
In vitro activity of nicotinamide/antileishmanial drug combinations
6
In vitro activity of nicotinamide/antileishmanial drug combinations Elodie Gazanion a , Baptiste Vergnes a , Marie Seveno a , Deborah Garcia a , Bruno Oury a , Khatima Ait-Oudhia c , Ali Ouaissi b , Denis Sereno a, ⁎ a IRD (Institut de Recherche pour le Développement), UR16 « Caractérisation et Contrôle des Populations de Vecteurs », Montpellier, France b INSERM CNRS UMR5235, Université Montpellier 2, France c Ecole Nationale Supérieure Vétérinaire, BP 161 Hassan Badi El-Harrach Alger, Algeria abstract article info Article history: Received 20 November 2009 Received in revised form 4 September 2010 Accepted 17 September 2010 Available online 25 September 2010 Keywords: Nicotinamide Antimonials Leishmania Drug combinations To improve the management of leishmaniasis, new drugs and/or alternative therapeutic strategies are required. Combination therapy of antileishmanial drugs is currently considered as one of the most rational approaches to lower treatment failure rate and limit drug resistance spreading. Nicotinamide (NAm), also known as vitamin B3 that is already is used in human therapy, exerts in vitro antileishmanial activity. Drug combination studies, performed on L. infantum axenic amastigotes, revealed that NAm significantly improves the antileishmanial activity of trivalent antimony in a synergistic manner while it shows additive activity with amphotericin B and slightly antagonizes pentamidine activity. NAm also significantly increases the toxicity of pentavalent antimony against the intracellular forms of L. infantum, L. amazonensis and L. braziliensis. The potential of NAm to be used as adjuvant during leishmaniasis chemotherapy is further discussed. © 2010 Elsevier Ireland Ltd. All rights reserved. 1. Introduction Leishmaniases are a complex of diseases caused by at least 17 different species of protozoan parasites belonging to the genus Leishmania. They are significant causes of morbidity and mortality in 88 countries around the world. The clinical manifestations range from self-healing cutaneous and mucocutaneous skin ulcers to a fatal visceral form, named visceral leishmaniasis or kala-azar. Basic treatment for all forms of leishmaniases consists of the administration of pentavalent antimony SbV (sodium stibogluconate Pentostam or meglumine antimoniate Glucantime) or more recently the liposomal formulation of amphotericin B in endemic Mediterranean countries and for immunocompromised patients [1]. Antimony unresponsiveness, reported in several parts of the world [2], involves various mechanisms, including host factors [3], variations in drug delivery [4] and decreases in parasite drug susceptibility [5–7]. Alternative drug options include amphotericin B, pentamidine, miltefosine, paromomycin and allopuri- nol. Their use is however limited because of their toxicity or declined efficacy [8]. Combination therapy between commercially available drugs that aimed to reduce cost, toxicity and duration of treatments, represents a promising rational alternative. Nicotinamide (NAm) and nicotinic acid are both NAD + precursors of the NAD + salvage biosynthetic pathway. NAm alone [9] or combined with immunomod- ulatory complexes [10] bears in vitro antileishmanial activity but it also exerts antiplasmodial activity at concentrations close to those found to be leishmaniostatic [11]. This vitamin is safe even administered at high dosage (6 g/day) in human [12]. As it is particularly cheap to produce, to distribute and deliver to human, this vitamin that can be orally administered, may thus be a valuable and rational candidate for combination therapy in areas where leishmaniases are endemic. In this paper, we used different approaches to investigate the capacity of NAm to improve in vitro the antileishmanial activity of clinically relevant antileishmanial compounds against axenic and intramacrophagic amastigotes of various Leishmania species. 2. Materials and methods 2.1. Materials Meglumine antimoniate (Glucantime, batch number 793-F) was supplied by Rhône Poulenc Specia. Nicotinamide, nicotinic acid, pentamidine, amphotericin B, potassium antimonyl tartrate trihy- drate (SbIII), phorbol myristate acetate (PMA), 3-(4,5-dimethlythin- zol-2-yl)-2,5-diphenyltetrazolium (MTT) were provided by Sigma. Propidium iodide was supplied by Molecular Probes. Penicillin, streptomycin and fetal calf serum (FCS) were provided by Lonza. 2.2. Parasites and cultures Axenically grown amastigotes of L. infantum (MHOM/MA/67/ ITMAP-263) were maintained in 25-cm 2 flasks, at 37 °C with 5% CO 2 Parasitology International 60 (2011) 19–24 ⁎ Corresponding author. IRD UR16, Institut National d'Hygiene, Av Ibn Batouba, 10000 Rabat Agdal, Maroc. E-mail address: [email protected] (D. Sereno). 1383-5769/$ – see front matter © 2010 Elsevier Ireland Ltd. All rights reserved. doi:10.1016/j.parint.2010.09.005 Contents lists available at ScienceDirect Parasitology International journal homepage: www.elsevier.com/locate/parint
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of In vitro activity of nicotinamide/antileishmanial drug combinations

Parasitology International 60 (2011) 19–24
Contents lists available at ScienceDirect
Parasitology International
j ourna l homepage: www.e lsev ie r.com/ locate /par in t
In vitro activity of nicotinamide/antileishmanial drug combinations
Elodie Gazanion a, Baptiste Vergnes a, Marie Seveno a, Deborah Garcia a, Bruno Oury a, Khatima Ait-Oudhia c,Ali Ouaissi b, Denis Sereno a,⁎a IRD (Institut de Recherche pour le Développement), UR16 « Caractérisation et Contrôle des Populations de Vecteurs », Montpellier, Franceb INSERM CNRS UMR5235, Université Montpellier 2, Francec Ecole Nationale Supérieure Vétérinaire, BP 161 Hassan Badi El-Harrach Alger, Algeria
⁎ Corresponding author. IRD UR16, Institut Nationa10000 Rabat Agdal, Maroc.
E-mail address: [email protected] (D. Sereno).
1383-5769/$ – see front matter © 2010 Elsevier Irelanddoi:10.1016/j.parint.2010.09.005
a b s t r a c t
a r t i c l e i n f oArticle history:Received 20 November 2009Received in revised form 4 September 2010Accepted 17 September 2010Available online 25 September 2010
Keywords:NicotinamideAntimonialsLeishmaniaDrug combinations
To improve the management of leishmaniasis, new drugs and/or alternative therapeutic strategies arerequired. Combination therapy of antileishmanial drugs is currently considered as one of the most rationalapproaches to lower treatment failure rate and limit drug resistance spreading. Nicotinamide (NAm), alsoknown as vitamin B3 that is already is used in human therapy, exerts in vitro antileishmanial activity. Drugcombination studies, performed on L. infantum axenic amastigotes, revealed that NAm significantlyimproves the antileishmanial activity of trivalent antimony in a synergistic manner while it shows additiveactivity with amphotericin B and slightly antagonizes pentamidine activity. NAm also significantlyincreases the toxicity of pentavalent antimony against the intracellular forms of L. infantum, L. amazonensisand L. braziliensis. The potential of NAm to be used as adjuvant during leishmaniasis chemotherapy isfurther discussed.
l d'Hygiene, Av Ibn Batouba,
Ltd. All rights reserved.
© 2010 Elsevier Ireland Ltd. All rights reserved.
1. Introduction
Leishmaniases are a complex of diseases caused by at least 17different species of protozoan parasites belonging to the genusLeishmania. They are significant causes of morbidity and mortality in88 countries around the world. The clinical manifestations range fromself-healing cutaneous andmucocutaneous skin ulcers to a fatal visceralform, named visceral leishmaniasis or kala-azar. Basic treatment for allforms of leishmaniases consists of the administration of pentavalentantimony SbV (sodium stibogluconate Pentostam or meglumineantimoniate Glucantime) or more recently the liposomal formulationof amphotericin B in endemic Mediterranean countries and forimmunocompromised patients [1]. Antimony unresponsiveness,reported in several parts of theworld [2], involves variousmechanisms,including host factors [3], variations in drug delivery [4] and decreasesin parasite drug susceptibility [5–7]. Alternative drug options includeamphotericin B, pentamidine, miltefosine, paromomycin and allopuri-nol. Their use is however limited because of their toxicity or declinedefficacy [8]. Combination therapy between commercially availabledrugs that aimed to reduce cost, toxicity and duration of treatments,represents a promising rational alternative. Nicotinamide (NAm) andnicotinic acid are both NAD+ precursors of the NAD+ salvagebiosynthetic pathway. NAm alone [9] or combined with immunomod-
ulatory complexes [10] bears in vitro antileishmanial activity but it alsoexerts antiplasmodial activity at concentrations close to those found tobe leishmaniostatic [11]. This vitamin is safe even administered at highdosage (6 g/day) in human [12]. As it is particularly cheap to produce, todistribute and deliver to human, this vitamin that can be orallyadministered, may thus be a valuable and rational candidate forcombination therapy in areas where leishmaniases are endemic. In thispaper, we used different approaches to investigate the capacity of NAmto improve in vitro the antileishmanial activity of clinically relevantantileishmanial compounds against axenic and intramacrophagicamastigotes of various Leishmania species.
2. Materials and methods
2.1. Materials
Meglumine antimoniate (Glucantime, batch number 793-F) wassupplied by Rhône Poulenc Specia. Nicotinamide, nicotinic acid,pentamidine, amphotericin B, potassium antimonyl tartrate trihy-drate (SbIII), phorbol myristate acetate (PMA), 3-(4,5-dimethlythin-zol-2-yl)-2,5-diphenyltetrazolium (MTT) were provided by Sigma.Propidium iodide was supplied by Molecular Probes. Penicillin,streptomycin and fetal calf serum (FCS) were provided by Lonza.
2.2. Parasites and cultures
Axenically grown amastigotes of L. infantum (MHOM/MA/67/ITMAP-263) were maintained in 25-cm2
flasks, at 37 °C with 5% CO2

20 E. Gazanion et al. / Parasitology International 60 (2011) 19–24
by weekly subpassages in a MAA20 cell-free medium supplementedwith 20% heat inactivated fetal calf serum [13]. Promastigote forms ofL. amazonensis (MHOM/BR/76/LTB-012) and L. braziliensis (MHOM/BR/75/M2904) were maintained at 26 °C in SDM-79 mediumsupplemented with 10% FCS [14].
2.3. Flow-cytometry analysis
Viability and growth of NAm-treated amastigotes were deter-mined by flow cytometry analysis. Briefly, a sample of cells (5 μl) wastaken daily from the culture and diluted into 500 μl of PBS (pH 7.2).Propidium iodide was added at a final concentration of 1 μg/ml andcells were analyzed on a flow cytometer (Facsalibur™; BectonDickinson) for 52 s. The parasite concentration was determinedusing standard curves where parasite concentrations were plottedas a function of mean number of cells counted by the cytometer after52 s [15].
2.4. DNA fragmentation analysis and quantification
Qualitative analysis of DNA fragmentation was performed byagarose gel electrophoresis. Amastigotes were seeded at5×106 cells/ml and grown for 24 h. Drugs were added to theculture medium and parasites were further incubated for 5 days.Total DNA was purified by the phenol-chloroform extractionmethod. Briefly, cells were pelleted and resuspended in lysis buffer(10 mM Tris–HCl, 5 mM EDTA, 0.5% SDS, 200 mM NaCl, pH 8). Themixture was then submitted to proteolysis in the presence of200 μg/ml proteinase K (Sigma) for 2 h at 60 °C. One volume ofphenol/chloroform/isoamylic alcohol (25:24:1) was added andsamples were mixed and centrifuged for 20 min at 10000×g.DNA from the aqueous phase was precipitated overnight at −20 °Cin the presence of 0.3 M NaCl (final concentration) and 2.5volumes of cold 100% ethanol. The DNA pellet was then washedtwice with cold 70% ethanol, air dried, dissolved in distilled waterand treated with 0.3 mg/ml RNase A (Invitrogen) for 1 h at 37 °C.Cell cycle analysis leading to the appearance and quantification ofDNA degradation was performed according to the methoddescribed by Alzate et al. [16] with small modifications. Afterincubation in cold 70% ethanol, cells were washed and resuspendedin 400 μl of PBS/50 mM EDTA/100 μg/ml RNase A. After the 30 minincubation at 37 °C, propidium iodide was added at a finalconcentration of 10 μg/ml and cells were immediately analyzedon the flow cytometer.
2.5. Anti-parasitic activity of nicotinamide/drug combinations anddetermination of the Combination Index (CI)
The synergy, additivity or antagonism between NAm andstandard antileishmanial compounds (trivalent antimony SbIII,pentamidine, amphotericin B) were determined for L. infantumaxenic amastigotes using the median effect method and themultiple drug-effect equation described by Chou and Talalay [17].The EC50, EC75 and EC90 of each compound alone or in combinationwere firstly determined using the MTT micromethod as previouslydescribed [13]. Briefly, parasites were seeded at 1×106 cells/ml in96-well plates and grown for 3 days in the presence of serialdilutions of drugs alone or in combination with NAm. Theinteraction between the combinations was then assessed by thecalculation of the Combination Index (CI) at 50%, 75% and 90%inhibition of cell growth and was defined as follows: CI 50%=[50%Effective Concentration (EC50) of partner drug in combination/EC50
of partner drug alone]+[EC50 of nicotinamide in combination/EC50
of nicotinamide alone]. CI values above 1.1 indicate antagonisticeffect, from 0.9 to 1.1 are indicative of an additive effect, 0.7 to 0.9
are moderately synergistic, 0.3 to 0.7 are synergistic and b0.3indicate strong synergism.
2.6. Drug efficacy assay on intracellular amastigotes
The effect of NAm/drug combinations on Leishmania spp. growthin a human leukemia monocyte cell line (THP-1 cells) was evaluatedaccording to the method previously described [18]. THP-1 cells werecultured in RPMI 1640 medium supplemented with 10% FCS, 2 mMglutamine, 100 IU of penicillin/ml, and 100 μg of streptomycin/ml.THP-1 cells in the log phase of growth were differentiated byincubation for 2 days in medium containing 20 ng/ml of PMA.Macrophages were then infected with stationary-phase L. infantumamastigotes at a parasite:macrophage ratio of 5:1 for 4 h at 37 °C with5% CO2. Infection with L. braziliensis or L. amazonensis stationary-phase promastigotes were performed at a parasite:macrophage ratioof 50:1 for 6 h at 34 °C with 5% CO2. Non-internalized parasites wereremoved and drugs were added to themedia. After 3 days for trivalentantimony (SbIII) or 5 days for meglumine antimoniate (SbV), infectedcells were washed and stained with Giemsa. To determine theefficiency of drug combinations against intracellular amastigotes theparasitic index (PI) was calculated as follow: % PI=(percentage ofinfected macrophages×number of intracellular parasites/macro-phage in treated wells)/(percentage of infected macrophages×num-ber of intracellular parasites/macrophage in untreated wells)×100.Alternatively, the cytotoxic effect of each drug alone or in combinationon non-infected macrophages was ascertained using trypan bluecoloration and after a 3-day period incubation for NAm/SbIIIcombination or a 5-day period incubation for NAm/SbV combination.
2.7. Statistical analyses
The efficiency of NAm/drug combinations against intracellularamastigotes was compared to the activity obtained with the drugsalone. Statistical analyses were performed with the Student's t testand a Pb0.05 considered statistically significant. Graphical illustra-tions and statistical analysis were performed using Prism software(GraphPad Software, Version 5).
3. Results
3.1. Toxicity of nicotinamide (NAm) towards axenic L. infantumamastigotes
In accordance with our previous results [9], NAm exerts aleishmaniostatic effect against axenic L. infantum amastigotes(Fig. 1A). The determination of EC50, EC75 and EC90 confirms theseobservations (Table 1). Many, if not all, antileishmanial drugs(pentamidine, amphotericin B, antimony and miltefosine) areknown to induce Leishmania death by a process reminiscent ofapoptosis [19]. DNA status analysis from NAm-treated amastigotesshowed that high concentrations (80 mM) induce genomic DNAdegradation into fragments of about 200 bp that are close to theoligonucleosome-sized fragments observed during apoptosis(Fig. 1B). Moreover, the quantification of cell death by cell cycleanalysis showed that parasites treated with 5 mM or 20 mM of NAmdisplayed only 10% and 13% of dead cells respectively, as monitored bythe occurrence of a hypodiploid population (M1). In concordancewith the DNA degradation visualized on agarose gel, this proportionrises 70% when 80 mM of NAm were added to the culture medium(Fig. 1C). These observations demonstrate that below 20 mM, NAmexerts a leishmaniostatic activity and induces cell death when higherconcentrations of NAm are used.

A B
C
Fig. 1. Antileishmanial activity of NAm. (A) L. infantum amastigote forms were incubated with increasing concentrations of NAm, viability was ascertained daily by FACS analysis.Results are the means of triplicate experiments. (B) DNA status of parasites treated with 0, 5, 20 or 80 mM NAm was visualized after agarose gel electrophoresis. Note thatoligonucleosomal DNA fragmentation was observed in parasites treated with 80 mM of NAm. (C) Measurement of DNA content of parasites treated with increasing concentrations ofNAm by FACS analysis. DNA degradation is monitored by the occurrence of the hypodiploid cell population (M1). The M2 population represents cellular aggregates.
21E. Gazanion et al. / Parasitology International 60 (2011) 19–24
3.2. Activity of NAm/antileishmanial drug combinations against L.infantum axenic amastigotes
We next tested the capacity of NAm to improve the cytotoxicity ofsome standard antileishmanial drugs (potassium antimony tartrate(SbIII), amphotericin B and pentamidine). Concentrations used forcombination studies were determined according to the EC50, EC75 andEC90 values of each compound (Table 1). Combination experimentswere performed as described in material and method section. Dose–response curves of pentamidine, amphotericin B or SbIII in thepresence of increasing concentrations of NAm are displayed in Fig. 2A.Combination index values (CI50%, CI75% and CI90%) calculated from
Table 1Susceptibility of axenic L. infantum amastigotes and THP-1 cell line to antileishmanialcompounds. EC50, EC75 and EC90 of each compound against axenic L. infantumamastigotes and the THP-1 cell line. Results are the mean of two to four independentexperiments each performed in triplicate.
Cell type Drugs EC50 EC75 EC90
AxenicL. infantum
Potassiumantimonytartrate (SbIII)
8.3±1.1 μg/ml 13.9±1.5 μg/ml 18.6±0.5 μg/ml
Pentamidine 1.0±0.2 μM 1.6±0.2 μM 2.5±0.7 μMAmphotericin B 0.2±0.1 μM 0.3±0.1 μM 0.4±0.1 μMNicotinamide 6.9±1.1 mM 14.1±2.5mM 19.3±0.9 mM
THP-1 cellline
Nicotinamide 20.1±3.3 mM 37.6±0.3 mM 50.2±4.6 mMPotassiumantimonytartrate (SbIII)
8.0±2.1 μg/ml ND ND
Glucantime®(SbV)
N100 μg/ml ND ND
these experiments, indicate that NAm antagonizes the effect ofpentamidine (CIN1.1), whereas the interaction between NAm andamphotericin B is additive (CI values close to 1) (Table 2). Interest-ingly, with CI values ranging from 0.66 to 0.79, the NAm/SbIIIcombination was found to be moderately synergistic, in all the fixedconcentrations used (Fig. 2A and Table 2). Moreover, this synergisticeffect appears to be restricted to the amide form of vitamin B3 sinceno synergism was found when NAm was replaced by its acidderivative, nicotinic acid (Table 2). These results were furtherconfirmed after qualitative and quantitative analyses of the DNAstatus from parasites treated with NAm or SbIII alone or incombination. As shown in Fig. 2B, each molecule alone did not affectparasite DNA integrity. In contrast, the addition of NAm (15 mM) toSbIII (10 μg/ml) in the culture medium led to a strong DNAdegradation. DNA status analysis also confirmed that the NAm/SbIIIcombination is approximately 3-fold more potent to induce L.infantum amastigote cell death than each of these molecules alone(Fig. 2C).
3.3. Activity of NAm/antimony combination against intracellularamastigotes of various Leishmania species
To further document the capacity of NAm to improve theantileishmanial activity of SbIII, we carried out experiments againstintracellular amastigotes. Toxicity of each compound against non-infected differentiated THP-1 monocytes was ascertained. We foundthat nicotinamide is toxic for THP-1 cells (EC50 of 20.1±3.3 mM; seeTable 1) at concentrations close to those previously reported for othermammalian cell types [20,21]. In our conditions, SbIII is highly toxicfor THP-1 cells with an EC50 of 8.0±2.1 μg/ml, contrary to SbV

CB
A
Fig. 2. Cytotoxic activity of NAm/drug combinations against axenic amastigotes. (A) Cell growth curves of L. infantum amastigotes treated with pentamidine, amphotericin B or SbIIIin absence of NAm (●) or in presence of 5 mMNAm (■), 10 mMNAm (♦) and 20 mMNAm (▲). Results are expressed as the mean of two independent experiments each performedin triplicate. (B) DNA integrity of parasites incubated with 15 mM NAm (N15), 10 μg/ml SbIII (S10) or both molecules (N15+S10). DNA was extracted by the phenol-chloroformmethod and visualized after agarose gel electrophoresis. (C) Quantification of DNA degradation after incubation of parasites with NAm or SbIII alone and in combination. DNAdegradation is monitored by the occurrence of the hypodiploid cell population (M1). The M2 population represents cellular aggregates.
22 E. Gazanion et al. / Parasitology International 60 (2011) 19–24
(Table 1). Two sets of experiments (1.25 μg/ml SbIII combined with2.5 mM NAm and 2.5 μg/ml SbIII with 5 mM NAm) were thenperformed. Toxicity of these combinations against non-infectedmacrophages was recorded in parallel and is given in Fig. 3 (lowerpanel). In both experiments, we found that the addition of NAm toSbIII improves significantly its capacity to reduce the intracellularproliferation of L. infantum (Fig. 3, higher panel). During theseexperiments, we confirmed that NAm alone was not toxic for the hostcells. However, we recorded a 14% reduction of THP-1 cell viability atthe highest SbIII concentration tested (Fig. 3, lower panel). Pentava-lent antimonial (SbV) is the form delivered during treatment of allforms of leishmaniasis. We thus investigated the antileishmanialactivity of some NAm/SbV (Glucantime) combinations againstintracellular amastigotes after a 5-day period incubation. To thisaim, we firstly determined the SbV concentration required to reduce
Table 2Effect of NAm or nicotinic acid combination with various antileishmanial compounds.Combination Index at 50%, 75% and 90% inhibition of growth (CI50% CI75% and CI90%respectively) were determined from dose–response curve experiments performed onaxenic L. infantum amastigotes according to the Chou and Talalay method.
CombinationIndex (CI)
NAm+Pentamidine
NAm+Amphotericin B
NAm+SbIII Nicotinicacid+SbIII
CI50% 1.13±0.33 0.94±0.04 0.79±0.06 1.01±0.16Effect Antagonistic Additive Moderately
synergisticAdditive
CI75% 1.15±0.08 1.00±0.09 0.66±0.12 NDEffect Antagonistic Additive SynergisticCI90% 1.26±0.10 1.00±0.06 0.67±0.06 NDEffect Antagonistic Additive Synergistic
Parasitic Index by about 50% and then tested the capacity of NAm(1.25 mM and 2.5 mM) to significantly improve the SbV activity. Theaddition of NAm to 10 μg/ml SbV significantly (Pb0.001) improves itsefficiency (Fig. 4A, upper panel) whatever the NAm concentrationused and without adverse effects for the host cell (Fig. 4A, lowerpanel). Remarkably, the NAm/SbV combinations are more efficientthan the use of 50 μg/ml SbV alone (data not shown). We thenevaluated the capacity of a single NAm dose (5 mM) to significantlyimprove SbV activity against L. amazonensis and L. braziliensisparasites, responsible for diffuse cutaneous and mucocutaneousleishmaniasis. In both cases, the addition of 5 mM NAm significantlyenhanced the intracellular SbV-mediated killing, as compared to NAmor SbV alone (Fig. 4B and C).
4. Discussion
The standard regimen for visceral leishmaniasis treatment consistsof intramuscular or intravenous injection of 20 mg SbV/kg/day for28 days, the treatment is lengthened to 40–60 days in the case ofrelapse [3,22]. However, these recommendations are still unsatisfac-tory since unresponsiveness and toxicity still occur [22]. While newantileishmanial compounds are continuously in development, drugcombination is currently considered as one of the most rationalalternatives to improve treatment efficiency, prevent drug resistanceand reduce treatment duration [3,23]. The use of vitamins to treatleishmaniasis has been raised following the relationship that has beenestablished between malnutrition and parasitic infections [24,25].We have recently demonstrated that nicotinamide (vitamin B3)exhibits a cytostatic activity against L. infantum parasites [9]. Here, we
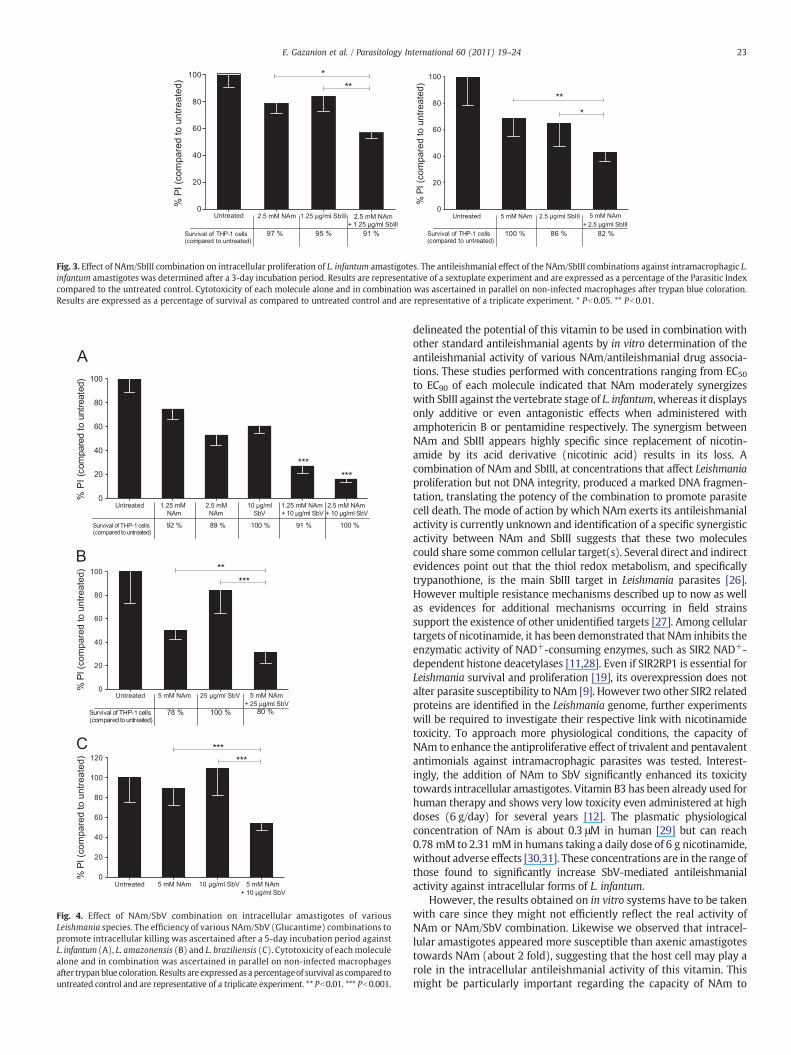
Fig. 3. Effect of NAm/SbIII combination on intracellular proliferation of L. infantum amastigotes. The antileishmanial effect of the NAm/SbIII combinations against intramacrophagic L.infantum amastigotes was determined after a 3-day incubation period. Results are representative of a sextuplate experiment and are expressed as a percentage of the Parasitic Indexcompared to the untreated control. Cytotoxicity of each molecule alone and in combination was ascertained in parallel on non-infected macrophages after trypan blue coloration.Results are expressed as a percentage of survival as compared to untreated control and are representative of a triplicate experiment. * Pb0.05. ** Pb0.01.
Fig. 4. Effect of NAm/SbV combination on intracellular amastigotes of variousLeishmania species. The efficiency of various NAm/SbV (Glucantime) combinations topromote intracellular killing was ascertained after a 5-day incubation period againstL. infantum (A), L. amazonensis (B) and L. braziliensis (C). Cytotoxicity of eachmoleculealone and in combination was ascertained in parallel on non-infected macrophagesafter trypanblue coloration. Results are expressedas apercentageof survival as compared tountreated control and are representative of a triplicate experiment. ** Pb0.01. *** Pb0.001.
23E. Gazanion et al. / Parasitology International 60 (2011) 19–24
delineated the potential of this vitamin to be used in combination withother standard antileishmanial agents by in vitro determination of theantileishmanial activity of various NAm/antileishmanial drug associa-tions. These studies performed with concentrations ranging from EC50to EC90 of each molecule indicated that NAm moderately synergizeswith SbIII against the vertebrate stage of L. infantum, whereas it displaysonly additive or even antagonistic effects when administered withamphotericin B or pentamidine respectively. The synergism betweenNAm and SbIII appears highly specific since replacement of nicotin-amide by its acid derivative (nicotinic acid) results in its loss. Acombination of NAm and SbIII, at concentrations that affect Leishmaniaproliferation but not DNA integrity, produced a marked DNA fragmen-tation, translating the potency of the combination to promote parasitecell death. The mode of action by which NAm exerts its antileishmanialactivity is currently unknown and identification of a specific synergisticactivity between NAm and SbIII suggests that these two moleculescould share some common cellular target(s). Several direct and indirectevidences point out that the thiol redox metabolism, and specificallytrypanothione, is the main SbIII target in Leishmania parasites [26].However multiple resistance mechanisms described up to now as wellas evidences for additional mechanisms occurring in field strainssupport the existence of other unidentified targets [27]. Among cellulartargets of nicotinamide, it has been demonstrated that NAm inhibits theenzymatic activity of NAD+-consuming enzymes, such as SIR2 NAD+-dependent histone deacetylases [11,28]. Even if SIR2RP1 is essential forLeishmania survival and proliferation [19], its overexpression does notalter parasite susceptibility to NAm [9]. However two other SIR2 relatedproteins are identified in the Leishmania genome, further experimentswill be required to investigate their respective link with nicotinamidetoxicity. To approach more physiological conditions, the capacity ofNAm to enhance the antiproliferative effect of trivalent and pentavalentantimonials against intramacrophagic parasites was tested. Interest-ingly, the addition of NAm to SbV significantly enhanced its toxicitytowards intracellular amastigotes. Vitamin B3 has been already used forhuman therapy and shows very low toxicity even administered at highdoses (6 g/day) for several years [12]. The plasmatic physiologicalconcentration of NAm is about 0.3 μM in human [29] but can reach0.78 mM to 2.31 mM in humans taking a daily dose of 6 g nicotinamide,without adverse effects [30,31]. These concentrations are in the range ofthose found to significantly increase SbV-mediated antileishmanialactivity against intracellular forms of L. infantum.
However, the results obtained on in vitro systems have to be takenwith care since they might not efficiently reflect the real activity ofNAm or NAm/SbV combination. Likewise we observed that intracel-lular amastigotes appeared more susceptible than axenic amastigotestowards NAm (about 2 fold), suggesting that the host cell may play arole in the intracellular antileishmanial activity of this vitamin. Thismight be particularly important regarding the capacity of NAm to

24 E. Gazanion et al. / Parasitology International 60 (2011) 19–24
significantly improve SbV activity. Pentavalent antimony that isknown to be poorly toxic for axenic amastigotes at concentrationspotentially achieved in sera of patients during chemotherapy (about20 μg/ml) displayed a strong, although host cell type dependenttoxicity against intramacrophagic amastigotes, with the EC50 rangingfrom 2 μg/ml to 29 μg/ml [32]. Thus, to more precisely define theantileishmanial activity of NAm and NAm/SbV combinations, it will beessential to perform in vitro tests using a more relevant host cellsystem, like peripheral blood monocyte-derived macrophages. Thisinformation will also help to shed light on the potential of a vitaminB3 supplement to decrease the susceptibility towards Leishmaniainfection in vivo. When considering in vivo administration of NAm, thecapacity of the host to shift NAm in its metabolic pathway(s) prior tosynergize with antimony might limit the use of this vitamin. Finally,immunomodulatory effects of NAm reported elsewhere in individuals[33,34] may also indirectly influence the efficiency of the NAm/SbVcombination.
In conclusion, our results demonstrated the existence of synergismbetween NAm and SbIII against Leishmania amastigotes and that NAmimproves significantly the antileishmanial activity of SbIII and SbVagainst intracellular parasites at concentrations reached in humans.Thus, NAm will be an interesting molecule aimed to reduce costassociated to antimonial treatment in low-income areas.
Acknowledgments
We are grateful to Dr. Francine Pratlong and Pr Jean-Pierre Dedetfrom the Centre National de Référence des Leishmania for providing L.amazonensis and L. braziliensis strains. This study was supported bygrant (AAP2008-Nicomed) from the Expertise and ValorisationDepartment of IRD (DEV). E.G. is supported by a Higher Educationand Research Ministerial grant (France). We thank Caroline Harris forthe English revision of the manuscript.
References
[1] Gradoni L, Soteriadou K, Louzir H, Dakkak A, Toz SO, Jaffe C, et al. Drug regimens forvisceral leishmaniasis in Mediterranean countries. Trop Med Int Health 2008;13(10):1272–6.
[2] Sundar S, Rai M. Advances in the treatment of leishmaniasis. Curr Opin Infect Dis2002;15(6):593–8.
[3] Croft SL, Sundar S, Fairlamb AH. Drug resistance in leishmaniasis. Clin MicrobiolRev 2006;19(1):111–26.
[4] Nieto J, Alvar J, Mullen AB, Carter KC, Rodríguez C, San Andrés MI, et al.Pharmacokinetics, toxicities, and efficacies of sodium stibogluconate formulationsafter intravenous administration in animals. Antimicrob Agents Chemother2003;47(9):2781–7.
[5] Faraut-Gambarelli F, Piarroux R, Deniau M, Giusiano B, Marty P, Michel G, et al. Invitro and in vivo resistance of Leishmania infantum to meglumine antimoniate: astudy of 37 strains collected from patients with visceral leishmaniasis. AntimicrobAgents Chemother 1997;41(4):827–30.
[6] Rojas R, Valderrama L, Valderrama M, Varona MX, Ouellette M, Saravia NG.Resistance to antimony and treatment failure in human Leishmania (Viannia)infection. J Infect Dis 2006;193(10):1375–83.
[7] Lira R, Sundar S, Makharia A, Kenney R, Gam A, Saraiva E, et al. Evidence that thehigh incidence of treatment failures in Indian kala-azar is due to the emergence ofantimony-resistant strains of Leishmania donovani. J Infect Dis 1999;180(2):564–7.
[8] Sundar S, Chatterjee M. Visceral leishmaniasis – current therapeutic modalities.Indian J Med Res 2006;123(3):345–52.
[9] Sereno D, Alegre AM, Silvestre R, Vergnes B, Ouaissi A. In vitro antileishmanialactivity of nicotinamide. Antimicrob Agents Chemother 2005;49(2):808–12.
[10] Al-Mulla Hummadi Y, Najim R, Al-Bashir N. Leishmania major and Leishmaniatropica: I the in vitro effects of an immunomodulator, S2-Complex. Exp Parasitol2005;111(1):47–54.
[11] Prusty D, Mehra P, Srivastava S, Shivange AV, Gupta A, Roy N, et al. Nicotinamideinhibits Plasmodium falciparum Sir2 activity in vitro and parasite growth. FEMSMicrobiol Lett 2008;282(2):266–72.
[12] Knip M, Douek IF, Moore WP, Gillmor HA, McLean AE, Bingley PJ, et al. Safety ofhigh-dose nicotinamide: a review. Diabetologia 2000;43(11):1337–45.
[13] Sereno D, Lemesre JL. Axenically cultured amastigote forms as an in vitro modelfor investigation of antileishmanial agents. Antimicrob Agents Chemother1997;41(5):972–6.
[14] Brun R, Schonenberger M. Cultivation and in vitro cloning of procyclic cultureforms of Trypanosoma brucei in a semi-defined medium. Acta Trop 1979;36(3):289–92.
[15] Vergnes B, Sereno D, Tavares J, Cordeiro-da-Silva A, Vanhille L, Madjidian-SerenoN, et al. Targeted disruption of cytosolic SIR2 deacetylase discloses its essentialrole in Leishmania survival and proliferation. Gene 2005;363:85–96.
[16] Alzate JF, Arias AA, Moreno-Mateos D, Alvarez-Barrientos A, Jiménez-Ruiz A.Mitochondrial superoxide mediates heat-induced apoptotic-like death in Leish-mania infantum. Mol Biochem Parasitol 2007;152(2):192–202.
[17] Chou TC, Talalay P. Quantitative analysis of dose-effect relationships: thecombined effects of multiple drugs or enzyme inhibitors. Adv Enzyme Regul1984;22:27–55.
[18] Sereno D, Roy G, Lemesre JL, Papadopoulou B, Ouellette M. DNA transformation ofLeishmania infantum axenic amastigotes and their use in drug screening.Antimicrob Agents Chemother 2001;45(4):1168–73.
[19] Shaha C. Apoptosis in Leishmania species & its relevance to disease pathogenesis.Indian J Med Res 2006;123(3):233–44.
[20] Chong ZZ, Lin SH, Maiese K. The NAD+ precursor nicotinamide governs neuronalsurvival during oxidative stress through protein kinase B coupled to FOXO3a andmitochondrial membrane potential. J Cereb Blood Flow Metab 2004;24(7):728–43.
[21] Crowley CL, Payne CM, Bernstein H, Bernstein C, Roe D. The NAD+precursors,nicotinic acid and nicotinamide protect cells against apoptosis induced by amultiple stress inducer, deoxycholate. Cell Death Differ 2000;7(3):314–26.
[22] Jha TK. Drug unresponsiveness & combination therapy for kala-azar. Indian J MedRes 2006;123(3):389–98.
[23] Bryceson A. A policy for leishmaniasis with respect to the prevention and controlof drug resistance. Trop Med Int Health 2001;6(11):928–34.
[24] Cerf BJ, Jones TC, Badaro R, Sampaio D, Teixeira R, Johnson JrWD. Malnutrition as arisk factor for severe visceral leishmaniasis. J Infect Dis 1987;156(6):1030–3.
[25] Maciel BL, Lacerda HG, Queiroz JW, Galvão J, Pontes NN, Dimenstein R, et al.Association of nutritional status with the response to infection with Leishmaniachagasi. Am J Trop Med Hyg 2008;79(4):591–8.
[26] Ouellette M, Drummelsmith J, Leprohon P, El Fadili K, Foucher A, Vergnes B, et al.In: Myler PJ, Fasel N, editors. Drug Resistance in Leishmania. Leishmania: After TheGenomeNorwich, UK: Horizon Press; 2007.
[27] Vergnes B, Gourbal B, Girard I, Sundar S, Drummelsmith J, Ouellette M. Aproteomics screen implicates HSP83 and a small kinetoplastid calpain-relatedprotein in drug resistance in Leishmania donovani clinical field isolates bymodulating drug-induced programmed cell death. Mol Cell Proteomics 2007;6(1):88–101.
[28] Bitterman KJ, Anderson RM, Cohen HY, Latorre-Esteves M, Sinclair DA. Inhibitionof silencing and accelerated aging by nicotinamide, a putative negative regulatorof yeast sir2 and human SIRT1. J Biol Chem 2002;277(47):45099–107.
[29] Revollo JR, GrimmAA, Imai S. The regulation of nicotinamide adenine dinucleotidebiosynthesis by Nampt/PBEF/visfatin in mammals. Curr Opin Gastroenterol2007;23(2):164–70.
[30] Horsman MR, Høyer M, Honess DJ, Dennis IF, Overgaard J. Nicotinamidepharmacokinetics in humans and mice: a comparative assessment and theimplications for radiotherapy. Radiother Oncol 1993;27(2):131–9.
[31] Bernier J, Stratford MR, Denekamp J, Dennis MF, Bieri S, Hagen F, et al.Pharmacokinetics of nicotinamide in cancer patients treated with acceleratedradiotherapy: the experience of the Co-operative Group of Radiotherapy of theEuropean Organization for Research and Treatment of Cancer. Radiother Oncol1998;48(2):123–33.
[32] Seifert K, Escobar P, Croft SL. In vitro activity of anti-leishmanial drugs againstLeishmania donovani is host cell dependent. J Antimicrob Chemother 2010;65(3):508–11.
[33] Osar Z, Samanci T, Demirel GY, Damci T, Ilkova H. Nicotinamide effects oxidativeburst activity of neutrophils in patients with poorly controlled type 2 diabetesmellitus. Exp Diabesity Res 2004;5(2):155–62.
[34] Ungerstedt JS, Blömback M, Söderström T. Nicotinamide is a potent inhibitor ofproinflammatory cytokines. Clin Exp Immunol 2003;131(1):48–52.




















