable component of the Rpd3 histone deacetylase complex, in- volved in transcriptional regulation of PHO5; C-terminus has similarity to human candidate tumor suppres-sor p33(ING1) and its isoform ING3
2.07
CAGLOF04829g YPR154W PINS Protein that induces appearance of 2.05 [PIN-1-] prion when overproduced
CAGLOL11110g YML057W CMP2 Calcineurin A; one isoform (the other is CNA1) of the cat- alytic subunit of calcineurin, a
2.05
Ca4--Ficalmodulin-regulated pro- tein phosphatase which regulates Cr. 1p (a stress-response transcrip-tion factor), the other calcineurin subunit is CNB1
CAGLOE04070g YHL039W - Description not mapped 2.05 CAGL00O2541g YLR399C BDF1 Protein involved in transcription
initiation at TATA-containing pro- moters; associates with the basal transcription factor TFIID; con- tains two bromodomains; corre- sponds to the C-terminal region of mammalian TAF1; redundant with
2.04
Bdf2p CAGLOK11946g - - Description not mapped 2.04 CAGLOF01111g YOL032W OP110 Protein with a possible role in
phospholipid biosynthesis, based on inositol-excreting phenotype of the null mutant and its suppression by exogenous choline
2.04
CAGL0004917g YJR109C CPA 2 Large subunit of carbamoyl phos- phate synthetase, which catalyzes a step in the synthesis of citrulline, an arginine precursor
2.03
CAGL0005357g YDL042C SIRS Conserved NAD4- dependent his- tone deacetylase of the Sirtuin fam- ily involved in regulation of lifes- pan; plays roles in silencing at
2.02
HML, HMR, telomeres, and the rDNA locus; negatively regulates initiation of DNA replication
CAGL0C04785g YJR115W - Description not mapped 2.02 CAGLOK02013g -- Description not mapped 2.02 CAGLOE04510g YOR022C - Description not mapped 2.01 CAGL0109592g YOR303W CPAI Small subunit of carbamoyl phos-
phate synthetase, which catalyzes a step in the synthesis of citrulline, an arginine precursor; translation-ally regulated by an attenuator pep-tide encoded by YOR302W within the CPAI mRNA 5'-leader
2
continued on next page

Appendix D. Chapter 3 Supplementary Data 223
continued from previous page CC ORF SC ORF gene name Description r WT.S - WT.0

Appendix D. Chapter 3 Supplementary Data 224
Table D.4: Candida glabrata transcripts repressed in wild-type cells following transient exposure to 1M sorbitol for 15 minutes. Repressed transcripts are defined
as having log2(Cy5) - log2(Cy3) < -2.0 and Benjamini-Hochberg corrected p-values < 0.05. Estimated log-ratios correspond to the WT.S - WT.0 effect described in Table D.2
CG ORF SC ORF gene name Description WT.S - WT.0
CAGLOH01089g YDR287W INM2 Inositol monophosphatase, involved in biosynthesis of inositol; enzy- matic activity requires magnesium ions and is inhibited by lithium and sodium ions; inml inm2 double mu-tant lacks inositol auxotrophy
-3.41
CAGL0L10384g YOR064C YNGI Subunit of the NuA3 histone acetyl- transferase complex that acetylates histone H3; contains PHD finger domain that interacts with methy-lated histone H3, has similarity to the human tumor suppressor ING1
-3.38
CAC LOH09064g YHR128W FUR/ Uracil phosphoribosyltransferase, synthesizes UMP from uracil; involved in the pyrimidine salvage pathway
-2.97
CAGLOL05500g YJL 122W ALB1 Shuttling pre-60S factor; involved in the biogenesis of ribosomal large subunit; interacts directly with
-2.91
Arxlp; responsible for Tif6p recy-cling defects in absence of Reilp
CAGL0000737g YLR222C UTP13 Nucleolar protein, component of the small subunit (SSU) processome containing the U3 snoRNA that is involved in processing of pre-18S rRNA
-2.91
CAGLOK09416g YGL211W NCS6 Protein required for thiolation of the uridine at the wobble position of Gln, Lys, and Glu tRNAs; has a role in urmylation and in invasive and pseudohyphal growth; inhibits replication of Brome mosaic virus in
-2.81
S. cerevisiae CAGL0000352g YGR277C - Description not mapped -2.81 CAGL0106743g YER145C FTR1 High affinity iron permease involved
in the transport of iron across the plasma membrane; forms complex with Fet3p; expression is regulated by iron
-2.74
CAGL0106809g YER142C MAGI 3-methyl-adenine DNA glycosylase involved in protecting DNA against alkylating agents; initiates base ex-cision repair by removing damaged bases to create abasic sites that are subsequently repaired
-2.71
CAGLOG04411g YJL109C UTPIO Nucleolar protein, component of the small subunit (SSU) processome containing the U3 snoRNA that is involved in processing of pre-18S rRNA
-2.7
CAGLOK03795g YMR128W ECM16 Essential DEAR-box ATP- dependent RNA helicase specific to the U3 snoRNP, predominantly nucleolar in distribution, required for 18S rRNA synthesis
-2.63
continued on next page

Appendix D. Chapter 3 Supplementary Data 225
continued from previous page CG ORF SC ORF gene name Description WT.S - WT.0
CAGLOG00484g CAGLOA01914g
YGR272C YHR099W
- TRA I
Description not mapped Subunit of SAGA and NuA4 histone acetyltransferase complexes; inter- acts with acidic activators (e.g., Gal4p) which leads to transcription activation; similar to human TR-
-2.59 -2.58
RAP, which is a cofactor for c-Myc mediated oncogenic transformation
CAGLOG07953g YNR037C RSMI;) Mitochondrial ribosomal protein of the small subunit, has similarity to
-2.53
E. coli S19 ribosomal protein CAGLOI09900g YOR319W HSH49 U2-snRNP associated splicing fac-
tor with similarity to the mam-malian splicing factor SAP49; pro-posed to function as a U2-snRNP assembly factor along with Hsh155p and binding partner Cuslp; con- tains two RNA recognition motifs
-2.51
(RRM) CAGLOD05940g YGR175C ERG1 Squalene epoxidase, catalyzes the
epoxidation of squalene to 2,3- oxidosqualene; plays an essential role in the ergosterol-biosynthesis pathway and is the specific target of the antifungal drug terbinafine
-2.48
CAGLOKO6281g YDR454C GUK1 Guanylate kinase, converts GMP to -2.48 GDP; required for growth and man-nose outer chain elongation of cell wall N-linked glycoproteins
CAGL0104026g YDL053C PBP4 Pbplp binding protein, interacts strongly with Pablp-binding pro-tein 1 (Pbplp) in the yeast two-hybrid system; also interacts with
-2.47
Lsml2p in a copurification assay CAGLOF01793g YLR056W ERGS C-5 sterol desaturase, catalyzes the
introduction of a C-5(6) double bond into episterol, a precursor in ergosterol biosynthesis; mutants are viable, but cannot grow on non- fermentable carbon sources
-2.43
CAGL0101826g YHR148W IMP3 Component of the 55LT processome, which is required for pre-18S rRNA processing, essential protein that interacts with MpplOp and me- diates interactions of Imp4p and
-2.42
MpplOp with U3 snoRNA CAGLOL03025g YKR025W RPC37 RNA polymerase III subunit C37 -2.4 CAGLOH04301g YML078W CPR3 Mitochondrial peptidyl-prolyl cis-
trans isomerase (cyclophilin), cat-alyzes the cis-trans isomerization of peptide bonds N-terminal to proline residues; involved in protein refold-ing after import into mitochondria
-2.4
CAGLOK02233g YER126C NSA2 Protein constituent of 66S pre- ribosomal particles, contributes to processing of the 27S pre-rRNA
-2.38
CAGLOM08448g YKL165C MCD4 Protein involved in glycosylphos- phatidylinositol (GPI) anchor syn- thesis; multimembrane-spanning protein that localizes to the endo-plasmic reticulum; highly conserved among eukaryotes
-2.32
continued on next page

Appendix D. Chapter 3 Supplementary Data 226
continued from previous page CG ORF SC ORF gene name Description WT.S - WT.0
CAGLOH05533g YPL082C MOT] Essential abundant protein involved in regulation of transcription, re- moves Sptl5p (TBP) from DNA via its C-terminal ATPase activity, forms a complex with TBP that binds TATA DNA with high affin-ity but with altered specificity
-2.31
CAGL00O2211g YELO4OW UTR2 Cell wall protein that functions in the transfer of chitin to beta(1-
-2.27
6)glucan; putative chitin transgly-cosidase; glycosylphosphatidylinos-itol (GPI)-anchored protein local-ized to the bud neck; has a role in cell wall maintenance
CAGLOE05324g YOR330C MIP1 Catalytic subunit of the mitochon- drial DNA polymerase; conserved
-2.24
C-terminal segment is required for the maintenance of mitochondria] genome.
CAGLOD00748g YDL067C COX9 Subunit VIIa of cytochrome c oxi- dose, which is the terminal member of the mitochondrial inner mem-brane electron transport chain
-2.18
CAGLOB00638g YCL047C - Description not mapped -2.16 CAGLOM09086g YJR0924V BUD4 Protein involved in bud-site selec-
tion and required for axial budding pattern; localizes with septins to bud neck in mitosis and may con-stitute an axial landmark for next round of budding; potential Cdc28p substrate
-2.16
CAGLOK04477g YGRO6OW ERG25 C-4 methyl sterol oxidase, cat- alyzes the first of three steps re-quired to remove two C-4 methyl groups from an intermediate in er- gosterol biosynthesis; mutants ac- cumulate the sterol intermediate
-2.15
4,4-dimethylzymosterol CAGLOG08646g YIL122W POG1 Putative transcriptional activa-
tor that promotes recovery from pheromone induced arrest; inhibits both alpha-factor induced G1 arrest and repression of CLN1 and CLN2 via SCB/MCB promoter elements; potential Cdc28p substrate; SBF regulated
-2.12
CAGLOL13156g YLR073C - Description not mapped -2.1 CAGLOJ02442g YIL022W TL)144 Peripheral mitochondrial mem-
brane protein involved in mito- chondria] protein import, tethers essential chaperone Ssclp to the translocon channel at the matrix side of the inner membrane
-2.09
CAGLOL01991g YKL033W TTI1 Putative protein of unknown func- tion; subunit of the ASTRA com- plex which is part of the chro- matin remodeling machinery; sim- ilar to S. pombe Ttilp; detected in highly purified mitochondria in high-throughput studies
-2.09
continued on next page

Appendix D. Chapter 3 Supplementary Data 227
continued from previous page CG ORF SC ORF gene name Description WT.S - WT.0
CAGL00O2739g YAL008W FUN.14 Mitochondria) protein of unknown function
-2.05
CAGLOL01551g YML052W SUR7 Putative integral membrane pro- tein; component of eisosomes; asso- ciated with endocytosis, along with
-2.04
PiIlp and Lsplp; sporulation and plasma membrane sphingolipid con-tent are altered in mutants
CAGLOL08470g - - Description not mapped -2.03

Appendix D. Chapter 3 Supplementary Data 228
Table D.5: Profile of transcripts involved in metabolism in wild-type, hog)
and stel 1 cells. Key to abbreviations: CG= Candida glabrata open reading frame ID;
SC=Saccharomyces cerevisiae open reading frame ID; WT=wild-type C. glabrata cells;
hog 1 =hog 1 C. glabrata strain; stel 1 =stel 1 C. glabrata strain. The digits in each column
indicate whether the transcript is induced (1), repressed (-1) or not differentially expressed (0) after 15 minutes exposure to 1M sorbitol. All descriptions are inferred by sequence
homology between C. glabrata and S. cerevisiae open reading frames.
CG ORF SC ORF Gene name Description WT hog) stet 1
CAGLOM07381g YPL053C KTR6 Probable mannosylphosphate transferase involved in the syn- thesis of core oligosaccharides in protein glycosylation path-way; member of the KRE2/MNT1 mannosyltransferase family
1 1 0
CAGL0104554g YBRO14C GRX7 Cis-golgi localized monothiol glutaredoxin; more similar in ac- tivity to dithiol than other monothiol glutaredoxins; involved in the oxidative stress response; does not bind metal ions; func-tional overlap with GRX6
1 0 0
CAGLOG01166g YLR348C DIC1 Mitochondrial dicarboxylate carrier, integral membrane pro- tein, catalyzes a dicarboxylate-phosphate exchange across the inner mitochondrial membrane, transports cytoplasmic dicar-boxylates into the mitochondrial matrix
1 0 0
CAGLOK11022g YDR236C FMN1 Riboflavin kinase, phosphorylates riboflavin to form riboflavin monophosphate (FMN), which is a necessary cofactor for many enzymes; localizes to microsomes and to the mitochondrial. in-ner membrane
1 0 1
CAGL0004917g YJR1O9C CPA 2 Large subunit of carbamoyl phosphate synthetase, which cat- alyzes a step in the synthesis of citrulline, an arginine precursor
1 1 1
CAGLOH05137g YPL061W A LD6 Cytosolic aldehyde dehydrogenase, activated by Mg2+ and uti- lizes NADP+ as the preferred coenzyme; required for conver-sion of acetaldehyde to acetate; constitutively expressed; lo-cates to the mitochondrial outer surface upon oxidative stress
1 0 0
CAGL00O2233g YER042W If XR 1 Peptide methionine sulfoxide reductase, reverses the oxidation of methionine residues; involved in oxidative damage repair, providing resistance to oxidative stress and regulation of lifes-pan
1 0 0
CAGLOI09592g YOR303W CPA I Small subunit of carbamoyl phosphate synthetase, which cat- alyzes a step in the synthesis of citrulline, an arginine pre-cursor; translationally regulated by an attenuator peptide en-coded by YOR302W within the CPA1 mRNA 5'-leader
1 0 0
CAGLOG04763g YOR107W RGS2 Negative regulator of glucose-induced cAMP signaling; directly activates the GTPase activity of the heterotrimeric G protein alpha subunit Gpa2p
1 1 1
CAGL0005137g YOL059W GPD2 NAD-dependent glycerol 3-phosphate dehydrogenase, homolog of Gpdlp, expression is controlled by an oxygen-independent signaling pathway required to regulate metabolism under anoxic conditions; located in cytosol and mitochondria
1 0 0
CAGLOF08107g YGR244C LSC2 Beta subunit of succinyl-CoA ligase, which is a mitochon- drial enzyme of the TCA cycle that catalyzes the nucleotide-dependent conversion of succinyl-CoA to succinate
1 0 0
CAGLOM12034g YALO3SW CDCI9 Pyruvate kinase, functions as a homotetramer in glycolysis to convert phosphoenolpyruvate to pyruvate, the input for aero-bic (TCA cycle) or anaerobic (glucose fermentation) respira-tion
1 1 0
CAGLOF06941g YGL062W PYCI Pyruvate carboxylase isoform, cytoplasmic enzyme that con- verts pyruvate to oxaloacetate; highly similar to isoform Pyc2p but differentially regulated; mutations in the human homolog are associated with lactic acidosis
1 0 0
CAGLOG01540g YNL036W NCE103 Carbonic anhydrase; poorly transcribed under aerobic condi- tions and at an undetectable level under anaerobic conditions; involved in non-classical protein export pathway
1 1 1
continued on next page

Appendix D. Chapter 3 Supplementary Data 229
continued from previous page CC ORF SC ORF Gene Name Description WT hogI ste11
CAGLOM14047g YMR318C ADH6 NADPH-dependent medium chain alcohol dehydrogenase with broad substrate specificity; member of the cinnamyl family of alcohol dehydrogenases; may be involved in fusel alcohol syn-thesis or in aldehyde tolerance
1 0 0
CAGLOH06523g YIL145C PANG Pantothenate synthase, also known as pantoate-beta-alanine ligase, required for pantothenic acid biosynthesis, deletion causes pantothenic acid auxotrophy, homologous to E. coli panC
1 1 1
CAGLOK03509g YMR110C HFDI Putative fatty aldehyde dehydrogenase, located in the mito- chondrial outer membrane and also in lipid particles; has simi-larity to human fatty aldehyde dehydrogenase (FALDH) which is implicated in Sjogren-Larsson syndrome
1 1
CAGL0F01111g YOL032W OPII0 Protein with a possible role in phospholipid biosynthesis, based on inositol-excreting phenotype of the null mutant and its sup-pression by exogenous choline
1 0 0
CAGLOG05313g YDR072C IPT1 Inositolphosphotransferase, involved in synthesis of mannose- 1 1 1 (inositol-P)2-ceramide (M(IP)2C), the most abundant sph- ingolipid;, can mutate to resistance to the antifungals sy- ringomycin E and DmAMP1 and to K. lactis zymocin
CAGLOI01100g YOR120W GCY1 Putative NADP(+) coupled glycerol dehydrogenase, pro- posed to be involved in an alternative pathway for glycerol catabolism; member of the aldo-keto reductase (AKR) family
1 0 0

Appendix D. Chapter 3 Supplementary Data 230
Table D.6: Profile of transcripts involved in protein degradation in wild-type, hogl and sten cells. Key to abbreviations: CG=Candida glabrata open reading frame
ID; SC=Saccharomyces cerevisiae open reading frame ID; WT=wild-type C. glabrata cells;
hogl=hogl C. glabrata strain; stell=stel 1 C. glabrata strain. The digits in each column
indicate whether the transcript is induced (1), repressed (-1) or not differentially expressed
(0) after 15 minutes exposure to 1M sorbitol. All descriptions are inferred by sequence
homology between C. glabrata and S. cerevisiae open reading frames.
CG ORF SC ORF Gene name Description WT hog 1 stell
CAGLOD05082g YLL039C UBLI Ubiquitin, becomes conjugated to proteins, marking them for selective degradation via the ubiquitin-26S proteasome system; essential for the cellular stress response; encoded as a polyu-biquitin precursor comprised of 5 head-to-tail repeats
1 0 0
CAGLOL09152g YFROO4W RPN11 Metalloprotease subunit of the 19S regulatory particle of the 1 0 0 265 proteasome lid; couples the deubiquitination and degrada-tion of proteasome substrates
CAGLOK09526g YDR394W RPT3 One of six ATPases of the 19S regulatory particle of the 26S proteasome involved in the degradation of ubiquitinated sub-strates; substrate of N-acetyltransferase B
1 0 1
CAGLOL07238g YDL097C RPN6 Essential, non-ATPase regulatory subunit of the 265 protea- some lid required for the assembly and activity of the 26S proteasome; the human homolog (S9 protein) partially rescues
1 1 1
Rpn6p depletion CAGLOH07007g YML092C PRES Alpha 2 subunit of the 20S proteasome 1 0 1 CAGLOM06545g YKL124W SSH4 Specificity factor required for Rsp5p-dependent ubiquitination
and sorting of cargo proteins at the multivesicular body; iden-tified as a high-copy suppressor of a SHR3 deletion, increasing steady-state levels of amino acid permeases
1 1 1
CAGLOE02299g YOL013C HRD1 Ubiquitin-protein ligase required for endoplasmic reticulum- associated degradation (ERAD) of misfolded proteins; geneti-cally linked to the unfolded protein response (UPR); regulated through association with Hrd3p; contains an 112 ring finger
1 0 1
CAGL0108239g YER021W RPN3 Essential, non-ATPase regulatory subunit of the 26S protea- some lid, similar to the p58 subunit of the human 26S pro-teasome; temperature-sensitive alleles cause metaphase arrest, suggesting a role for the proteasome in cell cycle control
1 1 0
CAGLOJ09108g YDL19OC UFD2 Ubiquitin chain assembly factor (E4) that cooperates with a ubiquitin-activating enzyme (El), a ubiquitin-conjugating en-zyme (E2), and a ubiquitin protein ligase (E3) to conjugate ubiquitin to substrates; also functions as an E3
1 1 0
CAGLOL05434g YKR042W UTH1 Mitochondria] outer membrane and cell wall localized SUN family member required for mitochondria] autophagy; involved in the oxidative stress response, life span during starvation, mitochondrial biogenesis, and cell death
1 1 1
CAGLOA02046g YOR018W ROD] Membrane protein that binds the ubiquitin ligase Rsp5p via its 2 PY motifs; overexpression confers resistance to the GST substrate o-dinitrobenzene,zinc, and calcium; proposed to reg-ulate the endocytosis of plasma membrane proteins
1 1 1

Appendix D. Chapter 3 Supplementary Data 231
Table D.7: Profile of transcripts involved in protein trafficking in wild-type, hogl
and stel 1 cells. Key to abbreviations: CG=Candida glabrata open reading frame ID;
SC=Saccharomyces cerevisiae open reading frame ID; WT=wild-type C. glabrata cells;
hogl =hogl C. glabrata strain; stel 1 =stel 1 C. glabrata strain. The digits in each column
indicate whether the transcript is induced (1), repressed (-1) or not differentially expressed
(0) after 15 minutes exposure to 1M sorbitol. All descriptions are inferred by sequence
homology between C. glabrata and S. cerevisiae open reading frames.
CG ORF SC ORF Gene name Description WT hog1 stel 1
CAGLOL11572g YHR103W SBE22 Protein involved in the transport of cell wall components from the Golgi to the cell surface; similar in structure and function-ally redundant with Sbe2p; involved in bud growth
1 1 1
CAGLOL05126g YKL080W VMA 5 Subunit C of the eight-subunit VI peripheral membrane do- main of vacuolar HA--ATPase (V-ATPase), an electrogenic pro-ton pump found throughout the endomembrane system; re-quired for the VI domain to assemble onto the vacuolar mem-brane
1 0 0
CAGLOI10010g YGR142W BTN2 v-SNARE binding protein that facilitates specific protein re- trieval from a late endosome to the Golgi; modulates arginine uptake, possible role in mediating pH homeostasis between the vacuole and plasma membrane H(-1-)-ATPase
1 1 1
CAGLOH03729g YNLOO6W LST8 Protein required for the transport of amino acid permease 1 0 0 Gap1p from the Golgi to the cell surface; component of the TOR signaling pathway; associates with both Tor1p and Tor2p; contains a WD-repeat
CAGLOJ00583g YHR028C DA P2 Dipeptidyl aminopeptidase, synthesized as a glycosylated pre- cursor; localizes to the vacuolar membrane; similar to Stel3p
1 0 0
CAGLOM01782g YBRO80C SEC18 ATPase required for the release of Sec17p during the 'priming' step in homotypic vacuole fusion and for ER to Golgi trans-port; homolog of the mammalian NSF
1 0 0
CAGL0B03619g YEL060C PRB1 Vacuolar proteinase B (yscB), a serine protease of the subtil- isin family; involved in protein degradation in the vacuole and required for full protein degradation during sporulation
1 0 1
CAGLOB01991g YDR126W S WEI Palmitoyltransferase that acts on the SNAREs Snclp, Syn8p, Tlglp and likely on all SNAREs; member of a family of pu-tative palmitoyltransferases containing an Asp-His-His-Cys-cysteine rich (DHIJC-CRD) domain; may have a role in vacuole fusion
1 0 0
CAGLOL06028g YGL181W GTS1 Protein that localizes to the nucleus and is involved in tran- scription regulation; also localizes to actin patches and plays a role in endocytosis; N-terminus contains an ARF-GAP domain and C-terminus contains a Gln-rich domain
1 0 0
CAGL000320Ig YLL040C VPS13 Protein of unknown function; heterooligomeric or ho- mooligomeric complex; peripherally associated with mem- branes; homologous to human COH1; involved in sporulation, vacuolar protein sorting and protein-Golgi retention
1 1 1
CAGLOK00539g YPL259C APMI Mul-like medium subunit of the clathrin-associated protein complex (AP-1); binds clathrin; involved in clathrin-dependent
1 1 0
Golgi protein sorting CAGLOJ01001g YJL036W SNX4 Sorting nexin, involved in retrieval of late-Golgi SNAREs from
post-Golgi endosomes to the trans-Golgi network and in cyto-plasm to vacuole transport; contains a PX phosphoinositide- binding domain; forms complexes with Snx41p and with
1 1 0
Atg2Op CAGLOK02761g YHLOO2W HSEI Subunit of the endosomal Vps27p-Hselp complex required for
sorting of ubiquitinated membrane proteins into intralumenal vesicles prior to vacuolar degradation, as well as for recycling of Golgi proteins and formation of lumenal membranes
1 0 1
continued on next page

Appendix D. Chapter 3 Supplementary Data 232
continued from previous page CG ORF SC ORF Gene Name Description WT hogl step
CAGLOJ11704g YMR004W MVP1 Protein required for sorting proteins to the vacuole; overpro- duction of Mvplp suppresses several dominant VPS1 muta-tions; Mvplp and Vpslp act in concert to promote membrane traffic to the vacuole
1 0 0

Appendix D. Chapter 3 Supplementary Data 233
Table D.8: Profile of transcripts involved in the transcriptional regulation in wild-type, hogl and stell cells. Key to abbreviations: CG=Candida glabrata open reading
frame ID; SC=Saccharomyces ccrevisiae open reading frame ID; WT=wild-type C. glabrata
cells; hagl=hogl C. glabrata strain; stell=stell C. glabrata strain. The digits in each
column indicate whether the transcript is induced (1), repressed (-1) or not differentially
expressed (0) after 15 minutes exposure to 1M sorbitol. All descriptions are inferred by
sequence homology between C. glabrata and S. cerevisiae open reading frames.
CG ORF SC ORF Gene name Description WT hog! ate!!
CAGLOI04246g YGL162W S UT/ Transcription factor of the Zn[III2Cys6 family involved in sterol uptake; involved in induction of hypoxic gene expres-sion
1 1 0
CAGLOM06875g YNL025C SSN8 Cyclin-like component of the RNA polymerase II holoenzyme, involved in phosphorylation of the RNA polymerase II C-terminal domain; involved in glucose repression and telomere maintenance
1 0 1
CAGL0005357g YDL042C SIR2 Conserved NAD+ dependent histone deacetylase of the Sirtuin family involved in regulation of lifespan; plays roles in silencing at HML, HMR, telomeres, and the rDNA locus; negatively regulates initiation of DNA replication
1 0 0
CAGLOD05368g YBR253W SR136 Subunit of the RNA polymerase II mediator complex; asso- ciates with core polymerase subunits to form the RNA poly-merase II holoenzyme; essential for transcriptional regulation
11 0
CAGL0CO2541g YLR399C DDF1 Protein involved in transcription initiation at TATA- containing promoters; associates with the basal transcription factor TFIID; contains two bromodomains; corresponds to the
1 1 0
C-terminal region of mammalian TAF1; redundant with Bdf2p CAGLOD05434g YFRO65W ROX I Heme-dependent repressor of hypoxic genes; contains an HMG
domain that is responsible for DNA bending activity 1 1 0
CAGLOH07557g YGL254W FZF1 Transcription factor involved in sulfite metabolism, sole iden- tified regulatory target is SSU1, overexpression suppresses sulfite-sensitivity of many unrelated mutants due to hyperac-tivation of SSU1, contains five zinc fingers
1 1 0
CAGLOL06028g YGL181W GTSI Protein that localizes to the nucleus and is involved in tran- scription regulation; also localizes to actin patches and plays a role in endocytosis; N-terminus contains an ARF-GAP domain and C-terminus contains a Gln-rich domain
1 0 0
CAGLOD05478g YGR186W TFG1 TFIIF (Transcription Factor II) largest subunit; involved in both transcription initiation and elongation of RNA poly-merase II; homologous to human RAP74
1 1 0
CAGL0G06556g YNL097C PH023 Probable component of the Rpd3 histone deacetylase complex, involved in transcriptional regulation of PHO5; C-terminus has similarity to human candidate tumor suppressor p33(ING1) and its isoform ING3
1 0 0
CAGLOH08173g YOR028C CIN5 Basic leucine zipper (bZIP) transcription factor of the yAP-1 family, mediates pleiotropic drug resistance and salt tolerance; nuclearly localized under oxidative stress and sequestered in the cytoplasm by Lot6p under reducing conditions
1 1 1
CAGLOF09119g YHR207C SETS Zinc-finger protein of unknown function, contains one canoni- cal and two unusual fingers in unusual arrangements; deletion enhances replication of positive-strand RNA virus
1 0 0
CAGLOH04631g YML007W YAPI Basic leucine zipper (bZIP) transcription factor required for oxidative stress tolerance; activated by H202 through the mul-tistep formation of disulfide bonds and transit from the cyto-plasm to the nucleus; mediates resistance to cadmium
1 1 0

Appendix D. Chapter 3 Supplementary Data 234
Table D.9: Profile of induced transcripts conserved between S. cerevisiae (Gasch
et al., and C. glabrata after exposure to hyperosmotic stress. Key to abbreviations:
CG=Candida glabrata open reading frame ID; SC=Saccharornyces cerevisiae open reading
frame ID. All descriptions are inferred by sequence homology between C. glabrata and S.
cerevisiae open reading frames. S. cerevisiae data were obtained from a study by Gasch et
a/.(230) in which S. cerevisiae cells were treated with 1M sorbitol.
CG ORF SC ORF Gene name Description WT
CAGLOI10010g YGR142W BTN2 v-SNARE binding protein that facilitates specific protein re- trieval from a late endosome to the Golgi; modulates arginine uptake, possible role in mediating pH homeostasis between the vacuole and plasma membrane H(+)-ATPase
5.55
CAGLOG04433g YJL108C PRM10 Pheromone-regulated protein, predicted to have 5 transmem- brane segments; induced by treatment with 8-methoxypsoralen and UVA irradiation
5.14
CAGLOE00803g YDR171W HSP42 Small heat shock protein (sHSP) with chaperone activity; forms barrel-shaped oligomers that suppress unfolded protein aggregation; involved in cytoskeleton reorganization after heat shock
3.82
CAGLOG03883g YLL026W HSP/0.4 Heat shock protein that cooperates with Ydjlp (Hsp40) and 3.61 Ssalp (Hsp70) to refold and reactivate previously denatured, aggregated proteins; responsive to stresses including: heat, ethanol, and sodium arsenite; involved in [PSI-I-] propagation
CAGLOH08173g YOR028C CIN5 Basic leucine zipper (bZIP) transcription factor of the yAP-1 family, mediates pleiotropic drug resistance and salt tolerance; nuclearly localized under oxidative stress and sequestered in the cytoplasm by Lot6p under reducing conditions
3.49
CAGLOG01540g YNL036W NCE103 Carbonic anhydrase; poorly transcribed under aerobic condi- tions and at an undetectable level under anaerobic conditions; involved in non-classical protein export pathway
3.47
CAGLOH08844g YMR173W DDR48 DNA damage-responsive protein, expression is increased in re- sponse to heat-shock stress or treatments that produce DNA lesions; contains multiple repeats of the amino acid sequence
3.06
NNNDSYGS CAGLOH08844g YMR173W DDR48 DNA damage-responsive protein, expression is increased in re-
sponse to heat-shock stress or treatments that produce DNA lesions; contains multiple repeats of the amino acid sequence
3.06
NNNDSYGS CAGLOH08844g YlvIR173W DDR48 DNA damage-responsive protein, expression is increased in re-
sponse to heat-shock stress or treatments that produce DNA lesions; contains multiple repeats of the amino acid sequence
3.06
NNNDSYGS CAGLOH08844g YMR173W DDR48 DNA damage-responsive protein, expression is increased in re-
sponse to heat-shock stress or treatments that produce DNA lesions; contains multiple repeats of the amino acid sequence
3.06
NNNDSYGS CAGLOI01100g YOR120W GCY1 Putative NADP(+) coupled glycerol dehydrogenase, pro-
posed to be involved in an alternative pathway for glycerol catabolism; member of the aldo-keto reductase (AKR) family
2.56
CAGLOH05137g YPL061W ALD6 Cytosolic aldehyde dehydrogenase, activated by Mg2+ and uti- lizes NADP+ as the preferred coenzyme; required for conver-sion of acetaldehyde to acetate; constitutively expressed; lo-cates to the mitochondrial outer surface upon oxidative stress
2.53
CAGLOM06347g YBR183W YPC1 Alkaline ceramidase that also has reverse (CoA-independent) ceramide synthase activity, catalyzes both breakdown and syn-thesis of phytoceramide; overexpression confers fumonisin B1 resistance
2.52
CAGLOJ04158g YOR220W RCN2 Protein of unknown function; green fluorescent protein (GFP)- fusion protein localizes to the cytoplasm and is induced in re-sponse to the DNA-damaging agent MMS; phosphorylated in response to alpha factor
2.35
continued on next page

Appendix D. Chapter 3 Supplementary Data 235
continued from previous page CC ORF SC ORF Gene Name Description WT.S - WT.0
CAGLOF07513g YKL093W MBR1 Protein involved in mitochondrial functions and stress re- sponse; overexpression suppresses growth defects of hap2, hap3, and hap4 mutants
2.16

Appendix D. Chapter 3 Supplementary Data 236
Table D.10: Profile of induced transcripts conserved between S. cerevisiae
(O'Rourke & Herskowitz 1M sorbitol) and C. glabrata after exposure to hy-
perosmotic stress. Key to abbreviations: CG=Candida glabrata open reading frame ID;
SC=Saccharomyces cerevisiae open reading frame ID. All descriptions are inferred by se-
quence homology between C. glabrata and S. cerevisiae open reading frames. S. cerevisiae
data were obtained from a study by O'Rourke & Herskowitz (518) in which S. cerevisiae
cells were treated with 1M sorbitol
CG ORF SC ORF Gene name Description WT
CAGLOI10010g YGR142W BTN2 v-SNARE binding protein that facilitates specific protein re- trieval from a late endosome to the Golgi; modulates arginine uptake, possible role in mediating pH homeostasis between the vacuole and plasma membrane H(+)-ATPase
5.55
CAGLOG04433g YJLI08C PRM1O Pheromone-regulated protein, predicted to have 5 transmem- brane segments; induced by treatment with 8-methoxypsoralen and UVA irradiation
5.14
CAGLOE00803g YDR171W HSP42 Small heat shock protein (sHSP) with chaperone activity; forms barrel-shaped oligomers that suppress unfolded protein aggregation; involved in cytoskeleton reorganization after heat shock
3.82
CAGLOG03883g YLL026W HSPI 04 Heat shock protein that cooperates with Ydjlp (Hsp40) and 3.61 Ssalp (Hsp70) to refold and reactivate previously denatured, aggregated proteins; responsive to stresses including: heat, ethanol, and sodium arsenite; involved in [PSI-] propagation
CAGLOF08745g YLR327C TMA I 0 Protein of unknown function that associates with ribosomes 3.53 CAGLOG01540g YNL036W NCE103 Carbonic anhydrase; poorly transcribed under aerobic condi-
tions and at an undetectable level under anaerobic conditions; involved in non-classical protein export pathway
3.47
CAGLOH08844g YMR173W DDR48 DNA damage-responsive protein, expression is increased in re- sponse to heat-shock stress or treatments that produce DNA lesions; contains multiple repeats of the amino acid sequence
3.06
NNNDSYGS CAGLOE05632g YOR348C PUT4 Proline permease, required for high-affinity transport of pro-
line; also transports the toxic proline analog azetidine- 2.85
2-carboxylate (AzC); PUT4 transcription is repressed in ammonia-grown cells
CAGLOJ04158g YOR220W RCN2 Protein of unknown function; green fluorescent protein (GFP)- fusion protein localizes to the cytoplasm and is induced in re-sponse to the DNA-damaging agent MMS; phosphorylated in response to alpha factor
2.35
CAGLOF01111g YOL032W OPIIO Protein with a possible role in phospholipid biosynthesis, based on inositol-excreting phenotype of the null mutant and its sup-pression by exogenous choline
2.04

Appendix D. Chapter 3 Supplementary Data 237
Table D.11: Profile of induced transcripts conserved between S. cerevisiae (Rep
et al.,) and C. glabrata after exposure to hyperosmotic stress. Key to abbrevia-
tions: CG=Candida glabrata open reading frame ID; SC=Saccharomyces cerevisiae open
reading frame ID. All descriptions are inferred by sequence homology between C. glabrata
and S. cerevisiae open reading frames. S. cerevisiae data were obtained from a study by
Rep et al. (581) in which S. cerevisiae cells were treated with 0.71\4 NaCl.
CG 0 RF SC ORE Gene name Description WT.S - WT.0 CAGL00O2321g YER037W PHM8 Protein of unknown function, expression is induced by low
phosphate levels and by inactivation of Pho85p 5.41
CAGLOG04433g YJL108C PR 114.10 Pheromone-regulated protein, predicted to have 5 transmem- brane segments; induced by treatment with 8-methoxypsoralen and UVA irradiation
5.14
CAGLOE00803g YDR171W HSP42 Small heat shock protein (sHSP) with chaperone activity; forms barrel-shaped oligomers that suppress unfolded protein aggregation; involved in cytoskeleton reorganization after heat shock
3.82
CAGLOG03883g YLL026W HSP/04 Heat shock protein that cooperates with Ydjlp (Hsp40) and 3.61 Ssalp (Hsp70) to refold and reactivate previously denatured, aggregated proteins; responsive to stresses including: heat, ethanol, and sodium arsenite; involved in [PSI-I-1 propagation
CAGLOF08745g YLR327C TMA 10 Protein of unknown function that associates with ribosomes 3.53 CAGLOH08173g YOR028C CINS Basic leucine zipper (bZIP) transcription factor of the yAP-1
family, mediates pleiotropic drug resistance and salt tolerance; nuclearly localized under oxidative stress and sequestered in the cytoplasm by Lot6p under reducing conditions
3.49
CAGLOG01540g YNL036W NCE 103 Carbonic anhydrase; poorly transcribed under aerobic condi- tions and at an undetectable level under anaerobic conditions; involved in non-classical protein export pathway
3.47
CAGLOM12474g YIL055C - - 3.46 CAGLOH08844g YMR173W DDR48 DNA damage-responsive protein, expression is increased in re-
sponse to heat-shock stress or treatments that produce DNA lesions; contains multiple repeats of the amino acid sequence
3.06
NNNDSYGS CAGLOE05632g YOR348C PUT4 Proline permease, required for high-affinity transport of pro-
line; also transports the toxic proline analog azetidine- 2.85
2-carboxylate (AzC); PUT4 transcription is repressed in ammonia-grown cells
CAGLOL00803g YER054C GIP2 Putative regulatory subunit of the protein phosphatase Glc7p, involved in glycogen metabolism; contains a conserved motif
2.62
(GVNK motif) that is also found in Gaclp, Pig1p, and Pig2p CAGLOK12034g YDR040C ENA 1 P-type ATPase sodium pump, involved in Na+ and Li+ efflux
to allow salt tolerance 2.61
CAGLOH05137g YPL061W ALD6 Cytosolic aldehyde dehydrogenase, activated by Mg2+ and uti- lizes NADP+ as the preferred coenzyme; required for conver-sion of acetaldehyde to acetate; constitutively expressed; lo-cates to the mitochondrial outer surface upon oxidative stress
2.53
CAGLOJ04466g YLR414C - - 2.5 CAGLOE06424g YKL150W MCR/ Mitochondrial NADH-cytochrome b5 reductase, involved in er-
gosterol biosynthesis 2.42
CAGLOJ04158g YOR220W RCN2 Protein of unknown function; green fluorescent protein (GFP)- fusion protein localizes to the cytoplasm and is induced in re-sponse to the DNA-damaging agent MMS; phosphorylated in response to alpha factor
2.35
CAGLOI04554g YBRO14C GRX7 Cis-golgi localized monothiol glutaredoxin; more similar in ac- tivity to dithiol than other monothiol glutaredoxins; involved in the oxidative stress response; does not bind metal ions; func-tional overlap with GRX6
2.34
continued on next page

Appendix D. Chapter 3 Supplementary Data 238
continued from previous page CG ORF SC ORF Gene Name Description WT.S - WT.0
CAGLOB03619g YEL060C PRB1 Vacuolar proteinase B (yscB), a serine protease of the subtil- isin family; involved in protein degradation in the vacuole and required for full protein degradation during sporulation
2.09

Appendix D. Chapter 3 Supplementary Data 239
Table D.12: Profile of induced transcripts conserved between S. cerevisiae
(O'Rourke & Herskowitz 0.5M KC1) and C. glabrata after exposure to hy-
perosmotic stress. Key to abbreviations: CG.Candida glabrata open reading frame ID;
SC=Saccharomyces cerevisiae open reading frame ID. All descriptions are inferred by se-
quence homology between C. glabrata and S. cerevisiae open reading frames. S. cerevisiae
data were obtained from a study by O'Rourke & Herskowitz (518) in which S. cerevisiae
cells were treated with 0.5M KC1.
CG ORF SC ORF Gene name Description WT CAGLOI10010g YGR142W HTN2 v-SNARE binding protein that facilitates specific protein re-
trieval from a late endosome to the Golgi; modulates arginine uptake, possible role in mediating pH homeostasis between the vacuole and plasma membrane 1-1(+)-ATPase
5.55
CAGLOG04433g YIL108C PRI V I 10 Pheromone-regulated protein, predicted to have 5 transmem- brane segments; induced by treatment with 8-methoxypsoralen and UVA irradiation
5.14
CAGLOM06963g YNR034W SOL 1 Protein with a possible role in tRNA export; shows similar- ity to 6-phosphogluconolactonase non-catalytic domains but does not exhibit this enzymatic activity; homologous to Sol2p, Sol3p, and Sol4p
4.31
CAGLOE00803g YDR171W LISP42 Small heat shock protein (sHSP) with chaperone activity; forms barrel-shaped oligomers that suppress unfolded protein aggregation; involved in cytoskeleton reorganization after heat shock
3.82
CAGLOG03883g YLL026W HSP104 Heat shock protein that cooperates with Ydjlp (Hsp40) and 3.61 Ssalp (Hsp70) to refold and reactivate previously denatured, aggregated proteins; responsive to stresses including: heat, ethanol, and sodium arsenite; involved in EPSI4-1 propagation
CAGLOF08745g YLR327C TMAIO Protein of unknown function that associates with ribosomes 3.53 CAGLOG01540g YNL036W NCE103 Carbonic anhydrase; poorly transcribed under aerobic condi-
tions and at an undetectable level under anaerobic conditions; involved in non-classical protein export pathway
3.47
CAGLOJ00539g YHR030C SLT2 Serine/threonine MAP kinase involved in regulating the main- tenance of cell wall integrity and progression through the cell cycle; regulated by the PKC1-mediated signaling pathway
3.13
CAGLOE05632g YOR348C PUT4 Proline permease, required for high-affinity transport of pro- line; also transports the toxic proline analog azetidine-
2.85
2-carboxylate (AZC); PUT4 transcription is repressed in ammonia-grown cells
CAGLOK12034g YDR040C ENA I P-type ATPase sodium pump, involved in Na+ and Li-{- efflux to allow salt tolerance
2.61
CAGLOI01100g YOR120W GCYI Putative NADP(+) coupled glycerol dehydrogenase, pro- posed to be involved in an alternative pathway for glycerol catabolism; member of the aldo-keto reductase (AKR) family
2.56
CAGLOHD5137g YPL061W ALD6 Cytosolic aldehyde dehydrogenase, activated by Mg2+ and uti- lizes NADP+ as the preferred coenzyme; required for conver-sion of acetaldehyde to acetate; constitutively expressed; lo-cates to the mitochondrial outer surface upon oxidative stress
2.53
CAGLOM06347g YBR183W YPCI Alkaline ceramidase that also has reverse (CoA-independent) ceramide synthase activity, catalyzes both breakdown and syn-thesis of phytoceramide; overexpression confers fumonisin B1 resistance -
2.52
CAGLOL10186g YOR052C -2.45 CAGLOE06424g YKL150W MCR / Mitochondrial NADH-cytochrome b5 reductase, involved in er-
gosterol biosynthesis 2.42
CAGLOJ04158g YOR220W RCN2 Protein of unknown function; green fluorescent protein (GFP)- fusion protein localizes to the cytoplasm and is induced in re-sponse to the DNA-damaging agent MMS; phosphorylated in response to alpha factor
2.35
continued on next page

Appendix D. Chapter 3 Supplementary Data 240
continued from previous page CG ORF SC ORF Gene Name Description WT.S - WT.0
CAGL0104554g
CAGLOF01111g
Yl3R014C
YOL032W
GRX7
OPI10
Cis-golgi localized monothiol glutaredoxin; more similar in ac- tivity to dithiol than other monothiol glutaredoxins; involved in the oxidative stress response; does not bind metal ions; func-tional overlap with GRX6 Protein with a possible role in phospholipid biosynthesis, based on inositol-excreting phenotype of the null mutant and its sup-pression by exogenous choline
2.34
2.04

Appendix D. Chapter 3 Supplementary Data 241
Table D.13: C. glabrata hyperosmotic stress repressed transcripts that depend on a functional putative SHO1 signalling branch. C. glabrata transcripts induced in
wild-type, but not hogl or stell cells following exposure to transient hyper-osmotic shock (1M sorbitol, 15 minutes). repressed transcripts are defined as having log2(Cy5) - log2(Cy3) > 2.0 and Benjamini-Hochberg corrected p-values < 0.05. Estimated log-ratios correspond to the WT.S - WT.0 in Table D.2
CG ORF SC ORF Gene name Description WT.S - WT.0
CAGLOE04554g NORBH - Description not mapped 4.96 CAGLOM06963g YNR034W SOLI Protein with a possible role in tRNA export; shows similar-
ity to 6-phosphogluconolactonase non-catalytic domains but does not exhibit this enzymatic activity; homologous to Sol2p, Sol3p, and Sol4p
4.31
CAGLOL05016g YKL072W STB6 Protein that binds Sin3p in a two-hybrid assay 3.64 CAGLOF09119g YHR207C SET5 Zinc-finger protein of unknown function, contains one canoni-
cal and two unusual fingers in unusual arrangements; deletion enhances replication of positive-strand RNA virus
3.38
CAGLOM12320g YAL053W FLC2 Putative FAD transporter; required for uptake of FAD into endoplasmic reticulum; involved in cell wall maintenance
3.28
CAGLOL09251g NORBH - Description not mapped 3.17 CAGL00O2233g YER042W MXR1 Peptide methionine sulfoxide reductase, reverses the oxidation
of methionine residues; involved in oxidative damage repair, providing resistance to oxidative stress and regulation of lifes-pan
3.14
CAGLOD05082g YLL039C UBI4 Ubiquitin, becomes conjugated to proteins, marking them for selective degradation via the ubiquitin-26S proteasome system; essential for the cellular stress response; encoded as a polyu-biquitin precursor comprised of 5 head-to-tail repeats
2.99
CAGLOF08107g YGR244C LSC2 Beta subunit of succinyl-CoA ligase, which is a mitochon- drial enzyme of the TCA cycle that catalyzes the nucleotide-dependent conversion of succinyl-CoA to succinate
2.96
CAGLOB04257g YBRO31W RPL4A N-terminally acetylated protein component of the large (60S) ribosomal subunit, nearly identical to Rpl4Bp and has similar-ity to E. coli L4 and rat L4 ribosomal proteins
2.96
CAGLOJ01595g YPRO15C - Description not mapped 2.91 CAGLOJ00385g YHR049W FSH1 Putative serine hydrolase that localizes to both the nucleus
and cytoplasm; sequence is similar to S. cerevisiae Fsh2p and 2.89
Fsh3p and the human candidate tumor suppressor OVCA2 CAGLOJ03256g YER075C PTPS Phosphotyrosine-specific protein phosphatase involved in the
inactivation of mitogen-activated protein kinase (MAPK) dur-ing osmolarity sensing; dephosporylates Hog1p MAPK and regulates its localization; localized to the cytoplasm
2.83
CAGL0005137g YOL059W GPD2 NAD-dependent glycerol 3-phosphate dehydrogenase, homolog of Gpdlp, expression is controlled by an oxygen-independent signaling pathway required to regulate metabolism under anoxic conditions; located in cytosol and mitochondria
2.81
CAGLOF09163g YER182W FMP10 Putative protein of unknown function; the authentic, non- tagged protein is detected in highly purified mitochondria in high-throughput studies
2.74
CAGLOG01166g YLR34SC DICI Mitochondrial dicarboxylate carrier, integral membrane pro- tein, catalyzes a dicarboxylate-phosphate exchange across the inner mitochondria] membrane, transports cytoplasmic dicar-boxylates into the mitochondria] matrix
2.7
CAGLOF06941g YGL062W PYCI Pyruvate carboxylase isoform, cytoplasmic enzyme that con- verts pyruvate to oxaloacetate; highly similar to isoform Pyc2p but differentially regulated; mutations in the human homolog are associated with lactic acidosis
2.66
CAGLOL00803g YER054C GIP2 Putative regulatory subunit of the protein phosphatase Glc7p, involved in glycogen metabolism; contains a conserved motif
2.62
(GVNK motif) that is also found in Gaclp, Pig1p, and Pig2p continued on next page

Appendix D. Chapter 3 Supplementary Data 242
continued from previous page CG ORF SC ORF Gene name Description WT.S - WT.0
CAGLOM01782g YBRO80C SEC18 ATPase required for the release of Secl7p during the 'priming' step in homotypic vacuole fusion and for ER to Golgi trans-port; homolog of the mammalian NSF
2.59
CAGLOS01353g YMR089C YTA /2 Component, with Afg3p, of the mitochondrial inner membrane m-AAA protease that mediates degradation of misfolded or unassembled proteins and is also required for correct assembly of mitochondrial enzyme complexes
2.57
CAGLOIO1100g YOR120W GCY1 Putative NADP(+) coupled glycerol dehydrogenase, pro- posed to be involved in an alternative pathway for glycerol catabolism; member of the aldo-keto reductase (AKR) family
2.56
CAGLOB01991g YDR126W S WF/ Palmitoyltransferase that acts on the SNAREs Snclp, Syn8p, Tlglp and likely on all SNAREs; member of a family of pu-tative palmitoyltransferases containing an Asp-His-His-Cys-cysteine rich (DHHC-CRD) domain; may have a role in vacuole fusion
2.55
CAGLOF07095g YGL122C NA B2 Nuclear polyadenylated RNA-binding protein required for nu- clear mRNA export and poly(A) tail length control; binds nuclear pore protein MIplp; autoregulates mRNA levels; re-lated to human hnRNPs; nuclear localization sequence binds
2.54
Kap104p CAGLOH05137g YPL061.W ALD6 Cytosolic aldehyde dehydrogenase, activated by Mg2+ and uti-
lizes NADP+ as the preferred coenzyme; required for conver-sion of acetaldehyde to acetate; constitutively expressed; lo-cates to the mitochondria] outer surface upon oxidative stress
2.53
CAGLOM06347g YBR183W YPCI Alkaline ceramidase that also has reverse (CoA-independent) ceramide synthase activity, catalyzes both breakdown and syn-thesis of phytoceramide; overexpression confers fumonisin B1 resistance
2.52
CAGLOL09152g YFR0O4W RPN11 Metalloprotease subunit of the 19S regulatory particle of the 2.52 26S proteasome lid; couples the deubiquitination and degrada-tion of proteasome substrates
CAGLOH03729g YNLOO6W LST8 Protein required for the transport of amino acid permease 2.48 Gaplp from the Golgi to the cell surface; component of the TOR signaling pathway; associates with both Torlp and Tor2p; contains a WD-repeat
CAGLOL00605g YKL 190W CNB1 Calcineurin B; the regulatory subunit of calcineurin, a 2.46 Ca++/calmodulin-regulated type 2B protein phosphatase which regulates Crzlp (a stress-response transcription factor), the other calcineurin subunit is encoded by CNA1 and/or CMP1
CAGLOE06424g YKL150W 11,1 CR 1 Mitochondrial NADII-cytochrome b5 reductase, involved in er- gosterol biosynthesis
2.42
CAGLOJ03322g YER080W A IA12 Putative protein of unknown function; the authentic, non- tagged protein is detected in highly purified mitochondria in high-throughput studies
2.4
CAGLOM10263g YCLOO2C - Description not mapped 2.39 CAGLOL06028g YGL181W GTS1 Protein that localizes to the nucleus and is involved in tran-
scription regulation; also localizes to actin patches and plays a role in endocytosis; N-terminus contains an ARF-GAP domain and C-terminus contains a Gln-rich domain
2.38
CAGLOM14047g YMR318C ADH6 NADPH-dependent medium chain alcohol dehydrogenase with broad substrate specificity; member of the cinnamyl family of alcohol dehydrogenases; may be involved in fusel alcohol syn-thesis or in aldehyde tolerance
2.36
CAGL0004499g YDLOO6W PTC1 Type 2C protein phosphatase (PP2C); inactivates the os- mosensing MAPK cascade by dephosphorylating Hoglp; muta-tion delays mitochondria] inheritance; deletion reveals defects in precursor tRNA splicing, sporulation and cell separation
2.35
CAGLOI04554g YBRO14C GRX7 Cis-golgi localized monothiol glutaredoxin; more similar in ac- tivity to dithiol than other monothiol glutaredoxins; involved in the oxidative stress response; does not bind metal ions; func-tional overlap with GRX6
2.34
continued on next page

Appendix D. Chapter 3 Supplementary Data 243
continued from previous page CG ORF SC ORF Gene name Description WT.S - WT.0
CAGLOD04752g YPR127W Description not mapped 2.33 CAGLOB00836g YCL033C - Description not mapped 2.33 CAGLOGO6886g YJL017W - Description not mapped 2.3 CAGLOA03058g YDR379W RGA2 GTPase-activating protein for the polarity-establishment pro-
tein Cdc42p; implicated in control of septin organization, pheromone response, and haploid invasive growth; regulated by Pho85p and Cdc28p
2.27
CAGL0000363g YJL174W KRE9 Glycoprotein involved in cell wall beta-glucan assembly; null mutation leads to severe growth defects, aberrant multibudded morphology, and mating defects
2.27
CAGLOJ11506g YNL192W CHS1 Chitin synthase I, requires activation from zymogenic form in order to catalyze the transfer of N-acetylglucosamine (GIcNAc) to chitin; required for repairing the chitin septum during cy-tokinesis; transcription activated by mating factor
2.25
CAGLOG04873g YLR371W ROM2 GDP/GTP exchange protein (GEP) for Rho1p and Rho2p; mutations are synthetically lethal with mutations in roml, which also encodes a GEP
2.25
CAGLOA04587g YBL082C ALG3 Dolichol-P-Man dependent alpha(1-3) mannosyltransferase, involved in the synthesis of dolichol-linked oligosaccharide donor for N-linked glycosylation of proteins
2.23
CAGLOM11242g YMR226C TMA29 NADP(+)-dependent dehydrogenase; acts on serine, L-allo- threonine, and other 3-hydroxy acids; green fluorescent protein fusion protein localizes to the cytoplasm and nucleus; may in-teract with ribosomes, based on co-purification experiments
2.21
CAGLOJ04136g YOR221C MGT/ Predicted malonyl-CoA:ACP transferase, putative component of a type-II mitochondrial fatty acid synthase that produces intermediates for phospholipid remodeling
2.19
CAGLOG08382g YDR195W REFS RNA-binding protein involved in the cleavage step of mRNA 2.17 3'-end formation prior to polyadenylation, and in snoRNA maturation; part of holo-CPF subcomplex APT, which asso-ciates with 3'-ends of snoRNA- and mRNA-encoding genes
CAGLOF07513g YKL093W MBR1 Protein involved in mitochondrial functions and stress re- sponse; overexpression suppresses growth defects of hap2, hap3, and hap4 mutants
2.16
CAGLOL05126g YKLOSOW VMA5 Subunit C of the eight-subunit V1 peripheral membrane do- main of vacuolar H-I--ATPase (V-ATPase), an electrogenic pro-ton pump found throughout the endomembrane system; re-quired for the V1 domain to assemble onto the vacuolar mem-brane
2.15
CAGL0107513g YOL100W PKH2 Serine/threonine protein kinase involved in sphingolipid- mediated signaling pathway that controls endocytosis; acti-vates Ypklp and Ykr2p, components of signaling cascade re-quired for maintenance of cell wall integrity; redundant with
2.15
Pkhlp CAGLOI00418g YGL055W OLEI Delta(9) fatty acid desaturase, required for monounsaturated
fatty acid synthesis and for normal distribution of mitochon-dria
2.13
CAGLOJ00583g YHR028C DAP2 Dipeptidyl aminopeptidase, synthesized as a glycosylated pre- cursor; localizes to the vacuolar membrane; similar to Ste13p
2.12
CAGLOF00583g YDR517W GRH1 Acetylated, cis-golgi localized protein involved in ER to Golgi transport; homolog of human GRASP65; forms a complex with the coiled-coil protein Bug1p; mutants are compromised for the fusion of ER-derived vesicles with Golgi membranes
2.12
CAGLOK01287g YGRO74W SMD1 Core Sm protein Sm DI; part of heteroheptameric complex 2.11 (with Smblp, Smd2p, Smd3p, Smelp, Smx3p, and Smx2p) that is part of the spliceosomal Ul, U2, U4, and US snRNPs; homolog of human Sm D1
CAGL00O2717g YAL009W SPOT Putative regulatory subunit of Nemlp-Spo7p phosphatase holoenzyme, regulates nuclear growth by controlling phospho-lipid biosynthesis, required for normal nuclear envelope mor-phology, premeiotic replication, and sporulation
2.1
C AG L0J11704g YMR004W MVP1 Protein required for sorting proteins to the vacuole; overpro- duction of Mvplp suppresses several dominant VPS1 muta-tions; Mvplp and Vpslp act in concert to promote membrane traffic to the vacuole
2.08
continued on next page

Appendix D. Chapter 3 Supplementary Data 244
continued from previous page CC ORF SC ORF Gene name Description WT .S - WT. U
CAGLOG06556g YNL097C PH02.9 Probable component of the Rpd3 histone deacetylase complex, involved in transcriptional regulation of PHO5; C-terminus has similarity to human candidate tumor suppressor p33(ING1) and its isoform ING3
2.07
CAGLOE04070g YHL039W - Description not mapped 2.05 CAGLOFO1111g YOL032W OP/I0 Protein with a possible role in phospholipid biosynthesis, based
on inositol-excreting phenotype of the null mutant and its sup-pression by exogenous choline
2.04
CAGL0005357g YDL042C SIRS Conserved NAD+ dependent histone deacetylase of the Sirtuin family involved in regulation of lifespan; plays roles in silencing at HML, HMR, telomeres, and the rDNA locus; negatively regulates initiation of DNA replication
2.02
CAGLOK02013g NORBH - Description not mapped 2.02 CAGL0109592g YOR303W CPA1 Small subunit of carbamoyl phosphate synthetase, which cat-
alyzes a step in the synthesis of citrulline, an arginine pre-cursor; translationally regulated by an attenuator peptide en-coded by YOR302W within the CPA1 mRNA 5'-leader
2

Appendix D. Chapter 3 Supplementary Data 245
Table D.14: C. glabrata hyperosmotic stress induced transcripts that are inde-
pendent of HOG1 and STE11. C. glabrata transcripts induced in wild-type, hogl and
stell cells following exposure to transient hyper-osmotic shock. Induced transcripts are defined as having log2(Cy5) - log2(Cy3) > 2.0 and Benjamini-Hochberg corrected p-values < 0.05. Estimated log-ratios correspond to the WT.S - WT.U, HOG.S - HOG.0 and STE.S - STE.0 effects described in Table D.2
CG ORF SC ORF Gene name Description WT.S - WT.0 HOGS - HOG.0 STE.S - STE.0
CAGLOI10010g YGR142W BTN2 v-SNARE binding protein that fa- cilitates specific protein retrieval from a late endosome to the Golgi; modulates arginine uptake, possi- ble role in mediating pH homeosta-sis between the vacuole and plasma membrane H(-1-)-ATPase
5.55 5.26 6.39
CAGL00O2321g YER037W PHM8 Protein of unknown function, ex- pression is induced by low phos- phate levels and by inactivation of
5.41 2.93 2.86
Pho85p CAGLOG04433g YJL108C PRMIO Pheromone-regulated protein, pre-
dicted to have 5 transmembrane segments; induced by treatment with 8-methoxypsoralen and UVA irradiation
5.14 4.41 4.12
CAGLOM09713g YMR152W YIM1 Protein of unknown function; null mutant displays sensitivity to DNA damaging agents; the authentic, non-tagged protein is detected in highly purified mitochondria in high-throughput studies -
4.9 4.7 4.16
CAGLOK08338g YOR385W -4.57 5.14 3.71 CAGLOA02816g YDR368W YPRI NADPH-dependent aldo-keto
reductase, reduces multi- ple substrates including 2- methylbutyraldehyde and D,L- glyceraldehyde, expression is induced by osmotic and oxidative stress; functionally redundant with other aldo-keto reductases
4.43 2.1 2.19
CAGLOJ10846g YHR071W PCL5 Cyclin, interacts with and phos- phorylated by Pho85p cyclin- dependent kinase (Cdk), induced by Gcn4p at level of transcrip- tion, specifically required for
4.09 4.3 3.39
Gcn4p degradation, may be sensor of cellular protein biosynthetic capacity
CAGL0105390g YPL026C SKS1 Putative serine/threonine protein kinase; involved in the adaptation to low concentrations of glucose in-dependent of the SNF3 regulated pathway
3.97 3.69 3.22
CAGLOM08822g YDR258C HSP7R Oligomeric mitochondrial matrix chaperone that cooperates with
3.93 3.91 4.94
Ssclp in mitochondrial thermotol-erance after heat shock; able to pre-vent the aggregation of misfolded proteins as well as resolubilize pro-tein aggregates
continued on next page

Appendix D. Chapter 3 Supplementary Data 246
continued from previous page CG ORF SC ORF Gene name Description WT.S - WT.0 HOG.S - HOG.0 STE.S - STE.0
CAGLOH08151g NORBH - - 3.86 3.67 3.1 CAGLOE00803g YDR171W HSP42 Small heat shock protein (sIISP)
with chaperone activity; forms barrel-shaped oligomers that sup-press unfolded protein aggregation; involved in cytoskeleton reorganiza-tion after heat shock
3.82 4.38 4.02
CAGLOG03883g YLL026W HSP/04 Heat shock protein that cooper- ales with Ydjlp (Hsp40) and Ssalp
3.61 3.51 3.38
(Hsp70) to refold and reactivate previously denatured, aggregated proteins; responsive to stresses in-cluding: heat, ethanol, and sodium arsenite; involved in [PSI-I-] propa-gation
CAGLOH06523g YIL145C PANG Pantothenate synthase, also known as pantoate-beta-alanine ligase, re-quired for pantothenic acid biosyn-thesis, deletion causes pantothenic acid auxotrophy, homologous to E. coli panC
3.6 3.23 2.82
CAGLOF08745g YLR327C TMAIO Protein of unknown function that associates with ribosomes
3.53 4.7 2.16
CAGLOH08173g YOR028C CIN5 Basic leucine zipper (bZIP) tran- scription factor of the yAP-1 fam- ily, mediates pleiotropic drug re- sistance and salt tolerance; nucle- arly localized under oxidative stress and sequestered in the cytoplasm by
3.49 2.14 2.61
Lot6p under reducing conditions CAGLOG01540g YNL036W NCE103 Carbonic anhydrase; poorly tran-
scribed under aerobic conditions and at an undetectable level under anaerobic conditions; involved in non-classical protein export path-way
3.47 2.7 2.44
CAGLOK00715g YGR213C RTA1 Protein involved in 7- aminocholesterol resistance; has seven potential membrane-spanning regions; expression is induced un-der both low-heme and low-oxygen conditions
3.3 5.14 3.11
CAGLOG00858g YLR332W MID2 0-glycosylated plasma membrane protein that acts as a sensor for cell wall integrity signaling and activates the pathway; interacts with Rom2p, a guanine nucleotide exchange factor for Rholp, and with cell integrity pathway protein
3.23 4.33 3.14
Zeolp CAGLOG00242g YGR281W YOR1 Plasma membrane ATP-binding
cassette (ABC) transporter, mul- tidrug transporter mediates export of many different organic anions including oligomycin; similar to hu-man cystic fibrosis transmembrane receptor (CFTR)
3.21 3.25 3.28
CAGLOD03916g YHR034C PIHI Protein of unresolved function; may function in protein folding and/or rRNA processing, interacts with a chaperone (Ilsp82p), two chromatin remodeling factors (Rvblp, Rvb2p) and two rRNA processing factors
3.1 2.88 3.05
(Rrp43p, Nop58p) continued on next page

Appendix D. Chapter 3 Supplementary Data 247
continued from previous page CG ORF SC ORF Gene name Description WT.S - WT.1.1 HOG.S - HOG.0 STE.S - STE.0
CAGLOH08844g
CAGLOE03025g CAGLOM02255g
YMR173W
NORBH YPL152W
DDR48
- RRD2
DNA damage-responsive protein, expression is increased in response to heat-shock stress or treatments that produce DNA lesions; contains multiple repeats of the amino acid sequence NNNDSYGS - Activator of the phosphoty- rosyl phosphatase activity of PP2A,peptidyl-prolyl cis/trans- isomerase; regulates G1 phase progression, the osmoresponse, microtubule dynamics; subunit of the Tap42p-Pph2lp-Rrd2p complex
3.06
2.94 2.92
2.32
2.92 3.37
2.39
3.06 3.12
CAGLOG04763g YOR1071V RGS2 Negative regulator of glucose- induced cAMP signaling; directly activates the GTPase activity of the heterotrimeric G protein alpha subunit Gpa2p
2.84 3.3 2.39
CAGLOL05434g YKR042W UTHI Mitochondria' outer membrane and cell wall localized SUN family mem-ber required for mitochondrial au-tophagy; involved in the oxidative stress response, life span during starvation, mitochondrial biogene-sis, and cell death
2.83 4.39 3.62
CAGLOG07601g YBR287W - - 2.79 2.43 3.46 CAGLOA02046g YOR018W ROD] Membrane protein that binds the
ubiquitin ligase Rsp5p via its 2 PY motifs; overexpression confers re- sistance to the GST substrate o- dinitrobenzene,zinc, and calcium; proposed to regulate the endocyto-sis of plasma membrane proteins
2.79 2.92 3.7
CAGLOL07480g YBRO66C NRG2 Transcriptional repressor that me- diates glucose repression and nega-tively regulates filamentous growth; has similarity to Nrglp
2.64 2.49 3.15
CAGL0003201g YLL040C VPS13 Protein of unknown function; het- erooligomeric or homooligomeric complex; peripherally associated with membranes; homologous to human COH1; involved in sporula-tion, vacuolar protein sorting and protein-Golgi retention
2.63 2.76 3.01
CAGLOJ10296g YNL077W A PJ/ Putative chaperone of the HSP40 2.62 4.21 2.7 (DNAJ) family; overexpression interferes with propagation of the 1Psi-1-] prion; the authentic, non-tagged protein is detected in highly purified mitochondria in high-throughput studies
CAGLOK12034g YDR040C ENA1 P-type ATPase sodium pump, in- volved in Nal- and Li-t- efflux to al-low salt tolerance
2.61 3.29 2.39
CAGLOI07007g YGL219C Ill DM34 Mitochondrial outer membrane pro- tein required for normal mito- chondrial morphology and inher- itance; localizes to dots on mi- tochondrial surface near mtDNA nucleoids; ubiquitination target of
2.55 3.07 2.16
Mdm30p; interacts genetically with MDM31 and MDM32
continued on next page

Appendix D. Chapter 3 Supplementary Data 248
continued from previous page CG ORF SC ORF Gene name Description WT.S - WT. U HOGS - HOG.0 STE.S - STE. U
CAGLOA02024g YOL025W LAG2 Protein involved in determination of longevity; LAG2 gene is preferen-tially expressed in young cells; over-expression extends the mean and maximum life span of cells -
2.52 2.84 2.64
CAGLOJ11176g YNL176C -2.51 3.31 2.64 CAGLOL04708g YGR111W - - 2.48 2.28 2.3 CAGLOL11572g YHR103W SDE22 Protein involved in the transport
of cell wall components from the 2.43 2.56 2.29
Golgi to the cell surface; similar in structure and functionally redun-dant with Sbe2p; involved in bud growth
CAGLOG05313g YDR072C IP TI Inositolphosphotransferase, in- volved in synthesis of mannose-
2.34 2.65 2.15
(inositol-P)2-ceramide (M(IP)2C), the most abundant sphingolipid;, can mutate to resistance to the antifungals syringomycin E and DmAMP1 and to K. lactis zymocin
CAGLOK03509g YMR110C HFD 1 Putative fatty aldehyde dehydro- genase, located in the mitochon- dria) outer membrane and also in lipid particles; has similarity to hu-man fatty aldehyde dehydrogenase
2.3 3.32 2.26
(FALDH) which is implicated in Sjogren-Larsson syndrome
CAGLOA02321g YDR345C HXT3 Low affinity glucose transporter of the major facilitator superfamily, expression is induced in low or high glucose conditions
2.28 2.36 2.11
CAGLOL07238g YDL097C RP N6 Essential, non-ATPase regulatory subunit of the 26S proteasome lid required for the assembly and activ-ity of the 265 proteasome; the hu-man homolog (S9 protein) partially rescues Rpn6p depletion
2.25 2.01 2.76
CAGLOM06545g YKL124W SSH4 Specificity factor required for 2.24 2.69 2.21 Rsp5p-dependent ubiquitination and sorting of cargo proteins at the multivesicular body; identified as a high-copy suppressor of a SHR3 deletion, increasing steady-state levels of amino acid permeases
CAGLOK12892g YFLO47W RC D2 GTPase-activating protein 2.2 3.62 2.14 (RhoGAP) for Cdc42p and Rho5p
CAGLOL10912g YOR273C TPO4 Polyamine transport protein, rec- ognizes spermine, putrescine, and spermidine; localizes to the plasma membrane; member of the major fa-cilitator superfamily
2.12 2.27 3.14
CAGLOA03806g YOR298W MUMS Protein of unknown function in- volved in the organization of the outer spore wall layers; has similar-ity to the tafazzins superfamily of acyltransferases
2.11 2.2 2.97
CAGLOJ09328g YDL127W PCL2 G1 cyclin, associates with Pho85p cyclin-dependent kinase (Cdk) to contribute to entry into the mitotic cell cycle, essential for cell morpho-genesis; localizes to sites of polar-ized cell growth
2.09 3.68 3.02
continued on next page

Appendix D. Chapter 3 Supplementary Data 249
continued from previous page CG ORF SC ORF Gene name Description WT.S - WT.0 HOG.S - HOG.0 STE.S - STE.0
CAGLOL11110g YML057W CAIP2 Calcineurin A; one isoform (the other is CNA1) of the cat- alytic subunit of calcineurin, a
2.05 2.68 2.74
Ca+-1-/calmodulin-regulated pro- tein phosphatase which regulates Crzlp (a stress-response transcrip-tion factor), the other calcineurin subunit is CNB1
CAGL0004917g YJR109C CPA2 Large subunit of carbamoyl phos- phate synthetase, which catalyzes a step in the synthesis of citrulline, an arginine precursor
2.03 2.93 3.31
CAGL0004785g YJR115W - - 2.02 2.81 2.63

Appendix D. Chapter 3 Supplementary Data 250
Table D.15: HOG/-dependent hyperosmotic stress induced C. glabrata tran-scripts. C. glabrata transcripts induced in wild-type and stel 1 cells, but not hog] cells
following exposure to transient hyper-osmotic shock (1M sorbitol, 15 minutes). Induced transcripts are defined as having log2(Cy5) - log2(Cy3) > 2.0 and Benjamini-Hochberg cor-rected p-values < 0.05. Estimated log-ratios correspond to the WT.S - WT.0 in Table D.2
CG ORF SC ORF Gene Name Description WT.S - WT.0 STE.S - STE.0
CAGL0I02684g YMR2S5C NGL2 Protein involved in 5.8S rRNA pro- cessing; Ccr4p-like RNase required for correct 3'-end formation of 5.8S rRNA at site E; similar to Ngllp and Ngl3p
3.2 2.19
CAGLOK02761g YHLOO2W FISE1 Subunit of the endosomal Vps27p- 3.2 3.01 Hselp complex required for sorting of ubiquitinated membrane proteins into intralumenal vesicles prior to vacuolar degradation, as well as for recycling of Golgi proteins and for-mation of lumenal membranes - CAGLOH02519g YMR253C -2.65 3.46
CAGL0G07689g YKR014C YPT52 GTPase, similar to Ypt5lp and 2.48 2.47 Ypt53p and to mammalian rab5; re-quired for vacuolar protein sorting and endocytosis
CAGLOL10186g YOR052C - - 2.45 2.92
CAGLOE02299g YOL013C HRD1 Ubiquitin-protein ligase required for endoplasmic reticulum- associated degradation (ERAD) of misfolded proteins; genetically linked to the unfolded protein response (UPR); regulated through association with Hrd3p; contains an H2 ring finger
2.31 2.18
CAGLOG05049g YDR063W AIM7 Putative protein of unknown func- tion; green fluorescent protein
2.28 2.75
(GFP)-fusion protein localizes to the cytoplasm and nucleus; YDR063W is not an essential gene
CAGL0104774g YB R036C CSG2 Endoplasmic reticulum membrane protein, required for mannosylation of inositolphosphorylceramide and for growth at high calcium concen-trations
2.25 3.48
CAGLOH07007g YML092C PRE8 Alpha 2 subunit of the 20S protea- some
2.23 2.03
CAGLOM09108g YJR091C JSNI Member of the Puf family of 2.21 2.07 RNA-binding proteins, interacts with mRNAs encoding membrane-associated proteins; overexpression suppresses a tub2-150 mutation and causes increased sensitivity to beno-myl in wild-type cells
CAGLOK11022g YDR236C FAIN1 Riboflavin kinase, phosphory- lates riboflavin to form riboflavin monophosphate (FMN), which is a necessary cofactor for many enzymes; localizes to microsomes and to the mitochondria] inner membrane
2.18 2.74
continued on next page

Appendix D. Chapter 3 Supplementary Data 251
continued from previous page CC ORF SC ORF Gene name Description WT.S - WT.0 STE.S - STE.0
CAGLOD01474g
CAGL0M06875g
CAGL0D03322g
YBR108W
YNL025C
YLR023C
AIMS
SSN8
IZH3
Protein interacting with Rys167p; YBR108W is not an essential gene Cyclin-like component of the RNA polymerase II holoenzyme, involved in phosphorylation of the RNA polymerase II C-terminal domain; involved in glucose repression and telomere maintenance Membrane protein involved in zinc metabolism, member of the four-protein IZII family, expression in- duced by zinc deficiency; deletion reduces sensitivity to elevated zinc and shortens lag phase, overexpres-sion reduces Zap1p activity
2.17
2.15
2.12
2.39
2.38
2.01
CAGL00O2673g YLR392C - - 2.11 2.03
CAGLOL09889g YKL025C PAN3 Essential subunit of the Pan2p- 2.09 2.68
Pan3p poly(A)-ribonuclease com- plex, which acts to control poly(A) tail length and regulate the stoi-chiometry and activity of postrepli-cation repair complexes
CAGL0K09526g YDR394W RPTS One of six ATPases of the 19S reg- ulatory particle of the 26S protea-some involved in the degradation of ubiquitinated substrates; substrate of N-acetyltransferase B
2.09 2.11
CAGLOB03619g YEL060C PRI31 Vacuolar proteinase B (yscB), a ser- ine protease of the subtilisin fam-ily; involved in protein degradation in the vacuole and required for full protein degradation during sporula-tion
2.09 2.17
CAGLOF04829g YPR154W PINS Protein that induces appearance of 2.05 3.37 (PIN-k] prion when overproduced
CAGLOE04510g YOR022C - 2.01 2.29

Appendix D. Chapter 3 Supplementary Data 252
Table D.16: STE11-dependent hyperosmotic stress induced C. glabrata tran-
scripts. C. glabrata transcripts induced in wild-type and hog] cells, but not stell cells
following exposure to transient hyper-osmotic shock (1M sorbitol, 15 minutes). Induced
transcripts are defined as having log2(Cy5) - log2(Cy3) > 2.0 and Benjamini-Hochberg cor-
rected p-values < 0.05. Estimated log-ratios correspond to the WT.S - WT.0 in Table D.2
CG ORF SC ORF Gene Name Description WT.S - WT.0 HOGS - HOG . U
CAGLOM12474g YIL055C - - 3.46 3.07
CAGLOM07381g YPL053C KTR6 Probable mannosylphosphate transferase involved in the syn- thesis of core oligosaccharides in protein glycosylation pathway; member of the KRE2/MNT1 mannosyltransferase family
3.45 3.54
CAGLOJ00539g YHR030C SLT2 Serine/threonine MAP kinase in- volved in regulating the mainte- nance of cell wall integrity and pro-gression through the cell cycle; reg-ulated by the PKC1-mediated sig-naling pathway
3.13 4.02
CAGLOK00539g YPL259C APMI Mul-like medium subunit of the clathrin-associated protein complex
3.1 3.96
(AP-1); binds clathrin; involved in clathrin-dependent Golgi protein sorting
CAGLOM12034g YAL038W CDC19 Pyruvate kinase, functions as a ho- motetramer in glycolysis to convert phosphoenolpyruvate to pyruvate, the input for aerobic (TCA cycle) or anaerobic (glucose fermentation) respiration
3.09 4.32
CAGL0104246g YGL162W S UT/ Transcription factor of the 3.04 4.21 Zn[II]2Cys6 family involved in sterol uptake; involved in induction of hypoxic gene expression
CAGLOH04631g YMLOO7W YAP1 Basic leucine zipper (bZIP) tran- scription factor required for oxida- tive stress tolerance; activated by
2.98 3.6
H202 through the multistep forma-tion of disulfide bonds and transit from the cytoplasm to the nucleus; mediates resistance to cadmium
CAGLOD05368g YBR253W SRB6 Subunit of the RNA polymerase II mediator complex; associates with core polymerase subunits to form the RNA polymerase II holoen- zyme; essential for transcriptional regulation
2.97 2.55
CAGLOE05632g YOR348C PUT4 Proline permease, required for high- affinity transport of proline; also transports the toxic proline ana-log azetidine-2-carboxylate (AzC);
2.85 3.55
PUT4 transcription is repressed in ammonia-grown cells
CAGLOJ00253g YGRO23W MTL1 Protein with both structural and functional similarity to Mid2p, which is a plasma membrane sensor required for cell integrity signaling during pheromone-induced morpho-genesis; suppresses rgdl null muta-tions
2.83 2.86
continues on next page

Appendix D. Chapter 3 Supplementary Data 253
continued from previous page CG ORF SC ORF Gene name Description WT.S - WT.0 HOG.S - HOG.0
CAGLOM01628g
CAGLOH09812g
YDR389W
YDROO6C
S AC7
SOK1
GTPase activating protein (GAP) for Rho1p, involved in signaling to the actin cytoskeleton, null muta-tions suppress tort mutations and temperature sensitive mutations in actin; potential Cdc28p substrate Protein whose overexpression sup- presses the growth defect of mu-tants lacking protein kinase A ac-tivity; involved in cAMP-mediated signaling; localized to the nucleus; similar to the mouse testis-specific protein PBS13
2.77
2.77
4.08
3.42
CAGLOH07557g YGL254W FZF/ Transcription factor involved in sul- fite metabolism, sole identified reg-ulatory target is SSU1, overexpres-sion suppresses sulfite-sensitivity of many unrelated mutants due to hy-peractivation of SSU1, contains five zinc fingers
2.68 2.24
CAGLOM11066g YGR109C CLB6 B-type cyclin involved in DNA replication during S phase; acti- vates Cdc28p to promote initiation of DNA synthesis; functions in for- mation of mitotic spindles along with Clb3p and C1b4p; most abun-dant during late G1
2.64 2.23
CAGLOE00649g YCR079W PTC6 Mitochondrial protein phosphatase of type 2C with similarity to mam- malian PP1Ks; involved in mi- tophagy; null mutant is sensitive to rapamycin and has decreased phos-phorylation of the Pdal subunit of pyruvate dehydrogenase
2.53 4.06
CAGL0,104466g YLR414C - - 2.5 2.92 CAGLOA04081g YLR194C - 2.48 3.88 CAGLOK00891g YGR205W - - 2.42 2.53 CAGLOJ09108g YDL190C UFD2 Ubiquitin chain assembly fac-
tor (E4) that cooperates with a ubiquitin-activating enzyme (El), a ubiquitin-conjugating enzyme
2.39 2.07
(E2), and a ubiquitin protein ligase (E3) to conjugate ubiquitin to substrates; also functions as an E3
CAGLOD05434g YPRO65W ROX1 Heme-dependent repressor of hy- poxic genes; contains an HMG do-main that is responsible for DNA bending activity
2.38 2.58
CAGLOJ04158g YOR220W RCN2 Protein of unknown function; green fluorescent protein (GFP)-fusion protein localizes to the cytoplasm and is induced in response to the
2.35 2.1
DNA-damaging agent MMS; phos-phorylated in response to alpha fac-tor
CAGL0004433g YDL003W MOD 1 Essential subunit of the cohesin complex required for sister chro- matid cohesion in mitosis and meio-sis; apoptosis induces cleavage and translocation of a C-terminal frag- ment to mitochondria; expression peaks in S phase
2.22 2.41
continues on next page

Appendix D. Chapter 3 Supplementary Data 254
continued from previous page CG ORF SC ORF Gene name Description WT.S - WT.0 HOG.S - HOG.11
CAGL0005093g CAGLOM03839g CAGL0108239g
YOL057W YNL305C YER021W
- -
RPN3
- - Essential, non-ATPase regulatory subunit of the 26S proteasome lid, similar to the p58 subunit of the hu-man 26S proteasome; temperature-sensitive alleles cause metaphase ar-rest, suggesting a role for the pro-teasome in cell cycle control
2.21 2.2 2.2
3.05 2.94 2.55
CAGLOM04081g YNL317W PFS2 Integral subunit of the pre-mRNA cleavage and polyadenylation factor
2.2 3.07
(CPF) complex; plays an essential role in mRNA 3'-end formation by bridging different processing factors and thereby promoting the assem-bly of the processing complex
CAGLOH09130g YKL201C MNN4 Putative positive regulator of mannosylphosphate transferase
2.14 2.63
(Mnn6p), involved in manno- sylphosphorylation of N-linked oligosaccharides; expression in- creases in late-logarithmic and stationary growth phases
CAGLOM08206g YJL171C - 2.14 3.13
CAGLOD02948g YJL034W KAR2 ATPase involved in protein import into the ER, also acts as a chap-erone to mediate protein folding in the ER and may play a role in ER export of soluble proteins; regulates the unfolded protein response via interaction with Ire1p
2.12 2.29
CAGLOK05995g YLR260W LCB5 Minor sphingoid long-chain base ki- nase, paralog of Lcb4p responsible for few percent of the total activ-ity, possibly involved in synthesis of long-chain base phosphates, which function as signaling molecules
2.12 2.35
CAGLOJ01001g YJL036W SNX4 Sorting nexin, involved in retrieval of late-Golgi SNAREs from post-
2.1 2.84
Golgi endosomes to the trans- Golgi network and in cytoplasm to vacuole transport; contains a PX phosphoinositide-binding do- main; forms complexes with Snx4lp and with Atg2Op
CAGLOG01034g YGR032W GSC2 Catalytic subunit of 1,3-beta- glucan synthase, involved in formation of the inner layer of the spore wall; activity positively regulated by Rho1p and negatively by Smklp; has similarity to an alternate catalytic subunit, Fkslp
2.08 2.73
(Gsclp) CAGLOD05478g YGR186W TFG1 TFIIF (Transcription Factor II)
largest subunit; involved in both transcription initiation and elonga-tion of RNA polymerase II; homol-ogous to human RAP74
2.08 3.07
CAGL00O2541g YLR399C BDF1 Protein involved in transcription initiation at TATA-containing pro- moters; associates with the basal transcription factor TFIID; con- tains two bromodomains; corre- sponds to the C-terminal region of mammalian TAF1; redundant with
2.04 2.25
Bdf2p continues on next page

Appendix D. Chapter 3 Supplementary Data 255
continued from previous page CG ORF SC ORF Gene name Description WT.S - WT.0 HOG.S - HOG.0
CAGLOK11946g I NORBH I - I - 2.04 3.9

Appendix D. Chapter 3 Supplementary Data 256
Table D.17: C. glabrata hyperosmotic stress repressed transcripts that depend on a functional putative SHO1 signalling branch. C. glabrata transcripts repressed in
wild-type, but not hogl or sten cells following exposure to transient hyper-osmotic shock (1M sorbitol, 15 minutes). repressed transcripts are defined as having log2(Cy5) - log2(Cy3) < -2.0 and Benjamini-Hochberg corrected p-values < 0.05. Estimated log-ratios correspond to the WT.S - WT.0 in Table D.2
CG ORF SC ORF Gene Name Description WT.S - WT.0
CAGLOL08470g NORBH - Description not mapped -2.03 CAGLOL01551g YML052W SUR7 Putative integral membrane protein; component of eisosomes;
associated with endocytosis, along with Pillp and Lsplp; sporulation and plasma membrane sphingolipid content are al-tered in mutants
-2.04
CAGL00O2739g YAL008W FUNI4 Mitochondria] protein of unknown function -2.05
CAGLO/02442g YIL022W TIM44 Peripheral mitochondria] membrane protein involved in mito- chondrial protein import, tethers essential chaperone Ssclp to the translocon channel at the matrix side of the inner mem-brane
-2.09
CAGLOL01991g YKL033W TTI1 Putative protein of unknown function; subunit of the ASTRA complex which is part of the chromatin remodeling machin-ery; similar to S. pombe Ttilp; detected in highly purified mitochondria in high-throughput studies
-2.09
CAGLOL13156g YLR073C - Description not mapped -2.1
CAGL0G08646g YIL122W POC/ Putative transcriptional activator that promotes recovery from pheromone induced arrest; inhibits both alpha-factor induced
-2.12
G1 arrest and repression of CLN1 and CLN2 via SCB/MCB promoter elements; potential Cdc28p substrate; SBF regulated
CAGLOK04477g YGR060W ERG25 C-4 methyl sterol oxidase, catalyzes the first of three steps required to remove two C-4 methyl groups from an intermedi-ate in ergosterol biosynthesis; mutants accumulate the sterol intermediate 4,4-dimethylzymosterol
-2.15
CAGLOB00638g YCL047C - Description not mapped -2.16
CAGLOD00748g YDL067C COX9 Subunit VIIa of cytochrome c oxidase, which is the terminal member of the mitochondria] inner membrane electron trans-port chain
-2.18
CAGL00O2211g YELO4OW UTR2 Cell wall protein that functions in the transfer of chitin to beta(1-6)glucan; putative chitin transglycosidase; glyco- sylphosphatidylinositol (GPI)-anchored protein localized to the bud neck; has a role in cell wall maintenance
-2.27
CAGLOH05533g YPL082C MO T/ Essential abundant protein involved in regulation of transcrip- tion, removes Sptl5p (TBP) from DNA via its C-terminal AT-
-2.31
Pase activity, forms a complex with TBP that binds TATA DNA with high affinity but with altered specificity
CAGLOM08448g YKL165C MCD4 Protein involved in glycosylphosphatidylinositol (GPI) anchor synthesis; multimembrane-spanning protein that localizes to the endoplasmic reticulum; highly conserved among eukaryotes
-2.32
CAGLOK02233g YER126C NSA2 Protein constituent of 66S pre-ribosomal particles, contributes to processing of the 27S pre-rRNA
-2.38
CAGLOL03025g YKR025W RPC37 RNA polymerase III subunit C37 -2.4
CAGLOI01826g YHR148W IMPS Component of the SSU processome, which is required for pre- -2.42 18S rRNA processing, essential protein that interacts with MpplOp and mediates interactions of Imp4p and MpplOp with U3 snoRNA
CAGLOD05940g YGR175C ERG1 Squalene epoxidase, catalyzes the epoxidation of squalene to -2.48 2,3-oxidosqualene; plays an essential role in the ergosterol- biosynthesis pathway and is the specific target of the antifun-gal drug terbinafine
CAGLOK06281g YDR454C GUK1 Guanylate kinase, converts GMP to GDP; required for growth and mannose outer chain elongation of cell wall N-linked gly-coproteins
-2.48
continued on next page

Appendix D. Chapter 3 Supplementary Data 257
continued from previous page CG ORF SC ORF Gene name Description WT.S - WT.0
CAGL0109900g YOR3I9W HSH49 U2-snRNP associated splicing factor with similarity to the mammalian splicing factor SAP49; proposed to function as a
-2.51
U2-snRNP assembly factor along with Hsh155p and binding partner Cus1p; contains two RNA recognition motifs (RRM)
CAGLOA01914g YHR099W TRAI Subunit of SAGA and NuA4 histone acetyltransferase corn- plexes; interacts with acidic activators (e.g., Gal4p) which leads to transcription activation; similar to human TRRAP, which is a cofactor for c-Myc mediated oncogenic transforma-tion
-2.58
CAGLOG00484g YGR272C - Description not mapped -2.59 CAGL0106809g YER142C MAGI 3-methyl-adenine DNA glycosylase involved in protecting DNA
against alkylating agents; initiates base excision repair by re-moving damaged bases to create abasic sites that are subse-quently repaired
-2.71
CAGLOK09416g YGL211W NCS6 Protein required for thiolation of the uridine at the wobble position of Gln, Lys, and Glu tRNAs; has a role in urmylation and in invasive and pseudohyphal growth; inhibits replication of Brome mosaic virus in S. cerevisiae
-2.81
CAGLOLO5500g YJL122W ALBI Shuttling pre-60S factor; involved in the biogenesis of riboso- mal large subunit; interacts directly with Arxlp; responsible for Tif6p recycling defects in absence of Reilp
-2.91
CAGL0000737g YLR222C UTP13 Nucleolar protein, component of the small subunit (SSU) pro- cessome containing the U3 snoRNA that is involved in process-ing of pre-18S rRNA
-2.91
CAGLOH09064g YHR128W PURI Uracil phosphoribosyltransferase, synthesizes UMP from uracil; involved in the pyrimidine salvage pathway
-2.97
CAGLOL10384g YOR064C YNG1 Subunit of the NuA3 histone acetyltransferase complex that acetylates histone H3; contains PHD finger domain that inter-acts with methylated histone I-13, has similarity to the human tumor suppressor ING1
-3.38
CAGLOH01089g YDR287W INM2 Inositol monophosphatase, involved in biosynthesis of inositol; enzymatic activity requires magnesium ions and is inhibited by lithium and sodium ions; inml inm2 double mutant lacks inositol auxotrophy
-3.41

Appendix D. Chapter 3 Supplementary Data 258
Table D.18: HOG1-dependent hyperosmotic stress repressed C. glabrata tran-
scripts. C. glabrata transcripts repressed in wild-type and stell cells, but not hogl cells following exposure to transient hyper-osmotic shock (1M sorbitol, 15 minutes). repressed transcripts are defined as having log2(Cy5) - log2(Cy3) < -2.0 and Benjamini-Hochberg corrected p-values < 0.05. Estimated log-ratios correspond to the WT.S - WT.0 in Table D.2
CG ORF SC ORF Gene Name Description WT.S - WT.0 STE.S - STE.0
CAGLOM09086g YJR092W BUD4 Protein involved in bud-site selec- tion and required for axial budding pattern; localizes with septins to bud neck in mitosis and may con-stitute an axial landmark for next round of budding; potential Cdc28p substrate
-2.16 -3.74
CAGLOE05324g YOR330C MIP1 Catalytic subunit of the mitochon- drial DNA polymerase; conserved
-2.24 -3.57
C-terminal segment is required for the maintenance of mitochondrial genome.
CAGLOH04301g YML078W CPB3 Mitochondrial peptidyl-prolyl cis- trans isomerase (cyclophilin), cat-alyzes the cis-trans isomerization of peptide bonds N-terminal to proline residues; involved in protein refold-ing after import into mitochondria
-2.4 -3.31
CAGLOG07953g YNR037C RSM19 Mitochondrial ribosomal protein of the small subunit, has similarity to
-2.53 -3.36
E. coli S19 ribosomal protein CAGLOK03795g YMR128W ECM/6 Essential DEAD-box ATP-
dependent RNA helicase specific to the 1J3 snoRNP, predominantly nucleolar in distribution, required for 18S rRNA synthesis
-2.63 -3.71
CAGLOG04411g YJL109C UTP/ 0 Nucleolar protein, component of the small subunit (SSU) processome containing the U3 snoRNA that is involved in processing of pre-18S rRNA
-2.7 -3.39
CAGL0106743g YER145C FTR1 High affinity iron permease involved in the transport of iron across the plasma membrane; forms complex with Fet3p; expression is regulated by iron
-2.74 -2.54

Appendix D. Chapter 3 Supplementary Data 259
Table D.19: STE11-dependent hyperosmotic stress repressed C. glabrata tran-
scripts. C. glabrata transcripts repressed in wild-type and hogl cells, but not stel 1 cells following exposure to transient hyper-osmotic shock (1M sorbitol, 15 minutes). repressed transcripts are defined as having log2(Cy5) - log2(Cy3) > 2.0 and Benjamini-Hochberg cor-rected p-values < 0.05. Estimated log-ratios correspond to the WT.S - WT.0 in Table D.2
CG ORF SC ORF Gene Name Description WT.S - WT.0 HOGS - HOG.0 CAGLOF01793g YLR056W ERG3 C-5 sterol desaturase, catalyzes the
introduction of a C-5(6) double bond into episterol, a precursor in ergosterol biosynthesis; mutants are viable, but cannot grow on non- fermentable carbon sources
-2.43 -2.73
CAGLOG00352g YGR277C - - -2.81 -3.46

Appendix D. Chapter 3 Supplementary Data 260
Table D.20: S. cerevisiae transcripts whose hyperosmotic stress dependent in-duction is affected by HOG1 deletion. Data obtained from Rep et al., (581)
CG ORF SC ORF Gene name Description CAGLOI09878g YOR317W FAA 1 Long chain fatty acyl-CoA synthetase with a preference for C12:0-C16,0
fatty acids; involved in the activation of imported fatty acids; localized to both lipid particles and mitochondrial outer membrane; essential for stationary phase
CAGLOG05984g YHR139C SPS100 Protein required for spore wall maturation; expressed during sporula-tion; may be a component of the spore wall
CAGLOL11374g YML070W DAK I Dihydroxyacetone kinase, required for detoxification of dihydroxyace-tone (DHA); involved in stress adaptation
CAGLOD06688g YOR374W ALD4 Mitochondrial aldehyde dehydrogenase, required for growth on ethanol and conversion of acetaldehyde to acetate; phosphorylated; activity is K+ dependent; utilizes NADP+ or NAD+ equally as coenzymes; ex-pression is glucose repressed
CAGLOG06028g YHR137W AR09 Aromatic aminotransferase II, catalyzes the first step of tryptophan, phenylalanine, and tyrosine catabolism
CAGLOB02563g YML128C MSC1 Protein of unknown function; mutant is defective in directing meiotic recombination events to homologous chromatids; the authentic, non-tagged protein is detected in highly purified mitochondria and is phos-phorylated
CAGLOE05280g YOL151W GRES 3-methylbutanal reductase and NADPH-dependent methylglyoxal re-ductase (D-lactaldehyde dehydrogenase); stress induced (osmotic, ionic, oxidative, heat shock and heavy metals); regulated by the HOG path-way
CAGLOK08844g YHL021C AIM17 Putative protein of unknown function; the authentic, non-tagged pro-tein is detected in highly purified mitochondria in high-throughput studies
CAGLOB00286g YCL064C CHA1 Catabolic L-serine (L-threonine) deaminase, catalyzes the degradation of both L-serine and L-threonine; required to use serine or threonine as the sole nitrogen source, transcriptionally induced by serine and threo-nine
CAGLOK12034g YDR040C ENA I P-type ATPase sodium pump, involved in Na-F and Li+ efflux to allow tolerance _salt
CAGLOJ01331g YMR090W - CAGLOG03883g YLL026W HSP/04 Heat shock protein that cooperates with Ydjlp (Hsp40) and Ssalp
(Hsp70) to refold and reactivate previously denatured, aggregated pro- teins; responsive to stresses including: heat, ethanol, and sodium ar- senite; involved in [PSI+, propagation
CAGLOE06380g YKL151C - - CAGLOI10516g YGR130C - - CAGLOG03773g YLL023C - - CAGLOJ04202g YFLO14W HSPI2 Plasma membrane localized protein that protects membranes from des-
iccation; induced by heat shock, oxidative stress, osmostress, station-ary phase entry, glucose depletion, oleate and alcohol; regulated by the HOG and Ras-Pka pathways
CAGLOG04433g YJL108C PRIV110 Pheromone-regulated protein, predicted to have 5 transmembrane seg-ments; induced by treatment with 8-methoxypsoralen and UVA irradi-ation
CAGLOH02101g YHR087W RTC3 Protein of unknown function involved in RNA metabolism; has struc-tural similarity to SBDS, the human protein mutated in Shwachman- Diamond Syndrome (the yeast SBDS ortholog = SDO1); null mutation suppresses cdc13-1 temperature sensitivity
CAGLOF04807g YIL136W 0M45 Protein of unknown function, major constituent of the mitochondrial outer membrane; located on the outer (cytosolic) face of the outer mem-brane
CAGLOM11660g YIL053W 1311132 Constitutively expressed isoform of DL-glycerol-3-phosphatase; in- volved in glycerol biosynthesis, induced in response to both anaerobic and, along with the Hor2p/Gpp2p isoform, osmotic stress
C AG LOI07865g YOL084W PHM7 Protein of unknown function, expression is regulated by phosphate lev-els; green fluorescent protein (GFP)-fusion protein localizes to the cell periphery and vacuole
continued on next page
Appendix D. Chapter 3 Supplementary Data 261
continued from previous page CG ORF SC ORF Gene Name Description
CAGLOK12958g YML131W - - CAGLOG05335g YDR074W TPS2 Phosphatase subunit of the trehalose-6-phosphate syn-
thase/phosphatase complex, which synthesizes the storage carbo- hydrate trehalose; expression is induced by stress conditions and repressed by the Ras-cAMP pathway
CAGLOF08085g YGR243W FMP43 Putative protein of unknown function; expression regulated by osmotic and alkaline stresses; the authentic, non-tagged protein is detected in highly purified mitochondria in high-throughput studies
CAGLOG02915g YGR086C DILI Primary component of eisosomes, which are large immobile cell cortex structures associated with endocytosis; null mutants show activation of Pkclp/Ypklp stress resistance pathways; detected in phosphorylated state in mitochondria
CAGLOF07579g YKL096W CWP1 Cell wall mannoprotein, linked to a beta-1,3- and beta-1,6-glucan het-eropolymer through a phosphodiester bond; involved in cell wall orga-nization
CAGLOL07656g YMLOO4C GL 0 1 Monomeric glyoxalase I, catalyzes the detoxification of methylglyoxal (a by-product of glycolysis) via condensation with glutathione to pro-duce S-D-lactoylglutathione; expression regulated by methylglyoxal lev-els and osmotic stress
CAGLOF07777g YMR170C AL D2 Cytoplasmic aldehyde dehydrogenase, involved in ethanol oxidation and beta-alanine biosynthesis; uses NAD+ as the preferred coenzyme; ex-pression is stress induced and glucose repressed; very similar to Ald3p
CAGLOI01122g YHR1O4W GRE3 Aldose reductase involved in methylglyoxal, d-xylose and arabinose metabolism; stress induced (osmotic, ionic, oxidative, heat shock, star-vation and heavy metals); regulated by the HOG pathway
Table D.21: Betweenness centrality measures and degree of tube sta- tions in the London Underground network London underground tube- map data sourced from http: //research. cs . queensu . ca/-daver/235/C1352963146/ E20070302133910/index.html were used to build a network of stations and edges be-tween stations. The degree of each station is the number of adjacent stations i.e. those that are one stop away. The betweenness centrality CB(v) is defined in section (3.3.6). High centrality scores indicate that a node can reach others on a relatively short path or that a node lies on a large proportion of shortest paths connecting other nodes. In the case of the London Underground tube-map, Baker Street is the most central node and King's Cross, although it has a high degree is ranked five places below Baker Street in terms of betweenness centrality.
Station Degree CB(v) Baker Street 7 0.34 Green Park 6 0.33
Bank 7 0.3 Waterloo 5 0.29 Stratford 5 0.22
Liverpool Street 6 0.19 King's Cross St Pancras 7 0.19
Turnham Green 5 0.11 Oxford Circus 6 0.1
Shadwell 5 0.06

(b)
(c)
YEPD + 1 M SORBITOL CGTFKOLIB Plate 1
Appendix D. Chapter 3 Supplementary Data 262
062001 C „2,03B1 PO81 14161 H052 GAL83 SASS 5EFI RTG2 KARA TO54
81103 11102 MEW NAP1 Han RFXI EIL I LTA'S CUP. GLIY3 MI53 POP2
TOPS- like (Rot RTG1 HST1 45111
ATO)
SFP1-
(not A1.01 SSAAS 11051 11051 PP131 BRE2 31104 46 00?
TUPI 5000 HGGI SWIS SBA HIRI SHF6 8I14101 STP2 AHC1 ADRI
TYE7 6AL11 YAP] SITS ARCOT 5131 CADJ 11574 TSPI MCI 1
0.4081 51134 hISSI 1 0021 GCHS SWDl X501 45610 FLOE G273 5362
IIII• P11023 YOX1 GIS 1 POGI A5G1 FX211 AFT2 TOSS SP01 WARS #101
%V- 1032 . ARGR1 FIFE RTF1 HAP2 0051 0533 11102 STBS COPY
(a)
YEPD CGTFKOLib Plate 1
• 4111•• 4116••••• •••••••• • 6 II 41 41 41 • 41 41 44 41 41 40 41 ••
4114
• 40 ED • 0 • • ',10 • • • • • • IA •••••••• • •••• • 111 411111•• 4Il • • 11 • • fir 41 • • • 410 11104141•64.0•41 0.41
Figure D.1: Solid agar phenotype assay screen of CGTFKO1ib plate 1 on hyper-tonic agar (1 M Sorbitol). Frozen glycerol stocks of plate 1 of the CGTFKOLib arrayed in 96 well format (a) were defrosted by a Bunsen flame, subcultured (18 hours, 37°C, 180 rpm), washed twice in distilled water (3000 rpm, 5 minutes) and replicated (104 cells) onto solid YEPD agar or solid YEPD agar + 1 M sorbitol and incubated for 24 hours. Feint red squares highlight mutant strains relevant to hyperosmotic stress and/or SAPk pathways. Experiments performed by Lauren Ames

•••••••••fie ••••••••••••
•••••••••111 ••••••••••• +60 • • • • • ••• • •
• • • • • * 411 zi • • • • 11111 • • • iik • 81 41 • 1 0 41 41 0 • ••
(b)
YEPD + 1 M SORBITOL CGTFKOLIB Plate 2
* II • 0 • 114\ 041000111/00 411•••••• • • • *GOO* • • • ••••• o• • • • ••• ►•••••••• 410•••
(c)
Appendix D. Chapter 3 Supplementary Data 263
(111001 C20C" SUT1 CST6 5111 MO 15115 MCMI 1-041 Han 1 1013
ZAP1 HOU III PIP2 GISI San 11142 MOSS
10,
5502 WTI SAU 1 RINI1
5614 EMl1 4.61- like
ADA2 STP1 5514-11e GAT1 1464 METH YAPS MI2.2 NAPS
SUM LY514 HOU 0013 WTI SAPS OW StS3 auto SiP0 2211' YHPI LIVE
YAM AS11 GT51 TIM MCI 44312 14.401 .61 11111 PUTS sprz CHOI
*PHI SSIO 0501 IOW IIIASET31 if AIM SPT3 UNIZI SPP1
3052 TA114 CAR SRL DAM ACF2 SPAM 5053 CALI SSW FAPI SON/
0464 MEI Rira VAPilikr 0444
(a)
YEPD CGTFKOLib Plate 2
Figure D.2: Solid agar phenotype assay screen of CGTFKOlib plate 2 on hyper-tonic agar (1 M Sorbitol). Frozen glycerol stocks of plate 1 of the CGTFKOLib arrayed in 96 well format (a) were defrosted by a Bunsen flame, subcultured (18 hours, 37°C, 180 rpm), washed twice in distilled water (3000 rpm, 5 minutes) and replicated (104 cells) onto solid YEPD agar or solid YEPD agar + 1 M sorbitol and incubated for 24 hours. Feint red squares highlight mutant strains relevant to hyperosmotic stress and/or SAPk pathways. Experiments performed by Lauren Ames

ver.812C Yogi SW
YGFI133W Y13191,03W
YEN75C o 0 8 c o
0 o 0 o
° SO 43-V-43
YLL09W Y011906C
YPRgI36W
__1'0.55.11.R22 S ..................... _
i i i 1000 2000 3000
arbitrary gene Index
4000 500 1 000 1500 0000 2500 3000
arbitrary gene index
012
Appendix D. Chapter 3 Supplementary Data 264
z
S. bayanus dblidS
YILV4W
YILI,F3C 0
;PAW/
vp111,55c ° o
° o
-7-
S. mikatae dblidS
YP147W
VIL1.;4W
V.1,4,70W
Velr_ig51W rr
1000 2000 3000 40011 1000
2000 3000 4000
arbitrary gene Index arbitrary gene Index
(a)
(b)
S. paradoxus dNIdS C. glabrata dNIdS
(c)
(d)
Figure D.3: Calculated dN/dS > rates for Saccharomyces bayanus (A), Sac-charomyces mikatae (B), Saccharomyces paradoxus (C) and C. glabrata (D). Genome sequences of Saccharomyces paradoxus, Saccharomyces bayanus, Saccharomyces mikatae and C. glabrata were obtained along with pre-computed orthologue lists and used to construct a table of ORFs that are conserved between S. cerevisiae and the other Sac-charomycotina. In order to calculate the ratio of non-synonymous substitution rate to synonymous substitution rate (dN/dS), nucleic acid sequences were first translated to their amino acid sequences using translfas (147; 148) and the default universal codon tables. This step is performed to preserve the alignment of codons and permits the use of substitution matrices, such as those described in, for the estimation of evolutionary rates. Multiple align-ment of conserved Saccharomycotina proteins was performed using clustalw (723; 689) and putgaps (147; 148) was used to create a gapped, in-frame codon alignment of nucleic acids for subsequent estimation of dN/dS using the Creevey-McInerney method. The average C. glabrata dN/dS (0.32) is significantly greater than dN/dS for S. paradoxus (mean = 0.15, p < 0.05, t = 31.54, df = 4355), S. bayanus (mean = 0.15, p < 0.05, t = 32.00, df = 4105) and S. mikatae (mean = 0.16, p < 0.05, t = 30.00, df = 4581). A larger proportion of C. glabrata orthologue have dN/dS greater than 1. The dN/dS > 1 threshold is depicted by a horizontal red, dashed line at dN/dS = 1

Appendix D. Chapter 3 Supplementary Data 265
Table D.22: C. glabrata ORFs with dN/dS > 1. Genome sequences of Saccharomyces paradoxus, Saccharomyces bayanus, Saccharomyces mikatae and C. glabrata were obtained along with pre-computed orthologue lists and used to construct a table of ORFs that are conserved between S. cerevisiae and the other Saccharomycotina. In order to calculate the ratio of non-synonymous substitution rate to synonymous substitution rate (dN dS), nucleic acid sequences were first translated to their amino acid sequences using translfas (147; 148) and the default universal codon tables. This step is performed to preserve the alignment of codons and permits the use of substitution matrices, such as those described in, for the estimation of evolutionary rates. Multiple alignment of conserved Saccharomycotina proteins was performed using clustalw (723; 689) and putgaps (147; 148) was used to create a gapped, in-frame codon alignment of nucleic acids for subsequent estimation of dN/dS using the Creevey--McInerney method. The average C. glabrata dNI dS (0.32) is significantly greater than dN/dS for S. paradoxus (mean = 0.15, p < 0.05, t = 31.54, df = 4355), S. bayanus (mean = 0.15, p < 0.05, t = 32.00, df = 4105) and S. mikatae (mean = 0.16, p < 0.05, t = 30.00, df = 4581). A larger proportion of C. glabrata orthologue have dN dS greater than 1.
homologous SC ORF Gene Name Description YALOO5C SSA1 ATPase involved in protein fold-
ing and nuclear localization signal (NLS)-directed nuclear transport; member of heat shock protein 70 (HSP70) family; forms a chaper- one complex with Ydjlp; localized to the nucleus, cytoplasm, and cell wall
YAL047C SPC7.2 Component of the cytoplasmic Tub4p (gamma-tubulin) complex, binds spindle pole bodies and links them to microtubules; has roles in astral microtubule formation and stabilization
YBR127C VMA2 Subunit B of the eight-subunit Y1 peripheral membrane domain of the vacuolar H-1-ATPase (V-ATPase), an electrogenic proton pump found throughout the endomembrane sys- tem; contains nucleotide binding sites; also detected in the cytoplasm
YCL036W GFD2 Protein of unknown function, iden-tified as a high-copy suppressor of a dbp5 mutation
YCR024C SLM5 Mitochondrial asparaginyl-tRNA synthetase
YCR087C-A Putative protein of unknown func- tion; green fluorescent protein (GFP)-fusion protein localizes to the nucleolus; YCR087C-A is not an essential gene
continued on next page

Appendix D. Chapter 3 Supplementary Data 266
continued from previous page homologous SC ORF Gene Name Description
YDL155W
YDL237W
YDR071C
YDR075W
YDR103W
YDR137W
YDR168W
YDR350C
YDR452W
YDR463W
CLB3
AIMS
PA A 1
PPH3
STE5
RGP1
CD C37
ATP22
PPN1
STPI
B-type cyclin involved in cell cy-cle progression; activates Cdc28p to promote the G2/M transition; may be involved in DNA replication and spindle assembly; accumulates dur-ing S phase and G2, then targeted for ubiquitin-mediated degradation Putative protein of unknown function, required for respiratory growth; YDL237W is not an essential gene Polyamine acetyltransferase; acety- lates polyamines (e.g. putrescine, spermidine, spermine) and also aralkylamines (e.g. tryptamine, phenylethylamine); may be in- volved in transcription and/or DNA replication Catalytic subunit of an evolution-arily conserved protein phosphatase complex containing Psy2p and the regulatory subunit Psy4p; required for cisplatin resistance; involved in activation of GIn3p Pheromone-response scaffold pro- tein; binds Ste11p, Ste7p, and Fus3p kinases, forming a MAPK cascade complex that interacts with the plasma membrane and Ste4p-Stel8p; allosteric activator of Fus3p that facilitates Ste7p-mediated ac-tivation Subunit of a Golgi membrane ex-change factor (Riclp-Rgplp) that catalyzes nucleotide exchange on Ypt6p Essential HspOOp co-chaperone; necessary for passage through the START phase of the cell cycle; stabilizes protein kinase nascent chains and participates along with IfspOOp in their folding Mitochondrial inner membrane pro-tein required for assembly of the FO sector of mitochondria' FIFO ATP synthase, which is a large, evolu-tionarily conserved enzyme complex required for ATP synthesis Vacuolar endopolyphosphatase with a role in phosphate metabolism; functions as a ho- modimer Transcription factor, undergoes proteolytic processing by SPS (Ssylp-Ptr3p-Ssy5p)-sensor com- ponent Ssy5p in response to extracellular amino acids; activates transcription of amino acid perme-ase genes and may have a role in tRNA processing
continued on next page

Appendix D. Chapter 3 Supplementary Data 267
continued from previous page homologous SC ORF Gene Name Description
YER058W
YER168C
YER175C
YFRO12W
YFRO36W
YGL110C
YGL112C
YGL153W
YGRO13W
YGRO78C
YGR133W
YGR199W
PETI17
CCAI
TMT1
CDC26
CUES
TAPS
PEXI4
SNU7I
PACIO
PEX4
PMTG
Protein required for assembly of cy-tochrome c oxidase ATP (CTP):tRNA-specific tRNA nucleotidyltransferase; different forms targeted to the nucleus, cytosol, and mitochondrion are generated via the use of multiple transcriptional and translational start sites Trans-aconitate methyltransferase, cytosolic enzyme that catalyzes the methyl esterification of 3- isopropylmalate, an intermediate of the leucine biosynthetic pathway, and trans-aconitate, which inhibits the citric acid cycle Putative protein of unknown func-tion Subunit of the Anaphase- Promoting Complex/Cyclosome (APC/C), which is a ubiquitin- protein ligase required for degra- dation of anaphase inhibitors, including mitotic cyclins, during the metaphase/anaphase transition Protein of unknown function; has a CUE domain that binds ubiquitin, which may facilitate intramolecular monoubiquitination Subunit (60 kDa) of TFIID and SAGA complexes, involved in tran-scription initiation of RNA poly-merase II and in chromatin modi-fication, similar to histone H4 Peroxisomal membrane peroxin that is a central component of the peroxisomal protein import machinery; interacts with both PTS1 (Pex5p) and PTS2 (Pex7p), peroxisomal matrix protein signal recognition factors and membrane receptor Pexl3p Component of Ul snRNP required for mRNA splicing via spliceosome; yeast specific, no metazoan counter-part Part of the heteromeric co- chaperone GimC/prefoldin com- plex, which promotes efficient protein folding Peroxisomal ubiquitin conjugating enzyme required for peroxisomal matrix protein import and peroxi-some biogenesis Protein 0-mannosyltransferase, transfers mannose from dolichyl phosphate-D-mannose to protein serine/threonine residues of secre- tory proteins; reaction is essential for cell wall rigidity; member of a family of mannosyltransferases
continued on next page

Appendix D. Chapter 3 Supplementary Data 268
continued from previous page homologous SC ORF Gene Name Description
YHROO5C
YHR032W
YIL018W
YIL044C
YIL066C
YIL079C
YIR031C
YJL110C
YJL176C
GPA 1
RPL2B
AGE2
RNR3
A IR I
DA L7
GZ.F3
SW13
GTP-binding alpha subunit of the heterotrimeric G protein that cou-ples to pheromone receptors; nega-tively regulates the mating pathway by sequestering G(beta)gamma and by triggering an adaptive response; activates Vps34p at the endosome Putative protein of unknown func- tion; putative substrate of the cAMP-dependent protein kinase (PKA) Protein component of the large (60S) ribosomal subunit, identical to Rpl2Ap and has similarity to E. coli L2 and rat L8 ribosomal pro-teins; expression is upregulated at low temperatures ADP-ribosylation factor (ARF) GTPase activating protein (GAP) effector, involved in Trans-Golgi- Network (TGN) transport; contains C2C2142 cysteine/histidine motif One of two large regulatory sub-units of ribonucleotide-diphosphate reductase; the RNR complex cat-alyzes rate-limiting step in dNTP synthesis, regulated by DNA repli-cation and DNA damage checkpoint pathways via localization of small subunits Zinc knuckle protein, involved in nuclear RNA processing and degra- dation as a component of the TRAMP complex; stimulates the poly(A) polymerase activity of Pap2p in vitro; functionally redun-dant with Air2p Malate synthase, role in allan- toin degradation unknown; expres-sion sensitive to nitrogen catabo-lite repression and induced by allo-phanate, an intermediate in allan-toin degradation GATA zinc finger protein and Da180p homolog that negatively regulates nitrogen catabolic gene expression by competing with Gat1p for GATA site binding; func- tion requires a repressive carbon source; dimerizes with Da180p and binds to Tor1p Subunit of the SWI/SNF chromatin remodeling complex, which regu- lates transcription by remodeling chromosomes; required for tran- scription of many genes, including ADH1, ADH2, GALI, HO, INC)]. and SUC2
continued on next page
Appendix D. Chapter 3 Supplementary Data 269
continued from previous page homologous SC ORF Gene Name Description
YJR006W
YJR136C
YKL201C
YKR039W
YKR055W
YLL029W
YLL036C
YLL043W
YLRO09W
YLR369W
YLR438C-A
POL31
TTI2
MNN4
GAP1
RHO4
FRA I
PRP19
FPS1
RLP24
SSQI
LSM7
DNA polymerase III (delta) sub-unit, essential for cell viability; in-volved in DNA replication and DNA repair Putative protein of unknown func-tion; subunit of the ASTRA com-plex which is part of the chromatin remodeling machinery; similar to S. pombe Tti2p; may interact with Rsm23p; GFP-fusion protein local-izes to the cytoplasm Putative positive regulator of mannosylphosphate transferase (Mnn6p), involved in manno- sylphosphorylation of N-linked oligosaccharides; expression in- creases in late-logarithmic and stationary growth phases General amino acid permease; lo-calization to the plasma membrane is regulated by nitrogen source Non-essential small GTPase of the Rho/Rac subfamily of Ras-like pro-teins, likely to be involved in the es-tablishment of cell polarity Protein involved in negative regula-tion of transcription of iron regulon; forms an iron independent complex with Fra2p, Grx3p, and Grx4p; cy-tosolic; mutant fails to repress tran-scription of iron regulon and is de-fective in spore formation Splicing factor associated with the spliceosome; contains a U-box, a motif found in a class of ubiquitin ligases Plasma membrane channel, mem-ber of major intrinsic protein (MIP) family; involved in efflux of glyc- erol and in uptake of acetic acid and the trivalent metalloids arsen-ite and antimonite; phosphorylated by Hoglp MAPK under acetate stress Essential protein with similarity to Rp124Ap and Rp12413p, associated with pre-60S ribosomal subunits and required for ribosomal large subunit biogenesis Mitochondrial hsp70-type molecu-lar chaperone, required for assem-bly of iron/sulfur clusters into pro-teins at a step after cluster synthe-sis, and for maturation of Yfhlp, which is a homolog of human frataxin implicated in Friedreich's ataxia Lsm (Like Sm) protein; part of het-eroheptameric complexes (Lsm2p-7p and either Lsmlp or Sp): cyto-plasmic Lsmlp complex involved in mRNA decay; nuclear Lsm8p com-plex part of U6 snRNP and pos-sibly involved in processing tRNA, snoRNA, and rRNA
continued on next page

Appendix D. Chapter 3 Supplementary Data 270
continued from previous page homologous SC ORF Gene Name Description
YLR449W
YML041C
YML120C
YMR291W
YNL099C
YNR003C
YNR004W
YOL022C
YOR006C
YOR104W
YORHOW
YOR160W
FP./1.4
VPS71
ND//
OCA I
RPC34
PIN2
TFC7
A I TR 10
Peptidyl-prolyl cis-trans isomerase (PPIase) (proline isomerase) local-ized to the nucleus; catalyzes iso-merization of proline residues in hi-stones H3 and H4, which affects ly-sine methylation of those histones Nucleosome-binding component of the SWR1 complex, which exchanges histone variant H2AZ (Htzlp) for chromatin-bound hi- stone H2A; required for vacuolar protein sorting NADHaMiquinone oxidoreductase, transfers electrons from NADH to ubiquinone in the respiratory chain but does not pump protons, in contrast to the higher eukaryotic multisubunit respiratory complex I; phosphorylated; homolog of human AMID Putative kinase of unknown func- tion; green fluorescent protein (GFP)-fusion protein localizes to the cytoplasm and nucleus; YMR291W is not an essential gene Putative protein tyrosine phos- phatase, required for cell cycle ar-rest in response to oxidative dam-age of DNA RNA polymerase III subunit C34; interacts with TFIIIB70 and is a key determinant in pol III recruit-ment by the preinitiation complex Putative protein of unknown func- tion; haploid disruptant exhibits cold-sensitive growth and elongated buds Cytoplasmic protein of unknown function; essential gene in S288C background, while tsr4 null muta-tions in a CEN.PK2 background confer a reduced growth rate Putative protein of unknown func- tion; green fluorescent protein (GFP)-fusion protein localizes to both the cytoplasm and the nucleus Protein that induces appearance of [PIN-I-] prion when overproduced; predicted to be palmitoylated One of six subunits of the RNA polymerase III transcription initia-tion factor complex (TFIIIC); part of the TauA globular domain of TFIIIC that binds DNA at the BoxA promoter sites of tRNA and similar genes Nuclear import receptor, mediates the nuclear localization of proteins involved in mRNA-nucleus export; promotes dissociation of mRNAs from the nucleus-cytoplasm mRNA shuttling protein Npl3p
continued on next page

Appendix D. Chapter 3 Supplementary Data 271
continued from previous page homologous SC ORF Gene Name Description
YOR247W SRL1 Mannoprotein that exhibits a tight association with the cell wall, required for cell wall stability in the absence of GPI-anchored mannoproteins; has a high serine-threonine content; expression is in-duced in cell wall mutants

Appendix D. Chapter 3 Supplementary Data 272
Table D.23: Profile of induced transcripts conserved between S. cerevisiae sten (O'Rourke & Herskowitz 0.5M KC1, 10 minutes) and C. glabrata after exposure to hyperosmotic stress. Key to abbreviations: CG= Candida glabrata open reading frame
ID; SC=Saccharomyces cerevisiae open reading frame ID. All descriptions are inferred by
sequence homology between C. glabrata and S. cerevisiae open reading frames. S. cerevisiae data were obtained from a study by O'Rourke & Herskowitz (518) in which S. cerevisiae cells were treated with 0.5M KC1.
CC ORF SC ORF Gene name Description WT V1 V2 V3 V4 V5
CAGLOI10010g YGR142W BTN2 v-SNARE binding protein that facilitates specific protein re- trieval from a late endosome to the Golgi; modulates arginine uptake, possible role in mediating pH homeostasis between the vacuole and plasma membrane H(+)-ATPase
5.55
CAGL0CO2321g YER037W PHM8 Protein of unknown function, expression is induced by low phosphate levels and by inactivation of Pho85p
5.41
CAGLOG04433g YJL108C PRM10 Pheromone-regulated protein, predicted to have 5 transmem- brave segments; induced by treatment with 8-methoxypsoralen and UVA irradiation
5.14
CAGLOM06963g YNR034W SOLI Protein with a possible role in tRNA export; shows similar- ity to 6-phosphogluconolactonase non-catalytic domains but does not exhibit this enzymatic activity; homologous to Sol2p, Sol3p, and Sol4p
4.31
CAGLOM08822g YDR258C HSP78 Oligomeric mitochondrial matrix chaperone that cooperates with Ssclp in mitochondrial thermotolerance after heat shock; able to prevent the aggregation of misfolded proteins as well as resolubilize protein aggregates
3.93
CAGLOE00803g YDR171W HSP42 Small heat shock protein (sHSP) with chaperone activity; forms barrel-shaped oligomers that suppress unfolded protein aggregation; involved in cytoskeleton reorganization after heat shock
3.82
CAGLOG03883g YLL026W HSP/04 Heat shock protein that cooperates with Ydjlp (Hsp40) and 3.61 Ssalp (Hsp70) to refold and reactivate previously denatured, aggregated proteins; responsive to stresses including: heat, ethanol, and sodium arsenite; involved in [PSI-H propagation
CAGLOF08745g YLR327C TMAIO Protein of unknown function that associates with ribosomes 3.53 CAGLOH08173g YOR028C CIN5 Basic leucine zipper (bZIP) transcription factor of the yAP-1
family, mediates pleiotropic drug resistance and salt tolerance; nuclearly localized under oxidative stress and sequestered in the cytoplasm by Lot6p under reducing conditions
3.49
CAGLOG01540g YNL036W NCE103 Carbonic anhydrase; poorly transcribed under aerobic condi- tions and at an undetectable level under anaerobic conditions; involved in non-classical protein export pathway
3.47
CAGLOM12474g YIL055C - Description not mapped 3.46 CAGLOH08844g YMR173W DDR48 DNA damage-responsive protein, expression is increased in re-
sponse to heat-shock stress or treatments that produce DNA lesions; contains multiple repeats of the amino acid sequence
3.06
NNNDSYGS CAGLOE05632g YOR348C PUT4 Proline permease, required for high-affinity transport of pro-
line; also transports the toxic proline analog azetidine- 2.85
2-carboxylate (AzC); PUT4 transcription is repressed in ammonia-grown cells
CAGLOF09163g YER182W FM -PIO Putative protein of unknown function; the authentic, non- tagged protein is detected in highly purified mitochondria in high-throughput studies
2.74
CAGLOL07480g YBRO66C NRG2 Transcriptional repressor that mediates glucose repression and negatively regulates filamentous growth; has similarity to
2.64
Nrglp CAGLOL00803g YER054C GIP2 Putative regulatory subunit of the protein phosphatase Glc7p,
involved in glycogen metabolism; contains a conserved motif 2.62
(GVNK motif) that is also found in Gac1p, Pig1p, and Pig2p continued on next page

Appendix D. Chapter 3 Supplementary Data 273
continued from previous page CG ORF SC ORF Gene Name Description WT.S - WT.0
CAGLOK12034g YDR040C ENA1 P-type ATPase sodium pump, involved in Na+ and Li+ efflux to allow salt tolerance
2.61
CAGLOI01100g YOR12OW GCYI Putative NADP(+) coupled glycerol dehydrogenase, pro- posed to be involved in an alternative pathway for glycerol catabolism; member of the aldo-keto reductase (AKR) family
2.56
CAGLOH05137g YPLO61W ALD6 Cytosolic aldehyde dehydrogenase, activated by Mg2+ and uti- lizes NADP+ as the preferred coenzyme; required for conver-sion of acetaldehyde to acetate; constitutively expressed; lo-cates to the mitochondrial outer surface upon oxidative stress
2.53
CAGLOM06347g YBR183W YPC/ Alkaline ceramidase that also has reverse (CoA-independent) ceramide synthase activity, catalyzes both breakdown and syn-thesis of phytoceramide; overexpression confers fumonisin B1 resistance
2.52
CAGLOJ04466g YLR414C - - 2.5 CAGLOL10186g YOR052C - Description not mapped 2.45 CAGLOB06424g YKL150W - Mitochondrial NADH-cytochrome b5 reductase, involved in er-
gosterol biosynthesis 2.42
CAGLOJ04158g YOR22OW RCN2 Protein of unknown function; green fluorescent protein (GFP)- fusion protein localizes to the cytoplasm and is induced in re-sponse to the DNA-damaging agent MMS; phosphorylated in response to alpha factor
2.35
CAGLOI04554g YBRO14C GRX7 Cis-golgi localized monothiol glutaredoxin; more similar in ac- tivity to dithiol than other monothiol glutaredoxins; involved in the oxidative stress response; does not bind metal ions; func-tional overlap with GRX6
2.34
CAGLOM06545g YKL124W SSH4 Specificity factor required for Rsp5p-dependent ubiquitination and sorting of cargo proteins at the multivesicular body; iden-tified as a high-copy suppressor of a SHR3 deletion, increasing steady-state levels of amino acid permeases
2.24
CAGLOL10912g YOR273C TPO4 Polyamine transport protein, recognizes spermine, putrescine, and spermidine; localizes to the plasma membrane; member of the major facilitator superfamily
2.12
CAGLOFO1111g YOL032W 01'110 Protein with a possible role in phospholipid biosynthesis, based on inositol-excreting phenotype of the null mutant and its sup-pression by exogenous choline
2.04

274
Appendix E
Chapter 4 Supplementary Data

411101•1•11100., Atu203 t130 aRNA2 CCrIl A1u293 IC a A
03- (Afu29330 - Afu293.t60) + totRNA
Appendix E. Chapter 4 Supplementary Data 275
31- (Aftt293.t0 - Afu293.t60) + ARNA1
c..ortrast, 32 - (Afu293.t0 - Afu293.t60) aRNA2
co,trast2
contrast,- aRNA2 aRNA1
contrast2- totRNA - aRNA2
contrast3- totRNA - aRNA1
Figure E.1: Schematic representation of hybridisations performed in the ampli-fication control study. Boxes represent the tissue from which the RNA was isolated and used as a template for reverse transcription and concomitant fluorophore labelling. The open and shaded arrows demonstrate that samples were dye-swapped to account for preferential fluorophore labelling in some transcripts. In all cases, labelled cDNA from A. fumigatus strain 237 at time = 0 minutes (Afu237.To ) was co-hybridised with equivalent material derived from the RNA isolated from A. fumigatus strain 237 at time = 60 minutes (Afu237.t60). The top row of boxes shows co-hybridisation of cDNA derived from RNA sam-ples that had been diluted and subjected to 1 round of mRNA amplification prior to reverse transcription and fluorophore labelling. The second row of boxes shows co-hybridisation of samples that had been diluted and subjected to 2 rounds of mRNA amplification, and the third row of boxes shows co-hybridisation of samples that did not undergo mRNA amplifi-cation prior to reverse transcription. We investigated the relative abundance of transcripts at To minutes and T60 minutes, and used a linear regression framework to identify am-plification protocols that provided evidence for differences in those relative abundances, flagging those genes that showed significant difference in relative abundance after linear amplification. The treatment effects (i3i , /32 and 33) and contrasts (contrasts , contrast2 and contrast3 ) estimated in the linear model are also depicted.

Appendix E. Chapter 4 Supplementary Data 276
-3.0
0.0
3.0
4 t1 C
U 0
4 4 m htu2/13110: cytochresee c kfu4/01110: 14-alpha sterol deeethylthe C1)151 "hfu4r0/4110, coproporphythnopes III oridase, putative" 'hfu4q15140: nitochondrial phosphate carrier protein (Kiel), MO/0740 li•olpha sterol &Aoki'''. Cip5111 Ifu11105600: conserved hypothetical protein kfuSgOt230: citrate synthase CitA "Aft101120: fatty acid hydroxylue, putative" 1E0603400' eor.served hnethetieal protein "Afulr110/0 pyruvite decarboxylase Pdck, putative" 'Mu400100 :hoe! chain dehydrogenase Oast), putative" "AN201100: wane acid manse (Dip , putative" Afudg10720: alpha-ketollutazate-dependent twine &ionic's.: ifuSg13060: 001275 dada protein "Afu1g02310: actin cortical patch, protein. Sue, putative hfuSg01220, conserved hypothetical protein Aing142$0 flavia-hnding aoloorypenne-like protein "Afu3g0940: 2,3-diketo-S•aethylthio-1-phosphopentone phosphatase, putat: IftelgOOda cell surface pnteir., putative" hfulq16440 hypothetical protein Afulg01690 conserved hypothetical protein "If:409110. suralefferredosin•like (oily protein Apdl, putative" kfu1g00350• conserved hypothetical protein "Afu301010: pyravate decirboxylase Pdch, ?native "Afulg064/0 . hest :hock protein !I:pH-like, putative "Aft:40116D: tartrate dehydrogenate, putative' Afu3g10310 conserved hypothetical protein "AfulgISPSO Elwin-binding aotooxygerase, putative "Aftag09330 tipe-like antibiotic response protein, putative AftrtgOPIPO S-netosyliaetbiorine-sterol-C- sethyltreateferae 'Aft:401140. VS Raltidrag tresporter, putative' AfuSg01820: estratellulas ?reline-rich protein 'Aft13,09890: extractllalu thmattin Wain protein, putative' 1(001960: VI anchored cell rill protein, putative "1:n902700: KV antidrug transporter, putative' "X614900240 . eine-conturing alcohol &hydrogens, putative' Afir4q13510 isocitrate irate AtuD hfu4g00150 conserved hypothetical protein 'A6502100 VS aultietral transporter, putative' Afu1g13310 isocitrite lyase kcu "Afseg01000. potusiwattivated aldehyde dehydrogenase 1114 putative" hfaSg011130 conserved hypothetical protein `Afu3g09110 sucruelierredorin-like fussily protein Apt11, putative' "UuSg01920 GPI anchored protein, putative' Afulg05100. "AfuSg00710 ZIA pessease, putative "kfulg0100 isomayl alcohol oxidant, putative' 'Aft:204200 4.hydeorypher.ylpyrovate dioxygenne, putative "Ain/01240 tint-containing alcohol dedydrogenue, putative' "Aft151012$0 ondontactase, putative' Afu410190 arkyria repeat protein "kfu3I00110 short eltair, dekdraprese rktsel , putative' Afu3g11680 lysophospholipue Pill kfu6111190. conserved hypothetical peoteta A10;0161 bifunctional eatalase-perexidue Catl 'AfuSg03330 ktyph-like protein, putative' 11003010 phosphite-ripreszille ho/phosphate cotransporter Phe$9, 'hfulq46020 skra folly transcriptional regulator, putative 'At p21313 ferric-chelate eeductase Int), putative 1E15100120 ZAI folly acetyltronsferase, putative Afull14550 Hz superoxide dismutose
Figure E.2: Comparative analysis of A. fumigates gene expression datasets. A pan-experimental comparison of A. furnigatus gene expression comparing log2 ratios ob-tained during host adaptation (mice); exposure to neutrophils(neut), increased expression in parental strain versus AlaeA mutant, acid shift (acid), iron starvation (iron), oxygen depletion (anaer) and oxidative stress (H202) for various genes. The colour bar indi-cates the range of loge expression ratios, grey bars indicate genes from which signals were undetectable. Experimental conditions are described in the materials and methods sec-tion. LaeA dataset take from (539). Comparative analysis performed with TM4 software http://www.jcvi.org/cms/research/software/.

Appendix E. Chapter 4 Supplementary Data 277
10.00
5.00
0.00
-5.00
-10.00
-15.00
..-Liti Lij 1 I
Figure E.3: Concordance between estimates of relative abundance from triplicate qPCR measurements (yellow) and microarray experiments (grey) for selected A. fumigates ORFs. Oligo sequences are given in Table E.12

6 ' 8
ci : 2 I0o C
l',_
`4'
Iiiiiiiiiiillill
O
a -
1—
Appendix E. Chapter 4 Supplementary Data 278
Foreground Intensity Boxplots of Cy5 Channel Foreground Intensity Boxplots of Cy3 Channel
1 2
3 4 5 8 8 4 5 7 8
Slide tilde
(a)
(b)
Background Intensity Boxplols of Cy5 Channel Background Intensity Boxplots of Cy3 Channel
8 _ -
8
a a a
a 82 3 a I I
4441-1-11.1: 8
O -
1 2 3 4 5 6 7 8
Side
(C)
I III I
1 2 3 4 5 5 7
Sbde
(d)
Figure E.4: Box and whisker plots of Cy5TM and Cyr intensities for slides hy-bridized with amplified and non-amplified RNA. Foreground intensities are shown for Cy5rm and Cye in a) and b). Background intensities are shown for Cy5TM and Cy3
TM
in c) and d). Slides 1-6 (blue) represent slides that were hybridized with cDNA derived from amplified RNA. Slides 7-8 (orange) represent slides that were hybridized with cDNA derived from unamplified RNA.

Appendix E. Chapter 4 Supplementary Data 279
a) To
a
difference
(a)
aRNA 2 v totRNA
R. 0.75
log2
ratio
totR
NA
a?
-5
5
10
log2 ratio aRNA 2
(b)
Figure E.5: Genes rejected in the amplification control experiment transposed onto correlation plots. Plot of the difference in log2 ratio between the amplified (aRNA2) and the non-amplified material (totRNA) for the loge ratio for the two samples vs. the p-value for each of the retained genes a). Correlation of log2 ratios between non-amplified and amplified material. The genes in the "conservative set" are coloured gold, the genes in the "rejected set" are coloured black and the genes in the "undetermined set" are coloured grey b). Sets are defined in the materials and methods and follow criteria originally described in (509).

Appendix E. Chapter 4 Supplementary Data 280
Table E.1: RNA Samples and associated amplification factors. Key to abbrevia-
tions: ID = sample identifier; tRNA = total RNA, mRNA = messenger RNA; aRNAi = RNA after 1 round of amplification; aRNA2 = RNA after 2 rounds of amplification; AF = amplification factor.
ID Strain tRNA ng mRNA ng aRNA1 mg aRNA2 tig AF To Overnight broth
culture 1000 20 25.95 184.76 9.2 X103
T60 Overnight broth plus media shift for 60 minutes
1000 20 87.33 174.60 8.7 X 103
A (in vivo)1 323 13 11.08 251.14 2.0 x 104 B (in vivo)2 130 3 6.25 164.02 6.3 X 104 C (in vivo)3 240 10 9.02 172.63 1.8 x 104 D (in vivo)4 800 64 19.27 177.11 2.8 x 103 E (in vivo)5 110 2 12.27 258.39 1.2 X 105 F (in vitro) 1136 23 3.72 186.96 8.2 X 103 G (in vitro) 1000 20 56.00 566.00 2.8 X 104

Appendix E. Chapter 4 Supplementary Data 281
Table E.2: Filtering per array in the amplification study. Filtering criterion A refers
to spots flagged by the TIGR spotfinder software. Filtering criterion B excludes spots whose
foreground intensity is less than twice its background intensity in either the Cy5Th or Cy3TM
channel
Array Number 1 2 3 4 5 6 Criterion A 1895 2080 1867 3630 8079 7151
6.33% 6.94% 6.23% 12.11% 26.97% 23.90% Criterion B 2147 2008 1687 1827 6584 6197
7.17% 6.70% 5.63% 6.10% 21.98% 20.69% Sum 4042 4088 3554 5457 14663 13348
13.49% 13.65% 11.87% 18.22% 48.95% 44.56%
Table Found at http : //ukpmc . ac .uk/picrender . cgi?artid=16630688thlobnarne=ppat
1000154.s006.doc

Appendix E. Chapter 4 Supplementary Data 282
Table E.3: Over-represented Gene Ontology (GO) terms amongst genes having increased transcript abundance.
GO ID GO Term List hits List size Pop hits Pop
GO:0019538 protein metabolism 174 561 781 4
GO:0044267 cellular protein metabolism 159 561 695 4
G0:0044260 cellular macromolecule metabolism 168 561 775 4
G0:0043170 macromolecule metabolism 254 561 1383 4
GO:0042254 ribosome biogenesis and assembly 51 561 161 4
GO:0007028 cytoplasm organization and biogenesis 51 561 161 4
GO:0044237 cellular metabolism 366 561 2251 4
GO:0007046 ribosome biogenesis 46 561 140 4
GO:0016072 rRNA metabolism 41 561 118 4
GO:0006364 rRNA processing 40 561 115 4
GO:0044238 primary metabolism 334 561 2028 4
GO:0006457 protein folding 23 561 50 4
00:0006412 protein biosynthesis 72 561 302 4
GO:0008152 metabolism 394 561 2570 4
GO:0043037 translation 30 561 96 4
00:0016070 RNA metabolism 58 561 247 4
G0:0009059 macromolecule biosynthesis 74 561 346 4
GO:0006365 35S primary transcript processing 18 561 46 4
GO:0006413 translational initiation 17 561 42 4
166 RNA processing 43 561 175 4
GO:0030490 processing of 20S pre-rRNA 15 561 37 4
G0:0006996 organelle organization and biogenesis 100 561 522 4
GO:0009058 biosynthesis 123 561 696 4 mitochondria] electron transport, ubiquinol to cytochrome c 5 561 6 4
GO:0001302 replicative cell aging 6 561 9 4
GO:0000027 ribosomal large subunit assembly and maintenance 11 561 28 4
00:0007569 cell aging 6 561 11 4
GO:0006359 regulation of transcription from RNA polymerase III promoter 4 561 5 4
GO:0006360 transcription from RNA polymerase I promoter 7 561 15 4
G0:0009987 cellular process 461 561 3265 4
G0:0043283 biopolymer metabolism 151 561 930 4
00:0042257 ribosomal subunit assembly 12 561 37 4
GO:0000128 flocculation 3 561 3 4
GO:0006356 regulation of transcription from RNA polymerase I promoter 3 561 3 4
GO:0044249 cellular biosynthesis 105 561 616 4
GO:0042255 ribosome assembly 13 561 43 4
G0:0043412 biopolymer modification 73 561 406 4
00:0050875 cellular physiological process 454 561 3222 4
GO:0007568 aging 6 561 13 4
G0:0006783 heme biosynthesis 6 561 13 4
00:0042168 hems metabolism 6 561 13 4
GO:0000154 rRNA modification 6 561 13 4
GO:0006628 mitochondrial translocation 7 561 17 4
GO:0006508 proteolysis 27 561 125 4
00:0006778 porphyrin metabolism 6 561 14 4
GO:0006779 porphyrin biosynthesis 6 561 14 4
GO:0006626 protein targeting to mitochondrion 6 561 14 4
GO:0006461 protein complex assembly 19 561 79 4
G0:0016337 cell-cell adhesion 4 561 7 4 00:0009060 aerobic respiration 13 561 48 4 GO:0045333 cellular respiration 13 561 48 4 GO:0016043 cell organization and biogenesis 135 561 849 4
GO:0006696 ergosterol biosynthesis 11 561 38 9
GO:0008204 ergosterol metabolism 11 561 38 4
GO:0007155 cell adhesion 5 561 11 4
G0:0042273 ribosomal large subunit biogenesis 5 561 11 4
G0:0018193 peptidyl-amino acid modification 5 561 11 4
G0:0006091 generation of precursor metabolites and energy 32 561 161 4
conti

Appendix E. Chapter 4 Supplementary Data 283
continued from previous page GO ID GO Term List hits List size Pop hit:
GO:0006464 protein modification 64 561 372 mitochondrial electron transport, succinate to ubiquinone 2 561 2
GO:0042126 nitrate metabolism 2 561 2 GO:0042128 nitrate assimilation 2 561 2 GO:0044257 cellular protein catabolism 20 561 92 GO:0046148 pigment biosynthesis 7 561 22 GO:0042440 pigment metabolism 7 561 22 GO:0009451 RNA modification 9 561 32 GO:0051603 proteolysis during cellular protein catabolism 19 561 SE GO:0016126 sterol biosynthesis 11 561 42 GO:0015980 energy derivation by oxidation of organic compounds 24 561 12C GO:0006383 transcription from RNA polymerase III promoter 7 561 24 GO:0019941 modification-dependent protein catabolism 18 561 87 GO:0043632 modification-dependent macromolecule catabolism 18 561 87 GO:0000002 mitochondria] genome maintenance 5 561 15 GO:0000283 establishment of cell polarity (sensu Saccharomyces) 5 561 15 GO:0016125 sterol metabolism 11 561 47 GO:0006139 nucleobase, nucleoside, nucleotide and nucleic acid metabolism 127 561 835 GO:0006694 steroid biosynthesis 11 561 45 GO:0030163 protein catabolism 21 561 105 GO:0042798 protein neddylation during NEDD8 class-dependent protein catabolism 2 561 2 GO:0045041 protein import into mitochondrial intermembrane space 2 561 G0:0018195 peptidyl-arginine modification 2 561 00:0019942 NEDD8 class-dependent protein catabolism 2 561 GO:0006534 cysteine metabolism 2 561 GO:0016574 histone ubiquitination 2 561 GO:0006549 isoleucine metabolism 2 561 2 GO:0006839 mitochondrial transport 6 561 21 GO:0006512 ubiquitin cycle 22 561 119 GO:0006511 ubiquitin-dependent protein catabolism 17 561 85 GO:0043285 biopolymer catabolism 31 561 177 137 tRNA aminoacylation 6 561 22 G0:0006418 tRNA aminoacylation for protein translation 6 561 22 GO:0006403 RNA localization 8 561 32 G0:0046112 nucleobase biosynthesis 4 561 12 00:0006399 tRNA metabolism 11 561 52 GO:0008202 steroid metabolism 11 561 52 GO:0044265 cellular macromolecule catabolism 31 561 181 00:0019856 pyrimidine base biosynthesis 3 561 8 GO:0006743 ubiquinone metabolism 3 561 E GO:0016571 historic methylation 3 561 5 GO:0043414 biopolymer methylation 5 561 15 00:0032259 methylation 5 561 15 GO:0006090 pyruvate metabolism 7 561 25 00:0006730 one-carbon compound metabolism 7 561 25 G0:0008151 cell growth and/or maintenance 4 561 13 G0:0015931 nucleobase, nucleoside, nucleotide and nucleic acid transport 8 561 35 00:0007005 mitochondrion organization and biogenesis 9 561 41 GO:0006080 mamma metabolism 2 561 4 G0:0006097 glyoxylate cycle 2 561 4 G0:0016074 snoRNA metabolism 2 561 4 GO:0006835 dicarboxylic acid transport 2 561 4 G0:0096487 glyoxylate metabolism 2 561 9
deadenylation-dependent decapping 2 561 4 Molecular Function 0 0 0 C G0:0003754 chaperone activity 17 568 35 GO:0003743 translation initiation factor activity 16 568 34 G0:0008135 translation factor activity, nucleic acid binding 20 568 5C GO:0045182 translation regulator activity 21 568 55 G0:0016627 oxidoreductase activity, acting on the CH-CH group of donors 11 568 19 00:0030515 snoRNA binding 11 568 21 GO:0003723 RNA binding 27 568 10C

Appendix E. Chapter 4 Supplementary Data 284
continued from previous page GO ID
GO Term
GO:0003735 GO:0003724 GO:0016628 oxidoreductase activity,
GO:0004680 GO:0004004 GO:0008186 GO:0004682 GO:0016274 GO:0016273 GO:0004176 GO:0008026 GO:0016876 GO:0016875 GO:0004812 GO:0003773 GO:0008369 GO:0004175 GO:0004386 GO:0008565 GO:0004691 GO:0004690 GO:0008398 GO:0008177 GO:0008097 GO:0004739 GO:0016635 oxidoreductase activity, acting on the
GO:0000246 GO:0004827 GO:0000339 GO:0004738 GO:0008121 GO:0016681 oxidoreductase activity, acting on c
GO :0003925 GO:0016679 oxidoreduc
GO:0015078 G 0:0004722 GO :0016705 oxidoreductase activity, acting on I
GO :0008170 GO :0003676 GO :0005198 GO:0016709 oxidoreductase activity, acting on paired donors, with incorporation or reduction of molecular oxygen, NAI
GO:0008233 GO :0008757 GO :0016251 GO:0019202 GO:0000248 GO:0016661 oxidore
GO:0000171 GO:0047956 GO:0005294 GO:0000102 GO:0019206 GO:0004526 GO:0000049 GO:0016624 oxidoreductase activity, a
GO:0016741 GO:0004674 GO:0016455 GO:0015077 GO:0003674 GO:0008168 GO:0042054

Appendix E. Chapter 4 Supplementary Data 285
continued from previous page GO ID GO Term List hits List size Pop hits F
GO:0019205 nucleobase, nucleoside, nucleotide kinase activity 3 568 8 GO:0016491 oxidoreductase activity 68 568 441 G0:0016215 CoA desaturase activity 2 568 4 GO:0015191 L-methionine transporter activity 2 568 4 00:0004768 stearoyl-CoA 9-desaturase activity 2 568 4 G0:0030276 clathrin binding 2 568 4 GO:0005310 dicarboxylic acid transporter activity 2 568 4 GO:0004128 cytochrome-b5 reductase activity 2 568 4 60:0004721 phosphoprotein phosphatase activity 7 568 30 GO:0016830 carbon-carbon lyase activity 8 568 36 GO:0004497 monooxygenase activity 6 568 25 Cellular component 0 0 0 0 GO:0044429 mitochondria! part 65 526 222 GO:0031974 membrane-enclosed lumen 96 526 395 G0:0043233 organelle lumen 96 526 395 G0:0043234 protein complex 149 526 711 GO:0005730 nucleolus 40 526 130 GO:0005739 mitochondrion 95 526 426 GO:0030529 ribonucleoprotein complex 62 526 243 GO:0043228 non-membrane-bound organelle 96 526 442 GO:0043232 intracellular non-membrane-bound organelle 96 526 442 GO:0005759 mitochondrial matrix 31 526 93 GO:0031980 mitochondrial lumen 31 526 93 G0:0005852 eukaryotic translation initiation factor 3 complex 9 526 12 GO:0031966 mitochondria! membrane 31 526 97 GO:0044455 mitochondrial membrane part 13 526 24 GO:0005622 intracellular 441 526 2891 GO:0044424 intracellular part 438 526 2868 GO:0044452 nucleolar part 17 526 39 GO:0005761 mitochondria! ribosome 17 526 41 G0:0005840 ribosome 40 526 151 00:0000313 organellar ribosome 17 526 43 GO:0005746 mitochondrial electron transport chain 8 526 12 GO:0005743 mitochondrial inner membrane 25 526 80 GO:0019866 organelle inner membrane 25 526 82 GO:0031981 nuclear lumen 60 526 270 GO:0005740 mitochondrial envelope 34 526 129 GO:0005732 small nucleolar ribonucleoprotein complex 10 526 23 GO:0016282 eukaryotic 43S preinitiation complex 15 526 47 GO:0005844 polysome 4 526 5 GO:0045285 ubiquinol-cytochrome-c reductase complex 4 526 5 00:0005750 respiratory chain complex III (sensu Eukaryota) 4 526 5 G0:0045275 respiratory chain complex III 4 526 5
proteasome regulatory particle, lid subcomplex (sensu Eukaryota) 5 526 8 G0:0031967 organelle envelope 37 526 167 GO:0031975 envelope 37 526 167 00:0005956 protein kinase CK2 complex 3 526 3 GO:0005762 mitochondrial large ribosomal subunit 9 526 24 GO:0044422 organelle part 177 526 1086 GO:0044446 intracellular organelle part 177 526 1086 GO:0000315 organellar large ribosomal subunit 9 526 25 GO:0005737 cytoplasm 306 526 2010 GO:0005763 mitochondrial small ribosomal subunit 6 526 14 G0:0043229 intracellular organelle 335 526 2228 GO:0043226 organelle 335 526 2228 G0:0045254 pyruvate dehydrogenase complex 3 526 4 GO:0000314 organellar small ribosomal subunit 6 526 15 GO:0015934 large ribosomal subunit 16 526 65 GO:0005845 mRNA cap complex 2 526 2 GO:0005749 respiratory chain complex II (sensu Eukaryota) 2 526 2 GO:0045283 fumarate reductase complex 2 526 2 GO:0045273 respiratory chain complex II 2 526 2
Co

Appendix E. Chapter 4 Supplementary Data 286
continued from previous pare GO ID GO Term List hits List size Pop hits
GO:0045257 succinate dehydrogenase complex (ubiquinone) 2 526 2
GO:0045281 succinate dehydrogenase complex 2 526 2 GO:0044464 cell part 490 526 3436
GO:0005742 mitochondrial outer membrane translocase complex 3 526 5
GO:0005741 mitochondrial outer membrane 6 526 17 GO:0044444 cytoplasmic part 194 526 1254 GO:0031968 organelle outer membrane 6 526 18
00:0019867 outer membrane 6 526 18
GO:0005623 cell 490 526 3444 GO:0015935 small ribosomal subunit 12 526 50
proteasome core complex, beta-subunit complex (sensu Eukaryota) 3 526 6
GO:0030663 COPI coated vesicle membrane 3 526 6 GO:0030126 COPI vesicle coat 3 526 6 00:0005736 DNA-directed RNA polymerase I complex 4 526 10
00:0000502 proteasome complex (sensu Eukaryota) 8 526 29 G0:0030677 ribonuclease P complex 2 526 3
GO:0000172 ribonuclease MRP complex 2 526 3 proteasome regulatory particle, base subcomplex (sensu Eukaryota) 2 526 3
GO:0005655 nucleolar ribonuclease P complex 2 526 3 GO:0012506 vesicle membrane 6 526 22
GO:0030120 vesicle coat 6 526 22 00:0030659 cytoplasmic vesicle membrane 6 526 22 GO:0030662 coated vesicle membrane 6 526 22 GO:0043231 intracellular membrane-bound organelle 301 526 2061 GO:0043227 membrane-bound organelle 301 526 2061 GO:0031090 organelle membrane 51 526 307
G0:0005839 proteasome core complex (sensu Eukaryota) 4 526 13 GO:0019897 extrinsic to plasma membrane 4 526 13 G0:0012510 trans-Golgi network transport vesicle membrane 2 526 4
GO:0030130 clathrin coat of trans-Golgi network vesicle 2 526 4 GO:0030121 AP-1 adaptor complex 2 526 4
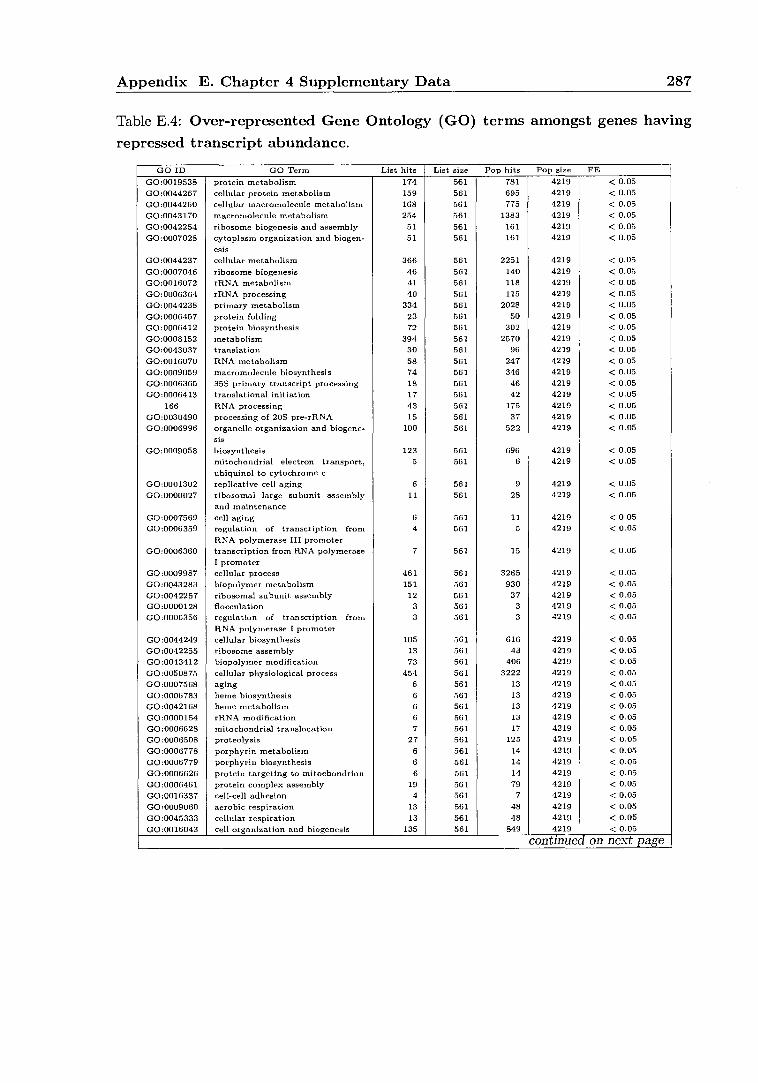
Appendix E. Chapter 4 Supplementary Data 287
Table E.4: Over-represented Gene Ontology (GO) terms amongst genes having repressed transcript abundance.
GO ID GO Term List hits List size Pop hits Pop size FE GO:0019538 protein metabolism 174 561 781 4219 < 0.05 GO:0044267 cellular protein metabolism 159 561 695 4219 < 0.05 GO:0044260 cellular macromolecule metabolism 168 561 775 4219 < 0.05 GO:0043170 macromolecule metabolism 254 561 1383 4219 < 0.05 GO:0042254 ribosome biogenesis and assembly 51 561 161 4219 < 0.05 GO:0007028 cytoplasm organization and biogen-
esis 51 561 161 4219 < 0.05
GO:0044237 cellular metabolism 366 561 2251 4219 < 0.05 00:0007046 ribosome biogenesis 46 561 140 4219 < 0.05 GO:0016072 rRNA metabolism 41 561 118 4219 < 0.05 GO:0006364 rRNA processing 40 561 115 4219 < 0.05 GO:0044238 primary metabolism 334 561 2028 4219 < 0.05 GO:0006457 protein folding 23 561 50 4219 < 0.05 GO:0006412 protein biosynthesis 72 561 302 4219 < 0.05 00:0008152 metabolism 394 561 2570 4219 < 0.05 GO:0043037 translation 30 561 96 4219 < 0.05 GO:0016070 RNA metabolism 58 561 247 4219 < 0.05 00:0009059 macromolecule biosynthesis 74 561 346 4219 < 0.05 GO:0006365 35S primary transcript processing 18 561 46 4219 < 0.05 GO:0006413 translational initiation 17 561 42 4219 < 0.05
166 RNA processing 43 561 175 4219 < 0.05 60:0030490 processing of 20S pre-rRNA 15 561 37 4219 < 0.05 G0:0006996 organelle organization and biogene-
sis 100 561 522 4219 < 0.05
GO:0009058 biosynthesis 123 561 696 4219 < 0.05 mitochondrial electron transport, ubiquinol to cytochrome c
5 561 6 4219 < 0.05
00:0001302 replicative cell aging 6 561 9 4219 < 0.05 00:0000027 ribosomal large subunit assembly
and maintenance 11 561 28 4219 < 0.05
GO:0007569 cell aging 6 561 11 4219 < 0.05 GO:0006359 regulation of transcription from 4 561 5 4219 < 0.05
RNA polymerase III promoter G0:0006360 transcription from RNA polymerase 7 561 15 4219 < 0.05
I promoter GO:0009987 cellular process 461 561 3265 4219 < 0.05 GO:0043283 biopolymer metabolism 151 561 930 4219 < 0.05 GO:0042257 ribosomal subunit assembly 12 561 37 4219 < 0.05 GO:0000128 flocculation 3 561 3 4219 < 0.05 GO:0006356 regulation of transcription from 3 561 3 4219 < 0.05
RNA polymerase I promoter GO:0044249 cellular biosynthesis 105 561 616 4219 < 0.05 GO:0042255 ribosome assembly 13 561 43 4219 < 0.05 GO:0043412 biopolymer modification 73 561 406 4219 < 0.05 GO:0050875 cellular physiological process 454 561 3222 4219 < 0.05 GO:0007568 aging 6 561 13 4219 < 0.05 GO:0006783 heme biosynthesis 6 561 13 4219 < 0.05 GO:0042168 heme metabolism 6 561 13 4219 < 0.05 GO:0000154 rRNA modification 6 561 13 4219 < 0.05 60:0006628 mitochondrial translocation 7 561 17 4219 < 0.05 GO:0006508 proteolysis 27 561 125 4219 < 0.05 GO:0006778 porphyrin metabolism 6 561 14 4219 < 0.05 GO:0006779 porphyrin biosynthesis 6 561 14 4219 < 0.05 GO:0006626 protein targeting to mitochondrion 6 561 14 4219 < 0.05 GO:0006461 protein complex assembly 19 561 79 4219 < 0.05 GO:0016337 cell-cell adhesion 4 561 7 4219 < 0.05 GO:0009060 aerobic respiration 13 561 48 4219 < 0.05 00:0045333 cellular respiration 13 561 48 4219 < 0.05 00:0016043 cell organization and biogenesis 135 561 849 4219 < 0.05
continued on next page

Appendix E. Chapter 4 Supplementary Data 288
continued from previous page GO ID GO Term List hits List size Pop hits Pop size FE
G0:0006696 ergosterol biosynthesis 11 561 38 4219 < 0.05 GO:0008204 ergosterol metabolism 11 561 38 4219 < 0.05 00:0007155 cell adhesion 5 561 11 4219 < 0.05 GO:0042273 ribosomal large subunit biogenesis 5 561 11 4219 < 0.05 00:0018193 peptidyl-amino acid modification 5 561 11 4219 < 0.05 GO:0006091 generation of precursor metabolites
and energy 32 561 161 4219 < 0.05
GO:0006464 protein modification 64 561 373 4219 < 0.05 mitochondrial electron transport, succinate to ubiquinone
2 561 2 4219 < 0.05
G0:0042126 nitrate metabolism 2 561 2 4219 < 0.05 00:0042128 nitrate assimilation 2 561 2 4219 < 0.05 00:0044257 cellular protein catabolism 20 561 93 4219 < 0.05 GO:0046148 pigment biosynthesis 7 561 22 4219 < 0.05 GO:0042440 pigment metabolism 7 561 22 4219 < 0.05 GO:0009451 RNA modification 9 561 32 4219 < 0.05 GO:0051603 proteolysis during cellular protein
catabolism 19 561 88 4219 < 0.05
GO:0016126 sterol biosynthesis 11 561 43 4219 < 0.05 G0:0015980 energy derivation by oxidation of
organic compounds 24 561 120 4219 < 0.05
GO:0006383 transcription from RNA polymerase 7 561 24 4219 < 0.05 III promoter
GO:0019941 modification-dependent protein catabolism
18 561 87 4219 < 0.05
GO:0043632 modification-dependent macro- molecule catabolism
18 561 87 4219 < 0.05
GO:0000002 mitochondrial genome maintenance 5 561 15 4219 < 0.05 GO:0000283 establishment of cell polarity (sensu 5 561 15 4219 < 0.05
Saccharomyces) G0:0016125 sterol metabolism 11 561 47 4219 < 0.05 GO:0006139 nucleobase, nucleoside, nucleotide
and nucleic acid metabolism 127 561 839 4219 < 0.05
G0:0006694 steroid biosynthesis 11 561 48 4219 < 0.05 G0:0030163 protein catabolism 21 561 109 4219 < 0.05 GO:0042798 protein neddylation during NEDD8
class-dependent protein catabolism 2 561 3 4219 < 0.05
GO:0045041 protein import into mitochondrial intermembrane space
2 561 3 4219 < 0.05
G0:0018195 peptidyl-arginine modification 2 561 3 4219 < 0.05 GO:0019942 NEDD8 class-dependent protein
catabolism 2 561 3 4219 < 0.05
GO:0006534 cysteine metabolism 2 561 3 4219 < 0.05 00:0016574 histone ubiquitination 2 561 3 4219 < 0.05 GO:0006549 isoleucine metabolism 2 561 3 4219 < 0.05 G0:0006839 mitochondrial transport 6 561 21 4219 0.0504 GO:0006512 ubiquitin cycle 22 561 116 4219 0.0509 GO:0006511 ubiquitin-dependent protein
catabolism 17 561 85 4219 0.0523
00:0043285 biopolymer catabolism 31 561 177 4219 0.0614 137 tRNA aminoacylation 6 561 22 4219 0.0618
GO:0006418 tRNA aminoacylation for protein translation
6 561 22 4219 0.0618
GO:0006403 RNA localization 8 561 33 4219 0.0625 60:0046112 nucleobase biosynthesis 4 561 12 4219 0.0638 GO:0006399 tRNA metabolism 11 561 52 4219 0.0762 GO:0008202 steroid metabolism 11 561 52 4219 0.0762 GO:0044265 cellular macromolecule catabolism 31 561 181 4219 0.0783 00:0019856 pyrimidine base biosynthesis 3 561 8 4219 0.0783 GO:0006743 ubiquinone metabolism 3 561 8 4219 0.0783 G0:0016571 histone methylation 3 561 8 4219 0.0783 GO:0043414 biopolymer methylation 5 561 18 4219 0.0799 GO:0032259 methylation 5 561 18 4219 0.0799 GO:0006090 pyruvate metabolism 7 561 29 4219 0.0805
continued on next page

Appendix E. Chapter 4 Supplementary Data 289
continued from previous page GO ID GO Term List hits List size Pop hits Pop size FE
GO:0006730 one-carbon compound metabolism 7 561 29 4219 0.0805 GO:0008151 cell growth and/or maintenance 4 561 13 4219 0.0828 00:0015931 nucleobase, nucleoside, nucleotide
and nucleic acid transport 8 561 35 4219 0.0838
00:0007005 mitochondrion organization and biogenesis
9 561 41 4219 0.0853
00:0006080 mannan metabolism 2 561 4 4219 0.0881 GO:0006097 glyoxylate cycle 2 561 4 4219 0.0881 GO:0016074 snoRNA metabolism 2 561 4 4219 0.0881 00:0006835 dicarboxylic acid transport 2 561 4 4219 0.0881 GO:0046487 glyoxylate metabolism 2 561 4 4219 0.0881
deadenylation-dependent decap- ping
2 561 4 4219 0.0881
Molecular Function 0 0 0 0 0 < 0.05 GO:0003754 chaperone activity 17 568 35 4324 < 0.05 GO:0003743 translation initiation factor activity 16 568 34 4324 < 0.05 G0:0008135 translation factor activity, nucleic
acid binding 20 568 50 4324 < 0.05
GO:0045182 translation regulator activity 21 568 55 4324 < 0.05 00:0016627 oxidoreductase activity, acting on
the CH-CH group of donors 11 568 19 4324 < 0.05
00:0030515 snoRNA binding 11 568 21 4324 < 0.05 GO:0003723 RNA binding 27 568 100 4324 < 0.05 00:0003735 structural constituent of ribosome 28 568 112 4324 < 0.05 G0:0003724 RNA helicase activity 13 568 39 4324 < 0.05 GO:0016628 oxidoreductase activity, acting on
the CH-CH group of donors, NAD or NADP as acceptor
4 568 5 4324 < 0.05
GO:0004680 casein kinase activity 4 568 5 4324 < 0.05 00:0004004 ATP-dependent RNA helicase ac-
tivity 10 568 27 4324 < 0.05
GO:0008186 RNA-dependent ATPase activity 10 568 27 4324 < 0.05 GO:0004682 protein kinase CIC2 activity 3 568 3 4324 < 0.05 GO:0016274 protein-arginine N-
methyltransferase activity 3 568 4 4324 < 0.05
GO:0016273 arginine N-methyltransferase activ- ity
3 568 4 4324 < 0.05
GO:0004176 ATP-dependent peptidase activity 3 568 4 4324 < 0.05 GO:0008026 ATP-dependent helicase activity 11 568 40 4324 < 0.05 GO:0016876 ligase activity, forming aminoacyl-
tRNA and related compounds 10 568 35 4324 < 0.05
00:0016875 ligase activity, forming carbon- oxygen bonds
10 568 35 4324 < 0.05
G0:0004812 aminoacyl-tRNA ligase activity 10 568 35 4324 < 0.05 GO:0003773 heat shock protein activity 4 568 8 4324 < 0.05 GO:0008369 obsolete molecular function 38 568 204 4324 < 0.05 GO:0004175 endopeptidase activity 13 568 52 4324 < 0.05 GO:0004386 helicase activity 16 568 69 4324 < 0.05 60:0008565 protein transporter activity 9 568 31 4324 < 0.05 G0:0004691 cAMP-dependent protein kinase ac-
tivity 2 568 2 4324 < 0.05
G0:0004690 cyclic nucleotide-dependent protein kinase activity
2 568 2 4324 < 0.05
GO:0008398 sterol 14-demethylase activity 2 568 2 4324 < 0.05 GO:0008177 succinate dehydrogenase 2 568 2 4324 < 0.05
(ubiquinone) activity G0:0008097 5S rRNA binding 2 568 2 4324 < 0.05 GO:0004739 pyruvate dehydrogenase (acetyl-
transferring) activity 2 568 2 4324 < 0.05
GO:0016635 oxidoreductase activity, acting on the CH-CH group of donors, quinone or related compound as acceptor
2 568 2 4324 < 0.05
continued on next page

Appendix E. Chapter 4 Supplementary Data 290
continued from previous page GO ID GO Term List hits List size Pop hits Pop size FE
00:0000246 delta24(24-1) sterol reductase ac- tivity
2 568 2 4324 < 0.05
GO:0004827 proline-tRNA ligase activity 2 568 2 4324 < 0.05 GO:0000339 RNA cap binding 2 568 2 4324 < 0.05 GO:0004738 pyruvate dehydrogenase activity 2 568 2 4324 < 0.05 GO:0008121 ubiquinol-cytochrome-c reductase
activity 3 568 5 4324 < 0.05
GO:0016681 oxidoreductase activity, acting on diphenols and related substances as donors, cytochrome as acceptor
3 568 5 4324 < 0.05
GO:0003925 small monomeric GTPase activity 3 568 5 4324 < 0.05 GO:0016679 oxidoreductase activity, acting on
diphenols and related substances as donors
3 568 5 4324 < 0.05
GO:0015078 hydrogen ion transporter activity 5 568 13 4324 < 0.05 00:0004722 protein serine/threonine phos-
phatase activity 6 568 18 4324 < 0.05
GO:0016705 oxidoreductase activity, acting on paired donors, with incorporation or reduction of molecular oxygen
6 568 18 4324 < 0.05
GO:0008170 N-rnethyltransferase activity 6 568 18 4324 < 0.05 GO:0003676 nucleic acid binding 73 568 448 4324 < 0.05 00:0005198 structural molecule activity 31 568 168 4324 < 0.05 00:0016709 oxidoreductase activity, acting on
paired donors, with incorporation or reduction of molecular oxygen, NAD or NADH as one donor, and incorporation of one atom of oxy-gen
3 568 6 4324 < 0.05
GO:0008233 peptidase activity 23 568 119 4324 < 0.05 GO:0008757 S-adenosylmethionine-dependent
methyltransferase activity 12 568 53 4324 < 0.05
00:0016251 general RNA polymerase II tran- scription factor activity
9 568 36 4324 < 0.05
GO:0019202 amino acid kinase activity 2 568 3 4324 < 0.05 GO:0000248 C-5 sterol desaturase activity 2 568 3 4324 < 0.05 GO:0016661 oxidoreductase activity, acting on
other nitrogenous compounds as donors
2 568 3 4324 < 0.05
GO:0000171 ribonuclease MRP activity 2 568 3 4324 < 0.05 GO:0047956 glycerol dehydrogenase (NADP+)
activity 2 568 3 4324 < 0.05
GO:0005294 neutral L-amino acid porter activity 2 568 3 4324 < 0.05 G0:0000102 L-methionine porter activity 2 568 3 4324 < 0.05 GO:0019206 nucleoside kinase activity 2 568 3 4324 < 0.05 GO:0004526 ribonuclease P activity 2 568 3 4324 < 0.05 GO:0000049 tRNA binding 2 568 3 4324 < 0.05 GO:0016624 oxidoreductase activity, acting on
the aldehyde or oxo group of donors, disulfide as acceptor
2 568 3 4324 < 0.05
GO:0016741 transferase activity, transferring one-carbon groups
15 568 73 4324 < 0.05
00:0004674 protein serine/threonine kinase ac- tivity
11 568 49 4324 < 0.05
00:0016455 RNA polymerase II transcription mediator activity
3 568 7 4324 0.0525
GO:0015077 monovalent inorganic cation trans- porter activity
5 568 17 4324 0.0617
GO:0003674 molecular! unction 549 568 4124 4324 0.0691 GO:0008168 methyltransferase activity 14 568 71 4324 0.075 GO:0042054 histone methyltransferase activity 3 568 8 4324 0.076 G0:0019205 nucleobase, nucleoside, nucleotide
kinase activity 3 568 8 4324 0.076
continued on next page

Appendix E. Chapter 4 Supplementary Data 291
continued from previous page GO ID GO Term List hits List size Pop hits Pop size FE
GO:0016491 oxidoreductase activity 68 568 441 4324 0.0792 GO:0016215 CoA desaturase activity 2 568 4 4324 0.0862 GO:0015191 L-methionine transporter activity 2 568 4 4324 0.0862 GO:0004768 stearoyl-CoA 9-desaturase activity 2 568 4 4324 0.0862 GO:0030276 clathrin binding 2 568 4 4324 0.0862 GO:0005310 dicarboxylic acid transporter activ-
ity 2 568 4 4324 0.0862
GO:0004128 cytochrome-b5 reductase activity 2 568 4 4324 0.0862 GO:0004721 phosphoprotein phosphatase activ-
ity 7 568 30 4324 0.0889
GO:0016830 carbon-carbon lyase activity 8 568 36 4324 0.0907 GO:0004497 monooxygenase activity 6 568 25 4324 0.0998
Cellular component 0 0 0 0 0 < 0.05 GO:0044429 mitochondrial part 65 526 222 3787 < 0.05 GO:0031974 membrane-enclosed lumen 96 526 395 3787 < 0.05 GO:0043233 organelle lumen 96 526 395 3787 < 0.05 GO:0043234 protein complex 149 526 711 3787 < 0.05 GO:0005730 nucleolus 40 526 130 3787 < 0.05 GO:0005739 mitochondrion 95 526 426 3787 < 0.05 GO:0030529 ribonucleoprotein complex 62 526 243 3787 < 0.05 GO:0043228 non-membrane-bound organelle 96 526 442 3787 < 0.05 GO:0043232 intracellular non-membrane-bound
organelle 96 526 442 3787 < 0.05
GO:0005759 mitochondrial matrix 31 526 93 3787 < 0.05 GO:0031980 mitochondrial lumen 31 526 93 3787 < 0.05 GO:0005852 eukaryotic translation initiation
factor 3 complex 9 526 12 3787 < 0.05
GO:0031966 mitochondrial membrane 31 526 97 3787 < 0.05 GO:0044455 mitochondria] membrane part 13 526 24 3787 < 0.05 GO:0005622 intracellular 441 526 2891 3787 < 0.05 00:0044424 intracellular part 438 526 2868 3787 < 0.05 GO:0044452 nucleolar part 17 526 39 3787 < 0.05 00:0005761 mitochondrial ribosome 17 526 41 3787 < 0.05 00:0005840 ribosome 40 526 151 3787 < 0.05 GO:0000313 organellar ribosome 17 526 43 3787 < 0.05 GO:0005746 mitochondrial electron transport
chain 8 526 12 3787 < 0.05
GO:0005743 mitochondrial inner membrane 25 526 80 3787 < 0.05 GO:0019866 organelle inner membrane 25 526 82 3787 < 0.05 GO:0031981 nuclear lumen 60 526 270 3787 < 0.05 GO:0005740 mitochondrial envelope 34 526 129 3787 < 0.05 GO:0005732 small nucleolar ribonucleoprotein
complex 10 526 23 3787 < 0.05
GO:0016282 eukaryotic 43S preinitiation com- plex
15 526 47 3787 < 0.05
GO:0005844 polysome 4 526 5 3787 < 0.05 GO:0045285 ubiquinol-cytochrome-c reductase
complex 4 526 5 3787 < 0.05
GO:0005750 respiratory chain complex III (sensu 4 526 5 3787 < 0.05 Eukaryota)
GO:0045275 respiratory chain complex III 4 526 5 3787 < 0.05 proteasome regulatory particle, lid subcomplex (sensu Eukaryota)
5 526 8 3787 < 0.05
GO:0031967 organelle envelope 37 526 167 3787 < 0.05 GO:0031975 envelope 37 526 167 3787 < 0.05 GO:0005956 protein kinase CK2 complex 3 526 3 3787 < 0.05 GO:0005762 mitochondria] large ribosomal sub-
unit 9 526 24 3787 < 0.05
GO:0044422 organelle part 177 526 1086 3787 < 0.05 GO:0044446 intracellular organelle part 177 526 1086 3787 < 0.05 GO:0000315 organellar large ribosomal subunit 9 526 25 3787 < 0.05 GO:0005737 cytoplasm 306 526 2010 3787 < 0.05 00:0005763 mitochondrial small ribosomal sub-
unit 6 526 14 3787 < 0.05
continued on next page

Appendix E. Chapter 4 Supplementary Data 292
continued from previous page GO ID GO Term List hits List size Pop hits Pop size FE
00:0043229 intracellular organelle 335 526 2228 3787 < 0.05 GO:0043226 organelle 335 526 2228 3787 < 0.05 GO:0045254 pyruvate dehydrogenase complex 3 526 4 3787 < 0.05 GO:0000314 organellar small ribosomal subunit 6 526 15 3787 < 0.05 GO:0015934 large ribosomal subunit 16 526 65 3787 < 0.05 GO:0005845 mRNA cap complex 2 526 2 3787 < 0.05 00:0005749 respiratory chain complex II (sensu 2 526 2 3787 < 0.05
Eukaryota) GO:0045283 fumarate reductase complex 2 526 2 3787 < 0.05 G0:0045273 respiratory chain complex II 2 526 2 3787 < 0.05 00:0045257 succinate dehydrogenase complex 2 526 2 3787 < 0.05
(ubiquinone) GO:0045281 succinate dehydrogenase complex 2 526 2 3787 < 0.05 GO:0044464 cell part 490 526 3436 3787 < 0.05 GO:0005742 mitochondria] outer membrane
translocase complex 3 526 5 3787 < 0.05
00:0005741 mitochondria] outer membrane 6 526 17 3787 < 0.05 G0:0044444 cytoplasmic part 194 526 1254 3787 < 0.05 GO:0031968 organelle outer membrane 6 526 18 3787 < 0.05 GO:0019867 outer membrane 6 526 18 3787 < 0.05 GO:0005623 cell 490 526 3444 3787 < 0.05 GO:0015935 small ribosomal subunit 12 526 50 3787 < 0.05
proteasome core complex, beta- subunit complex (sensu Eukaryota)
3 526 6 3787 < 0.05
GO:0030663 COPI coated vesicle membrane 3 526 6 3787 < 0.05 GO:0030126 COPI vesicle coat 3 526 6 3787 < 0.05 GO:0005736 DNA-directed RNA polymerase I
complex 4 526 10 3787 < 0.05
GO:0000502 proteasome complex (sensu Eukary- ota)
8 526 29 3787 < 0.05
GO:0030677 ribonuclease P complex 2 526 3 3787 0.0525 GO:0000172 ribonuclease MRP complex 2 526 3 3787 0.0525
proteasome regulatory particle, base subcomplex (sensu Eukaryota)
2 526 3 3787 0.0525
GO:0005655 nucleolar ribonuclease P complex 2 526 3 3787 0.0525 GO:0012506 vesicle membrane 6 526 22 3787 0.0736 GO:0030120 vesicle coat 6 526 22 3787 0.0736 GO:0030659 cytoplasmic vesicle membrane 6 526 22 3787 0.0736 GO:0030662 coated vesicle membrane 6 526 22 3787 0.0736 G0:0043231 intracellular membrane-bound or-
ganelle 301 526 2061 3787 0.0895
Table E.5: A. fumigatus genes having increased transcript abundance, relative to laboratory culture, in the murine lung and S. cerevisiae orthologs regulated by rapamycin-mediated TOR kinase inhibition
Accesion SC ORF Description Afu4g13660 YBRO43C Multidrug transporter required for resistance to quinidine, barban, cis-
platin, and bleomycin Afu1g15520 YBR208C Urea amidolyase, contains both urea carboxylase and allophanate hy-
drolase activities, degrades urea to CO2 and NH3; expression sensitive to nitrogen catabolite repression.
Afu7g06380 YBR299W Maltase (alpha-D-glucosidase), inducible protein involved in maltose catabolism; encoded in the MAL3 complex locus; functional in genomic reference strain S288C
Afulg06150 YCL064C Catabolic L-serine (L-threonine) deaminase, catalyzes the degradation of both L-serine and L-threonine; required to use serine or threonine as the sole nitrogen source.
Afulg10780 YDR019C T subunit of the mitochondria] glycine decarboxylase complex, required for the catabolism of glycine to 5,10-methylene-THF.
Afu2g04260 YDR421W Zinc finger transcriptional activator of the Zn2Cys6 family; activates transcription of aromatic amino acid catabolic genes in the presence of aromatic amino acids
continued on next page

Appendix E. Chapter 4 Supplementary Data 293
continued from previous page Accesion SC ORF Description
Afu7g06060
Afu1g17370
Afu5g08090
Afulg17150
Afu3g07850
Afu5g13810
Afu5g01520
Afu6g03540
Afu8g04760
Afu2g17300
Afu7g00850 Afu1g09690
Afu7g04290
Afu6g07720
Afu1g12620 Afu3g02300
Afu7g04970
Afu4g14640 Afu7g04710
Afu7g00830
Afu2g02480 Afu6g08570
Afu7g01090
Afu7g01000
Afulg01600
Afu1g04780
YEL065W
YFLO14W
YFLO59W
YGR288W
YHR028C
YHR112C
YIL121W
YIR031C
YIR032C
YIR038C
YIR042C Y.IL087C
YKR039W
YKR097W
YKR105C YLR142W
YLR214W
YMR319C YNL023C
YNR064C
YOL048C YOR346W
YOR348C
YOR374W
YOR386W
YPL147W
Ferrioxamine B transporter, member of the ARN family of transporters that specifically recognize siderophore-iron chelates; transcription is in-duced during iron deprivation. Plasma membrane localized protein that protects membranes from des-iccation; induced by heat shock, oxidative stress, osmostress, stationary phase entry, glucose depletion. Member of a stationary phase-induced gene family; transcription of SNZ2 is induced prior to diauxic shift, and also in the absence of thi-amin in a Thi2p-dependent manner;. MAL-activator protein, part of complex locus MALI.; nonfunctional in genomic reference strain S288C Dipeptidyl aminopeptidase, synthesized as a glycosylated precursor; lo-calizes to the vacuolar membrane; similar to Stel3p Putative protein of unknown function; green fluorescent protein (GFP)-fusion protein localizes to the cytoplasm Multidrug transporter required for resistance to quinidine, barban, cis-platin, and bleomycin; may have a role in potassium uptake; member of the major facilitator superfamily. Malate synthase, role in allantoin degradation unknown; expression sen-sitive to nitrogen catabolite repression and induced by allophanate, an intermediate in allantoin degradation Ureidoglycolate hydrolase, converts ureidoglycolate to glyoxylate and urea in the third step of allantoin degradation; expression sensitive to nitrogen catabolite repression ER associated glutathione S-transferase capable of homodimerization; expression induced during the diauxic shift and throughout stationary phase. Putative protein of unknown function; YIR042C is a non-essential gene tRNA ligase, required for tRNA splicing; composed of three essential domains containing the phosphodiesterase, polynucleotide kinase, and ligase activities. General amino acid permease; localization to the plasma membrane is regulated by nitrogen source Phosphoenolpyruvate carboxykinase, key enzyme in gluconeogenesis, catalyzes early reaction in carbohydrate biosynthesis, glucose represses transcription. Putative transporter of the Major Facilitator Superfamily (MFS) Proline oxidase, nuclear-encoded mitochondrial protein involved in uti-lization of proline as sole nitrogen source; PUT1 transcription is induced by Put3p in the presence of proline Ferric reductase and cupric reductase, reduces siderophore-bound iron and oxidized copper prior to uptake by transporters; expression induced by low copper and iron levels Low-affinity Fe(II) transporter of the plasma membrane Protein that binds to Fprlp (FKBP12), conferring rapamycin resistance by competing with rapamycin for Fprlp binding; has similarity to pu-tative transcription factors. Epoxide hydrolase, member of the alpha/beta hydrolase fold family; may have a role in detoxification of epoxides Putative protein of unknown function Deoxycytidyl transferase, forms a complex with the subunits of DNA polymerase zeta, Rev3p and Rev7p; involved in repair of abasic sites in damaged DNA Proline permease, required for high-affinity transport of proline; also transports the toxic proline analog azetidine-2-carboxylate (AzC);. Mitochondrial aldehyde dehydrogenase, required for growth on ethanol and conversion of acetaldehyde to acetate; activity is K+ dependent. DNA photolyase involved in photoreactivation, repairs pyrimidine dimers in the presence of visible light; induced by DNA damage; regu-lated by transcriptional repressor Rphlp Subunit of a heterodimeric peroxisomal ATP-binding cassette trans-porter complex (Pxalp-Pxa2p), required for import of long-chain fatty acids into peroxisomes.
continued on next page

Appendix E. Chapter 4 Supplementary Data 294
continued from previous page Accesion SC ORF Description
Afu2g08800
Afufig03730
Afu3g02280
YPL265W
YPROO2W
YPRO26W
Dicarboxylic amino acid permease, mediates high-affinity and high-capacity transport of L-glutamate and L-aspartate; also a transporter for Gln, Asn, Ser, Ala, and Gly Mitochondria] protein that participates in respiration, induced by di-auxic shift; homologous to E. coli PrpD, may take part in the conversion of 2-methylcitrate to 2-methylisocitrate Acid trehalase required for utilization of extracellular trehalose
Table E.6: A. fumigatus genes having decreased transcript abundance, relative to laboratory culture, in the murine lung and S. cerevisiae orthologs regulated by rapamycin-mediated TOR kinase inhibition
Accesion SC ORF Description Afu1g05560 YAL036C Member of the DRG family of GTP-binding proteins; interacts with
translating ribosomes and with Tma46p Afu8g05580 YBL015W Acetyl-coA hydrolase, primarily localized to mitochondria; required for
acetate utilization and for diploid pseudohyphal growth Afu1g07470 YBL078C Protein required for autophagy; modified by the serial action of Atg4p,
Atg7p, and Atg3p, and conjugated at the C terminus with phos-phatidylethanolamine,.
Afulg06190 YBRO34C Nuclear SAM-dependent mono- and asymmetric arginine dimethylating methyltransferase that modifies hnRNPs, including Npl3p and Hrplp.
Afulg14110 YBR154C RNA polymerase subunit ABC27, common to RNA polymerases I, II, and III; contacts DNA and affects transactivation
Afu6g13600 YBR155W TPR-containing co-chaperone; binds both HspS2p (Hsp90) and Ssalp (Hsp70) and stimulates the ATPase activity of SSA1, is mutants reduce Hsp82p function.
Afu1g09770 YCL037C Cytoplasmic RNA-binding protein that associates with translating ri-bosomes; involved in heme regulation of Hap1p as a component of the HMC complex.
Afu5g02760 YCR034W Fatty acid elongase, involved in sphingolipid biosynthesis; acts on fatty acids of up to 24 carbons in length; mutations have regulatory effects on 1,3-beta-glucan synthase.
Afu5g05610 YCR063W Protein involved in bud-site selection; analysis of integrated high- throughput datasets predicts an involvement in RNA splicing.
Afu1g14220 YDL014W Nucleolar protein, component of the small subunit processome com-plex, which is required for processing of pre-18S rRNA; has similarity to mammalia fibrillarin
Afulg11170 YDL154W Protein of the MutS family, forms a dieter with Msh4p that facilitates crossovers between homologs during meiosis; Insh5-Y823H mutation confers tolerance to DNA alkylating agents.
Afu5g02160 YDL167C Protein of unknown function, rich in asparagine residues Afu5g02410 YDR021W Nucleolar protein required for maturation of 18S rRNA, member of the
eIF4A subfamily of DEAD-box ATP-dependent RNA helicases Afulg10310 YDR091C Essential iron-sulfur protein required for ribosome biogenesis and trans-
lation initiation;. Afu2g16820 YDR101C Shuttling pre-60S factor; involved in the biogenesis of ribosomal large
subunit biogenesis; interacts directly with Albl; responsible for Tif6 recycling defects in absence of Reil.
Afu6g13630 YDR124W Putative protein of unknown function; non-essential gene; expression is strongly induced by alpha factor
Afulg05070 YDR165W Subunit of a tRNA methyltransferase complex composed of Trm8p and Trm82p that catalyzes 7-methylguanosine modification of tRNA
Afu2g03930 YDR324C Nucleolar protein, component of the small subunit (SSU) processome containing the U3 snoRNA that is involved in processing of pre-18S rRNA
Afu5g10540 YELO11W Glycogen branching enzyme, involved in glycogen accumulation; green fluorescent protein (GFP)-fusion protein localizes to the cytoplasm in a punctate pattern
Afu6g09160 YEL012W Ubiquitin-conjugating enzyme that negatively regulates gluconeogen-esis by mediating the glucose-induced ubiquitination of fructose-1,6-bisphosphatase (FBPase).
continued on next page

Appendix E. Chapter 4 Supplementary Data 295
continued from previous page Accesion SC ORF Description
Afu2g08360
Afu4g07580
Afu5g07020
Afu3g13280
Afu7g05920
Afu2g06310
Afu5g06700
Afu2g04780
Afu6g11800
Afu3g13320
Afu6g07880
Afu2g16010
Afu8g04820
Afu5g13480
Afu2g12440
Afu4g07080
Afu6g11070
Afu6g13370
Afu6g11310
Afulg06810
Afu3g13480
Afulg04070
Afu8g01970
Afu5g13470
Afu7g02420
YEL021W
YER025W
YER036C
YFLOO2C
YGL055W
YGL078C
YGR123C
YGR145W
YGR213C
YGR214W
YHR016C
YHRO2OW
YHR062C
YIL061C
YIL103W
YIR026C
YJL033W
YJL109C
YJL130C
YJL200C
YJR007W
YJR047C
YJR153W
YKL009W
YKL029C
Orotidine-5'-phosphate (OMP) decarboxylase, catalyzes the sixth enzy-matic step in the de novo biosynthesis of pyrimidines, converting OMP into uridine monophosphate (UMP) Gamma subunit of the translation initiation factor eIF2, involved in the identification of the start codon; binds GTP when forming the ternary complex with GTP and tRNAi-Met ATPase of the ATP-binding cassette (ABC) family involved in 40S and 60S ribosome biogenesis, has similarity to Gcn20p. Putative ATP-dependent RNA helicase, nucleolar protein required for synthesis of 60S ribosomal subunits at a late step in the pathway; sed-iments with 66S pre-ribosomes in sucrose gradients Fatty acid desaturase, required for monounsaturated fatty acid synthe-sis and for normal distribution of mitochondria Putative ATP-dependent RNA helicase of the DEAD-box family in-volved in ribosomal biogenesis Protein serine/threonine phosphatase with similarity to human phos-phatase PPS; present in both the nucleus and cytoplasm; expressed during logarithmic growth Essential nucleolar protein of unknown function; contains WD repeats, interacts with MpplOp and Bfr2p, and has homology to Spblp Protein involved in 7-aminocholesterol resistance; has seven potential membrane-spanning regions Protein component of the small (40S) ribosomal subunit, nearly iden-tical to RpsOBp; required for maturation of 18S rRNA along with RpsOBp. Protein involved in the organization of the actin cytoskeleton; contains SH3 domain similar to Rys167p Protein of unknown function that may interact with ribosomes, based on co-purification experiments; has similarity to proline-tRNA ligase; YHRO2OW is an essential gene Subunit of both RNase MRP, which cleaves pre-rRNA, and nuclear RNase P, which cleaves tRNA precursors to generate mature 5' ends Component of Ul snRNP required for mRNA splicing via spliceosome; may interact with poly(A) polymerase to regulate polyadenylation; ho-molog of human U1 70K protein Protein required, along with Dph2p, Ktillp, Jjj3p, and Dphip, for syn-thesis of diphthamide, which is a modified histidine residue of transla-tion elongation factor 2 (Eft 1p or Eft2p). Protein phosphatase involved in vegetative growth at low temperatures, sporulation, and glycogen accumulation; transcription induced by low temperature and nitrogen starvation;. Putative nucleolar DEAD box RNA helicase; high-copy number sup-pression of a U14 snoRNA processing mutant suggests an involvement in 18S rRNA synthesis Nucleolar protein, component of the small subunit (SSU) processome containing the U3 snoRNA that is involved in processing of pre-18S rRNA Bifunctional carbamoylphosphate synthetase (CPSase)-aspartate tran-scarbamylase (ATCase), catalyzes the first two enzymatic steps in the de novo biosynthesis of pyrimidines Putative mitochondria] aconitase isozyme; similarity to Acolp, an aconitase required for the TCA cycle; expression induced during growth on glucose, by amino acid starvation via Gcn4p Alpha subunit of the translation initiation factor eIF2, involved in the identification of the start codon; phosphorylation of Ser51 is required for regulation of translation Translation initiation factor eIF-5A, promotes formation of the first peptide bond; similar to and functionally redundant with Hyp2. Endo-polygalacturonase, pectolytic enzyme that hydrolyzes the alpha-1,4-glycosidic bonds in the rhamnogalacturonan chains in pectins Protein involved in mRNA turnover and ribosome assembly, localizes to the nucleolus Mitochondria' malic enzyme, catalyzes the oxidative decarboxylation of malate to pyruvate, which is a key intermediate in sugar metabolism and a precursor for synthesis of several amino acids
continued on next page

Appendix E. Chapter 4 Supplementary Data 296
continued from previous page Accesion SC ORF Description
Afu6g04570 Afu6g02150
Afu5g09850 Afu2g02190
Afu3g12300
Afu1g03930
Afu2g12150
Afu3g06010
Afu2g13980
Afu6g12930
Afu2g13020
Afu3g04310
Afu6g02060
Afu4g09140
Afu5g05450
Afu4g12480
Afu3g08640
Afu2g16040
Afu6g13490
Afu6g02520
Afu2g02120
Afu3g05840
Afu3g06580
Afu8g03930
Afu3g05320
YKL081W YKL100C
YKL205W YLL035W
YLR061W
YLR067C
YLR106C
YLR186W
YLR276C
YLR304C
YLR348C
YLR409C
YLR43OW
YLR438W
YML063W
YML096W
YMR146C
YMR229C
YMR250W
YMR260C
YMR278W
YNL175C
YNL182C
YNL209W
YNL227C
Translation elongation factor EF-1 gamma Putative protein of unknown function with similarity to a human minor histocompatibility antigen; YKL100C is not an essential gene Nuclear pore protein involved in nuclear export of pre-tRNA Protein of unknown function, required for cell growth and possibly in-volved in rRNA processing; mRNA is cell cycle regulated Protein component of the large (60S) ribosomal subunit, has similarity to Rp122Bp and to rat L22 ribosomal protein Specific translational activator for the COX1 mRNA, also influences stability of intron-containing COX1 primary transcripts; located in the mitochondrial inner membrane Huge dynein-related AAA-type ATPase (midasin), forms extended pre-60S particle with the Rixl complex (Rix1p-Ipilp-Ipi3p), may mediate ATP-dependent remodeling of 60S subunits Protein required for the maturation of the 18S rRNA and for 40S ribo-some production; associated with spindle/microtubules; nuclear local-ization depends on physical interaction with Nopl4p ATP-dependent RNA helicase of the DEAD-box family involved in bio-genesis of the 60S ribosomal subunit Aconitase, required for the tricarboxylic acid (TCA) cycle and also inde-pendently required for mitochondrial genome maintenance; component of the mitochondria] nucleoid. Mitochondria] dicarboxylate carrier, integral membrane protein, cat-alyzes a dicarboxylate-phosphate exchange across the inner mitochon-drial membrane. Possible U3 snoRNP protein involved in maturation of pre-18S rRNA, based on computational analysis of large-scale protein-protein interac-tion data Putative helicase required for RNA polymerase II transcription termi-nation and processing of RNAs; homolog of Senataxin which causes Ataxia-Oculomotor Apraxia 2. L-ornithine transaminase (OTAse), catalyzes the second step of arginine degradation, expression is dually-regulated by allophanate induction and a specific arginine induction process. Ribosomal protein 10 (rp10) of the small (40S) subunit; nearly identical to RpslAp and has similarity to rat S3a ribosomal protein Putative protein of unknown function with similarity to asparagine syn-thetases; green fluorescent protein (GFP)-fusion protein localizes to the cytoplasm;. Subunit of the core complex of translation initiation factor 3(eIF3), which is essential for translation Protein required for the synthesis of both 18S and 5.8S rRNA; C-terminal region is crucial for the formation of 18S rRNA and N-terminal region is required for the 5.85 rRNA. Glutamate decarboxylase, converts glutamate into gamma- aminobutyric acid (GABA) during glutamate catabolism; involved in response to oxidative stress Translation initiation factor eIF1A, essential protein that forms a com-plex with Suilp (eIF1) and the 40S ribosomal subunit and scans for the start codon. Putative protein of unknown function; green fluorescent protein (GFP)-fusion protein localizes to the cytoplasm and nucleus; YMR278W is not an essential gene Protein of unknown function, localizes to the nucleolus and nucleo-plasm; contains an RNA recognition motif (RR/t4) and has similarity to Nopl2p. Essential component of the Rixl complex (Rix1p, Ipilp, Ipi3p) that is required for processing of ITS2 sequences from 35S pre-rRNA; highly conserved and contains WD40 motifs. Cytoplasmic ATPase that is a ribosome-associated molecular chaper-one, functions with J-protein partner Zuolp; may be involved in the folding of newly-synthesized polypeptide chains. Co-chaperone that stimulates the ATPase activity of Scalp, required for a late step of ribosome biogenesis; associated with the cytosolic large ribosomal subunit.
continued on next page

Appendix E. Chapter 4 Supplementary Data 297
continued from previous page Accesion SC ORF Description
Afu5g04240 YNL251C RNA-binding protein that interacts with the C-terminal domain of the RNA polymerase II large subunit (Rpo21p), required for transcription termination.
Afu1g07630 YNL255C Protein with seven cysteine-rich CCHC zinc-finger motifs, similar to human CNBP, proposed to be involved in the RAS/cAMP signaling pathway
Afu5g12310 YNL292W Pseudouridine synthase, catalyzes only the formation of pseudouridine- 55 (Psi55), a highly conserved tRNA modification, in mitochondria] and cytoplasmic tRNAs;.
Afu7g04130 YNL299W Poly (A) polymerase involved in nuclear RNA quality control based on, homology with Trf4p, genetic interactions with TRF4 mutants, physical interaction with Mtr4p (TRAMP subunit).
Afu2g05430 YNR012W Uridine/cytidine kinase, component of the pyrimidine ribonucleotide salvage pathway that converts uridine into UMP and cytidine into CMP.
Afu4g08930 YNR053C Putative GTPase that associates with pre-60S ribosomal subunits in the nucleolus and is required for their nuclear export and maturation
Afulg06690 YOR006C Putative protein of unknown function; green fluorescent protein (GFP)-fusion protein localizes to both the cytoplasm and the nucleus
Afulg16730 YOR243C Pseudouridine synthase, catalyzes pseudouridylation at position 35 in U2 snRNA, position 13 in cytoplasmic tRNAs, and position 35 in pre-tRNA(Tyr).
Afu4g07630 YOR272W Constituent of 66S pre-ribosomal particles, required for maturation of the large ribosomal subunit
Afulg02030 YOR361C Subunit of the core complex of translation initiation factor 3(eIF3), essential for translation.
Afu2g11810 YPL012W Protein required for export of the ribosomal subunits; associates with the RNA components of the pre-ribosomes; contains HEAT-repeats
Afu3g05570 YPL054W Zinc-finger protein of unknown function Afu7g02610 YPL126W LT3 snoRNP protein, component of the small (ribosomal) subunit (SSU)
processosome containing U3 snoRNA; required for the biogenesis of18S rRNA
Afu6g07490 YPL183C Cytoplasmic protein of unknown function Afu6g11510 YPL207W Protein required for the synthesis of wybutosine, a modified guano-
sine found at the 3'-position adjacent to the anticodon of phenylalanine tRNA.
Afu2g08780 YPRO95C Guanine nucleotide exchange factor (GEF) for Arf proteins; involved in vesicular transport; suppressor of ypt3 mutations; member of the Sec7-domain family
Afu4g07500 YPR137W Protein involved in pre-rRNA processing, associated with U3 snRNP; component of small ribosomal subunit (SSU) processosome; ortholog of the human U3-55k protein
Afu8g05430 YPR144C Nucleolar protein, forms a complex with Nopl4p that mediates matu-ration and nuclear export of 40S ribosomal subunits
Afu3g06540 YPR167C 3'-phosphoadenylsulfate reductase, reduces 3'-phosphoadenylyl sulfate to adenosine-3',5'-bisphosphate and free sulfite using reduced thiore-doxin as cosubstrate.

Appendix E. Chapter 4 Supplementary Data 298
Table E.7: A. fumigatus specific genes having elevated transcript abundance in the murine lung, relative to laboratory culture. The Affc-core set were defined as
A. fumigatus Af293 proteins that have orthologs in N. fischeri and A. clavatus. The Affc-
unique set is a sub-set of Affc-core proteins that do not have orthologs in A. terreus, A.
oryzae, A. nidulans or A. niger
Accesion Old Accession Protein name Location SignalP Process AFUA_IG00100 Afulg00100 MFS monocarboxylate transporter,
putative subtelomeric SignalP transport
AFUA_1G00450 Afulg00450 N-acetylglucosamine-6-phosphate deacetylase (NagA), putative
subtelomeric metabolism
AFUA_1G01580 Afulg01580 hypothetical protein unknown AFUA_1G05400 Afulg05400 hypothetical protein unknown AFUA_1G11030 Afulg11030 xylitol dehydrogenase metabolism AFUA_1G11240 Afulg11240 hypothetical protein unknown AFUA_1G15190 Afulg15190 hypothetical protein unknown AFUA_2G00490 Afu2g00490 glycosyl hydrolase, family 31 subtelomeric metabolism AFUA_2G00550 Afu2g00550 conserved hypothetical protein subtelomeric unknown AFUA_2G00700 Afu2g00700 hypothetical protein subtelomeric unknown AFUA_2G02330 Afu2g02330 hypothetical protein unknown AFUA_2G04530 Afu2g04530 hypothetical protein unknown AFUA_2G05300 Afu2g05300 hypothetical protein unknown AFUA_2G11530 Afu2g11530 hypothetical protein unknown AFUA_2G12740 Afu2g12740 methyltransferase, putative metabolism AFIJA_2017640 Afu2g17640 conserved hypothetical protein subtelomeric unknown AFUA_2G17910 Afu2g17910 conserved hypothetical protein subtelomeric SignalP unknown AFUA_2G18050 Afu2g18050 FAD binding oxidoreductase CpoX1 subtelomeric SignalP metabolism AFUA_2G18060 Afu2g18060 AdoMet:dimethylallyltryptophan subtelomeric metabolism
N-methyltransferase EasF AFUA_3G01060 Afu3g01060 hypothetical protein subtelomeric unknown AFIJA_3G01140 Afu3g01140 hypothetical protein subtelomeric unknown AFIJA_3001230 Afu3g01230 MFS sugar transporte, putative transport AFUA_3G01730 Afu3g01730 hypothetical protein unknown AFIJA_3G02110 Afu3g02110 MFS multidrug transporter, puta-
tive SignalP transport
AFIJA_3G02130 Afu3g02130 oxidoreductase, zinc-binding, puta- tive
metabolism
AFUA_3G02420 Afu3g02420 ThiJ/PfpI family transcriptional regulator, putative
transcriptional regulation
AFUA_3 G 03600 Afu3g03600 carboxylesterase, putative metabolism AFUA_3G03750 Afu3g03750 hypothetical protein unknown AFUA_3007090 Afu3g07090 hypothetical protein unknown AFUA_3G11880 Afu3g11880 hypothetical protein unknown AFUA_3G14740 Afu3g14740 conserved hypothetical protein subtelomeric unknown AFUA_4G00860 Afu4g00860 cell surface protein, putative subtelomeric cell wall biogenesis AFIJA_4006580 Afu4g06580 hypothetical protein unknown AFUA_4G06660 Afu4g06660 hypothetical protein unknown AFUA_4G08840 Afu4g08840 RING finger protein, putative SignalP unknown AFUA_4G13520 Afu4g13520 oxidoreductase, short-chain dehy-
drogenase/reductase family metabolism
AFIJA_5G00190 Afu5g00190 hypothetical protein subtelomeric unknown AFUA_5G00390 Afu5g00390 hypothetical protein subtelomeric unknown AFUA_5G00490 Afu5g00490 hypothetical protein subtelomeric unknown AFUA-5001410 Afu5g01410 hypothetical protein unknown AFLJA_5G01610 Afu5g01610 hypothetical protein unknown AFUA_5G02960 Afu5g02960 hypothetical protein unknown AFUA_5010350 Afu5g10350 conserved hypothetical protein SignalP unknown AFUA_5G12220 Afu5g12220 conserved hypothetical protein unknown AFLTA_5G13770 Afu5g13770 hypothetical protein unknown AFUA_5G14280 Afu5g14280 integral membrane protein subtelomeric unknown
continued on next page

Appendix E. Chapter 4 Supplementary Data 299
continued from previous page Accesion Old Accession Protein name Location SignalP Process
AFUA_5G14900 Afu5g14900 hypothetical protein subtelomeric unknown AFUA_6G00140 Afu6g00140 hypothetical protein subtelomeric SignalP unknown AFUA_6G00690 Afu6g00690 conserved hypothetical protein subtelomeric SignalP unknown AFUA_6G02290 Afu6g02290 hypothetical protein unknown AFUA_6003220 Afu6g03220 hypothetical protein unknown AFUA-6G07790 Afu6g07790 hypothetical protein unknown AFUA_6G10180 Afu6g10180 spherulin 4 family protein, putative unknown AFUA_6G10190 Afu6g10190 hypothetical protein unknown AFUA_6G11500 Afu6g11500 dipeptidase, putative metabolism AFUA_6G11950 Afu6g11950 hypothetical protein unknown AFUA-6G11990 Afu6g11990 hypothetical protein unknown AFUA_6G12100 Afu6g12100 nitrilase family protein metabolism AFUA-7G00790 Afu7g00790 hypothetical protein subtelomeric unknown AFUA_7G01020 Afu7g01020 hypothetical protein subtelomeric unknown AFUA_7G04560 Afu7g04560 hypothetical protein unknown AFUA_7G07130 Afu7g07130 hypothetical protein unknown AFUA_7G08450 Afu7g08450 ornithine decarboxylase, putative subtelomeric metabolism AFUA_8G00100 Afu8g00100 aspartate-tRNA ligase, putative subtelomeric metabolism AFUA_8G00490 Afu8g00490 PKS-like enzyme, putative subtelomeric metabolism AFUA_8000500 Afu8g00500 acetate-CoA ligase, putative subtelomeric metabolism AFUA_8G00810 Afu8g00810 hypothetical protein subtelomeric unknown AFUA_8G01460 Afu8g01460 hypothetical protein unknown AFUA_8G01650 Afu8g01650 hypothetical protein unknown AFUA_8G01950 Afu8g01950 hypothetical protein SignalP unknown AFUA_8G02260 Afu8g02260 neutral amino acid permease transport AFUA_8G05050 Afu8g05050 hypothetical protein unknown AFUA_8G05080 Afu8g05080 hypothetical protein unknown AFUA_8G05470 Afu8g05470 hypothetical protein unknown AFUA_8G05770 Afu8g05770 hypothetical protein unknown AFUA_8G05780 Afu8g05780 NACHT and Ankyrin domain pro-
tein signaling
AFUA_8G06640 Afu8g06640 ubiE/COQS methyltransferase, pu- tative
subtelomeric metabolism
Table E.8: A. fumigatus specific genes having repressed transcript abundance in the murine lung, relative to laboratory culture. The Affc-core set were defined
as A. fumigatus Af293 proteins that have orthologs in N. fischeri and A. clavatus. The
Affc-unique set is a sub-set of Affc-core proteins that do not have orthologs in A. terreus,
A. oryzae, A. nidulans or A. niger
Accesion Old Accession Protein name Location SignalP Process AFUAAG00270 Afulg00270 hypothetical protein subtelomeric unknown AFUA_1G00310 Afulg00310 class V chitinase, putative subtelomeric SignalP cell wall biogenesis AFUAAG01010 Afulg01010 polyketide synthase, putative metabolism AFUA_1G02220 Afu1g02220 hypothetical protein unknown AFUA_1G02960 Afu1g02960 conserved hypothetical protein unknown AFUA_1G03230 Afu1g03230 ABC multidrug transporter, puta-
tive transport
AFUA_1G03270 Afu1g03270 hypothetical protein unknown AFUA_1G17420 Afu1g17420 hypothetical protein subtelomeric unknown AFUA_2G01470 Afu2g01470 hypothetical protein unknown AFUA_2G05280 Afu2g05280 F-box domain protein unknown AFUA_2G06160 Afu2g06160 alpha-1,3-glucanase/mutanase, pu-
tative cell wall biogenesis
AFUA_2G09460 Afu2g09460 potassium transporter transport AFUA_2G16440 Afu2g16440 hypothetical protein unknown AFUA_3G02180 Afu3g02180 NRPS-like enzyme, putative metabolism AFUA_3007010 Afu3g07010 hypothetical protein unknown AFUA_3009120 Afu3g09120 hypothetical protein unknown AFUA_3G09510 Afu3g09510 3-oxoacyl-(acyl-carrier-protein) re-
duct ase metabolism
continued on next page

Appendix E. Chapter 4 Supplementary Data 300
continued from previous page Accesion Old Accession Protein name Location SignalP Process
AFUA_3009660 Afu3g09660 hypothetical protein unknown AFUA_3013710 Afu3g13710 GTP cyclohydrolase I, putative metabolism AFUA_3G13720 Afu3g13720 oxidoreductase, 20G-Fe(II) oxyge-
nose family, putative metabolism
AFUA_4G00510 Afu4g00510 hypothetical protein subtelomeric unknown AFUA_4G00850 Afu4g00850 hypothetical protein subtelomeric unknown AFUA_4G00880 Afu4g00880 hypothetical protein subtelomeric unknown AFUA_4G01430 Afu4g01430 hypothetical protein SignalP unknown AFUA_4G06810 Afu4g06810 hypothetical protein unknown AFUA_4G07920 Afu4g07920 hypothetical protein unknown AFUA_4G08860 Afu4g08860 hypothetical protein unknown AFUA_5G00170 Afu5g00170 extracellular serine threonine rich
protein, putative subtelomeric SignalP cell wall biogenesis
AFUA_5G00220 Afu5g00220 hypothetical protein subtelomeric unknown AFUA_5G00230 Afu5g00230 hypothetical protein subtelomeric unknown AFUA_5G00860 Afu5g00860 Ankyrin repeat protein subtelomeric unknown AFUA_5G01070 Afu5g01070 C6 transcription factor, putative subtelomeric transcriptional regulation AFUA_5G02710 Afu5g02710 hypothetical protein unknown AFUA_5G13870 Afu5g13870 mRNA export factor Mlo3, putative metabolism AFUA_5G15140 Afu5g15140 hypothetical protein subtelomeric unknown AFUA_6G00180 Afu6g00180 hypothetical protein subtelomeric unknown AFUA_6G00350 Afu6g00350 hypothetical protein subtelomeric unknown AFUA_6G03120 Afu6g03120 hypothetical protein unknown AFUA_6G09310 Afu6g09310 class V chitinase, putative SignalP cell wall biogenesis AFUA_6009340 Afu6g09340 hypothetical protein unknown AFUA_6G10150 Afu6g10150 hypothetical protein unknown AFUA_6G10350 Afu6g10350 hypothetical protein unknown AFUA_6G11630 Afu6g11630 FAD-dependent isoamyl alcohol ox-
idase, putative SignalP metabolism
AFUA_6G11700 Afu6g11700 hypothetical protein unknown AFUA_7G00400 Afu7g00400 hypothetical protein subtelomeric unknown AFUA_7G04750 Afu7g04750 hypothetical protein unknown AFUA_7G05710 Afu7g05710 hypothetical protein unknown AFUA_7G06000 Afu7g06000 hypothetical protein unknown AFUA_7G07040 Afu7g07040 hypothetical protein unknown AFIJA_7G07060 Afu7g07060 hypothetical protein unknown AFUA_7G07140 Afu7g07140 hypothetical protein subtelomeric unknown AFUA_7G08250 Afu7g08250 C6 finger domain protein, putative subtelomeric transcriptional regulation AFUA_7G08380 Afu7g08380 hypothetical protein subtelomeric unknown AFUA_7G08470 Afu7g08470 peroxisomal copper amine oxidase,
putative subtelomeric metabolism
AFUA_7G08520 Afu7g08520 hypothetical protein subtelomeric unknown AFIJA_SG02110 Afu8g02110 hypothetical protein unknown AFUA_8G06150 Afu8g06150 serine-threonine rich protein, puta-
tive unknown
AFIJA_8G06370 Afu8g06370 monooxygenase, putative metabolism
Table E.9: Affc-specific genes having elevated transcript abundance in the murine lung, relative to laboratory culture. The Affc-core set were defined as A. fumigatus Af293 proteins that have orthologs in N. fischeri and A. clavatus. The Affc-unique set is a sub-set of Affc-core proteins that do not have orthologs in A. terreus, A. oryzae, A. nidulans
or A. niger
Accesion Old Accession Protein name Location SignalP Process AFUA_1G04160
AFUAAG05910 AFUA_1G11370 AFUA_IG13410 AFUA_1G16740 AFUA_1G17370 AFUAAG17600
Afulg04160
Afulg05910 Afulg11370 Afulg13410 Afu1g16740 Afu1g17370 Afulg17600
aspartate aminotransferase, puta- tive conserved hypothetical protein GMC oxidoreductase, putative conserved hypothetical protein conserved hypothetical protein HSP9/12 family heat shock protein conserved hypothetical protein
subtelomeric subtelomeric
signalP
metabolism
unknown metabolism
unknown unknown unknown unknown
continued on next page

Appendix E. Chapter 4 Supplementary Data 301
continued from previous page Accesion Old Accession Protein name Location SignalP Process
AFUA_2G00580 Afu2g00580 conserved hypothetical protein subtelomeric unknown AFUA_2G00990 Afu2g00990 glycerophosphoryl diester phospho-
diesterase family protein subtelomeric metabolism
AFUA_2G02500 AFUA_2C12780
Afu2g02500 Afu2g12780
conserved hypothetical protein von Willebrand domain protein
unknown
AFUA_2G12850 Afu2g12850 1,3-beta-glucanosyltransferase Ge13 signalP cell wall biogenesis AFUA_2G17420 Afu2g17420 Pfs and NB-ARC domain protein subtelomeric signaling AFUA_2G17650 Afu2g17650 DUF907 domain protein subtelomeric signalP unknown AFUA_3G01130 Afu3g01130 cell wall protein, putative subtelomeric signalP cell wall biogenesis AFUA_3G01320 Afu3g01320 homocysteine S-methyltransferase,
putative metabolism
AFUA_3G02800 Afu3g02800 lipase/esterase, putative metabolism AFUA_3G12220 Afu3g12220 ABC transporter, putative transport AFUA_3G12610 Afu3g12610 conserved hypothetical protein unknown AFUA_3G13130 Afu3g13130 UBE domain protein metabolism AFUA_3G13290 Afu3g13290 conserved hypothetical protein unknown AFUA_3G14770 Afu3g14770 NAD dependent
epimerase/dehydratase family protein
subtelomeric signalP metabolism
AFUA_3G15080 Afu3g15080 conserved hypothetical protein subtelomeric unknown AFUA_4G01360 Afu4g01360 MFS transporter of unkown speci-
ficity transport
AFUA_4G01370 Afu4g01370 pyoverdine/dityrosine biosynthesis family protein
signaling
AFUA_4C01392 Afu4g01390 C6 transcription factor, putative transcriptional regulation AFUA_4G01470 Afu4g01470 C6 finger domain protein, putative transcriptional regulation AFUA_4G08390 Afu4g08390 conserved hypothetical protein unknown AFUA_4G13950 Afu4gI3950 GNAT family acetyltransferase, pu-
tative subtelomeric metabolism
AFUA_4014170 Afu4g14170 conserved hypothetical protein subtelomeric signalP unknown AFUA_4G14180 Afu4g14180 conserved hypothetical protein subtelomeric signalP unknown AFUA_4G14200 Afu4g14200 conserved hypothetical protein subtelomeric signalP unknown AFUA_5G00810 Ant500810 conserved hypothetical protein subtelomeric unknown AFUA_5G01130 Afu5g01130 conserved hypothetical protein subtelomeric unknown. AFUA.5G01300 Afu5g01300 integral membrane protein signalP unknown AFUA_5G08180 Afu5g08180 cell wall protein, putative signalP cell wall biogenesis AFUA.5G09130 Afu5g09130 polysaccharide deacetylase family
protein metabolism
AFUA-5G09440 Afu5g09440 amino acid permeate, putative transport. AFUA_6G00620 Afu6g00620 GPI anchored hypothetical protein subtelomeric cell wall biogenesis AFIJA-6003550 Afu6g03550 conserved hypothetical protein unknown AFUA_6004280 Afu6g04280 integral membrane protein signalP unknown AFUA-6G07260 Afu6g07260 purine-cytosine permease, putative transport AFUA_6G07920 Afu6g07920 acetyltransferase, GNAT family
family metabolism
AFUA_6G08500 Afu6g08500 phosphoglycerate mutate family protein
metabolism
AFUA_6008650 Afu6g08650 conserved hypothetical protein unknown AFUA_6G12150 Afu6g12150 bZIP transcription factor, putative transcriptional regulation AFUA_7G00690 Afu7g00690 aminotransferase, putative subtelomeric metabolism AFUA_7001000 Afu7g01000 aldehyde dehydrogenate, putative subtelomeric metabolism AFUA_7G04990 Afu7g04990 dUTP diphosphatase, putative uracil metabolism AFUA_7G05650 Afu7g05650 XYPPX repeat family protein unknown AFUA_8G00580 Afu8g00580 glutathione S-transferase, putative subtelomeric metabolism. AFUA_8000710 Afu8g00710 secreted antimicrobial peptide, pu-
tative subtelomeric signalP unknown
AFUA_8G01770 Afu8g01770 conserved hypothetical protein signalP unknown AFUA_8G05930 Afu8g05930 conserved hypothetical protein unknown
Table E.10: Affc-specific genes having repressed transcript abundance in the murine lung, relative to laboratory culture. The Affc-core set were defined as A.
fumigates Af293 proteins that have orthologs in N. fischeri and A. clavatus. The Affc-
unique set is a sub-set of Affc-core proteins that do not have orthologs in A. terreus, A.
oryzae, A. nidulans or A. niger

Appendix E. Chapter 4 Supplementary Data 302

Appendix E. Chapter 4 Supplementary Data 303
Accesion Old Accession Protein name Location SignalP Process
AFUA_1G12220 Afulg12220 conserved hypothetical protein unknown
AFITA_1G13120 Afulg13120 MIT/CorA family metal ion trans- porter
unknown
AFUA_2G02650 Afu2g02650 conserved hypothetical protein unknown
AFUA_3G09260 Afu3g09260 conserved hypothetical protein SignalP unknown
AFUA_3G12680 Afu3g12680 conserved hypothetical protein unknown AFUA_4G08890 Afu4g08890 aldo-keto reductase family protein metabolism
AFUA_5G10940 Afu5g10940 conserved hypothetical protein unknown
AFUA_6G00500 Afu6g00500 chitosanase, putative subtelomeric SignalP cell wall biogenesis
AFUA_7G06440 Afu7g06440 F-box domain protein unknown
AFUA_8G01820 Afu8g01820 hypothetical protein unknown
Table S6 Found at http : //ukpmc . ac .uk/picrender . cgi?artid=1663068&blobname=
ppat . 1000154 . s006 . doc Table S7 Found at http : //ukpmc . ac .uk/picrender . cgi?artid=1663068&blobnarne=
ppat 1000154 . s010.xls
Table E.11: Nitrogen starvation gene clusters
Cluster Number Accession Description log2 ratio Sub-telomeric Murine gene cluster
1 Afulg00500 FMN dependent dehydrogenase, putative
6.56
ZZ
z ZZ
z Z
ZZ
ZZ
ZZ
z ZZX
ZZ
ZZ
ZZ
ZZ
Z. <
Afulg00530 thermoresistant gluconokinase fam- ily protein
5.50
Afulg00550 conserved hypothetical protein 4.13 Afulg00560 Pfs domain protein 4.24 Afulg00570 Tel-mariner transposase, putative 2.25
2 Afu1g03280 maltose permease, putative 2.05 4 Afulg03300 carboxylesterase, putative 2.27 4
Afu1g03330 hypothetical protein 4.98 Afulg03340 hypothetical protein 3.75 4 Afu1g03350 alpha-1,3-glucanase, putative 2.41 4 Afu1g03360 hypothetical protein 3.00
3 Afulg16020 hypothetical protein 3.82 Afulg16030 conserved hypothetical protein 4.82 Afulg16050 conserved serine-proline rich pro-
tein 5.07
Afulg16060 conserved hypothetical protein 6.13 Afulg16080 hypothetical protein 5.04 Afulg16110 arsenic methyltransferase Cyt19,
putative 3.16
Afu1g16130 copper-transporting ATPase, puta- tive
3.71
Afulg16170 hypothetical protein 2.71 4 Afu3g02520 MSF multidrug transporter, puta-
tive 4.76
Afu3g02550 conserved hypothetical protein 4.91 Afu3g02570 polyketide synthase, putative 2.48 Afu3g02600 hypothetical protein 4.54 Afu3g02610 MFS transporter, putative 3.85 Afu3g02640 nucleoside-diphosphate-sugar
epimerase family protein 2.24
5 Afu3g03280 FAD binding monooxygenase, puta- tive
2.50 33
Afu3g03290 hypothetical protein 2.89 Afu3g03330 mitochondria] enoyl reductase, pu-
t at ive 6.03
Afu3g03350 nonribosomal peptide synthase, pu- tative
5.84 33
Afu3g03360 hypothetical protein 5.29 6 Afu3g13620 conserved hypothetical protein 2.38
continued on next page

Appendix E. Chapter 4 Supplementary Data 304
continued from previous page Cluster Number Accession Description log2 ratio Sub-telomeric Murine gene cluster
Afu3g13640 extracellular serine-rich protein 2.12 N Afu3g13650 integral membrane protein 2.17 N Afu3g13660 Ctr copper transporter family pro-
tein 3.61 N
Afu3g13670 siderochrome-iron transporter, pu- tative
5.30 N
Afu3g13680 conserved hypothetical protein 5.04 N Afu3g13700 transferase family protein 3.47 N
7 Afu4g00490 conserved hypothetical protein 4.21 Y Afu4g00510 hypothetical protein 3.72 Y Afu4g00530 conserved hypothetical protein 3.40 Y Afu4g00550 membrane transport protein, puta-
tive 4.09 Y
Afu4g00580 hypothetical protein 2.78 Y Afu4g00590 monooxygenase, putative 4.31 Y Afu4g00600 integral membrane protein 4.32 Y Afu4g00610 aryl-alcohol dehydrogenase, puta-
tive 4.67 Y
8 Afu4g14000 tripeptidyl peptidase A 3.51 Y Afu4g14040 Hsp70 family protein 4.50 Y 44 Afu4g14060 conserved hypothetical protein 2.52 Y Afu4g14070 glycosyl transferase, putative 5.61 Y 44
Afu4g14080 cell surface antigen spherulin 4, pu- tative
2.37 Y 44
Afu4g14090 UDP-glucose 4-epimerase 3.36 Y 44
9 Afu4g14160 hypothetical protein 3.97 Y Afu4g14170 hypothetical protein 3.39 Y 44
Afu4g14180 hypothetical protein 2.58 Y 44
Afu4g14190 hypothetical protein 2.47 Y Afu4g14230 MFS transporter, putative 3.17 Y Afu4g14240 0-methyltransferase 2.95 Y Afu4g14250 hypothetical protein 2.62 Y Afu4g14290 conserved hypothetical protein 2.40 Y
10 Afu5g03690 CRAL/TRIO domain protein 2.33 N Afu5g03710 conserved hypothetical protein 2.93 N Afu5g03750 WW domain protein 3.78 N Afu5g03760 class III chitinase ChiAl 2.92 N Afu5g03800 high-affinity iron permease 2.19 N
CaFTR2 11 Afu5g14860 cytochrome P450 4.64 Y
Afu5g14870 protein kinase, putative 5.27 Y Afu5g14880 hypothetical protein 5.67 Y Afu5g14900 hypothetical protein 5.56 Y Afu5g14920 hypothetical protein 3.39 Y. Afu5g14930 conserved hypothetical protein 4.72 Y Afu5g14940 cell surface metalloreductase 5.13 Y
(FreA), putative Afu5g14960 hypothetical protein 5.83 Y Afu5g14970 conserved hypothetical protein 4.82 Y Afu5g14990 hypothetical protein 4.36 Y Afu5g15010 arsenite permease (ArsB), putative 3.42 Y Afu5g15040 hypothetical protein 3.06 Y Afu5g15070 AhpC/TSA family thioredoxin per-
oxidase, putative 2.41 Y
Afu5g15080 Rho-associated protein kinase, pu- tative
2.71 Y
12 Afu6g00230 isoflavone reductase family protein 4.53 Y Afu6g00260 phosphatidylserine decarboxylase
family protein 3.98 Y
Afu6g00280 isoflavone reductase family protein 2.02 Y Afu6g00290 aminotransferase, putative 2.27 Y Afu6g00310 serine carboxypeptidase (CpdS),
putative 2.48 Y
continued on next page

Appendix E. Chapter 4 Supplementary Data 305
continued from previous page Cluster Number Accession Description log2 ratio Sub-telomeric Murine gene cluster
13 Afu6g00550 glycosyl transferase family 8 family, putative
3.19
zzzzzzz z Z
>.>
.?.
>..)-.}.Z
ZZ
ZZ
Afu6g00560 hypothetical protein 2.76
Afu6g00600 hypothetical protein 2.22 55
Afu6g00610 hypothetical protein 2.25
Afu6g00620 GPI anchored hypothetical protein 2.71
Afu6g00630 MFS transporter, putative 2.19 55
Afufig00640 integral membrane protein 3.14 55
14 Afu6g03390 glutathione S-transferase family protein
2.86
Afu6g03410 2.15
Afu6g03420 trehalose synthase, putative 2.21
Afu6g03430 C6 finger domain protein, putative 3.62 58
Afu6g03440 fructosyl amine 5.72 58
Afu6g03450 conserved hypothetical protein 4.33 58
Afu6g03460 hypothetical protein 4.00 58
Afu6g03470 ABC multidrug transporter, puta- tive
3.49 58
Afu6g03480 nonribosomal peptide synthetase, putative
3.19 58
Afu6g03490 phenol 2-monooxygenase, putative 4.08 58
15 Afu7g08520 hypothetical protein 6.35
Afu7g08530 hypothetical protein 4.07 Afu7g08540 ankyrin repeat protein 4.41
Afu7g08560 hypothetical protein 2.10
Afu7g08590 hypothetical protein 5.07 Afu7g08600 hypothetical protein 3.96
16 Afu8g06050 salicylate hydroxylase, putative 3.49
Afu8g06060 conserved hypothetical protein 4.18
Afu8g06070 hypothetical protein 4.21
Afu8g06090 amino acid permease, putative 3.32
Afu8g06100 integral membrane protein 3.19
Table E.12: Oligonucleotides used in qPCR
Oligo Name Oligo Sequence
Afu3g03080F ATACTIGGCGCATCGICTGG Afu3g0308OR ACGGCCACGTTGAGAATGAA Afu2g07770F TACGATGTCGTTCGCATGCT Afu2g0777OR TCACAGGATGACGCACTTGAT Afu3g12060F GTGTIGAAGAGGCCCTCAAG Afu3g1206OR GGGCCAGATATCGTTAGCAAT Afulg12310F CTATGCCCGCAAGCATTTCA Afu lgl 231 0 R TCTTGGAGGACATCGATCCAT Aful g16890F CCAGATGATGTGCCCGTGAA Afulg16890R CGGCTGGAATAGATTCCTTC Afu2g07910F TTIGGGTTCTATGCTGGGAT Afu2g0791OR GCTTCTGCCTCTCCTGAAGTA Afu2g06210F AGTTCGGCCGGCTTTGGTA Afu2g0621OR TGAATACAAAGGGGACGCAA Afulg15970F GGATTGATTGCAATCCCAGG Afulg1597OR TAATCACCGACCATCGATTG Afu2g05360F CGGATTTCGATTCACGAAGC Afu2g05360R TGGTATACATGCTCTCCGTGT Afu2g1499 OF TCGACCTCATGTACTCGAAG Afu2g 1499 OR CCTTCCTCCTCTGGTTCCATA Afu6g09610F ATGCGCCAT9CTCGAGATCA Afu6g096 lOR GCGAGACCTTCAAGGCATT Afu6g09620F CGACGATAAGTTCGCGCCGT Afu6g0962OR AAAATCTOCCGGICCGTGAAA Afu6g09630F TCTITGAACCCTATGATCCG Afu6g0963OR AGTAGGTTGCACAATCGAGCC

E.1 Dataset Si reference 306
E.1 Dataset S1 reference
Found at http : //ukpmc . ac . uk/picrender . cgi?artid=1663068&blobname=ppat .1000154 .
s013 .xls
E.2 Dataset S2 reference
Found at http : //ukpmc . ac .uk/picrender. cgiTartid=1663068&blobname=ppat . 1000154 .
s014.xis
E.3 Dataset S3 reference
Found at http://ukpmc.ac.uk/picrender . cgilart id=1663068&blobname=ppat .1000154.
s015 . xls
E.4 Dataset S4 reference
Found at http: //ukpmc . ac .uk/picrender . cgi?artid=1663068&blobname=ppat . 1000154 .
s016.xls

307
Appendix F
Chapter 5 Supplementary Data
Table F.1: Summary of null mutant CGTFKOLib strain growth relative to his3
parental strain in YEPD and YEPD + 5 - 10 pg m1-1 fluconazole..
Gene Name YEPD % Control
YEPD % SEM
5 pg ml-' fluconazole % Control
5 pg ml-' fluconazole SEM
10 pg ml-1 fluconazole % Control
10 pg ml-1 fluconazole SEM
ASF1 13.91 2.67 16.42 4.19 34.03 1.53
POPS 17.95 5.84 9.42 5.18 9.02 13.94 SNF6 21.13 4.26 8.33 2.25 9.02 5.07
SPT3 21.64 5.73 17.88 4.33 26.05 5.48 GCN5 38.13 5.54 10.93 4.83 14.49 11.15
SNF5 42.15 10.73 31.20 8.04 56.51 15.64 NGGI 44.48 5.96 12.70 4.02 17.53 7.64 SSN6 53.85 2.87 75.27 14.12 81.45 66.53 RTF1 55.12 6.41 46.09 14.75 45.11 43.83
TAFf4 61.76 2.98 42.47 9.86 49.16 1.77 ADA2 67.45 12.59 21.28 3.80 36.13 6.67
HAT2 68.63 8.82 49.45 18.46 41.68 24.08 SFLI 69.73 7.30 93.88 10.50 106.30 5.98 EMIT 70.95 11.04 84.84 12.87 176.05 2.29 SSN8 72.50 22.60 75.80 18.85 182.77 25.54 RPD3 72.84 4.79 72.83 6.08 87.55 45.68 GLN3 74.42 17.92 76.11 18.27 108.77 69.38 AR080 76.49 11.71 61.14 27.55 50.70 37.99 CTI6 78.00 11.80 48.28 21.00 36.85 35.50 SPT8 78.40 15.92 55.93 16.21 67.47 24.24 SSN2 79.17 7.42 90.86 23.93 91.60 17.97 MSNI 79.98 8.71 52.40 21.85 48.92 31.92 MKS1 80.07 15.21 96.31 20.83 105.46 23.74 CST6 80.15 6.30 97.86 8.22 204.20 25.54
SRB8 81.02 6.24 53.91 8.00 34.94 12.80
HOST 81.89 13.64 57.70 26.35 49.30 66.02 M1G/ 83.24 4.60 63.84 10.51 54.38 8.15
FKH2 83.97 18.76 92.61 20.59 210.50 39.12
BA S1 84.05 15.83 89.70 15.13 203.57 40.92 TECI 85.92 21.25 96.60 15.45 170.59 10.97 SFP1-like (not OT- tholoque)
86.18 14.50 62.07 21.56 53.88 40.11
SUT2 87.55 5.77 108.84 19.16 224.79 53.70
SINS 88.09 5.47 97.39 5.52 121.98 6.17
STP2 88.40 9.86 100.84 18.05 128.97 13.72
continued on next page

Appendix F. Chapter 5 Supplementary Data 308
continued from previous page Gene Name YEPD %
Control YEPD % SEM
5 pg m1-1 fluconazole % Control
5 pg ml-1 fluconazole SEM
10 pg m1-1 fluconazole % Control
10 ttg ml-1 fluconazole SEM
GAT1 89.91 1.60 101.36 14.45 171.43 36.37 PHO2 90.15 12.09 105.83 22.16 241.39 54.89 ASG1 90.55 10.15 86.46 13.45 84.50 29.22 HS T3 91.02 8.38 80.32 15.73 71.66 35.25
ACE2 91.13 17.71 105.93 14.74 212.82 0.39 SPO1 91.74 7.65 95.46 18.26 130.88 20.23
MSN4 91.94 4.25 111.76 17.46 211.97 48.97 CG2001 92.03 15.96 103.50 9.62 100.00 30.59 CRZI 92.03 19.90 109.91 19.12 223.74 0.76 SNF4-like 92.43 4.41 107.48 21.11 133.19 24.31
FAP1 93.00 14.58 113.61 16.69 226.89 1.66
PHO23 93.17 11.06 93.61 21.41 131.77 57.17 STP/ 93.25 7.37 110.20 8.56 187.61 27.15
PUT3 93.49 13.39 112.73 16.80 207.14 41.19
YAP6 93.82 26.64 117.30 26.47 224.79 14.34
RFX1 93.88 13.84 82.00 13.72 72.30 59.19
MSN2 94.12 5.12 96.47 11.26 127.06 21.77
SWD/ 94.36 4.92 97.56 6.60 100.76 22.64
FKH1 94.44 8.72 94.79 17.03 126.30 42.09 SPP I 94.79 6.56 107.19 10.59 78.15 13.22
SIPI 95.00 4.38 92.68 18.02 122.74 20.97 CATS 95.04 16.27 112.93 18.80 151.68 10.40
/ME/ 95.36 16.39 107.87 13.75 139.92 17.06
RTG2 95.63 10.07 100.76 19.43 80.94 7.04 RIM101 95.71 6.74 98.91 11.65 134.43 26.67
SSN3 95.85 10.37 83.09 12.67 151.47 30.63
.51P4 96.01 16.59 117.69 18.64 197.06 31.21
GAL83 96.27 4.71 102.61 2.82 85.13 4.05 SUT/ 96.58 26.40 117.01 28.90 209.87 52.87
OAFI 96.58 10.07 120.80 10.55 237.82 35.44 RGT1 96.66 3.93 117.20 18.34 163.24 32.49
SMP1 96.66 14.75 117.40 15.50 223.95 33.32
TUP1 96.82 6.45 108.33 17.11 124.65 57.94 HA P1-like 96.83 14.13 77.16 10.94 42.86 0.59
LYS 14 96.83 3.68 100.29 8.53 110.92 3.87
GIS2 96.83 6.90 119.24 16.25 206.72 39.23
EMI5 96.91 14.06 116.72 28.56 159.03 41.60
YRR1 96.91 6.57 119.14 6.45 102.10 5.46
MBP1 97.06 6.59 104.79 12.59 144.09 47.80 DAL80 97.15 8.41 120.89 24.60 202.10 44.99 GAL 1 1 97.22 8.03 84.95 17.56 74.97 36.94
DAL82 97.40 17.10 115.16 14.72 211.34 22.27
HCM1 97.40 8.06 117.20 10.66 221.22 28.76
HOT I 97.46 2.03 96.72 12.01 136.72 14.53
BRE2 97.46 14.34 100.76 16.54 107.12 47.91
KA R4 97.54 11.24 94.45 18.58 118.81 33.08
ASK10 97.54 13.73 100.17 16.73 139.90 27.52
RPH1 97.56 21.82 76.97 17.76 102.73 12.10
RTG1 97.62 7.12 87.80 19.52 75.22 26.88
SDS3 97.64 15.35 106.12 17.23 184.03 42.39
HOS2 97.70 4.64 101.35 10.54 128.08 20.26 GZF3 97.86 16.61 100.42 10.69 108.26 5.76
HDA 1 97.97 11.54 118.17 20.74 176.47 43.53 MCM1-like. 98.05 8.07 117.40 23.93 182.98 45.01
FZF1 98.09 8.57 102.02 15.69 108.39 34.41
YR A/ / 98.21 26.55 109.91 20.85 155.46 4.80 HAP2 98.25 13.54 99.58 31.33 146.12 45.56
HMS1 98.33 10.05 99.07 22.18 141.68 36.12
SKN7 98.33 14.08 103.20 19.90 132.53 44.06
HIR1 98.33 4.56 104.21 9.94 106.23 28.94
continued on next page
Appendix F. Chapter 5 Supplementary Data 309
continued from previous page Gene Name YEPD %
Control YEPD % SEM
5 og ml-' fluconazole % Control
5 og ml-1 fluconazole SEM
10 og m1-1 fluconazole % Control
10 og ml-' fluconazole SEM
YAPS 98.37 12.15 104.96 15.52 151.05 22.04
ND T80 98.45 12.66 117.49 12.78 222.06 37.49
HIR2 98.49 4.43 106.56 12.98 151.72 26.05 CHA4 98.62 15.32 117.40 13.41 183.82 11.50
CG2001 98.65 8.45 74.10 25.38 65.69 46.33
PPR I 98.65 9.28 102.69 20.07 144.09 62.49
UMEI 99.02 20.02 110.50 15.14 187.18 4.17 AZF I-LIKE 99.10 9.30 119.24 12.62 189.29 29.53 TEC1-L IKE 99.13 3.37 94.28 8.99 70.78 27.82
TOSS 99.13 11.97 98.74 20.76 139.26 30.66 SIPS 99.19 12.87 117.49 17.15 168.07 29.77
STE 12-LIKE 99.19 9.19 119.24 22.42 220.38 44.92
PIPS 99.35 35.39 120.41 38.24 216.39 85.41 XBP1 99.36 10.98 94.20 15.70 93.14 23.34 SASS 99.43 13.35 116.62 19.78 234.24 44.06
ASH1 99.44 4.41 80.24 11.33 82.59 15.09 AFT2 99.44 11.53 100.25 22.55 112.07 108.38 IMPS' 99.52 17.18 86.29 28.99 ' 75.73 82.36
MO T3 99.59 5.69 114.87 19.10 196.22 44.17 Z MS 1 99.67 7.61 120.21 18.53 210.71 46.72
CUPS 99.92 5.23 101.18 6.09 99.11 22.90 HST4 99.92 5.18 103.20 12.68 141.55 22.78
SOK 2 99.92 11.33 114.38 9.78 166.39 1.13 his3 100.00 ' 0.00 100.00 0.00 100.00 0.00
his3 100.00 0.00 100.00 0.00 100.00 0.00 TUP1 -lac (not ortholog)
100.08 9.02 103.62 24.95 160.99 197.00
PDR1 100.16 5.13 28.34 5.07 35.96 12.52 POG1 100.16 10.78 101.77 20.18 141.55 30.84 GCRS 100.24 19.19 98.07 21.96 129.35 43.64
CAD I 100.24 4.24 105.97 14.86 106.23 30.63
NOT3 100.24 26.37 112.54 25.40 236.34 35.83 SPT2 100.33 6.99 114.77 12.12 224.37 46.25
SASS 100.40 13.54 82.84 26.96 63.91 62.41 SKOI 100.41 7.15 100.00 15.10 91.39 13.79 HAA I 100.41 9.94 118.46 25.71 224.58 54.54 SEE/ 100.48 1.33 98.32 0.46 129.73 24.14 /MG/ 100.49 23.58 115.35 25.69 219.75 5.29
HAP5 100.65 9.04 119.34 9.00 96.01 1.18 GTS1 100.73 14,54 108.45 7.29 142.44 3.89 THIS 100.73 14.88 114.38 14.75 200.00 8.22 SW15 100.87 14.28 102.02 29.53 131.51 42.54
RME1 100.90 5.38 116.81 10.75 125.84 4.50 YA P7 100.95 5.73 99.33 15.08 125.67 11.29 INOS 101.19 8.51 88.14 18.43 76.37 42.23 CUPS 101.19 13.38 103.03 20.24 120.46 35.80 METS8 101.22 15.77 114.77 10.62 169.12 12.08 MET32 101.27 4.80 99.07 3.91 95.04 2.02 TSPI 101.27 10.85 105.55 12.55 129.73 20.71 SNF4 101.43 9.98 106.64 20.14 128.46 37.26
HA P4 101.46 10.43 116.13 21.82 180.46 40.52 RLM I 101.67 7.19 99.92 16.79 124.02 40.45 SASS 102.12 21.25 118.46 28.01 212.61 59.91
CHD1 102.12 5.20 120.21 5.35 176.05 25.11 HA PI 102.22 10.47 96.55 15.54 85.01 32.51
ZAP I 102.28 23.67 101.36 19.54 104.62 39.10
A HC1 102.46 0.99 105.30 3.96 117.15 2.25
MCM/ 102.62 21.13 100.76 40.07 101.78 57.48
GIS I 102.70 10.20 103.11 19.60 132.53 32.28
WARI 102.70 5.87 104.63 8.55 127.45 17.89 continued on next page

Appendix F. Chapter 5 Supplementary Data 310
continued from previous page Gene Name YEPD %
Control YEPD % SEM
5 pg ml - I fluconazole % Control
5 pg ml -1 fluconazole SEM
10 pg m1-1 fluconazole % Control
10 pg ml-1 fluconazole SEM
MIG2 102.77 '5.16 115.16 16.30 175.84 26.65 EZL I 102.94 5.05 109.42 18.64 144.98 60.25 HIR3 103.09 12.66 108.94 19.31 179.62 42.30 STE12 103.10 4.53 100.34 13.67 86.66 27.35 UPC2 103.10 7.21 104.29 12.38 146.63 21.96 YOX1 103.18 7.80 92.85 8.96 95.55 45.31 HAC1 103.25 5.22 110.50 10.73 167.65 20.26 HATI 103.25 12.07 119.53 18.33 226.47 50.15 CAL//-like 103.34 5.94 54.33 10.66 62.90 5.96 ST135 103.42 9.04 104.54 17.37 124.02 24.91 HALO 103.66 7.32 123.32 11.58 210.71 42.16 MET3I 103.82 4.87 117.78 6.31 219.75 38.32 RDS1 103.89 7.75 97.22 11.53 133.04 17.74 DA L81 103.97 10.18 110.34 23.61 155.91 49.06 H033 103.99 6.97 93.97 15.58 118.70 16.28 ARC81 104.37 5.84 103.62 16.46 133.42 39.36 ADR1 104.45 12.77 99.50 21.48 107.37 21.67 MILS 104.45 3.85 101.68 6.38 80.05 21.09 HS TI 104.53 16.79 105.30 20.66 133.42 33.95 RDS2 104.64 30.59 120.70 21.05 245.38 20.72
FL O8 104.69 12.98 112.45 26.33 154.89 52.86 YAP3-like 104.80 10.88 115.94 7.77 216.39 29.89 HA P3 104.80 4.64 121.28 15.18 80.46 1.93 HS T2 105.04 12.67 117.78 26.33 210.08 54.73
PHO4 105.16 9.54 104.21 15.07 120.84 12.90 SFPI 105.29 22.08 95.92 19.42 173.95 4.35 STB1 105.29 32.50 113.80 35.65 219.12 18.97 TYE7 105.40 5.96 108.49 15.90 143.33 26.19
S/H4 106.02 26.35 114.87 24.86 188.24 55.97
TOS4 106.04 5.00 103.11 13.15 97.84 11.97
SPT4 106.92 21.30 115.65 18.12 158.40 28.25 MSS1 / 107.23 14.76 116.32 27.27 155.27 72.20
RTG3 107.97 12.96 114.48 18.12 159.45 4.30

Appendix F. Chapter 5 Supplementary Data 311
Table F.2: Summary of null mutant strains showing insufficent growth in YEPD (0-11.5 hours) for nonlinear regression based estimation of midpoint and upper asymptote
CG ORF SC ORF Gene Name Description CAGLOB01441g YNL330C RPD3 Histone deacetylase; regulates tran-
scription and silencing CAGL00O2277g YER040W GLN3 transcriptional activator of
nitrogen-regulated genes CAGL0003399g YNR052C POP2 transcription factor (putative) CAGLOD01364g YBR112C SSN6 General repressor of transcription CAGLOE00693g YDR176W NGG1 Involved in glucose repression of
GAL4p-regulated transcription; transcription factor; genetic and mutant analyses suggest that Ngglp (Ada3p) is part of two tran- scriptional adaptor/HAT (histone acetyltransferase complexes, the 0.8 MD ADA complex and the 1.8 MD SAGA complex
CAGLOE03718g YHL025W SNF6 RNA polymerase II transcription factor activity
CAGLOF08283g YGR252W GCN5 Ada2/Gcn5/Ada3 transcription ac-tivator complex
CAGLOH07711g YGL244W RTF1 Subunit of the RNA polymerase II-associated Pafl complex; directly or indirectly regulates DNA-binding properties of Sptl5p and relative activities of different TATA ele- ments
CAGLOL02123g YBR289W SNF5 RNA polymerase II transcription factor activity
CAGLOL05412g YJL115W ASF1 Nucleosome assembly factor, in- volved in chromatin assembly and disassembly, anti-silencing protein that causes derepression of silent loci when overexpressed
CAGLOM01540g YDR392W SPTS transcription factor
Table F.3: Summary of null mutant CGTFKOLib strains showing insufficent growth in YEPD 5//gm1-1 (0-11.5 hours) for nonlinear regression based es- timation of midpoint and upper asymptote. Strains above the line show a general
growth defect in YEPD and can also be found in Table F.2
CG ORF SC ORF Gene Name Description CAGLOE03718g YHL025W SNF6 RNA polymerase II transcription
factor activity CAGL0C03399g YNR052C POPS transcription factor (putative) CAGLOF08283g YGR252W GCN5 Ada2/Gcn5/Ada3 transcription ac-
tivator complex CAGLOE00693g YDR176W NGG1 Involved in glucose repression of
GAL4p-regulated transcription; transcription factor; genetic and mutant analyses suggest that Ngglp (Ada3p) is part of two tran- scriptional adaptor/HAT (histone acetyltransferase complexes, the 0.8 MD ADA complex and the 1.8 MD SAGA complex
continued on next page

Appendix F. Chapter 5 Supplementary Data 312
continued from previous page CG ORF SC ORF Gene Name Description
CAGLOL05412g
CAGLOM01540g CAGL0L02123g
CAGLOH07711g
CAGLOE00627g CAGLOB01441g
CAGLOD01364g CAGL00O2277g
YJL115W
YDR392W YBR289W
YGL244W
YCRO81W YNL330C
YBR112C YER040W
ASFI
SPT3 SNF5
RTF1
SRB8 RPD3
SSNG GLN3
Nucleosome assembly factor, in- volved in chromatin assembly and disassembly, anti-silencing protein that causes derepression of silent loci when overexpressed transcription factor RNA polymerase II transcription factor activity Subunit of the RNA polymerase II-associated Pafl complex; directly or indirectly regulates DNA-binding properties of Spt15p and relative activities of different TATA ele- ments transcription factor complex Histone deacetylase; regulates tran-scription and silencing General repressor of transcription transcriptional activator of nitrogen-regulated genes
CAGLOK06193g
CAGLOA00451g
CAGLOM02739g CAGL0001485g CAGLOB03575g CAGLOE02079g CAGLOF01837g CAGLOD01430g
CAGLOF03025g
CAGLOA01628g
CAGLOM06875g CAGLOL11880g
CAGLOL12650g CAGLOL02585g
CAGLOL10472g
YDR448W
YGL013C
YPL129W YPL181W YEL056W YOL116W YLR055C YPRO68C
YDR421W
YGL035C
YNL025C YER169W
YPL042C YKR099W
YDR443C
ADA2
PDR1
TAF14 OT/6 HAT2 MSNI SPT8 HOST
ARO8O
MIGI
SSN8 RPH1
SSN3 BAS/
SSN2
transcription factor, member of ADA and SAGA, two transcrip- tional adaptor/HAT (histone acetyltransferase)complexes zinc finger transcription factor of the Zn(2)-Cys(6) binuclear cluster domain type transcription factor TFIID complex transcription factor binding Histone acetyltransferase subunit 2, transcriptional activator transcription factor Putative class I histone deacetylase (HDAC) with sequence similarity to Hdalp, Rpd3p, Hos2p, and Hos3p; deletion results in increased histone acetylation at rDNA repeats; specific RNA polymerase II tran-scription factor activity transcription factor involved in glu- cose repression; C2H2 zinc finger protein which resembles the mam-malian Egr and Wilms tumour pro-teins transcription factor complex JmjC domain-containing histone demethylase which can specifi- cally demethylate H3K36 tri- and dimethyl modification states; tran-scriptional repressor of PHR1 transcription factor complex Myb-related transcription factor in-volved in regulating basal and in-duced expression of genes of the purine and histidine biosynthesis pathways Required for stable association of SrblOp-Srbllp kinase with RNA polymerase holoenzyme; regulates YGP 1 epxression; component of RNA polymerase II holoenzyme and Kornberg's mediator (SRB) sub- complex; transcription factor
continued on next page

Appendix F. Chapter 5 Supplementary Data 313
continued from previous page CG ORF SC ORF Gene Name Description
CAGLOK05841g YLR256W-like HARI-like Zinc finger transcription factor in-volved in the complex regulation of gene expression in response to levels of heme and oxygen
Table F.4: Summary of null mutant CGTFKOLib strains showing insufficent growth in YEPD 10pgm1-1 (0-11.5 hours) for nonlinear regression based estimation of midpoint and upper asymptote. Strains above the line show a general
growth defect in YEPD and can also be found in Table F.2
CC ORF SC ORF Gene Name Description CAGL0003399g YNR052C POP2 transcription factor (putative) CAGLOF08283g YGR252W GCN5 Ada2/Gcn5/Ada3 transcription ac-
tivator complex CAGLOE00693g YDR176W NGGI Involved in glucose repression of
GAL4p-regulated transcription; transcription factor; genetic and mutant analyses suggest that Ngglp (Ada3p) is part of two tran- scriptional adaptor/HAT (histone acetyltransferase complexes, the 0.8 MD ADA complex and the 1.8 MD SAGA complex
CAGLOK06193g YDR448W ADA2 transcription factor, member of ADA and SAGA, two transcrip- tional adaptor/HAT (histone acetyltransferase)complexes
CAGLOL02123g YBR289W SNF5 RNA polymerase II transcription factor activity
CAGLOA00451g YGL013C PDRI zinc finger transcription factor of the Zn(2)-Cys(6) binuclear cluster domain type
CAGL0001485g YPL181W CTI6 transcription factor binding CAGLOB03575g YEL056W HAT2 Histone acetyltransferase subunit 2 CAGLOE03718g YHL025W SNF6 RNA polymerase II transcription
factor activity—Function— CAGL0D01430g YPRO68C HOS1 Putative class I histone deacetylase
(HDAC) with sequence similarity to Hdalp, Rpd3p, Hos2p, and Hos3p; deletion results in increased histone acetylation at rDNA repeats;
CAGLOF03025g YDR421W AR080 specific RNA polymerase II tran-scription factor activity
CAGLOA01628g YGL035C MIG1 transcription factor involved in glu- cose repression; C2H2 zinc finger protein which resembles the mam-malian Egr and Wilms tumour pro-teins
CAGLOF01837g YLR055C SPT8 transcription factor CAGLOD01364g YBR112C SSN6 General repressor of transcription CAGLOB03421g YLR256W HAPI Heme-responsive zinc finger tran-
scription factor of the Zn(2)-Cys(6) binuclear cluster domain type; redox sensing regulator of gene expression (activates CYC1, CYC7, CYP3, CYB2, CTT1, COR2, ROX1, ERGS, ERG11, SOD2 and YHB1; represses HEM13)
continued on next page

Appendix F. Chapter 5 Supplementary Data 314
continued from previous page CG ORF SC ORF Gene Name Description CAGLOB01441g
CAGL00O2277g
CAGLOE02079g
YNL330C
YER040W
YOL116W
RPD3
GLN3
MSNI
Histone deacetylase; regulates tran-scription and silencing transcriptional activator of nitrogen-regulated genes transcriptional activator
CAGLOM01540g CAGLOL05412g
CAGLOK05841g
CAGLOM02739g
CAGLOE00627g CAGLOH07711g
CAGLOM02585g CAGLOJ04400g
CAGLOD00682g
CAGLOJ06182g
CAGLOJ10472g
CAGLOK09900g
CAGLOL04400g
CAGLOL11880g
CAGLOA04235g
CAGLOL03916g
YDR392W YJL115W
YLR256W-like
YPL129W
YCR081W YGL244W
YPL138C YI3L021C
YLR403W
YNL167C
YDR443C
YOR358W
YOR162C
YER169W
YBL052C
YOR113W
SPT3 ASF1
HAP1-like
TAF14
SR138 RTF1
SPP1 HAP3
SFP1-like (not orthologuc)
SKO1
SSNS
HAP5
YRR1
RPH1
SASS
AZFI-LIKE
transcription factor Nucleosome assembly factor, in- volved in chromatin assembly and disassembly, anti-silencing protein that causes derepression of silent loci when overexpressed Zinc finger transcription factor in-volved in the complex regulation of gene expression in response to levels of heme and oxygen transcription factor TFIID com- plex—Component— transcription factor complex Subunit of the RNA polymerase II-associated Pafl complex; directly or indirectly regulates DNA-binding properties of Sptl5p and relative activities of different TATA ele- ments transcriptional activator activity transcriptional activator protein of CYCI (component of HAP2/HAP3 heteromer) transcription factor activity Func-tion Basic leucine zipper (bZIP) tran-scription factor of the ATF/CREB family that forms a complex with Tuplp and Ssn6p to both activate and repress transcription; cytoso- lie and nuclear protein involved in the osmotic and oxidative stress re-sponses Required for stable association of SrblOp-Srbllp kinase with RNA polymerase holoenzyme; regulates YGP1 epxression; component of RNA polymerase II holoenzyme and Kornberg's mediator (SRB) sub- complex; transcription factor CCAAT-binding transcription fac-tor component (along with Hap2p and Hap3p) Zn2-Cys6 zinc-finger transcription factor that activates genes involved in multidrug resistance; paralog of Yrrlp, acting on an overlapping set of target genes JmjC domain-containing histone demethylase which can specifi- cally demethylate H3K36 tri- and dimethyl modification states; tran-scriptional repressor of PHR1 Protein with histone acetyltrans- ferase activity Zinc-finger transcription factor, in-volved in induction of CLN3 tran-scription in response to glucose; ge-netic and physical interactions in-dicate a possible role in mitochon-dria] transcription or genome main-tenance
continued on next page

Appendix F. Chapter 5 Supplementary Data 315
continued from previous page CG ORF SC ORF Gene Name Description CAGLOJ05060g
CAGL0.110274g
CAGL0107183g
CAGLOK11902g CAGLOF04081g
CAGLOH08239g CAGLOJ06974g
CAGLOF00803g
CAGL0005335g
CAGLOH06677g CAGLOB01947g
CAGLOK04543g
CAGL00O2519g
CAGLOD00462g CAGLOK07161g
CAGLOG08844g
CAGLOH02145g
YJL056C
YNL076W
YOR140W
YDR034C YBRO83W
YOR025W YPL116W
YOLO51W
YOL067C
YIL154C YDR123C
YGRO63C
YER028C
YKL185W YGL115w
YIL13OW
YHR084W
ZAPI
MKS1
SFL1
LYSI4 TEC1-LIKE
HST3 HOS3
GALII
RTG1
IMP2' INO2
SPT4
MIG3
ASH1 SNR4-like
ASG/
STEI2
Zinc-regulated transcription factor, binds to zinc-responsive promoter elements to induce transcription of certain genes in the presence of zinc; regulates its own transcrip- tion; contains seven zinc-finger do-mains Negative regulator of nitrogen metabolism transcription factor with domains homologous to Hsflp and to myc oncoprotein; required for normal cell surface assembly and floccu- lence transcriptional activator activity transcription factor of the TEA/ATTS DNA-binding do- main family, regulator of Tyl expression Protein similar to Sir2p Trichostatin A-insensitive homod-imeric histone deacetylase (HDAC) with specificity in vitro for histones H3, H4, H2A, and H2B; similar to Hdalp, Rpd3p, Hos1p, and Hos2p; Component of the Mediator com- plex; interacts with RNA poly- merase II and the general transcrip-tion factors to form the RNA poly-merase II holoenzyme; affects tran-scription by acting as target of ac-tivators and repressors transcription factor (bHLH) in- volved in interorganelle communi-cation between mitochondria, per-oxisomes, and nucleus transcription factor transcription factor required for derepression of inositol-choline- regulated genes involved in phos- pholipid synthesis; potential Cdc28p substrate; helix-loop-helix protein Protein involved in chromatin structure that influences expression of many genes transcription factor activity Func-tion GATA-type transcription factor Protein involved in derepression of glucose-repressed genes, acts with Snflp Proposed transcriptional activator, member of the Gal4p family of zinc cluster proteins transcription factor that is acti- vated by a MAP kinase signaling cascade, activates genes involved in mating or pseudohyphal/invasive growth pathways; cooperates with Teclp transcription factor to reg- ulate genes specific for invasive growth
continued on next page

Appendix F. Chapter 5 Supplementary Data 316
continued from previous page CG ORF SC ORF Gene Name Description CAGLOM09405g
CAGLOL06028g CAGLOK08756g CAGLOL12650g
CAGLOM03025g
CAGLOB02651g
CAGLOG07249g
CAGLOL04576g
CAGL0108085g
CAGLOM07634g
CAGLOK12540g
CAGLOJ07942g
CAGLOK08668g
CAGLOM01716g
CAGLOE02805g CAGLOK07634g
CAGLOE04884g
CAGLOM09955g
CAGLOG04389g
CAGLOK09372g
YBL103C
YGL181W YIR018W YPL042C
YMR280C
YDR253C
YML027W
YOR172W
YOL071W
YMRO16C
YEL031W
YNL257C
YIR017C
YBRO83W
YBLOO8W YFLO21W
YDR216W
YLR403W
YJL110C
YGL209W
RTG3
GTSI YAPS SSN3
CATS
MET32
YOX1
YRMI
EMI5
SOK2
HAC1
SIPS
MET28
TEC1
HIRI GAT1
ADRI
SFP1
GZF3
MIG2
Basic helix-loop-helix-leucine zip- per (bHLH/Zip) transcription fac-tor that forms a complex with an-other bHLH/Zip protein, Rtglp, to activate the retrograde (RTG) and TOR pathways transcriptional activator activity bZIP transcription factor transcription factor com- plex—Component— specific RNA polymerase II tran-scription factor activity specific RNA polymerase II tran-scription factor activity Homeodomain-containing tran- scriptional repressor, binds to Mcmlp and to early cell cycle boxes (ECBs) in the promoters of cell cycle-regulated genes expressed in M/G1 phase; expression is cell cycle-regulated; potential Cdc28p substrate Zn2-Cys6 zinc-finger transcription factor that activates genes involved in multidrug resistance; paralog of Yrrlp, Non-essential protein of unknown function required for transcrip- tional induction of the early meiotic-specific transcription factor IME1, also required for sporulation Protein that can when overex- pressed suppress mutants of cAMP-dependent protein kinase; displays homologies to several transcription factors bZIP transcription factor (ATF/CREB1 homolog) that regulates the unfolded-protein response, via UPRE binding, and membrane biogenesis; ER stress- induced splicing pathway utilizing Ire1p, Trllp and Ada5p facilitates efficient Haclp synthesis transcriptional activator, interact- ing with Snflp transcriptional activator in the Cbflp-Met4p-Met28p complex Transcription factor required for full Tyl epxression, Tyl-mediated gene activation, and haploid in- vasive and diploid pseudohyphal growth Histone transcription inhibitor transcriptional activator with GATA-1-type Zn finger DNA- binding motif transcription factor activity Func-tion transcription factor activity Func-tion specific RNA polymerase II tran-scription factor activity specific RNA polymerase II tran-scription factor activity
continued on next page

Appendix F. Chapter 5 Supplementary Data 317
continued from previous page CG ORF SC ORF Gene Name Description CAGLOK05797g
CAGLOL11770g
CAGLOJ03454g
CAGLOM00286g CAGLOK08624g
CAGLOM06875g
CAGLOI10769g
CAGLOM 11440g
CAGLOM06699g CAGLOM02563g CAGLOD02926g
CAGLOK03003g
CAGLOL03377g
CAGLOH05621g CAGLOL06842g CAGLOL03157g
CAGLOL02585g
CAGLOG05379g CAGLOI04246g
CAGLOM06831g
YDR512C
YER164W
YNL021W
YJR140C YKL109W
YNL025C
YMR043w
YLR098C
YIL084C YPL139C YLR015W
YMR070W
YJL089W
YPL089C YDL080C YKR034W
YKR099W
YNL199C YGL162W
YNL027W
EMII
CHDI
HDAI
HIR3 HAP4
SSN8
MCM/-like
CHA4
SDS3 UME1 BRE2
MOTS
SIP4
RLMI THIS
DAL80
BASI
GDR2 SUT1
CRZ1
Non-essential protein of unknown function required for transcrip- tional induction of the early meiotic-specific transcription factor IME1, also required for sporulation Nucleosome remodeling factor that functions in regulation of transcrip-tion elongation; component of both the SAGA and SILK complexes Component of histone deacetylase A, 75 kDa subunit Histone transcription regulator 3 transcriptional activator protein of CYC1 (component of HAP2/HAP3 heteromer) transcription factor corn- plex—Component— Transcription factor involved in cell-type-specific transcription and pheromone response DNA binding transcriptional acti-vator of CHA1 Transcriptional regulator Negative regulator of meiosis Subunit of the COMPASS (Set1C) complex, which methylates his- tone H3 on lysine 4 and is re- quired in transcriptional silencing near telomeres; involved in telom-ere maintenance; Nuclear transcription factor with two Cys2-His2 zinc fingers; involved in repression of a subset of hypoxic genes by Roxlp, repression of sev-eral DAN/TIR genes during aerobic growth, and repression of ergosterol biosynthetic genes specific RNA polymerase II tran-scription factor activity transcriptional activator activity transcriptional activator activity Negative regulator of genes in mul- tiple nitrogen degradation path- ways; member of the GATA-binding family, forms homodimers and het-erodimers with Dehlp Myb-related transcription factor in-volved in regulating basal and in-duced expression of genes of the purine and histidine biosynthesis pathways transcription factor specific RNA polymerase II tran-scription factor activity transcription factor that activates transcription of genes involved in stress response; nuclear localiza- tion is positively regulated by calcineurin-mediated dephosphory-lation
continued on next page

Appendix F. Chapter 5 Supplementary Data 318
continued from previous page CG ORF SC ORF Gene Name Description CAGLOM06919g YNL023C FAP1 transcription factor homolog; sim-
ilarity to Drosophila melanogaster shuttle craft protein; similarity to human NEX1 protein; similar-ity to human DNA-binding protein tenascin
CAGLOK04257g YGRO44C RME1 Transcriptional repressor of meiosis in non-a/alpha cells
CAGLOM09042g YJR094C /ME/ Transcription factor required for sporulation, positive regulator of IME2 and many sporulation genes

Appendix F. Chapter 5 Supplementary Data 319
Table F.5: Summary of hierarchical clustering membership with associated me-dian growth rates in YEPD f fluconazole (5 10/./g m1-1). Cluster Cl and C5
containing fluconazole sensitive and growth defect strains respectively are discussed in the
main text and are depicted in 5.4
Cluster Members YEPD 5pgm1-1 Fluc (hrs) lOugml-1 Flue (hrs)
1 pdrl, migl, hos2, hat2, cti6, ashl, sfpl, sfpl, hosl, srb8, ,nsnl, aro80, sste2, hapl-like, hap4, rph1, .sar13, sfpl -like
10.11 15.00 15.00
2 ga183, hst1, pho4, tupl, swi5, heel, shcl, adr1, yap7, cad1, tspl, mcml, snf4, rbp1, pho23, OM rlml, fzfl, sut1, cst6, hcm1, pip2, gist, sir4, rgtl, haal, spt2, mct31, dig1, redt80, smp1, crzl, fapl, msn4
9.20 10.01 9.00
3 sass, ino2, met32, hapl, mig3, rtg1, bra, ga111, tecl-likea, 8711(11, yor1, asgl, ste12, imp2', hst3, ATCC
8.90 10.79 15.0
2001, 41,13, cha4, cmi5, mcml-like, hdal, hap3, zap/, .skol, hos3, sip3, mots, renel, spt4, 8144-like, gat1, met28, yaps, mia, hap5, lys14, hac1, da180, sip4, azfl-like, yrrl, yrrn/, gtsl, this, chill, tec1, umel, sppl, cats, sok2, imc1, rtg3, sf11
4 sell, kar4, rfr1, cup9, idol, rint101, hst4, msn2, ask10, gist, tos8, wail, hot1, stb5, hat1, znes1, stc12-like, da182
9.20 12.00 7.26
5 Itg2, rpd3, 91713, pop2, SSTE6, nggl, 8710, spt8, gen5, rt11, taf14, treks1, ada2, snf5, asf1, .spt3
15.00 14.88 14.93
6 tos4, sin3, gzf3, gcr2, fkh2, sas2, emil, basl, sass, pho2, put3, hir3, odes
11.26 12.54 9.76
7 ez1/, °aft, mbpl, pprl, stp2, tye7, stip1, da181, mss11, upc2, skn7, flo8, pogl, fkh1, spot, guilt-like, arg81, hap.2, hinst, hir2, cup2, hal9, stpl, not3, hst2, sut2, rds2, stbl, ace2, yap6, yaps-like, tupl-like
8.25 9.45 7.23
Table F.6: Effect of timepoint and growth condition on Z factor. The Z factor
statistic, is commonly used in high throughput screening assays as a quality metric (769)
and is defined in equation 5.4. Values for Z were calculated for cells grown in YEPD (YEPD
Z) or YEPD supplemented with fluconazole (5pg iul-1) (Fluconazole Z).
Timepoint (hrs) YEPD Z Fluconazole Z
1.5 -1.22 -1.85
4 -1.75 -2.07 6 -1.16 -1.61 8 -0.54 -1.12
10 0.13 -0.43
11.5 0.84 0.29

Appendix F. Chapter 5 Supplementary Data 320
Table F.7: Fitted parameter values for a linear model describing optical density at 11.5 hours as a function of strain genotype, growth condition (YEPD or YEPD supplemented with fluconazole (5,ag ii1-1)) and the interaction between growth condition and genotype (equation 5.3). Plate 1 of 2. Results are for strains
contained on the first of two library plates of the CGTFKOlib. The residual standard
error of the fit was 0.159 on 576 degrees of freedom, with a multiple R2= 0.9864, adjusted
R2=0.9819 , F-statistic= 218.2 on 192 and 576 degrees of freedom, p < 2.2e - 16. The
genotype effect is of the form straincoden where n is the number of the strain in the plate.
The interaction term is of the form straincodesn:fluconazole5ugml-1. Note that significant
interactions (p < 0.05) are reported for strain 4 (pdrl), 38 (srb8), 39 (nggl), 65 (gcn5) and
85 (ga111-like) on this plate. Strain 97 is a dummy code representing the only blank well
on this plate
Effect Average 0D600 S.E.M t value p straincodes97 0.101 0.079 1.28 0.201 straincodes2 1.272 0.079 16.04 < 2e-16 straincodesl 1.274 0.079 16.074 < 2e-16 straincodes4 1.246 0.079 15.718 < 2e-16 straincodes5 1.193 0.079 15.043 < 2e-16 straincodes6 1.277 0.079 16.103 < 2e-16 straincodes7 1.222 0.079 15.412 < 2e-16 straincodes8 1.301 0.079 16.402 < 2e-16 straincodes9 1.274 0.079 16.062 < 2e-16 straincodes10 1.144 0.079 14.425 < 2e-16 straincodesll 1.265 0.079 15.958 < 2e-16 straincodes12 1.342 0.079 16.929 < 2e-16
straincodesl3 1.117 0.079 14.091 < 2e-16 straincodesl4 1.325 0.079 16.711 < 2e-16
straincodesl5 1.259 0.079 15.876 < 2e-16
straincodesl6 1.245 0.079 15.702 < 2e-16 straincodes17 1.29 0.079 16.273 < 2e-16
straincodesl8 1.227 0.079 15.475 < 2e-16 straincodesl9 1.324 0.079 16.702 < 2e-16
straincodes20 1.314 0.079 16.576 < 2e-16 straincodes21 1.242 0.079 15.667 < 2e-16
straincodes22 0.897 0.079 11.313 < 2e-16 straincodes23 1.343 0.079 16.944 < 2e-16
straincodes24 0.411 0.079 5.184 < 2e-16
straincodes25 1.314 0.079 16.572 < 2e-16 straincodes26 1.292 0.079 16.292 < 2e-16
straincodes27 1.246 0.079 15.712 < 2e-16
straincodes28 1.278 0.079 16.112 < 2e-16 straincodes29 1.306 0.079 16.465 < 2e-16
straincodes30 1.308 0.079 16.5 < 2e-16 straincodes3l 0.917 0.079 11.565 < 2e-16
straincodes32 1.24 0.079 15.642 < 2e-16 straincodes33 1.243 0.079 15.674 < 2e-16
straincodes34 1.242 0.079 15.667 < 2e-16
straincodes35 1.176 0.079 14.838 < 2e-16
straincodes36 1.268 0.079 15.992 < 2e-16 straincodes37 1.234 0.079 15.57 < 2e-16
straincodes38 1.209 0.079 15.245 < 2e-16
straincodes39 0.796 0.079 10.046 < 2e-16
straincodes40 1.274 0.079 16.074 < 2e-16
continued on next page

Appendix F. Chapter 5 Supplementary Data 321
continued from previous page Effect Average ()Dom S.E.M t value p straincodes4l 1.29 0.079 16.267 < 2e-16 straincodes42 1.129 0.079 14.242 < 2e-16 straincodes43 1.229 0.079 15.497 < 2e-16 straincodes44 0.411 0.079 5.184 < 2e-16 straincodes45 1.193 0.079 15.049 < 2e-16 straincodes46 1.172 0.079 14.778 < 2e-16 straincodes47 1.29 0.079 16.267 < 2e-16 straincodes48 1.257 0.079 15.85 < 2e-16 straincodes49 1.289 0.079 16.263 < 2e-16 straincodes50 1.289 0.079 16.263 < 2e-16 straincodes5l 1.241 0.079 15.652 < 2e-16 straincodes52 1.038 0.079 13.091 < 2e-16 straincodes53 1.302 0.079 16.418 < 2e-16 straincodes54 1.156 0.079 14.586 < 2e-16 straincodes55 1.235 0.079 15.573 < 2e-16 straincodes56 1.263 0.079 15.929 < 2e-16 straincodes57 1.271 0.079 16.036 < 2e-16 straincodes58 1.215 0.079 15.321 < 2e-16 straincodes59 1.283 0.079 16.178 < 2e-16 straincodes60 1.183 0.079 14.927 < 2e-16 straincodes61 1.294 0.079 16.32 < 2e-16 straincodes62 1.272 0.079 16.049 < 2e-16 straincodes63 1.39 0.079 17.531 < 2e-16 straincodes64 1.314 0.079 16.576 < 2e-16 straincodes65 0.888 0.079 11.2 < 2e-16 straincodes66 1.315 0.079 16.582 < 2e-16 straincodes67 1.221 0.079 15.399 < 2e-16 straincodes68 1.255 0.079 15.831 < 2e-16 straincodes69 1.2 0.079 15.138 < 2e-16 straincodes70 1.386 0.079 17.48 < 2e-16 straincodes7l 1.069 0.079 13.489 < 2e-16 straincodes72 1.227 0.079 15.478 < 2e-16 straincodes73 1.171 0.079 14.769 < 2e-16 straincodes74 1.306 0.079 16.465 < 2e-16 straincodes75 1.286 0.079 16.222 < 2e-16 straincodes76 1.241 0.079 15.649 < 2e-16
straincodes77 1.222 0.079 15.418 < 2e-16 straincodes78 1.261 0.079 15.907 < 2e-16 straincodes79 1.259 0.079 15.882 < 2e-16 straincodes80 1.318 0.079 16.629 < 2e-16 straincodes81 1.192 0.079 15.037 < 2e-16 straincodes82 1.236 0.079 15.589 < 2e-16 straincodes83 1.314 0.079 16.569 < 2e-16 straincodes84 1.284 0.079 16.197 < 2e-16 straincodes85 1.272 0.079 16.046 < 2e-16 straincodes86 1.217 0.079 15.346 < 2e-16
straincodes87 1.274 0.079 16.074 < 2e-16 straincodes88 1.257 0.079 15.853 < 2e-16 straincodes89 1.082 0.079 13.646 < 2e-16 straincodes90 1.224 0.079 15.444 < 2e-16 straincodes9l 1.243 0.079 15.68 < 2e-16 straincodes92 1.096 0.079 13.823 < 2e-16 straincodes93 1.213 0.079 15.299 < 2e-16 straincodes94 1.284 0.079 16.194 < 2e-16
straincodes95 1.326 0.079 16.721 < 2e-16 straincodes96 1.227 0.079 15.472 < 2e-16 fluconazole5ugml-1 0.012 0.112 0.103 0.918 straincodes2:fluconazole5ugml-1 -0.045 0.159 -0.284 0.777 straincodesl:fluconazole5ugml-1 -0.157 0.159 -0.987 0.324 straincodes4:fluconazole5ugml-1 -0.846 0.159 -5.332 0 straincodes5ffluconazole5ugm1-1 -0.183 0.159 -1.151 0.25 straincodes6:fluconazole5ugnal-1 -0.117 0.159 -0.739 0.46
continued on next page

Appendix F. Chapter 5 Supplementary Data 322
continued from previous page Effect Average OD600 S.E.M t value p straincodes7:fluconazole5ugm1-1 -0.143 0.159 -0.899 0.369 straincodes8:fluconazole5ugml-1 -0.115 0.159 -0.722 0.471 straincodes9:fluconazole5ugm1-1 -0.159 0.159 -1 0.318 straincodes10:fluconazole5ugm1-1 -0.171 0.159 -1.08 0.281 straincodes11:fluconazole5ugm1-1 -0.124 0.159 -0.784 0.434 straincodes12:fluconazole5ugml-1 -0.224 0.159 -1.414 0.158 straincodesl3:fluconazole5ugml-1 -0.043 0.159 -0.273 0.785 straincodesl4:fluconazole5ugml-1 -0.092 0.159 -0.58 0.562 straincodes15:fluconazole5ugm1-1 -0.157 0.159 -0.988 0.323 straincodes16:fluconazole5ugm1-1 -0.154 0.159 -0.97 0.333 straincodesl7:fluconazole5ugml-1 -0.217 0.159 -1.368 0.172 straincodes18:fluconazole5ugm1-1 -0.184 0.159 -1.162 0.246 straincodes19:fluconazole5ugm1-1 -0.039 0.159 -0.248 0.805 straincodes20:fluconazole5ugm1-1 -0.288 0.159 -1.818 0.07 straincodes21:fluconazole5ugm1-1 -0.138 0.159 -0.869 0.385 straincodes22:fluconazole5ugm1-1 -0.02 0.159 -0.126 0.9 straincodes23:fluconazole5ugm1-1 -0.156 0.159 -0.984 0.326 straincodes24:fluconazole5ugm1-1 -0.183 0.159 -1.154 0.249 straincodes25:fluconazole5ugml-1 -0.056 0.159 -0.355 0.723 straincodes26:fluconazole5ugml-1 -0.064 0.159 -0.404 0.687 straincodes27:fluconazole5ugm1-1 -0.057 0.159 -0.359 0.719 straincodes28:fluconazole5ugm1-1 -0.161 0.159 -1.012 0.312 straincodes29:fluconazole5ugml-1 -0.207 0.159 -1.304 0.193 straincodes30:fluconazole5ugml-1 -0.088 0.159 -0.552 0.581 straincodes31:fluconazole5ugm1-1 0.064 0.159 0.405 0.686 straincodes32:fluconazole5ugm1-1 -0.151 0.159 -0.952 0.341 straincodes33:fluconazole5ugm1-1 -0.125 0.159 -0.785 0.433 straincodes34:fluconazole5ugm1-1 -0.022 0.159 -0.139 0.89 straincodes35:fluconazole5ugml-1 -0.09 0.159 -0.566 0.572 straincodes36:fluconazole5ugm1-1 -0.077 0.159 -0.487 0.626 straincodes37:fluconazole5ugm1-1 -0.011 0.159 -0.066 0.947 straincodes38:fluconazole5ugml-1 -0.407 0.159 -2.567 0.011 straincodes39:fluconazole5ugml-1 -0.556 0.159 -3.503 0 straincodes40:fluconazole5ugm1-1 -0.083 0.159 -0.52 0.603 straincodes4l:fluconazole5ugml-1 -0.2 0.159 -1.258 0.209 straincodes42:fluconazole5ugm1-1 -0.097 0.159 -0.612 0.541 straincodes43:fluconazole5ugm1-1 -0.045 0.159 -0.285 0.775 straincodes44:fluconazole5ugm1-1 -0.224 0.159 -1.411 0.159 straincodes45:fluconazole5ugm1-1 -0.108 0.159 -0.678 0.498 straincodes46:fluconazole5ugml-1 -0.048 0.159 -0.303 0.762 straincodes47:fluconazole5ugm1-1 -0.105 0.159 -0.661 0.509 straincodes48:fluconazole5ugml-1 -0.104 0.159 -0.657 0.511 straincodes49:fluconazole5ugm1-1 -0.022 0.159 -0.137 0.891 straincodes50:fluconazole5ugm1-1 -0.041 0.159 -0.255 0.799 straincodes51:fluconazole5ugm1-1 -0.111 0.159 -0.698 0.485 straincodes52:fluconazole5ugm1-1 -0.287 0.159 -1.811 0.071 straincodes53:fluconazole5ugm1-1 -0.189 0.159 -1.189 0.235 straincodes54:fluconazole5ugm1-1 -0.015 0.159 -0.091 0.927 straincodes55:fluconazole5ugml-1 -0.024 0.159 -0.151 0.88 straincodes56:fluconazole5ugm1-1 -0.1 0.159 -0.631 0.529 straincodes57:fluconazole5ugm1-1 -0.148 0.159 -0.935 0.35 straincodes58:fluconazole5ugm1-1 -0.108 0.159 -0.681 0.496 straincodes59:fluconazole5ugm1-1 -0.053 0.159 -0.336 0.737 straincodes60:fluconazole5ugm1-1 -0.082 0.159 -0.514 0.607 straincodes61:fluconazole5ugml-1 -0.002 0.159 -0.014 0.989 straincodes62:fluconazole5ugm1-1 -0.053 0.159 -0.334 0.738 straincodes63:fluconazole5ugm1-1 -0.067 0.159 -0.421 0.674 straincodes64:fluconazole5ugml-1 -0.127 0.159 -0.798 0.425 straincodes65:fluconazole5ugml-1 -0.603 0.159 -3.801 0 straincodes66:fluconazole5ugm1-1 -0.067 0.159 -0.424 0.672 straincodes67:fluconazole5ugm1-1 -0.136 0.159 -0.858 0.391 straincodes68:fluconazole5ugm1-1 -0.093 0.159 -0.586 0.558
continued- on next page

Appendix F. Chapter 5 Supplementary Data 323
continued from previous page Effect Average OD900 S.E.M t value p straincodes69:fluconazole5ugm1-1 -0.063 0.159 -0.396 0.692 straincodes70:fluconazole5ugm1-1 -0.041 0.159 -0.259 0.796 straincodes71:fluconazole5ugm1-1 -0.044 0.159 -0.279 0.78 straincodes72:fluconazole5ugm1-1 -0.073 0.159 -0.462 0.644 straincodes73:fluconazole5ugml-1 -0.035 0.159 -0.218 0.828 straincodes74:fluconazole5ugml-1 -0.127 0.159 -0.799 0.424 straincodes75:fluconazole5ugm1-1 -0.145 0.159 -0.916 0.36 straincodes76:fluconazole5ugm1-1 -0.132 0.159 -0.834 0.405 straincodes77:fluconazole5ugml-1 -0.111 0.159 -0.702 0.483 straincodes78:fluconazole5ugm1-1 -0.063 0.159 -0.396 0.692 straincodes79:fluconazole5ugm1-1 -0.061 0.159 -0.385 0.701 straincodes80:fluconazole5ugm1-1 -0.078 0.159 -0.493 0.622 straincodes81:fluconazole5ugm1-1 -0.108 0.159 -0.679 0.497 straincodes82:fluconazole5ugm1-1 -0.076 0.159 -0.476 0.634 straincodes83:fluconazole5ugm1-1 -0.094 0.159 -0.59 0.556 straincodes84:fluconazole5ugm1-1 -0.059 0.159 -0.37 0.711 straincodes85:fluconazole5ugm1-1 -0.732 0.159 -4.614 0 straincodes86:fluconazole5ugm1-1 -0.012 0.159 -0.073 0.942 straincodes87:fluconazole5ugml-1 -0.063 0.159 -0.396 0.692 straincodes88:fluconazole5ugm1-1 -0.065 0.159 -0.407 0.684 straincodes89:fluconazole5ugm1-1 -0.111 0.159 -0.702 0.483 straincodes90:fluconazole5ugm1-1 0.018 0.159 0.114 0.91 straincodes91:fluconazole5ugm1-1 0.002 0.159 0.016 0.987 straincodes92:fluconazole5ugm1-1 -0.018 0.159 -0.115 0.908 straincodes93:fluconazole5ugm1-1 -0.063 0.159 -0.394 0.694 straincodes94:fluconazole5ugml-1 0 0.159 0 1 straincodes95:fluconazole5ugm1-1 -0.039 0.159 -0.246 0.806 straincodes96:fluconazole5ugm1-1 -0.054 0.159 -0.337 0.736

Appendix F. Chapter 5 Supplementary Data 324
Table F.8: Fitted parameter values for a linear model describing optical density at 11.5 hours as a function of strain genotype, growth condition (YEPD or YEPD supplemented with fluconazole (5/.ig p1-1) and the interaction between growth condition and genotype (equation 5.3). Plate two of two. Results are for strains contained on the second of two library plates of the CGTFKO1ib. The residual standard error of the fit was 0.183 on 576 degrees of freedom, with a multiple R2= 0.9802,
adjusted R2=0.9735 , F-statistic= 148.1 on 192 and 576 degrees of freedom, p < 2.2e - 16. The genotype effect is of the form straincoden where n is the number of the strain in the plate. The interaction term is of the form straincodesn:fluconazole5ugm1-1. Note that significant interactions (p < 0.05) are reported for strain 27 (hap)-like), 28 (ada2) and 61
(rphl) on this plate. Strain codes 93-97 inclusive are blank wells on this plate
Effect Average ()Dm) S.E.M t value p
straincodes97 0.063 0.092 0.69 0.49 straincodes2 1.264 0.092 13.796 0
straincodesl 1.259 0.092 13.747 0 straincodes4 1.254 0.092 13.687 0
straincodes5 1.071 0.092 11.695 0 straincodes6 1.026 0.092 11.193 0
straincodes7 1.274 0.092 13.905 0 straincodes8 1.097 0.092 11.979 0 straincodes9 1.192 0.092 13.013 0
straincodes10 1.119 0.092 12.208 0
straincodesll 1.25 0.092 13.643 0 straincodesl2 1.28 0.092 13.97 0
straincodesl3 1.217 0.092 13.288 0 straincodesl4 1.289 0.092 14.071 0
straincodesl5 1.238 0.092 13.518 0
straincodes16 1.257 0.092 13.719 0
straincodes17 1.205 0.092 13.157 0 straincodes18 1.143 0.092 12.478 0 straincodesl9 0.863 0.092 9.414 0 straincodes20 0.851 0.092 9.294 0 straincodes2) 1.051 0.092 11.474 0 straincodes22 1.246 0.092 13.602 0
straincodes23 1.138 0.092 12.418 0
straincodes24 1.282 0.092 13.987 0
straincodes25 1.326 0.092 14.475 0 straincodes26 1.115 0.092 12.172 0
straincodes27 1.266 0.092 13.823 0 straincodes28 0.92 0.092 10.039 0 straincodes29 1.208 0.092 13.182 0 straincodes30 1.192 0.092 13.015 0
straincodes3l 1.195 0.092 13.04 0
straincodes32 1.249 0.092 13.635 0
straincodes33 1.309 0.092 14.284 0 straincodes34 1.271 0.092 13.872 0
straincodes35 1.246 0.092 13.605 0
straincodes36 1.305 0.092 14.246 0 straincodes37 1.324 0.092 14.451 0
straincodes38 1.291 0.092 14.088 0 straincodes39 1.271 0.092 13.872 0 straincodes40 1.274 0.092 13.905 0
straincodes4) 1.239 0.092 13.52 0 straincodes42 0.492 0.092 5.37 0
continued on next page

Appendix F. Chapter 5 Supplementary Data 325
continued from previous page Effect Average OD600 S.E.M t value p straincodes43 0.891 0.092 9.719 0 straincodes44 1.217 0.092 13.286 0 straincodes45 1.195 0.092 13.043 0 straincodes46 1.144 0.092 12.481 0 straincodes47 1.274 0.092 13.908 0 straincodes48 1.291 0.092 14.091 0 straincodes49 1.244 0.092 13.578 0 straincodes50 0.32 0.092 3.495 0.001 straincodes5l 1.271 0.092 13.875 0 straincodes52 1.285 0.092 14.022 0 straincodes53 1.043 0.092 11.378 0 straincodes54 1.205 0.092 13.152 0 straincodes55 1.285 0.092 14.02 0 straincodes56 1.211 0.092 13.215 0 straincodes57 1.155 0.092 12.612 0 straincodes58 1.251 0.092 13.659 0 straincodes59 1.248 0.092 13.618 0 straincodes60 1.29 0.092 14.082 0 straincodes6l 1.22 0.092 13.318 0 straincodes62 1.256 0.092 13.709 0 straincodes63 1.194 0.092 13.032 0 straincodes64 1.28 0.092 13.97 0 straincodes65 1.265 0.092 13.807 0 straincodes66 1.185 0.092 12.934 0 straincodes67 1.244 0.092 13.578 0 straincodes68 1.265 0.092 13.807 0 straincodes69 0.272 0.092 2.969 0.003 straincodes70 1.173 0.092 12.8 0 straincodes7l 1.259 0.092 13.741 0 straincodes72 1.169 0.092 12.759 0 straincodes73 1.277 0.092 13.935 0 straincodes74 1.076 0.092 11.744 0 straincodes75 1.263 0.092 13.782 0 straincodes76 1.22 0.092 13.321 0 straincodes77 1.279 0.092 13.957 0
straincodes78 1.122 0.092 12.243 0 straincodes79 1.276 0.092 13.93 0 straincodes80 1.071 0.092 11.687 0 straincodes81 1.204 0.092 13.146 0 straincodes82 0.929 0.092 10.14 0
straincodes83 1.218 0.092 13.299 0 straincodes84 1.296 0.092 14.145 0 straincodes85 1.179 0.092 12.874 0 straincodes86 1.241 .0.092 13.548 0 straincodes87 1.278 0.092 13.951 0 straincodes88 1.143 0.092 12.478 0 straincodes89 1.303 0.092 14.222 0 straincodes90 1.285 0.092 14.025 0 straincodes9l 1.272 0.092 13.883 0 straincodes92 1.187 0.092 12.958 0 straincodes93 0.07 0.092 0.761 0.447 straincodes94 0.066 0.092 0.718 0.473
straincodes95 0.067 0.092 0.734 0.463 straincodes96 0.068 0.092 0.748 0.455 conditionsB 0.001 0.13 0.004 0.997 straincodes2:conclitionsB -0.059 0.183 -0.323 0.747 straincodesl:conditionsB -0.08 0.183 -0.437 0.663 straincodes4:conditionsB -0.054 0.183 -0.296 0.767 straincodes5:conditionsB 0.027 0.183 0.147 0.883 straincodes6:conditionsB -0.028 0.183 -0.154 0.878 straincodes7:conditionsB -0.064 0.183 -0.349 0.727 straincodes&conditionsB -0.085 0.183 -0.464 0.643
continue. on next page

Appendix F. Chapter 5 Supplementary Data 326
continued from previous page Effect Average 00600 S.E.M t value p
str aincodes9:conditionsB -0.11 0.183 -0.599 0.549 straincodes10:conditionsB -0.035 0.183 -0.191 0.849 straincodes11:conditionsB -0.051 0.183 -0.277 0.782 straincodes12:conditionsB -0.083 0.183 -0.453 0.651 straincodesl3:conditionsB 0.006 0.183 0.03 0.976 straincodesl4:conditionsB -0.202 0.183 -1.104 0.27 straincodes15:conditionsB -0.134 0.183 -0.734 0.463 straincodes16:conditionsB -0.022 0.183 -0.117 0.907 straincodesl7:conditionsB -0.067 0.183 -0.367 0.714 straincodesl8:conditionsB -0.034 0.183 -0.186 0.853 straincodes19:conditionsB -0.046 0.183 -0.25 0.803 straincodes20:conditionsB -0.02 0.183 -0.109 0.913 straincodes21:conditionsB -0.286 0.183 -1.559 0.119 straincodes22:conditionsB -0.151 0.183 -0.825 0.409 straincodes23:conditionsB -0.113 0.183 -0.618 0.537 straincodes24:conditionsB -0.084 0.183 -0.458 0.647 straincodes25:conditionsB -0.024 0.183 -0.128 0.898 straincodes26:conditionsE -0.067 0.183 -0.366 0.715 straincodes27:conditionsB -0.42 0.183 -2.293 0.022 straincodes28:conditionsB -0.599 0.183 -3.27 0.001 straincodes29:conditionsB -0.062 0.183 -0.34 0.734 straincodes30:conditionsB -0.167 0.183 -0.911 0.362 straincodes31:conditionsB -0.224 0.183 -1.222 0.222 straincodes32:conditionsB -0.187 0.183 -1.023 0.307 straincodes33:conditionsB -0.116 0.183 -0.636 0.525 straincodes34:conditionsB -0.246 0.183 -1.342 0.18 straincodes35:conditionsB -0.105 0.183 -0.573 0.567 straincodes36:conditionsB -0.031 0.183 -0.171 0.865 straincodes37:conditionsB -0.067 0.183 -0.363 0.717 straincodes38:conditionsB -0.109 0.183 -0.592 0.554 straincodes39:conditionsB -0.101 0.183 -0.554 0.58 straincodes40:conditionsS -0.116 0.183 -0.63 0.529 straincodes41:conditionsB -0.2 0.183 -1.09 0.276 straincodes42:conditionsB -0.182 0.183 -0.995 0.32 straincodes43:conditionsB -0.057 0.183 -0.308 0.758 straincodes44:conditionsB -0.15 0.183 -0.817 0.414
straincodes45:conditionsB -0.109 0.183 -0.596 0.551 straincodes46:conditions8 -0.131 0.183 -0.715 0.475
straincodes47:conditionsB -0.075 0.183 -0.409 0.682 straincodes48:conditionsB -0.028 0.183 -0.15 0.881 straincodes49:conditionsB -0.087 0.183 -0.473 0.636 straincodes50:conditionsB 0.04 0.183 0.218 0.827 straincodes51:conditionsB -0.042 0.183 -0.226 0.821 straincodes52:conditionsB -0.134 0.183 -0.734 0.463 straincodes53:conditionsB -0.147 0.183 -0.804 0.422 straincodes54:conditionsB -0.115 0.183 -0.63 0.529 straincodes55:conditionsB -0.181 0.183 -0.989 0.323 straincodes56:conditionsB -0.17 0.183 -0.925 0.355
straincodes57:conditionsB -0.014 0.183 -0.075 0.94 straincodes58:conditionsB -0.136 0.183 -0.742 0.458
straincodes59:conditionsB -0.151 0.183 -0.821 0.412 straincodes60:conditionsB 0.017 0.183 0.09 0.928 straincodes61:conditionsB -0.366 0.183 -1.999 0.046 straincodes62:conditionsB -0.027 0.183 -0.146 0.884 straincodes63:conditionsB -0.257 0.183 -1.405 0.16 straincodes64:conditionsB -0.103 0.183 -0.563 0.573 straincodes65:conditionsB -0.092 0.183 -0.502 0.616 straincodes66:conditionsB -0.15 0.183 -0.817 0.414 straincodes67:conditionsB -0.122 0.183 -0.663 0.508 straincodes68:conditionsB -0.21 0.183 -1.147 0.252 straincodes69:conditionsB -0.051 0.183 -0.277 0.782 straincodes70:conditionsB -0.072 0.183 -0.392 0.696
continued on next page

Appendix F. Chapter 5 Supplementary Data 327
continued from previous page Effect Average 0D600 S.E.M t value p straincodes71:conditionsB -0.076 0.183 -0.416 0.677 straincodes72:conditionsB -0.03 0.183 -0.162 0.871 straincodes73:conditionsB 0.03 0.183 0.161 0.872 straincodes74:conditionsB -0.272 0.183 -1.486 0.138 straincodes75:conditionsB -0.031 0.183 -0.168 0.867 straincodes76:conditionsB -0.098 0.183 -0.533 0.594 straincodes77:conditionsB -0.033 0.183 -0.18 0.857 straincodes78:conditionsB 0.002 0.183 0.012 0.99 straincodes79:conditionsB -0.061 0.183 -0.333 0.739 straincodes80:conditionsB -0.038 0.183 -0.206 0.837 straincodes81:conditionsB -0.027 0.183 -0.149 0.882 straincodes82:conditionsB -0.159 0.183 -0.87 0.384 straincodes83:conditionsB -0.04 0.183 -0.218 0.827 straincodes84:conditionsB -0.039 0.183 -0.213 0.832 straincodes85:conditionsB 0.066 0.183 0.362 0.718 straincodes86:conditionsB -0.011 0.183 -0.059 0.953 straincodes87:conditionsB -0.022 0.183 -0.119 0.906 straincodes88:conditionsB -0.07 0.183 -0.379 0.705 straincodes89:conditionsB -0.065 0.183 -0.353 0.724 straincodes90:conditionsB -0.039 0.183 -0.21 0.834 straincodes9I:conditionsB 0.002 0.183 0.014 0.989 straincodes92:conditionsB -0.103 0.183 -0.563 0.573 straincodes93:conditionsB -0.003 0.183 -0.016 0.987 straincodes94:conditionsB -0.001 0.183 -0.007 0.995 straincodes95:conditionsB -0.001 0.183 -0.004 0.997 straincodes96:conditionsB -0.003 0.183 -0.018 0.986

(a) tO (b) t2
(e) t8 (f) t10
1
(g) t12 (h) t14
3 • 6 .6 7 • 10 11 12 7 1 10 11 12
1 3 0
(c) t4 (d) t6
5 8 7 • 1 18 11 12 1 2 1 • .1 8 7 8 9 10 11 12
Appendix F. Chapter 5 Supplementary Data 328
Figure F.1: Assessment of pipette channel cross contamination on a robotic plat-form. Two fold serial dilutions of a C. glabrata culture (0D600=0.5) were dispensed into column 1 (rows A-H inclusive) and 7 (rows A-E inclusive) of a 96 well deep well plate. Columns 2-6 and 8-12 contained growth medium (YEPD) without cells. Samples (10p/1) were taken from the deep-well plate every 2 hours for 14 hours and transferred to a mi-crotitre plate containing 90 pd of sodium azide (final concentration 100mM, final OD600 = 0.1 x initial inoculation OD600) to stop cell growth. Wells lacking inoculum were inspected

Appendix F. Chapter 5 Supplementary Data 329
CG2001
CG2001 his3
0
0 10 15 20
5 10
1(hr.) t (hrs)
(a) ATCC CG2001
(b) his.
PDR1
MIG1
_
-
0 > _ 8
/ 1
•
0 5 10 15 20
5 10 20
t(hrs) t (hrs)
(c) pdrl
(d) migl
Figure F.2: Example growth curves for four strains following incubation and sampling on a robotic platform. Glycerol stocks of each library strain, collectively maintained in two 96-well microtitre plates at -80°C, were sampled (5p,1) using a multi-channel pipette and dispensed into corresponding sterile, round-bottomed 96-well plate containing 150/21 of YEPD broth. Library strains were sub-cultured to stationary phase for 18 hours in an orbital shaker (30°C, 180 RPM) before commencing the growth curve assay. Cells from each library plate were transferred (final OD600=0.2) into square bottom, deep well plates (Anachem) containing a final volume of 500 µl of YEPD and incubated at (37°C, 180 RPM), rather than 30°C (the temperature used in the manual assay). The robot was programmed to take 100 samples of cells every two hours and dispense them into microtitre plates containing 900 of sodium azide (final concentration of 100mM). The 8-tip pipette head was washed five times with a mixture of isopropanol and copper sulphate (100mM) and rinsed with distilled water before each move to a new column on the deep well plate. Plates were stored at room temperature and 0D600 measurements where made using a Multiskan Ascent absorbance plate-reader (Thermo Scientific, U.S.A).

330
Appendix G
Transcription profiling of C. glabrata serum stress responses
G.1 Chapter overview
This truncated chapter is a record of the current status of some of the environmental stress transcription profiling I have performed. The scope of this work was large and has not yet been fully summarized (including oxidative - 1mM tBOOH, osmotic stress - 0.5 M KC1 and carbon starvation 0.01% glucose w/v), partially due to time constraints and also because of uncertainties surrounding the use of orthologous integrated molecular interaction networks, which we intended to use to identify conserved orthologous active modules. For example, to date we do not have a C. glabrata protein interaction network, so the well documented health warnings that come with protein interaction data (excellently reviewed in (265)) are further compounded by assuming that orthologous proteins have the same connections in related species. Furthermore, current protein interaction data take no account of how compartmentalization, and dynamical expression and transcription may affect the structure of these ostensibly scale-free networks (47; 49; 48; 328; 329; 330).
The concept of a co-ordinated, general stress response or environmental stress response (subsequently called the environmental stress response - ESR) emerged after the effects of normally lethal doses of one stress were sublethal after pre-treatment with a sub-lethal dose of a different stress (467; 468). Furthermore, genes that were specifically thought to protect the cell against heat shock were shown to be induced under a variety of independent stressors. Analysis of the upstream promoter region of these genes identified a conserved sequence element that became known as the stress response element (STRE), which was later shown to be the consensus sequence binding site for the zinc-finger transcription fac-tors, Msn2p and Msn4p (615; 615; 81). The ESR was originally described using microarray

Appendix G. Transcription profiling of C. glabrata serum stress responses 331
expression analysis in S. cerevisiae by the independent groups Gasch et al and Causton et al. In combination these groups investigated the transcriptional responses to a range of environmental cues including heat shock (230), extremes of pH (118), oxidative stress (hydrogen peroxide (118) or menadione induced (230)), hyper-osmotic stress (NaC1 (118) or sorbitol (230; 118) induced), hypo-osmotic shock (230), amino acid starvation (230), nitrogen depletion (230), and sub-optimal carbon sources (230).
It had been proposed that this co-ordinated transcription programme is part of a con-served protective response to protect critical features of cellular physiology in budding yeast (229). However, initial transcriptional studies in C. albicans did not reveal a general stress response (191) (in oxidative, hyperosmotic stress and a 23 - 27°C temperature shift) nor was any cross protection observed when cells pre-treated with a mild heat shock were sub-sequently exposed to oxidative stress. In a contradictory study, Smith et al., (639) did demonstrate HOG/ -dependent cross-protection to 0.3M NaC1 after pre-treatment with a sublethal dose of H202. A subsequent systematic comparison of general stress response data sets of C. albicans, S. pombe and S. cerevisiae (192) did reveal a general stress re-sponse with almost 50-fold fewer genes than in S. cerevisiae. While there is evidence for a conserved ESR in the model fungi S. cervisiae and S.pombe (229), the evidence for such a comprehensive ESR in C. albicans is unconvincing. At the start of this study, the general stress response of C. glabrata had not been investigated, and the relevance of the ESR in opportunistic fungal pathogens is still unclear. This work was also put on hold after a similar publication addressed the specific question of whether or not an ESR is detectable in C. glabrata (589).
The following section is a summary of transcription profiling performed when C. glabrata cells were exposed to culture medium containing foetal calf serum (10% w/v), an environ-mental cue that is often used as a morphogenetic inducer in C. albicans. In this instance, we used synthetic medium supplemented with foetal calf serum to approximate the nitrogen availability that may be found in the blood. Blood products have been used in transcrip-tional profiling studies in C. albicans (210), and would have been a much better option to address this specific question. This is another reason why the following analysis has been relegated to the supplementary data section.
G.2 Specific methods
C. glabrata ATCC 2001 cells were re-streaked onto YEPD agar from frozen glycerol stocks 3 days prior to the microarray experiment. On the day prior to the experiment one colony was picked from the plate and used to inoculate 50 ml of synthetic complete medium (supplemented with 100mM succinic acid and buffered to pH 4.0) which was incubated

G.3 Transcriptional response of C. glabrata cells exposed to 10% Foetal Calf Serum (FCS) 332
in an orbitol shaking incubator for 18 hours (37° C, 180 rpm). Cells were harvested by centrifugation (3000 rpm, 5 minutes), resuspended to a volume of 50 ml and washed twice with sterile distilled water before estimating the cell density using a multiskan ascent at absorbance plate reader at OD600. The experiment was initialized by inoculating 1 L of synthetic complete medium in a 2 L flask with cells to a final OD600 = 0.2. Cells were incubated for 4 hours (0D600 = 0.8) and harvested using a Sorvall SLA-3000 ultracentrifuge rotor (suitable for large volume harvesting), washed with sterile distilled water and then subcultured in synthetic culture medium (reference sample) or synthetic complete medium supplemented with 10% foetal calf serum (FCS). The stress treated sub-culture and one control culture (untreated) were both incubated for a further sixty minutes before harvesting by centrifugation (3000 rpm, 5 minutes), snap-freezing in liquid nitrogen and storage at -80°C. RNA Isolation, cDNA labelling, microarray hybridizations and data analyses are fully described in the the materials and methods section (Chapter 2).
G.3 Transcriptional response of C. glabrata cells exposed to 10% Foetal Calf Serum (FCS)
Transcription profiling of cells exposed to 10% (v/v) FCS for one hour identified 501 in-duced transcripts (Supplementary Table G.1) and 448 repressed (Supplementary Table G.2) transcripts (approximately 10% of the genome in each case). These data were summarized by searching for over-represented GO component, process and function terms. Furthermore, repressed and induced transcript sets were combined and filtered to retain genes with S. cervisiae orthologues so that the differentially transcribed set could be mapped to KEGG pathways, which summarize gene expression in the context of the metabolic and signalling pathways curated for the closely related model organism. These tools provided insight into the effects of serum exposure (10% (v/v) FCS, 1 hour) on orthologous metabolic pathways, RNA metabolism and ribosome biogenesis, in addition to the activation of important sig-nalling pathways involved in nutrient and stress sensing. For brevity, all gene descriptions are taken from the functional annotation in SGD (131) and refer to references on the re-spective summary page of each gene. If it is deemed appropriate to write this section fully, proper citations will be included.
In particular, these analyses reveal an over-representation of induced transcripts local-ized in the nucleolus (Supplementary Table G.3) involved in rRNA and mRNA process-ing, ribosome biogenesis and assembly and also in the subunits of DNA directed RNA polymerases I and III (Supplementary Table G.4). The over-representation of transcripts involved in tRNA modification in combination with the above points to a cell that is up-regulating transcription and translation processes relative to the synthetic complete medium

Appendix G. Transcription profiling of C. glabrata serum stress responses 333
reference at the same time-point. Although C. glabrata and S. cerevisiae have different metabolic capabilities (e.g. the C. glabrata genome has lost genes involved in galactose and sucrose assimilation, phosphate, nitrogen and sulfur metabolism, as well as thiamine, pyridoxine and nicotinic acid biosynthesis (348)), mapping of transcript fold-changes to the orthologous global S. cerevisiae metabolic map can be useful to generate hypotheses regard-ing the metabolic adaptation that occurs following exposure to serum, an environmental cue that may approximate the nutrient availability during a bloodstream infection. Doing so reveals a co-ordinated change in gene expression, where repressed and induced transcripts are located to specific areas of the global metabolic map (Supplementary Figure G.4). In general this global view indicates that enzymes catalyzing reactions in fatty acid biosynthe-sis, starch and sucrose metabolism, glycerophospholipid metabolism, pyruvate metabolism, citrate cycle, glyoxylate metabolism and oxidative phosphorylation are repressed on expo-sure to serum. In contrast, induced genes are predominantly involved in other pathways including nucleotide metabolism, nitrogen metabolism, amino acid metabolism, the pentose phosphate shunt, the urea cycle, C5-branched dibasic acid metabolism, high mannose N-glycan biosynthesis and cyanoaminoacid metabolism. This high-level view of metabolism at a single time-point is of course an over-simplification of the dynamic nature of metabolism and gives no quantitative summary of reactant fluxes, however the hypothetical repression of C. glabrata energy production pathways which utilize glucose, and the concomitant hy-pothetical induction of C. glabrata pathways involved in amino acid metabolism indicate that glucose is not primarily used in the production of energy in serum treated cells.
The induction of genes whose products catalyze amino acid biosynthetic reactions, in conjunction with the up-regulation of permeases that transport such nutrients into the cell (e.g. the high-affinity glutamine permease GNP1 - also a transporter of leucine, threonine, cysteine and methionine, the dicarboxylic amino acid permease DIP5, the high-affinity histidine permease HIP1, a polyamine and urea transporter DUR3, an ammonium permease involved in regulation of pseudohyphal growth MEP2) and a cytosolic aspartate aminotransferase, involved in nitrogen metabolism AAT2 and CHA1 a catabolic L-serine and L-threonine deaminase which is required to use serine or threonine as the sole nitrogen source, indicates that a programme of nitrogen compound assimilation and metabolism is orchestrated by C. glabrata in response to serum exposure. Indeed, transcripts encoding catalytic enzymes in several amino acid metabolic pathways were identified. For example eight genes encoding enzymes of the phenylalanine, tyrosine and tryptophan biosynthesis pathways are induced (Supplementary Figure G.5). Such aromatic acid biosynthesis requires conversion of phosphoenolpyruvate or erythrose-4-phosphate precursors for the production of chorismate, the branch point of aromatic acid biosynthesis which is either converted to an-thranilate for tryptophan biosynthesis or to prephenate for tyrosine or phenylalanine biosyn-

G.3 Transcriptional response of C. glabrata cells exposed to 10% Foetal Calf Serum (FCS) 334
thesis. The induction of enzymes catalyzing precursor condensation (EC 2.5.1.54 ARO3), chorismate synthesis (EC 4.2.3.5 ARO2), anthranilate synthesis (EC 4.1.3.27 TRPE) and transamination (EC 2.6.1.1 AAT1 and EC 2.6.1.57 ARO8) suggest that phenylalanine, ty-rosine and tryptophan biosynthesis are all enhanced on serum exposure. The net effect of en-zyme differential transcription on valine, leucine and isoleucine is more difficult to interpret because both biosynthetic and catabolic enzymes are co-induced (Supplementary Figures G.6 and G.7). However, co-induction of enzymes catalyzing the biosynthesis (EC 4.3.1.19 CHAl, EC 2.2.1.6 ILV6,EC 4.2.1.9 , EC 2.3.3.13 ILV3) and subsequent degradation (EC 2.6.1.42 BAT1) of these amino acids may suggest that isoleucine, leucine and valine catabo-lites are preferred. Although this interpretation is confounded by the dual role of BAT1 in both synthesis and degradation of these amino acids, the concomitant repression of NAME (EC 6.1.1.4) which catalyzes leucine aminoacyl-tRNA biosynthesis further supports the no-tion that leucine biosynthesis is channeled to downstream metabolic reactions rather than into biopolymer production at the ribosome. Inspection of the arginine/proline metabolic map (Supplementary Figure G.8) and alanine/aspartate/glutamate metabolic map (Supple-mentary Figure G.9) reveals induction of enzymes that may increase flux through the citrate cycle, including arginosuccinate synthase (EC 6.3.4.5 ARG1) which catalyzes the formation of L-argininosuccinate from citrulline and L-aspartate leading to the production of fumarate a substrate in the citrate cycle. The induction of aspartate aminotransferase (EC 2.6.1.1 AAT1), L-asparaginase (EC 3.5.1.1 ASP1) and glutamine hydrolysing asparagine synthase (EC 6.3.4.5 ASNE) may lead to an increased L-arginosuccinate production and, following by, increased fumarate production. Induction of bifunctional carbamoylphosphate synthetase (EC 6.3.5.5 URA2) may also lead to an increased production of carabomyl phosphate, a precursor for the urea cycle after conversion to L-citrulline by ornithine carbamoyltrans-ferase (EC 2.1.3.3 ARG3). The concomitant repression of other enzymes that catalyze the catabolism of L-Glutamate (EC 1.2.1.16 UGAE, EC 2.6.1.19 UGA1, EC 4.1.1.15 GAD1, EC 1.4.1.2 GDH2 and EC 1.5.1.12 PUTS) suggests that the production of carabomyl phos-phate and subsequent increased flux though the urea cycle to produce fumarate rather than succinate is favoured.
Analysis of carbon catabolite pathways shows that the majority of glycolytic and elec-tron transport enzymes are repressed after exposure to serum (Supplementary Figures ??, G.12 and G.13). Notably, pyruvate kinase (EC 2.1.7.40 CDC19), acetyl CoA synthetase (EC6.2.1.1 A CS/ ), aldehyde dehydrogenase (EC 1.2.1.5 ALD3) and hexokinase (EC 2.7.1.1 GLK1) are all repressed whilst fructose-1,6-bisphosphatase I (EC 3.1.3.11 FBP1) Biotin, Co A and panthenoic acid biosynthetic genes are also induced (Supplementary Figure G.14 and Supplementary Figure G.15), suggesting a reduced production of pyruvate and an increased production of acetyl Co A. Induced carnitine transporter transcription and carnitine syn-

Appendix G. Transcription profiling of C. glabrata serum stress responses 335
thesis (EC 1.2.1.3 ALD5) with concomitant reduced fatty acid biosynthesis (Supplementary Figure G.16) suggest that unsaturated fatty acid oxidation is a preferred energy produc-tion pathway (this process requires acyl Co A and carnitine for uptake of fatty acids into the mitochondrion). This hypothetical decrease in carbon catabolism and carbon storage (Supplementary Figure G.17) indicates that carbon sources are diverted away from energy production pathways, which is perplexing as a glucose shortage is unlikely with the chosen experimental conditions (culture medium contains 2% (w/v) glucose and optical density of harvested cells was below saturation (0D600 = 0.45)).
Why are carbon catabolic energy production pathways down-regulated in favour of the hypothetical amino acid and fatty acid degradation proposed above? One explanation might be that carbon compounds are diverted to high mannose N-glycan biopolymer pro-duction in the cell wall. Exposure to serum leads to the differential transcription of genes whose products are either located in the cell wall or are involved in the modification of cell wall carbohydrates. For example, mannan polymerase II complex 1VINN10 subunit (EC 2.4.1 MNN10), the alpha-1,6-mannosyltransferase involved in cell wall mannan biosynthe-sis (HOC1) and the alpha-1,2-mannosyltransferase responsible for addition of the second alpha-1,2-linked mannose of the branches on the mannan backbone of oligosaccharides (EC 2.4.1 MNN5) are induced whilst mannosylphosphate transferase (EC 2.7.8 MNN6) is re-pressed (Supplementary Figure G.18). Furthermore, alpha-1,3-mannosyltransferase, which adds the fourth and fifth alpha-1,3-linked mannose residues to 0-linked glycans during pro-tein 0-glycosylation (EC 2.4.1 MNT3) is also induced (Supplementary Figure G.19) as is the v-snare SEC20 required for N- and 0-glycosylation in the Golgi (Supplementary Figure G.20). The induction of EXG1 EXG2 genes encoding exo-1,3-beta-glucanase and CWP1 a cell wall mannoprotein, linked to a beta-1,3- and beta-1,6-glucan provides further evidence in support of the hypothesis that serum treated C. glabrata cells undergo a programme of high mannose N-glycan biosynthesis and assembly in the cell wall. In addition, the induc-tion of UTR2 a chitin transglycosylase that functions in the transfer of chitin to beta(1-6) and beta(1-3) glucans in the cell wall suggest that newly synthesized and assembled beta glucans are also cross-linked to chitin. Further to the hypothetical reduction in carbon catabolism and increased amino acid catabolism, the induction of many transporters in the plasma membrane also provides clues regarding the nutritional demands of the serum treated cells. The co-induction of the genes encoding a putative divalent metal ion trans-porter involved in iron homeostasis (SMF3), a low affinity iron transporter (FET4) and a siderophore-iron chelate uptake transporter (ARN1) suggests that iron uptake is increased in this environmental perturbation. Furthermore, the co-induction of the biotin transporter gene VTH1 and biotin synthesis pathways, strongly suggest a nutritional requirement for this vitamin at this stage of growth in cells exposed to serum. Other vitamin transporter

G.3 Transcriptional response of C. glabrata cells exposed to 10% Foetal Calf Serum (FCS) 336
genes, whose protein products are located in the plasma membrane are also induced (the vitamin B2 transporter MGRS and the vitamin B6 transporter TPN6, which is a cofactor required for amino acid biosynthesis, whose upregulation is described above).
Cells exposed to foetal calf serum (10% v/v) also demonstrate differential transcrip-tion in non-metabolic functions relative to untreated cells, notably in the cell cycle (Figure G.22), a meiosis-like (Figure G.21) pathway and a MAPK signalling pathway (Figure G.23) inferred by orthology to S. cerevisiae. Inspection of the cell cycle pathway (Figure G.22) reveals serum-specific regulation of G1 phase (YOX1, PCL1 and CLN1), S phase (APC4 and CDC6) and 02 /M phase transcripts (RAD24, CHK1, CDC55 and PF2A). Although it is difficult to assess the dynamic nature of the cell cycle from one time point and also from dealing with transcripts in isolation rather than in an integrated mathematical model, speculatively one might suggest that the induction of YOX1 and CLN1 indicates the cells can progress from G1 phase to G2 phase. Intriguingly, genes involved in orthologous mating, filamentation and sporulation pathways of S. cerevisiae are induced after exposure to serum. Two genes encoding pheromone regulated S. cerevisiae orthologues (a transcription factor required for gene regulation in response to pheromones, Kar4p and Prm3p, a pheromone-regulated protein required for nuclear envelope fusion during karyogamy) in addition to a cAMP dependent protein kinase involved in regulating vegetative growth (Tpk2p), and two genes encoding Mat al orthologues (CAGLOB00242g/YCRO4OW and CAGLOB01243g/ YCR040W) indicate an activation of pathways whose orthologous functions in S. cerevisiae are to regulate the mating response pathway. Mapping of regulated transcripts to ortholo-gous MAPK pathways (Figure G.23) is consistent with this. For example, the concomitant induction of SHO1 and TEC1 and repression of MSN2 suggests an activation of the fil-amentation response signalling cascade rather than the hyperosmotic stress pathway that also lies downstream of Sholp. The induction of BEM1, a gene encoding a protein involved in establishing cell polarity and morphogenesis is also induced and two genes encoding sporulation specific genes (DIT1 and TPK2) was also observed in serum treated cells.

Appendix G. Transcription profiling of C. glabrata serum stress responses 337
Ad
6
no normalization
1 2 3 4 5 8 7 8 9 11 13 15
log2(Cy6 / Cy3)
(a)
RG densities
10
15
Intensity
(c)
loess + quantile normalization
2
1 2 3 4 5 6 7 8 9 11 13 15
log2(Cy5 / Cy3)
(b)
RG densities
o
5 10
15
Intensity
(d)
0 o
Figure G.1: Exploratory data analysis plots for C. glabrata environmental stress microarray hybridizations. The effects of normalization procedures on background cor-rected intensity distributions prior to linear modelling analysis. Box and whisker plots of M values (log2(Cy5TM) - log2(Cy3Tm )) are shown for each slide before normalization (a) and after lowess normalization and background correction (b). Coloured boxes depict the en-vironmental stress added in each experiment. Black boxes = synthetic complete + 10% (v/v) foetal calf serum (FCS), red boxes = synthetic medium + 0.01% dextrose (w/v), blue boxes = synthetic complete + 0.5M KC1 and blue boxes = synthetic complete + 1mM tertiary butyl hydroperoxide (t-BOOH). The pattern of boxes repeats four times, account-ing for each of the four biological replicates. The Cy5
TM (red) and Cy3TM (green) intensity
density distributions are plotted before (c) and after normalization (d). Both channels are comparable after normalization.
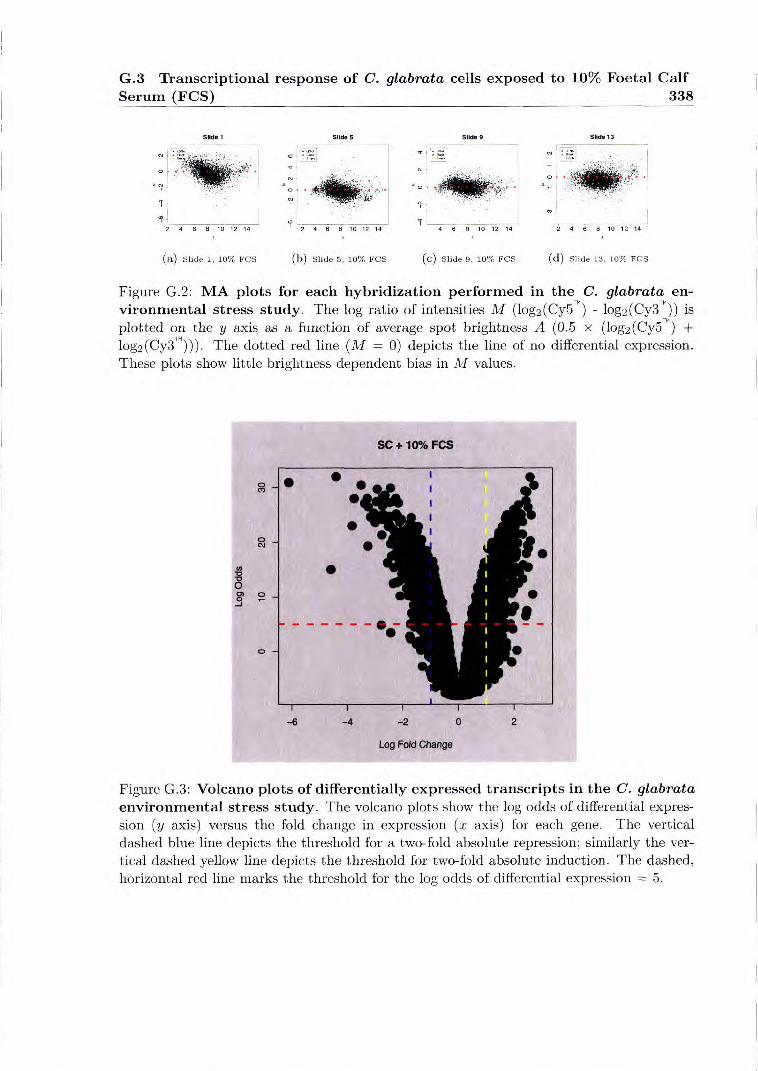
6 8 10 12 14 2 4 8 8 10 12 14
0
2 4 6 8 10 12 14
• •
•
SC + 10% FCS
0
01 -0
0 C0 0 0 _J
O
-6 -4 -2
0
2
Log Fold Change
G.3 Transcriptional response of C. glabrata cells exposed to 10% Foetal Calf Serum (FCS) 338
Slide 1
Slide 5
Slide 9
Slide 13
2 4 6 8 10 1.2 14
(a) Slide 1, 10% FCS (b) Slide 5, 10% FCS (C) Slide 9, 10% FCS (d) Slide 13, 10% FCS
Figure G.2: MA plots for each hybridization performed in the C. glabrata en-vironmental stress study. The log ratio of intensities M (log2 (Cy5TM ) - log2(Cy3T )) is plotted on the y axis as a function of average spot brightness A (0.5 x (log2(Cy5''') log2(Cy3TM))). The dotted red line (M = 0) depicts the line of no differential expression. These plots show little brightness dependent bias in M values.
Figure G.3: Volcano plots of differentially expressed transcripts in the C. glabrata environmental stress study. The volcano plots show the log odds of differential expres-sion (y axis) versus the fold change in expression (x axis) for each gene. The vertical dashed blue line depicts the threshold for a two-fold absolute repression; similarly the ver-tical dashed yellow line depicts the threshold for two-fold absolute induction. The dashed, horizontal red line marks the threshold for the log odds of differential expression = 5.

Appendix G. Transcription profiling of C. glabrata serum stress responses 339
TNT
11
1
Figure G.4: Genes differentially transcribed following exposure to 10% FCS (1 hour) mapped to the orthologous global metabolic map of S. cerevisiae. Cells were incubated for 4 hours (0D600 = 0.8) and harvested using a Sorvall SLA-3000 ultra-centrifuge rotor (suitable for large volume harvesting), washed with sterile distilled water and then subcultured in synthetic culture medium (reference sample) or synthetic com-plete medium supplemented 10% foetal calf serum (FCS). RNA Isolation, cDNA labelling, microarray hybridizations and data analyses are fully described in the the materials and methods section (Chapter 2). Pathways containing induced and repressed genes are coloured black and dark blue respectively.

1 2 6.1571 1 2,6.19
Tymaine yieC Isoquinoline alkaloid biosynthesis)
1 '5 1 (- Puronnycin biosynthesis
( Tyrosine metabolism ) TIVIICStanDia
PHENYLALANINE, TYROSINE AND TRYPTOPHAN BIOSYNTHESIS
Panama phosphate pethoray
L-Aspirtale 4-semiakiehyle
6-Deoxy-5-ketofructose phosOiale
7P-2-Dehydra-3-cleory-D arabwo-heptatate
04-1 QuiC 1-10.0stale rho- mate Protocatechua*
Olycolysis}-- i> Phosphoenol-pyrwate
• CEIZI A• T 3-De®te 142.1.101
3-Deh
OEM 11.199251 1111221
2-Anutio-3,7-chdeoxy-D-threo-liept-6-olosonic acid
111.121521 Photosyntliesis)-
1
; EEC D-Erythro 4-phosp1
seate
Indole i' 9*142.1.20
-1"( Itynthesbiosynthesis1 14.2.1281 be
1
1 ll! ! (3-1ncloy1)- jj. L-Tryplophan 4-142.1201-1e0 81rrol
# P11.9hle
(azet) — I V
14.1.1.481
Indole alkaloid) iosynthesis
E 1445-Phosiho 8-D-nloos anthianila
(-Acridone alkaloid biosynthesis }it
(- Phynyialenme metabolism )
4 [Biosylithesisofhopine,
pyrinkadine, rec and pipeiiiline allealciels
onne 1
1 4 !
(Phenyl panoid) i L binsyn sis
00400 8128109 (c) Kanehise Laboratories
5-0-(1-Carboxyriny1)- posynthesis of ilinlielrnt'n117471ides
1
Rotate biosynthesis
EMS (Ubiquinone and other tert:enced-cpanone biosynthesis
4-H n private
• 3-phosphoshikarriabe
11.3.1.121 26.1.57
Pre sine 0
14.2.1.511 .0 1
mere_ 14.2.1.911I alatine 11.141611 1
1 V
1 2.6..1 .1
♦ 1-(2-C
° 14!cliaTiggifriulose 5-phosphale
PAPP 0 hathreculate
*12 4.2 1812-1004-14.13.27 c6,6.....te T 1
15499 51 Prephenste
Phenyl.-pyrwete
2.6.1.1 2.6.1.5 1 1 2.61.9 12.61.571
114 201
41r
126.151
Shlimate
Slukimate 3-phosphate
G.3 Transcriptional response of C. glabrata cells exposed to 10% Foetal Calf Serum (FCS) 340
Figure G.5: Genes differentially transcribed following exposure to 10% FCS (1 hour) mapped to the orthologous phenylalanine, tyrosine and trytophan biosyn-thetic pathway of S. cerevisiae. Cells were incubated for 4 hours (0D600 = 0.8) and har-vested using a Sorvall SLA-3000 ultracentrifuge rotor (suitable for large volume harvesting), washed with sterile distilled water and then subcultured in synthetic culture medium (ref-erence sample) or synthetic complete medium supplemented 10% foetal calf serum (FCS). RNA Isolation, cDNA labelling, microarray hybridizations and data analyses are fully de-scribed in the the materials and methods section (Chapter 2). Induced and repressed genes are coloured yellow and dark blue respectively.

PHENYLALANINE, TYROSINE AND TRYPTORIAN BIOSYNTHESIS
4.1.1.51
( Phynyialanine metabolism ) 0 Piety/way 0
Ithigainorie and other 4-H terpenoid-gasonebiosyntlems ) ydioxy- phenylpyruvate
( Pentonphosphide) pathway
Photosynthesis)- 1
Quints
111.1.241 2-Athino-3,7-cluteoxy- 1119925 111 2s2 I D-6areo-1M,1- 6-ulosomc acid
,Lha ,D-EzYthose . mg] • nu] 1 4 1 - 1-
7F-2-Dehyds, 1 4211.13.1-11DrYthmluiwr (- Glyrolysis )-- — — -1> 3-deozy-
6-Deoxy-5-ketufructose Phosphoenol- D-arabuth- pynivate
, _ 111 1 n,,o„dm_, 1-phospmle heptouate
41-1 1 QuiC -m0 stikima --,t-i,"- Proton:decimate
L-Aspartata 4-sereialdelmnle •
lird:1:6-142.1 20 f 4,C Ptensostainoue 1 1
biosynthesis LEEE3
1 ; (3-Indoy1)- 1 L-Tryrtophan 0.6-142J 2:11÷ 4 phos 31•Yuem phate
(LYttbteoft}F—j1 2-C 14.1.1.431
Indo
‘1, 1-(
ie alkaloid ° '-fhvdtrrIrTrnbulose
biosynthesis 5- oh* PAPP 0
13Ngnb-Fhwosj) h° anduanilS
(Acridone alkoloidbiosynthesis
Shikimata
Slultimate 3-phosphate
125.1.191
t 5-0-(1-Cothoryvino0-3-phosphoshithinere
c Biosynthesis of ) no$iderueosormarrquolos
4
Polete bicsrth.
CE€E1 Anthrenilate 94-14.1.3 2T I-0 Chorisraate
15.4_99.51 Prephenate
(Biospfitesis of hapane, "1 • pyirshr.dine, nicotme and 1 ppencline andante , 1
(gitegCtin'90 .0 rhecLk.nyi 1.1416.1 1 1
1
( Tyrosine metabolism )
1 1 4.120 1 Trosias
Isom-timbre alkaloid biosynthesis)
2.611 2.6.1.5
2.6.19 2.6.1.57
1 1 ( Pthranycinbiosynthesis )
bioPhesnynVsP2's
00400 8078/09 (c)Kanehise Laborablies
Appendix G. Transcription profiling of C. glabrata serum stress responses 341
Figure G.6: Genes differentially transcribed following exposure to 10% FCS (1 hour) mapped to the orthologous valine, leucine and isoleucine biosynthetic pathway of S. cerevisiae. Cells were incubated for 4 hours (0D600 = 0.8) and harvested using a Sorvall SLA-3000 ultracentrifuge rotor (suitable for large volume harvesting), washed with sterile distilled water and then subcultured in synthetic culture medium (reference sample) or synthetic complete medium supplemented 10% foetal calf serum (FCS). RNA Isolation, cDNA labelling, microarray hybridizations and data analyses are fully described in the the materials and methods section (Chapter 2). Induced and repressed genes are coloured yellow and dark blue respectively.

0(S)-3-Hydroxy isobutyrate
2-Methyl-acetorreetyl-CoA
123.1161
•
04-142.1.171-00 3-Hydroxy- isovaleryl-00A I 6.1 -.l 11.ViethYlbui-2-en°34-C°A
+3 Mathyl-glutaconyl-CoA
14.2.1.181 (8)-3-Hydroxy-3-methytglutaryl-CoA v
MEE IBM
Thpp 2-Methyl- 1-11 xyb DPP
Lipoarnide-E
•
VALINE, LEUCINE AND ISOLEUCINE DEGRADATION
V aline, leucine and ) isolerxine biosynthesis
3R)-I3-Leucine ,p-xetoisocaproate 011-1 1.4.1.9 I-1100 0 L-V aline
2.6.1.42
•
1.2.4.4
L-Isoleucine
2.6.1.421 V(S)-3-Methyl-2-osoicentanoate
MEM
4-Methyl-2-oxopentanoate
3-Methyl-2-oxobutaitoate •
EEO
Lipoaniide-E
342-1•ktIrfil-Obutenoy1)-
dihydrolipoarnide-E
Branched chain fatty acid
3-Methyibutanoyl-CoA0 0
13.99.2
0 Acetoricette
1 1.2.13 1
thesis of type II )(1- -- I pol tide backbone 1 04—L5.4.99.21 I S ta: cinyl-Co A
c Biosynthesis of 12-, 14- and ,- —Lt
) ) 16-membered raernolides
.
00280 8.128.09 (c) Ranehisa Laboratories
•
11.3 99.21 1399.12 1 3 99 3 1.3.99.3
trans-2-Meilt3O- 0 but-2-enoyl-CoA
(S)--ldmxisob
3uty
Hril•CoA
2.33.10
4.1.3.4
01 2.6 1 18 V Proicanoyl-CoA
PO=
CCitrate cycle)
Boos
3-(2-Methyl-0 peopangy0-
th ydrofipoaude-E Dihyclro-lipoantide-E
OLsobntyryl CoA (S)-2-Meth3d- butenoyl-CoA
0 18 )-3.Hydrons .2-meth-CoAi
hcetoacetyl-CoA 0.4-1
23.19 Acetyl-CoA
I (8)-3-Amino-psi , asobutanoate 126.1
1 1 V
• 0-112.1.271 22 I (sSeLIVatelt2h4fialoe nate
1.2.3.1 1 Pyrimidine metabolism
1
1
16.4.1.3 I c PrneVaTiVeitt )
,IrtR)-Methyl- rnalonyi-CoA i
OMethylinalonate
9 2c)tioMeyln ttA 011-1 5 1.99 11
1399.121 11.3.99.-1
is 0 Metheczylyi-CoA
L.Leucine le7 0 5.4.3.7
1.4.19 2.6.14 12.6.1.421
• -Methyl- 1_h h xypropylo EIMP
11.8..41 Dihyito- 1 lipocattide-E 0
Liproatnide-E
I 2 44 3-Methyl-1,1lathoxybutyl- 0 mpe
G.3 Transcriptional response of C. glabrata cells exposed to 10% Foetal Calf Serum (FCS) 342
Figure G.7: Genes differentially transcribed following exposure to 10% FCS (1 hour) mapped to the orthologous valine, leucine and isoleucine degradation pathway of S. cerevisiae. Cells were incubated for 4 hours (0D600 = 0.8) and harvested using a Sorvall SLA-3000 ultracentrifuge rotor (suitable for large volume harvesting), washed with sterile distilled water and then subcultured in synthetic culture medium (reference sample) or synthetic complete medium supplemented 10% foetal calf serum (FCS). RNA Isolation, cDNA labelling, microarray hybridizations and data analyses are fully described in the the materials and methods section (Chapter 2). Induced and repressed genes are coloured yellow and dark blue respectively.

iakoadr miA®
1311 1
0-13 1433 541Acm.
L04.4 'rr"*
5313
193141 1512
• MEI
n.14216 m!*1 )
ERIE
1143221 1161331 1.61,1
11=1'.
L' sI 11.119 111 I Lavenzazor• 113111 .2
1141331
157
I P..al
I N. I rG
F.r,
u.=.1
• VIIE7 •
pm =I • 4.11E1 ,,Por..
L211=
310 1=
1]13 Ace
*so- Wm.
Aisco...• I
I3 331 123.111
I ARGINICE MD ellouNE METABOLISM 1
Daippilm• 1141cepoc
970a EEO •
112341 1,111•1w__.•41 )
I ADNp 4q I
tralarea.
12,3,1 [1141331
a off,
Rrin3s9
0-- 01-1 2331
13113
]3101 MO • ELM •
/113
161 if r,;,1 11". -t 31-- 231_,
riteig11
133.3.1
• [D
Appendix G. Transcription profiling of C. glabrata serum stress responses 343
Figure G.8: Genes differentially transcribed following exposure to 10% FCS (1 hour) mapped to the orthologous arginine and proline metabolic pathway of S. cerevisiae. Cells were incubated for 4 hours (0D600 = 0.8) and harvested using a Sorvall SLA-3000 ultracentrifuge rotor (suitable for large volume harvesting), washed with sterile distilled water and then subcultured in synthetic culture medium (reference sample) or synthetic complete medium supplemented 10% foetal calf serum (FCS). RNA Isolation, cDNA labelling, microarray hybridizations and data analyses are fully described in the the materials and methods section (Chapter 2). Induced and repressed genes are coloured yellow and dark blue respectively.
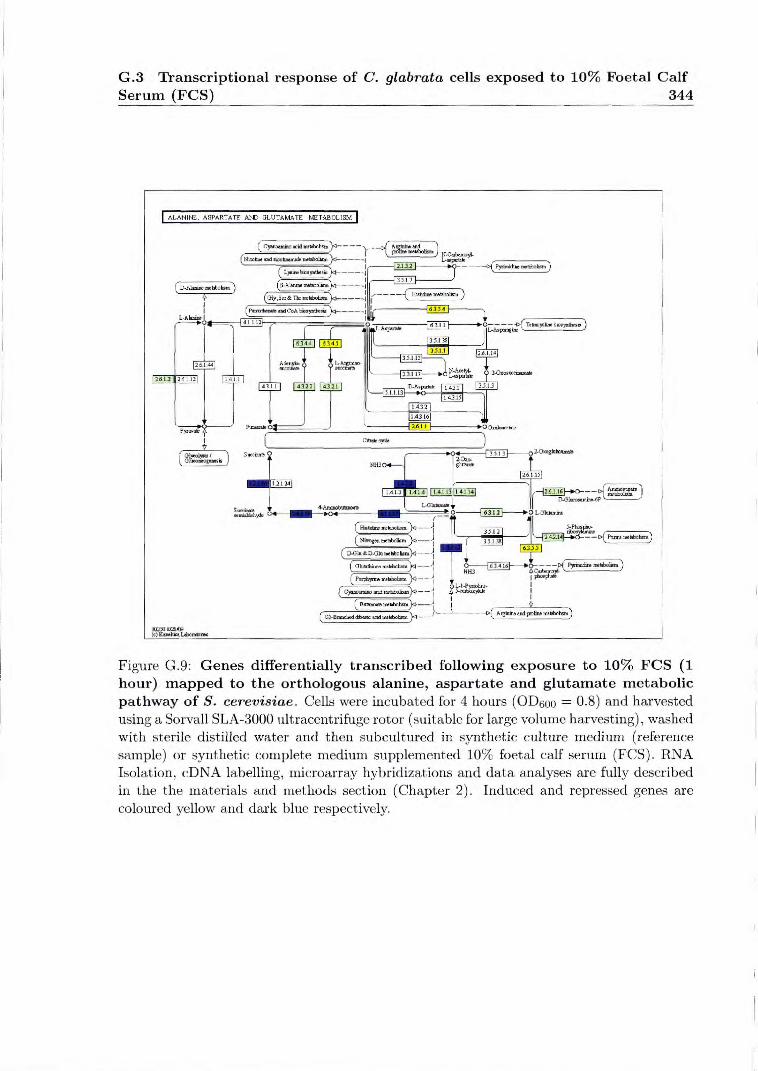
ALANINE. ASP ARTATE AND GLUTAMATE METABOLISM
1 26.11 14
Cr1ounine acid mete1c1c6.}.-- — ,==,11611()
(1,16)166 cad motabeaucle notebalca6}6---- Ls - P'nk —DK Prialid66 noticohcrn )
Lywinebicoyetl(es1)(6----
(13-Alcrace maticolicra)o---- C D-Alanitembl)obc1. ) 11.1bdue cnstisolima ) (Gly,Scr & Th:
(Pactiottenes 161051 biosyntocic — LAS I 41.1121
X63441
Aden*.
Ciente cede
6711 1 0 -1>(Tettacychne Innyansars L-Aspaii L-Acpc6bs )
16163
uni 11141.141 ...__mwttcrbusrancle L-GlAuare 4-116.1"
D-Glecosemic.61
‘-- 11'r 0 --I 63.11 I 0 1.,,Iniusine
01166666 61.616ohem.)cl (1,
Pc(66)61. nebbobera )0--1 Cycooanuco col reetabolom)./ — —
I
- Are
CS-Branci6oldcbanc end (6611)horn )(3 — ad polhu .1*ohm
(Rist616 6.66bolam ).3 -- p. ( Nitrogen metabolism )--1 LEM
V L-1-Pynohne-4 3-63.6.zylale
-)NOIT-1 6 3 416 — melba=
4
X6334 1-
Pyncede
GTuvaeogenii
26.
2 6 1 121
00250 1621614 (c) Kula. LAor6onm
G.3 Transcriptional response of C. glabrata cells exposed to 10% Foetal Calf Serum (FCS) 344
Figure G.9: Genes differentially transcribed following exposure to 10% FCS (1 hour) mapped to the orthologous alanine, aspartate and glutamate metabolic pathway of S. cerevisiae. Cells were incubated for 4 hours (0D600 = 0.8) and harvested using a Sorvall SLA-3000 ultracentrifuge rotor (suitable for large volume harvesting), washed with sterile distilled water and then subcultured in synthetic culture medium (reference sample) or synthetic complete medium supplemented 10% foetal calf serum (FCS). RNA Isolation, cDNA labelling, microarray hybridizations and data analyses are fully described in the the materials and methods section (Chapter 2). Induced and repressed genes are coloured yellow and dark blue respectively.

LYSINE DEGRADATION
6-Acetemielo-7osolesanome
2.6.121 D-Lyika
MEI • ECHE •
20+1 41-126.139 le&11-112.1.311--04 5 W PB
tmelelehy6 1-2-Ammo./..11.51101
9,491. 6-m
1-Fiyoritkim
LP s 141.
NEU EIFEEI 21,Ao.tyl-
23132 EICLIEEI 611.erba¢7 .-
1513 4
• Amin 7611
•
2.1.1A3i • MD 6--
EEMI
lEICI ECIE:1 • 0 5-Phooltrato 'Merino
5.0olecloeybsyleaine
6-Ammo-2-ovate:teem
(Pectioey
odline and cepholooyatins), Nnthecis
5-Asoiao-pertemeate
• ECM 011111=1/3 0
higfteyla hecar 3117 0-1632.-1-9.0
(1.-risetoosymn..-)
---- SAM meYl>iliem )
EMI •
rdleti790.1;elleh")'
Clove®
N-4.13413••••
Acetyl-GA? 13a Patio.
Me
le4s-
4Trunethyl moteemilmemmete
1141111-1.0 Coring •
heyermotte 2,5-Dtemorm 5-orsa hexane A
5112
Aceternylo 2-oxottemoste
5.1.191 5.1.1.51 3,5-Domno. 5-Ataino3 -
ocoltennoak 4.1.11 Mai ,154.3.2 • EMI •
1.5.1.7
.61-Pilyricleine-2-taktorylete
1.4314
Proteo-H. 16-.1.1**4
• EMI • MHO • MEI • Tytelltyl.
0 Glwme MotetItyltestne
47 21.142 21159 2.1160
EMU
2.1.143 211 21160
9031030109 Kuoeltiasitosehmee
Appendix G. Transcription profiling of C. glabrata serum stress responses 345
Figure G.10: Genes differentially transcribed following exposure to 10% FCS (1 hour) mapped to the orthologous lysine degradation pathway of S. cerevisiae. Cells were incubated for 4 hours (0D600 = 0.8) and harvested using a Sorvall SLA-3000 ultracentrifuge rotor (suitable for large volume harvesting), washed with sterile distilled water and then subcultured in synthetic culture medium (reference sample) or synthetic complete medium supplemented 10% foetal calf serum (FCS). RNA Isolation, cDNA la-belling, microarray hybridizations and data analyses are fully described in the the materials and methods section (Chapter 2). Induced and repressed genes are coloured yellow and dark blue respectively.

1.2.1.3
Ac+etyl-CoA
Acetate 04-
Starch awl sucrose ) metabolism
4
I GLYCOLYSIS 1 GLUCONEOGENESIS
• 3.1 39 2.7 1 69 D-Glucose
(extracellular)
1.4.1.1.321 -t>0
acaloacetate 4.1.1.49
1.2 7 1 ThPP •
1.2.4.1 e2thircl-ThPxpp-
1.1.1.1 1 1 99.8 Acetaldeh
• 41-11.1.1.271-11■0 1--Lactate Fynivate
CPropanoate metabolism)
2 7 2 3
g ce-D-Glacose-1P 3.1.3.10
ct-D-Glucose p 2312 27163
P-D-Glucos 23.1.63
Arbutim6P Arbutin (eximagar) L.) 12.71691—IP Salkin
(extraceltdar) kJ-12.7 1 691-10 S alicin-6P
-r> -D-Glucose-6P
5.13.15 5.3.1.9
O B-D-Glucose-6P
12 1 86 p-D-Fructose- 1,6P 2
3.2.186 4.1.2.13
elyceralciehyde-3P 04-1 5.3.1.11-0.0
Glycerone-P
(3_D-Frucbse-6P
1.2 1 12 1.2.1.59 Glycerate-1,3P 2
1.2 7.5 1.2.7.6 1 2 1 g
iCathort fixation in photosynthetic organisms
•
Private metabolism)
00010 8128/09 (c) 1Canehisa Laboratories
1.1 1 2 0 Ethanol
DAG
0 5 -Acetyl-
1.2.4.1 • 4.1.1.1
• 4.1.1.1
•
04-1 1.8.1.4 1-11. Dihydro- Lmoamide-E
Citrate cycle
2.7.1 41
•
Pentose PIT.sphate rammer
Phosphoenol-prmrate
G.3 Transcriptional response of C. glabrata cells exposed to 10% Foetal Calf Serum (FCS) 346
Figure G.11: Genes differentially transcribed following exposure to 10% FCS (1 hour) mapped to the orthologous glycolysis pathway of S. cerevisiae. Cells were incubated for 4 hours (0D600 = 0.8) and harvested using a Sorvall SLA-3000 ultracentrifuge rotor (suitable for large volume harvesting), washed with sterile distilled water and then subcultured in synthetic culture medium (reference sample) or synthetic complete medium supplemented 10% foetal calf serum (FCS). RNA Isolation, cDNA labelling, microarray hybridizations and data analyses are fully described in the the materials and methods section (Chapter 2). Induced and repressed genes are coloured yellow and dark blue respectively.

P32,1vale
t ----111/ 31
1122.2
123.61 S-Acetyl..
ermila-E celale Format
Ozaloscetele ?
0 Acely1CoA l
0 Acetillciekycie
L-Makte --KCzkale cycle)
(It)-2-Etleyirnalar
Buterrate metal:ohm A'ElY1-C°A
—1A(Laucute baosynlhems ) 3-Caboxy-3-hydroxy- 4-methylmtanoate
Homottret —M( Lymne basythasis )
1113.12.4
1,2,994
Melonyl-Co —14" Fet.yacielbiosyathams ) ofkebmbodies
2-Propylmilate
1. lac Washy& 313,tine, serim end Propane-1,24E A ,theanine
— — Olycerone-P
(31yeolysia \ ® MED
EEO • lisostalmead-Prwde
141131 4.1 1.32 4.1138
Mythylglyoxal
stIonedicaboxylate
P YRUVATE METABOLISM
3 7 9.2 1 1 , ??111
6.41.1 1
11.40
123.3 2.3121
Acetyl-P fOr
272.12 2.711 1
Tb.PP
127.1 1
30620 10121139 (c) Kanebisalahcastories
(motnax euvi Wilrelnide etabolism
IVAN% lanrim ami isoletrine biosynthesis
11127 115 .9 7
4.113
Acetyl-
• -
p 2.1.131
41-11.3.1.491-Ct.
D-Leclakieh3de
Appendix G. Transcription profiling of C. glabrata serum stress responses 347
Figure G.12: Genes differentially transcribed following exposure to 10% FCS (1 hour) mapped to the orthologous pyruvate metabolism pathway of S. cerevisiae. Cells were incubated for 4 hours (0D600 = 0.8) and harvested using a Sorvall SLA-3000 ultracentrifuge rotor (suitable for large volume harvesting), washed with sterile distilled water and then subcultured in synthetic culture medium (reference sample) or synthetic complete medium supplemented 10% foetal calf serum (FCS). RNA Isolation, cDNA la-belling, microarray hybridizations and data analyses are fully described in the the materials and methods section (Chapter 2). Induced and repressed genes are coloured yellow and dark blue respectively.

G.3 Transcriptional response of C. glabrata cells exposed to 10% Foetal Calf Serum (FCS) 348
Figure G.13: Genes differentially transcribed following exposure to 10% FCS (1 hour) mapped to the orthologous oxidative phosphorylation pathway of S. cerevisiae. Cells were incubated for 4 hours (0D600 = 0.8) and harvested using a Sorvall SLA-3000 ultracentrifuge rotor (suitable for large volume harvesting), washed with sterile distilled water and then subcultured in synthetic culture medium (reference sample) or synthetic complete medium supplemented 10% foetal calf serum (FCS). RNA Isolation, cDNA labelling, microarray hybridizations and data analyses are fully described in the the materials and methods section (Chapter 2). Induced and repressed genes are coloured yellow and dark blue respectively.

BIOTIN METABOLISM
Panelate 0 , Pimelovl-CoA rnino- 6.2.1.141-0-0 2.3.1.471---10.? toAxononanoate
Biotint- Biotinyl-CoA 5'-Ale
Detinobiotin ?4H 6.3.3.3
2.8.1.6
6 2 1 11
2 6 1 62
_011r 7,8-Diainino-noranoate
0 4 6 2.1 11 •
degradation
► 0— 3.4 - -
Holo- [cathoxylase]
• 0
Biotin J. .1.12 •
L-Lysine
Trope, .pericline anc0 id
iosynthesis
6.3.4.10 6.3.4.9
6.3.4.11 6.3.4.15
6 .3.4.10 6.3.4.9
6 3 4 15 6 3 4 11
00780 911/09 (c) Kanehisa Laboratories
Appendix G. Transcription profiling of C. glabrata serum stress responses 349
Figure G.14: Genes differentially transcribed following exposure to 10% FCS (1 hour) mapped to the orthologous biotin biosynthesis pathway of S. cerevisiae. Cells were incubated for 4 hours (0D600 = 0.8) and harvested using a Sorvall SLA-3000 ultracentrifuge rotor (suitable for large volume harvesting), washed with sterile distilled water and then subcultured in synthetic culture medium (reference sample) or synthetic complete medium supplemented 10% foetal calf serum (FCS). RNA Isolation, cDNA la-belling, microarray hybridizations and data analyses are fully described in the the materials and methods section (Chapter 2). Induced and repressed genes are coloured yellow and dark blue respectively.

PANTOTHENATE AND CoA BIOSYNTHESIS Olscolres )
•r7 0__H 2.2.1.6 ,i krif ietBIE
Eynerate
16.1.6 4 10.0 3-Methyl-7- 3.6.1.41 L. V aline obobutarbab
2-Debycleopentoete
(F1)-3,3-Dimethyhnalax
(R)-P oaken* 13-4-Phos bthenate Cysteme and metloonme
L-Cystaine l metaboborn (R).Panbata
(R)-4-Phospbc pantotkamo}4-1.-crieMe
Pareathatiot
Alm" ( Propenotee Uresil N-Cesbarno
metabolism
P-ATM
(- Mamma, aspadale and )_ — glutamate membolisna
kuspho-ranlethem
• 0
Adenosine 3',5'-berpltosplele
Alb-Wp] Dephospho-CoA 0-1111241 0
Acyl-canter Protein Peatetheme
A • L-Aepartate
00770 9,1J99 (c) )(audio(' Laboratone
G.3 Transcriptional response of C. glabrata cells exposed to 10% Foetal Calf Serum (FCS) 350
Figure G.15: Genes differentially transcribed following exposure to 10% FCS (1 hour) mapped to the orthologous pantothenoate and CoA biosynthesis pathway of S. cerevisiae. Cells were incubated for 4 hours (0D600 = 0.8) and harvested using a Sorvall SLA-3000 ultracentrifuge rotor (suitable for large volume harvesting), washed with sterile distilled water and then subcultured in synthetic culture medium (reference sample) or synthetic complete medium supplemented 10% foetal calf serum (FCS). RNA Isolation, cDNA labelling, microarray hybridizations and data analyses are fully described in the the materials and methods section (Chapter 2). Induced and repressed genes are coloured yellow and dark blue respectively.

'FAIT? ACID BIOSYNTHESIS-I
—(Caalf orb Acell.Caq,
ACP 0—, ASV
rpm IimmOL_Ctwil (F) Fr
Wear aid
COI ErIrcuboryl.
fonS1398109 (c)Kalehin Lakesieres
Appendix G. Transcription profiling of C. glabrata serum stress responses 351
Figure G.16: Genes differentially transcribed following exposure to 10% FCS (1 hour) mapped to the orthologous unsaturated fatty acid biosynthesis biosynthe-sis pathway of S. cerevisiae. Cells were incubated for 4 hours (0D600 = 0.8) and har-vested using a Sorvall SLA-3000 ultracentrifuge rotor (suitable for large volume harvesting), washed with sterile distilled water and then subcultured in synthetic culture medium (ref-erence sample) or synthetic complete medium supplemented 10% foetal calf serum (FCS). RNA Isolation, cDNA labelling, microarray hybridizations and data analyses are fully de-scribed in the the materials and methods section (Chapter 2). Induced and repressed genes are coloured yellow and dark blue respectively.

• MEI • ME • UDP-0 UDP
2.4.1.14, qd-Defielose.SP
UDP glazer IIIIII 2419
1.43111.43.6P Pnicbse
241.3 HPoSu. C7clen.4.4.44. ? Mabee Nxtrawkalma)
•1431k4Inlxie 5E40. 04
Colulose 111111111111,13.2.1.201 w Amy
,4p-D-01.44
( Nob* athglacutotele °444c" vetronverafors 2 1 31 J-4;81. 0-0-21owewtsich
)4L l'-1=2"'; PD
041 5 4 2 61 lk 4 pl-D-Ohicose-IP
1 311....LH41 3.2.1.93 1.0
3.Ketormase04--i I 159133 324 32 1 20 1 10,,,,,,,, 4--74-1.1 13
32 1 4 r 1 2.4.11 2413 1 241.131
0 D.Frodnee D-Oh.... 3.2 139 4-124.1341--
32138 1.3-13-Glocen ADP-glucaa
,41- 2.4.135 ...--1.0--1.1.1.21 (-12.41.121--' LI 6 1211 2.7.1.4 1 Cellthiose 37779 4 eligkeavose
al-P.71341,_,..— 4---) ci-D-0lums-1P
Dexem `-1 32.1.1
Khoo
( =kande gnu Intlabola. In Y4. CDP-gheese Anon mgu and EEO ‘-11=—.04 yie-D-Oluecte
2.7.1.10 v.17.01ucose-
•
.00-trecose
A.Dreennse.6P
0 kornakee
CtIldeiose Callcblreoss
o-D-Olunote4F Cellahophose Cellereninse Csa3those 0-13 2 1 741-Pa-13 a 1341+
Collotexam • EMI • EOM • MEI •
02500 8128.0? (01(8E313 4e 1.413ondo
I STARCH AND SUCROSE METABOLISM I Pwlut
Arnim eagv eN -q
Pod* 3 2 1101
Wit:" DOracta7oMir(
Satcoe4P uct.,:;;12 1.1591- s 4.
.!lestrwellira) sw.lross- 6P 32.1.251
4-0-011rose.64
13.D.Frucime
1321.741
D PrstosyrDa • EMS • ECOLCI
G.3 Transcriptional response of C. glabrata cells exposed to 10% Foetal Calf Serum (FCS) 352
Figure G.17: Genes differentially transcribed following exposure to 10% FCS (1 hour) mapped to the orthologous orthologous starch and sucrose metabolism pathway of S. cerevisiae. Cells were incubated for 4 hours (0D600 = 0.8) and harvested using a Sorvall SLA-3000 ultracentrifuge rotor (suitable for large volume harvesting), washed with sterile distilled water and then subcultured in synthetic culture medium (reference sample) or synthetic complete medium supplemented 10% foetal calf serum (FCS). RNA Isolation, cDNA labelling, microarray hybridizations and data analyses are fully described in the the materials and methods section (Chapter 2). Induced and repressed genes are coloured yellow and dark blue respectively.

HIGH-MANNOSE TYPE N-GLYCAN BIOSYNTHESIS
AIN1410 ANP1
MNNI I HOCI MNN9
MNN2 MUN5 1-11.0-111111F0•0-114NN1 Ho Maud 6Manal , MNN9
14N59 I. 6 2Manal MNN9 Manal 3Manal MNN2
LiN51 MNN5Manal MN 5 Manal Manal 3Manal 2Mana1 \
WW1 !Manal Man al 3Mana1 2Mana1 MNP2 149 Manal
mand / \
MNN1 WINN5 MNN9 Man al 3Manal 2Manal MNN2 6 1 Mani11-4GINAc 31— 4G1cNAc PI—Pun MNN1 MN145 MIII46
2 Mahal ] 6Manal / 3
Man al 3Man I —P., MNN9 OCHI 01111'51 5Man al MNN2
Mimi
Manal 3Manal / MNI41 MNI45 Man al 3Manal — 2Manal
MNN1
00513 8128109 (c)Kanehisa Laboralosien
• EMI 2.41232 OCH1
I1INN9 VANI1
C N-Olyranbionyallesis — —1>V-1321113 0-1 mow 1—ro
Appendix G. Transcription profiling of C. glabrata serum stress responses 353
Figure G.18: Genes differentially transcribed following exposure to 10% FCS (1 hour) mapped to the orthologous high mannose type N-glycan biosynthesis pathway of S. cerevisiae. Cells were incubated for 4 hours (0D600 = 0.8) and harvested using a Sorvall SLA-3000 ultracentrifuge rotor (suitable for large volume harvesting), washed with sterile distilled water and then subcultured in synthetic culture medium (reference sample) or synthetic complete medium supplemented 10% foetal calf serum (FCS). RNA Isolation, cDNA labelling, microarray hybridizations and data analyses are fully described in the the materials and methods section (Chapter 2). Induced and repressed genes are coloured yellow and dark blue respectively.

I 0-I'.NNOSYL GLYCAN BIOSYNTHESIS
Mania 3Mano(
Mani) I 2 hula n 15 .1
Man a t---- 2 Man a I/
Galpo:
3 3 2rulan a 2Mano( 1 2 2
/
Galp0:
2 Man Ser/Thr 2
GloNAo15
00514 412/09 (c) Kanads& Laboratories
G.3 Transcriptional response of C. glabrata cells exposed to 10% Foetal Calf Serum (FCS) 354
Figure G.19: Genes differentially transcribed following exposure to 10% FCS (1 hour) mapped to the orthologous orthologous o-mannosyl glycan biosynthesis pathway of S. cerevisiae. Cells were incubated for 4 hours (0D600 = 0.8) and harvested using a Sorvall SLA-3000 ultracentrifuge rotor (suitable for large volume harvesting), washed with sterile distilled water and then subcultured in synthetic culture medium (reference sample) or synthetic complete medium supplemented 10% foetal calf serum (FCS). RNA Isolation, eDNA labelling, microarray hybridizations and data analyses are fully described in the the materials and methods section (Chapter 2). Induced and repressed genes are coloured yellow and dark blue respectively.

PM !.plasma membrane)
I SNARE INTERACTIONS IN VESICULAR TRANSPORT
I Ykt6 LE date endosome)
Golgi body Sts
Sb:17 S Cc] 8 Use1
ER (ondoplasmic Miniboom) Sy-p7
S tc11 TGN
(trans Golgi network) STX6
Bost See22
I Batt
S Gast Ykt6
SNAP29
S ec20
Nucleus
04130 3/31/09 (c) Kanehisa Laboratories
apical PM (apical plasma membrane)
11 IVAMP4
RE (recycling endoseme)
VAMP?
I vAivrps (muscle)
IVAMP4
EE putty endeseme)
lysosome
Appendix G. Transcription profiling of C. glabrata serum stress responses 355
Figure G.20: Genes differentially transcribed following exposure to 10% FCS (1 hour) mapped to the orthologous orthologous v-snare vesicular trafficking pathway of S. cerevisiae. Cells were incubated for 4 hours (0D600 = 0.8) and harvested using a Sorvall SLA-3000 ultracentrifuge rotor (suitable for large volume harvesting), washed with sterile distilled water and then subcultured in synthetic culture medium (reference sample) or synthetic complete medium supplemented 10% foetal calf serum (FCS). RNA Isolation, cDNA labelling, microarray hybridizations and data analyses are fully described in the the materials and methods section (Chapter 2). Induced and repressed genes are coloured yellow and dark blue respectively.

MEICOSIS - yeast
CI 02 MI egx
Moine 0N4 Mtge C.1101iViat
BULL,
1=1
I PA
Am..= Sml 15me3
014
• s,2 I
= \ I/
=i M.77 MT
PR.1lc •j_il, 131..1 1 M I 7.1" F-1 C'I'' 1. \
EM rni NMI ', \ N.,: \ ' -1 4
C., 0 L.Np, + l
ir RopplogAza toy/ Span, dikearbly
.,-. I
?meek.. DNA reptialion
MP
PW•ltee Water LbStre
F Hcarolvwcltroacsom
CO3 liotrentattvetion O Noe amt
\
0 ',Iwo.
DtlyopoIii
Awl II
irtureni --' Act
moue. FrOglegn011. A
Y. MC..Ir.
ORC (0421 Taropm NICICIAnwalrooacare campkal Meinlemce) compler
Wm) Ora One,
oma Oad
G.3 Transcriptional response of C. glabrata cells exposed to 10% Foetal Calf Serum (FCS) 356
Figure G.21: Genes differentially transcribed following exposure to 10% FCS (1 hour) mapped to the orthologous meiosis pathway of S. cerevisiae. Cells were incubated for 4 hours (0D600 = 0.8) and harvested using a Sorvall SLA-3000 ultracentrifuge rotor (suitable for large volume harvesting), washed with sterile distilled water and then subcultured in synthetic culture medium (reference sample) or synthetic complete medium supplemented 10% foetal calf serum (FCS). RNA Isolation, cDNA labelling, microarray hybridizations and data analyses are fully described in the the materials and methods section (Chapter 2). Induced and repressed genes are coloured yellow and dark blue respectively.

r I
Spoe. chetpout DNA danve clack,mt UnNecled kireActers MSb.dspea DNA Mew ow,
•
hCV but
2412.4 025
C/01 Da4
Orel LAO Idan5
Mom]
GA 02
411111111111.6_1±11%11 q11114111.01. WI°
II ET
PAIR
I .21 I at I
CELL CYCLE
PA
5211 17=1 EMI IEZ7311 +1,
lizzrE3 Ell DC ehmo. o RAC I
r 'hog
wi
DNA ECE
Pb4 PIA]
I Rw3 I
1 aw Kt. It
TmecrAton of,20,*2
1444113A4 d. 0014AA2
S2 SN,4
ONA OAS
DNA S-Neda palsies
mp. ral F4A01944
sce,
ONG (000/44AgAtos AMA (2242-21424.0322 , 1442114241441 care,.
04111 Iona".
Appendix G. Transcription profiling of C. glabrata serum stress responses 357
Figure G.22: Genes differentially transcribed following exposure to 10% FCS (1 hour) mapped to the orthologous cell cycle pathway of S. cerevisiae. Cells were incubated for 4 hours (0D600 = 0.8) and harvested using a Sorvall SLA-3000 ultracentrifuge rotor (suitable for large volume harvesting), washed with sterile distilled water and then subcultured in synthetic culture medium (reference sample) or synthetic complete medium supplemented 10% foetal calf serum (FCS). RNA Isolation, cDNA labelling, microarray hybridizations and data analyses are fully described in the the materials and methods section (Chapter 2). Induced and repressed genes are coloured yellow and dark blue respectively.

MAPI MANGE MAPK
MAR{ SIGNALING PATHWAY
04011 3,31,99 (c) Kam hien Labotatoties
S te4 Stel$
G.3 Transcriptional response of C. glabrata cells exposed to 10% Foetal Calf Serum (FCS) 358
Figure G.23: Genes differentially transcribed following exposure to 10% FCS (1 hour) mapped to the orthologous MAPK signalling pathways of of S. cerevisiae. Cells were incubated for 4 hours (0D600 = 0.8) and harvested using a Sorvall SLA-3000 ultracentrifuge rotor (suitable for large volume harvesting), washed with sterile distilled water and then subcultured in synthetic culture medium (reference sample) or synthetic complete medium supplemented 10% foetal calf serum (FCS). RNA Isolation, cDNA la-belling, microarray hybridizations and data analyses are fully described in the the materials and methods section (Chapter 2). Induced and repressed genes are coloured yellow and dark blue respectively.

Appendix G. Transcription profiling of C. glabrata serum stress responses 359
Table G.1: Candida glabrata transcripts induced in ATCC 2001 following ex-posure to Synthetic Complete + 10% (v/v) foetal calf serum (FCS) Induced transcripts are defined as having log2(Cy5) - log2(Cy3) > 2.0 and Benjamini-Hochberg corrected p-values < 0.05.
CG ORF SC ORF gene name Description Log Ratio CAGLOI09108g YNL125C ESBP6 Protein with similarity to monocar-
boxylate permeases, appears not to be involved in transport of mono-carboxylates such as lactate, pyru-vate or acetate across the plasma membrane
8.15
CAGLOF00407g YJR070C LIA1 Deoxyhypusine hydroxylase, a 6.6 HEAT-repeat containing metal- loenzyme that catalyses hypusine formation; binds to and is re- quired for the modification of Hyp2p (eIF5A); complements S. pombe mmdl mutants defective in mitochondrial positioning
CAGLOG09086g - - Description not mapped 6.39 2cox3 - Description not mapped 6.21 rner - Description not mapped 6.06
CAGLOL03828g YNL111C CYB5 Cytochrome b5, involved in the sterol and lipid biosynthesis path-ways; acts as an electron donor to support sterol C5-6 desaturation
5.98
CAGL0003289g YLL048C YBT1 Transporter of the ATP-binding cassette (ABC) family involved in bile acid transport; similar to mam-malian bile transporters
5.77
CAGLOI00484g YLR300W EXG1 Major exo-1,3-beta-glucanase of the cell wall, involved in cell wall beta-glucan assembly; exists as three dif-ferentially glycosylated isoenzymes
5.69
CAGLOL10868g YOR271C FSF1 Putative protein, predicted to be an alpha-isopropylmalate carrier; be- longs to the sideroblastic-associated protein family; non-tagged protein is detected in purified mitochon- dria; likely to play a role in iron homeostasis
5.68
CAGLOK00605g YJL194W CDC6 Essential ATP-binding protein re- quired for DNA replication, compo- nent of the pre-replicative complex
5.61
(pre-RC) which requires ORC to as-sociate with chromatin and is in turn required for Mcm2-7p DNA as-sociation; homologous to S. pombe Cdcl8p
- - Description not mapped 5.54 CAGLOA00511g - - Description not mapped 5.54
- - Description not mapped 5.5 CAGLOE00792g YCL037C SR09 Cytoplasmic RNA-binding protein
that associates with translating ri-bosomes; involved in heme regula-tion of Hap1p as a component of the
5.45
HMC complex, also involved in the organization of actin filaments; con-tains a La motif
continued on next page

G.3 Transcriptional response of C. glabrata cells exposed to 10% Foetal Calf Serum (FCS) 360
continued from previous page CG ORF SC ORF gene name Description Log Ratio
CAGL0G01848g CAGLOG04741g
- YNL104C
- LEU4
Description not mapped Alpha-isopropylmalate synthase
5.38 5.32
(2-isopropylmalate synthase); the main isozyme responsible for the first step in the leucine biosynthesis pathway
CAGL0003531g - - Description not mapped 5.29 CAGLOG08668g YNL066W SUN4 Cell wall protein related to glu-
canases, possibly involved in cell wall septation; member of the SUN family
5.03
CAGLOF01485g YER011W TIR 1 Cell wall mannoprotein of the 5 Srplp/Tiplp family of serine- alanine-rich proteins; expression is downregulated at acidic pH and induced by cold shock and anaer-obiosis; abundance is increased in cells cultured without shaking
CAGLOM03487g YNR003C RPC34 RNA polymerase III subunit C34; 4.94 Be70 and is a
key determinant in pol III recruit-ment by the preinitiation complex
siown
Description
otinthTmFaIpHpd
CAGLOE06094g - - 4.93 CAGLOJ01848g YPRO10C RPA 135 RNA polymerase I subunit A135 4.79 CAGLOK01155g YGR079W - Description not mapped 4.69 CAGLOJ10252g YNL075W IMP4 Component of the SSU processome,
which is required for pre-18S rRNA processing; interacts with MpplOp; member of a superfamily of proteins that contain a sigma(70)-like motif and associate with RNAs
4.59
CAGLOJ01265g YMR093W UTP 15 Nucleolar protein, component of the small subunit (SSU) processome containing the U3 snoRNA that is involved in processing of pre-18S rRNA
4.5
CAGLOI11011g YARO5OW F LO 1 Lectin-like protein involved in floc- culation, cell wall protein that binds to mannose chains on the surface of other cells, confers floc-forming ability that is chy- motrypsin sensitive and heat resis-tant; similar to FloSp
4.48
CAGLOK00781g YGR210C - Description not mapped 4.47 CAGLOL03872g YNL113W RPC 19 RNA polymerase subunit, common
to RNA polymerases I and III 4.46
CAGLOJ06666g YML108W - Description not mapped 4.37 CAGLOLO8294g YIL14OW A XL 2 Integral plasma membrane protein
required for axial budding in hap-loid cells, localizes to the incipient bud site and bud neck; glycosylated by Pmt4p; potential Cdc28p sub-strate
4.34
CAGLOH01589g - - Description not mapped 4.29 CAGL0G08624g YIL121W QDR2 Multidrug transporter of the major
facilitator superfamily, required for resistance to quinidine, barban, cis-platin, and bleomycin; may have a role in potassium uptake
4.24
CAGLOG04499g YJL 105W SET4 Protein of unknown function, con- tains a SET domain
4.2
CAGLOG02409g YKR092C SRN° Nucleolar, serine-rich protein with a role in preribosome assembly or transport; may function as a chap-erone of small nucleolar ribonucle- oprotein particles (snoRNPs); im- munologically and structurally to rat Nopp140
4.15
continued on next page

Appendix G. Transcription profiling of C. glabrata serum stress responses 361
continued from previous page CG ORF SC ORF gene name Description Log Ratio
CAGLOL02937g CAGLOA04037g
CAGLOE02343g
-- YLR196W
YOLO1OW
PWPI
RCL I
Description not mapped Protein with WD-40 repeats in- volved in 'RNA processing; asso- ciates with trans-acting ribosome biogenesis factors; similar to beta-transducin superfamily Subunit of U3-containing 90S pre- ribosome processome complex in-volved in production of 18S rRNA and assembly of small ribosomal subunit; similar to RNA terminal phosphate cyclase-like proteins but RNA cyclase activity not detected
4.13 4.12
4.12
CAGLOL06072g YER130C COM2 Protein of unknown function 4.11 CAGL0G05874g - - Description not mapped 4.1 CAGLOE04092g YHL040C ARN1 Transporter, member of the ARN
family of transporters that specif- ically recognize siderophore-iron chelates; responsible for uptake of iron bound to ferrirubin, fer- rirhodin, and related siderophores
4.09
CAGLOL00671g YER056C FCY2 Purine-cytosine permease, mediates purine (adenine, guanine, and hy-poxanthine) and cytosine accumu-lation
4.08
CAGLOD05918g YGR177C ATF2 Alcohol acetyltransferase, may play a role in steroid detoxification; forms volatile esters during fermen-tation, which is important in brew-ing
4.06
CAGLOK02233g YER126C NSA2 Protein constituent of 665 pre- ribosomal particles, contributes to processing of the 27S pre-rRNA
4.05
CAGLOM11660g YIL053W RHR2 Constitutively expressed isoform of DL-glycerol-3-phosphatase; in- volved in glycerol biosynthesis, in-duced in response to both anaerobic and, along with the Hor2p/Gpp2p isoform, osmotic stress
4.04
CAGLOI08613g YHLO16C DUR3 Plasma membrane transporter for both urea and polyamines, expres-sion is highly sensitive to nitrogen catabolite repression and induced by allophanate, the last interme- diate of the allantoin degradative pathway
4
CAGLOD05060g YGR123C PPTI Protein serine/threonine phos- phatase with similarity to human phosphatase PP5; present in both the nucleus and cytoplasm; ex- pressed during logarithmic growth; computational analyses suggest roles in phosphate metabolism and rRNA processing
3.96
CAGLOF00187g YMR319C FEZ{ Low-affinity Fe(II) transporter of the plasma membrane
3.94
CAGLOF02013g YFLO10C WWM1 WW domain containing protein of unknown function; binds to
3.94
Mcalp, a caspase-related protease that regulates H202-induced apop- tosis; overexpression causes Gi phase growth arrest and clonal death that is suppressed by overex-pression of MCA1
continued on next page

G.3 Transcriptional response of C. glabrata cells exposed to 10% Foetal Calf Serum (FCS) 362
continued from previous page CG ORF SC ORF gene name Description Log Ratio
CAGL01102937g YJL069C UTPI8 Possible U3 snoRNP protein in- volved in maturation of pre-18S rRNA, based on computational analysis of large-scale protein- protein interaction data
3.93
CAGL0107689g YOL092W - Description not mapped 3.91 CAGL0003369g - - Description not mapped 3.9 CAGL0109746g YOR307C SLY4/ Protein involved in ER-to-Golgi
transport 3.9
CAGLOD04180g YIL091C - Description not mapped 3.88 CAGLOM07227g YHR196W UTP9 Nucleolar protein, component of
the small subunit (SSU) processome containing the U3 snoRNA that is involved in processing of pre-18S rRNA
3.88
CAGLOE02035g YOL119C MCH4 Protein with similarity to mam- malian monocarboxylate perme- ases, which are involved in trans-port of monocarboxylic acids across the plasma membrane; mutant is not deficient in monocarboxylate transport
3.86
CAGLOL07678g YPL266W DIM1 Essential 18S rRNA dimethy- lase (dimethyladenosine trans- ferase), responsible for conserved m6(2)Am6(2)A dimethylation in
3.86
3'-terminal loop of 18S rRNA, part of 90S and 40S pre-particles in nu- cleolus, involved in pre-ribosomal RNA processing
CAGLOJ11286g YNL182C 1P13 Essential component of the Rixl complex (Rix1p, Ipilp, Ipi3p) that is required for processing of ITS2 sequences from 35S pre-rRNA; highly conserved and contains
3.85
WD40 motifs; Rixl complex as- sociates with Mdnlp in pre-60S ribosomal particles
CAGLOB02343g YML116W ATRI Multidrug efflux pump of the major facilitator superfamily, required for resistance to aminotriazole and 4-nitroquinoline-N-oxide
3.84
CAGLOL04950g YMR049C ERD1 Protein required for maturation of the 25S and 5.8S ribosomal RNAs; constituent of 66S pre-ribosomal particles; homologous to mam- malian Bopl
3.79
CAGL0003938g YIL014W MNT3 Alpha-1,3-mannosyltransferase, adds the fourth and fifth alpha-
3.77
1,3-linked mannose residues to 0-linked glycans during protein 0-glycosylation
5i — Ugh - Description not mapped 3.75 CAGLOE01155g -- Description not mapped 3.75 CAGLOM01056g YDR339C FCF1 Putative PINc domain nuclease re-
quired for early cleavages of 35S pre-rRNA and maturation of 18S rRNA; component of the SSU
3.75
(small subunit) processome in- volved in 40S ribosomal subunit biogenesis; copurifies with Faflp
continued on next page

Appendix G. Transcription profiling of C. glabrata serum stress responses 363
continued from previous page CG ORF SC ORF gene name Description Log Ratio
CAGLOG04763g YORIO7W RGS2 Negative regulator of glucose- induced cAMP signaling; directly activates the GTPase activity of the heterotrimeric G protein alpha subunit Gpa2p
3.74
CAGLOL09581g YBR252W DUT1 dUTPase, catalyzes hydrolysis of dUTP to dUMP and PPi, thereby preventing incorporation of uracil into DNA during replication; crit-ical for the maintenance of genetic stability
3.73
CAGLOI06006g YJL148W RPA 34 RNA polymerase I subunit A34.5 3.71 CAGLOD03124g YLLOO8W DRS1 Nucleolar DEAD-box protein re-
quired for ribosome assembly and function, including synthesis of 60S ribosomal subunits; constituent of
3.69
66S pre-ribosomal particles CAGLOE02233g YNL289W PCL1 Pho85 cyclin of the Pc11,2-like sub-
family, involved in entry into the mitotic cell cycle and regulation of morphogenesis, localizes to sites of polarized cell growth
3.65
CAGLOK09416g YGL211W NCS6 Protein required for thiolation of the uridine at the wobble position of GIn, Lys, and Glu tRNAs; has a role in urmylation and in invasive and pseudohyphal growth; inhibits replication of Brome mosaic virus in
3.58
S. cerevisiae CAGLOL10890g YOR272W YTM1 Constituent of 66S pre-ribosomal
particles, required for maturation of the large ribosomal subunit
3.58
CAGLOJ00869g YKL144C RPC25 RNA polymerase III subunit C25, required for transcription initiation; forms a heterodimer with Rpc17p; paralog of Rpb7p
3.57
CAGLOJ09592g YDL2O9C CWG2 Protein involved in pre-mRNA splicing, component of a complex containing Ceflp; interacts with
3.51
Prpl9p; contains an RNA recog- nition motif; has similarity to S. pombe Cwf2p
CAGLOK09262g YCR059C YIH1 Protein that inhibits activation of 3.46 Gcn2p, an eIF2 alpha subunit pro-tein kinase, by competing for Gcnlp binding, thus impacting gene ex-pression in response to starvation; has sequence and functional simi-larity to the mouse IMPACT gene
CAGLOK06391g YDR462W MRPL28 Mitochondrial ribosomal protein of the large subunit
3.45
CAGLOG05093g YDR061W - Description not mapped 3.43 CAGLOM12430g YIL053W RHR2 Constitutively expressed isoform
of DL-glycerol-3-phosphatase; in- volved in glycerol biosynthesis, in-duced in response to both anaerobic and, along with the Hor2p/Gpp2p isoform, osmotic stress
3.43
9„, tp6 - - Description not mapped 3.41 CAGLOB01232g YLR186W EMG 1 Member of the alpha/beta knot
fold methyltransferase superfamily; required for maturation of 18S rRNA and for 40S ribosome pro- duction; interacts with RNA and with S-adenosylmethionine; asso- ciates with spindle/microtubules; forms homodimers
3.39
continued on next page

G.3 Transcriptional response of C. glabrata cells exposed to 10% Foetal Calf Serum (FCS) 364
continued from previous page CG ORF SC ORF gene name Description Log Ratio
CAGLOJ09746g YDL201W TRM8 Subunit of a tRNA methyltrans- ferase complex composed of Trm8p and Trm132p that catalyzes 7- methylguanosine modification of tRNA
3.38
CAGLOE00363g - - Description not mapped 3.37 CAGLOJ07766g YNL248C RPA4O RNA polymerase I subunit A49 3.37 CAGLOI09592g YOR303W CPAI Small subunit of carbamoyl phos-
phate synthetase, which catalyzes a step in the synthesis of citrulline, an arginine precursor; translation-ally regulated by an attenuator pep-tide encoded by YOR302W within the CPA1 mRNA 5'-leader
3.36
CAGLOF00253g YKL211C TRP3 Bifunctional enzyme exhibiting both indole-3-glycerol-phosphate synthase and anthranilate synthase activities, forms multifunctional hetero-oligomeric anthranilate synthase:indole-3-glycerol phos- phate synthase enzyme complex with Trp2p
3.35
6i — CglII - - Description not mapped 3.32 CAGLOF02431g YJL200C ACO2 Putative mitochondrial aconitase
isozyme; similarity to Acolp, an aconitase required for the TCA cycle; expression induced during growth on glucose, by amino acid starvation via Gcn4p, and repressed on ethanol
3.31
CAGLOH07975g YDL051W LHP1 RNA binding protein required for maturation of tRNA and U6 snRNA precursors; acts as a molecular chaperone for RNAs transcribed by polymerase III; homologous to hu-man La (SS-B) autoantigen
3.31
CAGL0105764g YNR009W NRMI Transcriptional co-repressor of 3.3 MBF (MCB binding factor)- regulated gene expression; Nrmlp associates stably with promoters via MBF to repress transcription upon exit from 01 phase
2 - Description not mapped 3.29 CAGLOB04543g YOR100C CRC.1 Mitochondrial inner membrane car-
nitine transporter, required for carnitine-dependent transport of acetyl-CoA from peroxisomes to mi-tochondria during fatty acid beta-oxidation
3.29
CAGL0.102398g YIL020C HIS6 Phosphoribosyl-5-amino- 3.29 1-phosphoribosy1-4- imidazolecarboxiamide isomerase, catalyzes the fourth step in his- tidine biosynthesis; mutations cause histidine auxotrophy and sensitivity to Cu, Co, and Ni salts
CAGLOJ00473g YHR052W C/C/ Essential protein that interacts with proteasome components and has a potential role in proteasome substrate specificity; also copurifies with 66S pre-ribosomal particles
3.28
continued on next page
Appendix G. Transcription profiling of C. glabrata serum stress responses 365
continued from previous page CG O RF SC ORF gene name Description Log Ratio
CAGLOI03608g YDL153C SASIO Essential subunit of U3-containing 3.27 Small Subunit (SSU) processome complex involved in production of 18S rRNA and assembly of small ri-bosomal subunit; disrupts silencing when overproduced
CAGLOK06215g YDR449C UTP6 Nucleolar protein, component of tpliescsmripatlliosnubnintitm(aSpSpUe)dprocessome containing the U3 snoRNA that is involved in processing of pre-18S rRNA
3.26
I,ox2 - - 3.2 CAGLOB00220g YLR186W EMG1 Member of the alpha/beta knot
fold methyltransferase superfamily; required for maturation of 18S rRNA and for 40S ribosome pro-duction; interacts with RNA and with S-adenosylmethionine, asso- ciates with spindle/microtubules; forms homodimers
3.24
CAGLOF04983g YLR276C DBP9 ATP-dependent RNA helicase of the DEAD-box family involved in biogenesis of the 60S ribosomal sub-unit
3.24
CAGLOK09284g YCR057C PWP2 Conserved 90S pre-ribosomal corn- ponent essential for proper endonu-cleolytic cleavage of the 35 S rRNA precursor at AO, Al, and A2 sites; contains eight WD-repeats; PWP2 deletion leads to defects in cell cy-cle and bud morphogenesis
3.24
CAGL0004279g YBROO2C RER2 Cis-prenyltransferase involved in dolichol synthesis; participates in endoplasmic reticulum (ER) pro- tein sorting
3.22
CAGLOL10648g YMR199W CLN1 G1 cyclin involved in regulation of the cell cycle; activates Cdc28p ki-nase to promote the GI to S phase transition; late G1 specific expres-sion depends on transcription factor complexes, MBF (Swi6p-Mbp1p) and SBF (Swi6p-Swi4p)
3.22
CAGLOM05621g YPL057C SUR/ Probable catalytic subunit of a mannosylinositol phosphorylce- ramide (MIPC) synthase, forms a complex with probable regulatory subunit Csg2p; function in sphin-golipid biosynthesis is overlapping with that of Cshlp
3.21
CAGLOM05775g YPL211W NIP7 Nucleolar protein required for 60S ribosome subunit biogenesis, con- stituent of 66S pre-ribosomal par- ticles; physically interacts with
3.21
Nop8p and the exosome subunit Rrp43p
CAGLO.J07590g - - Description not mapped 3.2 CAGLOK09460g YOR145C PNO I Essential nucleolar protein required
for pre-18S rRNA processing, in-teracts with Dim1p, an 18S rRNA dimethyltransferase, and also with
3.2
Nob1p, which is involved in protea-some biogenesis; contains a KH do-main
continued on next page

G.3 Transcriptional response of C. glabrata cells exposed to 10% Foetal Calf Serum (FCS) 366
continued from previous page CG ORF SC ORF gene name Description Log Ratio
CAGL0004213g CAGLOB02387g
YBROO5WP YMR285C
- NGL2
Description not mapped Protein involved in 5.8S rRNA pro- cessing; Ccr4p-like RNase required for correct 3'-end formation of 5.8S rRNA at site E; similar to Ngllp and Ngllp
3.19 3.18
CAGLOE00341g -- Description not mapped 3.18 CAGLOG06248g YAL025C MAK16 Essential nuclear protein, con-
stituent of 665 pre-ribosomal par- ticles; required for maturation of
3.17
25S and 5.8S rRNAs; required for maintenance of M1 satellite double-stranded RNA of the L-A virus
3,yb -- Description not mapped 3.16 CAGLOH02057g YHR089C GAR1 Protein component of the H/ACA
snoRNP pseudouridylase complex, involved in the modification and cleavage of the 18S pre-rRNA
3.15
CAGL0.110912g YHR065C RRP3 Protein involved in rRNA process- ing; required for maturation of the
3.15
35S primary transcript of pre-rRNA and for cleavage leading to ma- ture 18S rRNA; homologous to eIF- 4a, which is a DEAD box RNA-dependent ATPase with helicase ac-tivity
CAGLOE05522g YOR342C - Description not mapped 3.14 CAGLOF04499g YBL042C FUJI High affinity uridine permease, lo-
calized to the plasma membrane; not involved in uracil transport
3.14
CAGLOB04125g YPR110C RPC40 RNA polymerase subunit, common to RNA polymerase I and III
3.12
CAGLOG09108g YPL199C - Description not mapped 3.12 CAGLOH00935g -- Description not mapped 3.12 CAGLOH08019g YNL207W R102 Essential serine kinase involved in
the processing of the 20S pre-rRNA into mature 18S rRNA; has similar-ity to Rio1p
3.12
8,,, tp8 -- Description not mapped 3.1 CAGLOB00242g YCRO4OW MATALPHAI Transcriptional co-activator in-
volved in regulation of mating- type-specific gene expression; targets the transcription factor
3.1
Mcmlp to the promoters of alpha- specific genes; one of two genes encoded by the MATalpha mating type cassette
CAGLOE00979g YDR165W TRM82 Subunit of a tRNA methyltrans- ferase complex composed of Trm8p and TrmS2p that catalyzes 7- methylguanosine modification of tRNA
3.1
CAGL0107975g YOL077C BRXI Nucleolar protein, constituent of 3.1 66S pre-ribosomal particles; deple- tion leads to defects in rRNA pro- cessing and a block in the assem- bly of large ribosomal subunits; possesses a sigma(70)-like RNA- binding motif
continued on next page

Appendix G. Transcription profiling of C. glabrata serum stress responses 367
continued from previous page CG ORF SC ORF gene name Description Log Ratio
CAGLOB02057g YDR130C FINI Spindle pole body-related inter- mediate filament protein, forms cell cycle-specific filaments between spindle pole bodies in mother and daughter cells, able to self- assemble, expression induced dur-ing S/G2, localization cell-cycle de-pendent
3.09
CAGLOK09570g YNL130C CPT] Cholinephosphotransferase, re- quired for phosphatidylcholine biosynthesis and for inositol- dependent regulation of EPT1 transcription
3.08
CAGLOJ00715g YOR101W RASI GTPase involved in G-protein sig- naling in the adenylate cyclase activating pathway, plays a role in cell proliferation; localized to the plasma membrane; homolog of mammalian RAS proto-oncogenes
3.07
CAGLOL06138g YGL186C TPN1 Plasma membrane pyridoxine (vi- tamin B6) transporter; member of the purine-cytosine permease sub-family within the major facilitator superfamily; proton symporter with similarity to Fcy21p, Fcy2p, and
3.07
Fcy22p CAGLOH04939g YLR377C FBPI Fructose-1,6-bisphosphatase, key
regulatory enzyme in the gluco- neogenesis pathway, required for glucose metabolism; undergoes either proteasome-mediated or autophagy-mediated degradation depending on growth conditions; interacts with Vid3Op
3.06
CAGLOK12210g YBR104W YMC2 Mitochondrial protein, putative in- ner membrane transporter with a role in oleate metabolism and glu-tamate biosynthesis; member of the mitochondria] carrier (MCF) fam-ily; has similarity with Ymclp
3.05
CAGLOP00561g YJR063W RPA12 RNA polymerase I subunit Al2.2; contains two zinc binding domains, and the N terminal domain is re-sponsible for anchoring to the RNA pol I complex
3.04
CAGLOM02871g YJL186W MNN5 Alpha-1,2-mannosyltransferase, re- sponsible for addition of the sec-ond alpha-1,2-linked mannose of the branches on the mannan backbone of oligosaccharides, localizes to an early Golgi compartment
3.04
CAGLOM14113g YLR237W TH17 Plasma membrane transporter re- sponsible for the uptake of thi- amine, member of the major facil-itator superfamily of transporters; mutation of human ortholog causes thiamine-responsive megaloblastic anemia
3.04
continued on next page

G.3 Transcriptional response of C. glabrata cells exposed to 10% Foetal Calf Serum (FCS) 368
continued from previous page CG ORF SC ORF gene name Description Log Ratio
CAGLOB03773g
CAGLOF09207g
YGRI91W
YHR208W
HIP/
BAT1
High-affinity histidine permease, also involved in the transport of manganese ions Mitochondria] branched-chain amino acid aminotransferase, ho- molog of murine ECA39; highly expressed during logarithmic phase and repressed during stationary phase
3.03
3.03
CAGLOF08371g YGR26OW TNA1 High affinity nicotinic acid plasma membrane permease, responsible for uptake of low levels of nico- tinic acid; expression of the gene increases in the absence of ex- tracellular nicotinic acid or para-aminobenzoate (PABA)
3.02
CAGLOL10516g YKR081C RPF2 Essential protein involved in the processing of pre-rRNA and the as-sembly of the 60S ribosomal sub-unit; interacts with ribosomal pro- tein L11; localizes predominantly to the nucleolus; constituent of 66S pre-ribosomal particles
3.02
CAGLOH10010g - - Description not mapped 3.01 CAGLOJ03476g YCR072C RSA4 WD-repeat protein involved in ribo-
some biogenesis; may interact with ribosomes; required for maturation and efficient intra-nuclear transport or pre-60S ribosomal subunits, lo-calizes to the nucleolus
3.01
CAGLOE02893g YBL014C RRN6 Protein involved in the transcrip- tion of 35S rRNA genes by RNA polymerase I; component of the core factor (CF) complex also com-posed of Rrnllp, Rrn7p and TATA-binding protein
2.99
CAGLOK01771g YAR035W YAT1 Outer mitochondrial carnitine acetyltransferase, minor ethanol- inducible enzyme involved in transport of activated acyl groups from the cytoplasm into the mito-chondrial matrix; phosphorylated
2.99
CAGLOL03806g YNL11.0C NOP15 Constituent of 66S pre-ribosomal particles, involved in 60S ribosomal subunit biogenesis; localizes to both nucleolus and cytoplasm
2.99
2,, ar 1 - - Description not mapped 2.96 CAGLOF01023g YOL041C NOP12 Nucleolar protein, required for pre- 2.96
25S rRNA processing; contains an RNA recognition motif (RRM) and has similarity to Nopl3p, Nsrlp, and putative orthologs in Drosophila and S. pombe
CAGLOF04103g YBL028C - Description not mapped 2.96 CAGLOI00770g YMR144W - Description not mapped 2.96 CAGLOF05709g YDR184C ATC1 Nuclear protein, possibly involved
in regulation of cation stress re- sponses and/or in the establishment of bipolar budding pattern
2.95
CAGLOJ03916g YPL145C KES1 Member of the oxysterol binding protein family, which includes seven yeast homologs; involved in nega-tive regulation of Secl4p-dependent
2.95
Golgi complex secretory functions, peripheral membrane protein that localizes to the Golgi complex
continued on next page

Appendix G. Transcription profiling of C. glabrata serum stress responses 369
continued from previous page CG ORF SC ORF gene name Description Log Ratio
CAGLOL03025g CAGLOD04884g
YKR025W YPR137W
RPC37 RRP9
RNA polymerase III subunit C37 Protein involved in pre-rRNA pro- cessing, associated with U3 snRNP; component of small ribosomal sub-unit (SSU) processosome; ortholog of the human U3-55k protein
2.95 2.94
CAGLOH01309g YDR294C DPL1 Dihydrosphingosine phosphate lyase, regulates intracellular levels of sphingolipid long-chain base phosphates (LCBPs), degrades phosphorylated long chain bases, prefers C16 dihydrosphingosine-l- phosphate as a substrate
2.94
CAGLOL02849g YOR207C RET1 Second-largest subunit of RNA polymerase III, which is responsi-ble for the transcription of tRNA and 5S RNA genes, and other low molecular weight RNAs
2.91
CAGLOJ02222g YER002W NOP16 Constituent of 665 pre-ribosomal particles, involved in 60S ribosomal subunit biogenesis
2.9
CAGLOB01243g YCRO4OW MATALPHA1 Transcriptional co-activator in- volved in regulation of mating- type-specific gene expression; targets the transcription factor
2.89
Mcm1p to the promoters of alpha- specific genes; one of two genes encoded by the MATalpha mating type cassette
CAGLOL03846g - - Description not mapped 2.89 CAGLOD01606g YPRO58W YMC1 Mitochondrial protein, putative in-
ner membrane transporter with a role in oleate metabolism and glu-tamate biosynthesis; member of the mitochondrial carrier (MCF) fam-ily; has similarity with Ymc2p
2.88
CAGLOL13354g YLR237W THI7 Plasma membrane transporter re- sponsible for the uptake of thi- amine, member of the major facil-itator superfamily of transporters; mutation of human ortholog causes thiamine-responsive megaloblastic anemia
2.88
CAGLOM08074g YJR030C - Description not mapped 2.88 CAGLOM13563g YMR292W GOT1 Evolutionarily conserved non-
essential protein present in early 2.86
Golgi cisternae that may be in- volved in ER-Golgi transport at a step after vesicle tethering to Golgi membranes, exhibits membrane topology similar to that of Sft2p
CAGLOF08019g YGR239C PEX21 Peroxin required for targeting of peroxisomal matrix proteins con-taining PTS2; interacts with Pex7p; partially redundant with PexlSp
2.84
CAGLOM04631g YLR147C SMD3 Core Sm protein Sm D3; part of heteroheptameric complex (with
2.84
Smblp, Smdlp, Smd2p, Smelp, Smx3p, and Smx2p) that is part of the spliceosomal Ul, U2, U4, and U5 snRNPs; homolog of human Sm D3
continued on next page

G.3 Transcriptional response of C. glabrata cells exposed to 10% Foetal Calf Serum (FCS) 370
continued from previous page CG ORF SC ORF gene name Description Log Ratio
CAGLOM05599g YBR162C TOS1 Covalently-bound cell wall protein of unknown function; identified as a cell cycle regulated SBF target gene; deletion mutants are highly resistant to treatment with beta-
2.84
1,3-glucanase; has sequence similar-ity to YJL171C
CAGL0001441g YPL183C RTTIO Cytoplasmic protein with a role in regulation of Tyl transposition
2.83
CAGL00O2211g YELO40W UTR2 Cell wall protein that functions in the transfer of chitin to beta(1-
2.83
6)glucan; putative chitin transgly-cosidase; glycosylphosphatidylinos-itol (GPI)-anchored protein local-ized to the bud neck; has a role in cell wall maintenance
CAGLOH08712g YBR200W BEM1 Protein containing SI13-domains, involved in establishing cell polarity and morphogenesis; functions as a scaffold protein for complexes that include Cdc24p, Ste5p, Ste20p, and
2.83
Rsr 1p CAGLOJ00385g YHR049W FSH1 Putative serine hydrolase that lo-
calizes to both the nucleus and cy-toplasm; sequence is similar to S. cerevisiae Fsh2p and Fsh3p and the human candidate tumor suppressor
2.83
OVCA2 CAGLOJ00649g YHR025W THR1 Homoserine kinase, conserved pro-
tein required for threonine biosyn- thesis; expression is regulated by the GCN4-mediated general amino acid control pathway
2.81
CAGLOL02079g YBR291C GTP1 Mitochondria] inner membrane cit- rate transporter, member of the mi-tochondrial carrier family
2.81
CAGLOB00352g YCL059C KRR1 Essential nucleolar protein required for the synthesis of 18S rRNA and for the assembly of 40S ribosomal subunit
2.8
CAGLOF02849g YDR412W RRPI7 Component of the pre-60S pre- ribosomal particle; required for cell viability under standard (aerobic) conditions but not under anaerobic conditions
2.8
CAGL0106743g YER145C FTR1 High affinity iron permease involved in the transport of iron across the plasma membrane; forms complex with Fet3p; expression is regulated by iron
2.8
CAGLOA03014g YDR376W ARH1 Oxidoreductase of the mitochon- drial inner membrane, involved in cytoplasmic and mitochondrial iron homeostasis and required for activ-ity of Fe-S cluster-containing en-zymes; one of the few mitochondrial proteins essential for viability
2.79
CAGLOD01694g YPRO54W SMK1 Middle sporulation-specific mitogen-activated protein kinase
2.79
(MAPK) required for production of the outer spore wall layers; negatively regulates activity of the glucan synthase subunit Gsc2p
continued on next page

Appendix G. Transcription profiling of C. glabrata serum stress responses 371
continued from previous page CC ORF SC ORF gene name Description Log Ratio
CAGL0G01210g
CAGLOK10186g
YLR351C
YDR076W
NITS
RAD55
Nit protein, one of two proteins in S. cerevisiae with similarity to the Nit domain of NitFhit from fly and worm and to the mouse and human Nit protein which interacts with the Fhit tumor suppressor; nitrilase su-perfamily member Protein that stimulates strand ex- change by stabilizing the binding of Rad5lp to single-stranded DNA; involved in the recombinational re- pair of double-strand breaks in DNA during vegetative growth and meiosis; forms heterodimer with
2.79
2.79
Rad57p CAGLOM06435g YBR186W PCH2 Nucleolar component of the
pachytene checkpoint, which pre- vents chromosome segregation when recombination and chromo- some synapsis are defective; also represses meiotic interhomolog recombination in the rDNA
2.79
CAGLOM12122g YAL036C RBGI Member of the DRG family of GTP- binding proteins; interacts with translating ribosomes and with
2.79
Tma46p CAGLOF02695g YDR405W MRP20 Mitochondrial ribosomal protein of
the large subunit 2.78
CAGLOK01485g YOL066C R1132 DRAP deaminase, catalyzes the third step of the riboflavin biosyn-thesis pathway; cytoplasmic tRNA pseudouridine synthase involved in pseudouridylation of cytoplasmic tRNAs at position 32
2.78
CAGLOK08360g YKR056W TRM2 tRNA methyltransferase, 5- methylates the uridine residue at position 54 of tRNAs and may also have a role in tRNA stabilization or maturation; endo- exonuclease with a role in DNA repair
2.77
0,,. tp9 - - Description not mapped 2.76 CAGL00O2519g YER028C MIG3 Probable transcriptional repressor
involved in response to toxic agents such as hydroxyurea that inhibit ri-bonucleotide reductase; phosphory-lation by Snflp or the Meclp path-way inactivates Mig3p, allowing in-duction of damage response genes
2.76
CAGLOM10219g YKLOO8C LACI Ceramide synthase component, in- volved in synthesis of ceramide from C26(acyl)-coenzyme A and dihydrosphingosine or phytosphin- gosine, functionally equivalent to
2.76
Laglp CAGLOA04323g YEL057C PTH2 One of two (see also PTill)
mitochondrially-localized peptidyl- tRNA hydrolases; negatively regulates the ubiquitin-proteasome pathway via interactions with ubiquitin-like ubiquitin-associated proteins; dispensable for cell growth
2.75
continued on next page

G.3 Transcriptional response of C. glabrata cells exposed to 10% Foetal Calf Serum (FCS) 372
continued from previous page CG ORF SC ORF gene name Description Log Ratio
CAGLOB03883g YPRO58W YMC1 Mitochondrial protein, putative in- ner membrane transporter with a role in oleate metabolism and glu-tamate biosynthesis; member of the mitochondrial carrier (MCF) fam-ily; has similarity with Yme2p
2.75
CAGL0003575g YJR151C DAN4 Cell wall mannoprotein with simi- larity to Tirlp, Tir2p, Tir3p, and
2.75
Tir4p; expressed under anaero- bic conditions, completely repressed during aerobic growth
CAGLOL00341g YKL172W EBP2 Essential protein required for the maturation of 25S rRNA and 60S ri-bosomal subunit assembly, localizes to the nucleolus; constituent of 66S pre-ribosomal particles
2.75
CAGLOH02079g YHR088W RPFI Nucleolar protein involved in the assembly of the large ribosomal subunit; constituent of 66S pre- ribosomal particles; contains a sigma(70)-like motif, which is thought to bind RNA
2.74
CAGLOII10142g YDR035W ARO.? 3-deoxy-D-arabino-heptulosonate- 2.74 7-phosphate (DAHP) synthase, catalyzes the first step in aromatic amino acid biosynthesis and is feedback-inhibited by phenylala- nine or high concentration of tyrosine or tryptophan
CAGLOI05500g YER099C PRS2 5-phospho-ribosyl-1(alpha)- pyrophosphate synthetase, syn- thesizes PRPP, which is required for nucleotide, histidine, and tryp- tophan biosynthesis; one of five related enzymes, which are active as heteromultimeric complexes
2.74
CAGLOK00627g YJL193W Description not mapped 2.74 CAGL0.103278g YER077C Description not mapped 2.73 CAGLOB02926g - - Description not mapped 2.72 CAGLOI02354g YHR169W DBP8 ATPase, putative RNA helicase of
the DEAD-box family; component of 90S preribosome complex in- volved in production of 18S rRNA and assembly of 40S small riboso-mal subunit; ATPase activity stim-ulated by association with Esp2p
2.72
CAGLOK04477g YGRO6OW ERG25 C-4 methyl sterol oxidase, cat- alyzes the first of three steps re-quired to remove two C-4 methyl groups from an intermediate in er- gosterol biosynthesis; mutants ac- cumulate the sterol intermediate
2.72
4,4-dimethylzymosterol CAGLOM06941g YNL022C Description not mapped 2.72
- Description not mapped 2.71 CAGL0001617g - - Description not mapped 2.71 CAGLOE01903g YOL125W TRAD 3 2'-O-methyltransferase responsible
for modification of tRNA at posi- tion 4; exhibits no obvious simi- larity to other known methyltrans-ferases
2.71
continued on next page

Appendix G. Transcription profiling of C. glabrata serum stress responses 373
continued from previous page CG ORF SC ORF gene name Description Log Ratio
CAGLOF05753g YDR182W CDC1 Putative lipid phosphatase of the endoplasmic reticulum; shows
2.71
Mn2+ dependence and may affect Ca2+ signaling; mutants display actin and general growth defects and pleiotropic defects in cell cycle progression and organelle distribution
CAGLOJ01045g YJL033W fICA.4 Putative nucleolar DEAD box RNA helicase; high-copy number sup- pression of a U14 snoRNA process-ing mutant suggests an involvement in 18S rRNA synthesis
2.71
CAGLOJ02816g YER049W TPA1 Protein of unknown function; inter- acts with Sup45p (eRF1), Sup35p
2.7
(eRF3) and Pablp; has a role in translation termination efficiency, mRNA poly(A) tail length and mRNA stability
CAGLOJ11066g YLROO2C NOC3 Protein that forms a nuclear corn- plex with Noc2p that binds to
2.7
665 ribosomal precursors to me- diate their intranuclear transport; also binds to chromatin to promote the association of DNA replication factors and replication initiation
CAGLOG05610g YDL219W DTD1 D-Tyr-tRNA(Tyr) deacylase, func- tions in protein translation, may af-fect nonsense suppression via alter-ation of the protein synthesis ma-chinery; ubiquitous among eukary-otes
2.69
CAGLOG07106g YML022W APTI Adenine phosphoribosyltrans- ferase, catalyzes the formation of AMP from adenine and 5- phosphoribosylpyrophosph ate; involved in the salvage pathway of purine nucleotide biosynthesis
2.69
CAGLOI10670g YPR144C NOC4 Nucleolar protein, forms a complex with Nopl4p that mediates matura-tion and nuclear export of 40S ribo-somal subunits
2.69
CAGLOG01991g YOR056C NO131 Essential nuclear protein involved in proteasome maturation and syn-thesis of 40S ribosomal subunits; re-quired for cleavage of the 205 pre-rRNA to generate the mature 18S rRNA
2.68
CAGLOE04224g YBRO71W - Description not mapped 2.67 CAGLOI05016g YER090W TR P2 Anthranilate synthase, catalyzes
the initial step of tryptophan biosynthesis, forms multifunctional hetero-oligomeric anthranilate synthase.indole-3-glycerol phos- phate synthase enzyme complex with Trp3p
2.67
CAGLOI09438g YBR271W - Description not mapped 2.67 CAGLOE02409g YOLOO7C CS/2 Protein of unknown function; green
fluorescent protein (GFP)- fusion protein localizes to the mother side of the bud neck and the vacuole;
2.66
YOLOO7C is not an essential gene continued on next page

G.3 Transcriptional response of C. glabrata cells exposed to 10% Foetal Calf Serum (FCS) 374
continued from previous page CG ORF SC ORF gene name Description Log Ratio
CAGLOF01177g CAGLOG10043g
CAGLOJ10010g
YOL036W YPR187W
YNL062C
- RP026
GCD10
Description not mapped RNA polymerase subunit ABC23, common to RNA polymerases I, II, and III; part of central core; similar to bacterial omega subunit Subunit of tRNA (1- methyladenosine) methyltrans- ferase with Gcd14p, required for the modification of the adenine at position 58 in tRNAs, espe- cially tRNAi-Met; first identified as a negative regulator of GCN4 expression
2.66 2.66
2.66
CAGLOJ05412g YGL099W LSG1 Putative GTPase involved in 60S ribosomal subunit biogenesis; re- quired for the release of Nmd3p from 60S subunits in the cytoplasm
2.65
CAGLOJ10802g - - Description not mapped 2.64 CAGLOL13024g YHR121W LSM12 Protein of unknown function that
may function in RNA processing; interacts with Pbplp and Pbp4p and associates with ribosomes; con-tains an RNA-binding LSM domain and an AD domain; GFP-fusion protein is induced by the DNA-damaging agent MMS
2.63
CAGLOK08030g YPRO91C - Description not mapped 2.62
CAGLOD03344g YLR024C UDR2 Cytoplasmic ubiquitin-protein lig- ase (E3); required for ubiquityla- tion of Rpn4p; mediates formation of a Mublp-Ubr2p-Rad6p complex
2.61
CAGL0106490g - - Description not mapped 2.61
CAGLOE01925g YOL124C TRMI1 Catalytic subunit of an adoMet- dependent tRNA methyltransferase complex (Trmllp-Trm112p), re- quired for the methylation of the guanosine nucleotide at position
2.6
10 (m2G10) in tRNAs; contains a THUMP domain and a methyl-transferase domain
CAGLOE05126g YGL155W CDC43 Beta subunit of geranylgeranyl- transferase type I, catalyzes ger- anylgeranylation to the cysteine residue in proteins containing a C-terminal CaaX sequence ending in
2.6
Leu or Phe; has substrates impor-tant for morphogenesis
CAGL0107315g YOR131C - Description not mapped 2.6 CAGLOJ10604g YDR437W GPI19 Subunit of GPI-G1cNAc trans-
ferase involved in synthesis of N-acetylglucosaminyl phos- phatidylinositol (GIcNAc-PI), which is the first intermediate in glycosylphosphatidylinositol (GPI) anchor synthesis, shares similarity with mammalian PIG-P
2.6
CAGLOL02799g - - Description not mapped 2.6 CAGLOM05797g YGL063W PUS2 Mitochondrial tRNA:pseudouridine
synthase; acts at positions 27 and 2.6
28, but not at position 72; effi- ciently and rapidly targeted to mi- tochondria, specifically dedicated to mitochondria) tRNA modifica-tion
continued on next page

Appendix G. Transcription profiling of C. glabrata serum stress responses 375
continued from previous page CG ORF SC ORF gene name Description Log Ratio
CAGLOM10197g
CAGLOM13519g
YKL009W
YMR290C
MRT4
HAS1
Protein involved in mRNA turnover and ribosome assembly, localizes to the nucleolus ATP-dependent RNA helicase; lo- calizes to both the nuclear periph-ery and nucleolus; highly enriched in nuclear pore complex fractions; constituent of 66S pre-ribosomal particles
2.6
2.6
CAGLOD05610g YLL010C PSR1 Plasma membrane associated protein phosphatase involved in the general stress response; re- quired along with binding partner
2.59
Whi2p for full activation of STRE-mediated gene expression, possibly through dephosphorylation of Msn2p
CAGLOE03245g YGR159C NSRI Nucleolar protein that binds nu- clear localization sequences, re- quired for pre-rRNA processing and ribosome biogenesis
2.59
CAGL0001639g YBR247C ENP1 Protein associated with U3 and U14 snoRNAs, required for pre-rRNA processing and 40S ribosomal sub-unit synthesis; localized in the nu-cleus and concentrated in the nucle-olus
2.58
CAGL0107799g YBR154C RPB5 RNA polymerase subunit ABC27, common to RNA polymerases I, II, and III; contacts DNA and affects transactivation
2.58
CAGLOJ02948g YER060W FC1,11./ Putative purine-cytosine permease, very similar to Fcy2p but cannot substitute for its function
2.57
CAGLOD03410g YJL025W RRN7 Protein involved in the transcrip- tion of 35S rRNA genes by RNA polymerase I; component of the core factor (CF) complex also com-posed of Rrnllp, Rrn6p and TATA-binding protein
2.54
CAGL0,110890g YHRO66W SSF1 Constituent of 66S pre-ribosomal particles, required for ribosomal large subunit maturation; function-ally redundant with Ssf2p; member of the Brix family
2.54
CAGLOK02211g YER127W LCP5 Essential protein involved in matu- ration of 18S rRNA; depletion leads to inhibited pre-rRNA processing and reduced polysome levels; local-izes primarily to the nucleolus
2.54
- - Description not mapped 2.53 CAGL00O2497g YER029C SMR/ Core Sm protein Sm B; part
of heteroheptameric complex (with 2.53
Smdlp, Smd2p, Smd3p, Smelp, Smx3p, and Smx2p) that is part of the spliceosomal Ul, 112, U4, and 1.15 snRNPs; homolog of human Sm B and Sm B'
CAGLOJ09614g YDL208W NHP2 Nuclear protein related to mam- malian high mobility group (HMG) proteins, essential for function of
2.53
H/ACA-type snoRNPs, which are involved in 18S rRNA processing
continued on next page

G.3 Transcriptional response of C. glabrata cells exposed to 10% Foetal Calf Serum (FCS) 376
continued from previous page CG ORF SC ORF gene name Description Log Ratio
CAGL0G07535g YOR294W RRS1 Essential protein that binds ribo- somal protein L11 and is required for nuclear export of the 60S pre-ribosomal subunit during ribosome biogenesis; mouse homolog shows altered expression in Huntington's disease model mice
2.52
CAGL0003355g YNR054C ESF2 Essential nucleolar protein involved in pre-18S rRNA processing; binds to RNA and stimulates ATPase ac-tivity of Dbp8; involved in assembly of the small subunit (SSU) proces-some
2.51
CAGL0004961g YJR106W ECM27 Non-essential protein of unknown function
2.51
CAGLOD02018g - - Description not mapped 2.51 CAGLOHOO11Og YIR019C MUC/ GPI-anchored cell surface glyco-
protein (flocculin) required for pseudohyphal formation, invasive growth, flocculation, and biofllms; transcriptionally regulated by the
2.51
MAPK pathway (via Stel2p and Teclp) and the cAMP pathway (via Flo8p)
CAGLOH10252g YBRO61C TRM7 2'-O-ribose methyltransferase, methylates the 2'-O-ribose of nu-cleotides at positions 32 and 34 of the tRNA anticodon loop
2.51
CAGLOJ03388g YDL175C AIR2 RING finger protein that interacts with the arginine methyltransferase
2.51
Hmtlp; may regulate methylation of Npl3p, which modulates Npl3p function in mRNA processing and export; has similarity to Air1p
CAGLOJ11110g YJR003C - Description not mapped 2.51 CAGLOK09042g YOR252W TMA15 Protein of unknown function that
associates with ribosomes 2.51
CAGLOE03069g - - Description not mapped 2.5 CAGLOI10747g YPR138C MEPS Ammonium permease of high ca-
pacity and low affinity; belongs to a ubiquitous family of cytoplasmic membrane proteins that transport only ammonium (NII4-1-); expres- sion is under the nitrogen catabo-lite repression regulation ammonia permease
2.5
CAGLOJ11242g YNL180C RHOS Non-essential small GTPase of the 2.5 Rho/Rac subfamily of Ras-like pro-teins, likely involved in protein ki- nase C (Pkclp)-dependent signal transduction pathway that controls cell integrity
CAGLOM05687g YGL067W NPY1 NADH diphosphatase (pyrophos- phatase), hydrolyzes the pyrophos-phate linkage in NADH and re- lated nucleotides; localizes to per- oxisomes
2.5
CAGLOF07359g YGL117W - Description not mapped 2.49 CAGLOF07557g YKL095W YJU2 Essential protein required for pre-
mRNA splicing; associates tran- siently with the spliceosomal NTC
2.49
(nineteen complex) and acts after Prp2p to promote the first catalytic reaction of splicing
continued on next page

Appendix G. Transcription profiling of C. glabrata serum stress responses 377
continued from previous page CG ORF SC ORE gene name Description Log Ratio
CAGLOG10219g YHR211W FLO5 Lectin-like cell wall protein (floc- culin) involved in flocculation, binds to mannose chains on the surface of other cells, confers floc-forming ability that is chy- motrypsin resistant but heat labile; similar to Flo1p
2.49
CAGLOJ00297g YHR045W - Description not mapped 2.49 CAGL0D05588g YLL011W SOFT Essential protein required for bio-
genesis of 40S (small) ribosomal subunit; has similarity to the beta subunit of trimeric G-proteins and the splicing factor Prp4p
2.48
CAGLOI02508g YIIR175W CTR2 Putative low-affinity copper trans- porter of the vacuolar membrane; mutation confers resistance to toxic copper concentrations, while over-expression confers resistance to cop-per starvation
2.48
CAGLOM01716g YBRO83W TECI Transcription factor required for fpuellscTriypltioenpx.rissmioanp,pTedyl-mediated gene activation, and haploid in- vasive and diploid pseudohyphal growth; TEA/ATTS DNA-binding domain family member
2.48
CAGLOA03905g - - 2.47 CAGLOG01254g YGL202W ARO8 Aromatic aminotransferase I, ex-
pression is regulated by general con-trol of amino acid biosynthesis
2.47
CAGLONI11462g YLR099C ICT1 Lysophosphatidic acid acyltrans- ferase, responsible for enhanced phospholipid synthesis during or- ganic solvent stress; null displays increased sensitivity to Calcofluor white; highly expressed during or-ganic solvent stress
2.47
CAGLOA00385g YGL01.6W KAP1.22 Karyopherin beta, responsible for import of the Toalp-Toa2p complex into the nucleus; binds to nucleo-porins Nuplp and Nup2p; may play a role in regulation of pleiotropic drug resistance
2.45
CAGL0I00724g YOR306C MCH5 Plasma membrane riboflavin trans- porter; facilitates the uptake of vitamin B2; required for FAD- dependent processes; sequence simi-larity to mammalian monocarboxy- late permeases, however mutants are not deficient in monocarboxy-late transport
2.45
CAGLOM03949g YNL310C Z/M/ 7 Heat shock protein with a zinc fin- ger motif; essential for protein im- port into mitochondria; may act with Paml8p to facilitate recogni-tion and folding of imported pro-teins by Ssc1p (mtHSP70) in the mitochondrial matrix
2.45
CAGLOM04477g YLR141W RRN5 Protein involved in transcription of rDNA by RNA polymerase I; tran-scription factor, member of UAF
2.45
(upstream activation factor) family along with Rrn9p and RrnlOp
continued on next page

G.3 Transcriptional response of C. glabrata cells exposed to 10% Foetal Calf Serum (FCS) 378
continued from previous page CG ORF SC ORF gene name Description Log Ratio
CAGLOA00627g CAGLOA03212g
YAR002CA YDR384C
- ATO3
Description not mapped Plasma membrane protein, regu- lation pattern suggests a possible role in export of ammonia from the cell; phosphorylated in mito- chondria; member of the TC 9.B.33
2.44 2.44
YaaH family of putative trans- porters
CAGLOM08096g YOR208W PTP2 Phosphotyrosine-specific protein phosphatase involved in the in- activation of mitogen-activated protein kinase (MAPK) during osmolarity sensing; dephospory- lates Hog1p MAPK and regulates its localization; localized to the nucleus
2.44
CAGLOA04257g YER088C DOT6 Protein involved in rRNA and ribo- some biogenesis; binds polymerase
2.43
A and C motif; subunit of the RPD3L histone deacetylase com- plex; similar to Tod6p; has chro- matin specific SANT domain; in-volved in telomeric gene silencing and filamentation
CAGLOF06853g YKL021C MA KI 1 Protein involved in an early, nude- o] ribosomal subunit biogenesis; essential for cell growth and replication of killer M1 dsRNA
2.43
CAGLOD00396g - -
60S.tmd ofonn
virus; contains four beta-transducin repeats
r) easr crs itpetpi appe
2.42 CAGLOD00880g YDLO6OW TSR1 Protein required for processing of 2.42
20S pre-rRNA in the cytoplasm, associates with pre-40S ribosomal particles
CAGLOF06809g YIR026C YVH1 Protein phosphatase involved in vegetative growth at low tempera-tures, sporulation, and glycogen ac- cumulation; transcription induced by low temperature and nitrogen starvation; member of the dual- specificity family of protein phos-phatases
2.42
CAGLOJ11154g YNL175C HOPIS Protein of unknown function, local- izes to the nucleolus and nucleo-plasm; contains an RNA recogni-tion motif (RRM) and has similar-ity to Nopl2p, which is required for processing of pre-18S rRNA
2.42
CAGLOA02431g YDR349C YPS7 Putative GPI-anchored aspartic protease, located in the cytoplasm and endoplasmic reticulum
2.41
CAGLOB02970g - - Description not mapped 2.41 CAGL0102398g YHR170W NMD3 Protein involved in nuclear export
of the large ribosomal subunit; acts as a Crmlp-dependent adapter pro-tein for export of nascent ribosomal subunits through the nuclear pore complex
2.41
continued on next page

Appendix G. Transcription profiling of C. glabrata serum stress responses 379
continued from previous page CG ORF SC ORF gene name Description Log Ratio
CAGLOJ00517g
CAGLOJ09702g CAGLOM00308g CAGLOM00572g CAGLOG07018g CAGLOL04928g
YHR031C
-- --
YJR129C YML018C YMR047C
RRM3
- NUP116
DNA helicase involved in rDNA replication and Tyl transposition; relieves replication fork pauses at telomeric regions; structurally and functionally related to Pif1p Description not mapped Description not mapped Description not mapped Description not mapped Subunit of the nuclear pore corn- plex (NPC) that is localized to both sides of the pore; contains a repeti-tive GLFG motif that interacts with mRNA export factor Mex67p and with karyopherin Kap95p; homolo-gous to Nup100p
2.41
2.41 2.41 2.41 2.4 2.4
CAGLOM01430g YDR324C UTP4 Subunit of U3-containing 90S pre- ribosome and Small Subunit (SSU) processome complexes involved in
2.4
Production of 18S rRNA and as- sembly of small ribosomal subunit; member of t-Utp subcomplex in- volved with transcription of 35S rRNA transcript
CAGL00O2255g YER041W YEN1 Holliday junction resolvase; ho- molog of human GEN1 and has sim-ilarity to S. cerevisiae endonuclease
2.39
Rthlp; potentially phosphorylated by Cdc28p
CAGLOG02761g YIL103W DPH1 Protein required, along with 2.39 Dph2p, Ktillp, Jjj3p, and Dph5p, for synthesis of diphthamide, which is a modified histidine residue of translation elongation factor 2 (Eft1p or Eft2p); may act in a complex with Dph2p and Ktillp
CAGLOI05544g YER095W RAD51 Strand exchange protein, forms a helical filament with DNA that searches for homology; involved in the recombinational repair of double-strand breaks in DNA dur-ing vegetative growth and meiosis; homolog of Dmclp and bacterial
2.39
RecA protein CAGLOJ10208g YNL072W RNH201 Ribonuclease H2 catalytic sub-
unit, removes RNA primers dur- ing Okazaki fragment synthesis; homolog of RNAse HI (the S. cerevisiae homolog of mammalian
2.39
RNAse HII is RNH1); related to human AGS4 that causes Aicardi- Goutieres syndrome
CAGLOM11110g YDR087C RRP1 Essential evolutionarily conserved nucleolar protein necessary for bio-genesis of 60S ribosomal subunits and processing of pre-rRNAs to ma-ture rRNAs, associated with several distinct 66S pre-ribosomal particles
2.39
CAGLOD05874g YJR002W MPPIO Component of the SSU processome and 90S preribosome, required for pre-18S rRNA processing, interacts with and controls the stability of
2.38
Imp3p and Imp4p, essential for vi-ability; similar to human Mpp lOp
continued on next page

G.3 Transcriptional response of C. glabrata cells exposed to 10% Foetal Calf Serum (FCS) 380
continued from previous page CG ORF SC ORF gene name Description Log Ratio
CAGLOG02585g YIL094C LYSI2 Homo-isocitrate dehydrogenase, an NAD-linked mitochondria] enzyme required for the fourth step in the biosynthesis of lysine, in which homo-isocitrate is oxidatively de- carboxylated to alpha-ketoadipate
2.38
CAGLOL02035g YKL029C MAE! Mitochondrial malic enzyme, cat- alyzes the oxidative decarboxyla- tion of malate to pyruvate, which is a key intermediate in sugar metabolism and a precursor for syn-thesis of several amino acids
2.38
CAGLOH02189g YIVIR269W TMA23 Nucleolar protein of unknown func- tion implicated in ribosome bio- genesis; TMA23 may be a fungal-specific gene as no homologs have been yet identified in higher eukary-otes
2.37
CAGLOH07051g YDR321W ASP1 Cytosolic L-asparaginase, involved in asparagine catabolism
2.37
CAGLOH07469g YFRO19W FABI 1-phosphatidylinositol-3-phosphate 2.36 5-kinase; vacuolar membrane kinase that generates phosphatidylinos- itol (3,5)P2, which is involved in vacuolar sorting and homeostasis
CAGLOK06171g - - Description not mapped 2.36 CAGLOK11506g YKL113C RAD27 5' to 3' exonuclease, 5' flap endonu-
ment processing and maturation as well as for long-patch base-excision
cpleeascsrei,ptrieociunirstd rppOedkazaki frag-
repair; member of the S. pombe
2.36
RAD2/FEN1 family CAGLOL07590g - - mapped 2.36 CAGLOM09801g YLR285W NNT1 Putative nicotinamide N-
methyltransferase, has a role in rDNA silencing and in lifespan determination
2.36
CAGLOA03476g YLR034C SMF3 Putative divalent metal ion trans- porter involved in iron homeosta- sis; transcriptionally regulated by metal ions; member of the Nramp family of meta/ transport proteins
2.35
CAGL00O2981g YCR048W ARE1 Acyl-CoA:sterol acyltransferase, isozyme of Are2p; endoplasmic reticulum enzyme that contributes the major sterol esterification activity in the absence of oxygen
2.35
CAGLOG03003g YGRO9OW UTP22 Possible U3 snoRNP protein in- volved in maturation of pre-18S rRNA, based on computational analysis of large-scale protein- protein interaction data
2.35
CAGLOH06017g YBROO8C FLR1 Plasma membrane multidrug trans- porter of the major facilitator su- perfamily, involved in efflux of fluconazole, diazaborine, benomyl, methotrexate, and other drugs
2.35
CAGLOK06919g YBR227C MCX1 Mitochondrial matrix protein; pu- tative ATP-binding chaperone with non-proteolytic function; similar to bacterial CIpX proteins
2.35
CAGL0003003g YCR051W - Description not mapped 2.34 CAGLOK01287g YGRO74W SMD1 Core Sm protein Sm Dl; part
of heteroheptameric complex (with 2.34
Smblp, Smd2p, Smd3p, Smelp, Smx3p, and Smx2p) that is part of the spliceosomal Ul, U2, U4, and U5 snRNPs; homolog of human Sm D1
continued on next page

Appendix G. Transcription profiling of C. glabrata serum stress responses 381
continued from previous page CG ORF SC ORF gene name Description Log Ratio
CAGLOF03091g CAGLOG03443g
-- YER110C KAP123
Description not mapped Karyopherin beta, mediates nuclear import of ribosomal proteins prior to assembly into ribosomes and im-port of histones H3 and H4; local- izes to the nuclear pore, nucleus, and cytoplasm; exhibits genetic in-teractions with RAIL
2.33 2.33
CAGLOG07249g YML027W YOX1 Homeodomain-containing tran- scriptional repressor, binds to
2.33
Mcm1p and to early cell cycle boxes (ECBs) in the promoters of cell cycle-regulated genes expressed in M/G1 phase; expression is cell cycle-regulated; potential Cdc28p substrate
CAGLOH05731g YPL094C SEC62 Essential subunit of Sec63 corn- plex (Sec63p, Sec62p, Sec66p and Sec72p); with Sec61 com- plex, Kar2p/BiP and Lhslp forms a channel competent for SRP- dependent and post-translational
2.33
SRP-independent protein targeting and import into the ER
CAGLOL10714g YMR202W ERG2 C-8 sterol isomerase, catalyzes the isomerization of the delta-8 dou- ble bond to the delta-7 position at an intermediate step in ergosterol biosynthesis
2.33
CAGLOM08756g YDR261C EXG2 Exo-1,3-beta-glucanase, involved in cell wall beta-glucan assembly; may be anchored to the plasma mem-brane via a glycosylphosphatidyli-nositol (GPI) anchor
2.33
CAGL01103465g - - Description not mapped 2.32 CAGLOH10626g YARO5OW FLO1 Lectin-like protein involved in floc-
culation, cell wall protein that binds to mannose chains on the surface of other cells, confers floc-forming ability that is chy- motrypsin sensitive and heat resis-tant; similar to Flo5p
2.32
CAGL0105676g YNFt011C PRP2 RNA-dependent ATPase in the 2.32 DEAR-box family, required for acti-vation of the spliceosome before the first transesterification step in RNA splicing
CAGLOK09922g YOR359W VTSI Post-transcriptional gene regulator, RNA-binding protein containing a
2.32
SAM domain; shows genetic inter-actions with Vtilp, which is a v- SNARE involved in cis-Golgi mem-brane traffic
CAGLOL06314g YDR101C ARX1 Shuttling pre-60S factor; involved in the biogenesis of ribosomal large subunit biogenesis; interacts di- rectly with Albl; responsible for
2.32
Tif6 recycling defects in absence of Reil; associated with the ribosomal export complex
continued on next page

G.3 Transcriptional response of C. glabrata cells exposed to 10% Foetal Calf Serum (FCS) 382
continued from previous page CG ORF SC ORF gene name Description Log Ratio
CAGLOL13332g
CAGLOA01199g
YAL063C
YPL265W
FLOP
DIPS
Lectin-like protein with similarity to Flo1p, thought to be expressed and involved in flocculation Dicarboxylic amino acid permease, mediates high-affinity and high- capacity transport of L-glutamate and L-aspartate; also a transporter for Gln, Asn, Ser, Ala, and Gly
2.32
2.31
CAGLOE02673g YOR004W UTP23 Essential nucleolar protein that is a component of the SSU (small sub-unit) processome involved in 40S ribosomal subunit biogenesis; has homology to PING domain protein
2.31
Fcflp, although the PING domain of Utp23p is not required for function
CAGLOF06963g YGLO61C DUOI Essential subunit of the Daml corn- plex (aka DASH complex), couples kinetochores to the force produced by MT depolymerization thereby aiding in chromosome segregation; is transferred to the kinetochore prior to mitosis
2.31
CAGLOF08129g YGR245C SDAI Highly conserved nuclear protein required for actin cytoskeleton or- ganization and passage through
2.31
Start, plays a critical role in G1 events, binds Nap1p, also involved in 60S ribosome biogenesis
CAGL0G03597g YER118C SHOT Transmembrane osmosensor, par- ticipates in activation of both the Cdc42p- and MAP kinase- dependent filamentous growth pathway and the high-osmolarity glycerol response pathway
2.31
CAGLOJ02376g YIL019W FAFI Protein required for pre-rRNA pro- cessing and 40S ribosomal subunit assembly
2.31
CAGLOL05566g YJL125C GCDI4 Subunit of tRNA (1- methyladenosine) methyltrans- ferase, with GcdlOp, required for the modification of the adenine at position 58 in tRNAs, espe- cially tRNAi-Met; first identified as a negative regulator of GCN4 expression
2.31
CAGLOM03751g YNL299W TRFS Poly (A) polymerase involved in nuclear RNA quality control based on: homology with Trf4p, genetic interactions with TRF4 mutants, physical interaction with Mtr4p
2.31
(TRAMP subunit), and by direct assay; disputed role as a sigma DNA polymerase
CAGLOM06457g YBR187W GDTI Putative protein of unknown func- tion; expression is reduced in a gcrl null mutant; GFP-fusion protein lo- calizes to the vacuole; expression pattern and physical interactions suggest a possible role in ribosome biogenesis
2.31
continued on next page

Appendix G. Transcription profiling of C. glabrata serum stress responses 383
continued from previous page CG ORF SC ORF gene name Description Log Ratio
CAGLOG05544g CAGLOH01595g
- YDR527W
- RBA50
Description not mapped Protein involved in transcription; interacts with RNA polymerase II subunits Rpb2p, Rpb3, and
2.3 2.3
Rpbllp; has similarity to human RPAP 1
CAGLOI07777g YOL088C IlIPD2 Member of the protein disulfide iso- merase (PDI) family, exhibits chap-erone activity; overexpression sup-presses the lethality of a pdil dele-tion but does not complement all
2.3
Pdilp functions; undergoes oxida-tion by Erolp
CAGLOL08998g YFLOO1W DEG1 Non-essential tRNA:pseudouridine synthase, introduces pseu- douridines at position 38 or 39 in tRNA, important for mainte- nance of translation efficiency and normal cell growth, localizes to both the nucleus and cytoplasm
2.3
CAGLOB00286g YCL064C CHA I Catabolic L-serine (L-threonine) deaminase, catalyzes the degra- dation of both L-serine and L- threonine; required to use serine or threonine as the sole nitrogen source, transcriptionally induced by serine and threonine
2.29
CAGLOB03663g - - Description not mapped 2.29 CAGLOD04796g YPR131C NAT3 Catalytic subunit of the NatB N-
terminal acetyltransferase, which catalyzes acetylation of the amino-terminal methionine residues of all proteins beginning with Met-Asp or
2.29
Met-Glu and of some proteins be-ginning with Met-Asn or Met-Met
CAGLOH01441g YDR300C PRO1 Gamma-glutamyl kinase, catalyzes the first step in proline biosynthesis
2.29
CAGLOJ04576g YKL191W DPH2 Protein required, along with 2.29 Dphlp, Ktillp, Jjj3p, and Dph5p, for synthesis of diphthamide, which is a modified histidine residue of translation elongation factor 2 (Eftlp or Eft2p); may act in a complex with Dphlp and Ktillp
CAGLOL06710g YOL092W - Description not mapped 2.29 CAGLOD01892g YLR027C AAT2 Cytosolic aspartate aminotrans-
ferase, involved in nitrogen metabolism; localizes to perox- isomes in oleate-grown cells
2.28
CAGLOH06545g YIL146C ECM37 Non-essential protein of unknown function
2.28
CAGLOI02926g YDR496C PUF6 Pumilio-homology domain protein that binds ASH1 mRNA at PUF consensus sequences in the 3' UTR and represses its translation, result-ing in proper asymmetric localiza-tion of ASH1 mRNA
2.28
CAGLOI02860g YOR112W CEX1 Cytoplasmic component of the nuclear aminoacylation-dependent tRNA export pathway; interacts with nuclear pore component
2.27
Nup116p; copurifies with tRNA export receptors Loslp and Msn5p, as well as eIF-la and the RAN GTPase Gsplp
continued on next page

G.3 Transcriptional response of C. glabrata cells exposed to 10% Foetal Calf Serum (FCS) 384
continued from previous page CG ORF SC ORF gene name Description Log Ratio
CAGL0103806g YDL141W BPL1 Biotin:apoprotein ligase, covalently modifies proteins with the addi- tion of biotin, required for acetyl-
2.27
CoA carboxylase (Acclp) holoen-zyme formation
CAGLOJ03014g YIL056W VHR1 Transcriptional activator, required for the vitamin H-responsive ele-ment (VHRE) mediated induction of VHT1 (Vitamin H transporter) and BI05 (biotin biosynthesis inter-mediate transporter) in response to low biotin concentrations
2.27
CAGLOJ05390g - - Description not mapped 2.27 CAGLOL02871g YOR206W NOC2 Protein that forms a nucleolar corn-
plex with Mak21p that binds to 2.27
90S and 66S pre-ribosomes, as well as a nuclear complex with Noc3p that binds to 66S pre-ribosomes; both complexes mediate intranu- clear transport of ribosomal precur-sors
CAGLOL11308g YML067C ERV4/ Protein localized to COPII-coated vesicles, forms a complex with
2.27
Erv46p; involved in the membrane fusion stage of transport; has ho-mology to human ERGIC2 (PTX1) protein
CAGLOM02805g YPL126W NANI U3 snoRNP protein, component of the small (ribosomal) subunit
2.27
(SSU) processosome containing U3 snoRNA; required for the biogene-sis ofl8S rRNA
CAGLOM13915g YMR310C - Description not mapped 2.27 CAGLOA02673g YDR361C BCP1 Essential protein involved in nu-
clear export of Mss4p, which is a lipid kinase that generates phos- phatidylinositol 4,5-biphosphate and plays a role in actin cytoskele- ton organization and vesicular transport
2.26
CAGL0103542g YDL156W CMRI Putative protein of unknown func- tion; protein sequence contains three WD domains (WD-40 repeat); green fluorescent protein (GFP)- fusion protein localizes to the cyto-plasm and nucleus
2.26
CAGLOJ05984g YNL141W AAH1 Adenine deaminase (adenine amino- hydrolase), converts adenine to hy-poxanthine; involved in purine sal-vage; transcriptionally regulated by nutrient levels and growth phase;
2.26
Aahlp degraded upon entry into quiescence via SCF and the protea-some
CAGLOL13156g YLR073C - Description not mapped 2.26 CAGLOA01177g YPL268W PLC/ Phospholipase C, hydrolyzes phos-
phatidylinositol 4,5-biphosphate 2.2
(PIP2) to generate the signal- ing molecules inositol 1,4,5- triphosphate (IP3) and 1,2- diacylglycerol (DAG); involved in regulating many cellular processes
continued on next page

Appendix G. Transcription profiling of C. glabrata serum stress responses 385
continued from previous page CG O RF SC ORF gene name Description Log Ratio
CAGLOI00198g CAGLOL05610g
-- YJL127C SPTIO
Description not mapped Putative histone acetylase, sequence-specific activator of histone genes, binds specifically and highly cooperatively to pairs of UAS elements in core histone promoters, functions at or near the
2.2 2.2
TATA box CAGLOL10846g YCR047C B UD23 Protein involved in bud-site selec-
tion; diploid mutants display a ran-dom budding pattern instead of the wild-type bipolar pattern
2.2
CAGLOM06589g YDR020C DAS2 Putative protein of unknown func- tion; not an essential gene; weak similarity with uridine kinases and with phosphoribokinases
2.2
- - Description not mapped 2.24 CAGL0E04334g YHROO7C ERG11 Lanosterol 14-alpha-demethylase,
catalyzes the C-14 demethylation of lanosterol to form 4,4"-dimethyl cholesta-8,14,24-triene-3-beta-ol in the ergosterol biosynthesis path- way; member of the cytochrome
2.24
P450 family CAGLOF03261g - - Description not mapped 2.24 CAGLOH09724g YER006W NUG1 GTPase that associates with nu-
clear 60S pre-ribosomes, required for export of SOS ribosomal sub-units from the nucleus
2.24
CAGL0105588g YNR015W SMMI Dihydrouridine synthase, member of a family of dihydrouridine syn- thases including Duslp, Smm1p, Dus3p, and Dus4p; modifies uridine residues at position 20 of cytoplas-mic tRNAs
2.24
CAGLOM03905g YNL308C KRII Essential nucleolar protein required for 40S ribosome biogenesis; phys- ically and functionally interacts with Krrlp
2.24
CAGLOH04235g YML080W DUSI Dihydrouridine synthase, member of a widespread family of con- served proteins including Smmlp, Dus3p, and Dus4p; modifies pre- tRNA(Phe) at U17
2.23
CAGL0109834g - Description not mapped 2.23 CAGLOJ07986g YNL260C Description not mapped 2.23 CAGLOL06116g YGL185C - Description not mapped 2.23 CAGLOF03443g YMR274C RCE1 Type II CAAX prenyl protease in-
volved in the proteolysis and matu-ration of Ras and the a-factor mat-ing pheromone
2.22
CAGLOF08525g YGROO1C AML 1 Putative protein of unknown func- tion with similarity to methyltrans-ferase family members; green flu-orescent protein (GFP)-fusion pro-tein localizes to the cytoplasm; re-quired for replication of Brome mo-saic virus in S. cerevisiae
2.22
CAGLOI03080g YEL021W URA3 Orotidine-5'-phosphate (OMP) de- carboxylase, catalyzes the sixth en- zymatic step in the de novo biosyn- thesis of pyrimidines, converting
2.22
OMP into uridine monophosphate (UMP); converts 5-FOA into 5- fluorouracil, a toxic compound
continued on next page

G.3 Transcriptional response of C. glabrata cells exposed to 10% Foetal Calf Serum (FCS) 386
continued from previous page CG ORF SC ORF gene name Description Log Ratio
CAGLOJ00957g YLROO9W RLP24 Essential protein with similarity to 2.22 Rp124Ap and Rp124Bp, associated with pre-60S ribosomal subunits and required for ribosomal large subunit biogenesis
CAGLOJ10846g YHR071W PCL.5 Cyclin, interacts with and phos- phorylated by Pho85p cyclin- dependent kinase (Cdk), induced by Gcn4p at level of transcrip- tion, specifically required for
2.22
Gcn4p degradation, may be sensor of cellular protein biosynthetic capacity
CAGLOF04433g YBL039C URA 7 Major CTP synthase isozyme (see also URA8), catalyzes the ATP- dependent transfer of the amide nitrogen from glutamine to UTP, forming CTP, the final step in de novo biosynthesis of pyrimidines; involved in phospholipid biosynthe-sis
2.21
CAGLOJ05698g YNL151C RPC31 RNA polymerase III subunit C31; contains HMG-like C-terminal do-main
2.21
CAGLOK128O4g YLR074C BUD20 Protein involved in bud-site selec- tion; diploid mutants display a ran-dom budding pattern instead of the wild-type bipolar pattern
2.21
CAGLOM02409g YPL146C NOP53 Nucleolar protein; involved in bio- genesis of the 60S subunit of the ri-bosome; interacts with rRNA pro-cessing factors Cbf5p and Nop2p; null mutant is viable but growth is severely impaired
2.21
CAGLOD04202g YIL090W ICE2 Integral ER membrane protein with type-III transmembrane domains; mutations cause defects in cortical
2.2
ER morphology in both the mother and daughter cells
CAGLOD05104g -- Description not mapped 2.2 CAGL0110879g YPL112C PEX25 Peripheral peroxisomal membrane
peroxin required for the regula- tion of peroxisome size and main- tenance, recruits GTPase Rho1p to peroxisomes, induced by oleate, interacts with homologous protein
2.2
Pex27p CAGLOJ01463g YKL096W CWPI Cell wall mannoprotein, linked to a
beta-1,3- and beta-1,6-glucan het-eropolymer through a phosphodi-ester bond; involved in cell wall or-ganization
2.2
CAG LOB 00462g YC L055W KAR4 Transcription factor required for gene regulation in repsonse to pheromones; also required during meiosis; exists in two forms, a slower-migrating form more abun-dant during vegetative growth and a faster-migrating form induced by pheromone
2.19
continued on next page

Appendix G. Transcription profiling of C. glabrata serum stress responses 387
continued from previous page CG ORF SC ORF gene name Description Log Ratio
CAGL0000737g YLR222C UTP13 Nucleolar protein, component of the small subunit (SSU) processome containing the U3 snoRNA that is involved in processing of pre-18S rRNA
2.19
CAGLOH08415g YDR045C RPC// RNA polymerase III subunit C11; mediates pol III RNA cleavage ac-tivity and is important for termina-tion of transcription; homologous to
2.19
TFIIS CAGLOL03333g YKR044W UIP5 Protein of unknown function
that interacts with IJIplp, a Ubl 2.19
(ubiquitin-like protein)-specific protease for Smt3p protein conju-gates
CAGLOD02838g YLL034C RIX7 Putative ATPase of the AAA fam- ily, required for export of pre- ribosomal large subunits from the nucleus; distributed between the nucleolus, nucleoplasm, and nuclear periphery depending on growth con-ditions
2.18
CAGLOM07425g YMR0281V TAP42 Essential protein involved in the 2.18 TOR signaling pathway; physically associates with the protein phos-phatase 2A and the SIT4 protein phosphatase catalytic subunits
CAGLOM09493g YMR160W - Description not mapped 2.18 CAGLOA00913g YLR320W MMS22 Protein that acts with Mmslp in
a repair pathway that may be in-volved in resolving replication inter-mediates or preventing the damage caused by blocked replication forks; required for accurate meiotic chro-mosome segregation
2.17
CAGLOH01023g YDR279W RNH202 Ribonuclease H2 subunit, required for RNase H2 activity; related to human AGS2 that causes Aicardi-
2.17
Goutieres syndrome CAGLOJ05896g YNL158W PGAI Essential component of GPI-
mannosyltransferase II, responsible for second mannose addition to
2.17
GPI precursors as a partner of Gpil8p; required for maturation of Gas1p and Pho8p; has synthetic genetic interations with secretory pathway genes
CAGLOK11231g YDR245W MNNIO Subunit of a Golgi mannosyl- transferase complex also contain- ing Anplp, Mnn9p, Mnnllp, and
2.17
Hoc1p that mediates elongation of the polysaccharide mannan back- bone; membrane protein of the mannosyltransferase family
CAGLOG00264g YGR280C PXR1 Essential protein involved in rRNA and snoRNA maturation; competes with TLC1 RNA for binding to
2.16
Est2p, suggesting a role in negative regulation of telomerase; human ho-molog inhibits telomerase; contains a G-patch RNA interacting domain
continued on next page

G.3 Transcriptional response of C. glabrata cells exposed to 10% Foetal Calf Serum (FCS) 388
continued from previous page CG ORF SC ORF gene name Description Log Ratio
CAGLOJ03146g
CAGLOK03927g
CAGLOL00759g
CAGLOM09691g
CAGL0001111g CAGLOK01463g
CAGLOM09911g
CAGLOB04169g
CAGLOD06050g
CAGLOF02563g
CAGLOF02871g
YER070W
YMR134W
YER055C
YMR153W
- YDL042C
YLR405W
YPR112C
YJR054W
YDR399W
YDR414C
RNR1
ERG29
11151
NUP53
- SIR2
DUS4
MRD/
ERAIG
HPT1
ERD1
One of two large regulatory sub- units of ribonucleotide-diphosphate reductase; the RNR complex cat-alyzes rate-limiting step in dNTP synthesis, regulated by DNA repli-cation and DNA damage checkpoint pathways via localization of small subunits Protein of unknown function that may be involved in iron metabolism; mutant bm-8 has a growth defect on iron-limited medium that is comple-mented by overexpression of Yfhlp; shows localization to the ER; highly conserved in ascomycetes ATP phosphoribosyltransferase, a hexameric enzyme, catalyzes the first step in histidine biosynthesis; mutations cause histidine auxotro-phy and sensitivity to Cu, Co, and Ni salts; transcription is regulated by general amino acid control Subunit of the nuclear pore corn-
pherin Kap121p or with Nup170p via overlapping regions of Nup53p,
ape with karyo-interacts), tmpd
involved in activation of the spindle
ppleesxcr(iNPi.C. no
checkpoint mediated by the Mad1p-Mad2p complex
pt
Conserved NAD+ dependent his- tone deacetylase of the Sirtuin fam-ily involved in regulation of lifes- pan; plays roles in silencing at HML, HMR, telomeres, and the rDNA locus; negatively regulates initiation of DNA replication Dihydrouridine synthase, member of a widespread family of conserved proteins including Smmlp, Duslp, and Dus3p Essential conserved protein that is part of the 90S preribosome; re- quired for production of 18S rRNA and small ribosomal subunit; con-tains five consensus RNA-binding domains Vacuolar protein of unknown func- Lion; potential Cdc28p substrate Dimeric hypoxanthine-guanine phosphoribosyltransferase, cat- alyzes the formation of both inosine monophosphate and guano-sine monophosphate; mutations in the human homolog HPRT1 can cause Lesch-Nyhan syndrome and Kelley-Seegmiller syndrome Predicted membrane protein re- quired for the retention of lume-nal endoplasmic reticulum proteins; mutants secrete the endogenous ER protein, BiP (Kar2p)
2.16
2.16
2.16
2.16
2.15 2.15
2.15
2.14
2.14
2.14
2.14
continued on next page

Appendix G. Transcription profiling of C. glabrata serum stress responses 389
continued from previous page CG ORF SC ORF gene name Description Log Ratio
CAGLOF07535g
CAGLOI02948g
YKL094W
YDR498C
YJU3
SEC20
Serine hydrolase with sequence similarity to monoglyceride lipase (MGL), localizes to lipid particles Membrane glycoprotein v-SNARE involved in retrograde transport from the Golgi to the ER; required for N- and 0-glycosylation in the
2.14
2.14
Golgi but not in the ER; forms a complex with the cytosolic Tip2Op
CAGLOK00957g YOR200C ELP2 Subunit of Elongator complex, which is required for modification of wobble nucleosides in tRNA; target of Kluyveromyces lactis zymocin
2.14
CAGLOB04961g YLR172C DPH5 Methyltransferase required for syn- thesis of diphthamide, which is a modified histidine residue of trans-lation elongation factor 2 (Eft1p or
2.13
Eft2p); not essential for viability; GFP-Dph5p fusion protein localizes to the cytoplasm
CAGLOD00220g YAL059W ECM1 Protein of unknown function, local- ized in the nucleoplasm and the nu- cleolus, genetically interacts with
2.13
MTR2 in 60S ribosomal protein subunit export
CAGLOE05016g YGL148W AR02 Bifunctional chorismate syn- thase and flavin reductase, catalyzes the conversion of 5- enolpyruvylshikimate 3-phosphate
2.13
(EPSP) to form chorismate, which is a precursor to aromatic amino acids
CAGLOG07843g YGR103W NOP7 Nucleolar protein involved in rRNA processing and 60S ribosomal subunit biogenesis; constituent of several different pre-ribosomal particles; required for exit from
2.13
Gisub2,0j/subi, and the initiation of cell proliferation
CAGLOI02596g -- Description not mapped 2.13 CAGLOK04917g - - Description not mapped 2.13 CAGLOL04114g YNL124W NAF1 RNA-binding protein required
for the assembly of box H/ACA snoRNPs and thus for pre-rRNA processing, forms a complex with
2.13
Shqlp and interacts with II/ACA snoRNP components Nhp2p and Cbf5p; similar to Gar1p
CAGL0001595g YER248C HIS7 Imidazole glycerol phosphate synthase (glutamine amidotrans- ferase:cyclase), catalyzes the fifth and sixth steps of histidine biosynthesis and also produces
2.12
5-aminoimidazole-4-carboxamide ribotide (AICAR), a purine precur- sor
CAGLOG05445g YDL226C GCS1 ADP-ribosylation factor GTPase activating protein (ARF GAP), involved in ER-Golgi transport; shares functional similarity with
2.12
Glo3p continued on next page

G.3 Transcriptional response of C. glabrata cells exposed to 10% Foetal Calf Serum (FCS) 390
continued from previous page CC ORF SC ORF gene name Description Log Ratio
CAGLOK06787g YBR218C PYC2 Pyruvate carboxylase isoform, cyto- plasmic enzyme that converts pyru-vate to oxaloacetate; highly similar to isoform Pyclp but differentially regulated; mutations in the human homolog are associated with lactic acidosis
2.12
CAGLOK10780g YML056C titfD4 Inosine monophosphate dehydroge- nase, catalyzes the first step of
2.12
GMP biosynthesis, member of a four-gene family in S. cerevisiae, constitutively expressed
CAGLOM06699g YIL084C SDS3 Component of the Rpd3p/Sin3p deacetylase complex required for its structural integrity and catalytic activity, involved in transcriptional silencing and required for sporula-tion; cells defective in SDS3 display pleiotropic phenotypes
2.12
CAGLOH03905g YCLOO9C MVO Regulatory subunit of acetolactate synthase, which catalyzes the first step of branched-chain amino acid biosynthesis; enhances activity of the Ily2p catalytic subunit, local- izes to mitochondria
2.11
CAGLOH05115g YLR385C SWC7 Protein of unknown function, corn- ponent of the Swrlp complex that incorporates Htzlp into chromatin
2.11
CAGL0101254g YHR111W UBA4 Protein that activates Urmlp be- fore its conjugation to proteins
2.11
(urmylation); one target is the thioredoxin peroxidase Ahplp, sug-gesting a role of urmylation in the oxidative stress response
CAGLOK03465g YMR108W ILV2 Acetolactate synthase, catalyses the first common step in isoleucine and valine biosynthesis and is the target of several classes of in- hibitors, localizes to the mitochon-dria; expression of the gene is under general amino acid control
2.11
CACLOL08976g YFLOO2C SPB4 Putative ATP-dependent RNA heli- case, nucleolar protein required for synthesis of 60S ribosomal subunits at a late step in the pathway; sedi-ments with 66S pre-ribosomes in su-crose gradients
2.11
CAGLOL10678g YOR276W CAF20 Phosphoprotein of the mRNA cap- binding complex involved in trans- lational control, repressor of cap- dependent translation initiation, competes with eIF4G for binding to eIF4E
2.11
CAGL0003960g YIL014W MNT3 Alpha-1,3-mannosyltransferase, adds the fourth and fifth alpha-
2.1
1,3-linked mannose residues to 0-linked glycans during protein 0-glycosylation
CAGLOG05742g YDL213C NOP6 Putative RNA-binding protein im- plicated in ribosome biogenesis; contains an RNA recognition mo-tif (RRM) and has similarity to hy-drophilins; NOP6 may be a fungal-specific gene as no homologs have been yet identified in higher eukary-otes
2.1
continued on next page

Appendix G. Transcription profiling of C. glabrata serum stress responses 391
continued from previous page CG ORF SC ORF gene name Description Log Ratio
CAGLOH02607g YOL052C SPE2 S-adenosylmethionine decarboxy-
of spermidine and spermine; cells lacking Spe2p require spermine or spermidine for growth in the
liesesc,rirpetqlenclotfomr atphpeedbiosynthesis 2.1
presence of oxygen but not when grown anaerobically
CAGLOK07183g - - 2.1 7i — C91111 - - Description not mapped 2.09
CAGLOA03674g YBR141C - Description not mapped 2.09 CAGLOB03993g YJR016C ILVS Dihydroxyacid dehydratase, cat-
alyzes third step in the common pathway leading to biosynthesis of branched-chain amino acids
2.09
CAGLOE01573g YNL313C - Description not mapped 2.09 CAGLOF00297g YJR075W HOC1 Alpha-1,6-mannosyltransferase in-
volved in cell wall mannan biosyn-thesis; subunit of a Golgi-localized complex that also contains Anplp, Mnn9p, Mnnllp, and MnnlOp; identified as a suppressor of a cell lysis sensitive pkcl-371 allele
2.09
CAGLOH05995g YAL049C AIMS Cytoplasmic protein of unknown function, potential Hsp82p interac-tor; YAL049C is not an essential gene
2.09
CAGLOH08910g YGL138C - Description not mapped 2.09 CAGL0109350g YBR267W REI1 Cytoplasmic pre-60S factor; re-
quired for the correct recycling of shuttling factors Albl, Arxl and
2.09
Tif6 at the end of the ribosomal large subunit biogenesis; involved in bud growth in the mitotic signaling network
CAGLOJ01375g YMRO88C VBAI Permease of basic amino acids in the vacuolar membrane
2.09
CAGLOJ03344g YER082C UTP7 Nucleolar protein, component of the small subunit (SSU) processome containing the XJ3 snoRNA that is involved in processing of pre-18S rRNA
2.09
CAGLDL05500g YJL 122W ALB1 Shuttling pre-60S factor; involved in the biogenesis of ribosomal large subunit; interacts directly with
2.09
Arxlp; responsible for Tif6p recy- ,..
cling defects in absence of Reilp CAGLOM00946g YLR435W TSR2 Protein with a potential role in pre-
rRNA processing 2.09
- - Description not mapped 2.08 CAGLOD03652g YMR179W SPT21 Protein required for normal tran-
scription at several loci including 2.08
HTA2-HTB2 and HHF2-HHT2, but not required at the other histone loci; functionally related to SptlOp; involved in telomere maintenance
CAGLOD03960g YHR036W BRL1 Essential nuclear envelope integral membrane protein identified as a suppressor of a conditional mu- tation in the major karyopherin, CRM1; homologous to and interacts with Brr6p, a nuclear envelope pro-tein involved in nuclear export
2.08
continued on next page

G.3 Transcriptional response of C. glabrata cells exposed to 10% Foetal Calf Serum (FCS) 392
continued from previous page CG ORF SC ORF gene name Description Log Ratio
CAGLOE01067g YDR161W - Description not mapped 2.08 CAGLOG04983g YLR363WA - Description not mapped 2.08 CAGLOH01375g YDR297W SUR2 Sphinganine C4-hydroxylase, catal-
yses the conversion of sphinganine to phytosphingosine in sphingolipid biosyntheis
2.08
CAGLOI10648g YGR124W ASN2 Asparagine synthetase, isozyme of 2.08 Asnlp; catalyzes the synthesis of L-asparagine from L-aspartate in the asparagine biosynthetic pathway
CAGLOJ01727g YIR019C MUC1 GPI-anchored cell surface glyco- protein (flocculin) required for pseudohyphal formation, invasive growth, flocculation, and biofilms; transcriptionally regulated by the
2.08
MAPK pathway (via Stel2p and Tee1p) and the cAMP pathway (via Flo8p)
CAGLOK02497g YPRO16C TIF6 Constituent of 66S pre-ribosomal particles, has similarity to hu- man translation initiation factor 6
2.08
(eIF6); may be involved in the bio-genesis and or stability of 60S ribo-somal subunits
CAGLOL08756g YPL012W RRP12 Protein required for export of the ribosomal subunits; associates with the RNA components of the pre-ribosomes; contains HEAT-repeats
2.08
CAGLOM12815g YIL073C SP022 Meiosis-specific protein essential for chromosome synapsis, involved in completion of nuclear divisions dur-ing meiosis; induced early in meiosis
2.08
CAGLOG08019g YDR090C - Description not mapped 2.07 CAGLOG08195g YLR168C UPS2 Putative protein of unknown func-
tion; similar to Ups1p, Ydr185Cp and to human PRELI; GFP-tagged protein localizes to mitochondria; required for wild-type respiratory growth
2.07
CAGLOH07183g YHRO7OW TRM5 tRNA(m(1)G37)methyltransferase, methylates a tRNA base adjacent to the anticodon that has a role in prevention of frameshifting; highly conserved across Archaea, Bacteria, and Eukarya
2.07
CAGLOI08195g YER020W GPA2 Nucleotide binding alpha subunit of the heterotrimeric G protein that interacts with the receptor Gprlp, has signaling role in response to nutrients; green fluorescent protein
2.07
(GFP)-fusion protein localizes to the cell periphery
CAGLOK07436g YMR241W YHM2 Mitochondria] DNA-binding pro- tein, component of the mitochon-drial nucleoid structure, involved in mtDNA replication and segregation of mitochondria] genomes; member of the mitochondrial carrier protein family
2.07
continued on next page

Appendix G. Transcription profiling of C. glabrata serum stress responses 393
continued from previous page CG ORF SC ORF gene name Description Log Ratio
CAGLOL02915g YOR204W DEDI ATP-dependent DEAD (Asp-Glu- 2.07 Ala-Asp)-box RNA helicase, re- quired for translation initiation of all yeast mRNAs; mutations in hu-man DEAD-box DBY are a frequent cause of male infertility
CAGLOB02145g YHR149C SKG6 Integral membrane protein that lo- calizes primarily to growing sites such as the bud tip or the cell periphery; potential Cdc28p sub- strate; Skg6p interacts with Zdslp and Zds2p
2.06
CAGLOD06534g -- Description not mapped 2.06 CAGLOF07645g YKL099C UTPII Subunit of U3-containing Small 2.06
Subunit (SSU) processome complex involved in production of 18S rRNA and assembly of small ribosomal subunit
CAGL0G01947g YKR071C DRE2 Protein of unknown function re- quired for iron-sulfur cluster assem-bly and sister chromatid cohesion; mutation displays synthetic lethal interaction with the po13-13 allele of CDC2
2.06
CAGLOH09152g YJR059W PTK2 Putative serine/threonine protein kinase involved in regulation of ion transport across plasma membrane; enhances spermine uptake
2.06
CAGLOJ03212g YER073W ALD5 Mitochondrial aldehyde dehydro- genase, involved in regulation or biosynthesis of electron transport chain components and acetate for-mation; activated by K+; utilizes
2.06
NADP+ as the preferred coenzyme; constitutively expressed
CAGLOL11660g YOR116C RP031 RNA polymerase III subunit C160, part of core enzyme; similar to bac-terial beta-prime subunit
2.06
CAGLOA03608g YOL155C HPF1 Haze-protective mannoprotein that reduces the particle size of aggre-gated proteins in white wines
2.05
C AG LOE100572g - - Description not mapped 2.05 CAGLOE02937g YGL029W CGRI Protein involved in nucleolar in-
tegrity and processing of the pre-rRNA for the 60S ribosome subunit; transcript is induced in response to cytotoxic stress but not genotoxic stress
2.05
CAGLOI03014g YNL282W POPS Subunit of both RNase MRP, which cleaves pre-rRNA, and nu-clear RNase P, which cleaves tRNA precursors to generate mature 5' ends
2.05
CAGL0107997g YOL076W MDM20 Non-catalytic subunit of the NatB 2.05 N-terminal acetyltransferase, which catalyzes N-acetylation of proteins with specific N-terminal sequences; involved in mitochondrial inheri- tance and actin assembly
CAGLOJ06028g YNL142W MEP2 Ammonium permease involved in regulation of pseudohyphal growth; belongs to a ubiquitous family of cytoplasmic membrane proteins that transport only ammonium
2.05
(NH4+); expression is under the ni-trogen catabolite repression regula-tion
continued on next page

G.3 Transcriptional response of C. glabrata cells exposed to 10% Foetal Calf Serum (FCS) 394
continued from previous page CG ORF SC ORF gene name Description Log Ratio
CAGLOL04290g
CAGLOB04191g CAGL0001419g
YOR156C
YPR114W YPL184C
TIFI1
- MRN1
SUMO ligase, catalyzes the covalent attachment of SUMO (Smt3p) to proteins; involved in maintenance of proper telomere length Description not mapped RNA-binding protein proposed to be involved in translational regu- lation; binds specific categories of mRNAs, including those that con-tain upstream open reading frames (uORFs) and internal ribosome en-try sites (IRES)
2.05
2.04 2.04
CAGLOD02596g YOL146W PSF3 Subunit of the GINS complex 2,04 (Sld5p, Psflp, Psf2p, Psf3p), which is localized to DNA replication ori-gins and implicated in assembly of the DNA replication machinery
CAGLOH04917g YDR088C SLUT RNA splicing factor, required for 2.04 ATP-independent portion of 2nd catalytic step of spliceosomal RNA splicing; interacts with Prpl8p; contains zinc knuckle domain
CAGLOH10384g YDL111C RRP4 2 Exosome non-catalytic core compo- nent; involved in 3'-5' RNA process-ing and degradation in both the nu-cleus and the cytoplasm; has simi-larity to E. coli RNase PH and to human hRrp42p (EXOSC7)
2.04
CAGL0105148g YDL174C DLD1 D-lactate dehydrogenase, oxidizes 2.04 D-lactate to pyruvate, transcription is heme-dependent, repressed by glucose, and derepressed in ethanol or lactate; located in the mitochon-dria' inner membrane
CAGLOK02937g YKL027W - Description not mapped 2.04 CAGLOK11319g YDR249C - Description not mapped 2.04 CAGL0005115g YOL058W ARG1 Arginosuccinate synthetase,
catalyzes the formation of L- argininosuccinate from citrulline and L-aspartate in the arginine biosynthesis pathway; potential
2.03
Cdc28p substrate CAGLOF02651g YDR403W DIT1 Sporulation-specific enzyme re-
quired for spore wall maturation, involved in the production of a soluble LL-dityrosine-containing precursor of the spore wall; tran- scripts accumulate at the time of prospore enclosure
2.03
CAGLOG09229g YPL192C PRM3 Pheromone-regulated protein re- quired for karyogamy; localizes to the inner membrane of the nuclear envelope
2.03
CAGLOH05973g YPL108W - Description not mapped 2.03 CAGLOK03861g YMR131C RRB1 Essential nuclear protein involved
in early steps of ribosome biogene-sis; physically interacts with the ri-bosomal protein Rpl3p
2.03
CAGLOL08206g YCR035C RI1P43 Exosome non-catalytic core compo- nent; involved in 3'-5' RNA process-ing and degradation in both the nu-cleus and the cytoplasm; has simi-larity to E. coli RNase PH and to human hRrp43p (01E2, EXOSCS)
2.03
continued on next page

Appendix G. Transcription profiling of C. glabrata serum stress responses395
continued from previous page CG ORF SC ORF gene name Description Log Ratio
CAGLOM01892g
CAGLOM09845g
YBR076W
YLR409C
ECM8
UTP21
Non-essential protein of unknown function Subunit of U3-containing 90S pre- ribosome and Small Subunit (SSU) processome complexes involved in production of 18S rRNA and as- sembly of small ribosomal sub- unit; synthetic defect with STI1
2.03
2.03
Hsp90 cochaperone; human ho- molog linked to glaucoma
CAGL0B04785g YCL010C SGF29 Probable subunit of SAGA histone acetyltransferase complex
2.02
CAGLOF03553g YPL170W DAP1 Heme-binding protein involved in regulation of cytochrome P450 pro-tein Ergl 1p; damage response pro-tein, related to mammalian mem-brane progesterone receptors; mu-tations lead to defects in telomeres, mitochondria, and sterol synthesis
2.02
CAGLOG04477g - - Description not mapped 2.02 CAGLOG06578g YNL099C OCA1 Putative protein tyrosine phos-
phatase, required for cell cycle ar-rest in response to oxidative dam-age of DNA
2.02
CAGLOH03773g YNLOO2C RLP7 Nucleolar protein with similarity to large ribosomal subunit L7 proteins; constituent of 66S pre- ribosomal particles; plays an essen- tial role in processing of precur- sors to the large ribosomal subunit
2.02
RNAs CAGLOJ04070g YOR224C RPB8 RNA polymerase subunit ABC14.5,
common to RNA polymerases I, II, and III
2.02
CAGL0001287g YIL114C POR2 Putative mitochondrial porin 2.01 (voltage-dependent anion channel), related to Porlp but not required for mitochondrial membrane per-meability or mitochondrial osmotic stability
CAGLOF01793g YLR056W ERGS C-5 sterol desaturase, catalyzes the introduction of a C-5(6) double bond into episterol, a precursor in ergosterol biosynthesis; mutants are viable, but cannot grow on non- fermentable carbon sources
2.01
CAGLOG08294g YLR243W Description not mapped 2.01 CAGLOH00440g YJR124C - Description not mapped 2.01 CAGLOJ09460g YIL079C AIR/ RING finger protein that inter-
acts with the arginine methyltrans-ferase Hmtlp to regulate methy-lation of Npl3p, which modulates
2.01
Npl3p function in mRNA process- ing and export; has similarity to Air2p
CAGLOK05753g YDR508C GNP1 High-affinity glutamine permease, also transports Leu, Ser, Thr, Cys, Met and Asn; expression is fully de-pendent on Grrlp and modulated by the Ssylp-Ptr3p-Ssy5p (SPS) sensor of extracellular amino acids
2.01
continued on next page

continued from previous page CG ORF SC ORF gene name Description Log Ratio
Description not mapped Description not mapped Protein required for mismatch re-pair in mitosis and meiosis, forms a complex with Msh2p to repair both single-base
Nucleolar protein required for ex-port of tRNAs from the nucleus; also copurifies with the small sub-unit (SSU) processome containing the U3 snoRNA that is involved in processing of pre-18S rRNA
MSH6
UTP8
YML053C
YDR097C
YGR128C
2.01 2.01
insertion-deletion mispairs; potentially phosphorylated by I
2
CAGLOL01573g GAGLOL09361g CAGLOG08129g
2 CAGL0110560g
G.3 Transcriptional response of C. glabrata cells exposed to 10% Foetal Calf Serum (FCS) 396

Appendix G. Transcription profiling of C. glabrata serum stress responses 397
Table G.2: Candida glabrata transcripts repressed in ATCC 2001 following ex-posure to Synthetic Complete + 10% (v/v) foetal calf serum (FCS) repressed transcripts are defined as having log2(Cy5) - log2(Cy3) < -2.0 and Benjamini-Hochberg corrected p-values < 0.05.
CG ORF SC ORF gene name Description Log Ratio CAGLOE04884g YDR216W ADR1 Carbon source-responsive zinc-
finger transcription factor, required for transcription of the glucose- repressed gene ADH2, of peroxi-somal protein genes, and of genes required for ethanol, glycerol, and fatty acid utilization
-9.77
CAGL0107249g YDR389W SAC7 GTPase activating protein (GAP) for Rho1p, involved in signaling to the actin cytoskeleton, null muta-tions suppress tort mutations and temperature sensitive mutations in actin; potential Cdc28p substrate
-9.63
CAGLOB01875g NORBH - Description not mapped -9.48 CAGLOJ03080g NORBH - Description not mapped -8.95 CAGLOG-09977g YPR184W GDB1 Glycogen debranching enzyme
containing glucanotranferase and alpha-1,6-amyloglucosidase ac- tivities, required for glycogen degradation; phosphorylated in mitochondria
-8.27
CAGLOK03421g YMR105C PGM2 Phosphoglucomutase, catalyzes the conversion from glucose-1- phosphate to glucose-6-phosphate, which is a key step in hexose metabolism; functions as the ac- ceptor for a Glc-phosphotransferase
-8.04
CAGLOE06380g YKL151C Description not mapped -8 CAGLOA02002g YFRO17C - Description not mapped -7.75 CAGLOH02387g YMR261C TPS8 Regulatory subunit of trehalose- -7.61
6-phosphate synthase/phosphatase complex, which synthesizes the storage carbohydrate trehalose; ex-pression is induced by stress con-ditions and repressed by the Ras-cAMP pathway
CAGLOI06050g YJL153C INOI Inositol 1-phosphate synthase, involved in synthesis of inositol phosphates and inositol-containing phospholipids; transcription is coregulated with other phospho- lipid biosynthetic genes by Ino2p and Ino4p, which bind the CASINO
-69.05
DNA element CAGLOF04719g YLR258W GSY2 Glycogen synthase, similar to -6.89
Gsylp; expression induced by glucose limitation, nitrogen star- vation, heat shock, and stationary phase; activity regulated by cAMP- dependent, Snflp and Pho85p kinases as well as by the Gaclp- Glc7p phosphatase
continued on next page

G.3 Transcriptional response of C. glabrata cells exposed to 10% Foetal Calf Serum (FCS) 398
continued from previous page CG ORF SC ORF gene name Description Log Ratio
CACLOJ04202g YFLO14W HSP12 Plasma membrane localized protein that protects membranes from des-iccation; induced by heat shock, ox-idative stress, osmostress, station-ary phase entry, glucose depletion, oleate and alcohol; regulated by the HOG and Ras-Pka pathways
-6.76
CAGLOF00605g YCLO4OW GLK1 Glucokinase, catalyzes the phos- phorylation of glucose at C6 in the first irreversible step of glucose metabolism; one of three glucose phosphorylating enzymes; expres- sion regulated by non-fermentable carbon sources
-6.73
CAGLOE05610g YAL038W CDC19 Pyruvate kinase, functions as a ho- motetramer in glycolysis to convert phosphoenolpyruvate to pyruvate, the input for aerobic (TCA cycle) or anaerobic (glucose fermentation) respiration
-6.67
CAGLOL06094g YGL184C STR3 Cystathionine beta-lyase, converts cystathionine into homocysteine
-6.66
CAGLOF02717g NORBH - Description not mapped -6.54 CAGLOF08107g YGR244C LSC2 Beta subunit of succinyl-CoA lig-
ase, which is a mitochondrial en- zyme of the TCA cycle that catalyzes the nucleotide-dependent conversion of succinyl-CoA to suc-cinate
-6.5
CAGLOJ01699g NORBH - Description not mapped -6.27 CAGLOK05687g YHR179W OYEZ Widely conserved NADPH oxidore-
ductase containing flavin mononu- cleotide (F MN), homologous to
-5.89
Oye3p with slight differences in lig- and binding and catalytic prop- erties; may be involved in sterol metabolism
CAGLOHO1111g YDR286C - Description not mapped -5.76 CAGLOK05813g NORBH - Description not mapped -5.67 CAGLOF04807g YIL136W OM45 Protein of unknown function, ma-
jor constituent of the mitochondrial outer membrane; located on the outer (cytosolic) face of the outer membrane
-5.63
CAGLOK03459g NORBH _ Description not mapped -5.54 CAGLOK07480g YMR105C PGM2 Phosphoglucomutase, catalyzes
the conversion from glucose-1- phosphate to glucose-6-phosphate, which is a key step in hexose metabolism; functions as the ac- ceptor for a Glc-phosphotransferase
-5.46
CAGLOL01925g YKL035W UGP1 UDP-glucose pyrophosphorylase -5.43 (UGPase), catalyses the reversible formation of UDP-Glc from glucose 1-phosphate and UTP, involved in a wide variety of metabolic pathways, expression modulated by Pho85p through Pho4p
continued on next page
Appendix G. Transcription profiling of C. glabrata serum stress responses 399
continued from previous page CG ORF SC ORF gene name Description Log Ratio
CAGLOB01078g CAGLOL00957g
YLR177W YER048C
- CAJ1
Description not mapped Nuclear type II J heat shock protein of the E. coli dnaJ family, contains a leucine zipper-like motif, binds to non-native substrates for presenta-tion to Ssa3p, may function during protein translocation, assembly and disassembly
-5.42 -5.31
CAGLOB01925g YDR122W KIN1 Serine/threonine protein kinase in- volved in regulation of exocytosis; localizes to the cytoplasmic face of the plasma membrane; closely re-lated to Kin2p
-5.29
CAGLOG05698g YDL215C GDH2 NAD(+)-dependent glutamate de- hydrogenase, degrades glutamate to ammonia and alpha-ketoglutarate; expression sensitive to nitrogen catabolite repression and intracellu- lar ammonia levels
-5.15
CAGLOD06512g YLR310C CDC25 Membrane bound guanine nu- cleotide exchange factor (GEF or
-5.08
GDP-release factor); indirectly regulates adenylate cyclase through activation of Ras1p and Ras2p by stimulating the exchange of GDP for GTP; required for progression through G1
CAGL0004587g YJR098C Description not mapped -5.03 CAGLOH0826Ig YOR019W - Description not mapped -5 CAGLOL04598g YLR270W DCS1 Non-essential hydrolase involved in
mRNA decapping, may function in a feedback mechanism to regulate deadenylation, contains pyrophos- phatase activity and a HIT (his- tidine triad) motif; interacts with neutral trehalase Nth1p
-4.92
CAGLOK04301g YGRO52W FMP48 Putative protein of unknown func- tion; the authentic, non-tagged pro- tein is detected in highly purified mitochondria in high-throughput studies; induced by treatment with
-4.9
8-methoxypsoralen and UVA irradi-ation
CAGLOE05148g YGL156W AMS1 Vacuolar alpha mannosidase, in- volved in free oligosaccharide (fOS) degradation; delivered to the vac-uole in a novel pathway separate from the secretory pathway
-4.82
CAGLOF07029g YGL125W MET13 Major isozyme of methylenete- trahydrofolate reductase, cat- alyzes the reduction of 5,10- methylenetetrahydrofolate to
-4.8
5-methyltetrahydrofolate in the methionine biosynthesis pathway
CAGLOI01980g YNL257C SIPS Protein that activates transcrip- tion through interaction with DNA-bound Snflp, C-terminal region has a putative leucine zipper motif; po-tential Cdc28p substrate
-4.79
CAGLOM03377g YEL011W GLC3 Glycogen branching enzyme, in- volved in glycogen accumulation; green fluorescent protein (GFP)- fusion protein localizes to the cyto-plasm in a punctate pattern
-4.79
continued on next page

G.3 Transcriptional response of C. glabrata cells exposed to 10% Foetal Calf Serum (FCS) 400
continued from previous page CC ORF SC ORF gene name Description Log Ratio
CAGLOE00803g YDR171W HSP42 Small heat shock protein (sHSP) with chaperone activity; forms barrel-shaped oligomers that sup-press unfolded protein aggregation; involved in cytoskeleton reorganiza-tion after heat shock
-4.72
CAGLOL11902g YER170W ADK2 Mitochondrial adenylate kinase, catalyzes the reversible synthesis of GTP and AMP from GDP and
-4.71
ADP; may serve as a back-up for synthesizing GTP or ADP depending on metabolic conditions; 3' sequence of ADK2 varies with strain background
CAGLOL00473g YMR187C - Description not mapped -4.66 CAGL0107359g YOL108C INN Transcription factor required for
derepression of inositol-choline- regulated genes involved in phos- pholipid synthesis; forms a com- plex, with Ino2p, that binds the inositol-choline-responsive element through a basic helix-loop-helix domain
-4.62
CAGLOH08778g YPRO42C PUF2 Member of the PUF protein fam- ily, which is defined by the pres-ence of Pumilio homology domains that confer RNA binding activity; preferentially binds mRNAs encod-ing membrane-associated proteins
-4.49
CAGLOB02860g YLR356W - Description not mapped -4.47 CAGLOE01837g YDR144C AI KC7 GPI-anchored aspartyl protease -4.45
(yapsin) involved in protein pro- cessing; shares functions with Yap3p and Kex2p
CAGLOF03267g YDR326C YSP2 Protein involved in programmed cell death; mutant shows resistance to cell death induced by amiodarone or intracellular acidification
-4.41
CAGLOM04763g YOR289W - Description not mapped -4.35 CAGLOM07403g YPL052W OAZ I Regulator of ornithine decarboxy-
lase (Spelp), antizyme that binds to Spelp to regulate ubiquitin- independent degradation; riboso- mal frameshifting during synthe- sis of Oazlp and its ubiquitin- mediated degradation are both polyamine-regulated
-4.34
CAGLOH02585g YMR250W GAD1 Glutamate decarboxylase, con- verts glutamate into gamma- aminobutyric acid (GABA) during glutamate catabolism; involved in response to oxidative stress
-4.31
CAGLOK00583g YCR034W FEN1 Fatty acid elongase, involved in sphingolipid biosynthesis; acts on fatty acids of up to 24 carbons in length; mutations have regulatory effects on 1,3-beta-glucan synthase, vacuolar ATPase, and the secretory pathway
-4.3
continued on next page

Appendix G. Transcription profiling of C. glabrata serum stress responses 401
continued from previous page CG ORF SC ORF gene name Description Log Ratio
CAGLOJ09812g YBR126C TPS1 Synthase subunit of trehalose-6- phosphate synthase/phosphatase complex, which synthesizes the storage carbohydrate trehalose; also found in a monomeric form; expression is induced by the stress response and repressed by the
-4.2
Ras-cAMP pathway CAGLOE01881g YLR120C YPS1 Aspartic protease, attached to the
plasma membrane via a glyco- sylphosphatidylinositol (GPI) an- chor
-4.15
CAGLOM04367g YLR133W CKII Choline kinase, catalyzing the first step in phosphatidylcholine synthesis via the CDP-choline
-4.15
(Kennedy pathway); exhibits some ethanolamine kinase ac- tivity contributing to phos- phatidylethanolamine synthesis via the CDP-ethanolamine pathway
CAGLOJ06050g YNL160W YGP1 Cell wall-related secretory gly- coprotein; induced by nutrient deprivation-associated growth ar- rest and upon entry into stationary phase; may be involved in adapta-tion prior to stationary phase entry; has similarity to Sps100p
-4.13
CAGLOB01947g YDR123C ING2 Component of the heteromeric -4.12 Ino2p/Ino4p basic helix-loop- helix transcription activator that binds inositol/choline-responsive elements (ICREs), required for derepression of phospholipid biosynthetic genes in response to inositol depletion
CAGLOD00198g YAL060W BDH1 NAD-dependent (R,R)-butanediol dehydrogenase, catalyzes oxida- tion of (R,R)-2,3-butanediol to
-4.12
(3R)-acetoin, oxidation of meso- butanediol to (3S)-acetoin, and reduction of acetoin; enhances use of 2,3-butanediol as an aerobic carbon source
CAGLOI05874g YIL053W RHR2 Constitutively expressed isoform of DL-glycerol-3-phosphatase; in- volved in glycerol biosynthesis, in-duced in response to both anaerobic and, along with the Hor2p/Gpp2p isoform, osmotic stress
-4.11
CAGLOH06699g YIL155C GUT2 Mitochondrial glycerol-3-phosphate dehydrogenase; expression is re- pressed by both glucose and cAMP and derepressed by non-fermentable carbon sources in a Snflp, Rsflp, Hap2/3/4/5 complex dependent manner
-4.1
CAGLOH06281g YAL016W TPD3 Regulatory subunit A of the het- erotrimeric protein phosphatase 2A, which also contains regulatory sub- unit Cdc55p and either catalytic subunit Pph2lp or Pph22p; re- quired for cell morphogenesis and for transcription by RNA poly- merase III
-4.09
continued on next page

G.3 Transcriptional response of C. glabrata cells exposed to 10% Foetal Calf Serum (FCS) 402
continued from previous page CG ORF SC ORF gene name Description Log Ratio
CAGLOJ01870g YGL167C PMR1 High affinity Ca2+/Mn2+ P-type -4.05 ATPase required for Ca2+ and Mn2+ transport into Golgi; in- volved in Ca2-1- dependent protein sorting and processing; mutations in human homolog ATP2C1 cause acantholytic skin condition Hailey- Hailey disease
CAGLOG05335g YDR074W TPS2 Phosphatase subunit of the trehalose-6-phosphate syn- thase/phosphatase complex, which synthesizes the storage carbohydrate trehalose; expression is induced by stress conditions and repressed by the Ras-cAMP pathway
-4.04
CAGL0000275g YDR533C HSP31 Possible chaperone and cysteine protease with similarity to E. coli Hsp31; member of the DJ-
-4.01
1/ThiJ/PfpI superfamily, which in- cludes human DJ-1 involved in Parkinson's disease; exists as a (timer and contains a putative metal-binding site
CAGLOL10186g YOR052C Description not mapped -3.96 CAGLOH02541g YMR252C - Description not mapped -3.95 CAGLOE05654g YPL206C PGC1 Phosphatidyl Glycerol phospho-
lipase C; regulates the phos- phatidylglycerol (PG) content via a phospholipase C-type degradation mechanism; contains glycerophos- phodiester phosphodiesterase motifs
-3.81
CAGLOL02827g YOR208W PTPS Phosphotyrosine-specific protein phosphatase involved in the in- activation of mitogen-activated protein kinase (MAPK) during osmolarity sensing; dephospory- lates Hog1p MAPK and regulates its localization; localized to the nucleus
-3.75
CAGLOI10516g YGR130C Description not mapped -3.72 CAGL01102893g YJL070C - Description not mapped -3.65 CAGLOH08558g YPRO49C ATG11 Adapter protein for pexophagy and
the cytoplasm-to-vacuole targeting -3.65
(Cvt) pathway; directs receptor- bound cargo to the phagophore as- sembly site (PAS) for packaging into vesicles; required for recruiting other proteins to the (PAS)
CAGL0003223g YLL041C SDH2 Iron-sulfur protein subunit of succi- nate dehydrogenase (Sdhlp, Sdh2p, Sdh3p, Sdh4p), which couples the oxidation of succinate to the trans-fer of electrons to ubiquinone
-3.63
CAGLOF02519g YJL206C Description not mapped -3.63 CAGLOG03245g YKR018C - Description not mapped -3.63 CAGLOH05775g YPL097W MSYI Mitochondrial tyrosyl-tRHA syn-
thetase -3.61
CAGLOL12012g YER175C TMTI Trans-aconitate methyltransferase, cytosolic enzyme that catalyzes the methyl esterification of 3- isopropylmalate, an intermediate of the leucine biosynthetic pathway, and trans-aconitate, which inhibits the citric acid cycle
-3.58
continued on next page

Appendix G. Transcription profiling of C. glabrata serum stress responses 403
continued from previous page CG ORF SC ORF gene name Description Log Ratio
CAGLOF05071g YLR284C EC11 Peroxisomal delta3,delta2-enoyl- -3.53 CoA isomerase, hexameric protein that converts 3-hexenoyl-CoA to trans-2-hexenoyl-CoA, essential for the beta-oxidation of unsaturated fatty acids, oleate-induced
CAGLOI09482g YBR273C UBX7 UBX (ubiquitin regulatory X) domain-containing protein that interacts with Cdc48p
-3.53
CAGLOJ08976g NORBH - Description not mapped -3.53 CAGLOL06820g YMR139W RIM11 Protein kinase required for sig-
nal transduction during entry into meiosis; promotes the formation of the Imelp-Ume6p complex by phosphorylating Imelp and Ume6p; shares similarity with mammalian glycogen synthase kinase 3-beta
-3.51
CAGLOH10362g YDL110C TMA17 Protein of unknown function that associates with ribosomes; het- erozygous deletion demonstrated increases in chromosome instability in a rad9 deletion background; pro-tein abundance is decreased upon intracellular iron depletion
-3.5
CAGLOL10582g YMR196W Description not mapped -3.5 CAGLOM08552g NORBH - Description not mapped -3.47 CAGLOI04246g YGL162W SUT1 Transcription factor of the -3.44
Zn[II]2Cys6 family involved in sterol uptake; involved in induction of hypoxic gene expression
CAGLOM02981g YMR278W PCMS Phosphoglucomutase, catalyzes interconversion of glucose-1- phosphate and glucose-6-phospate; transcription induced in response to stress; green fluorescent protein
-3.43
(GFP)-fusion protein localizes to the cytoplasm and nucleus; non-essential
CAGLOH04037g YOR178C CACI Regulatory subunit for Glc7p type- -3.42 1 protein phosphatase (PP1), teth-ers Glc7p to Gsy2p glycogen syn-thase, binds Hsflp heat shock tran-scription factor, required for induc-tion of some HSF-regulated genes under heat shock
CAGLOF03817g YMR210W - Description not mapped -3.41 CAGLOB02431g YML120C MDI1 NADFLubiquinone oxidoreductase,
transfers electrons from NADH to ubiquinone in the respiratory chain but does not pump protons, in contrast to the higher eukaryotic multisubunit respiratory complex I; phosphorylated; homolog of human
-3.4
AMID CAGLOK02629g YNL134C - Description not mapped -3.4 CAGLOK04367g YGRO55W MUPI High affinity methionine perme-
ase, integral membrane protein with -3.4
13 putative membrane-spanning re-gions; also involved in cysteine up-take
continued on next page

G.3 Transcriptional response of C. glabrata cells exposed to 10% Foetal Calf Serum (FCS) 404
continued from previous page CC ORF SC ORF gene name Description Log Ratio
CAGLOH03289g CAGLOH06259g
YGL082W YAL017W
- PSK1
Description not mapped One of two (see also PSK2) PAS domain containing S/T protein ki- nases; coordinately regulates pro- tein synthesis and carbohydrate metabolism and storage in response to a unknown metabolite that re-flects nutritional status
-3.39 -3.39
CAGLOF02101g YFLOO7W BLMIO Proteasome activator subunit; found in association with core particles, with and without the
-3.38
19S regulatory particle; required for resistance to bleomycin, may be involved in protecting against oxidative damage; similar to mammalian PA200
C AG LOM12474g YIL055C - Description not mapped -3.37 CAGLOA01221g YPR192W AQYI Spore-specific water channel that
mediates the transport of water across cell membranes, developmen-tally controlled; may play a role in spore maturation, probably by al-lowing water outflow, may be in-volved in freeze tolerance
-3.36
CAGLOE06138g YPL231W FAS2 Alpha subunit of fatty acid syn- thetase, which catalyzes the syn-thesis of long-chain saturated fatty acids; contains beta-ketoacyl reduc-tase and beta-ketoacyl synthase ac-tivities; phosphorylated
-3.35
CAGLOH02695g YJL137C GLG2 Self-glucosylating initiator of glyco- gen synthesis, also glucosylates n- dodecyl-beta-D-maltoside; similar to mammalian glycogenin
-3.34
CAGLOD01782g YGL197W MDS3 Protein with an N-terminal kelch- like domain, putative negative reg- ulator of early meiotic gene expres- sion; required, with Pmdlp, for growth under alkaline conditions
-3.32
CAGLOF04917g YOR178C GACI Regulatory subunit for Glc7p type- -3.29 1 protein phosphatase (PP1), teth-ers Glc7p to Gsy2p glycogen syn-thase, binds Hsflp heat shock tran-scription factor, required for induc-tion of some HSF-regulated genes under heat shock
CAGL0005027g YAR035W YAT1 Outer mitochondrial carnitine acetyltransferase, minor ethanol- inducible enzyme involved in transport of activated acyl groups from the cytoplasm into the mito-chondrial matrix; phosphorylated
-3.27
CAGLOL00649g YAL054C ACSI Acetyl-coA synthetase isoform which, along with Acs2p, is the nuclear source of acetyl-coA for histone acetlyation; expressed during growth on nonfermentable carbon sources and under aerobic conditions
-3.27
continued on next page

Appendix G. Transcription profiling of C. glabrata serum stress responses 405
continued from previous page CG ORF SC ORF gene name Description Log Ratio
CAGLOG05830g
CAGLOM11792g
YHR146W
YAL031C
CRP]
GIP4
Protein that binds to cruciform DNA structures Cytoplasmic Glc7-interacting pro- tein whose overexpression relocal-izes Glc7p from the nucleus and prevents chromosome segregation; potential Cdc28p substrate
-3.26
-3.24
CAGLOM13761g YMR302C YME2 Integral inner mitochondria] mem- brane protein with a role in maintaining mitochondrial nucleoid structure and number; mutants ex-hibit an increased rate of mitochon-drial DNA escape; shows some se-quence similarity to exonucleases
-3.24
CAGLOM08822g YDR258C HSP78 Oligomeric mitochondrial matrix chaperone that cooperates with
-3.22
Ssclp in mitochondria] thermotol-erance after heat shock; able to pre-vent the aggregation of misfolded proteins as well as resolubilize pro-tein aggregates
CAGLOK00231g YKL215C - Description not mapped -3.21 CAGLOA04455g YBL066C SEFI Putative transcription factor, has
homolog in Kluyveromyces lactis -3.2
CAGLOA00957g YLR324W PEX30 Peroxisomal integral membrane protein, involved in negative reg- ulation of peroxisome number; partially functionally redundant with Pex31p; genetic interac- tions suggest action at a step downstream of steps mediated by
-3.19
Pex28p and Pex29p CAGLOKOOS91g YGR205W Description not mapped -3.19 CAGLOG07601g YBR287W - Description not mapped -3.18 CAGLOF07975g YGR237C - Description not mapped -3.17 CAGLOG02717g YIL099W SGA1 Intracellular sporulation-specific
glucoamylase involved in glyco- gen degradation; induced during starvation of a/a diploids late in sporulation, but dispensable for sporulation
-3.17
CAGLOJ05830g YNL144C Description not mapped -3.14 CAGLOK03663g NORBH - Description not mapped -3.14 CAGLOK08624g YKL109W HAP4 Subunit of the heme-activated,
glucose-repressed Hap2p/3p/4p/5p -3.14
CCAAT-binding complex, a tran-scriptional activator and global reg-ulator of respiratory gene expres-sion; provides the principal activa-tion function of the complex
CAGLOF00363g YJR073C OPI3 Phospholi pi d methyltransferase -3.1 (methylene-fatty-acyl-phospholipid synthase), catalyzes the last two steps in phosphatidylcholine biosynthesis
CAGLOG07062g YML020W - Description not mapped -3.1 CAGLOH01287g YDR293C SSD1 Protein with a role in maintenance
of cellular integrity, interacts with components of the TOR pathway; ssdl mutant of a clinical S. cere-visiae strain displays elevated vir-ulence
-3.09
continued on next page

G.3 Transcriptional response of C. glabrata cells exposed to 10% Foetal Calf Serum (FCS) 406
continued from previous page CG ORF SC ORF gene name Description Log Ratio
CAGLOF03839g CAGLOD01270g CAGLOF07513g
YMR209C NORBH
YKL093W -
MBR1
Description not mapped Description not mapped Protein involved in mitochondrial functions and stress response; over-expression suppresses growth de-fects of hap2, hap3, and hap4 mu-tants
-3.08 -3.07 -3.07
CAGLOK10824g YLR149C - Description not mapped -3.07 CAGLOG03289g YBL075C SSA3 ATPase involved in protein folding
and the response to stress; plays a role in SRP-dependent cotransla-tional protein-membrane targeting and translocation; member of the heat shock protein 70 (HSP70) fam-ily; localized to the cytoplasm
-3.06
CAGLOM13255g YKL065C YETI Endoplasmic reticulum transmem- brane protein; may interact with ri- bosomes, based on co-purification experiments; homolog of human
-3.06
BAP31 protein CAGLOG04829g YML017W PSP2 Asn rich cytoplasmic protein that
contains RGG motifs; high-copy suppressor of group II intron- splicing defects of a mutation in
-3.02
MRS2 and of a conditional muta-tion in POLL (DNA polymerase al-pha); possible role in mitochondrial mRNA splicing
CAGLOI01012g YOR036W PEP12 Target membrane receptor (t- -3.02 SNARE) for vesicular intermedi- ates traveling between the Golgi apparatus and the vacuole; controls entry of biosynthetic, endocytic, and retrograde traffic into the prevacuolar compartment; syntaxin
CAGLOG09020g YPL203W TPK2 cAMP-dependent protein kinase catalytic subunit; promotes vegeta-tive growth in response to nutrients via the Ras-cAMP signaling pathway; inhibited by regulatory subunit Bcy1p in the absence of cAMP; partially redundant with
-3.01
Tpklp and Tpklp CAGLOJ06468g YMR261C TPS3 Regulatory subunit of trehalose- -3.01
6-phosphate synthase/phosphatase complex, which synthesizes the storage carbohydrate trehalose; ex-pression is induced by stress con-ditions and repressed by the Ras-cAMP pathway
CAGLOH08888g YPL221W FLC1 Putative FAD transporter; required for uptake of FAD into endoplas-mic reticulum; involved in cell wall maintenance
-3
CAGLOK03575g YMR114C - Description not mapped -3 CAGLOI06644g YDR077W SEDI Major stress-induced structural -24.11
GPI-cell wall glycoprotein in stationary-phase cells, associates with translating ribosomes, possi-ble role in mitochondrial genome maintenance; ORF contains two distinct variable minisatellites
continued on next page
Appendix G. Transcription profiling of C. glabrata serum stress responses 407
continued from previous page CG ORF SC ORF gene name Description Log Ratio
CAGLOM11000g CAGLOJ04004g CAGLOL10912g
NORBH YOR228C YOR273C
- -
TPO4
Description not mapped Description not mapped Polyamine transport protein, rec- ognizes spermine, putrescine, and spermidine; localizes to the plasma membrane; member of the major fa-cilitator superfamily
-21.21 -2.99 -2.99
CAGLOI01408g YJR048W CYC1 Cytochrome c, isoform 1; electron carrier of the mitochondrial inter-membrane space that transfers elec-trons from ubiquinone-cytochrome c oxidoreductase to cytochrome c oxidase during cellular respiration
-2.96
CAGLOL11880g YER169W RPH1 JmjC domain-containing histone demethylase which can specif- ically demethylate H3K36 tri- and dimethyl modification states; transcriptional repressor of PHR1;
-2.95
Rphlp phosphorylation during DNA damage is under control of the MEC1-RAD53 pathway
CAGLOD06600g YLL019C KNSI Nonessential putative protein ki- nase of unknown cellular role; member of the LAMMER fam- ily of protein kinases, which are serine/threonine kinases also ca- pable of phosphorylating tyrosine residues
-2.94
CAGLOM02211g YPL154C PEP4 Vacuolar aspartyl protease (pro- teinase A), required for the post-translational precursor maturation of vacuolar proteinases; important for protein turnover after oxidative damage; synthesized as a zymogen, self-activates
-2.94
CAGLOB03619g YEL060C PR131 Vacuolar proteinase B (yscB), a ser- ine protease of the subtilisin fam-ily; involved in protein degradation in the vacuole and required for full protein degradation during sporula-tion
-2.93
CAGLOF04213g YBL030C PETS Major ADP/ATP carrier of the mi- tochondrial inner membrane, ex- changes cytosolic ADP for mito-chondrially synthesized ATP; phos- phorylated; required for viability in many common lab strains carry-ing a mutation in the polymorphic
-2.93
SAL1 gene CAGLOH03971g YCROO4C YCP4 Protein of unknown function, has
sequence and structural similar- ity to flavodoxins; predicted to be pahnitoylated; the authentic, non-tagged protein is detected in highly purified mitochondria in high-throughput studies
-2.93
CAGLOB02838g YGRO55W MUPI High affinity methionine perme- ase, integral membrane protein with
-2.92
13 putative membrane-spanning re-gions; also involved in cysteine up-take
continued on next page

G.3 Transcriptional response of C. glabrata cells exposed to 10% Foetal Calf Serum (FCS) 408
continued from previous page CG ORF SC ORF gene name Description Log Ratio
CAGL0C05357g CAGLOK03245g CAGLOL12914g CAGLOD04026g
YNL200C NORBH
YMR031C YGRO19W
-
- UGA1
Description not mapped Description not mapped Description not mapped Gamma-aminobutyrate (GABA) transaminase (4-aminobutyrate aminotransferase) involved in the
-2.92 -2.92 -2.92 -2.91
4-aminobutyrate and glutamate degradation pathways; required for normal oxidative stress tolerance and nitrogen utilization
CAGLOG02915g YGRO86C PILL Primary component of eisosomes, which are large immobile cell cor-tex structures associated with en-docytosis; null mutants show acti-vation of Pkclp/Ypklp stress resis-tance pathways; detected in phos-phorylated state in mitochondria
-2.91
CAGLOI03190g YEL024W RIP/ Ubiquinol-cytochrome-c reductase, a Rieske iron-sulfur protein of the mitochondria] cytochrome bc1 complex; transfers electrons from ubiquinol to cytochrome cl during respiration
-2.91
CAGLOL10494g YOR070C GYPI Cis-golgi GTPase-activating pro- tein (GAP) for the Rab family members Yptlp (in vivo) and for
-2.91
Yptlp, Sec4p, Ypt7p, and Ypt5lp (in vitro); involved in vesicle dock-ing and fusion
CAGLOI01100g YOR120W GCYI Putative NADP(+) coupled glyc- erol dehydrogenase, proposed to be involved in an alternative pathway for glycerol catabolism; member of the aldo-keto reductase (AKR) fam-ily
-2.9
CAGL0101496g NORBH - Description not mapped -2.9 CAGL0107447g YOL103W ITR2 Myo-inositol transporter with
strong similarity to the major myo-inositol transporter Itrlp, member of the sugar transporter superfam-ily; expressed constitutively
-2.88
CAGLOK05137g YPRO26W ATH1 Acid trehalase required for utiliza- tion of extracellular trehalose
-2.87
CAGLOL04268g YOR155C ISN1 Inosine 5'-monophosphate (IMP)- specific 5'-nucleotidase, catalyzes the breakdown of IMP to inosine, does not show similarity to known
-2.87
5'-nucleotidases from other organ-isms
CAGLOE04548g NORBH Description not mapped -2.86 CAGLOK04037g NORBH - Description not mapped -2.86 CAGLOL00825g YIL044C AGE2 ADP-ribosylation factor (ARF) -2.86
GTPase activating protein (GAP) effector, involved in Trans-Golgi- Network (TGN) transport; contains C2C2H2 cysteine/histidine motif
CAGLOA04829g YGL253W HXK2 Hexokinase isoenzyme 2 that cat- alyzes phosphorylation of glucose in the cytosol; predominant hex- okinase during growth on glucose; functions in the nucleus to repress expression of HXK1 and GLK1 and to induce expression of its own gene
-2.85
continued on next page

Appendix G. Transcription profiling of C. glabrata serum stress responses 409
continued from previous page CG ORF SC ORF gene name Description Log Ratio
CAGLOK12892g
CAGLOL06006g
YFLO47W
YGL180W
RGD2
ATGI
GTPase-activating protein (RhoGAP) for Cdc42p and Rho5p Protein ser/thr kinase required for vesicle formation in autophagy and the cytoplasm-to-vacuole targeting (Cvt) pathway; structurally re- quired for phagophore assembly site formation; during autophagy forms a complex with Atgl3p and Atgl7p
-2.85
-2.85
CAGLOI06094g YJL155C FBP26 Fructose-2,6-bisphosphatase, re- quired for glucose metabolism
-2.84
CAGLOM06413g YPL084W BROI Cytoplasmic class E vacuolar pro- tein sorting (VPS) factor that coor-dinates deubiquitination in the mul-tivesicular body (MVB) pathway by recruiting Doa4p to endosomes
-2.84
CAGLOMI2793g YER079W - Description not mapped -2.84 CAGLOH01089g YDR287W INM2 Inositol monophosphatase, involved
in biosynthesis of inositol; enzy- matic activity requires magnesium ions and is inhibited by lithium and sodium ions; inml inm2 double mu-tant lacks inositol auxotrophy
-2.83
CAGLOL03982g YNL117W MLSI Malate synthase, enzyme of the glyoxylate cycle, involved in uti- lization of non-fermentable carbon sources; expression is subject to carbon catabolite repression; local-izes in peroxisomes during growth in oleic acid medium
-2.83
CAGLOK04719g YNL208W - Description not mapped -2.82 CAGLOM07381g YPL053C KTR6 Probable mannosylphosphate
transferase involved in the syn- thesis of core oligosaccharides in protein glycosylation pathway; member of the KRE2/MNT1 mannosyltransferase family
-2.82
CAGLOD00528g YKL182W FASI Beta subunit of fatty acid syn- thetase, which catalyzes the syn-thesis of long-chain saturated fatty acids; contains acetyltransacylase, dehydratase, enoyl reductase, mal- onyl transacylase, and palmitoyl transacylase activities
-2.81
CAGLOK10868g YDR256C CTA1 Catalase A, breaks down hydrogen peroxide in the peroxisomal ma- trix formed by acyl-CoA oxidase
-2.8
(Pox1p) during fatty acid beta- oxidation
CAGL0000110g YKR102W FLO10 Lectin-like protein with similarity to Flo1p, thought to be involved in flocculation
-2.78
CAGLOH03619g YNL011C - Description not mapped -2.78 CAGLOH08844g YMR173W DDR48 DNA damage-responsive protein,
expression is increased in response to heat-shock stress or treatments that produce DNA lesions; contains multiple repeats of the amino acid sequence NNNDSYGS
-2.77
CAGLOI05610g YNR014W - Description not mapped -2.77 CAGLOA04477g YBL067C UBP13 Putative ubiquitin carboxyl-
terminal hydrolase, ubiquitin- specific protease that cleaves ubiquitin-protein fusions
-2.76
continued on next page

G.3 Transcriptional response of C. glabrata cells exposed to 10% Foetal Calf Serum (FCS) 410
continued from previous page CC ORF SC ORF gene name Description Log Ratio
CAGLOE03388g YJL024C APS3 Small subunit of the clathrin- associated adaptor complex AP-3, which is involved in vacuolar pro- tein sorting; related to the sigma subunit of the mammalian clathrin
-2.76
AP-3 complex; suppressor of loss of casein kinase 1 function
CAGLOJ06270g YEL176W - Description not mapped -2.76 CAGLOL10780g YNR016C ACCI Acetyl-CoA carboxylase, biotin
containing enzyme that catalyzes the carboxylation of acetyl-CoA to form malonyl-CoA; required for de novo biosynthesis of long-chain fatty acids
-2.76
CAGLOF06325g YMR053C STBd Protein that interacts with Sin3p in a two-hybrid assay and is part of a large protein complex with Sin3p and Stblp
-2.75
CAGLOA02211g YDR343C HXT6 High-affinity glucose transporter of the major facilitator superfamily, nearly identical to Hxt7p, expressed at high basal levels relative to other
-2.74
HXTs, repression of expression by high glucose requires SNF3
CAGLOE04554g NORBH - Description not mapped -2.74 CAGLOF02255g YFR003C YP11 Inhibitor of the type I protein
phosphatase Glc7p, which is in- volved in regulation of a variety of metabolic processes; overproduc-tion causes decreased cellular con-tent of glycogen
-2.74
CAGLOH08063g YNL202W SPS19 Peroxisomal 2,4-dienoyl-CoA re- ductase, auxiliary enzyme of fatty acid beta-oxidation; homodimeric enzyme required for growth and sporulation on petroselineate medium; expression induced during late sporulation and in the presence of oleate
-2.74
CAGLOK08668g YIR017C MET28 Basic leucine zipper (bZIP) tran- scriptional activator in the Cbflp-
-2.74
Met4p-Met28p complex, partici- pates in the regulation of sulfur metabolism
CAGLOG10153g YDR529C QCR7 Subunit 7 of the ubiquinol cytochrome-c reductase com- plex, which is a component of the mitochondrial inner membrane electron transport chain; oriented facing the mitochondria] matrix;
-2.73
N-terminus appears to play a role in complex assembly
CAGLOK00803g YGR209C TRX2 Cytoplasmic thioredoxin isoenzyme of the thioredoxin system which protects cells against oxidative and reductive stress, forms LMA1 com-plex with Pbi2p, acts as a cofactor for Tsalp. required for ER-Golgi transport and vacuole inheritance
-2.73
continued on next page

Appendix G. Transcription profiling of C. gtabrata serum stress responses 411
continued from previous page CG ORF SC ORF gene name Description Log Ratio
CAGLOH06721g
CAGLOH04851g
YIL156W
YML016C
UBP7
PPZI
Ubiquitin-specific protease that cleaves ubiquitin-protein fusions Serine/threonine protein phos- phatase Z, isoform of Ppz2p; involved in regulation of potassium transport, which affects osmotic stability, cell cycle progression, and halotolerance
-2.72
-2.71
CAGLOB03685g YCROO4C YCP4 Protein of unknown function, has sequence and structural similar- ity to flavodoxins; predicted to be palmitoylated; the authentic, non-tagged protein is detected in highly purified mitochondria in high-throughput studies
-2.69
CAGLOD00704g YDL072C YET3 Protein of unknown function: YET3 null mutant decreases the level of secreted invertase; homolog of hu-man BAP31 protein
-2.69
CAGL0,107150g YALO51W OAF1 Oleate-activated transcription fac- tor, acts alone and as a heterodimer with Pip2p; activates genes in- volved in beta-oxidation of fatty acids and peroxisome organization and biogenesis
-2.69
CAGLOL06204g YGL191W COX13 Subunit Via of cytochrome c oxi- dase, which is the terminal member of the mitochondrial inner mem-brane electron transport chain; not essential for cytochrome c oxidase activity but may modulate activity in response to ATP
-2.69
CAGLOE06116g YPL230W US lil Putative transcription factor con- taining a C2H2 zinc finger; muta-tion affects transcriptional regula-tion of genes involved in growth on non-fermentable carbon sources, re-sponse to salt stress and cell wall biosynthesis
-2.68
CAGL0106765g YDR069C D0.4.4 Ubiquitin isopeptidase, required for recycling ubiquitin from proteasome-bound ubiquitinated intermediates, acts at the late endosome/prevacuolar compart- ment to recover ubiquitin from ubiquitinated membrane proteins en route to the vacuole
-2.68
CAGLOE04004g YHL036W MUP3 Low affinity methionine permease, similar to Muplp
-2.67
CAGL0100748g YMR145C NDEI Mitochondrial external NADH dehydrogenase, a type II
-2.67
NAD(P)H:quinone oxidoreduc- tase that catalyzes the oxidation of cytosolic NADH; Ndelp and Nde2p provide cytosolic NADH to the mitochondrial respiratory chain
CAGLOI08591g YER158C - Description not mapped -2.67 CAGLOK03509g YMR110C HFDI Putative fatty aldehyde dehydro-
genase, located in the mitochon- dria] outer membrane and also in lipid particles; has similarity to hu-man fatty aldehyde dehydrogenase
-2.67
(FALDH) which is implicated in Sjogren-Larsson syndrome
continued on next page

G.3 Transcriptional response of C. glabrata cells exposed to 10% Foetal Calf Serum (FCS) 412
continued from previous page CG ORF SC ORF gene name Description Log Ratio
CAGLOM13189g
CAGLOB03069g
YKL062W
YLR354C
MSN4
TALI
Transcriptional activator related to Msn2p; activated in stress condi- tions, which results in translocation from the cytoplasm to the nucleus; binds DNA at stress response ele-ments of responsive genes, inducing gene expression Transaldolase, enzyme in the non- oxidative pentose phosphate path- way; converts sedoheptulose 7- phosphate and glyceraldehyde 3- phosphate to erythrose 4-phosphate and fructose 6-phosphate
-2.67
-2.66
CAGLOE00869g YOL048C - Description not mapped -2.65 CAGL0G09449g YGRI89C CRH1 Putative chitin transglycosidase,
cell wall protein that functions in the transfer of chitin to beta(1-
-2.65
6)glucan; localizes to sites of polar-ized growth; expression is induced under cell wall stress conditions
CAGLOE02981g YGR149W - Description not mapped -2.64 CAGLOJ07304g YNL223W ATG4 Conserved cysteine protease re-
quired for autophagy; cleaves Atg8p to a form required for autophago-some and Cvt vesicle generation
-2.64
CAGLOJOS316g YNL277W MET2 L-homoserine-O-acetyltransferase, catalyzes the conversion of ho- moserine to 0-acetyl homoserine which is the first step of the methionine biosynthetic pathway
-2.64
CAGLOL08932g YPLOO4C LSP1 Primary component of eisosomes, which are large immobile patch structures at the cell cortex asso-ciated with endocytosis, along with
-2.64
PiIlp and Sur7p; null mutants show activation of Pkclp/Ypklp stress resistance pathways
CAGLOM05467g YBR204C - Description not mapped -2.64 CAGLOM13783g YMR304W UBP15 Ubiquitin-specific protease that
may play a role in ubiquitin precursor processing
-2.64
CAGLOCO3113g YLR270W DCSI Non-essential hydrolase involved in mRNA decapping, may function in a feedback mechanism to regulate deadenylation, contains pyrophos- phatase activity and a HIT (his- tidine triad) motif; interacts with neutral trehalase Nth1p
-2.62
CAGLOG01738g YGRO86C PIL1 Primary component of eisosomes, which are large immobile cell cor-tex structures associated with en-docytosis; null mutants show acti-vation of Pkclp/Ypklp stress resis-tance pathways; detected in phos-phorylated state in mitochondria
-2.61
CAGL0G05049g YDR063W AIM7 Putative protein of unknown func- tion; green fluorescent protein
-2.61
(GFP)-fusion protein localizes to the cytoplasm and nucleus; YDR063W is not an essential gene
continued on next page

Appendix G. Transcription profiling of C. glabrata serum stress responses 413
continued from previous page CG ORF SC ORF gene name Description Log Ratio
CAGLOK06083g NORBH - Description not mapped -2.61 CAGLOL02717g YOR215C A111441 Putative protein of unknown func- -2.61
Lion; the authentic protein is de-tected in highly purified mitochon- dria in high-throughput studies; YOR215C is not an essential gene
CAGLOM12551g YIL057C RGI2 Putative protein of unknown func- -2.61 Lion; expression induced under car-bon limitation and repressed under high glucose
CAGLOB01595g NORBH - Description not mapped -2.6 CAGLOL07744g YCR011C ADPI Putative ATP-dependent permease
of the ABC transporter family of proteins
-2.6
CAGLOL08426g YPR151C SUE/ Mitochondrial protein required for degradation of unstable forms of cy-tochrome c
-2.6
CAGLOL10406g YOR065W CYTI Cytochrome cl, component of the mitochondria] respiratory chain; expression is regulated by the heme-activated, glucose-repressed
-2.6
Hap2p/3p/4p/5p CCAAT-binding complex
CAGLOF04521g YBL043W ECM13 Non-essential protein of unknown function; induced by treatment with 8-methoxypsoralen and UVA irradiation
-2.59
CAGLOH05621g YPL089C RLM/ MADS-box transcription factor, component of the protein kinase
-2.59
C-mediated MAP kinase pathway involved in the maintenance of cell integrity; phosphorylated and activated by the MAP-kinase Sit2p
CAGLOJ04488g YLR417W VrS36 Component of the ESCRT-II corn- plea; contains the GLUE (GRAM
-2.59
Like Ubiquitin binding in EAP45) domain which is involved in interac-tions with ESCRT-I and ubiquitin-dependent sorting of proteins into the endosome
CAGL0005533g YDL237W LRC1 Putative protein of unknown func- tion; YDL237W is not an essential gene
-2.58
CAGLOM06325g YPL089C RLMI MADS-box transcription factor, component of the protein kinase
-2.58
C-mediated MAP kinase pathway involved in the maintenance of cell integrity; phosphorylated and activated by the MAP-kinase Slt2p
CAGLOA02233g YDR343C HXT6 High-affinity glucose transporter of the major facilitator superfamily, nearly identical to Hxt7p, expressed at high basal levels relative to other
-2.57
HXTs, repression of expression by high glucose requires SNF3
CAGLOF04565g YBL045C COR/ Core subunit of the ubiquinol- cytochrome c reductase complex
-2.57
(bc1 complex), which is a com- ponent of the mitochondrial inner membrane electron transport chain
CAGLOJ04268g YBL015W ACHI Acetyl-coA hydrolase, primarily lo- calized to mitochondria; phospho-rylated; required for acetate utiliza-tion and for diploid pseudohyphal growth
-2.57
continued on next page

G.3 Transcriptional response of C. glabrata cells exposed to 10% Foetal Calf Serum (FCS) 414
continued from previous page CG ORF SC ORF gene name Description Log Ratio
CAGLOK07678g CAGLOK12760g CAGLOF07777g
NORBH YEL042C YMR17OC
- -
ALD2
Description not mapped Description not mapped Cytoplasmic aldehyde dehydroge- nase, involved in ethanol oxida- tion and beta-alanine biosynthesis; uses NAD+ as the preferred coen-zyme; expression is stress induced and glucose repressed; very similar to Ald3p
-2.57 -2.56 -2.55
CAGLOL01265g YJR049C UTRI ATP-NADH kinase; phosphorylates both NAD and NADH; active as a hexamer; enhances the activity of ferric reductase (Fre1p)
-2.55
CAGLOL03498g YJL094C KHA 1 Putative K+/I-1-1- antiporter with a probable role in intracellular cation homeostasis, localized to Golgi vesi-cles and detected in highly purified mitochondria in high-throughput studies
-2.55
CAGLOB03201g YKR011C - Description not mapped -2.54 CAGLOF04631g YBL049W MOH/ Protein of unknown function, has
homology to kinase Snf7p; not required for growth on nonfer- mentable carbon sources; essential for viability in stationary phase
-2.54
CAGLOG01100g YLR345W - Description not mapped -2.53 CAGLOM03839g YNL305C Description not mapped -2.53 CAGLOF04697g YLR257W - Description not mapped -2.52 CAGL0104994g YER091C METS Cobalamin-independent methionine
synthase, involved in amino acid biosynthesis; requires a minimum of two glutamates on the methylte-trahydrofolate substrate, similar to bacterial metE homologs
-2.52
CAGLOJ00803g YJL042W MHP1 Microtubule-associated protein in- volved in assembly and stabiliza- tion of microtubules; overproduc- tion results in cell cycle arrest at
-2.52
G2 phase; similar to Drosophila protein MAP and to mammalian MAP4 proteins
CAGL0C04763g YCLO01W liERI Protein involved in retention of membrane proteins, including
-2.51
Secl2p, in the ER; localized to Golgi; functions as a retrieval receptor in returning membrane proteins to the ER
CAGL0,102200g YIROO1C SGNI Cytoplasmic RNA-binding protein, contains an RNA recognition mo- tif (RRM); may have a role in mRNA translation, as suggested by genetic interactions with genes en-coding proteins involved in transla-tional initiation
-2.51
CAGLOK06237g YDR452W PPN1 Vacuolar endopolyphosphatase with a role in phosphate metabolism; functions as a ho- modimer
-2.51
CAGLOL06182g YGL190C CDC55 Non-essential regulatory subunit B of protein phosphatase 2A, which has multiple roles in mitosis and protein biosynthesis; involved in regulation of mitotic exit; found in the nucleus of most cells, also at the bud neck and at the bud tip
-2.51
continued on next page

Appendix G. Transcription profiling of C. glabrata serum stress responses 415
continued from previous page CG ORF SC ORF gene name Description Log Ratio
CAGLOH10472g
CAGL0004191g
YDL115C
YBROO6W
IWR1
UGA2
Protein of unknown function, dele- tion causes hypersensitivity to the K1 killer toxin Succinate semialdehyde dehydroge- nase involved in the utilization of gamma-aminobutyrate (GABA) as a nitrogen source; part of the 4-aminobutyrate and glutamate degradation pathways; localized to the cytoplasm
-2.5
-2.49
CAGLOH10428g YDL113C A TG20 Sorting nexin family member required for the cytoplasm-to- vacuole targeting (Cvt) pathway and for endosomal sorting; has a
-2.49
Phox homology domain that binds phosphatidylinositol-3-phosphate; interacts with Snx4p; potential Cdc28p substrate
CAGL0105236g YIL033C BCY1 Regulatory subunit of the cyclic -2.49 AMP-dependent protein kinase (PICA), a component of a signaling pathway that controls a variety of cellular processes, including metabolism, cell cycle, stress response, stationary phase, and sporulation
CAGLOA03564g YLR039C RIC1 Protein involved in retrograde transport to the cis-Golgi network; forms heterodimer with Rgplp that acts as a GTP exchange factor for
-2.48
Ypt6p; involved in transcription of rRNA and ribosomal protein genes
CAGLOJ04158g YOR220W RCN2 Protein of unknown function; green fluorescent protein (GFP)-fusion protein localizes to the cytoplasm and is induced in response to the
-2.48
DNA-damaging agent MMS; phos-phorylated in response to alpha fac-tor
CAGLOK03003g YMR070W MOTS Nuclear transcription factor with two Cys2-His2 zinc fingers; involved in repression of a subset of hypoxic genes by Roxlp, repression of sev-eral DAN/TIR genes during aerobic growth, and repression of ergosterol biosynthetic genes
-2.48
CAGLOA03542g NORBH - Description not mapped -2.47 CAGL00O2321g YER037W PHMS Protein of unknown function, ex-
pression is induced by low phos- phate levels and by inactivation of
-2.47
Pho85p CAGL0005137g YOL059W GPD2 NAD-dependent glycerol 3-
phosphate dehydrogenase, homolog of Gpdlp, expression is controlled by an oxygen-independent signal-ing pathway required to regulate metabolism under anoxic con- ditions; located in cytosol and mitochondria
-2.47
continued on next page

G.3 Transcriptional response of C. glabrata cells exposed to 10% Foetal Calf Serum (FCS) 416
continued from previous page CG ORF SC ORF gene name Description Log Ratio
CAGLOF04741g YOL016C CMK2 Calmodulin-dependent protein kinase; may play a role in stress response, many CA++/calmodulan dependent phosphorylation sub- strates demonstrated in vitro, amino acid sequence similar to
-2.47
Cmklp and mammalian Cam Kinase II
CAGLOI00880g YMR137C P502 Required for a post-incision step in the repair of DNA single and double-strand breaks that result from interstrand crosslinks pro- duced by a variety of mono- and bi-functional psoralen derivatives; in-duced by UV-irradiation
-2.47
CAGLOK09350g YGL208W SIP2 One of three beta subunits of the -2.47 Snfl serine/threonine protein ki- nase complex involved in the re- sponse to glucose starvation; null mutants exhibit accelerated aging; N-myristoylprotein localized to the cytoplasm and the plasma mem-brane
CAGLOB03817g Y.111008W Description not mapped -2.46 CAGLOD02266g YOR352W - Description not mapped -2.45 CAGLOK02783g YKL007W CAPI Alpha subunit of the capping pro-
rein (CP) heterodimer (Cap1p and -2.45
Cap2p) which binds to the barbed ends of actin filaments prevent- ing further polymerization; local- ized predominantly to cortical actin patches
CAGLOL00803g YER054C GIP2 Putative regulatory subunit of the protein phosphatase Glc7p, in- volved in glycogen metabolism; contains a conserved motif (GVNK motif) that is also found in Gaclp, Pig1p, and Pig2p
-2.45
CAGLOF03707g YOR267C HRK1 Protein kinase implicated in activa- tion of the plasma membrane H(4-)-
-2.44
ATPase Pmalp in response to glu-cose metabolism; plays a role in ion homeostasis
CAGLOI07821g YOL087C DUF1 Putative protein of unknown func- tion; green fluorescent protein
-2.44
(GFP)-fusion protein localizes to the cytoplasm; deletion mutant is sensitive to various chemicals in-cluding phenanthroline, sanguinar-ine, and nordihydroguaiaretic acid
CAGLOJ02134g YILOO2C INP51 Phosphatidylinositol 4,5- bisphosphate 5-phosphatase, synaptojanin-like protein with an
-2.44
N-terminal Sacl domain, plays a role in phosphatidylinositol 4,5- bisphosphate homeostasis and in endocytosis; null mutation confers cold-tolerant growth
continued on next page

Appendix G. Transcription profiling of C. glabrata serum stress responses 417
continued from previous page CO ORF SC ORF gene name Description Log Ratio
CAGLOJ05962g CAGLOM13937g
YNL155W YMR311C
- GLC8
Description not mapped Regulatory subunit of protein phos- phatase 1 (Glc7p), involved in glycogen metabolism and chromo-some segregation; proposed to reg-ulate Glc7p activity via conforma- tional alteration; ortholog of the mammalian protein phosphatase in-hibitor 2
-2.44 -2.44
CAGL0CO2189g YER043C SAH1 S-adenosyl-L-homocysteine hy- drolase, catabolizes S-adenosyl-L- homocysteine which is formed after donation of the activated methyl group of S-adenosyl-L-methionine
-2.43
(AdoMet) to an acceptor CAGLOI10054g YGR143W SKN1 Protein involved in sphingolipid
biosynthesis; type II membrane pro-tein with similarity to Kre6p
-2.43
C AG LOK09900g YOR358W HAP5 Subunit of the heme-activated, glucose-repressed Hap2/3/4/5
-2.43
CCAAT-binding complex, a tran- scriptional activator and global regulator of respiratory gene ex- pression; required for assembly and DNA binding activity of the complex
CAGLOE05588g YOR346W REV1 Deoxycytidyl transferase, forms a complex with the subunits of DNA polymerase zeta, Rev3p and Rev7p; involved in repair of abasic sites in damaged DNA
-2.42
CAGLOM03025g YMR280C CATS Zinc cluster transcriptional acti- vator necessary for derepression of a variety of genes under non-fermentative growth conditions, ac-tive after diauxic shift, binds car-bon source responsive elements
-2.42
CAGL0G08712g YIL125W KGD1 Component of the mitochondria' alpha-ketoglutarate dehydrogenase complex, which catalyzes a key step in the tricarboxylic acid (TCA) cycle, the oxidative decarboxyla- tion of alpha-ketoglutarate to form succinyl-CoA
-2.41
CAGLOL06600g YGR232W NAS6 Regulatory, non-ATPase subunit of the 26S proteasome; homolog of the human oncoprotein gankyrin, which interacts with the retinoblastoma tumor suppressor (Rb) and cyclin-dependent kinase 4/6
-2.41
CAGLOA01782g YDR343C HXT6 High-affinity glucose transporter of the major facilitator superfamily, nearly identical to Hxt7p, expressed at high basal levels relative to other
-2.4
HXTs, repression of expression by high glucose requires SNF3
CAGLOK08338g YOR385W - Description not mapped -2.4 CAGLOB00814g YCL034W LSB5 Protein of unknown function; binds -2.39
Las17p, which is a homolog of human Wiskott-Aldrich Syndrome protein involved in actin patch as-sembly and actin polymerization
continued on next page

G.3 Transcriptional response of C. glabrata cells exposed to 10% Foetal Calf Serum (FCS) 418
continued from previous page CG ORF SC ORF gene name Description Log Ratio
CAGLOJ06380g CAGLOK08734g CAGLOJ10428g
YDL130WA YIR014W YKL092C
- -
BUD2
Description not mapped Description not mapped GTPase activating factor for Rsrlp/Budlp required for both axial and bipolar budding patterns; mutants exhibit random budding in all cell types
-2.39 -2.39 -2.38
CAGLOM03179g YJL082W IML2 Protein of unknown function; the authentic, non-tagged protein is de-tected in highly purified mitochon-dria in high-throughput studies
-2.38
CAGLOM04125g YNL320W - Description not mapped -2.38 CAGL0004785g YJR115W - Description not mapped -2.37 CAGL0105830g YNROO6W VPS27 Endosomal protein that forms a
complex with Hselp; required for recycling Golgi proteins, forming lumenal membranes and sorting ubiquitinated proteins destined for degradation; has Ubiquitin Inter- action Motifs which bind ubiquitin
-2.37
(Ubi4p) CAGLOK09878g YNL094W APPI Protein of unknown function, in-
teracts with Rvs161p and Rvs167p; computational analysis of protein- protein interactions in large-scale studies suggests a possible role in actin filament organization
-2.37
CAGLOD03982g YHR037W PUT2 Delta-1-pyrroline-5-carboxylate de- hydrogenase, nuclear-encoded mito-chondrial protein involved in uti- lization of proline as sole nitro- gen source; deficiency of the hu-man homolog causes HPII, an au-tosomal recessive inborn error of metabolism
-2.36
CAGLOI01122g YHRIO4W GRE.? Aldose reductase involved in methylglyoxal, d-xylose and ara- binose metabolism; stress induced
-2.36
(osmotic, ionic, oxidative, heat shock, starvation and heavy metals); regulated by the HOG pathway
CAGLOL04818g YG111.17C - Description not mapped -2.36 CAGL0G02849g YPL186C UIP4 Protein that interacts with Ulplp, a -2.35
Ubl (ubiquitin-like protein)-specific protease for Smt3p protein conju-gates; detected in a phosphorylated state in the mitochondrial outer membrane; also detected in ER and nuclear envelope
CAGLOH05291g YPLO7OW MUKI Cytoplasmic protein of unknown function; computational analysis of large-scale protein-protein interac-tion data suggests a possible role in transcriptional regulation
-2.35
CAGLOK10626g YFRO15C GSY1 Glycogen synthase with similar- ity to Gsy2p, the more highly expressed yeast homolog; expres- sion induced by glucose limitation, nitrogen starvation, environmental stress, and entry into stationary phase
-2.35
continued on next page

Appendix G. Transcription profiling of C. glabrata serum stress responses 419
continued from previous page CC ORF SC ORF gene name Description Log Ratio
CAGLOL05258g CAGLOH02959g
NORBH YGL096W
- TOSS
Description not mapped Homeodomain-containing protein and putative transcription factor found associated with chromatin; target of SBF transcription factor; induced during meiosis and under cell-damaging conditions; similar to Cup9p transcription factor
-2.34 -2.33
CAGL0102420g YHR171W ATG7 Autophagy-related protein and dual specificity member of the
-2.33
El family of ubiquitin-activating enzymes; mediates the conjugation of Atgl2p with Atg5p and Atg8p with phosphatidylethanolamine, required steps in autophagosome formation
CAGLOJ04906g YJL049W - Description not mapped -2.33 CAGLOM05951g YKR049C FMP46 Putative redox protein containing
a thioredoxin fold; the authen- tic, non-tagged protein is detected in highly purified mitochondria in high-throughput studies
-2.33
CAGLOF07051g YGL124C MON1 Protein required for fusion of cvt- vesicles and autophagosomes with the vacuole; associates, as a com-plex with Cczlp, with a perivacuo-lar compartment; potential Cdc28p substrate
-2.32
CAGL0102662g YMR284W YKU70 Subunit of the telomeric Ku corn- plex (Yku70p-Yku80p), involved in telomere length maintenance, struc-ture and telomere position effect; relocates to sites of double-strand cleavage to promote nonhomologous end joining during DSB repair
-2.32
CAGL0104972g NORBH - Description not mapped -2.32 CAGL0106182g YKL164C PIR1 0-glycosylated protein required for
cell wall stability; attached to the cell wall via beta-1,3-glucan; medi-ates mitochondrial translocation of
-2.32
Apnlp; expression regulated by the cell integrity pathway and by Swi5p during the cell cycle
CAGLOE01353g YLR130C ZRT2 Low-affinity zinc transporter of the plasma membrane; transcription is induced under low-zinc conditions by the Zap1p transcription factor
-2.31
CAGLOF06061g YMR041C ARA2 NAD-dependent arabinose dehydro- genase, involved in biosynthesis of dehydro-D-arabinono-1,4-lactone; similar to plant L-galactose dehy-drogenate
-2.31
CAGL0110582g YGR127W - Description not mapped -2.31 CAGLOK03113g YIIL014C YLF2 Protein of unknown function, has
weak similarity to E. coli GTP- binding protein gtpl; the authen-tic, non-tagged protein is detected in highly purified mitochondria in high-throughput studies
-2.31
continued on next page

G.3 Transcriptional response of C. glabrata cells exposed to 10% Foetal Calf Serum (FCS) 420
continued from previous page CG ORF SC ORF gene name Description Log Ratio
CAGLOH02491g CAGLOH03025g
NORBH YGL094C
- PANS
Description not mapped Essential subunit of the Pan2p- Pan3p poly(A)-ribonuclease com- plex, which acts to control poly(A) tail length and regulate the stoi-chiometry and activity of postrepli-cation repair complexes
-2.3 -2.3
CAGLOI00836g YMR140W SIPS Protein of unknown function; in- teracts with both the Reglp/Glc7p phosphatase and the Snflp kinase
-2.3
CAGLOK08800g YMR009W ADI1 Acireductone dioxygenease involved in the methionine salvage path- way; ortholog of human MTCBP-
-2.3
1; transcribed with YMR010W and regulated post-transcriptionally by RNase III (Rntlp) cleavage; ADI1 mRNA is induced in heat shock con-ditions
CAGL0001771g YBR241C - Description not mapped -2.29 CAGLOK10714g YOR018W ROD1 Membrane protein that binds the
ubiquitin ligase Rsp5p via its 2 PY motifs; overexpression confers re- sistance to the GST substrate o- dinitrobenzene,zinc, and calcium; proposed to regulate the endocyto-sis of plasma membrane proteins
-2.29
CAGLOG06952g YJL012C VTC4 Vacuolar membrane protein in- volved in vacuolar polyphosphate accumulation; functions as a regu-lator of vacuolar H-F-ATPase activ-ity and vacuolar transporter chap-erones; involved in non-autophagic vacuolar fusion
-2.27
CAGLOL05236g YKL085W AIDH1 Mitochondria] malate dehydroge- nase, catalyzes interconversion of malate and oxaloacetate; involved in the tricarboxylic acid (TCA) cy-cle; phosphorylated
-2.27
CAGLOG01342g YNL054W VAC7 Integral vacuolar membrane protein involved in vacuole inheritance and morphology; activates Fab1p kinase activity under basal conditions and also after hyperosmotic shock
-2.26
CAGLOG04081g YLROO4C THI73 Putative plasma membrane perme- ase proposed to be involved in carboxylic acid uptake and re- pressed by thiamine; substrate of
-2.26
Dbf2p/Moblp kinase; transcription is altered if mitochondrial dysfunc-tion occurs
CAGLOJ08294g YNL275W DORI Boron efflux transporter of the plasma membrane; binds HCO3-, I- , Br-, NO3- and Cl-; has similar- ity to the characterized boron efflux transporter A. thaliana BOR1
-2.26
CAGLOI00660g YLR289W GUF1 Mitochondria] matrix GTPase that associates with mitochondrial ri- bosomes; important for transla- tion under temperature and nutri-ent stress; may have a role in trans-lational fidelity; similar to bacterial
-2.25
LepA elongation factor continued on next page

Appendix G. Transcription profiling of C. glabrata serum stress responses 421
continued from previous page CG ORF SC ORF gene name Description Log Ratio
CAGLOI00902g
CAGLOI07139g
YMR136W
YOR142W
GAT2
LSC1
Protein containing GATA family zinc finger motifs; similar to Gln3p and Dal80p; expression repressed by leucine Alpha subunit of succinyl-CoA lig- ase, which is a mitochondrial en- zyme of the TCA cycle that catalyzes the nucleotide-dependent conversion of succinyl-CoA to suc-cinate; phosphorylated
-2.25
-2.25
CAGLOJ04180g YFLO13C IESI Subunit of the IN080 chromatin re- modeling complex
-2.25
CAGLOM13541g YMR291W - Description not mapped -2.25 CAGLOB01639g YDR137W RGP1 Subunit of a Golgi membrane ex-
change factor (Riclp-Rgplp) that catalyzes nucleotide exchange on
-2.24
Ypt6p CAGLOH06765g YIL159W BNRI Formin, nucleates the formation of
linear actin filaments, involved in cell processes such as budding and mitotic spindle orientation which require the formation of polarized actin cables, functionally redundant with BNI1
-2.24
CAGLOI08305g YER024W YAT2 Carnitine acetyltransferase; has similarity to Yatlp, which is a car-nitine acetyltransferase associated with the mitochondrial outer mem-brane
-2.24
CAGLOJ08613g YOR088W - Description not mapped -2.24 CAGLOM08206g YJL171C - Description not mapped -2.24 CAGLOD00660g YDL073W - Description not mapped -2.23 CAGLOE00583g YCR083W TRX3 Mitochondria! thioredoxin, highly
conserved oxidoreductase required to maintain the redox homeostasis of the cell, forms the mitochondrial thioredoxin system with Trr2p, re-dox state is maintained by both
-2.23
Trr2p and Glrlp CAGLOF01397g YOR014W RTS1 B-type regulatory subunit of pro-
tein phosphatase 2A (PP2A); ho-molog of the mammalian B' subunit of PP2A
-2.23
CAGLOG09603g YOR186W - Description not mapped -2.23 CAGL0104620g YBRO42C CST26 Protein of unknown function, af-
fects chromosome stability when overexpressed
-2.23
CAGLOM02255g YPL152W REDS Activator of the phosphoty- rosy) phosphatase activity of
-2.23
PP2A,peptidyl-prolyl cis/trans- isomerase; regulates GI phase progression, the osmoresponse, microtubule dynamics; subunit of the Tap42p-Pph2lp-Rrd2p complex
CAGLOI10494g YPR149W NCE102 Protein of unknown function; contains transmembrane domains; involved in secretion of proteins that lack classical secretory sig- nal sequences; component of the detergent-insoluble glycolipid- enriched complexes (DIGs)
-2.22
continued on next page

G.3 Transcriptional response of C. glabrata cells exposed to 10% Foetal Calf Serum (FCS) 422
continued from previous page CG ORF SC ORF gene name Description Log Ratio
CAGLOL07128g YDL091C UBX3 UBX (ubiquitin regulatory X) domain-containing protein that interacts with Cdc48p, green fluorescent protein (GFP)-fusion protein localizes to the cytoplasm in a punctate pattern
-2.22
CAGLOB01727g YDR109C - Description not mapped -2.21 CAGLOL08668g YPL015C HST2 Cytoplasmic member of the silent-
ing information regulator 2 (Sir2) family of NAD(+)-dependent pro-tein deacetylases; modulates nude-olar (rDNA) and telomeric silenc- ing; possesses NAD(+)-dependent histone deacetylase activity in vitro
-2.2
CAGLOB02035g YDR129C SAC6 Fimbrin, actin-bundling pro- tein; cooperates with Scplp
-2.19
(calponin/transgelin) in the orga- nization and maintenance of the actin cytoskeleton
CAGLOF02513g NORBH - Description not mapped -2.19 CAGLOH03113g YGLO9OW LIF1 Component of the DNA ligase IV
complex that mediates nonhomol-ogous end joining in DNA double-strand break repair; physically in-teracts with Dn14p and Nejlp; ho-mologous to mammalian XRCC4 protein
-2.19
CAGL0109812g YOR311C DGK1 Diacylglycerol kinase, localized to the endoplasmic reticulum
-2.19
(ER); overproduction induces enlargement of ER-like mem- brane structures and suppresses a temperature-sensitive slyl muta- tion; contains a CTP transferase domain
CAGLOK07315g NORBH - Description not mapped -2.19 CAGLOL04378g YOR161C PNS 1 Protein of unknown function; has
similarity to Torpedo californica tCTL1p, which is postulated to be a choline transporter, neither null mutation nor overexpression affects choline transport
-2.19
CAGLOB00748g YCL039W GID7 Protein of unknown function, involved in proteasome-dependent catabolite inactivation of fructose-
-2.18
1,6-bisphosphatase; contains six WD40 repeats; computational analysis suggests that Gid7p and Mohlp have similar functions
CAGLOG05247g YDR069C DOA4 Ubiquitin isopeptidase, required for recycling ubiquitin from proteasome-bound ubiquitinated intermediates, acts at the late endosome/prevacuolar compart- ment to recover ubiquitin from ubiquitinated membrane proteins en route to the vacuole
-2.18
continued on next page

Appendix G. Transcription profiling of C. glabrata serum stress responses 423
continued from previous page CG ORF SC ORF gene name Description Log Ratio
CAGLOH05137g YPL061W ALD6 Cytosolic aldehyde dehydrogenase, activated by Mg2+ and utilizes NADP+ as the preferred coenzyme; required for conversion of acetalde-hyde to acetate; constitutively ex- pressed; locates to the mitochon- dria] outer surface upon oxidative stress
-2.18
CAGLOH08437g YBRO97W VPS15 Myristoylated serine/threonine protein kinase involved in vacuo- lar protein sorting; functions as a membrane-associated complex with Vps34p; active form recruits
-2.18
Vps34p to the Golgi membrane; interacts with the GDP-bound form of Gpalp
CAGLOG07689g YKR014C YPT52 GTPase, similar to Ypt5lp and -2.17 Ypt53p and to mammalian rab5; re-quired for vacuolar protein sorting and endocytosis
CAGLOJ05940g YHRI35C YCK1 Palmitoylated plasma membrane- bound casein kinase I isoform; shares redundant functions with
-2.17
Yck2p in morphogenesis, proper septin assembly, endocytic traffick-ing; provides an essential function overlapping with that of Yck2p
CAGLOJ08118g YNL265C ISTI Protein with a positive role in the multivesicular body sorting path-way; functions and forms a com-plex with Did2p; recruitment to en-dosomes is mediated by the Vps2p-
-2.17
Vps24p subcomplex of ESCRT-III; also interacts with Vps4p
CAGLOL03135g YKR031C SPO/4 Phospholipase D, catalyzes the hydrolysis of phosphatidylcholine, producing choline and phospha- tidic acid; involved in Secl4p- independent secretion; required for meiosis and spore formation; dif- ferently regulated in secretion and meiosis
-2.17
CAGLOB01100g NORBH Description not mapped -2.16 CAGLOB02563g YML128C MSC/ Protein of unknown function; mu-
taut is defective in directing mei- otic recombination events to ho- mologous chromatids; the authen-tic, non-tagged protein is detected in highly purified mitochondria and is phosphorylated
-2.16
CAGLOG08932g YOL018C TLG2 Syntaxin-like t-SNARE that forms a complex with Tlglp and Vtilp and mediates fusion of endosome-derived vesicles with the late Golgi; binds Vps45p, which prevents
-2.16
T1g2p degradation and also facili-tates t-SNARE complex formation
CAGL0102046g YPR127W - Description not mapped -2.16 CAGLOL1111Og YML057W CMP2 Calcineurin A; one isoform (the
other is CNA1) of the cat- alytic subunit of calcineurin, a
-2.16
Ca+-1-/calmodulin-regulated pro- tein phosphatase which regulates Crzlp (a stress-response transcrip- tion factor), the other calcineurin subunit is CNB1
continued on next page

G.3 Transcriptional response of C. glabrata cells exposed to 10% Foetal Calf Serum (FCS) 424
continued from previous page CG ORF SC ORF gene name Description Log Ratio
CAGLOM10956g CAGLOG01474g
CAGLOH04983g
YCR073WA YNL042W
YDL055C
- BOPS
PSAI
Description not mapped Protein of unknown function, po- tential Cdc28p substrate; over- production confers resistance to methylmercury GDP-mannose pyrophospho- rylase (mannose-1-phosphate guanyltransferase), synthesizes
-2.16 -2.15
-2.15
GDP-mannose from GTP and mannose-l-phosphate in cell wall biosynthesis; required for normal cell wall structure
CAGL0106072g Y.11.154C VPS35 Endosomal subunit of membrane- associated retromer complex re- quired for retrograde transport; re-ceptor that recognizes retrieval sig-nals on cargo proteins, forms sub-complex with Vps26p and Vps29p that selects cargo proteins for re-trieval
-2.15
CAGLOL00583g YPL230W USVI Putative transcription factor con- taining a C2112 zinc finger; muta-tion affects transcriptional regula-tion of genes involved in growth on non-fermentable carbon sources, re-sponse to salt stress and cell wall biosynthesis
-2.15
CAGLOL09933g YOR042W CUES Protein containing a CUE domain that binds ubiquitin, which may fa-cilitate intramolecular monoubiqui-tination; green fluorescent protein
-2.15
(GFP)-fusion protein localizes to the cytoplasm in a punctate pattern
CAGLOE02695g YOR005C DNL4 DNA ligase required for non- homologous end-joining (NHEJ), forms stable heterodimer with re-quired cofactor Liflp, interacts with
-2.14
Nejlp; involved in meiosis, not es-sential for vegetative growth
CAGLOG07711g YKR016W A111128 Mitochondrial protein of unknown function; null mutation displays a synthetic slow growth defect in combination with a null mutation in
-2.14
GEM1, which encodes a mitochon-drial GTPase
CAGLOM02013g YPL166W ATG29 Autophagy-specific protein that is required for recruitment of other ATG proteins to the pre- autophagosomal structure (PAS); interacts with Atgl7p and localizas to the PAS in a manner interde-pendent with Atgl7p and Cis1p; not conserved
-2.14
CAGL0001397g YIL107C PFK26 6-phosphofructo-2-kinase, inhibited by phosphoenolpyruvate and sn- glycerol 3-phosphate, has negligi- ble fructose-2,6-bisphosphatase ac-tivity, transcriptional regulation in-volves protein kinase A
-2.13
continued on next page

Appendix G. Transcription profiling of C. glabrata serum stress responses 425
continued from previous page CG ORF SC ORF gene name Description Log Ratio
CAGLOD02464g YNL325C FIG4 Phosphatidylinositol 3,5- bisphosphate (PtdIns[3,51P) phosphatase; required for efficient mating and response to osmotic shock; physically associates with and regulated by Vacl4p; contains a SAC1-like domain
-2.13
CAGLOE06006g YPL224C MMT2 Putative metal transporter involved in mitochondrial iron accumulation; closely related to Mmtlp
-2.13
CAGLOH10120g YBRO56W - Description not mapped -2.13 CAGL0101034g YOR035C SH.E.4 Protein containing a UCS (UNC- -2.13
45/CRO1/SHE4) domain, binds to myosin motor domains to regulate myosin function; involved in endo-cytosis, polarization of the actin cy-toskeleton, and asymmetric mRNA localization
CAGL0105962g YJLI46W IDS2 Protein involved in modulation of -2.13 Ime2p activity during meiosis, ap-pears to act indirectly to promote Ime2p-mediated late meiotic func-tions; found in growing cells and de-graded during sporulation
CAGL0110384g YGR138C TPO2 Polyamine transport protein spe- cific for spermine; localizes to the plasma membrane; transcription of
-2.13
TPO2 is regulated by Haalp; mem-ber of the major facilitator super-family
CAGLOL13178g YLR077W FMP25 Putative protein of unknown func- tion; the authentic, non-tagged pro- tein is detected in highly purified mitochondria in high-throughput studies
-2.13
CAGLOM02387g YPL147W PXA1 Subunit of a heterodimeric peroxi- somal ATP-binding cassette trans-porter complex (Pxalp-Pxa2p), re- quired for import of long-chain fatty acids into peroxisomes; simi-larity to human adrenoleukodystro-phy transporter and ALD-related proteins
-2.13
CAGLOM11902g YAL034C FUNI9 Non-essential protein of unknown function; expression induced in re-sponse to heat stress
-2.13
CAGLOD04070g YGRO21W - Description not mapped -2.12 CAGLOD06204g YGL045W RIMS Protein of unknown function, in-
volved in the proteolytic activation of RimlOip in response to alkaline pH; has similarity to A. nidulans
-2.12
Pall"; essential for anaerobic growth CAGLOG10131g YPR191W QCR2 Subunit 2 of the ubiquinol
cytochrome-c reductase com- plex, which is a component of the mitochondrial inner membrane electron transport chain; phospho- rylated; transcription is regulated by Hap1p, Hap2p/Hap3p, and heme
-2.12
continued on next page

G.3 Transcriptional response of C. glabrata cells exposed to 10% Foetal Calf Serum (FCS) 426
continued from previous page CG ORF SC ORF gene name Description Log Ratio
CAGL01104631g YML007W YAP1 Basic leucine zipper (bZIP) tran- scription factor required for oxida- tive stress tolerance; activated by H202 through the multistep forma-tion of disulfide bonds and transit from the cytoplasm to the nucleus; mediates resistance to cadmium
-2.12
CAGLOH05049g YLR382C NAM2 Mitochondria' leucyl-tRNA syn- thetase, also has a direct role in splicing of several mitochondria] group I introns; indirectly re- quired for mitochondrial genome maintenance
-2.12
CAGLOI02904g YOR110W TFC7 One of six subunits of the RNA polymerase III transcription initia-tion factor complex (TFIIIC); part of the TauA globular domain of
-2.12
TFIIIC that binds DNA at the BoxA promoter sites of tRNA and similar genes
CAC LOL04686g YGRHOW - Description not mapped -2.12 CAGLOK01903g NORBH - Description not mapped -2.11 CAGLOK07590g YKL129C MYO3 One of two type I myosins; lo-
calizes to actin cortical patches; deletion of MY03 has little effect on growth, but myo3 myo5 dou-ble deletion causes severe defects in growth and actin cytoskeleton orga-nization
-2.11
CAGLOK10252g YOR124C UBP2 Ubiquitin-specific protease that re- moves ubiquitin from ubiquitinated proteins, cleaves at the C termi- nus of ubiquitin fusions; capable of cleaving polyubiquitin and pos-sesses isopeptidase activity
-2.11
CAGLOL03674g YJL103C GSMI Putative zinc cluster protein of un- known function; proposed to be in-volved in the regulation of energy metabolism, based on patterns of expression and sequence analysis
-2.11
CAGLOL03938g YNL115C - Description not mapped -2.11 CAGLOM01760g YOR153W PDR5 Plasma membrane ATP-binding
cassette (ABC) transporter, multidrug transporter actively reg-ulated by Pdrlp; also involved in steroid transport, cation resistance, and cellular detoxification during exponential growth
-2.11
CAGLOM09537g YBL095W - Description not mapped -2.11 CAGLOA03718g YGL206C CHC1 Clathrin heavy chain, subunit of
the major coat protein involved in intracellular protein transport and endocytosis; two heavy chains form the clathrin triskelion structural component; the light chain (CLC1) is thought to regulate function
-2.1
CAGL0004631g YJR100C AIM25 Putative protein of unknown func- tion; non-tagged protein is de- tected in purified mitochondria in high-throughput studies; similar to murine NORA; YJR100C is not an essential gene
-2.1
continued on next page

Appendix G. Transcription profiling of C. glabrata serum stress responses 427
continued from previous page CG ORF SC ORF gene name Description Log Ratio
CAGLOD02134g YMR115W MGRS Subunit of the mitochondrial (mt) i- AAA protease supercomplex, which degrades misfolded mitochondrial proteins; forms a subcomplex with
-2.1
Mgrlp that binds to substrates to facilitate proteolysis; required for growth of cells lacking mtDNA
CAGLOG04873g YLR371W ROM2 GDP/CTP exchange protein (GEP) for Rholp and Rho2p; mutations are synthetically lethal with muta-tions in roml, which also encodes a
-2.1
GEP CAGL0103652g YDL149W ATG9 Transmembrane protein involved in
forming Cvt and autophagic vesi-cles; cycles between the phagophore assembly site (PAS) and other cy- tosolic punctate structures, not found in autophagosomes; may be involved in membrane delivery to the PAS
-2.1
CAGLOJ07612g YNL241C ZWF1 Glucose-6-phosphate dehydroge- nase (G6PD), catalyzes the first step of the pentose phosphate pathway; involved in adapting to oxidatve stress; homolog of the human G6PD which is deficient in patients with hemolytic anemia
-2.1
CAGLOK11275g YDR247W VHS1 Cytoplasmic serine/threonine pro- tein kinase; identified as a high- copy suppressor of the synthetic lethality of a sis2 sit4 double mu- tant, suggesting a role in G1/S phase progression; homolog of
-2.1
Skslp CAGLOE01287g YDR148C KGD2 D ihydrolipoyl transsuccinylase,
component of the mitochondrial alpha-ketoglutarate dehydrogenase complex, which catalyzes the oxidative decarboxylation of alpha- ketoglutarate to succinyl-CoA in the TCA cycle; phosphorylated
-2.09
CAGLOE04400g YFIR011W DMA Probable mitochondrial seryl-tRNA synthetase, mutant displays in- creased invasive and pseudohyphal growth
-2.09
CAGLOH09812g YDROO6C SOK1 Protein whose overexpression sup- presses the growth defect of mu-tants lacking protein kinase A ac-tivity; involved in cAMP-mediated signaling; localized to the nucleus; similar to the mouse testis-specific protein PBS13
-2.09
CAGLOJ00429g YHR051W COX6 Subunit VI of cytochrome c oxidase, which is the terminal member of the mitochondrial inner membrane elec-tron transport chain; expression is regulated by oxygen levels
-2.09
CAGLOL02167g YKROO9C FOX2 Multifunctional enzyme of the per- oxisomal fatty acid beta-oxidation pathway; has 3-hydroxyacyl-CoA dehydrogenase and enoyl-CoA hy-dratase activities
-2.09
continued on next page

G.3 Transcriptional response of C. glabrata cells exposed to 10% Foetal Calf Serum (FCS) 428
continued from previous page CG ORF SC ORF gene name Description Log Ratio
CAGLOA01672g YGR009C SEC9 t-SNARE protein important for fu- sion of secretory vesicles with the plasma membrane; similar to but not functionally redundant with
-2.08
Spo20p; SNAP-25 homolog CAGL0B00946g YCL028W RNQI [PIN(+)] prion, an infectious pro-
tein conformation that is generally an ordered protein aggregate
-2.08
CAGL0104796g YB R037C SCO/ Copper-binding protein of the mito- chondrial inner membrane, required for cytochrome c oxidase activity and respiration; may function to de-liver copper to cytochrome c oxi-dase; has similarity to thioredoxins
-2.08
CAGLOLI0604g YMR197C VTII Protein involved in cis-Golgi mem- brane traffic; v-SNARE that inter-acts with two t-SNARES, Sed5p and Pepl2p; required for multiple vacuolar sorting pathways
-2.08
CAGL01-100979g YPL236C ENV7 -2.07 CAGL0100682g YMR150C IMPI Catalytic subunit of the mitochon-
drial inner membrane peptidase complex, required for maturation of mitochondrial proteins of the inter-membrane space; complex contains
-2.07
Imp1p and Imp2p (both catalytic subunits), and Som1p
CAGLOJ06820g YPL123C RNYI RNAse; member of the T(2) family of endoribonucleases
-2.07
CAGLOL06798g YDL078C MDH3 Peroxisomal malate dehydrogenase, catalyzes interconversion of malate and oxaloacetate; involved in the glyoxylate cycle
-2.07
CAGLOM04565g YLR144C ACF2
-
Intracellular beta-1,3- endoglucanase, expression is induced during sporulation; may have a role in cortical actin cytoskeleton assembly
-2.07
CAGLOH00682g YMR196W - Description not mapped -2.06 CAGL0109.504g YBR274W CHK I Serine/threonine kinase and DNA
damage checkpoint effector, me- diates cell cycle arrest via phos- phorylation of Pdslp; phosphory- lated by checkpoint signal trans- ducer Meclp; homolog of S. pombe and mammalian Chkl checkpoint kinase
-2.06
CAGL0,104796g YLR427W MAG2 Cytoplasmic protein of unknown function, predicted by computa- tional analysis of interaction data to encode a DNA-3-methyladenine glycosidase II that catalyzes the hy-drolysis of alkylated DNA
-2.06
CAGLOJ11704g YMR004W MVPI Protein required for sorting pro- teins to the vacuole; overproduction of Mvplp suppresses several domi-nant VPS1 mutations; Mvplp and
-2.06
Vpslp act in concert to promote membrane traffic to the vacuole
continued on next page

Appendix G. Transcription profiling of C. glabrata serum stress responses 429
continued from previous page CG ORF SC ORF gene name Description Log Ratio
CAGLOMOS492g YKL164C PH11 O-glycosylated protein required for cell wall stability; attached to the cell wall via beta-1,3-glucan; medi-ates mitochondrial translocation of
-2.06
Aprilp; expression regulated by the cell integrity pathway and by Swi5p during the cell cycle
CAGL0G06182g YHR131C - Description not mapped -2.05 CAGLOI01672g YJR036C HUL4 Protein with similarity to hect do-
main E3 ubiquitin-protein ligases, not essential for viability
-2.05
CAGLOJ09768g YDL200C MGT/ DNA repair methyltransferase (6- -2.05 O-methylguanine-DNA methylase) involved in protection against DNA alkylation damage
CAGLOM08360g YJL165C HAL5 Putative protein kinase; overex- pression increases sodium and lithium tolerance, whereas gene disruption increases cation and low pH sensitivity and impairs potassium uptake, suggesting a role in regulation of Trklp and/or
-2.05
Trk2p transporters CAGLOB01859g YDR118W APC4 Subunit of the Anaphase- -2.04
Promoting Complex/Cyclosome (APC/C), which is a ubiquitin- protein ligase required for degra- dation of anaphase inhibitors, including mitotic cyclins, during the metaphase/anaphase transition
CAGLOD01672g YPRO55W SECS Essential 121kDa subunit of the exocyst complex (Sec3p, Sec5p, Sec6p, Sec8p, SeclOp, Secl5p, Exo70p, and Exo84p), which has the essential function of mediat- ing polarized targeting of secretory vesicles to active sites of exocytosis
-2.04
CAGLOG03883g YLL026W HSP/04 Heat shock protein that cooper- ates with Ydjlp (Hsp40) and Ssalp
-2.04
(Hsp70) to refold and reactivate previously denatured, aggregated proteins; responsive to stresses in-cluding: heat, ethanol, and sodium arsenite; involved in [PSI+] propa-gation
CAGLOI08745g YGR170W PSD2 Phosphatidylserine decarboxylase of the Golgi and vacuolar mem-branes, converts phosphatidylserine to phosphatidylethanolamine
-2.04
CAGLOK12914g YFLO48C EMP47 Integral membrane component of endoplasmic reticulum-derived
-2.04
COPII-coated vesicles, which function in ER to Golgi transport
CAGL0M13343g YHR183W GND1 6-phosphogluconate dehydrogenase -2.04 (decarboxylating), catalyzes an NADPH regenerating reaction in the pentose phosphate pathway; required for growth on D-glucono- delta-lactone and adaptation to oxidative stress
continued on next page

G.3 Transcriptional response of C. glabrata cells exposed to 10% Foetal Calf Serum (FCS) 430
continued from previous page CG ORF SC ORF gene name Description Log Ratio
CAGLOD05632g CAGLOK09482g
NORBH YOR147W
- MDM32
Description not mapped Mitochondrial inner membrane pro- tein with similarity to Mdm31p, required for normal mitochondria' morphology and inheritance; in- teracts genetically with MMM1,
-2.03 -2.03
MDM10, MDM12, and MDM34 CAGLOL06512g NORI3H - Description not mapped -2.03 CAGLOF06567g YMR064W AEP1 Protein required for expression of
the mitochondrial OLD gene encod-ing subunit 9 of Fl-FO ATP syn-thase
-2.02
CAGLOG05467g YNL199C GCR2 Transcriptional activator of genes involved in glycolysis; interacts and functions with the DNA-binding protein Gcrlp
-2.02
CAGL0I00638g YLR290C - Description not mapped -2.02 CAGLOI01210g YIIR109W CTI111 Cytochrome c lysine methyltrans-
ferase, trimethylates residue 72 of apo-cytochrome c (Cyc1p) in the cytosol; not required for normal res-piratory growth
-2.02
CAGLOK02145g YER13OC COM2 Protein of unknown function -2.02 CAGLOL06160g YGL187C COX4 Subunit IV of cytochrome c oxidase,
which is the terminal member of the mitochondrial inner membrane elec-tron transport chain; N-terminal 25 residues of precursor are cleaved during mitochondria' import; phos-phorylated
-2.02
CAGLOL06292g YDR100W TVP1.5 Integral membrane protein local- ized to late Golgi vesicles along with the v-SNARE TIg2p
-2.02
CAGLOL08712g YPL014W - Description not mapped -2.02 CAGLOM03135g YJL084C A LY2 Protein proposed to regulate the
endocytosis of plasma membrane proteins; interacts with the cyclin
-2.02
Pcl7p and ubiquitin ligase Rsptp; phosphorylated by the cyclin-CDK complex, Pc17p-Pho85p; mRNA is cell cycle regulated, peaking in M phase
CAGLOM11682g YLI1108C - Description not mapped -2.02 CAGLOB04367g YBRO24W SCO2 Protein anchored to the mitochon-
drial inner membrane, similar to -2.01
Scolp and may have a redundant function with Scolp in delivery of copper to cytochrome c oxidase; in-teracts with Cox2p
CAGLOE05918g YPL219W PCL8 Cyclin, interacts with Pho85p cyclin-dependent kinase (Cdk) to phosphorylate and regulate glycogen synthase, also activates
-2.01
PhoS5p for Glc8p phosphorylation CAGLOG03179g YGRO97W ASK 10 Component of the RNA polymerase -2.01
II holoenzyme, phosphorylated in response to oxidative stress; has a role in destruction of Ssntp, which relieves repression of stress-response genes
continued on next page

Appendix G. Transcription profiling of C. glabrata serum stress responses 431
continued from previous page CG ORF SC ORF gene name Description Log Ratio
CAGL0105522g
CAGLOK03377g CAGLOK11990g
YER098W
YMR102C YBRO59C
UBP9
- AKLI
Ubiquitin carboxyl-terminal hy- drolase, ubiquitin-specific protease that cleaves ubiquitin-protein fusions Description not mapped Ser-Thr protein kinase, member (with Ark1p and Prklp) of the Ark kinase family; involved in endocyto-sis and actin cytoskeleton organiza-tion
-2.01
-2.01 -2.01
CAGLOL02431g YIL005W EPS1 ER protein with chaperone and co- chaperone activity, involved in re-tention of resident ER. proteins; has a role in recognizing proteins tar-geted for ER-associated degrada-tion (ERAD), member of the pro-tein disulfide isomerase family
-2.01
CAGLOL07634g YML002W - Description not mapped -2.01 CAGLOL12276g YLR090W XDJI Putative chaperone, homolog of E.
coli DnaJ, closely related to Ydjlp; the authentic, non-tagged protein is detected in highly purified mito-chondria in high-throughput stud-ies
-2.01
CAGL0104026g YOR227W HERI Protein of unknown function re- quired for proliferation or remod-eling of the ER that is caused by overexpression of Hmg2p; may in-teract with ribosomes, based on co-purification experiments
-2
CAGLOL03267g YKR039W GAPI General amino acid permease; lo- calization to the plasma membrane is regulated by nitrogen source
-2
CAGLOL03960g YNL116W DMA2 Protein involved in ubiquitin liga- tion; plays a role in regulating spin-dle position and orientation; func-tionally redundant with Dmalp; or-thologous to human RNF8 protein, also has sequence similarity to hu-man Chfr.
-2
CAGLOL11968g YER173W RAD24 Checkpoint protein, involved in the activation of the DNA damage and meiotic pachytene checkpoints; sub-unit of a clamp loader that loads
-2
Radl7p-Mec3p-Ddclp onto DNA; homolog of human and S. pombe Rad17 protein
CAGLOF08745g YLR327C TMAIO Protein of unknown function that associates with ribosomes
-14.24
CAGLOH04279g NORBH - Description not mapped -13.58 CAGLOH02101g YHROS7W RTC3 Protein of unknown function in-
volved in RNA metabolism; has structural similarity to SBDS, the human protein mutated in
-11.37
Shwachman-Diamond Syndrome (the yeast SBDS ortholog = SDO1); null mutation suppresses cdc13-1 temperature sensitivity
continued on next page

G.3 Transcriptional response of C. glabrata cells exposed to 10% Foetal Calf Serum (FCS) 432
continued from previous page CC ORF SC ORF gene name Description Log Ratio
CAGLOF04895g YPR160W GPH1 Non-essential glycogen phosphory- lase required for the mobilization of glycogen, activity is regulated by cyclic AMP-mediated phospho-rylation, expression is regulated by stress-response elements and by the HOG MAP kinase pathway
-10.49
Table G.3: Over-represented GO component terms for C. glabrata transcripts induced by 10% FCS (w/v).
Process Term p , nucleolus < 1 X 10—' nuclear lumen < 1 x 10-5 cell part < 1 x 10-5 cell < 1 x 10-5 membrane-enclosed lumen < 1 x 10-5 organelle lumen < 1 X 10-5 nucleus < 1 x 10-5 nuclear part < 1 x 10-5 membrane-bound organelle < 1 x 10-5 intracellular membrane-bound organelle < 1 x 10-5 nucleolar part < 1 x 10-5 intracellular < 1 x 10-5 non-membrane-bound organelle < 1 X 10-5 intracellular non-membrane-bound organelle < 1 x 10-5 intracellular part < 1 x 10-5 intracellular organelle < 1 x 10-5 organelle < 1 x 10-5 RNA polymerase complex < 1 x 10-5 DNA-directed RNA polymerase III complex < 1 x 10-5 DNA-directed RNA polymerase I complex < 1 x 10-5 organelle part < 1 x 10-5 intracellular organelle part < 1 x 10-5 small nucleolar ribonucleoprotein complex < 1 x 10-5 preribosome < 1 x 10-5 small subunit processome < 1 X 10-5 nucleolar preribosome < 1 x 10-5
Table GA: Over-represented GO process terms for C. glabrata transcripts in-duced by 10% FCS (w/v).
Process Term p ribosome biogenesis < 1 x 10—' cytoplasm organization and biogenesis < 1 X 10-5 ribosome biogenesis and assembly < 1 x 10-5 rRNA processing < 1 x 10-5 rRNA metabolism < 1 x 10-5 RNA processing < 1 x 10-5 RNA metabolism < 1 X 10-5 35S primary transcript processing < 1 x 10-5 nucleobase, nucleoside, nucleotide and nucleic acid metabolism < 1 x 10-5 organelle organization and biogenesis < 1 x 10-5 ribosomal large subunit biogenesis < 1 x 10-5 processing of 20S pre-rRNA < 1 x 10-5 cell organization and biogenesis < 1 x 10-5 biopolymer metabolism < 1 X 10-5 cellular process < 1 x 10-5 cellular physiological process < 1 x 10-5 primary metabolism < 1 x 10-5 physiological process < 1 x 10-5 transcription from RNA polymerase I promoter < 1 X 10-5
continued on next page

Appendix G. Transcription profiling of C. glabrata serum stress responses 433
continued from previous page Process Term p
metabolism < 1 x 10-5 tRNA modification < 1 x 10-5 cellular metabolism < 1 x 10-5 transcription from RNA polymerase III promoter < 1 x 10-5 ribosome assembly < 1 x 10-5 amino acid and derivative metabolism < 1 x 10-5 tRNA methylation < 1 X 10-5 amine biosynthesis < 1 x 10-5 nitrogen compound biosynthesis < 1 x 10-5 amino acid biosynthesis < 1 X 10-5 ribosomal large subunit assembly and maintenance < 1 X 10-5 amino acid metabolism < 1 X 10-5 tRNA metabolism < 1 x 10-5 nitrogen compound metabolism < 1 x 10-5 ribosomal subunit assembly < 1 X 10-5 processing of 27S pre-rRNA < 1 X 10-5 ribosomal large subunit export from nucleus < 1 x 10-5 macromolecule metabolism < 1 x 10-5 amine metabolism < 1 X 10-5
Table G.5: Over-represented GO function terms for C. glabrata transcripts in-duced by 10% FCS.
Function Term p snoRNA binding < 1 x 10-5 DNA-directed RNA polymerase activity < 1 X 10-5 catalytic activity < 1 X 10-5 tRNA methyltransferase activity < 1 X 10-5 RNA methyltransferase activity < 1 x 10-5 RNA binding < 1 X 10-5 transferase activity < 1 x 10-5 methyltransferase activity < 1 X 10-5 nucleotidyltransferase activity < 1 x 10-5 transferase activity, transferring one-carbon groups < 1 X 10-5 ATP-dependent RNA helicase activity < 1 X 10-5 RNA helicase activity < 1 x 10-5 RNA-dependent ATPase activity < 1 X 10-5 rRNA primary transcript binding < 1 X 10-5 S-adenosylmethionine-dependent methyltransferase activity < 1 X 10-5
Table G.6: Over-represented GO component terms for C. glabrata transcripts repressed by 10% FCS (w/v).
Process Term p cytoplasm < 1 X 10-5 cell < 1 X 10-5 cell part < 1 X 10-5 intracellular < 1 X 10-5 intracellular part < 1 X 10-5 mitochondrion < 1 X 10-5 cytoplasmic part < 1 X 10-5 mitochondrial electron transport chain < 1 x 10-5 respiratory chain complex III (sensu Eukaryota) < 1 X 10-5 respiratory chain complex III < 1 x 10-5 ubiquinol-cytochrome-c reductase complex < 1 x 10-5 protein serine/threonine phosphatase complex < 1 x 10-5
Table G.7: Over-represented GO process terms for C. glabrata transcripts re-pressed by 10% FCS (w/v).
Process Term p generation of precursor metabolites and energy < 1 X 10-5
continued on next page

G.3 Transcriptional response of C. glabrata cells exposed to 10% Foetal Calf Serum (FCS) 434
continued from previous page Process Term P
energy derivation by oxidation of organic compounds < 1 x 10-5
carbohydrate metabolism < 1 x 10-5
cellular carbohydrate metabolism < 1 x 10-5
energy reserve metabolism < 1 x 10-5 main pathways of carbohydrate metabolism < 1 X 10-5 glycogen metabolism < 1 x 10-5
monosaccharide metabolism < 1 x 10-5 glucan metabolism < 1 X 10-5 response to stress < 1 X 10-5 hexose metabolism < 1 x 10-5
alcohol metabolism < 1 X 10-5 carbohydrate catabolism < 1 x 10-5 cellular carbohydrate catabolism < 1 X 10-5 phosphorus metabolism < 1 X 10-5 phosphate metabolism < 1 x 10-5 acetyl-CoA metabolism < 1 x 10-5 response to stimulus < 1 x 10-5 organic acid metabolism < 1 x 10-5
carboxylic acid metabolism < 1 x 10-5
glucose metabolism < 1 x 10-5 monosaccharide catabolism < 1 X 10-5 polysaccharide metabolism < 1 x 10-5
cellular polysaccharide metabolism < 1 x 10-5 glycogen biosynthesis < 1 X 10-5 aerobic respiration < 1 X 10-5 alcohol catabolism < 1 x 10-5 response to external stimulus < 1 X 10-5 response to extracellular stimulus < 1 X 10-5 response to nutrient levels < 1 X 10-5 ATP synthesis coupled electron transport < 1 X 10-5 ATP synthesis coupled electron transport (sensu Eukaryota) < 1 X 10-5 cellular respiration < 1 x 10-5
electron transport < 1 x 10-5 glucan biosynthesis < 1 x 10-5
glucose catabolism < 1 x 10-5 carbohydrate biosynthesis < 1 x 10-5 tricarboxylic acid cycle < 1 x 10-5 acetyl-CoA catabolism < 1 x 10-5
Table G.8: Over-represented GO function terms for C. glabrata transcripts re-pressed by 10% FCS.
Function Term p catalytic activity < 1 x 10-5
oxidoreductase activity < 1 x 10-5 kinase activity < 1 x 10-5 phosphotransferase activity, alcohol group as acceptor < 1 X 10-5 enzyme regulator activity < 1 x 10-5
phosphoric monoester hydrolase activity < 1 X 10-5 phosphoric ester hydrolase activity < 1 x 10-5 cysteine-type peptidase activity < 1 X 10-5

435
References
[1] A DE LAS PENAS, PAN, S.-J., NO, I. C., ALDER, J., CREDO, R., AND COMMACK, B. P. Virulence-related surface glycoproteins in the yeast pathogen Candida glabrata are encoded in subtelomeric clusters and subject to RAP-I and SIR-dependent transcriptional
silencing. Genes and Development 17 (2003), 2245-2258.
[2] ABE, F., AND HIRAM, T. Mechanistic role of ergosterol in membrane rigidity and cycloheximide resistance in Saccharomyces cerevisiae.
Biochirn Biophys Acta 1788, 3 (2009 Mar), 743-752.
[3] ADAMSON, I. Y., KING, G. M., AND BOWDEN, D. H. Collagen breakdown during acute lung injury. Thorax 43, 7 (Jul 1988), 562-568.
[4] AGATENSI, L., FRANCIII, F., MONDELLO, F., BEVILACQUA, R. L., CEDDIA, T., DE BERNARDIS, F., AND CASSONE, A. Vaginopathic and proteolytic
Candida species in outpatients attending a gynaecology clinic. J Clin Pathol 44, 10 (Oct 1991), 826-830.
[5] AGNAN, J., KORCII, C., AND SELITRENNIKOEE, C. Cloning heterologous genes: problems and approaches. Fungal Genet Bird 21, 3 (Jun
1997), 292-301.
[6] AIIN, S., AND TANKSLEY, S. D. Comparative linkage maps of the rice and maize genomes. Proc Nail Acad Sci U S A 90, 17 (1993),
7980-7984.
[7] AISNER, J., WIERNIK, P. H., AND SCIIIMPFF, S. C. Treatment of invasive aspergillosis: relation of early diagnosis and treatment to response. Ann Intern Med 86, 5 (1977 May), 539-543.
[8] ALARCO, A. M., AND RAYMOND, M. The bZip transcription factor Cap1p is involved in multidrug resistance and oxidative stress
response in Candida allocans. J Bacterial 181, 3 (Feb 1999), 700-708.
[9] ALBERT, R., AND BARABASI, A. L. Statistical mechanics of complex networks. Reviews of Modern Physics 74, 1 (Jan 2002), 47-97.
[10] ALBERT, R., JEONG, H., AND BARABASI, A. Error and attack tolerance of complex networks. Nature 406,6794 (Jul 2000), 378-382.
[11] ALBERTYN, J., HOHMANN. S., AND PRIOR, B. A. Characterization of the osmotic-stress response in Saccharomyees cc:miasma: osmotic
stress and glucose repression regulate glycerol-3-phosphate dehydrogenase independently. Curr Genet 25, 1 (Jan 1994), 12-18.
[12] ALBERTYN, J., HOHMANN, S., THEVELEIN, J. M., AND PRIOR, B. A. GPD1, which encodes glycerol-3-phosphate dehydrogenase, is essential
for growth under osmotic stress in Saccharomyces ccrevisiae, and its expression is regulated by the high-osmolarity glycerol response
pathway. Mol Cell Biol 14, 6 (1994 Jun), 4135-4144.
[13] ALEPUZ, P. M., DE NADAL. E., ZAPATER, M AMMERER, G., AND POSAS, F. Osmostress-induced transcription by HOT1 depends on a
Hogl-mediated recruitment of the RNA Pol II. EMBO J 22, 10 (2003 May 15), 2433-2442.
[14] ALONSO-MONGE, R., NAVARRO-GARCIA, F., MOLERO, G., DIEZ-OREJAS, R., GUSTIN, M„ PLA, J., SANCHEZ, M., AND NOMBELA, C. Role of the mitogen-activated protein kinase Hog1p in morphogenesis and virulence of Candida albicans. J Bacterial 181, 10 (1999 May),
3058-3068.
[15] ALONSO-MONGE, R., NAVARRO-GALICIA, F., ROMAN, E., NEGREDO, A. I., EISMAN, B., NOMBELA, C., AND PLA, J. The Hogl mitogen-activated protein kinase is essential in the oxidative stress response and chlamydospore formation in Candida alhicans. Eakaryot Cell 2, 2
(2003 Apr), 351-361.
[16] ALTSCIIUL, S. F., Gtsa, W., MILLER, W., MYERS, E. W., AND LIPMAN. D. J. Basic local alignment search tool. Journal of Molecular Biology
215 (1990), 403-410.
[17] AMAAR, Y. G., AND MOORE, M. M. Mapping of the nitrate-assimilation gene cluster (crnA-niiA-niaD) and characterization of the nitrite reductase gene (niiA) in the opportunistic fungal pathogen Aspergillus fumigatus. Cure Genet 33, 3 (Mar 1998), 206-215.
[18] AMITANI, R., TAYLOR, G., ELEZIS, E. N., LLEWELLYN-JONES, C.. MITCHELL, J., Kum, F., COLE, P. J., AND WILSON, R. Purification and characterization of factors produced by Aspergillus fumigatus which affect human ciliated respiratory epithelium. Infect Irntnun
63, 9 (Sep 1995), 3266-3271.

REFERENCES 436
[19] AMOROS, M., AND ESTRUCII, F. Hsflp and Msn2/4p cooperate in the expression of Saccharomyces cerevisiae genes HSP26 and HSP104 in a gene and stress type-dependent manner. Mal Microbiol 39, 6 (2001 Mar), 1523-1532.
[20] ANDERSON. J. B., RICKER, N., AND SIRJUSINGII, C. Antagonism between two mechanisms of antifungal drug resistance. Eukaryot Cell 5,8 (2006 Aug), 1243-1251.
[21] ANDRADE, A. C., DEL SORBO, G., VAN NISTELROOY, J. G., AND WAARD, M. A. The ABC transporter AtrB from Aspergillus nidulans mediates resistance to all major classes of fungicides and some natural toxic compounds. Microbiology 146 ( Pt 8) (Aug 2000),
1987-1997.
[22] ANGELICHI0, M. J., AND CAMILLI, A. In vivo expression technology. Infection and Immunity 70, 12 (2002), 6518-6523.
[23] ANGERMAYR, K., PARSON, W., STOFFLER, G., AND HAAS, H. Expression of atrC - encoding a novel member of the ATP binding cassette transporter family in Aspergillus nidulans - is sensitive to cycloheximide. Biochim Biophys Acta 1453, 2 (Feb 1999), 304-310.
[24] ANNAIX, V., BOUCHARA, J. P., LARCIIER, G., CIIABASSE, D., AND TRONCHIN, G. Specific binding of human fibrinogen fragment D to Aspergsllus furnigatus conidia. Infect Immun 60, 5 (1992 May), 1747-1755.
[25] AOUIDA, M., PAGE, N., LEDUC, A., PETER, M., AND RAMOTAR, D. A genome-wide screen in Saccharomyces cerevisiae reveals altered
transport as a mechanism of resistance to the anticancer drug bleomycin. Cancer Res 64, 3 (Feb 2004), 1102-1109.
[26] AOUIDA, M., TOUNEKTI, 0., LEDUC, A., BELFIADJ, 0., MIR, L., AND RAMOTAR, D. Isolation and characterization of Saccharomyces cerevisiae mutants with enhanced resistance to the anticancer drug bleomycin. Gurr Genet 45, 5 (May 2004), 265-272.
[27] APARICIO, 0., GEISBERC. J. V., SEKINGER, E., YANG, A., MOQTADERI, Z., AND STRUHL, K. Chromatin immunoprecipitation for determining the association of proteins with specific genomic sequences in vivo. Carr Protoc Mel HMI Chapter 21 (2005 Feb), Unit 21.3.
[28] ARENDORE, T. M.. AND WALKER, D. M. Oral candidal populations in health and disease. Br Dent .7 147, 10 (1979 Nov 20), 267-272.
[29] AkeNooRP, T. M., AND WALKER, D. M. The prevalence and intra-oral distribution of Candida albicans in man. Arch Oral Biol 25, 1 (1980), 1-10.
[30] ARKIN, A., Ross, J.. AND MCADAMS, H. H. Stochastic kinetic analysis of developmental pathway bifurcation in phage lambda-infected escherichia coli cells. Genetics 149, 4 (Aug 1998), 1633-1648.
[31] ARMSTRONG-JAMES, D. Systematic analysis of pH adaptation in fungi. PhD thesis, Imperial College, London, England, February
2007.
[32] ARMSTRONG-JAMES, D. P. H., TURNBULL, S. A., TEO, I., STARK, J., ROGERS, N. J., ROGERS, T. R. F., BIGNELL. E., AND HAYNES, K. Impaired interferon-gamma responses, increased interleukin-17 expression, and a tumor necrosis factor-alpha transcriptional program in
invasive aspergillosis. J Infect Dis 200, 8 (2009 Oct 15), 1341-1351.
[33] ARST, H. N., AND PENALVA, M. A. pH regulation in Aspergillus and parallels with higher eukaryotic regulatory systems. Trends Genet 19, 4 (Apr 2003), 224-231.
[34] ARST, H. N. J., AND COVE, D. J. Nitrogen metabolite repression in Aspergillus nidulans. Mol Gen Genet 126, 2 (1973 Nov 2), 111-141.
[35] ASIIBURNER, M., BALL, C. A., BLAKE, J. A., BOTSTEIN, D., BUTLER, H., CHERRY, J. M., DAVIS, A. P., DOLINSKI, K., DWIGHT, S. S., ErriG, J. T., HARRIS, M. A., H0.1,, D. P., ISSEL-TARVER, L., KASARSKIS. A., LEWIS, S., MATESE, J. C., RICHARDSON, J. E., RINGWALD, M., ROBIN, G. M.. AND SHERLOCK, G. Gene ontology: tool for the unification of biology. The Gene Ontology Consortium. Nat Genet 25, 1 (2000 May),
25-29.
[36] ASKEW, D. S. Aspergillus fumigatus: virulence genes in a street-smart mold. Curr Opin Microbial 11, 4 (2008 Aug), 331-337.
[37] ASSMANN, S. M., AND ALBERT, R. Discrete dynamic modeling with asynchronous update, or how to model complex systems in the absence of quantitative information. Methods Mol Biol 553 (2009), 207-225.
[38] ASTIR, A. M., AND HASLAM, J. NI. The manipulation of cellular cytochrome and lipid composition in a haem mutant of Saccharomyces cerevisiae. Biochetri J 166, 2 (1977 Aug 15), 275-285.
[39] AUSUBEL, F. M., BRENT, R., KINGSTON, R. E.. MOORE, D. D., SEIDMAN, J. G., SMITH, J. A., AND STIWIIL, A., Eds. Current Protocols in
Molecular Biology. John Wiley and Sons, 1987.
[40] AYENI, O., RIEDERER, K. NI., WILSON, F. M., AND KHATIB, R. Clinicians' reaction to positive urine culture for Candida organisms.
Mycoses 42,4 (1999), 285-289.
[41] BADER, J. S., CHAUDHURI, A., ROTHBERC, J. M., AND CHANT, J. Gaining confidence in high-throughput protein interaction networks.
Nat Biotechnol 22, 1 (2004 Jan), 78-85.
[42] BARN, Y.-S.. KOJIMA, K., Cox, G. NI., AND HEITMAN, J. Specialization of the HOG pathway and its impact on differentiation and virulence of Cryptococcus neoformans. Mol Biol Cell 16, 5 (2005 May), 2285-2300.

REFERENCES 437
[43] BAILLY, E., AND REED, S. I. Functional characterization of RPN3 uncovers a distinct 195 proteasomal subunit requirement for ubiquitin-dependent proteolysis of cell cycle regulatory proteins in budding yeast. Mol Cell Riot 19, 10 (1999 Oct), 6872-6890.
[44] BALCIUNAS, D., AND RONNE, H. Three subunits of the RNA polymerase II mediator complex are involved in glucose repression. Nucleic Acids Res 23, 21 (1995 Nov 11), 4421-4425.
[45] BALE, M., YANG. C., AND PFALLER, M. A. Evaluation of growth characteristics on blood agar and eosin methylene blue agar for the identification of Candida (Torulopsis) glabrata. Diagnostic Microbiology and Infectious Disease 28 (1997), 65-67.
[46] BALZ1, E., WANG, NI., LETERME, S., VAN DYCK, L., AND GOFFEAU, A. PDR5, a novel yeast multidrug resistance conferring transporter controlled by the transcription regulator PDR1. J Biol Chem 269, 3 (1994 Jan 21), 2206-2214.
[47] BARABASI, A., AND ALBERT, R. Emergence of scaling in random networks. Science 286, 5439 (1999 Oct 15), 509-512.
[48] BARABASI, A.-L. Scale-free networks: a decade and beyond. Science 325, 5939 (2009 Jul 24), 412-413.
[49] BARABASI, A.-L., AND OLTVAI, Z. N. Network biology: understanding the cell's functional organization. Nat Rev Genet 5, 2 (2004 Feb), 101-113.
[50] BARDWELL, L. A walk-through of the yeast mating pheromone response pathway. Peptides 26, 2 (2005 Feb), 339-350.
[51] BARENCO, NI., BREWER, D., PAPOULI, E., TOMESCU, D., CALLARD, R., STARK, J., AND HUBANK, NI. Dissection of a complex transcriptional response using genome-wide transcriptional modelling. Mol Syst Bird 5 (2009), 327.
[52] BARENCO, NI., TOMESCU, D., BREWER, D., CALLARD, R., STARK, J., AND HUBANK, M. Ranked prediction of p53 targets using hidden variable dynamic modeling. Genoine Biol 7,3 (2006), R25.
[53] BARRETT, J. P., VARDULAKI, K. A., CONLON, C.. COOKE, J., DAZA-RAMIREZ, P., EVANS, E. G. V., HAWKEY, P. NI., HERBRECIIT, R., MARKS, D. I., MORALEDA, J. NI., PARK, G. R., SENN, S. J., AND VISCOLI, C. A systematic review of the antifungal effectiveness and tolerability of amphotericin B formulations. Clin Ther 25, 5 (2003 May), 1295-1320.
[54] BARRY, J. D., GINGER, NI. L., BURTON, P., AND MCCULLOCH, R. Why are parasite contingency genes often associated with telomeres? lot J Parasitol 33, 1 (2003 Jan), 29-45.
[55] BAUDIN, A., OZIER-KALOGEROPOULOS, 0., DENOUEL, A., LACROUTE, F., AND CULLIN, C. A simple and efficient method for direct gene deletion in Saccharomyces ccrevistae. Nucleic Acids Res 21, 14 (1993 Jul 11), 3329-3330.
[56] BEAUVAIS, A., MONOD, NI., DEBEAUPUIS, J. P., DIAQUIN, M., KOBAYASHI, H., AND LATGE, J. P. Biochemical and antigenic characterization of a new dipeptidyl-peptidase isolated from Aspergillus fumigatus. S Biol Chem 272, 10 (Mar 1997), 6238-6244.
[57] BEAUVAIS, A., MONOD, NI., WYN1GER, J., DEBEAUPUIS, J. P., GROUZMANN, E.. BRAKCII, N., SVAB, J., HOVANESSLAN, A. G., AND LATGE, J. P. Dipeptidyl-peptidase IV secreted by Asyeryillus fumigatus, a fungus pathogenic to humans. Infect hninun 65, 8 (Aug 1997), 3042-3047.
[58] BECK, T., AND HALL, M. N. The TOR signalling pathway controls nuclear localization of nutrient-regulated transcription factors. Nature 402, 6762 (1999 Dec 9), 689-692.
[59] BEEVER. R. E., AND DEMPSEY, G. P. Function of rodlets on the surface of fungal spores. Nature 272, 5654 (1978 Apr 13), 608-610.
[60] BENNETT, C. B., LEWIS, L. K., KARTIIIKEYAN, G., LOBACIIEV, K. S., JIN, Y. H., STERLING, J. F., SNIPE, J. R., AND RESNICK, NI. A. Genes required for ionizing radiation resistance in yeast. Nat Genet 29, 4 (Dec 2001), 426-434.
[61] BENNETT, R. J., AND JOHNSON, A. D. Mating in Candida albicans and the search for a sexual cycle. Annu Rev M crobiol 59 (2005), 233-255.
[62] BENSEN, E. S., MARTIN, S. J., Li, NI., BERMAN, J., AND DAVIS, D. A. Transcriptional profiling in Cundida albicans reveals new adaptive responses to extracellular pH and functions for Rim101p. Mal Microbial 54, 5 (2004 Dec), 1335-1351.
[63] BERMAN, J., AND SUDBERY, P. E. Candida albicans: a molecular revolution built on lessons from budding yeast. Nat Rev Genet 3, 12 (Dec 2002), 918-930.
[64] BERROUANE, Y. F., HERWALDT, L. A., AND PFALLER, NI. A. Trends in antifungal use and epidemiology of nosocomial yeast infections in a university hospital. Journal of Clinical Microbiology 37 (1999), 531-537.
[65] BIIALLA, U. S., AND IYENGAR, R. Emergent properties of networks of biological signaling pathways. Science 288, 5400 (Jan 1999), 381-387.
[66] BIGNELL, E., NEGRETE-URTASAN, S., CALCAGNO, A. NI., HAYNES, K., JR, H. N. A., AND ROGERS, T. The A.spergillus pH-responsive tran-scription factor PacC regulates virulence. Molecular Microbiology 55, 4 (2005), 1072-1084.

REFERENCES 438
1671 BIRCH, M., DENNING, D. W., AND ROBSON, G. D. Comparison of extracellular phospholipase activities in clinical and environmental Aspergillus furnigatus isolates. Med Mycol 42, 1 (Feb 2004), 81-86.
[68] BIRSE, C. E., IRWIN. N. Y., FONZI, W. A., AND SHEPHERD, P. S. Cloning and characterization of ECE/, a gene expressed in association with cell elongation of the dimorphic pathogen Canida albicans. Infection and Immunity 61 (1993), 3648-3655.
[69] BISSINGER, P. H., AND KUCHLER, K. Molecular cloning and expression of the Saccharomyces cm-cuisine STS1 gene product. a yeast ABC transporter conferring mycotoxin resistance. J Biol Chem 269, 6 (1994 Feb 11), 4180-4186.
[70] BLYTII, W., GRANT, I. W., BLACKADDER, E. S., AND GREENBERG, M. Fungal antigens as a source of sensitization and respiratory disease in scottish maltworkers. Clin Allergy 7,6 (1977 Nov), 549-562.
[71] BODENMILLER, B., CAMPBELL, D., GERRITS, B., LAM, H., JOVANOVIC, NI., PICOTTI, P., SCHLAPBACH, R., AND AEBERSOLD, R. Phosphopep-a database of protein phosphorylation sites in model organisms. Nat Bietechnal 26, 12 (2008 Dec), 1339-1340.
[72] BODENMILLER, B., MALMSTROM, J., GERRITS, B., CAMPBELL, D., LAM, H., SCHMIDT, A., RINNER, 0., MUELLER, L. N., SHANNON, P. T., FEDRIOLI, P. G., PANSE, C., LEE, H.-K., SCHLAPBACII, R., AND AEBERSOLD, R. Phosphopep-a phosphoproteome resource for systems biology research in drosophila kc167 cells. Mel Syst Biol 3 (2007), 139.
[73] BoK, .7. W., BALAJEE, S. A., MARR, K. A., ANDES, D., NIELSEN, K. F., FRISVAD, J. C., AND KELLER, N. P. LaeA, a regulator of morphogenetic fungal virulence factors. Eukaryot Cell 4, 9 (Sep 2005), 1574-1582.
[74] BOK, J. W., CHUNG, D., BALAJEE, S. A., MARR, K. A., ANDES, D., NIELSEN, K. F., FRISVAD, J. C.. KIRBY, K. A., AND KELLER, N. P. GIiZ, a transcriptional regulator of gliotoxin biosynthesis, contributes to Aspergillus fumigatus virulence. Infect Immure 74,12 (Dec 2006), 6761-6768.
[75] BOK, J. W., AND KELLER, N. P. LaeA, a regulator of secondary metabolism in Aspergillus spp. Eukaryot Cell 3, 2 (Apr 2004), 527-535.
[76] BORIC, P.. HOLM, L., AND SANDER, C. The immunoglobulin fold. Structural classification, sequence patterns and common core. J Mel Biel 242, 4 (1994 Sep 30), 309-320.
[77] BORNEMAN, A. R., GIANOULIS, T. A., ZHANG, Z. D., H., ROZOWSKY, J., SERINGIIAUS, M. R., WANG, L. Y., GERSTEIN, M., AND SNYDER, NI. Divergence of transcription factor binding sites across related yeast species. Science 317, 5839 (2007 Aug 10), 815-819.
[78] DOUCHARA, J. P., BOUALI, A., TRONCHIN, G., ROBERT, R., CHABASSE, D., AND SENET, J. M. Binding of fibrinogen to the pathogenic Aspergillus species. J Med Vet Mycol 26, 6 (1988), 327-334.
[79] BOUCHARA, J. P., SANCIIEZ, M., CHEYAILLER, A.. MAROT-LEBLOND, A., LISSITZKY, J. C., TRONCHIN, G., AND CHABASSE, D. Sialic acid-dependent recognition of laminin and fibrinogen by Aspergillus fumigates conidia. Infect Immure 65, 7 (1997 Jul), 2717-2724.
[80] BOVERIS, A., CADENAS, E., AND STOPPANI, A. 0. Role of ubiquinone in the mitochondrial generation of hydrogen peroxide. Biochem J 156, 2 (1976 May 15), 435-444.
[81] BOY-MARCOTTE, E., PERROT, M., BUSSEREAU, F., BOUCIIER[E, H., AND JACQUET, N. Msn2p and Msn4p control a large number of genes induced at the diauxic transition which are repressed by cyclic AMP in Saccharemyces cerevisiae. J Bacterial 180, 5 (Mar 1998), 1044-1052.
[82] BOYER, J., BADIS, C., FAIRHEAD, C., TALLA, E., HANTRAYE, F., FABRE, E., FISCHER, G., HENNEQUIN, C., KOSZUL, R., LAFONTAINE, I., OZIER-KALOGEROPOULOS, 0., RICCHETTI, M., RICHARD, G.-F., THIERRY, A., AND DUJON, B. Large-scale exploration of growth inhibition caused by overexpression of genomic fragments in Saccharomyces cerevisiae. Genorne Biol 5,9 (2004), R72.
[83] BOYLE, E. I., WEND, S., GOLLUB, J., JIN, H., BOTSTEIN, D., CHERRY, J. NI., AND SIIERLOCK, G. GO::TermFinder-open source software for accessing Gene Ontology information and finding significantly enriched Gene Ontology terms associated with a list of genes. Bioinformatics 20, 18 (2004 Dec 12), 3710-3715.
[84] BRACKEN, C. P., WHITELAW, M. L., AND PEET, D. J. The hypoxia-inducible factors: key transcriptional regulators of hypoxic responses. Cell Mot Life Sci 60, 7 (Jul 2003), 1376-1393.
[85] BRANCO, M. R., MARINHO, H. S., CYRNE, L., AND ANTUNES, F. Decrease of H202 plasma membrane permeability during adaptation to H202 in Saccharomyces cerevisiae. J Biol Chem 279, 8 (Feb 2004), 6501-6506.
[86] BRAUN, B. R., AND JOHNSON, A. D. Control of filament formation in Candida albicans by the transcriptional repressor TUP1. Scimulf! 277, 5322 (1997 Jul 4), 105-109.
[87] BRAUN, B. R., AND JOHNSON, A. D. TUP1, CPH1 and EFG1 make independent contributions to filamentation in Candida albicans. Genetics 155, 1 (2000 May), 57-67.
[88] BRAUN, B. R., KADOSII, D., AND JOHNSON, A. D. NRG1, a repressor of filamentous growth in C.albicans, is down-regulated during filament induction. EMBO J 20, 17 (2001 Sep 3), 4753-4761.

REFERENCES 439
[80] BREWSTER, J. L., DE VALOIR, T., DWYER, N. D., WINTER, E., AND DUSTIN, M. C. An osmosensing signal transduction pathway in yeast. Science 259, 5102 (1993 Mar 19), 1760-1763.
[90] BROMLEY, 1. M., AND DONALDSON, K. Binding of Aspergillus fumigatus spores to lung epithelial cells and basement membrane proteins: relevance to the asthmatic lung. Thorax 51, 12 (1996 Dec), 1203-1209.
[91] BROWN, A. J., BERTRAM, G., FELDMANN, P. J., PEGGIE, M. W., AND SWOBODA, R. K. Codon utilisation in the pathogenic yeast, Candida albicans. Nucleic Acids Res 19, 15 (Aug 1991), 4298.
[92] BROWN, A. J., AND DOW, N. A. Regulatory networks controlling Candida albicans morphogenesis. Trends Microbiol 7,8 (1999 Aug), 333-338.
[93] BROWN, J. A., SHERLOCK, G., MYERS, C. L., BURROWS, N. M., DENG, C., Wu, H. I., MCCANN, K. E., TROYANSKA, 0. G., AND BROWN, 3. M. Global analysis of gene function in yeast by quantitative phenotypic profiling. Molecular Systems Biology 2 (2006), 2006.0001 (epub).
[94] BROWN, J. S., AUFAIJVRE-BROWN, A., AND HOLDEN, D. W. Insertional mutagenesis of Aspergillus fumigatus. Mol Gen Genet 259, 3 (Aug 1998), 327-335.
[95] BROWN, M. J., WORTHY, S. A., FLINT, J. D., AND MULLER, N. L. Invasive aspergillosis in the immunocompromised host: utility of computed tomography and bronchoalveolar lavage. Clin Radiol 53, 4 (1998 Apr), 255-257.
[96] BRUN, S., Areal", C., LIMA, 0., FILMON, R., BERGES, T., CHABASSE, D., AND BOUCIIARA, J.-P. Relationships between respiration and susceptibility to azole antifungals in Candida glabratu. Antimicrob Agents Chemother 47, 3 (2003 Mar), 847-853.
[97] BRUN, S., BERGES, T., POUPARD, P., VAUZELLE-MOREAU, C., RENTER, G., CHABASSE, D., AND BOLICHARA, J.-P. Mechanisms of azole resistance in petite mutants of Candida glabrata. Antimicrob Agents Chemother 48, 5 (2004 May), 1788-1796.
[98] BUNDOCK, P., VAN ATTIKUM, H., DEN DULK-RAS, A., AND HOOYKAAS, P. J. J. Insertional mutagenesis in yeasts using T-DNA from Agrobacteriunt tun/chic/ens. Yeast 19, 6 (Apr 2002), 529-536.
[99] HUNGER, J., WESTPHAL, G., MONNICII, A., HINNENDAHL, B., HALLIER, E., AND MULLER, M. Cytotoxicity of occupationally and environmen-tally relevant mycotoxins. Toxicology 202, 3 (Oct 2004), 199-211.
[100] BURKIIOLDER, A. C., AND HARTWELL, L. H. The yeast alpha-factor receptor: structural properties deduced from the sequence of the STE2 gene. Nucleic Acids Res 13, 23 (Dec 1985), 8463-8475.
[101] CACCIAPUOTI, A., LOEBENBERG, D., CORCORAN, E., MENZEL, F. J., Moss, E. L. J., NORRIS, C., MICIIALSKI, M., RAYNOR, K., HALPERN, J., MENDRICK, C., ARNOLD, B., ANTONACCI, B., PARMECIANI, R., YAROSII-TONIAINE, T., MILLER, G. H., AND HARE, R. S. In Intro and to num activities of SCH 56592 (posaconazole), a new triazole antifungal agent, against A.spergillus and Candida. Antirnicrob Agents Chemother 44, 8 (2000 Aug), 2017-2022.
[102] CAIRNEY. J. W. G. Basidiomycete mycelia in forest soils: dimensions, dynamics and roles in nutrient distribution. Mycol Res 109, Pt 1 (Jan 2005), 7-20.
[103] CALCAGNO, A.-M., BIGNELL, E., Rocens, T. R., CANEDO, M., MUIILSCHLECEL, F. A., AND HAYNES, K. Candida glabrata STE20 is involved in maintaining cell wall integrity and adaptation to hypertonic stress, and is required for wild-type levels of virulence. Yeast 21, 7 (2004 May), 557-568.
[104] CALCAGNO, A.-M.. BIGNELL, E.. ROGERS, T. R., JONES, M. D., MUIILSCIILEGEL, F. A., AND HAYNES, K. Candida glabrata STE11 is involved in adaptation to hypertonic stress, maintenance of wild-type levels of filamentation and plays a role in virulence. Med Mycol 43, 4 (2005), 355-364.
[106] CALCAGNO, A.-M., BIGNELL, E., WARN, P., JONES, M. D., DENNING, D. W., MMILSCHLEGEL, F. A.. ROGERS, T. R., AND HAYNES, K. Candida glabrata stel2 is required for wild-type levels of virulence and nitrogen starvation induced filamentation. Mol Microbiol 50, 4 (2003 Nov), 1309-1318.
[106] CALCAGNO-PIZARELLI, A. M. Stel 1, Stel2 and Ste20: important mediators of dimorphism, the response to hypertonic stress, cell wall integrity and virulence in the pathogenic fungus Candida glabrata. PhD thesis, Imperial College, 2004.
[107] CALDERONE, R. A. Candida and candidiasis. American Society for Microbiology Press, 2001.
[108] CALDERONE, R. A., AND WADSWORTH, E. Characterization of mannoproteins from a virulent Candida albicans strain and its derived, avirulent strain. Rev Infect Dis 10 Supp/ 2 (1988 Jul-Aug), S423-7.
[109] CALVO, A. M., WILSON, R. A., BOK, 1. W., AND KELLER, N. P. Relationship between secondary metabolism and fungal development. Micrvbiol Mol Biol Rev 66, 3 (Sep 2002), 447-459.
[110] CANNON, R. D., LAMPING, E., HOLMES, A. R., N111,11, K., BARET, P. V., KENIYA, M. V., TANABE, K., NIEMI, M., DOFFEAU, A., AND MONK. D. C. Efflux-mediated antifungal drug resistance. Clin Microbiol Rev 22, 2 (2009 Apr), 291-321.

REFERENCES 440
[111] CARDENAS, M. E., CUTLER, N. S., LORENZ, M. C., DI COMO, C. J., AND HEITMAN, J. The TOR signaling cascade regulates gene expression in response to nutrients. Genes Dee 13, 24 (Dec 1999), 3271-3279.
[112] CASADEVALL, A., AND PIROFSKI, L.-A. "anti-virulence" genes—further muddling the lexicon? response from arturo casadevall and liise-anne pirofski. Trends Microbiol 11, 9 (2003 Sep), 413-414.
[113] CASADEVALL, A., ROSAS, A. L., AND NOSANCHUK, J. D. Melanin and virulence in Cryptucoccus neoformans. Corr Opm Microbiol 3, 4 (Aug 2000), 354-358.
[114] CASS1LL, D., STUY, A., AND BUCK, R. Emergent properties of food distribution among fire ant larvae. J Theor Biol 195, 3 (Dec 1998), 371-381.
[115] CASSONE, A., DE BERNARD'S, F., TOROSANTUCCI, A., TACCONELLI, E., TUMBARELLO, M., AND CAUDA, R. In 'intro and in men anticanclidal activity of human immunodeficiency virus protease inhibitors. J Infect Dis 180, 2 (Aug 1999), 448-453.
[116] CASTANO, I., KAUR, R., PAN, S., CREDO, R., DE LAS PENAS, A., GITO, N., BIERY, M. C., CRAIG, N. L., AND CORMACK, B. P. Tn7-based genome-wide random insertional mutagenesis of Candida glabrata. Garonne Research 13 (2003), 905-915.
[117] CASTANO, I., PAN, ZUPANCIC, M., HENNEQUIN, C., DIMON, B., AND CORN/ACK, B. P. Telomere length control and transcriptional regulation of subtelomeric adhesins in Candida glabrata. Mol Microbiol 55, 4 (2005 Feb), 1246-1258.
[118] CAL'STON, H. C., REN, B., KOH, S. S., HARBISON, C. T., KANIN, E.. JENNINGS, E. G., LEE, T. I., TRUE, H. L., LANDER, E. S., AND YOUNG, R. A. Remodeling of yeast genome expression in response to environmental changes. Mol Biol Cell 12, 2 (2001 Feb), 323-337.
[119] CELIO, G. J., PADAMSEE, M., DENTINGER, B. T. M., BAUER, R., AND MCLAUGHLIN, D. J. Assembling the fungal tree of life: constructing the structural and biochemical database. Mycologta 98, 6 (2006), 850-859.
[120] CHAMILOS, G., AND KONTOYIANNIS, D. P. Update on antifungal drug resistance mechanisms of Aspergillu.s fumigator. Drug Resist Updat 8, 6 (2005 Dec), 344-358.
[121] CHANG, Q., GR1EST, T. A., HARTER, T. M., AND PETRASII, J. M. Functional studies of aldo-keto reductases in Saccharomyces cerevisiae. Biachint Biophys Acta 1773, 3 (2007 Mar), 321-329.
[122] CIIANG, Q., HARTER, T. M., RIKIMARU, L. T., AND PETRASII, J. M. Aldo-keto reductases as modulators of stress response. Chem Biol Interact 143-144 (2003 Feb 1), 325-332.
[123] CHANG, Y. C., BIEN. C. M., LEE, H., ESPENSHADE, P. J., AND KWON-CIIUNG, K. J. Srelp, a regulator of oxygen sensing and sterol homeostasis, is required for virulence in Cryptococcus neoformans. Mal Microbial 64, 3 (May 2007), 614-629.
[124] CIIARLIER, C., HART, E., LEFORT, A., HIRAM), P., DROMER, F., DENNING, D. W., AND LORTIIOLARY, 0. Fluconazole for the management of invasive candidiasisi where do we stand after 15 years? J Antinticrob Chernother 57,3 (2006 Mar), 384-410.
[125] CHAVES, G. M., BATES, S., MACCALLUM, D. M., AND ODDS, F. C. Candida albicans GRX2, encoding a putative glutaredoxin, is required for virulence in a murine model. Genet Mol Res 6, 4 (2007), 1051-1063.
[126] CIIEETIIAM, J., Sloan, D. A., DA SILVA DANTAS, A., DORIS, K. S., PATTERSON, M. J., BRUCE, C. R., AND QUINN. .T. A single MAPKKK regulates the Hogl MAPK pathway in the pathogenic fungus Candida albican.s. Mol Dial Cell 18, 11 (Nov 2007), 4603-4614.
[127] CHELSTOWSKA, A., AND BUTOW, R. A. RTG genes in yeast that function in communication between mitochondria and the nucleus are also required for expression of genes encoding peroxisomal proteins. J Biol Chern 270, 30 (1995 Jul 28), 18141-18146.
[128] CHEN, K. C., CALZONE, L., CSIKASZ-NACY, A., CROSS, F. R., NOVAK, B., AND TYSON, J. J. Integrative analysis of cell cycle control in budding yeast. Mol Biol Cell 15, 8 (2004 Aug), 3841-3862.
[129] CHEN, M. H., SHEN, Z. M., BOBIN, S., KAHN, P. C., AND LIPKE, P. N. Structure of Saccharomyces cerevisiae alpha-agglutinin. Evidence for a yeast cell wall protein with multiple immunoglobulin-like domains with atypical disulfides. J Blot Chem 270,44 (1995 Nov 3), 26168-26177.
[130] CIIEN, Y., ZIIU, J., LUM, P. Y., YANG, X., PINTO, S., MACNEIL, D. J., ZHANG, C., LAMB, J., EDWARDS, S., SIEBERTS, S. K., LEONARDSON, A., CASTELLINI, L. W., WANG, S., CHAMPY, M.-F., ZIIANG, B., EMILSSON, V., Doss, S., GIIAZALPOUR, A., HORVATH, S., DRAKE, T. A., LUSIS, A. J., AND SCHADT, E. E. Variations in DNA elucidate molecular networks that cause disease. Nature 452, 7186 (2008 Mar 27), 429-435.
[131] CHERRY, M., ADLER, C., BALL, C., CHERVITZ, S. A., DWIGHT, S. S., HESTER, E. T., .11A, Y., JUVIK, G., ROE, T., SCHROEDER, M., WENG, S., AND BOTSTE1N, D. Saccharornyces genome database. Nucleic Acids Research 26 (1998), 73-80.
[132] CHIN, C. S., Ho. P. L., AND YUEN, K. Y. Simultaneous Aspergillos ,fischeri and Herpes simplex pneumonia in a patient with multiple myeloma. Scand J Infect Dis 30, 2 (1998), 190-191.
[133] CHO, T., Sunon, M., TANAKA, T., NAKAMISIIA, Y., CHIBANA, H., AND KAMINISIII, H. Isolation and expression of a gene (CG/7/) regulated during the yeast-hyphal transformation in Candida albtcans. Biochimica et Biophysica Ada 1517 (2001), 288-292.

REFERENCES 441
[134] CHRISTIE, K. R., WENG, S., BALAKRISIINAN, R., COSTANZO, M. C., DOLINSKI, K., DWIGHT, S. S., ENGEL, S. R., FEIERBACH, B., FISK. D. G., HIRSCIIMANN, J. E., HONG, E. L., ISSEL-TARVER, L., NASH, R., SETIIURAMAN, A., STARR, B., THEESFELD, C. L., ANDRADA, R., BINKLEY, G., DONG, Q., LANE, C., SCHROEDER, M., BOTSTEIN, D., AND CHERRY, J. M. Saccharoniyees genome database SOD provides tools to identify and analyze sequences from Saccharornyees and related sequences from other organisms. Nucleic Acids Research 32 (2004), D311—D314.
[135] G., TARICANI, L., STANGLE, W., AND YOUNG, P. G. Insertional mutagenesis based on illegitimate recombination in Schszosaccha- romyces pornbe. Nucleic Acids Res 28, 11 (Jun 2000), E53.
[136] CLARK, K. L., FELDNIANN, P. J., DIGNARD, D., LAROCQUE, R., BROWN, A. J., LEE, M. G., THOMAS, D. Y., AND WHITEWAY, M. Constitutive activation of the Saccharomyces ce.rev siae mating response pathway by a MAP kinase kinase from Candida albicans. Mot Gen Genet 249, 6 (1995 Dec 20), 609-621.
[137] CLINE, M. S., SMOOT, M., CERAMI, E., KUCHINSKY, A., LANDYS. N., WORKMAN, C., CHRISTMAS, R., AVILA-CARIPILO, I., CREECII, M., GROSS, B., HAMPERS, K., ISSERLIN, R., KELLEY, R., KILLCOYNE, S., LOTIA, S., MAERE, S., MORRIS, J., ONO, K., PAVLOVIC, V., PI00, A. R., VAILAYA, A., WANG, P.-L., ADLER, A., CONKLIN, B. R., HOOD, L., KUIPER, M., SANDER, C., SCIIMULEVICII, I., SCHWIKOWSKI, B., WARNER, G. J., IDEKER, T., AND BADER, G. D. Integration of biological networks and gene expression data using cytoscape. Nat Protoc 2,10 (2007), 2366-2382.
[13S] CLINICAL AND LABORATORY STANDARDS INSTITUTE. Reference Method for broth dilution antifungal susceptibility testing of yeasts; approved standard - Third Edition, 3 ed. Clinical and Laboratory Standard Institute, 940 West Valley Road, Suite 1400, Wayne, Pennsylvania 19087-1898,2008.
[139] CORMACK, B. P., AND FALKOW, S. Efficient homologous and illegitimate recombination in the opportunistic yeast pathogen Candida glabrata. Genetics 151 (1999), 979-987.
[140] CORMACK, B. P., GIIORI, N., AND FALKOW, S. An adhesin of the yeast pathogen Candida glabrata mediating adherence to human epithelial cells. Science 258 (1999), 578-581.
[141] CORMACK, B. P., GHORI, N., AND FALKOW, S. An adhesin of the yeast pathogen Candida glabrata mediating adherence to human epithelial cells. Science 285, 5427 (1999 Jul 23), 578-582.
[142] COSTANZO, M., BARYSIINIKOVA, A., BELLAY, J., KIM, Y., SPEAR, E. D., SEVIER, C. S., DING, H., Koti, J. L. Y., TOUFIGIII, K., MOSTAFAVI, S., PRINZ, J., STONGE, R. P., VANDERSLUIS, B., ALIZADEII, S., BAHR, S., BROST, R. L., CIIEN, Y., COKOL, M., DESIIPANDE, II., LI, Z., LI, Z.-Y., LIANG, W., MARBACK, M., PAW, J., SAN LUIS, ft-J., SIIUTERIQI, E., TONG, A. H. Y., VAN DYK, N., WALLACE, I. M., WHITNEY, J. A., WEIRAUCH, M. T., ZIIONG, G., ZIIU, H., HOURY, W. A., BRUDNO, M.. RAGIBIZADEll, S., PAPP, B., PAL, C., ROTH, F. P., GIAEVER, G., NISLOW, C., TROYANSKAYA, 0. G., TYERS, M., BUSSEY, H., BADER, G. D., GINGRAS, A.-C., MORRIS, Q. D., KIM, P. M., KAISER, C. A., MYERS, C. L., ANDREWS, B. J., AND BOONE, C. The genetic landscape of a cell. Received as personal communication., November 2009.
[143] COSTENOBLE. R., VALADI, H., GUSTAFSSON, L., NIKLASSON, C., AND FRANZEN, C. J. Microaerobic glycerol formation in Saccharomyees cerevisiae. Yeast 16, 16 (2000 Dec), 1483-1495,
[144] COVE, D. J. The induction and repression of nitrate reductase in the fungus Aspergillus nidufans. Biochim Biophys Acta 113, 1 (Jan 1966), 51-56.
[145] CRAMER, R. A. J., GAMCSIK, M. P., BROOKING, R. M., NAJVAR, L. K., KIRKPATRICK, W. R., PATTERSON, T. F., BALIBAR, C. J., GRAYBILL, J. R., PERFECT, J. R., ABRAHAM, S. N., AND STEINBACH, W. J. Disruption of a nonribosomal peptide synthetase in Aspergillus furnigatus eliminates gliotoxin production. Eukaryot Cell 5, 6 (Jun 2006), 972-980.
[146] CRAMER, R. A. J., GAMCSIK, M. P., BROOKING. R. M., NAJVAR, L. K., KIRKPATRICK, W. R., PATTERSON, T. F., BALIBAR, C. J., GRAYBILL, J. R., PERFECT, J. R., ABRAHAM, S. N., AND STEINBACH, W. J. Disruption of a nonribosomal peptide synthetase in Aspergillus furnigatus eliminates gliotoxin production. Eukaryot Cell 5, 6 (2006 Jun), 972-980.
[147] CREEVEY, C. J., AND MCINERNEY, J. 0. An algorithm for detecting directional and non-directional positive selection, neutrality and negative selection in protein coding dna sequences. Gene 300, 1-2 (Oct 2002), 43-51.
[148] CREEVEY, C. J., AND MCINERNEY, J. 0. Crann: detecting adaptive evolution in protein-coding dna sequences. Bioinforrnatics 19, 13 (Sep 2003), 1726.
[149] CRISP, R. J., ADKINS, E. M., KIMMEL, E., AND KAPLAN, J. Recruitment of Tuplp and Cti6p regulates heme-deficient expression of Aftlp target genes. EMBO J 25, 3 (2006 Feb 8), 512-521.
[150] CSANK, C., AND HAYNES, K. Candida glabrata displays pseudohyphal growth. FEMS Microbiology Letters 189 (2000), 115-120.
[151] CSIKASZ-NAGY, A. Computational systems biology of the cell cycle. B of Bioinforrn 10, 4 (2009 Jul), 424-434.
[1521 CULLEN, P. J., SABRAGII, W. J., GRAHAM, E., TRICK, M. M., VAN OLDEN, E. K., NEAL, C., DELROW, J., BARDWELL, L. , AND SPRAGUE, G. F. J. A signaling mucin at the head of the Cdc42- and MAPK-dependent filamentous growth pathway in yeast. Genes Dev 18, 14 (2004
Jul 15), 1695-1708.
[153] CULLEN, P.1., SABRAGII, W..T., GRAHAM, E., TRICK, M. M., VAN OLDEN, E. K., NEAL, C., DELROW, J., BARDWELL, L. , AND SPRAGUE, G. F. J. A signaling mucin at the head of the Cdc42- and MAPK-dependent filamentous growth pathway in yeast. Genes Do, 18, 14 (2004 Jul 15), 1695-1708.

REFERENCES 442
[1541 DA SILVA FERREIRA, M. E., CAPELLARO, J. L., DOS REIS MARQUES, E., MALAVAZI, I., PERLIN, D., PARK, S., ANDERSON, J. B., COLOMBO, A. L., ARTHINGTON-SKAGGS, B. A., GOLDMAN, M. H. S., AND GOLDMAN, G. H. In vitro evolution of itraconazole resistance in AspergtIlus furnigatus involves multiple mechanisms of resistance. Antirnicrob Agents Chernother 48, 11 (Nov 2004), 4405-4413.
[155] DAIC110, K., MARUYAMA, H., SUZUKI, A., UENO, M., URITANI, M., AND IJSIIIMARU, T. The ergosterol biosynthesis inhibitor zaragozic acid promotes vacuolar degradation of the tryptophan permease Tat2p in yeast. Biochint Btophys Acta 1768, 7 (2007 Jul), 1681-1690.
[156] DASBACII, E. J., DAVIES, G. M., AND TEUTSCH, S. NI. Burden of aspergillosis-related hospitalizations in the United States. Clin Infect Dis 31, 6 (2000 Dec), 1524-1528.
[157] awls, D. Adaptation to environmental pH in Candida uthicarts and its relation to pathogenesis. Curr Genet 44, 1 (Oct 2003), 1-7.
[158] DE BERNARDIS, F., SULLIVAN, P. A., AND CASSONE, A. Aspartyl proteinases of Candida alhicans and their role in pathogenicity. Med Mycol 39, 4 (Aug 2001), 303-313.
[159] DE JESUS FERREIRA, M. C., BAO, X., LAIZE, V., AND HOIIMANN, S. Transposon mutagenesis reveals novel loci affecting tolerance to salt stress and growth at low temperature. Cure Genet 40, 1 (Aug 2001), 27-39.
[160] DE KRULIFF, B., AND DEMEL, R. A. Polyene antibiotic-sterol interactions in membranes of Acholeplasma laidlawii cells and lecithin liposomes. 3. molecular structure of the polyene antibiotic-cholesterol complexes. Biochirin Biophys Acta 339, 1 (1974 Feb 26), 57-70.
[1611 DE NOBEL, J. G., Kus, F. M., PRIEM, J., MUNNIK, T., AND VAN DEN ENDE, H. The glucanase-soluble mannoproteins limit cell wall porosity in Saccharomyces cerevisiae. Yeast 6, 6 (1990 Nov-Dec), 491-499.
[162] DE SILVA, E., THORNE, T., INGRAM, P., ACRAFIOTI, I., SWIRE, J., WIUF, C., AND STUMPF, M. P. H. The effects of incomplete protein interaction data on structural and evolutionary inferences. BMC Biol 4 (2006), 39.
[1631 DEFONTAINE, A., BOUCHARA, J. P., DECLERK, P., PLANCIIENAULT, C., CIIABASSE, D., AND HALLET, J. N. In vitro resistance to azoles associated with mitochondria] DNA deficiency in Candida ylabrata. J Med Microbiol 48, 7 (1999 Jul), 663-670.
[164] DEHART, D. J., AGWU, D. E., JULIAN, N. C., AND WASIIIIURN, R. G. Binding and germination of Aspergillus fumigatus conidia on cultured A549 pneumocytes. J Infect Dts 175, 1 (1997 Jan), 146-150.
[165] DEL SORBO, G., ANDRADE, A. C., VAN NISTELROOY, J. G., VAN KAN, J. A., BALZI, E., AND DE WAARD, M. A. Multidrug resistance in Aspergillus nululans involves novel ATP-binding cassette transporters. Mol Gen Genet 254, 4 (Apr 1997), 417-426.
[1661 D'ENFERT, C. Selection of multiple disruption events in Aspergillus furnigatus using the orotidine-5'-decarboxylase gene, pyrG, as a unique transformation marker. Carr Genet 30, 1 (Jun 1996), 76-82.
[167] Dimie, S. Z.. VASIC, B., CHARALAMBOUS, C. D., AND PALANIVELU, R. Robust control of uncertain context-sensitive probabilistic boolean networks. IET Syst Biol 3, 4 (2009 Jul), 279-295.
[1681 DENNING, D. W. Invasive aspergillosis. Clin Infect Dis 26, 4 (1998 Apr), 781-803.
[169] DENNING, D. W., FOLLANSBEE, S. E., SCOLARO, M., NORRIS, S., EDELSTEIN, H., AND STEVENS, D. A. Pulmonary aspergillosis in the acquired immunodeficiency syndrome. N Engl J Med 324, 10 (1991 Mar 7), 654-662.
[170] DENNING, D. W., WARD, P. N., FENELON, L. E., AND BENBOW, E. W. Lack of vessel wall elastolysis in human invasive pulmonary aspergillosis. Infect Intraun 60, 12 (Dec 1992), 5153-5156.
[171] DENNISON, P. M. J., RANISDALE, M., MANSON, C. L., AND BROWN, A. J. P. Gene disruption in Candida a/le/cans using a synthetic, codon-optirnised Cre-loxP system. Fungal Genet Riot 42, 9 (Sep 2005), 737-748.
[172] DEFROST, D., YAO, L., SORMANI, R., MOREAU, M., LETERREUX, G., NICOLAI, M., BEDU, M., RORAGLIA, C., AND MEYER, C. The Arabidousts TOR kinase links plant growth, yield, stress resistance and mRNA translation. EMDO Rep 8, 9 (Sep 2007), 864-870.
[173] DEUTSCHBAUER, A. M., WILLIAMS, R. M., CIID, A. hi., AND DAVIS, R. W. Parallel phenotypic analysis of sporulation and postgermination growth in Saccharornyces cerevisiae. Proceedings of the National Academy of Science USA 99 (2002), 155300-15535.
[174] DEVORE-CARTER, D., KAR, S., VELLUCCI, V., BHATTACHERJEE, V., DOMANSKI, P., AND HOSTETTER, M. K. Superantigen-like effects of a Candida albicans polypeptide. J Infect Dis 197, 7 (2008 Apr 1), 981-989.
[175] DI FIORE, P. P., PoLo, S., AND HOFMANN, K. When ubiquitin meets ubiquitin receptors: a signalling connection. Nat Rev Mol Cell Biol 4, 6 (2003 Jun), 491-497.
[176] DM, L., HAYEK, P., SADEK, H., BEYROUTHY, B., AND KIIALAF, R. A. The Candida alincans Ddr48 protein is essential for filamentation, stress response, and confers partial antifungal drug resistance. Med Set Monit 14, 6 (2008 Jun), BR113-121.
[177] DIXON, D. M., POLAK, A., AND SZANISZLO, P. J. Pathogenicity and virulence of wild-type and melanin-deficient Wangiclla derrnatitidis. J Med Vet Mycol 25, 2 (Apr 1987), 97-106.

REFERENCES 443
[178] DIXON, S. J., FEDYSIIYN, Y., Knit, .1. L. Y., PRASAD, T. S. K., CHAHIVAN, C., CEIBA, G., TOUPIGHI, K., BARYSIINIKOVA, A., HAYLES, J., HOE, K.-L., KIM, D.-U., PARK, H.-0., MYERS, C. L., PANDEY, A., DUROCIIER, D,, ANDREWS, B. J., AND BOONE, C. Significant conservation of synthetic lethal genetic interaction networks between distantly related eukaryotes. Proc Nati Aced Set U S A 105, 43 (2008 Oct
28), 16653-16658.
[179] DOERING, T. L., NOSANCHUK, J. D., ROBERTS, W. K., AND CASADEVALL, A. Melanin as a potential cryptococcal defence against microbi-cidal proteins. Med Mycol 37, 3 (Jun 1999), 175-181.
[180] DOMERGUE, R., CASTANO, I., DE LAS PENAS, A., ZUI'ANCIC, M., LOCKATELL, V., HEBEL, J. R., .TOIINSON, D., AND CORMACK, B. P. Nicotinic acid limitation regulates silencing of Candida adhesins during UTI. Science 308, 5723 (May 2005), 866-870.
[181] DONOFRIO, N. M., On, Y., LUNDY, R., PAN, H., BROWN, D. E., JEONG, J. S., COUCIILAN, S., MITCHELL, T. K., AND DEAN, R. A. Global gene expression during nitrogen starvation in the rice blast fungus, Magnaporthe grisea. Fungal Genet Biol 43, 9 (Sep 2006), 605-617.
[182] DUJON, B., SIIERMAN, D., FISCIIER, G., DURRENS, P., CASAREGOLA, S., LAFONTAINE, I., DE MONTIGNY, J., MARCK, C., NEUVEGLISE, C., TALLA, E., GOFFARD, N., FRANCIEUL, L., AiGi.c, M., ANTHOUARD, V., BABOUR, A., BARBE, V., BARNAY, S., BLANCIIIN, S., BECKERICII, J.-M., BEYNE, E., BLEYKASTEN, C., BOISRAME, A., BOYER, J., CATTOLICO, L., CONFANIOLERI, F., DE DARUVAR, A., DESPONS. L., FABRE, E., FAIRHEAD, C., FERRY-DUMAZET, H., GROPPI, A., HANTRAYE, F., HENNEQUIN, C., JAUNIAUX, N., JOYET, P., KACHOURI, R., KERREST, A., KoszuL, R., LEMAIRE, M., LESTER, I., MA, L., MULLER, H., NICAUD, J.-M., NIKOLSKI, M., OZTAS, S., OZIER-KALOGEROPOULOS, O., PELLENZ, S., POTIER, S., RICHARD, G.-F., STRAUB, M.-L., SULEAU, A., SWENNEN, D., TEKA1A, F., WESOLOVVSKI-LOUVEL, M., WESTHOF, E., WIRTH, B., ZENIOU-MEYER, M., ZIVANOVIC, I., BOLOTIN-FUIEDHARA, M., THIERRY, A., BOUCIIIER, C., CAILDRON, B., SCARPELLI, C., GAILLARDIN, C., WE1SSENBACH, J., W1NCKER, P., AND SOUCIET, .1.-L. Genome evolution in yeasts. Nature 4.90,6995 (2004), 35-44.
[183] EBERWINE. J. Amplification of mRNA populations using aRNA generated from immobilized oligo(dT)-T7 primed cDNA. Btotech-nigues 20, 4 (1996 Apr), 584-591,
[184] EBINA, K., SAKAGAMI, H., YOKOTA, K., AND KoNno, R. Cloning and nucleotide sequence of cDNA encoding Asp-hemolysin from Aspergillos fumigates. Bionirim Biophys Acta 1219, 1 (Sep 1994), 148-150.
[185] EJZYKOWICZ, D. E., CUNIIA, M. M., ROZENTAL, S., SOLIS, N. V., GRAVELAT, F. N., Sin:Pram), D. C., AND FILLER, S. G. The Aspergillus fumigates transcription factor Ace2 governs pigment production, conidiation and virulence. Mel Microbiol 72, 1 (Apr 2009), 155-169.
[186] EL-NESHAWY, S., AND SIIETAIA, Y. M. H. Biocontrol capability of Candida spp. against Botrytis rot of strawberries with respect to fruit quality. Acta Horticutturae (ISHS) 604 (2003), 727-733.
[187] ELLIS, If. E., CLINK, H., ERNST, P., Haunt, M. A., PADMOS, A., SPENCE, D., KALIN, M., HUSSAIN QADRI, S. M., BURNIE, J., AND GREER, W. Controlled study of fiuconazole in the prevention of fungal infections in neutropenic patients with haematological malignancies and bone marrow transplant recipients. Eur .1 Clm Mierobiol Infect Dis 13, 1 (1994 Jan), 3-11.
[188] EMERY, H. S., SHELBURNE. C. P., BOWMAN, J. P., FALLON, P. G., SCHULZ, C. A., AND JACOBSON, E. S. Genetic study of oxygen resistance and melanization in Cryptococcus neoformans. Infect Immun 62, 12 (Dec 1994), 5694-5697.
[189] ENACHE-ANGOULVANT, A., AND HENNEQUIN, C. Invasive Saccharomyces infection: a comprehensive review. Min. Infect Dis 41, 11 (2005 Dec 1), 1559-1568.
[190] ENOEBRECIIT, J. Cell signaling in yeast sporulation. Biochem Biophys Res Common 306, 2 (2003 Jun 27), 325-328.
[191] ENJALBERT, B., NANTEL, A., AND WHITEWAY, M. Stress-induced gene expression in Candida a/bieares: Absence of a general stress response. Mel Riot Cell 14, 4 (2003 Apr), 1460-1467.
[192] ENJALBERT, B., SMITH, D. A., CORNELL, M. J., ALAM, I., NICIIOLLS, S., BROWN, A. J. P., AND QUINN, J. Role of the Hogl stress-activated protein kinase in the global transcriptional response. Mal HMI Cell 17, 2 (2006 Feb), 1018-1032.
[193] Ertn'os, P., AND RENYI, A. Random graphs. Academic Press, London, England, 1960.
[194] ESPINEL-INGROPF, A., MCGINNIS, M. R., PINCUS, D. H., GOLDSON, P. R., AND KERKERING, T. M. Evaluation of the API 20C yeast identifi-cation system for the differentiation of some dematiaceous fungi. J Clin Microbiol 27, 11 (1989 Nov), 2565-2569.
[195] FALKOW, S. Molecular Koch's postulates applied to microbial pathogenicity. Rev Infect Dis 10 Suppl 2 (Jul-Aug 1988), S274-6.
[196] FARMAN, M. L. Telomeres in the rice blast fungus Magnaporthe oryzac: the world of the end as we know it. FEMS Microbiol Lett
273, 2 (2007 Aug), 125-132.
[197] FEDOROVA, N. D., KIIALDI, N., .TOARDAR, V. S., MAIM R., AMEDEO, P., ANDERSON, M. J., CRABTREE, J., SILVA, J. C., BADGER, J. H., ALRARRAQ, A., ANGIUOLI, S., HUSSEY, H., BOWYER, P., COTTY, P. J., DYER, P. S., EGAN, A., GALENS, K., FRASER-LIGGETT, C. M., HAAS, B. J., INMAN, J. M., KENT, R., LEMIEUX, S., MALAVAZI, I., ORVIS, J., ROEMER, T., RONNING, C. M., SUNDARAM, J. P., SUTTON, G., TURNER, G., VENTER, J. C., WHITE, 0. R., WHITTY, B. R., YOUNGMAN, P., WOLFS. K. H., GOLDMAN, G. H., WORTMAN, J. R., JIANG, B., DENNING, D. W., AND MERMAN,
W. C. Genomic islands in the pathogenic filamentous fungus Aspergillus fumigates. PLoS Genet 4, 4 (Apr 2008), e1000046.
[198] FELSENSTEIN, J. Phylogenies from molecular sequences: inference and reliability. Annu Rev Genet 22 (1988), 521-565.

REFERENCES 444
11991 FERA, M. T., LA CAMERA, E., AND DE SARRO, A. New triazoles and echinocandins: mode of action, in vitro activity and mechanisms of resistance. Expert Rev Anti Infect Ther 7,8 (Oct 2009), 981-998.
[200] FERNANDES, L., RODRIGUES-POUSADA, C., AND STRUHL, K. Yap, a novel family of eight bZIP proteins in Saccharomyces cerevisiac with distinct biological functions. Mod Cell Bud 17, 12 (1997 Dec), 6982-6993.
12011 FERRIGNO, P., POSAS, F., KOEPP, D., SAITO, H., AND SILVER, P. A. Regulated nucleo/cytoplasmic exchange of HOG1 MAPK requires the importin beta homologs NMD5 and XPO1. EMBO J 17, 19 (Oct 1998), 5606-5614.
[202] FIDEL, P. L. J. Immunity to Candida. Oral Dis 8 Suppl 2 (2002), 69-75.
[203] FINLAY, B. B., AND FALKOW, S. Common themes in microbial pathogenicity. Microbial Rev 53, 2 (Jun 1989), 210-230.
[204] FINLAY, B. B., AND FALKOW, S. Common themes in microbial pathogenicity revisited. Microbial Mal Biol Rev 61, 2 (Jun 1997), 136-169.
[205] FITZPATRICK, D. A., LOGUE, M. E., STARCH, J. E., AND BUTLER, G. A fungal phylogeny based on 42 complete genomes derived from supertree and combined gene analysis. LOW Evol Biol 6 (2006), 99.
[206] FLANAGAN, P. M., KELLEHER. R. J. R., SAYRE, M. H., TSCIIOCIINER, H., AND KORNBERG, R. D. A mediator required for activation of RNA polymerase II transcription in vitro. Nature 350, 6317 (1991 Apr 4), 436-438.
[207] FLING, NI. E., KOPF, J., TAMARKIN, A., GORMAN, J. A., SMITH, IL A., AND KOLTIN, Y. Analysis of a Candida albicans gene that encodes a novel mechanism for resistance to benomyl and methotrexate. Molecular and General Genetics 227 (1991), 318-329.
[208) FONZI, W. A., AND IRWIN, M. Y. Isogenic strain construction and gene mapping in Candida albicans. Genetics 134, 3 (1993 Jul), 717-728.
[209] FORTUNATO, S. Community detection in graphs.
[210] FRADIN, C., KRETSCIIMAR, M., NICIITERLEIN, T., GAILLARDIN, C., D'ENFERT, C., AND HURE, D. Stage-specific gene expression of Candida albicans in human blood. Mod Microbiol 47, 6 (Mar 2003), 1523-1543.
[211] FRANKE, K., NGUYEN, M., HARTL, A., DAIISE, H.-M., VOGL, G., WURZNER, R., ZIPFEL, P. F., KUNKEL, W., AND ECK, R. The vesicle transport protein Vaclp is required for virulence of Candida albicans. Microbiology 152, Pt 10 (2006 Oct), 3111-3121.
[212] FRASER, V. J., JONES, M., DUNKEL, J., STORFER, S., MEDOFF, G., AND DUNAGAN, W. C. Candidemia in a tertiary care hospital: edpidemi-ology, risk factors, and predictors of mortality. Clinical Infectious Diseases 15, 3 (1992), 414-421.
[213] FRICKER, M. D., LEE, J. A., BERBER, D. P., TLALKA, M., HYNES, J., DARRAII, P. R., WATKINSON, S. C., AND BODDY, L. Imaging complex nutrient dynamics in mycelia] networks. J Microsc 231, 2 (2008 Aug), 317-331.
[214] FRIDKIN, S. K., AND JARVIS, W. R. Epidemiology of nosocomial fungal infections. Clinical Microbiology Reviews 9, 4 (1996), 499-511.
[215] FRISVAD, J. C., ANDERSEN, B., AND TIIRANE, U. The use of secondary metabolite profiling in chemotaxonomy of filamentous fungi. Mycol Res 112, Pt 2 (2008 Feb), 231-240.
[216] FROIINER, I. E., BOURGEOIS, C., YATSYK, K., MAJER, 0., AND KUCIILER, K. Candida albicans cell surface superoxide dismutases degrade host-derived reactive oxygen species to escape innate immune surveillance. Mol Microbial 71, 1 (Jan 2009), 240-252.
[217] FROSCO, M., CHASE, T. J., AND MACMILLAN, J. D. Purification and properties of the elastase from Aspergillus furnigatus. Infect /rnmun 60, 3 (Mar 1992), 728-734.
[218] Frtosco, M., FARED. C., CHASE, T. .T., AND MACMILLAN, J. D. Inhibition of Aspergillus furnigatus elastase with monoclonal antibodies produced by using denatured elastase as an immunogen. Infect /7117re tat 60, 3 (Mar 1992), 735-741.
[219] FROSCO, M. B., CHASE, T. J., AND MACMILLAN, J. D. The effect of elastase-specific monoclonal and polyclonal antibodies on the virulence of Aspergillus fumigatus in immunocompromised mice. Mycopathologia 125, 2 (Feb 1994), 65-76.
[220] Fu, Y., BIEG, G., FONZ1, W. A., BELANGER, P. H., EDWARDS, J. E. J., AND FILLER, S. G. Expression of the Candida albicans gene ALS1 in Saccharomyces cerevisiae induces adherence to endothelial and epithelial cells. Infect Intinun 66, 4 (1998 Apr), 1783-1786.
[221] FUKIICIII. Y., KUDO. Y., KUMAGAI, T., EBINA, K., AND YOKOTA, K. Oxidized low density lipoprotein inhibits the hemolytic activity of Asp-hemolysin from Aspergillus furnigatus. FEMS Microbial Lett 167, 2 (Oct 1998), 275-280.
[222] FURUCIII, T., ISIIIKAWA, H., MICRA, N., ISIIIZUKA, M., KAJIYA, K., KUCE, S., AND NAGANUMA, A. Two nuclear proteins, Cin5 and Ydr259c, confer resistance to cisplatin in Saccharomyces cerevisuic. Mol Pharrnacal 59, 3 (2001 Mar), 470-474.

REFERENCES 445
[223] GALAGAN, J. E., CALVO, S. E., CUOMO, C., MA, L.-J., WORTMAN, I BATZOGLOU, S., LEE, S.-1, BASTURKMEN, M., SPEVAK, C. C., CLUTTER- BUCK, J., KAPITONOV, V., JURKA, J., SCAZZOCCI110, C., FARMAN, M., BUTLER, J., PURCELL, S., HARRIS, S., BRAL'S, G. H., DRAHT, 0., BUSCH, S., D'ENFERT, C., BOUCIIIER, C., GOLDMAN, G. H., BELL-PEDERSEN, D., GRIFFITHS-JONES, S., DOONAN, J. H., Yu, J., VIENKEN, K., PAIN, A., FREITAG, M., SELKER, E. U., ARCHER, D. B., PENALVA, M. A., OAKLEY, B. R., MOMANY, M., TANAKA, T., KUMAGAI, T., ASAI, K., MACHIDA, M., NIERMAN, W. C., DENNING. D. W., CADDICK, M., HYNES, M., PAOLETTI, M., FISCHER, R., MILLER, B., DYER, P., SACIIS, M. S., OSMANI, S. A., AND BIRREN, B. W. Sequencing of Aspergillus nidulans and comparative analysis with A. fumigatus and A. oryzac. Nature 438, 7071 (2005 Dec 22), 1105-1115.
[224] GALE, C. A., BENDEL, C. M., MCCLELLAN, M., HAUSER, M., BECKER, J. M., BERMAN, J., AND HOSTETTER, M. K. Linkage of adhesion, filamentous growth, and virulence in Candida albicans to a single gene, INT1. Science 279, 5355 (1998 Feb 27), 1355-1358.
[225] GAME, J. C., BIRRELL, G. W., BROWN, J. A., SIIIRATA, T., BACCARI, C., CHU, A. M., WILLIAMSON. M. S., AND BROWN, J. M. Use of a genome-wide approach to identify new genes that control resistance of Saccharomyces cerevisiae to ionizing radiation. Radiat Res 160, 1 (Jul 2003), 14-24.
[226] GARCIA-GIMENO. M. A., AND STRUM., K. Acal and Aca2, ATF/CREB activators in Saccharomyces cerevisiae, are important for carbon source utilization but not the response to stress. Mid Cell Bud 20, 12 (2000 Jun), 4340-4349.
[227] GARREAU, II., HASAN, R. N., RENAULT, G., ESTRUCH, F., BOY-MARCOTTE, E.. AND JACQUET, M. Hyperphosphorylation of Msn2p and Msn4p in response to heat shock and the diauxic shift is inhibited by cAMP in Saccharomyces cerevisiae. Microbiology 146 ( Pt 9) (2000 Sep), 2113-2120.
[228] GARREAU, H., HASAN, R. N., RENAULT, G., ESTRUCH, F., BOY-MARCOTTE, E., AND JACQUET, M. Hyperphosphorylation of Msn2p and Msn4p in response to heat shock and the diauxic shift is inhibited by cAMP in Saccharomyces rerevisiae. Microbiology 146 ( Pt 9) (2000 Sep), 2113-2120.
[229] GASCII, A. P. Comparative genomics of the environmental stress response in ascomycete fungi. Yeast 24, 11 (Nov 2007), 961-976.
[230] GASCII, A. P., SPELLMAN, P. T., KAO, C. M., CARNIEL-HAREL, 0., EISEN, M. B., STORZ, G., BOTSTEIN, D., AND BROWN, P. 0. Genomic expression programs in the response of yeast cells to environmental changes. Mol Biol Cell 11, 12 (2000 Dec), 4241-4257.
[231] GAUR, N. K., AND KLOTZ, S. A. Expression, cloning, and characterization of a Candida albicans gene, ALAI, that confers adherence properties upon Saccharomyces cerevisiae for extracellular matrix proteins. Infect Thernun 65, 12 (1997 Dec), 5289-5294.
[232] GAVIN, A., BOSCOE, M., KRAUSE, R., GRAND!, P., MARZIOCII, M., BAUER, A., SCHULTZ, J., LEUTWEIN, C., BOUWMEESTER, T., KUSTER, B.. AND SUPERTI-FURGA, G. Functional organization of the yeast proteome by systematic analysis of protein complexes. In Faseb Journal (9650 ROCKVILLE PIKE, BETHESDA, MD 20814-3998 USA, MAR 20 2002), vol. 16, FEDERATION AMER SOC EXP BIOL, pp. A523—A523.
[233] GAVIN, A.-C., ALOY, P., GRAND], P., KRAUSE, R., BOESCHE, M., MARZIOCII, M., RAU, C., JENSEN, L. J., BASTUCK, S., DUMPELFELD, B., EDELMANN, A., HEURTIER, M.-A., HOFFMAN, V., HOEFERT, C., KLEIN, K., HUDAK, M., MICRON, A.-M., SCHELDER, M., SCIIIRLE, M., REMOR, M., RUN, T., HOOFER, S., BAUER, A., BOUWMEESTER, T., CASARI, G., DREWES, G., NEUBAUER, G., RICK, J. M., KUSTER, B., BORK, P., RUSSELL, R. B., AND SUPERTI-FURGA, G. Proteome survey reveals modularity of the yeast cell machinery. Nature 440, 7084 (2006 Mar 30), 631-636.
[234] GEIS, P. A., WHEELER, M. H., AND SZANISZLO, P. J. Pentaketide metabolites of melanin synthesis in the dernatiaceous fungus Warigiella dermatitidi.s. Arch Mierobiol 187, 4 (Apr 1984), 324-328.
[235] GENOVA, M. L., PICII, M. M., BIONDI, A., BERNACCHIA, A., FALASCA, A., BOVINA, C., FORMIGGINI, G., PARENT] CASTELLI, G., AND LENAZ, G. Mitochondrial production of oxygen radical species and the role of Coenzyme Q as an antioxidant. Exp Biol Med (Maywood) 228, 5 (2003 May), 506-513.
[236] GERSUK, G. M., UNDERHILL, D. M., ZOO, L., AND MARR. K. A. Dectin-1 and TLRs permit macrophages to distinguish between different Aspergillus fumigatus cellular states. J Immunol 176, 6 (Mar 2006), 3717-3724.
[237] GIIANNOUM, M., AND AIM ELTEEN, K. Correlative relationship between proteinase production, adherence and pathogenicity of various strains of candida albicans. J Med Vet Mycol 24, 5 (Oct 1986), 407-413.
[238] GIIANNOUM. M. A., AND RICE, L. B. Antifungal agents: mode of action, mechanisms of resistance, and correlation of these mechanisms with bacterial resistance. Clin Microbiol Rev 12, 4 (1999 Oct), 501-517.
[239] GIAEVER, G., CIIU, A., NI, L., CONNELLY, C., RILES, L., VERONNEAU, S., Dow, S., LLICAU-DANILA, A., ANDERSON, K., ANDRE; B., ARKIN, A. P., ASTROMOFF, A., EL BAKKOURY, M., BANOIIAM, a., BENITO, R., BRACIIAT, S., CAMPANARO, S., CURTISS, M., D.AVIS, K., DEUTSCHBAUER, A., ENTIAN, K.-D., FLAHERTY, P., FOURY, F., GARFINKEL, D. J., GERSTEIN, M., GOTTE, D., GULDENER, I., HEGEMANN, J. H., HEMPEL. S., HERMAN, Z., JARAMILLO, D. F., KELLY, D. E., KELLY, S. L., KOTTER, P., LABONTE, D., LAMB, D. C., LAN, N., LIANG, H., LIAO, H., LIU, L., Luo, C., LUSSIER, M., MAO, R., MENARD, P., LOON 001, S., REVEULTA, J. L., ROBERTS, C. J., ROSE, M., ROSS-MACDONALD, P., SCHERENS, B., SCHIMNIACK, G., SHAFER, B., SHOEMAKER, D. D.. SOOKHAI-MAIIADEO, S., STORMS, R. K., STRATIIERN, J. N., VALLE, G., VOET, M., VOLCKAERT, G., WANG, C.-Y., WARD, T. R., WILIIELMY, J., WINZELER, E. A., YANG, Y., YEN, G., YOUNGMAN, E., Ye, K., HUSSEY, H., BOEKE, J. D., SNYDER, M., PHILIPPSEN, P., DAVIS, R. W., AND JOHNSTON, M. Functional profiling of the Saccharomyces cerevisiae. genome. Nature 418 (July 2002), 387-391.

REFERENCES 446
[240] GIAEVER, G., FLAIIERTY, P., KUMM, J., PROCTOR, M., NISLOW, C., JARAMILLO, D. F., CIILI, A. M., JORDAN, M. I., ARKIN, A. P., AND DAVIS, R. W. Chemogenomic profiling: identifying the functional interactions of small molecules in yeast. Proc Natl Aced Sci U S A 101, 3 (Jan 2004), 793-798.
[241] GIL, M. L., PENALVER, M. C., LOPEZ-RIBOT, J. L., O'CONNOR, J. E., AND MARTINEZ, J. P. Binding of extracellular matrix proteins to Aspergillus fumigates conidia. Infect itirmun 64, 12 (1996 Dec), 5239-5247.
[242] GIMENO, C. J., AND FINK, G. R. Induction of pseudohyphal growth by overexpression of PHD1, a Saccharomyces cerevisiae gene related to transcriptional regulators of fungal development. Mol Cell Biol 14, 3 (1994 Mar), 2100-2112.
[243] GIOT, L., BADER, J. S., BROUWER, C., CHAUDHURI, A., KUANG, B., LI, Y., HAO, Y. L., 001, C. E., GODWIN, 13., VITOLS, E., VIJAYADAMODAR, G., POCIIART, P., MACIIINENI, H., WELSH, NI., KONG, Y., ZERHUSEN, B., MALCOLM, R., VARRONE, Z., COLLIS, A., MINTO, M., BURGESS. S., IVICDANIEL, L., STIMPSON, E., SPRIGGS, F., WILLIAMS. J., NEL'RATII, K., botur, N., AGEE, NI., VOSS, E., FURTAK, K., RENZULLI, R., AANENSEN, N., CARROLLA, S., BICKELHAUPT, E., LAZOVATSKY, Y., DASILVA, A., ZIIONG, J., STANYON, C. A., FINLEY, R. L. J., WHITE, K. P., BRAVERMAN, NI., JARVIE, T., GOLD, S., LEACH, NI., KNIGHT, J., SHINIKETS, R. A., MCKENNA, M. P., CHANT, J., AND ROTIIBERG, J. NI. A protein interaction map of Drosophila melanogaster. Science 302, 5651 (Dec 2003), 1727-1736.
[244] GIRMENIA, C. New generation azole antifungals in clinical investigation. Expert Opin Investig Drugs 18, 9 (Sep 2009), 1279-1295.
[245] GLICKMAN, NI. H., AND 11hvEll, D. Proteasome plasticity. FEBS Lett 579, 15 (2005 Jun 13), 3214-3223.
[246] GOFFEAU, A., HARRELL, 13. G., BUSSEY, H., DAVIS, R. W., DUJON, B., FELDMANN, H., GALIBERT, F., HOIIEISEL, J. D., JACQ, C., JOHNSTON, LOUIS, E. J., Muwes, H. W., MURAKAMI, Y., PHILIPPSEN, P., TETTELIN, H., AND OLIVER, S. G. Life with 6000 genes. Science 274, 5287 (1996 Oct 25), 546,563-7.
[247] GOLA, S., MARTIN. R WALTIIER, A., DUNKLER, A., AND WENDLAND, J. New modules for FOR-based gene targeting in Candida albicans: rapid and efficient gene targeting using 100 bp of flanking homology region. Yeast 20, 16 (Dec 2003), 1339-1347.
[248] GOLDBERG, A. L. Protein degradation and protection against misfolded or damaged proteins. Nature 426, 6968 (2003 Dec 18), 895-899.
[249] GOLLUB, E. G., Lir, K. P., DAYAN, J., ADLERSRERG, NI., AND SPRINSON, D. B. Yeast mutants deficient in heme biosynthesis and a heme mutant additionally blocked in cyclization of 2,3-oxidosqualene. J Biol Chem 252, 9 (1977 May 10), 2846-2854.
[250] GOMEZ-LOPEZ, A., GARCIA-EFFRON, G., MELLADO, E., MONZON, A., RODRICUEZ-TUDELA, J. L., AND CUENCA-ESTRELLA, M. In vitro activities of three licensed antifungal agents against spanish clinical isolates of Aspergillus .spp. Antimicrob Agents Chemother 47, 10 (2003 Oct), 3085-3088.
[251] GONZALEZ, NI. NI., DIEZ-OREJAS, R., MOLERO, G., ALVAREZ. A. NI., PLA, J., NOMBELA, C., AND SANCHEZ-PEREZ, NI. Phenotypic characteriza-tion of a Candida albicans strain deficient in its major exoglucanase. Microbiology 143 ( Pt 9) (1997 Sep), 3023-3032.
[252] CORNER, W., DURCHSCHLAG, E., MARTINEZ-PASTOR, M. T., ESTRUCII, F., AMMERER, G., HAMILTON. B., Run, H., AND SCIIULLER, C. Nuclear localization of the C2H2 zinc finger protein Msn2p is regulated by stress and protein kinase A activity. Genes Dev 12, 4 (1998 Feb 15), 586-597.
[253] Cow, N. A. R., BROWN, A. J. P., AND ODDS, F. C. Fungal morphogenesis and host invasion. Curr Opin M erobuil 5,4 (2002 Aug), 366-371.
[254] GRANGER, B. L., FLENNIKEN, NI. L., DAVIS, D. A., MITCHELL, A. P., AND CUTLER, J. E. Yeast wall protein 1 of Candida Mbicans. Microbiology 151, Pt 5 (2005 May), 1631-1644.
[255] GREGORI, C., SCHULLER, C., ROETZER, A., SCHWARZMULLER, T., AllINICREIZ, G., AND KUCHLER, K The high-osmolarity glycerol response pathway in the human fungal pathogen Candida glabrata strain ATCC 2001 lacks a signaling branch that operates in baker's yeast. Eukaryot Cell 6, 9 (2007 Sep), 1635-1645.
[256] GROLL, A. H., PISCITELLI, S. C., AND WALSII, T. J. Clinical pharmacology of systemic antifungal agents: a comprehensive review of agents in clinical use, current investigational compounds, and putative targets for antifungal development. Advances in Pharma-cology 44 (1998), 343-500.
[257] GRUNENFELDER, B., AND WINZELER, E. A. Treasures and traps in genome-wide data sets: case examples from yeast. Nat Rev Genet 3, 9 (Sep 2002), 653-661.
[258] GUARRO, J., AND STCHIGEL, A. NI. Developments in fungal taxonomy. Clin Microbiol Rev 12, 3 (1999), 454-500.
[259] GUDLAUGSSON, 0., GILLESPIE, S., LEE, K., VANDE BERG, J., HU, J., MESSER, S., HERWALDT, L., PFALLER, M., AND DIEKEMA, D. Attributable mortality of nosocomial candidemia, revisited. Olin Infect D28 37, 9 (2003 Nov 1), 1172-1177.
[260] GUREVICH, NI., DEVINS, G. If., AND RODIN, G. M. Stress response syndromes and cancer: conceptual and assessment issues. Psycho-somatics 43, 4 (Jul-Aug 2002), 259-281.

REFERENCES 447
[261] DUSTIN, M. C., ALBERTYN, J., ALEXANDER, M., AND DAVENPORT, K. MAP kinase pathways in the yeast Saceliaromyces cercnisiae. Microbiol Mol Biol Rev 62,4 (1998 Dec), 1264-1300.
[262] GUTENKUNST. R. N., WATERFALL, J. J., CASEY, F. P., BROWN, K. S., MYERS, C. R., AND SETIINA, J. P. Universally sloppy parameter sensitivities in systems biology models. PLaS Comput Biol 3, 10 (2007 Oct), 1871-1878.
[263] HACBERC, A. A., SCHULT, D. A., AND STEWART, P. J. Exploring network structure, dynamics and function using networkx. In Proceedings of the 7th Python in Science Conference (ScaPy2008) (Pasadena, CA USA, Aug 2008), G. Varoquaux, T. Vaught, and J. Millman, Eds., pp. 11-15.
[264] HAGLUND, K., AND DIKIC, I. Ubiquitylation and cell signaling. EMBO J 24, 19 (2005 Oct 5), 3353-3359.
[265] HAKES, L., PINNEY, J. W., ROBERTSON, D. L., AND LOVELL, S. C. Protein-protein interaction networks and biology—what's the connection? Nat Biotechnol 26, 1 (2008 Jan), 69-72.
[266] HAMILTON, A. J., HOLDOM, M. D., AND HAY, R. J. Specific recognition of purified Cu,Zn superoxide dismutase from Aspergillus furnigatus by immune human sera. J Clin Microbiol 33, 2 (Feb 1995), 495-496.
[267] HAMPSEY, M. A review of phenotypes in Saccharomyces cerevistiae. Yeast 13 (1997), 1099-1133.
1268] HAMPSEY, M. Molecular genetics of the RNA Polymerase II general transcriptional machinery. Microbial Mol BM! Rev 62, 2 (1998
Jun), 465-503.
[269] HAN, S. J., LEE, Y. C., Gin. B. S., RYU, G. H., PARK, S. J., LANE, W. S., AND KIM, Y. J. Activator-specific requirement of yeast mediator proteins for RNA Polymerase H transcriptional activation. Mol Cell Biol 19, 2 (1999 Feb), 979-988.
[270] HANSEN, I. A., ATTARDO, G. NI., ROY, S. G., AND RAIKIIEL, A. S. Target of rapamycin-dependent activation of s6 kinase is a central step in the transduction of nutritional signals during egg development in a mosquito. J Biol Chem 280, 21 (May 2005), 20565-20572.
[271] HART, G. T., RAMANI, A. K., AND MARCOTTE, E. M. How complete are current yeast and human protein-interaction networks? Genomc Bird 7, 11 (2006), 120.
[272] HARTWELL, L. H., HOPFIELD. J. J., LEIBLER, S., AND MURRAY, A. W. From molecular to modular cell biology. Nature. 402,6761 Suppl (1999 Dec 2), C47-52.
[273] HATA, K., KIMURA, J., MIKI, H., TOYOSAWA, T., MORIYAMA, NI., AND KATSU, K. Efficacy of ER-30346, a novel oral triazole antifungal agent, in experimental models of aspergillosis, candidiasis, and cryptococcosis. Antimierob Agents Chemother 40, 10 (1996 Oct),
2243-2247.
[274] HAUGEN, A. C., KELLEY, R., COLLINS, J. B., TUCKER, C. J.. DENG, C., AFSIIARI, C. A., BROWN, J. M., IDEKER, T., AND VAN HOUTEN, B. Integrating phenotypic and expression profiles to map arsenic-response networks. Genome Biol 5,12 (2004), R95.
[275] HAUSER, C. R., GIULIANI, N. W., AND BOYNTON, J. E. Molecular Biology of Chlarnydomonas: Chloroplasts and Mitochondria. Kluwer Academic Publishers, 1998, ch. Regulation of chloroplast translation.
[276] HAWSER, S. P., AND DOUGLAS, L..1. Resistance of Candida albicans biofilms to antifungal agents in vitro. Antamicnob Agents Chemother 30, 9 (1995 Sep), 2128-2131.
[277] HAYASHI, A., FUJIOKA. S., NUKINA, NI., KAWANO, T., SIIIMADA, A., AND KIMURA, Y. Fumiquinones A and B, nematicidal quinones produced by Aspergillus furnigatus. Biosci Brotechnol Biochem 71, 7 (Jul 2007), 1697-1702.
[278] HAYNES, K. Virulence in Candida species. Trends in Microbiology 9, 12 (December 2001), 591-596.
[279] HEARN, V. M., WILSON, E. V., AND MACKENZIE, D. W. Analysis of Aspergillus fumigatus catalases possessing antigenic activity. J Med Microbiol 36, 1 (Jan 1992), 61-67.
[2801 HEITMAN, 1., MOVVA, N. R., AND HALL, M. N. Targets for cell cycle arrest by the immunosuppressant rapamycin in yeast. Science 253, 5022 (Aug 1991), 905-909.
[281] HENSEL, NI., ARST, H. N. J., AUFAUVRE-BROWN, A., AND HOLDEN, D. W. The role of the Aspergillus furnigatus areA gene in invasive
pulmonary aspergillosis. Mol Cen Genet 258, 5 (Jun 1998), 553-557.
[282] HENSEL, M., SHEA, 1. E., GLEESON, C., JONES, M. D., DALTON, E., AND HOLDEN, D. W. Simultaneous identification of bacterial virulence genes by negative selection. Science 260, 5222 (1995), 400-403.
[283] HERBRECIIT, B., DENNINC, D. W., PATTERSON, T. F., BENNETT. J. E., GREENE, R. E., OESTMANN, J.-W., KERN, W. V., MARR, K. A., RIBAUD, P., LORTIIOLARY, 0., SYLVESTER, R., RUBIN, R. H., WINCARD, J. R., STARK, P., DURAND, C., CAILLOT, D., THIEL, E., CHANDRASEKAR, P. H., HODCES, M. R., SCIILAMM, H. T., TROKE, P. F., AND DE PAUW, B. Voriconazole versus amphotericin B for primary therapy of invasive aspergillosis. N Engl J Mcd 347, 6 (Aug 2002), 408-415.

REFERENCES 448
[284] HERRANZ, S., RODRIGUEZ, J. M., BUSSINK, H.-J., SANCIIEZ-FERRERO, J. C., ARST, H. N. J.. PENALVA, M. A., AND VINCENT, 0. Arrestin-related proteins mediate pH signaling in fungi. Proc Nail Acad Sci U S A 102, 34 (Aug 2005), 12141-12146.
[285] HICKE, L., AND DUNN, R Regulation of membrane protein transport by ubiquitin and ubiquitin-binding proteins. Annu Rev Cell Dev Baal 19 (2003), 141-172.
[286] HICKMAN, M. J., AND WINSTON, F. Heme levels switch the function of Hapl of Saccharomyces cerevisiae between transcriptional
activator and transcriptional repressor. Mal Cell Riot 27, 21 (2007 Nov), 7414-7424.
[287] HILLENMEYER, M. E., FUNG, E., WILDENIIAIN, J., PIERCE, S. E., HOON, S., LEF,, W., PROCTOR, M., ST ONCE, R. P., TYERS, M., KOLLER, D., ALTMAN, R. B., DAVIS, R. W., NISLOW, C., AND GIAEVER, G. The chemical genomic portrait of yeast: uncovering a phenotype for all genes. Science 320, 5874 (2008), 362-365.
[288] HITCHCOCK, C. A., DICKINSON, K., BROWN, S. B., EVANS, E. G., AND ADAMS, D. J. Interaction of azole antifungal antibiotics with cytochrome P-450-dependent 14 alpha-sterol demethylase purified from Candida albicans. Biocherri J 266,2 (1990 Mar 1), 475-480.
[289] HOBERG, K. A., CHILAR, R. L., AND CALDERONE, R. A. Characterization of cerulenin-resistant mutants of Candida albicans. Infect Immure 51, 1 (1986 Jan), 102-109.
[290] HOCHBERG, Y., AND BENJAMINI, Y. More powerful procedures for multiple significance testing. Slat Med 9, 7 (1990 Jul), 811-818.
[291] HOFFMAN. R. iHOP, a new gene and protein analysis tool. Cancer Dial Titer 6, 1 (2007 Jan), 7-8.
[292] HOIIMANN, S. Osmotic stress signaling and osmoadaptation in yeasts. Micrabiol Mol Biol Rev 66, 2 (2002 Jun), 300-372.
[293] HOHMANN. S. The yeast systems biology network: mating communities. Curr Opirc Biotechnol 16, 3 (2005 Jun), 356-360.
[294] HOLZ, R. W. The effects of the polyene antibiotics nystatin and amphotericin B on thin lipid membranes. Ann N Y Acad Sci 235, 0 (1974 May 10), 469-479.
[295] HOSACK, D. A., DENNIS, G. J., SIIERMAN, B. T., LANE, H. C., AND LEMPICKI, R. A. Identifying biological themes within lists of genes with ease. G CTIOME Biot 4, 10 (2003), R70.
[296] HOTT1GER, T., DE VIRGILIO, C., HALL, M. N., BOLLER, T., AND WIEMKEN, A. The role of trehalose synthesis for the acquisition of thermotolerance in yeast. II. physiological concentrations of trehalose increase the thermal stability of proteins in vitro. Eur J Bioeliera 219, 1-2 (1994 Jan 15), 187-193.
[297] HOUNSA, C. G.. BRANDT, E. V., THEVELEIN, J.. HOHNIANN, S., AND PRIOR, B. A. Role of trehalose in survival of Saccharomyces cerevisiae under osmotic stress. Microbiology 144 ( Pt 3) (1998 Mar), 671-680.
[298] HOWARD, S. J., WEBSTER, I., MOORE, C. B., GARDINER, R. E., PARK, S., PERLIN, D. S., AND DENNING, D. W. Multi-azole resistance in Aspergillus fumigatus. Int J Antimicmb Agents 28, 5 (2006 Nov), 450-453.
[299] HOWLETT, B. J., IDNURM, A., AND HEITMAN, J. Fungal pathogenesis: gene clusters unveiled as secrets within the Ustilago maydis code.
Curr Biol 17, 3 (Feb 2007), R87-90.
[300] HOVER, L. L., AND HECHT, J. E. The ALS6 and ALS7 genes of Candida albicans. Yeast 16, 9 (2000 Jun 30), 847-855.
[301] HOVER, L. L., AND HECHT, J. E. The ALS5 gene of Candida albicans and analysis of the AlsSp N-terminal domain. Yeast 18, 1 (2001
Jan 15), 49-60.
[302] HOVER, L. L., PAYNE, T. L., BELL. M., MYERS. A. M., AND SCHERER, S. Candida albicans ALS3 and insights into the nature of the ALS gene family. Curl- Gcnet 33, 6 (1998 Jun), 451-459.
[303] HOVER, L. L., PAYNE, T. L., AND HECIIT, J. E. Identification of Candida albicans ALS2 and ALSO and localization of als proteins to
the fungal cell surface. J Bacterial 180, 20 (1998 Oct), 5334-5343.
[304] HOVER, L. L., SCHERER, S., SIIATZMAN, A. R., AND Loft, G. P. Candida albicans ALS1: domains related to a Saccharomyces cerevisiae sexual agglutinin separated by a repeating motif. Mol Microbial 15, 1 (1995 Jan), 39-54.
[305] HROMATKA, B. S., NOBLE, S. M., AND JOHNSON. A. D. Transcriptional response of Candida albicans to nitric oxide and the role of the Y11.131 gene in nitrosative stress and virulence. Mal Bird Cell 16, 10 (2005 Oct), 4814-4826.
[306] HSUEH. P.-R., LAU, Y.-J., CFIUANG, Y.-C., WAN, J.-H., HUANG, W.-K., SI1YR, J.-M., YAN, J.-J., Yu, K.-W., \Vu, J.-J., Ku, W.-C., YANG, Y.-C., LIU, Y.-C., TENC, L.-J., Liu, C.-Y., AND UM, K.-T. Antifungal susceptibilities of clinical isolates of Candida species, Cryptococcus
neoformans, and Aspergillus species from Taiwan: surveillance of multicenter antimicrobial resistance in Taiwan program data from 2003. Antirnicrob Agents Chernother 49, 2 (2005 Feb), 512-517.
[307] HUANG, M.-E.. RIO, A.-G., NICOLAS, A., AND KOLODNER, R. D. A genome-wide screen in Saccharomyces cerevisine for genes that suppress the accumulation of mutations. Pro, Natl Arad Sci U S A 100, 20 (Sep 2003), 11529-11534.

REFERENCES 449
[308] HUFINAGLE, G. B., CIEEN, G. II., CURTIS, J. L., MCDoNALD, R. A., STREETER, R. M., AND TOEWS. G. B. Down-regulation of the afferent phase of T cell-mediated pulmonary inflammation and immunity by a high melanin-producing strain of Cryptococcus neoformans.
J Immunol 155, 7 (Oct 1995), 3507-3516.
[309] HUGHES, A. L., TODD, B. L., AND ESPENSHADE, P. J. SREBP pathway responds to sterols and functions as an oxygen sensor in fission yeast. Cell 120, 6 (Mar 2005), 831-842.
[310] HULL, C. M., AND JOHNSON, A. D. Identification of a mating type-like locus in the asexual pathogenic yeast Candida albicans. Science
285, 5431 (Aug 1999), 1271-1275.
[311] IBRAHIM, A. S., SPELLBERG, B. J., AVANESIAN, V., FU, Y., AND EDWARDS, J. E. J. The anti-Candida vaccine based on the recombinant N-terminal domain of Alslp is broadly active against disseminated candidiasis. Infect Immun 74, 5 (2006 May), 3039-3041.
[312] IBRAHIM-GRANET, 0., DUBOURDEAU, NI., LATGE, J.-P., AVE, P., HUERRE, M., DRAKIIAGE, A. A., AND BROCK, M. Methylcitrate synthase from Aspergtilus furnigatus is essential for manifestation of invasive aspergillosis. Cell Microbiol 10, 1 (Jan 2008), 134-148.
[313] IDEKER, T., GALITSKI, T., AND HooD, L. A new approach to decoding life: systems biology. Annu Rev Cenomics Hum Genet 2 (2001),
343-372.
[314] IDEKEtt. T., OZIER, 0., SCHWIKOWSKI, B., AND SIEGEL, A. F. Discovering regulatory and signalling circuits in molecular interaction networks. Bioinformatics 18, S1 (2002), S233—S240.
[315] IKNER, A., AND SEHOZAKI, K. Yeast signaling pathways in the oxidative stress response. Mutat Res 569, 1-2 (2005 Jan 6), 13-27.
[316] IRAQUI, I., GARCIA-SANCHEZ, S., AUfiEftT, S., DIMMER, F., GIIIGO, J.-M.. D'ENFERT, C., AND JANBON, G. The Yaklp kinase controls expression of adhesins and biofilm formation in Candida glabrata in a Sir4p-dependent pathway. Mol Microbial 55, 4 (2005 Feb), 1259-1271.
[317] IRAQUI, I., VESSERS, S., ANDRE, B., AND URRESTARAZU, A. Transcriptional induction by aromatic amino acids in Saccharottryces cerevisiae. Mol Cell Btol 19, 5 (May 1999), 3360-3371.
[318] ITO, T., CHIBA, T., OZAWA, R., YOSHIDA, M., HATTORI, M., AND SAKAKI, Y. A comprehensive two-hybrid analysis to explore the yeast protein interactome. Proc Nat( Acad Set U S A 98, 8 (2001 Apr 10), 4569-4574.
[319] IWAMOTO, K., AND SHIRAIWA, Y. Salt-regulated mannitol metabolism in algae. Mar Biotechnol (NY) 7,5 (2005 Sep-Oct), 407-415.
[320] IWASE, M., AND TOEI-E, A. Nisi encoded by YNL078W: a new neck protein of Saccharotrryces cerevistae. Genes Genet Syst 76, 5 (2001
Oct), 335-343.
[321] JACOBSON, E. S., AND EMERY, H. S. Catecholamine uptake, melanization, and oxygen toxicity in Cryptococcus neafarmans. J Bacterial
173, 1 (Jan 1991), 401-403.
[322] JACOBSON, E. S., AND TINNELL, S. B. Antioxidant function of fungal melanin. J Bacterial 175, 21 (Nov 1993), 7102-7104.
[323] JAIIN, B., BOUKHALLOUK, F., LOTZ, J., LANGFELDER, K., WANNER, G., AND BRAKEIAGE, A. A. Interaction of human phagocytes with pigmentless Aspergillus connlia. Infect Immun 68, 6 (Jun 2000), 3736-3739.
[324] JANBON, G., HIMMELREICH, U.. MOYRAND, F., IMPROVISI, L., AND DROMER, F. Caslp is a membrane protein necessary for the 0-acetylation of the Cryptococcus neoformans capsular polysaccharide. Mol Microbial 42, 2 (Oct 2001), 453-467.
[325] JATON-OGAY, K., PARIS, S., HUERRE, NI., QUADRONI, NI., FALCHETTO, B., TOGNI, G., LATGE, J. P., AND MONOD, M. Cloning and disruption of the gene encoding an extracellular metalloprotease of Aspergillus fumigatus. Mol Microbial 14, 5 (Dec 1994), 917-928.
[326] .TATON-OGAY, K., SUTER, M., CRAMER', R., FALCHETTO, R., FATIII, A., AND MONOD, M. Nucleotide sequence of a genomic and a cDNA clone encoding an extracellular alkaline protease of AspergtIlus furnigatus. FEMS Microbial Lett 71, 2 (Apr 1992), 163-168.
[327] JATON-OGAY, K., SUTER, NI., CRAMER', R., FALCHETTO, R., FATIEL A., AND MONOD, NI. Nucleotide sequence of a genomic and a cDNA clone encoding an extracellular alkaline protease of Aspergillus fumtgatus. FEMS Microbial Lett 71, 2 (Apr 1992), 163-168.
[328] Jcoso, H., MASON, S. P., BARABASI, A. L., AND OLTVAI, Z. N. Lethality and centrality in protein networks. Nature 411, 6833 (May
2001), 41-42.
[329] JEONG, H., TOMBOR, B., ALBERT, R., OLTVAL Z. N., AND BARABASI. A. L. The large-scale organization of metabolic networks, Nature
407,6804 (Oct 2000), 651-654.
[330] JEONG, H., TONI/JR, B., ALBERT, B., OLTVAL Z. N., AND BARABASI, A.-L. The large-scale organization of metabolic networks. Nature
407 (2000), 651-654.
[331] JERNIGAN, C., AND M!STREE, B. Project CAYDAR.
[332] JEN, H., MCCAFFERY, J. NI., AND GROTE, E. Ergosterol promotes pheromone signaling and plasma membrane fusion in mating yeast. J Cell Biol 180, 4 (2008 Feb 25), 813-826.

REFERENCES 450
[333] JIN, Y. H., DUNLAP, P. E., MeBRIDE, S. J., AL-REFAI, H., BUSHEL, P. R., AND FREEDMAN, J. H. Global transcriptome and deletome profiles of yeast exposed to transition metals. PLoS Genet 4, 4 (2008 Apr), e1000053.
[334] JONES, G. M., STALKER, J., HUMPIIRAY, S., WEST, A., COX, T., ROGERS, J., DUNHAM, I., AND PRELICIL G. A systematic library for comprehensive overexpression screens in Saccharornyces cereeisiae. Nat Methods 5,3 (2008 Mar), 239-241.
[335] JONES, P. M., AND GEORGE, A. M. The abc transporter structure and mechanism: perspectives on recent research. Cell Mol Life Set 61, 6 (2004 Mar), 682-699.
[336] JONES, S., WHITE, G., AND HUNTER, P. R. Increased phenotypic switching in strains of Candida albicans associated with invasive
infections. Journal of Clinical Microbiology 32, 11 (November 1994), 2869-2870.
[337] KADOSII, D., AND STRUIIL, K. Histone deacetylase activity of Rpd3 is important for transcriptional repression in viva. Genes Des 12, 6 (1998 Mar 15), 797-805.
[338] KAISER, P., MONCOLLIN, V., CLARICE, D. J., WATSON, M. H., BEItTOLAET, B. L., REED, S. I., AND BAILLY, E. Cyclin-dependent kinase and Cks/Sucl interact with the proteasome in yeast to control proteolysis of M-phase targets. Genes Dee 13, 9 (1999 May 1),
1190-1202.
[339] KAMA, R., ROBINSON, M., AND GERST, J. E. Btn2, a Hookl ortholog and potential Batten disease-related protein, mediates late endosome-Golgi protein sorting in yeast. Mol Cell Bud 27, 2 (2007 Jan), 605-621.
[340] KAMAI, Y., KUBOTA. M., KAUAI, Y., HOSOKAWA, T., FUKUOKA, T., AND FILLER, S. G. Contribution of Candida albicans ALS1 to the
pathogenesis of experimental oropharyngeal candidiasis. Infect human 70, 9 (2002 Sep), 5256-5258.
[341] KAMRAN, M., CALCACNO, A.-M., FINDON, H., BIGNALL, E., JONES, M. D., WARN, P., HOPKINS, P., DENNING, D. W., BUTLER, G., ROGERS, T., hlflitsoiliEGEL, F. A., AND HAYNES, K. Inactivation of transcription factor gene A CE2 in the fungal pathogen Candida glabrata results
in hypervirulence. Eukaryotic Cell 3, 2 (2004), 546-552.
[342] KANIRAN, M. F. The identification and isolation of putative virulence determinants. PhD thesis, Imperial College, University of
London, London, 2003.
[343] KANEIIISA, M., ARAKI, M., GOTO, S., HATTORI, M., HIRAKAAA'A, M., ITOH, M., KATAYAMA, T.. KAWASIIIMA, S., OKUDA, S., TOKIMATSU, T., AND YAMANISIII, Y. Kegg for linking genomes to life and the environment. Nucleic Acids Res 36, Database issue (2008 Jan), D480-4.
[344] KAO, R., AND DAVIES, J. Fungal ribotoxins: a family of naturally engineered targeted toxins? Biochern Cell Biol 73, 11-12 (Nov-Dec
1995), 1151-1159.
[345] KAO. R., AND DAVIES, J. Molecular dissection of mitogillin reveals that the fungal ribotoxins are a family of natural genetically engineered ribonucleases. J Biol Cheers 274, 18 (Apr 1999), 12576-12582.
[346] KAPTEYN, J. C., HOVER, L. L., HECHT, J. E., MULLER, W. H., ANDEL, A., VERKLEIJ, A. J., MAKAROW, M., VAN DEN ENDE, H., AND KLIS, F. M. The cell wall architecture of Candida albicans wild-type cells and cell wall-defective mutants. Mol Microbiol 35, 3 (2000 Feb),
601-611.
[347] KAUR, R., CASTANO, I., AND CORMACK, 13. P. Functional genomic analysis of fluconazole susceptibility in the pathogenic yeast Candida
glabrata: roles of calcium signaling and mitochondria. Antirnicroh Agents Chemother 48, 5 (2004 May), 1600-1613.
[348] KAUR, R., DOMERGUE, R., ZUPANCIC, M. L., AND CORMACK, 13. P. A yeast by any other name: Candida glabrata and its interaction with
the host. Curr Opin Microbiol 8, 4 (2005), 378-384.
[349] KELLEIIER, R. J. R., FLANAGAN, P. M., AND KORNBERG, R. D. A novel mediator between activator proteins and the RNA polymerase II transcription apparatus. Cell 61, 7 (1990 Jun 29), 1209-1215.
[350] KELLER, N., Box, J., CHUNG, D., PERRIN, R. NI AND SHWAB, E. K. LaeA, a global regulator of Aspergillus toxins. Medical Mycology 44
(September 2006), S83-585.
[351] KELLER, N. P., TURNER, G., AND BENNETT, J. W. Fungal secondary metabolism - from biochemistry to genomics. Nat Rev Microinot
3, 12 (Dec 2005), 937-947.
[352] KELLIS, hi., BIRREN, B. NV., AND LANDER, E. S. Proof and evolutionary analysis of ancient genome duplication in the yeast Saccha-
rontyces cerevisine. Nature 428,6983 (2004), 617-624.
[353] KEMP, H. A., AND SPRAGUE, G. F. J. Far3 and five interacting proteins prevent premature recovery from pheromone arrest in the budding yeast Saccharoneyees ceremsiac. Mol Cell Rio( 23, 5 (Mar 2003), 1750-1763.
[354] KErauncE, D. The eukaryotic microbial cell. Cambridge University press, Cambridge, England, 1980, ch. The plasma membrane of
Candida albicans and its role in the action of antifungal drugs.
[355] KERRIDGE, D., KOII, T. Y., MARRIOT, hl. S., AND GALE, E. F. Microbiology and plant protoplasts. Churchill Livingston, London, England,
1976, ch. Microbiology and plant protoplasts, pp. 23-38.

REFERENCES 451
[356] KIECHLE, M., MANIVASAKARI, P., ECKARDT-SCIIUPP, F'., SCIIIESTL, R. H., AND FRIEDL, A. A. Promoter-trapping in Saccharomyces cerevisiae
by radiation-assisted fragment insertion. Nucleic Acids Res 30, 24 (Dec 2002), e136.
[357] KIM, K. S., ROSENKRANTZ, M. S., AND GUARENTE, L. Saccharumyces cerevisiae contains two functional citrate synthase genes. Mol Cell Biel 6,6 (1986 Jun), 1936-1942.
[358] KING, R. D., LEE, J. C., AND MORRIS, A. L. Adherence of Candida albican.s and other Candida species to mucosal epithelial cells. Infect Immun 27, 2 (1980 Feb), 667-674.
[359] KITANO, H. Computational systems biology. Nature 420,6912 (2002), 206-210.
[360] KL[EWER, S. A., GOODWIN, B., AND WILLSON, T. M. The nuclear pregnane X receptor: a key regulator of xenobiotic metabolism. Endocr Rev 23, 5 (Oct 2002), 687-702.
[361] KLIPP, E., NORDLANDER, B., KRUGER, R., GENNEMARK, P., ANI) HOHMANN, S. Integrative model of the response of yeast to osmotic shock. Nat Biotechnol 23, 8 (2005), 975-982.
[362] KLOTZ, S. A., DRUTZ, D. J., HARRISON, J. L., AND HUPPERT, M. Adherence and penetration of vascular endothelium by Candida yeasts.
Infect Immun 42, 1 (Oct 1983), 374-384.
[363] KNIGHT, C., AND PINNEY, J. Making the right connections: biological networks in the light of evolution. Btocssays 31, 10 (2009 Aug
31), 1080-1090.
[364] KNUTSEN, A. P., HUTCIIESON, P. S., SLAVIN. R. G., AND KURUP, V. P. IgE antibody to Aspergilluq fumigatus recombinant allergens in cystic fibrosis patients with allergic bronchopulmonary aspergillosis. Allergy 59, 2 (Feb 2004), 198-203.
[365] KOLATTUKUDY, P. E.. LEE, J. D., ROGERS, L. M., ZIMMERMAN, P., CESELSKI, S., Fox, B., STEIN, B., AND COPELAN, E. A. Evidence for possible involvement of an elastolytic serine protease in aspergillosis. Infect Immun 61, 6 (Jun 1993), 2357-2368.
[366] KOLOTILA, M. P., SMITH, C. W., AND ROGERS, A. L. Candidacidal activity of macrophages from three mouse strains as demonstrated by a new method: neutral red staining. J Med Vet Mycol 25,5 (Oct 1987), 283-290.
[367] KOMAGATA, D., FUJITA, S., YAMASIIITA, N., SAITO, S., AND MORINO, T. Novel neuritogenic activities of pseurotin A and penicillic acid. J Antibiot (Tokyo) 49, 9 (Sep 1996), 958-959.
[368] KOMEILI, A., WEDAMAN, K. P., O'SHEA, E. K., AND POWERS, T. Mechanism of metabolic control. target of rapamycin signaling links nitrogen quality to the activity of the Rtgl and Rtg3 transcription factors. J Cell Biol 151, 4 (2000 Nov 13), 863-878.
[369] KONTOYIANNIS, D. P. Modulation of fluconazole sensitivity by the interaction of mitochondria and Erg3p in Saccharrnayces cerevisiae. J Antimicroh Chemother 46, 2 (2000 Aug), 191-197.
[370] KONTOYIANNIS, D. P. Towards more effective antifungal therapy: the prospects of combination therapy. British Journal of Haema-tology 126 (2004), 165.
[371] KONTOYIANNIS, D. P., AND BODEY, G. P. Invasive aspergillosis in 2002: an update. Eur J Chu Microbrol Infect Dts 21, 3 (2002 Mar), 161-172.
[372] KOTHARY, M. H., CHASE, T. J., AND MACMILLAN, J. D. Correlation of elastase production by some strains of Asperyillus fumigatus with ability to cause pulmonary invasive aspergillosis in mice. Infect Immure 43,1 (Jan 1984), 320-325.
[373] KommtY, M. H., CHASE, T. J., AND MACMILLAN, J. D. Correlation of elastase production by some strains of Aspergtllus fumigatus with
ability to cause pulmonary invasive aspergillosis in mice. Infect Immun 43, 1 (Jan 1984), 320-325.
[374] KRAPPMANN, S., BIGNELL, E. M., REICIIARD, U., ROGERS, T., HAYNES, K., AND BRADS, G. H. The Aspergillus fumigatus transcriptional
activator CpcA contributes significantly to the virulence of this fungal pathogen. Mol Microbial 52,3 (May 2004), 785-799.
[375] KRAPPMANN, S., AND BRAUS, G. H. Nitrogen metabolism of Aspergillus and its role in pathogenicity. Med Mycol 43 Suppl 1 (May
2005), S31-40.
[376] KRETSCHMAR, M., FELK, A., STAIR, P., SCIIALLER, M.. HESS, D., CALLAPINA, M., MORSCHHAUSER, J., SCHAFER, W., KORTING, H. C., HOF, H., RUBE, B., AND NICIITERLEIN, T. Individual acid aspartic proteinases (Saps) 1-6 of Candida albican.s are not essential for invasion and colonization of the gastrointestinal tract in mice. Microh Pathog 32, 2 (2002 Feb), 61-70.
[377] KUDO, Y., OOTANI, T., KUMAGAI, T., FUKUCIII, Y., ERIN, K., AND YOKOTA, K. A novel oxidized low-density lipoprotein-binding protein, Asp-hemolysin, recognizes lysophosphatidylcholine. Biol Pharm Bull 25, 6 (Jun 2002), 787-790.
[378] KULLBERG, B. J., SOBEL, J. D., RUIINKE, M., PAPPAS, P. G., VISCOLI, C., REX, J. H., CLEARY, J. D., RUBINSTEIN, E., Client-It, L. W. P.. BROWN, J. M., SCIILAMM, H. T., OBORSKA, I. T., HILTON, F., AND HODGES, M. R. Voriconazole versus a regimen of amphotericin B followed by fluconazole for candidaemia in non-neutropenic patients: a randomised non-inferiority trial. Lancet 366, 9495 (Oct 2005), 1435-1442.

REFERENCES 452
[379] KUPFAHL, C., HEINEKAMP, T., GEGINAT, G., RUPPERT, T., HARTL, A., Hoc, H., AND BRAKIIAGE, A. A. Deletion of the GliP gene of Aspergillus furnigatus results in loss of gliotoxin production but has no effect on virulence of the fungus in a low-dose mouse infection model. Mot Microbial 62, 1 (Oct 2006), 292-302.
[380] KWAST, K. E., BURKE, P. V., BROWN, K., AND POYTON, R. 0. REO1 and ROX1 are alleles of the same gene which encodes a transcrip-tional repressor of hypoxic genes in Saccharorreyce.s cerevisiae. Curr Genet 32, 6 (1997 Dec), 377-383.
[381] KWAST, K. E., BURKE, P. V., AND POYTON, R. 0. Oxygen sensing and the transcriptional regulation of oxygen-responsive genes in yeast. J Exp Blot 201, Pt 8 (1998 Apr), 1177-1195.
[382] liwox-Cituxu, K. J., POLACHECK, I., AND POPKIN, T. .T. Melanin-lacking mutants of Cryptococcus neoformans and their virulence for mice. J Bacterial 150, 3 (Jun 1982), 1414-1421.
[383] KWON-CHUNG, K. J., AND RHODES, J. C. Encapsulation and melanin formation as indicators of virulence in Cryptococcus neciformans. Infect Inernun 51, 1 (Jan 1986), 218-223.
[384] LACIIKE, S. A., Jetty, S., K DANIELS, AND SOLL, D. R. Phenotypic switching and filamentation in Candida glabrata. Microbiology 148 (2002), 2661-2674.
[385] LACIIKE, S. A., SRIKANTIIA, T., TSAI, L. K., DANIELS, K., AND SOLL, D. R. Phenotypic switching in Candida glabrata involves phase-specific regulation of the metallothionein gene MT-I and the newly discovered hemolysin gene HLPi. Infection and Immunity 68, 2 (February 2000), 884-895.
[386] LAI, H.-Y., TAM, M. F., TANG, R.-B., Cituu, H., CIIANG, C.-Y., TSAI, J.-J., AND SHEN, H.-D. cDNA cloning and immunological characteri-zation of a newly identified enolase allergen from Penicillium citrinum and Aspergillus furnigatus. Int Arch Allergy Immunol 127, 3 (Mar 2002), 181-190.
[387] LALOUX, I., DUBOIS, E., DEWERCIIIN, M., AND JACOBS, E. TEC1, a gene involved in the activation of Tyl and Tyl-mediated gene expression in Saccharomyces cerevisiae: cloning and molecular analysis. Mol Cell Biol 10, 7 (1990 Jul), 3541-3550.
[388] LAMB, T. NI., AND MITCHELL, A. P. The transcription factor RimlOip governs ion tolerance and cell differentiation by direct repression of the regulatory genes NRG1 and SMP1 in Saccharomyce.s cerevisiae. Mal Cell Biol 23, 2 (2003 Jan), 677-686.
[389] LAMP, B., MOUTAOUAKIL, M., LATGE, J. P., AND DAVIES, .1. Secretion of a potential virulence factor, a fungal ribonucleotoxin, during human aspergillosis infections. Mal Microbial 5, 7 (Jul 1991), 1811-1815.
[390] LANDRY, J., SUTTON, A., TAFROV, S. T., HELLER, R. C., STEBBINS. J., PILLUS, L., AND STERNGLANZ, R. The silencing protein sir2 and its homologs are NAD-dependent protein deacetylases. Proc Nail Acad Sci U S A 97, 11 (May 2000), 5807-5811.
[391] LANGFELDER, K., JAHN, B., GEIIRINGER, H., SCIIMIDT, A., WANNER, G., AND BRAKIIAGE, A. A. Identification of a polyketide synthase gene (pksP) of Aspergillus furnagatus involved in conidial pigment biosynthesis and virulence. Med Microbial Immunol 187, 2 (Oct 1998), 79-89.
[392] LARSSON, K., ANSELL, R., ERIKSSON, P., AND ADLER, L. A gene encoding sn-glycerol 3-phosphate dehydrogenase (NADI-) complements an osmosensitive mutant of Saccharamyces cerevisiae. Mal Microbial 10, 5 (1993 Dec), 1101-1111.
[393] LASLO, P., SPOONER, C. J., WARMPLASII, A., LANCKI, D. W., LEE, H.-J., SCIAMMAS, R., GANTNEIt, B. N., DINNER, A. R., AND S[NGH, H. Multilineage transcriptional priming and determination of alternate hematopoietic cell fates. Cell 126, 4 (Aug 2006), 755-766,
[394] LATGE, J. P. Aspergillus fumigates and aspergillosis. Clin Microbial Rev 12, 2 (Apr 1999), 310-350.
[395] LAU, G., AND HAMER, J. Regulatory genes controlling mpg1 expression and pathogenicity in the rice blast fungus Magriaporthe grisea. Plant Cell 8, 5 (May 1996), 771-781.
[396] LAZEBNIK, Y. Can a biologist fix a radio? - or, what i learned while studying apoptosis, (cancer cell. 2002 sep;2(3):179-82). Biochemistry (Most) 69, 12 (2004), 1403-1406.
[397] LEBERER, E., ZIEGELBAUER, K., SCHMIDT, A., HARCUS, D., DIGNARD, D., Asti, J., JOHNSON, L., AND THOMAS, D. Y. Virulence and hyphal formation of Candida alhicans require the Ste20p-like protein kinase CaCla4p. Curr Riot 7,8 (1997 Aug 1), 539-546.
[398] LEE, J. D., AND KOLATTUKUDY, P. E. Molecular cloning of the cDNA and gene for an elastinolytic aspartic proteinase from Asperyillus fumigatus and evidence of its secretion by the fungus during invasion of the host lung. Infect Iminun 63, 10 (Oct 1995), 3796-3803.
[399] LEE, T. I., AND YOUNG, R. A. Transcription of eukaryotic protein-coding genes. Anna Rev Genet 34 (2000), 77-137.
[400] LEE, Y. C., PARK, J. M., MIN, S., HAN, S. J., AND KIM, Y. J. An activator binding module of yeast RNA Polymerase II holoenzyme. Med Cell Biol /9,4 (1999 Apr), 2967-2976.
[401] LEFRANCOIS, P., EUSKIRCIIEN, G. M., AUERBACII, R. K., ROZOWSKY, J., GIBSON, T., YELLMAN, C. M., GERSTEIN, M., AND SNYDER, M. Efficient yeast ChIP-Seq using multiplex short-read DNA sequencing. BMC Cenomics 10 (2009), 37.

REFERENCES 453
[402] LELANDAIS, G., TANTY, V., GENEIX, C., ETCHEREST, C., JACQ, C., AND DEVAUX, F. Genome adaptation to chemical stress: clues from comparative transcriptomics in Saccharotnyces cerevisiae and Candida glabrata. Genome Bull 9, 11 (2008 Nov 24), R164.
[403] LENAZ, G., BOVINA, C., D'AURELIO, M., FATO, R., FORMIGGINI, G., GENOVA, M. L., GIULIANO, G., MERLO PICH, M., PAOLUCCI, U., PAR-ENT' CASTELLI, G., AND VENTURA, B. Role of mitochondria in oxidative stress and aging. Ann N Y Acad Sea 959 (2002 Apr), 199-213.
[404] LESACE, G., SHAPIRO, J., SPECIIT, C. A., SDICU, A.-M., MENARD, P., HUSSEIN, S., TONG, A. H. Y., BOONE, C., AND BUSSEY, H. An interactional network of genes involved in chitin synthesis in Saccharomyces cerevisiae. BMC Genetics 6, 8 (2005).
[405] LETOVSKY, S., AND KASH', S. Predicting protein function from protein/protein interaction data: a probabilistic approach. Bioinfor-matics 19 Suppl 1 (2003), i197-204.
[406] LEWIS, R., KLEPSER, M., AND PFALLER, M. Combination systemic antifungal therapy for cryptococcosis, candidiasis and aspergillosis. Journal of infectious Diseases Phormacotherapy 3 (1999), 61-83.
[407] LI, S., ARMSTRONG, C. M., DENTIN, N., GE, H., MII.STEIN, S., BoXEM, M., VIDALAIN, P.-0., HAN, .T.-D. J., CIIESNEAU, A., HAO, T., GOLDBERG, D. S.. LI. N., MARTINEZ, M., REAL, J.-F., LAMESCH, P., Xu, L., TEWARI, M., WONG, S. L., ZIIANG, L. V., BERRIZ, G. F., JACOTOT, L., VAGLIO, P., REBOUL, J., HIROZANE-KISIIIKAWA, T., LI, Q., GABEL, H. W., ELEWA, A., BAUMGARTNER, B., ROSE, D. J., Yu, H., BOSAK, S., SEQUERRA, R., FRASER, A., MANGO, S. E., SAXTON, W. M., STROME, S., VAN DEN HEUVEL, S., PIANO, F., VANDF,NHAUTE, J., SARDET, C., GERSTEIN. M., DOUCETTE-STAMM, L., GUNSALUS, K. C., HARPER, .T. W., Cusick, M. E., ROTH, F. P., HILL, D. E., AND VIDAL, M. A map of the interactome network of the metazoan C. elegans. Science 303, 5657 (Jan 2004), 540-543.
[408] LIAO, X., AND Burow, R. A. RTG1 and RTG2: two yeast genes required for a novel path of communication from mitochondria to the nucleus. Cell 72, 1 (1993 Jan 15), 61-71.
[409] LIN, C. H., N1ACGURN, J. A., Cott, T., STEFAN, C. J., AND Emu, S. D. Arrestin-related ubiquitin-ligase adaptors regulate endocytosis and protein turnover at the cell surface. Cell 135, 4 (2008 Nov 14), 714-725.
[410] LIN, H.. NGUYEN, P., AND VANCURA, A. Phospholipase C interacts with Sgdlp and is required for expression of GPD1 and osmore-sistance in Saccharomyces cereristiae. Mol Genet Genornics 267, 3 (2002 May), 313-320.
[411] LIN, S. J., SCIIRANZ, J., AND TEUTSCII, S. M. Aspergillosis case-fatality rate: systematic review of the literature. C/tin Infect Dia 32, 3 (2001 Feb 1), 358-366.
[412] LINDAMOOD, S., HEATHERLY, R., KANTARCIOGLU, M., AND THURAISINGHAM, B. Inferring private information using social network data. In WWW '09: Proceedings of the 18th international conference on World wide web (2009), pp. 1145-1146.
[413] LIPKE, P. N., WOJCIECIIOWICZ, D., AND KURJAN, J. AG alpha 1 is the structural gene for the Saccharomyces cerevisiae alpha-agglutinin, a cell surface glycoprotein involved in cell-cell interactions during mating. Mol Cell Biol 9, 8 (1989 Aug), 3155-3165.
[414] LIU, H., KOHLER, J., AND FINK, G. R. Suppression of hyphal formation in Candida &Weans by mutation of a STE12 homolog. Science 266,5191 (1994 Dec 9), 1723-1726.
[415] LIE, H., KOHLER, J.. AND FINK, G. R. Suppression of hyphal formation in Candida albmans by mutation of a STE12 homolog. Science 266, 5191 (1994 Dec 9), 1723-1726.
[416] Liu, H., STYLES, C. A., AND FINK, G. R. Elements of the yeast pheromone response pathway required for filamentous growth of diploids. Science 262, 5140 (1993 Dec 10), 1741-1744.
[417] Liu, H. Y., BADARINARAYANA, V., AUDINO, D. C., RAPPSILBER, J., MANN, M., AND DENIS, C. L. The NOT proteins are part of the CCR4 transcriptional complex and affect gene expression both positively and negatively. EMBO J 17, 4 (1998 Feb 16), 1096-1106.
[418] LIU, L., TEWARI, R. P., AND WILLIAMSON, P. R. Lactase protects Cryptococcus neoforrnans from antifungal activity of alveolar macrophages. Infect /DEMUR 67, 11 (Nov 1999), 6034-6039.
[419] Liu, T. T., LEE, R. E. B., BARKER, K. S., LEE, R. E., WEI, L., HOMAYOUNI, R., AND ROGERS, P. D. Genome-wide expression profiling of the response to azole, polyene, echinocandin, and pyrimidine antifungal agents in Candida albicarts. Antimicrob Agents Chernother 49 , 6 (2005 Jun), 2226-2236.
[420] LONIAL, S., WILLIAMS, L., CARRUM, G., OSTROWSKI, M., AND MCCARTIIY, P. J. Nermartorya filchers: an invasive fungal pathogen in an allogeneic bone marrow transplant patient. Bone Marrow Transplant 19, 7 (1997 Apr), 753-755.
[421] LORENZ, NI. C., BENDER, J. A.. AND FINK, G. R. Transcriptional response of Candida albicans upon internalization by macrophages. Enkaryot Cell 3, 5 (2004 Oct), 1076-1087.
[422] LORENZ, R. T., AND PARKS, L. W. Involvement of heme components in sterol metabolism of Saccharornyces cerevisiae. Lipids 26, 8 (1991 Aug), 598-603.
[423] LOSS, 0. Perturbation of pH and calcium signalling as a means to prevent Aspergillus infection. PhD thesis, Imperial College, Department of Microbiology, Division of Investigative Science, Imperial College London SW7 2AZ, July 2009.

REFERENCES 454
[424] LUCCHINI, S., THOMPSON, A., AND HINTON, J. C. Microarrays for microbiologists. Microbiology 147, Pt 6 (2001 Jun), 1403-1414.
[425] LUIKENHUIS, S.. PERRONE, G., DAWES, I. W., AND GRANT, C. M. The yeast Saccharomyces cerevisiae contains two glutaredoxin genes that are required for protection against reactive oxygen species. Mel Biel Cell 9,5 (1998 May), 1081-1091.
[426] LUPETTI, A., DANES!, R., CAMPA, M., DEL TACCA. M., AND KELLY, S. Molecular basis of resistance to azole antifungals. Trends Mel Med 8, 2 (2002 Feb), 76-81.
[427] LUTFIYYA, L. L., IYER, V. R., DERist, J., DEVIT, M. J., BROWN, P. 0., AND JOHNSTON, M. Characterization of three related glucose repressors and genes they regulate in Saccharomyees cerevisiae. Genetics 150, 4 (1998 Dec), 1377-1391.
[428] LUYTEN, K., ALBERTYN, J., SKIBBE, W. F., PRIOR, B. A., RAMOS, J., THEVELEIN, J. M., AND HOHMANN, S. Fpsl, a yeast member of the MIP family of channel proteins, is a facilitator for glycerol uptake and efflux and is inactive under osmotic stress. EMBO J 14, 7 (Apr
1995), 1360-1371.
[429] MA, Y., QIAO. J., LID, W., WAN, Z., WANG. X., CALDERONE, R., AND LI, R. The Shol sensor regulates growth, morphology, and oxidant adaptation in Aspergillus fumigates but is not essential for development of invasive pulmonary aspergillosis. Infect Immure 76, 4
(Apr 2008), 1695-1701.
[430] MAcCALLEAL D. M., FINDON, II., KENNY, C. C., BUTLER. G., HAYNES, K., AND ODDS, F. C. Different consequences of ACE2 and SWI5 gene disruptions for virulence of pathogenic and nonpathogenic yeasts. Infect Iminun 74, 9 (Sep 2006), 5244-5248.
[431] MACHIDA, M., ASAI, K., SANO, M., TANAKA, T., KUMAGAI, T., TERM, G., KUSLIMOTO, K.-1, ARIMA, T., AKITA, 0., KASIIIWAGI, Y., ABE, K., GOMI, K., HORIUCIII, H., KITAMOTO, K., KOBAYASHI, T., TAKEUCHI, M., DENNING, D. W., GALAGAN, J. E., NIERMAN, W. C., Yu, J., ARCHER, D. B., BENNETT, J. W., BHATNAGAR, D., CLEVELAND, T. E., FEDOROVA, N. D., G070it, 0., HORIKAWA, H., HOSOYAMA, A., ICHINOMIYA, M., IGARASHI, R., IWASHITA, K., JUVVADI, P. R., KATO, M., KATO, Y., KIN, T., KOKUBUN, A., MAEDA, H., MAEYAMA, N., MARUY.4MA, J.-I, NAGASAKI, H., NAKAJIMA, T., ODA, K., OKADA, K., PAULSEN, I., SAKAMOTO, K., SAWANO, T., TAKAIIASIII, M., TAKASE, K., TERABAYASHI, Y., WORTMAN, J. R.,
YAMAHA, 0., YAMAGAT.A, Y., ANAZAWA, H., HATA, Y., KOIDE, Y., KOMORI, T., KovAmA, Y., MINETOKI, T., SUIIARNAN, S., TANAKA, A., IsoNo, K., KUHARA, S., OGASAWARA, N., AND KIKUCHI, H. Genome sequencing and analysis of Aspergillus oryzae. Nature 438, 7071 (2005 Dec
22), 1157-1161.
[432] MACKENZIE, K. F., SINGII, K. K., AND BROWN, A. D. Water stress plating hypersensitivity of yeasts: protective role of trehalose in Saccharomyces cerevisiae. J Gen Microbiol 134, 6 (1988 Jun), 1661-1666.
[433] MACPHERSON, S., LAROCIIELLE, M:, AND TURCOTTE, B. A fungal family of transcriptional regulators: the zinc cluster proteins. Microbiol Mel Biol Rev 70, 3 (Sep 2006), 583-604.
[434] MACPHERSON. S., LAROCHELLE. M., AND TURCOTTE, B. A fungal family of transcriptional regulators: the zinc cluster proteins. Microbaol Mot Dial Rev 70, 3 (2006 Sep), 583-604.
[435] MAEDA, T., TAKEKAWA, M., AND SAITO, H. Activation of yeast PBS2 MAPKK by MAPKKKs or by binding of an SH3-containing
osmosensor. Science 269, 5223 (1995 Jul 28), 554-558.
[436] MAERKER, C., ROHDE, M., BRAKIIAGE, A. A., AND BROCK, M. Methylcitrate synthase from Asperyillas fumigatus. Propionyl-CoA affects
polyketide synthesis, growth and morphology of conidia. FEBS J 272, 14 (Jul 2005), 3615-3630.
[437] MAGO, N., AND KHULLER, G. K. Influence of lipid composition on the sensitivity of Candida albicans to antifungal agents. Indian J Biochern Biophys 26, 1 (1989 Feb), 30-33.
[438] MAIDAN, M. M.. DE BOP, L., SERNEELS, J., EXLER, S., Rupp, S., TOURNU, H., TIIEVELEIN, J. M.. AND VAN DI.ICK, P. The G protein-coupled receptor Gprl and the G, protein Gpa2 act through the cAMP-protein kinase A pathway to induce morphogenesis in Candida albicans. Mel Biol Cell 16, 4 (2005 Apr), 1971-1986.
[439] MALTA, S., GRUNDMANN, A., LI, X., Li, S.-M., AND TURNER, G. Identification of a hybrid PKS/NRPS required for pseurotin A biosyn-thesis in the human pathogen Aspergillus fumigatus. Chembiochein 8, 14 (Sep 2007), 1736-1743.
[440] MALFIOTRA, J. D., AND KAUFMAN, R. J. Endoplasmic reticulum stress and oxidative stress: a vicious cycle or a double-edged sword? Antioxid Redox Signal 9, 12 (2007 Dec), 2277-2293.
[441] MAMNUN, Y. M., PANDJMTAN, R., MAIIE, Y., DELAHODDE, A., AND KUCHLER, K. The yeast zinc finger regulators Pdrlp and Pdr3p control pleiotropic drug resistance (PDR) as homo- and heterodimers in vivo. Mot Microbiol 46, 5 (2002 Dec), 1429-1440.
[442] MANAVATHU, E. K., VAZQUEZ, J. A., AND CFIANDRASEKAIL, P. H. Reduced susceptibility in laboratory-selected mutants of Aspergillus fumigatus to itraconazole due to decreased intracellular accumulation of the antifungal agent. Int J Antimicrob Agents 12, 3 (Aug
1999), 21.3-219.
[443] MARECHAL, P. A., AND GERVAIS, P. Yeast viability related to water potential variation: influence of the transient phase. Applied microbiology and biotechnology 42, 4 (1994), 617-622.

REFERENCES 455
[4441 MARKARYAN, A., MOROZOVA, I., Yu, H., AND KOLATTUKUDY, P. E. Purification and characterization of an elastinolytic metalloprotease from Aspergillus fumrgatus and immunoelectron microscopic evidence of secretion of this enzyme by the fungus invading the murine lung. Infect Immun 62, 6 (Jun 1994), 2149-2157.
[445] MARSTON, A. L., Diam, W.-H., Snail, H., AND AMON, A. A genome-wide screen identifies genes required for centromeric cohesion. Science 303, 5662 (Feb 2004), 1367-1370.
[446] MARTINEZ-PASTOR, M. T., MARCHLER, G., SCHULLER, C., IAIARCHLER-BAUER, A., Ruts, H., AND ESTRUCII, F. The Saccharomyces cerevtsiae zinc finger proteins Msn2p and Msn4p are required for transcriptional induction through the stress response element (STRE). EMBO J 15, 9 (1996 May 1), 2227-2235.
[447] MATTERN, I. E., PUNT, P. J., AND VAN DEN HONDEL, C. A. NJJ A vector of Aspergillus transformation conferring phleomycin
resistance. Funal Genetics Newsletter, 25 (35).
[448] MATTISON, C. P., AND OTA, I. M. Two protein tyrosine phosphatases, Ptp2 and Ptp3, modulate the subcellular localization of the Hogl MAP kinase in yeast. Genes Day 14, 10 (2000 May 15), 1229-1235.
[449] MATrisoN, C. P., SPENCER, S. S., KRESGE, K. A., LEE, J., AND OTA, I. M. Differential regulation of the cell wall integrity mitogen-activated protein kinase pathway in budding yeast by the protein tyrosine phosphatases Ptp2 and Ptp3. Mot Cell Biol 19, 11
(1999 Nov), 7651-7660.
[450] MAYSER, P., WENZEL, M., KRAMER, 11-4., KINDLER, B. L. J. SPITELLER, P., AND HAASE, G. Production of indole pigments by Candida ylabrata. Med Mycol 45, 6 (Sep 2007), 519-524.
[451] McANuurv, R. J. Methods for measuring hydroxyproline and estimating to vivo rates of collagen synthesis and degradation. Methods Mot Med 117 (2005), 189-207.
[452] MCCLEAN, M. N., MODY, A., BROACH, J. R., AND RAMANATHAN, S. Cross-talk and decision making in MAP kinase pathways. Nat Genet
39, 3 (2007 Mar), 409-414.
[4531 MCDONAGII, A., FEDOROVA, N. D., CRABTREE, J., Yu, Y., Km, S., CIIEN, D., Loss, 0., CAIRNS, T., GOLDNIAN, G., ARMSTRONG-JAMES, D., HAYNES, K., HAAS, H., SCIIRETTL, M., MAY, G., NIERMAN, W. C.. AND BIGNELL, E. Sub-telomere directed gene expression during initiation of invasive aspergillosis. PLoS Pathog 4, 9 (2008), e1000154.
[454] McKEnnow, J. H., SUN, E., ROSENTHAL, P. J AND BOUVIER, J. The proteases and pathogenicity of parasitic protozoa. Annu Rev Microbiol 47 (1993), 821-853.
[455] McLAuctiLix, D., HIBBETT, D.. LUTZONI, F., SPATAFORA, J., AND VILGALYS, R. The search for the fungal tree of life. Trends Microbial (2009 Sep 24).
[456] MCNEIL, M. M., NASH, S. L., HAJJEII, R. A., PHELAN, M. A., CONN, L. A., PLIKAYTIS, B. D., AND WARNOCK, D. W. Trends in mortality due to invasive mycotic diseases in the United States, 1980-1997. Clin Infect Dis 33, 5 (2001 Sep 1), 641-647.
[457] MEDINA, M. Genomes, phylogeny, and evolutionary systems biology. Proc Natl Acad Sci U S A 102 Suppl 1 (2005 May 3), 6630-6635.
[4581 .MEHRA, S., CHARANIYA, S., TAKANO, E., AND HU, W.-S. A bistable gene switch for antibiotic biosynthesis: the butyrolactone regulon in streptomyces coelicolor. PLoS One 3, 7 (2008), e2724.
[459] MEIKLE, A. J., REED, R. H., AND GADD. G. M. Osmotic adjustment and the accumulation of organic solutes in whole cells and protoplasts of Saccharomyces cerevisiae. J Gen Microbiol 134, 11 (1988 Nov), 3049-3060.
[460] MELLADO, E.. GARCIA-EFFRON, G., BUITRAGO, M. J., ALCAZAR-FI'OLI, L., CUENCA-ESTRELLA, M., AND RODRIGUEZ-TUDELA, J. L. Targeted gene disruption of the 14-alpha sterol demethylase (cyp5la) in Aspergillus fumigates and its role in azole drug susceptibility. Antimicrob
Agents Chernother 49, 6 (Jun 2005), 2536-2538.
[461] MENDIZABAL, I., RIOS, G., MULCT, J. M., SERRANO, R., AND DE LARRINOA, I. F. Yeast putative transcription factors involved in salt tolerance. FEES Lett 425,2 (1998 Mar 27), 323-328.
[462] MESECKE, N., SPANCI, A., DEPONTE, M., AND HERRMANN, J. M. A novel group of glutaredoxins in the cis-Golgi critical for oxidative stress resistance. Mal Dial Cell 19, 6 (2008 Jun), 2673-2680.
[463] MILGRAM, S. The small world. Psychology today 1 (1967), 60.
[464] MILO, R SIIEN-ORR, S., ITZKOVITZ, S., KASIITAN, N., CIIKLOSKII, D., AND ALON, U. Network motifs: Simple building blocks of complex networks. Science 298 (2002), 824-827.
[465] MISHRA, K., AND SHORE, D. Yeast Ku protein plays a direct role in telomeric silencing and counteracts inhibition by Rif proteins.
Carr Blot 9, 19 (Oct 1999), 1123-1126.
[466] MISSALL, T. A., LODGE, J. K., AND McEwEN, J. E. Mechanisms of resistance to oxidative and nitrosative stress: implications for fungal survival in mammalian hosts. Eukaryot Cell 3, 4 (2004 Aug), 835-846.

REFERENCES 456
[467] MITCHEL, R. E., AND MORRISON, D. P. Heat-shock induction of ionizing radiation resistance in Saccharensyces cerevisiae, and corre-lation with stationary growth phase. Radiat Res .90,2 (1982 May), 284-291.
[468] MITCIIEL, R. E., AND MORRISON, D. P. Heat-shock induction of ultraviolet light resistance in Saccharornyees cerevisiae. Radiat Res 96, 1 (1983 Oct), 95-99.
[469] MITCHELL, A. P. Dimorphism and virulence in Candida albicans. Curr Opin Microbial 1, 6 (1998 Dec), 687-692.
[470] MITCHELL, C. G., SLIGHT, J., AND DONALDSON, K. Diffusible component from the spore surface of the fungus Aspergillus furnigatus which inhibits the macrophage oxidative burst is distinct from gliotoxin and other hyphal toxins. Thorax 52, 9 (Sep 1997), 796-801.
[4711 MIWA, T., TAKAGI, Y., SIIINOZAKI, M., YUN, C.-W., SCIIELL, W. A., PERFECT, J. R., KUMAGAI, H., AND TANIAKI, H. Gprl, a putative G-protein-coupled receptor, regulates morphogenesis and hypha formation in the pathogenic fungus Candida albicans. Eukaryot Cell 3, 4 (2004 Aug), 919-931.
[472] MIZUNO, T., AND HARASHIMA, S. Gault is a general activator of basal transcription, whose activity is regulated by the general repressor Sin4 in yeast. Mol Genet Genornies 269, 1 (2003 Apr), 68-77.
[473] MOLLAPOUR, M., AND PIPER, P. W. Hogl mitogen-activated protein kinase phosphorylation targets the yeast Fps1 aquaglyceroporin for endocytosis, thereby rendering cells resistant to acetic acid. Mol Cell Biol 27,18 (2007 Sep), 6446-6456.
[474] MONOD, M., AND BORG-VON ZEPEL1N, M. Secreted proteinases and other virulence mechanisms of Candida albicans. Chem Immunol 81 (2002), 114-128.
[475] MONOD, M., PARIS, S., SARFATI, 4., JATON-OGAY, K., AVE, P., AND LATGE, J. P. Virulence of alkaline protease-deficient mutants of Aspergillus fumigatus. FEMS Microbial Lett 106, 1 (Jan 1993), 39-46.
[476] MONOD, M., TOGNI, G., RAHALISON, L., AND FRENK, E. Isolation and characterisation of an extracellular alkaline protease of A spergillus furnigatus. J Med Microbial 35, 1 (Jul 1991), 23-28.
[477] MONTEIRO, P. T., MENDES, N. D., TEIXEIRA, M. C., D'OREY, S., TENREIRO, S., MIRA, N. P., PAIS, H., FRANCISCO, A. P., CARVALIIO, A. M., LOURENCO, A. B., SA-CORREIA, I., OLIVEIRA, A. L., AND FREITAS, A. T. YEASTRACT-DISCOVERER: new tools to improve the analysis of transcriptional regulatory associations in Saccharornyccs cerevisiae. Nucleic Acids Res 36, Database issue (2008 Jan), D132-6.
[478] MORENO, M. A., IBRAIIIM-GRANET, 0., VICENTEFRANQUEIRA, AMICH, J., AVE, P., LEAI., F., LATGE, J.-P., AND CALERA, J. A. The regulation of zinc homeostasis by the ZafA transcriptional activator is essential for Aspergillus furnigatus virulence. Mol Microbial 64, 5 (Jun 2007), 1182-1197.
[479] MORGAN, J., MELTZER, M. I., PLIKAYTIS, B. D., SOFAIR, A. N.. HUM-WHITE, S., WILCOX, S., HARRISON, L. H., SEABERG, E. C., HAJJEll, R. A., AND TEUTSCH, S. M. Excess mortality, hospital stay, and cost due to candidemia: a case-control study using data from population-based candidemia surveillance. Infect. Control Hasp Epidemiol 26, 6 (2005 Jun), 540-547.
[480] MORIMOTO, R. I. Regulation of the heat shock transcriptional response: cross talk between a family of heat shock factors, molecular chaperones, and negative regulators. Genes Dev 12, 24 (1998 Dec 15), 3788-3796.
[481] MORRIS, N. R., AND ENOS, A. P. Mitotic gold in a mold: Aspergillus genetics and the biology of mitosis. Trends Genet 8, 1 (1992 Jan), 32-37.
[482] MORRIS-JONES, R., GOMEZ, B. L., DIEZ, S., !IRAN, M., MORRIS-JONES, S. D., CASADEVALI.. A., NOSANCHUK, J. D., AND HAMILTON, A. J. Synthesis of melanin pigment by Candida albicans in vitro and during infection. Infect Immun 73, 9 (Sep 2005), 6147-6150.
[483] MORSCIIIIAUSER. J.. MICHEL, S., AND STAIR, P. Sequential gene disruption in Candida albicans by FLP-mediated site-specific recombi-nation. Mel Microbial 32, 3 (May 1999), 547-556.
[484] MoSen, H. U., AND FINK, G. R. Dissection of filamentous growth by transposon mutagenesis in Sacchararnyces cerevisiae. Genetics 145, 3 (Mar 1997), 671-684.
14851 MOUTAOUAKIL, M., MONOD, M., PREVOST, XL C., BOUCHAHA, J. P., PARIS, S., AND LATGE, J. P. Identification of the 33-kDa alkaline protease of Aspergillus furnigatus in vitro and in vivo. J Med Microbial 39, 5 (Nov 1993), 393-399.
[486] MULLBACIIER, A., AND EICIINER, R. D. Immunosuppression in intro by a metabolite of a human pathogenic fungus. Proc Natl Acad Sex U S A 81, 12 (Jun 1984), 3835-3837.
[487] MULLER, H., HENNEQUIN, C., GALLAUD, J., DUJON, B., AND FAIRI1EAD, C. The asexual yeast Candida glabrata maintains distinct a and a haploid mating types. Eukaryot Cell 7,5 (May 2008), 848-858.
[488] MUNRO, C. A., SELVAGGINI, S., DE BRUME, I., WALKER, L., LENARDON, M. D., GERSSEN, B., MILNE, S., BROWN, A. J. P., AND Gow, N. A. R. The PKC, HOG and Ca2+ signalling pathways co-ordinately regulate chitin synthesis in Candida albicans. Mal Microbial 63, 5 (2007 Mar), 1399-1413.

REFERENCES 457
[489] MYER, V. E., AND YOUNG, R. A. RNA Polymerase II holoenzymes and subcomplexes. J Brod Chem 273, 43 (1998 Oct 23), 27757-27760.
[490] NAGAHASIII, S., LUSSIER, M., AND HUSSEY, H. Isolation of Candida glabrata homologs of the Saccharornyces at:rearm:re KRE9 and KNH1 genes and their involvement in cell wall beta-1,6-glucan synthesis. .1 Bacterial 180, 19 (Oct 1998), 5020-5029.
[491] NAGLIK, J. R., CHALLACOMBE, S. J., AND HUBE, B. Candida albicans secreted aspartyl proteinases in virulence and pathogenesis. Microbial Mel Biol Rev 67, 3 (2003 Sep), 400-428.
[492] NAKAYAMA, H., TANABE, K., BARD, M., HODGSON, W., WU, S., TAKEMORI, D., AOYAMA, T., KUMARASWAMI, N. S., METZLER, L., TAKANO, Y., CHIBANA, H., AND NIIMI, M. The Candida glabrata putative sterol transporter gene CgA USI protects cells against azoles in the presence of serum. J Antimicrob Chernother 60, 6 (2007 Dec), 1264-1272.
[493] NASCIMENTO, A. M., GOLDMAN, G. 1-1., PARK, S., MARRAS, S. A. E., DELMAS, G., OZA, U., LOLANS, K., Duta.Ev, M. N., MANN, P. A., AND PERLIN, D. S. Multiple resistance mechanisms among Aspergillus fumigates mutants with high-level resistance to itraconazole. Antimicrob Agents Chernother 47, 5 (May 2003), 1719-1726.
[494] NATIONAL COMMITTEE FOR CLINICAL LABORATORY STANDARDS. Reference method for broth dilution antifungal susceptibility testing of yeasts. proposed standard m27-p.
[495] NEWMAN, M. E. J., AND GIRVAN, NI. Finding and evaluating community structure in networks. Phys Rev E Stat Nonlin Soft Matter Phys 69, 2 Pt 2 (2004 Feb), 026113.
[496] NGUYEN, M. H., PEACOCK, J. E., MORRIS, A. J., TANNER, D. C., NGUYEN, M. L., SNYDMAN, D. R., WAGENER, M. M., RINALDI, M. G., AND YU, V. L. The changing face of candidemia: emergence of non-Candida athicans species and anti-fungal resistance. American Journal of Medicine 100, 6 (1996), 617-623.
[497] NalivEN, N. H., Suit, S.-0., AND BLACKWELL, M. Five novel Candida species in insect-associated yeast clades isolated from Neuroptera and other insects. Mycologia 99,6 (2007 Nov-Dec), 842-858.
[498] Ni. L., AND SNYDER, M. A genomic study of the bipolar bud site selection pattern in Saccharornyces cereviaiae. Mod Hied Cell 12, 7 (Jul 2001), 2147-2170.
[499] NIERMAN, W. C., PAIN, A., ANDERSON, M. J., WORTMAN, J. R., KIM, H. S., ARROYO, J., BERRIMAN, M., ABE, K., ARCHER, D. B., BERMEJO, C., BENNETT, J., BOWYER, P., CIIEN, D., COLLINS, M., COULSEN, a., DAVIES, R., DYER, P. S., FARMAN, M., FEDEROVA, N., FEDEROVA, N., FELDBLYUM, T. V., FISCIIER, R., FOSKER, N., FRASER, A., GARCIA, J. L., GARCIA, M. J., GOBLE, A., GOLDMAN, G. H., GOMI, K., GRIFFITIIS-JONES, S., GWILLIAM, R., HAAS, B., HAAS, H., HARRIS, D., HORIUCIII, H., HUANG, J., HUMPHREY. S., JIMENEZ, J.. KELLER, N., KHOURI, H., KITAMOTO, K., KOBAYASHI, T., KONZACK, S., KULKARNI, R., KUMAGI, T., LAFTON, A.. LATGE, J.-P., LI, W., LORD, A., LU, C., MAJOROS, W. H., MAY, G. S., MILLER, B. L., MOIIAMOUD, Y., MOLINA. M., MONOD, M., MOUYNA, I., MULLIGAN, S., MURPHY, L., O'NEtt., S., PAULSEN, I., PE, M. A., PERTEA, M., Putout:, C., PRITCIIARD, B. L., QUAIL, M. A., RABBINOWITSCH, E., RAWLINS, N., RAJANDREAM, M.-A., RECIIARD, U., RENAUD, H., ROBSON, G. D., RODRIGUEZ DE CORDOBA, S., NA, J. M. R.-P., RONNING, C. M., RUTTER, S., SALZBERG, S. L., SANCHEZ, M., SANCHEX-FIRIRERO, J. C., SAUNNDERS, D., SEEGER, K., SQUARES, Ft., SQUARES, S., TAKEUCHI, NI., TEKAIA, F., TURNER, G., VAZQUEZ DE ALDANA, C. R., WEIDMAN, J., WHITE, 0., WOODWARD, J., Yu, J.-H., FRASER, C., GALAGAN, J. E., ASAI, K., MACHIDA, M., HALL, N., DARRELL, B., AND DENNING, D. W. Genomic sequence of the pathogenic and allergenic filamentous fungus Aspergillus furnigatus. Nature 438 (December 2005), 1151-1156.
[500] NisinEAwA, A., MENDEZ, B., JIGANII, Y., AND DEAN, N. Identification of a Candida glabrata homologue of the S. cerevisicre VRC4 gene, encoding the Golgi GDP-mannose transporter. Yeast 19, 8 (Jun 2002), 691-698.
[501] NOBLE, D. Modeling the heart—from genes to cells to the whole organ. Science 295, 5560 (2002 Mar 1), 1678-1682.
[502] NOBLE, S. M., AND JOHNSON, A. D. Strains and strategies for large-scale gene deletion studies of the diploid human fungal pathogen Candida albicans. Eukaryot Cell 4, 2 (2005 Feb), 298-309.
[503] NOMEIR, A. A., PRAMANIK, R. N., HEIMARK, L., BENNETT, F., VEALS, J., BARTNER, P., HILBERT, M., SAKSENA, A., MCNAMARA, P., GIRIJAVALLAB-ILAN, V., GANGULY, A. K., LOVEY, R., PIKE, R., WANG, H., LIE, Y.-T., KUMARI, P., KORFMACIIER, W., LIN, C.-C., CACCIAPUOTI, A., LOEBENBERG, D., HARE, R., MILLER, G., AND PICKETT, C. Posaconazole (Noxafil, SCH 56592), a new azole antifungal drug, was a discovery based on the isolation and mass spectral characterization of a circulating metabolite of an earlier lead (SCH 51048). J Mass Spectrum 43, 4 (2008 Apr), 509-517.
[504] NORBECK, J., AND BLOMBERG, A. Metabolic and regulatory changes associated with growth of Saccharomyces cereeristae in 1.4 M NaCl. evidence for osmotic induction of glycerol dissimilation via the dihydroxyacetone pathway. J BIM Chem 272, 9 (1997 Feb 28), 5544-5554.
[505] NORBECK, J., PAIILMAN, A. K., AKIITAR, N., BLOMBERG, A., AND ADLER, L. Purification and characterization of two isoenzymes of DL-glycerol-3-phosphatase from Saccharontyces cerevisrae. identification of the corresponding GPP1 and GPP2 genes and evidence for osmotic regulation of Gpp2p expression by the osmosensing mitogen-activated protein kinase signal transduction pathway. J Butt Chem 271, 23 (Jun 1996), 13875-13881.
[506] NOSANCIIUK, J. D., VALADON, P., FELD:MESSER, M., AND CASADEVALL, A. Melanization of Cryptococcus neoforrnans in murine infection. Mel Cell Brod 19, 1 (Jan 1999), 745-750.

REFERENCES 458
[507] NUCCI, M., AND MARE, K. A. Emerging fungal diseases. Clin Infect Dis 41, 4 (2005 Aug 15), 521-526.
[508] NYGAARD, V., AND HOVIG, E. Options available for profiling small samples: a review of sample amplification technology when combined with microarray profiling. Nucleic Acids Res 34, 3 (2006), 996-1014.
[509] NYGAARD, V., LOLAND, A., HOLDEN, M., LANGAAS, M., RUE, H.. Lin, F., MYKLEBOST, 0., FODSTAD, 0., HOVIG, E., AND SMITH-SORENSEN, B. Effects of mRNA amplification on gene expression ratios in cDNA experiments estimated by analysis of variance. BMC Genentics
4, 1 (Mar 2003), 11.
[510) O'BRIAN, G. R., FAKHOURY, A. M., AND PAYNE, G. A. Identification of genes differentially expressed during aflatoxin biosynthesis in Aspergillus flavus and Aspergillus parasiticus. Fungal Genet Dint 39, 2 (Jul 2003), 118-127.
[511] ODDS, F. C., Gow, N. A. It., AND BROWN, A. J. P. Fungal virulence studies come of age. Genome Biology 2, 3 (2001), 1009.1-1009.4.
[512] OGAWA, N., AND OSIIIMA, Y. Functional domains of a positive regulatory protein, PHO4, for transcriptional control of the phosphatase regulon in Sacchammyces cerevisiae. Mol Cell Bird 10, 5 (1990 May), 2224-2236.
[513] O'GORMAN, C., FULLER, H., AND DYER, P. Discovery of a sexual cycle in the opportunistic fungal pathogen Aspergillus fumigatus. Nature (2008 Nov 30).
[514] OGRYDZIAK, D. M. Yeast extracellular proteases. Crit Rev Biotechnol 13, 1 (1993), 1-55.
[515] 01INO, S. Evolution by Gene Duplication. George Allen and Unwin, 1970.
[516] O'ROURKE, S. M., AND HERSKOWITZ, I. The Hogl MAPK prevents cross talk between the HOG and pheromone response MAPK pathways in Saccharcanyces cerevisiac. Genes Dev 12, 18 (1998 Sep 15), 2874-2886.
[517) O'ROURKE, S. M., AND HERSKOWITZ, I. A third osmosensing branch in Saccharomyces cerevissae requires the Msb2 protein and functions in parallel with the Shol branch. Mot Cell alai 22, 13 (2002 Jul), 4739-4749.
[518] O'ROURKE, S. M., AND HERSKOWITZ, I. Unique and redundant roles for HOG MAPK pathway components as revealed by whole-genome expression analysis. Mot Binl Cell 15, 2 (2004 Feb), 532-542.
[519] OSMANI, S. A., MAY, G. S., AND MORRIS, N. R. Regulation of the mRNA levels of nimA, a gene required for the 02-11/1 transition in Aspergillus nidulans. J Cell Biol 104, 6 (Jun 1987), 1495-1504.
[520] OSTERL011, A., AND BRELOER, M. Heat shock proteins: linking danger and pathogen recognition. Med Microbiol Immunol 197,1 (2008 Mar), 1-8.
[521] OSTERLOH, A., VEIT, A., GESSNER, A., FLEISCHER, B., AND BRELOER, M. Hsp60-mediated T cell stimulation is independent of TLR4 and IL-12. Int Immunol 20, 3 (2008 Mar), 433-443.
[522] PALLA, G., DERENYI, I., FARKAS, I., AND VICSEK, T. Uncovering the overlapping community structure of complex networks in nature and society. Nature 435, 7043 (2005), 814-818.
[523] PANACKAL, A. A., GRIBSKOV, J. L., STAAB, .1. F., KIRBY, K. A., 111NAtm, M., AND MAAR, K. A. Clinical significance of azole antifungal drug cross-resistance in Candida ttlabrata. Journal of Clinical Microbiology 44, 5 (2006), 1740-1745.
[524] PANEPINTO, J. C., OLIVER, B. G., FORTWENDEL, J. R., SMITH, D. L. H., ASKEW, D. S., AND RHODES, J. C. Deletion of the Aspergillus fumigatus gene encoding the Ras-related protein RhbA reduces virulence in a model of invasive pulmonary aspergillosis. Infect Innmun 71, 5 (May 2003), 2819-2826.
[525] PAPA, S., AND SKULACHEV, V. P. Reactive oxygen species, mitochondria, apoptosis and aging. Mol Cell Biochern 174, 1-2 (1997 Sep), 305-319.
[526] PARDO, J., URBAN, C., GALVEZ, E. M., EKERT, P. G., MULLER, U., KWON-CHUNG, J., LOBIGS, M., MULLBACHER, A., WALLICII, R,, HORNER, C., AND SIMON, NI. NI. The mitochondria[ protein Bak is pivotal for gliotoxin-induced apoptosis and a critical host factor of Aspergillus fumigatus virulence in mice. J Cell Biol 174, 4 (Aug 2006), 509-519.
[527] PARENICOVA, L., SKOUBOE, P., FRISVAD, J., SAMSON, R. A., ROSSEN, L., TEN HOOK-SUYKERBUYK, M., AND VISSER, S. Combined molecular and biochemical approach identifies Aspergillus japonicus and Aspergillus aculeatus as two species. Appl Ennimit Microbial 67, 2 (2001 Feb), 521-527.
[528] PARIS, S., DEBEAUPU1S,J.-P., CRAMER!, IL, CAREY, M., CHARLES, F., PREVOST, M. C., SCIIMITT, C., PHILIPPE, B., AND LATGE, J. P. Conidial hydrophobins of Aspergillus fumigatus. Appl Environ Microbiol 69, 3 (2003 Mar), 1581-1588.
[5291 PARK, H., MYERS, C. L., SHEPPARD, D. C., PIIAN, Q. T., SANCHEZ, A. A., E EDWARDS, J.. AND FILLER, S. G. Role of the fungal Ras-protein kinase A pathway in governing epithelial cell interactions during oropharyngeal candidiasis. Cell Microbiol 7, 4 (2005 Apr), 499-510.

REFERENCES 459
[530] PARRISII, J. K., VISCIDO, S. V., AND GRUNBAUM, D. Self-organized fish schools: an examination of emergent properties. Biol Bull 202, 3 (Jun 2002), 296-305.
[5311 PARBOIL J. L., ENLALRERT, B., AND FRANCOIS, J. STRE- and cAMP-independent transcriptional induction of Saccharomyces cerevisiae GS Y2 encoding glycogen synthase during diauxic growth on glucose. Yeast 15, 14 (1999 Oct), 1471-1484.
[532] PASCUAL-AHUIR, A., SERRANO, R., AND PROFT, M. The Skolp repressor and Gcn4p activator antagonistically modulate stress-regulated transcription in Saccharomyces ccrevisiac. Mol Cell Biol 21, 1 (2001 Jan), 16-25.
[533] PAULSSON, J. Summing up the noise in gene networks. Nature 427,6973 (Jan 2004), 415-418.
[534] PEL, H. J., DE WINDE. J. H., ARCHER. D. B., DYER, P. S., HOFMANN, G., SCIIAAI', P. J., TURNER, G., DE Vales, R. P., ALBANG, R., ALBERMANN, K., ANDERSEN, M. R., BENDTSEN, J. D., BENEN, J. A. E., VAN DEN BERG, M., BREESTRAAT, S., CADDICK, M. X., CONTRERAS, R., CORNELL, M., COUTINHO, P. M., DANCIIIN, E. G. J., DEBETS, A. J. M., DEKKER, P., VAN DUCK, P. W. M., VAN DIJK, A., DIJKHUIZEN. L., DRIESSEN, A. J. M., D'ENFERT, C., GEYSENS, S., GOOSEN, C., GROOT, G. S. P., DE GROOT, P. W. J., GUILLEMETTE, T., HENRISSAT, B., HEMMER, NI., VAN DEN HOMBERGII, J. P. T. W., VAN DEN HONDEL, C. A. M. J. J., VAN DER HEIJDEN, R. T. J. M., VAN DER KAAIJ, R. NI., KLIS, F. NI., KOOLS, H. J., KUBICEK, C. P., VAN KUYK, P. A., LAUBER, J., LU, X., VAN DER MAAREL, M. J. E. C., MEULENHERG, R., MENKE, H., MORTIMER, M. A., NIELSEN, J., OLIVER, S. G., OLSTIIOORN, M., PAL, K., VAN PELL N. N. M. E., RAM, A. F. J., RINAS, U., ROUBOS, J. A., SALT, C. M. J., SCIIMOLL, M., SUN, J., USSERY, D., VARGA, J., VERVECKEN, W., VAN DE VONDERVOORT, P. J. J., WEDLER, H., WOSTEN, H. A. B., ZENG, A.-P., VAN DOYEN, A. J. J., VISSER, J., AND STAM, H. Genome sequencing and analysis of the versatile cell factory Aspergillus niger cbs 513.88. Nat Biotechnol 25,2 (2007 Feb), 221-231.
[535] PEMBERTON, L. F., AND BLOBEL, G. Characterization of the Wtm proteins, a novel family of Saccharomyces cerevisiac transcriptional modulators with roles in meiotic regulation and silencing. Mol Cell Biol 17, 8 (1997 Aug), 4830-4841.
[536] PENALVA, M. A., TILBURN, .1., BIGNELI„ E., AND ARST, H. N. J. Ambient pH gene regulation in fungi: making connections. Trends Microbial 16, 6 (Jun 2008), 291-300.
[5371 PEPELINJAK. S., SLOBODNJAK, Z., SEGVIC, M., PERAICA, M., AND PAVLOVIC, NI. The ability of fungal isolates from human Lung aspergilloma to produce mycotoxins. Horn Exp Toxtcol 23, 1 (Jan 2004), 15-19.
[538] PEREZ, P., AND CALONGE, T. NI. Yeast protein kinase C. J Btochem 132, 4 (2002 Oct), 513-517.
[539] PERRIN, R. M., FEDOROVA, N. D., BOK, J. W., CRAMER, R. A., WORTMAN, J. It., KIM, H. S., NIERNIAN, W. C., AND KELLER, N. P. Transcriptional regulation of chemical diversity in Aspergillus furnigatus by LaeA, PLoS Pathog 3, 4 (Apr 2007), e50.
[5401 PETERSON, C. L., ZHAO, Y., AND CIIAIT, B. T. Subunits of the yeast SWI/SNF complex are members of the actin-related protein (ARP) family. J Biol Chem 273, 37 (1998 Sep 11), 23641-23644.
[5411 PFALLER, N. A., AND DIEKEMA, D. J. Rare and emerging opportunistic fungal pathogens: concern for resistance beyond Candida alhicans and Aspergillus furnigatus. J Olin. Microbial 42, 10 (2004 Oct), 4419-4431.
[542] PFALLER, M. A., AND DIEKEMA, D. J. Epidemiology of invasive candidiasis: a persistent public health problem. Clin Microbiol Rev 20, 1 (2007), 133-163.
[543] PFALLER, NI. A., DIEKEMA, D. J., RINALDI, M. G., BARNES, R., Hu, B., VESELOV, A. V., TIRABOSCHI, N., NAGY, E., GIBBS, D. L., AND GLOBAL ANTIFUNGAL SURVEILLANCE GROUP. Results from the ARTEMIS DISK global antifungal surveillance study: a 6.5-year analysis of susceptibilities of Candida and other yeast species to fluconazole and voriconazole by standardized disk diffusion testing. Journal of Clinical Microbiology 43, 12 (2005), 5848-5859.
[544] PFALLER, NI. A., MESSES, S. A., MILLS, K., BOLMSTROM, A., AND JONES, R. N. Evaluation of E-test method for determining posaconazole MICs for 314 clinical isolates of Candida species. J Clin Microbiol 39, 11 (2001 Nov), 3952-3954.
[545] PLA, 3., PEREZ-DIAZ, R. NI., NAvAtto-GAituiA, F., SANuiluz, NI., AND NOMBELA, C. Cloning of the Candida slbicans HIS1 gene by direct complementation of a C. albicans histidine auxotroph using an improved double-ARS shuttle vector. Gene 165, 1 (Nov 1995), 115-120.
[546] POLACHECK, I., PLATT, Y., AND ARONOVITCH, J. Catecholamines and virulence of Cryptococcus neoforrnans. Infect Emerson 58, 9 (Sep 1990), 2919-2922.
[5471 POMERENINCI, J. R., SONTAG, E. D., AND FERRELL, J. E. J Building a cell cycle oscillator: hysteresis and bistability in the activation of cdc2. Nat Cell Biol 5,4 (Apr 2003), 346-351.
[548] PONTECORVO, G.. ROPER, J. A., HEMMONS, L. NI., MACDONALD, K. D., AND BUFTON, A. W. J. The genetics of Asperyillus nidulans. Adv Genet 5 (1953), 141-238.
[549] PORTER, NI. A., ONNELA, J.-P., AND MUCHA, P. J. Communities in networks. Notices of the American Mathematical Society 56, 9 (2009), 1082-1097.
[550] POSAS, F., CHAMBERS, J. R., HEYMAN, J. A., HOEFFLER, J. P., DE NADAL, E., AND ARINO, J. The transcriptional response of yeast to saline stress. J Dial Chem 275, 23 (2000 Jun 9), 17249-17255.

REFERENCES 460
[551] POSAS, F., AND SAITO, H. Osmotic activation of the HOG MAPK pathway via Stellp MAPKKK: scaffold role of Pbs2p MAPKK. Science 276, 5319 (1997 Jun 13), 1702-1705.
[552] POSAS, F., AND SAITO, H. Activation of the yeast SSK2 MAP kinase kinase kinase by the SSK1 two-component response regulator. EMBO J 17, 5 (1998 Mar 2), 1385-1394.
[553] POSAS, F., WURGLER-MURPHY, S. M., MAEDA, T., WITTEN, E. A., THAI, T. C., AND SAITO, H. Yeast HOG1 MAP kinase cascade is regulated by a multistep phosphorelay mechanism in the SLN1-YPD1-SSK1 "two-component" osmosensor. Cell 86, 6 (1996 Sep 20), 865-875.
[554] POWERS, T., DILOVA, L, CIIEN, C. Y., AND WEDAMAN, K. Yeast TOR signaling: a mechanism for metabolic regulation. Cure Top Microbial Immunol 279 (2004), 39-51.
[555] PRAIILAD, V., AND MORIMOTO, R. I. Integrating the stress response: lessons for neurodegenerative diseases from C. eteaans. Trends Cell Biol 19, 2 (Feb 2009), 52-61.
[556] PRASAD, T., CIIANDRA, A., MUKHOPAIIIIYAY, C. K., AND PRASAD, R. Unexpected link between iron and drug resistance of Candida spp.: iron depletion enhances membrane fluidity and drug diffusion, leading to drug-susceptible cells. Antirntcrob Agents Chemother 50, 11 (2006 Nov), 3597-3606.
[557] PROFT, M., PASCUAL-AHUIR, A., DE NADAL, E., ARINO, J., SERRANO, R., AND POSAS, F. Regulation of the Skol transcriptional repressor by the Hogl MAP kinase in response to osmotic stress. EMBO J 20, 5 (2001 Mar 1), 1123-1133.
[558] PROFT, M., AND SERRANO, R. Repressors and upstream repressing sequences of the stress-regulated ENA1 gene in Saccharomvces cerevisiae: bZIP protein Skolp confers HOG-dependent osmotic regulation. Mol Cell Bird 19, 1 (1999 Jan), 537-546.
[559] PTACEK, J., DEVGAN, G., MICHAUD, G., Znu, H., Zoo, X., FASOLO, J., Guo, H., JONA, G., HREITKREUTZ, A., SOPKO, R., MCCARTNEY, R. R., SCHMIDT, M. C., RACIIIDI, N., LEE, S.-J., MAIL, A. S., MENG, L., STARK, M. J. R., STERN, D. F., DE VIRGILIO, C., TYERS, M., ANDREWS, B., GEFLSTEIN, M., SCHWEITZER, B., PREDKI, P. F.. AND SNYDER, M. Global analysis of protein phosphorylation in yeast. Nature 438,7068 (2005 Dec 1), 679-684.
[560] PUIG, S., LAU, M., AND TIIIELE, D. J. Cti6 is an Rpd3-Sin3 histone deacetylase-associated protein required for growth under iron- limiting conditions in Saccharomyces cerevisiae. J Biol Chem 279, 29 (2004 Jul 16), 30298-30306.
[561] PUNT, P. J., OLIVER, R. P., DINGEMANSE, M. A., POUWELS, P. H., AND VAN DEN BONDEL, C. A. M. J. J. Transformation of Aspergillus based on the hygromycin B resistance marker from Escherichia coli. Gene (56), 117-124.
[562] R DEVELOPMENT CORE TEAM. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria, 2007. ISBN 3-900051-07-0.
[5631 RAITT, D. -0IINSON, A. L., ERKINE, A. M., MAKING, K., MORGAN, B., GROSS, D. S., AND JOHNSTON, L. H. The Skn7 response regulator of Saccharornyces cerevisiae interacts with Hsfl in vivo and is required for the induction of heat shock genes by oxidative stress. Mot Biol Cell 11, 7 (2000 Jul), 2335-2347.
[564] RAITT, D. C., POSAS, F., AND SAITO, H. Yeast Cdc42 GTPase and Ste20 PAK-like kinase regulate Shol-dependent activation of the Hogl MAPK pathway. EMBO J 19, 17 (2000 Sep 1), 4623-4631.
[565] RAM, A. F. J., AND KLIS, F. M. Identification of fungal cell wall mutants using susceptibility assays based on calcofluor white and congo red. Nat Protoc 1, 5 (2006), 2253-2256.
[566] RAmAciiAxtutms, H., BANERJEE, B., GREENBERGER, P. A., KELLY, K. J., FINK, J. N., AND KURIA', V. P. Role of C-terminal cysteine residues of Aspergillus furnigatus allergen Asp f 4 in immunoglobulin E binding. Min Diagn Lab Immunol 11, 2 (Mar 2004), 261-265.
(567] RAMAGE, G., VANDE \VALLE, K., WICKES, B. L., AND LOPEZ-11180T, J. L. Biofilm formation by Candida dubl niensis. J Clin M crobiot 39, 9 (2001 Sep), 3234-3240.
[568] RAMIREZ, F., SCHLICKER. A., ASSENOV, Y., LENGAUER, T., AND ALBRECHT, M. Computational analysis of human protein interaction networks. Proteomics 7,15 (2007 Aug), 2541-2552.
[569] BATUSFINY, A. V., RAMSEY, S. A., BODA, 0., WAN, Y., SMITH, J. J., AND AITCHISON, J. D Control of transcriptional variability by overlapping feed-forward regulatory motifs. Biophys J 95, 8 (2008 Oct), 3715-3723.
[570] RAYNER, A. D. M., AND GRIFFITHS, G. S. The growing fungus. Chapman and Hall, 1994, ch. Mycelial interconnectedness.
[571] REICHARD, U., BUTTNER, S., EIFFERT, H., STAIB, F., AND BUCHEL, R. Purification and characterisation of an extracellular serine proteinase from Aspergillus furnigatus and its detection in tissue. J Med Microbial 33, 4 (Dec 1990), 243-251.
[572] REICHARD, U., EIFFERT, H., AND RUCHEL, R. Purification and characterization of an extracellular aspartic proteinase from Aspergillus fumigates. J Med Vet Mycol 32, 6 (Dec 1994), 427-436.

REFERENCES 461
[573] REiciimin, U., MONOD, M., ODDS, F., AND RUCHEL, R. Virulence of an aspergillopepsin-deficient mutant of Aspergillus fumigates and evidence for another aspartic proteinase linked to the fungal cell wall. J Med Vet Mycol 35, 3 (May-Jun 1997), 189-196.
[574] REICHARD, U., MONOD, M., AND MICHEL, R. Molecular cloning and sequencing of the gene encoding an extracellular aspartic proteinase from Aspergillus furnigatus. FEMS Microbiol Lett 130, 1 (Jul 1995), 69-74.
[575] REISER, V., RAITT, D. C., AND SAITO, H. Yeast osmosensor Slnl and plant cytokinin receptor Crel respond to changes in turgor pressure. J Cell Biol 161, 6 (2003 Jun 23), 1035-1040.
[576] REMENTERIA, A., LOPEZ-MOLINA, N., LUDWIG, A., VIVANCO, A. D., BIKANDI, J., PONTON, J., AND GARAIZAR, J. Genes and molecules involved in Aspergillus funeigatus virulence. Rev lberoarn Micol 22, 1 (Mar 2005), 1-23.
[577] REN, B., ROBERT, F., WYRICK, J. J., APARICIO, 0., JENNINGS. E. G., SIMON, I., ZEITLINGER, J., SCHREIBER, J., HANNETT, N., KANIN, E., VOLKERT, T. L., WILSON, C. J., But., S. P., AND YOUNG, R. A. Genome-wide location and function of DNA binding proteins. Science 290, 5500 (2000 Dec 22), 2306-2309.
[578] REP, M., KRANTZ, M., THEVELEIN, J. M., AND HOIIMANN, S. The transcriptional response of Saccharomyces cerevisiae to osmotic shock. Hot1p and Msn2p/Msn4p are required for the induction of subsets of high osmolarity glycerol pathway-dependent genes. J Biol Chem 275, 12 (2000 Mar 24), 8290-8300.
[579] REP, M., PROFT, M., REMIZE, F., TAMAS, M., SERRANO, R., THEVELEIN, J. M., AND HOFMANN, S. The Saccharomyces ceremsioe Skolp transcription factor mediates HOG pathway-dependent osmotic regulation of a set of genes encoding enzymes implicated in protection from oxidative damage. Mol Microbiol 40, 5 (2001 Jun), 1067-1083.
[580] REP, M., PROFT, M., REMIZE, F., TAMAS, M., SERRANO, R., THEVELEIN, J. M., AND HOHMANN, S. The Saccharomyces corevismo Skolp transcription factor mediates HOG pathway-dependent osmotic regulation of a set of genes encoding enzymes implicated in protection from oxidative damage.skolp transcription factor mediates hog pathway-dependent osmotic regulation of a set of genes encoding enzymes implicated in protection from oxidative damage. Mol Microbiol 40, 5 (2001 Jun), 1067-1083.
[581] REP, M., REISER, V., GARTNER, U., THEVELEIN, 1. M., HOHNIANN, S., ANIMERER, G., AND RUIs, H. Osmotic stress-induced gene expression in Saccharionyces GCVO/2526Q requires Msnlp and the novel nuclear factor Hot1p. Mai Cell Blot 19, 8 (1999 Aug), 5474-5485.
[582] REUSS, 0., VIK, A., KOLTER, R., AND MORSCHHAUSER, J. The SAT1 flipper, an optimized tool for gene disruption in Candida a/bican.s. Gene 341 (Oct 2004), 119-127.
1583] REX, J. H., BENNETT, J. E., GALLIN, J. I., MALECH, H. L., DECARLO, E. S., AND MELNICK, D. A. In vivo interferon-gamma therapy augments the in vitro ability of chronic granulomatous disease neutrophils to damage Aspergillus hyphae. Infect Die 163, 4 (1991 Apr), 849-852.
[584] RHODES, J. C. Aspergillus furnigatus: growth and virulence. Med Mycol 44 Suppl 1 (2006 Sep), S77-81.
[585] RHODES, J. C., BODE, R. B., AND MCCUAN-KIRSCH, C. M. Elastase production in clinical isolates of Aspergillus. Diagn Microbiol Infect Dis 10, 3 (Jul 1988), 161-170.
[586] RHODES, J. C., POLACHECK. I., AND KWON-CHUNG, K. J. Phenoloxidase activity and virulence in isogenic strains of Cryptococcus neoforrnans. Infect Immun 36, 3 (Jun 1982), 1175-1184.
[587] RIVERA, M. C., JAIN, R., MOORE, J. E., AND LAKE, J. A. Genomic evidence for two functionally distinct gene classes. Proc Nati Acad Sci U S A 95, 11 (1998 May 26), 6239-6244.
[588] RODRIGUEZ-TUDELA, J. L., ARENDRUP, M. C., CUENCA-ESTRELLA, M., DONNELLY, J. P., AND LASS-FLORL, C. Eucast breakpoints for antifun-gals. Drug News Perspect 23, 2 (2010 Mar), 93-97.
[589] ROETZER, A., GREGORI, C., JENNINGS, A. M., QUINTIN, J., FERRANDON, D., BUTLER, G., KUCIILER, K., AMMERER, G., AND SCIIULLER, C. Candida gtahroto environmental stress response involves Saccharomyces cnverrisine Msn2/4 orthologous transcription factors. Mel Microbiol 69, 3 (2008 Aug), 603-620.
[590] ROMAN, T., TUNCUZ-ZAWISLAK, T.-J., SCHEFFER, S. G., AND HARMSEN, J. Network node ad targeting. US Patent and Trademark Office. Patent application 20080162260, July 2008.
[591] Bonus, M., ALBERT, NI., KELLER, N. P. AND KEMPKEN, F. Secondary chemicals protect mould from fungivory. Riot Lett 3, 5 (Oct 2007), 523-525.
[592] ROKAS, A., AND HITTINGER, C. T. Transcriptional rewiring: the proof is in the eating. Carr Biol 17, 16 (2007 Aug 21), R626-8.
[593] ROMAN, E., ARANA, D. M., NOMBELA, C., ALONSO-MONDE, R., AND PLA, J. Map kinase pathways as regulators of fungal virulence. Trends Microbiol 15, 4 (2007 Apr), 181-190.
[594] ROSAS, A. L., NOSANCHUK, J. D., FELDMESSER, NI., Cox, G. M., MCDADE, H. C., AND CASADEVALL, A. Synthesis of polymerized melanin by Cryptococcus neoformans in infected rodents. Infect Irnmun 68, 5 (May 2000), 2845-2853.

REFERENCES 462
[595] ROSS-MACDONALD, P., COELIIO, P. S. R., ROEMER, T., AGARWAL, S., KUMAR, A., JANSEN, R., CHEUNG, K.-H., SIIEEIIAN, A., SYMONIATIS, D., UMANSKY, L., HEIDTMAN, M., NELSON, F. K., IWASAKI, H., HALER, K., GERSTEIN, M., MILLER, P., ROEDER, G. S., AND SNYDER, M. Large-scale analysis of the yeast genome by transposon tagging and gene disruption. Nature 402, 413-418 (1999).
[596] RUBIN-BEIERANO, I., FRASER, I., GRISAFI, P., AND FINE, G. R. Phagocytosis by neutrophils induces an amino acid deprivation response in Saccharomyces cerevisiac and. Candida albicuns. Proc Nati Acad Sci U S A 100, 19 (Sep 2003), 11007-11012.
[597] RUCIIEL, FL, ZIMMERMAN, F., BONINC-STUTZER, B., AND HELMCIIEN, U. Candidiasis visualised by proteinase-directed immmunofluores-cence. Virchows Archive A: Pathological Anatomy and Histopathology 419 (1991), 199-202.
[598] RUCKENSTUHL, C., EIDENBERGER, A., LANG, S., AND TURNOWSKY, F. Single amino acid exchanges in FAD-binding domains of squalene epoxidase of Saccharomyces cerevistae lead to either loss of functionality or terbinafine sensitivity. Burchent Sae Trans 33, Pt 5 (2005 Nov), 1197-1201.
[599] SABATELLI, F., PATEL, R., MANN, P. A., MENDIUCK, C. A., Norms, C. C., HARE, R., LOEHENDERG, D., BLACK, T. A., AND MeNicooLAN, P. M. In vitro activities of posaconazole, fluconazole, itraconazole, voriconazole, and amphotericin B against a large collection of clinically important molds and yeasts. Antincicrob Agents Chentother 50, 6 (2006 Jun), 2009-2015.
[600] SAIER, M. H. J., BEATTY, J. T., GOFFEAU, A., HARLEY, K. T., HEIJNE, W. H., HUANG, S. C., JACK, D. L., JAIIN, P. S., LEW, K., LW, J.. PAO, S. S., PAULSEN, I. T., TSENG, T. T., AND VIRK, P. S. The major facilitator superfamily. J Mol Microbiol Biotechnol 1, 2 (1999 Nov), 257-279.
[601] SAIKI, R. K., GELFAND, D. H., STOFFEL, S., SCHARF, S. J., HIGUCIII, R., HORN, G. T., Mums, K. B., AND EytAco, H. A. Primer-directed enzymatic amplification of DNA with a thermostable DNA polymerase. Science 239, 4839 (1988), 487-491.
[602] SALAS, S. D., BENNETT, J. E., KtvoN-CfluNc, K. J., PERFECT, J. R., AND WILLIAMSON, P. R. Effect of the lactase gene CNLAC1, on virulence of Crypt OCOCCRI, neoformans. J Exp Med 184, 2 (Aug 1996), 377-386.
[603] SAMBROOK, J., AND RUSSELL, D. W. Molecular cloning: a Laboratory Manual, vol. 1. Cold Spring Harbor Laoratory press, 2001.
[604] SANATI, H., BELANGER, P., PRATT', R., AND GIIANNOUM, M. A new triazole, voriconazole (UK-109,496), blocks sterol biosynthesis in Candida albicans and Candida krusei. Antirnicroh Agents Chernother 41, 11 (1997 Nov), 2492-2496.
[605] SANATI, H., RAMOS, C. F., BAYER, A. S., AND GIIANNOUM, M. A. Combination therapy with ampotericin B and fluconazole against invasive candidiasis in neutropenic-mouse and infective-endocarditis rabbit models. Antunicrobial Agents and Chemotherapeutics 41, 6 (1997), 1345-1348.
[606] SANGLARD, D., 'SCHER, F., CALABRESE, D., MAJCIIERCZYK, P. A., AND Blue, J. The ATP binding cassette transporter gene CgCDR1 from Candida giabrata is involved in the resistance of clinical isolates to azole antifungal agents. Antimicrobial agents and chemotherapy 43, 11 (November 1999), 2753-2765.
[607] SANTANGELO, G. M. Glucose signaling in Saccharomyces cerevisiae. Mmrobiol Mol Biol Rev 70, 1 (2006 Mar), 253-282.
[608] SANTOS, M. A., AND TUITE, M. F. The CUG codon is decoded in vivo as serine and not leucine in Candida albicans. Nucleic Acids Research 23 (1995), 1481-1486.
[609] SATO, N., KAWAHARA, H., TOFI-E, A., AND MAEDA, T. Phosphorelay-regulated degradation of the yeast Ssklp response regulator by the ubiquitin-proteasome system. Mol Cell Biol 23, 18 (2003 Sep), 6662-6671.
[610] SCHADT, E. E. Molecular networks as sensors and drivers of common human diseases. Nature 461, 7261 (2009 Sep 10), 218-223.
[611] SCHAFFNER, A., DOUGLAS, II., AND BRAUDE, A. Selective protection against conidia and against mycelia by polymorphonuclear phagocytes in resistance to Aspergillus. observations on these two lines of defense in vivo and in vitro with human and mouse phagocytes. J Clin Invest 69, 3 (Mar 1982), 617-631.
[612] SCHALLER, M., REIN, M., KORTING, H. C., BAUR, S., HAMM, G., MONOD, NI., BEINHAUER, S., AND HUBS, B. The secreted aspartyl proteinases Sap]. and Sap2 cause tissue damage in an in vitro model of vaginal candidiasis based on reconstituted human vaginal epithelium. Infect /moray 71, 6 (2003 Jun), 3227-.3234.
[613] SCHELL W. A., DE ALMEIDA, G. M., DODGE, R. K., OKONOGI, K., AND PERFECT, J. R. In vitro and in vivo efficacy of the triazole tak-187 against cryptococcus neoformans. Arctimicrob Agents Chernother 42,10 (1998 Oct), 2630-2632.
[614] SCIIERENS, B., AND GOFFEAU, A. The uses of genome-wide yeast mutant collections. Genome Biology 5 (2004), 229.
[615] SCHMITT, A. P., AND MCENTEE, K. Msn2p, a zinc finger DNA-binding protein, is the transcriptional activator of the multistress response in Saccharomyces cerevisian. Pro,: Natl Acad Sei U S A 93, 12 (1996 Jun 11), 5777-5782.
[616] Smorr, H. J., BLEVINS, A., SOBECK, K., AND ARMSTRONG, D. Aspergillus species from hospital air and from patients. Mycoses .93, 11-12 (1990 Nov-Dec), 539-541.

REFERENCES 463
[617] SCHOEBERL, B., PACE, E. A., FITZGERALD, J. B., HARMS, B. D., XU, L., N1E, L., LINGGI, B.. KALRA, A., PARAGAS, V., BL'KIIALID, R., GRANTCIIAROVA, V., KOHLI, N., WEST, K. A., LESZCZYNIECKA, M., FELDHAUS, M. J., KL'DLA, A. J., AND NIELSEN, U. B. Therapeutically targeting ErbB3: a key node in ligand-induced activation of the ErbB receptor-PI3K axis. Sci Signal 2, 77 (2009), ra31.
[618] SCIIRETTL, M., BIGNALL, E., KRAGL, C., JOECIIL, C., ROGERS, T., JR, H. N. A., HAYNES, K., AND HAAS, H. Siderophore biosynthesis but not reductive iron assimilation is essential for Aspergillus fumigatus virulence. Journal of Experimental Medicine 200, 9 (November 2004), 1213-1219.
[619] SCIIRETTL, M., BIGNELL, E., KRAGL, C., SABIIIA, Y., Loss, 0., EISENDLE, M., WALLNER, A., ARST, H. N. J., HAYNES, K., AND HAAS, H. Distinct roles for intra- and extracellular siderophores during Aspergillus fumigates infection. PLoS Pathog 3, 9 (Sep 2007), 1195-1207.
[620] SCHWEIZER, A., RUPP, S., TAYLOR, B. N., ROLLINGHOFF, M., AND SCHROPPEL, K. The TEA/ATTS transcription factor CaTeclp regulates hyphal development and virulence in Candida albicans. Mol Microbiol 38, 3 (2000 Nov), 435-445.
[621] SENEV1RATNE, C. J., „RN, L., AND SAMARANAYAKE, L. P. Biofilm lifestyle of Candida: a mini review. Oral Dis 14, 7 (2008 Oct), 582-590.
[6221 SETIADI, E. R., DOEDT, T., COTTIER, F., NOFFZ, C., AND ERNST, J. F. Transcriptional response of Candida albicans to hypoxia: linkage of oxygen sensing and Efglp-regulatory networks. J Mol Biol 361, 3 (2006 Aug 18), 399-411.
[6231 SHAH ALAM BIIUIYAN, M., ECKSTEIN, J., BARBUCH, R., AND BARD, M. Synthetically lethal interactions involving loss of the yeast ERG24: the sterol C-14 reductase gene. Lipids 42, 1 (2007 Feb), 69-76.
[6241 SHANNON, P., MARKIEL, A., OZIER, 0., BALICA, N. S., WANG, J. T., RAMAGE, D., AMIN, N., SCIIWIKOWSKI, B., AND IDEKER, T. Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Res 13, 11 (2003), 2498-2504.
[625] SIIARAN, R., UL1TSKY, I., AND SIIAMIR, R. Network-based prediction of protein function. Mol Syst Biol 3 (2007), 88.
[626] SHARKEY, L. L., LIAO, W.-L., GHOSII, A. K., AND FONZI, W. A. Flanking direct repeats of hisG alter URA3 marker expression at the HIVP1 locus of Candida albicans. Microbiology 151, Pt 4 (2005 Apr), 1061-1071.
[627] SHAW, P. J., TONON1, G., GREENSPAN, R. J., AND ROBINSON, D. F. Stress response genes protect against lethal effects of sleep deprivation in Drosophila. Nature 417, 6886 (May 2002), 287-291.
[628] SIIEN, J., GUO, W., AND KOHLER, J. R. CaNAT1, a heterologous dominant selectable marker for transformation of Candida albicans and other pathogenic Candida species. Infect Immure 73, 2 (Feb 2005), 1239-1242.
[629] SIIEN-ORIL S. S., M11,0, R., MANGAN, S., AND ALON, U. Network motifs in the transcriptional regulation network of E.schcrichia coli. Nat Genet 31, 1 (2002), 64-68.
[6301 SHERMAN, D., DL'RRENS, P., IRACNE, F., BEYNE, E., NIKOLSKI, M., AND SOL'CIET, J.-L. Genolevures complete genomes provide data and tools for comparative genomics of hemiascomycetous yeasts. Nucleic Acids Res 34, Database issue (2006 Jan 1), D432-5.
[631] SIIINODA, J., AND KIKUCHI, Y. Rodl, an arrestin-related protein, is phosphorylated by Snfl-kinase in Saccharomyees cerevisiae. Biochem Biophys Res Common 364, 2 (Dec 2007), 258-263.
[632] SHINOHARA, C., HASUMI, K., AND ENDO, A. Inhibition of oxidized low-density lipoprotein metabolism in macrophage J774 by helvolic acid. Biochim Biophy.s Acta 1167, 3 (Apr 1993), 303-306.
[633] SHOCK, T., THOMPSON, J., YATES JR, R., AND MADIIANI, H. Hogl MAP kinase interrupts signal transduction between the Kssl MAP kinase and the Teel transcription factor to maintain pathway specificity. Enkaryot Cell (2009 Feb 13).
[634] SHOEMAKER, D. D., LASIIKARI, D. A., MORRIS, D., M1TTNIANN, M., AND DAVIS, R. W. Quantitative phenotypic analysis of yeast deletion mutants using a highly parallel molecular bar-coding strategy. Nat Genet 14, 4 (1996 Dec), 450-456.
[635] SIMON, H. A. The architecture of complexity. Proceedings fo the American Philosophical Society 106, 6 (December 1962), 467-482.
[636] SIRAKOVA, T. D., MARKARYAN, A., AND KOLATTUKUDY, P. E. Molecular cloning and sequencing of the cDNA and gene for a novel elastinolytic metalloproteinase from Aspergillus fumigatus and its expression in Escherichm coli. Infect Immure 62, 10 (Oct 1994), 4208-4218.
[637] SLAUCH, J. M., MAIIAN, M. J., AND MEKANOLOS, J. J. In vivo expression technology for selection of bacterial genes specifically induced in host tissues. Methods in Enzymology 235 (1994), 481-492.
[638] SMITH. A., WARD, M. P.. AND GARRETT, S. Yeast PKA represses Msn2p/Msn4p-dependent gene expression to regulate growth, stress response and glycogen accumulation. EMBO J 17, 13 (1998 Jul 1), 3556-3564.
[639] SMITH, D. A., NICHOLLS, S., MORGAN, B. A., BROWN, A. J. P., AND QUINN, J. A conserved stress-activated protein kinase regulates a core stress response in the human pathogen Candida albicans. Mol 131o1 Cell /5,9 (Sep 2004), 4179-4190.
[640] SMITH, J. M., TANG, C. M., VAN NOORDEN, S.. AND HOLDEN, D. W. Virulence ofAspergillus fumigatus double mutants lacking restriction and an alkaline protease in a low-dose model of invasive pulmonary aspergillosis. Infect Immun 62, 12 (Dec 1994), 5247-5254.

REFERENCES 464
[641] SMITH, R. L., AND JOHNSON, A. D. Turning genes off by Ssn6-Tupl: a conserved system of transcriptional repression in eukaryotes. Trends 13 .ochent Sci 25, 7 (2000 Jul), 325-330.
[642] SMYTH, G. K. Linear models and empirical bayes methods for assessing differential expression in microarray experiments. Stat Appl Genet Mol Biol 3 (2004), Article3.
[643] SMYTH, G. K. Bioinforsnatics and Computational Biology Solutions using R and bioconductor. Springer, New York, 2005, ch. LIMMA: linear models for microarray data, pp. 397-420.
[644] SMYTH, G. K., MICHAUD, J., AND SCOTT, H. S. Use of within-array replicate spots for assessing differential expression in microarray experiments. Bioinformaties 21, 9 (2005), 2067-2075.
[645] SOBEL, J. D. Vulvovaginitis. When Candida becomes a problem. Dermatol Clin 16, 4 (1998 Oct), 763-768.
[646] SOLLNER, S., SCHOBER, M., WAGNER, A., PREM, A., LORKOVA, L., PALFEY, B. A., GROLL, M., AND MACIIEROUX, P. Quinone reductase acts as a redox switch of the 20s yeast proteasome. EMBO Rep 10, 1 (2009 Jan), 65-70.
[647] SOONG, T. W., YONC, T. F., RAMANAN, N., AND WANG, Y. The Candida albicans antiporter gene CNH1 has a role in Na+ and H+ transport, salt tolerance, and morphogenesis. Microbiology 146 ( Pt 5) (2000 May), 1035-1044.
[648] SORGER, P. K. A reductionist's systems biology: opinion. Curr Opin Cell Biol 17, 1 (2005 Feb), 9-11.
[649] SORIANI, F. M., MALAVAZI, I., DA SILVA FERREIRA, M. E., SAVOLDI, M., VON ZESKA KRESS, M. R., DE SOUZA GOLDMAN, M. H., LOSS, O.. BIGNELL, E., AND GOLDMAN, G. H. Functional characterization of the Aspergillus fumigatus CRZ1 homologue, CrzA. Mol Microbiol 67, 6 (Mar 2008), 1274-1291.
[650] SORIANI, F. M., MARTINS, V. P., MAGNANI, T., TUDELLA, V. G., CURTI, C., AND UYEMURA, S. A. A PMR/ -like calcium ATPase of Asp:minus fumigatus: cloning, identification and functional expression in S. eerevisiae. Yeast 22, 10 (Jul 2005), 813-824.
[651] SPANAKIS, E. K., APERIS, G., AND MYLONAKIS, E. New agents for the treatment of fungal infections: clinical efficacy and gaps in coverage. Clin Infect Dis 43, 8 (Oct 2006), 1060-1068.
[652] SPIKES, S., XL', R., NGUYEN, C. K., CIIAMILOS, G., KONTOYIANNIS, D. P., JACOBSON, R. H., EJZYKOWICZ, D. E., CHIANG, L. Y., FILLER, S. G., AND MAY, G. S. Gliotoxin production in Asperyillus fmnigatus contributes to host-specific differences in virulence. J Infect Dis 197, 3 (Feb 2008), 479-486.
[653] SPREER. A., RUCIIEL, R., AND REICHARD, U. Characterization of an extracellular subtilisin protease of Rhizopus inicrosporus and evidence for its expression during invasive rhinoorbital mycosis. Med Mycot 44, 8 (Dec 2006), 723-731.
[654] SPRING, J. Vertebrate evolution by interspecific hybridisation—are we polyploid? FEBS Lett 400, 1 (1997), 2-8.
[655] STAAB, J. F., HEADWAY, S. D., FIDEL, P. L., AND SUNDSTROM, P. Adhesive and mammalian transglutaminase substrate properties of Candida albicans Hwp1. Science 283 (1999), 1535-1538.
[656] STAAB, J. F., FERRER, C. A., AND SUNDSTROM, P. Developmental expression of a tandemly repeated, proline-and glutamine-rich amino acid motif on hyphal surfaces on Candida albicans. Dial Chem 271, 11 (1996 Mar 15), 6298-6305.
[657] STANZANI, M., ORCIULO, E., LEWIS, R., KANTOYIANNIS, D. P., DIMITRIOS, P., MARTINS, S. L. R., ST JOHN, L. S., AND KOMANDURI, K. V. Aspergillus fumigatus suppresses the human cellular immune response via gliotoxin-mediated apoptosis of monocytes. Blood 105, 6 (2005 Mar 15), 2258-2265.
[658] STATFIOPOULOS-GERONTIDF.S, A., Guo, J. J AND OVERT, M. S. Yeast calcineurin regulates nuclear localization of the Crzlp transcription factor through dephosphorylation. Genes Dev 13, 7 (1999 Apr 1), 798-803.
[659] STEINMETZ. L. M., SCIIARFE, C., DEUTSCHBAUER, A. M., MOKRANJAC, D., HERMAN, Z. S., JONES, T., Clic, A. M., GIAEVER, G., PROKISCH, H., OEFNER, P. J., AND DAVIS, R. W, Systematic screen for human disease genes in yeast. Nat Genet 31, 4 (Aug 2002), 400-404.
[660] STEVENS, D. A., KAN, V. L., JUDSON, M. A., MORRISON, V. A., DUMMER, S., DENNING, D. W., BENNETT, J. E., WALSH, T. J., PATTERSON, T. F., AND PANKEY, G. A. Practice guidelines for diseases caused by Aspemillus. Infectious Diseases Society of America. Clin Infect Dis 30, 4 (2000 Apr), 696-709.
[661] STIBIUS, K. B., AND SNEPPEN, K. Modeling the two-hybrid detector: experimental bias on protein interaction networks. Biophys J 93, 7 (2007 Oct 1), 2562-2566.
[662] STOCK, A. M., ROBINSON, V. L., AND GOUDREAU, P. N. Two-component signal transduction. Annu Rev Biochem 69 (2000), 183-215.
[663] STOLDT, V. R., SONNEBORN, A., LEUKER, C. E., AND ERNST, J. F. Efglp, an essential regulator of morphogenesis of the human pathogen Candida albicans, is a member of a conserved class of bHLH proteins regulating morphogenetic processes in fungi. EMBO J 16, 8 (Apr 1997), 1982-1991.

REFERENCES 465
[664] STRANGE, K., EMMA, F., PAREDES, A., AND MORRISON, R. Osmoregulatory changes in myo-inositol content and Na+f myo-inositol cotransport in rat cortical astrocytes. Clio 12, 1 (1994 Sep), 35-43.
[665] STROGATZ, S. H. Exploring complex networks. Nature 410, 6825 (2001 Mar 8), 268-276.
[666] STUMPF, M. P. H., WWI', C., AND MAY, R. M. Subnets of scale-free networks are not scale-free:. Proceedings of the National Academy of Sciences 102, 12 (March 2005), 4221-4224.
[667] SUDBERY, P. E. The germ tubes of Candida albicans hyphae and pseudohyphae show different patterns of septin ring localization. Mal Microbial 41, 1 (2001 Jul), 19-31.
[668] SUGAR, A. M. Antimicrobial agents, vol. 1. Elsevier Science Publishers, Amsterdam, The Netherlands, 1986, ch. The polyene macrolide antifungal drugs, pp. 229-244.
[669] SUGUI, J. A., PARDO, J., CIIANG, Y. C., MULLBACHER, A., ZAREMBER, K. A., GALVEZ, E. M., BRINSTER, L., ZERFAS, P., GALLIN, J. I., SIMON, M. M., AND KWON-CHUNG, K. J. Role of laeA in the Regulation of albl, gliP, conidial morphology, and virulence in Aspergillus fumigator. Eukaryot Cell 6, 9 (2007 Sep), 1552-1561.
[670] Scout, J. A., PARDO, J., CIIANG, Y. C., ZAREMBER, K. A., NARDONE, G.. GALVEZ, E. M., MULLBACHER, A., GALLIN, J. I., SIMON, M. M.. AND 'OXON-CHUNG, K. J. Gliotoxin is a virulence factor of Aspergillus fumtgatus: gliP deletion attenuates virulence in mice immunosuppressed with hydrocortisone. Eukaryot Cell 6, 9 (Sep 2007), 1562-1569.
[671] SUKIIAN, A. The invasion-associated type III secretion system of Salmonella typhimu urn: common and unique features. Cell Mal Life Sc. 57, 7 (Jul 2000), 1033-1049.
[672] SUMMERBELL, R. C., DE REFENTIGNY, L., CIIARTRAND, C., AND ST GERMAIN, G. Graft-related endocarditis caused by Ncosartorya fischeri var. spinosa. J Min Microbial 30, 6 (1992 Jun), 1580-1582.
[673] SUNDSTROM, P., CUTLER, J. E.. AND STAAB, J. F. Re-evaluation of the role of HWP/ in systemic candidiasis by use of Candida albicans strains with selectable marker URA3 targeted to the EN01 locus. Infect Irnmun 70, 6 (2002 Jun), 3281-3283.
[674] SUSEK. R. E., AND LINDQUIST, S. Transcriptional derepression of the Sacchamrnyces cerevisiae HSP26 gene during heat shock. Mot Cell Biol 10, 12 (1990 Dec), 6362-6373.
[675] TAKAHASHI, T., SMAIGI, H., AND ITO, K. Identification of genes required for growth under ethanol stress using transposon mutagenesis in Saccharomyces cerevisiae. Mol Genet Genomics 265, 6 (Aug 2001), 1112-1119.
[676] TAKESHIGE, K., AND MINAKAMI. S. NADH- and NADPH-dependent formation of superoxide anions by bovine heart submitochondrial particles and NADH-ubiquirtone reductase preparation. Biochern J 180, 1 (1979 Apr 15), 129-135.
[677] TALIBI, D., AND RAYMOND, M. Isolation of a putative Candida albicans transcriptional regulator involved in pleiotropic drug resistance by functional complementation of a pdrl pdr3 mutation in Saccharomyces cerevisiae. J Bacteriol 181, 1 (Jan 1999), 231-240.
[678] TAMAS, M. J., LL'YTEN, K., SUTHERLAND, F. C., HERNANDEZ, A., ALBERTYN, 1., VALADI, H., LI, H., PRIOR, B. A., KILIAN, S. G., RAMOS, J., GUSTAFSSON, L., THEVELEIN, J. M., AND HOHMANN, S. Fpslp controls the accumulation and release of the compatible solute glycerol in yeast osmoregulation. Mol Microbial 31, 4 (1999 Feb), 1087-1104.
[679] TAMURA, GU, Y., WANG, Q., YAMADA, T., ITO, K., AND SIIIMOI, H. A hapl mutation in a laboratory strain of Saccharomyces cerevisiae results in decreased expression of ergosterol-related genes and cellular ergosterol content compared to sake yeast. J Biasci Biaeng 98, 3 (2004), 159-166.
[680] TANG, C. NI., COVEN, J., AND HOLDEN, D. W. An Aspergillus fumigates alkaline protease mutant constructed by gene disruption is deficient in extracellular elastase activity. Mol Microbial 6, 12 (Jun 1992), 1663-1671.
[681] TANG, C. M., COHEN, J KRAUSZ, T., VAN NOORDEN, S., AND HOLDEN, D. W. The alkaline protease of Aspergillus fumigator is not a virulence determinant in two murine models of invasive pulmonary aspergillosis. Infect Bunion 61, 5 (May 1993), 1650-1656.
[682] TANNY, J. C., AND MOAZED, D. Coupling of histone deacetylation to NAD breakdown by the yeast silencing protein Sir2: Evidence for acetyl transfer from substrate to an NAD breakdown product. Proc Nat! Acad Set U S A 98, 2 (Jan 2001), 415-420.
[683] TAO, W. A., WOLLSCIIEID, B., O'BRIEN, R., ENG, J. K., LI, X.-J., BODENMILLER, B., WATTS, J. D., HOOD, L.. AND AEBERSOLD, R. Quantitative phosphoproteome analysis using a dendrimer conjugation chemistry and tandem mass spectrometry. Nat Methods 2, 8 (2005 Aug), 591-598.
[684] TATEBAYASHI, K., YAMAMOTO, K., TANAKA, K., TOMIDA, T., MARUOKA, T., KASUKAWA, E., AND SAITO, H. Adaptor functions of Cdc42, Ste50, and Shol in the yeast osmoregulatory HOG MAPK pathway. EMBO J 25, 13 (2006 Jul 12), 3033-3044.
[685] TEICIIERT, S., WOTTAWA, NI., SCHONIG, B., AND TUDZYNSKI, B. Role of the Fusarium fujikuroi TOR kinase in nitrogen regulation and secondary metabolism. Eukaryot Cell 5, 10 (Oct 2006), 1807-1819.

REFERENCES 466
[6861 TEIXEIRA, M. C., MONTEIRO, P., JAIN, P., TENREIRO, S., FERNANDES, A. Ft.. MIRA, N. P., ALENQUER, M., FREITAS. A. T., OLIVEIRA, A. L., AND SA-CORREIA, I. The YEASTRACT database: a tool for the analysis of transcription regulatory associations in Saccharornyces cerevisiae. Nucleic Acids Res 34, Database issue (2006 Jan 1), ID446-51.
[687] THAKUR, J. K., ARTHANARI, H., YANG, F., PAN, S.-J., FAN, X., BREGER, J., FRUEII, D. P., GULSHAN. K., L[, D. K., MYLONAKIS, E., STRUM., K., MOYE-ROWLEY, W. S., CORMACK, B. P., WAGNER, G., AND NAAR, A. M. A nuclear receptor-like pathway regulating multidrug resistance in fungi. Nature 452, 7187 (2008 Apr 3), 604-609.
[688] THEWES, S., KRETSCIIGAR, M., PARK, H., SCHALLER, M., FILLER, S. G., AND RUBE, B. In vivo and ex vivo comparative transcriptional profiling of invasive and non-invasive Candida albicans isolates identifies genes associated with tissue invasion. Mal Microbial 63, 6 (Mar 2007), 1606-1628.
[669] THOMPSON, J. D., GIBSON, T. J., AND HIGGINS, D. G. Multiple sequence alignment using clustalw and clustalx. Curr Protoc Bioin or-rnatics Chapter 2 (Aug 2002), Unit 2.3.
[690] THORNE, N. P. Single-channel normalisation and analysis of two-colour cDNA microurray data. PhD thesis, The Walter and Eliza Hall Institute of Medical Research, Department of Medical Biology and Department of Mathematics and Statistics, 2004.
[691] TOBIAS, T. S., JUNGWIRTH, H., AND KUCIILER, K. Creation of a hss3 C. glabrata strain. ORF deletion checked by PCR and Southern blot analysis., August 2009.
[692] TONG, A. H., EVANGELISTA, M., PARSONS, A. B., Xu, H., BADER, G. D., PAGE, N., ROBINSON, M., RAGIIIBIZADEll, S., HOGUE, C. W., HUSSEY, H., ANDREWS, B., TYERS, M., AND BOONE, C. Systematic genetic analysis with ordered arrays of yeast deletion mutants. Science 29, 5550 (Dec 2001), 2364-2368.
[693] TONG, A. H. Y., LESACE, G., BADER, G. D., DING, H., XU, H., XIN, X., YOUNG, J., BERM, G. F., BROST, R. L., CHANG, M., CHEN, Y., CHENG, X., CIIUA, G., FRIESEN, H., GOLDBERG, D. S., HAYNES, J., HUMPHRIES, C., HE, G., HUSSEIN, S., KE, L., KROGAN, N., Li, Z., LEVINSON, J. N., LU, H., MENARD, P., MUNYANA, C., PARSONS, A. B., RYAN, 0., TONIKIAN, R., ROBERTS, T., SDICU, A.-NI., SHAPIRO, J., SHEIKH, B., SUTER, B., WONG, S. L., ZIIANG, L. V., Zuv, H., KURD, C. G., MUNRO, S., SANDER, C., RINE, J., GREENBLATT, J., PETER, NI., BRETSCIIER, A., BELL, G., ROTH, F. P., BROWN, G. W., ANDREWS, B., HUSSEY, H., AND BOONE, C. Global mapping of the yeast genetic interaction network. Science 303, 5659 (Feb 2004), 808-813.
[694] TORA, L. A unified nomenclature for TATA box binding protein (TBP)-associated factors (TAFs) involved in RNA polymerase II transcription. Genes Dev 16, 6 (2002 Mar 15), 673-675.
[695] TREGER, J. NI., SCHMITT, A. P., SIMON„T. R., AND MUENTEE, K. Transcriptional factor mutations reveal regulatory complexities of heat shock and newly identified stress genes in Sacchammuces cerentsise. J Biol Chem 273, 41 (1998 Oct 9), 26875-26879.
[696] TSAI, H. F., CHANG, Y. C., WASIIBURN, R. G., WHEELER, M. H., AND KWON-CIIUNG, K. J. The developmentally regulated albl gene of Aspergillus fumigatus: its role in modulation of conidial morphology and virulence. J Bacterial 180, 12 (Jun 1998), 3031-3038.
[697] TSAI, H.-F., KROL, A. A., SARTI, K. E., AND BENNETT, J. E. Candida glabrata PDR1, a transcriptional regulator of a pleiotropic drug resistance network, mediates azole resistance in clinical isolates and petite mutants. Antimicrob Agents Chernother 50, 4 (2006 Apr), 1384-1392.
[698] TSAI, II. F., WASHBURN, R. G., CHANG, Y. C., AND KWON-CHUNG, K. J. Aspergillus funtigatus arpl modulates conidial pigmentation and complement deposition. Mol Microbial 26, 1 (Oct 1997), 175-183.
[699] TSUBOUCIII, H. Sustained activation of epidermal growth factor receptor in cholangiocarcinoma: a potent therapeutic target? J Hepatol 41, 5 (Nov 2004), 859-861.
[700] TSUKAMOTO, Y., KATO, J., AND IKEDA, H. Silencing factors participate in DNA repair and recombination in Saceharomyces ceretrisiae. Nature 388, 6645 (Aug 1997), 900-903.
[701] TSUNAWAKI, S.. YOSHIDA, L. S., NISHIDA, S., KOBAYASHI, T., AND SIIILIOYAMA, T. Fungal metabolite gliotoxin inhibits assembly of the human respiratory burst NADPH oxidase. Infect Irrimun 72, 6 (Jun 2004), 3373-3382.
[702] 'Dieu, B. B., LI, H., AND JOHNSON, A. D. Evolution of eukaryotic transcription circuits. Science 319, 5871 (2008 Mar 28), 1797-1799.
[703] TUDER, R. NI.. YUN, J. H., BHUNIA, A. AND FIJALKOWSKA, I. Hypoxia and chronic lung disease. J Mol Med 85, 12 (Dec 2007), 1317-1324.
[704] TURNER-WARWICK, M. Aspergillus fumigatus and lung disease. Pastgrad Med J 55, 647 (1979 Sep), 642-644.
[705] UETZ, P., GIOT, L., CAGNEY, G.. MANSFIELD, T. A., JUDSON, R. S., KNIGHT, J. R., LOCKSHON, D., NARAYAN, V., SRINIVASAN, NI.. POCHART, P., QURESIII-EMILI, A., LI, Y., GouwiN, B., CONOVER, D., KALBFLEISCII, T., VIJAYADAMODAR, G.. YANG, NI., JOHNSTON, M., FIELDS, S., AND ROTIIBERG, J. NI. A comprehensive analysis of protein-protein interactions in Saccharamyces cerovisme. Nature 403, 6770 (2000 Feb 10), 623-627.
[706] UNIEBAYASHI, K., AND NAKANO, A. Ergosterol is required for targeting of tryptophan permease to the yeast plasma membrane. J Cell Thal 161, 6 (2003 Jun 23), 1117-1131.

REFERENCES 467
[707] UPPULURI, P., NETT, J., HEITMAN, J., AND ANDES, D. Synergistic effect of calcineurin inhibitors and fluconazole against Candida albicans biofilms. Antimicrob Agents Chemother 52, 3 (2008 Mar), 1127-1132.
[708] VAIIEDI, G., IVAN0v, I. V., AND DOUGHERTY, E. R. Inference of boolean networks under constraint on bidirectional gene relationships. IET Syst Biol 3, 3 (2009 May), 191-202.
[709] VALIANTE, V., HEINEKARIP, T., JAIN, R., HARTI., A., AND BRAKIIAGE, A. A. The mitogen-activated protein kinase MpkA of Asp Talus fumigatus regulates cell wall signaling and oxidative stress response. Fungal Genet Dial 45, 5 (May 2008), 618-627.
[710] VALIANTE, V., JAIN, R., HEINEKAMP, T., AND BRAKUAGE, A. The MpkA MAP kinase module regulates cell wall integrity signaling and pyomelanin formation in Aspergillus furnigatus. Fungal Genet Biol (Aug 2009).
[711] VAN DEN BOSSCIIE, H., MartICItaL, P., ODDS, F. C.. LE JEUNE, L., AND COENE, M. C. Characterization of an azole-resistant Candida glabrata isolate. Antimicrob Agents Chemother 36, 12 (1992 Dec), 2602-2610.
[712] VAN DEN BOSSCIIE, H., WILLEMSENS, G., Coots, W., MARICHAL, P., AND LAUWERS, W. Hypothesis on the molecular basis of the antifungal activity of N-substituted imidazoles and triazoles. Biochem Soc Trans 11, 6 (1983 Dec), 665-667.
[713] VAN DER HORST, C. M., SAAR, M. S.. CLOUD. G. A., lIstimh, R. J., GRAYBILL, J. R., SOBEL, J. D., JOHNSON, P. C., TUAZO, C. U., KERKERING, T.. MOSKOVITZ, B. L., POWDERLY, W. G., AND DISMUKES, W. E. Treatment of cryptococcal meningitis associated with the acquired immune deficiency syndrome. New England Journal of Medicine 337 (1997), 15-21.
[714] VAN DER WALT, J. P. Sexually active strains of Candida albicans and Cryptococcus albidus. Antonie Van Leenwenhoek 38, 3 (1967), 246-256.
[715] VAN DRCIGEN, F., AND PETER, M. MAP kinase cascades: scaffolding signal specificity. Curt- Dial 12, 2 (2002 Jan 22), R53-5.
[716] VAN NOORT, V., SNEL, B., AND HUYNEN, M. A. Predicting gene function by conserved co-expression. Trends Genet 19, 5 (May 2003), 238-242.
[717] VAN ROSSUM, G. Python reference manual. Report CS-R9525, pub-CWI, pub-CWI:adr, Apr 1995.
[718] VAZQUEZ-TORRES, A., JONES-CARSON, J., AND BALISII, E. Peroxynitrite contributes to the candidacidal activity of nitric oxide-producing macrophages. Infect frrirnuti 64, 8 (Aug 1996), 3127-3133.
[719] VERMITSKY, J.-P., EARHART, K. D., SMITH, W. L., HOMAYOUNI, R., EDLIND, T. D.. AND ROGERS, P. D. Pdrl regulates multidrug resistance in Candida glabrata: gene disruption and genome-wide expression studies. Mal Microbial 61, 3 (2006 Aug), 704-722.
[720] VON FIEF, M., ROOS, N., SCHULTEN, R., HF.SSE, M., ZUHLSDORE, M., AND VAN DE Loo, J. Pulmonary aspergillosis: early diagnosis improves survival. Respiration 62, 6 (1995), 341-347.
[721] VON MERING, C., KRAUSE, R.. SNEL, B., CORNELL, M.. OLIVER, S. G., FIELDS, S., AND BORK, P. Comparative assessment of large-scale data sets of protein-protein interactions. Nature .417, 6887 (2002 May 23), 399-403.
[722] WALKER, L. A., MUNRO, C. A., DE BRUIJN, I., LENARDON, M. D., MCKINNON, A., AND Gow, N. A. R. Stimulation of chitin synthesis rescues Candida albicans from echinocandins. PLoS Pathog 4, 4 (2008 Apr), e1000040.
[723] WALLACE, I. M., O'SULLIVAN, 0., AND HIGGINS, D. G. Evaluation of iterative alignment algorithms for multiple alignment. Bannforrnatias 21, 8 (Apr 2005), 1408-1414.
[724] WALSH, T. J., GROLL, A., HIEMENZ, J., FLEMING, R., ROILIDES, E., AND ANAISSIE, E. Infections due to emerging and uncommon medically important fungal pathogens. Clin Microbial Infect 10 Suppl 1 (2004 Mar), 48-66.
[725] WANG, Y., AISEN, P., AND CASADEVALL, A. Cryptococcus neoformans melanin and virulence: mechanism of action. Infect Immun 63, 8 (Aug 1995), 3131-3136.
[726] WANG, Y., AND CASADEVALL, A. Growth of Cryptococcus neoformans in presence of L-dopa decreases its susceptibility to amphotericin B. Antimicrob Agents Chernother 38, 11 (Nov 1994), 2648-2650.
[727] WARD, M. P., GIMENO, C. J., FINK, G. R., AND GARRETT, S. SOK2 may regulate cyclic AMP-dependent protein kinase-stimulated growth and pseudohyphal development by repressing transcription. Mol Cell Biol 15, 12 (1995 Dec), 6854-6863.
[728] WARENDA, A. J., KAUFFMAN, S., SIIERRILL, T. P., BECKER, J. M., AND KONOPKA, J. B. Candida albicans septin mutants are defective for invasive growth and virulence. Infect frnmun 71, 7 (2003 Jul), 4045-4051.
[729] WARMKA, J., HANNEMAN, J., LEE, J., AMIN, D., AND OTA, I. Ptcl, a type 2C Ser/Thr phosphatase, inactivates the HOG pathway by dephosphorylating the mitogen-activated protein kinase Hogl. Mal Cell Biol 21, 1 (2001 Jan), 51-60.
[730] WARRINGER, J., ERICSON, E., FERNANDEZ, L., NERMAN, 0., AND BLOMBERG, A. High-resolution yeast phenomics resolves different physi-ological features in the saline response. Proc Natl Acad Sci U S A 100, 26 (Dec 2003), 15724-15729.

REFERENCES 468
[731] WATANABE, A., KAMEI, K., SEKINE, T., WAKu, NI., NISHIMURA, K., MIYAJI, M., AND KURIYAMA, T. Immunosuppressive substances in Aspergillus fumigates culture filtrate. J Infect Cheinother 9,2 (Jun 2003), 114-121.
[732] WATSON, A. D., EDMONDSON, D. G., BONE, J. R., MuKAI, Y., Yu, Y., Du, W., STILLMAN, D. J., AND ROTH, S. Y. Ssn6-Tupl interacts with class I histone deacetylases required for repression. Genes Dev 14, 21 (2000 Nov 1), 2737-2744.
[733] WATTS, D. J., AND STROGATZ, S. H. Collective dynamics of 'small-world' networks. Nature 393, 6684 (1998 Jun 4), 440-442.
[734] WEIC, M., HAYNES, K., ROGERS, T. R., Kt:nal, 0., FitoSOL M., AND MUHLSCHLEGEL, F. A. A GAS-like gene family in the pathogenic fungus Candida glabrata. Microbiology 147, Pt 8 (2001 Aug), 2007-2019.
[735] WEINER, N. Cybernetics or Control and Communication in the Animal and the Machine. MIT Press, Cambridge, MA, 1948.
[736] WENZEL, R. P., AND GENNINGS. C. Bloodstream infections due to Candida species in the intensive care unit: identifying especially high-risk patients to determine prevention strategies. Clio Infect Dis 41 Suppl 6 (2005 Sep 15), S389-93.
[737] WESTFALL, P. J., PATTERSON, J. C., CHEN, R. E., AND TIIORNER, J. Stress resistance and signal fidelity independent of nuclear MAPK function. Proc Nat! Acad Sci U S A 10,5, 34 (Aug 2008), 12212-12217.
[738] WHEELER, M. H., AND BELL, A. A. Melanins and their importance in pathogenic fungi. Curr Top Med Mycol 2 (1988), 338-387.
[739] WHEELER, R. T., AND FINK, G. R. A drug-sensitive genetic network masks fungi from the immune system. PLoS Pathog 2,4 (Apr 2006), e35.
[740] WHITE, T., AND PAGUREK, B. Emergent behaviour and mobile agents. In In Proc. of the Workshop on Mobile Agents in the Context of Competition and Cooperati on at Autonomous Agents (1999).
[741] WHITE, T. C. The presence of an 15.467K amino acid substitution and loss of allelic variation correlate with an azole-resistant lanosterol 14-alpha demethylase in Candida albicans. Antimicrob Agents Chemother 41, 7 (1997 Jul), 1488-1494.
[742] WIESER, R., ADAM, G., WAGNER, A., SCHULLER, C., MARCIILER, G., RUIS, H., KRAWIEC, Z., AND BILINSKI, T. Heat shock factor-independent heat control of transcription of the CTTI gene encoding the cytosolic catalase T of Sacchammyces ccrevisiae. J Riot Chem 266, 19 (1991 Jul 5), 12406-12411.
[743] WILCZYNSKI, B., AND FURLONG, E. Challenges for modeling global gene regulatory networks during development: Insights from drosophila. Deo Biol (2009 Oct 27).
[744] WILLSON, T. M., AND KLIEWER, S. A. PXR, CAR and drug metabolism. Nat Rev Drug Discov 1, 4 (Apr 2002), 259-266.
[745] WILSON, L. S., REYES, C. M., STOLPMAN, NI., SPECKMAN, J., ALLEN, K., AND BENEY, J. The direct cost and incidence of systemic fungal infections. Value Health 5, 1 (2002 Jan-Feb), 26-34.
[746] WILSON, R. B., DAVIS, D., ENLOE, B. M., AND ;MITCHELL, A. P. A recyclable Candida albicans URA3 cassette for PCR product-directed gene disruptions. Yeast 16, 1 (Jan 2000), 65-70.
[747] WILSON, W. A., WANG, Z., AND ROACH, P..1. Systematic identification of the genes affecting glycogen storage in the yeast Saccharomyces cerciiisiaci implication of the vacuole as a determinant of glycogen level. Mal Cell Proteamics 1, 3 (Mar 2002), 232-242.
[748] WINGARD, J. R. infections due to resistant Candida species in patients with cancer who are receiving chemotherapy. Clinical Infectious Diseases SI (August 1994), S49—S53.
[749] WINZELER, E. A., SHOEMAKER, D. D., ASTROMOFF, A., LIANG, H., ANDERSON, K., ANDRE, B., BANGIIAM, R., BENITO, R., BOEKE, J. D., HUSSEY, H., Ciiu, A. M., CONNELLY, C., DAVIS, K., DIETRICH, F., WIIELAN-DOW, S., EL-BAKKOURY, M., FOURY, F., FRIEND, S. H., GENTALEN, E., GIAEVER, G., HEGEMANN, J. H., JONES, T., LAUB, hi., LIAO, H., LIEBUNDGUTII, N., LOCKHART, D. J., LUCAU-DANILA, A., LUSSIER, M., M'RABET, N., MENARD, P., MITTMANN, M., PAI, C., REBISCHL'NC, C., REVUELTA„T. L., RILES, L., ROBERTS, C. J., ROSS-MACDONALD, P., SCHERENS, B., SNYDER, M., SOOKHAI-MAHADEO, S., STORMS, R. K., VERONNEAY, S., VOET, NI., VOLCKAERT, G., WARD, T. R., WYSOCKI, R., YEN, G. S., YU, K., ZIMMERMAN, K., PHILIPPSEN, P., JOHNSTON, NI., AND DAVIS, R. W. Functional characterization of the S. cerevistiac genome by gene deletion and parallel analysis. Science 5423 (August 285), 901-906.
[750] WIRSCHING, S., MICHEL, S., AND MORSCHHAUSER, J. Targeted gene disruption in Candida albicans wild-type strains: the role of the MDR1 gene in fluconazole resistance of clinical Candida albicans isolates. Mot Microbial 36, 4 (May 2000), 856-865.
[751] WISPLINGHOFF, H., BISCHOFF, T., TALLENT, S. NI., SEIFERT, H., WENZEL, R. P., AND EDMOND, M. B. Nosocomial bloodstream infections in us hospitals: analysis of 24,179 cases from a prospective nationwide surveillance study. Clin Infect Dis 39, 3 (2004 Aug 1), 309-317.
[752] WoLFE, K. H., AND Siiiums, D. C. Molecular evidence for an ancient duplication of the entire yeast genome. Nature 387, 6634 (1997), 708-713.
[7531 Wocuioup, J., HUBER, R., POIILERS, D., KOCZAN, D., GUTIIKE, R., KINNE, R. W., AND GAIISNIANN, U. Adapted boolean network models for extracellular matrix formation. BMC Syst Riot 3 (2009), 77.

REFERENCES 469
[754] WONG, S.. FARES, M. A., ZIMMERMAN, W., BUTLER, G., AND WOLFE, K. H. Evidence from comparative genomics for a complete sexual cycle in the "asexual" pathogenic yeast Candida glabrata. Gcnonic Biology 4, 2 (2003), R10.
[755] WORKMAN, C. T., MAK, H. C., MCCUINE, S., TAGNE, J.-B., AGARWAL, M., OZIER, 0., BEGLEY, T. J., SAMSON, L. D., AND IDEKER, T. A systems approach to mapping DNA damage response pathways. Science 312, 5776 (2006), 1054-1059.
[756] We, J., SUKA, N., CARLSON, M.. AND GRUNSTEIN, M. Tupl utilizes histone H3/H2B-specific HDA1 deacetylase to repress gene activity in yeast. Mol Cell 7, 1 (2001 Jan), 117-126.
[757] WURGLER-MURPIIY, S. M., MAEDA. T., WITTEN, E. A., AND SAITO, H. Regulation of the Saccharornyces cerevimae HOG1 mitogen-activated protein kinase by the PTP2 and PTP3 protein tyrosine phosphatases. Mol Cell Biol 17, 3 (1.997 Mar), 1289-1297.
[758] WYSONG, D. R., CHRISM, L., SUGAR, A. M., ROBBINS, P. W., AND DIAMOND, R. D. Cloning and sequencing of a Candida albicans catalase gene and effects of disruption of this gene. Infect Iranrun 66, 5 (May 1998), 1953-1.961.
[759] XIONG, W., AND FERRELL, J. E. J. A positive-feedback-based bistable "memory module" that governs a cell fate decision. Nature 426, 6965 (Nov 2003), 460-465.
[760] Xu, X.-L., LEE, R. T. H., FANG, H.-M., WANG, Y.-M., LI, R., Zott, H., Diu, Y., AND WANG, Y. Bacterial peptidoglycan triggers Candida albicans hyphal growth by directly activating the adenylyl cyclase Cyr 1p. Cell Host Microbe 4, 1 (2008 Jul 17), 28-39.
[761] YAAKOV, G., DUCH, A., GARCIA-RUBIO, M., CLOTET, J., JIMENEZ, J., AGUILERA, A., AND POSAS, F. The stress-activated protein kinase Hogl mediates S phase delay in response to osmostress. Mid Biol Cell 20, 15 (2009 Aug), 3572-3582.
[762] YANG, Y. H., DUDOIT, S., Lee, P., LIN, D. M., PENG, V., NGAI, J., AND SPEED, T. P. Normalization for cDNA microarray data: a robust composite method addressing single and multiple slide systematic variation. Nucleic Acids Res 30, 4 (2002 Feb 15), e15.
[763] YANG, Z. PAML 4: phylogenetic analysis by maximum likelihood. Mol Biol Evol 24, 8 (2007 Aug), 1586-1591.
[764] YOOK, S.-H., OLTVAI. Z. N., AND BARABASI, A.-L. Functional and topological characterization of protein interaction networks. Pro-temnics 4, 4 (2004 Apr), 928-942.
[765] YOGA, J.-H., GWAK, G.-Y., LEE, H.-S., BRONK, S. F., WERNEBURG, N. W., AND GORES, G. J. Enhanced epidermal growth factor receptor activation in human cholangiocarcinoma cells. J Hepatol 41, 5 (Nov 2004), 808-814.
[766] Yosti/DA, Y., AND AOYAMA, Y. Sterol 14 alpha-demethylase and its inhibition: structural considerations on interaction of azole antifungal agents with lanosterol 14 alpha-demethylase (p-450(14dm)) of yeast. Biocherre Sac Trans 19, 3 (1991 Aug), 778-782.
[767] YOUNG, C., MAPES, J., HANNEMAN, J., AL-ZARBAN, S., AND OTA, I. Role of Ptc2 type 2C Ser/Thr phosphatase in yeast high-osmolarity glycerol pathway inactivation. Enkaryot Cell 1, 6 (2002 Dec), 1032-1040.
(768] ZAMAN, Z., ANSARI, A. Z., KOH, S. S., YOUNG, R., AND PTASIINE, M. Interaction of a transcriptional repressor with the RNA polymerase II holoenzyme plays a crucial role in repression. Proc Natl Acad Sri U S A 98, 5 (2001 Feb 27), 2550-2554.
[769] ZHANG, J., CIIUNG, T., AND OLDENBURG, K. A simple statistical parameter for use in evaluation and validation of high throughput screening assays. J Biornol Screen 4, 2 (1999), 67-73.
[770) ZHAO, X., Oil. S.-H., YEATER, K. M., AND HOYER, L. L. Analysis of the Candida albicans Als2p and Als4p adhesins suggests the potential for compensatory function within the ALS family. Microbiology 151, Pt 5 (2005 May), 1619-1630.
[771] ZITOMER, R. S. CARRICO, P., AND DECKERT, J. Regulation of hypoxic gene expression in yeast. Kidney Jut 51, 2 (1997 Feb), 507-513.
[772] ZITOMER, R. S., AND LOWRY, C. V. Regulation of gene expression by oxygen in Saccharamyces cerrevisiae. Microbial Rev 56, 1 (1992 Mar), 1-11.
[773] ZMEILI, 0. S., AND SOUBANI, A. 0. Pulmonary aspergillosis: a clinical update. QJM 100, 6 (Jun 2007), 317-334.
[774] Zoc, X., PENG, T., AND PAN, Z. Modeling specificity in the yeast MAPK signaling networks. J Theor Biol 250, 1 (2008 Jan 7), 139-155.




















