
Prevalence of trypanosomes, salivary gland hypertrophy virus ...

Cancer Risk Related to Mammary Gland Structureand DevelopmentJOSE RUSSO,1* YUN FU HU, ISMAEL D.C.G. SILVA,2 AND IRMA H. RUSSO1
1Breast Cancer Research Laboratory, Fox Chase Cancer Center, Philadelphia, Pennsylvania 191112Fellow, Fundacao de Amparo a Pesquisa do Estado de Sao Paulo (FAPESP) Brasil
ABSTRACT The breast undergoes dramatic changes in size, shape, and function in associationwith growth, reproduction, and post-menopausal regression. Those changes impact women’s life-time breast cancer risk. An early first full-term pregnancy exerts a protective effect, emphasizingthe need for understanding the role of reproductive influences on breast development and on cancerinitiation and progression, and providing a paradigm for developing preventive strategies based onphysiological principles. Even though the cause of breast cancer and the ultimate mechanismsthrough which an early pregnancy protects from cancer development remain largely unknown, alikely explanation for this protection has been provided by experimental in vivo and in vitro models.These studies have led to the conclusions that cancer initiation requires the interaction of acarcinogen with an undifferentiated and highly proliferating mammary epithelium, whereas dif-ferentiation of the mammary gland inhibits carcinogenic initiation. The process of mammary glanddifferentiation is the result of complex interactions of ovarian, pituitary, and placental hormones,which in turn induce inhibition of cell proliferation, downregulation of estrogen and progesteronereceptors, activation of specific genes, such as inhibin, mammary derived growth factor inhibitorand a serpin-like gene, and expression of extracellular matrix proteins in the normal breast. Cellimmortalization and transformation are associated with the expression of ferritin H and S100Pprotein, which serve as markers of cancer initiation. Comparative studies of normal and neoplasticbreast development have unraveled similarities with experimental models that validate the ex-trapolation of findings for testing hypotheses on the initiation and progression of breast cancer.Microsc. Res. Tech. 52:204–223, 2001. © 2001 Wiley-Liss, Inc.
INTRODUCTIONThe breast is a bilateral organ that in the female
undergoes dramatic changes in size, shape, and func-tion in association with infantile growth, puberty, preg-nancy, lactation, and post-menopausal regression(MacMahon et al., 1970; Russo and Russo, 1987a,1996). The breast is also the source of the most fre-quently diagnosed malignancy in the female popula-tion (Chu et al., 1999; Greenlee et al., 2000). Theknowledge that the risk of developing breast cancer isheavily influenced by endocrinological and reproduc-tive influences (Russo and Russo, 1987a,b, 1996) indi-cates that a thorough understanding of how the endo-crinological milieu, especially that created by preg-nancy, influences the development of this organ isrequired. It has long been known that the risk of breastcancer shows an inverse relationship with early parity(De Waard and Trichopoulos, 1988; Henderson et al.,1993; MacMahon et al., 1970; McGregor et al., 1977;Rosner et al., 1994; Russo and Russo, 1987b, 1996).Case control studies have demonstrated that breastcancer risk increases with the age at which a womanbears her first child. This protection seems to be in-versely related to the length of the interval betweenmenarche and the first pregnancy, because the breastcancer risk increases when this interval is lengthenedover 14 years (Rosner et al., 1994). Therefore, to beprotective, pregnancy has to occur before age 30; in-deed, women first becoming pregnant after that ageappear to have a risk above that of nulliparous women
(Rosner et al., 1994). Multiparity confers additionalprotection (Lambe et al., 1996). The protective effect,however, remains largely limited to the first birth, andpersists at all subsequent ages, even in women olderthan 75 years of age (MacMahon et al., 1970; Rosner etal., 1994).
The cause of breast cancer and the ultimate mecha-nisms through which an early first full-term pregnancyprotects this organ from cancer development remainlargely unknown. Nevertheless, a likely explanationfor the protective effect of pregnancy has been providedby studies performed in an experimental animal modelbased on the induction of mammary carcinomas byadministration of chemical carcinogens to rats. Thesestudies have led to the conclusion that cancer initiationrequires the interaction of the carcinogen with an un-differentiated and highly proliferating mammary epi-thelium. Differentiation of the mammary gland, suchas that induced by full-term pregnancy, inhibits carci-
Yun Fu Hu’s present address is 54 Loveton Circle, Sparks, MD 21152.Ismael D.C.G. Silva’s present address is UNIFESP- Escola Paulista de Medi-
cina, Disciplina de Ginecologia, Rua Napoleao de Barros, 715, 70 Andar, SaoPaulo, Brasil.
Contract grant sponsor: National Cancer Institute; Contract grant numbers:CA64896, CA67238; Contract grant sponsor: NIEHS; Contract grant number:ESO7280.
*Correspondence to: Jose Russo, M.D., F.C.A.P., Director, Breast Cancer Re-search Laboratory, Fox Chase Cancer Center, 7701 Burholme Avenue, Philadel-phia, PA 19111. E-mail: J_Russo @ fccc.edu
Received 20 July 2000; accepted in revised form 31 July 2000
MICROSCOPY RESEARCH AND TECHNIQUE 52:204–223 (2001)
© 2001 WILEY-LISS, INC.

nogenic initiation (Russo and Russo, 1987a,b, 1996;Russo et al., 1988, 1989, 1991, 1992, 1994). The processof mammary gland differentiation is the result of com-plex interactions of ovarian, pituitary, and placentalhormones (Ciocca et al., 1982), which in turn induce inthe mammary epithelium activation of specific genessuch as inhibin (Russo and Russo, 1994a, 1995), mam-mary derived growth factor inhibitor (Hu et al, 1997b)and a Serpin-like gene (Hu et al., 1998). The role ofthese genes in the protection that an early full-termpregnancy confers to women is a matter that needsfurther investigation.
There is no explanation for the higher risk to developmalignancies exhibited by nulliparous and late parouswomen. The fact that experimentally induced rat mam-mary carcinomas develop only when the carcinogeninteracts with the undifferentiated and highly prolifer-ating mammary epithelium of young nulliparous rats(Russo and Russo 1987a,b, 1996; Russo et al., 1982,1988, 1989, 1991, 1992, 1994), suggests that the breastof late parous and of nulliparous women might exhibitsome of the undifferentiated and/or cell proliferativecharacteristics that predispose the tissue to undergoneoplastic transformation in experimental models.Comparative studies of breast development in womenand rodents indicate that there are similarities be-tween them that validate the extrapolation of findingsfrom the experimental model to the human situation(Russo and Russo, 1996; Russo et al., 1991; Toy andRusso, 1981, 1985). Based on the fact that the under-standing of the initiation and progression of the carci-nogenic process requires a thorough knowledge of thenormal development of the target organ, this manu-script briefly reviews the stages of mammary glanddevelopment, the changes occurring during pregnancyand at post-lactation, and at menopause. This basicinformation is complemented with the analysis of cellproliferation and hormone receptors in relation tobreast structure, expression of extracellular matrixproteins and angiogenesis in the normal breast, forfinalizing with the correlation between breast struc-ture and cancer development and the expression offerritin H (Higgy et al., 1997) and S100P protein (Hu etal., 1998) as marker of cancer initiation.
PRENATAL AND PERINATALDEVELOPMENT
The mammary gland parenchyma arises from a sin-gle epithelial ectodermal bud. Most authors agree onthe successive stages of development of the mammarygland during the embryonic and fetal stages. However,there are variations in nomenclature and in the exacttime of appearance of each structure because there arevariations in the age of the embryo, depending uponwhether it is measured from the estimated time ofconception, the last missed menstrual period, or thelength of the embryo. Because of difficulties in estab-lishing precisely the day of conception, we have corre-lated herewith the phases of mammary gland develop-ment with embryonal or fetal length. The intrauterinedevelopment of the mammary gland has been dividedinto 10 different stages, beginning at the ridge stage inthe 5-mm embryo and terminating in the end vesiclestage in the newborn (Table 1).
The breast of the newborn is composed of 6–10straight ductal structures opening into the nipple.These ducts end in short, convoluted, and slightly di-lated ductules that form primitive lobules (Figs. 1a andc). The ductal system is composed of hollow tubulesending in short ductules lined by one layer of epithelialand one of myoepithelial cells (Figs. 1b and d). Theepithelial cells have eosinophilic cytoplasm that isfinely vacuolated due to the presence of lipid droplets.They exhibit typical apocrine secretion that is not con-fined to the primitive alveolar structures, but involvesthe whole ductal system, which appears dilated, secre-tion filled, and lined by a secretory-type epithelium.These observations suggest that secretory activity is ageneralized response of all the mammary epithelium tothe maternal hormonal levels. The secretory activity ofthe newborn gland subsides within 3 to 4 weeks post-partum (Russo and Russo, 1987a).
POSTNATAL DEVELOPMENTThe literature on the development of the breast dur-
ing childhood is scanty. It is generally accepted, how-ever, that it does little more than to keep pace with thegeneral growth of the body until the approach of pu-berty. Breast development can be measured by differ-ent parameters, i.e., external appearance of the breast(Geithner et al., 1998; Hergenroeder et al., 1999), orinternal structure, namely determination of mammarygland area, volume, degree of branching, or degree oflobular development, whose appearance indicates thelevel of differentiation of the gland.
The adolescent period begins with the first signs ofsexual change at puberty and terminates with sexualmaturity. Puberty in the female sets in between theages of 10 and 12 years. With the approach of puberty,the rudimentary mammae begin to show growth activ-ity both in the glandular tissue and in the surroundingstroma. Glandular increase is due to the growth anddivision of small bundles of primary and secondaryducts (Fig. 1e–h). They grow and divide partly dichot-omously (from the Greek word dichotomos, or repeatedbifurcation) and partly sympodially (from Greek syn 1podion base, involving the formation of an apparentmain axis from successive secondary axes), on a dichot-omous basis. The ducts grow, divide, and form club-shaped terminal end buds. Terminal end buds giveorigin to new branches, or alveolar buds, which arestructures morphologically more developed than theterminal end bud, but yet more primitive than theductules or acini that originate from them, and char-acterize the more developed or fully functional lobules.
TABLE 1. Stages of prenatal development of the human breast1
1. Ridge stage (less than 5-mm embryo)2. Milk hill stage (more than 5.5-mm embryo)3. Mammary disc stage (;10- to 11-mm embryo)4. Globule type stage (11.0- to 25.0-mm embryo)5. Cone stage (25- to 30-mm embryo)6. Budding stage (30- to 68-mm embryo)7. Indentation stage (68 mm to 10 cm)8. Branching stage (10-cm fetus)9. Canalization stage (20 and 32 weeks of gestation)
10. End-vesicle stage (newborn)1Reprinted with permission from reference Russo and Russo (1998).
205CANCER RISK AND MAMMARY GLAND STRUCTURE
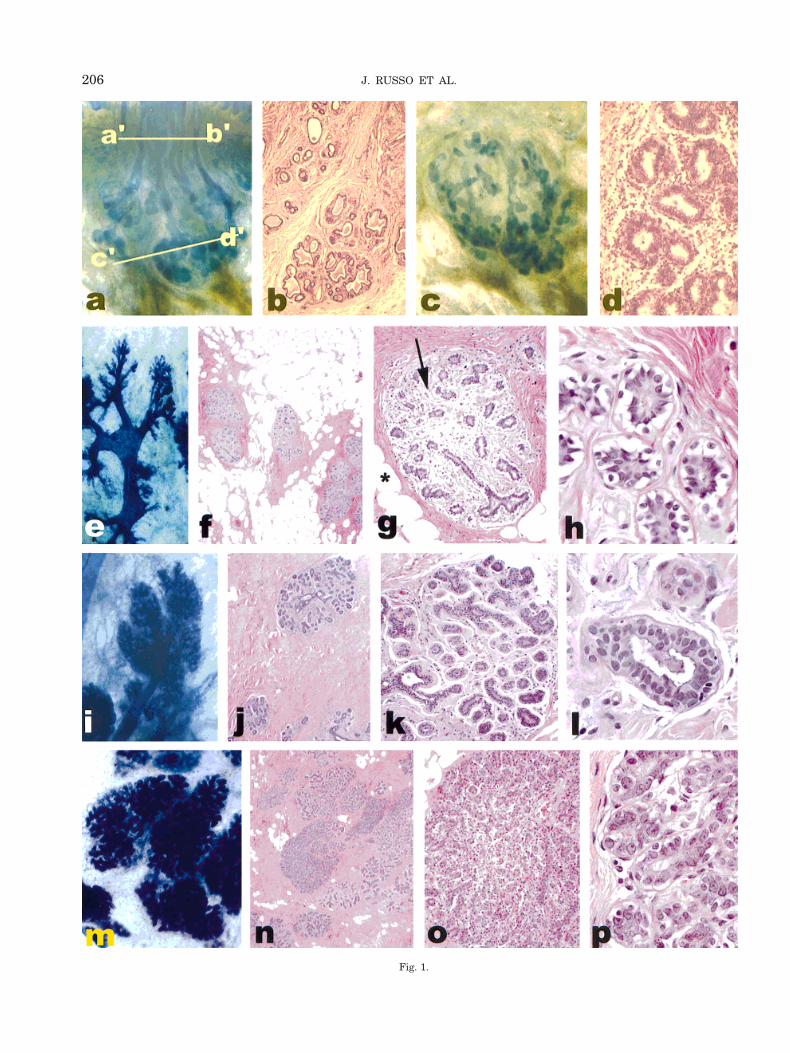
Fig. 1.
206 J. RUSSO ET AL.

Ductules that originate from alveolar buds cluster ingroups of 10 to 11 around a terminal duct, forming thelobule type 1 (Lob 1) or virginal lobule (Figs. 1f–h).Lobule formation in the female breast occurs within1 to 2 years after onset of the first menstrual period.The differentiation of the mammary gland is a gradualprocess that takes many years and varies greatly fromwoman to woman. In some cases, if pregnancy does notsupervene, full differentiation is never attained (Russoand Russo, 1987a,b; Russo et al., 1992).
It is acknowledged that hormonal influences play asignificant role in breast development; however, theeffect of their fluctuations during the menstrual cycleon parenchymal proliferation has not been definitivelyelucidated. The normal breast epithelium undergoescyclic variations of DNA synthesis, as determined innormal breast samples cultured in the presence of 3H-thymidine. Although cell proliferation and cell deathseem balanced in maintaining the equilibrium of theresting breast, the stimulus induced by ovarian hor-mones during a menstrual cycle results in a positivebalance. Accordingly, each ovulatory cycle fostersmammary development with budding of new struc-tures that never fully return to the starting point of thepreceding cycle. This process continues until about age35, when it reaches a plateau that remains constantuntil the initiation of menopause, when the regressiveprocess begins (Russo and Russo, 1987a).
The normal breast of adult women contains threeidentifiable types of lobules: The previously describedLob 1, and two additional ones, the lobule type 2 (Lob 2)(Fig. 1i–l), and lobule type 3 (Lob 3) (Fig. 1m–p). Thetransition from Lob 1 to Lob 2, and of this to Lob 3 is agradual process of sprouting of new ductules, whichincrease in number from approximately 11 found inLob 1 to 47 and 81 in Lob 2 and Lob 3, respectively(Table 2). Although the increased number of ductulesresults in a concomitant increase in lobular size, eachindividual ductule becomes smaller, as indicated by thereduced transversal diameter and reduced number of
cells per cross section (Table 2). The breast paren-chyma of nulliparous women is composed of undiffer-entiated structures such as terminal ducts and Lob 1,which constitute the predominant structures through-out the lifespan. Lob 2 are present in moderate num-bers during the early years, but sharply decrease afterage 23. This observation suggests that a certain per-centage of Lob 1 might have progressed to Lob 2. Thenumber of Lob 3 is significantly lower, since they areonly occasionally seen. In parous women, a history of afull-term pregnancy between the ages of 14 to 20 yearscorrelates with a significant increase in the number ofLob 3, which remains as the predominant structureduring the early reproductive years. Their number de-creases after the fourth decade of life, and more mark-edly after menopause, when they involute to Lob 2 andLob 1 (Russo and Russo, 1987a).
PREGNANCYDuring pregnancy, the breast attains its maximum
development. It occurs in two distinctly dominantphases: an early phase of growth and a late phase oflobular differentiation. The growth of the early phase isdriven by proliferation of the distal elements of theductal tree, resulting in the formation of new branches,and of new ductules, or acini, that will form Lob 3 (Fig.1m–p). The intensity of budding and degree of lobuleformation goes beyond what has been observed in thevirginal breast. By the third month of pregnancy, thenumber of well-formed lobules exceeds the number ofprimitive budding stages; however, primitive buddingstages are still found. In newly formed lobules, theepithelial cells composing each acinus not only increasegreatly in number due to active cell division but theyalso increase in size mainly because of cytoplasmicenlargement (Russo and Russo, 1987a). During thesecond trimester of pregnancy, the lobules are furtherenlarged and increased in number. They surround theduct from which their central branch proceeds sothickly that the chief duct, the terminal or intralobularterminal duct, can no longer be recognized. The tran-sition between the terminal ducts and the buddingacini is gradual, making the histological distinctionbetween the two of them difficult, since both showevidence of early secretory activity. The definitivestructure of the ductal tree is essentially settled by theend of the first half of pregnancy. The mammarychanges that characterize the second half of pregnancyare chiefly continuation and accentuation of the secre-tory activity. Further progressive branching continues
Fig. 1. a: Whole mount preparation of the breast of a 2-week oldhuman female. a’-b’ indicates the region of the major lactiferous ducts,c’-d’ the portion of the mammary parenchyma distal to the nipple thatcontains primitive ducto-lobular structures (Toluidine blue, 32.5).b: Histological section at the level a’-b’ from a. The ductal structureshave dilated lumina filled with proteinaceous fluid (Hematoxylin andeosin [H & E], 34). c: Whole mount preparation of section c’-d’ thebreast shown in a (Toluidine blue, 32.5). d: Histological section of theducto-lobular structures shown in a and c. The ductules are lined bya bilayered epithelium. Proliferation takes place chiefly in the basalcells. The luminal cells exhibit secretory properties (H & E, 316).e: Breast tissue of an 18-year-old nulliparous woman. The wholemount preparation contains ducts and lobules type 1 (Lob 1) (Tolu-idine blue, 32.5). f–h: Histological sections of Lob 1 shown in e. f,lobules surrounded by perilobular connective tissue are separated byinterlobular fat (32.5); g, Lob 1 composed of ductules separated byintralobular loose connective tissue (arrow) (310); h, individualductules of the Lob 1 shown in 1g exhibit subnuclear vacuolization(340) (H & E). i: Lobules type 2 (Lob 2) found in the breast of a24-year-old nulliparous woman. Whole mount preparation (Toluidineblue, 32.5). j–l: Histological sections of the Lob 2 shown in i. j, theinterlobular fat has been replaced by dense connective tissue (32.5);k, (310); l (340) (H & E). m: Lobules type 3 (Lob 3) found in the breastof a 35-year-old parous woman. Whole mount preparation (Toluidineblue, 32.5). n–p: Histological sections of the Lob 3 shown in m,stained with H & E. Photographed at 32.5, 310, and 340, respectively.
TABLE 2. Characteristics of the lobular structuresof the human breast1
StructureLobular area
(mm2)2No. of ductules/
lobule3No. of cells/
cross section4
Lob 1 48 6 44 11.2 6 6.3 32.4 6 14.1Lob 2 60 6 26 47.0 6 11.7 13.1 6 4.8Lob 3 129 6 49 81.0 6 16.6 11.0 6 2.01Reprinted from Russo et al. (1992) with permission of the publisher.2Student’s t-tests were done for all possible comparisons. Lobular areas showedsignificant differences between lob 1 vs. lob 3 and lob 2 vs. lob 3 (P , 0.005).3The number of ductules per lobule was different (P , 0.01) in all the compar-isons.4The number of cells per cross section was significantly different in ductules oflob 1 vs. 2 and 3 (P , 0.01).
207CANCER RISK AND MAMMARY GLAND STRUCTURE

Fig. 2.
208 J. RUSSO ET AL.

with less prominent bud formation. At this time, theformation of true secreting units or acini, the differen-tiated structures, becomes more and more evident. Cellproliferation and formation of new acini are reduced toa minimum, and the lumina of those already formedbecome distended by accumulation of secretory mate-rial or colostrum (Figs. 2a–c). The secretory acinusformed during pregnancy is a terminal outgrowth thatmarks the end of glandular differentiation. However,just before and during parturition, there is a new waveof mitotic activity with an increase in the total DNA ofthe gland. During lactation, the process of growth anddifferentiation may be observed in the same lobuletype, side by side with the process of milk secretion(Russo and Russo, 1987a, 1994a,b, 1996; Russo et al.,1988).
POSTLACTATIONAL INVOLUTIONFrom mid-pregnancy on, a yellowish fluid containing
a high concentration of protein is secreted by the mam-mary alveolar epithelium into the lumen and may beexpelled from the nipple. Lactation starts after post-partum withdrawal of placental lactogen and sex ste-roids, which appear to prevent the action of prolactinon the mammary epithelium. Colostrum is secretedduring the first week postpartum, followed by a 2 to3 week period of transitional milk secretion, leading tothe secretion of mature milk (Hu et al., 1997b; Russoand Russo, 1987a).
No major morphological changes of the mammarygland are observed during lactation. The mammarylobules are enlarged and the acini have dilated luminafilled with granular, slightly basophilic material ad-mixed with fat. There is a significant variation in lob-ule size throughout the gland, suggestive of a variationin lactogenic activity from lobule to lobule. Milk issynthesized and released into the mammary acini andductal system, although it can be stored for up to
48 hours before the rate of milk synthesis and secretionbegins to decrease.
As long as milk is removed regularly from the mam-mary ductal tree, the alveolar cells continue to secretealmost indefinitely (Hu et al., 1997b; Russo and Russo,1987a). The accumulation of milk in the ductoacinarlumina and within the cytoplasm of the lactogenic ep-ithelial cells that occurs after weaning has an inhibi-tory effect on further milk synthesis. This effect isfollowed by a series of involutional changes in themammary gland consisting of a multifocal asynchro-nous process of reduction in volume of the secretoryepithelial cells and further inhibition of their secretoryactivity. It is considered that post-lactational regres-sion is due to two complementary mechanisms, cellautolysis, with collapse of acinar structures and nar-rowing of the tubules, and appearance of round cellinfiltration and phagocytes in and about the disinte-grating lobules, and finally, regeneration of the peri-ductal and perilobular connective tissue with renewedbudding and proliferation in the terminal tubules. Thepostpartum breast retains more glandular tissue thanif pregnancy or pregnancy and lactation had neveroccurred until pre-menopause, when involution begins(Hu et al., 1997b; Russo and Russo, 1987a).
MENOPAUSAL BREASTMenopause supervenes as the consequence of the
maturation or atresia of more than 99% of the 400,000follicles that are present in the ovaries of a female fetusof a gestational age of 5 months. Gonadotropin-releas-ing hormone secretion is also implicated in this phe-nomenon, indicating that a hypothalamic process isinvolved in the development of menopause. The mostcharacteristic sign of menopause is amenorrhea, whichis the result of the almost complete cessation of ovarianestrogen and progesterone production. The years lead-ing up to the final menstrual period, until menopausesets in generally at around age 51 years, constitute theperimenopause. During this time, many women ovu-late irregularly, either because the rise in estrogenduring the follicular phase is insufficient for triggeringa luteinizing hormone (LH) surge, or because the re-maining follicles are resistant to the ovulatory stimu-lus. The increase in human longevity occurring in oursociety has caused a considerable increment in thenumber of women that will live one third or more oftheir lives in the menopausal period, namely withoutnatural estrogen and progesterone. After menopause,the breast undergoes a regressive phenomenon both innulliparous and parous women. This regression ismanifested as an increase in the number of Lob 1, anda concomitant decline in the number of Lob 2 and Lob3. At the end of the fifth decade of life, the breast ofboth nulliparous and parous women is predominantlycomposed of Lob 1 (Russo and Russo, 1987a; Russo etal., 1992). These observations led us to conclude thatthe understanding of breast development requires ahorizontal study in which all the different phases ofgrowth are taken into consideration. For example, theanalysis of breast structures at a single given point,i.e., age 50 years, would lead us to conclude that thebreast of both nulliparous and parous women appearsidentical. However, the endocrinological and physiolog-ical changes triggered by pregnancy in prior years
Fig. 2. a–c: Histological sections of lobues type 4 (Lob 4) found in thebreast at the beginning of lactation. a, secretory lobules surroundedby scant perilobular connective tissue; a moderate amount of inter-lobular fat is still present (32.5); b, Lob 4 composed of tightly packedsecretory acini (310); c, individual acini exhibit a vacuolated cyto-plasm containing lipid droplets and proteinaceous material (340) (H& E). d: Lob 1 stained with factor VIII antibody for detecting endo-thelial cells. (3,39’-Diaminobenzidine [DAB] with H & E. 32.5.e: Higher magnification of the Lob 1 depicted in d. Blood vessels inbrown among the ductules counterstained with H & E, 340). f: Lob4 stained with factor VIII antibody. The number of vascular elementsis markedley reduced as the acini become distended by secretion.DAB-H & E, 34). g: Lob 2 stained with anti-tenascin antibody.DAB-H & E, photographed in dark field, 32.5). h: Lob 1 stained withanti-elastin antibody (DAB-H & E, 32.5). i: Lob 1 ductule containingKi67 positive cells (brown nuclei) (DAB), and estrogen receptor posi-tive cells (red purple nuclei) (Alkaline phosphatase [AP]) (H & Ecounterstain, 340). j: Lob 1 ductule containing Ki67 positive cells(brown nuclei) (DAB), and progesterone receptor positive cells (redpurple nuclei) (AP) (H & E counterstain, 340). k: Lob 3 of the humanbreast stained with an antibody against inhibin B (DAB), counter-stained with H&E (316). l: Lob 4 of the human breast stained with anantibody against inhibin B and counterstained with H&E (316). m: Insitu hybridization of a Lob 1 with a probe for a serpin-like moleculeshowed no signal over the background. (Dark field, 32.5). n: In situhybridization of a Lob 3 with a probe for a serpin-like moleculedetected a strong signal (Bright field with H&E counterstain, 32.5).o: In situ hybridization of the Lob 3 shown in n with a probe for aserpin-like molecule (Dark field, 32.5).
209CANCER RISK AND MAMMARY GLAND STRUCTURE

might have imprinted permanent biological and/orgenomic changes in the breast that might affect thepotential of this organ for undergoing neoplastic trans-formation, even though they might not be manifestedat a morphological level. Thus, from a qualitative pointof view, the regressive phenomenon occurring in thebreast at menopause differs in nulliparous and parouswomen. This postulate is supported by the facts that inthe total lobular population of the breast of nulliparouswomen, Lob 1 represent 65 to 80%, Lob 2 10 to 35%,and Lob 3 only 0 to 5%, and that the relative percent-age of these components is independent of age. In thebreast of parous women, on the other hand, Lob3 comprise 70 to 90% of the total lobular populationduring the years encompassed from post-lactationalinvolution to the 4th decade of life, when they start todecline as the number of Lob 1 and 2 progressivelyincrease (Fig. 3, Table 3). After menopause, the per-centages of the three types of lobules are similar innulliparous and parous women (Fig. 3). These lifetimeobservations led us to conclude that early parouswomen truly underwent lobule differentiation, whichwas evident at a younger age, whereas nulliparouswomen seldom reached the Lob 3, and never the Lob4 stages (Russo et al., 1992).
STEROID HORMONE RECEPTORS, CELLPROLIFERATION, AND ANGIOGENESIS IN
RELATION TO BREAST STRUCTUREThe importance of classifying the lobular structures
of the breast into four types based on their morpholog-ical characteristics, which are, in turn, an indication oftheir stage of development, has been further empha-sized by the analysis of their proliferative activity andcontent of steroid hormone receptors (Russo et al.,1998a, 1999). The proliferative activity of the breast is
determined utilizing a variety of parameters, i.e., countof the number of cells in mitosis (mitotic index), rate ofincorporation of 3H-thymidine as a marker of DNAsynthesis (DNA labeling index [DNA-LI]) (Calaf et al.,1982a,b, Russo et al., 1988), and more recently immu-nocytochemical methods that identify cells traversingthe cell cycle, and react positively with the proliferat-ing cell nuclear antigen (PCNA) or the nuclear anti-body Ki67 (Russo et al., 1998a, 1999). When adjustingfor specific differences in the parameters measured,there is good correlation among the various methodol-ogies mentioned above. For that reason we have uti-lized immunocytochemical detection of cells positive forKi67 for measuring the proliferative activity of normalbreast tissues obtained from reduction mammoplastiesperformed in 32 women free of mammary pathology.Seven of the women were nulliparous and 25 parous.The group of parous was slightly older than that ofnulliparous women (Table 3). Lob 1 had the highestproliferative activity in both nulliparous and parouswomen, although the proliferative index was twofoldhigher in the Lob 1 of nulliparous than in those ofparous women (P , 0.02), and higher than in Lob 2 ofthe same donors, and Lob 2 and Lob 3 of parous women(Table 3). These differences were not abrogated whenthe phases of the menstrual cycle were taken into con-sideration (Russo and Russo, 1997). These observationsled us to conclude that parity, in addition to exerting animportant influence in the lobular composition of thebreast, profoundly influences the proliferative activityof the mammary epithelium. Although after meno-pause the proliferative activity of the mammary epi-thelium decreases, making the differences in cell pro-liferative activity of the breast between nulliparousand parous women less pronounced, still parouswomen retain a lower rate of cell proliferation thannulliparous women.
Estrogens and progesterone are known to promoteproliferation and differentiation in the normal breastepithelium. Both steroids act intracellularly through areceptor that, when activated by its binding with thehormone, regulates the expression of specific genes(King, 1992; Kumar et al., 1987; Li et al., 1995). Thefact that estradiol acts locally in the mammary gland,stimulating DNA synthesis and promoting duct forma-tion, and that this effect is mediated by an estrogenreceptor (ER) has been confirmed by the developmentof an ERa knockout mouse in which branching of themammary gland is abrogated (Fuqua et al., 2000).However, the mechanism by which steroid hormonesexert their mitogenic and differentiation effects has notbeen clearly established (Aakvaag et al., 1991;Dell’aquilla et al., 1984; Dickson et al., 1986; Han andLiehr, 1995; Huff et al., 1988; Huseby et al., 1987;Katzenellenbogen et al., 1987; Markaverich et al.,1988; Page et al., 1983; Soto and Sonnenschein, 1987).One of the accepted mechanisms of action of steroidhormones postulates that cells proliferate in responseto a direct stimulus that results from the binding ofestradiol to its specific receptor (ER) and its interactionwith the DNA (King, 1992; Russo et al., 1998a). Mea-surements of the levels of ER and progesterone recep-tor (PgR) in the cytosol fraction of the normal breastusing standard biochemical techniques do not reflectthe true receptor content of the breast epithelium be-
Fig. 3. Percentage of lobular structures in the breast of premeno-pausal (Pre-Men.) and postmenopausal (Post-Men.) parous women,and combined pre and postmenopausal (Pre & Post-Men) nulliparouswomen. The content of lobules type 1 (Lob.1), lobules type 2 (Lob.2),and lobules type 3 (Lob.3) in the breast is expressed as a percentage.
210 J. RUSSO ET AL.

cause of the low cellularity of the tissue. The use ofmonoclonal antibodies that specifically recognize ERand PgR makes it possible to identify and to quantitateindividual cells expressing these nuclear receptors(Russo and Russo, 1997, Russo et al., 1998a, 1999). Theutilization of this technique has allowed us to deter-mine that the percentage of cells expressing these re-ceptors varies as a function of the degree of lobulardevelopment of the breast, and, therefore, of the type oflobular structure analyzed. Lob 1 consistently containa higher percentage of cells that express simulta-neously ER and PgR than Lob 2, and their numberprogressively decreases in Lob 3 and Lob 4. Theseobservations indicate that a progressive decrease in thepercentage of cells expressing steroid hormone recep-tors occurs as the structures become more differenti-ated. This trend parallels the progressive decrease incell proliferation. However, the cells that express thereceptors are not the same that express the Ki67 im-munocytochemical reaction (Fuqua et al., 2000; Russoet al., 1998a, 1999). These observations indicate thatestrogens might stimulate the synthesis of growth fac-tor(s), which in turn would be able to induce cell pro-liferation in Ki67 positive cells. Nevertheless, thesedata allowed us to conclude that degree of differentia-tion of the breast is an important determinant of theexpression of both ER and PgR, in addition to modu-lating the proliferative activity of the breast epithe-lium. Neither age nor parity history affects the per-centage of cells reacting for both receptors.
Using factor VIII as a marker of endothelial cells foridentifying blood vessels, the number of identifiablevascular spaces were counted in the intralobularstroma (Fig. 2d–f). The number of blood vessels (BV) inrelation to the number of ductules present in eachspecific lobule type was expressed as the AngiogenicIndex (AI) (Table 4).The AI was 1.64 in Lob 1, 0.84 inLob 2, and 0.55 in Lob 3. AI was significantly higher(P , 0.00000) in Lob 1 than in Lob 2 and Lob 3 (Table4). The differences in AI among the three differentlobule types analyzed was similar in nulliparous andparous women, and aging did not affect the differencesin AI among the different lobule types analyzed (Tables4 and 5).Our data indicated that there is a direct rela-tionship between the proliferative activity in the lobu-lar structures and the angiogenic index. Lob 1, whichare the less differentiated, are those with the highestproliferative and angiogenic indices.
EXPRESSION OF EXTRACELLULAR MATRIXPROTEINS IN THE NORMAL BREAST
The breast extracellular matrix cannot be consideredas a simple amorphous gel anymore. The study of its
molecular organization has revealed that it consists ofmultifunctional proteins that play important roles incell growth, adhesion, migration, and differentiation.Indeed, we know today that besides its mechanicalproperties, the extracellular matrix undergoes a vari-ety of biochemical changes caused by minimal varia-tions in the metabolism of the mammary parenchyma(Bonte, 1995; Shrevestha et al., 1996). In order to de-termine whether specific extracellular matrix proteinsare produced or repressed during the differentiation ofthe human mammary gland, we identified by immuno-histochemistry elastin and tenascin, two proteins thatplay important roles in the cell-cell and cell-substra-tum interactions, as well as in malignant transforma-tion (Chiquet-Ehrisman et al., 1986, Senior et al.,1984). The study of the structure and tissue distribu-tion of tenascin, as well as in vitro models, indicate thatit is a multifunctional glycoprotein participating in celladhesion, motility, and migration pathways, sheddingof epithelial cells from surfaces, promoting cell growth,demarcating tissue boundaries, angiogenesis, tissue re-modeling, and immune modulation (Yamada et al.,1976). Tenascin is largely involved in areas of epithe-lial-mesenchymal interactions during the process ofembryo development (Chiquet-Ehrisman et al., 1986),suggesting an important role during the differentiationprocess. The interactions of elastin with microfibrilarproteins and the demonstration that peptide segmentsof elastin elicit a chemotactic response by fibroblastsand monocytes (Senior et al., 1984) suggest that elastinhas domains with biologically important activitiesother than those related to its mechanical properties(Wrenn et al., 1986).
Immunocytochemical detection of elastin using ananti-elastin monoclonal antibody (Sigma ChemicalCompany, St. Louis, MO), allowed us to identify elastinaround ductular structures (Fig. 2g). The intensity ofthe reaction was similar in all the lobular structures.However, the number of lobules reacting against elas-tin varied. Lob 1 had the lowest percentage of reactiv-ity (10.35%) compared with 19.82% in Lob 2, 36.64% inLob 3, and 33.80% in Lob 4 (Table 6).Parity slightlyincreased the percentage of elastin positive Lob 1 andLob 3. However, the differences were not statisticallysignificant. Aging did not affect the percentage of lob-ules presenting positive elastin reaction (Table 7). Itwould be of importance to compare the presence ofelastin in a larger population of older parous and nul-liparous women in order to determine whether parityhad induced permanent changes, but unfortunately notenough cases in these two groups were available.
Tenascin was identified using a mouse IgG1mono-clonal antibody raised against purified human tenascin
TABLE 3. Influence of age and parity on the lobular composition and proliferative activity of the breast
No. ofcases Parity Donors’ age
Lobular structures and proliferative activity1
Lob 1 Lob 2 Lob 3 Lob 4Ducts
(positivecells)Lobules
Positivecells Lobules
Positivecells Lobules
Positivecells Lobules
Positivecells
7 Nulliparous 40.85 6 10.93 81.13 6 6.90 4.97 6 2.512 18.87 6 6.89 0.62 6 0.30 0 0.0 0 0 2.00 6 0.7525 Parous 48.76 6 11.00 66.84 6 7.55 1.56 6 0.292 23.10 6 6.37 0.85 6 0.31 8.03 6 4.67 0.22 6 0.18 66 0 1.47 6 0.191Lobules, percentage of lobules type 1 (Lob 1), type 2 (Lob 2), type 3 (Lob 3), and type 4 (Lob 4); positive cells, percentage of cells positive for Ki67.2Lob 1 Nulliparous vs. Lob 1 Parous, t 5 2.44 P , 0.02.
211CANCER RISK AND MAMMARY GLAND STRUCTURE

(Chemicon International Inc., CA). This antibody re-acted positively in the intralobular stroma of Lob 1,Lob 2, and Lob 3 of the human breast (Fig. 2h). Thepercentage of lobules reacting with tenascin rangedfrom 30% in Lob 3 to 89.25% in Lob 4 (Table 8). Thedifference in percentage of reactivity between Lob4 and the other three lobular types was highly signifi-cant. Parity and aging did not affect the percentage ofstructures reacting for tenascin. These findings indi-cate that the expression of elastin increases proportion-ally with the differentiation of the breast, since it isexpressed more highly in Lob 3, which are the moredifferentiated, than in the less differentiated Lob 1 andLob 2. Tenascin does not exhibit a linear relationshipwith differentiation, and overall age and parity do notaffect the expression of elastin and tenascin.
NOVEL DIFFERENTIATION-ASSOCIATEDSERPIN IS UP-REGULATED DURING
LOBULAR DEVELOPMENTIn order to determine whether the morphological
differences observed between Lob 1 and Lob 3 wereassociated with specific molecular changes, we com-pared by differential display technique the expressionof genes between two types of lobule that represent theextreme ends of the spectrum of differentiation in theresting breast. This analysis allowed us to identify twocDNA fragments, L342C and L346C, that were prefer-entially expressed in the differentiated Lob 3 of thehuman breast (Fig. 4). The differential expression ofthese genes was then confirmed by reverse Northernblot analysis, which showed that plasmid DNA con-taining either L342C or L346C cDNA was hybridized tothe labeled cDNA synthesized from Lob 3 RNA, but notto that from Lob 1 RNA (Fig. 5).
Sequence analysis indicated that L342C and L346Cconsisted of 665 and 416 nucleotides, respectively (Fig.6). Remarkably, the sequence of L346C was identical tothat of L342C except for a stretch of 249 nucleotides inL342C that was missing in L346C (Fig. 6). Search forhomology in the gene bank databases revealed thatL342C and L346C each were nearly identical to two
stretches of sequences in human a1-antichymotrypsin(a1-ACT), a serine protease inhibitor (serpin), acces-sion nos. K01500 and J05176, that have been previ-ously published (Chandra et al., 1983; Rubin et al.,1990) (Fig. 6). When the three sequences were aligned,it became apparent that L342C and L346C contained327-nt and 576-nt deletions, missing nucleotides 1041–1367 and 783–1367 of the a1-ACT, respectively (Fig. 4).These results suggested that these two differentiallyexpressed genes represented potentially alternativelyspliced transcripts of a novel serpin.
To differentiate the novel serpin from human a1-ACT and to detect isoforms of the novel serpin, anantisense riboprobe was synthesized using the RSA-linearized L342C plasmid DNA. The antisense ribo-probe was approximately 464 bp in full length andcontained 68 bp of vector sequences and covered theentire region that was missing in L346C (see Fig. 6).The riboprobe was predicted to protect two fragmentsin human a1-ACT transcripts, a 228-bp fragment pro-tected by its 39-end and a 168-bp fragment protected byits 59-end. These two fragments were expected to beequally well protected. In addition, if the novel serpinwas present as two isoforms in human breast cells, theriboprobe was predicted also to protect two fragments,a full-length 396-bp fragment found in L342C and a168-bp fragment found in L346C. Results from theRNase protection assay showed that the antisense ri-boprobe protected three fragments in human breastcells, namely, the full-length 396-bp fragment found inL342C and the 168-bp fragment expected for its iso-form as well as the 228-bp fragments apparently froma1-ACT (Fig. 7). Similarly, the same antisense ribo-probe protected three differently sized transcripts inliver, but not in full-term human placenta (Fig. 7).Yeast tRNA used as control did not protect any frag-ments from RNase digestion (Fig. 7) and a sense ribo-probe did not protect any fragments of RNA from thesame panel of cells (data not shown). These resultsindicated that the serpin we cloned from the differen-tiated human breast epithelial cells was novel anddifferent from human a1-ACT, and there were twoisoforms of the novel form present in human breastepithelial cells as well as in liver tissue.
The amount of various transcripts protected by theantisense riboprobe varied with the status of immor-talization and neoplastic transformation in the panel ofcells we studied. Protection of the full-length 396-bpfragments indicative of expression of the long tran-scripts of the novel serpin was high in primary culturesand S130 mortal cells, very low in the immortalized
TABLE 5. Effect of age on angiogenic index in parous women
No. of cases Donors’ age
Angiogenic index
Lob 1 Lob 2 Lob 3
4 30–39 1.54 6 0.30 0.75 6 0.03 0.59 6 0.1110 40–50 1.42 6 0.41 0.76 6 0.34 0.44 6 0.144 .50 1.88 6 0.58 0.97 6 0.02 0.47 6 0.02
TABLE 4. Angiogenic index in the normal breast
No. of cases Parity Donors’ age
Angiogenic index
Lob 1 Lob 2 Lob 3
48 Parous 1 Nulliparous 43.36 6 7.59 1.64 6 0.401,2 0.84 6 0.281,3 0.55 6 0.112,3
13 Nulliparous 40.16 6 8.32 1.66 6 0.354,5 0.82 6 0.194,6 0.57 6 0.095,6
35 Parous 44.36 6 7.29 1.63 6 0.427,8 0.84 6 0.317,9 0.55 6 0.138,9
1Lob 1 vs. Lob 2 t 5 9.18 P , 0.00000.2Lob 1 vs. Lob 3 t 5 10.49 P , 0.00000.3Lob 2 vs. Lob 3 t 5 3.76 P , 0.0004.4Lob 1 vs. Lob 2 t 5 6.00 P , 0.00001.5Lob 1 vs. Lob 3 t 5 6.59, P , 0.00001.
6Lob 2 vs. Lob 3 t 5 2.54 P , 0.02.7Lob 1 vs. Lob 2 t 5 7.16 P , 0.00000.8Lob 1 vs. Lob 3 t 5 8.21, P , 0.00000.9Lob 2 vs. Lob 3 t 5 2.98, P , 0.005.
212 J. RUSSO ET AL.

MCF-10F cells and their transformed derivatives (BP1and BP1E), and completely absent in T47D, BT20, andMCF-7, the three breast carcinoma cell lines examined(Fig. 7). In fact, the highest amount of the full-length396-bp fragments was observed in those cells derivedfrom tissues containing more differentiated structures(e.g. S#244, S#250). In contrast, expression of the hu-man a1-ACT, determined as the 228-bp fragments, wasapparent in the primary cells, the mortal S130, theimmortal MCF-10F cells, and chemically transformedderivatives (Fig. 7). Similarly, the 168-bp fragments,which could result from protection of either humana1-ACT transcripts or the shorter transcripts of thenovel serpin, were apparent in mortal and immortalhuman breast epithelial cells and the chemically trans-formed cells (Fig. 7). As controls of equal sampling,gene expression of b-actin appeared to be similar in allthe samples tested, as determined by the same proce-dure of RNase protection analysis (Fig. 7).
Results from Northern blot analysis confirmed theresults of RNase protection analysis, that the expres-sion of the novel serpin was high in two primary cul-tures of human breast epithelial cells (HBEC), moder-ate in the mortal normal breast epithelial cell lineS130, low in MCF-10F and HBL-100 cells (Chang et al.,1982; Caron et al., 1991), two immortalized normalbreast epithelial cell lines (Band et al., 1990, 1993), andin BP1and BP1E, two benzo[a]pyrene (BP)-trans-formed breast epithelial cell lines. It was absent in D3and D3-1, cell lines transformed with 7,12-dimethyl-benz[a]anthracene (DMBA), in T ras transfected BP1-Tras, and D3-1-Tras cells, and in MCF-7, T47D, andBT20 tumorigenic breast carcinoma cell lines (Fig. 8).
The predominant transcripts of the novel serpin wereapproximately 1.5–1.6 kb in size. The results re-mained the same whether the cells were cultured inlow calcium concentration (0.04 mM) (L) or standardcalcium concentration (1.05 mM) identified as high(H) (Fig. 8).
The physiological function of the novel serpin inhuman breast is completely unknown. It is clearly abiomarker of lobular differentiation because of itspreferential expression in the more differentiatedLob 3 as compared to Lob 1 of the human breast, asdetermined by differential display and reverseNorthern analyses. In fact, it is of interest to notethat expression of the novel serpin, as determined byRNase protection analyses, was higher in primarycultures established from breast tissues containingmore differentiated structures than those containingfewer of them. Since the degree of mammary lobulardifferentiation is a critical determinant of mammarysusceptibility to carcinogenesis (Russo et al., 1988),the preferential expression of the novel serpin inmore differentiated epithelium may confer protectionagainst breast cancer development, as has been pro-posed for other differentiation-associated genes (Huet al., 1998). Thus, it is reasonable to speculate thatthe silencing of serpin in Lob 1, known to be the siteof origin of ductal carcinomas (Russo et al., 1991),may render the mammary cells susceptible to eventsleading to carcinogenesis (Hu et al., 1998). This no-tion is supported by our evidence that the full-lengthtranscription of the novel serpin is indeed down reg-ulated during the process of immortalization andneoplastic transformation of HBEC in vitro. We have
TABLE 6. Influence of parity on elastin lobular reaction
No. cases Parity
Elastin reaction
Lob 1 Lob 2 Lob 3 Lob 4
44 Nulliparous 1 Parous 10.35 6 1.551,2,3 19.82 6 2.371 36.64 6 4.632,4 33.80 6 20.753,4
11 Nulliparous 7.37 6 1.745,6 19.36 6 5.925,7 28.71 6 7.746,7
33 Parous 11.50 6 2.038,9,10 19.92 6 2.648,11,12 40.60 6 5.618,11,13 84.52 6 4.5210,12,13
1Lob 1 vs. Lob 2, t 5 3.47, P , 0.0009.2Lob 1 vs. Lob 3, t 5 6.58, P , 0.00000001.3Lob 1 vs. Lob 4, t 5 2.89, P , 0.005.4Lob 3 vs. Lob 4, t 5 3.47, P , 0.001.5Lob 1 vs. Lob 2, t 5 2.58, P , 0.02.6Lob 1 vs. Lob 3, t 5 3.30, P , 0.004.7Lob 2 vs. Lob 3 N/S.
8Lob 1 vs. Lob 2, t 5 2.87, P , 0.006.9Lob 1 vs. Lob 3, t 5 7.91, P , 0.000000.10Lob 1 vs. Lob 4, t 5 9.75, P , 0.0000000.11Lob 2 vs. Lob 3, t 5 5.66, P , 0.000004.12Lob 2 vs. Lob 4, t 5 8.68, P , 0.000000.13Lob 3 vs. Lob 4, t 5 3.74, P , 0.002.
TABLE 7. Influence age on elastin lobular reaction
No. cases Donors’ age
Elastin reaction
Lob 1 Lob 2 Lob 3 Lob 4
17 20–40 14.53 6 3.57 20.02 6 3.73 40.91 6 5.76 84.52 6 4.5218 41–50 8.21 6 1.83 20.47 6 3.46 34.61 6 7.269 51–80 7.70 6 2.65 12.25 6 5.90 54.15 6 20.85
TABLE 8. Tenascin expression in lobular structures
No. of cases
Positive tenascin expression (%)
Lob 1 Lob 2 Lob 3 Lob 4
43 44.43 6 16.281,2,3 41.53 6 18.221 30.67 6 22.872,4 89.25 6 0.913,4
1Lob 1 vs. Lob 2, not significant.2Lob 1 vs. Lob 3, t 5 2.35, P , 0.02.
3Lob 1 vs. Lob 4, t 5 3.84, P , 0.00003.4Lob 3 vs. Lob 4, t 5 3.50, P , 0.0004.
213CANCER RISK AND MAMMARY GLAND STRUCTURE

evaluated the expression of the differently sizedtranscripts of the novel serpin in relation to lobulardifferentiation and found that Lob 1 has no expres-sion (Fig. 2m) whereas in Lob 3 serpin is abundantlyexpressed (Fig. 2m and 2o).
MAMMARY-DERIVED GROWTHINHIBITOR (MDGI)
Mammary-derived growth inhibitor (MDGI), whichsuppresses carcinogenesis in transfected human breastcancer cells (Huynh et al., 1995) and inhibits thegrowth of Ehrlich ascitis carcinoma cells, was first de-tected in ascitic fluid (Lehmann et al., 1977, 1979).MDGI belongs to a multigene family of fatty acid-bind-ing proteins (FABP) known to bind long-chain fattyacids, retinoids and eicosanoids (Veerkamp et al.,1991). The amino-acid sequence of MDGI and/or thegene encoding it have been sequenced in cattle (Bo-hmer et al., 1987), rat (Jones et al., 1988), mouse(Treuner et al., 1994; Tweedie and Edwards, 1989), andhuman heart (Borchers et al., 1990; Huynh et al.,1995). A 70 kDA antigen has been identified in mam-mary epithelial cells (Muller et al., 1989). The human
heart FABP3, that covers an 8-kb region of genomicDNA, is considered to be identical to MDGI and homo-logue to the bovine MDGI (Phelan et al., 1996). Thisgene has been localized to chromosome 1p32-35(Huynh et al., 1995), a locus that has been associatedwith frequent loss of heterozygosity in sporadic breastcancer (Bieche et al., 1993, 1994; Genuardi et al., 1989),suggesting that MDGI serves as a tumor suppressorgene.
MDGI, that has been shown to induce rodent mam-mary gland differentiation (Binas et al., 1992) and tosuppress human breast cancer cell growth in vitro, hasbeen cloned from bovine (Kurtz et al., 1991) and rodentmammary gland. We have cloned MDGI from culturedHBEC as a 640-bp fragment that contained a singleopen reading frame encoding a protein of 133 aminoacids identical to MDGI/FABP3. In situ hybridizationanalysis of paraffin-embedded normal human breasttissues revealed that HBEC-MDGI was highly ex-pressed in the most differentiated Lob 4, poorly ex-pressed in moderately differentiated Lob 3, and absentin the least differentiated Lob 1 and Lob 2 (Figs. 9a–c).In addition, HBEC-MDGI was not expressed in breasttissues that contained ductal hyperplasia, carcinoma insitu, or invasive carcinomas. Our results suggested
Fig. 4. Differential display between mRNA from differentiatedLob3 obtained from three different breast samples(250R, 250L, and126R) and undifferentiated Lob l (226R, 227R, and 244); cDNA bandsthat were preferentially expressed in the Lob3 are identified as L342Cand L346C (arrows).
Fig. 5. Reverse Northern analysis of the differentially expressedgenes L342C and L346C. Plasmid DNAs containing cDNA insertsthat were differentially displayed in the differential display gels wereblotted onto 2 Nynon1 membranes and hybridized to labeled Lob l andLob 3 cDNA probes that were reverse transcribed from total RNA of226R and 126R shown in Figure 4.
214 J. RUSSO ET AL.

that HBEC-MDGI is a biomarker of lobular differenti-ation in the human breast, and its expression is si-lenced in poorly differentiated lobules as well as in theearly and late stages of breast cancer progression (Huet al. 1997b).
In light of a strong correlation between MDGI ex-pression and lobular differentiation, and the fact thatthe degree of mammary differentiation is a criticaldeterminant of mammary susceptibility to carcinogen-esis (Russo et al., 1988), it is tempting to speculate thatthe protective effects of pregnancy against breast can-cer may result from expression of MDGI, which mightin turn be responsible for pregnancy-associated mam-mary differentiation, however, more studies in thisdirection are necessary for clarifying its role in breastcarcinogenesis.
SITE OF ORIGIN OF BREAST CANCERIt is not known when in the lifetime of a woman
breast cancer initiates. It is not known either what
specific agent or combination of agents and/or factorsare the cause of it (Malins et al., 1993, 1996). The factsthat late menarche and a full-term pregnancy com-pleted before age 24, or early full-term pregnancy, re-duce the risk of breast cancer development, whereasearly menarche, nulliparity, or late parity are associ-ated with a higher breast cancer incidence (McGregoret al., 1977; Russo et al., 1991) indicate that the periodencompassed between menarche and the first full-termpregnancy represents a window of high susceptibilityfor the initiation of breast cancer. This postulate issupported by clinical data indicating that exposure toionizing radiations at ages younger than 19 increasesthe incidence of breast cancer, whereas exposure atolder ages or after pregnancy does not increase cancerrisk (Land et al., 1980, McGregor et al., 1977). Eventhough the time of cancer initiation is not known, thesite of origin of ductal carcinoma, the most frequentbreast malignancy, has been identified to be the termi-nal ductal-lobular unit (TDLU) (Wellings et al., 1975;
Fig. 6. Comparison of nucleotide se-quences of L342C, L346C, and human al-antichymotrypsin (al-ACT; accession no.K01500 and J05176; 33, 34). Sequences withdifferent nucleotides or frameshift deletionsare boldfaced; the sequences of forward andreverse PCR primers used for differentialanalysis are underlined.
215CANCER RISK AND MAMMARY GLAND STRUCTURE

Wellings, 1980) or Lob 1 (Russo et al., 1991, 1998b).Lob 2 have been associated with the development oflobular carcinomas, whereas no association has beenfound between Lob 3 and the development of malignan-cies (Russo et al., 1991). In order to ascertain whetherLob 1 and Lob 2 are more susceptible than Lob 3 to
undergo neoplastic transformation, we have utilizednormal breast tissues obtained from reduction mam-moplasties for developing an in vitro system that re-produces the in vivo conditions of the breast epithe-lium. Fresh and sterile tissues were digested with col-lagenase and hyaluronidase utilizing procedures that
Fig. 7. RNase protection assays of thenovel serpin L342C (top) and, 8-actin (bot-tom) in cultured human breast epithelialcells, placenta, and liver tissues. The sizes ofthe protected fragments are indicated on theside of the bands; the samples used for anal-ysis are listed on top of the lanes. The sam-ples include 4 primary cultures of normalhuman breast epithelial cells (227, 244, 248,and 250), a mortal human breast epithelialcell line S#130 (130L and 130H), immortal-ized normal human breast epithelial cell lineMCF-10F, and their chemical carcinogen-transformed non-tumorigenic (BP1, D3,D3-1) and tumorigenic (BP1E, BP1-Tras, andD3-1-Tras) derived cells; a normal humanbreast epithelial cell line (HBL-100), and thebreast carcinoma cell lines MCF-7, T47D,BT20. Yeast tRNA with or without RNasedigestion was used as controls. RNase pro-tection analysis of b-actin expression servedas controls of equal sampling.
216 J. RUSSO ET AL.

are standard in our laboratory (Russo et al., 1993a,b,1996). Digested tissues release epithelial cells in aggre-gates, also called organoids, which represent the vari-ous ductal and lobular structures that have been pre-viously described. Organoids are classified by visualinspection under the microscope into Lob 1, Lob 2, orLob 3 by applying the same criteria developed for clas-sifying these structures in whole mount and his-topathological preparations. Each lobular type is se-lected by micromanipulation, plated, and cultured inorder to evaluate whether the in vitro behavior of theepithelial cells correlates with the specific type of lob-ule that originated them. Cells from Lob 1 and Lob 2,which in organ culture have shown to exhibit a highDNA-LI, attach to the dishes promptly and start grow-ing logarithmically, whereas cells from Lob 3, whichhave a lower DNA-LI, have a long lag phase before theyattach to the dish and start growing. The number ofdoublings per lobular unit was also higher in cellsderived from Lob 1 and Lob 2 than in those derivedfrom Lob 3 (Russo et al., 1989). For testing the suscep-tibility of the different types of lobules to be trans-formed by chemical carcinogens in vitro, a total of52 human breast samples were processed. Organoidsrepresenting Lob 1, Lob 2, and Lob 3 were plated, andwhen the cells reached their logarithmic phase ofgrowth they were treated with the chemical carcino-gens N-methyl-N-nitrosourea (NMU), DMBA, meth-yl-N- nitro-nitroso-guanidine (MNNG), or benz(a-)pyrene (BP) for 24 hours. The cells were followed upfor several passages until they exhibited changes in-dicative of neoplastic transformation, such as varia-tions in cell morphology, loss of contact inhibition, andanchorage independent growth. Carcinogen-treatedcells derived from Lob 1 or Lob 2 exhibited changes inshape that were the result of increased number ofsurface microvilli and decreased cell-cell interaction.The alteration in contact inhibition also abolished the
property to form domes, a characteristic of normalbreast epithelial cells grown in plastic flasks. Treatedcells showed increased ability to survive and to formcolonies in agar methocel, and exhibited multinucle-ation. These types of responses, however, were notobserved in breast epithelial cells derived from Lob3 (Russo et al., 1993). These observations led us toconclude that primary cultures of HBEC are suscepti-ble to be transformed in vitro by chemical carcinogens,however, the expression of phenotypes indicative ofneoplastic transformation depends upon the stage ofdevelopment of the breast and of the in vivo rate of cellproliferation (Hu et al., 1996, 1997a; Russo et al.,1993a,b). The findings that in vitro cells derived fromLob 1 and Lob 2 express more readily changes indica-tive of neoplastic transformation indicate that thesestructures are more susceptible to the transformingeffect of genotoxic agents, thus supporting the obser-vations that they are the site of origin of mammarycarcinomas; it also correlates with the lack of associa-tion of the Lob 3 with the development of malignantneoplasms (Russo et al., 1989, 1993a,b). Of greaterrelevance is the observation that the breast of nullipa-rous women contains more numerous Lob 1 and Lob2 than the breast of parous women, in which Lob3 predominates, further emphasizing the protective ef-fect of gland differentiation, which modulates the re-sponse of breast epithelial cells to carcinogens under invitro conditions.
An important concept that emerged from our study ofbreast development is that the TDLU, which had beenoriginally identified by Wellings (1980) as the site oforigin of ductal carcinoma, the most common breastmalignancy, represents a specific stage of developmentof the mammary parenchyma, the Lob 1 (Fig. 9d,e)(Russo et al., 1991; Wellings et al., 1975; Wellings,1980). This observation is supported by comparativestudies of normal breast tissues and of the non-tumoral
Fig. 8. Northern analysis of the novel serpin L346C in cultures ofhuman breast epithelial cells, as influenced by concentration of Ca11
in the culture medium. The panel of cells tested included: the primarycultures 244L and 248L; the mortal human breast epithelial cell lineS#130 (130), the immortalized normal human breast epithelial cellline MCF-10F, its chemical carcinogen-transformed non-tumorigenic(BP1, D3, D3-1) and tumorigenic (BPl-E, BP1-Tras, D3-1-Tras) deriv-
atives. The cells were grown in low (L) (0.04 mM) or comparativelyhigh (H) (1.05 mM) calcium concentrations. A normal human breastepithelial cell line (HBL-100) and three breast carcinoma cell lines(MCF-7, T47D, BT20) were cultured in regular medium to comparewith the results of RNase protection assay. Intensities of ethidiumbromide staining of 28S and 18S rRNA on the membrane served ascontrols of equal sample loading.
217CANCER RISK AND MAMMARY GLAND STRUCTURE

Fig. 9.
218 J. RUSSO ET AL.

parenchyma in cancer-bearing breasts obtained at au-topsy. This analysis revealed that the tumor-freebreast tissues obtained from cancer patients containeda significantly higher number of hyperplastic terminalducts, atypical Lob 1, and ductal carcinomas in situoriginated in Lob 1 than those breasts of women free ofbreast cancer. These observations indicated that theLob 1 is affected by preneoplastic as well as by neoplas-tic processes (Russo et al., 1992, 1994). The finding thatthe most undifferentiated normal structures originatethe most undifferentiated and aggressive type of breastneoplasm acquires relevance in the light of the knowl-edge that these structures are more numerous in thebreast of nulliparous women, who are, in turn, at ahigher risk of developing breast cancer. We concludedthat the Lob 1 found in the breast of nulliparouswomen never completed the process of differentiation,whereas the same structures in the breast of postmeno-pausal parous women did. Thus, although they appearmorphologically identical, they differ in their biologicaland functional experience (Russo et al., 1992).
The study of experimentally induced mammary can-cer has revealed that not only the undifferentiatedterminal end buds and Lob 1 are affected by carcino-gens. More differentiated lobular structures respond tocarcinogenic stimuli as well, although they originatetumors whose malignancy is inversely related to thedegree of differentiation of the affected structure, i.e.,Lob 2 originate lobular carcinomas in situ, (Fig. 9f,g),Lob 3 give rise to more benign breast lesions, such ashyperplastic lobules, cysts, fibroadenomas, and adeno-mas, and Lob 4 originate lactating adenomas (Russo etal., 1991). We concluded from these observations thateach specific compartment of the breast gives origin toa specific type of lesion.
DIFFERENTIAL EXPRESSION OF HUMANFERRITIN H CHAIN GENE AND
BREAST CANCERDefining the molecular mechanisms involved in im-
mortalization of HBEC may be crucial to the under-standing of the early events in the development ofbreast cancer (Hopfer et al., 1996; Hackett et al., 1977;Russo et al., 1996). Although it has not been demon-strated whether the expression of immortalization invitro correlates with definitive stages of in vivo cancerprogression (Kohn and Liotta, 1995; Newbold, 1985),
we postulate that it may represent preneoplasticstages, such as ductal hyperplasia (Russo et al., 1992,1993, 1996). A suitable in vitro model for testing thishypothesis has been provided by the immortalization ofthe human breast epithelial cell line MCF-10F. Thesecells, derived from mortal human mammary epithelialcells S-130, became spontaneously immortalized, with-out chemical or viral intervention (Soule et al., 1991;Tait et al., 1991). We utilized this model for identifyinggenes that may be critical to the process of immortal-ization (Rhyu, 1985) by performing subtractive hybrid-ization between the mortal HBEC S-130 and its de-rived immortal cell line MCF-10F. This procedure al-lowed us to isolate the cDNA clone (233 bp) that wascloned, purified, and sequenced. Sequence comparisonin the GenBank data base revealed it to be 100% ho-mologous at the nucleotide level to the coding sequence(exon 4) of human ferritin H chain gene (accessionM11146) (Higgy et al. 1997; Russo et al., 1998b). Theunderlined sequence showed 100% homology at theamino acid translation level to the terminal portion ofthe mature peptide (nucleotides 584–626) (Boyd et al.,1985; Costanzo et al., 1984). In situ hybridization usingan antisense riboprobe for ferritin H chain showed nosignal in normal breast lobules (Fig. 9h, l); an increasedlocalized signal was observed in areas with ductal hy-perplasia (Fig. 9i, m), areas with carcinoma in situ (Fig.9j, n), and areas with infiltrating ductal carcinoma(Fig. 9k,o). No signal was detected using a sense ribo-probe (data not shown). Normal breast tissue (Lob 1,Lob 2, and Lob 3) had no detectable signal for ferritinH. One case of mild, two of moderate, and four of severeductal hyperplasia showed an increase in signal inten-sity correlating with the degree of hyperplasia. Ductalcarcinoma in situ had more intense labeling signalwhile invasive ductal and lobular carcinoma showedthe highest levels of reactivity. These observations ledus to conclude that the increased ferritin H chain genetranscription might have contributed to the immortal-ization of MCF-10F cells through one or various of thefollowing possible mechanisms: either as acting as asource of iron, which is required by rapidly dividingcells for clonal expansion; participating in free radicalreactions leading to oxidative DNA damage and muta-tion (Frenkel et al., 1995; Guyton and Kensler, 1993);or affecting immune surface antigens, thus providingimmortal cells with growth advantage by escaping im-mune surveillance. These postulates support the dem-onstration that the increase of ferritin H chain genemay provide iron necessary for clonal selection anduncontrolled growth of cells (Klausner et al., 1993), andthat iron and its binding proteins participate in a va-riety of reactions required for cell proliferation (Anison,1982; Weinberg, 1981; Dognin and Crichton, 1973;Ponka et al., 1991; Reif, 1992). Iron is also critical forthe activity of the enzyme ribonucleotide reductase, arate-limiting step in DNA synthesis (Fan et al., 1996;Richard and Ehrenberg, 1983; Zhao et al., 1994). Fer-ritin has been shown to have an immunosuppressiveeffect on host immune response in cancer patients(Moroz et al., 1977; Keown and Descaps-Latscha,1983). Placental isoferritin (PLF), an acidic form offerritin, and its p43 super heavy chain, have beendescribed to be synthesized by breast cancer cells, butto be absent in the normal breast epithelium (Rosen et
Fig. 9. a–c: Expression of MDGI in normal breast tissues detectedby in situ hybridization (Dark field). The level of expression increasesproportionally with the differentiation of the lobules. a, poorly differ-entiated Lob 1; b, moderately-differentiated Lob 3, c, the most differ-entiated Lob 4, that exhibits the maximal expression (34). d: Wholemount of ductal carcinoma in situ (Toluidine blue, 3 2.5). e: Histo-logical section of ductal carcinoma in situ. (H & E, 32.5). f: Wholemount of lobular carcinoma in situ. (Toluidine blue, 3 2.5). g: Histo-logical section of lobular carcinoma in situ. (H & E, 32.5). h: Histo-logical section of Lob 1 (H, 32.5). i:Histological section of ductalhyperplasia (H, 32.5). j: Histological section of ductal carcinoma insitu (H, 32.5). k: Invasive carcinoma (H, 32.5). l–o: l, In situ hybrid-ization of ferritin H of the normal breast shown in h; m, ductalhyperplasia, n, carcinoma in situ, and o, invasive carcinoma (Darkfield). p–s: Immunocytochemical localization of S100P in paraffinsections of formalin fixed tissue. Normal breast (H & E, 32.5); q,ductal hyperplasia; r, ductal carcinoma in situ; s, invasive ductalcarcinoma (q, r, and s, DAB, with H counterstain, 340).
219CANCER RISK AND MAMMARY GLAND STRUCTURE

al., 1992). Breast cancer associated p43 induces alter-ations of the expression of cell surface molecules inbreast cancer cells, which could have an effect on themodulation of cancer cell adhesive interactions (Rosenet al., 1991). Cytokines such as tumor necrosis factor(TNF), interleukin 1a, and NF-kB family of transcrip-tion factors specifically induce synthesis of ferritin Hby a selective increase in ferritin H transcription(Kwak et al., 1995; Rosen et al., 1991). Since the im-mortalization of MCF-10F cells occurred spontane-ously, we have no explanation for the cause of theinduction of the ferritin H chain gene. The expressionof this gene may be either a consequence of the immor-talization of the cells, or it might have preceded thisphenomenon, acting as an inducer of the same. Ineither instance, the overexpression of the ferritin Hchain gene may prove to become a valuable marker ofcell immortalization and/or an early indicator of malig-nant transformation.
S100P CALCIUM-BINDING PROTEIN AS AMARKER OF CANCER INITIATION
Normal and differentiated HBEC have a limited abil-ity to divide. Cultured cells are mortal., i.e., undergo aprogressive cessation of in vitro growth, become senes-cent, and die (Bond et al., 1994; Briand et al., 1987;Russo et al., 1993; Soule et al., 1991). The isolation andcharacterization of the spontaneously immortalizedHBEC line MCF-10F (Soule et al., 1991; Tait et al.,1991), have provided a valuable tool for addressingspecific questions on alternative pathways that mightlead a mortal cell to immortalization. The immortaliza-tion of S130 to MCF-10F cells was associated with theexpression of a balanced reciprocal chromosomal trans-location t(3;9) (3p13:9p22) (Soule et al., 1991), acquisi-tion of independence from calcium concentration in themedium for in vitro growth (Tait et al., 1991), expres-sion of an insertional mutation of p53 in exon7 (Barnabas et al., 1995), stabilization of telomerelength (Higgy et al., 1998), and overexpression of H-ferritin (Higgy et al., 1997). In order to identifywhether new genes are involved in the process of cellimmortalization, we analyzed by differential displaytechnique (Liang and Pardee, 1992; Liang et al., 1994;Zhao et al., 1995) total RNA from the spontaneouslyimmortalized HBEC line MCF-10F for a vis-a-vis com-parison with total RNA from its mortal parental coun-terpart S130. This technique allowed us to identify a439-bp band that, when using the primers HT11C andHAP-6, was found to be differentially amplified in theimmortal cell line MCF-10F. After the search for ho-mologies in gene bank databases (GenBank1EMBL1DDBJ1PDB) was completed, the cDNA band, that wascalled 10F-D showed very high homology (99%) to theH. sapiens mRNA encoding for the calcium-bindingprotein S100P (Averbouch et al., 1996; Ilg et al., 1996;Silva et al., 2000). Northern blot analysis confirmed thedifferential expression of S100P. The level of S100Pexpression was increased 9–10-fold above those foundin the mortal S130 cells in all the immortal, chemicallytransformed and malignant cells tested, which in-cluded MCF-10F, BP1-E, D3-1, and T47D. The S100Plevels in these immortalized and malignant cells werealso higher than the levels found in primary cultures ofHBCE, samples 244 and 248. The expression of S100P
was also clearly up-regulated (2–20-fold) in invasiveductal carcinomas when compared to the adjacent nor-mal tissues (Fig. 9p–s). Notably, a much higher in-crease in S100P expression (20-fold) was observedwhen RNA was obtained from a poorly differentiated,lymph node-positive, ductal invasive carcinoma sam-ple. The levels of expression were also high in variousnormal tissues examined, such as full-term placenta(10–15-fold); it was low in lung, breast, and colon, andessentially absent in other tissues. Detection of S100Pby immunocytochemistry using monoclonal antibodiesrevealed that typical and atypical ductal hyperplasias,ductal carcinomas in situ, and invasive carcinomaswere positive. The S100P protein was localized in thecytoplasm of the epithelial cells, and tended to accu-mulate at the apical and supranuclear regions of theepithelium (Fig. 9a and r). Interestingly, morphologi-cally normal breast tissues adjacent to ductal hyper-plasia, two carcinomas in situ, and invasive carcino-mas, were slightly positive for S100P, whereas normalbreast tissues were negative. S100P is a calcium-bind-ing protein that was first isolated from human placenta(Becker et al., 1992; Emoto et al., 1992). It belongs to afamily of calcium-binding proteins that was initiallycharacterized as a group of abundant low molecularweight acidic proteins (10–12 kDa) that are highlyenriched in nervous tissue (Moore, 1965). S100 pro-teins are characterized by a common structural motif,the EF-hand domain, which consists of 12 aminoacidresidues and binds to calcium with high affinity andspecificity (Sherbet and Lakshmi, 1998). Calcium is notonly an essential component of cell membrane struc-tures influencing their viscosity and permeability, butis also a major second messenger in the control of avariety of biological processes, such as cell cycle pro-gression, differentiation, and cell death (Schafer andHeizmann, 1996). An increase in the concentration ofcalcium in the culture medium decreases cell prolifer-ation and induces cell differentiation in HBEC(McGrath and Soule, 1984; Soule and McGrath, 1985),whereas decreasing the concentration of calcium in theculture medium extends the lifespan of primary cul-tures and of mortal HBEC (Russo et al., 1993b). Theintracellular concentration of calcium is tightly con-trolled by a number of calcium-binding proteins, andan increase in calcium-binding proteins could conceiv-ably diminish the intracellular pool of free calcium(Schafer and Heizmann, 1996). The findings thatS100P is overexpressed during the process of cell im-mortalization appear to provide an explanation for ourprevious observation that the intracellular concentra-tion of free calcium is higher in mortal than in immor-tal cells (Ochieng et al., 1993). It is of interest thatS100P is preferentially expressed in immortal cells,whereas the mortal parental cells and cultures of pri-mary HBEC express low levels of S100P, regardless ofthe calcium concentration in the medium. These re-sults suggest that the S100P protein is very unlikely toplay any role in intracellular calcium buffering by ex-changing calcium among different cellular compart-ments. Instead, S100P protein might be one of themolecules involved in specific pathways of cell cyclecontrol whose imbalance might enable cells to escapefrom senescence, therefore, reaching the immortal cellstatus (Derventzi et al., 1996; Kim et al., 1994).
220 J. RUSSO ET AL.

SUMMARY AND CONCLUSIONSIn summary, the human breast undergoes a com-
plete series of changes from intrauterine life to senes-cence. These changes can be divided into two distinctphases; the developmental phase and the differentia-tion phase. The developmental phase includes the earlystages of gland morphogenesis, from nipple epitheliumto lobule formation. In lobule formation, both pro-cesses, development and differentiation, take place al-most simultaneously. For example, the progressivetransition of Lob 1 to Lob 2 and Lob 3, and of these toLob 4, requires active cell proliferation for the acquisi-tion of the cell mass necessary for milk secretion. Thislater process implies differentiation of the mammaryepithelium. Therefore, the presence of Lob 4 is themaximal expression of development and differentiationin the adult gland, whereas the presence of Lob 3 couldindicate that the gland has already been developed, butbecause their lobule type is not secreting milk, they arenot completely differentiated. It is important to pointout that the presence of proteins that are indicative ofmilk secretion, such as a-lactalbumin, casein, or milkfat globule membrane protein is also indicative of thebreast epithelium type of cellular differentiation. How-ever, only when all the other components of milk, (suchas lactose, a lactalbumin, casein, and milk fat) arecoordinately synthesized within the appropriate struc-ture can full differentiation of the mammary gland beacknowledged.
ACKNOWLEDGMENTSThis work was supported by National Cancer Insti-
tute grants CA64896 (to I.H.R.) and CA67238, andNIEHS grant ESO7280 (to J.R.). The authors thankthe skillful technical assistance of Mr. Xiang Ao.
REFERENCESAakvaag A, Utaacker E, Thorsen T, Lea OA, Lahooti H. 1991. Growth
control of human mammary cancer cells (MCF-7 cells) in culture:effect of estradiol and growth factors in serum containing medium.Cancer Res 50:7806–7810.
Anison P. 1982. Current concepts in iron metabolism. Clin Haematol11:241–257.
Averbouch L, Liang P, Kantoff PW, Pardee AB. 1996. Regulation ofS100P expression by androgen. Prostate 29:350–355.
Band V, Zajchowski D, Kulesa V, Sager R. 1990.. Human papillomavirus DNA immortalize normal human mammary epithelial cellsand reduce the growth factor requirements. Proc Natl Acad Sci USA87:463–467.
Band V, Dalal S, Delmolino L, Androphy EL. 1993. Enhanced degra-dation of p53 protein in HPV-6 and BPV-1 E-6 immortalized humanmammary epithelial cells. EMBO J 12:1847–1852.
Barnabas N, Moraes R, Calaf G, Estrada S, Russo J 1995. Role of p53in MCF-10F cell immoratlization and chemically-induced neoplas-tic transformation. Int J Oncol 7:1289–1296.
Becker T, Gerke V, Kube E, Weber K. 1992. S100P: a novel calcium-binding protein from human placenta. cDNA cloning, recombinantprotein expression and calcium-binding properties. Eur J Biochem207:541–547.
Bieche I, Champeme MH, Lidereau R. 1994. A tumor suppressor geneon chromosome 1p32-pter controls the amplification of MYC familygenes in breast cancer. Cancer Res 54:4274–4276.
Bieche I, Champeme MH, Matifas F, Cropp CS, Callahan R, LidereauR. 1993. Two distinct regions involved in 1p deletion in humanprimary breast cancer. Cancer Res 53:1990–1994.
Biemond P, Swaak AJG, Beindorf CM, Koster JF. 1986. Superoxidedependent and independent mechanisms of iron mobilization fromferritin by xanthine oxidase. Biochem J 239:169–173.
Binas B, Spitzer E, Zschiesche W, Erdmann B, Kurtz A, Muller T,Niemann C, Blenau W, Grosse R. 1992. Hormonal induction offunctional differentiation and mammary-derived growth inhibitor
expression in cultured mouse mammary gland explants. In VitroCell Dev Biol 28A:625–634.
Bohmer FD, Kraft R, Otto A, Wernstedt C, Hellman U, Kurtz A,Muller T, Rohde K, Etzold G, Lehmann W, Langen P, Heldin CH,Grosse R. 1987. Identification of a polypeptide growth inhibitorfrom bovine mammary gland. Sequence homology to fatty acid- andretinoid-binding proteins. J Biol Chem 262:15137–15143.
Bond JA, Willie FS, Wynford-Thomas D. 1994.. Escape from senes-cence in human diploid fibroblasts induced directly by mutant p53.Oncogene 7:1885–1888.
Bonte F. 1995. Constitution de la matrice extracellulaire normal etpathologique. Arch Anat Cytol Path 43:170–172.
Borchers T, Hojrup P, Nielsen SU, Roepstorff P, Spener F, Knudsen J.1990. Revision of the amino acid sequence of human heart fattyacid-binding protein. Mol Cell Biochem 98:127–133.
Boyd D, Vecoli C, Belcher DM, Jain SK, Drysdale JW. 1985. Struc-tural and functional relationships of human ferritin H and L chainsdeduced from cDNA clones. J Biol Chem 260:11755–11761.
Briand P, Petersen OW, van Dews B. 1987. A new diploid nontumori-genic human breast epithelial cell line isolated and propagated inchemically defined medium. In Vitro Cell Dev Biol 23:181–188.
Calaf G, Russo J 1993. Transformation of breast epithelial cells bychemical carcinogens. Carcinogenesis 14:483–492.
Calaf G, Martinez F, Russo IH Russo J. 1982a. Age-related variationsin growth kinetics of primary human breast cell cultures. Interna-tional Research Communication System (IRCS) 10:551–553.
Calaf G, Russo IH, Roi L Russo J. 1982b. Effect of estrogen on thelength of S-phase of human breast tissue in organ culture. Interna-tional Research Communication (IRCS) 10:307–308.
Caron de Frontemel C, Nardeux PC, Soressi T, et al. 1991. EpithelialHBL-100 cell line derived from milk of an apparently healthywoman harbors SV-40 genetic information. Exp Cell Res 16:83–94.
Chandra T, Stackhouse R, Kidd VJ, Robson KJ, Woo SL.1983. Sequence homology between human alpha 1-antichymotryp-sin, alpha 1-antitrypsin, and antithrombin III. Biochemistry 22:5055–5061.
Chang SE, Keen J, Lane EB, Taylor-Papadimitriou J. 1982. Estab-lishment and character-ization of SV-40 transformed human breastepithelial cell lines. Cancer Res 42:2040–2053.
Chiquet-Ehrisman R, Mackie EJ, Pearson CA, Sakakura T.1986. Tenascin: an extracellular matrix protein involved in tissueinteractions during fetal development and oncogenesis. Cell 47:131–139.
Chu KC, Tarone RE, Brawley OW. 1999. Breast cancer trends of blackwomen compared with white women. Archives of Family Medicine8:521–528.
Ciocca DR, Parente A, Russo J. 1982. Endocrinological milieu andsusceptibility of the rat mammary gland to carcinogenesis. Am JPathol 109:47–56.
Costanzo F, Santoro C, Colantuoni V, et al. 1984. Cloning and se-quencing of a full length cDNA coding for a human apoferritin Hchain: evidence for a multigene family. The EMBO J 3:23–27.
De Waard F, Trichopoulos D. 1988. A unifying concept of the etiologyof breast cancer. Int J Cancer 41:666–669.
Dell’aquilla ML, Pigott DA, Bonaquist DL, Gaffney EV. 1984. A factorfrom plasma derived human serum that inhibits the growth of themammary cell line MCF-7: characterization and purification. J NatlCancer Inst 72:291–298.
Derventzi A, Rattan SIS, Gonos E. 1996. Molecular links betweencellular mortality and immortality. Anti Cancer Res 16:2901–2910.
Dickson RB, Huff KK, Spencer EM, Lippman ME. 1986. Introductionof epidermal growth factor related polipeptides by 17a-estradiol inMCF-7 human breast cancer cells. Endocrinology 118:138–142.
Dognin J, Crichton RR. 1973. Mobilization of iron from ferritin frac-tions of defined iron content by biological reductants. FEBS Lett54:234–236.
Emoto Y, Kobayashi R, Akatsuba H, Hidaka H. 1992. Purification andcharacterization of a new member of the S100 protein family fromhuman placenta. Biochem Biophys Res Comm 182:1246–1253.
Fan H, Villegas C, Wright JA. 1996. A link between ferritin geneexpression and ribonucleotide reductase R2 protein, as demon-strated by retroviral vector mediated stable expression of R2 cDNA.FEBS Lett 382:145–148.
Frenkel K, Wei L, Wei H. 1995. 7,12-dimethylbenz [a] anthraceneinduces oxidative DNA modification in vivo. Free Rad Biol Med19:373–380.
Fuqua SA, Russo J, Shackney SE, Stearns ME. 2000. Estrogen, es-trogen receptors, and selective estrogen receptor modulators inhuman breast cancer. J Women’s Cancer 2:21–32.
221CANCER RISK AND MAMMARY GLAND STRUCTURE

Geithner CA, Woynarowska B, Malina RM. 1998. The adolescentspurt and sexual maturation in girls active and not active in sport.An Human Biol 25: 415–423.
Genuardi M, Tsihira H, Anderson DE, Saunders GF. 1989. Distaldeletion of chromosome Ip in ductal carcinoma of the breast. Am JHum Genet 45:73–82.
Greenlee RT, Murray T, Bolden S, Wingo PA. 2000. Cancer Statistics2000. CA Cancer J Clin 50: 7–33.
Guyton KZ, Kensler TW. 1993.. Oxidative mechanisms in carcinogen-esis. Br Med Bull 49:523–533.
Hackett AJ, Smith TS, Springer EL, et al. 1977. Two syngeneic celllines from human breast tissue: the aneuploid mammary epithelial(Hs 578T) and the diploid myoepithelial (Hs 578BsT) cell lines.J Natl Cancer Inst 58:1795–1806.
Han X, Liehr JG. 1995. Microsome mediated 8-hydroxylation of gua-nine bases of DNA by steroid estrogens: correlation of DNA damageby free radicals with metabolic activation to quinones. Carcinogen-esis 16:2571–2574.
Henderson BE, Ross RK, Pike MC. 1993. Hormonal chemopreventionof cancer in women. Science 9:633–638.
Hergenroeder AC, Hill RB, Wong WW, Sangi-Haghpeykar H, TaylorW. 1999. Validity of self-assessment of pubertal maturation inAfrican American and European American adolescents. J Adoles-cent Health 24:201–205.
Higgy NA, Salicioni A, Russo IH, Zhang PL, Russo J. 1997. Dif-ferential expression of human ferritin H chain gene in immortal hu-man breast epithelial MCF-10F cells. Mol Carcinog 20:332–339.
Higgy NA, Russo J, Mgbonyebi P, Salicioni AM, Russo IH.1998. Human chorionic gonadotropin inhibits telomerase activity inhuman breast epithelial cells in vitro. Proc Am Assoc Cancer Res39:541.
Hopfer U, Jacobberger JW, Gruenert DC, Eckert RL, Jat RS, WhitsettJA. 1996. Immortalization of epithelial cells. Am J Physiol 270:c1–c11.
Hu YF, Russo IH, Zalipsky U, Russo J. 1996. Lack of involvement ofbcl2 and cyclin D1 in the early phases of human breast epithelialcell transformation by environmental chemical carcinogens. ProcAm Assoc Cancer Res 37:1005a.
Hu YF, Russo IH, Zalipsky U, Lynch HT, Russo J. 1997a. Environ-mental chemical carcinogens induce transformation of breast epi-thelial cells from women with familial history of breast cancer. InVitro Cell Dev Biol 33: 495–498.
Hu YF, Russo IH, Ao X, Russo J. 1997b. Mammary derived growthinhibitor (MDGI) cloned from human breast epithelial cells is ex-pressed in fully differentiated lobular structures. Int J Oncol 11:5–11.
Hu Y-F, Silva IDCG, Russo IH, Ao X, Russo J. 1998. A novel serpingene cloned from differentiated human breast epithelial cells is apotential tumor suppressor. Proc Am Assoc Can Res 39:775.
Huff KK, Knabbe C, Lindsey R, Kaufman D, Bronzert D, LippmanME, Dickson RB. 1988. Multihormonal regulation of insulin-likegrowth factor-1-related protein in MCF-7 human breast cancercells. Mol Endocrinol 2:200–208.
Huseby RA, Maloney TM, McGrath CM. 1987. Evidence for a directgrowth-stimulating effect of estradiol on human MCF-7 cells invitro . Cancer Res 144:2654–2659.
Huynh HT, Larsson C, Narod S, Pollak M. 1995. Tumor suppressoractivity of the gene encoding mammary-derived growth inhibitor.Cancer Res 55:2225–2231.
Ilg EC, Schafer BW, Heizmann CW. 1996. Expression pattern of S100proteins in human tumors. Int J Cancer 68:325–332.
Jones PD, Carne A, Bass NM, Grigor MR. 1988. Isolation and char-acterization of fatty acid binding proteins from mammary tissue oflactating rats. Biochem J 251: 919–925.
Katzenellenbogen BS, Kendra KL, Norman MJ, Berthois Y. 1987.Proliferation, hormonal responsiveness and estrogen receptor con-tent of MCF-7 human breast cancer cells growth in the short-termand long-term absence of estrogens. Cancer Res 47:4355–4360
Keown P, Descaps-Latscha B. 1983. In vitro suppression of cell me-diated immunity by ferro-proteins and ferric salts. Cell Immunol80:257–266.
Kim NW, Piatyszek MA, Prouse KR, Harley CB, West MD, Ho PL.1994. Specific association of telomerase activity with immortal cellsand cancer. Science (Washington, DC) 266:2011–2015.
King RJB. 1992. Effects of steroid hormones and related compoundson gene transcription. Clin Endocrinol 36:1–14.
Klausner RD, Rouault TA, Harford JB. 1993. Regulating the fate ofmRNA: The control of cellular iron metabolism. Cell 72:19–28.
Kohn EC, Liotta LA. 1995. Molecular insights into cancer invasion:strategies for prevention and intervention. Cancer Res 55:1856–1862.
Kumar V, Stack GS, Berry M, Jin JR, Chambon P. 1987. Functionaldomains of the human estrogen receptor. Cell 51:941–951.
Kurtz A, Vogel F, Funa K, Helding CH, Grosse R. 1991. Developmen-tal regulation of mammary-derived growth inhibitor expression inbovine mammary tissue. J Cell Biol 110:1779–1789.
Kwak EL, Larochelle DA, Blaumont C, Torti SV, Torti FM. 1995. Roleof NF-kB in the regulation of ferritin H by tumor necrosis factor-a.J Biol Chem 270:15285–15293.
Lambe M, Hsieh C-C, Chan H-W, Ekbom A, Trichopoulos D, AdamiHO. 1996. Parity, age at first and last birth, and risk of breastcancer: a population-based study in Sweden. Breast Cancer ResTreat 38:305–311.
Land CE, Boice JD, Shore RE, Norman JE, Tokunaga M. 1980. Breastcancer risk from low-dose exposures to ionizing radiation: results ofparallel analysis of three exposed populations of women. J NatlCancer Inst 65:353–376.
Lehmann W, Graetz H, Schutt M, Langen P. 1977. Chalone-likeinhibition of Ehrlich ascites cell proliferation in vitro by an ultra-filtrate obtained from the ascitic fluid. Acta Biol Med Germ 36:K43–K52.
Lehmann W, Graetz H, Schutt M, Langen P. 1979. Antagonisticeffects of insulin and a negative growth regulator from ascites fluidon the growth of Ehrlich ascites carcinoma cells in vitro. Exp CellRes 119:396–399.
Li M, Spitzer E, Zschiesche W, Binas B, Parczyk K, Grosse R.1995. Antiprogestins inhibit growth and stimulate differentiationin the normal mammary gland. J Cell Physiol 164:1–8.
Liang P, Pardee AB. 1992. Differential display of eukaryotic mRNA bymeans of the polymerase chain reaction. Science (Washington DC)257:967–971.
Liang P, Zhu W, Zhang X, Guo Z, O’Connell RP, Averbouck L, WangF, Pardee AB. 1994. Differential display using one-base anchoredoligo-dT primers. Nucleic Acid Res 22:5763–5764.
MacMahon B, Cole P, Liu M, Lowe CR, Mirra AP, Ravin I, Har B,Salber EJ, Valaoras VG, Yuasa S. 1970. Age at first birth and breastcancer risk. Bull World Health Org 34:209–221.
Malins DC, Holmes EH, Polissar NL, Gunselman SJ. 1993. The eti-ology of breast cancer. Characteristic alteration in hydroxyl radical-induced DNA base lesions during oncogenesis with potential forevaluating incidence risk. Cancer 71:3036–3043.
Malins DC, Polissar NL, Gunselman SJ. 1996. Progression of humanbreast cancers to the metastatic state is linked to hydroxyl radical-induced DNA damage. Proc Natl Acad Sci USA 93:2557–2563.
Markaverich BM, Gregory RR, Alejandro MA, Clark JH, Johnson GA,Middleditch BS. 1988. Methyl p-hydroxphenyllactate. An inhibitorof cell growth and proliferation and an endogenous ligand for nu-clear type-11 binding sites. J Biol Chem 263:7203–7210.
McGrath CM, Soule HD. 1984. Calcium regulation of normal humanmammary epithelial cell growth in culture. In Vitro Cell Dev Biol20:652–662.
McGregor DH, Land CE, Choi K, Tokuoka S, Liu PI, Wakabayashi I,Beebe GW. 1977. Breast cancer incidence among atomic bomb sur-vivors, Hiroshima and Nagasaki 1950–1989. J Natl Cancer Inst59:799–811.
Moore BE. 1965. A soluble protein characteristic of the nervous sys-tem. Biochem Biophys Res Commun 19:739–744.
Moroz C, Lahat N, Biniaminov M, Ramot B. 1977. Ferritin on thesurface of lymphocytes of Hodgkin’s Disease patients: a possibleblocking substance removed by levamisole. Clin Exp Immunol 29:30–34.
Muller T, Kurtz A, Vogel F, Breter H, Schneider F, Engstroem U,Mieth M, Bohmer FD, Grosse R. 1989. A mammary-derived growthinhibitor (MDGI) related 70 kDa antigen identified in nuclei ofmammary epithelial cells. J Cell Physiol 138:415–423.
Newbold RF. 1985. Multistep malignant transformation of mamma-lian cells by carcinogens: Induction of immortality as a key event.In: Barret JC, Tennant RW, editors. Carcinogenesis. New York:Raven Press, Vol 9, p 17–28.
Ochieng J, Tahin QS, Booth CC, Russo J. 1993. Buffering of intracellularcalcium in response to increase levels in mortal., immortal and trans-formed human breast epithelial cells. J Cell Biochem 46: 250–254.
Page MJ, Field JK, Everett P, Green CD. 1983. Serum regulation ofthe estrogen responsiveness of the human breast cancer cell lineMCF-7. Cancer Res 43:1244–1250.
Phelan CM, Larsson C, Baird S, Futreal PA, Ruttledge MH, MorganK, Tonin P, Hung H, Korneluk RG, Pollak MN, Narod SA. 1996. Thehuman mammary-derived growth inhibitor (MDGI) gene: genomicstructure and mutation analysis in human breast tumors. Genom-ics 34:63–68.
Ponka P, Schulman HM, Cox TM. 1991. Iron metabolism in relation toheme synthesis. In: Dailey HA, editor. Biosynthesis of heme andchlorophyllus. New York: McGraw-Hill, p 393–405.
222 J. RUSSO ET AL.

Reif DW. 1992. Ferritin as a source of iron for oxidative damage. FreeRad Biol Med 12:417–427.
Rhyu MS. 1985. Telomeres, telomerase and immortality. J Natl Can-cer Inst 87:884–894.
Richard P, Ehrenberg A. 1983. Ribonucleotide reductase:a radicalenzyme. Science 221:514–519.
Rosen HR, Flex D, Stierer M, Moroz C. 1991. Monoclonal antibodyCM-H-9 detects placental isoferritin in the serum of patients withvisceral metastases of breast cancer. Cancer Lett 59:145–151.
Rosen HR, Moroz C, Reiner A, et al. 1992. Placental isoferritin asso-ciated p43 antigen correlates with features of high differentiation inbreast cancer. Br Cancer Res Treat 24:17–26.
Rosner B, Colditz GA, Willett WC. 1994. Reproductive risk factors ina prospective study of breast cancer: the nurses health study. Am JEpidemiol 139:819–835.
Rubin H, Wang ZM, Nickbarg EB, McLarney S, Naidoo N, Schoen-berger OL, Johnson JL, Cooperman BS. 1990. Cloning, expression,purification, and biological activity of recombinant native and vari-ant human a1-antichymotrypsin. J Biol Chem 265:1199–1207.
Russo IH, Russo J. 1994a. Role of hCG and inhibin in breast cancer.Int J Oncol 4:297–306.
Russo IH, Russo J. 1996. Mammary gland neoplasia in long-termrodent studies. Environ Health Perspect 104:938–967.
Russo J, Russo IH. 1987a. Development of human mammary gland.In: Neville MC, Daniel CW, editors. The mammary gland develop-ment, regulation, and function. New York: Plenum Press, p 67–93.
Russo J, Russo IH. 1987b. Biological and molecular bases of mam-mary carcinogenesis. Lab Invest 57:112–137.
Russo J, Russo IH. 1994b. Toward a physiological approach to breastcancer prevention. Cancer Epidemiology, Biomarkers and Preven-tion 3:353–364.
Russo J, Russo IH. 1995. Hormonally induced differentiation: A novelapproach to breast cancer prevention. J Cell Biochem 22:58–64.
Russo J, Russo IH. 1997. Role of differentiation in the pathogenesisand prevention of breast cancer. Endocrine-Related Cancer 4:1–15.
Russo J, Russo IH. 1998. Mammary gland development In: Knovile E,Neill JD, editors. Encyclopedia of Reproduction, New York: Aca-demic Press, Vol 3, pp 71–80.
Russo J, Tay LK, Russo IH. 1982. Differentiation of the mammarygland and susceptibility to carcinogenesis. Breast Cancer Res Treat2: 5–73.
Russo J, Reina D, Frederick J, Russo IH 1988a. Expression of pheno-typical changes by human breast epithelial cells treated with car-cinogens in vitro. Cancer Res 48:2837–2857.
Russo J, Frederick J, Ownby H, Fine G, Hussain M, Krickstein HT,Robbins TO, Rosenberg B. 1988b. Predictors of Recurrence andSurvival of Patients with Breast Cancer. In: Hickey RC, SaundersGF, Rivera DS, editors. The yearbook of cancer. Chicago: YearbookMedical Publishers, Inc. p 296–300.
Russo J, Mills MJ, Moussalli MJ, Russo IH. 1989. Influence of breastdevelopment and growth properties in vitro . In vitro Cell Dev Biol25:643–649.
Russo J, Gusterson BA, Rogers AE, Russo IH, Wellings SR, VanZwieten MJ. 1991. Comparative study of human and rat mammarytumorigenesis. Lab Invest 62:1–32.
Russo J, Rivera R Russo IH. 1992. Influence of age and parity on thedevelopment of the human breast. Breast Cancer Res Treat 23:211–218.
Russo J, Calaf G, Russo IH. 1993a. A critical approach to the malig-nant transformation of human breast epithelial cells. CRC CriticalRev Oncogen 4:403–417.
Russo J, Calaf G, Sohi N, Tahin Q, Zhang PL, Alvarado ME, EstradaS, Russo IH. 1993b. Critical steps in breast carcinogenesis. Ann NYAcad Sci 698:1–20.
Russo J, Romero AL., Russo IH . 1994. Architectural pattern of thenormal and cancerous breast under the influence of parity. J CancerEpidemiol Biomark Prev 3:219–224.
Russo J, Barnabas N, Higgy N, Salicioni AM, Wu YL, Russo IH.1996. Molecular basis of human breast epithelial cell transformation.In: Calvo F, Crepin M, Magdalenat H, editors. Breast cancer, advancesin biology and therapeutics. John Libbey Eurotext, Paris, p 33–43.
Russo J, Hu YF, Ao X, Grill C, Russo IH. 1998a. Critical appraisal ofestrogens as carcinogenic agents in the human breast. In:Birkhauser MH, Rozenbaum H, editors. Proc. IV. European Con-gress on Menopause. Paris: Editions ESKA, p 279–287.
Russo J, Yang X, Hu YF, Bove B, Huang Y, Silva IDCG, Tahin Q, WuY, Higgy N, Zekri A, Abeer B,, Russo IH. 1998b. Molecular andbiological mechanism of breast cancer. Frontiers Biosci 3:944–960.
Russo J, Ao X, Grill C, Russo IH. 1999. Pattern of distribution of cellspositive for estrogen receptor a and progesterone receptor in rela-tion to proliferating cells in the mammary gland. Breast Cancer ResTreat 53:217–227.
Schafer BW, Heizmann CW. 1996. The S100 family of EF-hand cal-cium-binding proteins: functions and pathology. TIBS 21:134–140.
Senior RM, Griffin I, Mechan RP, Wrenn DS, Prasd KU, Urry DW.1984. Val-Gly-Val-Ala-Pro-Gly, a repeating peptide in elastin, ischemotactic for fibroblasts and monocytes. J Cell Biol 99:870–874.
Sherbet GV, Lakshmi MS. 1998. A100A4 (MTS1) calcium bindingprotein in cancer growth, invasion and metastasis. Anti Cancer Res18:2415–2422.
Shrevestha P, Kusakabe M, Mori M. 1996. Tenascin in humanneoplasia: Immuno-histochemical observations using seven differ-ent clones of monoclonal antibodies. Int J Oncol 8:741–755.
Silva IDCG, Hu YF, Russo IH, Ao X, Salicioni AM, Yang X, Russo J.2000. S100P Ca21-binding protein overexpression is associated withimmortalization and neoplastic transformation of human breastepithelial cells in vitro and tumor progression in vivo. Int J Oncol16:231–240.
Soto AM, Sonnenschein C. 1987. Cell proliferation of estrogen-sensi-tive cells: the case for negative control. Endocr Rev 48:52–58.
Soule HD, McGrath CM. 1985. A simplified method for passage andlong-term growth of human mammary epithelial cells. In Vitro22:6–12.
Soule HD, Maloney TM, Wolman SR, Peterson WD, Brenz R, McGrathCM, Russo J, Pauley RJ, Jones RF, Brooks SC. 1991. Isolation andcharacterization of a spontaneously immortalized human breast epi-thelial cell line MCF 10. Cancer Res 50:6075–6086.
Soussi T, Legros Y, Lubin R, Ory K, Schlichtholz B. 1994. Multifac-torial analysis of p53 alterations in human cancer: a review. Int JCancer 57:1–9.
Stampfer MR, Bartley JC. 1984.. Induction of transformation andcontinuous cell lines from normal human mammary epithelial cellsafter benz[a]pyrene. Proc Natl Acad Sci USA 82:2394–2398
Tait L, Soule H, Russo J. 1991. Ultrastructural and immunocytochem-ical characterization of an immortalized human breast epithelialcell line, MCF-10. Cancer Res 50:6087–6094.
Tay LK, Russo J. 1981. 7,12-Dimethylbenz(a)anthracene (DMBA)induced DNA binding and repair synthesis in susceptible and non-susceptible mammary epithelial cells in culture. J. Natl Cancer Inst67:155–161.
Tay LK, Russo J. 1982. Formation and removal of 7,12-dimethylben-z(a)anthracene nucleic acid adducts in rat mammary epithelial cellswith different susceptibility to carcinogenesis. Carcinogenesis2:1327–1333.
Tay LK, Russo J. 1985. Effect of human chorionic gonadotropin on7,12-dimethylbenz(a) anthracene-induced DNA binding and repairsynthesis by rat mammary epithelial cells. Chem-Biol Interact 55:13–21.
Theil EC. 1987. Ferritin: structure, gene regulation, and cellularfunction in animals, plants and microorganisms. Ann Rev Biochem56:289–315.
Treuner M, Kozak CA, Gallahan D, Grosse R, Muller T. 1994. Cloningand characterization of the mouse gene encoding mammary-derivedgrowth inhibitor/heart-fatty acid-binding protein. Gene 147:237–242.
Tweedie S, Edwards Y. 1989. cDNA sequence for mouse heart fattyacid binding protein, H-FABP. Nucleic Acid Res 17:4374.
Veerkamp JH, Peeters RA, Maatman RG. 1991. Structural and func-tional features of different types of cytoplasmic fatty acid-bindingproteins. Biochim Biophys Acta 1081:1–24.
Weinberg ED. 1981. Iron and neoplasia. Biol Trace Elem Res 3:55–80.Wellings SR. 1980. Development of human breast cancer. Adv Cancer
Res 31:287–299.Wellings SR, Jensen HM, Marcum RG. 1975. An atlas of subgross
pathology of 16 human breasts with special reference to possibleprecancerous lesions. J Natl Cancer Inst 55:231–275.
Wrenn DS, GriffinGL, Senior RM, Mechan RP. 1986. Characteriza-tion of biologically active domain on elastin: Identification of mono-clonal antibody to a cell recognition site. Biochemistry 25:5172–5176.
Yamada KM, Yamada SS, Partan I. 1976. Cell surface protein par-tially restores morphology, adhesivenes and contact inhibition ofmovement of fibroblast. Proc Natl Acad Sci 73:1217–1221.
Yang Y, Spitzer E, Kenney N, Zschiesche W, Li M, Kromminga A,Muller T, Spener F, Lezius A, Veerkamp JH, Smith GH, SalomonDS, Grosse R. 1994. Members of the fatty acid binding proteinfamily are differentiation factors for the mammary gland. J CellBiol 127:1097–1109.
Zhao MJ, Jung L, Tanielian C, Mechin R. 1994. Kinetics of thecompetitive degradation of deoxyribose and other biomolecules byhydroxyl radicals produced by the Fenton reaction. Free RadicalsRes 20:345–363.
Zhao S, Ooi SL, Pardee AB. 1995. New primer strategy improvesprecision of differential display. Biotechniques 18:842–850.
223CANCER RISK AND MAMMARY GLAND STRUCTURE
Copyright © 2022 FDOKUMEN




















