
Locus coeruleus neurons encode the subjective difficulty of ...

BMP-2 and cAMP Elevation Confer Locus CoeruleusNeurons Responsiveness to MultipleNeurotrophic Factors
Julia Reiriz,1,2,3 Pontus C. Holm,1 Jordi Alberch,3 Ernest Arenas1
1 Department of Medical Biochemistry and Biophysics, Laboratory of Molecular Neurobiology,Karolinska Institute, Stockholm, Sweden
2 Department Infermeria Fonamental i Medicoquirurgica, Escola d’Infermeria, Universitat deBarcelona, Barcelona, Spain
3 Department Biologia Cel.lular i Anatomia Patologica, Facultat de Medicina, IDIBAPS,Universitat de Barcelona, Barcelona, Spain
Received 11 June 2001; accepted 3 October 2001
ABSTRACT: The locus coeruleus (LC) is a majortarget of several neurodegenerative disorders, includingParkinson’s and Alzheimer’s diseases. However, verylittle is known of the trophic requirements of LC neu-rons. In the present work, we have studied the biologicalactivity of neurotrophic factors from different familiesin E15 primary cultures of LC neurons. In agreementwith previous results, neurotrophin-3 (NT-3) and alsoglial cell line- derived neurotrophic factor (GDNF) in-creased the number of embryonic LC noradrenergicneurons in the presence of serum. In serum-free condi-tions, none of the factors tested, including NT-3, GDNF,neurturin, basic fibroblast growth factor (bFGF), orbone morphogenetic protein-2 (BMP-2), promoted thesurvival of tyrosine hydroxylase (TH)-immunoreactiveneurons at 6 days in vitro. However, when BMP-2 was
coadministered with any of these factors the number ofLC TH-positive neurons increased twofold. Similar re-sults were obtained by cotreatment of LC neurons withforskolin and NT-3, bFGF, or BMP-2. The strongesteffect (a fourfold increase in the number of TH-positivecells) was induced by cotreatment with forskolin,BMP-2, and GDNF. Thus, our results show that LCneurons require multiple factors for their survival anddevelopment, and suggest that activation of LC neuronsby bone morphogenetic proteins and cAMP plays adecisive role in conferring noradrenergic neuron re-sponsiveness to several trophic factors. © 2002 Wiley Peri-
odicals, Inc. J Neurobiol 50: 291–304, 2002; DOI 10.1002/neu.10034
Keywords: locus coeruleus cultures; bone morphogeneticproteins; neurotrophic factors; cyclic AMP; basic fibro-blast growth factor
INTRODUCTION
In the vertebrate central nervous system (CNS), neu-rons that synthesize noradrenaline are restricted to the
pontine and medullary tegmental regions. The locuscoeruleus (LC) contains about half of the total numberof noradrenaline-synthesizing neurons. The LC is sit-uated in the floor of the fourth ventricle at the dorso-lateral pontine tegmentum level and gives rise toextensively branched ascending and descending fibersystems that project throughout the CNS (Nieuwen-huys, 1985). LC cells undergo differentiation at about12 days of gestational age in the rat, considerablyearlier than cells in target areas such as hippocampusand cerebellum (Lauder and Bloom, 1974). Moreover,LC neurons express a unique pattern of transcriptionfactors, compared to other catecholamine-containing
J. Reiriz and P.C. Holm contributed equally to this work.Correspondence to: E. Arenas ([email protected]).Contract grant sponsor: European Union; contract grant num-
ber: BIO4-98-0112.Contract grant sponsor: Swedish MRC.Contract grant sponsor: Karolinska Institute.Contract grant sponsor: Spanish Ministerio de Educacion y
Ciencia (J.R.).© 2002 Wiley Periodicals, Inc.
291

neurons. While the homeobox Phox2a is expressedfrom E9 onwards, Phox2b is transiently expressedaround E10 in the mouse (Pattyn et al., 1997). Dif-ferentiation of LC neurons is also complete at anearlier time than neurons in the raphe nucleus orsubstantia nigra pars compacta. Innervation of targetareas by LC neurons also occurs at a relatively earlydevelopmental stage (Marshall et al., 1991). Verylittle is known of the trophic requirements of LCneurons. From all known neurotrophic factors, onlythe biological activities of the neurotrophins and theglial cell line-derived neurotrophic factor (GDNF)family of ligands have been characterized in the LC.Neurotrophin-3 (NT-3) was found to be a survivalfactor for LC noradrenergic neurons in vitro (Fried-man et al., 1993), and in vivo, in a 6-hydroxydopa-mine (6OHDA) lesion model (Arenas and Persson,1994). These results are consistent with the findingthat adult LC expresses p75 and trkC mRNA (Merlioet al., 1992), encoding tyrosine kinase receptors thatcan be activated by NT-3. However, homozygousmice carrying a deletion of the NT-3 gene showed noreduction in the number of neurons or in the level oftyrosine hydroxylase (TH) staining in the LC (Ernforset al., 1994; Farinas et al., 1994), suggesting that otherneurotrophic factors may be responsible for maintain-ing the survival and phenotype of central noradrener-gic neurons. One candidate is GDNF, a trophic factorfound to protect adult rat brain LC neurons from6OHDA-induced degeneration and to induce hyper-trophy as well as an increase in TH levels and sprout-ing of noradrenergic neurons (Arenas et al., 1995).GDNF mRNA is expressed in several targets of cen-tral noradrenergic neurons in the adult rat brain butnot in the LC (Arenas et al., 1995), and both c-ret andGFR�1 mRNAs, encoding the functional GDNF re-ceptor complex, are expressed by LC neurons (Truppet al., 1997; Glazner et al., 1998), suggesting thatGDNF may be a neurotrophic factor for LC norad-renergic neurons.
Other candidate trophic factors for noradrenergicneurons include the bone morphogenetic proteins(BMPs) and fibroblast growth factor families. BMPsconstitute a family of proteins involved in severalaspects of neuronal physiology (Reissmann et al.,1996; Shah et al., 1996; Mehler et al., 1997; Ebendalet al., 1998), including the survival and extension ofnerve processes from midbrain dopamine-containingneurons (Jordan et al., 1997; Reiriz et al., 1999) andthe control of the noradrenergic neurotransmitteridentity (Lo et al., 1999). On the other hand, the vastmajority of the cell bodies in the LC show basicfibroblast growth factor (bFGF) immunoreactivitywithin the cytoplasm and processes (Chadi et al.,
1993), and bFGF receptor mRNAs are expressed inthe LC (Wanaka et al., 1990), suggesting that bFGFmay provide a trophic support to LC noradrenergicneurons. Moreover, noradrenergic neurons of the LCdo not develop in zebra fish in the absence of FGF 8,bone morphogenetic protein-2 (BMP-2), or BMP-7(Guo et al., 1999), indicating that several factors arerequired for the development of noradrenergic neu-rons. Finally, we have examined whether increases incAMP may affect the responsiveness of LC noradren-ergic neurons to survival factors. Administration offorskolin to Phox2a overexpressing neural crest cellshas been reported to activate TH, and cotreatment ofthese cells with BMP-2 and forskolin dramaticallyinduced both TH and dopamine beta hydroxylase(DBH) (Lo et al., 1999). Moreover, application offorskolin, dibutyryl cAMP (dbcAMP), or other cAMPanalogues has been reported to increase the number ofTH immunoreactive LC neurons (Sklair-Tavron andSegal, 1993; Sklair-Tavron et al., 1994).
In the present work we have analyzed the actionsof representative growth factors of the above men-tioned families, including bFGF, NT-3, GDNF, neur-turin (NTN), and BMP-2, on LC neurons in culture.Our results show that LC neurons require multipletrophic factors for their survival and suggest thatBMP-2 and cAMP activation confer LC noradrener-gic neuron responsiveness to several survival factors.
METHODS
LC Cell Culture and Treatments
At E15 Sprague-Dawley pregnant rats (B&K Universal AB)were sacrificed by exposure to CO2 and soaked in 70%ethanol for 5 min. Embryos were removed under sterileconditions and kept in iced PBS/HEPES (Gibco) (10 mM)/glucose (0.33%) for microscopic dissection of the wholeproximal rombencephalic ring between the distal part of themesencephalic flexure and the proximal part of the pontineflexure (Specht et al., 1981), which includes the LC but notthe motor nuclei of the III, IV, VII, IX, X cranial nerves orthe solitary tract. The tissue was gently dissociated with afire-polished Pasteur pipette and plated on poly-D-lysinecoated 12 well-plates (Corning) at a density of 12.5 � 104
cells per cm2 (5 � 105 cells per well). Time elapsed be-tween the death of the mothers and plating the cells was�2.5 h. The cells were maintained for 18 h or 6 days ineither serum-free medium, which yields a relatively neuron-pure culture [MEM (Gibco) with HEPES (10 mM), B27(1/50; Gibco),L-glutamine (1 mM), penicillin-streptomy-cin], or in media containing 7.5% fetal calf serum (FCS),which promotes survival and proliferation of non-neuronalsupport cells. GDNF, NTN, NT-3, BMP-2, and bFGF wereadded to the wells just before plating the cells at a concen-
292 Reiriz et al.

tration of 30 ng/mL in serum-treated cultures. In serum-freecultures the concentration of trophic factors was the same asin cultures with serum, except for BMP-2, which was usedat a concentration of 1 ng/mL, as explained in the text. Allthe trophic factors were obtained from Pepro Tech exceptfor BMP-2, which was obtained from Genetics Institute. Aset of experiments was performed using forskolin (5 �M;Sigma) in serum-deprived cultures. Solutions used in tissueculture were sterilized by filtration through 0.22 �m filters(Millipore). Cultures were grown for 18 h or 6 days at 37°Cin a water saturated 5% CO2/95% air atmosphere incubatorwithout changing media or supplementing with factors.
Immunocytochemical ProceduresCultures were fixed in 4% paraformaldehyde in 0.1 Mphosphate buffer (pH 7.4) and processed with one of thefollowing antibodies, diluted in PBS containing 1% BSAand 0.3% Triton X-100: mouse anti-tyrosine hydroxylase(TH), 1:1000 (Incstar); rabbit anti-glial fibrillary acidic protein(GFAP), 1:500 (DAKO); mouse anti-5-bromodeoxyuridine(BrdU), 1:50 (DAKO). Incubations were carried out at 4°Covernight. After washing, cultures were incubated for 2 h withappropriate secondary antibodies [biotinylated horse anti-mouse IgG or goat antirabbit IgG (Vector)], 1:500, in the samedilution buffer as above. Immunostaining was visualized withthe Vector Laboratory ABC immunoperoxidase kit, using ei-ther gray (SG) or red (AEC) substrates.
BrdU Incorporation AssayTo determine whether cells were synthesizing DNA, cul-tures were incubated in 10 �M BrdU for a period of 4 h andthen fixed. The incorporation of BrdU into the nuclei ofcells that synthesized DNA was visualized using mouseanti-BrdU antibody (1/50; DAKO). Before processing withthe primary antibody, fixed cells were treated with 2 M HClfor 30 min.
Cell Count Assessment and StatisticalAnalysisAll TH-positive cells were counted in every well, in two tothree culture wells per experiment and two to three exper-iments per condition.
Data were analyzed statistically by one-way ANOVA.When p � .05, then the F-test was followed by the New-man-Keuls posthoc test to compare the statistical signifi-cance between experimental groups.
RESULTS
Serum-Containing Media Enhancesthe Survival and MorphologicalDifferentiation of LC TH-PositiveNeurons
TH, the rate-limiting enzyme in catecholamine bio-synthesis (Levitt et al., 1965), was used as a marker
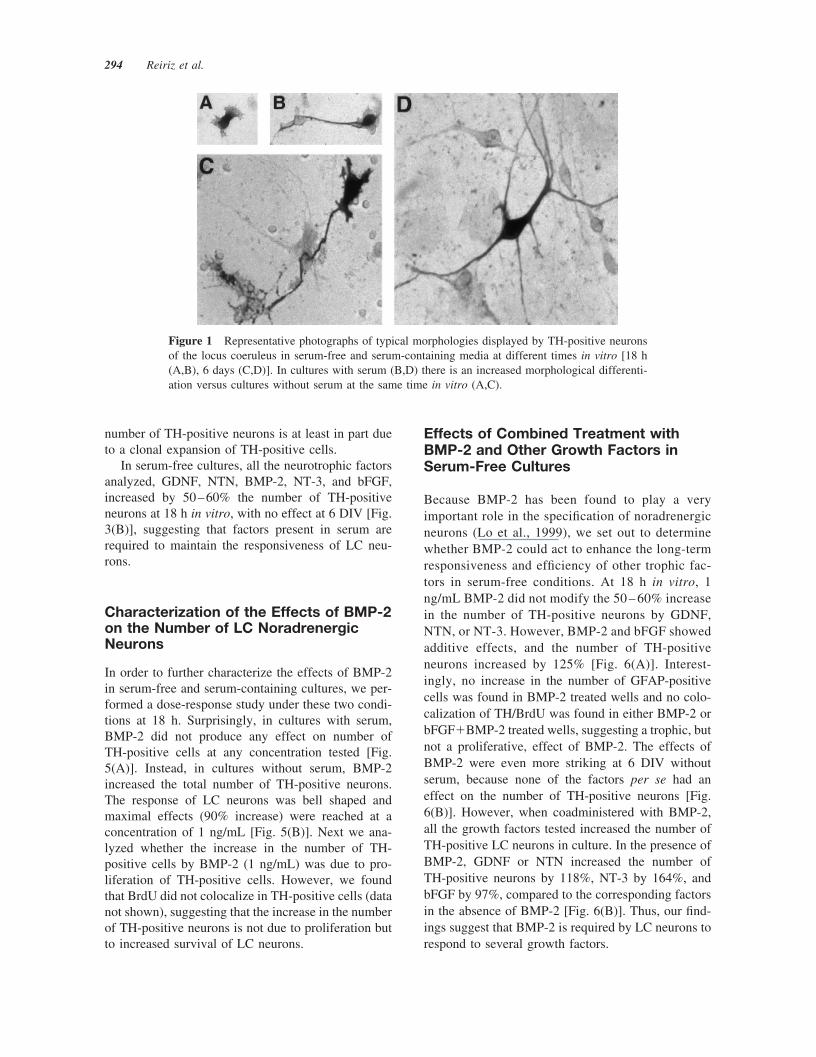
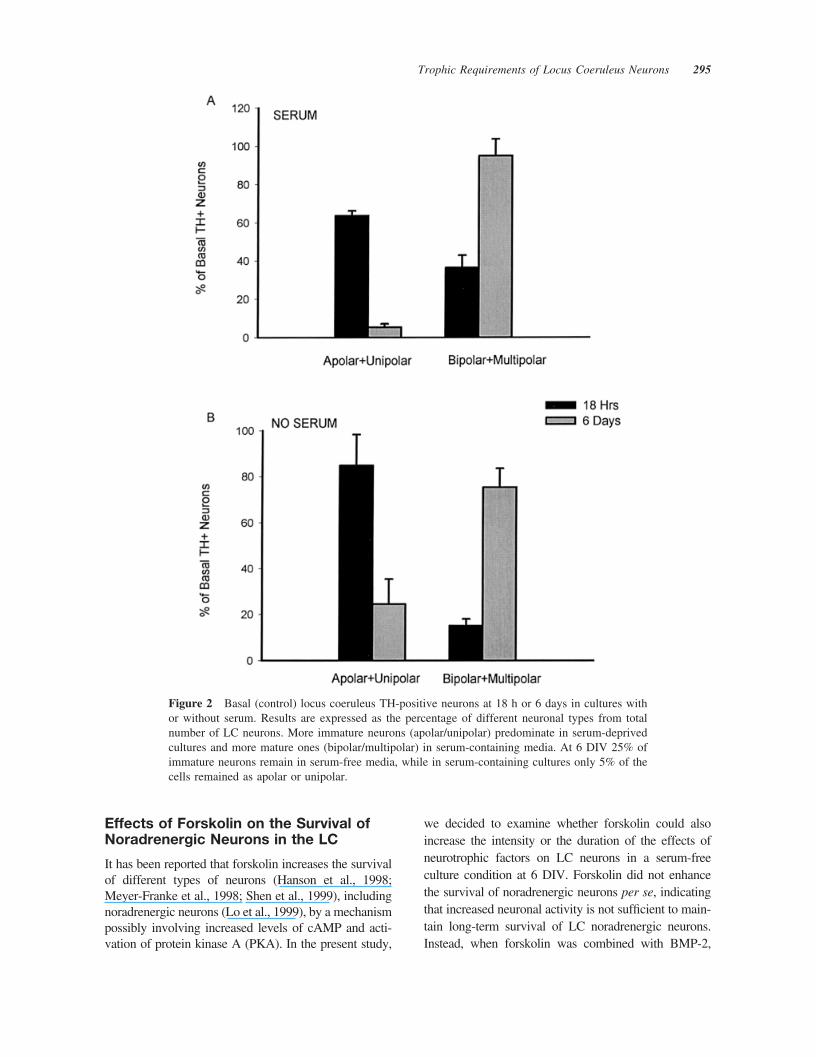
for noradrenergic cells of the LC. Colocalization ofTH-positive cells with both homeodomain proteinsPhox2a and Phox2b (Guo et al., 1999) supportedtheir identification as locus noradrenergic neurons(P. Holm and E. Arenas, unpublished data). In serum-containing cultures, the number of TH-positive cellsincreased from 44.33 � 3.79 (mean�S.D.) at 18 h to70.5 � 11.03 at 6 days in vitro (DIV) (p � .05 byANOVA). On the contrary, in serum-free culturesthere was a decrease from 29.67 � 9.77 TH-positivecells at 18 h to only 9.50 � 2.74 cells at 6 DIV (p� .001). Initially, TH-positive cells were small andpredominantly apolar and unipolar, with prominentgrowth cones [Fig. 1(A,B)]. Over the course of 6 DIV,bipolar and multipolar neurons increased by 60% ineither serum-free or serum-containing media, exhib-iting larger cell bodies and increased length, number,and complexity of neuronal processes [Fig. 1(C,D)].However, more mature neurons were observed inserum-containing than in serum-free cultures. At 18 hin vitro, 64% of the neurons in the presence of serum[Fig. 2(A)] and 85% of the neurons in serum-freecultures [Fig. 2(B)] were apolar or unipolar. At 6DIV, 95% of the neurons in the presence of serum[Fig. 2(A)] and 75% of the neurons in serum-freecultures [Fig. 2(B)] were bipolar or multipolar.
Effects of Neurotrophic Factors onthe Number and Phenotype of LCNoradrenergic Neurons In Vitro
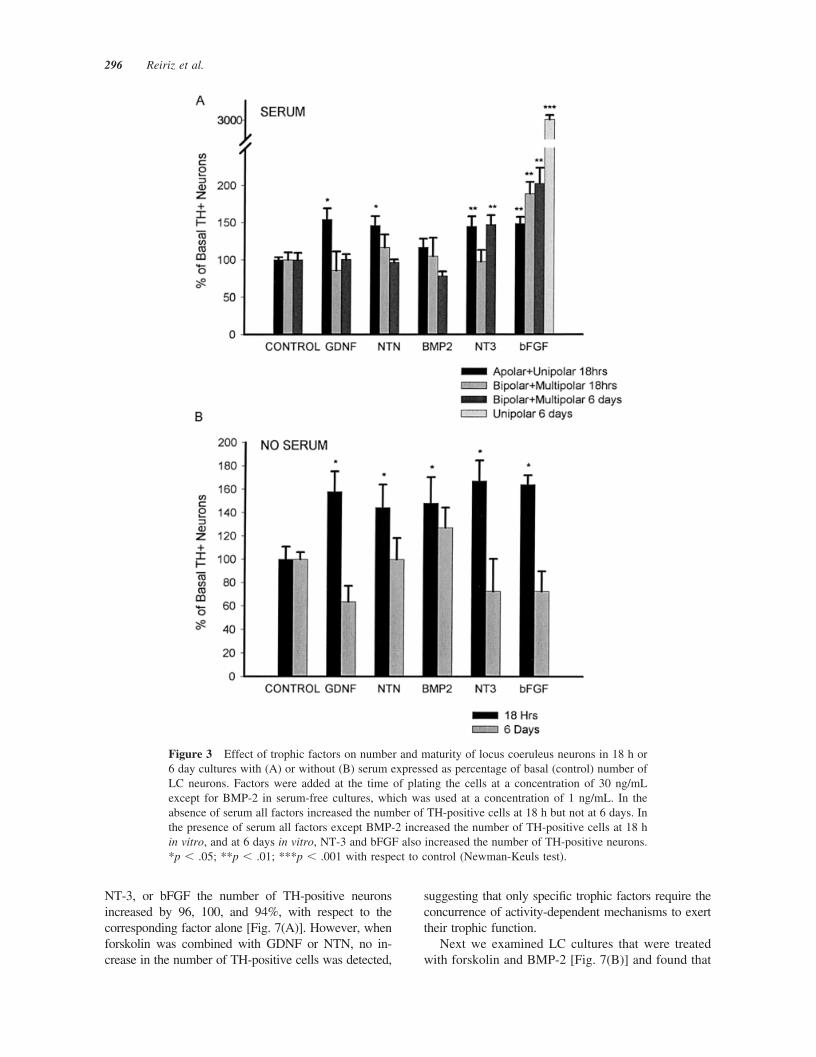
Following Loughlin et al. (1986), we considered bi-polar and multipolar TH-positive neurons as the ma-ture typical LC TH-positive cells and apolar andunipolar as the more immature ones. Our results showthat BMP-2 did not change the number or morphologyof LC neurons in the presence of serum [Fig. 3(A)]. Incontrast, other neurotrophic factors tested (bFGF,GDNF, NTN, and NT-3) increased by 50% the num-ber of apolar/unipolar TH-positive neurons at 18 h.However, the effects of GDNF or NTN were notsustained and only NT-3 and bFGF increased thenumber of TH-positive cells at 6 DIV. At this latetime point NT-3 increased bipolar/multipolar cells by50%, and bFGF increased by 30-fold the number ofimmature unipolar cells [Fig. 3(A)]. Interestingly,bFGF increased the total number of TH-positive neu-rons by 3.8-fold, from 70.5 (�11.03, n � 2) in serumalone to 269.75 (�21.13, n � 2). Moreover, the totalnumber of LC TH-positive cells also increased from18 h to 6 DIV and resulted in the formation of six toeight large clusters of unipolar TH-positive neurons inevery well (Fig. 4) , suggesting that the increase in the
Trophic Requirements of Locus Coeruleus Neurons 293
number of TH-positive neurons is at least in part dueto a clonal expansion of TH-positive cells.
In serum-free cultures, all the neurotrophic factorsanalyzed, GDNF, NTN, BMP-2, NT-3, and bFGF,increased by 50–60% the number of TH-positiveneurons at 18 h in vitro, with no effect at 6 DIV [Fig.3(B)], suggesting that factors present in serum arerequired to maintain the responsiveness of LC neu-rons.
Characterization of the Effects of BMP-2on the Number of LC NoradrenergicNeurons
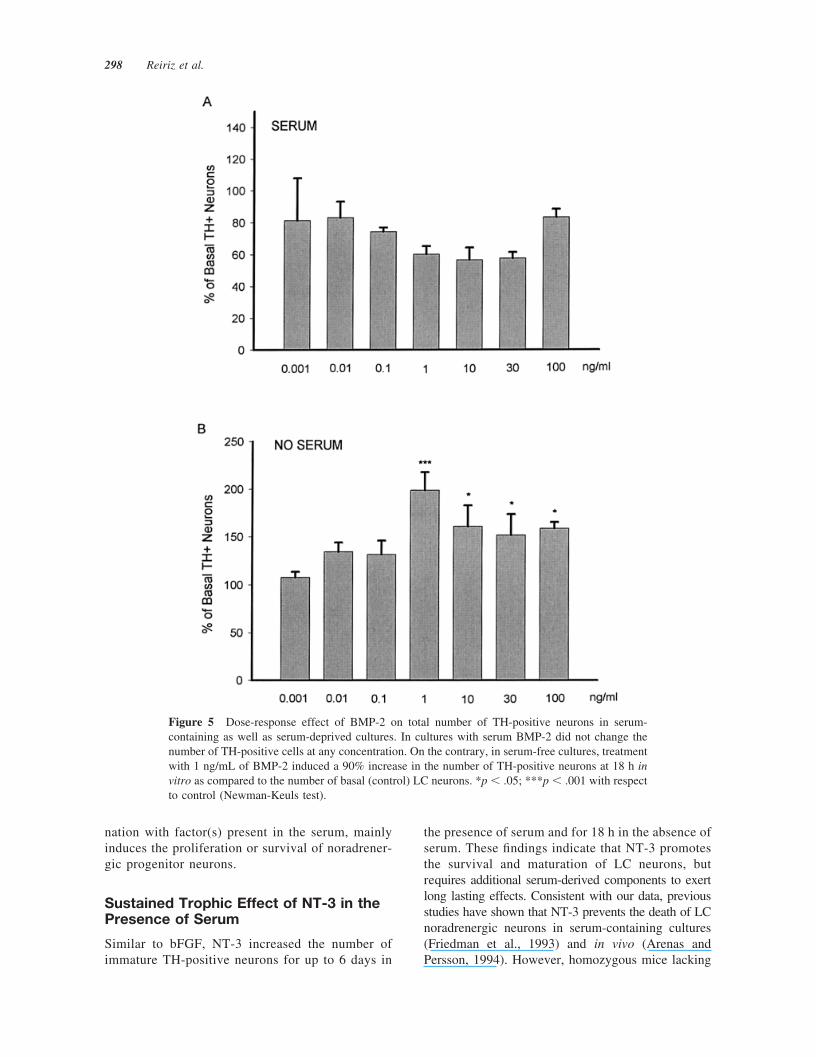
In order to further characterize the effects of BMP-2in serum-free and serum-containing cultures, we per-formed a dose-response study under these two condi-tions at 18 h. Surprisingly, in cultures with serum,BMP-2 did not produce any effect on number ofTH-positive cells at any concentration tested [Fig.5(A)]. Instead, in cultures without serum, BMP-2increased the total number of TH-positive neurons.The response of LC neurons was bell shaped andmaximal effects (90% increase) were reached at aconcentration of 1 ng/mL [Fig. 5(B)]. Next we ana-lyzed whether the increase in the number of TH-positive cells by BMP-2 (1 ng/mL) was due to pro-liferation of TH-positive cells. However, we foundthat BrdU did not colocalize in TH-positive cells (datanot shown), suggesting that the increase in the numberof TH-positive neurons is not due to proliferation butto increased survival of LC neurons.
Effects of Combined Treatment withBMP-2 and Other Growth Factors inSerum-Free Cultures
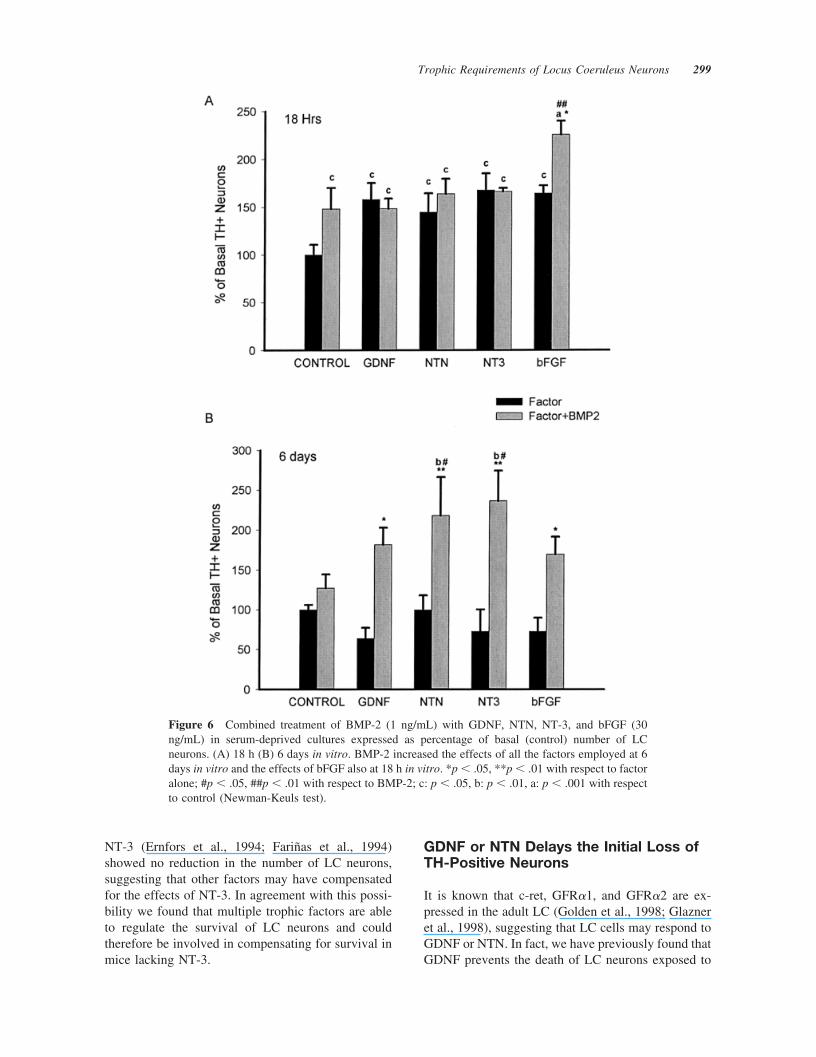
Because BMP-2 has been found to play a veryimportant role in the specification of noradrenergicneurons (Lo et al., 1999), we set out to determinewhether BMP-2 could act to enhance the long-termresponsiveness and efficiency of other trophic fac-tors in serum-free conditions. At 18 h in vitro, 1ng/mL BMP-2 did not modify the 50 – 60% increasein the number of TH-positive neurons by GDNF,NTN, or NT-3. However, BMP-2 and bFGF showedadditive effects, and the number of TH-positiveneurons increased by 125% [Fig. 6(A)]. Interest-ingly, no increase in the number of GFAP-positivecells was found in BMP-2 treated wells and no colo-calization of TH/BrdU was found in either BMP-2 orbFGF�BMP-2 treated wells, suggesting a trophic, butnot a proliferative, effect of BMP-2. The effects ofBMP-2 were even more striking at 6 DIV withoutserum, because none of the factors per se had aneffect on the number of TH-positive neurons [Fig.6(B)]. However, when coadministered with BMP-2,all the growth factors tested increased the number ofTH-positive LC neurons in culture. In the presence ofBMP-2, GDNF or NTN increased the number ofTH-positive neurons by 118%, NT-3 by 164%, andbFGF by 97%, compared to the corresponding factorsin the absence of BMP-2 [Fig. 6(B)]. Thus, our find-ings suggest that BMP-2 is required by LC neurons torespond to several growth factors.
Figure 1 Representative photographs of typical morphologies displayed by TH-positive neuronsof the locus coeruleus in serum-free and serum-containing media at different times in vitro [18 h(A,B), 6 days (C,D)]. In cultures with serum (B,D) there is an increased morphological differenti-ation versus cultures without serum at the same time in vitro (A,C).
294 Reiriz et al.
Effects of Forskolin on the Survival ofNoradrenergic Neurons in the LC
It has been reported that forskolin increases the survivalof different types of neurons (Hanson et al., 1998;Meyer-Franke et al., 1998; Shen et al., 1999), includingnoradrenergic neurons (Lo et al., 1999), by a mechanismpossibly involving increased levels of cAMP and acti-vation of protein kinase A (PKA). In the present study,
we decided to examine whether forskolin could alsoincrease the intensity or the duration of the effects ofneurotrophic factors on LC neurons in a serum-freeculture condition at 6 DIV. Forskolin did not enhancethe survival of noradrenergic neurons per se, indicatingthat increased neuronal activity is not sufficient to main-tain long-term survival of LC noradrenergic neurons.Instead, when forskolin was combined with BMP-2,
Figure 2 Basal (control) locus coeruleus TH-positive neurons at 18 h or 6 days in cultures withor without serum. Results are expressed as the percentage of different neuronal types from totalnumber of LC neurons. More immature neurons (apolar/unipolar) predominate in serum-deprivedcultures and more mature ones (bipolar/multipolar) in serum-containing media. At 6 DIV 25% ofimmature neurons remain in serum-free media, while in serum-containing cultures only 5% of thecells remained as apolar or unipolar.
Trophic Requirements of Locus Coeruleus Neurons 295
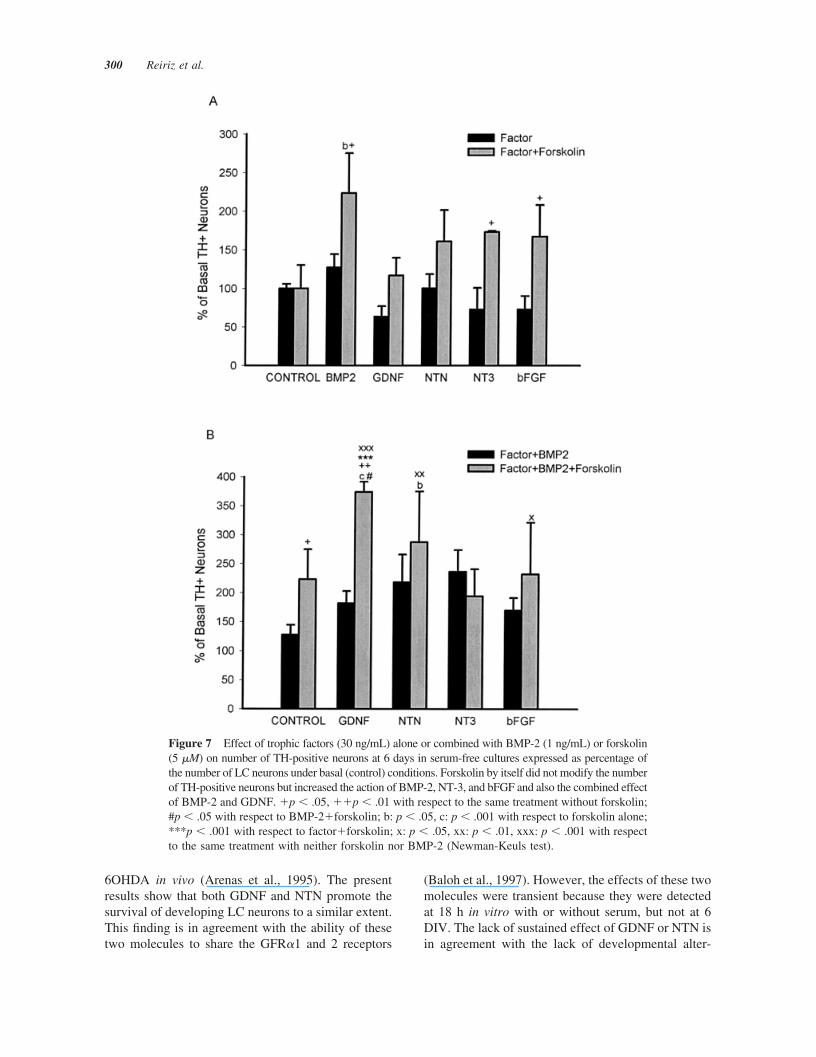
NT-3, or bFGF the number of TH-positive neuronsincreased by 96, 100, and 94%, with respect to thecorresponding factor alone [Fig. 7(A)]. However, whenforskolin was combined with GDNF or NTN, no in-crease in the number of TH-positive cells was detected,
suggesting that only specific trophic factors require theconcurrence of activity-dependent mechanisms to exerttheir trophic function.
Next we examined LC cultures that were treatedwith forskolin and BMP-2 [Fig. 7(B)] and found that
Figure 3 Effect of trophic factors on number and maturity of locus coeruleus neurons in 18 h or6 day cultures with (A) or without (B) serum expressed as percentage of basal (control) number ofLC neurons. Factors were added at the time of plating the cells at a concentration of 30 ng/mLexcept for BMP-2 in serum-free cultures, which was used at a concentration of 1 ng/mL. In theabsence of serum all factors increased the number of TH-positive cells at 18 h but not at 6 days. Inthe presence of serum all factors except BMP-2 increased the number of TH-positive cells at 18 hin vitro, and at 6 days in vitro, NT-3 and bFGF also increased the number of TH-positive neurons.*p � .05; **p � .01; ***p � .001 with respect to control (Newman-Keuls test).
296 Reiriz et al.

the only growth factor that selectively enhanced thenumber of TH-positive neurons over the effects ofBMP-2 plus forskolin (223%) was GDNF (373%).Thus, although forskolin, BMP-2, and GDNF were allinactive per se in serum-free conditions at 6 DIV, theycooperate to enhance the long-term survival of nor-adrenergic neurons. Moreover, because the effects offorskolin�BMP-2 and GDNF�BMP-2 are additive,our findings suggest that forskolin and GDNF utilizetwo separate mechanisms to increase the number ofTH-positive neurons, both of which are BMP-2-de-pendent. Finally, because the BrdU incorporation as-say showed no colocalization BrdU/TH in any of theforskolin-treated wells, our results indicate that theincrease in the number of TH-positive cells was notdue to TH precursor cell proliferation but to an in-crease in the expression of TH and/or a survival-promoting effect of these molecules.
DISCUSSION
The present study shows that neurotrophic factors fromdifferent families induce unique effects on LC cultures.These results suggest a specific function for each ofthese molecules in the maturation and survival of LCneurons and argue for a role for multiple neurotrophicfactors in the development of central noradrenergic neu-rons. In the next paragraphs the effects of the differentgrowth factors alone or in combination and in the pres-ence of elevated cAMP levels are considered.
bFGF Increases the Number of TH-Positive Cells Cultured in Serum
Our characterization of the LC cultures showed that thenumber of bipolar and multipolar TH-positive neuronsincreased over the course of 6 DIV, exhibiting larger cellbodies and increased length, number, and complexity ofneuronal processes. These changes resemble those ob-served during the development of LC in vivo (Specht etal., 1981; Loughlin et al., 1986), and suggest that LCneurons mature in vitro. Interestingly, addition of serumto the cultures increased the number of TH-positiveneurons and the maturation of their processes. Theseresults are in agreement with the finding that serumdeprivation induces apoptosis (Park et al., 1998; Hetmanet al., 2000) and suggest a trophic effect of serum.
In serum-containing cultures we found thatbFGF increased by four times the total number ofTH-positive neurons between 18 h and 6 days.Moreover, at 6 DIV the number of unipolar cellsincreased 30-fold with bFGF treatment. These find-ings indicate that factors present in the serum po-tentiated the effects of bFGF on the number andmaturation of noradrenergic LC neurons. The effectof the bFGF and serum was not mimicked by anyother defined culture condition, including cotreat-ment with BMP-2 and bFGF in serum-free cultures,suggesting that other factors, or a combination ofthem, may be involved in potentiating the effects ofbFGF. Moreover, the finding that the number ofundifferentiated unipolar TH-positive neurons in-creased over time suggested that bFGF, in combi-
Figure 4 Low magnification photomicrographs of TH-immunocytochemistry stained E15 LCprimary cultures treated for 6 days with serum alone (A) or with serum and bFGF (B). (A) Averagefield showing one TH-positive cell in the presence of serum. (B) Cluster of TH-positive neuronsappearing after bFGF treatment in the presence of serum.
Trophic Requirements of Locus Coeruleus Neurons 297
nation with factor(s) present in the serum, mainlyinduces the proliferation or survival of noradrener-gic progenitor neurons.
Sustained Trophic Effect of NT-3 in thePresence of Serum
Similar to bFGF, NT-3 increased the number ofimmature TH-positive neurons for up to 6 days in
the presence of serum and for 18 h in the absence ofserum. These findings indicate that NT-3 promotesthe survival and maturation of LC neurons, butrequires additional serum-derived components to exertlong lasting effects. Consistent with our data, previousstudies have shown that NT-3 prevents the death of LCnoradrenergic neurons in serum-containing cultures(Friedman et al., 1993) and in vivo (Arenas andPersson, 1994). However, homozygous mice lacking
Figure 5 Dose-response effect of BMP-2 on total number of TH-positive neurons in serum-containing as well as serum-deprived cultures. In cultures with serum BMP-2 did not change thenumber of TH-positive cells at any concentration. On the contrary, in serum-free cultures, treatmentwith 1 ng/mL of BMP-2 induced a 90% increase in the number of TH-positive neurons at 18 h invitro as compared to the number of basal (control) LC neurons. *p � .05; ***p � .001 with respectto control (Newman-Keuls test).
298 Reiriz et al.
NT-3 (Ernfors et al., 1994; Farinas et al., 1994)showed no reduction in the number of LC neurons,suggesting that other factors may have compensatedfor the effects of NT-3. In agreement with this possi-bility we found that multiple trophic factors are ableto regulate the survival of LC neurons and couldtherefore be involved in compensating for survival inmice lacking NT-3.
GDNF or NTN Delays the Initial Loss ofTH-Positive Neurons
It is known that c-ret, GFR�1, and GFR�2 are ex-pressed in the adult LC (Golden et al., 1998; Glazneret al., 1998), suggesting that LC cells may respond toGDNF or NTN. In fact, we have previously found thatGDNF prevents the death of LC neurons exposed to
Figure 6 Combined treatment of BMP-2 (1 ng/mL) with GDNF, NTN, NT-3, and bFGF (30ng/mL) in serum-deprived cultures expressed as percentage of basal (control) number of LCneurons. (A) 18 h (B) 6 days in vitro. BMP-2 increased the effects of all the factors employed at 6days in vitro and the effects of bFGF also at 18 h in vitro. *p � .05, **p � .01 with respect to factoralone; #p � .05, ##p � .01 with respect to BMP-2; c: p � .05, b: p � .01, a: p � .001 with respectto control (Newman-Keuls test).
Trophic Requirements of Locus Coeruleus Neurons 299
6OHDA in vivo (Arenas et al., 1995). The presentresults show that both GDNF and NTN promote thesurvival of developing LC neurons to a similar extent.This finding is in agreement with the ability of thesetwo molecules to share the GFR�1 and 2 receptors
(Baloh et al., 1997). However, the effects of these twomolecules were transient because they were detectedat 18 h in vitro with or without serum, but not at 6DIV. The lack of sustained effect of GDNF or NTN isin agreement with the lack of developmental alter-
Figure 7 Effect of trophic factors (30 ng/mL) alone or combined with BMP-2 (1 ng/mL) or forskolin(5 �M) on number of TH-positive neurons at 6 days in serum-free cultures expressed as percentage ofthe number of LC neurons under basal (control) conditions. Forskolin by itself did not modify the numberof TH-positive neurons but increased the action of BMP-2, NT-3, and bFGF and also the combined effectof BMP-2 and GDNF. �p � .05, ��p � .01 with respect to the same treatment without forskolin;#p � .05 with respect to BMP-2�forskolin; b: p � .05, c: p � .001 with respect to forskolin alone;***p � .001 with respect to factor�forskolin; x: p � .05, xx: p � .01, xxx: p � .001 with respectto the same treatment with neither forskolin nor BMP-2 (Newman-Keuls test).
300 Reiriz et al.

ations at birth in the LC of GDNF and NTN �/�mice (Moore et al., 1996; Pichel et al., 1996; Sanchezet al., 1996; Heuckeroth et al., 1999). Thus, our resultscan be interpreted as an effect of GDNF or NTN onmechanically damaged neurons in primary cultures.This interpretation would be in agreement with theeffects of GDNF in the lesioned adult LC (Arenas etal., 1995) and would argue for a predominant role ofGDNF and NTN in response to damage. However,these results do not rule out the possibility that GDNFand NTN may work as trophic factors regulatingdifferent aspects of the development of LC neuronsand even survival but in a redundant manner, asshown in the present study.
BMP-2 Prevents the Early Loss of LCNeurons Only in the Absence of Serum
Our data show that BMP-2 did not produce any effecton the number of LC neurons in the presence ofserum, suggesting that serum components interferewith BMP signaling. BMP signals are transmittedthrough the phosphorylation of Smad proteins (Hood-less et al., 1996) by the serine/threonine kinase activ-ity of the BMPR-I. However, Smads can also bephosphorylated at a different site by epidermal growthfactor (EGF) to mediate opposite effects on the nu-clear accumulation of Smad (Kretzschmar et al.,1997). Thus, factors such as EGF, which is present inserum, may explain the lack of effect of BMP-2 inserum- treated LC cultures. Because serum containsBMPs and the effects of BMP-2 are only detected at1 ng/mL, the addition of BMP-2 to serum wouldresult in higher BMP doses to which LC neurons donot respond.
Interestingly, BMP-2 clearly increased the numberof TH-positive cells in serum-deprived cultures, butonly at 18 h in vitro. These cells did not incorporateBrdU, suggesting that BMP-2-mediated increases inTH-positive neurons did not result from an increasedrate of cell division of TH precursors. Thus, ourfindings suggest that the levels of BMP-2 have to bevery tightly regulated in order to be able to promotethe survival of LC noradrenergic neurons.
BMP-2 Promotes the Long-TermSurvival of Multiple Trophic Factors
Because of the early role of BMPs in the induction ofthe noradrenergic phenotype (Lo et al., 1999) and thesurvival effects of BMP-2 on LC noradrenergic neu-rons (present results), we examined whether BMP-2could potentiate the effects of other growth factors,including GDNF, NTN, NT-3, and bFGF. Our results
show that only bFGF and BMP-2 had additive effectsat 18 h, and that at 6 days all the factors tested (NT-3,GDNF, NTN, and bFGF) showed additive effectswith BMP-2. We do not know what is the mechanismby which these interactions take place, but it is pos-sible that BMP-2 could act to induce or maintain thetrophic phenotype of LC noradrenergic neurons, in-cluding the expression of signaling components andreceptors for the different growth factors as it hasbeen reported for TrkC (Kobayashi et al., 1998;Zhang et al., 1998) or GFR�1 (Thang et al., 2000).BMPs have also been shown to interact with NF-�Bsubunits (Lopez-Rovira et al., 2000), which provideboth survival and differentiation signals to the cell(Foehr et al., 2000a, 2000b), and to act synergisticallyboth with neurotrophins and GDNF (Bengtsson et al.,1998). Thus, multiple mechanisms may account forthe potentiation of the effects of neurotrophic factorsby BMPs.
cAMP Activation Is Required to Maintainand/or Potentiate the Trophic Effectsof Neurotrophic Factors
Finally, we decided to test whether increased neuronalactivity induced by forskolin, which activates adeny-lyl cyclase and increases cAMP and neuronal respon-siveness to neurotrophic factors (Hanson et al., 1998),could increase the number of TH-positive neurons inLC cultures. It has been shown that cAMP can in-crease TH activity (Joh et al., 1978; Zigmond et al.,1989) and TH transcription (Kim et al., 1993a, 1993b,1994) in catecholaminergic neurons, and it can alsoincrease the number of TH-positive noradrenergiccells in the presence of serum (Sklair-Tavron andSegal, 1993; Sklair-Tavron et al., 1994, 1996). Inter-estingly, we found that in serum-free media, forskolinpotentiated the effect of BMP-2, NT-3, and bFGF butnot GDNF or NTN. This result is in contrast with theeffects of forskolin and BMP-2, which together en-hanced the effects of GDNF, NTN, and bFGF. Inter-estingly, the strongest increase in the number of TH-positive neurons in serum-free cultures was observedafter cotreatment with forskolin, BMP-2, and GDNF.The mechanism by which forskolin potentiates theeffects of growth factors on the number of TH-posi-tive neurons is not clear. Osaka and Sabban (1997)have shown that bFGF increases TH transcription inPC12 cells, suggesting that bFGF could regulate thelevels of TH in LC neurons. However, Dugan et al.(1999) found that cAMP promotes the survival of E15cortical neurons by activating MAPK in a Rap1/B-raf-dependent manner. Moreover, Meyer-Franke et al.(1998) reported that cAMP elevation rapidly induces
Trophic Requirements of Locus Coeruleus Neurons 301

a substantial increase in the number of TrkB receptorson the plasma membrane of retinal ganglion cells.This latter finding is very interesting because therecruitment of TrkB to the plasma membrane bycAMP constitute an activity-dependent mechanism toregulate neuronal responsiveness to neurotrophic fac-tors. Thus, in the future it will be important to deter-mine whether this type of mechanism could be uti-lized to regulate the membrane levels of other(tyrosine kinase) receptors and therefore explain thepotentiating effect of forskolin that we detect.
In summary, we found that serum increases thesurvival and maturation of LC neurons and that bFGFplus serum increases the total number of TH-positiveneurons. In serum-free cultures, none of the trophicfactors tested, BMP-2, GDNF, NTN, NT-3, or bFGF,promoted per se the long-term survival (6 days) ofTH-positive noradrenergic neurons. However, wefound that BMP-2 potentiated or induced the respon-siveness of LC neurons to GDNF, NTN, NT-3, andbFGF. Similarly, we also found that forskolin poten-tiated the effects of BMP-2, NT-3, and bFGF. Finally,when forskolin and BMP-2 were combined withGDNF, NTN, or bFGF, a further increase in thenumber of LC TH-positive neurons was detected,suggesting that an activity-dependant mechanism reg-ulates the responsiveness of LC neurons to neurotro-phic factors.
Thus, in conclusion, our data are consistent withthe idea that multiple trophic factors and activity-dependant mechanisms are necessary for the survivaland development of LC noradrenergic neurons.
We thank Lotta Skoog for secretarial help and AnnikaAhlsen for additional assistance. The authors are grateful toDr. Isidre Ferrer, Dr. Joan Serratosa, and Dr. Coral Sanfeliufor sharing their equipment with us.
REFERENCES
Arenas E, Persson H. 1994. Neurotrophin-3 prevents thedeath of adult central noradrenergic neurons in vivo.Nature 367:368–371.
Arenas E, Trupp M, Åkerud P, Ibanez C. 1995. GDNFprevents degeneration and promotes the phenotype ofbrain noradrenergic neurons in vivo. Neuron 15:1465–1473.
Baloh RH, Tansey MG, Golden JP, Creedon DJ, Heucker-oth RO, Keck CL, Zimonjic DB, Popescu NC, JohnsonEM, Milbrandt J. 1997. TrnR2, a novel receptor thatmediates Neurturin and GDNF signaling through Ret.Neuron 18:793–802.
Bengtsson H, Soderstrom S, Kylberg A, Charette MF, Eb-endal T. 1998. Potentiating interactions between morpho-
genetic protein and neurotrophic factors in developingneurons. J Neurosci Res 53:559–568.
Chadi G, Tinner B, Agnati LF, Fuxe K. 1993. Basic fibro-blast growth factor (bFGF, FGF-2) immunoreactivity ex-ists in the noradrenaline, adrenaline and 5-HT nerve cellsof the rat brain. Neurosci Lett 160:171–176.
Dugan LL, Kim JS, Zhang Y, Bart RD, Sun Y, HoltzmanDM, Gutmann DH. 1999. Differential effects of cAMP inneurons and astrocytes. J Biol Chem 274:25842–25858.
Ebendal T, Bengtsson H, Soderstrom S. 1998. Bone mor-phogenetic proteins and their receptors: potential func-tions in the brain. J Neurosci Res 51:139–146.
Ernfors P, Lee KF, Kucera J, Jaenisch R. 1994. Lack ofneurotrophin-3 leads to deficiencies in the peripheral ner-vous system and loss of limb propioceptive afferents. Cell77:503–512.
Farinas I, Jones KR, Backus C, Wang XY, Reichardt LF.1994. Severe and sympathetic deficits in mice lackingneurotrophin-3. Nature 369:658–661.
Foehr ED, Bohuslav J, Chen LF, DeNoronha C, GeleziunasR, Lin X, O’Mahony A, Greene WC. 2000a. The NF-�B-inducing kinase induces PC12 cell differentiation andprevents apoptosis. J Biol Chem 275:34021–34032.
Foehr ED, Lin X, O’Mahony A, Geleziunas R, Bradshaw RA,Greene WC. 2000b. NF-�B signaling promotes both cellsurvival and neurite process formation in nerve growthfactor-stimulated PC12 cells. J Neurosci 20:7556–7574.
Friedman WJ, Ibanez C, Hallbook F, Persson H, Cain LD,Dreyfus CF, Black IB. 1993. Differential actions of neu-rotrophins in the locus coeruleus and basal forebrain. ExpNeurol 119:72–78.
Glazner GW, Xiaojun M, Springer JE. 1998. Localization ofglial cell line-derived neurotrophic factor receptor alphaand c-ret mRNA in rat central nervous system. J CompNeurol 391:42–49.
Golden JP, Baloh RH, Kotzbauer PT, Lampe PA, OsbornePA, Milbrandt J, Johnson EM. 1998. Expression of neur-turin, GDNF and their receptors in the adult mouse CNS.J Comp Neurol 398:139–150.
Guo S, Brush J, Teraoka H, Goddard A, Wilson SW, Mul-lins MC, Rosenthal A. 1999. Development of noradren-ergic neurons in the zebrafish hindbrain requires BMP,FGF8, and the homeodomain protein Soulless/Phox 2a.Neuron 24:555–566.
Hanson MG, Shen S, Wiemelt AP, McMorris FA, BarresBA. 1998. Cyclic AMP elevation is sufficient to promotethe survival of spinal motor neurons in vitro. J Neurosci18:7361–7371.
Hetman M, Cavanaugh JE, Kimelman D, Xia Z. 2000. Role ofglycogen synthase kinase-3� in neuronal apoptosis inducedby trophic withdrawal. J Neurosci 20:2567–2574.
Heuckeroth RO, Enomoto H, Grider JR, Golden JP, HankeJA, Jackman A, Molliver DC, Bardgett ME, Snider WD,Johnson EM Jr, Milbrandt J. 1999. Gene targeting revealsa critical role for neurturin in the development and main-tenance of enteric, sensory, and parasympathetic neurons.Neuron 22:253–263.
Hoodless PA, Haerry T, Abdollah S, Stapleton M,
302 Reiriz et al.

O’Connor MB, Attisano L, Wrana L. 1996. MADR1, aMAD-related protein that functions in BMP-2 signalingpathways. Cell 85:489–500.
Joh TH, Park DH, Reis DJ. 1978. Direct phosphorylation ofbrain tyrosine hydroxylase by cyclic AMP-dependentprotein kinase: mechanism of enzyme activation. ProcNatl Acad Sci USA 75:4744–4748.
Jordan J, Bottner M, Schluesener HJ, Unsicker K, Kriegl-stein K. 1997. Bone morphogenetic proteins: neurotro-phic roles for midbrain dopaminergic neurons and impli-cations of astroglial cells. Eur J Neurosci 9:1699–1710.
Kim KS, Lee MK, Carroll J, Joh TH. 1993a. Both the basaland inducible transcription of the tyrosine hydroxylasegene are dependent upon a cAMP response element.J Biol Chem 268:15689–15695.
Kim KS, Park DH, Wessel TC, Song B, Wagner JA, JohTH. 1993b. A dual role for the cAMP-dependent proteinkinase in tyrosine hydroxylase gene expression. Proc NatlAcad Sci USA 90:3471–3475.
Kim KS, Tinti C, Song B, Cubells JF, Joh TJ. 1994. CyclicAMP-dependent protein kinase regulates basal and cyclicAMP-stimulated but not phorbolester-stimulated tran-scription of the tyrosine hydroxylase gene. J Neurochem63:834–842.
Kobayashi M, Fujii M, Kurihara K, Matsuoka I. 1998. Bonemorphogenetic protein-2 and retinoic acid induce neuro-trophin-3 responsiveness in developing rat sympatheticneurons. Mol Brain Res 53:206–217.
Kretzschmar M, Doody J, Massague J. 1997. OpposingBMP and EGF signaling pathways converge on theTGF-� family mediator Smad1. Nature 389:618–622.
Lauder JM, Bloom FE. 1974. Ontogeny of monoamineneurons in the locus coeruleus, raphe nuclei and substan-tia nigra of the rat. I. Cell differentiation. J Comp Neurol155:469–482.
Levitt M, Spector S, Sjoerdsma A, Udenfriend S. 1965.Elucidation of the rate-limiting step in norepinephrinebiosynthesis in the perfused guinea pig heart. J Path ExpTher 148:1–8.
Lo L, Morin X, Brunet JF, Anderson DJ. 1999. Specifica-tion of neurotransmitter identity by Phox2 proteins inneural crest stem cells. Neuron 22:693–705.
Lopez-Rovira T, Chalaux E, Rosa JL, Bartrons R, VenturaF. 2000. Interaction and functional cooperation of NF-�Bwith Smads. J Biol Chem 275:28937–28946.
Loughlin SE, Foote SL, Grzanna R. 1986. Efferent projec-tions of nucleus locus coeruleus: morphologic subpopu-lations have different efferent targets. Neuroscience 18:307–319.
Marshall KC, Christie MJ, Finlayson PG, Williams JT.1991. Developmental aspects of the locus coeruleus-nor-adrenaline system. Prog Brain Res 88:173–185.
Mehler MF, Mabie PC, Zhang D, Kessler JA. 1997. Bonemorphogenetic proteins in the nervous system. TrendsNeurosci 20:309–317.
Merlio JP, Ernfors P, Jaber M, Persson H. 1992. Molecularcloning of rat trkC and distribution of cells expressing
messenger RNAs for members of the trk family in the ratcentral nervous system. Neuroscience 51:513–532.
Meyer-Franke A, Wilkinson GA, Kruttgen A, Hu M, MunroE, Hanson MG, Reichardt LF, Barres BA. 1998. Depo-larization and cAMP elevation rapidly recruit TrkB to theplasma membrane of CNS neurons. Neuron 21:681–693.
Moore MW, Klein RD, Farinas I, Sauer H, Armanini M,Phillips H, Reichardt LF, Ryan AM, Carver-Moore K,Rosenthal A. 1996. Renal and neuronal abnormalities inmice lacking GDNF. Nature 382:76–79.
Nieuwenhuys R. 1985. Chemoarchitecture of the brain.New York: Springer-Verlag. p 19–30.
Osaka H, Sabban EL. 1997. Requirement for cAMP/cal-cium response element but not AP-1 site in fibroblastgrowth factor-2-elicited activation of tyrosine hydroxy-lase gene expression in PC12 cells. Mol Brain Res 49:222–228.
Park DS, Morris EJ, Stefanis L, Troy CM, Shelanski ML,Geller HM, Greene LA. 1998. Multiple pathways ofneuronal death induced by DNA-damaging agents, NGFdeprivation, and oxidative stress. J Neurosci 18:830–840.
Pattyn A, Morin X, Cremer H, Goridis C, Brunet JF. 1997.Expression and interactions of the two closely relatedhomeobox genes Phox2a and Phox2b during neurogen-esis. Development 124:4065–4075.
Pichel JG, Shen L, Sheng HZ, Granholm AC, Drago J,Grinberg A, Lee EJ, Huang SP, Saarma M, Hoffer BJ,Sariola H, Westphal H. 1996. Defects in enteric innerva-tion and kidney development in mice lacking GDNF.Nature 382:73–76.
Reiriz J, Espejo M, Ventura F, Ambrosio S, Alberch J.1999. Bone morphogenetic protein-2 promotes dissoci-ated effects on the number and differentiation of culturedventral mesencephalic dopaminergic neurons. J Neuro-biol 38:161–170.
Reissmann E, Ernsberger U, Francis-West PH, Rueger D,Brickell PM, Rohrer H. 1996. Involvement of bone mor-phogenetic protein-4 and bone morphogenetic protein-7in the differentiation of the adrenergic phenotype in de-veloping sympathetic neurons. Development 122:2079–2088.
Sanchez MP, Silos-Santiago I, Frisen J, He B, Lira SA,Barbacid M. 1996. Renal agenesis and the absence ofenteric neurons in mice lacking GDNF. Nature 382:70–73.
Shah NM, Groves AK, Anderson DJ. 1996. Alternativeneural crest cell fates are instructively promoted byTGF� superfamily members. Cell 85:331–343.
Shen S, Wiemelt AP, McMorris FA, Barres BA. 1999.Retinal ganglion cells lose trophic responsiveness afteraxotomy. Neuron 23:285–295.
Sklair-Tavron L, Nestler EJ, Segal M. 1994. Locus coer-uleus (LC)—target interaction and cAMP in control ofLC development. Brain Res Bull 35:397–402.
Sklair-Tavron L, Segal M. 1993. Neurotrophic effects ofcAMP generating systems on central noradrenergic neu-rons. Brain Res 614:257–269.
Trophic Requirements of Locus Coeruleus Neurons 303

Sklair-Tavron L, Shi WX, Lane SB, Harris HW, BunneyBS, Nestler EJ. 1996. Chronic morphine induces visiblechanges in the morphology of mesolimbic dopamine neu-rons. Proc Natl Acad Sci USA 93:1202–1207.
Specht LA, Pickel VM, Joh TH, Reis DJ. 1981. Light-microscopic immunocytochemical localization of ty-rosine hydroxylase in prenatal rat brain. I Early ontogeny.J Comp Neurol 199:233–253.
Thang SH, Kobayashi M, Matsuoka I. 2000. Regulation ofglial cell line-derived neurotrophic factor responsivenessin developing rat sympathetic neurons by retinoic acidand bone morphogenetic protein-2. J Neurosci 20:2917–2925.
Trupp M, Belluardo N, Funakoshi H, Ibanez CF. 1997.Complementary and overlapping expression of glial cell
line-derived neurotrophic factor (GDNF), c-ret proto-oncogene, and GDNF receptor-� indicates multiplemechanisms of trophic actions in the adult rat CNS.J Neurosci 17:3554–3567.
Wanaka A, Johnson EJ, Milbrandt J. 1990. Localization ofFGF receptor mRNA in the adult rat central nervoussystem by in situ hybridization. Neuron 5:267–281.
Zhang D, Mehler MF, Song Q, Kessler JA. 1998. Develop-ment of bone morphogenetic protein receptors in thenervous system and possible roles in regulating trkCexpression. J Neurosci 18:3314–3326.
Zigmond RE, Schwarzschild MA, Rittenhouse AR. 1989.Acute regulation of tyrosine hydroxylase by nerve activ-ity and by neurotransmitters via phosphorylation. AnnuRev Neurosci 12:415–461.
304 Reiriz et al.
Copyright © 2022 FDOKUMEN




















