
Calcium-mediated interactions regulate the subcellular localization of extracellular...
13
Calcium-mediated Interactions Regulate the Subcellular Localization of Extracellular Signal-regulated Kinases * Received for publication, November 2, 2007, and in revised form, February 8, 2008 Published, JBC Papers in Press, February 11, 2008, DOI 10.1074/jbc.M709030200 Dana Chuderland ‡1 , Goldie Marmor ‡1 , Alla Shainskaya § , and Rony Seger ‡2 From the ‡ Department of Biological Regulation and § Mass Spectroscopy Unit, The Weizmann Institute of Science, Rehovot 76100, Israel The subcellular localization of ERKs in cells, which is impor- tant for proper signaling, may be regulated through protein- protein interactions. We found that inactive ERK2 interacts with a large number of proteins through its cytosolic retention sequence/common docking domain, whereas the phospho- ERK2 interacts with only few substrates. Varying calcium con- centrations significantly modified the repertoire of ERK2-inter- acting proteins, of which many were identified. The effect of calcium on ERK interactions also influenced the localization of ERKs, as calcium chelators enhanced nuclear translocation, whereas elevated calcium levels prevented it. This effect of cal- cium was apparent upon lysophosphatidic acid stimulation, where ERKs translocation was delayed compared with that induced by EGF in a calcium-dependent manner. In vitro trans- location assay revealed that high calcium concentrations affect ERK translocation by preventing the shuttling machinery through the nuclear envelope, probably due to higher binding to nuclear pore proteins. These results are consistent with a model in which ERKs in quiescent cells are bound to several cytoplas- mic proteins. Upon stimulation, ERKs are phosphorylated and released from cytoplasmic anchors to allow shuttling toward the nucleus. This translocation is delayed when calcium levels are increased, and this modifies the localization of ERKs and, there- fore, also their spatiotemporal regulation. Thus, calcium regu- lates ERK localization, which is important for the compartmen- talization of ERKs with their proper substrates and thereby their signaling specificity. The ERK 3 cascade, which is a central and ubiquitous signal- ing pathway, utilizes Raf kinases MEKs and ERKs as its kinase components (1–3). Three isoforms of ERKs are currently known, ERK1, ERK2, and ERK1b/c (4 – 6), which upon extra- cellular stimulation are activated by MEK1 and MEK2. In rest- ing cells MEKs and ERKs seem to interact with each other due to several regions in both molecules. The main interaction site of MEK in ERK seems to be the cytosolic retention sequence (CRS)/common docking (CD) domain in the C terminus of ERKs (7, 8). This site, operating together with additional domains in ERKs, is responsible for the levels of ERK phospho- rylation (9) as well as subcellular localization. The CRS/CD domain of ERKs was reported to exert its functions mainly by interaction with many proteins including ERK substrates, and thereby the domain is responsible for the phosphorylation of many ERK targets. Interestingly, the regulators and the sub- strates may compete for the interaction with the CRS/CD domain and, thus, provide an additional facet to the regulation of ERK function (10). One of the key steps in the regulation of the ERK cascade is an inducible change in the subcellular localization of its compo- nents. In resting cells, the components of the ERK cascade are localized primarily in the cytoplasm, but this is rapidly changed upon activation when MEKs and ERKs translocate into the nucleus. Whereas ERKs are transiently retained in the nucleus for minutes to hours (11), MEKs are rapidly exported from that location due to their nuclear export signal (12, 13). The accu- mulation of ERKs in the nucleus is important for proper activity of the ERK cascade and its cellular responses (14). Over the past years ERKs were found to be retained in the cytoplasm of qui- escent cells by several proteins and organelles. One group of anchoring moieties confers an irreversible interaction (e.g. microtubules (15)), whereas another set allows a stimulus-de- pendent dissociation (MEKs (16), PTP-SL (17), KSR (18)). These interactions are a main cause of the homogeneous cyto- plasmic distribution of ERKs in resting cells. Upon stimulation, ERKs are detached from many of the cyto- plasmic anchors including MEKs by a mechanism that involves conformational change of the activation loop of ERKs (19), and this allows a rapid nuclear translocation. However, the mecha- nisms that regulate this translocation are not fully understood, as the ERKs do not have a nuclear localization signal in their sequence. Thus, several distinct mechanisms that may partici- pate in the translocation have been suggested (20), including direct interaction with nuclear pore proteins (NUPs (21, 22)). The presence of ERKs in the nucleus seems to be governed by two separate mechanisms; one is de novo synthesis of nuclear anchoring proteins (11), and the other is an exportin-mediated MEK-dependent nuclear export (23). Therefore, the subcellu- lar localization of ERKs is governed by several docking interac- * This work was supported by grants from the Israel Academy of Sciences and Humanities from the European Community’s Sixth Framework Program Project IST-2004-027265-SIMAP. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. 1 These authors are equal contributors. 2 An incumbent of the Yale Lewine and Ella Miller Lewine professorial chair for cancer research. To whom correspondence should be addressed. Tel.: 972- 8-9343602; Fax: 972-8-9344116; E-mail: [email protected]. 3 The abbreviations used are: ERK, extracellular signal-regulated kinase; pERK, phosphorylated ERK; actERK, activated ERK; Ab, antibody; CRS, cytoplasmic retention sequence; CD, common docking; MAPK, mitogen-activated pro- tein kinase; MKP, MAPK phosphatase: MEK, mitogen-activated protein (MAP) kinase/extracellular signal-regulated kinase kinase; NUP, nuclear pore protein; CaMK, calcium calmodulin kinase; hnRNP, heterogeneous nuclear ribonucleoprotein K; LPA, lysophosphatidic acid; FBS, fetal bovine serum; PBS, phosphate-buffered saline; GST, glutathione S-transferase. THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 283, NO. 17, pp. 11176 –11188, April 25, 2008 © 2008 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A. 11176 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 283 • NUMBER 17 • APRIL 25, 2008 at Weizmann Institute of Science on September 8, 2008 www.jbc.org Downloaded from
Transcript of Calcium-mediated interactions regulate the subcellular localization of extracellular...

Calcium-mediated Interactions Regulate the SubcellularLocalization of Extracellular Signal-regulated Kinases*
Received for publication, November 2, 2007, and in revised form, February 8, 2008 Published, JBC Papers in Press, February 11, 2008, DOI 10.1074/jbc.M709030200
Dana Chuderland‡1, Goldie Marmor‡1, Alla Shainskaya§, and Rony Seger‡2
From the ‡Department of Biological Regulation and §Mass Spectroscopy Unit, The Weizmann Institute of Science,Rehovot 76100, Israel
The subcellular localization of ERKs in cells, which is impor-tant for proper signaling, may be regulated through protein-protein interactions. We found that inactive ERK2 interactswith a large number of proteins through its cytosolic retentionsequence/common docking domain, whereas the phospho-ERK2 interacts with only few substrates. Varying calcium con-centrations significantlymodified the repertoire of ERK2-inter-acting proteins, of which many were identified. The effect ofcalcium on ERK interactions also influenced the localization ofERKs, as calcium chelators enhanced nuclear translocation,whereas elevated calcium levels prevented it. This effect of cal-cium was apparent upon lysophosphatidic acid stimulation,where ERKs translocation was delayed compared with thatinduced by EGF in a calcium-dependent manner. In vitro trans-location assay revealed that high calcium concentrations affectERK translocation by preventing the shuttling machinerythrough the nuclear envelope, probably due to higher binding tonuclear pore proteins. These results are consistent with amodelin which ERKs in quiescent cells are bound to several cytoplas-mic proteins. Upon stimulation, ERKs are phosphorylated andreleased fromcytoplasmic anchors to allow shuttling toward thenucleus. This translocation is delayed when calcium levels areincreased, and thismodifies the localization of ERKs and, there-fore, also their spatiotemporal regulation. Thus, calcium regu-lates ERK localization, which is important for the compartmen-talization of ERKswith their proper substrates and thereby theirsignaling specificity.
The ERK3 cascade, which is a central and ubiquitous signal-ing pathway, utilizes Raf kinases MEKs and ERKs as its kinasecomponents (1–3). Three isoforms of ERKs are currently
known, ERK1, ERK2, and ERK1b/c (4–6), which upon extra-cellular stimulation are activated by MEK1 and MEK2. In rest-ing cells MEKs and ERKs seem to interact with each other dueto several regions in both molecules. The main interaction siteof MEK in ERK seems to be the cytosolic retention sequence(CRS)/common docking (CD) domain in the C terminus ofERKs (7, 8). This site, operating together with additionaldomains in ERKs, is responsible for the levels of ERK phospho-rylation (9) as well as subcellular localization. The CRS/CDdomain of ERKs was reported to exert its functions mainly byinteraction with many proteins including ERK substrates, andthereby the domain is responsible for the phosphorylation ofmany ERK targets. Interestingly, the regulators and the sub-strates may compete for the interaction with the CRS/CDdomain and, thus, provide an additional facet to the regulationof ERK function (10).One of the key steps in the regulation of the ERKcascade is an
inducible change in the subcellular localization of its compo-nents. In resting cells, the components of the ERK cascade arelocalized primarily in the cytoplasm, but this is rapidly changedupon activation when MEKs and ERKs translocate into thenucleus. Whereas ERKs are transiently retained in the nucleusfor minutes to hours (11), MEKs are rapidly exported from thatlocation due to their nuclear export signal (12, 13). The accu-mulation of ERKs in the nucleus is important for proper activityof the ERK cascade and its cellular responses (14). Over the pastyears ERKs were found to be retained in the cytoplasm of qui-escent cells by several proteins and organelles. One group ofanchoring moieties confers an irreversible interaction (e.g.microtubules (15)), whereas another set allows a stimulus-de-pendent dissociation (MEKs (16), PTP-SL (17), KSR (18)).These interactions are a main cause of the homogeneous cyto-plasmic distribution of ERKs in resting cells.Upon stimulation, ERKs are detached frommany of the cyto-
plasmic anchors includingMEKs by amechanism that involvesconformational change of the activation loop of ERKs (19), andthis allows a rapid nuclear translocation. However, the mecha-nisms that regulate this translocation are not fully understood,as the ERKs do not have a nuclear localization signal in theirsequence. Thus, several distinct mechanisms that may partici-pate in the translocation have been suggested (20), includingdirect interaction with nuclear pore proteins (NUPs (21, 22)).The presence of ERKs in the nucleus seems to be governed bytwo separate mechanisms; one is de novo synthesis of nuclearanchoring proteins (11), and the other is an exportin-mediatedMEK-dependent nuclear export (23). Therefore, the subcellu-lar localization of ERKs is governed by several docking interac-
* This work was supported by grants from the Israel Academy of Sciences andHumanities from the European Community’s Sixth Framework ProgramProject IST-2004-027265-SIMAP. The costs of publication of this articlewere defrayed in part by the payment of page charges. This article musttherefore be hereby marked “advertisement” in accordance with 18 U.S.C.Section 1734 solely to indicate this fact.
1 These authors are equal contributors.2 An incumbent of the Yale Lewine and Ella Miller Lewine professorial chair for
cancer research. To whom correspondence should be addressed. Tel.: 972-8-9343602; Fax: 972-8-9344116; E-mail: [email protected].
3 The abbreviations used are: ERK, extracellular signal-regulated kinase; pERK,phosphorylated ERK; actERK, activated ERK; Ab, antibody; CRS, cytoplasmicretention sequence; CD, common docking; MAPK, mitogen-activated pro-tein kinase; MKP, MAPK phosphatase: MEK, mitogen-activated protein(MAP) kinase/extracellular signal-regulated kinase kinase; NUP, nuclearpore protein; CaMK, calcium calmodulin kinase; hnRNP, heterogeneousnuclear ribonucleoprotein K; LPA, lysophosphatidic acid; FBS, fetal bovineserum; PBS, phosphate-buffered saline; GST, glutathione S-transferase.
THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 283, NO. 17, pp. 11176 –11188, April 25, 2008© 2008 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A.
11176 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 283 • NUMBER 17 • APRIL 25, 2008
at Weizm
ann Institute of Science on S
eptember 8, 2008
ww
w.jbc.org
Dow
nloaded from

tions with proteins in the cytoplasm, nuclear pores, and thenucleus. Elucidation of all these interacting proteinsmay revealnew aspects of the regulation of signaling through the ERKcascade.Stimulation of ERK activity is induced by various extracellu-
lar agents, including growth factors and hormones (4). In manysystems, this stimulation is mediated or affected by elevation ofintracellular calcium levels. Calcium is a versatile second mes-senger that affects many signaling processes in the cell (24, 25).The involvement of elevated calcium in activation of the ERKcascade has been studied mainly in neurons but also in othercell types (26–28). Several molecular mechanisms have beenimplicated in the calcium-dependent transmission of signals toERKs. Thus, in EGF-stimulated vascular smooth muscle cells,elevated calcium levels induce activation of the ERK cascadethrough CaMKII, which is dependent on PYK2 and Src activa-tion (29).Moreover, PYK2 (30) and CaMKII (31) were reportedto stimulate ERKs upon calcium elevation independently ofeach other. Another calcium mediator that may induce ERKsactivation is CaMKI that was shown to operate upon depolar-ization of neuroblastoma cells. Finally, the activation of ERKsby calcium can bemediated by RAS-GRP, which is a Ras guanylnucleotide-releasing protein with calcium and diacylglycerolbinding motifs (32). These studies show that ERKs can beaffected by calcium and participate in the transmission of sig-nals that induce high calcium concentrations.During a study that was aimed at identifying ERK2 interac-
tions,we found that inactive ERK2 interactswithmanyproteinsthrough its CRS/CD domain, whereas pERK2 interacts withfewer targets.We then looked for factors thatmaymodulate theinteractions and found that changes in calcium concentrationsprofoundly modified the repertoire of bound proteins, whichincluded cytoskeletal and ribosomal proteins, ERKs effectors,and others. EGF treatment, which only slightly increases calciumconcentrations, did not affect the ERK2 interactions, suggestingthat the phosphorylation of ERKs and not of their substrates isimportant for the interaction. Because protein-protein interac-tions play a role in determining the subcellular localization ofERKs, we examined this distribution upon modification ofcalcium concentrations. We found that calcium chelatorsenhanced nuclear translocation, whereas calcium ionophoresprevented this translocation. A possible physiological effect ofcalcium on ERK signaling was shown with lysophosphatidicacid (LPA) that profoundly increases calcium concentration inRat1 cells. This increment delayed the nuclear translocation ofERKs as compared with translocation induced by EGF. Thiseffect of LPAoccurred in a calcium-dependentmanner as it wasmodified by calcium chelation. The molecular mechanism ofthe calcium effect is likely to be induced by inhibition of nuclearpore transport, as we found that ERKs are tightly bound toNUPs upon elevation of calcium concentration. We proposethat high calcium levels prevent the nuclear shuttling and,therefore, play an important role in compartmentalizing ERKs.This allows interactions of ERKs with their proper substratesunder varying physiological conditions and thereby determinesthe signaling specificity of the ERK cascade.
EXPERIMENTAL PROCEDURES
Reagents—Epidermal growth factor, ionomycin, bovineserum albumin, LPA, monoclonal anti-pERK and proteinkinase C and polyclonal anti-ERK, -MEK, -RSK, -KSR, -MKP1,-MKP3, p38MAPK, and -CaMKII antibodies (Abs) were fromSigma. Polyclonal anti-pElk-1, -Elk-1, -phosphorylated MEK,-phosphorylated RSK, -Lyn, -Fyn, -protein kinase D, -p27KIP,-peroxisome proliferator-activated receptor �, -c-Fos, �-arres-tin, and -actin Abs were from Santa Cruz Biotechnology, Inc.Anti-tubulin and vinculin Abs and purified vinculin were giftsfromDr. Benny Geiger from theWeizmann Institute, Rehovot,Israel, and hnRNP-K was a gift from Dr. Karol Bomsztyk fromthe University of Washington (Seattle, WA). BAPTA-AM,alkaline phosphatase-, peroxidase-, and lissamine rhodamine-conjugated secondary Abs were from Jackson Immunore-search. Digitonin was purchased from Calbiochem, nitro bluetetrazolium/5-bromo-4-chloro-3-indolyl phosphate was fromPromega, and the ECL kit was from Amersham Biosciences.Recombinant NUP153 was a gift from Dr. Michael Elbaumfrom the Weizmann Institute of Science. MEK1 was preparedas previously reported (12), and recombinant Elk1 was fromCell Signaling. Glutathione beads were from Amersham Bio-sciences, and Gelcode was from Pierce.Cell Culture, Stimulation, and Determination of ERK
Phosphorylation—Rat1 cells were cultured in Dulbecco’s mod-ified Eagle’s medium supplemented with 10% fetal bovineserum (FBS), 2 mM glutamine, and antibiotics. Subconfluentcells were serum-starved (0.1% FBS, 16 h) and stimulated withvarious stimuli. The treated cells were washed twice with phos-phate-buffered saline (PBS) and once with buffer A (50 mM
�-glycerophosphate, pH 7.3, 1 mM EDTA, 1 mM dithiothreitol,0.1 mM sodium orthovanadate). Cells were lysed in radioim-mune precipitation assay buffer (25 mM Tris, pH 7.4, 150 mM
KCl, 5mMEDTA, 1%Nonidet P-40, 0.5% sodiumdeoxycholate,and 0.1% SDS) for 5 min on ice. The lysates were centrifuged(20,000 � g, 15 min 4 °C). Similar amounts of proteins fromeach fraction were subjected to SDS-PAGE and Western blot-ting with relevant Abs and developed with either nitro bluetetrazolium/5-bromo-4-chloro-3-indolyl phosphate or ECL.Preparation of Immobilized ERK2 Columns—To prepare the
columns, cDNA of human ERK2 was inserted into the BamH1site of pGEX-2T (AP Biotech), fusing it to the C terminus ofGST.GST-316A-ERK2was prepared by inserting the appropri-ate cDNA (7) into the same sites instead of the wild type (wt)ERK2. BL21 bacteria were transformed with the GST-wtERK2,GST-316A-ERK2, and pGEX-2T constructs. For preparation ofimmobilized doubly phosphorylated, activated ERK2 (GST-actERK), theGST-wtERK2was coexpressedwith constitutivelyactiveMEK1 (�N-EE-MEK1 (12)) and then treated as the otherconstructs. Bacterial pellets were suspended in storage buffer (5�g/ml leupeptin, 10 �g/ml aprotinin, 1 �g/ml pepstatin, 1 mM
phenylmethylsulfonyl fluoride), lysed by sonication (7 times20 s), and then left shaking with 1% Triton X-100 at 4 °C for 30min. Glutathione-Sepharose 4B beads (Amersham Bio-sciences) were added to the lysed bacteria, and after 1 h ofincubation the beads were washed twice with PBS and oncewith 1 mg/ml BSA in storage buffer. The GST-ERK2 column
Calcium Affects ERK2 Interactions and Localization
APRIL 25, 2008 • VOLUME 283 • NUMBER 17 JOURNAL OF BIOLOGICAL CHEMISTRY 11177
at Weizm
ann Institute of Science on S
eptember 8, 2008
ww
w.jbc.org
Dow
nloaded from

contained mainly non-phosphorylated ERK2 but also mono-Tyr(P) ERK2. The immobilized GST-ERK2 was readily phos-phorylated by active MEK1 in vitro, indicating that the enzymewas properly folded. The GST-actERK2 column containedabout 65% doubly phosphorylated ERK2 but also mono-Tyr(P)and some (10%) non-phosphorylated ERK2. Moreover, theimmobilized GST-actERK2 demonstrated substantial activitytoward MBP (�200 nmol/min/mg), indicating proper folding.To initialize the experiments, the amount of glutathione-boundproteins in the column was equilibrated (�250 �g/0.5 mlbeads), and the columns were extensively washed with buffer Aand buffer A� 1.0 MNaCl to remove residual nonspecific bind-ing of bacterial proteins.Loading and Development of the GST-ERK2 Columns—Rat1
cells (2.5� 108 cells for each column)were serum-starved (0.1%FBS, 16 h) and then treated as described. Subsequently, the cellswerewashed twicewith PBS and oncewith bufferH (bufferA�protease inhibitors), and the cells were harvested into buffer H.Extracts (containing cytoplasmic and nuclear fractions) wereprepared by sonication (3 � 20 s) followed by centrifugation(20,000� g, 15 min), which yielded 3mg of total extract in 3mlbuffer H. The extracts were precleared on GST-glutathionebeads (0.5 ml) and loaded on the various columns describedabove at 0.5ml/min. The columnswerewashedwith 10 columnvolumes buffer A followed by 4 column volumes of 0.1 M NaClin buffer A to release proteins that do not bind at physiologicalsalt concentration. The remaining proteinswere elutedwith 0.2M NaCl in buffer A. The columns with BAPTA-AM and iono-mycin-treated extracts were developed similarly, but in keepingwith the intracellular calcium concentration, 1mM calciumwasadded to the ionomycin-treated extracts, whereas EGTA (1.5mM) was added to columns of BAPTA-AM-treated extracts.The eluates were subjected to SDS-PAGE followed by silverstaining or Western blotting.Mass Spectrometry—Column eluates were subjected to SDS-
PAGE, stained with Gelcode, and analyzed by the Mass Spec-trometry Unit of the Weizmann Institute of Science.Localization Studies—Rat1 cells were grown on glass cover-
slips under the conditions described above. After serum star-vation (16 h, 0.1% FBS), the cells were treated as above,washed twice with PBS, fixed with 3% paraformaldehyde for20min, and permeabilized with 0.2% Triton X-100 in PBS for5 min. Next, the cells were incubated with the relevant pri-mary and secondary Abs, and the fluorescence imaging wasperformed using Nikon (EFD-3) fluorescent microscope at�400 magnification.
RESULTS
Interaction of ERK2, but Not Its Phosphorylated Form, witha Large Number of Proteins Is Mediated by the CRS/CDDomain—Interaction of ERKs with various cytoplasmic pro-teins is important for their regulation and subcellular localiza-tion (14). Here we used a proteomic approach to identify pro-teins that interact with ERK2 as well as with its activated,doubly phosphorylated form (actERK2) and with ERK2mutated in its CRS/CD domain (316A-ERK2). To this end, weprepared several columns, including GST-ERK2, GST-actERK2,GST-316A-ERK2, andGST control, and used them to
fractionate treated Rat1 cells. Silver staining of eluates from theGST-ERK2 column loaded with extracts from non-stimulatedRat1 cells revealed about 50 protein bands of various molecularweights (Fig. 1A). The same protein distribution was observedin eluates of extracts from cells treated with EGF for 5, 15, and30 min (Fig. 1A, and data not shown). In contrast to the largenumber of proteins eluted from the GST-ERK2 column, onlytwo bands with molecular masses of 60 and 70 kDa and a fewother weak bands were detected in the eluates of GST-actERK2column loaded with extracts from non-stimulated as well asEGF-stimulated Rat1 cells. Interestingly, also the eluates of theGST-316A-ERK2 column contained only a small number ofproteins including the 60- and 70-kDa proteins seen in theGST-actERK2 and few other weak bands, different from thosein the eluates of the GST-actERK2 (Fig. 1). No proteins weredetected in the GST column eluates, demonstrating bindingspecificity.To verify that the proteins are bound through their interac-
tion with the residues mutated in the 316A-ERK2, we used abinding competition with CRS/CD-derived decapeptide (P2).When this peptide was added to the GST-ERK2 column duringthe loading anddeveloping procedures, it competed outmost ofthe bands seen in the regular columnunder the procedure used,whereas a control peptide derived from FAR1 (P1) did notchange the binding and elution (Fig. 1B). Taken together, theseresults indicate that ERK2 binds a large number of proteinsthrough its CRS/CD domain and that this binding may bereleased by the phosphorylation of ERK2. The fact that theshort-term treatment with EGF does not change the bindingindicates that the release is due to phosphorylation of ERKsand not due to changes in the binding proteins upon EGFstimulation.Changes in Calcium Concentrations Modify the Repertoire of
ERK2-binding Proteins—Althoughno changeswere detected inthe repertoire of ERK2-interacting proteins upon stimulationwith EGF (Fig. 1) and other mitogenic agents (not shown),changes in calcium concentrations did modify the interactions.Thus, Rat1 cells were treated for short time periods with eitherthe calcium ionophore, ionomycin, to increase intracellular cal-cium levels (from100nm to�2�M) or the intracellular calciumchelator BAPTA-AM to reduce intracellular calcium by about5-fold. The extracts that were yielded from these cells wereloaded on the GST columns, and the development of the col-umns proceeded as described above, except that 1 mM calciumwas added to the columns loaded with extracts from the iono-mycin-treated cells, and the development of columns loadedwith extracts from BAPTA-AM-treated cells was done in thepresence of EGTA (1.5 mM). Silver staining of eluates from theGST-ERK2 column loaded with extracts from BAPTA-AM-treated cells revealed a reduction in binding of many of theinteracting proteins, and some of them were not detected at all(Fig. 1C). On the other hand, the eluates from ionomycin-treated cells contained largely similar proteins to those seen ineluates of non-treated or EGF-treated extracts, although therelative intensity of the protein bands was different, and somenew proteins were detected. Interestingly, the GST-actERK2eluates of BAPTA-AM-treated cell extracts contained a signif-icant number of proteinswith an overall lower intensity as com-
Calcium Affects ERK2 Interactions and Localization
11178 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 283 • NUMBER 17 • APRIL 25, 2008
at Weizm
ann Institute of Science on S
eptember 8, 2008
ww
w.jbc.org
Dow
nloaded from
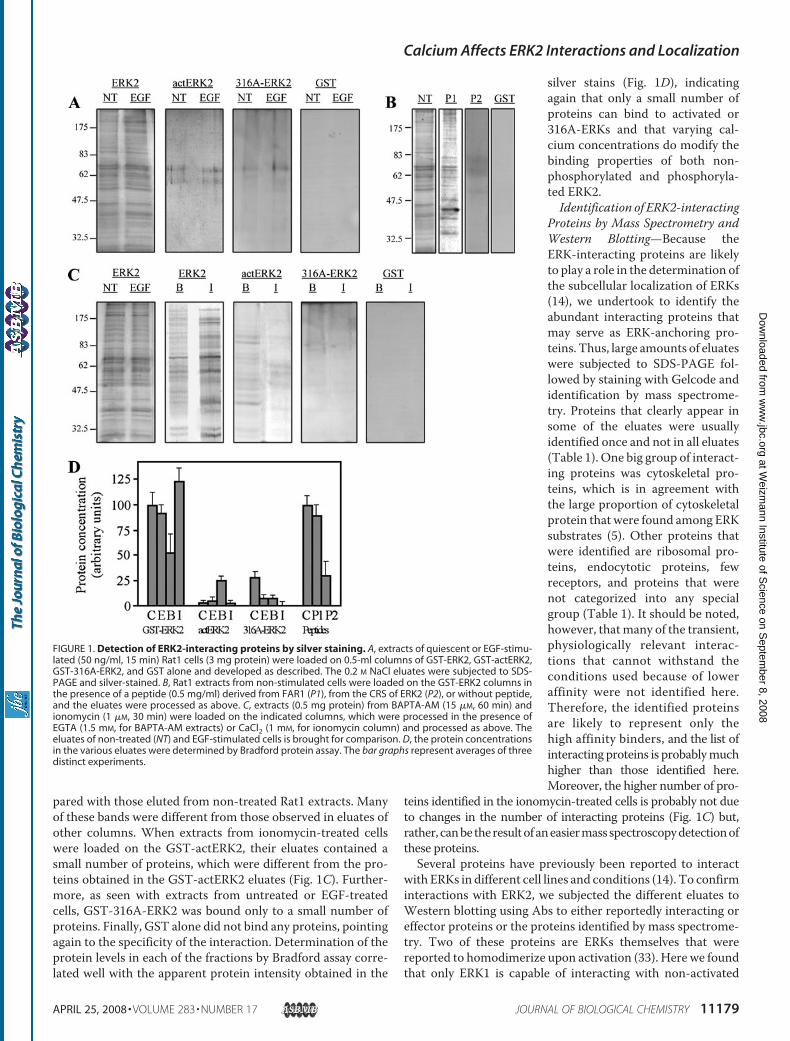
pared with those eluted from non-treated Rat1 extracts. Manyof these bands were different from those observed in eluates ofother columns. When extracts from ionomycin-treated cellswere loaded on the GST-actERK2, their eluates contained asmall number of proteins, which were different from the pro-teins obtained in the GST-actERK2 eluates (Fig. 1C). Further-more, as seen with extracts from untreated or EGF-treatedcells, GST-316A-ERK2 was bound only to a small number ofproteins. Finally, GST alone did not bind any proteins, pointingagain to the specificity of the interaction. Determination of theprotein levels in each of the fractions by Bradford assay corre-lated well with the apparent protein intensity obtained in the
silver stains (Fig. 1D), indicatingagain that only a small number ofproteins can bind to activated or316A-ERKs and that varying cal-cium concentrations do modify thebinding properties of both non-phosphorylated and phosphoryla-ted ERK2.Identification of ERK2-interacting
Proteins by Mass Spectrometry andWestern Blotting—Because theERK-interacting proteins are likelyto play a role in the determination ofthe subcellular localization of ERKs(14), we undertook to identify theabundant interacting proteins thatmay serve as ERK-anchoring pro-teins. Thus, large amounts of eluateswere subjected to SDS-PAGE fol-lowed by staining with Gelcode andidentification by mass spectrome-try. Proteins that clearly appear insome of the eluates were usuallyidentified once and not in all eluates(Table 1). One big group of interact-ing proteins was cytoskeletal pro-teins, which is in agreement withthe large proportion of cytoskeletalprotein that were found among ERKsubstrates (5). Other proteins thatwere identified are ribosomal pro-teins, endocytotic proteins, fewreceptors, and proteins that werenot categorized into any specialgroup (Table 1). It should be noted,however, thatmany of the transient,physiologically relevant interac-tions that cannot withstand theconditions used because of loweraffinity were not identified here.Therefore, the identified proteinsare likely to represent only thehigh affinity binders, and the list ofinteracting proteins is probablymuchhigher than those identified here.Moreover, the higher number of pro-
teins identified in the ionomycin-treated cells is probably not dueto changes in the number of interacting proteins (Fig. 1C) but,rather, canbe theresultofaneasiermass spectroscopydetectionofthese proteins.Several proteins have previously been reported to interact
with ERKs in different cell lines and conditions (14). To confirminteractions with ERK2, we subjected the different eluates toWestern blotting using Abs to either reportedly interacting oreffector proteins or the proteins identified by mass spectrome-try. Two of these proteins are ERKs themselves that werereported to homodimerize upon activation (33). Here we foundthat only ERK1 is capable of interacting with non-activated
FIGURE 1. Detection of ERK2-interacting proteins by silver staining. A, extracts of quiescent or EGF-stimu-lated (50 ng/ml, 15 min) Rat1 cells (3 mg protein) were loaded on 0.5-ml columns of GST-ERK2, GST-actERK2,GST-316A-ERK2, and GST alone and developed as described. The 0.2 M NaCl eluates were subjected to SDS-PAGE and silver-stained. B, Rat1 extracts from non-stimulated cells were loaded on the GST-ERK2 columns inthe presence of a peptide (0.5 mg/ml) derived from FAR1 (P1), from the CRS of ERK2 (P2), or without peptide,and the eluates were processed as above. C, extracts (0.5 mg protein) from BAPTA-AM (15 �M, 60 min) andionomycin (1 �M, 30 min) were loaded on the indicated columns, which were processed in the presence ofEGTA (1.5 mM, for BAPTA-AM extracts) or CaCl2 (1 mM, for ionomycin column) and processed as above. Theeluates of non-treated (NT) and EGF-stimulated cells is brought for comparison. D, the protein concentrationsin the various eluates were determined by Bradford protein assay. The bar graphs represent averages of threedistinct experiments.
Calcium Affects ERK2 Interactions and Localization
APRIL 25, 2008 • VOLUME 283 • NUMBER 17 JOURNAL OF BIOLOGICAL CHEMISTRY 11179
at Weizm
ann Institute of Science on S
eptember 8, 2008
ww
w.jbc.org
Dow
nloaded from
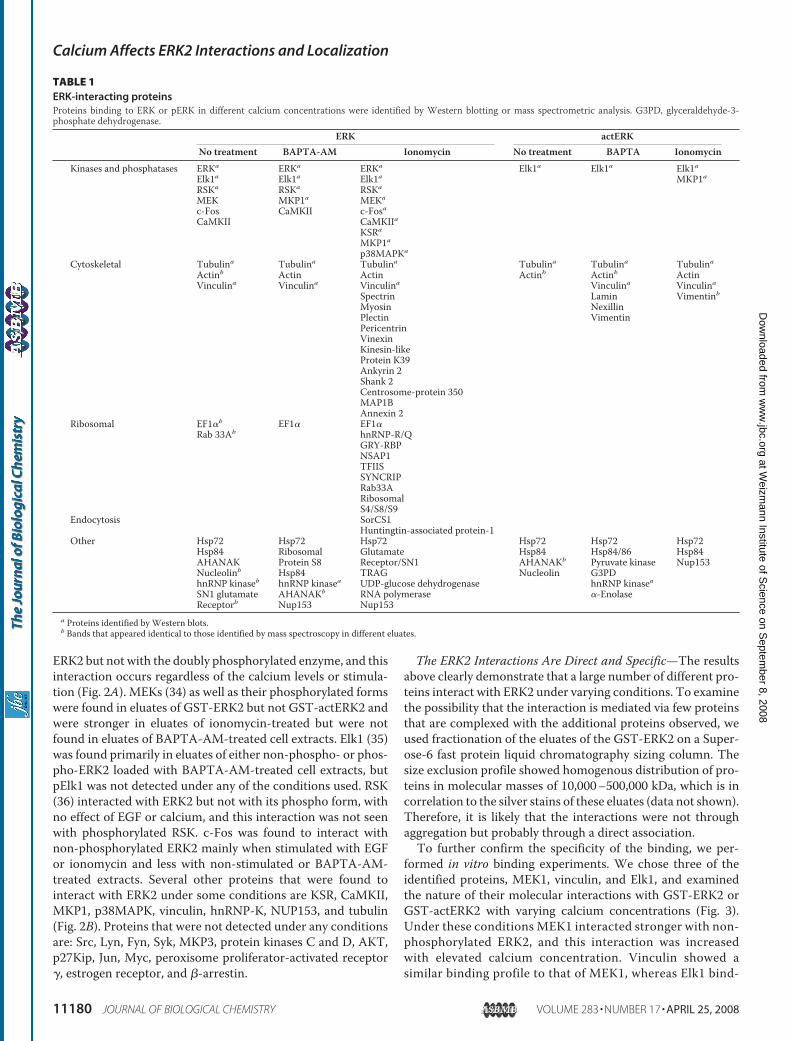
ERK2 but not with the doubly phosphorylated enzyme, and thisinteraction occurs regardless of the calcium levels or stimula-tion (Fig. 2A). MEKs (34) as well as their phosphorylated formswere found in eluates of GST-ERK2 but not GST-actERK2 andwere stronger in eluates of ionomycin-treated but were notfound in eluates of BAPTA-AM-treated cell extracts. Elk1 (35)was found primarily in eluates of either non-phospho- or phos-pho-ERK2 loaded with BAPTA-AM-treated cell extracts, butpElk1 was not detected under any of the conditions used. RSK(36) interacted with ERK2 but not with its phospho form, withno effect of EGF or calcium, and this interaction was not seenwith phosphorylated RSK. c-Fos was found to interact withnon-phosphorylated ERK2 mainly when stimulated with EGFor ionomycin and less with non-stimulated or BAPTA-AM-treated extracts. Several other proteins that were found tointeract with ERK2 under some conditions are KSR, CaMKII,MKP1, p38MAPK, vinculin, hnRNP-K, NUP153, and tubulin(Fig. 2B). Proteins that were not detected under any conditionsare: Src, Lyn, Fyn, Syk, MKP3, protein kinases C and D, AKT,p27Kip, Jun, Myc, peroxisome proliferator-activated receptor�, estrogen receptor, and �-arrestin.
The ERK2 Interactions Are Direct and Specific—The resultsabove clearly demonstrate that a large number of different pro-teins interact with ERK2 under varying conditions. To examinethe possibility that the interaction is mediated via few proteinsthat are complexed with the additional proteins observed, weused fractionation of the eluates of the GST-ERK2 on a Super-ose-6 fast protein liquid chromatography sizing column. Thesize exclusion profile showed homogenous distribution of pro-teins in molecular masses of 10,000–500,000 kDa, which is incorrelation to the silver stains of these eluates (data not shown).Therefore, it is likely that the interactions were not throughaggregation but probably through a direct association.To further confirm the specificity of the binding, we per-
formed in vitro binding experiments. We chose three of theidentified proteins, MEK1, vinculin, and Elk1, and examinedthe nature of their molecular interactions with GST-ERK2 orGST-actERK2 with varying calcium concentrations (Fig. 3).Under these conditions MEK1 interacted stronger with non-phosphorylated ERK2, and this interaction was increasedwith elevated calcium concentration. Vinculin showed asimilar binding profile to that of MEK1, whereas Elk1 bind-
TABLE 1ERK-interacting proteinsProteins binding to ERK or pERK in different calcium concentrations were identified by Western blotting or mass spectrometric analysis. G3PD, glyceraldehyde-3-phosphate dehydrogenase.
ERK actERKNo treatment BAPTA-AM Ionomycin No treatment BAPTA Ionomycin
Kinases and phosphatases ERKa ERKa ERKa Elk1a Elk1a Elk1aElk1a Elk1a Elk1a MKP1aRSKa RSKa RSKa
MEK MKP1a MEKa
c-Fos CaMKII c-FosaCaMKII CaMKIIa
KSRa
MKP1ap38MAPKa
Cytoskeletal Tubulina Tubulina Tubulina Tubulina Tubulina TubulinaActinb Actin Actin Actinb Actinb ActinVinculina Vinculina Vinculina Vinculina Vinculina
Spectrin Lamin VimentinbMyosin NexillinPlectin VimentinPericentrinVinexinKinesin-likeProtein K39Ankyrin 2Shank 2Centrosome-protein 350MAP1BAnnexin 2
Ribosomal EF1�b EF1� EF1�Rab 33Ab hnRNP-R/Q
GRY-RBPNSAP1TFIISSYNCRIPRab33ARibosomalS4/S8/S9
Endocytosis SorCS1Huntingtin-associated protein-1
Other Hsp72 Hsp72 Hsp72 Hsp72 Hsp72 Hsp72Hsp84 Ribosomal Glutamate Hsp84 Hsp84/86 Hsp84AHANAK Protein S8 Receptor/SN1 AHANAKb Pyruvate kinase Nup153Nucleolinb Hsp84 TRAG Nucleolin G3PDhnRNP kinaseb hnRNP kinasea UDP-glucose dehydrogenase hnRNP kinaseaSN1 glutamate AHANAKb RNA polymerase �-EnolaseReceptorb Nup153 Nup153
a Proteins identified by Western blots.b Bands that appeared identical to those identified by mass spectroscopy in different eluates.
Calcium Affects ERK2 Interactions and Localization
11180 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 283 • NUMBER 17 • APRIL 25, 2008
at Weizm
ann Institute of Science on S
eptember 8, 2008
ww
w.jbc.org
Dow
nloaded from

ing was detected under all conditions but was slightly higherin lower calcium concentrations. These results, which weresimilar with the endogenous proteins, indicate that the
interactions are direct and can be observed in the absence ofadditional proteins.Decreasing Intracellular Calcium Concentrations Enhances
Nuclear Translocation of ERKs in Rat1 Cells—As describedabove, the subcellular localization of ERKs may be regulated byprotein-protein interactions. We demonstrated here that cal-cium profoundly affects protein interactions of ERK2 (Figs. 1and 2), including proteins that may affect the subcellular local-ization of ERKs such as MEK1 and vinculin (Fig. 3) that directERKs to the cytoplasm (Ref. 7 and data not shown). Therefore,we hypothesized that calcium may determine the subcellularlocalization of ERKs and undertook to study the effects of vary-ing calcium concentrations on the translocation of ERKs intothe nucleus. As expected, ERKs were detected in the cytoplasmof resting Rat1 cells, whereas EGF stimulation resulted in trans-location of ERKs into the nucleus (Fig. 4A). Interestingly,BAPTA-AM caused a slow but steady nuclear accumulationthat peaked within 1 h and remained high for several hours.Pretreatment with BAPTA-AM before EGF stimulation signif-icantly enhanced the nuclear translocation of ERKs, which wasmaximal 5 min after EGF stimulation and remained high at alltimes tested.Because nuclear translocation of ERKs seems to be linked to
their TEY phosphorylation (19), it was important to study therole of this phosphorylation in BAPTA-AM-induced nuclearlocalization of ERKs. Western blotting with anti-pERK Abrevealed that treatment of cells with BAPTA-AM induces a lowlevel of ERKs phosphorylation (Fig. 4B). Moreover, pretreat-mentwithBAPTA-AMbefore stimulationwith EGF resulted inenhanced phosphorylation of ERKs. To determine whether theincreased ERKs translocation is a result of enhanced phospho-rylation, we defined activation conditions using EGF alone orBAPTA-AM plus EGF that produced equivalent amounts ofpERKs (Fig. 4C). Staining of these cells with anti-general ERKAb demonstrated that despite the comparable levels of ERKsactivation, the nuclear translocation of ERKs was much moreextensive in cells pretreated with BAPTA-AM than in thosestimulated with EGF alone (Fig. 4D). Thus, the nuclear translo-cation caused by decreased intracellular calcium concentra-tions is probably independent of the effect of reduced calciumon the activation of ERKs.Increasing Intracellular Calcium Concentrations Inhibits
Nuclear Translocation of ERKs—Next, the effect of increasingintracellular calcium concentrations on ERKs activation andsubcellular localization was examined. An increase in intracel-lular calcium concentrations was achieved by treatment of thecells with the calcium ionophores ionomycin and A23187 thathave previously been shown to induce a 10–15-fold elevation inthe amount of free calcium in Rat1 cells (37). However, neitherionomycin (Fig. 4E) nor A23187 (not shown) induced nucleartranslocation of ERKs under any of the conditions examined.Furthermore, the addition of ionomycin (Fig. 4E) or A23187(not shown) before EGF stimulation inhibited the EGF-inducednuclear translocation of ERKs in a dose-dependent manner,although some accumulation of ERKs was detected in theperinuclear region of the Rat1 cells. Interestingly, this inhibi-tion occurred despite substantial activation of ERKs by ionomy-cin or A23187, which was comparable with the EGF-induced
FIGURE 2. Identification of ERK2-interacting proteins by Western blot-ting. The eluates described in Fig. 1 were subjected to Western blot analysiswith the indicated Abs. The blots were developed as described. NT, not treat-ed; B, BAPTA-AM; I, ionomycin; E, EGF.
FIGURE 3. In vitro binding of MEK1, vinculin, and Elk1 to ERK2. 1 �g ofrecombinant GST-ERK2 was incubated with 0.5 �g of recombinant His-MEK1,purified vinculin, or recombinant Elk-1 (1 h at 23 °C) in buffer containing 20mM HEPES, pH 7.4, 20 mM MgCl2, 20 mM NaCl, 1 mM dithiothreitol, and 10mg/ml aprotinin with the indicated concentration of CaCl2. The beads werewashed 3 times with a buffer containing 20 mM HEPES, pH 7.4, 20 mM MgCl2,100 mM NaCl, and 1 mM dithiothreitol with the same CaCl2 concentrations.Then the beads were boiled in sample buffer and subjected to Western blot-ting with anti-MEKs, vinculin, and Elk-1 Abs. Total loading was detected bypolyclonal anti-GST Ab.
Calcium Affects ERK2 Interactions and Localization
APRIL 25, 2008 • VOLUME 283 • NUMBER 17 JOURNAL OF BIOLOGICAL CHEMISTRY 11181
at Weizm
ann Institute of Science on S
eptember 8, 2008
ww
w.jbc.org
Dow
nloaded from
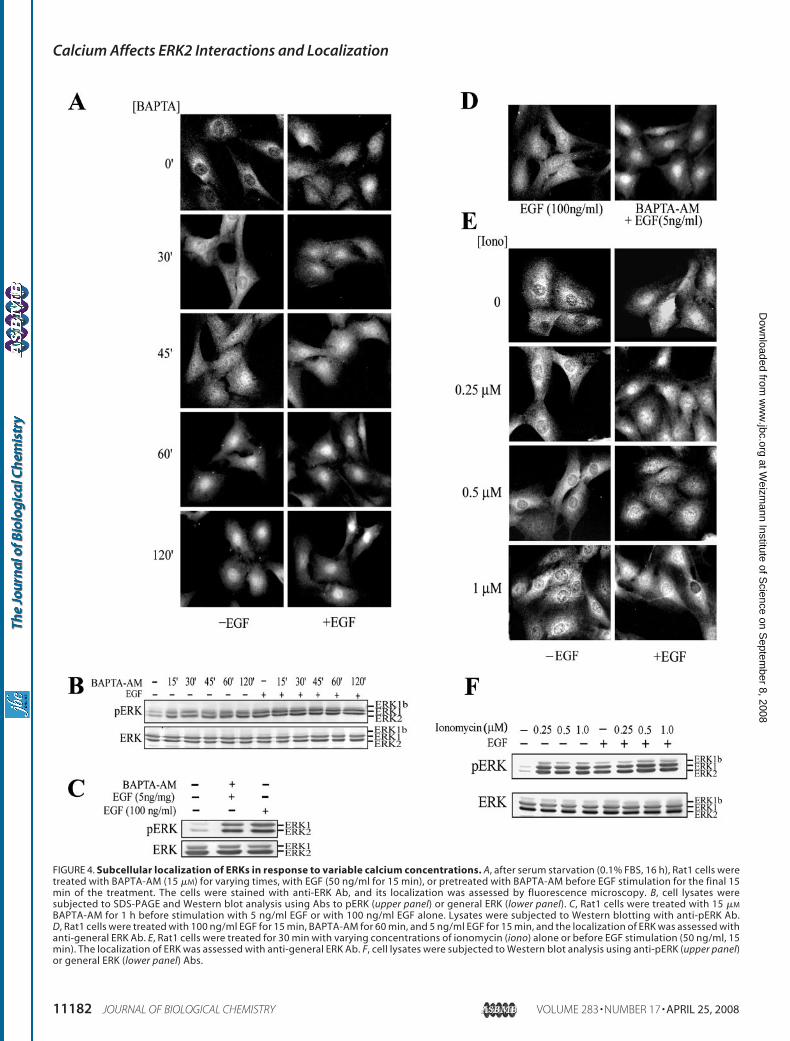
FIGURE 4. Subcellular localization of ERKs in response to variable calcium concentrations. A, after serum starvation (0.1% FBS, 16 h), Rat1 cells weretreated with BAPTA-AM (15 �M) for varying times, with EGF (50 ng/ml for 15 min), or pretreated with BAPTA-AM before EGF stimulation for the final 15min of the treatment. The cells were stained with anti-ERK Ab, and its localization was assessed by fluorescence microscopy. B, cell lysates weresubjected to SDS-PAGE and Western blot analysis using Abs to pERK (upper panel) or general ERK (lower panel). C, Rat1 cells were treated with 15 �M
BAPTA-AM for 1 h before stimulation with 5 ng/ml EGF or with 100 ng/ml EGF alone. Lysates were subjected to Western blotting with anti-pERK Ab.D, Rat1 cells were treated with 100 ng/ml EGF for 15 min, BAPTA-AM for 60 min, and 5 ng/ml EGF for 15 min, and the localization of ERK was assessed withanti-general ERK Ab. E, Rat1 cells were treated for 30 min with varying concentrations of ionomycin (iono) alone or before EGF stimulation (50 ng/ml, 15min). The localization of ERK was assessed with anti-general ERK Ab. F, cell lysates were subjected to Western blot analysis using anti-pERK (upper panel)or general ERK (lower panel) Abs.
Calcium Affects ERK2 Interactions and Localization
11182 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 283 • NUMBER 17 • APRIL 25, 2008
at Weizm
ann Institute of Science on S
eptember 8, 2008
ww
w.jbc.org
Dow
nloaded from
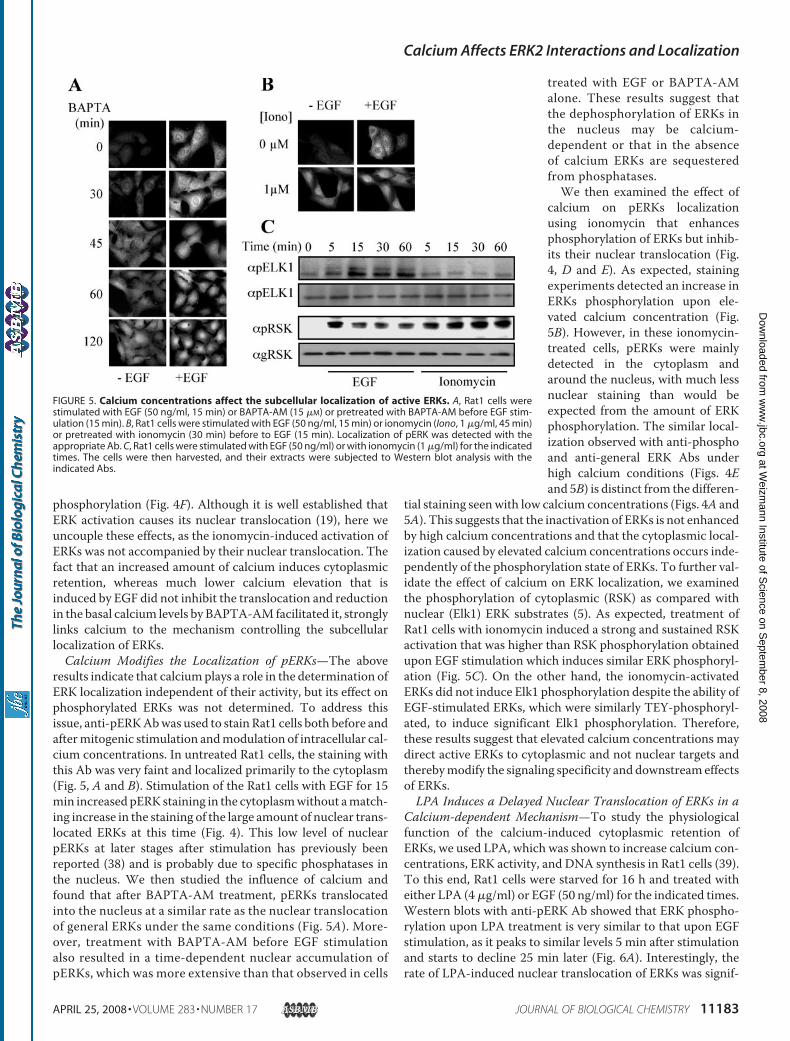
phosphorylation (Fig. 4F). Although it is well established thatERK activation causes its nuclear translocation (19), here weuncouple these effects, as the ionomycin-induced activation ofERKs was not accompanied by their nuclear translocation. Thefact that an increased amount of calcium induces cytoplasmicretention, whereas much lower calcium elevation that isinduced by EGF did not inhibit the translocation and reductionin the basal calcium levels by BAPTA-AM facilitated it, stronglylinks calcium to the mechanism controlling the subcellularlocalization of ERKs.Calcium Modifies the Localization of pERKs—The above
results indicate that calciumplays a role in the determination ofERK localization independent of their activity, but its effect onphosphorylated ERKs was not determined. To address thisissue, anti-pERKAbwas used to stain Rat1 cells both before andaftermitogenic stimulation andmodulation of intracellular cal-cium concentrations. In untreated Rat1 cells, the staining withthis Ab was very faint and localized primarily to the cytoplasm(Fig. 5, A and B). Stimulation of the Rat1 cells with EGF for 15min increased pERK staining in the cytoplasmwithout amatch-ing increase in the staining of the large amount of nuclear trans-located ERKs at this time (Fig. 4). This low level of nuclearpERKs at later stages after stimulation has previously beenreported (38) and is probably due to specific phosphatases inthe nucleus. We then studied the influence of calcium andfound that after BAPTA-AM treatment, pERKs translocatedinto the nucleus at a similar rate as the nuclear translocationof general ERKs under the same conditions (Fig. 5A). More-over, treatment with BAPTA-AM before EGF stimulationalso resulted in a time-dependent nuclear accumulation ofpERKs, which was more extensive than that observed in cells
treated with EGF or BAPTA-AMalone. These results suggest thatthe dephosphorylation of ERKs inthe nucleus may be calcium-dependent or that in the absenceof calcium ERKs are sequesteredfrom phosphatases.We then examined the effect of
calcium on pERKs localizationusing ionomycin that enhancesphosphorylation of ERKs but inhib-its their nuclear translocation (Fig.4, D and E). As expected, stainingexperiments detected an increase inERKs phosphorylation upon ele-vated calcium concentration (Fig.5B). However, in these ionomycin-treated cells, pERKs were mainlydetected in the cytoplasm andaround the nucleus, with much lessnuclear staining than would beexpected from the amount of ERKphosphorylation. The similar local-ization observed with anti-phosphoand anti-general ERK Abs underhigh calcium conditions (Figs. 4Eand 5B) is distinct from the differen-
tial staining seenwith low calcium concentrations (Figs. 4A and5A). This suggests that the inactivation of ERKs is not enhancedby high calcium concentrations and that the cytoplasmic local-ization caused by elevated calcium concentrations occurs inde-pendently of the phosphorylation state of ERKs. To further val-idate the effect of calcium on ERK localization, we examinedthe phosphorylation of cytoplasmic (RSK) as compared withnuclear (Elk1) ERK substrates (5). As expected, treatment ofRat1 cells with ionomycin induced a strong and sustained RSKactivation that was higher than RSK phosphorylation obtainedupon EGF stimulation which induces similar ERK phosphoryl-ation (Fig. 5C). On the other hand, the ionomycin-activatedERKs did not induce Elk1 phosphorylation despite the ability ofEGF-stimulated ERKs, which were similarly TEY-phosphoryl-ated, to induce significant Elk1 phosphorylation. Therefore,these results suggest that elevated calcium concentrations maydirect active ERKs to cytoplasmic and not nuclear targets andtherebymodify the signaling specificity and downstreameffectsof ERKs.LPA Induces a Delayed Nuclear Translocation of ERKs in a
Calcium-dependent Mechanism—To study the physiologicalfunction of the calcium-induced cytoplasmic retention ofERKs, we used LPA, which was shown to increase calcium con-centrations, ERK activity, and DNA synthesis in Rat1 cells (39).To this end, Rat1 cells were starved for 16 h and treated witheither LPA (4�g/ml) or EGF (50 ng/ml) for the indicated times.Western blots with anti-pERK Ab showed that ERK phospho-rylation upon LPA treatment is very similar to that upon EGFstimulation, as it peaks to similar levels 5 min after stimulationand starts to decline 25 min later (Fig. 6A). Interestingly, therate of LPA-induced nuclear translocation of ERKs was signif-
FIGURE 5. Calcium concentrations affect the subcellular localization of active ERKs. A, Rat1 cells werestimulated with EGF (50 ng/ml, 15 min) or BAPTA-AM (15 �M) or pretreated with BAPTA-AM before EGF stim-ulation (15 min). B, Rat1 cells were stimulated with EGF (50 ng/ml, 15 min) or ionomycin (Iono, 1 �g/ml, 45 min)or pretreated with ionomycin (30 min) before to EGF (15 min). Localization of pERK was detected with theappropriate Ab. C, Rat1 cells were stimulated with EGF (50 ng/ml) or with ionomycin (1 �g/ml) for the indicatedtimes. The cells were then harvested, and their extracts were subjected to Western blot analysis with theindicated Abs.
Calcium Affects ERK2 Interactions and Localization
APRIL 25, 2008 • VOLUME 283 • NUMBER 17 JOURNAL OF BIOLOGICAL CHEMISTRY 11183
at Weizm
ann Institute of Science on S
eptember 8, 2008
ww
w.jbc.org
Dow
nloaded from
icantly delayed as comparedwith that induced by EGF (Fig. 6B).The addition of BAPTA-AM to LPA before stimulation by LPAenhanced the rate of translocation, similarly to the rate inducedby EGF (Fig. 6C) withoutmodification of ERK activity (data notshown). Therefore, we suggest that elevated calcium concen-trations delay the rate of nuclear translocation and thereby
allow ERKs to first induce cytoplasmic processes and only later,their nuclear functions. These differences in localization caninduce distinct physiological functions without changes in therate or magnitude of ERKs phosphorylation or activity.Calcium Inhibits Nuclear Translocation of ERKs by Prevent-
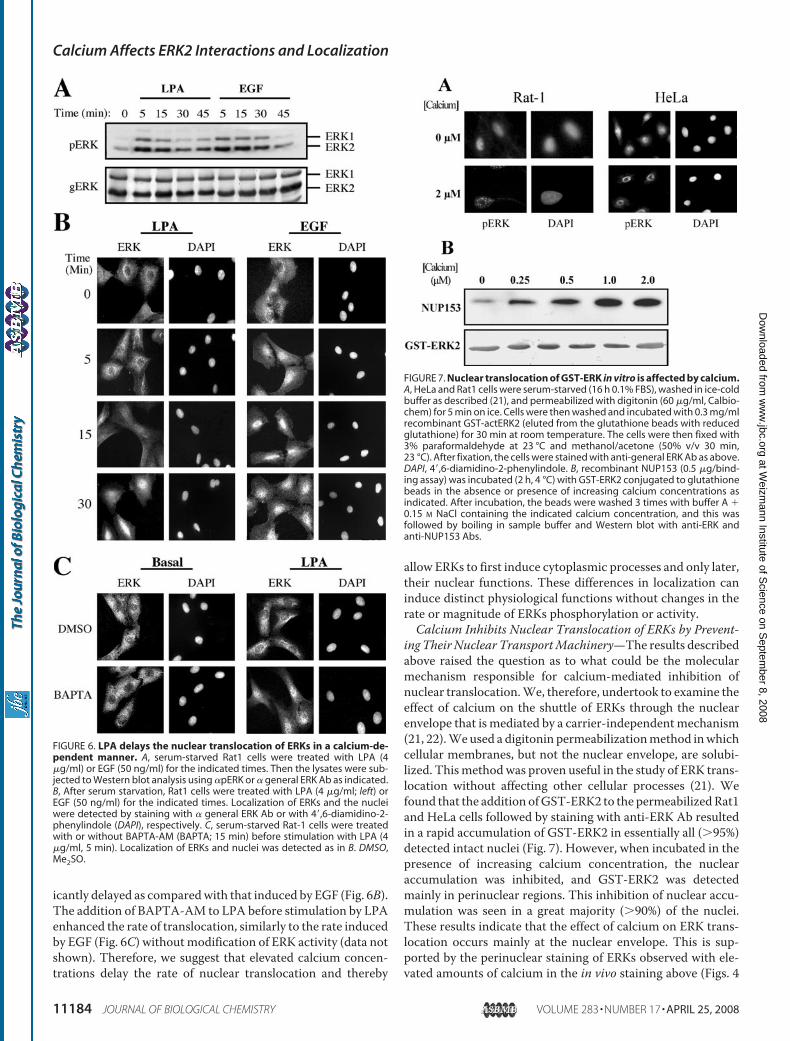
ing Their Nuclear TransportMachinery—The results describedabove raised the question as to what could be the molecularmechanism responsible for calcium-mediated inhibition ofnuclear translocation.We, therefore, undertook to examine theeffect of calcium on the shuttle of ERKs through the nuclearenvelope that is mediated by a carrier-independentmechanism(21, 22).Weused a digitonin permeabilizationmethod inwhichcellular membranes, but not the nuclear envelope, are solubi-lized. Thismethodwas proven useful in the study of ERK trans-location without affecting other cellular processes (21). Wefound that the addition ofGST-ERK2 to the permeabilizedRat1and HeLa cells followed by staining with anti-ERK Ab resultedin a rapid accumulation of GST-ERK2 in essentially all (�95%)detected intact nuclei (Fig. 7). However, when incubated in thepresence of increasing calcium concentration, the nuclearaccumulation was inhibited, and GST-ERK2 was detectedmainly in perinuclear regions. This inhibition of nuclear accu-mulation was seen in a great majority (�90%) of the nuclei.These results indicate that the effect of calcium on ERK trans-location occurs mainly at the nuclear envelope. This is sup-ported by the perinuclear staining of ERKs observed with ele-vated amounts of calcium in the in vivo staining above (Figs. 4
FIGURE 6. LPA delays the nuclear translocation of ERKs in a calcium-de-pendent manner. A, serum-starved Rat1 cells were treated with LPA (4�g/ml) or EGF (50 ng/ml) for the indicated times. Then the lysates were sub-jected to Western blot analysis using �pERK or � general ERK Ab as indicated.B, After serum starvation, Rat1 cells were treated with LPA (4 �g/ml; left) orEGF (50 ng/ml) for the indicated times. Localization of ERKs and the nucleiwere detected by staining with � general ERK Ab or with 4�,6-diamidino-2-phenylindole (DAPI), respectively. C, serum-starved Rat-1 cells were treatedwith or without BAPTA-AM (BAPTA; 15 min) before stimulation with LPA (4�g/ml, 5 min). Localization of ERKs and nuclei was detected as in B. DMSO,Me2SO.
FIGURE 7. Nuclear translocation of GST-ERK in vitro is affected by calcium.A, HeLa and Rat1 cells were serum-starved (16 h 0.1% FBS), washed in ice-coldbuffer as described (21), and permeabilized with digitonin (60 �g/ml, Calbio-chem) for 5 min on ice. Cells were then washed and incubated with 0.3 mg/mlrecombinant GST-actERK2 (eluted from the glutathione beads with reducedglutathione) for 30 min at room temperature. The cells were then fixed with3% paraformaldehyde at 23 °C and methanol/acetone (50% v/v 30 min,23 °C). After fixation, the cells were stained with anti-general ERK Ab as above.DAPI, 4�,6-diamidino-2-phenylindole. B, recombinant NUP153 (0.5 �g/bind-ing assay) was incubated (2 h, 4 °C) with GST-ERK2 conjugated to glutathionebeads in the absence or presence of increasing calcium concentrations asindicated. After incubation, the beads were washed 3 times with buffer A �0.15 M NaCl containing the indicated calcium concentration, and this wasfollowed by boiling in sample buffer and Western blot with anti-ERK andanti-NUP153 Abs.
Calcium Affects ERK2 Interactions and Localization
11184 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 283 • NUMBER 17 • APRIL 25, 2008
at Weizm
ann Institute of Science on S
eptember 8, 2008
ww
w.jbc.org
Dow
nloaded from

and 5). Thus, our results indicate that the inhibition of ERKtranslocation by elevated calcium concentration is mainlymediated by calcium-dependent inhibition of nuclear transportthrough the nuclear envelope.It was previously shown that ERKs translocate into the
nucleus through the nuclear pores at the nuclear envelope byinteraction with NUPs 153 and 214 (21, 22). Therefore, theaccumulation of ERKs at the nuclear envelope prompted us tostudy the effect of calcium on the interaction of active ERKswith NUPs. For this purpose we preformed an in vitro bindingassay between GST-actERK2 andHis-Nup153c in the presenceor absence of calcium. We found that the addition of calciumresulted in an increase in the interaction between the two pro-teins (Fig. 7C). Therefore, it is likely that under high calciumconcentration, the fully activated ERKs that are released fromtheir anchoring proteins do not translocate into the nucleusbecause of increasing irreversible interaction with NUPs. How-ever, because calciummay increase protein interactions of inac-tive and to some extent also active ERK2 with other proteins, wecannot exclude the possibility that the prevention of translocationby calcium is also mediated to some extent by increased cyto-plasmic retention by calcium-dependent anchoring proteins.
DISCUSSION
A plethora of extracellular stimuli are translated by cells intointracellular responses by a network of signaling pathways. Thisnetwork seems to regulate different and sometime even contra-dicting responses, raising the question of how the specificity ofthe signals is regulated. One way by which the specificity ofsignals can be secured is through protein-protein complexationand compartmentalization of special components of the net-work that bring about the right signals to their accurate desti-nations. Herewe show that indeed the interaction of ERK2withdifferent proteins may regulate its differential localization andfunction under distinct stimulation and cellular responses.Thus, we found that non-phosphorylated ERK2 interacts with alarge number of proteins through its CRS/CD domain. On theother hand, the doubly phosphorylated, active form of ERK2loses its binding properties and seems to interact with only asmall number of proteins that include its direct substrates.Most of the interactions identified are dependent on sufficientlevels of calcium, which seems to enhance mainly the multipleinteractions of the non-phosphorylated protein. Lower levels ofcalcium seem to weaken the binding of non-phosphorylatedERKs to many of the interacting proteins but may induce othertypes of interactions by active ERKs.Phosphorylation and activation of ERK2 as well as changes in
calcium concentrations not only modify the magnitude of pro-tein interaction to ERK2 but also affect the subcellular localiza-tion of the ERKs. Thus, we show here that an additional regu-lator of ERK translocation is varying calcium levels, whichwhenreduced, increase nuclear translocation, whereas elevated cal-cium levels prevent the translocation. The effects describedhere on the protein-protein interactions of ERK2 may explainthe regulation of the subcellular localization of ERKs accordingto the following model (Fig. 8); in resting cells, ERKs are local-ized in the cytoplasm due to calcium-dependent interactionswith a large number of proteins, which direct them to various
areas in the cytoplasm. Upon activation, ERKs detach frommost of their cytoplasmic anchoring proteins and are free tophosphorylate their substrates. The active ERK molecules arealso able at this stage to translocate into the nucleus where theyaccumulate by binding to a nuclear set of interacting proteins.Elevated calcium concentrations do not interfere with therelease of ERKs from the cytoplasmic anchoring proteins butseem to inhibit the penetration of the free pERK moleculesinto the nucleus, probably due to inhibition of the nucleartransport machinery. Consequently, active ERK moleculescan influence cytoplasmic processes for longer times butcannot induce short-term effects (e.g. transcription) in thenucleus. Finally, the reduction of calcium below physiologi-cal levels disrupts the calcium-dependent interactions ofERKs and allows non-phosphorylated ERKs to move intovarious cellular compartments where the ERKs interact witha different set of proteins in a calcium-independent manner.An important issue in this study was to determine the spec-
ificity of the interactions with ERK2. Several lines of evidencelead to the conclusion that most of the identified proteinsindeed bind directly to ERK2. Thus, we found the following. (a)Most of the proteins eluted from theGST-ERK2 column as freeproteins and not as aggregates, indicating that they were notbound through large protein complexes. (b) Some of the inter-actions identified (MEK1, vinculin, Elk1)were reproduced in anin vitro purified system in the absence of any other proteins. (c)No proteins were found to bind to either GST alone or even tothe GST-316A-ERK2, which is mutated in only three residueswithin the CRS/CD domain. (d) Many of the proteins exam-ined, including some that were reported as “sticky proteins”failed to bind to GST-ERK2. (e) Another method to determineprotein-protein interaction, the yeast two-hybrid system, alsorevealed a large number of ERK-interacting proteins (�100,data not shown). (f) It has previously been shown inmanyworksthat ERKs interact with substantial number of proteins includ-ing scaffolds, substrates, and others (5, 40). Taken together,these observations strongly indicate that most, if not all, of theidentified interactions are indeed through direct and specificbinding with ERK2.As mentioned above, several ERK-interacting proteins have
been identified over the past years, and therefore, it was impor-tant to analyze their interaction in the context of large numberof other interacting proteins. Thus, it was shown that the pro-tein interaction of ERKs is mediated by several regions includ-ingCRS/CD (7, 8), theDEFbinding domain (41), non-phospho-rylated activation loop residues (9), and others (for review, seeRef. 40). It was also shown that the interaction between ERKsand MEKs is reversed upon stimulation (19) and, thus, allowsnuclear translocation. Here we confirm these observations byshowing that the interaction of MEKs occurs only with non-phosphorylated ERK2 but not with the phosphorylated ERK2.We also show that the MEK-ERK interaction is dependent inpart on elevated calcium concentrations. Thus, our results sug-gest thatMEKs are not the only cytoplasmic anchoring proteinsof ERKs but rather belong to a group of CRS/CD interactorsthat keep ERKs in the cytoplasm.Homodimerization of active ERKmolecules was proposed to
participate in the mechanisms of translocation of ERKs (33).
Calcium Affects ERK2 Interactions and Localization
APRIL 25, 2008 • VOLUME 283 • NUMBER 17 JOURNAL OF BIOLOGICAL CHEMISTRY 11185
at Weizm
ann Institute of Science on S
eptember 8, 2008
ww
w.jbc.org
Dow
nloaded from

However, we found here that interaction between ERKs occursonly in their non-phosphorylated form, and calcium has only aminor effect on these interactions. These results together withthe fact that substitution of the homodimerization domain toalanines enhanced rather than slowed the nuclear translocationof ERKs (19) challenge the role of ERK dimerization in thenuclear translocation process. Another known interaction ofERKs is with the transcription factor Elk-1 (35). We found thatElk1 interaction occurs with unphosphorylated as well as phos-phorylated ERK2 but not with the phosphorylated formof Elk1.These results indicate that ERKs interact with Elk1 upon acti-vation, and the proteins are detached upon Elk1 phosphoryla-tion. In addition, we expected to find an interactionwithMAPKphosphatases (43) but foundMKP1 binding only after ionomy-cin treatment. The reason for the lack of MKPs interaction isprobably because they are only expressed upon stimulation(44). Thus, in the starved cells there were no MKPs, and theshort ionomycin treatmentwas sufficient to induce onlyMKP1.In contrast to most of the ERK-interacting proteins that
could bind to non-phosphorylated ERK2, there were only few
proteins that were found to interact under the conditionsexamined. One of these proteins is tubulin, which has previ-ously been shown to interact with a significant portion of ERKmolecules in many cells (15). This interaction was reported tobe only slightly reduced upon stimulation (15), and this is con-firmed here. Similar results were seen with actin, which wasalso reported to interactwith ERKs under some conditions (45).Therefore, tubulin, actin, and other cytoskeletal elements mayserve as cytoplasmic anchors of ERK2 even after stimulation,thus allowing some of the ERK molecules to execute essentialfunctions in the cytoskeleton. Such an anchoring effect could bemediated by vinculin, which we show here to strongly interactwith non-phosphorylated ERK2. Indeed, our data indicate thatthis protein, when overexpressed, can retain overexpressedERK2 in the cytoplasm (data not shown), supporting its role inthe regulation of ERKs localization. Finally, we found that pre-viously reported interactions with KSR (46), RSK (36), c-Fos(47), p38MAPK (48), vimentin (49), NUP153 (22), and thenewly discovered CaMKII and vinculin are increased by ele-vated calcium concentration. On the other hand, the interac-
FIGURE 8. Model representing ERKs localization upon stimulation and changes in calcium concentrations. Localization, phosphorylation, and interactionof ERKs are shown under the conditions studied in this work. For more details, see “Discussion.”
Calcium Affects ERK2 Interactions and Localization
11186 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 283 • NUMBER 17 • APRIL 25, 2008
at Weizm
ann Institute of Science on S
eptember 8, 2008
ww
w.jbc.org
Dow
nloaded from

tion with the nuclear hnRNP-K (50) seems to occur mainlywhen calcium is missing.The substantial effect of calcium on ERKs is likely to be of
high physiological significance. Here we show that calciumdelays the nuclear translocation of ERKs upon LPA treatmentin a calcium-dependent manner, suggesting that ERKs can firstexecute their cytoplasmic function and only at late stagesregulate nuclear processes. Indeed, we have shown that inthe presence of calcium, ERKs were able to activate theircytosolic substrate RSK for longer period of time, whereasthe phosphorylation of the nuclear substrate Elk1 was signifi-cantly reduced. This adds to the multiple effects that calciumcan play in activating the ERK cascade in various cell types (26).Another example that can be re-examined in light of this workis the pituitary, where calcium levels are increased dramaticallyupon gonadotropin-releasing hormone stimulation. Reducedcalcium concentrations only slightly decreased EGF or gonado-tropin-releasing hormone-induced activation of ERKs in thepituitary-derived �3T-1 cells (51), although calcium seems tobe required for the proper functioning of the ERK pathway inthese cells (52). We also found that in other pituitary-derivedcells termed L�T2, the translocation of ERKs into the nucleus isslow, starts only 30 min after activation, and is maximal about90 min after stimulation. These subcellular localizations corre-late with the function of ERKs in that initially (30 min) ERKsparticipate in the regulation of cytoplasmic exocitosis of LH orFSH (53), whereas ERK-regulated transcription can be detectedonly later (60–90 min (54). Thus, in these cells calcium regula-tion of ERK localization is important for proper outcome ofgonadotropin-releasing hormone signaling.The effect of calcium on the nuclear translocation of ERKs,
which is the main physiological response identified here, drewour attention to the molecular mechanism by which the pre-vention of nuclear translocation occurs. When addressing therole of calcium in nuclear translocation, it is important to notethat calcium may have a general role on the sliding of proteinsthrough nuclear pores (55, 56). The mechanism that mediatesthis effect on nuclear pores involves a release of calcium fromstores located in the cisternal space of ER that is adjacent to thenuclear envelope. This release of calcium induces a large con-formational change in the nuclear pores, which then affects freenuclear diffusion. Although this mechanism might be involvedin part in the prevention of ERKs translocation in our system, itcannot explain the effect on the stimulated, energy-dependenttranslocation observed here. Rather, this effect may be medi-ated by the effect of calcium on NUPs that has been previouslyshown as a crucial step in the translocation of ERKs into thenucleus (21, 22). Indeed, we found that ERK2 interact withNup153, and this interaction is regulated by elevated calciumconcentrations. It is, therefore, possible that high calcium con-centrations may affect both passive and active mechanisms ofERK translocation into the nucleus (42). Calcium-induced con-formational changes (55) may affect the diffusion of ERKs,which occurs mainly in resting cells, and increased associationof active ERKs with NUPs may explain the prevention of thestimulated translocation.In summary, we found that inactive ERK2 interacts with a
large number of proteins through its CRS/CD domain, whereas
the pERK2 interacts with only few proteins, including some ofits substrates. Changes in calcium concentrations affect proteininteraction of ERKs as well as their nuclear translocation. Wealso showed that translocation of ERKs induced by LPA isdelayed as compared with the translocation induced by EGF,and this delay is due to the elevated calcium concentrationsinduced by LPA. The prevention of translocation by high cal-cium probably occurs by inhibition of the transport of ERKsthrough the nuclear envelope. Together, these data are consist-ent with a model in which ERKs in resting cells are bound toseveral cytoplasmic proteins in a calcium-dependent manner.Upon stimulation, ERKs are phosphorylated and released fromthe cytoplasmic anchors to allow free shuttle within the cyto-plasm and toward the nucleus, which is inhibited by calcium.This is physiologically important because elevated calcium bysome stimuli, such as LPA, may change the localization andthereby the specificity of the ERK cascade. Thus, the subcellularlocalization of ERKs, which is influenced by calcium, and theCRS/CD-mediated interactions with many proteins appear tobe key regulators of the ERK cascade.
REFERENCES1. Imajo, M., Tsuchiya, Y., and Nishida, E. (2006) IUBMB Life 58, 312–3172. Avruch, J. (2007) Biochim. Biophys. Acta 1773, 1150–11603. Raman, M., Chen, W., and Cobb, M. H. (2007) Oncogene 26, 3100–31124. Seger, R., and Krebs, E. G. (1995) FASEB J. 9, 726–7355. Yoon, S., and Seger, R. (2006) Growth Factors 24, 21–446. Shaul, Y. D., and Seger, R. (2007)Biochim. Biophys. Acta 1773, 1213–12267. Rubinfeld, H., Hanoch, T., and Seger, R. (1999) J. Biol. Chem. 274,
30349–303528. Tanoue, T., and Nishida, E. (2002) Pharmacol. Ther. 93, 193–2029. Bendetz-Nezer, S., and Seger, R. (2007) J. Biol. Chem. 282, 25114–2512210. Bardwell, L. (2006) Biochem. Soc. Trans. 34, 837–84111. Lenormand, P., Brondello, J. M., Brunet, A., and Pouyssegur, J. (1998)
J. Cell Biol. 142, 625–63312. Jaaro, H., Rubinfeld, H., Hanoch, T., and Seger, R. (1997) Proc. Natl. Acad.
Sci. U. S. A. 94, 3742–374713. Fukuda,M., Gotoh, I., Gotoh, Y., andNishida, E. (1996) J. Biol. Chem. 271,
20024–2002814. Pouyssegur, J., Volmat, V., and Lenormand, P. (2002) Biochem. Pharma-
col. 64, 755–76315. Reszka, A. A., Seger, R., Diltz, C. D., Krebs, E. G., and Fischer, E. H. (1995)
Proc. Natl. Acad. Sci. U. S. A. 92, 8881–888516. Fukuda, M., Gotoh, Y., and Nishida, E. (1997) EMBO J. 16, 1901–190817. Blanco-Aparicio, C., Torres, J., and Pulido, R. (1999) J. Cell Biol. 147,
1129–113618. Morrison, D. K., and Davis, R. J. (2003)Annu. Rev. Cell Dev. Biol. 19, 91–11819. Wolf, I., Rubinfeld, H., Yoon, S., Marmor, G., Hanoch, T., and Seger, R.
(2001) J. Biol. Chem. 276, 24490–2449720. Ranganathan, A., Yazicioglu, M. N., and Cobb, M. H. (2006) J. Biol. Chem.
281, 15645–1565221. Matsubayashi, Y., Fukuda, M., and Nishida, E. (2001) J. Biol. Chem. 276,
41755–4176022. Whitehurst, A. W., Wilsbacher, J. L., You, Y., Luby-Phelps, K., Moore,
M. S., andCobb,M.H. (2002) Proc. Natl. Acad. Sci. U. S. A. 99, 7496–750123. Adachi, M., Fukuda, M., and Nishida, E. (2000) J. Cell Biol. 148, 849–85624. Berridge, M. J., Bootman, M. D., and Roderick, H. L. (2003)Nat. Rev. Mol.
Cell Biol. 4, 517–52925. Clapham, D. E. (2007) Cell 131, 1047–105826. Agell, N., Bachs, O., Rocamora, N., and Villalonga, P. (2002) Cell. Signal.
14, 649–65427. Cullen, P. J., and Lockyer, P. J. (2002) Nat. Rev. Mol. Cell Biol. 3, 339–34828. Xia, Z., Dudek, H., Miranti, C. K., and Greenberg, M. E. (1996) J. Neurosci.
16, 5425–5436
Calcium Affects ERK2 Interactions and Localization
APRIL 25, 2008 • VOLUME 283 • NUMBER 17 JOURNAL OF BIOLOGICAL CHEMISTRY 11187
at Weizm
ann Institute of Science on S
eptember 8, 2008
ww
w.jbc.org
Dow
nloaded from

29. Ginnan, R., and Singer, H. A. (2002) Am. J. Physiol. Cell Physiol. 282,754–761
30. Murasawa, S., Mori, Y., Nozawa, Y., Masaki, H., Maruyama, K., Tsutsumi,Y., Moriguchi, Y., Shibasaki, Y., Tanaka, Y., Iwasaka, T., Inada, M., andMatsubara, H. (1998) Hypertension 32, 668–675
31. Abraham, S. T., Benscoter,H.A., Schworer, C.M., and Singer,H.A. (1997)Circ. Res. 81, 575–584
32. Ebinu, J. O., Bottorff, D. A., Chan, E. Y., Stang, S. L., Dunn, R. J., and Stone,J. C. (1998) Science 280, 1082–1086
33. Khokhlatchev, A. V., Canagarajah, B.,Wilsbacher, J., Robinson,M., Atkin-son, M., Goldsmith, E., and Cobb, M. H. (1998) Cell 93, 605–615
34. Fukuda,M., Gotoh, I., Adachi,M., Gotoh, Y., andNishida, E. (1997) J. Biol.Chem. 272, 32642–32648
35. Rao, V. N., and Reddy, E. S. (1994) Oncogene 9, 1855–186036. Roux, P. P., Richards, S. A., and Blenis, J. (2003) Mol. Cell. Biol. 23,
4796–480437. Cook, S. J., Aziz, N., andMcMahon,M. (1999)Mol. Cell. Biol. 19, 330–34138. Yung, Y., Dolginov, Y., Yao, Z., Rubinfeld, H., Michael, D., Hanoch, T.,
Roubini, E., Lando, Z., Zharhary, D., and Seger, R. (1997) FEBS Lett. 408,292–296
39. Cook, S. J., and McCormick, F. (1996) Biochem. J. 320, 237–24540. Chuderland, D., and Seger, R. (2005)Mol. Biotechnol. 29, 57–7441. Lee, T., Hoofnagle, A. N., Kabuyama, Y., Stroud, J., Min, X., Goldsmith,
E. J., Chen, L., Resing, K. A., and Ahn, N. G. (2004)Mol. Cell 14, 43–5542. Adachi, M., Fukuda, M., and Nishida, E. (1999) EMBO J. 18, 5347–535843. Sun, H., Charles, C. H., Lau, L. F., and Tonks, N. K. (1993) Cell 75,
487–493
44. Camps, M., Chabert, C., Muda, M., Boschert, U., Gillieron, C., and Arkin-stall, S. (1998) FEBS Lett. 425, 271–276
45. Leinweber, B. D., Leavis, P. C., Grabarek, Z., Wang, C. L., and Morgan,K. G. (1999) Biochem. J. 1, 117–123
46. Yu, W., Fantl, W. J., Harrowe, G., andWilliams, L. T. (1998) Curr. Biol. 8,56–64
47. Murphy, L. O., Smith, S., Chen, R. H., Fingar, D. C., and Blenis, J. (2002)Nat. Cell Biol. 4, 556–564
48. Zhang, H., Shi, X., Hampong, M., Blanis, L., and Pelech, S. (2001) J. Biol.Chem. 276, 6905–6908
49. Perlson, E., Hanz, S., Ben-Yaakov, K., Segal-Ruder, Y., Seger, R., andFainzilber, M. (2005) Neuron 45, 715–726
50. Habelhah, H., Shah, K., Huang, L., Ostareck-Lederer, A., Burlingame,A. L., Shokat, K. M., Hentze, M.W., and Ronai, Z. (2001)Nat. Cell Biol. 3,325–330
51. Reiss, N., Levi, L. N., Shacham, S., Harris, D., Seger, R., andNaor, Z. (1997)Endocrinology 138, 1673–1682
52. Mulvaney, J. M., Zhang, T., Fewtrell, C., and Roberson,M. S. (1999) J. Biol.Chem. 274, 29796–29804
53. Naor, Z., Benard, O., and Seger, R. (2000) Trends Endocrinol. Metab. 11,91–99
54. Harris, D., Chuderland, D., Bonfil, D., Kraus, S., Seger, R., and Naor, Z.(2003) Endocrinology 144, 612–622
55. Lee, M. A., Dunn, R. C., Clapham, D. E., and Stehno-Bittel, L. (1998) CellCalcium 23, 91–101
56. Erickson, E. S.,Mooren, O. L.,Moore, D., Krogmeier, J. R., andDunn, R. C.(2006) Can. J. Physiol. Pharmacol. 84, 309–318
Calcium Affects ERK2 Interactions and Localization
11188 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 283 • NUMBER 17 • APRIL 25, 2008
at Weizm
ann Institute of Science on S
eptember 8, 2008
ww
w.jbc.org
Dow
nloaded from




















