Utilising Participatory Research Techniques For Community-Based Disaster Risk Assessment

This article was downloaded by: [Ibrahim Matter]On: 20 May 2013, At: 03:38Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registeredoffice: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK
Chemistry and EcologyPublication details, including instructions for authors andsubscription information:http://www.tandfonline.com/loi/gche20
Characterisation and identification ofcarbofuran-utilising bacteria isolatedfrom agricultural soilR. A.I. Abou-Shanab a , M. A. Khalafallah b , N. F. Emam c , M. A.Aly c , S. A. Abou-Sdera b & I. A. Matter ba Environmental Biotechnology Department, City of ScientificResearch and Technology Applications, Alexandria, Egyptb Agricultural Microbiology Department, National Research Center,Cairo, Egyptc Agricultural Microbiology Department, Cairo University, Cairo,EgyptPublished online: 04 Nov 2011.
To cite this article: R. A.I. Abou-Shanab , M. A. Khalafallah , N. F. Emam , M. A. Aly , S. A. Abou-Sdera & I. A. Matter (2012): Characterisation and identification of carbofuran-utilising bacteriaisolated from agricultural soil, Chemistry and Ecology, 28:2, 193-203
To link to this article: http://dx.doi.org/10.1080/02757540.2011.628317
PLEASE SCROLL DOWN FOR ARTICLE
Full terms and conditions of use: http://www.tandfonline.com/page/terms-and-conditions
This article may be used for research, teaching, and private study purposes. Anysubstantial or systematic reproduction, redistribution, reselling, loan, sub-licensing,systematic supply, or distribution in any form to anyone is expressly forbidden.
The publisher does not give any warranty express or implied or make any representationthat the contents will be complete or accurate or up to date. The accuracy of anyinstructions, formulae, and drug doses should be independently verified with primarysources. The publisher shall not be liable for any loss, actions, claims, proceedings,demand, or costs or damages whatsoever or howsoever caused arising directly orindirectly in connection with or arising out of the use of this material.

Chemistry and EcologyVol. 28, No. 2, April 2012, 193–203
Characterisation and identification of carbofuran-utilisingbacteria isolated from agricultural soil
R.A.I. Abou-Shanaba*, M.A. Khalafallahb, N.F. Emamc, M.A. Alyc, S.A. Abou-Sderab
and I.A. Matterb
aEnvironmental Biotechnology Department, City of Scientific Research and Technology Applications,Alexandria, Egypt; bAgricultural Microbiology Department, National Research Center, Cairo, Egypt;
cAgricultural Microbiology Department, Cairo University, Cairo, Egypt
(Received 16 December 2010; final version received 26 September 2011)
A total of 96 bacterial cultures were isolated from soil. Seventeen bacterial isolates were selected followingtheir cultivation on solid media containing 100 mg · L−1 carbofuran as the sole source of carbon andnitrogen. Of the 17 isolates, 10F, 11M, 17N, 23B and 26M were specifically chosen because of theirrelatively higher growth efficiency and genetic diversity based on Box-polymerase chain reaction analysis.These bacterial cultures had 16S rRNA gene sequences that were most similar to Acinetobacter baumannii(10F), Agrobacterium tumefaciens (11M), Ochrobactrum anthropi (17N), Escherichia coli (23B) andAgrobacterium tumefaciens (26M) with 97, 95, 93, 95 and 94% similarity in their 16S rDNA gene sequence,respectively. Degradation rates of carbofuran in soil inoculated with these isolates were 1.9, 1.5, 1.6, 1.7and 1.6 times, respectively, faster in comparison with uninoculated soil after 10 days of incubation. Themaximum degradation rates of carbofuran (45 and 91%) were detected in soil inoculated with A. baumannii(10F) after 10 and 20 days’ incubation, respectively. These data indicate that these isolates may have thepotential for use in bioremediation of pesticide contaminated soil.
Keywords: soil bacteria; Box-PCR; 16S rDNA; carbofuran; biodegradation
1. Introduction
Pesticides as synthetic organic compounds have been extensively used to protect and improvethe quality and quantity of food commodities, building materials, clothing and animal health, aswell as to combat diseases transmitted by insect vectors to man and animals [1]. Carbofuran,an N-methylcarbamate pesticide, is a soil-applied insecticide used extensively to control insectpests of corn, rice and other agricultural crops. Some N-methylcarbamate compounds and theirmetabolites might be sufficiently mobile in certain soils and are considered potential pollutantsof groundwater and surface water. The widespread agricultural use of pesticides has resulted inthese chemicals entering soil and water ecosystems [2,3]. Soil-applied pesticides reach surfacewater and groundwater by processes associated with run-off and leaching processes [4].
*Corresponding author. Email: [email protected]
ISSN 0275-7540 print/ISSN 1029-0370 online© 2012 Taylor & Francishttp://dx.doi.org/10.1080/02757540.2011.628317http://www.tandfonline.com
Dow
nloa
ded
by [
Ibra
him
Mat
ter]
at 0
3:38
20
May
201
3

194 R.A.I. Abou-Shanab et al.
The indiscriminate use of pesticides can cause toxic effects in non-target organisms [1]. Degra-dation of xenobiotic compounds by members of the soil microflora is an important means bywhich these compounds are removed from the environment, thus preventing them from becomingpollution problems. Bacteria, actinomycetes and fungi occupy a unique position in biologicalcycles and are essential for plant growth and soil fertility. Organic pesticides applied to soilmay be used as substrates by microorganisms and undergo degradation, resulting in the forma-tion of new compounds which may prove to be far more deleterious to plants than the originalmolecule [5]. Soil microorganisms which cause the breakdown of cellulose, nitrification, and theturnover of organic matter and other biological materials may also be adversely affected by pes-ticides [6]. Pesticides also affect reduction–oxidation enzymes, the virulence of bacteria and thenitrogen-fixing activity of Rhizobium [7]. Pesticide biodegradation by microorganisms has beendocumented in a wide range of habitats, including soils, sediments, surface and groundwater andsewage sludge [8]. The ubiquity of pesticide degradation suggests that bioremediation strategiesmay play an important role in the treatment of pesticide wastes [9]. For example, microorganismshave an important role in the metabolism of pesticides. Soil bacteria that utilise several pesti-cides have been isolated from soil. They include a metamitron-degrading Rhodococcus sp. [10],chloropyrifos-degrading Providencia stuartii, Pseudomonas diminuta and Pseudomonas putida[11,12], Isocarbophos-degrading Arthrobacter sp. [13], as well as carbofuran-degrading Pseu-domonas sp., Sphingomonas sp. and Burkholderia cepacia PCL3 [14–16]. Thus, the aim thisstudy was to isolate and characterise indigenous soil bacteria capable of utilising carbofuran as asource of carbon and nitrogen, evaluate the capability of bacterial isolates to degrade carbofuranin soil, and identify them by analysing the sequence of the gene encoding 16S rRNA.
2. Materials and methods
2.1. Chemicals
Carbofuran (75% active ingredient) was donated by Central Laboratory of Agriculture Pesti-cides, Agriculture Research Center (ARC), Ministry of Agriculture, Egypt. All solvents were ofhigh-performance liquid chromatography (HPLC) and analytical grade. Media used for bacterialisolation were high purity grade. The fine chemicals used for the molecular characterisation ofcarbofuran-utilising bacteria were purchased from Qiagen Inc., Promega and Fermentas.
2.2. Soil sampling and preparation
Soil samples were collected from Kafr El-Sheikh, Gharbia and Giza governorates and from newlycultivated soil at Noubaria region, Egypt. At each site, soil samples were collected from a depth of5–15 cm. The first soil sample was taken from wheat-cultivated soil at Kafr El-Sheikh which hasa long history (∼15 years) of pesticide application. The second soil was collected from Gharbiawhere clover has been grown with no pesticide application for at least 2 years. The third soil wastaken from a peanut field at Giza without pesticide application in the last year. Finally, the fourthsoil was collected from a horse bean cultivated soil in Noubaria region which had been treatedwith different pesticides (i.e. glyphosate, dimethoate, malathion, carbofuran and endosulfan) .Individually, soils from each site were mixed in large containers, dried at room temperatureand sieved. Soil mechanical analysis was carried out using the pipette method and the pH wasdetermined after mixing 1 g of soil in 2.5 mL water, according to Black et al. [17]. Organic carboncontent was measured by the rapid titration method and total nitrogen was determined usingthe Kjeldahl method [18]. The physical and chemical characteristics of the soils are presentedin Table 1.
Dow
nloa
ded
by [
Ibra
him
Mat
ter]
at 0
3:38
20
May
201
3

Chemistry and Ecology 195
Table 1. Selected characteristics of soil used in this study.
(%) (%)
Soil sample Clay Silt Sand Soil type OM TN pH
Kafr El-Sheikh 35 19 43 Clay loam 1.9 0.14 7.7Zifta 26 30 39 Clay loam 1.7 0.14 8.2El-Ayat 38 21 40 Clay loam 1.5 0.14 8.3Noubaria 11 1 87 Sandy 0.18 0.14 7.8
Note: OM, organic matter content; TN, total nitrogen.
2.3. Isolation and maintenance of carbofuran-utilising bacteria
Carbofuran-tolerant bacteria were obtained by enrichment culture in minimum mineral medium(MMM). MMM contained the following constitutes (in g · L−1): K2HPO4, 2.4; KH2PO4, 1.2;MgSO4 · 7H2O, 0.2; CaCl2 · 2H2O, 0.025 and Fe2(SO4)3, 0.008 [19]. The pH was adjusted to7.0, and the medium was autoclaved. Carbofuran was dissolved in methanol and sterilised byfiltration (0.22 μm pore size) prior to being added to the medium after autoclaving to preventthermal hydrolysis and the possible formation of degradation products. Ten grams of each soilsample was inoculated into 250-mL Erlenmeyer flasks containing 100 mL MMM amended with100 mg · L−1 of carbofuran. The inoculated flasks (three replicates) were incubated at 30 ◦C for7 days with constant shaking at 150 rpm.
Carbofuran-utilising bacteria were isolated and enumerated using the standard plate method.Aliquots (0.1 mL) of the 10-fold serial dilution of the above enrichment culture were spread onMMM agar plates amended with 100 mg · L−1 carbofuran as the sole carbon and nitrogen source.From each dilution, five plates were inoculated and incubated at 30 ◦C. Bacterial colonies werecounted every 2 days until maximal plate counts were obtained. Numbers were expressed as colonyforming units (cfu) · g soil−1. Bacterial isolates were streaked twice on MMM plates to ensurepurity of the cultures. Colonies with different morphological and cultural characteristics on themedium amended with carbofuran were chosen and stored at −80 ◦C in MMM broth containing50% glycerol for further studies.
2.4. Bacterial growth in presence of carbofuran
This experiment involved growing the bacteria in liquid MMM that contained 100 mg · L−1 car-bofuran as the sole source of carbon and nitrogen. Fifty millilitres of MMM broth (adjusted topH 7.0) were inoculated with 1 mL of mid-log phase cultures of each isolate. The cultures wereincubated by shaking at 30 ◦C for 6 days during which the growth of each isolate was determinedby viable cell enumeration as cfu · mL−1 by plating on the same media solidified with agar. Threereplicates were performed for each isolate.
2.5. Characterisation of carbofuran-utilising bacteria
Carbofuran-utilising bacteria were tested for their oxidase (1% tetramethyl p-phenylenediamine)and catalase (3% hydrogen peroxide) activities. Gram stain, shape and motility under the micro-scope were also observed. Subsequently, bacterial isolates were characterised by Box-PCR and16S rRNA gene sequence analysis. The DNA of 17 bacterial isolates was extracted from 10-mL bacterial cultures grown overnight in MMM according to the method described by Ausubelet al. [20].
Dow
nloa
ded
by [
Ibra
him
Mat
ter]
at 0
3:38
20
May
201
3

196 R.A.I. Abou-Shanab et al.
2.5.1. Box-PCR analysis
DNA amplification reactions of the bacterial isolates were conducted in an Eppen-dorf Mastercycler Gradient Thermocycler. The oligonucleotide primers BOXA1R (5′-CTACGGCAAGGCGACGCTGACG-3′) were used to amplify bacterial DNA [21]. DNA ampli-fication was carried out in a 50 μL reaction volume. DNA was amplified by mixing 1 μL ofthe template DNA, 5 μL of 10× polymerase reaction buffers, 4 μL of 25 mM MgCl2, 4 μL ofBOXA1R-primer, 4 μL of dNTPs (2.5 mM), 1 μL of 1 U · μL−1 Taq DNA polymerase and 31 μLwater per reaction. The PCR was initiated by incubating the reaction mixture at 95 ◦C for 10 min,followed by 30 cycles of 30 s at 94 ◦C, 1 min at 52 ◦C and 6.5 min at 65 ◦C. The reaction wasterminated with an extension step consisting of 16 min incubation at 65 ◦C. The PCR productswere analysed on 1.5% agarose gel and visualised by ultraviolet illumination after staining with0.5 μg · mL−1 ethidium bromide. The fingerprint patterns resulting from Box-PCR analyses werescored for each template DNA by recording the presence or absence of bands to construct arectangular binary matrix. The matrix was used to derive simple matching coefficients used inclustering analysis and the construction of dendrograms to illustrate the genetic relationshipsusing NTSYS-PC 2.0 [22].
2.5.2. 16S rRNA sequencing
Oligonucleotide primers with specificity for eubacterial 16S rRNA genes, primers 16Sa-F(5′-CGCTGGCGGCAGGCTTAACA-3′) and 16Sb-R (5′-CCAGCCGCAGGTTCCCCT-3′) wereused to amplify the 16S rRNA gene fragments with template DNA originating from bacterialculture and using PCR protocols described by van Berkum and Fuhrmann [23]. The followingconditions were used for DNA amplification: 35 cycles consisting of denaturation at 94 ◦C for0.5 min, annealing at 55 ◦C for 1 min and extension at 72 ◦C for 1.5 min, followed by a final exten-sion at 72 ◦C for 3 min. Amplified PCR products of the proper size (∼1500 bp fragment) wereconfirmed by electrophoresis of 10 μL subsamples through a 1% horizontal agarose gel contain-ing 0.5 μg · mL−1 ethidium bromide. PCR products were purified using QIAquick Spin columns(Qiagen Inc., Chatsworth, CA, USA). A Perkin–Elmer 377 DNA sequencer, in combination withDye Deoxy Terminator Cycle Sequencing Kit (Perkin–Elmer, Foster City, CA,N USA) was usedfor sequencing the purified PCR products as described previously [24]. A search of GenBankwith BLAST [25] was used to identify named bacterial species with 16S rRNA gene sequencessimilar to those of the isolates.
2.5.3. GenBank accession numbers
The partial sequences of 16S rDNA determined in this study have been deposited in the NationalCenter for Biotechnology Information (NCBI) database under accession no. GQ486031–486035.
2.6. Carbofuran degradation in soil
Bacterial cultures were grown in liquid MMM and harvested after 24 h of growth. Bacterial pelletswere washed twice with sterile distilled water. Bacterial suspensions in sterile distilled water wereadjusted to an absorbance at 600 nm of 0.5 (equivalent to ∼7.4 × 108 cfu · mL−1).
Equal amounts of sieved agricultural soil were collected from Gharbia governorate(150 g · flask−1) and added to a 500-mL Erlenmeyer flask, amended with carbofuran(10 mg · 100 g−1 dry soil) and thoroughly mixed to obtain uniform carbofuran distribution. Soilsamples were inoculated with 1 mL (7.4 × 108 cfu · mL−1) of each bacterial isolates individually
Dow
nloa
ded
by [
Ibra
him
Mat
ter]
at 0
3:38
20
May
201
3

Chemistry and Ecology 197
and the inoculums was thoroughly mixed into the soil with the addition of an appropriate amountof sterile distilled water required for moisture content adjustments to 50% of its water-holdingcapacity. Soil received the same amount of sterilised water without bacterial culture and the sameamount of insecticides solvent without carbofuran were used as the control. All flasks were incu-bated at 30 ◦C in the dark. Over time, soil samples were withdrawn in triplicate to evaluate thebacterial survival as well as percentage of carbofuran degradation.
2.6.1. Extraction of carbofuran from soil and analysis
Carbofuran was extracted from soil samples using the method described by Nelson and Cook[26]. Soils (50 g dry weight) were weighed into a 250-mL flask, 100 mL of dichloromethane wasadded, and the flask was shaken in the dark for 2 h at 25 ◦C on a rotary shaker (150 rpm). The soilmixture was filtered through a Buchner funnel under suction; the filter cake was washed threetimes with 20 mL of dichloromethane for each.All filtrates were passed through anhydrous sodiumsulfate and collected in a 250-mL flask and concentrated to ∼1 mL on a rotary evaporator, andsubsequently concentrated to dryness, then dissolved in 5 mL of petroleum ether, filtered (0.45 mmmembrane) and analysed by HPLC. HPLC analysis was performed using an Agilent 1100 modelinstrument with a diode array detector under the following conditions: cartridge column, Nova-Pak C18 (150 mm × 4.6 mm × 5 μm particle size); mobile phase, acetonitrile/water (70 : 30 byvolume); flow rate, 1 mL · min−1; detection wavelength, 247 nm; injection volume, 20 μL. Anexternal standard method was used for calibration.
3. Results and discussion
3.1. Isolation of carbofuran-utilising bacteria
Total culturable carbofuran-utilising bacteria were determined in soil samples collected fromNoubaria, Giza, Gharbia and Kafr El-Sheikh, with different histories of pesticide field treatment.Plates counts obtained were 50 × 104, 19 × 104, 18 × 104 and 4 × 104 cfu · g−1 dry soil, respec-tively. In total, 96 carbofuran-utilising bacterial cultures (28, 18, 21 and 29) were isolated fromKafr El-Sheikh, Gharbia, Giza and Noubria, respectively. The plate count data indicated that thenumbers of total viable bacteria in pesticide contaminated soils were higher than uncontaminatedsoil (soil with unknown pesticide history). Soil samples with a history of exposure to carbo-furan were consistently positive for the presence of carbofuran-utilising bacteria. Soil samplesobtained from areas with unknown history of carbofuran application contained few numbers ofviable carbofuran-degrading bacteria. The Kafr El-Sheikh soil with unknown history of carbofuranapplication harboured the lowest counts compared with other sites. These results are in agreementwith the hypothesis that application of pesticides promotes the evolution of microorganisms thatare capable of degrading these xenobiotic compounds in the soil [27].
3.2. Microbial growth in the presence of carbofuran
A total of 17 bacterial isolates, 8 from Kafr El-Sheikh (4M, 6M, 9M, 11M, 12M, 14M, 15M and26M); 4 from Gharbia (1B, 17B, 20B and 23B); 3 from Giza (17N, 18N and 20N), and 2 fromEl-Noubria (1F and 10F), were selected based on colony morphology (colourless, white, yellow-ish, etc.) and size (∼1.5 mm in diameter) to determine their survival in liquid MMM containing100 mg · L−1 carbofuran as a sole carbon and nitrogen source during 6 days of incubation. Theviable counts of most bacterial isolates reached a maximum after 2 days incubation and then tended
Dow
nloa
ded
by [
Ibra
him
Mat
ter]
at 0
3:38
20
May
201
3
198 R.A.I. Abou-Shanab et al.
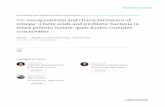
Table 2. Growth counts of bacterial isolates in MMM broth containing 100 mg · L−1 carbofuran as a carbon andnitrogen source.
Viable count × 105 cfu · mL−1 at regular intervals
Day
Isolate no. 0 1 2 3 4 5 6
4M 0.6 8 320 163 155 98 876M 0.5 181 329 400 438 424 4209M 4.2 126 268 269 274 268 25511M 0.1 564 1321 800 233 254 19012M 0.08 130 360 364 378 310 24414M 3.6 46 130 112 62 51 4815M 0.56 190 339 260 158 396 66526M 0.8 230 500 530 597 570 5321B 1 6 120 62 4 2 ND17B 0.6 189 410 320 200 154 13520B 2.2 72 143 194 228 248 28823B 4.5 324 820 586 308 310 33817N 0.4 98 480 210 156 119 8718N 0.04 0.2 0.3 7 17 11 220N 0.4 130 232 155 67 100 1381F 0.8 154 308 264 190 258 29010F 0.6 554 1243 765 468 413 380
Note: cfu, colony-forming unit.
to decrease with increasing incubation time (Table 2). The maximum viable count (13 × 107,12 × 107, 8 × 107, 5 × 107 and 2 × 107 cfu · mL−1) was obtained after 2 days’ incubation withbacterial isolates 11M, 10F, 23B, 26M and 17N, respectively. After 4 days’ incubation, isolates6M and 26M reached maximum counts of 4 × 107 and 6 × 107 cfu · mL−1, respectively, whereasisolates 20B and 15M showed a continued increase throughout incubation. Jilani and Khan [28]observed that the viable count of Pseudomonas (IES-Ps-1) exposed to different concentrationsand types of pesticides increased with time. The total viable count increased from 28 × 107 and93 × 107 to 197 × 107 cfu · mL−1 at 4, 6 and 24 h, respectively, showing that the culture enteredthe lag phase after 5 h and remain in that phase during 24 h of incubation.
López et al. [29] reported that the numbers of total viable bacteria in water samples amendedwith 10 and 50 μg · mL−1 pesticides were higher than in unamended controls. After treatment ofwater samples with aldrin, lindane, dimetoate, captan and atrazine at concentrations of 10 and50 μg · mL−1, bacterial populations increased significantly after 1 day, whereas when water sam-ples were amended with methidation, methylparation and diflubenzuron, the number of bacteriadid not exceed the control until day 7. However, after treatment with simazine at concentrationsof 10 and 50 μg · mL−1, bacterial microbiota did not increase until 14 days of incubation. Wesuggest that the decreasing counts only at extended times of incubation were due to the inhibitoryeffect of carbofuran byproducts. The increase in the biomass of bacterial cultures with carbofurancan be explained by the fact that this product constitutes an additional carbon and nitrogen sourcewhich allows the synthesis of new secondary metabolites favouring the production of microbialbiomass and in consequence supports faster use of carbofuran [30].
3.3. Characterisation of an isolated carbofuran-utilising bacterium
Box-PCR analyses of the 17 bacterial isolates that tolerated 100 mg · L−1 carbofuran led to aminimum of 8 and a maximum of 13 discrete visible bands ranging in size from 0.2 to 4.2 kb. Fromthe Box-PCR patterns of these cultures it was concluded that these isolates were more diverse.
Dow
nloa
ded
by [
Ibra
him
Mat
ter]
at 0
3:38
20
May
201
3
Chemistry and Ecology 199
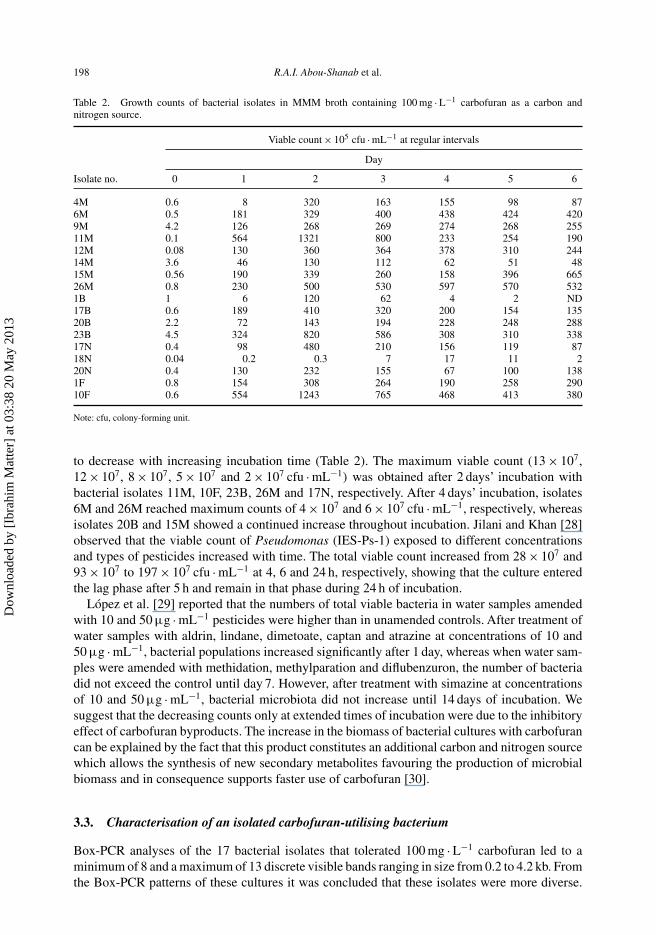
Figure 1. Dendogram showing the degree of similarity between Box-PCR fingerprints of carbofuran-utilising bacteria.
The profiles obtained (the presence or absence of band) were scored and a dendrogram wasgenerated using (STASTICA-6). The dendrogram derived from the Box-PCR dataset showedthat isolates 6M, 9M, 11M, 12M, 15M and 20B shared 100% similarity (Figure 1). Isolates4M, 18N, 14M and 26M shared 90% similarity, whereas isolates 1B, 20N and 23B had ∼86%similarity. Only isolates 1F, 10F, 17N and 17B shared a similarity level <83%. PCR-based methodsof fingerprinting take advantage of the presence of repetitive sequences that are interspersedthroughout the genome of diverse bacterial species. Amplification of the DNA sequences betweenprimers based on these repeated elements generate an array of differently sized DNA fragmentsfrom the genomes of individual strains. Separation of these fragments on agarose gels yieldshighly specific DNA fingerprints [21]. Genomic DNA fingerprinting using BOXA1R primer hasbeen found to be useful in differentiating between very closely related bacterial strains [21].
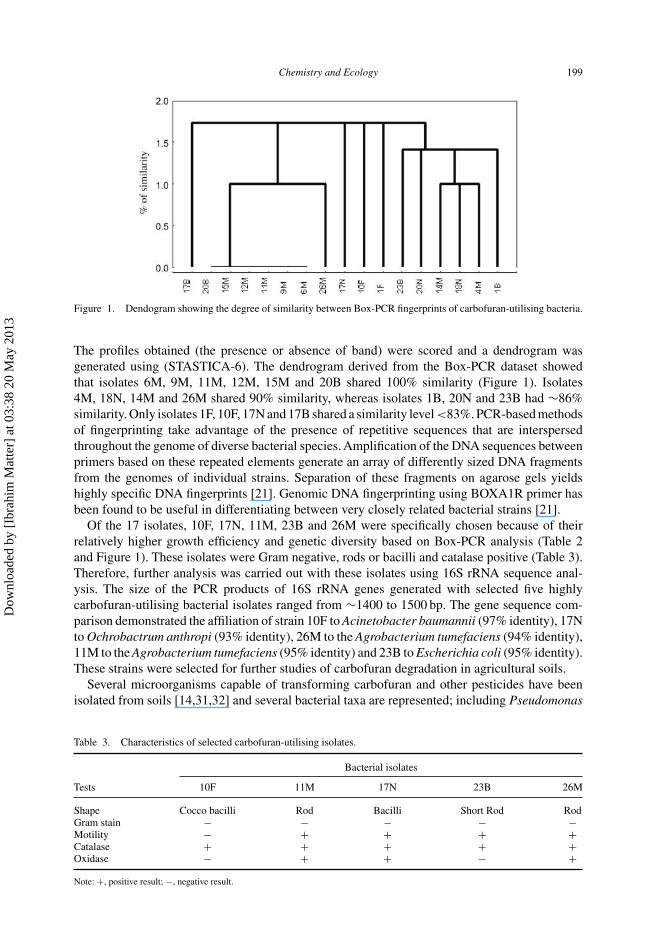
Of the 17 isolates, 10F, 17N, 11M, 23B and 26M were specifically chosen because of theirrelatively higher growth efficiency and genetic diversity based on Box-PCR analysis (Table 2and Figure 1). These isolates were Gram negative, rods or bacilli and catalase positive (Table 3).Therefore, further analysis was carried out with these isolates using 16S rRNA sequence anal-ysis. The size of the PCR products of 16S rRNA genes generated with selected five highlycarbofuran-utilising bacterial isolates ranged from ∼1400 to 1500 bp. The gene sequence com-parison demonstrated the affiliation of strain 10F to Acinetobacter baumannii (97% identity), 17Nto Ochrobactrum anthropi (93% identity), 26M to the Agrobacterium tumefaciens (94% identity),11M to the Agrobacterium tumefaciens (95% identity) and 23B to Escherichia coli (95% identity).These strains were selected for further studies of carbofuran degradation in agricultural soils.
Several microorganisms capable of transforming carbofuran and other pesticides have beenisolated from soils [14,31,32] and several bacterial taxa are represented; including Pseudomonas
Table 3. Characteristics of selected carbofuran-utilising isolates.
Bacterial isolates
Tests 10F 11M 17N 23B 26M
Shape Cocco bacilli Rod Bacilli Short Rod RodGram stain − − − − −Motility − + + + +Catalase + + + + +Oxidase − + + − +Note: +, positive result; −, negative result.
Dow
nloa
ded
by [
Ibra
him
Mat
ter]
at 0
3:38
20
May
201
3
200 R.A.I. Abou-Shanab et al.
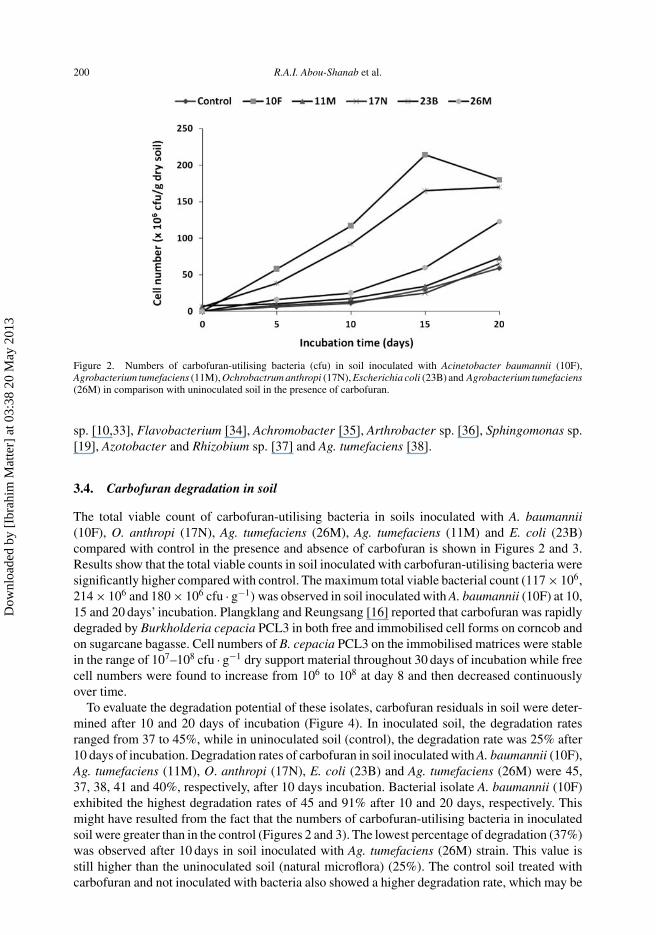
Figure 2. Numbers of carbofuran-utilising bacteria (cfu) in soil inoculated with Acinetobacter baumannii (10F),Agrobacterium tumefaciens (11M), Ochrobactrum anthropi (17N), Escherichia coli (23B) and Agrobacterium tumefaciens(26M) in comparison with uninoculated soil in the presence of carbofuran.
sp. [10,33], Flavobacterium [34], Achromobacter [35], Arthrobacter sp. [36], Sphingomonas sp.[19], Azotobacter and Rhizobium sp. [37] and Ag. tumefaciens [38].
3.4. Carbofuran degradation in soil
The total viable count of carbofuran-utilising bacteria in soils inoculated with A. baumannii(10F), O. anthropi (17N), Ag. tumefaciens (26M), Ag. tumefaciens (11M) and E. coli (23B)compared with control in the presence and absence of carbofuran is shown in Figures 2 and 3.Results show that the total viable counts in soil inoculated with carbofuran-utilising bacteria weresignificantly higher compared with control. The maximum total viable bacterial count (117 × 106,214 × 106 and 180 × 106 cfu · g−1) was observed in soil inoculated with A. baumannii (10F) at 10,15 and 20 days’ incubation. Plangklang and Reungsang [16] reported that carbofuran was rapidlydegraded by Burkholderia cepacia PCL3 in both free and immobilised cell forms on corncob andon sugarcane bagasse. Cell numbers of B. cepacia PCL3 on the immobilised matrices were stablein the range of 107–108 cfu · g−1 dry support material throughout 30 days of incubation while freecell numbers were found to increase from 106 to 108 at day 8 and then decreased continuouslyover time.
To evaluate the degradation potential of these isolates, carbofuran residuals in soil were deter-mined after 10 and 20 days of incubation (Figure 4). In inoculated soil, the degradation ratesranged from 37 to 45%, while in uninoculated soil (control), the degradation rate was 25% after10 days of incubation. Degradation rates of carbofuran in soil inoculated with A. baumannii (10F),Ag. tumefaciens (11M), O. anthropi (17N), E. coli (23B) and Ag. tumefaciens (26M) were 45,37, 38, 41 and 40%, respectively, after 10 days incubation. Bacterial isolate A. baumannii (10F)exhibited the highest degradation rates of 45 and 91% after 10 and 20 days, respectively. Thismight have resulted from the fact that the numbers of carbofuran-utilising bacteria in inoculatedsoil were greater than in the control (Figures 2 and 3). The lowest percentage of degradation (37%)was observed after 10 days in soil inoculated with Ag. tumefaciens (26M) strain. This value isstill higher than the uninoculated soil (natural microflora) (25%). The control soil treated withcarbofuran and not inoculated with bacteria also showed a higher degradation rate, which may be
Dow
nloa
ded
by [
Ibra
him
Mat
ter]
at 0
3:38
20
May
201
3
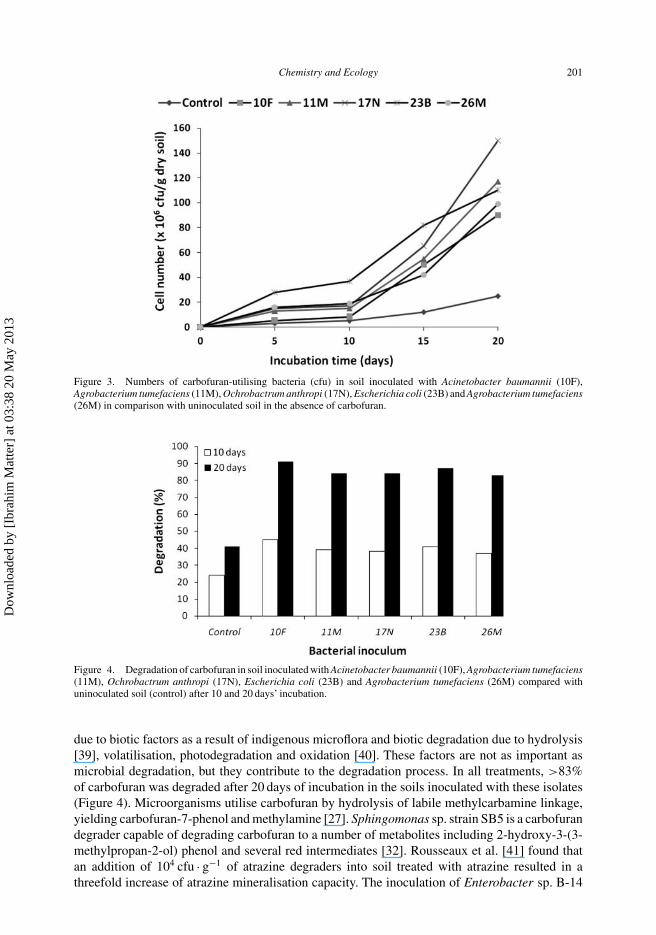
Chemistry and Ecology 201
Figure 3. Numbers of carbofuran-utilising bacteria (cfu) in soil inoculated with Acinetobacter baumannii (10F),Agrobacterium tumefaciens (11M), Ochrobactrum anthropi (17N), Escherichia coli (23B) and Agrobacterium tumefaciens(26M) in comparison with uninoculated soil in the absence of carbofuran.
Figure 4. Degradation of carbofuran in soil inoculated with Acinetobacter baumannii (10F), Agrobacterium tumefaciens(11M), Ochrobactrum anthropi (17N), Escherichia coli (23B) and Agrobacterium tumefaciens (26M) compared withuninoculated soil (control) after 10 and 20 days’ incubation.
due to biotic factors as a result of indigenous microflora and biotic degradation due to hydrolysis[39], volatilisation, photodegradation and oxidation [40]. These factors are not as important asmicrobial degradation, but they contribute to the degradation process. In all treatments, >83%of carbofuran was degraded after 20 days of incubation in the soils inoculated with these isolates(Figure 4). Microorganisms utilise carbofuran by hydrolysis of labile methylcarbamine linkage,yielding carbofuran-7-phenol and methylamine [27]. Sphingomonas sp. strain SB5 is a carbofurandegrader capable of degrading carbofuran to a number of metabolites including 2-hydroxy-3-(3-methylpropan-2-ol) phenol and several red intermediates [32]. Rousseaux et al. [41] found thatan addition of 104 cfu · g−1 of atrazine degraders into soil treated with atrazine resulted in athreefold increase of atrazine mineralisation capacity. The inoculation of Enterobacter sp. B-14
Dow
nloa
ded
by [
Ibra
him
Mat
ter]
at 0
3:38
20
May
201
3

202 R.A.I. Abou-Shanab et al.
(106 cfu · g−1) to soil with a low indigenous population of chlorpyrifos-degrading bacteria mixedwith 35 mg chlorpyrifos · kg soil−1 resulted in a higher degradation rate than was observed innon-inoculated soils [42]. Struthers et al. [43] found that inoculum levels of an Agrobacterimstrain as low as 105 cfu · g−1 into soils was adequate to rapidly degrade atrazine.
4. Conclusions
In view of these studies, it is suggested that bacterial strains isolated from these agricultural soilswere able to grow in the presence of carbofuran pesticides. Pesticide tolerance in these bacteriamay be important in the bioremediation of polluted environments by the pesticides.
Acknowledgements
The authors are very grateful to Prof. J. Scott Angle, College of Agricultural and Environmental Sciences, Athens, GA30602 USA for his valuable comment to improving this manuscript.
References
[1] C.S. Benimeli, M.J. Amoroso, A.P. Chaile, and G.R. Castro, Isolation of four aquatic Streptomycetes strains capableof growth on organochlorine pesticides, Bioresource Technol. 89 (2003), pp. 133–138.
[2] I.R. Hill and S.J.L. Wright, Pesticide Microbiology: Microbiological Aspects of Pesticide Behaviour in theEnvironment. Academic Press, London, 1978.
[3] R.L. Jones and R.C. Black, Monitoring aldicarb residues in Florida soil and water, Environ. Toxicol. Chem. 3(1984), pp. 9–20.
[4] A.G.S. Prado and C. Airoldi, The toxic effect on soil microbial activity caused by the free or immobilized pesticidediuron, Thermochim. Acta 394 (2002), pp. 155–162.
[5] N.S. Subba Rao, Soil Microorganisms and Plant Growth, 4th edn, IBP Publishing, Oxford, 1995.[6] K. Anu and S.K. Gosal, Effect of pesticide application on soil microorganisms, Arch. Agron. Soil Sci. 57 (2011),
pp. 569–596.[7] L.N. Paromenskaya, The symbiotic relations of rhizobium and leguminous plants exposed to herbicides, Byull. Vses.
Nauchno Issledovatelskogo Inst. Selskokhozyaistvennoi Microbiol. 32 (1979), pp. 11–13.[8] A. Nawab, A. Aleem, and A. Malik, Determination of organochlorine pesticides in agricultural soil with special
reference to -HCH degradation by Pseudomonas strains, Bioresource Technol. 88 (2003), pp. 41–46.[9] D.R. Shelton, S. Khader, J.S. Karns, and B.M. Pogell, Metabolism of twelve herbicides by Streptomyces,
Biodegradation 7 (1997), pp. 129–136.[10] N.R. Parekh, A. Hartman, M.P. Charnay, and J.C. Fournier, Diversity of carbofuran-degrading soil bacteria
and detection of plasmid-encoded sequences homologous to the mcd gene, FEMS Microbiol. Ecol. 17 (1995),pp. 149–160.
[11] N.L. Rani and D.L. Kumari, Degradation of methyl parathion by Pseudomonas putida, Can. J. Microbiol. 4 (1994),pp. 1000–1004.
[12] M.S. Rani, L.K.Vijaya, D.P. Suvarnalatha, R. Jaya Madhuri, S.Aruna, K. Jyothi, G. Narasimha, and K.Venkateswarlu,Isolation and characterization of a chlorpyrifos degrading bacterium from agricultural soil and its growth response,Afr. J. Microbiol. Res. 2 (2008), pp. 26–31.
[13] L. Rong, X. Guo, K. Chen, J. Zhu, S. Li, and J. Jiang, Isolation of an isocarbophos-degrading strain of Arthrobactersp. Sc1-2 and identification of the degradation pathway, J. Microbiol. Biotechnol. 19 (2009), pp. 1439–1446.
[14] N. Bano and J. Musarra, Characterization of a novel carbofuran degrading Pseudomonas sp. with collateralbiocontrol and plant growth promoting potential, FEMS Microbiol. Lett. 231 (2004), pp. 13–17.
[15] R.P. Myung, S. Lee, T. Han, B. Oh, J. Shin, and I. Kim, A new intermediate in the degradation of carbofuran bySphingomonas sp. strain SB5, J. Microbiol. Biotechnol. 16 (2006), pp. 1306–1310.
[16] P. Plangklang and A. Reungsang, Bioaugmentation of carbofuran residues in soil using Burkholderia cepacia PCL3adsorbed on agricultural residues, Int. Biodeterior. Biodegrad. 63 (2009), pp. 515–522.
[17] B.C. Black, D.D. Evans, J.I. White, L.E. Ensminger, and F.E. Clark, Methods of Soil Analysis, American Society ofAgronomy, Madison, WI, 1982.
[18] D.W. Nelson and L.E. Sommers, Total nitrogen analysis of soil and plant tissues, J. Ass. Off. Anal. Chem. 63 (1980),pp. 770–779.
[19] X. Feng, L.T. Ou, and A. Ogram, Plasmid mediated mineralization of carbofuran by Sphingomonas sp. strain CF06,Appl. Environ. Microbiol. 63 (1997), pp. 1332–1337.
Dow
nloa
ded
by [
Ibra
him
Mat
ter]
at 0
3:38
20
May
201
3

Chemistry and Ecology 203
[20] F.M. Ausubel, R. Brent, R.E. Kingston, D.D. Moore, J.G. Seidman, J.A. Smith, and K. Struhl, Short Protocols inMolecular Biology, 4th edn, Wiley, New York, 1999.
[21] J. Versalovic, M. Schneider, F.J. de Bruijn, and J.R. Lupski, Genomic fingerprinting of bacteria using repetitivesequence-based polymerase chain reaction, Methods Cell Biol. 5 (1994), pp. 25–40.
[22] F.J. Rohlf, NTSYS-PC Numerical Taxonomy and Multivariate Analysis System, Exeter Publishing, Setauket, NY,1988.
[23] P. van Berkum and J.J. Fuhrmann, Evolutionary relationships among the soybean bradyrhizobia reconstructed from16S rRNA gene and internally transcribed spacer region sequence divergence, Int. J. Syst. Evol. Microbiol. 50(2000), pp. 2165–2172.
[24] P. van Berkum, D. Beyene, and B.D. Eardly, Phylogenetic relationships among Rhizobium species nodulating thecommon bean (Phaseolus vulgarisL.), Int. J. Syst. Bacteriol. 46 (1996), pp. 240–244.
[25] S.F. Altschul, L.M. Thomas, A.S. Alejandro, Z. Jinghui, M. Webb, and J.L. David, Gapped BLAST and PSt-BLAST:a new generation of protein database search programs, Nucleic Acids Res. 25 (1997), pp. 3389–3402.
[26] T.R. Nelson and R.F. Cook, High pressure liquid chromatographic method for the determination of carbofuranresidues in soil and water, J. Agr. Food Chem. 27 (1979), pp. 1186–1188.
[27] G.R. Chaudhry and A.N. Ali, Bacterial metabolism of carbofuran, Appl. Environ. Microbiol. 54 (1988),pp. 1414–1419.
[28] S. Jilani and M.A. Khan, Isolation, characterization and growth response of pesticides degrading bacteria, J. Biol.Sci. 4 (2004), pp. 15–20.
[29] L.C. Lo’ Pez, B. Pozo, C. Rodelas, B. Calvo, M.V. Jua’ Rez, J. Martiñez-Toledo, and González-López, Identifi-cation of bacteria isolated from an oligotrophic lake with pesticide removal capacities, Ecotoxicology 14 (2005),pp. 299–312.
[30] M. Slaoui, M. Ouhssine, E. Berny, and M. Elyachioui, Biodegradation of the carbofuran by a fungus isolated fromtreated soil, Afr. J. Biotechnol. 6 (2007), pp. 419–423.
[31] J.S. Karns, W.W. Mulbry, J.O. Nelson, and P.C. Kearney, Metabolism of carbofuran by a pure bacterial culture,Pest. Biochem. Physiol. 25 (1986), pp. 211–217.
[32] I.S. Kim, J.Y. Ryu, H.G. Hur, M.B. Gu, S.D. Kim, and J.H. Shim, Sphingomonas sp. strain SB5 degrades carbofuranto a new metabolite by hydrolysis at the furanyl ring, J. Agr. Food Chem. 52 (2004), pp. 2309–2314.
[33] M. Slaoui, M. Ouhsssine, M. Massoui, and M. Elyachioui, Degradation du carbofuran par une bacterie du genrePseudomonas sp isolee a partir du sol, Sci Lett. 3 (2001), p. 3.
[34] S. Chapalamadugu and G.R. Chaudhry, Hydrolysis of carbaryl by a Pseudomonas sp. and construction of a microbialconsortium that completely metabolizes carbaryl, Appl. Environ. Microbiol. 57 (1991), pp. 744–750.
[35] G.R. Chaudhry, A.N. Ali, and W.B. Wheeler, Isolation of a methyl parathion-degrading Pseudomonas sp. thatpossesses DNA homologous to the opd gene from a Flavobacterium sp., Appl. Environ. Microbiol. 54 (1988),pp. 288–293.
[36] K. Ramanand, M.Sharmila, and N. Sethunathan, Mineralization of carbofuran by a soil bacterium, Appl. Environ.Microbiol. 54 (1988), pp. 2129–2133.
[37] S. Shafiani, and Abdul Mailk, Tolerance of pesticides and antibiotic resistance in bacteria isolated from wastewaterirrigated soil, World J. Microbiol. Biotechnol. 19 (2003), pp. 897–901.
[38] D. Smith, A. Sam, and D.E. Crowley, Cooperative catabolic pathways within an atrazine-degrading enrichmentculture isolated from soil, FEMS Microbiol. Ecol. 53 (2005), pp. 265–273.
[39] S.L. Trabue, A.V. Ogram, and L.T. Ou, Dynamics of carbofuran-degrading microbial communities in soil duringthree successive annual applications of carbofuran, Soil Biol. Biochem. 33 (2001), pp. 75–81.
[40] J.O. Lalah, S.O. Wandiga, and W.C. Dauterman, Mineralization, volatilization, and degradation of carbofuran insoil samples from Kenya, Bull. Environ. Contamin. Toxicol. 56 (1996), pp. 37–41.
[41] S. Rousseaux,A. Hartmann, B. Lagacherie, S. Piutti, F.Andreux, and G. Soulas, Inoculation of an atrazine-degradingstrain, Chelatobacter heintzii Cit1, in four different soils: Effects of different inoculum densities, Chemosphere 51(2003), pp. 569–576.
[42] B.K. Singh, A. Walker, J.A.W. Morgan, and D.J. Wright, Biodegrdation of chlorpyrifos by Enterbacter strain B-14and its use in bioremediation of contaminated soils, Appl. Environ. Microbiol. 70 (2004), pp. 4855–4863.
[43] J.K. Struthers, K. Jauachandran, and T.B. Moorman, Biodegradation of atrazine by Agrobacterium radiobacter J14aand use of this strain in bioremediation of contaminated soil, Appl. Environ. Microbiol. 64 (1998), pp. 3368–3375.
Dow
nloa
ded
by [
Ibra
him
Mat
ter]
at 0
3:38
20
May
201
3
Copyright © 2022 FDOKUMEN