
Tryptophan-derived secondary metabolites in Arabidopsis thaliana confer non-host resistance to...
13
Tryptophan-derived secondary metabolites in Arabidopsis thaliana confer non-host resistance to necrotrophic Plectosphaerella cucumerina fungi Andrea Sanchez-Vallet 1,† , Brisa Ramos 1,† , Pawel Bednarek 2 , Gemma Lo ´ pez 1 , Mariola Pis ´ lewska-Bednarek 2 , Paul Schulze-Lefert 2 and Antonio Molina 1,* 1 Centro de Biotecnologı ´a y Geno ´ mica de Plantas (UPM-INIA), Universidad Polite ´ cnica de Madrid, Campus Montegancedo, E-28223-Pozuelo de Alarco ´ n, Madrid, Spain, and 2 Department of Plant Microbe Interactions, Max Planck Institut fu ¨ r Zu ¨ chtungsforschung, Carl-von-Linne ´ -Weg 10, D-50829 Ko ¨ ln, Germany Received 1 March 2010; revised 23 March 2010; accepted 31 March 2010; published online 14 May 2010. * For correspondence (fax +34 917157721; e-mail [email protected]). † These authors contributed equally to this work. SUMMARY A defence pathway contributing to non-host resistance to biotrophic fungi in Arabidopsis involves the synthesis and targeted delivery of the tryptophan (trp)-derived metabolites indol glucosinolates (IGs) and camalexin at pathogen contact sites. We have examined whether these metabolites are also rate-limiting for colonization by necrotrophic fungi. Inoculation of Arabidopsis with adapted or non-adapted isolates of the ascomycete Plectosphaerella cucumerina triggers the accumulation of trp-derived metabolites. We found that their depletion in cyp79B2 cyp79B3 mutants renders Arabidopsis fully susceptible to each of three tested non- adapted P. cucumerina isolates, and super-susceptible to an adapted P. cucumerina isolate. This assigns a key role to trp-derived secondary metabolites in limiting the growth of both non-adapted and adapted necrotrophic fungi. However, 4-methoxy-indol-3-ylmethylglucosinolate, which is generated by the P450 monooxygenase CYP81F2, and hydrolyzed by PEN2 myrosinase, together with the antimicrobial camalexin play a minor role in restricting the growth of the non-adapted necrotrophs. This contrasts with a major role of these two trp-derived phytochemicals in limiting invasive growth of non-adapted biotrophic powdery mildew fungi, thereby implying the existence of other unknown trp-derived metabolites in resistance responses to non-adapted necrotrophic P. cucumerina. Impaired defence to non-adapted P. cucumerina, but not to the non- adapted biotrophic fungus Erysiphe pisi, on cyp79B2 cyp79B3 plants is largely restored in the irx1 background, which shows a constitutive accumulation of antimicrobial peptides. Our findings imply differential contribu- tions of antimicrobials in non-host resistance to necrotrophic and biotrophic pathogens. Keywords: indolglucosinolate, plant innate immunity, necrotrophic fungi, antifungal compounds, non-adapted fungi. INTRODUCTION Plants are constantly exposed to a wide variety of potentially pathogenic microorganisms, but only a small proportion, the host-adapted pathogens, are able to successfully colonize a plant species and cause disease. Host range changes are rare in nature, and for this reason the mechanisms underlying ‘durable resistance’ in non-host plants are of growing inter- est in plant–microbe interaction studies. Non-host plants were shown to respond to inoculation with non-adapted pathogens with inducible responses that appear to be shared with host defence responses in interactions with adapted pathogens (Heath, 2000; Thordal-Christensen, 2003). Non-host resistance in Arabidopsis confers immunity to non-adapted biotrophic fungi, such as powdery mildews that colonize barley or pea in nature (Collins et al., 2003; Lipka et al., 2005; Bednarek et al., 2009). Several Arabidopsis pen mutants (pen1, pen2 and pen3) show an increased penetration of these non-adapted biotrophic fungi in leaf epidermal cells, a process that is critical for nutrient uptake ª 2010 The Authors 115 Journal compilation ª 2010 Blackwell Publishing Ltd The Plant Journal (2010) 63, 115–127 doi: 10.1111/j.1365-313X.2010.04224.x
Transcript of Tryptophan-derived secondary metabolites in Arabidopsis thaliana confer non-host resistance to...

Tryptophan-derived secondary metabolites in Arabidopsisthaliana confer non-host resistance to necrotrophicPlectosphaerella cucumerina fungi
Andrea Sanchez-Vallet1,†, Brisa Ramos1,†, Paweł Bednarek2, Gemma Lopez1, Mariola Pislewska-Bednarek2,
Paul Schulze-Lefert2 and Antonio Molina1,*
1Centro de Biotecnologıa y Genomica de Plantas (UPM-INIA), Universidad Politecnica de Madrid, Campus Montegancedo,
E-28223-Pozuelo de Alarcon, Madrid, Spain, and2Department of Plant Microbe Interactions, Max Planck Institut fur Zuchtungsforschung, Carl-von-Linne-Weg 10, D-50829 Koln,
Germany
Received 1 March 2010; revised 23 March 2010; accepted 31 March 2010; published online 14 May 2010.*For correspondence (fax +34 917157721; e-mail [email protected]).†These authors contributed equally to this work.
SUMMARY
A defence pathway contributing to non-host resistance to biotrophic fungi in Arabidopsis involves the
synthesis and targeted delivery of the tryptophan (trp)-derived metabolites indol glucosinolates (IGs) and
camalexin at pathogen contact sites. We have examined whether these metabolites are also rate-limiting for
colonization by necrotrophic fungi. Inoculation of Arabidopsis with adapted or non-adapted isolates of the
ascomycete Plectosphaerella cucumerina triggers the accumulation of trp-derived metabolites. We found that
their depletion in cyp79B2 cyp79B3 mutants renders Arabidopsis fully susceptible to each of three tested non-
adapted P. cucumerina isolates, and super-susceptible to an adapted P. cucumerina isolate. This assigns a key
role to trp-derived secondary metabolites in limiting the growth of both non-adapted and adapted
necrotrophic fungi. However, 4-methoxy-indol-3-ylmethylglucosinolate, which is generated by the P450
monooxygenase CYP81F2, and hydrolyzed by PEN2 myrosinase, together with the antimicrobial camalexin
play a minor role in restricting the growth of the non-adapted necrotrophs. This contrasts with a major role of
these two trp-derived phytochemicals in limiting invasive growth of non-adapted biotrophic powdery mildew
fungi, thereby implying the existence of other unknown trp-derived metabolites in resistance responses to
non-adapted necrotrophic P. cucumerina. Impaired defence to non-adapted P. cucumerina, but not to the non-
adapted biotrophic fungus Erysiphe pisi, on cyp79B2 cyp79B3 plants is largely restored in the irx1 background,
which shows a constitutive accumulation of antimicrobial peptides. Our findings imply differential contribu-
tions of antimicrobials in non-host resistance to necrotrophic and biotrophic pathogens.
Keywords: indolglucosinolate, plant innate immunity, necrotrophic fungi, antifungal compounds,
non-adapted fungi.
INTRODUCTION
Plants are constantly exposed to a wide variety of potentially
pathogenic microorganisms, but only a small proportion, the
host-adapted pathogens, are able to successfully colonize a
plant species and cause disease. Host range changes are rare
in nature, and for this reason the mechanisms underlying
‘durable resistance’ in non-host plants are of growing inter-
est in plant–microbe interaction studies. Non-host plants
were shown to respond to inoculation with non-adapted
pathogens with inducible responses that appear to be shared
with host defence responses in interactions with adapted
pathogens (Heath, 2000; Thordal-Christensen, 2003).
Non-host resistance in Arabidopsis confers immunity to
non-adapted biotrophic fungi, such as powdery mildews
that colonize barley or pea in nature (Collins et al., 2003;
Lipka et al., 2005; Bednarek et al., 2009). Several Arabidopsis
pen mutants (pen1, pen2 and pen3) show an increased
penetration of these non-adapted biotrophic fungi in leaf
epidermal cells, a process that is critical for nutrient uptake
ª 2010 The Authors 115Journal compilation ª 2010 Blackwell Publishing Ltd
The Plant Journal (2010) 63, 115–127 doi: 10.1111/j.1365-313X.2010.04224.x

by these pathogens (Collins et al., 2003). The PEN1 syntaxin
is involved in a vesicle-mediated secretory defence pathway
that seems to be specific for biotrophic fungi (Collins et al.,
2003; Kwon et al., 2008). PEN2 is a myrosinase that accu-
mulates underneath powdery mildew contact sites, where
the enzyme initiates the metabolism of a group of trypto-
phan (trp)-derived compounds, known as indole glucosino-
lates (IGs), to release potential antimicrobial products (Lipka
et al., 2005; Bednarek et al., 2009). Specifically, 4-methoxy-
indol-3-ylmethyl glucosinolate (4MI3G), synthesized by
CYP81F2, is the biologically relevant substrate of PEN2
activity against non-adapted powdery mildews (Bednarek
et al., 2009; Pfalz et al., 2009). Formation of the PEN2-depen-
dent bioactive end product(s) relies on high glutathione
levels, as PAD2, encoding a c-glutamylcysteine synthetase
(Parisy et al., 2007), is indispensable for the formation of
indol-3-ylmethylamine (I3A) and raphanusamic acid (RA), as
well as extracellular growth termination of the non-adapted
powdery mildews (Bednarek et al., 2009). Interestingly,
under particular stress conditions the pad2-1 mutant, which
contains approximately 20% residual glutathione, accumu-
lates reduced levels of the trp-derived phytoalexin camalexin
(3-thiazol-2¢-yl-indole), IGs and aliphatic glucosinolates (Gs),
and is super-susceptible to infection by adapted fungi,
oomycete and bacteria (Glazebrook and Ausubel, 1994;
Roetschi et al., 2001; Schlaeppi et al., 2008; Bednarek et al.,
2009). PEN3 encodes a pleiotropic drug resistance (PDR)
ATP-binding cassette (ABC) transporter implicated in the
secretion of antimicrobial products, including those result-
ing from PEN2 activity (Stein et al., 2006). The PEN2/PEN3-
dependent extracellular defence is of particular interest
because this pathway restricts the growth of a broader
spectrum of microbial pathogens, including biotrophic and
necrotrophic fungi, and oomycetes (Lipka et al., 2005; Stein
et al., 2006; Bednarek et al., 2009; Maeda et al., 2009).
In addition to the PEN2/PEN3-dependent defence path-
way, the phytoalexin camalexin that is essential for Arabid-
opsis resistance to several adapted necrotrophic fungi
(Thomma et al., 1999a; van Wees et al., 2003; Bohman et al.,
2004; Denby et al., 2004; Kliebenstein et al., 2005; Glawisch-
nig, 2007; Van Baarlen et al., 2007), was found to be active in
the post-invasive defence barrier in non-host resistance to
powdery mildews (Bednarek et al., 2009). The pad3 mutant,
impaired in the last step of camalexin biosynthesis (Schu-
hegger et al., 2006), had the same infection phenotype as
wild-type plants, but the double mutant pen2 pad3 was not
only impaired in penetration resistance, but also supported
extensive hyphal growth of the non-adapted fungus Erysi-
phe pisi (Bednarek et al., 2009). The pen2 pad3 infection
phenotype was comparable with that of the double mutant
cyp79B2 cyp79B3, which is defective in the P450 monoox-
ygenase-catalyzed conversion of tryptophan to indole-3-
acetaldoxime (IAOX), a precursor of most known trp-derived
products, including IGs, such as 4MI3G and camalexin (Zhao
et al., 2002; Bednarek et al., 2009). Together, these data
indicate that an important component of Arabidopsis non-
host resistance to biotrophic powdery mildew fungi is the
accumulation and sequential action of PEN2-derived IGs
hydrolysis products and camalexin (Bednarek et al., 2009).
The majority of necrotrophic fungi, such as Botrytis
cinerea or Sclerotinia sclerotiorum, have a broad host range,
although for several well-studied necrothrophs, like Coch-
liobolus and Alternaria spp., a narrower host range has been
described (van Kan, 2006). Some ascomycete fungi, such as
the necrotrophs Plectosphaerella cucumerina or B. cinerea,
can proliferate and develop on dead and decaying tissues,
causing severe disease in a wide range of plants, including
Arabidopsis (Cramer and Lawrence, 2004; Bolton et al.,
2006; van Kan, 2006; Williamson et al., 2007).
Our current knowledge of the molecular basis of non-host
resistance to non-adapted necrotrophic pathogens is scarce.
By contrast, growth restriction of adapted necrotrophs in
Arabidopsis was found to be multigenic (Denby et al., 2004;
Llorente et al., 2005), and to depend on the precise regula-
tion of the ethylene (ET), jasmonate (JA), salicylic acid (SA)
or auxin signalling pathways, as well as the b subunit of
heterotrimeric G-protein (AGB1; Thomma et al., 1998,
1999b; Berrocal-Lobo et al., 2002; Ferrari et al., 2003;
Llorente et al., 2005; Hernandez-Blanco et al., 2007; Llorente
et al., 2008). Plant cell wall structure and composition are
also determinants for the success of necrotrophic fungal
colonization (Somerville et al., 2004). The irregular xylem
(irx) mutants, impaired in the cellulose synthase (CESA)
subunits specifically required for secondary cell wall
formation, showed enhanced resistance to both adapted
necrotrophic (e.g. P. cucumerina) and biotrophic (e.g.
Golovinomyces cichoracearum) fungi (Hernandez-Blanco
et al., 2007). The irx1-mediated broad spectrum resistance
may rely on the constitutive accumulation of antibiotic
compounds, as revealed by comparative transcriptome
analyses (Hernandez-Blanco et al., 2007).
In this study, we first identified three non-adapted P. cu-
cumerina isolates that were unable to colonize Arabidopsis
Col-0 wild-type plants. Using this pathosystem, we found
that in addition to the biosynthesis of camalexin and
CYP81F2/PEN2-derived metabolites, the engagement of
additional uncharacterized trp-derived compounds is
required for effective non-host resistance to necrotrophs.
The results presented here also suggest a differential
contribution of distinct sets of antimicrobials in non-host
resistance to necrotrophic and biotrophic pathogens.
RESULTS
Plectosphaerella cucumerina non-adapted isolates fail
to colonize Arabidopsis plants
We selected three P. cucumerina isolates (Pc1187, Pc2127
and Pc2125) that colonise Nicotiana tabacinum, Viola spp.
116 Andrea Sanchez-Vallet et al.
ª 2010 The AuthorsJournal compilation ª 2010 Blackwell Publishing Ltd, The Plant Journal, (2010), 63, 115–127
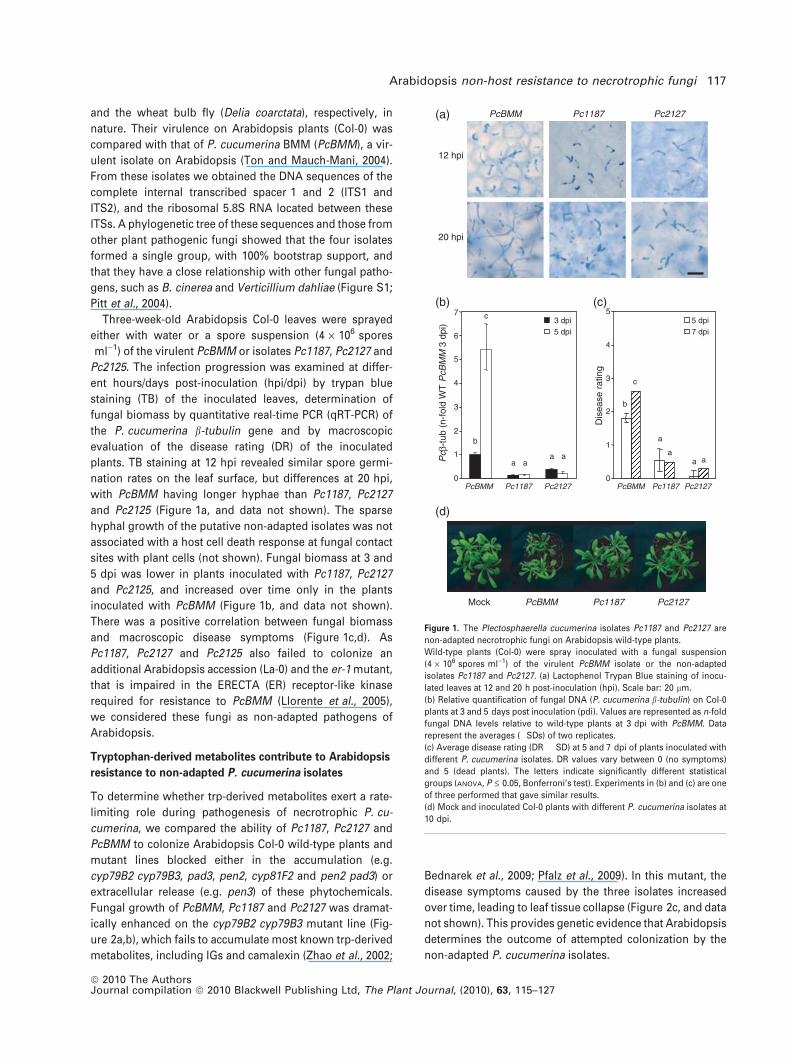
and the wheat bulb fly (Delia coarctata), respectively, in
nature. Their virulence on Arabidopsis plants (Col-0) was
compared with that of P. cucumerina BMM (PcBMM), a vir-
ulent isolate on Arabidopsis (Ton and Mauch-Mani, 2004).
From these isolates we obtained the DNA sequences of the
complete internal transcribed spacer 1 and 2 (ITS1 and
ITS2), and the ribosomal 5.8S RNA located between these
ITSs. A phylogenetic tree of these sequences and those from
other plant pathogenic fungi showed that the four isolates
formed a single group, with 100% bootstrap support, and
that they have a close relationship with other fungal patho-
gens, such as B. cinerea and Verticillium dahliae (Figure S1;
Pitt et al., 2004).
Three-week-old Arabidopsis Col-0 leaves were sprayed
either with water or a spore suspension (4 · 106 spores
ml)1) of the virulent PcBMM or isolates Pc1187, Pc2127 and
Pc2125. The infection progression was examined at differ-
ent hours/days post-inoculation (hpi/dpi) by trypan blue
staining (TB) of the inoculated leaves, determination of
fungal biomass by quantitative real-time PCR (qRT-PCR) of
the P. cucumerina b-tubulin gene and by macroscopic
evaluation of the disease rating (DR) of the inoculated
plants. TB staining at 12 hpi revealed similar spore germi-
nation rates on the leaf surface, but differences at 20 hpi,
with PcBMM having longer hyphae than Pc1187, Pc2127
and Pc2125 (Figure 1a, and data not shown). The sparse
hyphal growth of the putative non-adapted isolates was not
associated with a host cell death response at fungal contact
sites with plant cells (not shown). Fungal biomass at 3 and
5 dpi was lower in plants inoculated with Pc1187, Pc2127
and Pc2125, and increased over time only in the plants
inoculated with PcBMM (Figure 1b, and data not shown).
There was a positive correlation between fungal biomass
and macroscopic disease symptoms (Figure 1c,d). As
Pc1187, Pc2127 and Pc2125 also failed to colonize an
additional Arabidopsis accession (La-0) and the er-1 mutant,
that is impaired in the ERECTA (ER) receptor-like kinase
required for resistance to PcBMM (Llorente et al., 2005),
we considered these fungi as non-adapted pathogens of
Arabidopsis.
Tryptophan-derived metabolites contribute to Arabidopsis
resistance to non-adapted P. cucumerina isolates
To determine whether trp-derived metabolites exert a rate-
limiting role during pathogenesis of necrotrophic P. cu-
cumerina, we compared the ability of Pc1187, Pc2127 and
PcBMM to colonize Arabidopsis Col-0 wild-type plants and
mutant lines blocked either in the accumulation (e.g.
cyp79B2 cyp79B3, pad3, pen2, cyp81F2 and pen2 pad3) or
extracellular release (e.g. pen3) of these phytochemicals.
Fungal growth of PcBMM, Pc1187 and Pc2127 was dramat-
ically enhanced on the cyp79B2 cyp79B3 mutant line (Fig-
ure 2a,b), which fails to accumulate most known trp-derived
metabolites, including IGs and camalexin (Zhao et al., 2002;
Bednarek et al., 2009; Pfalz et al., 2009). In this mutant, the
disease symptoms caused by the three isolates increased
over time, leading to leaf tissue collapse (Figure 2c, and data
not shown). This provides genetic evidence that Arabidopsis
determines the outcome of attempted colonization by the
non-adapted P. cucumerina isolates.
(a)
(d)
Mock PcBMM Pc1187 Pc2127
12 hpi
20 hpi
PcBMM Pc2127Pc1187
Pcβ
-tub
(n-
fold
WT
PcB
MM
3 d
pi)
(c)(b)3 dpi5 dpi
PcBMM Pc1187 Pc2127 Pc1187PcBMM Pc21270
1
2
3
4
5
Dis
ease
rat
ing
5 dpi7 dpi
c
a
b
aa
0
1
2
3
4
5
6
7
aaaa
b
c
a
Figure 1. The Plectosphaerella cucumerina isolates Pc1187 and Pc2127 are
non-adapted necrotrophic fungi on Arabidopsis wild-type plants.
Wild-type plants (Col-0) were spray inoculated with a fungal suspension
(4 · 106 spores ml)1) of the virulent PcBMM isolate or the non-adapted
isolates Pc1187 and Pc2127. (a) Lactophenol Trypan Blue staining of inocu-
lated leaves at 12 and 20 h post-inoculation (hpi). Scale bar: 20 lm.
(b) Relative quantification of fungal DNA (P. cucumerina b-tubulin) on Col-0
plants at 3 and 5 days post inoculation (pdi). Values are represented as n-fold
fungal DNA levels relative to wild-type plants at 3 dpi with PcBMM. Data
represent the averages (�SDs) of two replicates.
(c) Average disease rating (DR � SD) at 5 and 7 dpi of plants inoculated with
different P. cucumerina isolates. DR values vary between 0 (no symptoms)
and 5 (dead plants). The letters indicate significantly different statistical
groups (ANOVA, P £ 0.05, Bonferroni’s test). Experiments in (b) and (c) are one
of three performed that gave similar results.
(d) Mock and inoculated Col-0 plants with different P. cucumerina isolates at
10 dpi.
Arabidopsis non-host resistance to necrotrophic fungi 117
ª 2010 The AuthorsJournal compilation ª 2010 Blackwell Publishing Ltd, The Plant Journal, (2010), 63, 115–127
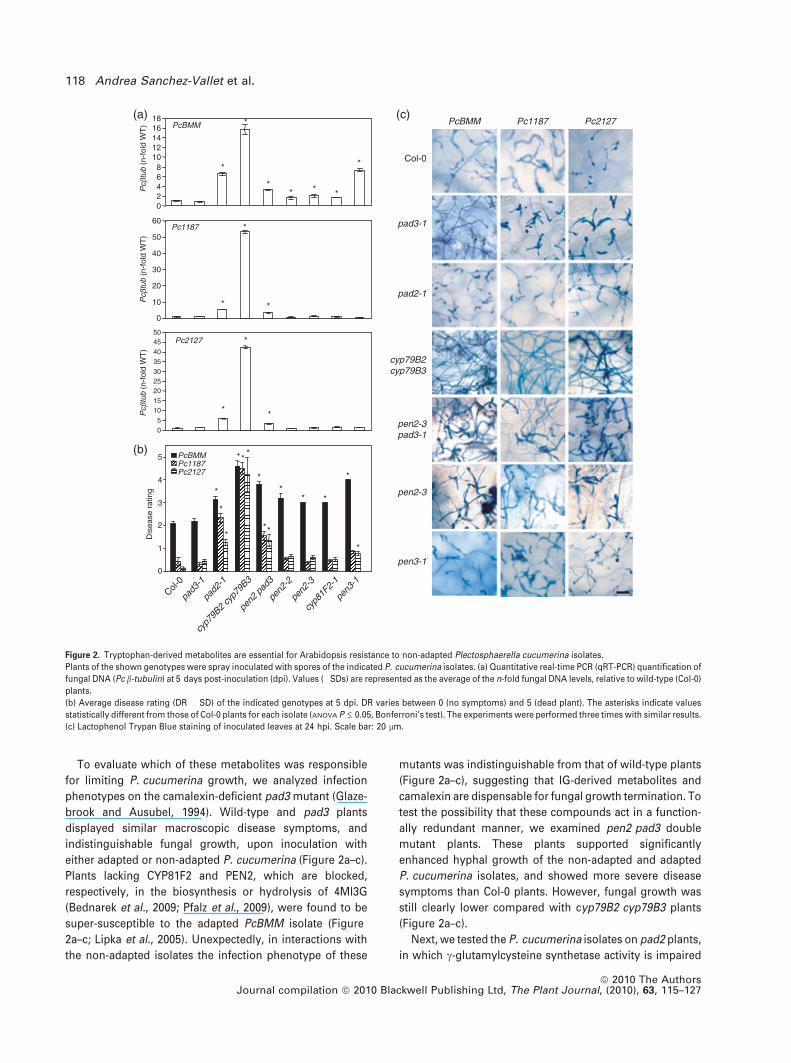
To evaluate which of these metabolites was responsible
for limiting P. cucumerina growth, we analyzed infection
phenotypes on the camalexin-deficient pad3 mutant (Glaze-
brook and Ausubel, 1994). Wild-type and pad3 plants
displayed similar macroscopic disease symptoms, and
indistinguishable fungal growth, upon inoculation with
either adapted or non-adapted P. cucumerina (Figure 2a–c).
Plants lacking CYP81F2 and PEN2, which are blocked,
respectively, in the biosynthesis or hydrolysis of 4MI3G
(Bednarek et al., 2009; Pfalz et al., 2009), were found to be
super-susceptible to the adapted PcBMM isolate (Figure
2a–c; Lipka et al., 2005). Unexpectedly, in interactions with
the non-adapted isolates the infection phenotype of these
mutants was indistinguishable from that of wild-type plants
(Figure 2a–c), suggesting that IG-derived metabolites and
camalexin are dispensable for fungal growth termination. To
test the possibility that these compounds act in a function-
ally redundant manner, we examined pen2 pad3 double
mutant plants. These plants supported significantly
enhanced hyphal growth of the non-adapted and adapted
P. cucumerina isolates, and showed more severe disease
symptoms than Col-0 plants. However, fungal growth was
still clearly lower compared with cyp79B2 cyp79B3 plants
(Figure 2a–c).
Next, we tested the P. cucumerina isolates on pad2 plants,
in which c-glutamylcysteine synthetase activity is impaired
(c)(a)
(b)
0
1
2
3
4
5
Col-0
pad3
-1
Dis
ease
rat
ing
PcBMMPc1187Pc2127
pen2
-2
*
pen2
-3
*
cyp8
1F2-
1
*
cyp7
9B2
cyp7
9B3
***
pen2
pad
3
*
**
pen3
-1
*
*
pad2
-1
*
*
*
PcBMM Pc1187 Pc2127
Col-0
cyp79B2cyp79B3
pen3-1
pad2-1
pen2-3
pad3-1
pen2-3pad3-1
05
101520253035404550
Pcb
tub
(n-f
old
WT
)P
cbtu
b (n
-fol
d W
T)
Pcb
tub
(n-f
old
WT
)
Pc2127
*
*
*
Pc1187
0
10
20
30
40
50
60
*
*
*
02468
1012141618
PcBMM
*
*
** * *
*
Figure 2. Tryptophan-derived metabolites are essential for Arabidopsis resistance to non-adapted Plectosphaerella cucumerina isolates.
Plants of the shown genotypes were spray inoculated with spores of the indicated P. cucumerina isolates. (a) Quantitative real-time PCR (qRT-PCR) quantification of
fungal DNA (Pc b-tubulin) at 5 days post-inoculation (dpi). Values (�SDs) are represented as the average of the n-fold fungal DNA levels, relative to wild-type (Col-0)
plants.
(b) Average disease rating (DR � SD) of the indicated genotypes at 5 dpi. DR varies between 0 (no symptoms) and 5 (dead plant). The asterisks indicate values
statistically different from those of Col-0 plants for each isolate (ANOVA P £ 0.05, Bonferroni’s test). The experiments were performed three times with similar results.
(c) Lactophenol Trypan Blue staining of inoculated leaves at 24 hpi. Scale bar: 20 lm.
118 Andrea Sanchez-Vallet et al.
ª 2010 The AuthorsJournal compilation ª 2010 Blackwell Publishing Ltd, The Plant Journal, (2010), 63, 115–127
and lower levels of trp derivatives accumulate upon infec-
tion with a diverse range of pathogens (Glazebrook and
Ausubel, 1994; Parisy et al., 2007; Schlaeppi et al., 2008;
Bednarek et al., 2009). Notably, pad2 plants supported
enhanced fungal growth of the adapted and non-adapted
isolates (Figure 2a–c). However, fungal growth limitation of
the latter isolates was essentially unaffected in pen3 plants
(Figure 2; Stein et al., 2006). This contrasts with a major
contribution of PEN3 in limiting hyphal growth of the
adapted PcBMM isolate (Figure 2a,b). In summary, these
results suggest that the accumulation and delivery of
IG-derived compounds and camalexin are required for full
resistance to non-adapted P. cucumerina. However, this
must involve other yet unknown trp-derived metabolites
than those acting against biotrophic non-adapted powdery
mildews (Bednarek et al., 2009).
Plectosphaerella cucumerina non-adapted isolates induce
the accumulation of trp-derived metabolites
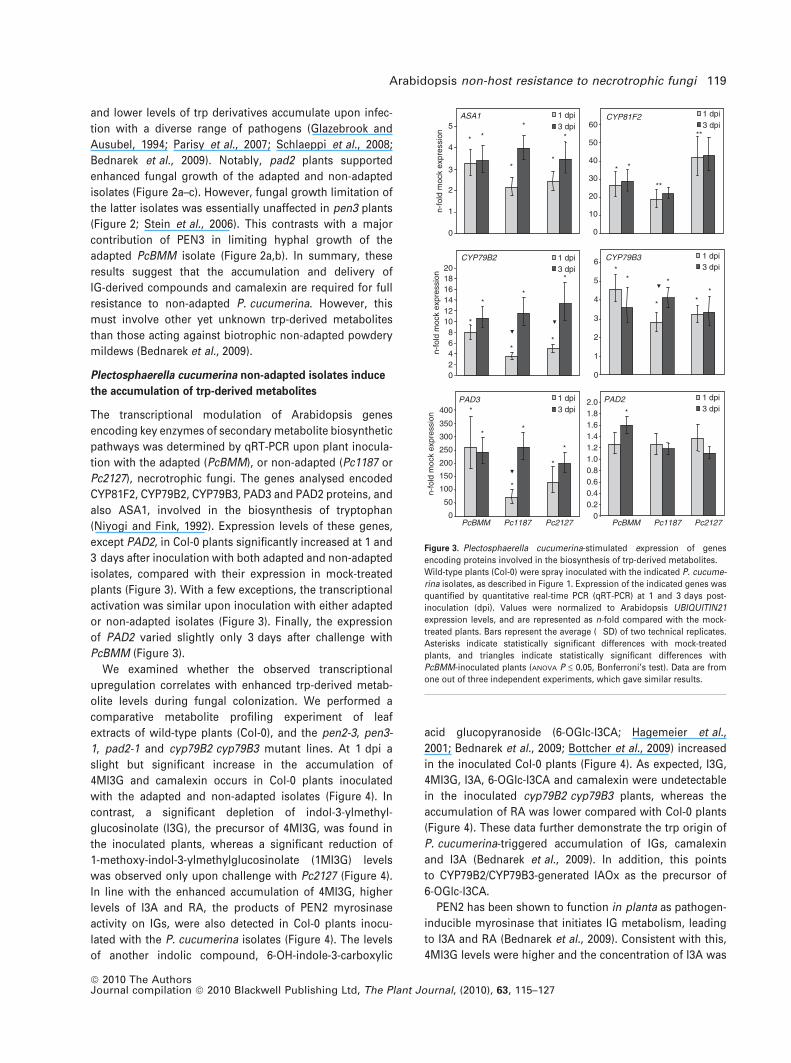
The transcriptional modulation of Arabidopsis genes
encoding key enzymes of secondary metabolite biosynthetic
pathways was determined by qRT-PCR upon plant inocula-
tion with the adapted (PcBMM), or non-adapted (Pc1187 or
Pc2127), necrotrophic fungi. The genes analysed encoded
CYP81F2, CYP79B2, CYP79B3, PAD3 and PAD2 proteins, and
also ASA1, involved in the biosynthesis of tryptophan
(Niyogi and Fink, 1992). Expression levels of these genes,
except PAD2, in Col-0 plants significantly increased at 1 and
3 days after inoculation with both adapted and non-adapted
isolates, compared with their expression in mock-treated
plants (Figure 3). With a few exceptions, the transcriptional
activation was similar upon inoculation with either adapted
or non-adapted isolates (Figure 3). Finally, the expression
of PAD2 varied slightly only 3 days after challenge with
PcBMM (Figure 3).
We examined whether the observed transcriptional
upregulation correlates with enhanced trp-derived metab-
olite levels during fungal colonization. We performed a
comparative metabolite profiling experiment of leaf
extracts of wild-type plants (Col-0), and the pen2-3, pen3-
1, pad2-1 and cyp79B2 cyp79B3 mutant lines. At 1 dpi a
slight but significant increase in the accumulation of
4MI3G and camalexin occurs in Col-0 plants inoculated
with the adapted and non-adapted isolates (Figure 4). In
contrast, a significant depletion of indol-3-ylmethyl-
glucosinolate (I3G), the precursor of 4MI3G, was found in
the inoculated plants, whereas a significant reduction of
1-methoxy-indol-3-ylmethylglucosinolate (1MI3G) levels
was observed only upon challenge with Pc2127 (Figure 4).
In line with the enhanced accumulation of 4MI3G, higher
levels of I3A and RA, the products of PEN2 myrosinase
activity on IGs, were also detected in Col-0 plants inocu-
lated with the P. cucumerina isolates (Figure 4). The levels
of another indolic compound, 6-OH-indole-3-carboxylic
acid glucopyranoside (6-OGlc-I3CA; Hagemeier et al.,
2001; Bednarek et al., 2009; Bottcher et al., 2009) increased
in the inoculated Col-0 plants (Figure 4). As expected, I3G,
4MI3G, I3A, 6-OGlc-I3CA and camalexin were undetectable
in the inoculated cyp79B2 cyp79B3 plants, whereas the
accumulation of RA was lower compared with Col-0 plants
(Figure 4). These data further demonstrate the trp origin of
P. cucumerina-triggered accumulation of IGs, camalexin
and I3A (Bednarek et al., 2009). In addition, this points
to CYP79B2/CYP79B3-generated IAOx as the precursor of
6-OGlc-I3CA.
PEN2 has been shown to function in planta as pathogen-
inducible myrosinase that initiates IG metabolism, leading
to I3A and RA (Bednarek et al., 2009). Consistent with this,
4MI3G levels were higher and the concentration of I3A was
0
50
100
150
200
250
300
350
400PAD3 1 dpi
3 dpi
*
*
*
*
*
*
00.20.40.60.81.01.21.41.61.82.0 PAD2
*
1 dpi3 dpi
PcBMM Pc1187 Pc2127 PcBMM Pc1187 Pc2127
n-fo
ld m
ock
expr
essi
onn-
fold
moc
k ex
pres
sion
0
1
2
3
4
5
6 CYP79B3
**
*
*
**
1 dpi3 dpi
02468
101214161820
CYP79B2
*
**
*
**
1 dpi3 dpi
n-fo
ld m
ock
expr
essi
on
0
10
20
30
40
50
60CYP81F2
*
**
**
*
1 dpi3 dpi
0
1
2
3
4
5ASA1
* *
*
*
*
*
1 dpi3 dpi
Figure 3. Plectosphaerella cucumerina-stimulated expression of genes
encoding proteins involved in the biosynthesis of trp-derived metabolites.
Wild-type plants (Col-0) were spray inoculated with the indicated P. cucume-
rina isolates, as described in Figure 1. Expression of the indicated genes was
quantified by quantitative real-time PCR (qRT-PCR) at 1 and 3 days post-
inoculation (dpi). Values were normalized to Arabidopsis UBIQUITIN21
expression levels, and are represented as n-fold compared with the mock-
treated plants. Bars represent the average (�SD) of two technical replicates.
Asterisks indicate statistically significant differences with mock-treated
plants, and triangles indicate statistically significant differences with
PcBMM-inoculated plants (ANOVA P £ 0.05, Bonferroni’s test). Data are from
one out of three independent experiments, which gave similar results.
Arabidopsis non-host resistance to necrotrophic fungi 119
ª 2010 The AuthorsJournal compilation ª 2010 Blackwell Publishing Ltd, The Plant Journal, (2010), 63, 115–127
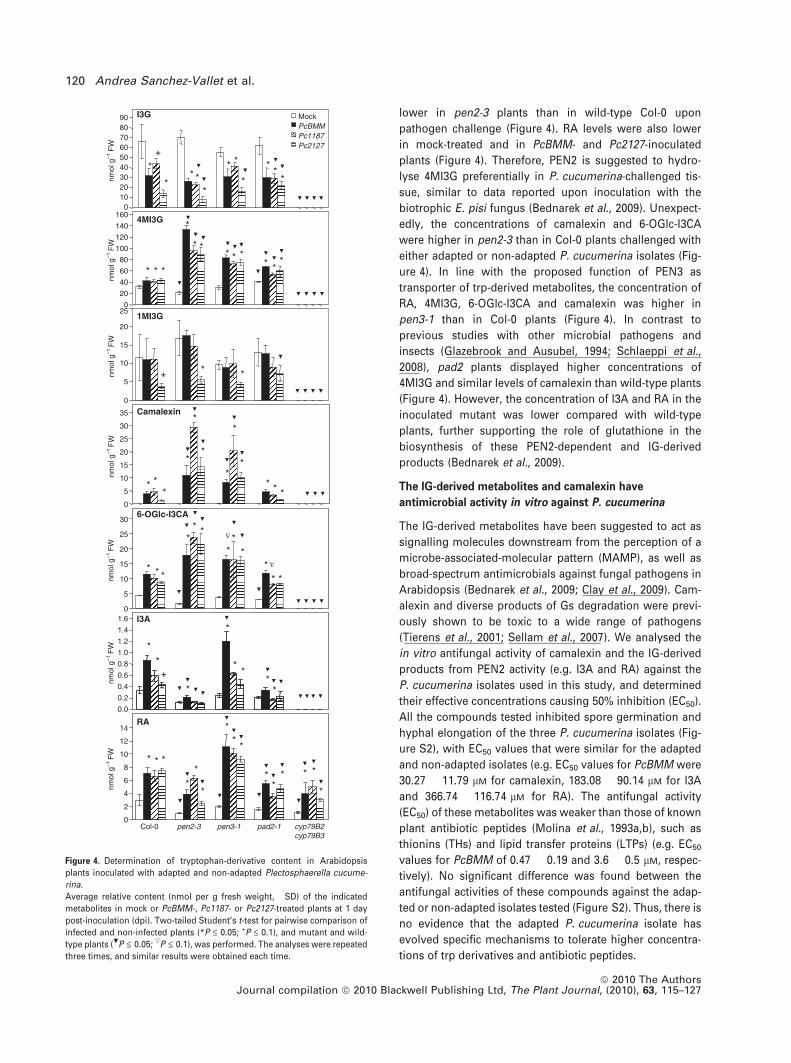
lower in pen2-3 plants than in wild-type Col-0 upon
pathogen challenge (Figure 4). RA levels were also lower
in mock-treated and in PcBMM- and Pc2127-inoculated
plants (Figure 4). Therefore, PEN2 is suggested to hydro-
lyse 4MI3G preferentially in P. cucumerina-challenged tis-
sue, similar to data reported upon inoculation with the
biotrophic E. pisi fungus (Bednarek et al., 2009). Unexpect-
edly, the concentrations of camalexin and 6-OGlc-I3CA
were higher in pen2-3 than in Col-0 plants challenged with
either adapted or non-adapted P. cucumerina isolates (Fig-
ure 4). In line with the proposed function of PEN3 as
transporter of trp-derived metabolites, the concentration of
RA, 4MI3G, 6-OGlc-I3CA and camalexin was higher in
pen3-1 than in Col-0 plants (Figure 4). In contrast to
previous studies with other microbial pathogens and
insects (Glazebrook and Ausubel, 1994; Schlaeppi et al.,
2008), pad2 plants displayed higher concentrations of
4MI3G and similar levels of camalexin than wild-type plants
(Figure 4). However, the concentration of I3A and RA in the
inoculated mutant was lower compared with wild-type
plants, further supporting the role of glutathione in the
biosynthesis of these PEN2-dependent and IG-derived
products (Bednarek et al., 2009).
The IG-derived metabolites and camalexin have
antimicrobial activity in vitro against P. cucumerina
The IG-derived metabolites have been suggested to act as
signalling molecules downstream from the perception of a
microbe-associated-molecular pattern (MAMP), as well as
broad-spectrum antimicrobials against fungal pathogens in
Arabidopsis (Bednarek et al., 2009; Clay et al., 2009). Cam-
alexin and diverse products of Gs degradation were previ-
ously shown to be toxic to a wide range of pathogens
(Tierens et al., 2001; Sellam et al., 2007). We analysed the
in vitro antifungal activity of camalexin and the IG-derived
products from PEN2 activity (e.g. I3A and RA) against the
P. cucumerina isolates used in this study, and determined
their effective concentrations causing 50% inhibition (EC50).
All the compounds tested inhibited spore germination and
hyphal elongation of the three P. cucumerina isolates (Fig-
ure S2), with EC50 values that were similar for the adapted
and non-adapted isolates (e.g. EC50 values for PcBMM were
30.27 � 11.79 lM for camalexin, 183.08 � 90.14 lM for I3A
and 366.74 � 116.74 lM for RA). The antifungal activity
(EC50) of these metabolites was weaker than those of known
plant antibiotic peptides (Molina et al., 1993a,b), such as
thionins (THs) and lipid transfer proteins (LTPs) (e.g. EC50
values for PcBMM of 0.47 � 0.19 and 3.6 � 0.5 lM, respec-
tively). No significant difference was found between the
antifungal activities of these compounds against the adap-
ted or non-adapted isolates tested (Figure S2). Thus, there is
no evidence that the adapted P. cucumerina isolate has
evolved specific mechanisms to tolerate higher concentra-
tions of trp derivatives and antibiotic peptides.
100
120
140
160
0
20
40
60
80
4MI3G
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
1.6 I3A
Col-0 pen2-3 pen3-1 pad2-1 cyp79B2cyp79B3
0
2
4
6
8
10
12
14
nmol
g–1
FW
nmol
g–1
FW
nmol
g–1
FW
nmol
g–1
FW
nmol
g–1
FW
nmol
g–1
FW
nmol
g–1
FW
RA
0
5
10
15
20
25
306-OGlc-I3CA
0
5
10
15
20
251MI3G
0
5
10
15
20
25
30
35 Camalexin
0102030405060708090 I3G
PcBMMPc1187Pc2127
Mock
Figure 4. Determination of tryptophan-derivative content in Arabidopsis
plants inoculated with adapted and non-adapted Plectosphaerella cucume-
rina.
Average relative content (nmol per g fresh weight, �SD) of the indicated
metabolites in mock or PcBMM-, Pc1187- or Pc2127-treated plants at 1 day
post-inoculation (dpi). Two-tailed Student’s t-test for pairwise comparison of
infected and non-infected plants (*P £ 0.05; +P £ 0.1), and mutant and wild-
type plants (.P £ 0.05; ,P £ 0.1), was performed. The analyses were repeated
three times, and similar results were obtained each time.
120 Andrea Sanchez-Vallet et al.
ª 2010 The AuthorsJournal compilation ª 2010 Blackwell Publishing Ltd, The Plant Journal, (2010), 63, 115–127
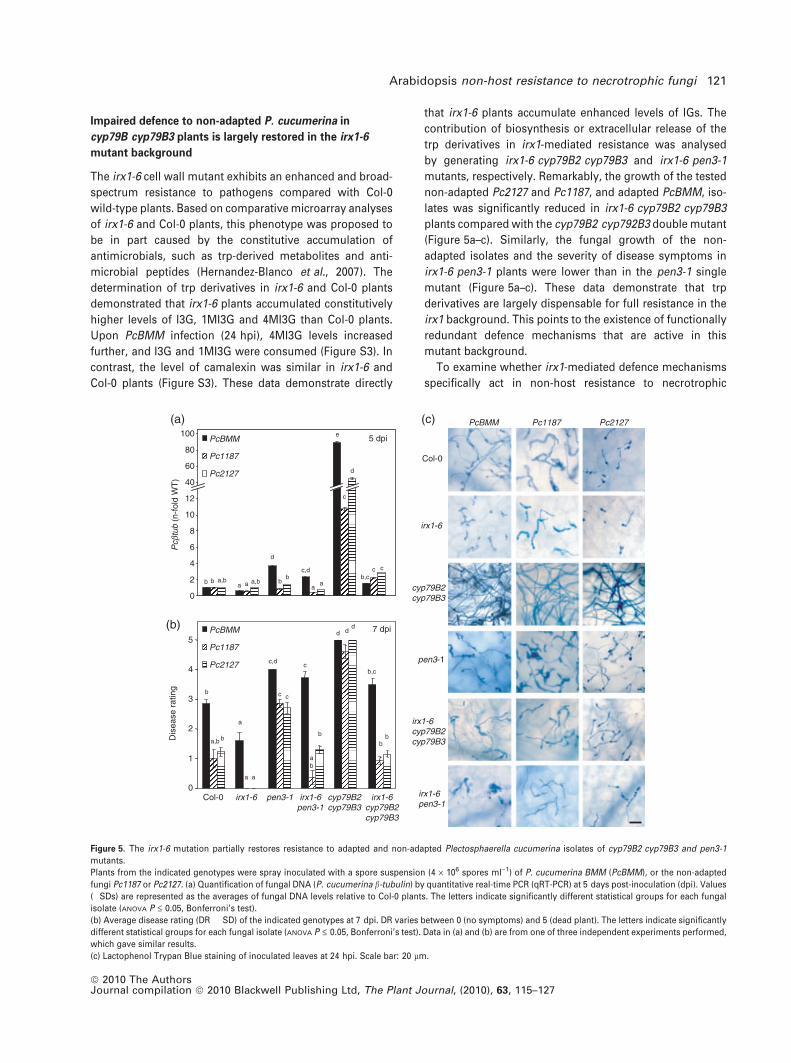
Impaired defence to non-adapted P. cucumerina in
cyp79B cyp79B3 plants is largely restored in the irx1-6
mutant background
The irx1-6 cell wall mutant exhibits an enhanced and broad-
spectrum resistance to pathogens compared with Col-0
wild-type plants. Based on comparative microarray analyses
of irx1-6 and Col-0 plants, this phenotype was proposed to
be in part caused by the constitutive accumulation of
antimicrobials, such as trp-derived metabolites and anti-
microbial peptides (Hernandez-Blanco et al., 2007). The
determination of trp derivatives in irx1-6 and Col-0 plants
demonstrated that irx1-6 plants accumulated constitutively
higher levels of I3G, 1MI3G and 4MI3G than Col-0 plants.
Upon PcBMM infection (24 hpi), 4MI3G levels increased
further, and I3G and 1MI3G were consumed (Figure S3). In
contrast, the level of camalexin was similar in irx1-6 and
Col-0 plants (Figure S3). These data demonstrate directly
that irx1-6 plants accumulate enhanced levels of IGs. The
contribution of biosynthesis or extracellular release of the
trp derivatives in irx1-mediated resistance was analysed
by generating irx1-6 cyp79B2 cyp79B3 and irx1-6 pen3-1
mutants, respectively. Remarkably, the growth of the tested
non-adapted Pc2127 and Pc1187, and adapted PcBMM, iso-
lates was significantly reduced in irx1-6 cyp79B2 cyp79B3
plants compared with the cyp79B2 cyp792B3 double mutant
(Figure 5a–c). Similarly, the fungal growth of the non-
adapted isolates and the severity of disease symptoms in
irx1-6 pen3-1 plants were lower than in the pen3-1 single
mutant (Figure 5a–c). These data demonstrate that trp
derivatives are largely dispensable for full resistance in the
irx1 background. This points to the existence of functionally
redundant defence mechanisms that are active in this
mutant background.
To examine whether irx1-mediated defence mechanisms
specifically act in non-host resistance to necrotrophic
cyp79B2cyp79B30
2
4
6
8
10
12
40
60
80
100
Pcb
tub
(n-f
old
WT
)
b aab
c,d
aaa,ba,b
d
bb b,c
c c
e
c
d
(c)(a)
irx1-6cyp79B2cyp79B3
Col-0 irx1-6 irx1-6pen3-1
pen3-1 cyp79B2cyp79B3
dd
d
0
1
2
3
4
5
Dis
ease
rat
ing
a
b
a
a,b
a
b
c
ab
b
b,c
bb
c,d
c c
7 dpiPcBMM
Pc1187
Pc2127
(b)
Col-0
irx1-6
irx1-6cyp79B2cyp79B3
irx1-6pen3-1
PcBMM Pc1187 Pc2127
pen3-1
5 dpiPcBMM
Pc1187
Pc2127
Figure 5. The irx1-6 mutation partially restores resistance to adapted and non-adapted Plectosphaerella cucumerina isolates of cyp79B2 cyp79B3 and pen3-1
mutants.
Plants from the indicated genotypes were spray inoculated with a spore suspension (4 · 106 spores ml)1) of P. cucumerina BMM (PcBMM), or the non-adapted
fungi Pc1187 or Pc2127. (a) Quantification of fungal DNA (P. cucumerina b-tubulin) by quantitative real-time PCR (qRT-PCR) at 5 days post-inoculation (dpi). Values
(�SDs) are represented as the averages of fungal DNA levels relative to Col-0 plants. The letters indicate significantly different statistical groups for each fungal
isolate (ANOVA P £ 0.05, Bonferroni’s test).
(b) Average disease rating (DR � SD) of the indicated genotypes at 7 dpi. DR varies between 0 (no symptoms) and 5 (dead plant). The letters indicate significantly
different statistical groups for each fungal isolate (ANOVA P £ 0.05, Bonferroni’s test). Data in (a) and (b) are from one of three independent experiments performed,
which gave similar results.
(c) Lactophenol Trypan Blue staining of inoculated leaves at 24 hpi. Scale bar: 20 lm.
Arabidopsis non-host resistance to necrotrophic fungi 121
ª 2010 The AuthorsJournal compilation ª 2010 Blackwell Publishing Ltd, The Plant Journal, (2010), 63, 115–127

P. cucumerina, we tested the infection phenotype of wild-
type Col-0 plants and various irx1-containing genotypes
with the non-adapted biotrophic fungus E. pisi (Bednarek
et al., 2009). As shown in Figure 6, E. pisi entry rates in Col-0
and irx1-6 mutant plants were statistically indistinguishable.
Of note, the E. pisi entry rates on irx1-6 pen3-1 and irx1-6
cyp79B2 cyp79B3 plants were significantly higher than
those observed in the corresponding pen3-1 and cyp79B2
cyp79B3 parental lines (Figure 6), which, as reported, sup-
ported higher entry rates than Col-0 plants (Stein et al., 2006;
Bednarek et al., 2009). These data suggest that irx1-medi-
ated defence against necrotrophic P. cucumerina negatively
interferes with pre-invasive non-host resistance responses
to the non-adapted biotrophic fungal pathogen.
Function of SA, JA and ET signalling pathways in
Arabidopsis non-host resistance to non-adapted
P. cucumerina fungi
In Arabidopsis, the SA, JA and ET signalling pathways are
essential to restrict the growth of the adapted PcBMM
isolate, as well as other necrotrophic fungal pathogens
(Berrocal-Lobo et al., 2002; Llorente et al., 2005). To test
whether any of these defence pathways contribute to the
early termination of fungal pathogenesis in interactions with
non-adapted P. cucumerina, we determined the expression
of defence marker genes of the SA (PR1), ET/JA (PDF1.2), ET
(PR4) and JA (THI2.1) signalling pathways upon inoculation
of wild-type plants. Their expression was upregulated at 1
and 3 dpi in Col-0 plants inoculated with either non-adapted
or adapted P. cucumerina (Figure 7a). We next determined
the expression of these genes in cyp79B2 cyp79B3 plants
following pathogen challenge. The activation of PR1,
PDF1.2, PR4 and THI2.1 at 1 dpi was retained in cy-
p79B2 cyp79B3 plants (Figure 7b). Of note, PR1 and PDF1.2
upregulation was even enhanced in the double mutant,
compared with Col-0 plants, which is consistent with the
idea that the expression level of these genes correlates with
the level of fungal colonization. Thus, the activation of the
Col-0 irx1-6 pen3-10
20
40
60
80
100
Fun
gal e
ntry
rat
e (%
)
a
a
b
irx1-6pen3-1
c,d
irx1-6cyp79B2cyp79B3
d
cyp79B2cyp79B3
c
Figure 6. Impaired defence of cyp79B2 cyp79B3 and pen3 single mutants to
the non-adapted biotrophic fungus Erysiphe pisi is enhanced by the irx1-6
mutation.
Frequency of E. pisi entry at interaction sites on Arabidopsis genotypes
scored 72 h after inoculation with conidiospores. Values are averages of two
experiments (�SDs). Letters indicate statistical differences with wild-type
plants (ANOVA P £ 0.05, Bonferroni’s test).
(a)
(b)
n-fo
ld m
ock
expr
essi
on
0
10
20
30
40 PR-4
*
*
*
*
*
*
0
THI2.1
10
20
30
40
50
60
70
* **
*
* *
PcBMM Pc1187 Pc2127
n-fo
ld m
ock
expr
essi
on
0
200
400
600
800
1000
1200 PDF1.2
*
*
*
*
*
*PcBMM Pc1187 Pc2127
0
20 000
40 000
60 000
80 000PR-1
*
**
*
*
*
1 dpi3 dpi
1 dpi3 dpi
1 dpi3 dpi
1 dpi3 dpi
n-fo
ld U
BC
21 e
xpre
ssio
n
PR1
0
2
4
6
8
10
12Col-0
cyp79B2cyp79B3
*
*
*
*
PDF1.2
0
10
20
30
40
50
60
70
80
*
* * THI2.1
0.00
0.05
0.10
0.15
0.20
0.25
**
Mock BMM 1187 2127 Mock BMM 1187 2127
90PR4
0
10
20
30
40
50
60
70
80
*
**
n-fo
ld U
BC
21 e
xpre
ssio
n
Pc Pc
Figure 7. Expression of ethylene (ET), jasmonate (JA) and salicylic acid
(SA)-related defence genes is induced in Arabidopsis upon infection with
non-adapted Plectosphaerella cucumerina fungi.
(a) Wild-type Col-0 plants were treated with the indicated P. cucumerina
isolates, as described in Figure 1. Expression of the indicated defence-related
genes was determined by quantitative real-time PCR (qRT-PCR) at 1 and
3 days post-inoculation (dpi). Values are represented as n-fold increased
expression, compared with mock-treated plants. Asterisks indicate expres-
sion values that are statistically different from those of mock-treated plants,
and triangles represent values that are statistically different from PcBMM-
challenged plants (ANOVA P £ 0.05, Bonferroni’s test).
(b) Expression of the indicated defence-related genes in Col-0 (grey bars)
and cyp79B2 cyp79B3 (black bars) plants 1 dpi with P. cucumerina isolates.
Expression values are normalized to UBIQUITIN21 (UBC21). Asterisks indicate
expression values statistically different from those of wild-type plants (ANOVA
P £ 0.05, Bonferroni’s test). These experiments were repeated three times
with similar results.
122 Andrea Sanchez-Vallet et al.
ª 2010 The AuthorsJournal compilation ª 2010 Blackwell Publishing Ltd, The Plant Journal, (2010), 63, 115–127

SA, JA and ET signalling pathways occurs independently of
the pathogen-inducible accumulation of trp-derived sec-
ondary metabolites, and is not impaired in cyp79B2 cyp79B3
mutants.
We also examined whether non-adapted P. cucumerina
were able to colonize Arabidopsis mutant lines blocked in
the synthesis of either SA (NahG) or JA (aos), or in ET
signalling (ein2-1). The Pc1187 and Pc2127 isolates failed to
colonize the NahG and ein2-1 plants, and caused slight
disease symptoms in aos mutant plants (Figure S4). As
described previously (Berrocal-Lobo et al., 2002; Llorente
et al., 2005), the severity of disease symptoms caused by the
adapted PcBMM in ein2-1, NahG and aos plants was higher
than that observed in Col-0 plants (Figure S4). These data
indicate that the SA and ET signalling pathways are either
dispensable for resistance to the non-adapted P. cucumerina
isolates or act in a functionally redundant manner, whereas
the JA signalling pathway may have a minor role in
restricting the pathogenesis of these non-adapted fungi.
DISCUSSION
Most of the characterized plant necrotrophic fungi have a
broad host range and can colonize phylogenetically diverse
plant species. However, for some necrotrophs specific,
compatible interactions with particular plant hosts have
been described (van Kan, 2006). The plant genetic basis
determining host restriction of necrotrophic fungi are
poorly understood. Here, we selected three isolates of the
ascomycete necrotrophic fungus P. cucumerina (Pc1187,
Pc2127 and Pc2125), which colonize different plant species
in nature, but fail to infect Arabidopsis Col-0 wild-type
plants after spore germination. This contrasts with the
sustained hyphal growth of the adapted, virulent
P. cucumerina (PcBMM) isolate (Berrocal-Lobo et al., 2002;
Ton and Mauch-Mani, 2004). Using this pathosystem, we
have shown that trp-derived metabolites are essential for
limiting Arabidopsis colonization by non-adapted and
adapted necrotrophic fungi. Notably, a comprehensive
depletion of trp-derived metabolites, as it occurs in cy-
p79B2 cyp79B3 plants, dramatically enhances the growth of
the non-adapted P. cucumerina isolates that sporulate on
leaf tissues, rendering these plants fully susceptible (Fig-
ure 2, and data not shown). This demonstrates that the
depletion of a single class of phytochemicals is sufficient to
render Arabidopsis plants as hosts for these pathogens.
This differs from a previous study in which the concurrent
impairment of pre-invasive (pen2) and post-invasive
(pad4 sag101) non-host resistance layers was necessary to
make Arabidopsis a host for the biotrophic E. pisi and
Blumeria graminis f.sp. hordei fungi (Lipka et al., 2005).
The function of trp-derived metabolites in plant defence
responses is supported by the observed upregulation of
genes encoding enzymes involved in their biosynthesis (e.g.
ASA1, CYP79B2, CYP79B3, CYP81F2 and PAD3) following
challenge with the non-adapted or adapted P. cucumerina
isolates (Figure 3). Accordingly, the accumulation of RA and
trp-derived metabolites, such as 4MI3G, I3A, 6-OGlc-I3CA
and camalexin, was enhanced in the inoculated plants
(Figure 4). The levels of these compounds were similar in
plants challenged with the adapted or non-adapted isolates,
although hyphal growth of the host-adapted PcBMM on
leaves was unrestricted, and the number of plant cells in
contact with this fungus was higher (Figure 4). In addition to
genes related to trp-derived metabolism, the expression of
marker genes of defence signalling pathways (Glazebrook,
2005; Robert-Seilaniantz et al., 2007; Grant and Jones, 2009),
such as PR-1, PR-4, PDF1.2 and THI2.1, was also upregulat-
ed upon inoculation with the non-adapted P. cucumerina
isolates, indicating that the SA, ET and JA pathways were
positively triggered by these pathogens. With a few excep-
tions, the immune responses activated by the non-adapted
isolates overlapped with those induced by the adapted
PcBMM, as the same subset of genes was similarly
upregulated upon infection. These results are in line with
those observed in Arabidopsis plants challenged with other
non-adapted fungi, such as powdery mildews, rusts and
Alternaria alternata (Zimmerli et al., 2004; Narusaka et al.,
2005; Shafiei et al., 2007). Consistent with the activation
of a defence response, a reduction of plant growth was
observed in the inoculated Col-0 wild-type plants (Fig-
ure 1e, and data not shown), indicating that activation of
non-host resistance entails an associated fitness cost, as
described for other immune responses (Zimmerli et al.,
2004).
The biosynthesis and targeted delivery of trp derivatives
are essential for Arabidopsis immune responses to non-
adapated and adapted powdery mildews (Bednarek et al.,
2009; Consonni et al., 2010). Specifically, PEN2 and CYP81F2
activities were demonstrated to be critical for the effective
resistance against non-adapted powdery mildews, other
non-adapted fungal pathogens, like Phytophthora infestans
and Magnaporthe grisea, as well as the adapted P. cucume-
rina BMM isolate (Lipka et al., 2005; Bednarek et al., 2009;
Maeda et al., 2009). Unexpectedly, we found that resistance
responses to the non-adapted P. cucumerina isolates were
unaffected in pen2 and cyp81F2 mutants (Figure 2). Simi-
larly, camalexin, which constitutes a post-invasive barrier
against non-adapted powdery mildews (Bednarek et al.,
2009), and mediates plant resistance to necrotrophic patho-
gens, such as Alternaria brassicicola and B. cinerea (Thom-
ma et al., 1999a; Kliebenstein et al., 2005; Nafisi et al., 2007),
was not essential for non-host resistance to the non-adapted
isolates (Figure 2). Although the disruption of the biosyn-
thetic pathway of camalexin in the pen2 background
(pen2 pad3 mutant) significantly enhanced the susceptibility
of the parental lines to non-adapted and host-adapted
necrotrophs (Figure 2), P. cucumerina growth on pen2 pad3
plants was still much lower than on cyp79B2 cyp79B3 plants.
Arabidopsis non-host resistance to necrotrophic fungi 123
ª 2010 The AuthorsJournal compilation ª 2010 Blackwell Publishing Ltd, The Plant Journal, (2010), 63, 115–127

This is in marked contrast with the comparable growth of
non-adapted biotrophic powdery mildews on these two
double mutants (Bednarek et al., 2009). This strongly sug-
gests that, in addition to camalexin and the PEN2-derived
products, other trp derivatives must be required for Arabid-
opsis non-host resistance to P. cucumerina (Figure 2). One
candidate group of compounds comprises indole-3-carbox-
ylic acid derivatives, of which 6-OGlc-I3CA has been shown
here to accumulate in wild-type plants upon inoculation with
the adapted and non-adapted P. cucumerina isolates (Fig-
ure 4). Indole-3-carboxylate derivatives could exert a role in
host cell wall fortification, as some of these have been
detected as pathogen-induced conjugates of the plant cell
wall (Hagemeier et al., 2001; Tan et al., 2004; Bednarek et al.,
2005). Unfortunately, mutants specifically impaired in the
accumulation of 6-OH-I3CA have not yet been identified.
Compelling evidence has been recently obtained for indole-
derived compounds in the establishment and maintenance
of systemic acquired immunity in Arabidopsis, thereby
underlining a key role of this compound class in diverse
forms of immunity (Truman et al., 2010).
As in non-host resistance to biotrophic powdery mildew
fungi, pad2 plants supported enhanced growth of the non-
adapted P. cucumerina isolates (Figure 2). Remarkably,
the concentrations of camalexin and IGs in the mutant
increased, as in the wild type, upon inoculation with the
non-adapted necrotrophs (Figure 4). This contrasts with the
reduction in the levels of camalexin, aliphatic Gs and IGs in
pad2 plants under particular stress conditions or pathogen
infection (Glazebrook and Ausubel, 1994; Roetschi et al.,
2001; Schlaeppi et al., 2008; Bednarek et al., 2009). The
reduced levels of these compounds are believed to explain
the enhanced susceptibility phenotype of this mutant to
non-adapted biotrophs and several adapted fungi, oomycete
and bacteria. The levels of PEN2-dependent metabolites, I3A
and RA, were reduced in P. cucumerina-challenged pad2-1
plants, further supporting the potential function of glutathi-
one as a cysteine donor in the synthesis of RA (Bednarek
et al., 2009). We hypothesize that the enhanced susceptibil-
ity of pad2 plants to non-adapted P. cucumerina isolates, in
conjunction with increases in the levels of camalexin and
iGSs, but full susceptibility of cyp79B2 cyp79B3 plants,
reflects the demand for glutathione in the biosynthesis
and/or metabolism of unknown resistance-conferring trp-
derived metabolites. Alternatively, glutathione may exert a
function in immunity that is completely independent from
trp metabolism.
PEN3 activity, which is essential for resistance to the non-
adapted powdery mildews and to P. cucumerina BMM
(Stein et al., 2006; Bednarek et al., 2009), is dispensable for
resistance to the tested non-adapted necrotrophs. However,
macroscopic lesions were more severe in pen3 plants
(Figure 2b). The pen3 plants also develop chlorotic lesions
upon E. cichoracearum inoculation that leads to the activa-
tion of the SA-signalling pathway (Stein et al., 2006;
Figure 6), and are presumably the result of an enhanced
intracellular accumulation of toxic compounds. Consistent
with this, pen3 plants inoculated with P. cucumerina
showed an increased level of trp-derived metabolites
(Figure 4).
The IG-derived metabolites have been suggested to act as
signalling molecules in MAMP-triggered immunity (Clay
et al., 2009). In addition, an insect-deterrent function for
IG-derived compounds and antimicrobial activity for cama-
lexin and diverse products of G metabolism have been
described (Tierens et al., 2001; Sellam et al., 2007). Here, we
have shown that RA, I3A and camalexin display antimicrobial
activity in vitro against the P. cucumerina isolates tested
(Figure S2). The EC50 values of these metabolites for adapted
and non-adapted isolates were similar, indicating that non-
host resistance to P. cucumerina cannot be explained by a
differential sensitivity of these fungal isolates to these
pathogen-induced compounds. Camalexin was the most
active of the compounds tested, with EC50 values compara-
ble with those described for A. brassicicola and Pseudomo-
nas syringae pv. maculicola (Figure S2; Rogers et al., 1996;
Thomma et al., 1999a). The combinatorial activity of these
metabolites and antimicrobial peptides (e.g. THs and LTPs)
against P. cucumerina isolates was tested, and found to be
additive rather than synergistic (not shown). This could
indicate that trp-derived metabolites inhibit fungal growth
by a mechanism similar to that described for plant antimi-
crobial peptides (Caaveiro et al., 1997). It remains a possi-
bility that the host-adapted P. cucumerina evolved
detoxification mechanisms that are specific to the unknown,
but functionally most important, trp-derived metabolites.
Alternatively, these unknown trp-derived metabolites may
have a novel defence signalling function, rather than an
in vivo antimicrobial activity. In this scenario, the host-
adapted PcBMM fungus may have evolved the means to
block the corresponding Arabidopsis immune response
pathway(s).
The relevance of antimicrobial compounds in plant
immunity may be supported by the broad-spectrum resis-
tance of the Arabidopsis irx1-6/cesa8 secondary cell wall
mutant. Defence activation in irx1-6 plants could result in an
enhanced accumulation of trp-derived metabolites and
antimicrobial peptides. Indeed, the irx1-6 plants accumu-
lated constitutively higher levels of I3G, 1MI3G and 4MI3G
than Col-0 plants, and upon P. cucumerina BMM infection,
4MI3G levels increased further (Figure S3). The biosynthesis
and delivery of IGs in irx1-6 plants was disrupted by
generating the irx1-6 cyp79B2 cyp79B3 and irx1-6 pen3-1
mutants, respectively. Remarkably, the growth of non-
adapted and adapted isolates in these mutants was almost
comparable with wild-type plants, pointing to the existence
of additional trp-independent resistance components in this
mutant background (Figure 5). In contrast, disrupting IRX1
124 Andrea Sanchez-Vallet et al.
ª 2010 The AuthorsJournal compilation ª 2010 Blackwell Publishing Ltd, The Plant Journal, (2010), 63, 115–127

in cyp79B2 cyp79B3 and pen3 backgrounds enhanced their
susceptibility to the non-adapted powdery mildew E. pisi
(Figure 6), indicating a negative crosstalk between the
trp-independent and -dependent defences.
Immune responses of wild-type Arabidopsis upon inocu-
lation with the non-adapted P. cucumerina isolates suggest
an enhanced activation of the SA, ET and JA signalling
pathways, that, like the b-subunit of heterotrimeric G-protein
(AGB1) and the ER receptor-like kinase are essential for
growth restriction of the host-adapted PcBMM isolate
(Berrocal-Lobo et al., 2002; Llorente et al., 2005). However,
the SA and ET pathways, and AGB1 and ER, were shown to
be dispensable or to act in a functionally redundant manner
in interactions with the non-adapted P. cucumerina isolates
(Figure S4, and data not shown). Similar results for
hormone signalling have been obtained for interactions
between Arabidopsis and Leptosphaeria maculans, whereas
non-host resistance to Magnaporthe oryzae was found to be
slightly impaired in the agb1 mutant (Bohman et al., 2004;
Park et al., 2009). Besides, the JA signalling pathway seems
to contribute to non-host resistance against P. cucumerina,
as shown by the slightly enhanced susceptibility of the aos
mutant, which is impaired in JA biosynthesis (Figure S4). JA
signalling controls the accumulation of iGSs upon treatment
of Arabidopsis with JAs or a culture filtrate from Erwinia
carotovora (Brader et al., 2001; Mikkelsen et al., 2003; Jost
et al., 2005). In line with these data, we found that the
constitutive expression of CYP79B2 in aos plants was lower
than in the Col-0 wild-type (not shown), suggesting that the
enhanced susceptibility of the aos mutant to non-adapted
P. cucumerina isolates could be in part explained by its
lower content of trp derivatives. These findings suggest that,
despite trp-derived metabolites being the main executioners
and/or signalling molecules of Arabidopsis resistance to
non-adapted P. cucumerina, a complex interplay of other
defence signalling pathways becomes engaged in the
control of these necrotrophic fungi.
EXPERIMENTAL PROCEDURES
Biological material and growth conditions
Arabidopsis thaliana plants were grown in phytochambers on asterilized mixture of soil and vermiculite (3:1; Hernandez-Blancoet al., 2007), with a 10-h day/14-h night photoperiod, a temperatureof 22�C day/20�C night and 50% relative humidity. Light intensitywas fixed to 120–150 lmol m)2 s)1, according to Weigel andGlazebrook (2002). The following lines, in the Col-0 background,were used: pen2-2, pen2-3 (Lipka et al., 2005), pen3-1 (Stein et al.,2006), cyp81F2-1 (Bednarek et al., 2009), cyp79B2 cyp79B3 (Zhaoet al., 2002), pad2-1 (Parisy et al., 2007), pad3-1 (Zhou et al., 1999),pen2-1 pad3-1 (Bednarek et al., 2009), irx1-6 (Hernandez-Blancoet al., 2007), NahG (Delaney et al., 1994) and ein2-1 (Guzman andEcker, 1990). The aos mutant is in Col-6 (gl1; Park et al., 2002). TheP. cucumerina BMM isolate was provided by Dr B. Mauch-Mani(University of Fribourg, Switzerland; Tierens et al., 2001), and theP. cucumerina isolates Pc2127, Pc1187 and Pc2125 were from theDSMZ collection (http://www.dsmz.de). In vitro fungal growth and
the collection of spores were carried out as reported by Hernandez-Blanco et al. (2007).
Fungal inoculation assays
Three-week-old Arabidopsis plants were inoculated with a sporesuspension (4 · 106 spores ml)1) of the different P. cucumerinaisolates. Disease progression was estimated by determining theaverage DR (0–5), trypan blue staining and relative quantification offungal DNA by qRT-PCR, as described by Sanchez-Rodriguez et al.(2009). Inoculation with E. pisi (Birmingham isolate) and determi-nation of entry rates was performed as described by Bednarek et al.(2009). For all of the fungal inoculations, at least three independentexperiments were performed. Statistical differences amongArabidopsis genotypes were determined by one-way analysis ofvariance and Bonferroni post hoc test, as previously reported(Sanchez-Rodriguez et al., 2009).
Gene expression analyses and fungal biomass
determination
For gene expression analysis, RNA extractions from P. cucumerina-infected or mock-treated plants were performed; for fungal DNAquantification, DNA from infected plants was extracted (Llorenteet al., 2005). Oligonucleotides were designed using PRIMER
EXPRESS v2.0 (Applied Biosystems, http://www.appliedbiosys-tems.com; Table S1). qRT-PCR analyses were performed as previ-ously reported, with 0.3 lM of each primer (Hernandez-Blanco et al.,2007), using the FS Universal SYBR GreenMasterRox (Roche, http://www.roche.com) and the described amplification conditions (San-chez-Rodriguez et al., 2009). The plant UBIQUITIN21 (At5G25760)gene was used to normalize and calculate the change in cyclethreshold (DCt) value. The relative expression ratio was determinedwith the equation 2)DDC
t (Rieu and Powers, 2009), using the relativequantification application of the sequence detector software (v1.4;Applied Biosystems; Rieu and Powers, 2009). The qRT-PCR resultsare mean values (�SDs) from two technical replicates. Differencesin expression ratios (DCt) among the samples were analysed byANOVA or Student’s t-test using STATGRAPHICS.
Quantification of trp derivatives
Plant samples were collected at 24 hpi and frozen in liquid nitrogen.Extraction and HPLC analysis of tryptophan derivatives was per-formed as previously described (Bednarek et al., 2009). Each linewas tested in at least three independent experiments.
In vitro antimicrobial tests
Inhibition tests of P. cucumerina spore were carried out in sterilemicrotiter plates, as described by Molina et al. (1993b). Differentconcentrations of purified secondary metabolites (Bednarek et al.,2009) and peptides (Molina et al., 1993a,b) were analysed. Plateswere incubated at 28�C, and pictures of hyphal growth were taken atdifferent time points.
Generation and selection if irx1-6 cyp79B2 cyp79B3 and
irx1-6 pen3-1 mutants
These mutants were generated by standard genetic crossesfollowed by identification of the mutant alleles. Genotyping of theirx1-6 and pen3-1 mutations were confirmed by sequencing themutations in the PCR products (irx1-6, 5¢-CATGTGCTGTTGGGT-AGGAATC-3¢ and 5¢-CATAGAGAATGTGTTTGATGATG-3¢; pen3-1,5¢-CTCGTCACTGATTATACTCTC-3¢ and 5¢-TGAGGTGAACGATTTG-TTGC-3¢). The T-DNA insertion mutations in cyp79B2 cyp79B3 wereidentified as described (Zhao et al., 2002).
Arabidopsis non-host resistance to necrotrophic fungi 125
ª 2010 The AuthorsJournal compilation ª 2010 Blackwell Publishing Ltd, The Plant Journal, (2010), 63, 115–127

Plectosphaerella cucumerina ITS1 and ITS2 GenBank
numbers
PcBMM, GU724979; Pc1187, GU724980; Pc2125, GU724981; Pc2127,GU724982.
ACKNOWLEDGEMENTS
Work in the laboratory of AM was supported by the Spanish Min-isterio de Ciencia e Innovacion (MICINN) [grants EUI2008-03728(BALANCE) and BIO2006-00488]. AS-V was a PhD fellow from theMinisterio de Educacion y Ciencia (MEC). BR was an Ayudante fromUniversidad Salamanca. Work in the laboratory of PS-L was sup-ported by the German Bundesministerium fur Bildung und For-schung (BMBF) (Plant-KBBE grant: BALANCE) and the Max PlanckSociety.
SUPPORTING INFORMATION
Additional Supporting Information may be found in the onlineversion of this article:Figure S1. Neighbour-joining tree of ribosomal DNA sequences(ITS1-5.8rRNA-ITS2) from Plectosphaerella cucumerina (Pc) iso-lates.Figure S2. Tryptophan-derived metabolites inhibit in vitro Plectosp-haerella cucumerina growth.Figure S3. Determination of tryptophan-derived metabolites con-tent in irx1-6 and wild-type plants upon infection by Plectosphae-rella cucumerina.Figure S4. Contribution of the ethylene (ET), jasmonate (JA) andsalicylic acid (SA) signalling pathways to Arabidopsis non-hostresistance against non-adapted Plectosphaerella cucumerina fungi.Table S1. Oligonucleotides used for quantitative real-time PCR(qRT-PCR) analyses.Please note: As a service to our authors and readers, this journalprovides supporting information supplied by the authors. Suchmaterials are peer-reviewed and may be re-organized for onlinedelivery, but are not copy-edited or typeset. Technical supportissues arising from supporting information (other than missingfiles) should be addressed to the authors.
REFERENCES
Bednarek, P., Schneider, B., Svatos, A., Oldham, N.J. and Hahlbrock, K. (2005)
Structural complexity, differential response to infection, and tissue speci-
ficity of indolic and phenylpropanoid secondary metabolism in Arabidop-
sis roots. Plant Physiol. 138, 1058–1070.
Bednarek, P., Pislewska-Bednarek, M., Svatos, A. et al. (2009) A glucosinolate
metabolism pathway in living plant cells mediates broad-spectrum anti-
fungal defense. Science, 323, 101–106.
Berrocal-Lobo, M., Molina, A. and Solano, R. (2002) Constitutive expression of
ETHYLENE-RESPONSE-FACTOR1 in Arabidopsis confers resistance to
several necrotrophic fungi. Plant J. 29, 23–32.
Bohman, S., Staal, J., Thomma, B.P.H.J., Wang, M. and Dixelius, C. (2004)
Characterisation of an Arabidopsis-Leptosphaeria maculans pathosys-
tem: resistance partially requires camalexin biosynthesis and is inde-
pendent of salicylic acid, ethylene and jasmonic acid signalling. Plant J.
37, 9–20.
Bolton, M.D., Thomma, B.P.H.J. and Nelson, B.D. (2006) Sclerotinia sclero-
tiorum (Lib.) de Bary: biology and molecular traits of a cosmopolitan
pathogen. Mol. Plant Pathol. 7, 1–16.
Bottcher, C., Westphal, L., Schmotz, C., Prade, E., Scheel, D. and Glawischnig,
E. (2009) The multifunctional enzyme CYP71B15 (PHYTOALEXIN DEFI-
CIENT3) converts cysteine-indole-3-acetonitrile to camalexin in the indole-
3-acetonitrile metabolic network of Arabidopsis thaliana. Plant Cell, 21,
1830–1841.
Brader, G., Tas, E. and Palva, E.T. (2001) Jasmonate-dependent induction of
indole glucosinolates in Arabidopsis by culture filtrates of the nonspecific
pathogen Erwinia carotovora. Plant Physiol. 126, 849–860.
Caaveiro, J.M., Molina, A., Gonzalez-Manas, J.M., Rodriguez-Palenzuela, P.,
Garcia-Olmedo, F. and Goni, F.M. (1997) Differential effects of five types of
antipathogenic plant peptides on model membranes. FEBS Lett. 410, 338–
342.
Clay, N.K., Adio, A.M., Denoux, C., Jander, G. and Ausubel, F.M. (2009)
Glucosinolate metabolites required for an Arabidopsis innate immune
response. Science, 323, 95–101.
Collins, N.C., Thordal-Christensen, H., Lipka, V. et al. (2003) SNARE-protein-
mediated disease resistance at the plant cell wall. Nature, 425, 973–977.
Consonni, C., Bednarek, P., Humphry, M., Francocci, F., Ferrari, S., Harzen, A.,
Ver Loren van Themaat, E. and Panstruga, R. (2010) Tryptophan-derived
metabolites are required for antifungal defence in the Arabidopsis thaliana
mlo2 mutant. Plant Physiol., 152, 1544–1561.
Cramer, R.A. and Lawrence, C.B. (2004) Identification of Alternaria brassici-
cola genes expressed in planta during pathogenesis of Arabidopsis thali-
ana. Fungal Genet. Biol. 41, 115–128.
Delaney, T.P., Uknes, S., Vernooij, B. et al. (1994) A central role of salicylic
acid in plant disease resistance. Science, 266, 1247–1250.
Denby, K.J., Kumar, P. and Kliebenstein, D.J. (2004) Identification of Botrytis
cinerea susceptibility loci in Arabidopsis thaliana. Plant J. 38, 473–486.
Domsch, K.H., Gams, W. and Anderson, T.H. (2007) Compendium of Soil
Fungi, 2nd edn. Eching: IHW Verlag.
Ferrari, S., Plotnikova, J.M., Lorenzo, G.D. and Ausubel, F.M. (2003) Arabid-
opsis local resistance to Botrytis cinerea involves salicylic acid and cama-
lexin and requires EDS4 and PAD2, but not SID2, EDS5 or PAD4. Plant J. 35,
193–205.
Glawischnig, E. (2007) Camalexin. Phytochemistry, 68, 401–406.
Glazebrook, J. (2005) Contrasting mechanisms of defense against biotrophic
and necrotrophic pathogens. Annu. Rev. Phytopathol. 43, 205–227.
Glazebrook, J. and Ausubel, F.M. (1994) Isolation of phytoalexin-deficient
mutants of Arabidopsis thaliana and characterization of their interactions
with bacterial pathogens. Proc. Natl. Acad. Sci. USA, 91, 8955–8959.
Grant, M.R. and Jones, J.D. (2009) Hormone (dis)harmony moulds plant
health and disease. Science, 324, 750–752.
Guzman, P. and Ecker, J.R. (1990) Exploiting the triple response of Arabid-
opsis to identify ethylene-related mutants. Plant Cell, 2, 513–523.
Hagemeier, J., Schneider, B., Oldham, N.J. and Hahlbrock, K. (2001) Accu-
mulation of soluble and wall-bound indolic metabolites in Arabidopsis
thaliana leaves infected with virulent or avirulent Pseudomonas syringae
pathovar tomato strains. Proc. Natl. Acad. Sci. USA, 98, 753–758.
Heath, M.C. (2000) Nonhost resistance and nonspecific plant defenses. Curr.
Opin. Plant Biol. 3, 315–319.
Hernandez-Blanco, C., Feng, D.X., Hu, J. et al. (2007) Impairment of cellulose
synthases required for Arabidopsis secondary cell wall formation enhances
disease resistance. Plant Cell, 19, 890–903.
Jost, R., Altschmied, L., Bloem, E. et al. (2005) Expression profiling of meta-
bolic genes in response to methyl jasmonate reveals regulation of genes of
primary and secondary sulfur-related pathways in Arabidopsis thaliana.
Photosynth. Res. 86, 491–508.
van Kan, J.A.L. (2006) Licensed to kill: the lifestyle of a necrotrophic plant
pathogen. Trends Plant Sci., 11, 247–253.
Kliebenstein, D.J., Rowe, H.C. and Denby, K.J. (2005) Secondary metabolites
influence Arabidopsis/Botrytis interactions: variation in host production
and pathogen sensitivity. Plant J. 44, 25–36.
Kwon, C., Neu, C., Pajonk, S. et al. (2008) Co-option of a default secretory
pathway for plant immune responses. Nature, 451, 835–840.
Lipka, V., Dittgen, J., Bednarek, P. et al. (2005) Pre- and postinvasion defenses
both contribute to nonhost resistance in Arabidopsis. Science, 310, 1180–
1183.
Llorente, F., Alonso-Blanco, C., Sanchez-Rodriguez, C., Jorda, L. and Molina,
A. (2005) ERECTA receptor-like kinase and heterotrimeric G protein from
Arabidopsis are required for resistance to the necrotrophic fungus Plec-
tosphaerella cucumerina. Plant J. 43, 165–180.
Llorente, F., Muskett, P., Sanchez-Vallet, A., Lopez, G., Ramos, B., Sanchez-
Rodriguez, C., Jorda, L., Parker, J. and Molina, A. (2008) Repression of the
auxin response pathway increases Arabidopsis susceptibility to necro-
trophic fungi. Mol. Plant, 1, 496.
Maeda, K., Houjyou, Y., Komatsu, T., Hori, H., Kodaira, T. and Ishikawa, A.
(2009) AGB1 and PMR5 contribute to PEN2-mediated preinvasion resis-
tance to Magnaporthe oryzae in Arabidopsis thaliana. Mol. Plant-Microbe
Interact. 22, 1331–1340.
126 Andrea Sanchez-Vallet et al.
ª 2010 The AuthorsJournal compilation ª 2010 Blackwell Publishing Ltd, The Plant Journal, (2010), 63, 115–127

Mikkelsen, M.D., Petersen, B.L., Glawischnig, E., Jensen, A.B., Andreasson, E.
and Halkier, B.A. (2003) Modulation of CYP79 genes and glucosinolate
profiles in Arabidopsis by defense signaling pathways. Plant Physiol. 131,
298–308.
Molina, A., Ahl Goy, P., Fraile, A., Sanchez-Monge, R. and Garcia-Olmedo, F.
(1993a) Inhibition of bacterial and fungal plant pathogens by thionins of
types I and II. Plant Sci. 92, 166–177.
Molina, A., Segura, A. and Garcia-Olmedo, F. (1993b) Lipid transfer proteins
(nsLTPs) from barley and maize leaves are potent inhibitors of bacterial and
fungal plant pathogens. FEBS Lett. 316, 119–122.
Nafisi, M., Goregaoker, S., Botanga, C.J., Glawischnig, E., Olsen, C.E., Halkier,
B.A. and Glazebrook, J. (2007) Arabidopsis cytochrome P450 monooxy-
genase 71A13 catalyzes the conversion of indole-3-acetaldoxime in cama-
lexin synthesis. Plant Cell, 19, 2039–2052.
Narusaka, Y., Narusaka, M., Seki, M., Ishida, J., Shinozaki, K., Nan, Y., Park, P.,
Shiraishi, T. and Kobayashi, M. (2005) Cytological and molecular analyses
of non-host resistance of Arabidopsis thaliana to Alternaria alternata. Mol.
Plant Pathol. 6, 615–627.
Niyogi, K.K. and Fink, G.R. (1992) Two anthranilate synthase genes in Ara-
bidopsis: defense-related regulation of the tryptophan pathway. Plant Cell,
4, 721–733.
Parisy, V., Poinssot, B., Owsianowski, L., Buchala, A., Glazebrook, J. and
Mauch, F. (2007) Identification of PAD2 as a gamma-glutamylcysteine
synthetase highlights the importance of glutathione in disease resistance
of Arabidopsis. Plant J. 49, 159–172.
Park, J.-H., Halitschke, R., Kim, H.B., Baldwin, I.T., Feldmann, K.A. and
Feyereisen, R. (2002) A knock-out mutation in allene oxide synthase results
in male sterility and defective wound signal transduction in Arabidopsis
due to a block in jasmonic acid biosynthesis. Plant J. 31, 1–12.
Park, J.-Y., Jin, J., Lee, Y.-W., Kang, S. and Lee, Y.-H. (2009) Rice blast fungus
(Magnaporthe oryzae) infects Arabidopsis via a mechanism distinct from
that required for the infection of rice. Plant Physiol. 149, 474–486.
Pfalz, M., Vogel, H. and Kroymann, J. (2009) The gene controlling the indole
glucosinolate modifier1 quantitative trait locus alters indole glucosinolate
structures and aphid resistance in Arabidopsis. Plant Cell, 21, 985–999.
Pitt, W.M., Goodwin, S.B., Ash, G.J., Cother, N.J. and Cother, E.J. (2004)
Plectosporium alismatis comb. nov. a new placement for the Alismataceae
pathogen Rhynchosporium alismatis. Mycol. Res., 108, 775–780.
Rieu, I. and Powers, S.J. (2009) Real-time quantitative RT-PCR: design, cal-
culations, and statistics. Plant Cell, 21, 1031–1033.
Robert-Seilaniantz, A., Navarro, L., Bari, R. and Jones, J.D.G. (2007) Patho-
logical hormone imbalances. Curr. Opin. Plant Biol. 10, 372–379.
Roetschi, A., Si-Ammour, A., Belbahri, L., Mauch, F. and Mauch-Mani, B.
(2001) Characterization of an Arabidopsis-Phytophthora pathosystem:
resistance requires a functional PAD2 gene and is independent of salicylic
acid, ethylene and jasmonic acid signalling. Plant J. 28, 293–305.
Rogers, E.E., Glazebrook, J. and Ausubel, F.M. (1996) Mode of action of the
Arabidopsis thaliana phytoalexin camalexin and its role in Arabidopsis-
pathogen interactions. Mol. Plant Microbe Interact. 9, 748–757.
Sanchez-Rodriguez, C., Estevez, J.M., Llorente, F., Hernandez-Blanco, C.,
Jorda, L., Pagan, I., Berrocal, M., Marco, Y., Somerville, S. and Molina, A.
(2009) The ERECTA receptor-like kinase regulates cell wall mediated
resistance to pathogens in Arabidopsis thaliana. Mol. Plant-Microbe
Interact. 22, 953–963.
Schlaeppi, K., Bodenhausen, N., Buchala, A., Mauch, F. and Reymond, P.
(2008) The glutathione-deficient mutant pad2-1 accumulates lower
amounts of glucosinolates and is more susceptible to the insect herbivore
Spodoptera littoralis. Plant J. 55, 774–786.
Schuhegger, R., Nafisi, M., Mansourova, M., Petersen, B.L., Olsen, C.E.,
Svatos, A., Halkier, B.A. and Glawischnig, E. (2006) CYP71B15 (PAD3) cat-
alyzes the final step in camalexin biosynthesis. Plant Physiol. 141, 1248–
1254.
Sellam, A., Iacomi-Vasilescu, B., Hudhomme, P. and Simoneau, P. (2007)
In vitro antifungal activity of brassinin, camalexin and two isothiocyanates
against the crucifer pathogens Alternaria brassicicola and Alternaria
brassicae. Plant Pathol. 56, 296–301.
Shafiei, R., Hang, C., Kang, J.-G. and Loake, G.J. (2007) Identification of loci
controlling non-host disease resistance in Arabidopsis against the leaf rust
pathogen Puccinia triticina. Mol. Plant Pathol. 8, 773–784.
Somerville, C., Bauer, S., Brininstool, G. et al. (2004) Toward a systems ap-
proach to understanding plant cell walls. Science, 306, 2206–2211.
Stein, M., Dittgen, J., Sanchez-Rodriguez, C., Hou, B.H., Molina, A.,
Schulze-Lefert, P., Lipka, V. and Somerville, S. (2006) Arabidopsis PEN3/
PDR8, an ATP binding cassette transporter, contributes to nonhost resis-
tance to inappropriate pathogens that enter by direct penetration. Plant
Cell, 18, 731–746.
Tan, J., Bednarek, P., Liu, J., Schneider, B., Svatos, A. and Hahlbrock, K. (2004)
Universally occurring phenylpropanoid and species-specific indolic
metabolites in infected and uninfected Arabidopsis thaliana roots and
leaves. Phytochemistry, 65, 691–699.
Thomma, B.P., Eggermont, K., Penninckx, I.A., Mauch-Mani, B., Vogelsang,
R., Cammue, B.P. and Broekaert, W.F. (1998) Separate jasmonate-depen-
dent and salicylate-dependent defense-response pathways in Arabidopsis
are essential for resistance to distinct microbial pathogens. Proc. Natl.
Acad. Sci. USA, 95, 15107–15111.
Thomma, B.P., Nelissen, I., Eggermont, K. and Broekaert, W.F. (1999a)
Deficiency in phytoalexin production causes enhanced susceptibility of
Arabidopsis thaliana to the fungus Alternaria brassicicola. Plant J. 19,
163–171.
Thomma, B.P.H.J., Eggermont, K., Tierens, K.F.M.J. and Broekaert, W.F.
(1999b) Requirement of functional ethylene-insensitive 2 gene for efficient
resistance of Arabidopsis to infection by Botrytis cinerea. Plant Physiol.
121, 1093–1101.
Thordal-Christensen, H. (2003) Fresh insights into processes of nonhost
resistance. Curr. Opin. Plant Biol. 6, 351–357.
Tierens, K.F., Thomma, B.P., Brouwer, M., Schmidt, J., Kistner, K., Porzel, A.,
Mauch-Mani, B., Cammue, B.P. and Broekaert, W.F. (2001) Study of the role
of antimicrobial glucosinolate-derived isothiocyanates in resistance of
Arabidopsis to microbial pathogens. Plant Physiol. 125, 1688–1699.
Ton, J. and Mauch-Mani, B. (2004) b-amino-butyric acid-induced resistance
against necrotrophic pathogens is based on ABA-dependent priming for
callose. Plant J. 38, 119–130.
Truman, W.M., Bennett, M.H., Turnbull, C.G.N. and Grant, M.R. (2010) Ara-
bidopsis auxin mutants are compromised in sytemic acquired resistance
and exhibit aberrant accumulation of various indolic compounds. Plant
Physiol. 152, 1562–1573.
Van Baarlen, P., Woltering, E.J., Staats, M. and Van Kan, J.A. (2007) Histo-
chemical and genetic analysis of host and non-host interactions of Ara-
bidopsis with three Botrytis species: an important role for cell death
control. Mol. Plant Pathol. 8, 41.
van Wees, S.C.M., Chang, H.-S., Zhu, T. and Glazebrook, J. (2003) Charac-
terization of the early response of Arabidopsis to Alternaria brassicicola
infection using expression profiling. Plant Physiol. 132, 606–617.
Weigel, D. and Glazebrook, J. (2002) Arabidopsis, a Laboratory Manual. New
York: Cold Spring Harbor.
Williamson, B., Tudzynski, B., Tudzynski, P. and Kan, J.A.L.V. (2007)
Botrytis cinerea: the cause of grey mould disease. Mol. Plant Pathol. 8,
561–580.
Zhao, Y., Hull, A.K., Gupta, N.R., Goss, K.A., Alonso, J., Ecker, J.R., Normanly,
J., Chory, J. and Celenza, J.L. (2002) Trp-dependent auxin biosynthesis in
Arabidopsis: involvement of cytochrome P450s CYP79B2 and CYP79B3.
Genes Dev. 16, 3100–3112.
Zhou, N., Tootle, T.L. and Glazebrook, J. (1999) Arabidopsis PAD3, a gene
required for camalexin biosynthesis, encodes a putative cytochrome P450
monooxygenase. Plant Cell, 11, 2419–2428.
Zimmerli, L., Stein, M., Lipka, V., Schulze-Lefert, P. and Somerville, S. (2004)
Host and non-host pathogens elicit different jasmonate/ethylene responses
in Arabidopsis. Plant J. 40, 633–646.
Arabidopsis non-host resistance to necrotrophic fungi 127
ª 2010 The AuthorsJournal compilation ª 2010 Blackwell Publishing Ltd, The Plant Journal, (2010), 63, 115–127

![FIBRILLIN4 Is Required for Plastoglobule Development and Stress Resistance in Apple and Arabidopsis 1[W][OA](https://static.fdokumen.com/doc/165x107/634098f928d04168ce02a616/fibrillin4-is-required-for-plastoglobule-development-and-stress-resistance-in-apple.jpg)


















