
Noradrenergic lesion of the locus coeruleus increases apomorphine-induced circling behavior and the...
11
Research Report Noradrenergic lesion of the locus coeruleus increases the firing activity of the medial prefrontal cortex pyramidal neurons and the role of α 2 -adrenoceptors in normal and medial forebrain bundle lesioned rats Yong Wang a , Qiao Jun Zhang b , Jian Liu a, ⁎ , Umar Ali a , Zhen Hua Gui a , Yan Ping Hui b , Tao Wang a , Li Chen a , Qiang Li a a Department of Physiology and Pathophysiology, School of Medicine, Xi'an Jiaotong University, Xi'an 710061, China b Department of Neurology, the Second Affiliated Hospital, Medical School of Xi'an Jiaotong University, Xi'an 710004, China ARTICLE INFO ABSTRACT Article history: Accepted 3 February 2010 Available online 11 February 2010 Degeneration of noradrenergic neurons in the locus coeruleus (LC) and dysfunction of the prefrontal cortex were regarded as playing a specific role in the occurrence of non-motor symptoms in Parkinson's disease. The present study examined the spontaneous firing rate and firing pattern of medial prefrontal cortex (mPFC) pyramidal neurons, and effects of α 2 -adrenoceptor agonist UK-14,304 and antagonist yohimbine on the neuronal activity in rats with 6-hydroxydopamine lesions of the LC, medial forebrain bundle (MFB) and with combined MFB and LC lesions. The firing rate of mPFC pyramidal neurons in rats with lesions of the LC and with combine LC and MFB lesions is significantly higher than that of normal and MFB-lesioned rats and the firing pattern of these neurons in rats with lesions of the LC and with combine LC and MFB lesions also changed significantly towards more regular compared with normal and MFB-lesioned rats. The local administration of UK-14,304 in the mPFC inhibited the firing activity of the pyramidal neurons in normal rats and rats with lesions of the LC, MFB and with combined LC and MFB lesions, while yohimbine increased the firing activity of the pyramidal neurons. These results indicate that the lesions of the LC lead to hyperactivity of mPFC pyramidal neurons in normal and MFB-lesioned rats, and the postsynaptic α 2 -adrenoceptors may partially mediate the inhibitory effects of LC-noradrenergic system on the firing activity of pyramidal neurons in the mPFC, suggesting that LC-noradrenergic system plays an important role in the functional disorders of mPFC in Parkinson's disease. © 2010 Elsevier B.V. All rights reserved. Keywords: Medial prefrontal cortex Locus coeruleus Parkinson's disease α 2 -adrenoceptor Extracellular recording 1. Introduction Parkinson's disease (PD) is one of the most common neurodegenerative diseases (Graeber et al., 1998). Although PD is characterized by the cardinal symptoms including resting tremor, bradykinesia, rigidity and postural instability, the non-motor features including emotional and cognitive deficits, are also widespread clinical manifestations (Remy BRAIN RESEARCH 1324 (2010) 64 – 74 ⁎ Corresponding author. Department of Physiology and Pathophysiology, School of Medicine, Xi'an Jiaotong University, Yan Ta Xi Lu 76, Xi'an 710061, China. Fax: +86 29 82656364. E-mail address: [email protected] (J. Liu). 0006-8993/$ – see front matter © 2010 Elsevier B.V. All rights reserved. doi:10.1016/j.brainres.2010.02.009 available at www.sciencedirect.com www.elsevier.com/locate/brainres
-
Upload
independent -
Category
Documents
-
view
6 -
download
0
Transcript of Noradrenergic lesion of the locus coeruleus increases apomorphine-induced circling behavior and the...

B R A I N R E S E A R C H 1 3 2 4 ( 2 0 1 0 ) 6 4 – 7 4
ava i l ab l e a t www.sc i enced i r ec t . com
www.e l sev i e r . com/ loca te /b ra i n res
Research Report
Noradrenergic lesion of the locus coeruleus increases the firingactivity of the medial prefrontal cortex pyramidal neurons andthe role of α2-adrenoceptors in normal and medial forebrainbundle lesioned rats
Yong Wanga, Qiao Jun Zhangb, Jian Liua,⁎, Umar Alia, Zhen Hua Guia, Yan Ping Huib,Tao Wanga, Li Chena, Qiang Lia
aDepartment of Physiology and Pathophysiology, School of Medicine, Xi'an Jiaotong University, Xi'an 710061, ChinabDepartment of Neurology, the Second Affiliated Hospital, Medical School of Xi'an Jiaotong University, Xi'an 710004, China
A R T I C L E I N F O
⁎ Corresponding author. Department of PhysiXi'an 710061, China. Fax: +86 29 82656364.
E-mail address: [email protected] (
0006-8993/$ – see front matter © 2010 Elsevidoi:10.1016/j.brainres.2010.02.009
A B S T R A C T
Article history:Accepted 3 February 2010Available online 11 February 2010
Degeneration of noradrenergic neurons in the locus coeruleus (LC) and dysfunction of theprefrontal cortex were regarded as playing a specific role in the occurrence of non-motorsymptoms in Parkinson's disease. The present study examined the spontaneous firingrate and firing pattern of medial prefrontal cortex (mPFC) pyramidal neurons, and effects ofα2-adrenoceptor agonist UK-14,304 and antagonist yohimbine on the neuronal activity inrats with 6-hydroxydopamine lesions of the LC, medial forebrain bundle (MFB) and withcombined MFB and LC lesions. The firing rate of mPFC pyramidal neurons in rats withlesions of the LC and with combine LC and MFB lesions is significantly higher than that ofnormal and MFB-lesioned rats and the firing pattern of these neurons in rats with lesions ofthe LC and with combine LC and MFB lesions also changed significantly towards moreregular comparedwith normal andMFB-lesioned rats. The local administration of UK-14,304in the mPFC inhibited the firing activity of the pyramidal neurons in normal rats and rats withlesions of the LC, MFB and with combined LC and MFB lesions, while yohimbine increased thefiring activity of the pyramidal neurons. These results indicate that the lesions of the LC lead tohyperactivityofmPFCpyramidalneurons innormalandMFB-lesioned rats, and thepostsynapticα2-adrenoceptorsmaypartiallymediate the inhibitory effects of LC-noradrenergic systemon thefiring activity of pyramidal neurons in themPFC, suggesting that LC-noradrenergic systemplaysan important role in the functional disorders of mPFC in Parkinson's disease.
© 2010 Elsevier B.V. All rights reserved.
Keywords:Medial prefrontal cortexLocus coeruleusParkinson's diseaseα2-adrenoceptorExtracellular recording
1. Introduction
Parkinson's disease (PD) is one of the most commonneurodegenerative diseases (Graeber et al., 1998). Although
ology and Pathophysiolog
J. Liu).
er B.V. All rights reserved
PD is characterized by the cardinal symptoms includingresting tremor, bradykinesia, rigidity and postural instability,the non-motor features including emotional and cognitivedeficits, are also widespread clinical manifestations (Remy
y, School of Medicine, Xi'an Jiaotong University, Yan Ta Xi Lu 76,
.
65B R A I N R E S E A R C H 1 3 2 4 ( 2 0 1 0 ) 6 4 – 7 4
et al., 2005; Schapira, 2008). However, the pathophysiologicalmechanism of these non-motor symptoms remains largelyunknown. Pathological studies revealed that apart from thedegeneration of nigrostriatal dopamine (DA) neurons, aprofound loss of noradrenaline (NA) neurons in the locuscoeruleus (LC) was demonstrated in PD patients (Zarow et al.,2003). Neuroimaging and postmortem studies have demon-strated that the presence of depression and dementia in PDwas associated with the significant loss of noradrenergicneurons in the LC and NA depletion in the cortex (Zweig et al.,1993; Bosboom et al., 2004; Remy et al., 2005). These studiesindicate that the LC-noradrenergic neurotransmitter systemmay have an important role in the pathogenesis of non-motorsymptoms in PD.
The prefrontal cortex (PFC) is involved in a large number ofhigher brain functions and controls the activity of subcorticalstructures (Fuster, 1997; Miller and Cohen, 2001). Dysfunctionwithin the PFC and its connected regions has been linked withthe pathophysiology of emotional and cognitive impairment,including non-motor symptoms in PD (Cummings, 1992;Ridderinkhof et al., 2004; Ramos and Arnsten, 2007). The PFCreceives dense noradrenergic innervations mainly from theipsilateral LC and expresses several types of adrenoceptors,including α1, α2, β1 and β2 subtypes, while the pre- andpostsynaptic α2-adrenoceptors are regarded as playing acrucial role in the activity of the PFC (Waterhouse et al.,1983; Arnsten and Goldman-Rakic, 1985; Ramos and Arnsten,2007). A diminution of NA levels in the PFC was observed in PDpatients (Scatton et al., 1983), and the receptor binding studyshowed an increase in α1- and β1-adrenergic receptors, whilethe putative presynaptic α2-adrenergic receptors decreased inthe PFC of PD patients (Cash et al., 1984). Importantly, clinicalstudy reported that loss of noradrenergic innervation in thefrontal cortex is associated with non-motor symptoms in PD
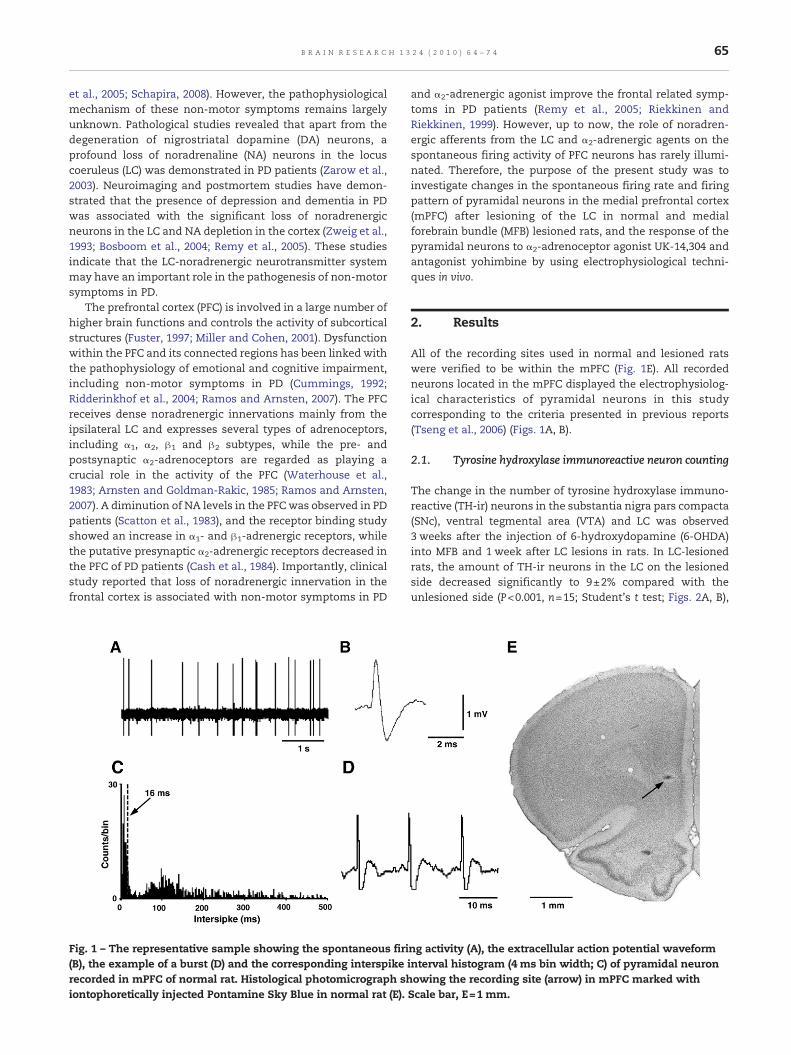
Fig. 1 – The representative sample showing the spontaneous firi(B), the example of a burst (D) and the corresponding interspikerecorded in mPFC of normal rat. Histological photomicrograph shiontophoretically injected Pontamine Sky Blue in normal rat (E).
and α2-adrenergic agonist improve the frontal related symp-toms in PD patients (Remy et al., 2005; Riekkinen andRiekkinen, 1999). However, up to now, the role of noradren-ergic afferents from the LC and α2-adrenergic agents on thespontaneous firing activity of PFC neurons has rarely illumi-nated. Therefore, the purpose of the present study was toinvestigate changes in the spontaneous firing rate and firingpattern of pyramidal neurons in the medial prefrontal cortex(mPFC) after lesioning of the LC in normal and medialforebrain bundle (MFB) lesioned rats, and the response of thepyramidal neurons to α2-adrenoceptor agonist UK-14,304 andantagonist yohimbine by using electrophysiological techni-ques in vivo.
2. Results
All of the recording sites used in normal and lesioned ratswere verified to be within the mPFC (Fig. 1E). All recordedneurons located in the mPFC displayed the electrophysiolog-ical characteristics of pyramidal neurons in this studycorresponding to the criteria presented in previous reports(Tseng et al., 2006) (Figs. 1A, B).
2.1. Tyrosine hydroxylase immunoreactive neuron counting
The change in the number of tyrosine hydroxylase immuno-reactive (TH-ir) neurons in the substantia nigra pars compacta(SNc), ventral tegmental area (VTA) and LC was observed3 weeks after the injection of 6-hydroxydopamine (6-OHDA)into MFB and 1 week after LC lesions in rats. In LC-lesionedrats, the amount of TH-ir neurons in the LC on the lesionedside decreased significantly to 9±2% compared with theunlesioned side (P<0.001, n=15; Student's t test; Figs. 2A, B),
ng activity (A), the extracellular action potential waveforminterval histogram (4 ms bin width; C) of pyramidal neuronowing the recording site (arrow) in mPFC marked withScale bar, E=1 mm.

Fig. 2 – Photomicrographs of TH immunocytochemicalstaining of the LC, SNc and VTA in rats with lesions of the LC(A, B), MFB (C, D) andwith combined LC andMFB lesions (E–H)showing the LC, SNc and VTA neurons on the injected side(left) compared with intact side (right). The SNc in rats withlesions of the MFB (C) andwith combined LC andMFB lesions(E) showing a total loss of TH-ir neurons, and the LC and VTAin rats with lesions of the LC (A), MFB (C) and with combinedLC and MFB lesions (E, G) showing a subtotal loss of TH-irneurons. MTN, medial terminal nucleus; 4V, the 4thventricle. Scale bar, A–H=1 mm.
Fig. 3 – Histograms showing the distribution of firing rates ofmPFC pyramidal neurons recorded in normal rats (n=75) andrats with lesions of the LC (n=74), MFB (n=72) and withcombined LC and MFB lesions (n=51; A), and comparisons ofmean firing rate of the neurons (B). *P<0.05, **P<0.01.
66 B R A I N R E S E A R C H 1 3 2 4 ( 2 0 1 0 ) 6 4 – 7 4
the TH-ir neuron counts in the SNc and the VTA on theinjected side were unaltered compared with the unlesionedside. In MFB-lesioned rats, the SNc on the lesioned sideshowed a total loss of TH-ir neurons compared with theunlesioned side, the amount of TH-ir neurons in the VTA onthe lesioned side decreased significantly to 12±3% comparedwith the unlesioned side (P<0.001, n=12; Student's t test;Figs. 2C, D), and the number of TH-ir neurons in the LC on thelesioned side decreased slightly to 97±3% compared with theunlesioned side (n=12). In rats with combined MFB and LClesions, the SNc on the lesioned side showed a total loss of TH-ir neurons compared with the unlesioned side, the amount ofTH-ir neurons in the VTA on the lesioned side decreasedsignificantly to 11±3% compared with the unlesioned side
(P<0.001, n=10; Student's t test; Figs. 2E, F), and the number ofTH-ir neurons in the LC on the lesioned side decreasedsignificantly to 9±3% compared with the unlesioned side(P<0.001, n=10; Student's t test; Figs. 2G, H).
2.2. Spontaneous firing activity of mPFC pyramidalneurons
In normal rats, firing rates of mPFC pyramidal neurons rangedfrom 0.47 to 6.40 spikes/s with a mean of 1.81±0.16 spikes/s(n=75; Figs. 3A, B). In 300 consecutive spikes, the number ofbursts varied from 2 to 81 with a mean of 39.47±2.35 bursts(Fig. 4A), and the mean number of single spike per burst was2.49±0.05 (Fig. 4B). The percentage of the spikes in bursts was32.46±1.94% (Fig. 4C), and the mean ISI coefficient of variationwas 1.43±0.05 (Fig. 4D).
In rats with lesions of the LC, the firing rate of mPFCpyramidal neurons varied from 0.81 to 9.69 spikes/s (Fig. 3A);the mean firing rate increased significantly to 4.02±0.27 spikes/s compared with normal rats (P<0.01, n=74;Holm–Sidak test; Fig. 3B). The number of bursts out of 300consecutive spikes varied from 2 to 78, showing a slightly
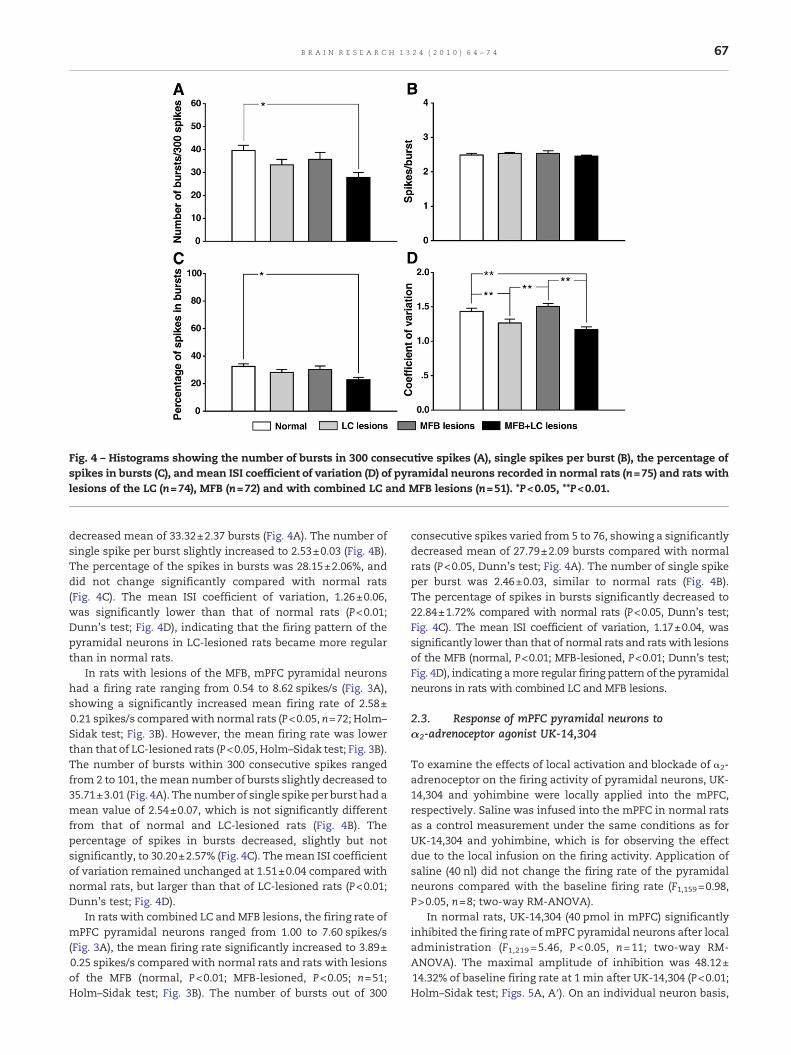
Fig. 4 – Histograms showing the number of bursts in 300 consecutive spikes (A), single spikes per burst (B), the percentage ofspikes in bursts (C), and mean ISI coefficient of variation (D) of pyramidal neurons recorded in normal rats (n=75) and rats withlesions of the LC (n=74), MFB (n=72) and with combined LC and MFB lesions (n=51). *P<0.05, **P<0.01.
67B R A I N R E S E A R C H 1 3 2 4 ( 2 0 1 0 ) 6 4 – 7 4
decreased mean of 33.32±2.37 bursts (Fig. 4A). The number ofsingle spike per burst slightly increased to 2.53±0.03 (Fig. 4B).The percentage of the spikes in bursts was 28.15±2.06%, anddid not change significantly compared with normal rats(Fig. 4C). The mean ISI coefficient of variation, 1.26±0.06,was significantly lower than that of normal rats (P<0.01;Dunn's test; Fig. 4D), indicating that the firing pattern of thepyramidal neurons in LC-lesioned rats became more regularthan in normal rats.
In rats with lesions of the MFB, mPFC pyramidal neuronshad a firing rate ranging from 0.54 to 8.62 spikes/s (Fig. 3A),showing a significantly increased mean firing rate of 2.58±0.21 spikes/s comparedwith normal rats (P<0.05, n=72; Holm–Sidak test; Fig. 3B). However, the mean firing rate was lowerthan that of LC-lesioned rats (P<0.05, Holm–Sidak test; Fig. 3B).The number of bursts within 300 consecutive spikes rangedfrom 2 to 101, themean number of bursts slightly decreased to35.71±3.01 (Fig. 4A). The number of single spike per burst had amean value of 2.54±0.07, which is not significantly differentfrom that of normal and LC-lesioned rats (Fig. 4B). Thepercentage of spikes in bursts decreased, slightly but notsignificantly, to 30.20±2.57% (Fig. 4C). Themean ISI coefficientof variation remained unchanged at 1.51±0.04 compared withnormal rats, but larger than that of LC-lesioned rats (P<0.01;Dunn's test; Fig. 4D).
In rats with combined LC and MFB lesions, the firing rate ofmPFC pyramidal neurons ranged from 1.00 to 7.60 spikes/s(Fig. 3A), the mean firing rate significantly increased to 3.89±0.25 spikes/s compared with normal rats and rats with lesionsof the MFB (normal, P<0.01; MFB-lesioned, P<0.05; n=51;Holm–Sidak test; Fig. 3B). The number of bursts out of 300
consecutive spikes varied from 5 to 76, showing a significantlydecreased mean of 27.79±2.09 bursts compared with normalrats (P<0.05, Dunn's test; Fig. 4A). The number of single spikeper burst was 2.46±0.03, similar to normal rats (Fig. 4B).The percentage of spikes in bursts significantly decreased to22.84±1.72% compared with normal rats (P<0.05, Dunn's test;Fig. 4C). The mean ISI coefficient of variation, 1.17±0.04, wassignificantly lower than that of normal rats and rats with lesionsof the MFB (normal, P<0.01; MFB-lesioned, P<0.01; Dunn's test;Fig. 4D), indicating amore regular firing pattern of the pyramidalneurons in rats with combined LC and MFB lesions.
2.3. Response of mPFC pyramidal neurons toα2-adrenoceptor agonist UK-14,304
To examine the effects of local activation and blockade of α2-adrenoceptor on the firing activity of pyramidal neurons, UK-14,304 and yohimbine were locally applied into the mPFC,respectively. Saline was infused into the mPFC in normal ratsas a control measurement under the same conditions as forUK-14,304 and yohimbine, which is for observing the effectdue to the local infusion on the firing activity. Application ofsaline (40 nl) did not change the firing rate of the pyramidalneurons compared with the baseline firing rate (F1,159=0.98,P>0.05, n=8; two-way RM-ANOVA).
In normal rats, UK-14,304 (40 pmol in mPFC) significantlyinhibited the firing rate of mPFC pyramidal neurons after localadministration (F1,219=5.46, P<0.05, n=11; two-way RM-ANOVA). The maximal amplitude of inhibition was 48.12±14.32% of baseline firing rate at 1 min after UK-14,304 (P<0.01;Holm–Sidak test; Figs. 5A, A′). On an individual neuron basis,

Fig. 5 – Representative firing rate histograms showing the local application of α2-adrenoceptor agonist UK-14,304 (40 pmol inmPFC) inhibited the firing rate of pyramidal neurons in normal rats (A) and rats with lesions of the LC (B), MFB (C) and withcombined LC and MFB lesions (D). PITHs showing the inhibitory effects of local application of UK-14,304 in the firing activityof pyramidal neurons in normal rats (A′, n=11) and rats with lesions of the LC (B′, n=10), MFB (C′, n=10) and with combinedLC and MFB lesions (D′, n=10). Arrows indicate time of administration of UK-14,304. #P<0.05, *P<0.01, **P<0.001 versusbaseline firing rate.
68 B R A I N R E S E A R C H 1 3 2 4 ( 2 0 1 0 ) 6 4 – 7 4
UK-14,304 decreased the firing rates of seven pyramidalneurons (63.64%) and did not alter the firing rates of fourpyramidal neurons (36.36%) in normal rats. In rats with lesionsof the LC, local administration of UK-14,304, at the same dose,inhibited the firing rate of all pyramidal neurons examined(F1,199=6.96, P<0.05, n=10; two-way RM-ANOVA). The maxi-mal decrease was 68.16±6.38% of baseline firing rate at 2 minafter administration (P<0.001; Holm–Sidak test; Figs. 5B, B′). Inrats with lesions of the MFB, UK-14,304 (40 pmol in mPFC)significantly inhibited the firing rate of the pyramidal neuronsafter local administration (F1,199=5.44, P<0.05, n=10; two-wayRM-ANOVA). The amplitude of inhibitionwas 50.42±13.84% ofbaseline firing rate at 2 min after UK-14,304 (P<0.001; Holm–Sidak test; Figs. 5C, C′). Of the ten pyramidal neuronsexamined in the lesioned rats, eight neurons (80%) decreasedand one neuron (10%) increased their firing rates, and one(10%) was unaltered after the administration. In rats withcombined LC and MFB lesions, local administration of UK-14,304, at same dose, inhibited the firing rate of all pyramidalneurons examined (F1,199=15.41, P<0.01, n=10; two-way RM-ANOVA). The maximal amplitude of inhibition was 66.17±22.31% of baseline firing rate at 1 min after UK-14,304(P<0.001; Holm–Sidak test; Figs. 5D, D′). The inhibitory effectof UK-14,304 began at the end of the infusion, and maintained4–6 min in different groups, respectively (Fig. 5). Post-hoccomparisons showed no significance in the normalized firingrate after the application of UK-14,304 between differentgroups (F3,409=0.46, P>0.05, two-way RM-ANOVA).
2.4. Response of mPFC pyramidal neurons toα2-adrenoceptor antagonist yohimbine
In normal rats, local administration of yohimbine (40 pmol inmPFC) significantly increased the firing rate of all pyramidal
neurons examined (F1,179=15.95, P<0.01, n=9; two-way RM-ANOVA). The maximal amplitude of increase was 135.27±19.33% of baseline firing rate at 2 min after yohimbine(P<0.001; Holm–Sidak test; Figs. 6A, A′). In rats with lesionsof the LC, yohimbine, at the same dose, did not change thefiring rate of pyramidal neurons for the 10 min after theadministration as a whole (F1,199=2.17, P>0.05, n=10; two-wayRM-ANOVA), whereas the firing rate of the pyramidal neuronssignificantly increased at first 2 min after yohimbine. Themaximal increase was 67.98±18.86% of baseline firing rate at1 min (P<0.01; Holm–Sidak test; Figs. 6B, B′). Of the tenpyramidal neurons examined, seven neurons (70%) increasedand three (30%) did not alter their firing rates after yohimbine.In rats with lesions of the MFB, yohimbine (40 pmol in mPFC)significantly increased the firing rate of the pyramidalneurons examined (F1,179=25.56, P<0.001, n=9; two-way RM-ANOVA). The maximal amplitude of increase was 81.89±11.40% of baseline firing rate at 2 min after yohimbine(P<0.001; Holm–Sidak test; Figs. 6C, C′). On an individualneuron basis, UK-14,304 increased the firing rates of eightpyramidal neurons (88.89%) and didn't alter the firing rate ofone pyramidal neuron (11.11%). In rats with combined LC andMFB lesions, local administration of yohimbine (40 pmol inmPFC) did not change the firing rate of the pyramidal neuronsexamined for the 10 min after the administration as a whole(F1,199=2.17, P>0.05, n=10; two-way RM-ANOVA). However, thefiring rate of the pyramidal neurons significantly increased atfirst 2 min after yohimbine. The maximal increase was 30.28±7.25% of baseline firing rate at 2 min (P<0.001; Holm–Sidaktest; Figs. 6D, D′). Of the ten pyramidal neurons examined, sixneurons (60%) increased and four (40%) did not alter theirfiring rates after the administration. The excitatory effect ofyohimbine started immediately after the end of the infusion,and continued 2–5 min in different groups, respectively (Fig. 6).
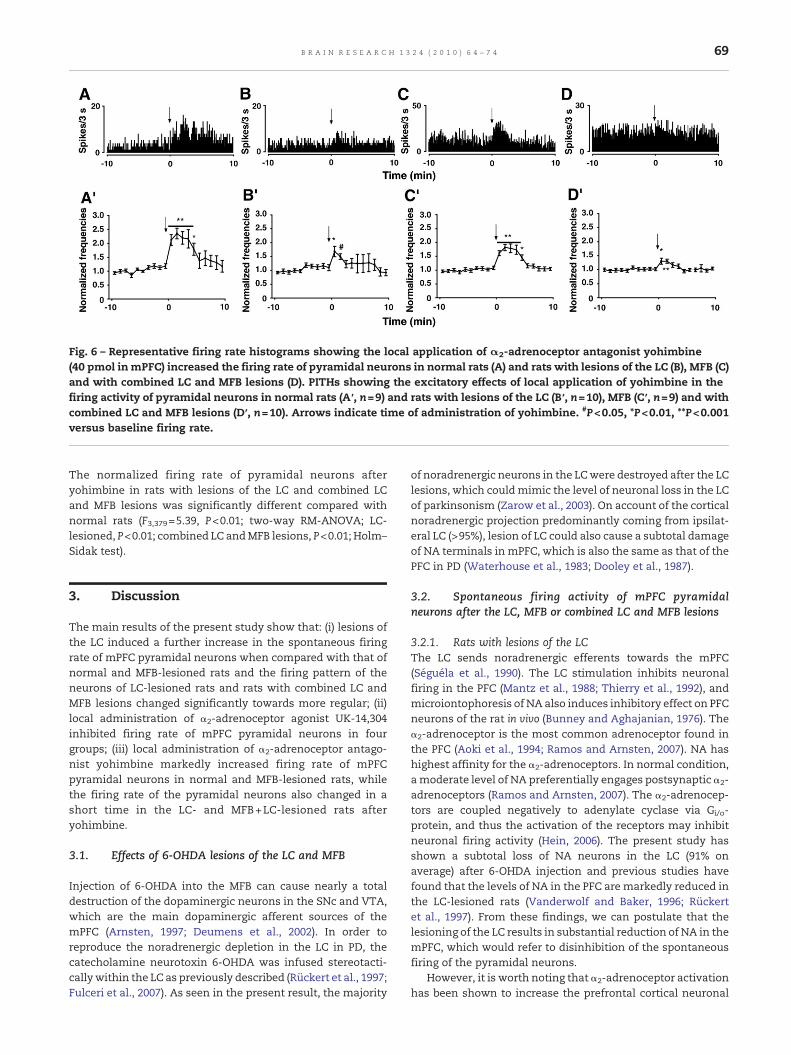
Fig. 6 – Representative firing rate histograms showing the local application of α2-adrenoceptor antagonist yohimbine(40 pmol in mPFC) increased the firing rate of pyramidal neurons in normal rats (A) and rats with lesions of the LC (B), MFB (C)and with combined LC and MFB lesions (D). PITHs showing the excitatory effects of local application of yohimbine in thefiring activity of pyramidal neurons in normal rats (A′, n=9) and rats with lesions of the LC (B′, n=10), MFB (C′, n=9) and withcombined LC and MFB lesions (D′, n=10). Arrows indicate time of administration of yohimbine. #P<0.05, *P<0.01, **P<0.001versus baseline firing rate.
69B R A I N R E S E A R C H 1 3 2 4 ( 2 0 1 0 ) 6 4 – 7 4
The normalized firing rate of pyramidal neurons afteryohimbine in rats with lesions of the LC and combined LCand MFB lesions was significantly different compared withnormal rats (F3,379=5.39, P<0.01; two-way RM-ANOVA; LC-lesioned, P<0.01; combined LC andMFB lesions, P<0.01; Holm–Sidak test).
3. Discussion
The main results of the present study show that: (i) lesions ofthe LC induced a further increase in the spontaneous firingrate of mPFC pyramidal neurons when compared with that ofnormal and MFB-lesioned rats and the firing pattern of theneurons of LC-lesioned rats and rats with combined LC andMFB lesions changed significantly towards more regular; (ii)local administration of α2-adrenoceptor agonist UK-14,304inhibited firing rate of mPFC pyramidal neurons in fourgroups; (iii) local administration of α2-adrenoceptor antago-nist yohimbine markedly increased firing rate of mPFCpyramidal neurons in normal and MFB-lesioned rats, whilethe firing rate of the pyramidal neurons also changed in ashort time in the LC- and MFB+LC-lesioned rats afteryohimbine.
3.1. Effects of 6-OHDA lesions of the LC and MFB
Injection of 6-OHDA into the MFB can cause nearly a totaldestruction of the dopaminergic neurons in the SNc and VTA,which are the main dopaminergic afferent sources of themPFC (Arnsten, 1997; Deumens et al., 2002). In order toreproduce the noradrenergic depletion in the LC in PD, thecatecholamine neurotoxin 6-OHDA was infused stereotacti-callywithin the LC as previously described (Rückert et al., 1997;Fulceri et al., 2007). As seen in the present result, the majority
of noradrenergic neurons in the LCwere destroyed after the LClesions, which couldmimic the level of neuronal loss in the LCof parkinsonism (Zarow et al., 2003). On account of the corticalnoradrenergic projection predominantly coming from ipsilat-eral LC (>95%), lesion of LC could also cause a subtotal damageof NA terminals in mPFC, which is also the same as that of thePFC in PD (Waterhouse et al., 1983; Dooley et al., 1987).
3.2. Spontaneous firing activity of mPFC pyramidalneurons after the LC, MFB or combined LC and MFB lesions
3.2.1. Rats with lesions of the LCThe LC sends noradrenergic efferents towards the mPFC(Séguéla et al., 1990). The LC stimulation inhibits neuronalfiring in the PFC (Mantz et al., 1988; Thierry et al., 1992), andmicroiontophoresis of NA also induces inhibitory effect on PFCneurons of the rat in vivo (Bunney and Aghajanian, 1976). Theα2-adrenoceptor is the most common adrenoceptor found inthe PFC (Aoki et al., 1994; Ramos and Arnsten, 2007). NA hashighest affinity for the α2-adrenoceptors. In normal condition,amoderate level of NA preferentially engages postsynaptic α2-adrenoceptors (Ramos and Arnsten, 2007). The α2-adrenocep-tors are coupled negatively to adenylate cyclase via Gi/o-protein, and thus the activation of the receptors may inhibitneuronal firing activity (Hein, 2006). The present study hasshown a subtotal loss of NA neurons in the LC (91% onaverage) after 6-OHDA injection and previous studies havefound that the levels of NA in the PFC are markedly reduced inthe LC-lesioned rats (Vanderwolf and Baker, 1996; Rückertet al., 1997). From these findings, we can postulate that thelesioning of the LC results in substantial reduction of NA in themPFC, which would refer to disinhibition of the spontaneousfiring of the pyramidal neurons.
However, it is worth noting that α2-adrenoceptor activationhas been shown to increase the prefrontal cortical neuronal

70 B R A I N R E S E A R C H 1 3 2 4 ( 2 0 1 0 ) 6 4 – 7 4
activity related to spatial working memory (Li et al., 1999). Theneuronal firing activity recorded in the study is not the sameas ours. Our present study was performed in anaesthetizedrats and the spontaneous firing of pyramidal neurons wasrecorded. This neuronal firing reflects the baseline activity ofthe neuron recorded. In contrast, stimulus-evoked neuronalactivities recorded in awake animal reflect event-relatedresponse of the neuron. It has been proposed that NA suppressspontaneous background firing while specifically enhancingthe capacity of prefrontal neurons to increase activity inresponse to specific stimulus and tuning the signal-to-noiseresponses (Winterer and Weinberger, 2003).
3.2.2. Rats with lesions of the MFBThe mPFC receives primarily dopaminergic afferents from theVTA and expresses both D1 and D2 receptors (Oades andHalliday, 1987; Vincent et al., 1995). The electrical or chemicalVTA stimulation inhibits spontaneous firing of pyramidalneurons in the PFC (Bernardi et al., 1982; Tseng et al., 2006).The inhibitory response observed in pyramidal neurons follow-ing VTA stimulation may involve DA acting directly onpostsynaptic D2 receptors or indirectly via activation ofGABAergic interneurons (Wang and O'Donnell, 2001; Tsengand O'Donnell, 2004; Tseng et al., 2006). The present study hasshown a marked reduction of DA neurons (89% on average) inthe VTA after 6-OHDA injection into the MFB of rats. Further-more, unilateral lesionof theMFBby6-OHDA led to thedecreasein the expression ofD1 andD2 receptors in the PFC of rats (Wanget al., 2005). These results suggest that the 6-OHDA lesion of theMFB causes the degeneration of DA neurons in the VTA andmarkedlydecreasesexpressionofDAreceptors in thepyramidalneurons of mPFC in rats, which would lead to an increase infiring activity of the pyramidal neurons.
3.2.3. Rats with combined LC and MFB lesionsThe present study show that the spontaneous firing activity ofthe pyramidal neurons in rats with combined LC and MFBlesions showed no significant change comparedwith rats witha simple LC lesions. Apart from the above-mentioned reasonsthat cause hyperactivity of the pyramidal neurons, anothercandidate could also be at the origin of the hyperactivity of thepyramidal neurons in rats with simple LC lesions. Severalstudies have demonstrated that the LC is a main source ofextracellular DA in the cortex, and the inhibition of the activityof the LC reducesDA release in the ipsilateralmPFC (Kawaharaet al., 2001; Devoto et al., 2003; Devoto et al., 2008). Further-more, NA depletion in the LC suppresses DA release in thenigrostriatal and mesolimbic system (Lategan et al., 1990;Lategan et al., 1992). The decreased DA content in the mPFCwould lead to disinhibition of the pyramidal neurons.
In addition, it is noteworthy that the firing pattern changedtowards more regular in LC-lesioned rats and rats withcombined LC and MFB lesions in this study. The firingsequences of mPFC pyramidal neurons in rats consist of twotypes of spikes, low frequency single spikes and spikesclusters known as bursts. In comparison with single spikes,bursts have been shown to improve the signal-to-noise ratioand transmit additional information about stimuli (Oswald etal., 2004). Previous studies have suggested that burst firing canbe generated subsequent to action potential backpropagation
into the dendritic tree (Williams and Stuart, 1999), while thegeneration of dendritic spike was promoted by activation ofα2-adrenoceptor (Barth et al., 2008). As seen in our results, theburst firing of mPFC pyramidal neurons didn't increase whilethe frequency of single spikes increased after LC lesion, whichis partially in line with the theory mentioned above.
3.3. Effects of α2-adrenoceptor agonist and antagonist infiring activity of mPFC pyramidal neurons after the LC, MFB orcombined LC and MFB lesions
Neuropharmacological investigations in the mPFC were per-formed to bring further understanding about the role of thenoradrenergic system and α2-adrenoceptor agents on sponta-neous activity of mPFC. UK-14,304 was used in the presentstudy for the higher selectivity to α2-adrenoceptor andintrinsic activity (van Veldhuizen et al., 1993; MacDonald etal., 1997). The present study shows that the local application ofthe α2-adrenoceptor agonist UK-14,304 induced a decrease inspontaneous firing activity of mPFC pyramidal neurons innormal rats and rats with lesions of the LC, MFB and withcombined LC and MFB lesions. The inhibition of pyramidalneurons inmPFC induced by local administration of UK-14,304most likely reflects the direct activation of postsynaptical α2-adrenoceptors in the pyramidal neurons recorded, because thelesions of the LC eliminate the possibility that the UK-14,304acts onpresynapticα2-adrenoceptors in themPFC.Thepresentresults imply that the beneficial effects of α2-adrenoceptoragonist on PFC function may partially attribute to theirinfluence on spontaneous firing activity of PFC and indicatethat α2-adrenoceptor agonist could reserve the effects ofnoradrenergic lesions on the spontaneous firing activity ofmPFC pyramidal neurons in rats. These results also show thatthe response of the pyramidal neurons to α2-adrenoceptorsstimulation is not significantly changed in rats with lesions ofthe LC and with combined LC and MFB lesions. Noradrenergiclesions remove the presynaptic α2-adrenoceptors. In previousreport, the undamaged postsynaptic α2-adrenoceptors re-spond to the loss of NA by slight increase in density (Dooleyet al., 1987; Ordway, 1995). However, the specific response ofthe postsynaptic α2-adrenoceptors and their intracellulardown-stream pathway to the change of NA loss is still unclear.According to the present data, the response of the pyramidalneurons to α2-adrenoceptors stimulation is not changed by LClesions. But further experiments would be required to clarifythe response of postsynaptic α2-adrenoceptors to NA deple-tion. Furthermore, the local administration of α2-adrenoceptorantagonist yohimbine increased the firing activity of thepyramidal neurons in the normal and MFB-lesioned rats,although the firing activity of the pyramidal neurons changedin a short time after yohimbine application in rats with lesionsof the LC and with combined LC and MFB lesions. The resultconfirmed that the inhibited effect of endogenousNA onmPFCpyramidal neurons is through the action on postsynaptic α2-adrenoceptors. It is worth noting that the outcomes ofyohimbine application were abated by LC lesion. In normalcondition, yohimbine exerts as a competitive antagonist ofendogenous NA and could disinhibitmPFC pyramidal neurons(Vilardaga et al., 2005). In LC-lesioned rats and rats withcombined LC and MFB lesions, noradrenergic afferents were

71B R A I N R E S E A R C H 1 3 2 4 ( 2 0 1 0 ) 6 4 – 7 4
destroyed and the NA content in synaptic clefts decreasedsignificantly. The inhibited effect of NA on postsynapticneurons reduced. That is why the outcomes of yohimbineinfusion were abated after LC lesions. Yohimbine also hasmoderate affinity for the 5-HT1A, 5-HT1B, 5-HT1D and 5-HT2
receptors. 5-HT1A and 5-HT2 receptors are founded in pyrami-dal neurons in PFC (Santana et al., 2004). It is worth notingthat theyohimbineactivates5-HT1A receptors,whereas it blocks5-HT2 receptors. Therefore, actions of yohimbine on 5-HT1A and5-HT2 receptors should inhibit the firingof pyramidal neurons inmPFC (Millan et al., 2000). So, the disinhibited influence ofyohimbine on pyramidal neurons inmPFC in the present studyshould be mediated by the action through α2-adrenoceptors.
In conclusion, the present study indicates that LC lesionsled to a further hyperactivity of the mPFC pyramidal neuronsin normal and MFB-lesioned rats and the pyramidal neuronsin mPFC fire with a more regular pattern after LC lesions. Thelocal application of α2-adrenoceptor agonist UK-14,304 andantagonist yohimbine in the mPFC decreased or increased thefiring activity of the pyramidal neurons in normal rats, andrats with lesions of the LC, MFB and with combined LC andMFB lesions, respectively. These results suggest LC-noradren-ergic neurotransmitter system plays an important role in thepathophysiology of PFC in PD, and α2-adrenoceptor agonistcan reserve the effects of noradrenergic lesions on thespontaneous firing activity of mPFC pyramidal neurons inrats after the LC lesions. Therefore, the present results mayhelp to bring understanding about the neurophysiologicalmechanisms involved in non-motor dysfunctions of PD.
4. Experimental procedures
4.1. Animals and drugs
The experiments were performed on adult male Sprague–Dawley rats weighing 220–360 g at the beginning of theexperiments, provided by the Experimental Animal Center ofShaanxi Province (Xi'an, Shaanxi, China). Animals were main-tainedona12 h light/darkcyclewith foodandwater availableadlibitum. The experimentswere carried out on four groups of rats:normal rats (n=31), rats with a 6-OHDA unilateral lesion of theLC (n=25), ratswitha 6-OHDAunilateral lesionof theMFB (n=23)and ratswithacombinedMFBandLC6-OHDA lesions (n=27).Allprocedures used in this study were performed in accordancewith the National Institute of Health Guide for the Care and Useof Laboratory Animals (NIH Publications No. 80-23) revised in1996, and according to the guideline of the Institutional AnimalCare andUseCommitteeof theUniversity.All effortsweremadeto minimize the number of animals used and their suffering.
The drugs used in this study were obtained from thefollowing sources: 6-OHDA hydrochloride, desipramine hy-drochloride, apomorphine hydrochloride, nomifensine male-ate and yohimbine hydrochloride from Sigma-Aldrich, MO,USA; UK-14,304 tartrate from Tocris Cookson, Ellisville, USA.
4.2. 6-OHDA lesions
MFB lesions were inflicted on the rats with 4% chloral hydrate(300 mg/kg, i.p.) anaesthetized and fixed to a stereotaxic
frame (SN-2N, Narishige, Tokyo, Japan). Thirty minutes priorto the injection of 6-OHDA, animals were pretreated withdesipramine (25 mg/kg, i.p.) in order to protect NA neurons.The 6-OHDA (12 μg free base in 4 μl ice cold saline containing0.01% ascorbic acid) was injected at the left MFB according tothe following stereotaxic coordinates (in mm) relative tobregma and dura: AP (anterior–posterior) −4.4; L (lateral) 1.1; D(dorsal–ventral) 7.8 (Paxinos and Watson, 2005). In order tominimize the damage to the brain and the drug diffusion, thesolution was infused through a glass micropipette connectedwith a 10 µl Hamilton microsyringe (World Precision Instru-ments, Sarasota, USA). After the injection, the micropipettewas left in place for an additional 5 min before being slowlywithdrawn. Two weeks post-surgery, rats were given apo-morphine (0.05 mg/kg, s.c.) and those exhibiting more than 20contralateral turns per 5 min were chosen for the furtherinvestigation (Wang et al., 2009). All rats used in this studyturned consistently toward the side contralateral to the sideof the lesion of more than 28 turns per 5 min (38±4 turns per5 min).
6-OHDA lesion of the LC was performed as previouslydescribed (Rückert et al., 1997; Fulceri et al., 2007). Animalswere anaesthetized with 4% chloral hydrate (300 mg/kg, i.p.),pretreated with nomifensine (10 mg/kg, i.p.) to protect DAneurons (Caillé et al., 1999), placed in a stereotaxic frame andinjectedwith 6-OHDA (12 μg/3 μl) into the left LC (AP −9.9, L 1.4,D 7.0; Paxinos and Watson, 2005). For the group of rats withcombined MFB and LC lesions, the LC was lesioned 2 weeksafter the injection of 6-OHDA into the MFB, as same asdescribed above.
4.3. Electrophysiological recordings
Electrophysiological recordings were performed 3 weeks afterlesions of the MFB and 1 week after lesions of the LC.Extracellular single unit recordings were made in ratsanaesthetized with 4% chloral hydrate (400 mg/kg, i.p.) andmounted in a stereotaxic frame as described previously (Wanget al., 2009). No recordings were made until the formeranaesthesia began to wear off, as indicated by sporadicwhisker movement. The animals remained anaesthetized toa moderate but stable level throughout the experiments byintravenous administration of additional doses of chloralhydrate (90 mg/kg·h) via a polyethylene tubing driven by aninfusion pump. Body temperaturewasmaintained at 37±0.5 °C,heart rate and papillary diameter monitored throughout theexperiment. Glass microelectrodes (8–12 MΩ) filled with 1%Pontamine Sky Blue in 0.5 M sodium acetate were directedstereotaxically to the mPFC (AP 2.8–3.4, L 0.6–0.8, D 1.5–5.0;PaxinosandWatson, 2005).Theneuronal firingswereamplified,bandpass-filtered using a pre-amplifier (AVB-11A, Nihon Koh-den, Tokyo, Japan), displayed on an oscilloscope (VC-11, NihonKohden, Tokyo, Japan) and stored in a computer equippedwithPowerLab digital data acquisition system (LabChart V6, Power-Lab, AD Instruments, Castle Hill, Australia) for off-line analysis.Only single-spike neuronal activity with a good signal-to-noiseratio (3:1) and stable firing were investigated. The mPFCpyramidal neuron recorded typically had a broad actionpotential (>1 ms) and an irregular firing pattern with burstactivity (Tseng et al., 2006).

72 B R A I N R E S E A R C H 1 3 2 4 ( 2 0 1 0 ) 6 4 – 7 4
4.4. The firing pattern analysis
The firing pattern of pyramidal neurons in the mPFC wasanalyzed and the following parameters were calculated:number of single spike per burst, number of bursts andpercentage of spikes in bursts. Burst firing was defined as thepresence of two or more spikes with short interspike interval(≤16 ms) (Figs. 1A, C, D), which determined from burstexamples and interspike interval (ISI) histograms (binwidth=4 ms) (Cocatre-Zilgien and Delcomyn, 1992; Bastianand Nguyenkim, 2001). The mean ISI coefficient of variationwas also calculated. The mean ISI coefficient of variationreflects the degree of regularity of neuronal firing, which isdefined as the ratio between S.D. and mean ISI (Wang et al.,2009). These parameters regarding the firing pattern weredetermined by 300 consecutive spikes.
4.5. Drug administration
Local administration of the α2-adrenoceptor agonist UK-14,304and antagonist yohimbine into the mPFC was performed via aglass micropipette connected via polyethylene tubing to a 1 µlmicrosyringe (air-tight Hamilton; World Precision Instru-ments, Sarasota, USA) driven by a syringe pump. Infusionmicropipettewas pulled from glass capillary and broken underamicroscope at an external diameter of 50 µm, and then glued150 µm above the tip of the recordingmicroelectrode. The totalvolume of 40 nl of UK-14,304, yohimbine or saline was infusedwithin 1 min. The rats were given 40 pmol UK-14,304 oryohimbine in 40 nl saline. The α2-adrenoceptor agonist UK-14,304 and antagonist yohimbine used in the study wereselected on the basis of previous reports and pilot experimentof the study (van Veldhuizen et al., 1993; MacDonald et al.,1997). Doses for local application of α2-adrenoceptor agentswere also selected on the basis of the pilot study thatidentified the minimum response concentration of eachdrug in normal rats. Drug application induced a mechanicaleffect, which usually induced a decrease in spike amplitude. Inmost cases, the spike amplitude recovered within 1 min.Therefore, the early effects of the infusion on the firing activityof a recorded neuron, which occurred during the first minuteafter injection, were not recorded. Only one neuron wasobserved per animal during the procedure of the localadministration.
4.6. Histology and Immunocytochemistry
At the end of each experiment, the recording site was markedby the ejection of Pontamine Sky Blue (−20 µA, 15 min). The ratwas given an overdose of urethane, and perfused with 150 mlnormal saline followed by 200 ml of 4% paraformaldehyde, thebrain was immediately removed and post-fixed in the samefixative for 4 h. It was then placed in phosphate bufferedsaline (PBS) containing 20% sucrose overnight. The brainswere frozen and cut into 40 µm thick coronal sections using acryostat. Cresyl violet staining of the sections mounted ongelatin-coated slides was used to determine the location ofrecording sites.
To determine the extent of DA or NA degeneration,sections of the SNc, VTA or LC from rats receiving 6-OHDA
injection were examined for immunocytochemical staining oftyrosine hydroxylase (TH). Briefly, the sections were preincu-bated with 3% bovine serum albumin in PBS containing 0.3%TritonX-100 for 30 min at room temperature and thenincubated at 4 °C for 48 h with rabbit polyclonal anti-THantibodies (1:1000, AB 152, Chemicon, CA, USA). Next, sectionswere incubated for 2 h with biotinylated anti-rabbit IgG (1:200,Chemicon, CA, USA), and incubated for 2 h with the avidin−biotin-peroxidase complex (1:100, Vector Laboratories, USA) atroom temperature. Finally they were exposed for 5−10 min atroom temperature to a solution of 0.05% 3,3′-diaminobemi-dine (Sigma-Aldrich, MO, USA) containing 0.02% H2O2, whichserved as chromogen in the subsequent visualization reaction.Rinses were performed between each step excluding theblocking solution step and the addition of the primaryantibody. Sections were rinsed, mounted onto gelatin-coatedslides, dehydrated, cleared in xylene and coverslipped.
After TH staining, counting of the DA neuron bodies in theSNc, VTA andNAneuron bodies in the LCwas carried out on therepresentative 3 sections per animal that best matched thespecified levels in the stereotaxic atlas of rat brain. A neuronwasconsidered when intact, round with clear nucleus and/orcytoplasm. The full extent of the structure in each section wasexamined in the lesioned and unlesioned sides. Only sections inwhich the medial and lateral parts of the SNc and VTA wereclearly separatedby themedial terminalnucleusof theaccessoryoptic tract level were selected for analysis of TH-ir neuronnumber. For assessment of VTA neuron counts, neurons werecounted lateral to theblack line (Figs. 2D, F). This line is defined tocoincidewith the tangent plane of the fasciculus retroflexus (Leeet al., 1996). Only ratswith a total loss of TH immunoreactivity inthe SNc anda subtotal loss (>80%) of TH immunoreactivity in theLC after specific lesionwere used to analyze electrophysiologicalrecordings.
4.7. Data treatment and statistical analysis
The firing rate was analyzed with one-way ANOVA. Post hocmultiple comparisons were made using the Holm–Sidak test.The mean ISI coefficient of variation, number of single spikeper burst, number of bursts per 300 spikes and percentage ofthe spikes in bursts were compared using the Kruskal−Wallisone-way ANOVA on ranks follow by Dunn's test for multiplecomparisons, as the data were not normally distributed.Comparisons of TH-ir neurons of the SNc, VTA and LC forlesioned and unlesioned sides in each group were assessedusing paired Student's t test.
The baseline activity of identified neuronswas recorded for10 min before any treatment. The effect of each drug on thefiring rate of pyramidal neurons inmPFCwas followed on-linewith constructions of peri-infusion time histogram (PITH,1 min bin width) 10 min after the local infusion, and then themean and S.D. of counts per bin was determined. Theexceeding or bellowing the mean baseline activity by 1 S.D.was considered a significant alteration for an individualneuron. Furthermore, we calculated the relative firing rate ineach neuron, which was defined as the ratio of firing rate ineachminute to themean basal firing rate. This value was usedto determine the averaged effects of drugs in mPFC pyramidalneuronswith different firing levels (Suzuki et al., 2002; Belujon

73B R A I N R E S E A R C H 1 3 2 4 ( 2 0 1 0 ) 6 4 – 7 4
et al., 2007). The differences of relative firing rate before andafter drug infusion and the differences among four groupsafter drug application were tested statistically by two-wayrepeated measures ANOVA (two-way RM-ANOVA) with posthoc multiple comparisons (Holm–Sidak test). Statistical anal-ysis was performed using SigmaStat for Windows Version 3.5,data are presented as mean±S.E.M and P values <0.05 wereconsidered to indicate statistical significance.
Acknowledgments
This study was supported by the National Natural ScienceFoundation of China (No. 30970954), and the Natural ScienceFoundation (SJ08C210) and Science and Technological Project[2008K15-01(9)] of Shaanxi Province, China.
R E F E R E N C E S
Aoki, C., Go, C., Venkatesan, C., Kurose, H., 1994. Perikaryal andsynaptic localization of alpha 2A-adrenergic receptor-likeimmunoreactivity. Brain Res. 650, 181–204.
Arnsten, A., 1997. Catecholamine regulation of the prefrontalcortex. J. Psychopharmacol. 11, 151–162.
Arnsten,A., Goldman-Rakic, P., 1985.Alpha2-adrenergicmechanismsin prefrontal cortex associated with cognitive decline in agednonhuman primates. Science 230, 1273–1276.
Barth, A., Vizi, E., Zelles, T., Lendvai, B., 2008. Alpha2-adrenergicreceptors modify dendritic spike generation via HCN channelsin the prefrontal cortex. J. Neurophysiol. 99, 394–401.
Bastian, J., Nguyenkim, J., 2001. Dendritic modulation of burst-likefiring in sensory neurons. J. Neurophysiol. 85, 10–22.
Belujon, P., Bezard, E., Taupignon,A., Bioulac, B., Benazzouz,A., 2007.Noradrenergic modulation of subthalamic nucleus activity:behavioral and electrophysiological evidence in intact and6-hydroxydopamine-lesioned rats. J. Neurosci. 27, 9595–9606.
Bernardi, G., Cherubini, E., Marciani, M.G., Mercuri, N., Stanzione,P., 1982. Responses of intracellularly recorded cortical neuronsto the iontophoretic application of dopamine. Brain Res. 245,267–274.
Bosboom, J., Stoffers, D., Wolters, E., 2004. Cognitive dysfunctionand dementia in Parkinson's disease. J. Neural Transm. 111,1303–1315.
Bunney, B.S., Aghajanian, G.K., 1976. Dopamine and norepinephrineinnervated cells in the rat prefrontal cortex: pharmacologicaldifferentiation usingmicroiontophoretic techniques. Life Sci. 19,1783–1789.
Caillé, S., Espejo, E., Reneric, J., Cador, M., Koob, G., Stinus, L., 1999.Total neurochemical lesion of noradrenergic neurons of thelocus ceruleus does not alter either naloxone-precipitated orspontaneous opiate withdrawal nor does it influence ability ofclonidine to reverse opiate withdrawal. J. Pharmacol. Exp. Ther.290, 881–892.
Cash, R., Ruberg, M., Raisman, R., Agid, Y., 1984. Adrenergicreceptors in Parkinson's disease. Brain Res. 322, 269–275.
Cocatre-Zilgien, J., Delcomyn, F., 1992. Identification of bursts inspike trains. J. Neurosci. Methods 41, 19–30.
Cummings, J., 1992. Depression and Parkinson's disease: a review.Am. J. Psychiatry 149, 443–454.
Deumens, R., Blokland, A., Prickaerts, J., 2002.Modeling Parkinson'sdisease in rats: an evaluation of 6-OHDA lesions of thenigrostriatal pathway. Exp. Neurol. 175, 303–317.
Devoto, P., Flore, G., Longu, G., Pira, L., Gessa, G., 2003. Origin ofextracellular dopamine from dopamine and noradrenaline
neurons in the medial prefrontal and occipital cortex. Synapse50, 200–205.
Devoto, P., Flore, G., Saba, P., Castelli, M., Piras, A., Luesu, W.,Viaggi, M., Ennas, M., Gessa, G., 2008. 6-Hydroxydopaminelesion in the ventral tegmental area fails to reduceextracellular dopamine in the cerebral cortex. J. Neurosci. Res.86, 1647–1658.
Dooley, D., Jones, G., Robbins, T., 1987. Noradrenaline- andtime-dependent changes in neocortical alpha 2- andbeta 1-adrenoceptor binding in dorsal noradrenergicbundle-lesioned rats. Brain Res. 420, 152–156.
Fulceri, F., Biagioni, F., Ferrucci, M., Lazzeri, G., Bartalucci, A., Galli,V., Ruggieri, S., Paparelli, A., Fornai, F., 2007. Abnormalinvoluntary movements (AIMs) following pulsatiledopaminergic stimulation: severe deterioration andmorphological correlates following the loss of locuscoeruleus neurons. Brain Res. 1135, 219–229.
Fuster, J.M., 1997. The Prefrontal Cortex: Anatomy, Physiology andNeuropsychology of the Frontal Lobe 3rd ed. Lippincott-William& Wilkins, Philadelphia.
Graeber, M., Grasbon-Frodl, E., Eitzen, U., Kösel, S., 1998.Neurodegeneration and aging: role of the second genome.J. Neurosci. Res. 52, 1–6.
Hein, L., 2006. Adrenoceptors and signal transduction in neurons.Cell Tissue Res. 326, 541–551.
Kawahara, H., Kawahara, Y., Westerink, B., 2001. Thenoradrenaline-dopamine interaction in the rat medialprefrontal cortex studied by multi-probe microdialysis.Eur. J. Pharmacol. 418, 177–186.
Lategan, A., Marien, M., Colpaert, F., 1990. Effects of locuscoeruleus lesions on the release of endogenous dopamine inthe rat nucleus accumbens and caudate nucleus as determinedby intracerebral microdialysis. Brain Res. 523, 134–138.
Lategan, A., Marien, M., Colpaert, F., 1992. Suppression ofnigrostriatal and mesolimbic dopamine release in vivofollowing noradrenaline depletion by DSP-4: a microdialysisstudy. Life Sci. 50, 995–999.
Lee, C., Sauer, H., Bjorklund, A., 1996. Dopaminergic neuronaldegeneration and motor impairments following axonterminal lesion by instrastriatal 6-hydroxydopamine in the rat.Neuroscience 72, 641–653.
Li, B., Mao, Z., Wang, M., Mei, Z., 1999. Alpha-2 adrenergicmodulation of prefrontal cortical neuronal activity related tospatialworkingmemory inmonkeys.Neuropsychopharmacology21, 601–610.
MacDonald, E., Kobilka, B., Scheinin, M., 1997. Genetargeting–homing in on alpha 2-adrenoceptor-subtypefunction. Trends Pharmacol. Sci. 18, 211–219.
Mantz, J., Milla, C., Glowinski, J., Thierry, A.M., 1988. Differentialeffects of ascending neurons containing dopamine andnoradrenaline in the control of spontaneous activity and ofevoked responses in the rat prefrontal cortex. Neuroscience 27,517–526.
Millan, M., Newman-Tancredi, A., Audinot, V., Cussac, D., Lejeune,F., Nicolas, J., Cogé, F., Galizzi, J., Boutin, J., Rivet, J., Dekeyne, A.,Gobert, A., 2000. Agonist and antagonist actions ofyohimbine as compared to fluparoxan at alpha(2)-adrenergicreceptors (AR)s, serotonin (5-HT)(1A), 5-HT(1B), 5-HT(1D) anddopamine D(2) and D(3) receptors. Significance for themodulation of frontocorticalmonoaminergic transmission anddepressive states. Synapse 35, 79–95.
Miller, E., Cohen, J., 2001. An integrative theory of prefrontal cortexfunction. Annu. Rev. Neurosci. 24, 167–202.
Oades, R.D., Halliday, G.M., 1987. Ventral tegmental (A10) system:neurobiology. 1. Anatomy and connectivity. Brain Res. Rev. 12,117–165.
Ordway, G., 1995. Effect of noradrenergic lesions on subtypes ofalpha 2-adrenoceptors in rat brain. J. Neurochem. 64,1118–1126.

74 B R A I N R E S E A R C H 1 3 2 4 ( 2 0 1 0 ) 6 4 – 7 4
Oswald, A., Chacron, M., Doiron, B., Bastian, J., Maler, L., 2004.Parallel processing of sensory input by bursts and isolatedspikes. J. Neurosci. 24, 4351–4362.
Paxinos, G., Watson, C., 2005. The Rat Brain in StereotaxicCoordinates, 5th ed. Academic Press, New York.
Ramos, B., Arnsten, A., 2007. Adrenergic pharmacology andcognition: focus on the prefrontal cortex. Pharmacol. Ther. 113,523–536.
Remy, P., Doder, M., Lees, A., Turjanski, N., Brooks, D., 2005.Depression in Parkinson's disease: loss of dopamine andnoradrenaline innervation in the limbic system. Brain 128,1314–1322.
Ridderinkhof, K., Ullsperger, M., Crone, E., Nieuwenhuis, S., 2004.The role of the medial frontal cortex in cognitive control.Science 306, 443–447.
Riekkinen, M., Riekkinen, P.J., 1999. Alpha2-adrenergic agonistclonidine for improving spatial working memory inParkinson's disease. J. Clin. Psychopharmacol. 19, 444–449.
Rückert, N., Bubser, M., Schmidt, W., 1997. 6-Hydroxydopaminelesion of locus coeruleus and the antiparkinsonian potential ofNMDA-receptor antagonists in rats. J. Neural Transm. 104,363–377.
Santana, N., Bortolozzi, A., Serrats, J., Mengod, G., Artigas, F., 2004.Expression of serotonin-1A and serotonin-2A receptors inpyramidal and GABAergic neurons of the rat prefrontal cortex.Cereb. Cortex 14, 1100–1109.
Scatton, B., Javoy-Agid, F., Rouquier, L., Dubois, B., Agid, Y., 1983.Reduction of cortical dopamine, noradrenaline, serotonin andtheir metabolites in Parkinson's disease. Brain Res. 275,321–328.
Schapira, A., 2008. Progress in Parkinson's disease. Eur. J. Neurol.15, 1.
Suzuki, Y., Jodo, E., Takeuchi, S., Niwa, S., Kayama, Y., 2002. Acuteadministration of phencyclidine induces tonic activation ofmedial prefrontal cortex neurons in freely moving rats.Neuroscience 114, 769–779.
Séguéla, P., Watkins, K.C., Geffard, M., Descarries, L., 1990.Noradrenaline axon terminals in adult rat neocortex: animmunocytochemical analysis in serial thin sections.Neuroscience 35, 249–264.
Thierry, A.M., Mantz, J., Glowinski, J., 1992. Influence ofdopaminergic and noradrenergic afferents on their targetcells in the rat medial prefrontal cortex. Adv. Neurol. 57,545–554.
Tseng, K.Y., Mallet, N., Toreson, K., Le Moine, C., Gonon, F.,O'Donnell, P., 2006. Excitatory response of prefrontal corticalfast-spiking interneurons to ventral tegmental areastimulation in vivo. Synapse 59, 412–417.
Tseng, K.Y., O'Donnell, P., 2004. Dopamine–glutamate interactionscontrolling prefrontal cortical pyramidal cell excitabilityinvolve multiple signaling mechanisms. J. Neurosci. 24,5131–5139.
van Veldhuizen, M., Feenstra, M., Heinsbroek, R., Boer, G., 1993. Invivo microdialysis of noradrenaline overflow: effects ofalpha-adrenoceptor agonists and antagonists measured bycumulative concentration–response curves. Br. J. Pharmacol.109, 655–660.
Vanderwolf, C., Baker, G., 1996. The role of brain noradrenaline incortical activation and behavior: a study of lesions of the locuscoeruleus, medial thalamus and hippocampus-neocortex andofmuscarinic blockade in the rat. Behav. Brain Res. 78, 225–234.
Vilardaga, J., Steinmeyer, R., Harms, G., Lohse, M., 2005. Molecularbasis of inverse agonism in a G protein-coupled receptor. Nat.Chem. Biol. 1, 25–28.
Vincent, S.L., Khan, Y., Benes, F.M., 1995. Cellular colocalization ofdopamine D1 and D2 receptors in the rat medial prefrontalcortex. Synapse 19, 112–120.
Wang, J., O'Donnell, P., 2001. D1 dopamine receptors potentiateNMDA-mediated excitability increase in layer V prefrontalcortical pyramidal neurons. Cereb. Cortex 11, 452–462.
Wang, Q., Wang, P., McLachlan, C., Wong, P., 2005. Simvastatinreverses the downregulation of dopamine D1 and D2 receptorexpression intheprefrontal cortexof6-hydroxydopamine-inducedParkinsonian rats. Brain Res. 1045, 229–233.
Wang, S., Zhang, Q.J., Liu, J., Wu, Z.H., Wang, T., Gui, Z.H., Chen, L.,Wang, Y., 2009. Unilateral lesion of the nigrostriatal pathwayinduces an increase of neuronal firing of the midbrain raphenuclei 5-HT neurons and a decrease of their response to5-HT1A receptor stimulation in the rat. Neuroscience 159,850–861.
Waterhouse, B., Lin, C., Burne, R., Woodward, D., 1983. Thedistribution of neocortical projection neurons in the locuscoeruleus. J. Comp. Neurol. 217, 418–431.
Williams, S., Stuart, G., 1999. Mechanisms and consequences ofaction potential burst firing in rat neocortical pyramidalneurons. J. Physiol. 521, 467–482.
Winterer, G., Weinberger, D.R., 2003. Cortical signal-to-noise ratio:insight into the pathophysiology and genetics ofschizophrenia. Clin. Neurosci. Res. 3, 55–66.
Zarow, C., Lyness, S.A., Mortimer, J.A., Chui, H.C., 2003. Neuronalloss is greater in the locus coeruleus than nucleus basalis andsubstantia nigra in Alzheimer and Parkinson diseases. Arch.Neurol. 60, 337–341.
Zweig, R., Cardillo, J., Cohen, M., Giere, S., Hedreen, J., 1993. Thelocus ceruleus and dementia in Parkinson's disease. Neurology43, 986–991.




















