
Jasmonates act positively in adventitious ... - BMC Plant Biology
10
RESEARCH ARTICLE Open Access Jasmonates act positively in adventitious root formation in petunia cuttings Sandra Lischweski 1,2 , Anne Muchow 1,3 , Daniela Guthörl 1,4 and Bettina Hause 1* Abstract Background: Petunia is a model to study the process of adventitious root (AR) formation on leafy cuttings. Excision of cuttings leads to a transient increase in jasmonates, which is regarded as an early, transient and critical event for rooting. Here, the role of jasmonates in AR formation on petunia cuttings has been studied by a reverse genetic approach. Results: To reduce the endogenous levels of jasmonates, transgenic plants were generated expressing a Petunia hybrida ALLENE OXIDE CYCLASE (PhAOC)-RNAi construct. The transgenic plants exhibited strongly reduced PhAOC transcript and protein levels as well as diminished accumulation of cis-12-oxo-phytodienoic acid, jasmonic acid and jasmonoyl-isoleucine after wounding in comparison to wild type and empty vector expressing plants. Reduced levels of endogenous jasmonates resulted in formation of lower numbers of ARs. However, this effect was not accompanied by altered levels of auxin and aminocyclopropane carboxylate (ACC, precursor of ethylene) or by impaired auxin and ethylene-induced gene expression. Neither activity of cell-wall invertases nor accumulation of soluble sugars was altered by jasmonate deficiency. Conclusions: Diminished numbers of AR in JA-deficient cuttings suggest that jasmonates act as positive regulators of AR formation in petunia wild type. However, wound-induced rise in jasmonate levels in petunia wild type cuttings seems not to be causal for increased auxin and ethylene levels and for sink establishment. Keywords: Adventitious root formation, Auxin, Cell wall invertase, Cuttings, Ethylene, Jasmonic acid, JA-isoleucine, Petunia hybrida Background Adventitious root (AR) formation in leafy stem cuttings is a crucial process for the propagation of many ornamental plant species. Vegetative propagation of plants relies on the ability of shoot cuttings to effectively generate such roots. There is, however, a dramatic variation between species in their capability to form AR [1]. During AR formation roots develop from non-root tissue, mostly aerial plant parts such as hypocotyls, leaves and stems [2]. The origin of ARs varies among plant species and organ or tissue they develop from [3]. ARs always develop from cells neighboring vascular tissues and can initiate from hypocotyl pericycle cells, phloem or xylem parenchyma cells, young secondary phloem cells, or interfascicular cambium cells. In Arabidopsis thaliana, a model often used to investigate AR formation, rooting is induced by pre-etiolation of intact seedlings. After supply with sugars or hormones or after transfer into light they develop roots at the intact hypocotyl [4–7]. In this model system, AR formation occurs without stresses that disrupt root-shoot correlative influences [4]. This is in contrast to leafy cuttings, which are subjected to severance from the donor plant accompanied by injury and the isolation from func- tional integrity of the whole plant conditions [8]. In both, pre-etiolated seedlings and leafy cuttings, AR formation is a complex process influenced by multiple endogenous and exogenous factors, including phytohormones, light, wounding, and stress [2]. Among the endogenous factors, the phytohormones are the most important modulators of AR formation—it has become obvious that auxin and ethylene (ET) play a central role, but they interact with * Correspondence: [email protected] 1 Department of Cell and Metabolic Biology, Leibniz Institute of Plant Biochemistry, Weinberg 3, D06120 Halle/Salle, Germany Full list of author information is available at the end of the article © 2015 Lischweski et al. Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated. Lischweski et al. BMC Plant Biology (2015) 15:229 DOI 10.1186/s12870-015-0615-1
-
Upload
khangminh22 -
Category
Documents
-
view
3 -
download
0
Transcript of Jasmonates act positively in adventitious ... - BMC Plant Biology

Lischweski et al. BMC Plant Biology (2015) 15:229 DOI 10.1186/s12870-015-0615-1
RESEARCH ARTICLE Open Access
Jasmonates act positively in adventitiousroot formation in petunia cuttings
Sandra Lischweski1,2, Anne Muchow1,3, Daniela Guthörl1,4 and Bettina Hause1*Abstract
Background: Petunia is a model to study the process of adventitious root (AR) formation on leafy cuttings. Excisionof cuttings leads to a transient increase in jasmonates, which is regarded as an early, transient and critical event forrooting. Here, the role of jasmonates in AR formation on petunia cuttings has been studied by a reverse geneticapproach.
Results: To reduce the endogenous levels of jasmonates, transgenic plants were generated expressing a Petuniahybrida ALLENE OXIDE CYCLASE (PhAOC)-RNAi construct. The transgenic plants exhibited strongly reduced PhAOCtranscript and protein levels as well as diminished accumulation of cis-12-oxo-phytodienoic acid, jasmonic acid andjasmonoyl-isoleucine after wounding in comparison to wild type and empty vector expressing plants. Reducedlevels of endogenous jasmonates resulted in formation of lower numbers of ARs. However, this effect was notaccompanied by altered levels of auxin and aminocyclopropane carboxylate (ACC, precursor of ethylene) or byimpaired auxin and ethylene-induced gene expression. Neither activity of cell-wall invertases nor accumulation ofsoluble sugars was altered by jasmonate deficiency.
Conclusions: Diminished numbers of AR in JA-deficient cuttings suggest that jasmonates act as positive regulatorsof AR formation in petunia wild type. However, wound-induced rise in jasmonate levels in petunia wild typecuttings seems not to be causal for increased auxin and ethylene levels and for sink establishment.
Keywords: Adventitious root formation, Auxin, Cell wall invertase, Cuttings, Ethylene, Jasmonic acid, JA-isoleucine,Petunia hybrida
BackgroundAdventitious root (AR) formation in leafy stem cuttings isa crucial process for the propagation of many ornamentalplant species. Vegetative propagation of plants relies onthe ability of shoot cuttings to effectively generate suchroots. There is, however, a dramatic variation betweenspecies in their capability to form AR [1]. During ARformation roots develop from non-root tissue, mostlyaerial plant parts such as hypocotyls, leaves and stems [2].The origin of ARs varies among plant species and organor tissue they develop from [3]. ARs always develop fromcells neighboring vascular tissues and can initiate fromhypocotyl pericycle cells, phloem or xylem parenchyma
* Correspondence: [email protected] of Cell and Metabolic Biology, Leibniz Institute of PlantBiochemistry, Weinberg 3, D06120 Halle/Salle, GermanyFull list of author information is available at the end of the article
© 2015 Lischweski et al. Open Access This arInternational License (http://creativecommonsreproduction in any medium, provided you gthe Creative Commons license, and indicate if(http://creativecommons.org/publicdomain/ze
cells, young secondary phloem cells, or interfascicularcambium cells. In Arabidopsis thaliana, a model oftenused to investigate AR formation, rooting is induced bypre-etiolation of intact seedlings. After supply with sugarsor hormones or after transfer into light they develop rootsat the intact hypocotyl [4–7]. In this model system, ARformation occurs without stresses that disrupt root-shootcorrelative influences [4]. This is in contrast to leafycuttings, which are subjected to severance from the donorplant accompanied by injury and the isolation from func-tional integrity of the whole plant conditions [8]. In both,pre-etiolated seedlings and leafy cuttings, AR formation isa complex process influenced by multiple endogenousand exogenous factors, including phytohormones, light,wounding, and stress [2]. Among the endogenous factors,the phytohormones are the most important modulators ofAR formation—it has become obvious that auxin andethylene (ET) play a central role, but they interact with
ticle is distributed under the terms of the Creative Commons Attribution 4.0.org/licenses/by/4.0/), which permits unrestricted use, distribution, andive appropriate credit to the original author(s) and the source, provide a link tochanges were made. The Creative Commons Public Domain Dedication waiverro/1.0/) applies to the data made available in this article, unless otherwise stated.

Lischweski et al. BMC Plant Biology (2015) 15:229 Page 2 of 10
one another; with other phytohormones and with environ-mental cues in complex networks [3].For Petunia hybrida, a model plant for AR develop-
ment on leafy cuttings, a three-phase mechanism waspostulated for the metabolic responses involved in ARformation [9] consisting of (i) establishment of a sink,(ii) a recovery phase and of (iii) a maintenance phase. Inthe first phase, the excision of cuttings leads to rapidand transient increase in the wound-phytohormonejasmonic acid (JA) and a continuous accumulation ofsoluble and insoluble carbohydrates [9]. It was hypothe-sized that wounding accompanied by the rise in JAinitiates the establishment of a sink tissue necessary tofacilitate subsequent AR formation. With that, wound-induced JA accumulation at the cutting stem base hasbeen regarded as an early, transient and critical event forrooting of Petunia cuttings. Remarkably, in Arabidopsisintact hypocotyls jasmonates negatively regulate adventi-tious rooting, and their homeostasis is under control ofauxin [10].Jasmonates are ubiquitously occurring signaling com-
pounds in plants and accumulate in response to biotic andabiotic stress as well as in development [11]. JA and itsmolecularly active metabolite (+)-7-iso-jasmonoyl isoleu-cine (JA-Ile) are lipid-derived compounds and are synthe-sized from α-linolenic acid by one of seven differentbranches of the lipoxygenase (LOX) pathway [12, 13].LOX and the two following enzymatic steps are located inthe plastids and involve the action of an ALLENE OXIDESYNTHASE and an ALLENE OXIDE CYCLASE (AOC)leading to formation of the intermediate cis-12-oxo-phyto-dienoic acid (OPDA). Further reactions occur in peroxi-somes and form JA, which is enzymatically converted toJA-Ile within the cytosol. JA-Ile was demonstrated to me-diate binding of the co-receptor proteins CORONATINEINSENSITIVE1 (COI1) and JASMONATE ZIM DO-MAIN (JAZ), thereby triggering JA responses [14–17].Within the JA pathway the AOC-catalyzed step is
regarded as the crucial step as here the exclusive forma-tion of the enantiomeric form occurring in naturalcyclopentanones like JA and JA-Ile is facilitated [18, 19].In petunia, the AOC is constitutively present in stem tis-sue, and has constantly high activity [9]. Additionally,PhAOC belongs to the wound-induced genes, and itstranscript levels increase following endogenous rise inJA/JA-Ile [9].To study the role of JA in AR formation in leafy cut-
tings of petunia by a reverse genetic approach, thecDNA of PhAOC was used to generate an RNAi con-struct for the stable transformation of plants to partiallysuppress PhAOC expression. Our data clearly show thatthis suppression markedly decreases the accumulation ofjasmonates as tested in leaves after mechanical wound-ing. Additionally, suppression of PhAOC caused a delay
in AR formation. To get insights into how diminishedJA levels might affect AR formation, stem bases of cut-tings from transgenic plants were used to comparativelyanalyze levels of IAA and aminocyclopropane carboxyl-ate (ACC, precursor of ET), but also transcript levels ofa cell wall invertase (CWI) encoding gene as well assugar contents.
Results and discussionAmong the lipid-derived compounds, octadecanoids andjasmonates have a crucial role in plant responses to bi-otic and abiotic stresses, but also in developmental pro-cesses [20]. JA was suggested to be one of severalendogenous factors regulating the formation of AR [4].For petunia cuttings, their excision is characterized by afast and transient increase in the level of JA followed byinduction of genes encoding JA biosynthesis genes andproteins involved in sink establishment, such as cell wallinvertases (CWIs) [9, 21].
Generation of transgenic plants with reduced jasmonatelevelsTo elucidate the role of jasmonates in formation of ARin petunia, the endogenous levels of jasmonates were re-duced by a transgenic approach. Transgenic plants exhi-biting an impaired JA biosynthesis were generated bythe RNAi-mediated knock-down of the expression of thegene encoding AOC. AOC of petunia is encoded by asingle copy gene (Additional file 1: Figure S1). A 180-bpfragment covering a middle part of the PhAOC-codingregion was used to generate the PhAOC-RNAi construct,which was introduced into petunia plants via A. tumefa-ciens mediated transformation. The expression ofPhAOC-RNAi is under control of the cauliflower mosaicvirus 35S promoter, thereby conferring constitutive ex-pression in all plant tissues. T1 plants were grown to-gether with wild-type plants and plants expressing theempty vector pHellsgate (pHell) as controls. Three trans-genic PhAOC–RNAi lines exhibiting significant reduc-tion of PhAOC transcripts (Fig. 1a) were selected. ThePhAOC–RNAi lines 4, 10 and 15 exhibited residualPhAOC transcript levels between 9 and 17 % in compari-son to the controls. This reduced level of PhAOC-tran-script caused a reduction of AOC protein contents toundetectable amounts as shown by immunoblot analyses(Additional file 1: Figure S2). As visualized by theimmuno cytological analysis, in wild type stems AOC islocated within plastids of internal phloem cells as well asof xylem parenchyma cells of the amphiphloic siphono-stele (Fig. 1b, upper micrograph).I In the PhAOC–RNAiline, however, the protein amount is below the detectionlimit (Fig. 1b, lower micrograph). Consequently, cuttingsof the PhAOC-RNAi lines exhibited a significant reduction
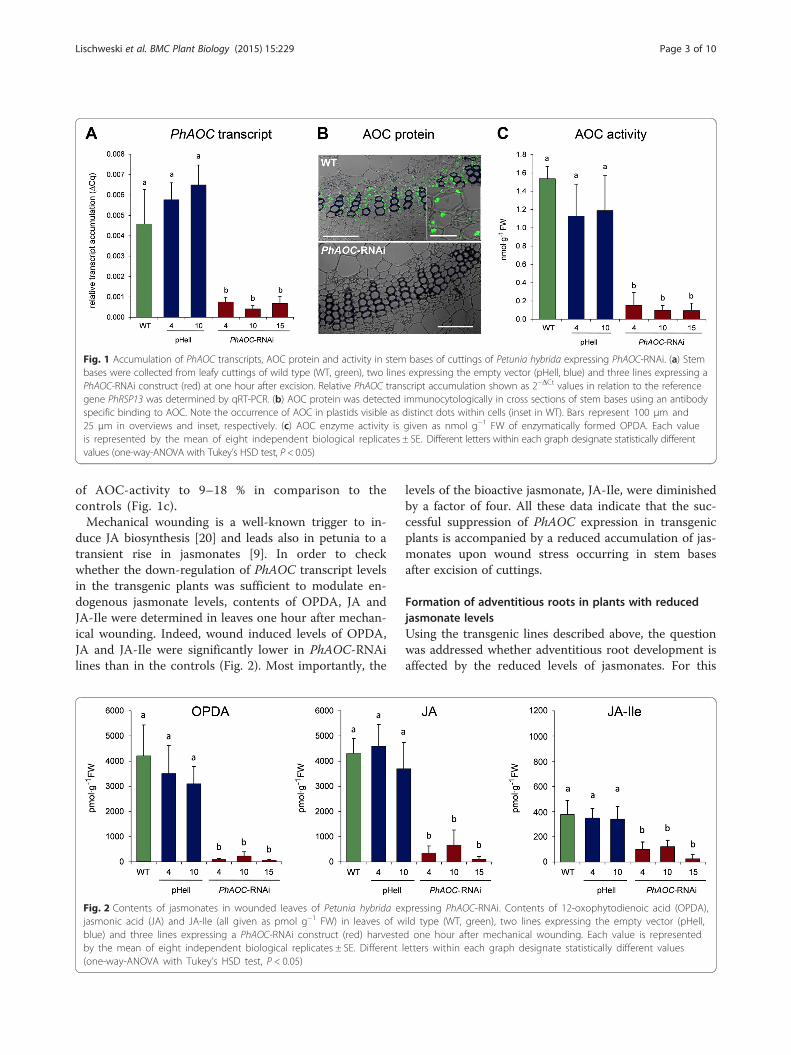
Fig. 1 Accumulation of PhAOC transcripts, AOC protein and activity in stem bases of cuttings of Petunia hybrida expressing PhAOC-RNAi. (a) Stembases were collected from leafy cuttings of wild type (WT, green), two lines expressing the empty vector (pHell, blue) and three lines expressing aPhAOC-RNAi construct (red) at one hour after excision. Relative PhAOC transcript accumulation shown as 2−ΔCt values in relation to the referencegene PhRSP13 was determined by qRT-PCR. (b) AOC protein was detected immunocytologically in cross sections of stem bases using an antibodyspecific binding to AOC. Note the occurrence of AOC in plastids visible as distinct dots within cells (inset in WT). Bars represent 100 μm and25 μm in overviews and inset, respectively. (c) AOC enzyme activity is given as nmol g−1 FW of enzymatically formed OPDA. Each valueis represented by the mean of eight independent biological replicates ± SE. Different letters within each graph designate statistically differentvalues (one-way-ANOVA with Tukey’s HSD test, P < 0.05)
Lischweski et al. BMC Plant Biology (2015) 15:229 Page 3 of 10
of AOC-activity to 9–18 % in comparison to thecontrols (Fig. 1c).Mechanical wounding is a well-known trigger to in-
duce JA biosynthesis [20] and leads also in petunia to atransient rise in jasmonates [9]. In order to checkwhether the down-regulation of PhAOC transcript levelsin the transgenic plants was sufficient to modulate en-dogenous jasmonate levels, contents of OPDA, JA andJA-Ile were determined in leaves one hour after mechan-ical wounding. Indeed, wound induced levels of OPDA,JA and JA-Ile were significantly lower in PhAOC-RNAilines than in the controls (Fig. 2). Most importantly, the
Fig. 2 Contents of jasmonates in wounded leaves of Petunia hybrida exjasmonic acid (JA) and JA-Ile (all given as pmol g−1 FW) in leaves of wblue) and three lines expressing a PhAOC-RNAi construct (red) harvesteby the mean of eight independent biological replicates ± SE. Different(one-way-ANOVA with Tukey’s HSD test, P < 0.05)
levels of the bioactive jasmonate, JA-Ile, were diminishedby a factor of four. All these data indicate that the suc-cessful suppression of PhAOC expression in transgenicplants is accompanied by a reduced accumulation of jas-monates upon wound stress occurring in stem basesafter excision of cuttings.
Formation of adventitious roots in plants with reducedjasmonate levelsUsing the transgenic lines described above, the questionwas addressed whether adventitious root development isaffected by the reduced levels of jasmonates. For this
pressing PhAOC-RNAi. Contents of 12-oxophytodienoic acid (OPDA),ild type (WT, green), two lines expressing the empty vector (pHell,d one hour after mechanical wounding. Each value is representedletters within each graph designate statistically different values

Lischweski et al. BMC Plant Biology (2015) 15:229 Page 4 of 10
purpose, leafy cuttings of all plant lines were transferredwithout any external additives to Perlite as neutral sub-strate. After 7, 14, and 21 days after excision (dpe) num-bers of root primordia and emerged adventitious rootswere determined (Fig. 3). Adventitious roots were notyet emerging from stem bases of all lines analyzed at 7dpe. However, significantly fewer root primordia weredetected in the stem base of cuttings of PhAOC-RNAiplants in comparison to wild type and the empty vectorcontrol plants (Fig. 3a). Numbers of primordia ofPhAOC-RNAi lines were reduced by 70–90 %. This ef-fect on root primordia was not detectable anymore atthe two later time points analyzed. At 14 dpe, adventi-tious roots emerged from all cuttings (Fig. 3b). Here,cuttings of PhAOC-RNAi plants developed significantlyfewer roots than cuttings of the controls: Root numbersof cuttings of PhAOC-RNAi line 4 and 15 were reduced
Fig. 3 Effect of down-regulation of PhAOC on rooting of Petunia hybrida cucounting primordia (a) and emerged roots (b) in cuttings of wild type (WTlines expressing a PhAOC-RNAi construct (red). Each value is represented byeach of three technical replicates (single cuttings). Different letters within eTukey’s HSD test, P < 0.05)
to 70 %, whereas the root number of cuttings of thePhAOC-RNAi line 10 was reduced to 50 % of that of thecontrols. The reduced number of roots emerged fromcuttings of PhAOC-RNAi plants was still visible at 21dpe, although to a lesser extent (Fig. 3). These data sug-gest that AR formation is delayed in cuttings from plantsexhibiting decreased JA levels resulting in a decreasednumber of roots at 14 and 21 dpe.These data show that JA deficiency resulted in lower
numbers of ARs, but the AR formation itself was notabolished. Therefore, wound-induced jasmonates mightfunction as an accelerator of AR formation in petuniaand might act therefore as positive regulators. This is inline with the proposed role of JA in AR formation in pe-tunia and in other species rooted under in vitro condi-tions. For potato (Solanum tuberosum) [22] and pea(Pisum sativum) [23] stem cuttings, and tobacco
ttings. Rooting was rated at 7, 14 and 21 days post excision (dpe) by, green), two lines expressing the empty vector (pHell, blue) and threethe mean of eight independent biological replicates (± SE) consistingach graph designate statistically different values (one-way-ANOVA with

Lischweski et al. BMC Plant Biology (2015) 15:229 Page 5 of 10
(Nicotiana tabacum) thin cell layers [24] it was shownthat in the presence of exogenous auxin, JA promotessynergistically AR formation. However, these data contrastwith data obtained on AR formation in Arabidopsis intacthypocotyls, where JA inhibits AR initiation through theCOI1 signaling pathway [10]. These apparent contradic-tions suggest that there might be either species-specificdifferences between Arabidopsis and other dicots in re-spect to AR formation or organ and cultivation-specificdifferences that may alter the balance between auxin andJA regulating AR formation [3]. Indeed, intact seedlings,de-rooted older plants and cuttings show significantdifferences, not only in root founding tissues, but also inauxin requirements, sensitivity, and rooting mutant phe-notypes [25]. This holds also true for de-rooted seedlingsof petunia, which showed reduced numbers of ARs aftertreatment with jasmonates (Additional file 2: Table S1).Treatments with OPDA, JA and JA-Ile at lower concen-trations (0.1–1.0 μM) did not changed AR numbers.Higher concentrations (10 μM) of all compounds, how-ever, significantly reduced the root number, whereas100 μM JA even completely inhibited AR formation. Incontrast to jasmonate treatments, de-rooted seedlingstreated with the auxin 2,4-dichlorophenoxy acetic acid(2,4-D) or ACC developed significantly more ARs thancontrol plants (Additional file 2: Table S1). Combinedapplication of JA and either 2,4-D or ACC showedthat both, 2.4-D and ACC, suppressed the inhibitoryeffect of JA on development of ARs, and JA did notdiminished the promoting effects of 2.4-D and ACC(Additional file 2: Table S2).
Effects of diminished jasmonate levels on role of auxinand ethylene in root formationIt is well accepted that ET and auxins play an importantrole in stimulating the process of AR formation throughthe different phases [4]. Therefore, the levels of IAA andACC, the precursor of ET, were determined in JA-deficient cuttings. In parallel, auxin-induced and ET-induced gene expression was monitored by determin-ation of transcript levels of a petunia GH3 homologue(PhDevA-20-C01) and ACC oxidase1 (PhACO1), respect-ively. In wild type cuttings, free IAA levels increasedtransiently about two-fold at 2 and 24 hpe followed byincreased transcript accumulation of PhDevA-20-C01(Additional file 1: Figure S3), whereas ACC levels in-creased about 200-fold within 24 hpe [21]. Therefore,free IAA and ACC levels as well as transcript accumula-tion of PhDevA-20-C01 and PhACO1 were measured instem bases of cuttings at 24 and 48 hpe, respectively(Fig. 4). There were no differences in free IAA contentand PhDevA-20-C01 transcript levels between PhAOC-RNAi lines and the controls (Fig. 4a, b). Similar resultswere obtained in respect to ACC levels and transcript
accumulation of the ET responsive gene PhACO1(Fig. 4c, d), which both also did not show significantdifferences between JA-deficient plants and wild type/transformation control.These results showed that diminished JA levels did not
change the levels and action of both, auxin and ET incuttings, although auxin, ET, and JA were found to becoordinately (cooperatively or antagonistically) regulatedor exhibit opposite effects on many plant responses [20].Auxin is considered as inductor of AR formation in leafycuttings – after detachment of the shoot, basipetal polartransport of auxin contributes to auxin accumulation inthe stem base [26]. The rise of free auxin in the basalstem very probably contributes to the early events of ARformation [1, 21, 27, 28]. Additionally, AR formation isalso dependent on the action of ET [29, 30], which issynthesized after wounding during the cutting process[27]. It is tempting to speculate that in the process ofAR formation JA acts independently from auxin and ET,since diminished JA levels in the transgenic cuttings de-layed the formation of AR without changing auxin andET levels and signaling. It cannot completely be ex-cluded that alterations in hormone levels and relatedgene expression might occur at other time points thanthose analyzed. This is, however, not very probable, sincethe time points used represent in each case the timepoint of the transient maximum of the respective hor-mone/transcript level in wild type cuttings (Additionalfile 1: Figure S3 and [21]).
Effects of diminished jasmonate levels on cell wallinvertase and sugar levels during root formationCWI is a key enzyme of the apoplastic phloem unload-ing of transported sucrose and links jasmonate actionwith primary metabolism [31]. It can establish a sinkfunction of a certain tissue and thus provide a mechan-ism for flexible and appropriate adjustment to a widerange of internal and external stimuli [32]. To test,whether altered jasmonate levels might affect the induc-tion of the gene encoding CWI and thereby influencingthe establishment of the sink [9], transcript levels of3CL9414 and CWI activity were determined at 4 hpeand 6 hpe, respectively (Table 1). Both time points repre-sent the respective maximum after excision of wild typecuttings [9]. Comparing cutting stem bases of PhAOC-RNAi with that of wild type and empty vectors controls,there were no differences in 3CL9414 transcript levelsand CWI activity. Additionally, the levels of glucose,fructose and sucrose were determined. The levels ofthese three compounds reached maximum levels in wildtype cuttings at 192 hpe, the time point at which firstdifferentiating root primordia are detectable within thestem base [9]. At this time point the significant reduc-tion in the number of root primordia was found for
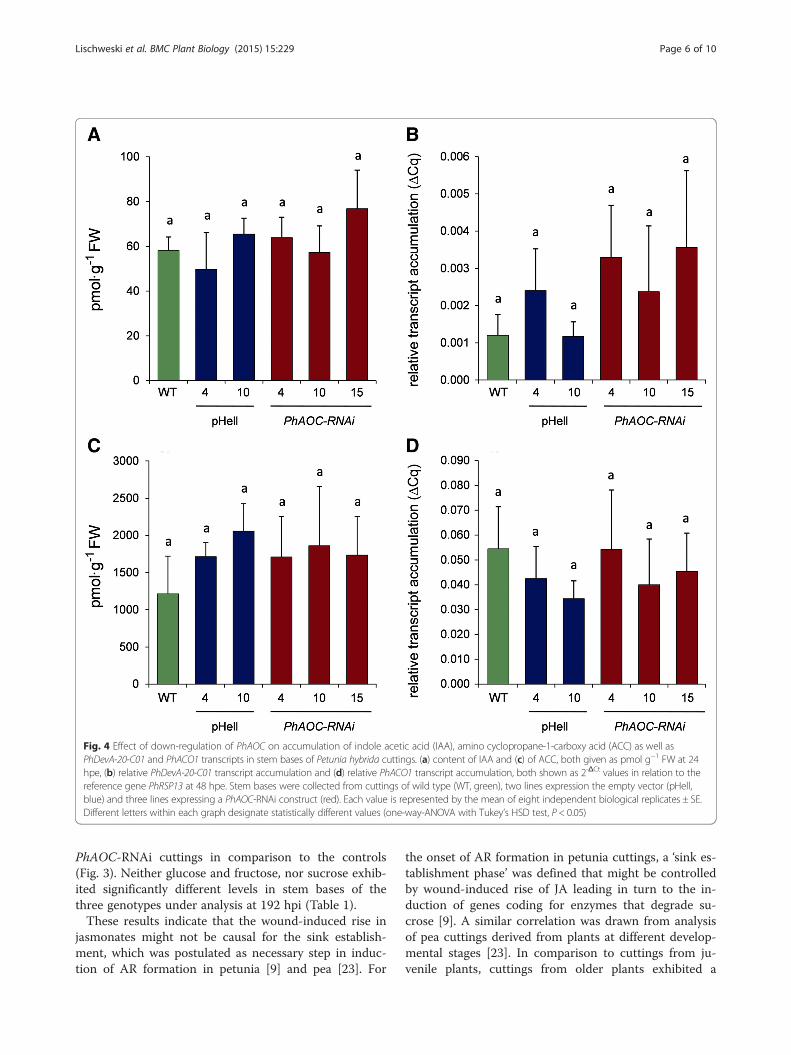
Fig. 4 Effect of down-regulation of PhAOC on accumulation of indole acetic acid (IAA), amino cyclopropane-1-carboxy acid (ACC) as well asPhDevA-20-C01 and PhACO1 transcripts in stem bases of Petunia hybrida cuttings. (a) content of IAA and (c) of ACC, both given as pmol g−1 FW at 24hpe, (b) relative PhDevA-20-C01 transcript accumulation and (d) relative PhACO1 transcript accumulation, both shown as 2-ΔCt values in relation to thereference gene PhRSP13 at 48 hpe. Stem bases were collected from cuttings of wild type (WT, green), two lines expression the empty vector (pHell,blue) and three lines expressing a PhAOC-RNAi construct (red). Each value is represented by the mean of eight independent biological replicates ± SE.Different letters within each graph designate statistically different values (one-way-ANOVA with Tukey’s HSD test, P < 0.05)
Lischweski et al. BMC Plant Biology (2015) 15:229 Page 6 of 10
PhAOC-RNAi cuttings in comparison to the controls(Fig. 3). Neither glucose and fructose, nor sucrose exhib-ited significantly different levels in stem bases of thethree genotypes under analysis at 192 hpi (Table 1).These results indicate that the wound-induced rise in
jasmonates might not be causal for the sink establish-ment, which was postulated as necessary step in induc-tion of AR formation in petunia [9] and pea [23]. For
the onset of AR formation in petunia cuttings, a ‘sink es-tablishment phase’ was defined that might be controlledby wound-induced rise of JA leading in turn to the in-duction of genes coding for enzymes that degrade su-crose [9]. A similar correlation was drawn from analysisof pea cuttings derived from plants at different develop-mental stages [23]. In comparison to cuttings from ju-venile plants, cuttings from older plants exhibited a
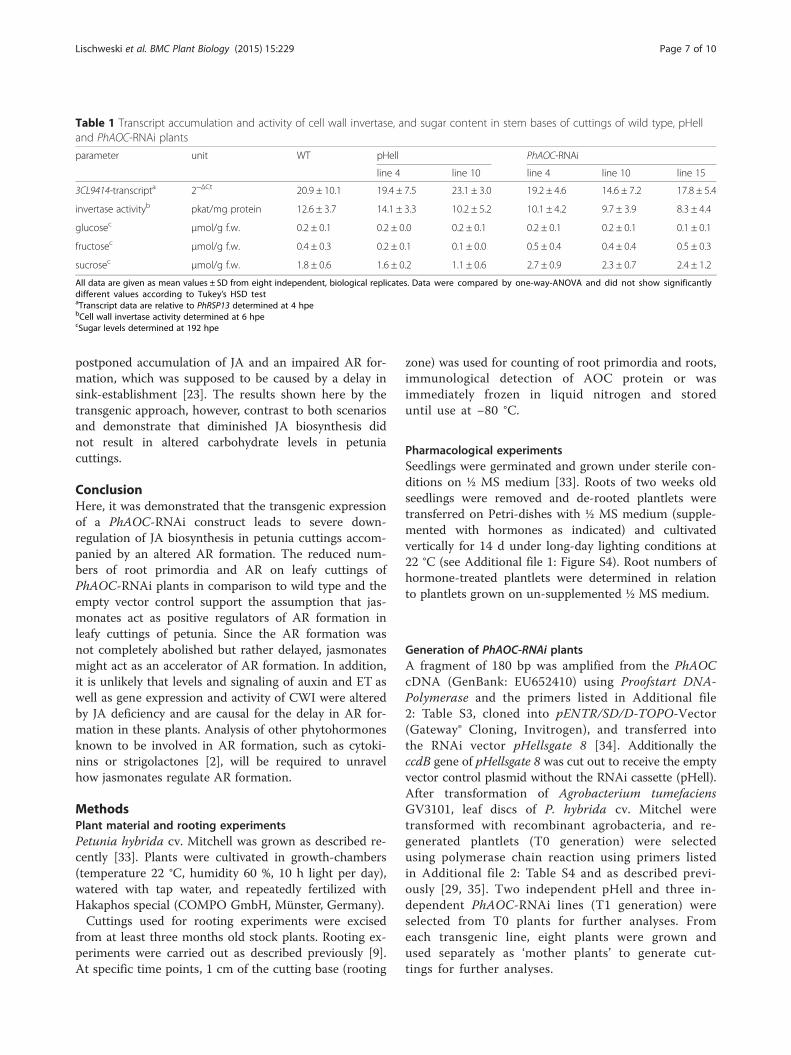
Table 1 Transcript accumulation and activity of cell wall invertase, and sugar content in stem bases of cuttings of wild type, pHelland PhAOC-RNAi plants
parameter unit WT pHell PhAOC-RNAi
line 4 line 10 line 4 line 10 line 15
3CL9414-transcripta 2−ΔCt 20.9 ± 10.1 19.4 ± 7.5 23.1 ± 3.0 19.2 ± 4.6 14.6 ± 7.2 17.8 ± 5.4
invertase activityb pkat/mg protein 12.6 ± 3.7 14.1 ± 3.3 10.2 ± 5.2 10.1 ± 4.2 9.7 ± 3.9 8.3 ± 4.4
glucosec μmol/g f.w. 0.2 ± 0.1 0.2 ± 0.0 0.2 ± 0.1 0.2 ± 0.1 0.2 ± 0.1 0.1 ± 0.1
fructosec μmol/g f.w. 0.4 ± 0.3 0.2 ± 0.1 0.1 ± 0.0 0.5 ± 0.4 0.4 ± 0.4 0.5 ± 0.3
sucrosec μmol/g f.w. 1.8 ± 0.6 1.6 ± 0.2 1.1 ± 0.6 2.7 ± 0.9 2.3 ± 0.7 2.4 ± 1.2
All data are given as mean values ± SD from eight independent, biological replicates. Data were compared by one-way-ANOVA and did not show significantlydifferent values according to Tukey’s HSD testaTranscript data are relative to PhRSP13 determined at 4 hpebCell wall invertase activity determined at 6 hpecSugar levels determined at 192 hpe
Lischweski et al. BMC Plant Biology (2015) 15:229 Page 7 of 10
postponed accumulation of JA and an impaired AR for-mation, which was supposed to be caused by a delay insink-establishment [23]. The results shown here by thetransgenic approach, however, contrast to both scenariosand demonstrate that diminished JA biosynthesis didnot result in altered carbohydrate levels in petuniacuttings.
ConclusionHere, it was demonstrated that the transgenic expressionof a PhAOC-RNAi construct leads to severe down-regulation of JA biosynthesis in petunia cuttings accom-panied by an altered AR formation. The reduced num-bers of root primordia and AR on leafy cuttings ofPhAOC-RNAi plants in comparison to wild type and theempty vector control support the assumption that jas-monates act as positive regulators of AR formation inleafy cuttings of petunia. Since the AR formation wasnot completely abolished but rather delayed, jasmonatesmight act as an accelerator of AR formation. In addition,it is unlikely that levels and signaling of auxin and ET aswell as gene expression and activity of CWI were alteredby JA deficiency and are causal for the delay in AR for-mation in these plants. Analysis of other phytohormonesknown to be involved in AR formation, such as cytoki-nins or strigolactones [2], will be required to unravelhow jasmonates regulate AR formation.
MethodsPlant material and rooting experimentsPetunia hybrida cv. Mitchell was grown as described re-cently [33]. Plants were cultivated in growth-chambers(temperature 22 °C, humidity 60 %, 10 h light per day),watered with tap water, and repeatedly fertilized withHakaphos special (COMPO GmbH, Münster, Germany).Cuttings used for rooting experiments were excised
from at least three months old stock plants. Rooting ex-periments were carried out as described previously [9].At specific time points, 1 cm of the cutting base (rooting
zone) was used for counting of root primordia and roots,immunological detection of AOC protein or wasimmediately frozen in liquid nitrogen and storeduntil use at −80 °C.
Pharmacological experimentsSeedlings were germinated and grown under sterile con-ditions on ½ MS medium [33]. Roots of two weeks oldseedlings were removed and de-rooted plantlets weretransferred on Petri-dishes with ½ MS medium (supple-mented with hormones as indicated) and cultivatedvertically for 14 d under long-day lighting conditions at22 °C (see Additional file 1: Figure S4). Root numbers ofhormone-treated plantlets were determined in relationto plantlets grown on un-supplemented ½ MS medium.
Generation of PhAOC-RNAi plantsA fragment of 180 bp was amplified from the PhAOCcDNA (GenBank: EU652410) using Proofstart DNA-Polymerase and the primers listed in Additional file2: Table S3, cloned into pENTR/SD/D-TOPO-Vector(Gateway® Cloning, Invitrogen), and transferred intothe RNAi vector pHellsgate 8 [34]. Additionally theccdB gene of pHellsgate 8 was cut out to receive the emptyvector control plasmid without the RNAi cassette (pHell).After transformation of Agrobacterium tumefaciensGV3101, leaf discs of P. hybrida cv. Mitchel weretransformed with recombinant agrobacteria, and re-generated plantlets (T0 generation) were selectedusing polymerase chain reaction using primers listedin Additional file 2: Table S4 and as described previ-ously [29, 35]. Two independent pHell and three in-dependent PhAOC-RNAi lines (T1 generation) wereselected from T0 plants for further analyses. Fromeach transgenic line, eight plants were grown andused separately as ‘mother plants’ to generate cut-tings for further analyses.

Lischweski et al. BMC Plant Biology (2015) 15:229 Page 8 of 10
Quantitative RT-PCR analysisRNA isolation and determination of transcript accumula-tions of PhAOC, PhACO1 (SGN-U207414) PhDevA-20-C01(SGN-U212243) as well as 3CL9414 (PhCWI) by qRT-PCR were done as described by [9] using Cytoplasmicribosomal protein S13 of P. hybrida (PhRSP13; SGN-U207968) as reference gene. This gene was selectedaccording to [36] and was tested in advance for eventranscript levels in stem bases of cuttings (Additionalfile 1: Figure S5). Each reaction mix contained a 15 ngRNA equivalent of cDNA and 1 pm gene-specificprimers. All assays were performed on at least eightbiological replicates in three technical replicates each.Relative gene expressions were calculated by the com-parative Cq method [37]. Real-time PCR primers weredesigned using Primer Express software (Applied Bio-systems, Warrington, UK). Primer sequences are givenin Additional file 2: Table S5.
Extraction of proteins, immunoblot analysis, and assay ofAOC activityProteins were extracted from 1 g of homogenizedplant material with 50 mM sodium phosphate buffer,pH 7, containing 2 % PVPP and 0,05 % Tween 20 asdescribed previously [38]. Resulting plants extractswere used for both, immunoblot analysis accordingto [38] as well as determination of AOC activity.The latter was performed according to [39] with thefollowing modifications. Protein extracts containing35 μg of total protein, recombinant HvAOS activity(4 nkat; [40], and sodium phosphate buffer (50 mM;pH 7.0) were combined in a final volume of 200 μl.The reaction was initiated by the addition of 40 nmol13(S)-HPOT. After incubation at 4 °C for 10 min,reaction was stopped by acidification. Me-OPDA wasadded as internal standard. Extraction with 2 ml ofdiethyl ether and evaporation of extract was performedfollowed by treatment with 0.2 M NaOH (in methanol) toactivate trans-isomerization of cis-(+)-OPDA. After incu-bation at 4 °C for 60 min, reaction was stopped byneutralization with 2 N HCl. The reaction mixtures wasextracted with 2 ml of diethyl ether, evaporated and sub-jected to HPLC using an Eurospher100 C18 4 mm column(Macherey-Nagel, Düren, Germany) and 75 % (v/v) solv-ent A (methanol) in solvent B (0.2 % acetic acid in H2O)at 1 ml/min. Fraction at Rt 12.0–13.4 min (OPDA) wassubsequently separated isocratically in cis- and trans-iso-mers by HPLC with an EC 200/4 Nucleotex beta-PMcolumn (Macherey-Nagel) and 65 % (v/v) solvent Ain solvent B at 1 ml/min. The absolute content ofOPDA was calculated using the internal standard.Percentage of enzymatic formed cis-OPDA was calcu-lated according to [41].
ImmunocytochemistryImmunocytochemical detection of AOC in stem baseswas performed as described [42]. Small pieces of stemswere fixed with 4 % (w/v) paraformaldehyde/0.1 % (v/v)Triton X-100 in phosphate buffered saline (135 mM,NaCl, 3 mM KCl, 1.5 mM KH2PO4, and 8 mMNa2HPO4) and embedded in polyethylene glycol 1500(Merck, Darmstadt, Germany). Cross-sections of 5 μmthickness were used for immunolabeling with therabbit polyclonal antibody raised against recombinantLeAOC [40] at a dilution of 1:1000. The use of pre-immune serum at the same dilutions served as a con-trol and revealed no signals. As secondary antibody,goat anti-rabbit IgG conjugated with AlexaFluor488(life technologies, Carlsbad, CA, USA) was used ac-cording to the manufacturer’s instructions. Sectionswere analyzed by confocal laser scanning microscopyusing a LSM510 META (Carl Zeiss GmbH, Jena,Germany).
Determination of primordia numberFor counting the primordia number, 0.5 cm of each cut-ting base was fixed in a solution of 2 % (v/v) formalde-hyde, 0,1 % (v/v) Triton X-100 in buffer (50 mMPIPES, 5 mM MgSO4, 5 mM EGTA, pH 6.9) at roomtemperature for two hours. Fixed stem segments werecut into 300 μm sections using a VT1000S microtome(Leica Instruments, Nussloch, Germany). Microscopicanalyses were performed using an Axioplan microscope(Carl Zeiss).
Determination of IAA, ACC, JA, JA-Ile and OPDAAbout 0.5 g FW of homogenised plant material (stembases of about 1 cm in length) pooled from at least threecuttings was extracted with 10 ml methanol. To quantifyIAA and ACC, [13C6]-IAA and [2H4]-ACC, respectively,were added as internal standards in appropriate amountsbefore extraction. The homogenate was filtered and wasplaced on a column filled with 3 ml DEAE-SephadexA25 (Amersham Pharmacia Biotech AB, Uppsala,Sweden). The column was washed with 3 ml metha-nol resulting after evaporation in extract E1. Subsequently,the column was washed with 3 ml of 0.1, 1.0, and 1.5 Macetic acid in methanol. The following elution with 4 ×3 ml of 3 M acetic acid in methanol resulted after evapor-ation in E2.To determine ACC content, E1 was processed as de-
scribed in [21]. Extract E2 was separated on preparativeHPLC using method 1 as described by [43]. Fraction atRt 10.5–11.9 min was evaporated, dissolved in 100 μlmethanol and methylated with 200 μl ethereal diazo-methane at 20 °C for 10 min. Afterwards the sample wasevaporated, dissolved in 70 μl CH3CN and IAA contentwas determined by GC-MS as described by [43].

Lischweski et al. BMC Plant Biology (2015) 15:229 Page 9 of 10
JA, JA-Ile and OPDA were determined as described by[43] using 1 g fresh weight of homogenized plant mater-ial per sample. For these determinations, leaves weremechanically wounded and harvested after 1 h.All determinations were done at least from eight inde-
pendent biological replicates, each represented by oneplant, from which cuttings were excised.
Determination of invertase activity and soluble sugarcontentsInvertase activity was measured as described previously[44]. Determination of soluble sugar contents was per-formed photometrically by a coupled enzymatic assay asdescribed by [44].
Statistical analysesStatistical analyses were performed using the softwareSPSS 17.0 (SPSS Inc., Chicago, IL, USA). All data setswere tested for normal distribution and homogeneity ofvariance using the Kolmogorov-Smirnov test and Levenetest, respectively. One-way-ANOVA was followed byTukey’s HSD test for significance.
Additional files
Additional file 1: Figure S1. Genome structure of PhAOC. Figure S2.Accumulation of AOC protein in stem bases of cuttings of Petuniahybrida expressing PhAOC-RNAi. Figure S3. Accumulation of indole aceticacid (IAA) and PhDevA-20-C01 transcripts in stem bases of Petunia hybridacuttings. Figure S4. AR formation in de-rooted seedlings of P. hybridatreated with different concentrations of jasmonic acid. Figure S5. qRT-PCRanalysis of putative reference genes used for transcript analyses in stembases of Petunia hybrida cuttings. (DOCX 1376 kb)
Additional file 2: Table S1. Number of ARs in de-rooted seedlings of P.hybrida wild-type treated with various concentrations of oxylipins, 2,4-D andACC. Table S2. Number of ARs in de-rooted seedlings of P. hybrida wild-typenon-treated or treated with jasmonic acid (JA) alone or in combination with2,4-D or ACC. Table S3. Primer sequences for cloning of 35S::PhAOC-RNAiinto pENTR. Table S4. Primer sequences for selection of transgenic plants.Table S5. Primer sequences of PhAOC, PhACO, Ph2-GH3, Ph3CL9414, PhRSP13used in quantitative real-time PCR. (DOCX 26 kb)
AbbreviationsACC: Amino cyclopropane-1-carboxy acid; AOC: Allene oxide cyclase;AR: Adventitious roots; CWI: Cell wall invertase; JA: Jasmonic acid;JA-Ile: Jasmonoyl isoleucine; OPDA: cis-12-oxo-phytodienoic acid; 2,4-D:2,4-dichlorophenoxy acetic acid.
Competing interestsThe authors declare that they have no competing interests.
Authors’ contributionsSL and BH designed the research. SL, DG and AM performed the researchand analyzed the data. BH wrote the manuscript with the assistance of SL.All authors read and approved the final manuscript.
Authors’ informationNot applicable.
Availabity of data and materialsNot applicable.
AcknowledgementsWe thank Birgit Ortel (IPB Halle) for help in IAA and ACC quantification. ClausWasternack and Alain Tissier (IPB Halle) are highly acknowledged for criticalreading of the manuscript.
FundingThis work was funded by the ‘Pakt für Forschung und Innovation’ of theLeibniz Association, Germany.
Author details1Department of Cell and Metabolic Biology, Leibniz Institute of PlantBiochemistry, Weinberg 3, D06120 Halle/Salle, Germany. 2Present address:Interdisziplinäres Stoffwechsel-Centrum, Charité, Augustenburger Platz 1,D13353 Berlin, Germany. 3Present address: IDT Biologika GmbH, AmPharmapark, D06861 Dessau-Roßlau, Germany. 4Present address: Institute ofPlant Biology, University of Zürich, Zollikerstrasse 107, CH-8008 Zürich,Switzerland.
Received: 4 May 2015 Accepted: 12 September 2015
References1. De Klerk G, Van der Krieken W, De Jong J. The formation of adventitious
roots: new concepts new possibilities. In Vitro Cell Dev Biol Plant.1999;35:189–99.
2. Verstraeten I, Schotte S, Geelen D. Hypocotyl adventitious rootorganogenesis differs from lateral root development. Front Plant Sci.2014;5:495.
3. Bellini C, Pacurar DI, Perrone I. Adventitious roots and lateral roots:Similarities and differences. Annu Rev Plant Biol. 2014;65(1):639–66.
4. Da Costa CT, De Almeida MR, Ruedell CM, Schwambach J, Maraschin FDS,Fett-Neto AG. When stress and development go hand in hand: Mainhormonal controls of adventitious rooting in cuttings. Front Plant Sci.2013;4:133.
5. Della Rovere F, Fattorini L, D’Angeli S, Veloccia A, Del Duca S, Cai G, et al.Arabidopsis SHR and SCR transcription factors and AUX1 auxin influx carriercontrol the switch between adventitious rooting and xylogenesis in plantaand in in vitro cultured thin cell layers. Ann Bot. 2015;115(4):617–28.
6. Della Rovere F, Fattorini L, D’Angeli S, Veloccia A, Falasca G, Altamura MM.Auxin and cytokinin control formation of the quiescent centre in theadventitious root apex of arabidopsis. Ann Bot. 2013;112(7):1395–407.
7. Takahashi F, Sato-Nara K, Kobayashi K, Suzuki M, Suzuki H. Sugar-inducedadventitious roots in Arabidopsis seedlings. J Plant Res. 2003;116(2):83–91.
8. Druege U. Involvement of carbohydrates in survival and adventitious rootformation of cuttings within the scope of global horticulture. In: Niemi K,Scagel C, editors. Adventitious root formation of forest trees andhorticultural plants – from genes to applications. Kerala: Research Signpost;2009. p. 187–208.
9. Ahkami AH, Lischewski S, Haensch K-T, Porfirova S, Hofmann J, RolletschekH, et al. Molecular physiology of adventitious root formation in Petuniahybrida cuttings: involvement of wound response and primary metabolism.New Phytol. 2009;181(3):613–25.
10. Gutierrez L, Mongelard G, Floková K, Păcurar DI, Novák O, Staswick P, et al.Auxin controls Arabidopsis adventitious root initiation by regulatingjasmonic acid homeostasis. Plant Cell. 2012;24(6):2515–27.
11. Clarke SM, Cristescu SM, Miersch O, Harren FJM, Wasternack C, Mur LAJ.Jasmonates act with salicylic acid to confer basal thermotolerance inArabidopsis thaliana. New Phytol. 2009;182(1):175–87.
12. Vick BA, Zimmerman DC. The biosynthesis of jasmonic acid: a physiologicalrole for plant lipoxygenase. Biochem Biophys Res Comm. 1983;111:470–7.
13. Feussner I, Wasternack C. The lipoxygenase pathway. Annu Rev Plant Biol.2002;53:275–97.
14. Chini A, Fonseca S, Fernandez G, Adie B, Chico JM, Lorenzo O, et al. The JAZfamily of repressors is the missing link in jasmonate signalling. Nature.2007;448(7154):666–71.
15. Thines B, Katsir L, Melotto M, Niu Y, Mandaokar A, Liu G, et al. JAZ repressorproteins are targets of the SCFCOI1 complex during jasmonate signalling.Nature. 2007;448(7154):661–5.
16. Fonseca S, Chini A, Hamberg M, Adie B, Porzel A, Kramell R, et al.(+)-7-iso-jasmonoyl-L-isoleucine is the endogenous bioactive jasmonate.Nat Chem Biol. 2009;5(5):344–50.

Lischweski et al. BMC Plant Biology (2015) 15:229 Page 10 of 10
17. Sheard LB, Tan X, Mao H, Withers J, Ben-Nissan G, Hinds TR, et al. Jasmonateperception by inositol-phosphate-potentiated COI1-JAZ co-receptor. Nature.2010;468(7322):400–5.
18. Laudert D, Schaller F, Weiler E. Transgenic Nicotiana tabacum andArabidopsis thaliana plants overexpressing allene oxide synthase. Planta.2000;211:163–5.
19. Hause B, Stenzel I, Miersch O, Maucher H, Kramell R, Ziegler J, et al.Tissue-specific oxylipin signature of tomato flowers - allene oxidecyclase is highly expressed in distinct flower organs and vascularbundles. Plant J. 2000;24:113–26.
20. Wasternack C, Hause B. Jasmonates: biosynthesis, perception, signaltransduction and action in plant stress response, growth anddevelopment. An update to the 2007 review in Annals of Botany. AnnBot. 2013;111:1021–58.
21. Druege U, Franken P, Lischewski S, Ahkami AH, Zerche S, Hause B, et al.Transcriptomic analysis reveals ethylene as stimulator and auxin asregulator of adventitious root formation in petunia cuttings. Front PlantSci. 2014;5:494.
22. Ravnikar M, Vilhar B, Gogala N. Stimulatory effects of jasmonic acid onpotato stem node and protoplast culture. J Plant Growth Regul.1992;11(1):29–33.
23. Rasmussen A, Hosseini SA, Hajirezaei M-R, Druege U, Geelen D. Adventitiousrooting declines with the vegetative to reproductive switch and involves achanged auxin homeostasis. J Exp Bot. 2015;66(5):1437–52.
24. Fattorini L, Falasca G, Kevers C, Mainero Rocca L, Zadra C, Altamura M.Adventitious rooting is enhanced by methyl jasmonate in tobacco thin celllayers. Planta. 2009;231(1):155–68.
25. da Rocha CL, Troleis J, Mastroberti AA, Mariath JEA, Fett-Neto AG.Distinct modes of adventitious rooting in Arabidopsis thaliana. Plant Biol.2012;14(1):100–9.
26. Garrido G, Ramón Guerrero J, Angel Cano E, Acosta M, Sánchez-Bravo J.Origin and basipetal transport of the IAA responsible for rooting ofcarnation cuttings. Physiol Plant. 2002;114(2):303–12.
27. Blakesley D, Davis TD. Auxin metabolism and adventitious root initiation. In:Haissig BE, editor. Biology of adventitious root formation. New York: PlenumPress; 1994. p. 143–53.
28. Sorin C, Bussell JD, Camus I, Ljung K, Kowalczyk M, Geiss G, et al. Auxin andlight control of adventitious rooting in Arabidopsis require ARGONAUTE1.Plant Cell. 2005;17(5):1343–59.
29. Clark DG, Gubrium EK, Barrett JE, Nell TA, Klee HJ. Root formation inethylene-insensitive plants. Plant Physiol. 1999;121(1):53–60.
30. Shibuya K, Barry KG, Ciardi JA, Loucas HM, Underwood BA, Nourizadeh S,et al. The central role of PhEIN2 in ethylene responses throughout plantdevelopment in Petunia. Plant Physiol. 2004;136(2):2900–12.
31. Roitsch T, González M-C. Function and regulation of plant invertases: sweetsensations. Trends Plant Sci. 2004;9(12):606–13.
32. Roitsch T, Balibrea M, Hofmann M, Proels R, Sinha A. Extracellular invertase:key metabolic enzyme and PR protein. J Exp Bot. 2003;54:513–24.
33. Klopotek Y, Haensch K-T, Hause B, Hajirezaei M-R, Druege U. Dark exposureof petunia cuttings strongly improves adventitious root formation andenhances carbohydrate availability during rooting in the light. J PlantPhysiol. 2010;167(7):547–54.
34. Wesley SV, Helliwell CA, Smith NA, Wang M, Rouse DT, Liu Q, et al.Construct design for efficient, effective and high-throughput gene silencingin plants. Plant J. 2001;27(6):581–90.
35. Meer IM. Agrobacterium-mediated transformation of Petunia leaf discs. In:Loyola-Vargas V, Vázquez-Flota F, editors. Plant Cell Culture Protocols, vol.318. New York: Humana Press; 2006. p. 265–72.
36. Mallona I, Lischewski S, Weiss J, Hause B, Egea-Cortines M. Validation ofreference genes for quantitative real-time PCR during leaf and flowerdevelopment in Petunia hybrida. BMC Plant Biol. 2010;10(1):4.
37. Schmittgen TD, Livak KJ. Analyzing real-time PCR data by the comparativeC(T) method. Nat Protoc. 2008;3:1101–8.
38. Stenzel I, Hause B, Maucher H, Pitzschke A, Miersch O, Ziegler J, et al.Allene oxide cyclase dependence of the wound response and vascularbundle-specific generation of jasmonates in tomato - amplification inwound signaling. Plant J. 2003;33:577–89.
39. Ziegler J, Hamberg M, Miersch O, Parthier B. Purification and characterization ofallene oxide cyclase from dry corn seeds. Plant Physiol. 1997;114(2):565–73.
40. Ziegler J, Stenzel I, Hause B, Maucher H, Hamberg M, Grimm R, et al.Molecular cloning of allene oxide cyclase: The enzyme establishing the
stereochemistry of octadecanoids and jasmonates. J Biol Chem.2000;275:19132–8.
41. Ziegler J, Wasternack C, Hamberg M. On the specificity of allene oxidecyclase. Lipids. 1999;34:1005–15.
42. Isayenkov S, Mrosk C, Stenzel I, Strack D, Hause B. Suppression of alleneoxide cyclase in hairy roots of Medicago truncatula reduces jasmonate levelsand the degree of mycorrhization with Glomus intraradices. Plant Physiol.2005;139(3):1401–10.
43. Miersch O, Neumerkel J, Dippe M, Stenzel I, Wasternack C. Hydroxylatedjasmonates are commonly occurring metabolites of jasmonic acid andcontribute to a partial switch-off in jasmonate signaling. New Phytol.2008;177(1):114–27.
44. Schaarschmidt S, Qu N, Strack D, Sonnewald U, Hause B. Local induction ofthe alc gene switch in transgenic tobacco plants by acetaldehyde. PlantCell Physiol. 2004;45(11):1566–77.
Submit your next manuscript to BioMed Centraland take full advantage of:
• Convenient online submission
• Thorough peer review
• No space constraints or color figure charges
• Immediate publication on acceptance
• Inclusion in PubMed, CAS, Scopus and Google Scholar
• Research which is freely available for redistribution
Submit your manuscript at www.biomedcentral.com/submit




















