
BMC Evolutionary Biology
12
BioMed Central Page 1 of 12 (page number not for citation purposes) BMC Evolutionary Biology Open Access Research article AFLP analysis reveals a lack of phylogenetic structure within Solanum section Petota Mirjam MJ Jacobs 1,2,6 , Ronald G van den Berg* 1,6 , Vivianne GAA Vleeshouwers 5,6 , Marcel Visser 5,6 , Rolf Mank 4,6 , Mariëlle Sengers 4,6 , Roel Hoekstra 3,6 and Ben Vosman 2,6 Address: 1 Biosystematics Group, Wageningen University and Research Centre, Wageningen, The Netherlands, 2 Plant Research International, Wageningen University and Research Centre, Droevendaalsesteeg 1, 6708PB Wageningen, The Netherlands, 3 Centre for Genetic Resources, Wageningen University and Research Centre, Wageningen, The Netherlands, 4 KEYGENE N.V., Wageningen, The Netherlands, 5 Plant Breeding, Wageningen University and Research Centre, Wageningen, The Netherlands and 6 Centre for BioSystems Genomics, P.O. Box 98, 6700 AB Wageningen, The Netherlands Email: Mirjam MJ Jacobs - [email protected]; Ronald G van den Berg* - [email protected]; Vivianne GAA Vleeshouwers - [email protected]; Marcel Visser - [email protected]; Rolf Mank - [email protected]; Mariëlle Sengers - [email protected]; Roel Hoekstra - [email protected]; Ben Vosman - [email protected] * Corresponding author Abstract Background: The secondary genepool of our modern cultivated potato (Solanum tuberosum L.) consists of a large number of tuber-bearing wild Solanum species under Solanum section Petota. One of the major taxonomic problems in section Petota is that the series classification (as put forward by Hawkes) is problematic and the boundaries of some series are unclear. In addition, the classification has received only partial cladistic support in all molecular studies carried out to date. The aim of the present study is to describe the structure present in section Petota. When possible, at least 5 accessions from each available species and 5 individual plants per accession (totally approx. 5000 plants) were genotyped using over 200 AFLP markers. This resulted in the largest dataset ever constructed for Solanum section Petota. The data obtained are used to evaluate the 21 series hypothesis put forward by Hawkes and the 4 clade hypothesis of Spooner and co-workers. Results: We constructed a NJ tree for 4929 genotypes. For the other analyses, due to practical reasons, a condensed dataset was created consisting of one representative genotype from each available accession. We show a NJ jackknife and a MP jackknife tree. A large part of both trees consists of a polytomy. Some structure is still visible in both trees, supported by jackknife values above 69. We use these branches with >69 jackknife support in the NJ jackknife tree as a basis for informal species groups. The informal species groups recognized are: Mexican diploids, Acaulia, Iopetala, Longipedicellata, polyploid Conicibaccata, diploid Conicibaccata, Circaeifolia, diploid Piurana and tetraploid Piurana. Conclusion: Most of the series that Hawkes and his predecessors designated can not be accepted as natural groups, based on our study. Neither do we find proof for the 4 clades proposed by Spooner and co-workers. A few species groups have high support and their inner structure displays also supported subdivisions, while a large part of the species cannot be structured at all. We believe that the lack of structure is not due to any methodological problem but represents the real biological situation within section Petota. Published: 14 May 2008 BMC Evolutionary Biology 2008, 8:145 doi:10.1186/1471-2148-8-145 Received: 20 December 2007 Accepted: 14 May 2008 This article is available from: http://www.biomedcentral.com/1471-2148/8/145 © 2008 Jacobs et al; licensee BioMed Central Ltd. This is an Open Access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/2.0 ), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of BMC Evolutionary Biology

BioMed CentralBMC Evolutionary Biology
ss
Open AcceResearch articleAFLP analysis reveals a lack of phylogenetic structure within Solanum section PetotaMirjam MJ Jacobs1,2,6, Ronald G van den Berg*1,6, Vivianne GAA Vleeshouwers5,6, Marcel Visser5,6, Rolf Mank4,6, Mariëlle Sengers4,6, Roel Hoekstra3,6 and Ben Vosman2,6Address: 1Biosystematics Group, Wageningen University and Research Centre, Wageningen, The Netherlands, 2Plant Research International, Wageningen University and Research Centre, Droevendaalsesteeg 1, 6708PB Wageningen, The Netherlands, 3Centre for Genetic Resources, Wageningen University and Research Centre, Wageningen, The Netherlands, 4KEYGENE N.V., Wageningen, The Netherlands, 5Plant Breeding, Wageningen University and Research Centre, Wageningen, The Netherlands and 6Centre for BioSystems Genomics, P.O. Box 98, 6700 AB Wageningen, The Netherlands
Email: Mirjam MJ Jacobs - [email protected]; Ronald G van den Berg* - [email protected]; Vivianne GAA Vleeshouwers - [email protected]; Marcel Visser - [email protected]; Rolf Mank - [email protected]; Mariëlle Sengers - [email protected]; Roel Hoekstra - [email protected]; Ben Vosman - [email protected]
* Corresponding author
AbstractBackground: The secondary genepool of our modern cultivated potato (Solanum tuberosum L.) consists of alarge number of tuber-bearing wild Solanum species under Solanum section Petota. One of the major taxonomicproblems in section Petota is that the series classification (as put forward by Hawkes) is problematic and theboundaries of some series are unclear. In addition, the classification has received only partial cladistic support inall molecular studies carried out to date.
The aim of the present study is to describe the structure present in section Petota. When possible, at least 5accessions from each available species and 5 individual plants per accession (totally approx. 5000 plants) weregenotyped using over 200 AFLP markers. This resulted in the largest dataset ever constructed for Solanum sectionPetota. The data obtained are used to evaluate the 21 series hypothesis put forward by Hawkes and the 4 cladehypothesis of Spooner and co-workers.
Results: We constructed a NJ tree for 4929 genotypes. For the other analyses, due to practical reasons, acondensed dataset was created consisting of one representative genotype from each available accession. Weshow a NJ jackknife and a MP jackknife tree. A large part of both trees consists of a polytomy. Some structure isstill visible in both trees, supported by jackknife values above 69. We use these branches with >69 jackknifesupport in the NJ jackknife tree as a basis for informal species groups. The informal species groups recognizedare: Mexican diploids, Acaulia, Iopetala, Longipedicellata, polyploid Conicibaccata, diploid Conicibaccata,Circaeifolia, diploid Piurana and tetraploid Piurana.
Conclusion: Most of the series that Hawkes and his predecessors designated can not be accepted as naturalgroups, based on our study. Neither do we find proof for the 4 clades proposed by Spooner and co-workers. Afew species groups have high support and their inner structure displays also supported subdivisions, while a largepart of the species cannot be structured at all. We believe that the lack of structure is not due to anymethodological problem but represents the real biological situation within section Petota.
Published: 14 May 2008
BMC Evolutionary Biology 2008, 8:145 doi:10.1186/1471-2148-8-145
Received: 20 December 2007Accepted: 14 May 2008
This article is available from: http://www.biomedcentral.com/1471-2148/8/145
© 2008 Jacobs et al; licensee BioMed Central Ltd. This is an Open Access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/2.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Page 1 of 12(page number not for citation purposes)

BMC Evolutionary Biology 2008, 8:145 http://www.biomedcentral.com/1471-2148/8/145
BackgroundThe secondary genepool of our modern cultivated potato(Solanum tuberosum L.) consists of a large number oftuber-bearing wild Solanum species which grow in varioushabitats from the southern states of the USA to the mostsouthern parts of Chile and Argentina. These wild speciesare important as a resource for valuable traits that can beused to improve the quality of the cultivars, includingresistance against important diseases like Phytophthorainfestans and potato cyst nematodes (Globodera spp.).Therefore it is no surprise that the wild relatives of the cul-tivated potato have since long drawn the attention ofmany plant breeders and botanists. To benefit most fromthe possibilities that the secondary genepool has to offer,it is necessary to have a good insight in the taxonomy. Theclassical treatments of potato taxonomy are from Correll[1], and Hawkes [2], later followed by reviews fromSpooner and Hijmans [3], Spooner and Salas [4], and vanden Berg and Jacobs [5].
There are two major taxonomic problems in the sectionPetota. First, many described species are extremely similarto each other and section Petota seems to be overclassified[5]. In many cases, potato species can only be distin-guished by means of multivariate analysis of quantitativecharacters and/or on the basis of geographic origin [6-9].
The main cause for these difficulties is the ability of manyspecies in section Petota to hybridize easily with other spe-cies [4]. Many species have been suspected to arise fromhybrid speciation. Other causes are high morphologicalsimilarity among species, and phenotypic plasticity in dif-ferent environments [3]. In recent reviews the number ofspecies is reduced due to increased insights in potato tax-onomy. Hawkes [2] recognized 227 tuber bearing species(7 cultivated species included) and 9 non-tuber-bearingspecies within section Petota. Spooner and Hijmans [3]recognized 203 tuber-bearing species including 7 culti-vated species. Finally, Spooner and Salas [4], reduced thenumber further to 189 species (including 1 cultivated spe-cies) in section Petota.
The second taxonomic problem is the series classification.Hawkes [2] classified section Petota into 19 tuber bearingseries plus two non-tuber bearing series that vary consid-erably in the number of species included. The boundariesbetween some series are unclear. As outlined earlier bySpooner et al. [10], the series classification of Hawkes andprevious authors has received only partial cladistic sup-port in any molecular study to date. The cpDNA RFLP datafrom Spooner and Sytsma [11], Castillo and Spooner[12], Rodriguez and Spooner [13], and Spooner and Cas-tillo [14] could only find support for a classification in 4clades.
The aim of the present study is to focus on the secondproblem and to describe the structure within sectionPetota. In the present study the largest number of speciesand accessions to date are examined in one simultaneousAFLP analysis. The obtained data are used for evaluationof the hypothesis put forward by Hawkes [2] that sectionPetota can be divided in 21 series and the hypothesis ofSpooner and Castillo [14], that the section consists of 4clades only.
AFLP has proven to be a useful method to solve phyloge-netic relationships at a low taxonomic level [15-17]. Theapplication of AFLP has many advantages. It produceshighly reproducible data [18], it does not need a priorisequence information and it has the ability of high reso-lution [17]. Because AFLP generates fragments at randomover the whole genome it avoids the problem that manysequence data based phylogeny reconstructions have, e.g.the generation of a gene tree instead of a species tree [15].
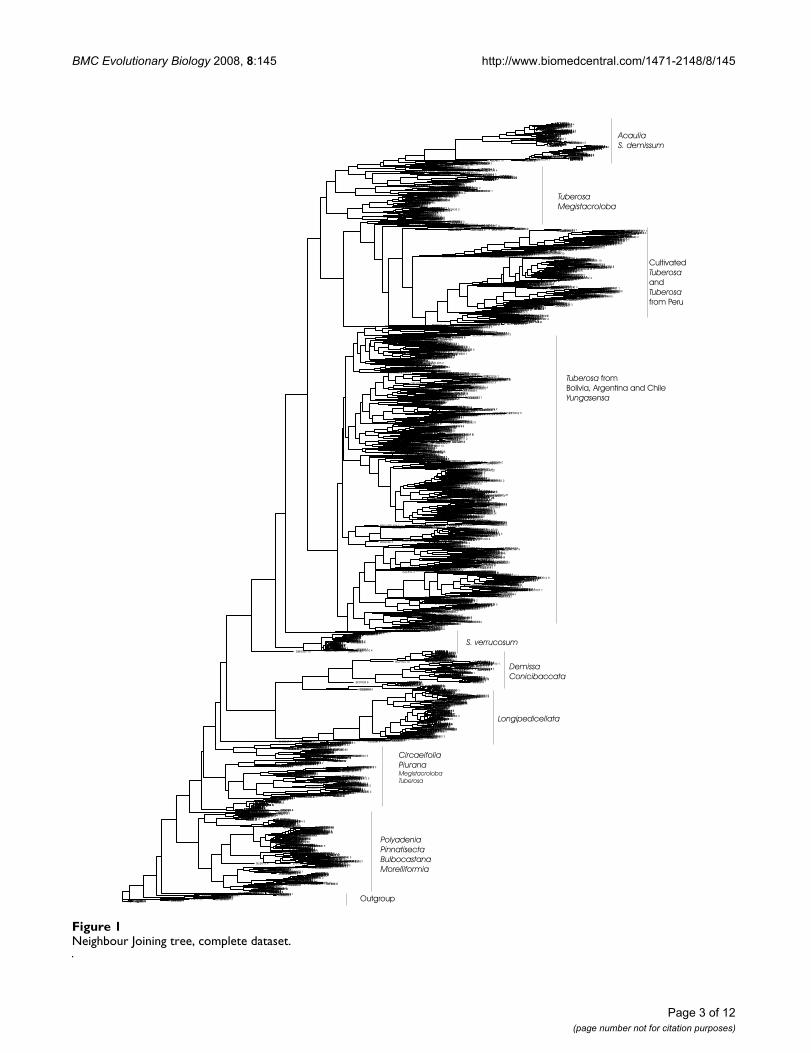
ResultsThe large dataset (4929 genotypes)Figure 1 shows the Neighbor Joining (NJ) tree of the 4929genotypes dataset. To describe the structure found in thisNJ tree, we differentiate between 3 levels of structure: theaccession level, the species level and the interspecies level.
At the accession level, the genotypes of the majority of theaccessions cluster together. Of those accessions that donot form complete clusters, in most cases only one geno-type deviates from the other 4 genotypes. In other cases,the accession was apparently so closely related with one ormore other accessions that their genotypes formed amixed group.
At the species level, 58 species or subspecies show consist-ency in their clustering, e.g. all accessions of a species clus-ter together. Nevertheless there are also many species (38in total) whose accessions did not cluster all together and48 species whose accessions were mixed with accessionsof other species. The latter was often the case with speciesthat occur in South America, the borders of many of thesespecies are not clearly recognizable from the NJ tree.
Above the species level, a few clusters of species groupscan be distinguished in the large NJ tree (but there is noindication on the statistical strength of the structureobserved). Roughly, the following groups can be found inthe NJ tree of the large dataset: 1) an outgroup with S.nigrum, S. chaparense, S. sitiens, and S. fraxinifolium 2)North and Central American diploid series Polyadenia,Pinnatisecta, Bulbocastana and Morelliformia, 3) Circaeifoliaand Piurana accessions, 4) Longipedicellata accessions, 5)Demissa and Conicibaccata accessions but without S. demis-sum and S. semidemissum, 6) S. verrucosum accessions, 7)
Page 2 of 12(page number not for citation purposes)
BMC Evolutionary Biology 2008, 8:145 http://www.biomedcentral.com/1471-2148/8/145
Page 3 of 12(page number not for citation purposes)
Neighbour Joining tree, complete datasetFigure 1Neighbour Joining tree, complete dataset.
ACL317 1ACL317 3ACL317 2ACL318 5ACL317 6ACL424 3ACL424 5ACL424 4ACL318 2ACL318 4ACL318 6PNE442 1PNE442 2PNE442 4PNE442 5PNE442 3BUK513 1BUK513 3BUK513 5BUK513 4BUK513 2ACL424 1ACL424 2ACL256 1ACL256 2ACL256 3ACL431 1ACL431 2ACL431 4ACL431 3ACL256 4ACL256 5ACL431 5ACL432 2ACL432 1ACL432 3ACL432 5ACL432 4ACL425 1ACL425 13ACL425 12ACL425 11ACL425 2ACL425 3 LPH681 1LPH681 2LPH681 3LPH681 4TAR861 1TAR861 2TAR861 5TAR861 3TAR861 4PNE443 1PNE443 2PNE441 1PNE441 2PNE441 3PNE441 5PNE441 4GRL604 5ACL426 1ACL426 2ACL426 3ACL426 4ACL426 5ACL427 1ACL427 2ACL427 3ALB301 1ALB301 2ALB301 3ALB301 4ALB301 5ACL430 6ACL430 7ACL430 9ACL430 1ACL430 2ACL430 3ACL430 4ACL430 5AEM436 1AEM436 2AEM436 3AEM436 5AEM436 4AEM435 1AEM435 5AEM435 3AEM435 4AEM435 2AEM434 1AEM434 2AEM434 3AEM434 5AEM434 4IOP669 5 IOP669 1IOP669 2IOP669 3IOP669 4AEM437 1AEM437 2AEM437 3 ALB461 1ALB461 2ALB461 4ALB461 3ALB462 1ALB462 5ALB462 2ALB462 3ALB462 4ALB464 1ALB464 3ALB464 4ALB464 2ALB464 5 JUZ173 1JUZ173 2JUZ173 4JUZ173 5JUZ173 3JUZ174 1JUZ174 2JUZ174 5JUZ175 1JUZ175 2JUZ175 4JUZ174 4JUZ174 3JUZ177 2JUZ175 3JUZ176 1JUZ176 3JUZ177 5JUZ177 1JUZ175 5JUZ176 2JUZ176 5JUZ177 4JUZ177 3JUZ176 4 DMS345 1DMS345 6DMS345 14DMS343 12MTP364 6DMS998 3DMS586 1DMS344 2DMS344 18CMM264 1CMM264 2CMM264 4CMM264 3CMM264 5DMS342 2DMS342 5DMS342 4BUK510 4BUK510 5BUK510 3DMS998 1DMS998 5DMS998 4DMS998 2DMS343 1DMS343 21DMS582 1DMS582 4DMS582 5DMS582 2DMS582 3DMS585 1DMS585 2DMS585 3DMS585 5DMS585 4DMS584 5DMS584 4DMS586 2DMS586 3SEM295 1SEM295 3SEM295 2UGT891 1BLG48 2DMS344 10DMS344 11DMS344 12DMS344 14MTP364 1MTP364 7MTP364 9MTP364 8MTP364 10MTP364 2MTP364 4MTP364 5DMS587 1DMS587 3DMS587 5DMS587 2DMS587 4TUQ299 1TUQ299 2BUK510 1BUK510 2AST114 5OPL748 2TUQ299 4TUQ299 5 EDN151 1EDN151 3EDN151 2EDN151 4EDN150 1EDN150 4EDN150 5EDN152 3EDN152 1EDN152 4EDN152 2EDN152 5EDN150 3EDN151 5EDN150 2TUQ299 3OPL748 1DMS584 1DMS584 2DMS584 3ACL430 8ACL430 10MGA699 1MGA696 1MGA696 2TOR702 1TOR702 5TOR702 2TOR702 4TOR702 3SCT805 1SCT805 2 SCT804 1SCT804 5SCT804 3SCT807 2SCT806 1SCT806 5SCT806 3SCT806 4SCT806 2SCT804 4SCT804 2SCT807 1SCT807 3SCT807 4SCT807 5MGA700 1MGA700 2TOR706 1TOR706 2TOR706 3TOR706 4MGA697 1MGA697 3MGA697 4MGA697 2MGA697 5TOR701 1TOR701 2TOR701 3TOR701 4TOR701 5TOR703 6TOR703 2TOR703 7TOR703 8 TOR703 9TOR703 10TOR703 3TOR703 4TOR703 1TOR278 1TOR278 2TOR278 3TOR705 6TOR705 7TOR705 8TOR705 3 TOR704 1TOR704 2TOR704 3TOR704 4TOR705 1TOR705 5TOR705 2TOR705 4TOR705 9TOR705 10SCT803 1SCT803 4SCT803 2SCT803 3SCT1061 AR50SCT803 5 SCT805 3SCT805 4 AJH47 1MLT729 1MLT729 3MLT729 4MLT729 5MLT725 1 MLT725 2MLT725 3MLT724 1MLT729 2ACH99 1ACH99 3ACH99 5ACH99 2ACH99 4 VIO394 1VIO394 3VIO394 4VIO394 10VIO394 2VIO394 5VIO394 6VIO394 9VIO394 7 VIO925 1VIO925 5VIO925 3VIO925 2VIO925 4VIO925 26VIO925 22VIO925 21VIO925 23VIO925 24VIO925 25VIOxYUN421 1VIOxYUN421 5VIOxYUN421 4VIOxYUN420 3VIOxYUN421 3VIOxYUN421 2VIOxYUN420 1VIOxYUN420 2VIO924 1VIO924 2VIO924 5VIO924 3VIO924 4VIO926 1VIO926 2VIO926 4VIO926 5VIO926 3LXS679 1LXS679 4LXS679 3LXS679 5LXS679 2SAN36 1SAN36 2SAN36 3SAN36 4SAN36 5SAN809 1SAN809 2SAN809 5SAN809 3SAN809 4LXS178 1LXS178 4LXS178 3LXS178 5LXS178 2 BUE7 1BUE7 2BUE7 4BUE7 5BUE7 3BUE8 1BUE8 4BUE8 3BUE8 5BUE8 2LGL685 1LGL685 3LGL685 5LGL685 2LGL685 4LGL179 1WBR253 1 WBR253 3WBR253 2WBR253 4 MED693 1MED693 5MED693 2MED692 1MED692 2MED692 4MED692 3MED695 1MED695 2MED695 5MED695 3MED695 4MED694 2MED694 4MED694 1MED691 1MED691 5MED691 2MED691 4MED691 3MED694 3 MED183 1MED183 2MED183 5MED183 4MED183 3MCQ186 1MCH185 5MED694 5SND93 1 SND808 1SND808 2SND808 3SND808 4SND808 5WBR254 1WBR254 2WBR300 1WBR300 2WBR300 3SND93 2SND93 3SND93 4 SND94 2SND94 1SND94 3SND94 5SND94 4 HCB170 2 HCB170 3HCB170 5HCB170 1HCB170 4HCB170 21NRS365 1 OKA368 1OKA368 9VNT896 1VNT896 4VNT896 3VNT896 5VNT896 2OKA740 9OKA740 1OKA740 2OKA740 6OKA740 7OKA740 8OKA740 10NRS365 3NRS365 6NRS365 2NRS365 7NRS365 4NRS365 9OKA368 2OKA368 3OKA368 6OKA368 8 NRS365 10NRS365 8NRS365 5NRS365 AR34HJT350 AR36NRS365 AR33OKA741 6OKA741 7OKA741 AR4OKA741 8OKA741 1OKA741 2OKA741 10OKA741 9OKA367 1OKA366 AR5OKA367 6OKA367 7OKA283 1OKA283 2OKA367 10OKA367 8OKA366 10OKA366 2OKA366 9OKA366 8OKA366 1OKA366 7OKA367 3OKA367 4OKA367 5VNT250 1VNT250 2VNT250 3VNT250 5VNT250 4VRN982 5OKA969 5OKA969 2OKA969 3OKA969 1OKA969 4VNT993 1VNT993 3VNT993 4VNT993 5VNT894 3VNT894 1VNT894 2VNT894 4VNT894 5VNT993 2 KTZ675 1KTZ675 3KTZ675 2KTZ675 4KTZ675 5 KTZ677 1KTZ677 2KTZ995 3KTZ995 5KTZ995 1 RCH35 2KTZ678 1KTZ678 4KTZ678 3KTZ678 2KTZ676 1KTZ676 3KTZ676 4KTZ676 5KTZ676 2KTZ678 5KTZ276 1KTZ276 2KTZ276 3KTZ276 5KTZ276 4RCH35 1RCH35 3RCH35 5RCH35 4KTZ275 1KTZ275 4KTZ275 5KTZ275 2KTZ275 3CMM577 1CMM577 2CMM576 1CMM1018 2CMM576 3CMM576 4CMM576 5CMM576 2CMM575 1CMM575 2CMM575 3CMM1017 1CMM1017 2CMM1018 1CMM1050 1CMM1050 2CMM1027 1CMM1039 2CMM1028 1CMM1028 2CMM1027 2CMM1039 1CMM1019 2CMM1019 1 CMM578 1CMM578 2MLM139 1MLM1038 1MLM1058 2MLM581 1MLM581 3MLM581 4MLM581 2MLM581 5MLM580 1MLM580 5MLM580 3MLM580 2MLM580 4MLM1045 1MLM1045 2MLM1038 2CMM265 1CMM265 3CMM265 2CMM265 4CMM265 5MLM266 2MLM266 1MLM1058 1HMP171 5 LPH684 1 LPH684 2LPH684 3LPH684 4LPH684 5VRG931 1VRG931 3VRG931 2VRG931 4VRG931 5CND531 2CND531 3MLT730 1MLT730 2MLT730 4MLT730 5 MLT731 1MLT731 4MLT731 22MTP190 5 SPL384 2SPL384 4SPL384 9SPL384 5ADG887 2 ADG887 6ADG887 8ADG887 4ADG887 10ADG887 5ADG887 1ADG887 9ADG887 7SPL384 3SPL384 6SPL384 8 ADGxSCR398 2ADGxSCR398 4ADGxSCR398 5MTP190 24 MGA184 3MRL187 1MTP190 1MTP190 2 MCQ186 2MCQ186 3MCQ186 4MCQ186 5MRL187 2MSP188 3MSP188 4MSP188 5MTP190 4MTP190 3AMB104 2AMB104 3 MGA184 1MGA184 2MGA184 4MGA184 5 ADGxCUR397 1ADGxCUR397 5ADGxCUR397 3ADGxCUR397 4ADGxCUR397 2TBR234 2TBR234 4TBR234 5TBR234 3TBR233 1TBR233 4TBR233 5TBR233 3TBR233 2ADG885 1ADG885 2ADG885 3ADG885 4ADG885 5 ADG241 4ADG242 5ADG242 3ADG242 1ADG242 4ADG242 2CND533 3CHA128 1CHA128 3CHA128 2CHA128 4 TBR232 1TBR232 2TBR232 4TBR232 5TBR232 3ADG238 4 ADG243 1ADG243 5ADG243 2ADG243 4ADG243 3ADG238 3ADG238 1ADG238 2ADG238 5AJH454 1AJH454 2AJH454 4AJH454 3AJH454 5ADG241 3 ADG241 1ADG241 2ADG241 5ADG235 1ADG235 4ADG235 3ADG235 2ADGxCUR395 2ADGxCUR395 4ADGxCUR395 5SCR847 1SCR847 2SCR847 4SCR847 5SCR847 3 CUR267 1CUR267 5CUR267 3CUR267 4CUR267 2CND533 1STN829 9STN829 7STN829 8CND533 2CND533 4CND533 5 ADG240 1ADG240 3ADG240 5ADG240 2ADG240 4VRG930 1VRG930 4VRG930 5VRG930 3VRG930 2 CUR142 1CUR142 4CUR142 2CUR142 3CUR142 5ADGxCUR395 1ADGxCUR395 3STN388 4STN388 8STN388 7STN829 6STN829 1STN829 3STN829 5STN829 4STN829 2AJH453 2 TBR883 1TBR883 2TBR883 4TBR883 3TBR883 5ADG889 1ADG889 3ADG889 4ADG889 2ADG235 5 ADG237 1ADG237 3ADG237 5ADG237 2ADG237 4SH 1SH 2SH 3SH 4SH 5TBR884 1TBR884 4TBR884 5TBR884 2TBR884 3ADG889 5 ADG239 1ADG239 4ADG239 2ADG239 3ADG239 5ADGxCUR396 1ADGxCUR396 4ADGxCUR396 5ADGxCUR396 22ADGxCUR396 25ADGxCUR396 21ADGxCUR396 24ADGxCUR396 2ADGxCUR396 3ADGxCUR396 23CUR141 1CUR141 4CUR141 3CUR141 5STN388 2ADG887 3CUR143 1CUR143 2CUR143 3CUR143 4CUR143 5WBR933 1WBR933 2WBR933 5WBR933 3WBR933 4CAN952 3SOU815 1SOU815 2GRL1052 2 SCR391 4ADG888 1ADG888 5ADG888 2ADG888 4ADG888 3AJH453 3AJH453 5AJH453 4 TBR234 1GON833 1GON833 4GON833 3 PHU371 1PHU371 6PHU372 5PHU372 6PHU371 7PHU371 5PHU372 3PHU372 9 PHU771 9PHU771 4PHU771 10PHU771 7PHU371 3PHU200 2PHU289 2PHU289 4PHU289 3PHU289 5CHA131 4CHA131 5PHU371 8PHU201 2PHU201 3PHU371 9PHU771 8 PHU774 1PHU774 4PHU774 2PHU774 5PHU201 1PHU773 1PHU200 4PHU773 3PHU773 4PHU773 5PHU200 3CHA131 2CHA131 3CUR140 1 CUR140 2CUR140 3CUR140 5CUR140 4GON220 1GON220 4GON220 2GON220 3GON220 5 PHU372 2PHU200 1PHU201 4PHU201 5PHU203 4PHU203 5PHU203 1PHU203 3SUB222 5SUB222 1 STN219 3STN219 5STN218 2STN219 1STN219 4STN219 2PHU372 10PHU771 3PHU771 5PHU771 6PHU372 4PHU372 7 SUB222 3PHU200 5PHU373 1CAN528 1CAN528 2PHU373 2PHU772 4PHU373 3CAN528 5CAN528 3CAN528 4PHU772 1PHU772 2PHU772 3PHU772 5PHU371 10PHU373 7PHU373 6PHU372 8PHU373 8PHU373 4PHU289 1PHU373 5PHU373 10PHU773 2STN387 5STN387 10STN387 4STN387 2PHU290 1PHU290 2PHU290 3PHU290 4PHU290 5GON834 1GON834 2STN387 6GON833 5GON831 6GON831 1GON831 4GON831 7GON831 2GON831 8GON831 3GON831 5CHA132 1 CHA132 2CHA132 4CHA132 5CHA132 3SUB222 2SUB222 4STN218 1STN218 5STN218 3STN218 4RH 1RH 3RH 2RH 4RH 5CHA130 4 CHA131 1CHA130 3PHU203 2CHA130 1CHA130 2CHA130 5PAM762 5STN387 1GON832 6GON832 9GON832 10GON831 9GON832 8GON832 7GON834 3GON834 4GON834 5VIO998 1VIO998 2PHU774 3GON831 10CHA129 1CHA129 5CHA129 3CHA129 4CHA129 2 SPLxSCR414 1SPLxSCR414 2SPLxSCR414 4SPLxSCR414 3SPLxSCR414 5MLT726 1MLT726 3MLT726 4MLT726 2AMB104 5 AMB104 1PAM762 1 PAM762 3PAM762 4PAM762 2 RAP292 1RAP292 4RAP292 3RAP292 2RAP292 5ABN423 1ABN423 2ABN423 4ABN423 3COP135 1COP135 2 MRN181 1MRN181 2MRN181 4MRN181 3MRN181 5LMB686 1LMB686 2CHI12 1CHI12 2CHI12 3CHI12 5COP306 1COP306 2COP306 5COP306 3COP306 4AMB104 4MRN182 1MRN182 3MRN182 2MRN182 4MRN182 5ORP29 1ORP29 2ORP29 3ORP29 5ORP29 4AML3 1AML3 5AML3 2AML3 3AML3 4COP572 1COP572 2COP572 4COP572 3COP572 5AMB467 1AMB467 3AMB467 2AMB467 5AMB467 4 ORP196 1ORP196 4ORP196 2ORP196 3ORP83 1ORP83 4ORP83 5ORP83 2ORP83 3 ORP84 1ORP84 5ORP84 4ORP84 3ORP84 2COP307 1COP307 5COP307 4COP307 2COP307 3VLR97 1VLR97 4VLR97 3VLR97 5VLR97 2BUK511 1BUK511 2BUK511 3BUK955 1BUK955 5BUK955 2BUK955 3BUK955 4AMB105 1AMB105 3AMB105 2AMB105 5 AMB105 4CND530 1CND530 4CND530 2CND530 3CND531 1CND531 5CND531 4CND532 1CND532 2CND532 3CAN951 3CAN951 4CAN951 1CAN526 2CAN526 3CAN526 4CAN952 1CAN952 4CAN952 5CAN952 2CAN526 5CAN526 1CHI12 4CAN260 1CAN260 2 CAN529 1CAN529 3CAN529 5CAN529 4CAN529 2COP134 1COP134 2MRN690 1MRN690 4MRN690 2MRN690 3MRN690 5MRN277 1MRN277 2MRN277 3MRN77 1MRN77 2CAN953 1CAN953 2CAN953 5CAN953 3CAN953 4 AYM5 1AYM5 5AYM5 2AYM5 4AYM5 3VLR893 1VLR893 3VLR893 5VLR893 2VLR893 4PAM288 1PAM288 5PAM288 2PAM288 4PAM288 3BUK971 1PAM764 4PAM764 1 BUK512 1BUK512 2BUK512 5BUK512 4BUK512 3BUK971 3BUK971 4BUK971 5BUK971 2PAM764 2PAM764 3PAM764 5PAM763 1PAM763 2PAM763 3PAM763 4PAM763 5CAN527 1CAN527 4CAN527 3CAN527 5CAN527 2BUK514 1BUK514 2BUK514 3 STN296 1STN296 4STN296 3BUK328 4BUK328 9MLT731 2MLT731 23MLT731 24MLT731 3MLT731 25MLT731 5MLT731 21MLT730 3MLT732 1MLT732 2MLT732 3 MLT363 10MLT363 6MLT363 9MLT363 5MLT363 4MLT363 2MLT363 3MLT722 1MLT722 3MLT722 5MLT722 2MLT722 4MLT727 1MLT727 3MLT727 4MLT727 5MLT727 2AMY302 1AMY302 5AMY302 4AMY302 2AMY302 3 AMY303 1AMY303 3AMY303 2AMY303 5AMY303 4MLT728 1 MLT728 3MLT728 2ORP756 1ORP756 3ORP756 2ORP756 4ORP756 5MLT723 1MLT723 2MLT723 4MLT723 5MLT723 3HRO309 1HRO309 3HRO309 5HRO309 4HRO309 2RAP796 5RAP796 2RAP796 4RAP796 1RAP796 3MTP190 21MTP190 22AJH453 1BUK329 4BUK329 AR16BUK329 AR15 MSP188 1KTZ995 2KTZ995 4PTR613 2 SCRxOPL412 1SCRxOPL412 3SCRxOPL412 2SCRxOPL412 5SCRxOPL412 4TND246 1 HMP171 1HMP171 2HMP171 3HMP171 4RCH210 2RCH210 4OPL749 2SCR391 1 SCR391 10SCR391 6SCR391 3SCR391 2 SCR844 8SCR844 2SCR844 5SCR844 3SCR844 7SCR844 1SCR844 4SCR850 1SCR850 3SCR850 2SCR850 4SCR850 5ADGxSCR400 3ADGxSCR400 4ADGxSCR400 1ADGxSCR400 2ADGxSCR400 5PTR617 1 PTR617 2PTR617 3PTR617 5PTR617 4SPL972 1SPL972 4SPL972 3SPL972 5SPL972 2GRR165 1 GRR165 2GRR165 4GRR165 3GRR165 5PLT789 5OPL1001 2ADGxSCR398 1ADGxSCR398 3 SCR845 7 SCR845 9SCR845 8SCR845 2SCR845 10SCR845 1HDM649 1HDM649 5 HDM649 3HDM649 4HDM649 2HDM352 8HDM352 1HDM352 10HDM352 2HDM352 3HDM352 6HDM352 4SCR849 7SCR849 9SCR849 1SCR849 3SCR849 4SCR849 2SCR849 5SCR849 AR53 SCR849 6SCR849 10SCR849 8LPH356 1LPH356 7LPH356 8LPH356 3LPH356 4LPH356 5LPH356 6 LPH683 6LPH683 7LPH683 8LPH683 1LPH683 2LPH683 9 TUQ43 1TUQ43 4TUQ43 2TUQ43 5LPH683 3SPL975 1RCHxMCD411 3BRCxSPL404 4 SPL820 2SPL820 3 SPLxLPH413 2SPLxLPH413 4SPLxLPH413 3SPL821 1SPL821 3SPL821 5SPL821 2SPL821 4SPL820 4SPL820 5BRC1020 1BRC1020 2 VRG928 1VRG928 3VRG928 5VRG928 2VRG928 4SPL820 1SPL973 1 SPL973 5SPL973 3SPL973 4SPL973 2VRG927 1VRG927 5VRG927 3VRG927 4VRG932 2VRG927 2VRG932 1VRG932 4SPL975 2SPL975 4SPL975 5SPL975 3 SPL816 1SPL816 2SPL816 4SPL816 5SPL816 3RAP381 2RAP381 6RAP381 8RAP381 9RAP381 4ARP110 1ARP110 4ARP110 5ARP110 2ARP110 3VRG932 5ADG886 1GRL1000 2LPHxSPL405 1LPHxSPL405 3SPLxLPH413 1LPH680 11LPH680 1LPH680 21LPH680 41LPH680 4LPH680 51LPH680 5LPH680 31LPH680 3 SPL382 1SPL382 10SPL819 2SPL819 5SPL382 2SPL382 8 SPL382 4SPL382 7SPL382 6SPL382 5SPL383 1SPL383 2SPL383 5SPL383 3SPL383 8 SPL383 10SPL383 6SPL383 4SPL818 6SPL818 1SPL818 4SPL818 5SPL818 10SPL818 3SPL818 7SPL818 8SPL818 9SPL818 2SPL819 1SPL819 4SPL819 3LPH357 10LPH357 5LPH357 9LPH357 6spp416 1SPL978 4SPL978 1SPL978 2SPL978 3SPL978 5spp416 3spp416 2GON830 2GON830 1GON830 3GON830 5GON833 2GON830 4ADG886 2ADG886 3ADG886 4ADG886 5BRC1026 1BRC1026 2LPH682 1LPH682 3LPH682 2LPH682 4 LPHxSPL405 2LPH682 5LPHxSPL405 4ALN455 1 ALN455 2ALN455 3ALN455 4ALN455 5SCR843 1SCR843 4SCR843 2SCR843 5BRC327 10BRC327 8BRC327 5BRC327 7BRC327 2BRC327 4BRC327 9 BRC505 1BRC505 4BRC505 3BRC505 2BRC507 1BRC507 4BRC507 3BRC507 5BRC505 5BRC506 1BRC506 2BRC506 3BRC506 5BRC506 4BRC1047 2BRC1025 1BRC1025 2VRG929 1VRG929 4VRG929 5VRG929 2VRG932 3VRG929 3 BRC509 5BRC1040 1BRC1040 2BRC507 2 BRC509 1BRC509 3BRC509 2BRC509 4SPG828 1SPG828 4SPG828 5SPG828 2OPL1041 1OPL1041 2OPL1057 1OPL1057 2 SCR846 7 GRL1008 1GRL1008 2GRL1033 1GRL1033 2GRL1009 1GRL1009 2 GRL1029 1GRL1029 2GRL1034 1GRL1034 2GRL1032 2VRN904 2GRL1032 1GRL1021 1SPG822 AR51GRL610 2GRL1044 1GRL1044 2GRL1005 1GRL1030 2GRL1005 2GRL1006 1GRL1006 2 GRL1014 1GRL1014 2GRL1013 1GRL1049 1GRL1049 2INM662 1INM662 3INM662 5INM662 2INM662 4INM1016 1INM1016 2GRL1030 1 HAW634 2HAW634 5HAW635 1HAW635 4HAW635 2HAW636 2HAW637 1HAW637 2HAW637 5HAW637 3HAW637 4HAW636 1HAW636 4HAW166 2HAW166 4HAW166 1HAW167 2HAW167 4HAW167 5HAW167 3HAW167 1HAW635 5HAW166 5 HAW634 3HAW636 5HAW638 1HAW634 4HAW635 3HAW638 3HAW638 2HAW638 4HAW636 3HAW638 5HAW166 3GRL605 AR52 OPL753 1OPL753 4OPL753 3OPL753 5OPL753 2IFD1007 1IFD1007 2IFD1023 1IFD1023 2IFD664 1IFD664 3IFD664 2IFD664 4IFD664 5 IFD667 2IFD665 3IFD665 1IFD665 2IFD665 4IFD665 5IFD667 1IFD667 5IFD667 4IFD667 3IFD666 2IFD666 3IFD666 4IFD666 5IFD666 1VID623 2VID623 3IFD668 1IFD668 2IFD668 3IFD668 4GRL1051 1GRL1051 2VID623 5 OPL754 1OPL754 5OPL754 3OPL754 4OPL754 2OPL1059 1OPL1059 2OPL1046 1OPL1046 2GRL1012 1GRL1012 2OPL1002 1OPL1004 2OPL1004 1OPL1024 1OPL1024 2OPL1002 2OPL1003 2OPL1031 1OPL1031 2OPL750 1OPL750 2OPL750 4OPL750 5OPL750 3OPL747 1OPL747 2OPL747 4OPL747 3OPL747 5GRL1054 1GRL1054 2GRL1010 1GRL1010 2 GRL1011 1GRL1011 2GRL1035 1GRL1035 2GRL604 1GRL604 4GRL604 3GRL604 2GRL1022 1GRL1022 2OPL1001 1GRL1021 2GRL1015 1GRL1015 2GRL1013 2GRL1036 2GRL1036 1 GRL347 2GRL347 7GRL347 9VID621 3VID621 1VID621 2VID621 4VID626 1VID626 2VID626 3VID626 4VID626 5GRL1043 1GRL1042 1 VID624 4VID624 5VID625 1VID625 5VID625 3VID625 4VID625 2GRL1042 2GRL1043 2 OPL752 1OPL752 2OPL752 3OPL751 1OPL749 1OPL749 3OPL749 4OPL749 5NRS736 1NRS736 2NRS736 5NRS736 3NRS736 4NRS737 4NRS737 1NRS737 3NRS737 5NRS737 2NRS735 1NRS735 4NRS735 2NRS735 5NRS735 3NRS281 1NRS281 2NRS281 3NRS281 4NRS281 5 VRN987 1VRN987 5VRN987 3VRN987 4VRN987 2 PTR613 1PTR613 5PTR613 3PTR613 4DDS588 1DDS588 2DDS588 4DDS588 5DDS588 3GRL608 1GRL608 2GRL608 3GRL608 4 GRL608 5GRL606 2GRL606 3GRL606 1 GRL609 2GRL609 5GRL609 3GRL609 4GRL610 4GRL610 5GRL610 3GRL610 1GRL611 1GRL611 2GRL611 4GRL611 3GRL611 5 GRL605 1GRL605 5GRL605 3GRL605 2GRL609 1GRL605 4HPS650 1HPS650 2HPS650 3HPS650 4HPS650 5 HPS652 1HPS652 5HPS652 2HPS652 3HPS652 4HPS653 1HPS653 4HPS653 3HPS653 5HPS653 2 INM659 1INM659 4INM660 1INM660 5INM660 4INM660 2INM660 3INM658 5INM659 5INM659 2INM659 3INM663 3INM663 4INM663 5INM657 5INM661 1INM661 3INM661 2INM663 2INM657 1INM657 4INM661 5INM657 2INM657 3INM661 4INM663 1 INM658 1INM658 3INM658 4INM658 2 PTR616 1PTR616 5PTR616 4PTR616 2PTR616 3 VID619 1VID619 2VID619 5VID619 3VID619 4 VID620 1VID620 4VID620 2VID620 5VID620 3VID622 1VID622 2 VID622 5VID622 4VID622 3VID624 1VID624 2VID624 3GRL1053 1GRL1053 2PTR615 5GRL1037 1AST472 1AST472 2AST472 21AST472 5AST472 24AST472 4AST472 3AST472 23BER494 1AST472 22AST114 1AST114 3AST114 4AST114 2AST475 5AST475 23AST475 22 AST474 1AST474 14AST474 4AST474 11AST474 12AST474 2AST474 15AST474 13AST474 5AST474 3AST475 1AST475 4AST475 3AST475 2AST475 21AST475 24BLV496 1BLV496 2BLV496 3BLV496 5BLV496 4AST476 4AST476 5AST476 1AST476 2AST476 3BLV499 1BLV499 2BLV499 3BLV499 4 BLV498 1BLV498 5BLV498 2BLV498 3BLV498 4AST472 25 UGT891 6UGT891 10UGT891 8UGT891 9UGT891 7UGT891 2UGT891 5UGT891 4UGT891 3GRL1000 1OPL1056 1OPL1056 2 HDM351 2HDM351 3HDM351 6HDM351 8HDM351 4HDM648 4HDM648 1HDM648 3HDM648 5HDM648 2 HDM645 1HDM645 2HDM645 5HDM645 4HDM645 3HDM168 1HDM168 21HDM168 4HDM168 22HDM168 25HDM168 23HDM168 2HDM168 3HDM168 5HDM168 24HDM646 1HDM646 2HDM646 3HDM646 4 HDM646 5 HDM647 1HDM647 5HDM647 2HDM647 3HDM647 4HPS169 1HPS169 2HPS169 3HPS169 5HPS169 4UGT248 1UGT248 2UGT248 4UGT248 5UGT248 3 UGT249 1 UGT249 2UGT249 3UGT249 4UGT249 5UGT892 1UGT892 4UGT892 2UGT892 3UGT892 5UGT44 1UGT44 4UGT44 2UGT44 3 PTR614 1PTR614 3PTR614 4PTR614 5PTR614 2PTR615 3PTR618 1PTR618 2PTR618 3PTR618 4PTR618 5GRL1048 1GRL1048 2PTRxLPH409 11PTRxLPH409 12PTRxLPH409 15PTRxLPH409 13PTRxLPH409 14PTR615 1PTR615 2PTR612 3PTR612 1PTR612 5PTR612 4PTR612 2PTR615 4PTRxLPH409 1PTRxLPH409 3PTRxLPH409 4PTRxLPH409 5PTRxLPH409 2SPL817 1SPL817 2SPL817 3SPL817 5SPL817 4BERxSPL402 3BRC1047 1VID623 1VID623 4OKA742 2 BRCxSPL403 1BRCxSPL404 3BRCxSPL403 2BRCxSPL403 4BRCxSPL403 5BRCxSPL403 3BRCxSPL404 5BRCxSPL404 2 AVL477 1AVL478 1AVL478 3AVL478 23AVL477 2AVL478 2AVL478 4AVL478 21AVL478 22AVL478 24AVL477 3AVL477 4AVL477 5AVL478 5AVL478 25 AVL479 1AVL479 3AVL479 4AVL479 2AVL479 5SCR848 1SCR848 4SCR848 2SCR848 3SCR848 5 ARP109 1ARP109 4ARP109 5ARP109 2ARP109 3SCR843 3 CJM122 1CND123 1CND123 5CND123 2CND123 3CND123 4ALN460 1 ALN460 2ALN460 5ALN460 3ALN460 4DDS144 2DDS144 1DDS144 5DDS144 3DDS144 4ALN320 10ALN320 7ALN320 5ALN320 2ALN320 4ALN320 9ALN320 1ALN320 6ALN320 8ALN257 1ALN257 4ALN257 5ALN257 2ALN257 3 DDS589 1DDS589 4DDS589 3DDS589 2DDS589 5ALN459 1ALN459 3ALN459 4ALN459 5ALN459 2 GND346 1GND346 9 GND346 2GND346 7GND346 8GND346 6GND346 10GND346 5GND346 4GND62 1GND62 2GND62 4GND62 5GND62 3GND16 1 GND16 2GND16 5GND16 3GND16 4GND270 1GND270 4GND270 5GND270 2GND270 3GND603 1GND603 2GND603 3GND603 4GND603 5ALN458 1ALN458 3ALN458 2ALN458 5ALN458 4ALN457 1ALN457 5ALN457 4ALN457 2ALN457 3GND163 1GND163 2GND163 5GND163 3GND163 4 BER324 2 CHM561 4CHM561 2CHM561 3BER324 AR21CHM561 1BER486 1BER486 4BER493 AR31BER494 2BER494 3BER494 5BER493 3BER494 4BER493 2BER493 6BER493 7BER493 1BER493 9BER487 9BER488 1BER488 4BER488 5BER488 2BER488 3BER486 2BER486 3BER487 6BER487 7BER487 8BER487 4BER487 5BER487 2BER487 3BERxSPL402 1BERxSPL402 4BERxSPL402 2BER490 1BER490 2BER490 3BER490 5BER490 4BER487 1BER493 5BER493 8BER493 AR32BER940 3BER940 2BER940 1BER489 1BER493 4BER941 1BER941 5BER941 4BER941 2BER941 3BER480 4TAR858 2BER480 1BER480 2BER480 5BER480 3 TAR857 2TAR857 1TAR857 3TAR857 5TAR857 4BER483 1BER483 4BER483 5BER483 2BER483 3 BER943 1BER943 2BER943 3BER943 4BER943 5BER322 3BER322 6BER322 7BER486 5BER322 5BER322 8BER939 1BER939 2BER939 4BER939 3BER481 1BER481 4BER481 2BER481 3 TAR858 1TAR858 4 TAR858 5TAR858 3BER482 1BER482 4BER482 2BER482 3BER482 5BER939 5TAR852 1TAR879 1TAR879 3TAR879 5TAR879 4TAR879 2BER323 1BER323 7BER323 2BER323 6BER323 10BER323 5BER323 9TAR864 5TAR864 2TAR867 3TAR854 3TAR852 4TAR874 4TAR852 2TAR852 3TAR852 5TAR225 1TAR225 4TAR225 3TAR225 5TAR225 2 BER485 1BER485 2BER485 5BER485 3BER485 4BER484 1BER484 2BER484 5BER484 3BER484 4BER944 1 CHC544 10CHC544 1CHC544 15CHC544 5CHC544 2CHC543 1CHC543 2CHC543 5CHC543 3CHC543 4DDS145 1DDS145 5DDS145 3DDS145 2DDS145 4DDS146 1DDS146 3DDS146 4DDS146 5DDS146 21DDS146 2DDS146 24DDS146 25DDS146 23DDS146 22ARZxHDM401 1ARZxHDM401 3 ARZxHDM401 21ARZxHDM401 2ARZxHDM401 4ARZxHDM401 22ARZxHDM401 24BER944 2BER944 4BER944 3BER944 5BER492 1BER492 5BER492 2BER492 3BER492 4ARZxHDM401 AR30ARZxHDM401 23ARZxHDM401 25CHC544 6CHC544 8CHC544 11CHC544 14CHC544 7CHC544 12CHC544 13CHC544 9 CHC338 1CHC338 3CHC338 8CHC338 10CHC338 2CHC338 5CHC338 9CHC338 6CHC338 7YUN255 1YUN255 2YUN255 4YUN255 5YUN255 3MLM579 1 CHC126 3CHC126 4HCB170 24MLM579 4MLM579 5 TAR853 1TAR853 5TAR853 4TAR853 2TAR853 3 TAR870 3TAR870 4TAR875 2TAR876 1TAR870 1TAR878 1 TAR871 1TAR871 4TAR871 2TAR871 5TAR878 2TAR878 3TAR870 5TAR870 2TAR877 2TAR874 1TAR874 3TAR877 5TAR877 4TAR877 1TAR877 3TAR876 2TAR876 3TAR876 5TAR224 1TAR224 3TAR224 5TAR224 2TAR224 4TAR859 1TAR859 3TAR859 4TAR859 5TAR859 2TAR865 1 TAR865 2TAR865 5TAR865 3TAR865 4TAR860 1TAR860 3TAR860 4TAR860 2TAR860 5TAR863 4TAR863 2 TAR863 3TAR863 5BER491 1BER491 2BER491 3BER491 4BER491 5TAR863 1TAR873 1TAR873 2TAR873 4TAR873 3TAR873 5 TAR864 4TAR864 25TAR864 21TAR864 24TAR864 23TAR864 1TAR864 3TAR875 5TAR864 22TAR862 5TAR862 2TAR862 3TAR862 1TAR862 4TAR872 4TAR872 5TAR872 2TAR871 3TAR878 4TAR878 5TAR872 3 TAR855 6TAR855 9TAR855 3TAR855 1 TAR855 8TAR854 5TAR855 10TAR855 4TAR855 5TAR855 2TAR854 1TAR854 2TAR854 4TAR855 7TAR856 1TAR875 1TAR875 4TAR875 3TAR874 2TAR874 5TAR876 4TAR867 1TAR867 2TAR867 4TAR867 5TAR866 1 TAR866 2TAR866 3TAR866 5TAR866 4 NCD280 1NCD280 3NCD280 4NCD280 5NCD280 2TAR869 1TAR868 5TAR392 2TAR392 5TAR392 3TAR392 6TAR392 8TAR868 10TAR868 7TAR868 9TAR869 4TAR868 4TAR868 1TAR868 8TAR868 AR61TAR869 2TAR869 3TAR868 6TAR868 AR62TAR868 2TAR868 3TAR869 5TAR856 2TAR856 4TAR856 3TAR856 5BRCxSPL404 1 TAR392 1SCR846 2SCR846 1TAR392 10SCR846 5SCR846 9SCR846 3SCR846 8SCR846 10SCR846 4SCR846 6 HPS651 1HPS651 2HPS651 4HPS651 3HPS651 5 MED693 3RAP800 1RAP800 4RAP800 5RAP800 3RAP800 2 RAP799 1RAP799 3RAP799 2RAP799 AR18RAP799 AR19RAP799 5RAP380 2RAP380 8 RAP792 2RAP792 4RAP792 3RAP792 5RAP790 1RAP790 2RAP790 5RAP790 3RAP790 4 RAP797 1RAP797 4RAP797 2RAP797 3RAP209 1RAP209 5RAP209 3RAP209 2RAP209 4RAP976 1RAP976 2RAP791 1RAP792 1 RAP791 2RAP791 5RAP791 3RAP791 4RAP801 3RAP801 1RAP801 2RAP801 5RAP801 4RAP798 1RAP798 5RAP798 2RAP798 4RAPxSPL410 4RAPxSPL410 3RAPxSPL410 2RAPxSPL410 1RAPxSPL410 5RAP291 1RAP291 2RAP798 3RAP794 1RAP794 4RAP794 3RAP794 5RAP794 2RAP208 1RAP208 2RAP208 5RAP208 3RAP208 4RAP793 5 RAP793 1RAP793 3RAP793 4RAP793 2 SPL974 1SPL974 3SPL974 4SPL974 2SPL974 5MCQ717 1 COL137 1COL137 5COL137 4COL137 2COL137 3VRN905 3 VRN905 7 VRN905 10OKA742 3 OKA742 1OKA742 4OKA744 1OKA744 5OKA745 1OKA745 3OKA745 2OKA745 5OKA745 4OKA744 4 OKA744 2OKA744 3OKA970 1OKA970 4OKA970 5OKA970 2OKA970 3 OKA743 1OKA743 3OKA743 4OKA743 5OKA743 2OKA739 3OKA739 4OKA739 5OKA739 1OKA739 2OKA746 1OKA746 3OKA746 5OKA746 4OKA746 2 MTP190 23MTP190 25MLM579 3 ADR470 1ADR470 2ADR470 3ADR470 5ADR470 4MLM579 2 CHC547 8CHC547 9CHC547 10CHC547 6CHC547 1CHC547 5CHC547 4CHC547 3CHC547 2CHC545 1CHC545 2CHC545 5CHQ51 1CHC545 4CHC551 1CHC551 3CHC551 2ARZ471 1ARZ471 5ARZ471 2ARZ471 3ARZ471 4 YUN98 1YUN936 4YUN935 1YUN936 3YUN936 2YUN935 3YUN936 1YUN936 5YUN935 4YUN935 2YUN935 5ARZ111 1ARZ111 5ARZ111 4ARZ111 2ARZ111 3ARZ113 2 ARZ4 11ARZ4 1ARZ4 5ARZ4 12ARZ4 13ARZ4 2ARZ4 4ARZ4 3TARxMCD419 1TARxMCD419 2TARxMCD419 5TARxMCD419 3TARxMCD419 4CHC548 1CHC548 24CHC549 2CHC548 2 CHC548 3CHC548 4CHC549 1CHC549 4CHC549 3CHC549 5CHC548 25CHC548 23CHC548 21CHC548 22GRL1055 1GRL1055 2OCR194 1OCR194 3OCR194 4OCR194 23OCR194 2OCR194 5OCR194 24 CHC546 4OCR194 21OCR194 25 ARZ113 1ARZ113 5ARZ113 3ARZ113 4CHC126 1CHC126 5CHC126 2 CHC125 1CHC125 2CHC125 5CHC125 3CHC125 4STL214 4TND246 2TND246 3TND246 5TND246 4 CHC127 1CHC127 2CHC127 3CHC127 4CHC127 5CHC550 1CHC550 4CHC550 2CHC550 5CHC550 3CHC546 1CHC546 5CHC546 2CHC546 3 YUN934 4YUN934 5YUN934 2YUN934 1YUN934 3 STL214 1STL214 2STL214 5STL811 5 STL811 1STL811 4STL811 2STL811 3STL214 3RZL802 5CHC338 4 OCR194 22 FLH601 1FLH601 5FLH601 2FLH601 3FLH601 4TARxARZ418 1TARxARZ418 3TARxARZ418 4TARxARZ418 2TARxARZ418 5 WBRxYUN422 1WBRxYUN422 5WBRxYUN422 2WBRxYUN422 3WBRxYUN422 4RZL802 1RZL802 4RZL802 2RZL802 3 MAG359 1MAG359 9MAG359 2MAG359 3MAG359 10MAG359 4MAG359 8 MAGxMCD406 2MAGxMCD406 4MAGxMCD406 3MAG75 1MAG76 1MAGxMCD406 5RCHxMCD411 1RCHxMCD411 5RCHxMCD411 2RCHxMCD411 4MAG688 1MAG688 3MAG688 2MAG688 4MAG688 5 MCD360 1MCD360 4MCD360 5MCD360 2MCD360 9MCD360 6MCD360 8MCD360 7GIG715 1GIG715 2GIG715 3GIG715 4GIG715 5GIG965 4 GIG712 6GIG712 2 GIG712 9GIG712 1GIG712 10GIG712 7GIG712 3 GIG957 1GIG957 3GIG957 5GIG957 2GIG712 4GIG712 5GIG712 8GIG957 4 GIG965 1GIG965 3GIG965 2GIG711 6GIG711 10GIG711 5GIG711 3GIG711 4GIG711 1GIG711 9GIG711 7GIG711 2GIG711 8GIG710 1GIG710 3GIG710 2GIG710 4GIG710 5GIG960 1GIG960 4GIG960 5GIG960 3GIG960 2MCD959 1MCD959 3MCD959 5MCD959 2MCD959 4GIG967 1GIG967 3GIG967 4GIG967 2GIG967 5GIG966 1GIG966 3GIG966 2GIG966 4GIG966 5GIG362 3GIG362 4GIG362 6GIG361 1 GIG361 10GIG361 3GIG361 9GIG361 2GIG361 5GIG361 8GIG361 4GIG361 7GIG361 6 MCD958 1MCD958 2MCD958 4MCD958 5MCD958 3GIG964 1GIG964 4GIG964 3GIG964 5GIG964 2 MCD708 6MCD708 5MCD708 1MCD708 3MCD708 7MCD708 8MCD708 9MCD708 10 MCD707 1MCD707 3MCD707 5MCD707 4MCD707 2MCD708 2GIG714 1GIG714 2GIG714 3GIG714 4GIG963 1 GIG963 5GIG963 2GIG963 4GIG963 3GIG962 1GIG962 5GIG962 2GIG962 3GIG962 4GIG714 5 MCD994 1MCD994 3MCD994 5GIG956 1GIG956 2GIG956 4GIG956 5GIG956 3GIG961 1GIG961 4GIG961 5GIG961 3GIG961 2MCD994 2MCD994 4 GIG713 1GIG713 2GIG713 3GIG713 4GIG713 5MCD360 10RCH210 1RCH210 3 RCH210 5MAGxMCD406 1RCH211 1RCH211 4RCH211 3RCH211 5RCH211 2VRN903 1VRN903 2VRN903 4VRN903 5VRN903 3 VRN905 6VRN905 9VRN904 6VRN904 9VRN904 10VRN904 8BAL906 2BAL906 4BAL906 1BAL906 3VRN985 4VRN904 7VRN985 1 VRN985 3VRN904 3VRN904 1VRN985 5VRN905 1VRN905 2VRN904 4BAL907 1 BAL907 4BAL907 5BAL907 2BAL907 3VRN985 2BAL908 1BAL908 2BAL908 4BAL908 5BAL908 3 HAN629 1HAN629 3HAN629 5HAN629 2HAN630 4HAN630 1HAN630 3HAN630 2HAN630 5HAN631 1HAN631 5HAN631 2HAN631 4HAN629 4HAN632 4HAN633 2HAN633 1HAN632 1HAN633 5HAN632 3HAN633 3HAN632 2HAN633 4HAN631 3HAN628 1HAN628 4HAN628 2HAN628 5HAN628 3HAN632 5VRG252 1VRG252 3VRG252 2VRG252 4VRG252 5VRN902 6VRN902 10VRN902 7VRN902 4VRN983 5VRN983 4VRN983 1VRN983 3VRN902 9VRN902 5VRN983 2VRN902 1VRN902 2VRN982 1VRN982 2VRN982 4VRN982 3CHC263 1CHC263 3CHC263 4CHC263 5SPG386 1SPG386 2SPG386 8SPG386 7SPG386 9SPG386 10SPG386 6SPG386 3SPG827 1SPG827 5SPG827 2SPG827 3SPG827 4 VNT895 6VNT895 10VNT895 8VNT895 7 VRN980 2VNT895 5VRN980 4VNT895 4VNT895 9VRN980 1VRN980 5VRN980 3 VRN901 6VRN901 7VRN901 10VRN901 8VRN898 10VRN898 6VRN898 7VRN979 1VRN979 3VRN979 2VRN901 4VRN901 5VRN984 1VRN984 2VRN984 4VRN984 3VRN984 5VRN901 9VRN897 1VRN897 4VRN897 2VRN897 3VRN897 5VRN900 4VRN900 2VRN900 21VRN898 2VRN901 3VRN901 1VRN900 22VRN900 24VRN900 25VRN904 5VRN901 2VRN900 5VRN898 8VRN900 1VRN900 3VRN898 9VRN900 23 VRN899 6VRN899 8VRN899 2VRN899 4VRN899 10VRN899 5VRN981 3VRN899 7VRN899 9VRN899 3VRN899 1VRN981 1VRN981 4VRN981 5VRN981 2VRN986 1VRN986 2VRN986 4VRN986 3GRL1037 2SPG824 3SPG824 4SPG824 5SPG824 1SPG824 2SPG823 1SPG823 4SPG823 3SPG823 5SPG823 2SPG822 1SPG822 2SPG822 3SPG822 5SPG822 4SPG385 6SPG385 7SPG385 9 SPG826 1SPG826 3SPG826 4SPG826 2SPG826 5SPG217 3SPG217 4SPG217 1SPG217 5SPG217 2SPG828 3 ARZ112 1ARZ112 2ARZ112 4 ARZ112 3ARZ112 5SCR844 6SCR845 6TAR872 1VRN902 8BAL906 5CHQ49 1TUQ43 3VER393 10VER393 4VER920 3VER920 4VER920 5VER920 1VER920 2VER920 8VER920 9VER920 10VER920 7VER912 1VER912 3VER912 5VER912 4VER912 2VER918 1VER918 3VER918 2VER918 5VER918 4VER921 1VER921 2VER921 5VER922 1VER922 3VER922 2VER922 4VER922 5VER921 3VER990 3VER921 4VER990 4VER990 5VER990 1VER990 2MCP23 5MCP23 4VER916 2VER916 5MCP23 2MCP23 3MCP23 1MCP74 3MCP74 5MCP74 1MCP74 2MCP74 4VER919 6VER919 7VER919 8VER917 1VER917 2VER917 3VER917 5VER917 4SPG825 1SPG825 2SPG825 4SPG825 3VER919 10VER919 2VER919 3VER919 4VER919 5VER916 3VER919 1VER923 3VER916 1VER916 4VER923 1VER923 4VER910 3VER910 5VER923 2VER923 5VER910 2VER916 AR17VER911 3VER911 4VER915 1VER915 2VER915 5VER915 4VER915 3VER909 3VER988 1VER988 4VER988 3VER988 2VER989 3VER988 5VER989 2VER989 4VER989 5VER911 1VER911 2VER911 5VER989 1VER909 4VER914 6VER914 1VER914 3VER914 4VER914 2VER914 5VER909 2VER909 5HDM644 1 VER909 1VER910 4VER910 1VER919 9VER914 8VER914 9 VER914 7VER914 10DMS585 10 ADR321 1ADR321 2ADR321 9ADR321 3ADR321 4ADR321 5ADR321 7ADR321 8ADR321 6BCP326 1BCP326 AR46BCP326 AR47BCP326 AR48IOP273 1BCP326 4IOP273 5BCP326 3IOP273 3IOP273 2BCP326 9BCP326 8BCP326 6BCP326 2IOP273 4BCP326 5BCP326 10BCP259 1BCP259 2BCP259 3BCP259 4BCP259 5IOP670 AR42IOP670 AR43IOP670 AR45IOP670 AR44IOP670 AR41BCP501 4BCP501 5BCP501 3BCP501 1BCP501 2GRR348 1GRR348 10GRR348 3GRR348 2GRR348 5GRR348 6GRR348 8GRR348 7GRR348 9GRR627 3GRR627 4GRR627 2GRR627 AR49GRR348 4GRR627 1 IOP670 1IOP670 2IOP670 4IOP670 5IOP670 3IOP671 1IOP671 5IOP671 3IOP671 4IOP671 2BCP504 10 COL136 1COL573 1COL573 2COL574 3COL574 1COL574 2COL574 4COL574 5COL573 3OTI30 1OTI30 2OTI30 3OTI30 4OTI30 5GAB17 1GAB17 3GAB17 2GAB17 4GAB17 5 AGF319 1AGF319 2AGF319 8AGF319 5AGF101 1AGF101 2AGF101 4AGF101 3OXC85 1OXC85 2AGF101 5FLH14 1FLH14 5FLH14 4FLH14 2FLH14 3FLH15 1FLH61 2FLH15 2FLH15 5FLH15 4FLH15 3FLH61 1FLH61 3FLH61 5FLH61 4 SUP223 1SUP223 2SUP223 3SUP223 5SUP223 4AGF450 1AGF450 2 OXC757 6OXC757 8OXC757 9OXC86 1OXC86 3OXC86 2OXC86 4OXC757 1OXC757 2OXC757 5 OXC32 1OXC32 2OXC32 3OXC32 4OXC32 5OXC757 3OXC86 5 LGC180 1LGC180 3LGC180 5LGC180 4LGC180 2LGC687 1LGC70 1LGC70 3LGC70 4LGC70 5LGC70 2LGC68 1LGC68 2LGC68 5LGC68 4LGC68 3LGC992 3LGC992 4LGC992 5LGC69 1LGC69 2LGC69 3LGC992 1LGC992 2TND244 1TND244 2TND247 5TND245 5TND247 1TND247 2TND247 4TND247 3TND245 1TND245 4TND245 2TND245 3MRL78 5MSP81 4MSP81 2MSP81 1ORO28 4SUC39 3ORO28 3ORO28 1ORO28 2ORO28 5SUC39 2SUC39 1MSP25 3MSP25 5MSP25 4MSP25 2MSP81 3MSP720 1SUC39 5MSP188 2MSP25 1SUC39 4PRM88 1PRM87 1BCP504 6 HOU655 10HOU272 1HOU272 2HOU272 3HOU272 5HOU271 1HOU271 4HOU271 5HOU271 2HOU272 4HOU271 3HOU654 1HOU654 3HOU655 1HOU655 5HOU654 2HOU654 5HOU654 4HOU655 2HOU654 6HOU654 7 SNK810 1SNK810 4SNK810 2SNK810 5SNK810 3SNK294 2SNK293 1SNK294 5SNK293 2SNK293 3SNK293 4SNK293 5SNK294 3SNK294 4SNK294 1SNK213 1SNK213 3SNK213 4SNK213 2SNK213 5SNK212 1SNK212 2SNK212 3SNK212 5SNK212 4HOU654 8HOU654 9 CHN554 2CHN554 4CHN554 5 STO390 1STO390 10STO390 6STO390 8STO390 5STO390 2STO390 7STO390 4STO221 5STO839 1STO839 2STO839 3STO839 4RAP799 AR20STO221 1STO840 1STO840 2STO221 4STO221 2STO221 3STO297 1STO297 3STO297 4STO297 5STO297 2STO390 3 STO839 5STO837 7STO837 9STO837 2STO837 8STO837 4STO837 1STO837 10 STO842 6STO842 8STO840 7STO842 10STO842 9STO842 7STO842 3STO842 5STO842 4STO840 10STO840 9STN296 2STO298 1STO298 4STO298 3STO298 5STO835 1STO835 2STO835 5STO835 3STO835 4STO298 2STO840 6STO841 3STO841 4STO840 8STO842 2STO841 1STO841 2STO840 3STO841 5STO838 6STO838 7STO838 8STO838 9STO838 10STO838 5BLG48 1MAT192 1 HJT349 3HJT349 4HJT349 5HJT349 8HJT349 6HJT640 2HJT640 5HJT640 3HJT640 4HJT640 1HJT350 1HJT350 10HJT350 AR35HJT350 8HJT350 AR37HJT350 9HJT350 6HJT350 3HJT350 7HJT350 5HJT350 4HJT350 2HJT641 1HJT641 3HJT641 5HJT641 AR38HJT641 AR39HJT641 AR40HJT643 1HJT643 3HJT643 4HJT643 2HJT641 2HJT641 4HJT639 1HJT639 3HJT639 2HJT639 5HJT639 4HJT639 21HJT639 23HJT639 24HJT639 22HJT639 25 PAM287 1PAM287 4PAM287 3PAM287 2PAM287 5CHC262 1CHC262 2CHC262 3CHC262 4CHC262 5PTA369 1PTA369 8PTA369 7PTA370 1PTA765 1PTA765 2PTA765 4PTA765 5PTA370 2PTA370 6PTA370 8PTA370 9PTA370 7PTA369 10PTA369 9PTA369 2HJT642 1HJT642 5HJT642 2HJT642 4HJT642 3PTA369 5PTA767 AR27PTA767 1PTA767 AR29PTA767 3PTA767 5PTA767 AR28PTA767 4PTA370 4PTA370 5GRL607 1FEN160 1FEN160 5FEN156 3FEN160 3FEN160 4FEN160 2AZN162 1AZN162 2AZN162 4AZN162 5AZN162 3AZN54 1AZN54 2AZN54 3AZN54 4AZN54 5AZN55 3AZN55 4AZN55 1AZN55 2AZN55 5AZN13 1AZN13 2AZN13 3AZN13 4AZN13 5AZN161 1AZN161 5AZN161 4AZN161 3AZN161 2LPS19 1LPS19 5LPS19 4LPS19 2LPS19 3FEN156 1FEN156 2FEN156 4FEN156 21FEN156 22FEN156 23FEN156 24FEN156 5FEN156 25FEN597 1FEN597 3FEN597 2FEN597 4FEN597 5 FEN157 1FEN157 4FEN157 5FEN157 2FEN157 3FEN158 1FEN158 3FEN158 4FEN158 2FEN158 5AZN56 1AZN56 2AZN56 3AZN56 4AZN56 5FEN159 1FEN159 2FEN159 4FEN159 5FEN159 3FEN598 1FEN598 5FEN598 4FEN598 3FEN598 2PLT378 1PLT378 2PLT378 3PLT378 10PLT378 4PLT378 5PLT378 7PLT378 8 PLT789 AR23PLT789 AR22PLT789 AR25PLT789 AR26PLT789 AR24PLT789 1PLT789 2PLT789 4PLT789 3PLT379 1PLT379 8PLT379 7PLT379 2PLT379 9PLT379 10PLT379 4PLT379 6PLT788 3PLT788 5PLT788 4PLT786 5PLT786 1PLT786 3PLT786 2PLT786 4PLT787 1PLT787 5PLT787 2PLT787 3PLT787 4PCS769 1PLT789 7PLT789 9STO389 1STO389 3STO389 10STO389 2STO389 4STO389 5STO389 7STO389 6STO389 9STO836 1STO836 2STO836 5STO836 4STO836 3GRL607 2GRL607 3GRL607 4GRL607 5 PTA765 3PTA765 6PTA765 7PTA765 8PTA765 9PTA767 7PTA767 10PTA767 8PTA765 10PTA767 6PLT788 1PLT788 2PTA767 9PTA766 1PTA766 4PTA766 5PTA766 2PTA766 8PTA766 10PTA766 9PTA766 3RAP799 4PTA766 6STO837 6FEN596 2FEN596 4FEN596 5FEN596 3FEN596 1CHC547 7 PLT789 8GRL1052 1VER920 6BCP504 9 HCB354 10HCB353 1HCB353 2HCB353 3HCB353 6HCB353 9HCB354 6HCB354 4HCB170 25HCB353 8HCB354 2HCB354 8HCB354 5HCB354 1HCB170 22HCB170 23HCB18 11HCB18 1HCB18 5HCB18 12HCB18 14HCB18 4HCB18 13HCB18 2HCB18 15HCB18 3SGR215 1SGR215 2SGR215 4SGR215 3SGR215 5SGR315 1SGR315 4SGR316 1SGR316 2SGR316 4SGR315 5SGR315 2SGR315 3SGR316 5SGR814 1SGR814 4SGR814 3SGR814 2SGR814 5ABN423 5 CAP335 1CAP335 2CAP335 10CAP335 8CAP335 6CAP335 3CAP335 5CAP335 4CAP335 7CAP534 1CAP334 1CAP334 5CAP534 2CAP534 5CAP534 3CAP334 2CAP334 4CAP534 4CAP537 1CAP538 1CAP538 2CAP538 4CAP538 3CAP261 1CAP261 2CAP261 3CAP261 4CAP261 5CAP535 1CAP535 2CAP535 3CAP535 4CAP535 5SST813 3CAP536 1CAP536 2QUM566 1QUM566 2QUM566 4QUM566 5QUM566 3 QUM340 5QUM340 7QUM340 6QUM567 2QUM567 1QUM567 3QUM341 5QUM341 10QUM341 4QUM341 9QUM341 7QUM341 1QUM341 3QUM341 6QUM341 8QUM565 1QUM565 3QUM565 2QUM565 4QUM565 5CRC563 1CRC563 3CRC563 4CRC563 5CRC563 2 CRC564 1CRC564 3CRC564 4CRC564 2CRC564 5 MSP721 1MSP721 4MSP721 2MSP721 3MSP721 5OXC757 7DCM147 1DCM149 1DCM149 2CHV11 2CHV11 3CHV11 1CHV11 4CHV11 5DCM308 1DCM308 3DCM308 4DCM308 2DCM308 5DCM148 1DCM148 3DCM148 5DCM148 4DCM148 2DCM148 21DCM148 24DCM148 22DCM148 25DCM148 23AGU305 1AGU305 2AGU305 4AGU305 3AGU305 5ACS100 1ACS100 5ACS100 2ACS100 3ACS100 4 MCQ716 5MCQ716 4MCQ718 2MCQ718 3MCQ718 5MCQ718 4MCQ716 2MCQ716 1MCQ716 3MCQ719 4MCQ719 1MCQ719 5MCQ719 2MCQ719 3 CHN1 1CHN1 5CHN1 3MCQ718 AR71MCQ718 AR74MCQ718 AR72MCQ718 AR73CHN1 2MCQ717 2MCQ717 3MCQ717 5MCQ717 4CHN1 4 MCQ718 1CHN552 1CHN552 4CHN552 5CHN552 25CHN552 23CHN552 22CHN552 24CHN552 21CHN552 3CHN552 2CHN553 1CHN553 4CHN553 5CHN553 2CHN553 3HCR311 1HCR311 2HCR311 4HCR311 5HCR311 3HCB170 11IMT172 1IMT64 3IMT172 2IMT172 4IMT172 5IMT172 3IMT64 1IMT64 5IMT64 4IMT64 2IMT63 1IMT63 2IMT63 3 PCS91 1PUR34 1PUR34 3PUR34 5PUR34 4 PUR34 2PUR206 1PUR206 2IRS312 1IRS312 3IRS312 4IRS312 5IRS312 2IRS66 1IRS313 1IRS313 2IRS313 5IRS313 4IRS313 3HRO310 1HRO310 2HRO310 3HRO310 4HRO310 5BLG115 1 BLG996 1BLG996 3BLG996 2BLG996 4BLG48 3BLG48 5BLG48 4CHQ50 1 ACG45 1ACG45 3ACG45 5ACG45 4 ACG448 1ACG45 2 CHM558 1CHM558 4CHM558 3CHM559 2CHM559 3CHM559 5CHM555 1CHM555 2CHM555 4CHM555 5CHM555 3 CHM559 1CHM559 4CHM558 2 CHM556 1CHM556 2CHM556 3CHM562 1CHM562 3CHM562 2CHM997 3CHM997 5CHM997 2CHM997 1PUR205 1PUR205 5CHM557 3CHM557 4CHM557 2CHM557 5CHM557 1PUR205 2PUR205 3PUR205 4CHM997 4 PCJ768 1PCJ768 5PCJ768 2PCJ768 3PCJ768 4PCJ33 1PCJ33 4PCJ33 2PCJ33 5PCJ33 3SOL216 1SOL216 2SOL216 3SOL216 4SOL216 5PCJ199 1PCJ199 4PCJ199 3PCJ199 5PCJ199 2PCJ90 3PCJ90 5PCJ90 4TUQ890 1TUQ890 3TUQ890 4TUQ890 5TUQ890 2TUQ96 1PCJ89 1PCJ90 2PCJ89 2PCJ89 5PCJ90 1PCJ89 3PCJ89 4MIN24 1MIN24 2MIN24 3MIN24 5MIN24 4ACP304 1ACP304 4ACP304 2ACP304 5ACP304 3SCB37 1 SCB37 2SCB37 4SCB37 3SCB37 5BLG6 1BLG6 2BLG6 4BLG6 5BLG6 3ABZ102 1ABZ102 2ABZ102 3ABZ103 4ABZ102 5ABZ102 4ABZ103 1ABZ103 2ABZ103 3ABZ103 5ABZ466 3ABZ466 1ABZ466 2ABZ466 5ABZ2 1ABZ466 4ABZ2 2ABZ2 3ABZ2 4ABZ2 5ETB591 1ETB591 3ETB591 4ETB591 5ETB594 1ETB594 2ETB591 2ETB593 4ETB593 3ETB595 1ETB595 5ETB595 3ETB595 4ETB593 1ETB595 2ETB593 2PLS760 1PLS759 5PLS760 5PLS759 3PLS760 3PLS759 4PLS759 1PLS760 2PLS197 1PLS197 5PLS197 4PLS197 3PLS197 2PLS198 2PLS198 1PLS198 3PLS198 5PLS198 4PLS760 4PLS759 2PLS761 1PLS284 5PLS285 1PLS286 3PLS761 4PLS284 2PLS286 5PLS285 2PLS285 3PLS284 3PLS285 5PLS286 1PLS285 4PLS284 1PLS286 2PLS284 4PLS286 4PLS761 5PLS761 2PLS761 3FRN600 1FRN59 2FRN600 3FRN600 2FRN600 4FRN600 5FRN599 1FRN599 2FRN59 1 FRN599 3FRN599 4FRN599 5NCD734 6NCD734 1NCD734 5NCD734 AR67NCD734 2NCD734 AR68NCD734 AR70NCD193 1NCD193 3NCD193 5NCD193 4NCD193 2MRL187 3MRL187 4MRL187 5 MRL78 1MRL78 2MRL78 3MRL79 1MRL79 2 MRL79 3CLR568 1CLR568 2CLR52 1CLR133 1CLR133 5CLR133 2CLR570 1CLR570 4CLR570 2CLR570 3CLR52 2CLR52 3CLR52 4CLR569 1CLR569 2CLR569 4CLR569 3CLR569 5CLR999 2CLR999 1LES358 10LES358 4LES358 3LES358 6LES358 5LES358 8LES358 7LES358 2LES358 9LES21 1LES21 3LES21 2LES21 4LES21 5 PLD377 10PLD377 3PLD377 2PLD785 2PLD783 1PLD783 3PLD783 6PLD783 10PLD784 1PLD784 2PLD784 4PLD784 3PLD784 5PLD376 1PLD376 5PLD783 5PLD782 4PLD785 1PLD785 4PLD785 3PLD782 1PLD376 2PLD376 6PLD783 2PLD376 8PLD785 5PLD376 10PLD376 3PLD782 9PLD376 4PLD782 10PLD782 7PLD783 7PLD207 1PLD207 4PLD207 2PLD207 3PLD207 5LES20 1LES20 2LES20 4LES20 3LES20 5PLD783 9PLD783 8 PLD782 8PNT776 10CAN951 2 BLB331 10BLB331 4BLB331 2BLB331 AR14BLB331 3BLB331 5BLB331 6BLB331 7BLB945 4BLB945 5BLB330 5BLB330 6BLB330 9BLB330 8BLB516 1BLB516 2BLB516 4BLB946 4BLB946 2BLB946 3BLB946 5BLB516 3BLB947 1DPH937 AR55BLB522 1BLB522 3BLB522 4BLB522 2BLB516 5BLB517 1BLB517 4BLB519 AR10BLB519 AR11BLB330 1BLB946 1BLB522 AR58BLB521 AR57DPH937 AR54BLB522 AR56DPH119 3DPH119 4BLB330 4BLB330 10BLB330 7BLB517 2BLB517 5BLB517 3BLB518 3BLB519 AR12BLB518 1BLB518 4BLB518 AR9BLB518 5BLB518 AR2BLB518 2BLB947 2BLB947 3BLB947 4BLB515 6BLB515 9BLB515 7BLB515 5BLB945 3BLB948 1BLB521 4BLB948 2BLB948 3BLB945 2BLB521 3BLB521 AR60BLB521 2BLB521 5BLB522 AR59BLB948 4BLB948 5BLB515 2BLB521 1BLB945 1BLB522 5BLB515 3 DPH119 1DPH119 5DPH119 2DPH9 1DPH9 2DPH9 4DPH9 5DPH9 3DPH10 1DPH10 5DPH10 4DPH10 2DPH10 3BLB515 8BLB515 10BLB515 1BLB520 1BLB520 2BLB520 5BLB520 3BLB520 25BLB520 4BLB520 23BLB520 24BLB520 22BLB520 21 PTT121 1PTT121 4PTT121 5PTT121 2PTT121 3 BLB523 1BLB523 4BLB523 AR3BLB523 AR1BLB523 2PTT120 3BLB525 2BLB525 3BLB525 5PTT120 1PTT120 4PTT120 5PTT120 2BLB523 5BLB524 AR8spp AR13BLB523 3BLB524 5BLB524 1BLB524 4BLB524 3BLB525 1BLB949 1BLB950 4BLB950 2BLB950 5BLB950 3BLB950 1BLB949 2BLB949 5BLB949 3BLB525 4BLB949 4 BLB524 2BLB524 AR6BLB524 AR7BLB523 AR63BLB523 AR64BST325 1BST325 2BST325 8BST325 3BST325 10BST325 9BST325 4BST325 6BST325 7 BST500 1BST500 4BST500 5BST500 2BST500 3NYR27 11NYR27 2NYR26 31NYR27 4NYR27 5NYR26 51NYR26 1BST117 1BST117 3BST117 4BST117 5BST117 2NYR27 41 NYR27 51NYR26 41NYR27 3NYR27 1NYR27 21NYR27 31NYR26 21NYR26 2NYR26 3NYR26 11 CPH542 1CPH542 2CPH542 4CPH542 5CPH542 3EHR153 1EHR153 2EHR153 3EHR153 5 BST118 1BST118 22 BST118 2BST118 3BST118 4BST118 23BST118 24BST118 21BST118 25BST118 5CPH124 1EHR155 2EHR155 22EHR155 3EHR155 23EHR155 21EHR155 24EHR155 4EHR155 25EHR155 5 EHR154 1EHR154 2EHR154 3EHR154 21EHR154 23EHR154 4EHR154 24EHR154 25EHR154 5BST258 1 BST258 3BST258 5BST258 2BST258 4EHR154 22BLB515 4 CPH336 1CPH336 5CPH336 8CPH336 2CPH337 9CPH337 1CPH337 5CPH337 10CPH336 4CPH336 7CPH337 8CPH336 10CPH336 9CPH337 3CPH337 AR75CPH541 1CPH541 4CPH541 2CPH541 3CPH541 5CPH539 2CPH539 3CPH337 2CPH539 1CPH539 5CPH539 4SMB92 1BST116 1JAM355 1JAM355 6JAM355 7JAM355 2JAM355 9JAM355 4JAM355 3JAM355 10JAM355 8BST116 4 BST116 2BST116 3BST116 5JAM673 1JAM673 2JAM274 1JAM274 2JAM274 5JAM274 3JAM274 4EHR268 3EHR268 1EHR268 2EHR268 4EHR268 5 JAM672 1JAM672 4JAM672 5JAM672 3JAM672 2JAM674 1JAM674 4JAM674 3JAM674 2JAM674 5 PNT375 1PNT375 5PNT375 3PNT776 1PNT777 2PNT375 9PNT778 1PNT781 1PNT374 4PNT780 3PNT375 4PNT778 2PNT778 3PNT375 6PNT375 2PNT375 10PNT375 8PNT778 4PNT374 7PNT776 5PNT775 2PNT776 2PNT780 1PNT780 4PNT780 2PNT780 5PNT775 1PNT374 10PNT374 3PNT374 2PNT778 5PNT374 5PNT374 8PNT374 9PNT781 4PNT775 10PNT776 7PNT781 2PNT781 3PNT779 1PNT779 2PNT779 4PNT779 3PNT779 5PNT776 9PNT776 6PNT775 8PNT775 9PNT204 1PNT204 5 PNT204 2PNT204 3PNT204 22PNT204 23PNT204 24PNT204 25PNT204 21PNT204 4 TRF880 3TRF880 5TRF880 9TRF880 10TRF880 7TRF880 4TRF880 1TRF880 6TRF231 1TRF880 2 TRF231 2TRF231 4TRF231 3TRF231 5TRF880 8TRF882 1TRF882 2TRF882 4TRF882 3TRF882 5MCH185 1MCH185 2MCH185 3MCH185 4TRF881 1TRF881 3TRF881 4TRF881 5TRF881 2MCH279 1MCH279 5MCH279 2MCH279 4MCH279 3TRN41 1TRN41 2TRN41 3TRN41 5TRN41 4TRN228 3TRN40 1TRN40 2TRN40 3TRN40 4TRN229 2TRN229 3TRN229 5TRN228 5TRN228 2TRN228 1TRN229 1TRN229 4TRN40 5TRN228 4TRN226 1TRN226 3TRN226 5TRN226 4TRN226 2TRN227 3TRN227 4TRN227 1TRN227 2TRN227 5STO838 3BCP504 7CNS332 1CNS332 7CNS333 1CNS333 10CNS333 3CNS333 5 FRX602 1FRX602 2FRX602 3FRX602 4FRX602 5VRN905 8 SIT812 1SIT812 5SIT812 2SIT812 4SIT95 3SIT95 5SIT95 1SIT95 2SIT812 3SIT38 1SIT38 2SIT38 5SIT38 4SIT38 3SIT95 4LYC71 2LYC72 1LYC72 2LYC71 5LYC71 3BCP504 8LYC314 1LYC314 4LYC314 5LYC314 2LYC314 3PNT775 7CHP339 1CHP339 9CHP339 7CHP339 2CHP339 3CHP339 8CHP339 4CHP339 6nig968 1nig968 5nig968 4nig968 3nig968 2
Acaulia
S. demissum
Cultivated
and
from Peru
Tuberosa
Tuberosa
Outgroup
Polyadenia
Pinnatisecta
Bulbocastana
Morelliformia
Circaeifolia
PiuranaMegistacroloba
Tuberosa
S. verrucosum
Demissa
Conicibaccata
Longipedicellata
Tuberosa
Megistacroloba
Tuberosa
Yungasensa
from
Bolivia, Argentina and Chile

BMC Evolutionary Biology 2008, 8:145 http://www.biomedcentral.com/1471-2148/8/145
Tuberosa from Bolivia, Argentina and Chile plus someaccessions from other series such as Yungasensa, 8) acces-sions from cultivated Tuberosa species and wild Tuberosafrom Peru, 9) accessions from Tuberosa and Megistacroloba,10) accessions from S. acaule (and its subspecies), S. albi-cans, S. demissum, S x semidemissum and S. edinense.
The condensed dataset (916 genotypes)Because of the size of the dataset, it proved impossible toanalyze it with cladistic methods nor to analyze it for sta-tistical support. A condensed dataset was created bychoosing a representative genotype from all the availableaccessions (see methods section for exact details). Thiscondensed dataset consisted of 916 genotypes.
A single ratchet parsimony search consisting of 200 itera-tions yielded a Maximum Parsimony (MP) tree of 9669steps. Furthermore, 20 individual independent ratchetsearches each consisting of 50 iterations also yielded a MPtree of 9669 steps.
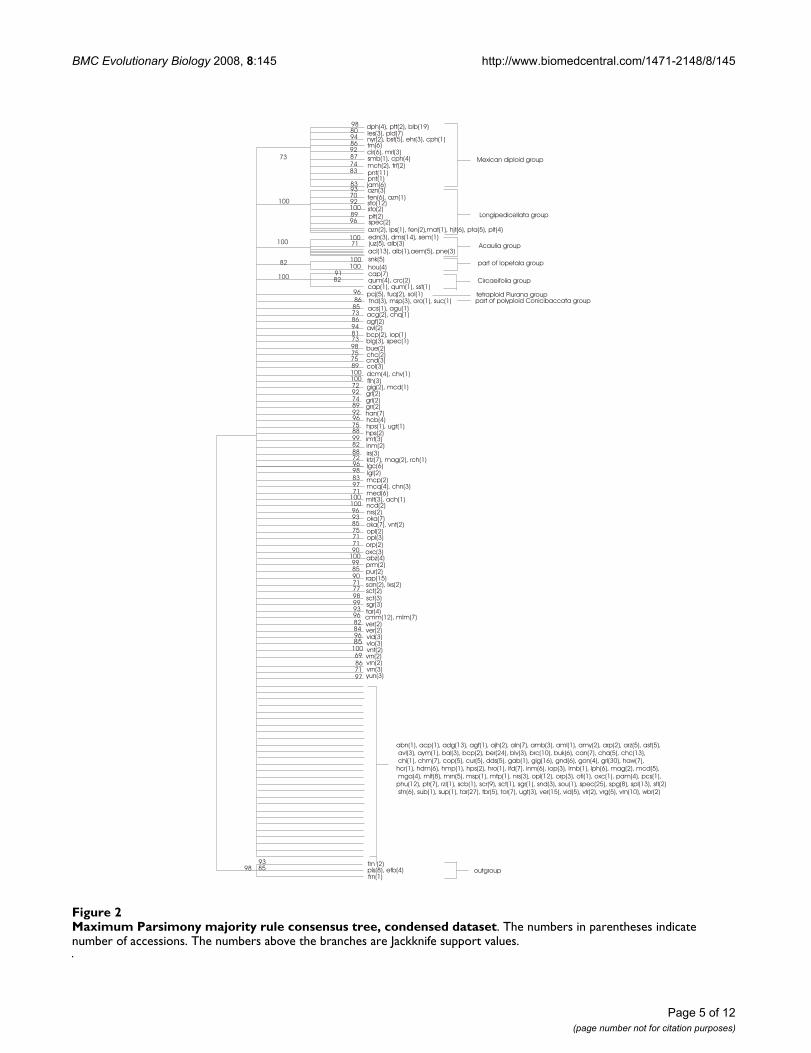
Figure 2 shows the schematised majority rule consensusNJ jackknife tree and Figure 3 shows the schematisedmajority rule consensus MP jackknife tree of the con-densed dataset. The strict consensus trees were manipu-lated in such a manner that not all the separate brancheswere represented but some were summarised. The sche-matised trees only show branches with more than 69 jack-knife support. The original majority rule consensus NJjackknife tree and majority rule consensus MP jackknifetree are available from the authors as supplemental data.
When comparing the NJ and the MP jackknife trees it isapparent that a large part of both trees consists of a poly-tomy. However, some structure is still visible in both trees,supported by jackknife values above 69. The followinggroups can be recognized in both the NJ jackknife tree andthe MP jackknife tree:
1) Mexican diploid species, with a jackknife support of 73for the MP tree and 99 for the NJ tree; the substructurefound within the Mexican and Northern American dip-loids is almost the same for both trees.
2) A group of tetraploid Mexican/North and CentralAmerican species belonging to series Longipedicellata, witha jackknife support of 100 in both trees.
3) A group consisting of accessions of S. acaule, S. demis-sum, and closely related species with a jackknife support of100 in the MP tree and 99 in the NJ tree.
4) A group consisting of the species belonging to seriesCircaeifolia, with a jackknife support of 100 in both trees.
5) A small group of accessions belonging to S. paucijugum,S. tuquerrense, and S. solisii, tetraploid species belonging tothe series Piurana, with a jackknife support of 96 in the NJtree and 92 in the MP tree.
There are also differences in group structure between thetwo trees. There are a number of groups that have goodjackknife support in the NJ tree but are not supported inthe MP jackknife tree:
1) A group of hexaploid Mexican species belonging toseries Demissa with a jackknife support of 79. In the MPtree only 2 species that are part of this group were foundin one small clade: S. schenckii and S. hougasii.
2) A group of accessions from species belonging to seriesConicibaccata has a jackknife support of 82 in the NJ jack-knife tree. In the MP jackknife tree the same accessions arepart of the polytomy. These clades represent the sub-groups found within the Conicibaccata group in the NJtree. Only one subgroup is not represented by a similarclade in the MP jackknife tree.
3) A group of species belonging to series Piurana has ajackknife support of 69 in the NJ tree. In the MP tree, thejackknife support was low, so this group collapsed and 4out of 5 supported subgroups found in the NJ jackknifetree are visible as supported separate small groups in theMP jackknife tree.
4) A group consisting of accessions from diploid speciesof series Conicibaccata, S. buesii, S. sandemannii and S. lax-issimum with jackknife support of 92.
5) A group which contains accessions of S. medians, S. san-demanii, S. weberbauerii and an unknown species with ajackknife support of 85.
DiscussionThe value of AFLPOne of the arguments against the use of AFLP is the possi-ble bias caused by homoplasy [17,19,20]. Non-identicalco-migrating bands in the AFLP fingerprints can contrib-ute noise instead of signal to the dataset without beingdetected. However, it is not likely that in the tuber-bearingwild potatoes homoplasy will cause many problemsbecause the species are all very closely related and homo-plasy becomes a problem when distantly related speciesare involved. Koopman [16] showed that in a set ofclosely related Lactuca species, sufficient phylogenetic sig-nal was present and concluded that in practice the influ-ence of possible limitations of AFLP, such as co-migrationof nonhomologous fragments is limited. However, hestresses that the conclusion only applies to datasets withclosely related species. Moreover, Kardolus [19] concludes
Page 4 of 12(page number not for citation purposes)
BMC Evolutionary Biology 2008, 8:145 http://www.biomedcentral.com/1471-2148/8/145
Page 5 of 12(page number not for citation purposes)
Maximum Parsimony majority rule consensus tree, condensed datasetFigure 2Maximum Parsimony majority rule consensus tree, condensed dataset. The numbers in parentheses indicate number of accessions. The numbers above the branches are Jackknife support values.

BMC Evolutionary Biology 2008, 8:145 http://www.biomedcentral.com/1471-2148/8/145
Page 6 of 12(page number not for citation purposes)
Neighbour Joining majority rule consensus tree, condensed datasetFigure 3Neighbour Joining majority rule consensus tree, condensed dataset. The numbers in parentheses indicate number of accessions. The numbers above the branches are Jackknife support values.

BMC Evolutionary Biology 2008, 8:145 http://www.biomedcentral.com/1471-2148/8/145
from his AFLP results that in Solanum section Petota theAFLP technique is suitable up to the species level. TheAFLP method has since then successfully been used inmore studies on potato taxonomy [21-24].
Status of groups within section PetotaNot all the groups found in this study have the same levelof cohesion or have the same level of demarcation. Somegroups have clear borders, while from others we can onlyvaguely recognize the contours. First, there is a number ofgroups that are always well supported, whether the analy-sis is done in a phenetic or phylogenetic way, see Figure 2and 3. This is the case for the group of Mexican diploidspecies, the group of Mexican tetraploids, the group of S.demissum and S. acaule, the group of S. circaeifolium, thegroup of S. commersonii and the group of S. schenckii andS. hougasii. Then there are groups that are not supportedin the MP jackknife tree (Figure 2) but that can be foundin both the original MP trees and NJ trees (not shown)and are supported in the NJ jackknife tree (Figure 3). Thisapplies to the group with Mexican hexaploid species, thegroup containing polyploid species belonging to seriesConicibaccata, the group containing diploid Piurana spe-cies, and the small groups of S. huancabambense, S. kurtz-ianum, S. medians, S. mochiquense, S. hannemanii, S. buesii,and S. paucijugum.
The largest part of the jackknife trees consists of a poly-tomy of species that does not seem to contain structure atall. If one was only to consider the structure shown in thejackknife trees, the conclusion would have to be thataccording to the results of the present AFLP analyses thelargest part of section Petota is without any taxonomicstructure.
However, it is possible to identify additional groups thatare present in many of the original NJ and MP trees, butdo not have enough support to be shown in the jackknifetrees. For example, in the 4929 dataset NJ tree a clusterrepresents the group of cultivated potatoes together withspecies of series Tuberosa from Peru. The groups that arefound in both the phenetic and phylogenetic analysis arestrong groups with clear borders. The exchange of geneticmaterial is most likely restricted to the members of thegroup. The groups with only low support in the MP aloneor in both trees are groups that probably share a consider-able amount of genetic material with genotypes outsidethe group. In a study of Jacobs, van den Berg and Vosman:Comparison of Chloroplast DNA and AFLP data fromSolanum section Petota reveals incongruencies between thedatasets, submitted, the incongruencies found betweenthe chloroplast data and the AFLP data suggest thathybridization occurs between species of different series insection Petota. For example, the composition of species ofthe clade representing the series Piurana in the chloroplast
tree is different from that of the clade representing thePiurana series in the AFLP tree.
The resulting groups also have implications for the theoryon EBN of Hawkes and Jackson [25]. EBN stands forEndosperm Balance Number and refers to a hypotheticalgenetic factor that would explain the success or failure ofcrosses due to the functioning or breakdown of theendosperm after fertilization. Crosses between specieswith the same EBN are generally successful and crossesbetween species with different EBN generally are not,independent of ploidy levels. Hawkes and Jackson [25]claim that there is a correlation between the EBN hypoth-esis and the evolution of the group of tuber-bearing Sola-num species. EBN 1 is found mainly in species that areconsidered to be close to the ancestors of the group: Mex-ican series Morelliformia, Bulbocastana, Pinnatisecta, andPolyadenia. The EBN 2 condition would have arisen as anisolating mechanism when potato species moved south-wards. The EBN 4 condition occurs in hexaploids whichare allopolyploids.
From the present results it is clear that there is no absoluterelationship between EBNs and the groups found. In thegroup which contains S. acaule, S. demissum, S. semidemis-sum and S. edinense, different ploidy levels and differentEBNs occur. This mixture of ploidy and EBN levels alsooccurs in the group with representatives of series Conici-baccata. The species S. moscopanum and S. tundalomenseboth are hexaploid and have EBN 4 and they form a groupor cluster together with other series Conicibaccata specieswhich are known to be tetraploid and have EBN 2.Although these tetraploid and hexaploid species fromseries Conicibaccata are mixed, the diploid series Conicibac-cata (EBN 2) species do form a separate cluster.
With regard to the overall structure of the section as foundin this study two main observations can be made. Thereseems to be a lack of supported structure, especially in theSouth American part of section Petota. Furthermore, thereis a lack of support for the relationships between the dif-ferent groups that were found in the NJ and MP trees. It isimportant to differentiate between these two phenomenabecause the causes underlying both cases could be differ-ent.
Lack of structure in South American part of section PetotaThe AFLP jackknife NJ tree and the jackknife MP tree inthis study shows a lack of structure or rather, an unre-solved structure for the part of the tree which containsSouth American species while the other part of the treeshows several well supported groups.
Kardolus et al. [19] mentioned that within series Tuberosadifferent genotypes of the same species are not always
Page 7 of 12(page number not for citation purposes)

BMC Evolutionary Biology 2008, 8:145 http://www.biomedcentral.com/1471-2148/8/145
grouped together and are scattered among genotypes fromother species. He claims that the cause of this phenome-non is not the lack of resolution of AFLP, but the overclas-sification of a group of species, the so-called brevicaule-complex. The cpDNA RFLP studies of Spooner and Sytsma[11], and Spooner and Castillo [14] also showed a lack ofsupport for a resolved structure within the group of SouthAmerican species, and the branch uniting all these specieshad a bootstrap support value of only 67.
Volkov et al. [26] compared the ETS region of rDNA for 30species of Solanum section Petota and found high boot-strap values for the branch uniting all the South Americanspecies in three different types of dendrogram (Maximumparsimony, Bayesian statistics and Neigbour Joining).However, the two subgroups within the South Americanclade that they distinghuished (variants C1 and C2) oftenshow polytomies and resolution within the groups ismostly lacking.
Outside the field of potato taxonomy, researchers havereported similar patterns. Hughes and Eastwood [27]report a low sequence divergence and lack of resolution inthe large Andean clade of the genus Lupinus. This wouldpoint at a rapid and recent diversification in the Andes.The authors also suggest that Lupinus is probably only oneexample of many plant radiations that followed the finaluplift of the Andes. They assume that many of these plantradiations are yet unknown. It is possible that the factorsunderlying the Lupinus diversification are also responsiblefor the Solanum section Petota diversification. According toHughes and Eastwood [27] these factors would be thelarge scale of the area over which the radiation extends,repeated fragmentation of high altitude habitats due toquaternary climate fluctuations, the extremely dissectedtopography, and the habitat heterogeneity.
Lack of support for relationships between different groupsExcept for the outgroup consisting of S. etuberosum, S.palustre and S. fernandezianum which connects to the mainbranch of the NJ jackknife and MP jackknife tree withrespectively 100 or 98 support value, none of the branchesconnecting two or more groups have jackknife support of69 or higher. That is the reason why in the schematizedjackknife NJ and jackknife MP trees these branches col-lapse in a polytomy. Contrastingly, the branches of thegroups that can be recognised within the polytomy dohave jackknife support, although not all species can be putin groups as discussed previously.
In the first study on the use of AFLP in Petota taxonomy byKardolus et al. [19], it proved also difficult to find boot-strap support for branches connecting the different groupsin section Petota. Bootstrap support above 70 were givenfor a NJ tree branch connecting the outgroup of S. etubero-
sum and S. brevidens, for a branch connecting the out-groups, and for the Mexican diploids and S. circaeifoliumand S. circaeifolium subspecies quimense with the otherpart of the tree. In the cpDNA RFLP studies on the SouthAmerican part of section Petota [14] only a few branchesconnecting the larger groups showed bootstrap supportabove 70. Clade 1, consisting of Mexican diploids (exceptS. cardiophyllum and S. bulbocastanum) is connected to theother clades with a bootstrap value of 87, and Clade 3(mainly accessions belonging to series Piurana) and Clade4 (the rest of section Petota) are connected to each otherwith a branch with 96 bootstrap support.
We can conclude from these previous results that it isindeed difficult to find good support for the backbonestructure of section Petota in general. This indicates thatour and previous results represent the real biological situ-ation in Solanum section Petota. Since the phylogenetic sig-nal is clearly present in our data as shown in the well-supported groups in the present study, the lack of struc-ture in parts of the tree is not caused by the lack of phylo-genetic signal in AFLP markers.
New informal species groups for Solanum section PetotaAs outlined in this paper and in other earlier studies, thereare no results that support the classification of sectionPetota in 21 series. Although a few of the series seem toform natural groups, the majority of the series as pro-posed by Hawkes [2] could not be found as separate clus-ters or clades. Our goal is to use the found structure in thepresent study at maximum for classifying the sectionPetota.
We propose to divide section Petota in informal speciesgroups, following the approach of Spooner et al. [10] whoconstructed 11 informal species groups for the North andCentral American species. They followed the approach ofWhalen [28] and Knapp [29,30] who applied a similarinformal species group classification. We will use thenames already used by Spooner et al. [10] if applicable,and add new groups that were not treated in their study.We chose to base the informal group classification on thegroups that are supported in the NJ jackknife tree. The NJjackknife tree shows more resolution relative to the MP.However, it would not be useful to consider every smallgroup that appears in the schematized tree as a biologi-cally meaningful group. Therefore, the choice for speciesgroups is restricted to groups of species that make sense inthe light of former studies and contain at least 3 species.We maintain the species group Verrucosa which containsonly one species, because this species group is already des-ignated by Spooner et al [10].
In total, the NJ jackknife tree can be partitioned into 10species groups. It would be possible to construct more
Page 8 of 12(page number not for citation purposes)

BMC Evolutionary Biology 2008, 8:145 http://www.biomedcentral.com/1471-2148/8/145
species groups based on the structure shown in the vari-ous trees made in the present study, but these groupswould then not be supported by bootstrap or jackknifesupports.
Although a closed classification following the rules of theBotanical Code is desirable, it seems in this case difficultto apply. In the present study, many species cannot beaccommodated in groups. These species do not automati-cally form a group themselves, but are intentionally leftunclassified.
We suggest recognizing the following informal speciesgroups as shown in the NJ jackknife tree (Figure 3):
Diploid Mexican groupThis group contains the species groups of Spooner et al.[10]: Pinnatisecta, Stenophyllidia, Trifida, Polyadenia,Morelliforme, and Bulbocastana. These species groupscan be recognized in the present study as separatebranches within the NJ cluster which represents this spe-cies group. In the present study we recognize a higher levelof group structure which contains all the mentioned spe-cies groups, because the detailed contents of each sub-group in our study (Figure 3) differs from the contentsfrom the species groups from Spooner et al. [10].
Acaulia groupIn our study this group contains 2 supported subgroups,one branch with jackknife support of 96 containing thespecies S. semidemissum, S. demissum and S. x edinense. Theother group shows a jackknife support of 98 and containsS. juzepczukii, S. albicans and the three subspecies S. acaulesubsp. acaule, S. acaule subsp. aemulans, S. acaule subsp.punae.
Iopetala groupThis group contains the species S. schenckii, S. hougasii,that form a strongly supported cluster together (jackknifesupport 100) and a cluster containing the species S. iopeta-lum, S. brachycarpum, S. guerreroense (jackknife support90). All species were formerly designated by Hawkes [2] toseries Demissa which also included the species S. demissumand closely related species. The species in our group arethe same as in the species group Iopetala designated bySpooner et al. [10]. They reduced the species S. brachycar-pum as a synonym of S. iopetalum.
Longipedicellata groupAs the name does suggest, this group contains species thatwere formerly placed by Hawkes [2] in the series of Longi-pedicellata. The species included in this group are S. fend-leri including S. fendleri subsp. arizonicum, S. stoloniferum,S. hjertingii,. S. papita, S. polytrichon, S. leptosepalum, and S.matehualae. The species S. leptosepalum, S. fendleri, S. pap-
ita, and S. polytrichon have been reduced as synonyms of S.stoloniferum [10]. The species S. matehualae is reduced assynonym of S. hjertingii [10].
Polyploid Conicibaccata groupThis group contains species placed there by Spooner et al.[10], complemented with South American species. Thespecies in this species group are mainly the same asHawkes [2] placed in series Conicibaccata. According to thepresent study the group consists of S. flahaultii, S. moscopa-num, S. orocense, S. sucubunense, S. tundalomense, S. oxycar-pum, S. longiconicum, S. garcia-barrigae, S. otites, S.oxycarpum, S. agrimonifolium, S. moscopanum, S. subspandu-ratum, S. paramoense, and S. colombianum.
Diploid Conicibaccata groupAlthough most of the series Conicibaccata can be put in thespecies group Conicibaccata there are a few species thatform a separate group. This group consists of the diploidspecies S. buesii, S. sandemanii, and S. laxissimum.
Diploid Piurana groupThis species group was not designated by Spooner et al.[10]. The name refers to the former series Piurana as thecontents of the group are roughly similar: S. piurae, S.acroglossum, S. blanco-galdosii, S. irosinum, S. chomat-ophilum, and S. paucissectum from series Piurana and S.chiquidenum from series Tuberosa.
Tetraploid Piurana groupThe situation as described before for the Conicibaccatagroup also applies partly for the Piurana group. There area few species from the formerly designated Piurana series[2] that form their own species group. This species groupcontains the tetraploid species S. paucijugum, S. tuquer-rense, and S. solisii.
Circaeifolia groupThis group consists of S. circaeifolium, S. soestii, S. capsi-cumbaccatum and S. circaeifolium subsp. quimense. The con-tents is conform Hawkes' series Circaeifolia.
Verrucosa groupThis group contains only 2 species; S. macropilosum and S.verrucosum. The species S. macropilosum was reduced to asynonym of S. verrucosum by Spooner et al. [10].
ConclusionAs far as we know, this paper treats the largest collectionof Solanum section Petota accessions ever analysed simul-taneously. All other previous studies used datasets thatincluded less variation and fewer species. Because of thethorough sampling, it is possible to propose speciesgroups without too many reservations. A number of spe-cies groups coincide with certain series recognized by
Page 9 of 12(page number not for citation purposes)

BMC Evolutionary Biology 2008, 8:145 http://www.biomedcentral.com/1471-2148/8/145
Hawkes [2]. However, most of the series that Hawkes andhis predecessors recognized, cannot be supported anylonger as natural groups, based on our current knowledge.The present study shows that the taxonomic structure ofSolanum section Petota is highly unbalanced. A few speciesgroups have high support and their inner structure dis-plays also supported subdivisions, while a large part of thespecies cannot be structured and they seem to be allequally related to each other and to the supported groups.
It might be difficult to accept that a part of genus Solanumsection Petota cannot be structured or subdivided. Weeven doubt that it would be possible to find more resolu-tion with other methods or more markers, and we con-sider it likely that the polytomy is indicative of the realsituation in section Petota. A relatively fast spread of tuber-bearing Solanum species over South America, due to thegeographic conditions in the Andes [27], combined withhigh levels of hybridisation may explain why the phyloge-netic links between species are so difficult to establish.
MethodsPlant MaterialIn total 951 accessions representing 196 different taxa,species, 15 subspecies and 17 hybrids were sampled. Wetried to include as many species as possible from variousgene banks. In principle, at least 5 accessions from eachavailable species and 5 individual plants per species(totally approx. 5000 genotypes) were included. Seedswere surface-sterilized and sown in vitro at 25°C. The col-lection of individual Solanum clones was grown in vitrofor at least 6 weeks on MS medium supplemented with20% sucrose [31] at 18°C. DNA was extracted from leafsaccording to the method described by Stewart and Via[32].
NomenclatureAdditional file 1 lists the species used and the accessionsrepresenting the species names according to the passportinformation from the gene bank. The labels used are notcorrected according to the synonymy in recent taxonomicrevisions for two reasons. First, we do not want to changean original label of an accession without actually checkingthe identity of that accession. Furthermore, by retainingthe original labels it is possible to check many hypotheseson the taxonomy of species. However, we have includedsome remarks about recent taxonomy changes in Addi-tional file 1. In some cases names/labels were corrected byus after preliminary AFLP results and visual inspection ofthe plant material in the greenhouse or on the field. If anaccession could be assigned to another species accordingto AFLP pattern and morphology, it was given the name ofthis species, if there were any doubts on the identificationthe species was given the label S. spec.. The accessions
which labels were changed are indicated in Additional file1.
AFLPThe samples were fingerprinted with two EcoRI/MseI AFLPprimer combinations: E32/M49 and E35/M48. The proto-col of Vos et al. [33] was used to generate AFLP fragments.Primer combination E32/M49 yielded 91 polymorphicbands and primer combination E35/M48 yielded 131bands. Keygene carried out the AFLP analysis on a Mega-BACE 2.1 and scored the bands using their proprietarysoftware. Bands were scored as dominant markers, so onlythe presence or the absence of a band was scored.
DatasetsThe dataset in this study originally contained 4929 geno-types. This large dataset was analyzed with NJ andUPGMA. Because of the size of the dataset, it provedimpossible to analyze it with cladistic methods nor toanalyze it for statistical support, even using the SARAsupercomputer (see below). It was sheer impossible for apersonal computer to do any further analyses apart fromthe NJ and UPGMA, and for the SARA computer cluster itwould have taken many months/years of computing time.
For further analysis a condensed dataset was created bycarefully choosing a representative genotype from all theavailable accessions. This condensed dataset consisted of916 genotypes. The condensed dataset was used in bothphenetic and cladistic analyses and in the resamplingmethods.
Besides choosing only one genotype per accession to rep-resent the accession in the condensed dataset, otheradjustments were made to create this dataset. All the 22known interspecific hybrid accessions were removed, 23other accessions were completely removed because of theextreme heterogeneity of the accession (possibly resultingfrom a mixture of species) in both the NJ and the UPGMAtrees. Species labels of 49 accessions were changed basedon their position in the NJ and/or UPGMA tree (notshown) and visual inspection of the plants in the experi-mental field or greenhouse in 2005 and 2006. In total 11outgroup accessions were removed because preliminaryAFLP results showed these outgroups to be too distant (S.sitiens, S. nigrum, S. chaparense, S. lycopersicoides, S. can-ense, S. fraxinifolium). The outgroup species S. etuberosum,S. palustre and S. fernandezianum were retained in the data-set.
Data analysisBoth the phenetic and the cladistic analyses were con-ducted using PAUP 4.0 Altivec [34] on the TERAS comput-ing cluster of SARA computing facilities in Amsterdam.For the 4929 phenetic analysis we used the total character
Page 10 of 12(page number not for citation purposes)

BMC Evolutionary Biology 2008, 8:145 http://www.biomedcentral.com/1471-2148/8/145
distance, for the 916 data set we used the NeiLi distance[35] to calculate the distance matrix. A Neighbor JoiningJackknife tree was calculated using 10.000 replicates.
The cladistic analysis heuristic searches were done byusing PRAP, Parsimony Ratchet Analyses using PAUP, aprogram that writes commands for PAUP. The commandsin PRAP describe how PAUP should carry out parsimonyratchet searches [36]. By using parsimony ratchet, asdescribed by Nixon [37], many tree islands are searchedinstead of thoroughly searching through each island.
For the MP jackknife analysis, we followed the conclu-sions drawn by Muller [36] that using random additionsequence instead of simple addition sequence has no ben-eficial effect on bootstrap or jackknife support. Also, ajackknife or bootstrap analysis using one heuristic searchsaving one tree per jackknife replicate and simple additionsequence, performed as good as or even better than ananalysis using 10 parsimony ratchet iterations using theshortest tree only or using a strict consensus tree of allshortest trees [36]. Therefore, we conducted a jackknifeMP analysis by performing 10.000 replicates using simpleaddition, and saving one shortest tree per replicate.
Authors' contributionsMJ carried out the phenetic and phylogenetic analyses anddrafted the manuscript, RVB and BV participated in coor-dination and design of the study and participated in draft-ing the manuscript, RH chose the plant material andparticipated in coordination and design of the study, VVand MV coordinated and carried out the growing of thematerial, MS and RM carried out the AFLP analyses. Allauthors read and approved of the final manuscript.
Additional material
AcknowledgementsThis project was (co)financed by the research programme of the Centre of Biosystems Genomics (CBSG) which is part of the Netherlands Genomics Initiative/Netherlands Organization for Scientific Research.
We thank SARA Computing and Networking Services in Amsterdam for the use and help with running the analyses on the TERAS computing cluster. The computing time and services at SARA were financed by a grant for pilot projects of the programme Scientific use Supercomputers of the Nether-lands Organization for Scientific Research, (NWO).
The acronym AFLP is a registered trademark (AFLP®) of Keygene N.V. and the AFLP® technology is covered by patents and patent applications of Key-Gene N.V.
References1. Correll DS: The potato and its wild relatives. In Contributions
from the Texas Research Foundation 4 Texas Research Foundation, Ren-ner, Texas; 1962.
2. Hawkes JG: The Potato, Evolution, Biodiversity and Genetic Resources Lon-don: Belhaven Press; 1990.
3. Spooner DM, Hijmans RJ: Potato systematics and germplasmcollecting, 1989–2000. Amer J of Potato Res 2001, 78:237-268.
4. Spooner DM, Salas A: Structure, biosystematics and geneticresources. In Handbook of Potato, production, improvement and post-harvest management Edited by: Gopal J, Khurana SMP. New York: TheHaworth Press; 2006:1-39.
5. Berg RG Van den, Jacobs MMJ: Molecular Taxonomy. Potato Biol-ogy and Biotechnology 2007:55-76.
6. Giannattasio RG, Spooner DM: A reexamination of speciesboundaries and hypotheses concerning Solanum megistac-rolobum and S. toralapanum (Solanum sect. Petota seriesMegistacroloba): Molecular data. Syst Bot 1994, 19:106-115.
7. Berg RG Van den, Miller JI, Ugarte ML, Kardolus JP, Villand J, NienhuisJ, Spooner DM: Collapse of morphological species in the wildpotato Solanum brevicaule complex (Solanaceae: sect.Petota). Am J Bot 1998, 85:92-109.
8. Berg RG Van den, Groendijk-Wilders N: Numerical analysis ofthe taxa of series Circaeifolia (Solanum sect. Petota). In Pro-ceedings of the International Conference Solanaceae IV: July 1994, AdelaideEdited by: Nee M, Symon DE, Lester RN, Jessop JP. The Royal Botan-ical gardens, Kew; 1999:213-226.
9. Kardolus JP: A biosystematic analysis of Solanum acaule. In PhDThesis Wageningen University; 1998.
10. Spooner DM, Berg RG Van den, Rodríguez A, Bamberg J, Hijmans RJ,Lara-Cabrera SI: Wild Potatoes (Solanum section Petota;Solanaceae) of North and Central America. In Systematic Bot-any Monographs Edited by: McPherson GD, Prather LA, Ranker TA,Reznicek AA. USA: The American Society of Plant Taxonomists;2004.
11. Spooner DM, Sytsma KJ: Reexamination of Series Relationshipsof Mexican and Central-American Wild Potatoes (SolanumSect Petota) – Evidence from Chloroplast DNA RestrictionSite Variation. Syst Bot 1992, 17:432-448.
12. Castillo RO, Spooner DM: Phylogenetic relationships of wildpotatoes, Solanum series Conicibaccata (Sect. Petota). Syst Bot1997, 22:45-83.
13. Rodríguez A, Spooner DM: Chloroplast DNA analysis of Sola-num bulbocastanum and S. cardiophyllum, and evidence forthe distinctiveness of S. cardiophyllum subsp. ehrenbergii(sect. Petota). Syst Bot 1997, 22:31-43.
14. Spooner DM, Castillo RT: Reexamination of series relationshipsof South American wild potatoes (Solanaceae : Solanum Sect.Petota): Evidence from chloroplast DNA restriction site var-iation. Am J B 1997, 84:671-685.
15. Despres L, Gielly L, Redoutet W, Taberlet P: Using AFLP toresolve phylogenetic relationships in a morphologicallydiversified plant species complex when nuclear and chloro-plast sequences fail to reveal variability. Mol Phylogenet Evol2003, 27:185-196.
16. Koopman WJM: Phylogenetic signal in AFLP Data Sets. Syst Biol2005, 54:197-217.
Additional file 1List of material used in the AFLP analyses. List of material used in the AFLP analyses. Symbols used in additional file 1. # recorded hybrid, removed in 916 dataset. $ complete accession removed in the 916 dataset because of conflicting positions in NJ tree. & removed outgroups in 916 dataset: S. lycopersicoides, S. nigrum, S. chaparense, S. sitiens, S. canense, S. fraxinifolium. * the label of this accession was changed in the 916 dataset after checking the position in the large NJ tree and check-ing morphology in the greenhouse/field. () the number in parentheses indicates the number of accessions used for the 916 analysis in case of removal or change of accessions. Abbreviations for Genebank source codes: CPC: Commonwealth Potato Collection, UK. CGN: Centre for Genetic Resources, the Netherlands. cgn: cgn receipt number, Centre for Genetic Resources, the Netherlands. PI: Plant Introduction number, USA. GLKS: Gross Lusewitz, Germany.Click here for file[http://www.biomedcentral.com/content/supplementary/1471-2148-8-145-S1.pdf]
Page 11 of 12(page number not for citation purposes)

BMC Evolutionary Biology 2008, 8:145 http://www.biomedcentral.com/1471-2148/8/145
Publish with BioMed Central and every scientist can read your work free of charge
"BioMed Central will be the most significant development for disseminating the results of biomedical research in our lifetime."
Sir Paul Nurse, Cancer Research UK
Your research papers will be:
available free of charge to the entire biomedical community
peer reviewed and published immediately upon acceptance
cited in PubMed and archived on PubMed Central
yours — you keep the copyright
Submit your manuscript here:http://www.biomedcentral.com/info/publishing_adv.asp
BioMedcentral
17. Meudt HM, Clarke AC: Almost Forgotten or latest Practice?AFLP applications, analyses and advances. Trends in Plant Sci-ence 2007, 12:106-117.
18. Jones CJ, Edwards KJ, Castaglione S, Winfield MO, Sala F, Wiel C vande, Bredemeijer G, Vosman B, Matthes M, Daly A, Brettschneider R,Bettini P, Buiatti M, Maestri E, Malcevschi A, Marmiroli N, Aert E, Vol-ckaert G, Rueda J, Linacero R, Vazquez A, Karp A: Reproducibilitytesting of RAPD, AFLP and SSR markers in plants by a net-work of European laboratories. Mol Breed 1997, 3:381-390.
19. Kardolus JP, van Eck HJ, Berg RG van den: The potential of AFLPsin biosystematics: a first application in Solanum taxonomy.Plant Systematics and Evolution 1998, 210:87-103.
20. Koopman WJM, Gort G: Significance Tests and Weighted Val-ues for AFLP Similarities, Based on Arabidopsis in SilicoAFLP Fragment Length Distributions. Genetics 2004,167:1915-1928.
21. Berg RG van den, Bryan G, del Rio A, Spooner DM: Reduction ofspecies in the wild potato Solanum section Petota series Lon-gipedicellata : AFLP, RAPD and chloroplast SSR data. TAG2002, 105:1109-1114.
22. Lara-Cabrera SI, Spooner DM: Taxonomy of North and CentralAmerican diploid wild potato (Solanum sect. Petota) species:AFLP data. Plant Syst Evol 2004, 248:129-142.
23. McGregor CE, van Treuren E, Hoekstra R, van Hintum Th JL: Anal-ysis of the wild potato germplasm of the series Acaulia withAFLPs: implications for ex situ conservation. TAG 2002,104:146-156.
24. Spooner DM, McLean K, Ramsay G, Waugh R, Bryan GJ: A singledomestication for potato based on multilocus amplified frag-ment length polymorphism genotyping. PNAS 2005,102:14694-14699.
25. Hawkes JG, Jackson MT: Taxonomic and evolutionary implica-tions of the Endosperm Balance Number hypothesis in pota-toes. TAG 1992, 84:180-185.
26. Volkov RA, Komarovs NY, Panchuk II, Hemleben V: Molecular evo-lution of rDNA external transcribed spacer and phylogeny ofsect. Petota (genus Solanum). Mol Phylogenet Evol 2003,29:187-202.
27. Hughes C, Eastwood R: Island radiation on a continental scale:Exceptional rates of plant diversification after uplift of theAndes. PNAS 2006, 103:10334-10339.
28. Whalen MD: Conspectus of species groups in Solanum subge-nus Leptostemonum. Gentes Herbarum 1984, 12:179-282.
29. Knapp S: A revision of Solanum thelopedium species group(section Anthoresis sensu Seithe, pro parte): Solanaceae.Bull Nat Hist Mus Lond 2000, 30:13-30.
30. Knapp S: A revision of the Solanum sessile species group (sec-tion Geminata pro parte: Solanaceae). Bot J of Linn Soc 1991,105:179-210.
31. Murashigi T, Skoog F: A revised medium for rapid growth andbio-assay with tobacco tissue cultures. Physiol Plant 1962,15:473-497.
32. Stewart CNJ, Via LE: A rapid CTAB DNA isolation techniqueuseful for RAPD fingerprinting and other PCR applications.Biotechniques 1993, 14:748-750.
33. Vos P, Hogers R, Bleeker M, Reijans M, Lee T Van de, Hornes M, Fri-jters A, Pot J, Peleman J, Kuiper M, Zabeau M: AFLP: A new tech-nique for DNA fingerprinting. Nucl Acids Res 1995,23:4407-4414.
34. Swofford DL: PAUP*4.0 Beta, Phylogenetic analysis UsingParsimony (* and other methods). In Software program Sinauerassociates, Sunderland, Massachusetts; 2002.
35. Nei M, Li WH: Mathematical model for studying genetic vari-ation in terms of restriction endonucleases. Proc Natl Acad Sci(USA) 1979, 76:5269-5273.
36. PRAP – parsimony ratchet analyses using PAUP* [http://www.nees.uni-bonn.de/downloads/PRAP]
37. Nixon KC: Parsimony Ratchet: A New Method for Rapid Par-simony Analysis. Cladistics 1999, 15:407-414.
Page 12 of 12(page number not for citation purposes)
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=8512694
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=8512694
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=7501463




















