
The Caulobacter crescentus DNA-(adenine-N6)-methyltransferase CcrM methylates DNA in a distributive...
9
The Caulobacter crescentus DNA-(adenine-N6)- methyltransferase CcrM methylates DNA in a distributive manner Razvan F. Albu, Tomasz P. Jurkowski and Albert Jeltsch* Biochemistry Laboratory, School of Engineering and Science, Jacobs University Bremen, Campus Ring 1, 28759 Bremen, Germany Received August 8, 2011; Revised and Accepted September 1, 2011 ABSTRACT The specificity and processivity of DNA methyl- transferases have important implications regarding their biological functions. We have investigated the sequence specificity of CcrM and show here that the enzyme has a high specificity for GANTC sites, with only minor preferences at the central position. It slightly prefers hemimethylated DNA, which repre- sents the physiological substrate. In a previous work, CcrM was reported to be highly processive [Berdis et al. (1998) Proc. Natl Acad. Sci. USA 95: 2874–2879]. However upon review of this work, we identified a technical error in the setup of a crucial experiment in this publication, which prohibits making any statement about the processivity of CcrM. In this study, we performed a series of in vitro experiments to study CcrM processivity. We show that it distributively methylates six target sites on the pUC19 plasmid as well as two target sites located on a 129-mer DNA fragment both in unmethylated and hemimethylated state. Reaction quenching experiments confirmed the lack of processivity. We conclude that the original state- ment that CcrM is processive is no longer valid. INTRODUCTION DNA methylation at position N 6 of adenine, or at position N 4 or C 5 of cytosine bases is a chemical modifi- cation of DNA present in a wide variety of prokaryotic and eukaryotic organisms (1,2). The methylation reaction is catalyzed by DNA methyltransferases (MTases) which employ S-adenosine-L-methionine (AdoMet) as methyl group donor. In bacteria, DNA methylation is most often associated with restriction-modification (RM) systems, which protect the bacterial cell against bacterio- phages (3). However, there exists a distinct class of bacterial DNA MTases, known as solitary MTases, which are not part of an RM system. The best known examples of solitary MTases are the Escherichia coli DNA adenine MTase (EcoDam) which recognizes GA TC sequences and regulates DNA repair, gene expres- sion and DNA replication (1,4), and the Caulobacter crescentus cell-cycle regulating MTase (CcrM) which methylates the adenine in GA NTC sites and has a central role in the regulation of the bacterial cell division cycle (5–7). Furthermore, CcrM is an essential protein in several a-Proteobacteria, including pathogens, which makes it a potential antibacterial drug target (8–10). One important property of DNA MTases is their processivity in the methylation of DNA molecules con- taining more than one target site. Processive enzymes stay bound to one DNA molecule after first turnover and methylate several target sites on that molecule without dissociation. Thereby, they directly convert unmethylated DNA into DNA modified at all target sites. Distributive enzymes, in contrast, always dissociate from the DNA after one methyl group transfer leading to an accumulation of methylation intermediates, i.e. DNA molecules that are modified at some but not all target sites. Since methylation intermediates are not released by processive enzymes, detection of the presence or absence of intermediates is the most direct and reliable experimen- tal approach in processivity analysis. The processivity of DNA methyltransferases has a strong impact on their bio- logical function, because DNA methylation is established in a radically different way by each type of enzyme. The EcoDam enzyme, for example, was shown to be highly processive, thus leading to efficient re-methylation of the GATC sites after DNA replication (11), although particu- lar flanking sequences were shown to reduce processivity (12). T4Dam was shown to be processive as well (13), while most of the methyltransferases associated with RM systems are distributive, which may help prevent the methylation of incoming phage DNA before its cleavage by restriction digestion (1,11). *To whom correspondence should be addressed. Tel: +49 421 200 3247; Fax: +49 421 200 3249; Email: [email protected] Nucleic Acids Research, 2011, 1–9 doi:10.1093/nar/gkr768 ß The Author(s) 2011. Published by Oxford University Press. This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/ by-nc/3.0), which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited. Nucleic Acids Research Advance Access published September 16, 2011 by guest on June 6, 2016 http://nar.oxfordjournals.org/ Downloaded from
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of The Caulobacter crescentus DNA-(adenine-N6)-methyltransferase CcrM methylates DNA in a distributive...

The Caulobacter crescentus DNA-(adenine-N6)-methyltransferase CcrM methylates DNA in adistributive mannerRazvan F. Albu, Tomasz P. Jurkowski and Albert Jeltsch*
Biochemistry Laboratory, School of Engineering and Science, Jacobs University Bremen, Campus Ring 1,28759 Bremen, Germany
Received August 8, 2011; Revised and Accepted September 1, 2011
ABSTRACT
The specificity and processivity of DNA methyl-transferases have important implications regardingtheir biological functions. We have investigated thesequence specificity of CcrM and show here that theenzyme has a high specificity for GANTC sites, withonly minor preferences at the central position. Itslightly prefers hemimethylated DNA, which repre-sents the physiological substrate. In a previouswork, CcrM was reported to be highly processive[Berdis et al. (1998) Proc. Natl Acad. Sci. USA 95:2874–2879]. However upon review of this work, weidentified a technical error in the setup of a crucialexperiment in this publication, which prohibitsmaking any statement about the processivity ofCcrM. In this study, we performed a series ofin vitro experiments to study CcrM processivity.We show that it distributively methylates six targetsites on the pUC19 plasmid as well as two targetsites located on a 129-mer DNA fragment both inunmethylated and hemimethylated state. Reactionquenching experiments confirmed the lack ofprocessivity. We conclude that the original state-ment that CcrM is processive is no longer valid.
INTRODUCTION
DNA methylation at position N6 of adenine, or atposition N4 or C5 of cytosine bases is a chemical modifi-cation of DNA present in a wide variety of prokaryoticand eukaryotic organisms (1,2). The methylation reactionis catalyzed by DNA methyltransferases (MTases) whichemploy S-adenosine-L-methionine (AdoMet) as methylgroup donor. In bacteria, DNA methylation is mostoften associated with restriction-modification (RM)systems, which protect the bacterial cell against bacterio-phages (3). However, there exists a distinct class of
bacterial DNA MTases, known as solitary MTases,which are not part of an RM system. The best knownexamples of solitary MTases are the Escherichia coliDNA adenine MTase (EcoDam) which recognizesGATC sequences and regulates DNA repair, gene expres-sion and DNA replication (1,4), and the Caulobactercrescentus cell-cycle regulating MTase (CcrM) whichmethylates the adenine in GANTC sites and has acentral role in the regulation of the bacterial cell divisioncycle (5–7). Furthermore, CcrM is an essential protein inseveral a-Proteobacteria, including pathogens, whichmakes it a potential antibacterial drug target (8–10).One important property of DNA MTases is their
processivity in the methylation of DNA molecules con-taining more than one target site. Processive enzymesstay bound to one DNA molecule after first turnoverand methylate several target sites on that moleculewithout dissociation. Thereby, they directly convertunmethylated DNA into DNA modified at all targetsites. Distributive enzymes, in contrast, always dissociatefrom the DNA after one methyl group transfer leading toan accumulation of methylation intermediates, i.e. DNAmolecules that are modified at some but not all targetsites. Since methylation intermediates are not released byprocessive enzymes, detection of the presence or absenceof intermediates is the most direct and reliable experimen-tal approach in processivity analysis. The processivity ofDNA methyltransferases has a strong impact on their bio-logical function, because DNA methylation is establishedin a radically different way by each type of enzyme. TheEcoDam enzyme, for example, was shown to be highlyprocessive, thus leading to efficient re-methylation of theGATC sites after DNA replication (11), although particu-lar flanking sequences were shown to reduce processivity(12). T4Dam was shown to be processive as well (13),while most of the methyltransferases associated withRM systems are distributive, which may help preventthe methylation of incoming phage DNA before itscleavage by restriction digestion (1,11).
*To whom correspondence should be addressed. Tel: +49 421 200 3247; Fax: +49 421 200 3249; Email: [email protected]
Nucleic Acids Research, 2011, 1–9doi:10.1093/nar/gkr768
� The Author(s) 2011. Published by Oxford University Press.This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0), which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Nucleic Acids Research Advance Access published September 16, 2011 by guest on June 6, 2016
http://nar.oxfordjournals.org/D
ownloaded from

In a publication of Berdis et al. (14), it was reported thatCcrM methylated DNA in a processive manner. The assayapplied in that work for detection of CcrM processivityprobed the methylation of GANTC sites through protec-tion against HindII (GTYRAC) cleavage at overlappingsites (Figure 2A). However, although the substrate con-tained two CcrM sites, only one of them was flanked by aHindII site. Therefore, only one GANTC site was beingprobed for methylation and no conclusion could be drawntoward processivity. It is clear that this error is not just atypographical mistake in the ‘Materials and Methods’section of the manuscript, because at the zero time pointin Figure 5 of the Berdis et al. (14) publication, the long51-mer HindII cleavage product was observed, which isindicative of the absence of cleavage at the second CcrMsite. Since no methylation can be present at this point, thisresult can only be explained by the absence of the secondHindII site. Thus, the issue of processivity of CcrM mustbe considered open and not resolved. In this work, wehave re-investigated the processivity of CcrM and showthat it methylates pUC19 plasmid DNA and a linear129-mer substrate in a distributive manner.
MATERIALS AND METHODS
Protein expression and purification
His6-tagged C. crescentus CcrM cloned into pET28 vectorwas kindly provided by Dr Xiaodong Cheng (EmoryUniversity). The pET28 plasmid encoding His-taggedCcrM was transformed into HMS174 (DE3) cells(Novagen). The cells were grown in LB medium untilmid-exponential phase (OD600 0.5–0.6) and expressionwas induced by addition of 1mM isopropyl-b-D-1-thiogalactopyranoside (IPTG), after which thecultures were incubated at room temperature for an add-itional 3 h. The cells were harvested by centrifugation at4300g for 15min, followed by washing with STE buffer(10mM Tris–HCl, pH 8.0, 100mMNaCl, 0.1mM EDTA)and an additional centrifugation step. The His-taggedCcrM was purified by affinity chromatography onNi-NTA Agarose (Qiagen) using sonication/washingbuffer (50mM HEPES, pH 7.5, 500mM NaCl, 0.1mMDTT, 10% glycerol, 20mM imidazol) and elution buffer(50mM HEPES, pH 7.5, 500mM NaCl, 0.1mM DTT,10% glycerol, 220mM imidazol). The eluate wasdialyzed in dialysis buffer I (50mM HEPES, pH 7.5,200mM NaCl, 0.1mM DTT, 10% glycerol) for 2–3 h,and in dialysis buffer II (50mM HEPES, pH 7.5,200mM NaCl, 0.1mM DTT, 60% glycerol) for an add-itional 12 h. The purified protein was at high concentra-tion (�100 mM) and contained only two minorcontaminants (Figure 1A).
DNA substrates
The 129-mer DNA substrate containing two CcrM targetsites which was used for processivity analysis wasproduced by PCR using the pBAD24 vector as template.The hemimethylated 129-mer HM substrate was obtainedby PCR using an in vitro synthesized DNA oligonucleo-tide as template. The M.TaqI methylation was carried out
at 65�C for 2 h, with 80 mM AdoMet and using NEB4buffer supplied by New England Biolabs. The methylatedDNA was purified using a standard DNA extraction kit(Macherey-Nagel NucleoSpin ExtractII). All substratesdescribed above are shown in Figure 2B–D. The 23-mersubstrates used for determining the relative preference forhemimethylated over unmethylated DNA and for thequenching experiment were obtained by heating of anequimolar (20 mM) mixture of complementary oligo-nucleotides to 95�C for 5min and allowing the mixtureto slowly cool down to room temperature. The qualityof the annealing procedure was assessed by polyacryl-amide gel electrophoresis.
23-mer: Bt_d(GGCAGCTACGAATCGCAACAGCT)23-mer revmet:_d(AGCTGTTGCGmATTCGTAGC
TGCC)23-mer_rev: d(AGCTGTTGCGATTCGTAGCTGCC)
In addition, 12 double-stranded 23-mer substrates wereused, in which the base pairs at positions 1, 3, 4 or 5 wereexchanged against all other base pairs. All substrates werebiotinylated and hemimethylated with the methylation inthe lower strand (except of variants at the fourth position,which did not contain an A in the lower strand). Thesesubstrates were used to investigate the sequence specificityof CcrM for the first, third, fourth and fifth position of theGANTC site.
DNA binding experiments
DNA binding was analyzed by using the nitrocellulosefilter-binding assay. For the experiment, radioactivelylabeled hemimethylated 129-mer DNA substrate wasprepared by phosphorylation of the hemimethylatedDNA with g-[P32]-ATP using T4 polynucleotide kinase(NEB) following the recommendations of the supplier.CcrM concentrations were varied between 0 and 10 mM,10 nM of DNA was used. The binding reactions wereincubated in binding buffer (50mM HEPES pH 7.5,50mM NaCl, 1mM EDTA and 500 mM DTT) supple-mented with 200 mM sinefungin (Sigma) for 30min atambient temperature. The nitrocellulose filter membrane(Macherey & Nagel, Duren, Germany) was soaked inbinding buffer for 30min. Afterwards the membrane wastransferred into the dot blot chamber (BioRad) and theslots were washed with binding buffer. The samples weretransferred into the wells of the dot blot apparatus using amultiple pipette, immediately sucked through the nitrocel-lulose filter membrane, and washed several times with100 ml of binding buffer. The membranes were dried andthe radioactivity of the spots analyzed using aPhosphorImager (Fuji). The results were fitted to theequation describing a bimolecular association equilibriumto determine the binding constant.
CcrM DNA methylation reactions for processivity analysis
All methylation kinetics were performed in a buffer con-taining 50mM HEPES pH 7.5, 50mM NaCl, 1mMEDTA and 500mM DTT, in the presence of 5 ng/mlBSA. The AdoMet (Sigma-Aldrich) cofactor was used ata concentration of 200 mM for the processivity
2 Nucleic Acids Research, 2011
by guest on June 6, 2016http://nar.oxfordjournals.org/
Dow
nloaded from
experiments. All methylation reactions were carried out atroom temperature and started by addition of CcrM.Methylation of pUC19 was carried out using 2mMCcrM and 600 nM pUC19. To assess the methylationstate of the pUC19 vector, a double digestion wascarried out using HinfI and NdeI (New EnglandBiolabs), the latter being used for linearization of thevector. The double digestion was carried out overnightat 37�C in buffer NEB4. The methylation reactions withthe unmethylated 129-mer were carried out using 1 mMDNA and 2 mM CcrM, with the hemimethylated129-mer HM 1 mM DNA and 0.5 mM CcrM were used.Aliquotes were taken at various time points and reactionswere stopped by shock freezing in liquid nitrogen andpurified using PCR purification kit (Macherey-NagelNucleoSpin Extract II). All restriction endonuclease treat-ments were carried out for at least 1 h under the appropri-ate conditions, as recommended by the provider. For the
quenching experiment, a standard methylation wasprepared and time points were collected and treated thesame way as described above, except that 3min afterstarting the reaction 10 mM double stranded 23-mer com-petitor substrate was added.
Methylation of oligonucleotide substrates usingradioactively labeled AdoMet
In order to study the methylation of the unmethylated andhemimethylated 23-mer substrates by CcrM, the in vitrobiotin/avidin methylation assay was performed, as previ-ously described (15). The methylation reactions were per-formed in a 40 ml total volume, under single turnoverconditions using 760 nM 3H-labeled AdoMet. Theenzyme and DNA concentrations are given in the maintext. The enzymatic activity was assessed by linear
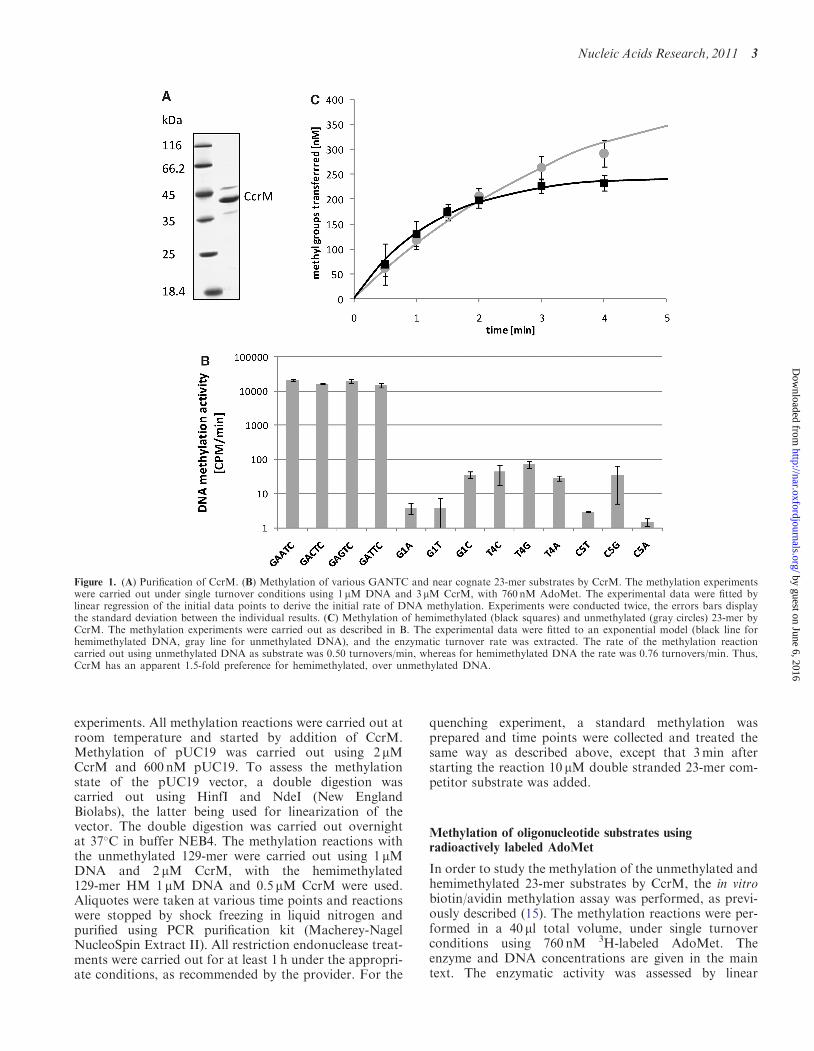
Figure 1. (A) Purification of CcrM. (B) Methylation of various GANTC and near cognate 23-mer substrates by CcrM. The methylation experimentswere carried out under single turnover conditions using 1mM DNA and 3 mM CcrM, with 760 nM AdoMet. The experimental data were fitted bylinear regression of the initial data points to derive the initial rate of DNA methylation. Experiments were conducted twice, the errors bars displaythe standard deviation between the individual results. (C) Methylation of hemimethylated (black squares) and unmethylated (gray circles) 23-mer byCcrM. The methylation experiments were carried out as described in B. The experimental data were fitted to an exponential model (black line forhemimethylated DNA, gray line for unmethylated DNA), and the enzymatic turnover rate was extracted. The rate of the methylation reactioncarried out using unmethylated DNA as substrate was 0.50 turnovers/min, whereas for hemimethylated DNA the rate was 0.76 turnovers/min. Thus,CcrM has an apparent 1.5-fold preference for hemimethylated, over unmethylated DNA.
Nucleic Acids Research, 2011 3
by guest on June 6, 2016http://nar.oxfordjournals.org/
Dow
nloaded from

regression of the initial data points or by fitting thereaction progress curve to the following exponentialequation:
CPM ¼ BL+F � ð1� e�k�tÞ
where BL indicates the background signal, F is the inten-sity factor, and k represents the catalytic rate constant,expressed as turnovers per minute. Calibration was donewith completely methylated DNA, incubated with CcrMfor several hours.
RESULTS
We have expressed His6-tagged CcrM in E. coli andpurified it using a Ni-NTA column (Figure 1A). We ini-tially investigated the specificity of CcrM by usingbiotinylated oligonucleotide substrates which wereincubated with CcrM and AdoMet, the methyl group ofwhich was radioactively labeled. After methylation, theDNA was purified using an avidin microplate and the in-corporation of radioactivity into the DNA was detected.
We used four 23-mer substrates with altered centralposition (N3) representing the different versions of theGANTC cognate sites. In addition, nine substrates wereused which contained variants of the GANTC targetsite in which one base pair was altered at the G1, T4 orC5 position (near-cognate sites). Substrates werehemimethylated in the lower strand, such that methylationof the upper strand was detected. The results shown inFigure 1B illustrate that CcrM has a high preference forthe GANTC sequence, because the best near-cognate sub-strate was methylated 200-fold less efficiently than theworst cognate one. Many of the near-cognate substrateswere methylated more than 1000-fold less efficiently. Atthe central position we observed only moderate variationsin the methylation rate which were close to the experimen-tal fluctuations. Since hemimethylated DNA is theproduct of DNA replication of methylated GANTCsites in vivo, it is the major physiological substrate ofCcrM. We compared the activity of CcrM usinghemimethylated and unmethylated DNA substratesindicating that CcrM methylates both substrates, but it
Figure 2. Various substrates used for studying CcrM processivity. (A) Substrate used by Berdis et al. (14) to study CcrM processivity, referred to asN6 60/66-mer. Two GANTC target sites are present, hemimethylated on the upper strand. HindII target sites (GTYRAC) coupled to CcrM targetsites were used to screen for methylation on the lower strand. However, only one of the two HindII sites is present, making it impossible to probe themethylation state of the second site. (B) The distribution of GANTC sequences (shown as HinfI target sequences) throughout the pUC19 plasmid.The position of each sequence is indicated relative to the plasmid’s replication origin. The vector contains a single NdeI target site, which was used inconjunction with HinfI for vector linearization, to facilitate viewing of the progression toward fully methylated state. (C) 129-mer substrate con-taining two CcrM target sites. The expected size of the fragments obtained after HinfI digestion of completely unmethylated, partially methylatedand fully methylated substrates are indicated. (D) 129-mer_HM substrate used to probe CcrM activity over hemimethylated GANTC sites.A M.TaqI methylation site (TCGA), as well as a HincII restriction site (GTYRAC) were linked to the GANTC site. M.TaqI-established methylationoccurs as shown earlier, creating two GANTC sites hemimethylated on the lower strand. CcrM-catalyzed methylation of the upper strand wasprobed through protection from HincII digestion, which is blocked by hemimethylation.
4 Nucleic Acids Research, 2011
by guest on June 6, 2016http://nar.oxfordjournals.org/
Dow
nloaded from
displayed a weak preference of �1.5-fold for methylationof hemimethylated DNA over the average of the methy-lation rates of both strands of the unmethylated DNA(Figure 1C).
Methylation of a plasmid substrate
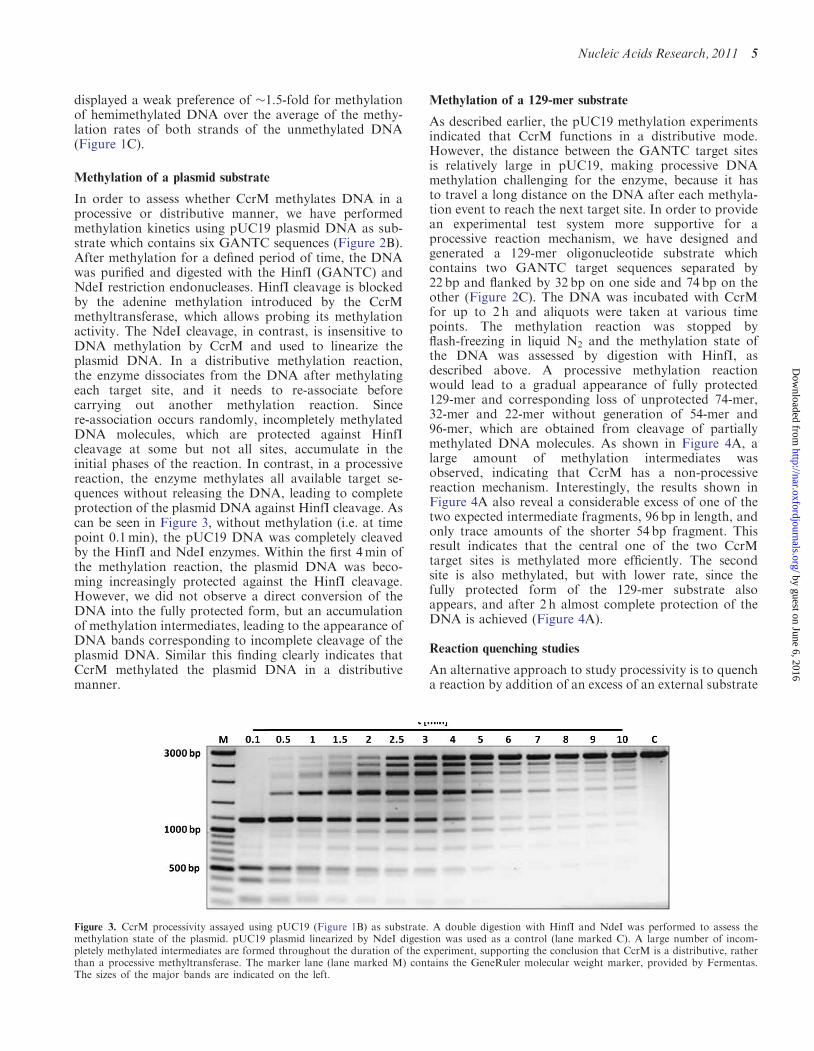
In order to assess whether CcrM methylates DNA in aprocessive or distributive manner, we have performedmethylation kinetics using pUC19 plasmid DNA as sub-strate which contains six GANTC sequences (Figure 2B).After methylation for a defined period of time, the DNAwas purified and digested with the HinfI (GANTC) andNdeI restriction endonucleases. HinfI cleavage is blockedby the adenine methylation introduced by the CcrMmethyltransferase, which allows probing its methylationactivity. The NdeI cleavage, in contrast, is insensitive toDNA methylation by CcrM and used to linearize theplasmid DNA. In a distributive methylation reaction,the enzyme dissociates from the DNA after methylatingeach target site, and it needs to re-associate beforecarrying out another methylation reaction. Sincere-association occurs randomly, incompletely methylatedDNA molecules, which are protected against HinfIcleavage at some but not all sites, accumulate in theinitial phases of the reaction. In contrast, in a processivereaction, the enzyme methylates all available target se-quences without releasing the DNA, leading to completeprotection of the plasmid DNA against HinfI cleavage. Ascan be seen in Figure 3, without methylation (i.e. at timepoint 0.1min), the pUC19 DNA was completely cleavedby the HinfI and NdeI enzymes. Within the first 4min ofthe methylation reaction, the plasmid DNA was beco-ming increasingly protected against the HinfI cleavage.However, we did not observe a direct conversion of theDNA into the fully protected form, but an accumulationof methylation intermediates, leading to the appearance ofDNA bands corresponding to incomplete cleavage of theplasmid DNA. Similar this finding clearly indicates thatCcrM methylated the plasmid DNA in a distributivemanner.
Methylation of a 129-mer substrate
As described earlier, the pUC19 methylation experimentsindicated that CcrM functions in a distributive mode.However, the distance between the GANTC target sitesis relatively large in pUC19, making processive DNAmethylation challenging for the enzyme, because it hasto travel a long distance on the DNA after each methyla-tion event to reach the next target site. In order to providean experimental test system more supportive for aprocessive reaction mechanism, we have designed andgenerated a 129-mer oligonucleotide substrate whichcontains two GANTC target sequences separated by22 bp and flanked by 32 bp on one side and 74 bp on theother (Figure 2C). The DNA was incubated with CcrMfor up to 2 h and aliquots were taken at various timepoints. The methylation reaction was stopped byflash-freezing in liquid N2 and the methylation state ofthe DNA was assessed by digestion with HinfI, asdescribed above. A processive methylation reactionwould lead to a gradual appearance of fully protected129-mer and corresponding loss of unprotected 74-mer,32-mer and 22-mer without generation of 54-mer and96-mer, which are obtained from cleavage of partiallymethylated DNA molecules. As shown in Figure 4A, alarge amount of methylation intermediates wasobserved, indicating that CcrM has a non-processivereaction mechanism. Interestingly, the results shown inFigure 4A also reveal a considerable excess of one of thetwo expected intermediate fragments, 96 bp in length, andonly trace amounts of the shorter 54 bp fragment. Thisresult indicates that the central one of the two CcrMtarget sites is methylated more efficiently. The secondsite is also methylated, but with lower rate, since thefully protected form of the 129-mer substrate alsoappears, and after 2 h almost complete protection of theDNA is achieved (Figure 4A).
Reaction quenching studies
An alternative approach to study processivity is to quencha reaction by addition of an excess of an external substrate
Figure 3. CcrM processivity assayed using pUC19 (Figure 1B) as substrate. A double digestion with HinfI and NdeI was performed to assess themethylation state of the plasmid. pUC19 plasmid linearized by NdeI digestion was used as a control (lane marked C). A large number of incom-pletely methylated intermediates are formed throughout the duration of the experiment, supporting the conclusion that CcrM is a distributive, ratherthan a processive methyltransferase. The marker lane (lane marked M) contains the GeneRuler molecular weight marker, provided by Fermentas.The sizes of the major bands are indicated on the left.
Nucleic Acids Research, 2011 5
by guest on June 6, 2016http://nar.oxfordjournals.org/
Dow
nloaded from

acting as competitor. Thereby, an enzyme working dis-tributively will be trapped and the reaction stopped. Incontrast, a processive enzyme should be able to finishthe methylation of the substrate it is bound to. In orderto implement this assay, the 129-mer methylation reactionwas performed as described and after 3min a 10-foldexcess of a competing 23-mer oligonucleotide containinga single, unmethylated GANTC target site was added. Ourresults show that before the addition of the competitorDNA, the 129-mer substrate was methylated with samekinetics as seen before (compare Figure 4A for methyla-tion without competitor and Figure 4B for methylationwith competitor). However, after addition of the competi-tor, the level of protection of the 129-mer DNA remainedconstant throughout the duration of the experiment, withno additional methylation taking place. Most importantly,the partially methylated intermediates were not convertedinto the fully methylated state indicating that the CcrMenzyme cannot move to the second site after methylationof one site of the 129-mer substrate. This behavior ischaracteristic of a distributive, rather than a processive,
methyltransferase confirming the conclusions from theprevious experiments.
Analysis of processivity on hemimethylated DNA
Although CcrM is able to efficiently methylate completelyunmodified substrates (Figures 1C, 3 and 4), it could beargued that processive methylation might occur on thepreferred substrate. To this end, a CcrM in vitro methyla-tion experiment was conducted using a hemimethylatedvariant of the 129-mer substrate (Figure 2D).Hemimethylation was introduced into the 129-mer HMsubstrate by using GTTGACTCGA sites which representoverlapping HincII (GTYRAC, position 1–6 of thesequence, inhibited by adenine methylation in eitherstrand), CcrM (GANTC, position 4–9 of the combinedsequence) and TaqI sites (TCGA, position 7–11 of thecombined sequence). The 129-mer HM DNA wasmethylated by M.TaqI at the TCGA adenine residues inboth strands. In the upper strand, the methylation isoutside of the CcrM site and it does not influenceGANTC methylation by CcrM. However, M.TaqI
Figure 4. CcrM processivity assays using the 129-mer DNA as substrate (Figure 2C) and conducted in the absence (A) and with the addition (B) ofcompetitor DNA. The expected running distance of partially protected intermediates is indicated on the right by asterisk. Undigested 129-mer DNAwas used as control, indicating the expected running distance of fully protected DNA. The marker lane (indicated by M) contained PCR markerprovided by New England Biolabs. The sizes of the bands are indicated on the left. (A) Initially all of the DNA is efficiently digested by HinfI (timepoint 0.1min). Increasing protection from HinfI digestion is established over time, with nearly complete protection being achieved after 120min. The96 bp intermediate is present in large amounts, as well as low levels of the 54 bp intermediate, indicating distributive methylation by CcrM, as well asa preference for one target sequence over the other. (B) Competitor DNA is added after 3min (indicated by the arrow). The protection state of the129-mer substrate remains the same after supplementation with competitor, suggesting dissociation of CcrM from incompletely methylated DNA.The weak band appearing in the last lanes at low molecular weights corresponds to the 23-mer competitor which has become methylated and,thereby, protected against HinfI cleavage.
6 Nucleic Acids Research, 2011
by guest on June 6, 2016http://nar.oxfordjournals.org/
Dow
nloaded from

methylation of the lower strand creates a hemimethylatedGANTC site. Completeness of the M.TaqI methylationwas confirmed by full resistance of the DNA againstR.TaqI cleavage after incubation with M.TaqI(Figure 5A). The hemimethylated state of all the CcrMsites was assayed by a control digestion with HinfI,which has the same recognition sequence as CcrM and isinhibited by hemimethylation of its target sequence. Asshown in Figure 5A, the 129-mer HM DNA wascompletely refractory to HinfI cleavage after incubationwith M.TaqI, indicating that all GANTC sites werehemimethylated. As expected, M.TaqI methylation didnot interfere with HincII cleavage because thehemimethylation resides outside of its recognitionsequence (Figure 5A).
In order to characterize the reaction process (cf.‘Discussion’ section), we determined the bindingconstant of CcrM to this substrate by nitrocellulose filterbinding after radioactive labeling of the hemimethylatedDNA (Figure 5B). We observed relatively weak bindingwith a binding constant of KAss=5.4� 105 M�1.
Next, this substrate containing hemimethylatedGANTC sites, which are methylated in the lower DNAstrand, was incubated with CcrM, which also leads to themethylation of the upper strand of the GANTC site. Since
the upper strand adenine of the GANTC site overlapswith the HincII site, and HincII is inhibited by adeninemethylation in either strand, the conversion ofhemimethylated to fully methylated CcrM sites can befollowed by protection against HincII cleavage. Asbefore, processive methylation would cause the direct con-version of the 22, 32 and 74 bp fragments into fully pro-tected 129-mer while appearance of 54 and 96 bpfragments which correspond to partially methylatedDNA would be expected in case of a distributivereaction mechanism. As shown in Figure 5C, appearanceof the 96-mer and 54-mer DNA fragments indeed wasobserved, which is indicative of distributive DNAmethylation. This finding confirms that CcrM is distribu-tive on unmethylated and hemimethylated DNA. Unlikethe results shown in Figure 4A, CcrM appears to have nopreference for either of the target sites on the 129-mer HMsubstrate, since the relative abundance of both intermedi-ate fragments is approximately the same throughout theexperiment. This is expected, since the inclusion of theHincII and TaqI sites on either side of both GANTCsites has placed them in an identical sequence context,whereas in the original 129-mer DNA, the flankingsequences of the two sites were different (compareFigure 2C and D).
Figure 5. (A) Quality control of M.TaqI methylated 129-mer HM DNA. Digestions were performed of 129-mer HM DNA, before and aftermethylation with M.TaqI (as indicated), using TaqI, HinfI and HincII. As expected, TaqI could not cleave DNA, after treatment with M.TaqI.HinfI was also unable to cleave methylated DNA, whereas HincII was unhindered by the M.TaqI treatment. Thus, since both HinfI and HincII areblocked by N6-adenine hemimethylation, the CcrM target sites present on the 129-mer HM substrate are both in hemimethylated state after M.TaqItreatment. (B) Binding of CcrM to the hemimethylated 129-mer analyzed by nitrocellulose filter binding. The insert shows one example of primarydata, the graph shows the quantitative analysis of one binding experiments. In two independent experiments the binding constant was determined as5.4 (±0.1)� 105M�1. (C) CcrM methylation of 129-mer HM DNA. Both the 96-mer and 54-mer methylation intermediates can be seen (indicated byasterisk), thus CcrM functions in a distributive manner on hemimethylated DNA. In both (A) and (B) the control lane (marked C) containedundigested 129-mer HM DNA. The marker lane (indicated by M) contained the same PCR marker (New England Biolabs) used in the previousexperiments, shown in Figure 3.
Nucleic Acids Research, 2011 7
by guest on June 6, 2016http://nar.oxfordjournals.org/
Dow
nloaded from

DISCUSSION
Caulobacter crescentus is an established model system tostudy cell cycle regulation and differentiation in bacteria.In a series of seminal papers in the 1990s, Shapiro andcoworkers (5,8,16–20) identified the CcrM DNAmethyltransferase and described its biological role in thecontrol of the cell cycle of Caulobacter. The enzymaticproperties of CcrM were studied by Benkovic andcoworkers (14,21), showing that CcrM is active asmonomer and dimer and it has some preference for methy-lation of hemimethylated targets. Here, we show thatCcrM has a high specificity for GANTC sites which issimilar to results obtained with T4Dam and EcoDam(22,23). At the central position, CcrM prefers GANTCsites 200-fold over the best near cognate substrate andhemimethylated targets are modified about 1.5-foldfaster than unmethylated. This slight preference of CcrMfor hemimethylated targets makes physiological sense,because replication of the Caulobacter chromosome gen-erates hemimethylated GANTC sites, which, therefore,are the main physiological substrate of CcrM. Inaddition, Berdis et al. (14) reported that CcrM methylatesDNA in a highly processive manner in a paper that ishighly cited. Unfortunately, due to a mistake in thedesign of the methylation substrates that has beendescribed above (Figure 2A), the conclusion of processiveDNA methylation was not justified. Berdis et al. (14)based their claim of processive DNA methylation alsoon a second observation, which is that CcrM methylateda substrate with two CcrM sites 2-fold faster than a singlesite substrate used at the same concentration. However,this experiment was not conclusive, because the two sitesubstrate offers twice the amount and concentration oftarget sites, which may explain the higher incorporationof radioactivity.The question if a DNA MTase acts in a processive or
distributive manner has important consequences for itsbehavior in the biological context of a living cell andstrongly influences the pathway of re-establishing DNAmethylation after DNA replication. Therefore, we havere-examined the processivity of CcrM and show thatCcrM distributively methylated six target sites on thepUC19 plasmid as well as two target sites located on a129-mer DNA fragment both in unmethylated andhemimethylated state. One technical challenge in detectingpotential processivity of CcrM was the relatively weakactivity of the enzyme, which may pretend a distributivereaction mechanism because of incomplete reactionprogress. However, this argument does not apply to ourstudy, since we observed complete methylation of the sub-strate in all our experiments (cf. Figures 3–5). Anotherpotential problem, that could obscure a processivereaction mechanism, is that two enzymes may bind onthe same substrate and methylate independently of eachother or block each others movement on the DNA. Toestimate if such problem could have occurred in our ex-periments, we determined the DNA binding constant ofCcrM to the hemimethylated 129-mer and found a KAss of5.4� 105M�1. In the corresponding methylation kineticswith the hemimethylated 129-mer, we used 1 mM DNA
and 0.5 mM CcrM, which corresponds to an average occu-pancy of the DNA of 7.5%. This result indicates that onaverage only 0.6% of the substrate molecules (=0.0752)had two CcrM molecules bound. Therefore, processiveDNA methylation should have been detectable underthese conditions. Since, the reaction quenching experi-ments confirmed the lack of processivity, we concludethat the original statement that CcrM is processive is nolonger valid.
Our new finding of a distributive methylation mechan-ism of CcrM can be interpreted by considering the bio-logical role of the enzyme. Since a processive enzymemethylates several sites after one DNA binding event, ittypically has long residence times on the DNA. Oneparadigm of this type is the EcoDam enzyme (11), whichshortly follows the replication fork and, therefore,probably acts on nascent DNA with few other proteinsbound. In contrast, distributive enzymes have to bind toand dissociate from the DNA for each methylation eventand typically have a short dwell time on the DNA. Thedistributive mechanism may be better suited for CcrM,because in C. crescentus DNA methylation happens afterDNA replication when other DNA binding proteins havehad enough time to re-bind to the DNA. Under such con-ditions, a processive DNA methylation may not be ideal,because bound proteins would act as roadblocks andprocessive MTases would be trapped due their slow dis-sociation from the DNA. Thereby the entire process ofDNA methylation could become impeded. On the otherhand, the necessity for multiple DNA binding events ofdistributive enzymes suggest that there may exist mechan-isms for targeting CcrM to the DNA, which so far havenot been discovered.
ACKNOWLEDGEMENTS
The authors gratefully acknowledge the gift of a CcrMexpression construct from Prof. X. Cheng.
FUNDING
This work has been supported by the DeutscheForschungsgemeinschaft (JE 252/2). Funding for openaccess charge: DFG (JE 252/2).
Conflict of interest statement. None declared.
REFERENCES
1. Jeltsch,A. (2002) Beyond Watson and Crick: DNA methylationand molecular enzymology of DNA methyltransferases.Chembiochem, 3, 274–293.
2. Cheng,X. (1995) Structure and function of DNAmethyltransferases. Annu. Rev. Biophys. Biomol. Struct., 24,293–318.
3. Pingoud,A. and Jeltsch,A. (2001) Structure and function of typeII restriction endonucleases. Nucleic Acids Res., 29, 3705–3727.
4. Wion,D. and Casadesus,J. (2006) N6-methyl-adenine: anepigenetic signal for DNA-protein interactions. Nat. Rev.Microbiol., 4, 183–192.
5. Reisenauer,A., Kahng,L.S., McCollum,S. and Shapiro,L. (1999)Bacterial DNA methylation: a cell cycle regulator? J. Bacteriol.,181, 5135–5139.
8 Nucleic Acids Research, 2011
by guest on June 6, 2016http://nar.oxfordjournals.org/
Dow
nloaded from

6. Reisenauer,A. and Shapiro,L. (2002) DNA methylation affects thecell cycle transcription of the CtrA global regulator inCaulobacter. EMBO J., 21, 4969–4977.
7. Collier,J., McAdams,H.H. and Shapiro,L. (2007) A DNAmethylation ratchet governs progression through a bacterial cellcycle. Proc. Natl Acad. Sci. USA, 104, 17111–17116.
8. Wright,R., Stephens,C. and Shapiro,L. (1997) The CcrM DNAmethyltransferase is widespread in the alpha subdivision ofproteobacteria, and its essential functions are conserved inRhizobium meliloti and Caulobacter crescentus. J. Bacteriol., 179,5869–5877.
9. Robertson,G.T., Reisenauer,A., Wright,R., Jensen,R.B., Jensen,A.,Shapiro,L. and Roop,R.M. 2nd (2000) The Brucella abortusCcrM DNA methyltransferase is essential for viability, and itsoverexpression attenuates intracellular replication in murinemacrophages. J. Bacteriol., 182, 3482–3489.
10. Benkovic,S.J., Baker,S.J., Alley,M.R., Woo,Y.H., Zhang,Y.K.,Akama,T., Mao,W., Baboval,J., Rajagopalan,P.T., Wall,M. et al.(2005) Identification of borinic esters as inhibitors of bacterialcell growth and bacterial methyltransferases, CcrM and MenH.J. Med. Chem., 48, 7468–7476.
11. Urig,S., Gowher,H., Hermann,A., Beck,C., Fatemi,M.,Humeny,A. and Jeltsch,A. (2002) The Escherichia coli dam DNAmethyltransferase modifies DNA in a highly processive reaction.J. Mol. Biol., 319, 1085–1096.
12. Peterson,S.N. and Reich,N.O. (2006) GATC flanking sequencesregulate Dam activity: evidence for how Dam specificity mayinfluence pap expression. J. Mol. Biol., 355, 459–472.
13. Zinoviev,V.V., Evdokimov,A.A., Malygin,E.G., Schlagman,S.L.and Hattman,S. (2003) Bacteriophage T4 Dam DNA-(N6-adenine)-methyltransferase. Processivity and orientation tothe methylation target. J. Biol. Chem., 278, 7829–7833.
14. Berdis,A.J., Lee,I., Coward,J.K., Stephens,C., Wright,R.,Shapiro,L. and Benkovic,S.J. (1998) A cell cycle-regulated adenine
DNA methyltransferase from Caulobacter crescentus processivelymethylates GANTC sites on hemimethylated DNA. Proc. NatlAcad. Sci. USA, 95, 2874–2879.
15. Roth,M. and Jeltsch,A. (2000) Biotin-avidin microplate assay forthe quantitative analysis of enzymatic methylation of DNA byDNA methyltransferases. Biol. Chem., 381, 269–272.
16. Zweiger,G., Marczynski,G. and Shapiro,L. (1994) A CaulobacterDNA methyltransferase that functions only in the predivisionalcell. J. Mol. Biol., 235, 472–485.
17. Stephens,C.M., Zweiger,G. and Shapiro,L. (1995) Coordinate cellcycle control of a Caulobacter DNA methyltransferase and theflagellar genetic hierarchy. J. Bacteriol., 177, 1662–1669.
18. Stephens,C., Reisenauer,A., Wright,R. and Shapiro,L. (1996) Acell cycle-regulated bacterial DNA methyltransferase is essentialfor viability. Proc. Natl Acad. Sci. USA, 93, 1210–1214.
19. Wright,R., Stephens,C., Zweiger,G., Shapiro,L. and Alley,M.R.(1996) Caulobacter Lon protease has a critical role in cell-cyclecontrol of DNA methylation. Genes Dev., 10, 1532–1542.
20. Reisenauer,A., Quon,K. and Shapiro,L. (1999) The CtrA responseregulator mediates temporal control of gene expression during theCaulobacter cell cycle. J. Bacteriol., 181, 2430–2439.
21. Shier,V.K., Hancey,C.J. and Benkovic,S.J. (2001) Identification ofthe active oligomeric state of an essential adenine DNAmethyltransferase from Caulobacter crescentus. J. Biol. Chem.,276, 14744–14751.
22. Horton,J.R., Liebert,K., Hattman,S., Jeltsch,A. and Cheng,X.(2005) Transition from nonspecific to specific DNA interactionsalong the substrate-recognition pathway of dammethyltransferase. Cell, 121, 349–361.
23. Horton,J.R., Liebert,K., Bekes,M., Jeltsch,A. and Cheng,X.(2006) Structure and substrate recognition of the Escherichia coliDNA adenine methyltransferase. J. Mol. Biol., 358, 559–570.
Nucleic Acids Research, 2011 9
by guest on June 6, 2016http://nar.oxfordjournals.org/
Dow
nloaded from




















