![Complex Formation of the Antiviral 9-[2-(Phosphonomethoxy)Ethyl]Adenine (PMEA) and of Its N 1, N 3, and N 7 Deaza Derivatives with Copper(II) in Aqueous Solution](https://static.fdokumen.com/doc/165x107/63436ec4f4cbfd5a7c05176a/complex-formation-of-the-antiviral-9-2-phosphonomethoxyethyladenine-pmea-and.jpg)
Calcium-Dependent NOX5 Nicotinamide Adenine Dinucleotide Phosphate Oxidase Contributes to Vascular...
12
Calcium dependent Nox5 NADPH oxidase contributes to vascular oxidative stress in human coronary artery disease Tomasz J Guzik, MD PhD 1,2 , Wei Chen, MD PhD 1 , Maria C. Gongora, MD 1 , Bartlomiej Guzik, MD PhD 2 , Heinrich E. Lob, PhD 1 , Deepa Mangalat, MD 1 , Nyssa Hoch, PhD 1 , Sergey Dikalov, PhD 1 , Pawel Rudzinski, MD PhD 2 , Boguslaw Kapelak, MD PhD 2 , Jerzy Sadowski, MD PhD 3 , and David G Harrison, MD, FACC 1 1 Division of Cardiology, Department of Medicine, Emory University School of Medicine, Atlanta, GA, USA 2 Departments of Pharmacology and Internal Medicine, Jagiellonian University School of Medicine, Cracow, Poland 3 Cardiovascular Surgery and Transplantation, Jagiellonian University School of Medicine, Cracow, Poland Abstract Objectives—To examine the expression and activity of the calcium dependent NADPH oxidase in human atherosclerotic coronary arteries. Background—The Nox based NADPH oxidases are major sources of reactive oxygen species (ROS) in human vessels. Several Nox homologs have been identified but their relative contribution to vascular ROS production in coronary artery disease (CAD) is unclear. Nox5 is a unique homolog in that it is calcium dependent and thus could be activated by vasoconstrictor hormones. Its presence has not yet been studied in human vessels. Methods—Coronary arteries from patients undergoing cardiac transplant with CAD or without CAD were studied. Nox5 was quantified and visualized using Western blotting, immunofluorescence and quantitative real-time PCR. Calcium dependent NADPH oxidase activity, corresponding greatly to Nox5 activity was measured by electron paramagnetic resonance. Results—Both western blotting and quantitative real time PCR indicated a marked increase in Nox5 protein and mRNA in CAD vs non CAD vessels. Calcium dependent NADPH driven production of reactive oxygen species in vascular membranes, reflecting Nox5 activity was increased 7 fold in CAD and correlated significantly with Nox5 mRNA levels among subjects. Immunofluorescence demonstrated that Nox5 was expressed in the endothelium in the early lesions and in vascular smooth muscle cells in the advanced in coronary lesions. Conclusions—These studies identify Nox5 as a novel, calcium dependent source of reactive oxygen species in atherosclerosis. Address for correspondence: David G Harrison MD FACC, Tomasz J Guzik MD PhD, 1639 Pierce Drive, Room 319 WMRB, Emory University Cardiology Division, Atlanta, GA 30322, Phone 404-727-3710, Fax 404 727-3585, e-mail: [email protected], tguzik@cm- uj.krakow.pl. There are no conflicts of interest to declare. Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain. NIH Public Access Author Manuscript J Am Coll Cardiol. Author manuscript; available in PMC 2009 November 25. Published in final edited form as: J Am Coll Cardiol. 2008 November 25; 52(22): 1803–1809. doi:10.1016/j.jacc.2008.07.063. NIH-PA Author Manuscript NIH-PA Author Manuscript NIH-PA Author Manuscript
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Calcium-Dependent NOX5 Nicotinamide Adenine Dinucleotide Phosphate Oxidase Contributes to Vascular...

Calcium dependent Nox5 NADPH oxidase contributes to vascularoxidative stress in human coronary artery disease
Tomasz J Guzik, MD PhD1,2, Wei Chen, MD PhD1, Maria C. Gongora, MD1, Bartlomiej Guzik,MD PhD2, Heinrich E. Lob, PhD1, Deepa Mangalat, MD1, Nyssa Hoch, PhD1, Sergey Dikalov,PhD1, Pawel Rudzinski, MD PhD2, Boguslaw Kapelak, MD PhD2, Jerzy Sadowski, MDPhD3, and David G Harrison, MD, FACC1
1 Division of Cardiology, Department of Medicine, Emory University School of Medicine, Atlanta, GA, USA2 Departments of Pharmacology and Internal Medicine, Jagiellonian University School of Medicine, Cracow,Poland 3 Cardiovascular Surgery and Transplantation, Jagiellonian University School of Medicine, Cracow,Poland
AbstractObjectives—To examine the expression and activity of the calcium dependent NADPH oxidase inhuman atherosclerotic coronary arteries.
Background—The Nox based NADPH oxidases are major sources of reactive oxygen species(ROS) in human vessels. Several Nox homologs have been identified but their relative contributionto vascular ROS production in coronary artery disease (CAD) is unclear. Nox5 is a unique homologin that it is calcium dependent and thus could be activated by vasoconstrictor hormones. Its presencehas not yet been studied in human vessels.
Methods—Coronary arteries from patients undergoing cardiac transplant with CAD or withoutCAD were studied. Nox5 was quantified and visualized using Western blotting, immunofluorescenceand quantitative real-time PCR. Calcium dependent NADPH oxidase activity, corresponding greatlyto Nox5 activity was measured by electron paramagnetic resonance.
Results—Both western blotting and quantitative real time PCR indicated a marked increase in Nox5protein and mRNA in CAD vs non CAD vessels. Calcium dependent NADPH driven production ofreactive oxygen species in vascular membranes, reflecting Nox5 activity was increased 7 fold inCAD and correlated significantly with Nox5 mRNA levels among subjects. Immunofluorescencedemonstrated that Nox5 was expressed in the endothelium in the early lesions and in vascular smoothmuscle cells in the advanced in coronary lesions.
Conclusions—These studies identify Nox5 as a novel, calcium dependent source of reactiveoxygen species in atherosclerosis.
Address for correspondence: David G Harrison MD FACC, Tomasz J Guzik MD PhD, 1639 Pierce Drive, Room 319 WMRB, EmoryUniversity Cardiology Division, Atlanta, GA 30322, Phone 404-727-3710, Fax 404 727-3585, e-mail: [email protected], [email protected] are no conflicts of interest to declare.Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customerswe are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resultingproof before it is published in its final citable form. Please note that during the production process errors may be discovered which couldaffect the content, and all legal disclaimers that apply to the journal pertain.
NIH Public AccessAuthor ManuscriptJ Am Coll Cardiol. Author manuscript; available in PMC 2009 November 25.
Published in final edited form as:J Am Coll Cardiol. 2008 November 25; 52(22): 1803–1809. doi:10.1016/j.jacc.2008.07.063.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Keywordsreactive oxygen species; nox5; NADPH oxidase; atherosclerosis; coronary artery disease
IntroductionAtherosclerosis and its risk factors increase vascular production of reactive oxygen species(ROS), which in turn contribute to many aspects of atherosclerotic lesion formation (1). Amajor source of ROS in atherosclerosis is the NADPH oxidases (2). The catalytic subunits ofthe NADPH oxidases are the Nox proteins, which are involved in transfer of electrons fromNADPH to heme groups and ultimately molecular oxygen to form superoxide (O2
•−) (2). TheNox based NADPH oxidases are major sources of ROS in human vasculature. There are varioushomologs of Nox proteins, and their expression in atherosclerotic lesions varies depending onlocation and stages of development (3,4). Particularly, the role of Nox2 and Nox4 and to thelesser extent Nox1 have been described in the context of human atherosclerosis and coronaryartery disease (3–5). The last discovered of these is Nox5, which is unique because it containsEF hand domains in the N-terminal region that bind calcium and permit activation of theenzyme by increases in intracellular calcium ([Ca]i) (6). Nox5 has been identified in immunetissues, testes, cancer cells (6) and recently in human endothelial cells (7). Although Nox5 hasbeen identified in endothelium, it’s activity and contribution to ROS production in nativeendothelial cells or human blood vessels remains unclear as all experiments were conductedon cells transfected with exogenous Nox5 (7). Thus the potential role of calcium-dependentreactive oxygen species (ROS) production in human coronary artery disease remains unknown.Interestingly, it has been reported that calcium channel antagonists reduce ROS production inhuman endothelial cells (8). These drugs have been demonstrated to be clinically beneficial inimproving clinical outcomes in patients with vascular diseases, particularly coronary arterydisease. While there are other potential calcium-dependent sources of ROS in endothelial cells,given a key role of NADPH oxidases, it is possible that Nox5 may contribute to oxidative stressin the setting of vascular disease. The aim of the present study was therefore to determine ifexpression and function of Nox5 is increased in human atherosclerotic coronary arteriescompared to non-atherosclerotic vessels.
MethodsPatients and Blood Vessels
Segments of human coronary arteries were obtained from explanted hearts of 26 patientsundergoing heart transplantation for end stage heart failure. Collection of tissues wasperformed as previously described (3). The Ethics Committee at the University of Cracowapproved collection of tissues. Informed consent was obtained from each subject.
Calcium dependent NADPH oxidase activityCalcium-dependent NADPH oxidase activity was measured from membrane fractions ofhuman coronary arteries using electron paramagnetic spin resonance by measuring productionof reactive oxygen species as described previously (9) upon stimulation with 100uM NADPH;in the presence and absence of 1mM Ca++. Membrane fractions were prepared by a 2-hourultracentrifugation of vascular homogenates at 200,000 g. Twenty μg of membrane fractionswere suspended in calcium free media A with 1 mmol/L AAP, 5 U/ml HRP, 50 U/ml Cu/Zn-SOD, and 1mmol/L PPH. The reaction mixture placed in 100 μl ESR capillary tube with 100□M NADPH and accumulation of the PP· nitroxide was measured using a Bruker EMX ESRspectrometer and a super-high Q microwave cavity as previously described (9). Allmeasurements were performed at 25°C using a Bruker Nitrogen Temperature Controller
Guzik et al. Page 2
J Am Coll Cardiol. Author manuscript; available in PMC 2009 November 25.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

system with the following ESR settings: microwave frequency 9.46 GHz, modulationamplitude 2 G, microwave power 10 dB, conversion time 1.3 s, time constant 5.2 s.Measurements were performed in both Ca++ free media and also in media A containing 1 mMCa++. The NADPH-driven ROS production in the presence of Ca++ was used to determine totalNADPH oxidase activity and the difference between total and Ca++ independent oxidaseactivity was calculated as Ca++ dependent NADPH oxidase activity.
Detection of Nox5 proteinTwenty □g of total protein from vascular homogenates was separated by 12% SDS-PAGE andtransferred to nitrocellulose membranes. Nox5 protein was detected using a rabbit polyclonalantibody raised against the Nox5 second extracellular loop peptideEASPFQFWELLLTTRPGIG. This is common to all Nox5 splice variants. Bands weredetected using chemiluminescence as previously described and analyzed using NIH Imagesoftware.
For detection of Nox5 protein in histological sections, immunofluorescence was performed onfrozen 7-μm OCT-embedded tissue sections as described previously. Nox5 was visualizedusing the rabbit polyclonal anti-Nox5 (1:100 dilution). Endothelial cells were identified usinga mouse monoclonal anti-CD31 (1:200 dilution). Appropriate secondary antibodies wereemployed. Sections treated with secondary antibodies alone did not show specific staining.Staining was visualized on a confocal microscope.
Detection of Nox5 mRNA by quantitative real-time RT-PCRFor detection of Nox5 mRNA, segments of coronary arteries were snap-frozen in Tri-reagentand total RNA isolated using the RNA easy kit with DNAse digestion (4). cDNA wassynthesized using High Capacity cDNA Reverse Transcription kit (Applied Biosystems) andwas subjected to quantitative PCR using the TaqMan ddCT method (Applied Biosystems) andABI 7500 Fast Real Time PCR system (Applied Biosystems). All primers and probes (Nox5:Hs00225846_m1 gene expression assay as well as an 18S endogenous control) were fromApplied Biosystems.
Suppression of Nox5 using small interfering RNANox5 siRNA OnTargetPlus™ (Dharmacon, US) and OnTarget Plus™siControl was pre-incubated in Eppendorf-tubes containing 0.572 ml Optimem and 24-μl Oligofectamine persample. siRNA was added at a final volume of 3 ml to pre-confluent human aortic endothelialcells (HAEC; Cambrex, MA) (final concentration of siRNA- 100 nmol/L). After 4–6 hours ofincubation at 37°C and 5% CO2 6 ml EBM2 media was added. This procedure was repeatedafter 48 hours.
Statistical analysisAll data are expressed as means ± SEM with n equal to the number of patients. Comparisonsbetween groups of patients or treatments were made using student t-test or Mann-Whitney Utest. Correlation between oxidase activity and Nox5 expression was assessed by Spearmancorrelation coefficient. Values of p < 0.05 were considered statistically significant.
ResultsCharacteristics of patients studied
Subjects included in this study had end-stage heart failure and were undergoing hearttransplantation. Fourteen individuals had coronary artery disease (CAD) and twelve patientshad either dilated (n=11) or hypertrophic cardiomyopathy (n=1), but did not have CAD. The
Guzik et al. Page 3
J Am Coll Cardiol. Author manuscript; available in PMC 2009 November 25.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

diagnosis of CAD was based on either a history of MI or on coronary angiography. As expected,demographic and clinical characteristics indicated that CAD patients had more risk factors foratherosclerosis than non-CAD patients and were more likely taking statins. Similarly, thepresence of CAD was associated with a greater incidence of prior MI, transient ischemic attack(TIA) and peripheral arterial disease, and typical CAD. The degree of left ventriculardysfunction, as reflected by the ejection fraction, was similar between the two groups (Table1).
Calcium-dependent NADPH oxidase activity in human endothelial cells and coronary arteriesAs an initial attempt to quantify Nox5 activity and protein levels in vascular segments, westudied human endothelial cells to validate our assay and antibody. Western blots using therabbit polyclonal antibody showed a single band at a molecular weight of ca. 70 kDa, in keepingwith the reported molecular the □ isoform of Nox5. Importantly, siRNA against Nox5 reducedthis band by 75%, while scrambled siRNA had no significant effect. Using ESR, we detectedNADPH-driven ROS production in membranes of human endothelial cells that was greater inthe presence of calcium than in its absence. The difference between these two, i.e. the calcium-dependent NADPH oxidase activity, was reduced by siRNA against Nox5. These data stronglysuggest that studies of membranes from human endothelial cells in the presence and absenceof calcium can reflect Nox5 activity. We recognize that calcium can activate protein kinase C,which can contribute to activation of Nox2 and perhaps other Nox proteins, by phosphorylationof p47phox (ref). In addition, calcium can serve to activate ROS from xanthine oxidase and themitochondria (ref), however these events require the presence of cytoplasmic proteins that wereabsent in this assay. The fact that siNox5 reduced the calcium-dependent NADPH oxidaseactivity to approximately the same degree as it reduced Nox5 protein levels suggests that Nox5is the source of ROS under the conditions of this assay.
As observed in endothelial cells, membrane preparations of all of studied coronary arteriesdemonstrated NADPH-driven ROS production both in the absence and in the presence ofcalcium. ROS production in the absence of calcium, reflecting the activity of Nox1, Nox2 andNox4 was significantly greater in membranes from CAD than in non-CAD subjects (Figure2A). Interestingly, the increase in calcium-dependent NADPH oxidase activity wassignificantly more pronounced, averaging 7–8 fold more in vascular membranes from CADsubjects compared to non-CAD subjects (Figure 2A; right panel).
Nox5 mRNA expression and correlation with calcium-dependent NADPH oxidase activityReal time PCR demonstrated the presence of Nox5 mRNA in all vascular segments, howeverits levels were much greater in homogenates of vessels from CAD as compared to non-CADsubjects (Figure 2B). Importantly there was a very significant relationship between these levelsof Nox5 mRNA and the calcium-dependent NADPH oxidase activity (Figure 2C). Thesefindings strongly suggest that, as in the case of cultured endothelial cells, in human coronaryarteries, Nox5 contributes to calcium-dependent NADPH oxidase activity.
Detection of Nox5 protein in human coronary arteriesTo quantify Nox5 protein, we performed Western blots on homogenates of coronary arteriesfrom CAD and non-CAD patients. As in cultured human endothelial cells, western blottingdemonstrated a single band at a molecular weight of 70 kDa, compatible with the beta isoformof Nox5, identical size to the band observed in endothelial cells and DU145 cells (Figure 3A).More importantly, this was increased by four fold in CAD compared to non-CAD segments.
To confirm this and to attempt to localize Nox5 in atherosclerotic lesions, we performedimmunofluorescence staining utilizing the antibody we developed. These studies corroboratedthat Nox5 in expression was very low in coronary arteries from subjects without CAD (Figure
Guzik et al. Page 4
J Am Coll Cardiol. Author manuscript; available in PMC 2009 November 25.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

4A), but that CAD vessels demonstrated specific staining. In the vessels from CAD subjectsbut in the absence of plaque, Nox 5 expression seemed most prominent in endothelial cells, asevidenced by co-staining with anti-CD31 (Figure 4B and C). As more progressed lesions, withmoderate neointima, Nox5 staining was most striking in the neointima although punctatestaining could also be observed in the inner aspects of the media (Figure 4D). The most complexlesions demonstrated intense co-staining of Nox5 with smooth muscle underlying advancedplaques (Figure 4E). In no instance did we observe co-localization of Nox5 with T cell markerCD3 or macrophage marker CD68 (data not shown).
DiscussionIn this report, we show that Nox5 protein and mRNA expression are markedly increased invessels of patients with CAD compared to non-CAD subjects. This is associated with increasedcalcium-dependent membrane NADPH oxidase activity, characteristic of Nox5. Indeed, wefind that the level of calcium-dependent NADPH oxidase activity correlates significantly withNox5 mRNA when compared between individual patients. Our immunostaining confirmed theincrease in Nox5 protein and shows that its presence seems to vary depending on the stage ofatherosclerosis.
Our study is the first to describe the presence of calcium dependent NADPH oxidase activityin human vasculature. We employed an electron spin resonance-based assay to detect calcium-dependent NADPH oxidase activity based on a modification of recently publishedmethodology (9). The conditions of this assay allow detection of hydrogen peroxide by takingadvantage of the formation of compound II from horseradish peroxidase, which in turn reactswith AAP, forming a radical that reacts with the nitroxide spin probe. In numerous preliminaryexperiments, we were unable to detect calcium-dependent superoxide formation frommembrane fractions of endothelial cells or human coronary arteries using ESR. This wassurprising, because Nox5, like other Nox proteins, contains heme groups that should performa one-electron reduction of oxygen, and should therefore form superoxide. It is possiblehowever, that Nox5 shares similarity with Nox4, which seems to predominantly releasehydrogen peroxide (10). This might be due to positive charges near the heme groups, whichretain the negatively charged superoxide until it undergoes spontaneous dismutation tohydrogen peroxide. It is also likely that the membranes we prepared contained extracellularsuperoxide dismutase, which is known to be present in large amounts in the vasculature (11).This enzyme binds to components of the extracellular matrix, which was almost certainlyincluded in our membrane preparations and could rapidly convert superoxide to hydrogenperoxide, preventing detection of the former. Prior reports have reported that Nox5 producesboth hydrogen peroxide and superoxide using fluorescent methods, however these studies usedoverexpression of the enzyme and did not quantitate the relative amounts of hydrogen peroxidevs. superoxide formed (7,12). Hydrogen peroxide plays an important role in cell signaling andalso contributes to atherosclerosis, as transgenic mice that overexpress catalase are protectedfrom lesion development (13). Thus, the formation of hydrogen peroxide by Nox5 could haveimportant implications for the genesis of vascular disease.
Our present observations have implications for mechanisms underlying alterations of vascularfunction in CAD. Vasoactive agonists increase [Ca]i both in the endothelial and vascularsmooth muscle cells. In endothelial cells, this promotes release of nitric oxide (NO), while invascular smooth muscle cells, increases in intracellular calcium trigger the contractileapparatus (14). The induction of Nox5 in the endothelium could negate release of NO from theendothelium by permitting the simultaneous production of superoxide and hydrogen peroxide.The former reacts with NO at a diffusion-limited rate, leading to loss of NO and formation ofperoxynitrite (15) and these mechanisms are enhanced in human atherosclerosis (16).Hydrogen peroxide can also participate in consumption of NO via reactions with peroxidases
Guzik et al. Page 5
J Am Coll Cardiol. Author manuscript; available in PMC 2009 November 25.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

(17). The induction of Nox5 in the vascular smooth muscle could also lead to loss of NO as itdiffuses into these cells. Moreover, reactive oxygen species such as peroxynitrite can alterfunction of the sarcoplasmic reticulum, leading to increases in vascular smooth muscle [Ca]i(18). Moreover, recent studies in Drosophila have shown that the fruit-fly homolog of Nox5can contribute to smooth muscle cell contraction in response to calcium flux (19). Together,these events would promote vasoconstriction and could alter vascular responses to vasoactivehormones.
The localization of Nox5 is of interest in human vessels. In early lesions, Nox5 seemed in manycases to co-localize with endothelial cells. It is conceivable that the cytokine milieu present inearly atherosclerosis could promote Nox5 expression in endothelial cells. Studies of cytokineregulation of Nox5 in endothelial cells would be informative in this regard. In moderatelyadvanced lesions, endothelial staining was less evident, however a large amount of Nox5 co-localized with vascular smooth muscle cells in sub-intimal regions. Complex regions showextensive Nox5 staining in the area of plaque. The presence of Nox5 in early lesions and itsloss in advanced lesions is reminiscent of the expression of the endothelial nitric oxide synthasein these settings, as it is present in early lesions and is lost in endothelial cells overlyingadvanced plaques (20).
In prior studies, xanthine oxidase, Nox4 and Nox2 were found to be expressed in coronaryarteries with CAD (3). In the conditions of our assays of isolated vascular membranes, it isunlikely that these enzymes would be activated by addition of calcium. In keeping with this,siRNA against Nox5 markedly inhibited calcium-dependent NADPH oxidase activity, whilecalcium-independent activity remained unchanged. As evident from figure 2A, the calcium-dependent activity was similar to the calcium-independent activity in CAD vessels, while innon-CAD membranes its contribution was much less. This is in keeping with Nox5 inductionin CAD. Thus, calcium-independent NADPH oxidase activity is increased approximately 2.5-fold, while the increase in calcium-dependent activity is 7-fold. This would indicate that theatherosclerotic milieu provides a potent stimulus for Nox5.
Our findings could have implications for therapy in the setting of CAD. Calcium channelantagonists are commonly employed in the treatment of angina and coronary artery diseaseand in general have been associated with improved symptoms and outcome (21). One of theirbeneficial effects would be to reduce activation of Nox5 in cells harboring L-type calciumchannels, including vascular smooth muscle cells in lesions. Other commonly employedvasodilators, including nitrovasodilators and phosphodiesterase inhibitors also reduceintracellular calcium. In this fashion, these agents might reduce activation of Nox5 in diseasedvessels and thus prevent oxidant injury.
AcknowledgementsThis study was supported by NIH HL390006 grant.
TJG is supported by the Polish Ministry for Higher Education.
References1. Glass CK, Witztum JL. Atherosclerosis the road ahead. Cell 2001;104:503–16. [PubMed: 11239408]2. Guzik TJ, Harrison DG. Vascular NADPH oxidases as drug targets for novel antioxidant strategies.
Drug Discov Today 2006;11:524–33. [PubMed: 16713904]3. Guzik TJ, Sadowski J, Guzik B, et al. Coronary artery superoxide production and nox isoform
expression in human coronary artery disease. Arterioscler Thromb Vasc Biol 2006;26:333–9.[PubMed: 16293794]
Guzik et al. Page 6
J Am Coll Cardiol. Author manuscript; available in PMC 2009 November 25.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

4. Sorescu D, Weiss D, Lassegue B, et al. Superoxide production and expression of nox family proteinsin human atherosclerosis. Circulation 2002;105:1429–35. [PubMed: 11914250]
5. Guzik T, Sadowski J, Kapelak B, et al. Systemic regulation of vascular NAD(P)H oxidase activity andnox isoform expression in human arteries and veins. Arterioscler Thromb Vasc Biol 2004;24:1614–20. [PubMed: 15256399]
6. Banfi B, Molnar G, Maturana A, et al. A Ca(2+)-activated NADPH oxidase in testis, spleen, and lymphnodes. J Biol Chem 2001;276:37594–601. [PubMed: 11483596]
7. BelAiba RS, Djordjevic T, Petry A, et al. NOX5 variants are functionally active in endothelial cells.Free Radic Biol Med 2007;42:446–59. [PubMed: 17275676]
8. Lob H, Rosenkranz AC, Breitenbach T, Berkels R, Drummond G, Roesen R. Antioxidant and nitricoxide-sparing actions of dihydropyridines and ACE inhibitors differ in human endothelial cells.Pharmacology 2006;76:8–18. [PubMed: 16220025]
9. Doughan AK, Harrison DG, Dikalov SI. Molecular Mechanisms of Angiotensin II MediatedMitochondrial Dysfunction. Linking Mitochondrial Oxidative Damage and Vascular EndothelialDysfunction. Circ Res. 2008in press
10. Martyn KD, Frederick LM, von Loehneysen K, Dinauer MC, Knaus UG. Functional analysis of Nox4reveals unique characteristics compared to other NADPH oxidases. Cell Signal 2006;18:69–82.[PubMed: 15927447]
11. Landmesser U, Spiekermann S, Dikalov S, et al. Vascular oxidative stress and endothelial dysfunctionin patients with chronic heart failure: role of xanthine-oxidase and extracellular superoxide dismutase.Circulation 2002;106:3073–8. [PubMed: 12473554]
12. Jagnandan D, Church JE, Banfi B, Stuehr DJ, Marrero MB, Fulton DJ. Novel mechanism of activationof NADPH oxidase 5. calcium sensitization via phosphorylation. J Biol Chem 2007;282:6494–507.[PubMed: 17164239]
13. Yang H, Roberts LJ, Shi MJ, et al. Retardation of atherosclerosis by overexpression of catalase orboth Cu/Zn-superoxide dismutase and catalase in mice lacking apolipoprotein E. Circ Res2004;95:1075–81. [PubMed: 15528470]
14. Forstermann U, Gath I, Schwarz P, Closs EI, Kleinert H. Isoforms of nitric oxide synthase - properties,cellular distribution and expressional control [review]. Biochem Pharm 1995;50:1321–1332.[PubMed: 7503779]
15. Gryglewski RJ, Palmer RM, Moncada S. Superoxide anion is involved in the breakdown ofendothelium-derived vascular relaxing factor. Nature 1986;320:454–456. [PubMed: 3007998]
16. Guzik TJ, West N, Pillai R, Taggart D, Channon KM. Nitric oxide modulates superoxide release andperoxynitrite formation in human blood vessels. Hypertension 2002;39:1088–1094. [PubMed:12052847]
17. Abu-Soud HM, Hazen SL. Nitric oxide is a physiological substrate for mammalian peroxidases. JBiol Chem 2000;275:37524–32. [PubMed: 11090610]
18. Adachi T, Weisbrod RM, Pimentel DR, et al. S-Glutathiolation by peroxynitrite activates SERCAduring arterial relaxation by nitric oxide. Nat Med 2004;10:1200–7. [PubMed: 15489859]
19. Ritsick DR, Edens WA, Finnerty V, Lambeth JD. Nox regulation of smooth muscle contraction. FreeRadic Biol Med 2007;43:31–8. [PubMed: 17561091]
20. Wilcox JN, Subramanian RR, Sundell CL, et al. Expression of multiple isoforms of nitric oxidesynthase in normal and atherosclerotic vessels. Arterioscler Thromb Vasc Biol 1997;17:2479–88.[PubMed: 9409218]
21. Nissen SE, Tuzcu EM, Libby P, et al. Effect of antihypertensive agents on cardiovascular events inpatients with coronary disease and normal blood pressure: the CAMELOT study: a randomizedcontrolled trial. Jama 2004;292:2217–25. [PubMed: 15536108]
Guzik et al. Page 7
J Am Coll Cardiol. Author manuscript; available in PMC 2009 November 25.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 1. Contribution of Nox5 to calcium-dependent NADPH oxidase activity in humanendothelial cellsPanel A. Example ESR spectra of nitroxide adduct formation by membranes prepared fromhuman aortic endothelial cells (HAEC) in Ca++ free media and in the presence of 1 mMCa++ (left) and the effects of small interfering RNA (siRNA) against Nox5 on calciumdependent signal (right). Panel B. Effects of siRNA Nox5 on Nox5 protein in HAECs; n=4experiments; Panel C. Average Ca++ dependent (top) and Ca++ independent (bottom) NADPHoxidase activity in the presence of control siRNA (black bars) and Nox5 siRNA (open bars);n=4; values are presented as mean +/−SEM; *-p<0.01 vs control siRNA.
Guzik et al. Page 8
J Am Coll Cardiol. Author manuscript; available in PMC 2009 November 25.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 2. Calcium dependent NADPH oxidase activity and Nox5 expression in coronary arterydiseasePanel A. Calcium independent (left; panel A) and calcium dependent (right; panel A) NADPHoxidase activity in human coronary arteries in relation to the presence of coronary artery disease(CAD). NADPH oxidase activity was measured by ESR as described in methods in membranesisolated from coronary arteries of subjects with (n=8) and without (n=8) CAD. Panel B. Nox5mRNA expression in coronary arteries from patients with (n=13) and without (n=11) CAD.TaqMan real time PCR analysis was performed using commercially available Gene expressionassays. Panel C. Relationship between Ca++ dependent NADPH oxidase activity and Nox5mRNA expression in studied coronary arteries. Data were expressed as mean+/−SEM. *-p<0.05 vs non CAD; **-p<0.01 vs non CAD.
Guzik et al. Page 9
J Am Coll Cardiol. Author manuscript; available in PMC 2009 November 25.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
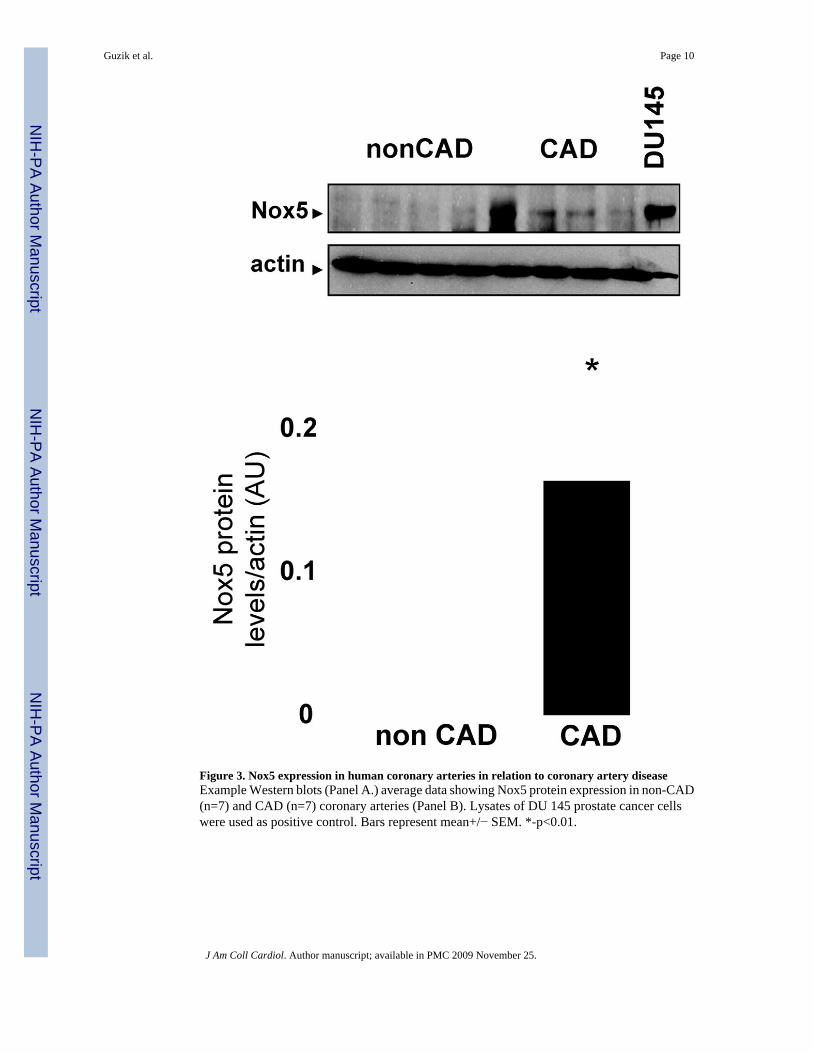
Figure 3. Nox5 expression in human coronary arteries in relation to coronary artery diseaseExample Western blots (Panel A.) average data showing Nox5 protein expression in non-CAD(n=7) and CAD (n=7) coronary arteries (Panel B). Lysates of DU 145 prostate cancer cellswere used as positive control. Bars represent mean+/− SEM. *-p<0.01.
Guzik et al. Page 10
J Am Coll Cardiol. Author manuscript; available in PMC 2009 November 25.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
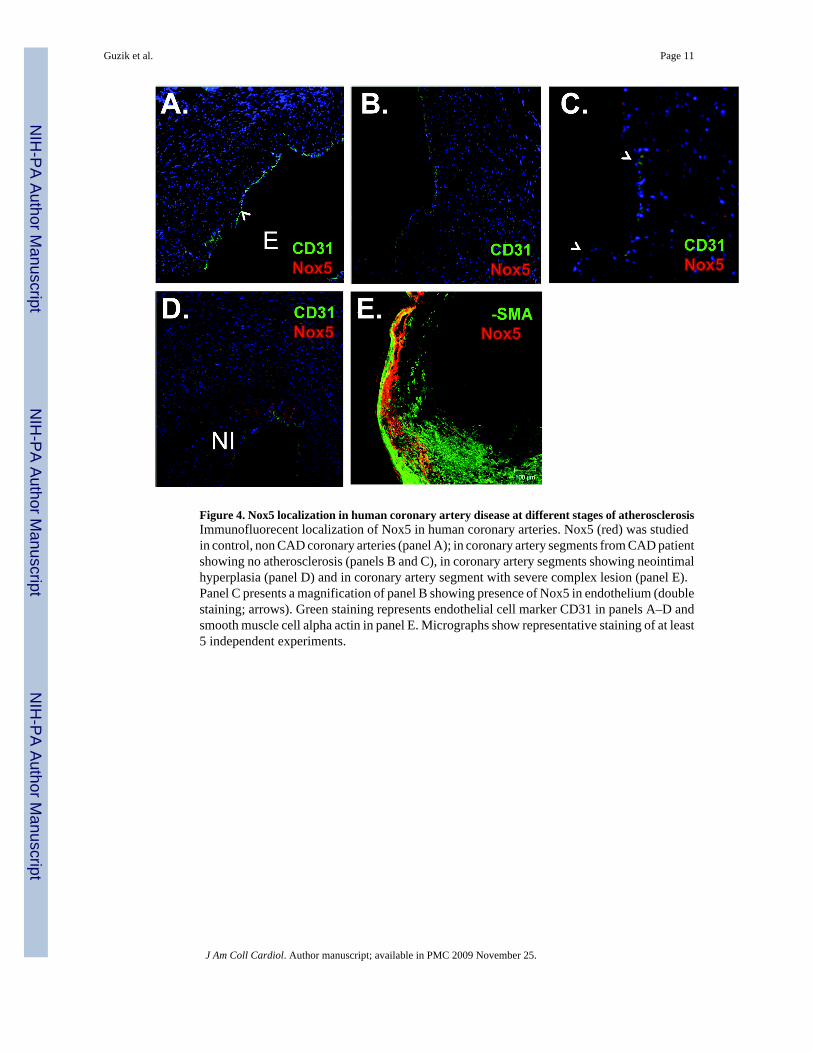
Figure 4. Nox5 localization in human coronary artery disease at different stages of atherosclerosisImmunofluorecent localization of Nox5 in human coronary arteries. Nox5 (red) was studiedin control, non CAD coronary arteries (panel A); in coronary artery segments from CAD patientshowing no atherosclerosis (panels B and C), in coronary artery segments showing neointimalhyperplasia (panel D) and in coronary artery segment with severe complex lesion (panel E).Panel C presents a magnification of panel B showing presence of Nox5 in endothelium (doublestaining; arrows). Green staining represents endothelial cell marker CD31 in panels A–D andsmooth muscle cell alpha actin in panel E. Micrographs show representative staining of at least5 independent experiments.
Guzik et al. Page 11
J Am Coll Cardiol. Author manuscript; available in PMC 2009 November 25.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Guzik et al. Page 12
Table 1Clinical characteristics of studied patients
No CAD CAD
N 12 14Age (mean ± SEM) 46.3±5.6 50.3±2.9Sex (M:F) 10:2 13:1
Risk factorsSmoking (n; %) 3 (25%) 4 (28%)Hypertension (n; %) 2 (16%) 12 (86%)*Diabetes (n; %) 0 (0%) 2 (15%)Hypercholesterolemia (n; %) 2 (16%) 9 (65%)*BMI 22.5±1.2 27.2±0.9*Cholesterol (mmol/L; mean±SEM) 4.9±0.7 5.2±0.6Fasting glucose (mmol/L; mean ±SEM) 5.0±0.3 5.6±0.2*
MI/CABG (n; %) 0 (0%) 11 (78%)*PAD, TIA (n; %) 0 (0%) 12 (86%)*Ejection fraction (%) 17.0 ± 7.2 18.2 ± 2.3
Medications Aspirin 3 (25%) 4 (28%) β-blocker 6 (50%) 7 (50%) HMG-CoA reductase inhibitor 0 (0%) 8 (55%)* Digoxin 4 (33%) 6 (42%) ACEI 9 (75%) 10 (70%)Ca++ channel blockers 0 (0%) 0 (%)
CAD- coronary artery disease; BMI - body mass index; PAD –peripheral arterial disease; TIA – transient ischemic attacks;
*- indicates p<0.05 vs no CAD group
J Am Coll Cardiol. Author manuscript; available in PMC 2009 November 25.




















