
Census of bacterial microbiota associated with the glacier ice ...
10
FEMS Microbiology Ecology, 91, 2015, fiv003 doi: 10.1093/femsec/fiv003 Advance Access Publication Date: 12 January 2015 Research Article RESEARCH ARTICLE Census of bacterial microbiota associated with the glacier ice worm Mesenchytraeus solifugus Takumi Murakami 1, ∗ , Takahiro Segawa 2, 3 , Dylan Bodington 1 , Roman Dial 4 , Nozomu Takeuchi 5 , Shiro Kohshima 6 and Yuichi Hongoh 1 1 Department of Biological Sciences, Tokyo Institute of Technology, Tokyo 152-8550, Japan, 2 National Institute of Polar Research, Tokyo 190-8518, Japan, 3 Transdisciplinary Research Integration Center, Tokyo 190-8518, Japan, 4 Department of Environmental Science, Alaska Pacific University, Alaska 99508, USA, 5 Department of Earth Sciences, Chiba University, Chiba 263-8522, Japan and 6 Wildlife Research Center, Kyoto University, Kyoto 606-8203, Japan ∗ Corresponding author: Department of Biological Sciences, Tokyo Institute of Technology, 2-12-1-W3-48, Ookayama, Meguro-ku, Tokyo 152-8550, Japan. Tel: +81-3-5734-2865; Fax: +81-3-5734-2946; E-mail: [email protected] One sentence summary: We analyzed the taxonomic composition of bacteria physically associated with the glacier-adapted worm Mesenchytraeus solifugus, and provided the first report of symbiosis between bacteria and the glacier invertebrates. Editor: Max H ¨ aggblom ABSTRACT The glacier ice worm, Mesenchytraeus solifugus, is a unique annelid, inhabiting only snow and ice in North American glaciers. Here, we analyzed the taxonomic composition of bacteria associated with M. solifugus based on the 16S rRNA gene. We analyzed four fixed-on-site and 10 starved ice worm individuals, along with glacier surface samples. In total, 1341 clones of 16S rRNA genes were analyzed for the ice worm samples, from which 65 bacterial phylotypes (99.0% cut-off) were identified. Of these, 35 phylotypes were closely related to sequences obtained from their habitat glacier and/or other components of cryosphere; whereas three dominant phylotypes were affiliated with animal-associated lineages of the class Mollicutes. Among the three, phylotype Ms-13 shared less than 89% similarity with database sequences and was closest to a gut symbiont of a terrestrial earthworm. Using fluorescence in situ hybridization, Ms-13 was located on the gut wall surface of the ice worms. We propose a novel genus and species, ‘Candidatus Vermiplasma glacialis’, for this bacterium. Our results raise the possibility that the ice worm has exploited indigenous glacier bacteria, while several symbiotic bacterial lineages have maintained their association with the ice worm during the course of adaptive evolution to the permanently cold environment. Keywords: glacier ecology; enchytraeid; symbiosis; 16S rRNA; gut bacteria INTRODUCTION On glaciers, which are extreme environments characterized by low temperature and poor nutrients, a variety of psychrophilic or psychrotolerant organisms form unique ecosystems (Ane- sio and Laybourn-Parry 2012). Many studies on their diversity, physiology and the impact of their activity on the surrounding environment have been reported (Takeuchi, Kohshima and Seko 2001; Napolitano and Shain 2004; Anesio et al., 2009; Cameron, Hodson and Osborn 2012; Lutz et al., 2014); however, most of these investigations have focused on microbial communities on Received: 10 October 2014; Accepted: 30 December 2014 C FEMS 2015. All rights reserved. For permissions, please e-mail: [email protected] 1 Downloaded from https://academic.oup.com/femsec/article/91/3/fiv003/433893 by guest on 07 February 2022
-
Upload
khangminh22 -
Category
Documents
-
view
3 -
download
0
Transcript of Census of bacterial microbiota associated with the glacier ice ...

FEMS Microbiology Ecology, 91, 2015, fiv003
doi: 10.1093/femsec/fiv003Advance Access Publication Date: 12 January 2015Research Article
RESEARCH ARTICLE
Census of bacterial microbiota associated with theglacier ice worm Mesenchytraeus solifugusTakumi Murakami1,∗, Takahiro Segawa2,3, Dylan Bodington1, Roman Dial4,Nozomu Takeuchi5, Shiro Kohshima6 and Yuichi Hongoh1
1Department of Biological Sciences, Tokyo Institute of Technology, Tokyo 152-8550, Japan, 2National Instituteof Polar Research, Tokyo 190-8518, Japan, 3Transdisciplinary Research Integration Center, Tokyo 190-8518,Japan, 4Department of Environmental Science, Alaska Pacific University, Alaska 99508, USA, 5Department ofEarth Sciences, Chiba University, Chiba 263-8522, Japan and 6Wildlife Research Center,Kyoto University, Kyoto 606-8203, Japan∗Corresponding author: Department of Biological Sciences, Tokyo Institute of Technology, 2-12-1-W3-48, Ookayama, Meguro-ku, Tokyo 152-8550, Japan.Tel: +81-3-5734-2865; Fax: +81-3-5734-2946; E-mail: [email protected] sentence summary: We analyzed the taxonomic composition of bacteria physically associated with the glacier-adapted worm Mesenchytraeussolifugus, and provided the first report of symbiosis between bacteria and the glacier invertebrates.Editor: Max Haggblom
ABSTRACT
The glacier ice worm, Mesenchytraeus solifugus, is a unique annelid, inhabiting only snow and ice in North American glaciers.Here, we analyzed the taxonomic composition of bacteria associated with M. solifugus based on the 16S rRNA gene. Weanalyzed four fixed-on-site and 10 starved ice worm individuals, along with glacier surface samples. In total, 1341 clones of16S rRNA genes were analyzed for the ice worm samples, from which 65 bacterial phylotypes (99.0% cut-off) were identified.Of these, 35 phylotypes were closely related to sequences obtained from their habitat glacier and/or other components ofcryosphere; whereas three dominant phylotypes were affiliated with animal-associated lineages of the class Mollicutes.Among the three, phylotype Ms-13 shared less than 89% similarity with database sequences and was closest to a gutsymbiont of a terrestrial earthworm. Using fluorescence in situ hybridization, Ms-13 was located on the gut wall surface ofthe ice worms. We propose a novel genus and species, ‘Candidatus Vermiplasma glacialis’, for this bacterium. Our resultsraise the possibility that the ice worm has exploited indigenous glacier bacteria, while several symbiotic bacterial lineageshave maintained their association with the ice worm during the course of adaptive evolution to the permanently coldenvironment.
Keywords: glacier ecology; enchytraeid; symbiosis; 16S rRNA; gut bacteria
INTRODUCTION
On glaciers, which are extreme environments characterized bylow temperature and poor nutrients, a variety of psychrophilicor psychrotolerant organisms form unique ecosystems (Ane-sio and Laybourn-Parry 2012). Many studies on their diversity,
physiology and the impact of their activity on the surroundingenvironment have been reported (Takeuchi, Kohshima and Seko2001; Napolitano and Shain 2004; Anesio et al., 2009; Cameron,Hodson and Osborn 2012; Lutz et al., 2014); however, most ofthese investigations have focused on microbial communities on
Received: 10 October 2014; Accepted: 30 December 2014C© FEMS 2015. All rights reserved. For permissions, please e-mail: [email protected]
1
Dow
nloaded from https://academ
ic.oup.com/fem
sec/article/91/3/fiv003/433893 by guest on 07 February 2022

2 FEMS Microbiology Ecology, 2015, Vol. 91, No. 3
glacier surfaces, while information on glacier invertebrates re-mains limited.
The glacier ice worm,Mesenchytraeus solifugus (family Enchy-traeidae, phylum Annelida), is the most dominant and largestmetazoan in North American maritime glaciers from Alaskato Oregon (Fig. S1, Supporting Information) (Shain et al., 2001;Hartzell et al., 2005). Mesenchytraeus solifugus is the only knownannelid that lives their entire life in glacier ice/snow, i.e. an en-vironment permanently at the freezing point. Physiological andprotein structure analyses have revealed that their metabolism,e.g. ATP synthesis capabilities, is adapted to cold environments(Farrell, Hohenstein and Shain 2004; Napolitano, Nagele andShain 2004; Tartaglia and Shain 2008; Marotta, Parry and Shain2009). A recent phylogeographic study has suggested that theirdistribution reflects past glacial dynamics (Dial et al., 2012). How-ever, little is known about their life cycle, nutrition and ecologi-cal niches in the glacier ecosystem.
Terrestrial earthworms play crucial roles in soil organic mat-ter decomposition and biogeochemical cycles in temperate re-gions (Curry and Schmidt 2007). It has been postulated that thegut microbiota of earthworms contributes significantly to theseactivities (Edwards and Fletcher 1988; Drake and Horn 2007).The majority of gut microorganisms harbored by earthwormsare derived from the soil ingested by the host; anaerobes stimu-lated by the gut environment participate in various bioactivitiessuch as fermentation of recalcitrant materials and denitrifica-tion (Karsten and Drake 1995; Thakuria et al., 2010; Wust, Hornand Drake 2011; Depkat-Jakob et al., 2013). Aquatic and semi-aquatic oligochaetes also have an impact on aquatic ecosystemsvia food webs and the formation of sediment structure (Martinet al., 2008). Given that it is a dominant fauna in the habitat, itis conceivable that the ice worm occupies a similar niche in theglacier ecosystem and has a significant impact on it.
In the present study, we aimed to clarify the microbial com-munity structure physically associatedwith the ice worm to elu-cidate the ice worm’s ecological niche and its nutrient sources.We performed both culture-dependent and -independent cen-suses of the bacterial microbiota based on 16S rRNA genes,whichwe comparedwith those of the glacier surface habitat andthose of two other enchytraeid worms. We also performed fluo-rescence in situ hybridization (FISH) analysis for a dominant gutbacterium and transmission electron microscopy (TEM) of theintestinal tract. Our results provide an insight into the relation-ship between invertebrates and bacteria on a glacier and shouldcontribute to further understanding of the glacier ecosystem.
MATERIALS AND METHODSSample collection
The field research was carried out at Byron Glacier and Hard-ing Icefield in the Kenai Peninsula, Alaska in August 2010, 2011and 2014 (Fig. S1, Supporting Information). Iceworms and glaciersurface ice/snow were collected with sterilized stainless steelscoops. A portion of samples was suspended in RNAlater solu-tion (Ambion, Austin, TX, USA) on site and stored at −30◦C untiluse. The remaining live ice worms were kept in melted glaciersurface ice at 4◦C. Collected samples were kept cold with theglacier ice during transportation.
Two other Enchytraeidae species were used for comparisonwith the ice worm. A snow worm, Mesenchytraeus sp., was col-lected from snow at Ozegahara, Japan in April 2012 and providedby Takaaki Torii (IDEA Consultants Inc.) (Fig. S2, Supporting In-formation). The collected worms were kept in melted snow at
4◦C in a laboratory. This species is active in snow during spring,and in summer, inhabits waterside soil (Torii 2012). A tiny earth-worm, Enchytraeus japonensis, reared in a laboratory, was pro-vided by Arata Tochinai (Hokkaido University).
To wash out ingested food materials, individual worms werestarved in Petri dishes filled with double-distilled water (DDW)at 4◦C for a period (ice worms: 1 week, 2 weeks, 1 month and2 months; snow worms: 3 days). DDWwas sterilized by ultravio-let (254 nm) irradiation for 30 min before use and changed every3 days for the first week, and weekly after that. Enchytraeus japo-nensis were kept in agar plates at room temperature for 3 weekswithout food.
DNA extraction
DNA was extracted from the entire body of individual wormsusing the Ultra Clean R© Soil DNA Isolation Kit (MO BIO Labora-tories, Carlsbad, CA, USA), according to the manufacturer’s pro-tocol. Prior to DNA extraction, the body surface of the wormswas washed twice by submerging them in DDW for 10 min. Theglacier surface samples were melted and 2 mL aliquots werecentrifuged at 4◦C and 17 000 × g for 15 min. The pellets weresuspended in 2 mL of phosphate-buffered saline (PBS) and cen-trifuged under the same conditions. The pellets were used forDNA extraction as above.
PCR, cloning and sequencing
PCR was performed with Ex-Taq polymerase (Takara,Shiga, Japan) and a Bacteria-specific primer pair, 27F-mix (5′-AGRGTTTGATYMTGGCTCAG-3′) and 1492R (5′-GGHTACCTTGTTACGACTT-3′) (Hongoh et al., 2007), to amplifynear-full-length 16S rRNA genes. DNA extracted from the sam-ples was diluted 10 times with TE buffer, and 10 μL of each wasused as a template for PCR in a 100 μL reaction. To minimizePCR drift, the reaction mixture was divided into four tubes andcombined after amplification (Polz and Cavanaugh 1998). Thereaction was performed in a C-1000 Thermal Cycler (Bio-Rad)and the following program was used: 1-min initial denaturationat 95◦C, 25 or 30 cycles of denaturation (15 s at 95◦C), annealing(30 s at 50◦C) extension (90 s at 72◦C) and a final 4-min extensionat 72◦C.
PCR products were purified using the MonoFasTM DNA Purifi-cationKit I (GL Sciences, Tokyo, Japan) according to themanufac-turer’s protocol. Clone libraries were prepared with the TOPO R©
TA cloning R© Kit for Sequencing (Invitrogen, Carlsbad, CA, USA)and Competent Quick DH5α (Toyobo, Osaka, Japan). The cloneswere chosen at random and the inserts were amplified withan M13 primer pair with the following program: 3-min initialdenaturation at 95◦C, 30 cycles of denaturation (15 s at 95◦C),annealing (30 s at 50◦C), extension (2 min at 72◦C) and a final5-min extension at 72◦C. The amplicons were purified with theMultiscreenTM PCR μ96 Filter Plate (Millipore, Billerica, MA, USA)and sequenced with T3 and T7 primers, using the Big Dye R©
Terminator Cycle Sequencing Kit ver. 3.1 (Applied Biosystems,Austin, TX, USA) in an ABI 3130xl or ABI 3730 Genetic Analyzer(Applied Biosystems).
Phylogenetic analysis
All sequences were subjected to identification of chimeras inUSEARCH 6.0 (Edgar et al., 2011) on the Ribosomal DatabaseProject website (http://fungene.cme.msu.edu/FunGenePipeline/chimera check/form.spr). After elimination of putative
Dow
nloaded from https://academ
ic.oup.com/fem
sec/article/91/3/fiv003/433893 by guest on 07 February 2022

Murakami et al. 3
chimeras, low-quality sequences and plastid 16S rRNA genesequences, the remaining ones were aligned with the SILVASSUref nr release111 database (Pruesse et al., 2007) using theSILVA Incremental Aligner in ARB software (Ludwig et al., 2004;Pruesse, Peplies and Glockner 2012). The alignment was man-ually corrected and sequences were assigned to phylotypesdefined by 99.0% sequence similarity criterion in the mothurprogram package ver. 1.33 with the furthest neighbor algorithm(Schloss et al., 2009). Sequences closely related to each phylo-type were retrieved from the GenBank nr database by BLASTsearches. The Chao1 richness, Simpson’s diversity index, Good’scoverage and rarefaction curve were calculated in mothur.
Unambiguously aligned 1049 nucleotide sites, correspond-ing to positions 110–1277 in Escherichia coli (J01695), were usedfor phylogenetic analysis of Mollicutes phylotypes. A maximum-likelihood (ML) tree was constructed in MEGA6 using the gen-eral time reversible (GTR) model and a gamma distribution (G)with 500 bootstrap resamplings (Tamura et al., 2013). A Bayesianinference of phylogeny was performed with the MrBayes soft-ware package ver. 3.2 using a GTR + G nucleotide substitutionmodel (Huelsenbeck and Ronquist 2001). The analysis was runfor 1 000 000 generations with a sample frequency of 100. Theinitial 250 000 generations were discarded as burn-in.
Unweighted UniFrac analysis implemented in the QIIME pro-gram package ver. 1.7.0 was conducted to measure bacterialcommunity structure similarities (Lozupone and Knight 2005;Caporaso et al., 2010). The robustness of sample clustering wasevaluated by jackknife sampling in QIIME.
Transmission electron microscopy
Sample preparation for TEM was conducted according to Shainet al. (2000). Briefly, an ice worm individual was dissected intothree parts with a sterile razor, pre-fixed with 2% paraformalde-hyde and 2.5% glutaraldehyde, and post-fixed with OsO4. Fixedspecimens were rinsed five times in 0.1 M cacodylate bufferand then dehydrated and embedded in EPONTM resin (Quetol651; Nisshin EM, Tokyo, Japan). Thin sections were observed us-ing an H-7500 transmission electronmicroscope (Hitachi, Tokyo,Japan).
FISH and cell enumeration
To detect Mollicutes phylotypes, the probe Tcutes1-825 (5′-CGCTGCTTATGCAACAACTAAT-3′, positions 825–855 in E. coli),which targets the Entomoplasmatales phylotypes Ms-05 and Ms-08, and the probe Tcutes2s-823 (5′-CGCTAGCCGACCTCTAATAC-3′, positions 823–844 in E. coli), which targets the Mycoplasmat-aceae phylotypeMs-13, were designed using the probe-designingfunction implemented in ARB and labeled with Texas Red attheir 5′ extremity. Specificity of the probe was checked us-ing the probeCheck web server (Loy et al., 2008) and it wasfound that each probe had four or more mismatches againstany other sequences in the SILVA database. The optimal tem-perature for hybridization was estimated in silico using the DI-NAMelt web server (Markham and Zuker 2005). A non-labeledhelper probe (5′-AGCGGCGAGACCGACTAT-3′) was also designedandusedwith the probe Tcutes2s-823 (Fuchs et al., 2000). To eval-uate the degree of non-specific binding of oligonucleotide probesto specimens, probe EUB338-III (5′-GCTGCCACCCGTAGGTGT-3′)(Daims et al., 1999) labeled with 6-carboxyfluorescein (6-FAM),mainly targeting verrucomicrobia, which we did not detect inthe clone analysis, was used.We employed the EUB338 probe (5′-GCTGCCTCCCGTAGGAGT-3′) labeled with 6-FAM to detect mostbacterial taxa (Amann et al., 1990).
Ice worm specimens preserved in RNAlater solution weredissected into three parts with a sterile razor and fixed in 4%paraformaldehyde at 4◦C overnight. After three washes in DDW,the specimens were sequentially treated with 10, 15 and 20% su-crose, embedded in Tissue-Tek R© OCT tissue-freezing medium(Sakura Finetechnical, Tokyo, Japan), and cut into 10-μm-thickcross-sections on a Leica CM 1850 cryostat (Leica Biosystems,Nußloch, Germany). The sections were placed on a MAS-coatedglass slide (Matsunami, Osaka, Japan) and treated with 0.25 NHCl at room temperature for 10 min. The sections were thenwashed with DDW and dehydrated in a series of 50, 80, 90 and100% ethanol. Alternatively, ice worm specimens were homoge-nized in 300 μL DDWwith a sterile plastic pestle, fixed and dehy-drated as above. Hybridizationwas conducted according toNodaet al. (2003) at 50◦C for 90 min. Specimens were enclosed usingSlowfade Gold with 4,6-diamidino-2-phenylindole (DAPI) (LifeTechnologies, Carlsbad, CA, USA) and observed under an Olym-pus BX51 epifluorescence microscope (Olympus, Tokyo, Japan).
To count the total number of prokaryotic cells associatedwith an ice worm individual, the entire body was homogenizedwith a sterile plastic pestle in 250 μL PBS after washing theworm surface as above. The homogenate was fixed with 4%paraformaldehyde overnight and centrifuged at 10 000 × g for10 min. The pellet was washed twice in DDW and then immo-bilized onto a polycarbonate hydrophilic membrane filter (Mil-lipore; pore size, 0.2 μm). The cells were stained with DAPI andcounted microscopically.
Bacterial isolation and identification
Individual ice worms were homogenized in 500 μL LB medium(1% Bacto Tryptone, 0.5% Bacto Yeast Extract, 1% NaCl and 0.1mM NaOH in 1 L of DDW) or its 100-fold dilution. The ho-mogenate was spread on either LB or 20-fold diluted LB agarplates and incubated at either 4◦C or −1◦C. Representativecolonies that exhibited unique morphologies and colors werepicked up and suspended in lysis buffer (20 mM Tris-HCl (pH8.0), 200 μg mL−1 proteinase K (Takara), 5 mM EDTA, 400 mMNaCl, and 0.3% SDS) at 55◦C for 1 h. The lysate was treated withAmpdirect R© Plus reagent (Shimadzu, Kyoto, Japan) and directlyused as a template for PCR with the primer set 27F-mix and1492R. The PCR conditions were as follows: an initial denatu-ration for 5 min at 94◦C, 30 cycles of denaturation (30 s at 94◦C),annealing (30 s at 52◦C), extension (150 s at 72◦C) and a final 7-min extension at 72◦C. Sequencing was performed as describedabove. The isolates were identified on the basis of their 16S rRNAgene sequences and stored in both glycerol solution and DMSOsolution at −80◦C.
Nucleotide sequence accession numbers
The nucleotide sequences reported in this study have beendeposited in DDBJ/EMBL/GenBank under the accession num-bers AB989660-AB991625 (clones), AB991626-AB991700 (iso-lates), AB991701-AB992193 (plastids) and LC012708 (18S rRNAgene of the snow worm Mesenchytraeus sp.).
RESULTSTaxonomic composition of ice worm-associatedbacteria
General featuresThe samples analyzed for the census of bacteria in this studyare given in Table 1. Two worm individuals were examined for
Dow
nloaded from https://academ
ic.oup.com/fem
sec/article/91/3/fiv003/433893 by guest on 07 February 2022
4 FEMS Microbiology Ecology, 2015, Vol. 91, No. 3
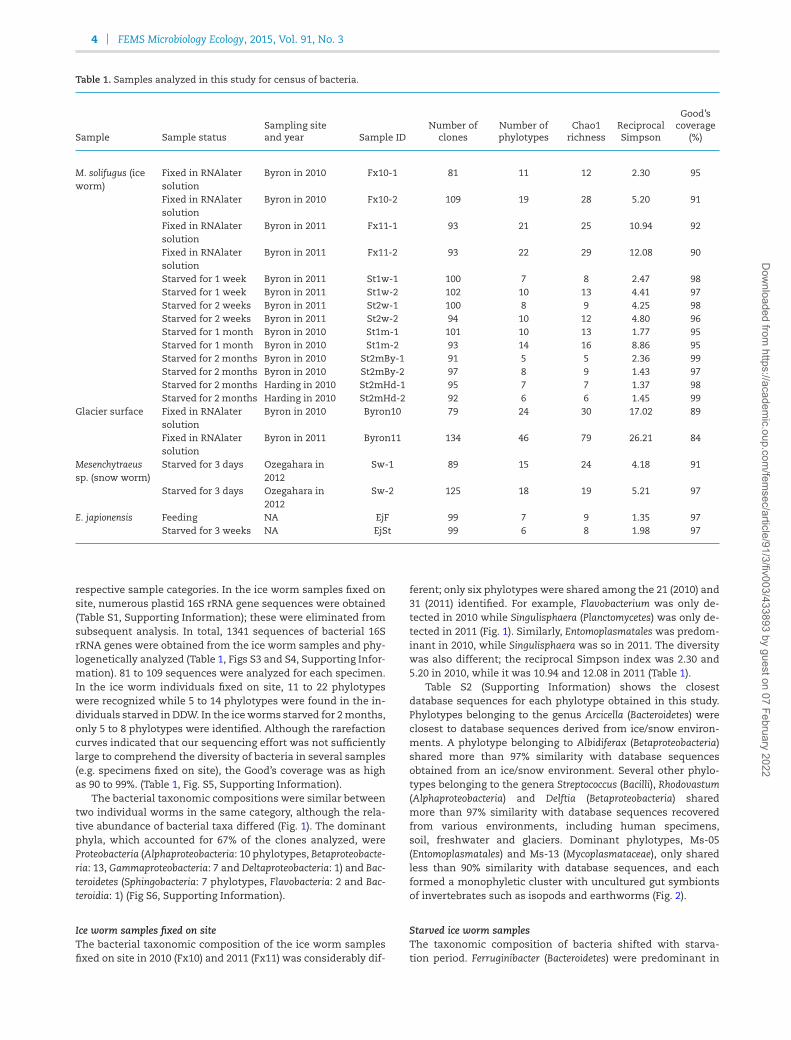
Table 1. Samples analyzed in this study for census of bacteria.
Sample Sample statusSampling siteand year Sample ID
Number ofclones
Number ofphylotypes
Chao1richness
ReciprocalSimpson
Good’scoverage
(%)
M. solifugus (iceworm)
Fixed in RNAlatersolution
Byron in 2010 Fx10-1 81 11 12 2.30 95
Fixed in RNAlatersolution
Byron in 2010 Fx10-2 109 19 28 5.20 91
Fixed in RNAlatersolution
Byron in 2011 Fx11-1 93 21 25 10.94 92
Fixed in RNAlatersolution
Byron in 2011 Fx11-2 93 22 29 12.08 90
Starved for 1 week Byron in 2011 St1w-1 100 7 8 2.47 98Starved for 1 week Byron in 2011 St1w-2 102 10 13 4.41 97Starved for 2 weeks Byron in 2011 St2w-1 100 8 9 4.25 98Starved for 2 weeks Byron in 2011 St2w-2 94 10 12 4.80 96Starved for 1 month Byron in 2010 St1m-1 101 10 13 1.77 95Starved for 1 month Byron in 2010 St1m-2 93 14 16 8.86 95Starved for 2 months Byron in 2010 St2mBy-1 91 5 5 2.36 99Starved for 2 months Byron in 2010 St2mBy-2 97 8 9 1.43 97Starved for 2 months Harding in 2010 St2mHd-1 95 7 7 1.37 98Starved for 2 months Harding in 2010 St2mHd-2 92 6 6 1.45 99
Glacier surface Fixed in RNAlatersolution
Byron in 2010 Byron10 79 24 30 17.02 89
Fixed in RNAlatersolution
Byron in 2011 Byron11 134 46 79 26.21 84
Mesenchytraeussp. (snow worm)
Starved for 3 days Ozegahara in2012
Sw-1 89 15 24 4.18 91
Starved for 3 days Ozegahara in2012
Sw-2 125 18 19 5.21 97
E. japionensis Feeding NA EjF 99 7 9 1.35 97Starved for 3 weeks NA EjSt 99 6 8 1.98 97
respective sample categories. In the ice worm samples fixed onsite, numerous plastid 16S rRNA gene sequences were obtained(Table S1, Supporting Information); these were eliminated fromsubsequent analysis. In total, 1341 sequences of bacterial 16SrRNA genes were obtained from the ice worm samples and phy-logenetically analyzed (Table 1, Figs S3 and S4, Supporting Infor-mation). 81 to 109 sequences were analyzed for each specimen.In the ice worm individuals fixed on site, 11 to 22 phylotypeswere recognized while 5 to 14 phylotypes were found in the in-dividuals starved in DDW. In the iceworms starved for 2months,only 5 to 8 phylotypes were identified. Although the rarefactioncurves indicated that our sequencing effort was not sufficientlylarge to comprehend the diversity of bacteria in several samples(e.g. specimens fixed on site), the Good’s coverage was as highas 90 to 99%. (Table 1, Fig. S5, Supporting Information).
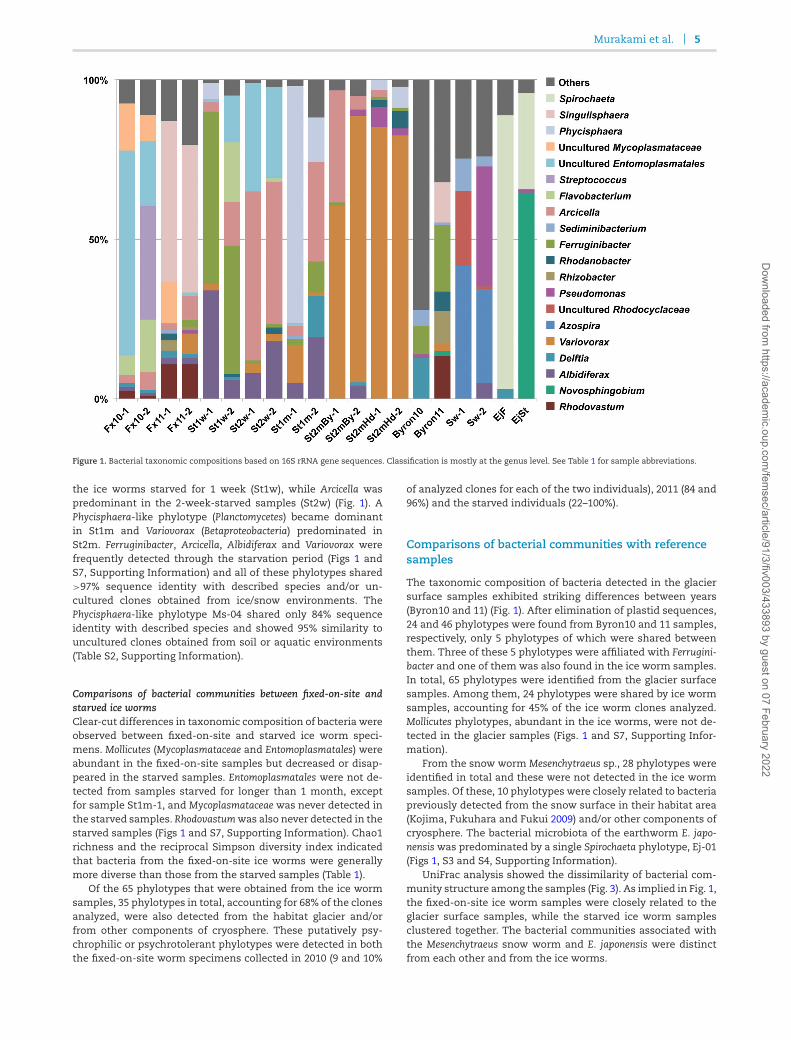
The bacterial taxonomic compositions were similar betweentwo individual worms in the same category, although the rela-tive abundance of bacterial taxa differed (Fig. 1). The dominantphyla, which accounted for 67% of the clones analyzed, wereProteobacteria (Alphaproteobacteria: 10 phylotypes, Betaproteobacte-ria: 13, Gammaproteobacteria: 7 and Deltaproteobacteria: 1) and Bac-teroidetes (Sphingobacteria: 7 phylotypes, Flavobacteria: 2 and Bac-teroidia: 1) (Fig S6, Supporting Information).
Ice worm samples fixed on siteThe bacterial taxonomic composition of the ice worm samplesfixed on site in 2010 (Fx10) and 2011 (Fx11) was considerably dif-
ferent; only six phylotypes were shared among the 21 (2010) and31 (2011) identified. For example, Flavobacterium was only de-tected in 2010 while Singulisphaera (Planctomycetes) was only de-tected in 2011 (Fig. 1). Similarly, Entomoplasmatales was predom-inant in 2010, while Singulisphaera was so in 2011. The diversitywas also different; the reciprocal Simpson index was 2.30 and5.20 in 2010, while it was 10.94 and 12.08 in 2011 (Table 1).
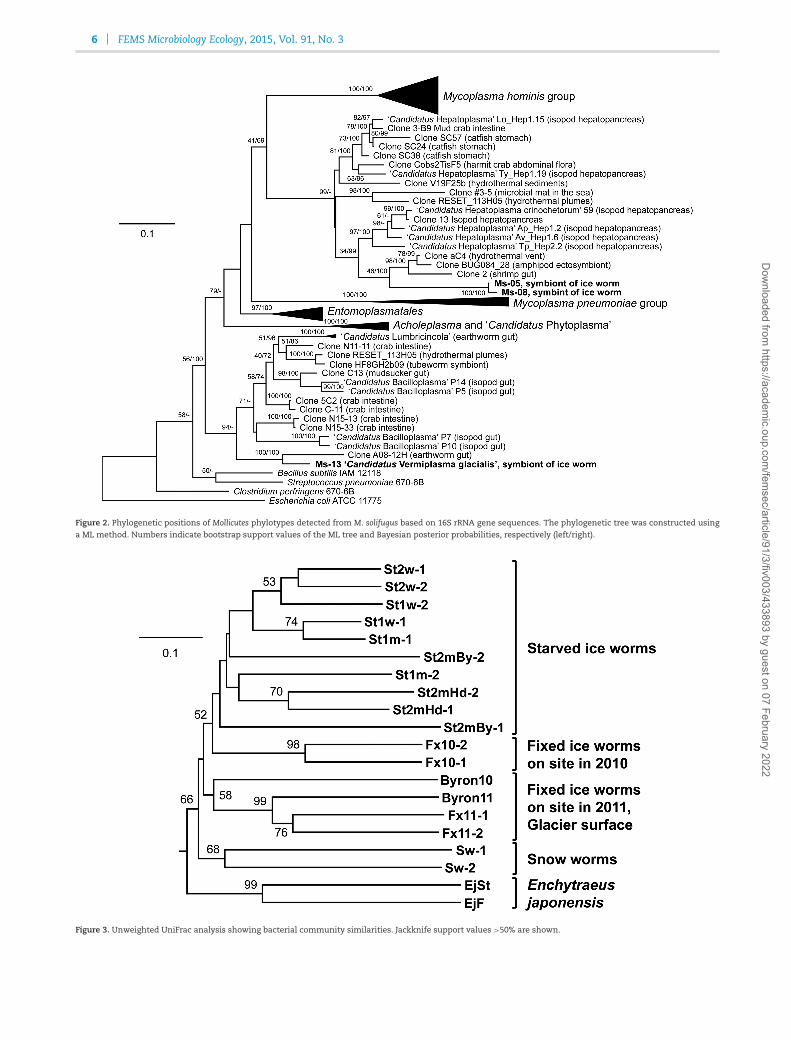
Table S2 (Supporting Information) shows the closestdatabase sequences for each phylotype obtained in this study.Phylotypes belonging to the genus Arcicella (Bacteroidetes) wereclosest to database sequences derived from ice/snow environ-ments. A phylotype belonging to Albidiferax (Betaproteobacteria)shared more than 97% similarity with database sequencesobtained from an ice/snow environment. Several other phylo-types belonging to the genera Streptococcus (Bacilli), Rhodovastum(Alphaproteobacteria) and Delftia (Betaproteobacteria) sharedmore than 97% similarity with database sequences recoveredfrom various environments, including human specimens,soil, freshwater and glaciers. Dominant phylotypes, Ms-05(Entomoplasmatales) and Ms-13 (Mycoplasmataceae), only sharedless than 90% similarity with database sequences, and eachformed a monophyletic cluster with uncultured gut symbiontsof invertebrates such as isopods and earthworms (Fig. 2).
Starved ice worm samplesThe taxonomic composition of bacteria shifted with starva-tion period. Ferruginibacter (Bacteroidetes) were predominant in
Dow
nloaded from https://academ
ic.oup.com/fem
sec/article/91/3/fiv003/433893 by guest on 07 February 2022
Murakami et al. 5
Figure 1. Bacterial taxonomic compositions based on 16S rRNA gene sequences. Classification is mostly at the genus level. See Table 1 for sample abbreviations.
the ice worms starved for 1 week (St1w), while Arcicella waspredominant in the 2-week-starved samples (St2w) (Fig. 1). APhycisphaera-like phylotype (Planctomycetes) became dominantin St1m and Variovorax (Betaproteobacteria) predominated inSt2m. Ferruginibacter, Arcicella, Albidiferax and Variovorax werefrequently detected through the starvation period (Figs 1 andS7, Supporting Information) and all of these phylotypes shared>97% sequence identity with described species and/or un-cultured clones obtained from ice/snow environments. ThePhycisphaera-like phylotype Ms-04 shared only 84% sequenceidentity with described species and showed 95% similarity touncultured clones obtained from soil or aquatic environments(Table S2, Supporting Information).
Comparisons of bacterial communities between fixed-on-site andstarved ice wormsClear-cut differences in taxonomic composition of bacteria wereobserved between fixed-on-site and starved ice worm speci-mens. Mollicutes (Mycoplasmataceae and Entomoplasmatales) wereabundant in the fixed-on-site samples but decreased or disap-peared in the starved samples. Entomoplasmatales were not de-tected from samples starved for longer than 1 month, exceptfor sample St1m-1, and Mycoplasmataceae was never detected inthe starved samples. Rhodovastumwas also never detected in thestarved samples (Figs 1 and S7, Supporting Information). Chao1richness and the reciprocal Simpson diversity index indicatedthat bacteria from the fixed-on-site ice worms were generallymore diverse than those from the starved samples (Table 1).
Of the 65 phylotypes that were obtained from the ice wormsamples, 35 phylotypes in total, accounting for 68% of the clonesanalyzed, were also detected from the habitat glacier and/orfrom other components of cryosphere. These putatively psy-chrophilic or psychrotolerant phylotypes were detected in boththe fixed-on-site worm specimens collected in 2010 (9 and 10%
of analyzed clones for each of the two individuals), 2011 (84 and96%) and the starved individuals (22–100%).
Comparisons of bacterial communities with referencesamples
The taxonomic composition of bacteria detected in the glaciersurface samples exhibited striking differences between years(Byron10 and 11) (Fig. 1). After elimination of plastid sequences,24 and 46 phylotypes were found from Byron10 and 11 samples,respectively, only 5 phylotypes of which were shared betweenthem. Three of these 5 phylotypes were affiliated with Ferrugini-bacter and one of them was also found in the ice worm samples.In total, 65 phylotypes were identified from the glacier surfacesamples. Among them, 24 phylotypes were shared by ice wormsamples, accounting for 45% of the ice worm clones analyzed.Mollicutes phylotypes, abundant in the ice worms, were not de-tected in the glacier samples (Figs. 1 and S7, Supporting Infor-mation).
From the snow worm Mesenchytraeus sp., 28 phylotypes wereidentified in total and these were not detected in the ice wormsamples. Of these, 10 phylotypes were closely related to bacteriapreviously detected from the snow surface in their habitat area(Kojima, Fukuhara and Fukui 2009) and/or other components ofcryosphere. The bacterial microbiota of the earthworm E. japo-nensiswas predominated by a single Spirochaeta phylotype, Ej-01(Figs 1, S3 and S4, Supporting Information).
UniFrac analysis showed the dissimilarity of bacterial com-munity structure among the samples (Fig. 3). As implied in Fig. 1,the fixed-on-site ice worm samples were closely related to theglacier surface samples, while the starved ice worm samplesclustered together. The bacterial communities associated withthe Mesenchytraeus snow worm and E. japonensis were distinctfrom each other and from the ice worms.
Dow
nloaded from https://academ
ic.oup.com/fem
sec/article/91/3/fiv003/433893 by guest on 07 February 2022
6 FEMS Microbiology Ecology, 2015, Vol. 91, No. 3
Figure 2. Phylogenetic positions of Mollicutes phylotypes detected from M. solifugus based on 16S rRNA gene sequences. The phylogenetic tree was constructed usinga ML method. Numbers indicate bootstrap support values of the ML tree and Bayesian posterior probabilities, respectively (left/right).
Figure 3. Unweighted UniFrac analysis showing bacterial community similarities. Jackknife support values >50% are shown.
Dow
nloaded from https://academ
ic.oup.com/fem
sec/article/91/3/fiv003/433893 by guest on 07 February 2022

Murakami et al. 7
Analysis of plastid sequences
We observed numerous cells of red-pigmented algae in thegut of the ice worms (Fig. S8, Supporting Information). Con-cordantly, we obtained abundant plastid sequences. Plastid se-quences were more abundant in samples collected in 2010than those in 2011, for both ice worms and glacier surface ice(Table S1, Supporting Information). When incorporated into theSILVA database, the majority were affiliated with the green al-gal family Trebouxiophyceae and the most dominant phylotypeshowed 93.9% sequence similarity to Pseudochlorella pringsheimii(Fig. S9, Supporting Information).
Microscopic observation of ice worm-associatedbacteria
Dense prokaryotic cells were observed in the lumen of ice wormdigestive tracts by TEM (Fig. 4a–c). Prokaryotic cells also abun-dantly existed within or proximal to the mucus layer that cov-ered the cilia of the gut epithelium (Fig. 4a and b). Severalprokaryotic cells were found in division form (Fig. 4d). No mi-crobial cells were found either on the epidermis or in the otherice worm tissues surveyed.
The ice worms fixed on site at Byron Glacier in 2011 wereused for FISH analysis. The cells of a dominant phylotype, Ms-13 (Mycoplasmataceae), were specifically detected on the surfaceof the gut epithelium in a cross-section (Fig. 5). They were also
Figure 4. Transmission electron micrographs of M. solifugus cross-sections. (a)
Prokaryotic cells found in the digestive tract. (b) Magnified image of regions prox-imal to the gut epithelium. (c) Prokaryotic cells contained in the gut lumen. (d)A prokaryotic cell in division: septum is indicated with a white arrow. ge: gutepithelium; c: cilia; ml: mucus layer; gl: gut lumen. Scale bars = 2 μm in (a) and
(c), and 500 nm in (b) and (d).
detected in a homogenized ice worm sample and cohybridizedwith the EUB338 probe but not with EUB338-III (Fig. S10, Sup-porting Information). We failed to detect the Entomoplasmatalesphylotypes Ms-05 and Ms-08 signals in any specimens.
The ice worm individuals collected on Byron Glacier andHarding Icefield in 2014 were kept alive at 4◦C for 1 month andthen used for enumeration of associated prokaryotic cells. Thenumber of prokaryotic cells per ice worm individual was esti-mated to be 1.7 × 105 ± 3.7 × 104 (Byron, n = 3) and 9.4 × 105 ±2.1 × 105 (Harding, n = 3) (mean ± SD).
Bacterial isolation
A total of 14 phylotypes were isolated by culture on LB platesat either 4◦C or −1◦C (Table S3, Supporting Information). Allof the phylotypes were affiliated with either Betaproteobacteria(Janthinobacterium: 2 phylotypes; Albidiferax, Actimicrobium, Glaci-imonas, Massilia, Polaromonas, Undibacterium and Variovorax: 1)or Gammaproteobacteria (Pseudomonas: 4 phylotypes and Rho-danobacter: 1). Of these, 6 phylotypes were also found in theculture-independent analysis of the ice worm samples, and11 phylotypes exhibited >97% sequence similarity with de-scribed species and/or uncultured clones obtained from othercomponents of cryosphere (Table S4, Supporting Information).
DISCUSSION
This is the first report specifically focusing on the bacterial mi-crobiota associated with invertebrates that inhabit a glacier en-vironment. We revealed that the majority of bacteria that iceworms harbored are closely related to those generally detectedin the components of cryosphere. This raises the possibility thatthe ice worm has exploited glacial bacteria as transient gut sym-bionts. This is analogous to the earthworms, where bacteria in-gested with soil act as transient gut symbionts and contributeto the host’s nutrition (Wust, Horn and Drake 2011).
Discrimination of transient and more tightly associated gutbacteria was attempted in the present study by starving the hostice worms. For example, phylotypes assigned to Rhodovastumwere frequently detected in both fixed-on-site ice worms andglacier surface samples, but not in starvedworm samples. Thesebacteria might be digested or pass through the digestive tractwithout intimate interaction with ice worms. In contrast, Albid-iferax, Variovorax and Arcicella phylotypes were detected in bothfixed-on-site and starved ice worms. The loss of diversity andpredominance of Variovorax in the samples starved for 2 months(St2m) might be attributable to the long starvation period thatwould have led to unhealthy gut microbiota. The difference intaxonomic compositions of bacteria between sampling years orindividuals may reflect the conditions, such as the abundanceof algae (Table S1, Supporting Information), of the collectionsites.
In earthworms, the taxonomic composition of gut bacteriadiffers in relative abundance from that of the surrounding soilbecause of differences in redox potential and nutrient concen-tration (Drake and Horn 2007). Similarly, the relative abundanceof putatively transient ice worm gut bacteria was distinct fromthe surrounding glacier surface ice/snow. While the ice wormsmight receive nutritional benefits from the transient gut bacte-ria, for glacier bacteria, the ice worm gut might be a hotspot forproliferation, given that organic matter is much concentrated inthe gut, compared to the oligotrophic glacier environment. Ac-cording to Segawa et al. (2010), the prokaryotic cell density on thesurface of a glacier in the Alaska Range is 106 cells mL−1. Thus,
Dow
nloaded from https://academ
ic.oup.com/fem
sec/article/91/3/fiv003/433893 by guest on 07 February 2022

8 FEMS Microbiology Ecology, 2015, Vol. 91, No. 3
Figure 5. In situ detection of theMollicutes phylotype Ms-13. (a–d) A cross-section of anM. solifugus specimen. (a) Phase-contrast image. The area with a black rectangleis magnified in (b–d). (b) Phase-contrast image. (c) DAPI-stained image. (d) Merged FISH image: Tcutes2s probe signal (red) and autofluorescence from worm tissues
(yellow). Arrowheads indicate phylotype Ms-13 cell signals. ep: epidermis; gl: gut lumen; ge: gut epithelium. (e–h) A phylotype Ms-13 cell in a homogenizedM. solifugus
tissue. (e) Phase-contrast image. (f) FISH image with the EUB338 probe with 6FAM signal. (g) FISH image with probe Tcutes2s specific to phylotype Ms-13 with TexasRed signal. (h) DAPI-stained image. Scale bars = 100 μm in (a), 10 μm in (b) and 2 μm in (e).
the density of prokaryotic cells associated with ice worms wasfar higher than that on the glacier surface.
In the culture-independent analysis of 16S rRNA genes, di-verse bacteria that were not obtained by the culture-dependentapproach were recovered, as in other environments (Singletonet al., 2003). Among them, phylotypes belonging to the classMol-licutes were dominant in several ice worm individuals. TheseMollicutes phylotypes were related to lineages that have tightand species-specific associations with the gut epithelium ormidgut gland of terrestrial earthworms and isopods (Single-ton et al., 2003; Wang et al., 2004, 2007; Kostanjsek, Strus andAvgustin 2007; Fraune and Zimmer 2008; Nechitaylo, Timmisand Golyshin 2009). In the present study, we found that the Mol-licutes phylotype Ms-13 was also localized on the gut wall sur-face of the ice worms. Thus, it seems likely that these Mollicuteslineages have maintained association with the host worm dur-ing the course of their evolution. Although the Mollicutes phy-lotypes obtained from the ice worms were not detected in theglacier surface samples in our Sanger sequencing analysis, weidentified a few V4 region sequences of 16S rRNA gene identi-
cal to those of Ms-13 and nearly identical to Ms-05 and Ms-08among the sequence reads generated by Choudhari et al. (2013)using an Illumina HiSeq from a Byron Glacier sample. This im-plies that the gut-associatedMollicutes phylotypes are depositedon the glacier surface with ice worm feces and re-infected toother individuals.
Previous observations have revealed that ice worms ingestalgae blooming on the surface of glaciers (Goodman 1971). Con-cordantly, we observed abundant algal cells in the icewormguts.Red-pigmented algae were dominant, and they morphologicallyresembled Chlamydomonas cf. nivalis that were found on the sur-face of glacier snow in Alaska (Takeuchi et al., 2006). We alsoidentified numerous plastid 16S rRNA genes of algae in the iceworm samples fixed on site. It is conceivable that ice worm gutmicrobiota, including those transiently acquired with ingestedfood, play a role in the digestion of recalcitrantmatter such as al-gal cell wall polysaccharides. For example, the phylotype Ms-01,one of themost frequently detected phylotypes, exhibited 98.0%sequence similarity with Variovorax paradoxus strains, whichparticipate in a wide variety of catabolic pathways, including
Dow
nloaded from https://academ
ic.oup.com/fem
sec/article/91/3/fiv003/433893 by guest on 07 February 2022

Murakami et al. 9
cellulolysis (Satola, Wubbeler and Steinbuchel 2013). Becausethe ice worm permanently resides in glacier ice/snow, exploita-tion of psychrophilic or psychrotolerant bacteria as gut sym-bionts might be important.
Our results provide basic information to elucidate the ecol-ogy and evolution of the ice worm and its associated bacteria.However, it remains unclear whether the detected bacterial se-quences represent truly active gut symbionts and what theirfunctional roles are. Future metagenomic and metatranscrip-tomic analyses of the ice worm gut microbiota should addressthese issues. We propose a novel genus and species for the Mol-licutes phylotype Ms-13.
Description of ‘Candidatus Vermiplasma glacialis’
Vermiplasma glacialis (ver. mi. plas′ma. L. merc. n. vermis worm;Gr. neut. n. plasma something formed or molded, N.L. neut.n. vermiplasma something from worm; gla. cia. lis. L. fem. adj.glacialis icy or glacial).
Cells are curved rods, 1.0–1.5 μm in length and specificallylocalized on the gut wall surface of the glacier ice worm M.solifugus. The assignment of ‘Candidatus Vermiplasma glacialis’is based on the 16S rRNA sequence (AB989845) and hybridiza-tion with the 16S rRNA-targeted oligonucleotide probe (5′-CGCTAGCCGACCTCTAATAC-3′).
SUPPLEMENTARY DATA
Supplementary data is available at FEMSEC online.
ACKNOWLEDGEMENTS
We thank Amy Dethlef, Melissa Becker, Yukihiko Ohnuma,Megumi Honda and Gerard Ganey for assistance with ice wormcollection. We also thank Takaaki Torii and Arata Tochinaifor providing reference worm samples, and Kenichi Watanabe,Keiko Ikeda, and Masaki Tamori for assisting with the experi-ments. Sequencing and TEM observation were done in Biomate-rial Analysis Center in Tokyo Institute of Technology. Sequencingwas also done in National Institute of Polar Research.
FUNDING
This study was financially supported by the Grant-in-Aid forScientific Research from the Japan Society for Promotion ofScience to S.K (22241005 and 26241020) and Y.H. (23117003) andthe NEXT program to Y.H.
Conflict of interest. None declared.
REFERENCES
Amann RI, Binder BJ, Olson RJ, et al. Combination of 16S rRNA-targeted oligonucleotide probes with flow cytometry for an-alyzing mixed microbial populations. Appl Environ Microb1990;56:1919–25.
Anesio AM, Hodson AJ, Fritz A, et al. High microbial activity onglaciers: importance to the global carbon cycle. Global ChangeBiol 2009;15:955–60.
Anesio AM, Laybourn-Parry J. Glaciers and ice sheets as a biome.Trends Ecol Evol 2012;27:219–25.
Cameron KA, Hodson AJ, Osborn AM. Structure and diversityof bacterial, eukaryotic and archaeal communities in glacialcryoconite holes from the Arctic and the Antarctic. FEMS Mi-crobiol Ecol 2012;82:254–67.
Caporaso JG, Kuczynski J, Stombaugh J, et al. QIIME allows anal-ysis of high-throughput community sequencing data. NatMethods 2010;7:335–6.
Choudhari S, Smith S, Owens S, et al.Metagenome sequencing ofprokaryotic microbiota collected from Byron Glacier, Alaska.Genome Announc 2013;1:e00099–13.
Curry JP, Schmidt O. The feeding ecology of earthworms—a re-view. Pedobiologia 2007;50:463–77.
Daims H, Bruhl A, Amann R, et al. The domain-specific probeEUB338 is insufficient for the detection of all Bacteria: devel-opment and evaluation of a more comprehensive probe set.Syst Appl Microbiol 1999;22:434–44.
Depkat-Jakob PS, Brown GG, Tsai SM, et al. Emission of nitrousoxide and dinitrogen by diverse earthworm families fromBrazil and resolution of associated denitrifying and nitrate-dissimilating taxa. FEMS Microbiol Ecol 2013;83:375–91.
Dial RC, Dial RJ, Saunders R, et al. Historical biogeography of theNorth American glacier ice worm, Mesenchytraeus solifugus(Annelida: Oligochaeta: Enchytraeidae). Mol Phylogenet Evol2012;63:577–84.
Drake HL, Horn MA. As the worm turns: the earthworm gut as atransient habitat for soilmicrobial biomes.Annu RevMicrobiol2007;61:169–89.
Edgar RC, Haas BJ, Clemente JC, et al. UCHIME improvessensitivity and speed of chimera detection. Bioinformatics2011;27:2194–200.
Edwards CA, Fletcher KE. Interactions between earthworms andmicroorganisms in organic-matter breakdown. Agr EcosystEnviron 1988;24:235–47.
Farrell AH, Hohenstein KA, Shain DH. Molecular adaptationin the ice worm, Mesenchytraeus solifugus: divergence ofenergetic-associated genes. J Mol Evol 2004;59:666–73.
Fraune S, Zimmer M. Host-specificity of environmentally trans-mitted Mycoplasma-like isopod symbionts. Environ Microbiol2008;10:2497–504.
Fuchs BM, Glockner FO, Wulf J, et al. Unlabeled helper oligonu-cleotides increase the in situ accessibility to 16S rRNA of fluo-rescently labeled oligonucleotide probes. Appl Environ Microb2000;66:3603–7.
GoodmanD. Ecological investigations of iceworms on CasementGlacier, Southeastern Alaska. Report No. 39, Institute of PolarStudies, Ohio State University Research Foundation, Colum-bus, 1971.
Hartzell PL, Nghiem JV, Richio KJ, et al. Distribution and phy-logeny of glacier iceworms (Mesenchytraeus solifugus andMes-enchytraeus solifugus rainierensis). Can J Zool 2005;83:1206–13.
Hongoh Y, Sato T, Dolan MF, et al. The motility symbiont of thetermite gut flagellate Caduceia versatilis is a member of the‘Synergistes’ group. Appl Environ Microb 2007;73:6270–6.
Huelsenbeck JP, Ronquist F. MRBAYES: Bayesian inference ofphylogenetic trees. Bioinformatics 2001;17:754–5.
Karsten GR, Drake HL. Comparative assessment of the aero-bic and anaerobic microfloras of earthworm guts and forestsoils. Appl Environ Microb 1995;61:1039–44
Kojima H, Fukuhara H, Fukui M. Community structure ofmicroorganisms associated with reddish-brown iron-richsnow. Syst Appl Microbiol 2009;32:429–37.
Kostanjsek R, Strus J, Avgustin G. ‘Candidatus Bacilloplasma,’ anovel lineage of Mollicutes associated with the hindgut wallof the terrestrial isopod Porcellio scaber (Crustacea: Isopoda).Appl Environ Microb 2007;73:5566–73.
Loy A, Arnold R, Tischler P, et al. probeCheck—a central resourcefor evaluating oligonucleotide probe coverage and specificity.Environ Microbiol 2008;10:2894–8.
Dow
nloaded from https://academ
ic.oup.com/fem
sec/article/91/3/fiv003/433893 by guest on 07 February 2022

10 FEMS Microbiology Ecology, 2015, Vol. 91, No. 3
Lozupone C, Knight R. UniFrac: a new phylogenetic methodfor comparing microbial communities. Appl Environ Microb2005;71:8228–35
Ludwig W, Strunk O, Westram R, et al. ARB: a software environ-ment for sequence data. Nucleic Acids Res 2004;32:1363–71.
Lutz S, Anesio AM, Jorge Villar SE, et al. Variations of algal com-munities cause darkening of a Greenland glacier. FEMS Mi-crobiol Ecol 2014;89:402–14.
Markham NR, Zuker M. DINAMelt web server for nucleic acidmelting prediction. Nucleic Acids Res 2005;33:W577–81.
Marotta R, Parry BR, Shain DH. Divergence of AMP deaminasein the ice wormMesenchytraeus solifugus (Annelida, Clitellata,Enchytraeidae). Int J Evol Biol 2009;2009:715086.
Martin P, Martinez-Ansemil E, Pinder A, et al. Global diversityof oligochaetous clitellates (‘Oligochaeta’; Clitellata) in fresh-water. Hydrobiologia 2008;595:117–27.
Napolitano MJ, Nagele RG, Shain DH. The ice worm, Mesenchy-traeus solifugus, elevates adenylate levels at low physiologicaltemperature. Comp Biochem Phys- A 2004;137:227–35.
Napolitano MJ, Shain DH. Four kingdoms on glacier ice: conver-gent energetic processes boost energy levels as temperaturesfall. Proc R Soc Lon B Bio 2004;271:S273–6.
Nechitaylo TY, Timmis KN, Golyshin PN. ‘Candidatus Lumbricin-cola’, a novel lineage of uncultured Mollicutes from earth-worms of family Lumbricidae. Environ Microbiol 2009;11:1016–26.
Noda S, Ohkuma M, Yamada A, et al. Phylogenetic position andin situ identification of ectosymbiotic spirochetes on protistsin the termite gut. Appl Environ Microb 2003;69:625–33
Polz MF, Cavanaugh CM. Bias in template-to-product ratios inmultitemplate PCR. Appl Environ Microb 1998;64:3724–30.
Pruesse E, Peplies J, Glockner FO. SINA: accurate high-throughput multiple sequence alignment of ribosomal RNAgenes. Bioinformatics 2012;28:1823–9.
Pruesse E, Quast C, Knittel K, et al. SILVA: a comprehensiveonline resource for quality checked and aligned ribosomalRNA sequence data compatible with ARB. Nucleic Acids Res2007;35:7188–96.
Satola B, Wubbeler JH, Steinbuchel A. Metabolic characteris-tics of the species Variovorax paradoxus. Appl Microbiol Biot2013;97:541–60.
Schloss PD, Westcott SL, Ryabin T, et al. Introducingmothur: open-source, platform-independent, community-
supported software for describing and comparing mi-crobial communities. Appl Environ Microb 2009;75:7537–41.
Segawa T, Takeuchi N, Ushida K, et al. Altitudinal changes in abacterial community on Gulkana Glacier in Alaska. MicrobesEnviron 2010;25:171–82.
Shain DH, Carter MR, Murray KP, et al. Morphologic character-ization of the ice worm Mesenchytraeus solifugus. J Morphol2000;246:192–7.
Shain DH, Mason TA, Farrell AH, et al. Distribution and behav-ior of ice worms (Mesenchytraeus solifugus) in south-centralAlaska. Can J Zool 2001;79:1813–21.
Singleton DR, Hendrix PF, ColemanDC, et al. Identification of un-cultured bacteria tightly associated with the intestine of theearthworm Lumbricus rubellus (Lumbricidae; Oligochaeta).Soil Biol Biochem 2003;35:1547–55.
Takeuchi N, Dial R, Kohshima S, et al. Spatial distributionand abundance of red snow algae on the Harding Ice-field, Alaska derived from a satellite image. Geophys Res Lett2006;33:L21502.
Takeuchi N, Kohshima S, Seko K. Structure, formation, and dark-ening process of albedo-reducing material (cryoconite) ona Himalayan glacier: a granular algal mat growing on theglacier. Arct Antarct Alp Res 2001;33:115–22
Tamura K, Stecher G, Peterson D, et al. MEGA6: molecularevolutionary genetics analysis version 6.0. Mol Biol Evol2013;30:2725–9.
Tartaglia LJ, Shain DH. Cold-adapted tubulins in the glacier iceworm, Mesenchytraeus solifugus. Gene 2008;423:135–41.
Thakuria D, Schmidt O, Finan D, et al. Gut wall bacteria of earth-worms: a natural selection process. ISME J 2010;4:357–66.
Torii T. Enchytraeidae (Annelida,Clitellata) from Ozegaharamire, central Japan. Low Temp Sci 2012;70:93–8.
Wang Y, Brune A, Zimmer M. Bacterial symbionts in the hep-atopancreas of isopods: diversity and environmental trans-mission. FEMS Microbiol Ecol 2007;61:141–52.
Wang Y, Stingl U, Anton-Erxleben F, et al.‘Candidatus Hepato-plasma crinochetorum,’ a new, stalk-forming lineage of Mol-licutes colonizing the midgut glands of a terrestrial isopod.Appl Environ Microb 2004;70:6166–72.
Wust PK, Horn MA, Drake HL. Clostridiaceae and Enterobacteri-aceae as active fermenters in earthworm gut content. ISMEJ 2011;5:92–106.
Dow
nloaded from https://academ
ic.oup.com/fem
sec/article/91/3/fiv003/433893 by guest on 07 February 2022




















