
Cloning of the human sodium-iodide symporter promoter and characterization in a differentiated human...
7
THYROID Volume 8, Number 1, 1998 Mary Ann Liebert, Inc. Cloning of the Human Sodium-Iodide Symporter Promoter and Characterization in a Differentiated Human Thyroid Cell Line, KAT-50 GOPALAKRISHNAN M. VENKATARAMAN, MUSTAFA YATIN, and KENNETH B. AIN ABSTRACT Elucidation of the regulation of human sodium-iodide symporter (hNIS) gene expression is critical to under- standing its effects on iodide concentration abilities of thyroid and thyroid carcinomas. To explore this issue, a 1.2-kb portion of the 5'-flanking region of the hNIS gene was isolated and characterized. Transient transfections with chimeric luciferase-reporter constructs into a differentiated human thyroid cell line, KAT-50, as well as non- thyroidal cells, defined an active promoter with tissue-specificity. Reverse-transcriptase polymerase chain reaction analysis for hNIS mRNA expression in normal human tissues was positive in thyroid, salivary gland, omentum, and gallbladder. KAT-50 cells expressed hNIS mRNA and were capable of thyrotropin-responsive iodide uptake in vitro. Despite the failure to exhibit iodide concentration in clinical anaplastic carcinoma tumors, 4 of 5 cell lines from this cancer phenotype expressed hNIS mRNA. Definition of the active promoter provides further in- sights and tools to uncover new approaches to use of radioiodine for therapy of thyroid carcinomas. INTRODUCTION Thyroid FOLLicuLAR cells have the unique ability to concentrate iodide, which is organified and utilized for the synthesis of thyroid hormones. Transport of iodide across the follicular cell membrane, in concert with inward movement of sodium (Na+) along its gradient, is an active process mediated by the tissue-specific sodium-iodide sym- porter (NIS). The cDNA sequence for the rat NIS gene was reported by Dai et al. (1) and the corresponding human cDNA sequence (hNIS) was delineated recently by Smanik et al. (2) as well as the intron-exon organization of the gene (3). Retention of the ability to concentrate iodide via NIS in carcinoma cells derived from thyroid follicular cells pro- vides an opportunity to destroy these cells with adminis- tered radioiodine. The efficacy of this approach relies on adequate expression of functional NIS. Unfortunately, NIS expression in human thyroid carcinoma cells can be vari- able or absent, leaving some carcinomas untreatable with this modality. To understand the regulation of expression of hNIS gene in normal and malignant thyroid tissues, it is essential to understand the regulation of the hNIS gene. We report the cloning and functional characterization of the hNIS gene promoter as well as the expression of this gene in diverse human tissues and human thyroid cancer cell lines. To elucidate the role of the promoter in tissue- specific hNIS expression, we evaluated the activity of chimeric promoter constructs in human thyroid and non- thyroidal cells. We utilized our human thyroid cell line, KAT-50, which was obtained from primary culture of the thyroid of a 3-year-old boy with benign follicular hyper- plasia, to demonstrate thyroid-specific promoter activity, because this is the only known human thyroid cell line that retains functional NIS expression and iodide-concentrating activity in vitro. MATERIALS AND METHODS Cell lines and human tissues KAT-50 is a cell line established by primary culture from the thyroid of a 3-year-old boy with benign follicular hy- perplasia. Cultures were treated for 4 weeks in media con- taining D-valine (4) and «s-4-hydroxy-L-proline (5) to en- Thyroid Cancer Research Laboratory, Medical Service, Veterans Affairs Medical Center, Lexington, Kentucky and the Department of In- ternal Medicine, University of Kentucky Medical Center, Lexington, Kentucky. 63
Transcript of Cloning of the human sodium-iodide symporter promoter and characterization in a differentiated human...

THYROIDVolume 8, Number 1, 1998Mary Ann Liebert, Inc.
Cloning of the Human Sodium-Iodide Symporter Promoterand Characterization in a Differentiated Human Thyroid
Cell Line, KAT-50
GOPALAKRISHNAN M. VENKATARAMAN, MUSTAFA YATIN, and KENNETH B. AIN
ABSTRACT
Elucidation of the regulation of human sodium-iodide symporter (hNIS) gene expression is critical to under-standing its effects on iodide concentration abilities of thyroid and thyroid carcinomas. To explore this issue, a
1.2-kb portion of the 5'-flanking region of the hNIS gene was isolated and characterized. Transient transfectionswith chimeric luciferase-reporter constructs into a differentiated human thyroid cell line, KAT-50, as well as non-
thyroidal cells, defined an active promoter with tissue-specificity. Reverse-transcriptase polymerase chain reactionanalysis for hNIS mRNA expression in normal human tissues was positive in thyroid, salivary gland, omentum,and gallbladder. KAT-50 cells expressed hNIS mRNA and were capable of thyrotropin-responsive iodide uptakein vitro. Despite the failure to exhibit iodide concentration in clinical anaplastic carcinoma tumors, 4 of 5 celllines from this cancer phenotype expressed hNIS mRNA. Definition of the active promoter provides further in-sights and tools to uncover new approaches to use of radioiodine for therapy of thyroid carcinomas.
INTRODUCTION
Thyroid FOLLicuLAR cells have the unique ability toconcentrate iodide, which is organified and utilized for
the synthesis of thyroid hormones. Transport of iodideacross the follicular cell membrane, in concert with inwardmovement of sodium (Na+) along its gradient, is an activeprocess mediated by the tissue-specific sodium-iodide sym-porter (NIS). The cDNA sequence for the rat NIS gene was
reported by Dai et al. (1) and the corresponding humancDNA sequence (hNIS) was delineated recently by Smaniket al. (2) as well as the intron-exon organization of thegene (3).
Retention of the ability to concentrate iodide via NIS incarcinoma cells derived from thyroid follicular cells pro-vides an opportunity to destroy these cells with adminis-tered radioiodine. The efficacy of this approach relies on
adequate expression of functional NIS. Unfortunately, NISexpression in human thyroid carcinoma cells can be vari-able or absent, leaving some carcinomas untreatable withthis modality. To understand the regulation of expressionof hNIS gene in normal and malignant thyroid tissues, itis essential to understand the regulation of the hNIS gene.
We report the cloning and functional characterization ofthe hNIS gene promoter as well as the expression of thisgene in diverse human tissues and human thyroid cancer
cell lines. To elucidate the role of the promoter in tissue-specific hNIS expression, we evaluated the activity ofchimeric promoter constructs in human thyroid and non-
thyroidal cells. We utilized our human thyroid cell line,KAT-50, which was obtained from primary culture of thethyroid of a 3-year-old boy with benign follicular hyper-plasia, to demonstrate thyroid-specific promoter activity,because this is the only known human thyroid cell line thatretains functional NIS expression and iodide-concentratingactivity in vitro.
MATERIALS AND METHODS
Cell lines and human tissuesKAT-50 is a cell line established by primary culture from
the thyroid of a 3-year-old boy with benign follicular hy-perplasia. Cultures were treated for 4 weeks in media con-
taining D-valine (4) and «s-4-hydroxy-L-proline (5) to en-
Thyroid Cancer Research Laboratory, Medical Service, Veterans Affairs Medical Center, Lexington, Kentucky and the Department of In-ternal Medicine, University of Kentucky Medical Center, Lexington, Kentucky.
63

64 VENKATARAMAN ET AL.
sure elimination of fibroblasts. Other human thyroid celllines were: AR081 and DRO90 (both from anaplastic car-
cinomas, provided by G.J.F. Juillard, University of Cali-fornia-Los Angeles School of Medicine); BHT-101(anaplastic carcinoma, provided by I. Pályi, National In-stitute of Oncology, Budapest, Hungary) (6); KAT-4 andKAT-18 (anaplastic carcinomas, from our laboratory) (7);NPA87 (papillary carcinoma, from Juillard); KAT-5 andKAT-10 (8) (both papillary carcinomas, from our labora-tory); KAK-1 (benign follicular adenoma, from our labo-ratory) (9); KAT-7 (benign follicular hyperplasia, from our
laboratory); and KAT-9 (derived from Hashimoto's thy-roiditis-afflicted follicular cells, from our laboratory). Ad-ditional cell lines include: HeLa, human epithelial cervicalcarcinoma; Hep G2, human hepatoma; and COS-1, SV-40-transformed African green monkey kidney cells. All ofthe thyroid-derived cell lines were grown in RPMI 1640with 10% fetal bovine serum (FBS), with the other celllines grown in Dulbecco's Modified Eagle's Medium(DMEM)/F12 (1:1) with 10% FBS (all from Life Tech-nologies, Gaithersburg, MD). Normal human tissues were
obtained from fresh surgical samples.Nucleic acid isolation and amplification (RT-PCR)
RNAs from normal tissues and cell lines were isolatedby acid-guanidinium-phenol-chloroform method (10).Complementary DNA (cDNA) was synthesized from 0.5fig of total RNA using MMLV reverse transcriptase (RT)with random-hexamer primers (Clontech, Palo Alto, CA).Each 50-/xL polymerase chain reaction (PCR) contained 60mM Tris HC1, pH 9.0, 15 mM ammonium sulfate, 3.5mM MgCl2 (for PAX-8 at 2 mM), 250 pM dNTPs, 2 pMprimers, 1 U AmpliTaq DNA polymerase (Perkin-Elmer,Norwalk, CT), 0.2 fig TaqStart Antibody (Clontech) and1% cDNA. /3-actin amplification (primers, Stratagene, LaJolla, CA) confirmed cDNA integrity, purity, and templateequivalence. PCR primers and product sizes are (upstream5' to 3'/downstream 5' to 3'): TG (thyroglobulin, 174 basepair [bp]), AACTTCAGTCTCTTTATTCAAAGTCTC/GCCATTTAAGATCTGCCAGAAGAG; TPO (thyroidperoxidase, 472/342 bp, splice variant), CCCCTAC-GAGTTAGGAGACGATGC/CATCCATGCCTGAGAG-TAACTACG; TSH-R (thyrotropin receptor, 367 bp),GGGTGCAACACGGCTGGTTT/CTGGGTTGTGCG-GATTTCGG; TTF1 (thyroid transcription factor-1, 257bp), CGCCGTACCAGGACACCATGAGC/GACAGGT-ACTTCTGTTGCTTGAAG; PAX-8 (329 bp), CACA-AGTCCAGCATTGCGGCACATC/CTGGAG-GGAAGTGCTGCTTATGGTCC; and hNIS (303 bp),CTGCCCCAGACCAGTACATGCC/TGACGGTGAAG-GAGCCCTGAAG. The hNIS primers amplify a region ofthe hNIS gene spanning four introns (3) to yield the 303-bp product with cDNA only and will not amplify genomicDNA. Amplification conditions are: denaturation 94°C X3 minutes; 40 cycles of 20 seconds at 94°C, 30 seconds at62°C (64°C for hNIS), and 60 seconds at 72°C; followedby 72°C for 7 minutes.
Iodide uptake in KAT-50 cellsKAT-50 cells (1 X 105/9.4 cm2) were grown in phenol
red-free RPMI 1640 with 10% FBS ( ± 0.1 nM bTSH), in
6-well plates. Medium was replenished on the fourth day.At the end of 7 days, cells in each well were washed with2 mL of buffer, containing: 10 mM HEPES, pH 8.3 (with5.5 mM glucose, 5.4 mM KC1, 1.3 mM CaCl2, 0.4 mMNa2HP04, and 0.44 mM KH2P04), and either 137 mMNaCl (buffer A) or 100 mM choline chloride (buffer B).After incubation in the same volume of the same buffer,supplemented with Na[125I](1.0 /xCi/2 mL) and 1.0 fiMNal, for 60 minutes, cell were briefly washed once withbuffer A, trypsinized, and gamma-counted. Trichloroaceticacid (10% v/v) was then added, contents centrifuged andthe pellet resuspended. Pellets, dried at 60°C, were weighedto normalize counts for cell mass.
Cloning and sequencing of NIS cDNA
Multiple sets of degenerate oligonucleotide primers,based on the rat NIS cDNA sequence (1), were used to am-
plify fragments of hNIS cDNA from random-hexamer-primed human thyroid cDNA. Two major amplificationproducts were sequenced and additional primers were syn-thesized and used for further RT-PCR amplifications to ob-tain 1.6 kb of hNIS cDNA sequence. From this, a 5'-prox-imal a-32P-labeled DNA probe was synthesized and usedto screen a human thyroid cDNA library (kindly providedby Basil Rapoport, VAMC, San Francisco, CA). Twelvedifferent positive cDNA clones were obtained, analyzed byrestriction enzyme digestion with electrophoresis on
agarose gels, and the three clones containing the longestupstream extent of hNIS gene were sequenced.
Cloning and sequencing of the promoterThe upstream regulatory region of the NIS was cloned
using the PromoterFinder™ DNA walking kit (Clontech)with a combination of Tth (Epicenter Tech., Madison, WI)and Vent (New England BioLabs, Beverly, MA) DNA poly-merases for long-distance PCR reaction (11). This kit con-
tains five "libraries" that are pools of DNA fragments ob-tained from digestion of genomic DNA derived fromhuman placenta, ligated to a proprietary DNA adapter thatprovides a PCR priming site. The first round of PCR usedgene specific primer (GSP-1) (5'-3') CCACGGCCTC-CATGAGGGCGGGTGCGGA and commercial adapter-primer-1. The gene-specific primers were chosen to benear the 5'-most end of our three cDNA clones (see priorsection). The second PCR round used GSP-2 TAG-GAATCTATGGGCTGTCGATGTCCGT and commercialadapter-primer-2. These PCR products (and plasmid DNAwhen applicable) were sequenced by fluorescence dye-ter-minator method (Model 3 73A, Perkin Elmer AppliedBiosystems, Foster City, CA) at the Macromolecular Struc-ture Analysis Facility, University of Kentucky. Promoterfragments were sequenced in both directions using syn-thetic oligonucleotide primers.
5' Rapid amplification of c-DNA ends (5'-RACE) todefine transcription start site
The transcription start site was mapped by a modifica-tion of the 5'-RACE method of Frohman (12), using hu-man thyroid cDNA ligated to a commercial adapter(Marathon-Ready™ cDNA, Clontech). Antisense primers

HUMAN THYROID Na+IODIDE SYMPORTER PROMOTER 65
(GSP-2; and RX1, GCTGGTGAGGCCCAGGCGGT) nation with either the GSP-2 or GSP-1 antisense primingwere used in combination with the sense adapter-primer in sequences.two separate PCR amplifications of the human thyroidcDNA. The lengths of these products were aligned to the T7 . j
.
/• .-
j- t^xta 111 Vector constructions and transtecttoncorresponding cDNA sequence map and compared withthe ends of the cDNA clones obtained screening the hu- Chimeric promoter-luciferase reporter constructs were
man thyroid cDNA library from Dr. Rapoport. In sepa- made by incorporating amplified promoter fragments inrate experiments, PCR amplifications using different sense the promoterless pGL3-Basic vector (Promega, Madison,primers, corresponding to DNA regions upstream of the WI), as sollows: A, —930 to —336; B, —1044 to —336;putative transcription start site were performed in combi- and C, —1108 to —336. The recombinant constructs were
gagtagctgg
gattacaggcatgtgccaccacgccccgctaatattagtatctttcatacagacaagatctcactatgttgctcagggtagtctcgaattctgggactcaaatgatcctcccacttcagcctcccaaagtgctgggattacaggcataagccatcatgcccggcctctgacgctgtttct -1501
ttcaacccccaggatttcagattccaccagcttatggagaagggaaccaagttcgagatgcgtgattgcccagaaagttggaggctgagctgagacttgaacccagagaccagaacctccagaggtcaaagtcctcctcctgggtcccccagagaagggccctgagatgacagctcgttggtcctcatggaagcgtgacccccccagtagactttctcccacacccaaccttggtttcctcate tata tgatagggacaagccagactctacctccctggtggtcatggtctccgct tat -1201
tcgggttcataaccttaaaggcccctcgcaccacctcagtgagccatttatgcctggcac
agggccaactctcagtgcatatctgcaaaggaaccaatgaatgaatgaatgaagtgacaa4*
atgaataaaggaataaatgaacgaggcacttatcatgtaccaggctttcgttaccacgtcccatttattcctctgaggcagggtctattttatccttgttacagatggggaaactaaggc
¿Accagggaggagcaaagtcttccccaagtatgtacccactcagaacttgagctctgaatgt -0901
ctcccacccagcttagcccaagagcggggttcagtgatgcccaccccctaaggctctaga
gaaagggggtaggcccacatgccagtttgggggtggtaaagccaggtaagttttctttat
gggtcccctgaaaccctgaaagtgaaccccagtcctgcatgaaagtgagctccccatagc
tcaaggtattcaagcacaatacggctttgagtgctgaagcaggctgtgcaggcttggata
gtgacatgccctctctgagcctcaatttccccacctgtcaacagcagacagcgacagctg -0601
tgatcaggggatcacagtgcatggggatgggtgggtgcatggggatggaggggcatttgg
gagccctccccgataccaccccctgcagccacccagatagcctgtcctggcctgtctgtc
ccagtccagggctgaaagggtgcgggtcctgcccgcccctaggtctggaggcggagtcgcJVA2
ggtgacccgggagcccaataaatctgcaacccacaatcacgagctgctcccgtaagcccc+VA1 *»" i .
aaggcgacctccagctgccagcgccgagcacagcgcccagggagagggacagacagccgg -0301
ctgcacgggacagcggaacccagagtgagaggggaggtggcaggacagacagacagcaggggcggacgcagagacagacagcggggacagggaggccgacacggacatcgacagcccata
gattcctaacccagggagccccggcccctctcgccgcttcccaccccagacggagcggggacaggctgccgagcatccccccacccgccctccccgtcctgcctcctcggcccctgccagctccccccgcccgagcacgcagggcgcccgaggacgcgctgggcctccgcacccgccctc -0001
ATOGAGGCCGTGGAGACCGGGGAACGGCCCACCTTCGGAGCCTGGGACTACGGGGTCTTT
FIG. 1. Sequence of the hNIS promoter. Capital letters indicate translated sequences, and numbering of sequences is rela-tive to the translation start site. Thin arrows denote the boundaries of the promoter portions of the chimeric transfection con-structs with the unlabeled arrow showing the common 3' end (position —336). Thin arrows A, B, and C represent the 5' endsof these respective promoter portions (Constructs: A at —930, B at —1044, C at —1108). Putative transcription factor bind-ing sites are underlined as: TTF1 (thin line), TTF2 (thick line), and PAX-8 (broken line). The 5' ends of the cDNA clone ofSmanik et al. (2) (thick SJ arrow) and our three clones (thick VA1 arrow, one clone, and thick VA2 arrow, two clones) aremarked as indicated.
66 VENKATARAMAN ET AL.
sequenced for verification. The negative control (NC) isthe pGL3-Basic vector alone and the positive control (PC)is the pGL3-Control vector with an SV-40 enhancer andpromoter. Specific promoter fragments were generated byhigh-fidelity-PCR using primers introducing Kpnl (5') andXhol (3') restriction sites bracketing the promoter regions.KAT-50 (1 X 106 cells/9 cm2 well), HeLa (4 X 105),HepG2 (2 X 105) and COS-1 (2 X 105) were transientlytransfected, after 24 hours, with 1.0 fig plasmid DNA/0.625 pg lipofectamine complex in serum-free and phenolred-free medium. After five hours, medium was replacedand cultures incubated for 48 hours at 37CC in 5% C02.Luciferase activity was measured using commercial Re-porter Lysis Buffer and the Luciferase Assay Substrate(both from Promega) on a microtiter plate luminometer(Model ML2250, Dynatech Laboratories, Chantilly, VA).
RESULTS
Cloning of the hNIS cDNA and location oftranscription start site
We sequenced the cDNA for the hNIS coding region andverified that it replicated the published sequence of Smaniket al. (2) with the addition of further upstream coding se-
quences as indicated in Figure 1. Our 5' RACE analysis ofthe adapter ligated human thyroid cDNA, with the adapterprimer and two different downstream antisense primers(GSP2 and RX1) generated products of approximately 250
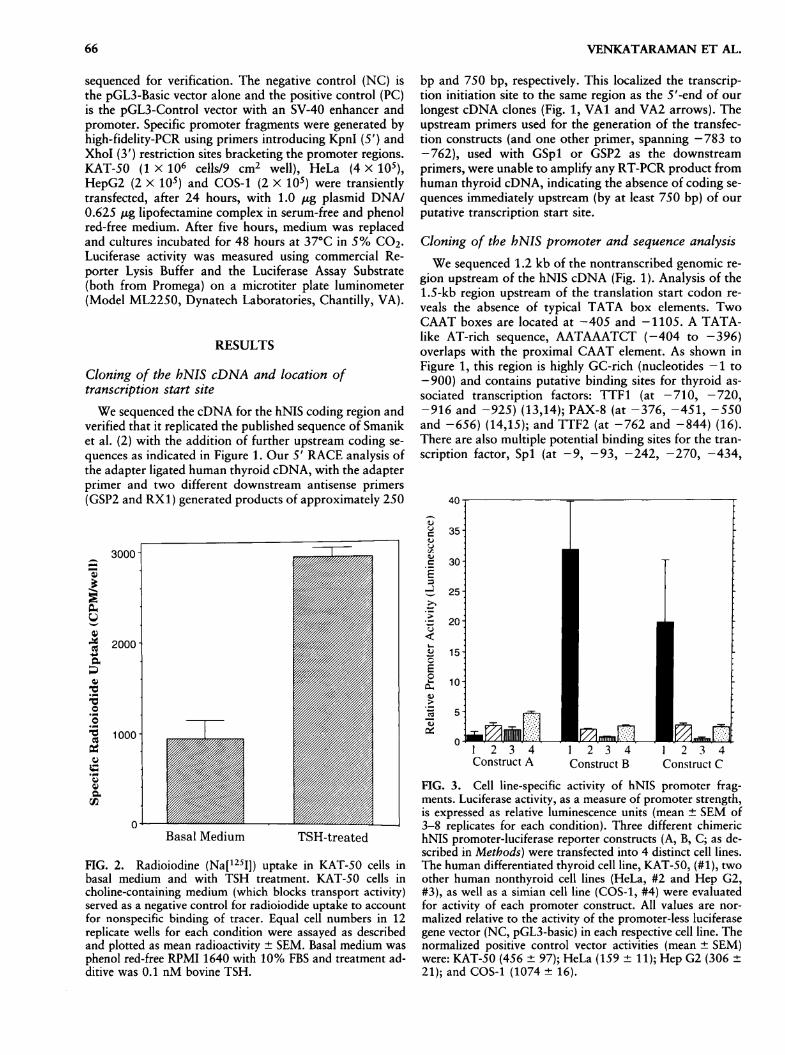
3000
OhU4)
aS30)
-a-5o
va
OSuS'oSa
ce
2000 i
1000
Basal Medium TSH-treated
FIG. 2. Radioiodine (Na[125Fj) uptake in KAT-50 cells inbasal medium and with TSH treatment. KAT-50 cells incholine-containing medium (which blocks transport activity)served as a negative control for radioiodide uptake to accountfor nonspecific binding of tracer. Equal cell numbers in 12replicate wells for each condition were assayed as describedand plotted as mean radioactivity ± SEM. Basal medium was
phenol red-free RPMI 1640 with 10% FBS and treatment ad-ditive was 0.1 nM bovine TSH.
bp and 750 bp, respectively. This localized the transcrip-tion initiation site to the same region as the 5'-end of our
longest cDNA clones (Fig. 1, VA1 and VA2 arrows). Theupstream primers used for the generation of the transfec-tion constructs (and one other primer, spanning —783 to
—
762), used with GSpl or GSP2 as the downstreamprimers, were unable to amplify any RT-PCR product fromhuman thyroid cDNA, indicating the absence of coding se-
quences immediately upstream (by at least 750 bp) of our
putative transcription start site.
Cloning of the hNIS promoter and sequence analysisWe sequenced 1.2 kb of the nontranscribed genomic re-
gion upstream of the hNIS cDNA (Fig. 1). Analysis of the1.5-kb region upstream of the translation start codon re-
veals the absence of typical TATA box elements. TwoCAAT boxes are located at -405 and -1105. A TATA-like AT-rich sequence, AATAAATCT (-404 to -396)overlaps with the proximal CAAT element. As shown inFigure 1, this region is highly GC-rich (nucleotides —1 to
—
900) and contains putative binding sites for thyroid as-
sociated transcription factors: TTF1 (at —710, —720,-916 and -925) (13,14); PAX-8 (at -376, -451, -550and -656) (14,15); and TTF2 (at -762 and -844) (16).There are also multiple potential binding sites for the tran-
scription factor, Spl (at -9, -93, -242, -270, -434,
oa.
12 3 4Construct A
2 3 4 12 3 4Construct B Construct C
FIG. 3. Cell line-specific activity of hNIS promoter frag-ments. Luciferase activity, as a measure of promoter strength,is expressed as relative luminescence units (mean ± SEM of3-8 replicates for each condition). Three different chimerichNIS promoter-luciferase reporter constructs (A, B, C; as de-scribed in Methods) were transfected into 4 distinct cell lines.The human differentiated thyroid cell line, KAT-50, (#1), twoother human nonthyroid cell lines (HeLa, #2 and Hep G2,#3), as well as a simian cell line (COS-1, #4) were evaluatedfor activity of each promoter construct. All values are nor-
malized relative to the activity of the promoter-less luciferasegene vector (NC, pGL3-basic) in each respective cell line. Thenormalized positive control vector activities (mean ± SEM)were: KAT-50 (456 ± 97); HeLa (159 ± 11); Hep G2 (306 ±21); and COS-1 (1074 ± 16).
HUMAN THYROID Na+-IODIDE SYMPORTER PROMOTER 67
-449, -537 and -861) (17). An unusual region of re-
peated GAAT units spans —1110 to -1057. In addition,a 14-nucleotide direct repeat element, GTGCATGG-GGATGG, separated by four bases, spans positions —574to -561 and -566 to -553.
Characterization of KAT-50 cell lineKAT-50 demonstrates log-phase growth at a doubling
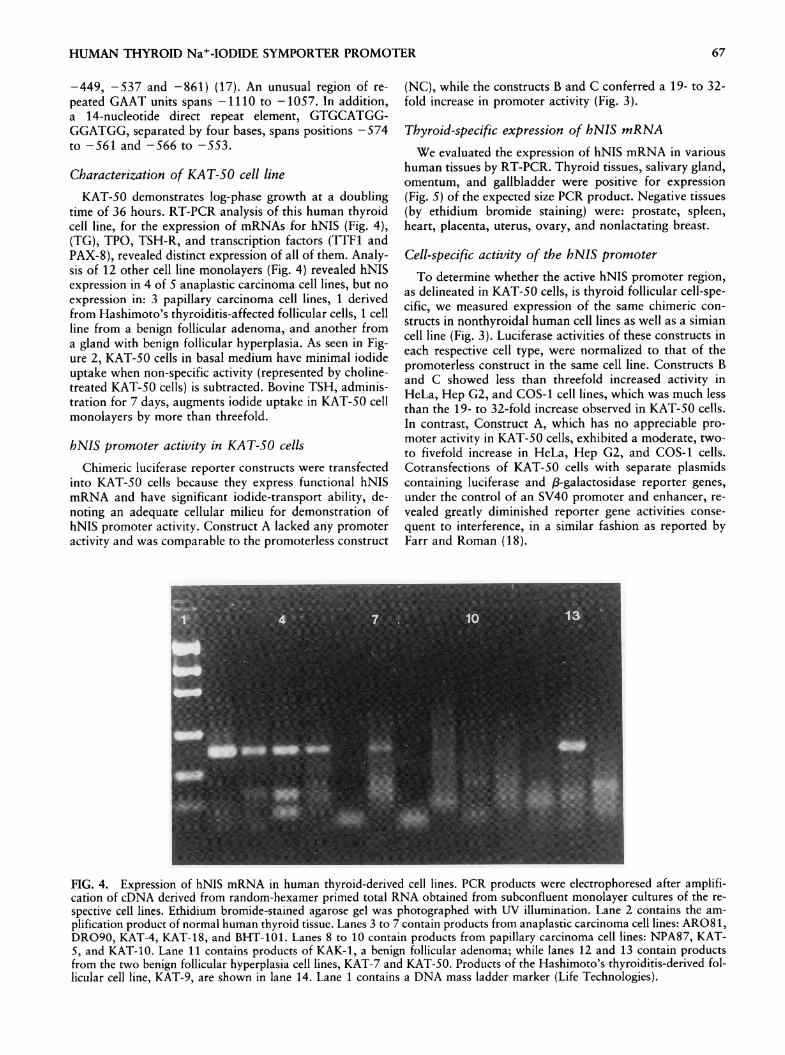
time of 36 hours. RT-PCR analysis of this human thyroidcell line, for the expression of mRNAs for hNIS (Fig. 4),(TG), TPO, TSH-R, and transcription factors (TTF1 andPAX-8), revealed distinct expression of all of them. Analy-sis of 12 other cell line monolayers (Fig. 4) revealed hNISexpression in 4 of 5 anaplastic carcinoma cell lines, but no
expression in: 3 papillary carcinoma cell lines, 1 derivedfrom Hashimoto's thyroiditis-affected follicular cells, 1 cellline from a benign follicular adenoma, and another froma gland with benign follicular hyperplasia. As seen in Fig-ure 2, KAT-50 cells in basal medium have minimal iodideuptake when non-specific activity (represented by choline-treated KAT-50 cells) is subtracted. Bovine TSH, adminis-tration for 7 days, augments iodide uptake in KAT-50 cellmonolayers by more than threefold.
hNIS promoter activity in KAT-50 cellsChimeric luciferase reporter constructs were transfected
into KAT-50 cells because they express functional hNISmRNA and have significant iodide-transport ability, de-noting an adequate cellular milieu for demonstration ofhNIS promoter activity. Construct A lacked any promoteractivity and was comparable to the promoterless construct
(NC), while the constructs B and C conferred a 19- to 32-fold increase in promoter activity (Fig. 3).
Thyroid-specific expression of hNIS mRNA
We evaluated the expression of hNIS mRNA in varioushuman tissues by RT-PCR. Thyroid tissues, salivary gland,omentum, and gallbladder were positive for expression(Fig. 5) of the expected size PCR product. Negative tissues(by ethidium bromide staining) were: prostate, spleen,heart, placenta, uterus, ovary, and nonlactating breast.
Cell-specific activity of the hNIS promoterTo determine whether the active hNIS promoter region,
as delineated in KAT-50 cells, is thyroid follicular cell-spe-cific, we measured expression of the same chimeric con-
structs in nonthyroidal human cell lines as well as a simiancell line (Fig. 3). Luciferase activities of these constructs ineach respective cell type, were normalized to that of thepromoterless construct in the same cell line. Constructs Band C showed less than threefold increased activity inHeLa, Hep G2, and COS-1 cell lines, which was much lessthan the 19- to 32-fold increase observed in KAT-50 cells.In contrast, Construct A, which has no appreciable pro-moter activity in KAT-50 cells, exhibited a moderate, two-to fivefold increase in HeLa, Hep G2, and COS-1 cells.Cotransfections of KAT-50 cells with separate plasmidscontaining luciferase and /3-galactosidase reporter genes,under the control of an SV40 promoter and enhancer, re-
vealed greatly diminished reporter gene activities conse-
quent to interference, in a similar fashion as reported byFarr and Roman (18).
FIG. 4. Expression of hNIS mRNA in human thyroid-derived cell lines. PCR products were electrophoresed after amplifi-cation of cDNA derived from random-hexamer primed total RNA obtained from subconfluent monolayer cultures of the re-
spective cell lines. Ethidium bromide-stained agarose gel was photographed with UV illumination. Lane 2 contains the am-
plification product of normal human thyroid tissue. Lanes 3 to 7 contain products from anaplastic carcinoma cell lines: AR081,DRO90, KAT-4, KAT-18, and BHT-101. Lanes 8 to 10 contain products from papillary carcinoma cell lines: NPA87, KAT-5, and KAT-10. Lane 11 contains products of KAK-1, a benign follicular adenoma; while lanes 12 and 13 contain productsfrom the two benign follicular hyperplasia cell lines, KAT-7 and KAT-50. Products of the Hashimoto's thyroiditis-derived fol-licular cell line, KAT-9, are shown in lane 14. Lane 1 contains a DNA mass ladder marker (Life Technologies).

VENKATARAMAN ET AL.
FIG. 5. Expression of hNIS mRNA in diverse normal human tissues. PCR products were electrophoresed after amplificationof cDNA derived from random-hexamer primed total RNA obtained from fresh surgical tissues. Ethidium bromide-stainedagarose gel was photographed with UV illumination. Lanes 2 to 12 contain amplification products of: thyroid, salivary gland,omentum, gallbladder, nonlactating breast, ovary, placenta, uterus, heart, prostate, and spleen, respectively. Lane 13 is a neg-ative control without target cDNA and lane 1 contains a DNA mass ladder marker (Life Technologies).
DISCUSSION
These results indicate successful identification of thehNIS gene promoter as well as delineation of a minimalessential regulatory sequence that confers both promoteractivity in differentiated human thyroid cells and cell-typespecificity. Thyroid-specific transcription factors (TTF1,TTF2, and PAX-8) have been associated with thyroid-spe-cific gene expression for TG, TSH-R, and TPO in numer-
ous species. Although Construct A contains potential bind-ing motifs for each of these transcription factors, this isnot sufficient for promoter activity in KAT-50 thyroid cellsand requires the participation of an additional 115 bp up-stream for activity of the chimeric constructs. We have not
yet identified a specific motif in this region to account forthis effect. The addition of 65 bp upstream of ConstructB, including the GAAT repeat region and the distal CAATbox (constituting Construct C) does not evoke additionalluciferase activity in our expression system.
Our independent analysis of the hNIS cDNA sequencewas the same as reported by Smanik et al. (2) with the ex-
ception of a longer untranslated upstream region. This re-
sult, in conjunction with the results of our 5'RACE analy-sis located the putative transcription start site at least 15bp upstream of the previously reported sequence. The rel-ative positions of the TATA-like sequence, the CAAT-box,and the Spl sites to this start site are in agreement withtheir consensus positions noted in the majority of verte-brate gene promoters (19).
The KAT-50 cell line is an important research tool forthe characterization of iodide transport in human thyroidcells. Because of its expression of the full repertoire ofknown thyroid-specific genes, its functional expression of
hNIS, as well as its demonstrated response to thyrotropin,it should prove to be as relevant to the study of humanthyrocyte biology as the FRTL-5 cell line has been for thestudy of rat thyrocyte biology (20).
RT-PCR for hNIS mRNA in 12 human thyroid cell linesrevealed expression of an appropriate-size product in 4 of5 anaplastic thyroid carcinoma cell lines. This is unexpected,considering the invariable failure of anaplastic tumors toconcentrate iodide in their human hosts. It is possible thatthis expressed hNIS mRNA is mutated or otherwise defec-tive consequent to splicing. Evaluation of iodide uptake inthese particular cell lines will be necessary to determine ifthis is associated with functional expression of symporterprotein. Smanik et al. (3) have demonstrated low and vari-able expression of hNIS mRNA in primary papillary carci-noma tissues while we were unable to see such expressionin 3 such cell lines. It is unclear whether this is related to
nonexpression in the original parent tumors or consequentto derivation of the cell lines or in vitro conditions.
In addition to expression of hNIS in normal human thy-roid tissues, we detected distinct expression in salivarygland, omentum, and gallbladder. Smanik et al. (3) havealso noted extrathyroidal expression in breast, colon, andovary. We have not been able to detect hNIS mRNA fromhuman breast or ovary by ethidium bromide staining. Clin-ically, patients subjected to radioiodine scanning demon-strate extrathyroidal uptake in salivary glands, gastric mu-
cosa, and intestines, with variable uptake in mammarytissues. The expression of hNIS in gallbladder is not un-
expected because this organ develops as an evagination ofthe embryonic gut (21) and can concentrate radioiodine indiagnostic scanning (22). Although clinical evidence of ra-
dioiodine uptake in human omentum has not been re-

HUMAN THYROID NaMODIDE SYMPORTER PROMOTER 69
ported, it would be difficult to discern from backgroundgut activity and is reasonable to expect because omentumderives from the embryonic mesogastrium (21).
Loss of hNIS expression in metastatic thyroid carcinomacreates a therapeutic dilemma because this renders ra-
dioiodine treatment ineffective and active systemicchemotherapies have not been found for this disease. Re-cent studies in vitro (23) and in vivo (24) suggest that lossof iodide transport is reversible with chemical agents, im-plying the absence of inactivating mutations. Further in-vestigations of hNIS gene regulation may elucidate thecause of loss of its activity and provide an approach to re-
store iodide transport.
ACKNOWLEDGMENTS
This work was supported by NCI Grant CA58935, VAMerit Review 596-0003, the Ephraim McDowell CancerResearch Foundation (Lexington, KY) and the LexingtonClinic Foundation (Lexington, KY). We thank S. Refetofffor helpful discussions and advice and K. D. Taylor for as-sistance with these experiments.
REFERENCES
1. Dai G, Levy O, Carrasco N 1996 Cloning and characteriza-tion of the thyroid iodide transporter. Nature 379:458^59.
2. Smanik PA, Liu Q, Furminger TL, Ryu K, Xing S, Mazza-ferri EL, Jhiang SM 1996 Cloning of the human sodium io-dide symporter. Biochem Biophys Res Comm 226:339-345.
3. Smanik PA, Ryu K-Y, Theil KS, Mazzaferri EL, Jhiang SM1997 Expression, exon-intron organization, and chromosomemapping of the human sodium iodide symporter. En-docrinology 138(8):3555-3558.
4. Gilbert SF, Migeon BR 1975 D-Valine as a selective agent fornormal human and rodent epithelial cells in culture. Cell5:11-17.
5. Kao WW-Y, Prockop DJ 1977 Proline analogue removes fi-broblasts from cultured mixed cell populations. Nature266:63-64.
6. Pályi I, Péter I, Daubner D, Vincze B, Lörincz I 1993 Estab-lishment, characterization and drug sensitivity of a new
anaplastic thyroid carcinoma cell line (BHT-101). VirchowsArchiv B Cell Pathol 63:263-269.
7. Aim KB, Tofiq S, Taylor KD 1996 Antineoplastic activity oftaxol against human anaplastic thyroid carcinoma cell linesin vitro and in vivo. J Clin Endocrinol Metab 81(10):3650-3653.
8. Ain KB, Taylor KD, Tofiq S, Venkataraman G 1997 So-matostatin receptor subtype expression in human thyroid andthyroid carcinoma cell lines. J Clin Endocrinol Metab82(6):1857-1862.
9. Ain KB, Taylor KD 1994 Somatostatin analogs affect prolif-eration of human thyroid carcinoma cell lines in vitro. J ClinEndocrinol Metab 78(5):1097-1102.
10. Chomczynski P, Sacchi N 1987 Singlee step method of RNA
isolation by acid guanidinium thiocyanate-phenol-chloroformextraction. Anal Biochem 162:156-159.
11. Cheng S, Fockler C, Barnes WM, Higuchi R 1994 Effectiveamplification of long targets from cloned inserts and humangenomic DNA. Proc Nati Acad Sei USA 91(12):5695-5699.
12. Frohman MA 1993 Rapid amplification of complementaryDNA ends for generation of full-length complementaryDNAs: Thermal RACE. Methods Enzymol 218:340-356.
13. Civitareale D, Lonigro R, Sinclair AJ, Di Lauro R 1989 Athyroid-specific nuclear protein essential for tissue-specific ex-
pression of the thyroglobulin promoter. EMBO J 8(9):2537-2542.
14. Civitariale D, Castelli MP, Falasca P, Saiardi A 1993Thyroid transcription factor 1 activates the promoter of thethyrotropin receptor gene. Mol Endocrinol 7(12):1589-1595.
15. Francis-Lang H, Zannini M, De Felice M, Berlingieri MT,Fusco A, Di Lauro R 1992 Multiple mechanisms of interfer-ence between transformation and differentiation in thyroidcells. Mol Cell Biol 12(12):5793-5800.
16. Santisteban P, Acebrón A, Polycarpou-Schwarz M, Di LauroR 1992 Insulin and insulin-like growth factor I regulate a thy-roid-specific nuclear protein that binds to the thyroglobulinpromoter. Mol Endocrinol 6(8):1310-1317.
17. Kadonaga JT, Coury AJ, Ladika, J, Tijian R 1988 Science242:1566-1570.
18. Farr A, Roman A 1992 A pitfall of using a second plasmidto determine transfection efficiency. Nucleic Acids Res20(4):920.
19. Bûcher P 1990 Weight matrix descriptions of four eukary-otic RNA polymerase II promoter elements derived from 502unrelated promoter sequences. J Mol Biol 212(4):563-578.
20. Bidey SP, Lambert A, Robertson WR 1988 Thyroid cellgrowth, differentiation and function in the FRTL-5 cell line:A survey. J Endocrinol 119:365-376.
21. Corliss CE 1976 Patten's Human Embryology: Elements ofClinical Development. McGraw-Hill Book Co, New York.
22. Achong DM, Oates E, Lee SL, Doherty FJ 1991 Gallbladdervisualization during post-therapy iodine-131 imaging of thy-roid carcinoma. J Nucl Med 32(12):2275-2277.
23. van Herle AJ, Agatep ML, Padua III DN, Totanes TL, Can-lapan DV, Van Herle HML, Juillard GJF 1990 Effects of 13cis-retinoic acid on growth and differentiation of human fol-licular carcinoma cells (UCLA RO 82 W-l) in vitro. J ClinEndocrinol Metab 71(3):755-763.
24. Simon D, Köhrle J, Schmutzler C, Mainz K, Reiners C, RoherH-D 1996 Redifferentiation therapy of differentiated thyroidcarcinoma with retinoic acid: Basics and first clinical results.Exp Clin Endocrinol Diabetes 104(Suppl 4):13-15.
Address correspondence to:Kenneth B. Ain, M.D.
Thyroid Nodule and Oncology Clinical ServiceDivision of Endocrinology and Molecular Medicine
Department of Internal Medicine, Room MN520University of Kentucky Medical Center
800 Rose Street,Lexington, Kentucky 40536-0084








![2-[( E )-2-(4Ethoxyphenyl)ethenyl]-1-methylquinolinium iodide dihydrate](https://static.fdokumen.com/doc/165x107/631e216b05964b686800aa35/2-e-2-4ethoxyphenylethenyl-1-methylquinolinium-iodide-dihydrate.jpg)











