
The impact of Felsenstein's "Phylogenies and the comparative method" on evolutionary biology
14
Jointly published by AkadØmiai Kiad, Budapest Scientometrics, and Springer, Dordrecht Vol. 62, No. 1 (2005) 5366 Received June 28, 2004 Address for correspondence: LUIS MAURICIO BINI Departamento de Biologia Geral, Instituto de CiŒncias Biolgicas Universidade Federal de GoiÆs, Goinia GO 74001-970, Brazil E-mail: [email protected] 01389130/2005/US $ 20.00 Copyright ' 2005 AkadØmiai Kiad, Budapest All rights reserved The impact of Felsensteins Phylogenies and the comparative method on evolutionary biology PRISCILLA CARVALHO, a,b,c JOS ALEXANDRE FELIZOLA DINIZ-FILHO, b,c LUIS MAURICIO BINI b a Graduate Program in Biology (Ecology), Instituto de CiŒncias Biolgicas, Universidade Federal de GoiÆs,Goinia (Brazil) b Departamento de Biologia Geral, Instituto de CiŒncias Biolgicas, Universidade Federal de GoiÆs, Goinia (Brazil) c Departamento de Biologia / MCAS-PROPE, Universidade Catlica de GoiÆs, Goinia, (Brazil) Felsensteins (1985) method of phylogenetic independent contrasts is probably the most commonly used technique in evolutionary biology to study adaptation of organisms to their environment, taking phylogeny into account. Here, we performed a scientometric evaluation of all 1462 articles that cited Felsenstein (1985) between 1985 and 2002, in order to analyze the impact of his comparative method on the evolutionary research program and what has been done since it. We found that Felsenteins (1985) article can be classified as a hot paper or a breakthrough contribution, since it was the most cited article from The American Naturalist published in 1985. Also, it can be considered as a citation classic, since it is the third more cited paper in The American Naturalist from 1945 to 2002. In general, papers that cited Felsenstein (1985) were published in high-impact journals, and most of them are theoretical articles indicating that biologists are aware of statistical and conceptual problems in dealing with comparative methods. Introduction The study of adaptation has been, since Darwins time, based on comparative method. Adaptation is a trait or organismal function that arises as a direct product of natural selection (inherited differential survival or reproductive ability), and it is usually measured by the correlation between traits or between traits and components of environmental variation across species. 1,2 For evolutionary biologists, thinking comparatively establishes the generality of evolutionary phenomena of adaptation, and it is fair to say that, even considering the existence of other evolutionary forces (genetic drift and mutation, acting at different time scales), adaptation and natural selection are usually considered the core of modern evolutionary synthesis. 3 Even today, most biologists are still influenced by the ultra-Darwinian view of evolution that dominated evolutionary thought during 1960s, the so-called panglossian paradigm. 4
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of The impact of Felsenstein's "Phylogenies and the comparative method" on evolutionary biology

Jointly published by Akadémiai Kiadó, Budapest Scientometrics,and Springer, Dordrecht Vol. 62, No. 1 (2005) 53–66
Received June 28, 2004Address for correspondence:LUIS MAURICIO BINIDepartamento de Biologia Geral, Instituto de Ciências BiológicasUniversidade Federal de Goiás, Goiânia GO 74001-970, BrazilE-mail: [email protected]
0138–9130/2005/US $ 20.00Copyright © 2005 Akadémiai Kiadó, BudapestAll rights reserved
The impact of Felsenstein’s “Phylogenies andthe comparative method” on evolutionary biologyPRISCILLA CARVALHO,a,b,c JOSÉ ALEXANDRE FELIZOLA DINIZ-FILHO,b,c
LUIS MAURICIO BINIb
a Graduate Program in Biology (Ecology), Instituto de Ciências Biológicas,Universidade Federal de Goiás,Goiânia (Brazil)
b Departamento de Biologia Geral, Instituto de Ciências Biológicas, Universidade Federal de Goiás,Goiânia (Brazil)
c Departamento de Biologia / MCAS-PROPE, Universidade Católica de Goiás, Goiânia, (Brazil)
Felsenstein’s (1985) method of phylogenetic independent contrasts is probably the mostcommonly used technique in evolutionary biology to study adaptation of organisms to theirenvironment, taking phylogeny into account. Here, we performed a scientometric evaluation of all1462 articles that cited Felsenstein (1985) between 1985 and 2002, in order to analyze the impactof his comparative method on the evolutionary research program and what has been done since it.We found that Felsentein’s (1985) article can be classified as a “hot paper” or a breakthroughcontribution, since it was the most cited article from The American Naturalist published in 1985.Also, it can be considered as a “citation classic”, since it is the third more cited paper in TheAmerican Naturalist from 1945 to 2002. In general, papers that cited Felsenstein (1985) werepublished in high-impact journals, and most of them are theoretical articles indicating thatbiologists are aware of statistical and conceptual problems in dealing with comparative methods.
Introduction
The study of adaptation has been, since Darwin’s time, based on comparativemethod. Adaptation is a trait or organismal function that arises as a direct product ofnatural selection (inherited differential survival or reproductive ability), and it is usuallymeasured by the correlation between traits or between traits and components ofenvironmental variation across species.1,2 For evolutionary biologists, thinkingcomparatively establishes the generality of evolutionary phenomena of adaptation, andit is fair to say that, even considering the existence of other evolutionary forces (geneticdrift and mutation, acting at different time scales), adaptation and natural selection areusually considered the core of modern evolutionary synthesis.3 Even today, mostbiologists are still influenced by the ultra-Darwinian view of evolution that dominatedevolutionary thought during 1960’s, the so-called “panglossian paradigm”.4

P. CARVALHO et al.: The impact of Felsenstein’s article
54 Scientometrics 62 (2005)
However, a critical problem occurs with this approach. It arises from the fact thatspecies are part of a hierarchically structured sequence of ancestrals and descendents(the phylogeny), and thus cannot be treated as independent data points for standardstatistical purposes. In other words, closely related species are not solely a product oftheir current environment, but have also inherited a certain proportion of theirphenotypes from a common ancestor.1,5,6 Thus, the relationship between two traitsmeasured in different species can arise because the species have common ancestors.Indeed, recent advances in systematic and evolutionary biology have shown theimportance of incorporating phylogenies into comparative studies in all areas ofbiology.7 Despite a few criticisms and limitations,8–10 the use of phylogeneticcomparative methods in cross-species studies has achieved a paradigmatic status.
To deal with non-independence of species due to phylogeny, two problems had to besolved first: (i) it was necessary to explicit know the historical relationships among taxa(phylogeny) and (ii) to develop methods that are able to incorporate these relationshipsin statistical models. The solution to these problems appeared during the 1970’s and1980’s. The methods necessary to reconstruct the phylogenetic relationships betweenorganisms started to be available especially after the pioneering work by W. Hennig inthe 1950’s and by the “operational approaches” to systematic developed in the 1960’sand 1970’s.11 It is now clear that the exponential growth in the phylogeneticreconstructions of different groups of organisms were due to molecular data that startedto be available mainly after the 1980’s 5 and, so, allowed for a wide use of comparativemethods. At the same time, in the beginning of 1980’s, evolutionary biologists startedto recognize the problems related to phylogenetic non-independence and to developdifferent analytical approaches to this problem.1,12,13
Felsenstein12 was the first to develop an explicit method for testing comparativerelationships based on the logic of comparing pairs of species that share a commonancestor (see Harvey and Pagel1 for discussion of previous attempts to overcome thisproblem). His phylogenetic independent contrasts (PIC) method was originally devisedas a way to address the problem of statistical dependence of comparative data. Underhis model, although the species data are not independent, the “contrasts”, or thedifference between pairs of species in the phylogeny, are independent of eachother,1,7,12,14 and can be used in standard statistical analysis. The method assumes aBrownian motion model of evolutionary change and requires that the true branchingphylogeny is known, including the branch lengths of the tree in units of expectedvariance of evolutionary change. This information allows the calculation of a set ofcomparisons among pairs of data points, each of which has the same expected mean andvariance under the null hypothesis. This method is frequently used by evolutionarybiologists, and other workers, following Felsenstein’s idea,12 have developed methodsthat can be applied to imperfectly resolved phylogenies.15,16

P. CARVALHO et al.: The impact of Felsenstein’s article
Scientometrics 62 (2005) 55
The large number of citations of Felsenstein’s paper since its publication in 1985and the many papers using his PIC method (see below) are initial indications of itsimportance in the field of evolutionary biology. However, there are now quantitativeindices to evaluate scientific activity and new methods and tools for assessing researchproductivity and trends, which allow us to evaluate objectively their importance.17–19
The citation and bibliographic data can be used to identify emerging specialties, newtechnologies and even the structure of research disciplines, and of science as a whole.These scientometric studies allow us to observe the progress of science and technologythrough the scientific literature and have been widely used due to the need of a globalview of research activities.17,20–23
One measurable output of research is a peer-reviewed article. In addition, one canmeasure the impact of these articles by determining how often they are cited. Thus, inthis paper we used a variety of datasets to analyze the impact of Felsenstein’scomparative method12 on evolutionary research and what has been done since itsoriginal publication. The analysis was based on all the papers that cited Felsenstein’sarticle12 between 1985 and 2002, and the abstracts of each paper were obtained usingthe database from the Institute for Scientific Information (ISI) (www.isiknowledge.com).For each paper we identified all authors, the country of the first author, the journal, theyear of publication, the taxonomic group (e.g. birds, mammals, reptiles, and others – seeFigure 3a) that were studied and the specific research field (classified into the followingcategories: physiology, behavior, co-evolution, life history and sexual selection). Forthe analysis of number of citations, the data were colleted in April, 2003.
Felsenstein (1985) as a “hot paper”
A total of 1462 articles cited Felsenstein12 between 1985 and 2002, and the numberof citations continuously increased from 1985 to 2002 (Figure 1a) (R2 = 0.96;F = 376.37; P < 0.001). Thus, there has been no stabilization of number of citations thatcould be interpreted as a “half-life” of the article;24,25 the influence of Felsenstein’spaper is still growing.
According to Garfield,17 a “hot paper” includes papers that have attracted more“attention” than other papers in the same journal published at the same time (i.e. aground-breaking work), allowing to monitor current breakthroughs at the forefront ofresearch in various specialties.26 To determine if Felsenstein12 can be considered a “hotpaper”, we analyzed how frequently all others articles published in 1985 in TheAmerican Naturalist were cited. The frequency of citation of these articles was recordedup to April, 2003 and ranged from 0 to 1522, with a median of 32 citations. Thedistribution of citation frequency was strongly right-skewed, and no other articles havebeen cited as much as Felsenstein12 (Figure 1b).

P. CARVALHO et al.: The impact of Felsenstein’s article
56 Scientometrics 62 (2005)
Figure 1. a) Citation counts of Felsenstein’s (1985) article from 1985 to 2002; b) histogram of the number ofcitations of the articles published in 1985 in The American Naturalist journal (the same year and the samejournal of Felsenstein’s article); c) histogram of the number of citations of the articles published between
1945 and 2002 in The American Naturalist journalData used in b) and c), were obtained in April 2003

P. CARVALHO et al.: The impact of Felsenstein’s article
Scientometrics 62 (2005) 57
Figure 1. d) mean impact factor of the journals that cited Felsenstein (1985) across the years; e) minimum andmaximum impact factor of the journals across the years; f) mean impact factor of the journals that cited
Felsentein (1985) by the first time

P. CARVALHO et al.: The impact of Felsenstein’s article
58 Scientometrics 62 (2005)
Kuhn27 stated that any branch of science cycles between long periods of “normalscience” when scientists follow a dominant tradition or “paradigm”, and short periodsof crisis and revolutionary change when the paradigm is shaken and ultimatelyreplaced.27,28 The papers that cited Felsenstein12 and that, in turn, were more cited, weretheoretical expansions and computational applications of the method, starting in 1992(see below). In this way, it is fair to say that Felsenstein’s article12 revolutionized thefield of comparative biology by developing a method to explicitly take into accountphylogenetic effects. Nevertheless, as more scientists become interested in a researchprogram, a new paradigm takes place, and a new generation of normal scientists beginsthe cycle again,28 and in this context new methods and corrections can appear.29–31
It is also worthwhile to compare the frequency of citation of Felsenstein’s paper12
with that one of all other 5226 articles published in The American Naturalist from 1945up to 2002. The results indicate that only two papers were more cited than Felsenstein’spaper12 (Figure 1c). Thus, if a large period of the history of the journal is considered wecan still demonstrate the impact of Felsenstein’s method12 in the general field ofevolutionary biology, and his paper can definitively designated as a “citation classic”. Inaddition, this result corroborate a general bibliometric pattern, as stated by Verbeek etal.18:...“most publications only contribute to addition of knowledge in a rather modestway, while only very few make a major contribution”.
Temporal dynamics of impact factorsThe journal’s impact factor (IF) has traditionally been used for ranking and
evaluating journals and as a surrogate for judging the quality and importance of anindividual researcher’s publications. The impact factor (IF) of journal X for a particularyear T is defined as the number of citations received in year T by all documentspublished in X in the years T-1 and T-2, divided by the number of documents publishedin X in the years T-1 and T-2.24,25,32 Thus, it is a ratio between citations and citableitems published. Despite some well deserved criticisms,25,26,33–35 we consider this indexas an estimator of a journal’s importance in the field of evolutionary biology. So, werecorded the impact factor of each journal (using the values of the year 2000 for ageneral reference), which published papers citing Felsenstein12 (225 journals).
In general, papers that cited Felsenstein12 were published in high-impact journals,and the average IF tends to decrease from 1985 to 2002 (R2 = 0.37; F = 9.5; P = 0.007;IF = 38.7 – 0.0189×year) (Figure 1d), although maximum values remained high andapproximately constant over the years (R2 = 0.14; F = 2.69; P = 0.12; Figure 1e). Thesame trend was observed when we record the IF of the journals that cited Felsenstein12
by the first time (R2 = 0.52; F = 17.33; P = 0.0007; IF= 89.9 – 0.0448×years)(Figure 1f). Journals with higher impact factor were the first to cite the article, and thiscan be interpreted as the beginning of the recognition of the importance of the method

P. CARVALHO et al.: The impact of Felsenstein’s article
Scientometrics 62 (2005) 59
for dealing statistically with the non-independence of closely related organisms inanalyses of causal relationships among morphological, ecological, physiological andbehavioral traits.
Paradigm expansion
We also analyzed the citation frequency and IF of all papers citing Felsenstein12 inorder to evaluate the expansion and improving of the PIC method in the evolutionarybiology field. Indeed, the original proposition has been discussed extensively andfurther developed by subsequent authors concerned with the difficulties of obtainingaccurate and complete information about both the phylogenetic relationships (the treestructure) and the process of phenotypic evolution (branch lengths in units of expectedcharacter change) of the traits under analysis.15,29,36,37
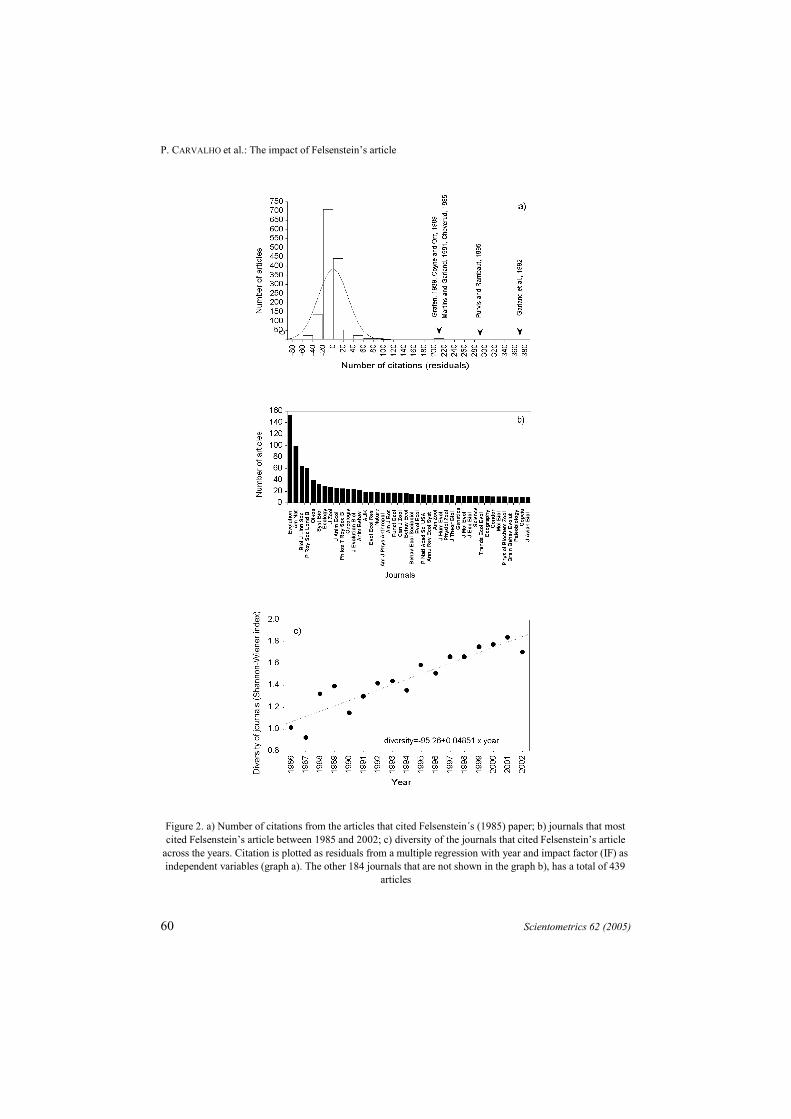
Since the number of citations was strongly dependent on the year and on the IF ofthe journals where the paper was published, it is necessary to take into account for theseeffects, avoiding highlighting studies that would be considered important developmentsof the original method only because they are older or were published, by unrelatedreasons, in high impact journals. On the other hand, this permits, for example, tounderline a paper published in a journal highly focused on evolutionary biology, whichcould have a low IF. We then regressed the number of citation (NC) of each article thatcited Felsentein12 against these two predictors (R2 = 0.26; NC = 7455.2 – 3.73×year +9.05×log IF). The residuals of this multiple regression allowed us to find out whichpaper has the largest repercussion, independently of the time when it was published andin which journal it was published (Figure 2a).
The articles that only applied PIC or other comparative methods to test specificadaptive hypotheses in any group of organisms are expected to be less cited, notbecause they are not “worth science”,38,39 but because their contribution is important to amore specific research area. On the other hand, theoretical-methodological articles areexpected to be widely cited, due to their wide application in different fields or becausethey improve on an established method. Indeed, we can highlight five highly citedarticles that improved and discussed the application of PIC as a comparative method(Figure 2a).
All these heavily cited articles are theoretical-methodological papers applying avariation of Felsenstein’s method,15 suggesting statistical improvements,29 proposingcomputer packages for analysis,40 or are about the comparison among distinctcomparative methods.36 The paper by Cheverud et al.13 also called attention to theproblem of species non-independence, but proposed a much more complex statisticalprocedure to take phylogenetic effects into account.
P. CARVALHO et al.: The impact of Felsenstein’s article
60 Scientometrics 62 (2005)
Figure 2. a) Number of citations from the articles that cited Felsenstein´s (1985) paper; b) journals that mostcited Felsenstein’s article between 1985 and 2002; c) diversity of the journals that cited Felsenstein’s article
across the years. Citation is plotted as residuals from a multiple regression with year and impact factor (IF) asindependent variables (graph a). The other 184 journals that are not shown in the graph b), has a total of 439
articles

P. CARVALHO et al.: The impact of Felsenstein’s article
Scientometrics 62 (2005) 61
The exception to these theoretical-methodological highly cited articles is the article byCoyne and Orr,41 in which Felsenstein’s method was applied to investigate the timecourse of speciation among pairs of closely related Drosophila species, a very importantissue for evolutionary theory per se at that time (which explains the high number ofcitations).
This confirms that Felsenstein’s original article12 can be classified as a hot paper anda citation classic (see above) and, consequently, other papers that proposed variationsand expansions of the original method were also widely cited.
Applications and distribution of studies: mapping comparative biology studies
The aim of mapping science is to display the foci of interest and attention thatprevail in a particular scientific area over a certain period of time. Based on the analysisof information from the scientific literature it is concerned with understanding both thestructure and the evolution of scientific (sub)fields.18
Although the papers that cited Felsenstein12 had contributors from 41 countries, halfof the scientific production originated from institutions in the USA, followed byEngland (13% of authors) and Canada (5.3%). The authors from 38 countriescontributed 31.7% of the papers published between 1985 and 2002.
During the surveyed period, Felsenstein12 was cited in papers published in 225different journals, but with a high concentration in a few ones. The dominant journalwas Evolution (10.5%), followed by The American Naturalist (7%) and BiologicalJournal of Linnean Society (4.3%) (Figure 2b). Indeed, these journals are usuallyconsidered as “top journals” in evolutionary biology.
We also measure the “diversity” of journals that published papers citingFelsenstein’s article12 in each year (1985 to 2002) using the Shannon-Wiener’s diversityindex (H’), widely used in ecological research.42 Gotelli and Graves43 defined diversityas a measure of both the number of species in the community and their relativeabundances. Some authors have also used ecological indexes in scientometric studies.44
A linear increase of H’ was detected (Figure 2c). Thus, PIC becomes more popularacross years. One important aspect contributing to this increase of popularity may berelated with the increase of phylogenies availability.
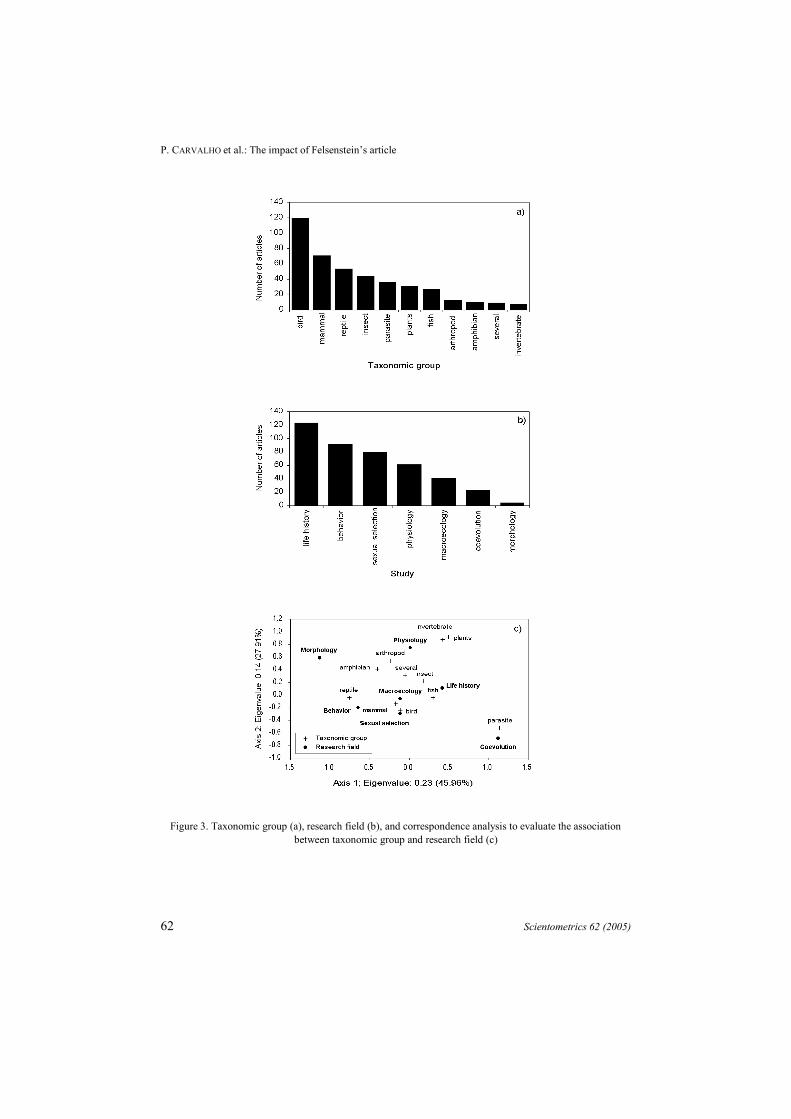
Out of the 1462 articles citing Felsenstein12, we estimate that 17% discuss themethod and its progress and do not deal with applications to study adaptive evolution inspecific group of organisms. Considering just the articles that really used thecomparative method (423 papers) and for which data is available, most applications ofPIC were in comparative studies of birds (28%), followed by mammals (17%)(Figure 3a). There is a clear bias toward studies in animals (92.5%) against plants(7.5%). This is expected, considering that phylogenies, as required for PIC and all othercomparative methods, are usually more frequently available for mammals and birds.
P. CARVALHO et al.: The impact of Felsenstein’s article
62 Scientometrics 62 (2005)
Figure 3. Taxonomic group (a), research field (b), and correspondence analysis to evaluate the associationbetween taxonomic group and research field (c)

P. CARVALHO et al.: The impact of Felsenstein’s article
Scientometrics 62 (2005) 63
We can also observe that evolutionary biologists were mainly interested in lifehistory (such as population structure, growth rate, longevity, fecundity) (29%), behavior(locomotor performance, prey escape strategies, migration) (21.5%) and sexualselection (sexual behavior, sexual dimorphism and sexual competition) (19%)(Figure 3b).
A correspondence analysis45 was performed to analyze graphically the relationshipsbetween the taxonomic group studied and the main research fields, based on thefrequency of studies in these categories (Table 1; Figure 3c). There is a significantassociation between taxonomic group studied and research field (X2 = 210.13; p<0.001).For example, studies on macroecology are more frequently focused in mammals andbirds, probably reflecting the paucity of macroecological data for other groups oforganisms.
Other interesting associations were also detected. Recent models of host-parasite co-evolution assume that parasites exert frequency-dependent selection on their hosts byfavoring rare alleles that may confer resistance against infection.46 Indeed, there is anincreasing interest about the immunological response of an organism (e.g. fishes andbirds) when it is affected by a parasite and the influence of this on the parasite richness.Thus, we were expecting the observed association between the studies about co-evolution and parasites. Also, the studies about behavior were concentrated on reptiles,mainly lizards (78%) with the great contributions of a few authors, such as Irschick(4.3%), Martins (3.5%) and Losos (2.6%). Physiological studies were mainly associatedwith plants.
Table 1. Number of studies that cited Felsenstein (1985) classified in research field and taxonomic groupResearch
fieldTaxonomicgroup
Morphology Co-evolution
Behavior Physiology Life history Macro-ecology
Sexualselection
Total
Reptile 2 0 29 7 6 4 6 54Bird 0 6 27 10 24 18 34 119Insect 0 2 4 10 14 5 10 45Mammal 0 2 21 7 19 5 17 71Parasite 0 11 0 0 19 1 5 36Fish 0 2 5 2 13 2 3 27Plants 0 1 0 13 15 2 0 31Amphibian 1 0 3 2 3 0 1 10Arthropod 1 0 2 4 3 0 3 13Several 0 0 1 2 2 3 1 9Invertebrate 0 0 0 3 4 1 0 8Total 4 24 92 60 122 41 80 423

P. CARVALHO et al.: The impact of Felsenstein’s article
64 Scientometrics 62 (2005)
Most of the research fields (such as sexual selection, macroecology and life history)are mainly concentrated on birds and mammals. Phylogenies are fundamental tocomparative biology. However, despite recent efforts, they are still unavailable formany groups of organisms. Although the number of articles reporting phylogeniesbased on gene-sequence information has been increasing exponentially since 1981, withmore than half appearing since 1996,5 our results indicate that the paucity of studies inother taxonomic groups can still be attributable to the lack of resolved phylogenies.
In short, as identified by Hoekstra et al.47 and Budilova et al.20 regarding ecologicalstudies, evolutionary biology theories and concepts are associated with their ownspecific types of organisms. The generality of these concepts and theories may be,therefore, questionable.
Concluding remarks
Citation data have been widely used to measure research quality and activity.26
Although publication and citation measures have been termed “lagging indicators”because they are an evaluation of work already completed, they also should be viewedas the scientific community’s opinion on the utility of earlier research. As such, thesedata tell us as much about current science and what is important in the eyes of today’sinvestigators.17 In this context, the main goal of the present study was to determine theinterest by the scientific community in Felsenstein’s article12 and the evolution of thecomparative methods since the original publication of PIC method.
Felsenstein’s paper12 achieves extraordinary citation frequencies in ecology andevolutionary biology, and it was the base for the development of several comparativemethods that are widely used today. Indeed, all our analyses indicate that Felsenstein’spaper12 can be surely classified as a “citation classic” or breakthrough contribution, withan enormous influence in the field. Ironically, Dr. Joseph Felsenstein has always beenmore concerned about phylogenetic reconstruction and not directly on the comparativeanalysis of adaptive process. Thus, the origin of the PIC method is still more interestingbecause it happened as an insight from someone looking comparative methods from aslight distinct perspective.
*
The authors are grateful to Dr. Bradford A. Hawkins (University of California) who critically revised themanuscript. J.A.F. Diniz-Filho and L.M. Bini are supported by the Conselho Nacional de DesenvolvimentoCientífico e Tecnológico (CNPq), the Coordenação para Aperfeiçoamento do Ensino Superior (CAPES) andby FUNAPE of the Universidade Federal de Goiás. P. Carvalho received a scholarship from CAPES.

P. CARVALHO et al.: The impact of Felsenstein’s article
Scientometrics 62 (2005) 65
Reference
1. HARVEY, P. H., PAGEL, M. D., The Comparative Method in Evolutionary Biology, Oxford UniversityPress, Oxford, 1991.
2. MARTINS, E. P., Adaptation and the comparative method, Trends in Ecology and Evolution, 15 (7)(2000) 296–299.
3. GOULD, S. J., The Structure of Evolutionary Theory, Harvard, 2002.4. GOULD, S. J., LEWONTIN, R. C., The spandrels of San Marco and the Panglossian paradigm: a critique of
the adaptationist program. Proceedings of the Royal Society of London Series B-Biological Sciences, 205(1979) 581–598.
5. PAGEL, M., Inferring the historical patterns of biological evolution, Nature, 401 (1999) 877–884.6. GARLAND, T. JR., IVES A. R., Using the past to predict the present: confidence intervals for regression
equations in phylogenetic comparative methods, The American Naturalist, 155 (3) (2000) 346–364.7. MARTINS, E. P., HANSEN, T. F., The statistical analysis of interspecific data: a review and evaluation of
phylogenetic comparative methods, In: MARTINS, E. P. (Ed.), Phylogenies and the Comparative Methodin Animal Behavior, Oxford University Press, Oxford, 1996, pp. 22–75.
8. RICKLEFS, R. E., STARCK, J. M., Applications of phylogenetically independent contrasts: a mixedprogress report, Oikos, 77 (1996) 167–172.
9. BJÖRKLUND, M., Are ‘comparative methods’ always necessary? Oikos, 80 (1997) 607–612.10. MAZER, S. J., Alternative approaches to the analysis of comparative data: compare and contrast,
American Journal of Botany, 85 (8) (1998) 1194–1199.11. FELSENSTEIN, J., Inferring Phylogenies, Sinauer Associates, Inc., Publishers, Sunderland, 2003.12. FELSENSTEIN, J., Phylogenies and the comparative method, The American Naturalist, 125 (1985) 1–15.13. CHEVERUD, J. M., DOW, M. M., LEUTENEGGER, W., The quantitative assessment of phylogenetic
constraints in comparative analyses: sexual dimorphism in body weight among primates, Evolution, 39(6) (1985) 1335–1351.
14. GITTLEMAN, J. L., LUH, H-K., On comparing comparative methods, Annual Review of Ecology andSystematics, 23 (1992) 383–404.
15. GRAFEN, A., The phylogenetic regression, Philosophical Transactions of the Royal Society of LondonSeries B-Biological Sciences, 326 (1989) 119–156.
16. PAGEL, M. D., HARVEY, P. H., Comparative methods for examining adaptation depend on evolutionarymodels, Folia Primatologica, 53 (1989) 203–220.
17. GARFIELD, E., Contract research services at ISI – citation analysis for governmental, industrial, andacademic clients, Essays of an Information Scientist, Vol. 15, 1992, pp. 75–83.
18. VERBEEK, A., DEBACKERE, K., LUWEL, M., ZIMMERMANN, E., Measuring progress and evolution inscience and technology. I: the multiple uses of bibliometric indicators, International Journal ofManagement Reviews, 4 (2) (2002) 179–211.
19. RESH, V. H., KOBZINA, N. G., A perspective on the key citations in freshwater benthic science, and thestudies that influenced them, Journal of the North American Benthological Society, 22 (3) (2003)341–351.
20. BUDILOVA, E. V., DROGALINA, J. A., TERIOKHIN, A. T., Principal trends in modern ecology and itsmathematical tools: an analysis of publications, Scientometrics, 39 (2) (1997) 147–157.
21. SUTTER, M., KOCHER, M. G., Power laws of research output: evidence for journals of economics,Scientometrics, 51 (2) (2001) 405–414.
22. DALPÉ, R., Bibliometric analysis of biotechnology, Scientometrics, 55 (2) (2002) 189–213.23. LETA, J., LEWISON, G., The contribution of women in Brazilian science: a case study in astronomy,
immunology and oceanography, Scientometrics, 57 (3) (2003) 339–353.24. MOED, H. F., VAN LEEUWEN, TH. N., REEDUK, J., A new classification system to describe the ageing of
scientific journals and their impact factors, Journal of Documentation, 54 (1998) 387–419.25. GLÄNZEL, W., MOED, H. F., Journal impact measures in bibliometric research, Scientometrics, 53 (2)
(2002) 171–193.

P. CARVALHO et al.: The impact of Felsenstein’s article
66 Scientometrics 62 (2005)
26. GARFIELD, E., WELLJAMS-DOROF, A., Citation data: their use as quantitative indicators for science andtechnology evaluation and policy-making, Science and Public Policy, 19 (5) (1992) 321–327.
27. KUHN, T., The Structure of Scientific Revolutions, University of Chicago Press, Chicago, 1962.28. RIGLER, F. H., PETERS, R. H., Excellence in Ecology: Science and Limnology, Ecology Institute,
Germany, 1995.29. GARLAND, T. JR., HARVEY, P. H., IVES, A. R., Procedures for the analysis of comparative data using
phylogenetically independent contrasts, Systematic Biology, 41 (1992) 18–32.30. DÍAZ-URIARTE, R., GARLAND, T. JR., Effects of branch length errors on the performance of
phylogenetically independent contrasts, Systematic Biology, 47 (1998) 654–672.31. GARLAND, T. JR., DÍAZ-URIARTE, R., Polytomies and phylogenetically independent contrasts:
examination of the bounded degrees of freedom approach, Systematic Biology, 48 (1999) 547–558.32. GARFIELD, E., Citation analysis as a tool in journal evaluation, Science, 178 (1972) 471–479.33. JONES, A. W., Impact factors of forensic science and toxicology journals: what do the numbers really
mean? Forensic Science International, 133 (2003) 1–8.34. COLQUHOUN, D., Challenging the tyranny of impact factors, Nature, 423 (2003) 479.35. LAWRENCE, P. A., The politics of publication, Nature, 422 (2003) 259–261.36. MARTINS, E. P., GARLAND, T. JR., Phylogenetic analyses of the correlated evolution of continuous
characters: a simulation study, Evolution, 45 (3) (1991) 534–557.37. MCKITRICK, M. C., Phylogenetic constraint in evolutionary theory: has it any explanatory power?
Annual Review of Ecology and Systematics, 24 (1993) 307–330.38. NUHFER, E. B., Science, citation, and funding, Science, 251 (1991) 1409.39. TAINER, J. A., Science, citation, and funding, Science, 251 (1991) 1408.40. PURVIS, A., RAMBAUT, A., Comparative-analysis by independent contrasts (CAIC) – an apple-macintosh
application for analyzing comparative data, Computer applications in the bioscience, 11 (1995) 247–251.41. COYNE, J. A., ORR, H. A., Patterns of speciation in Drosophila, Evolution, 43 (2) (1989) 362–381.42. MAGURRAN, A. E., Ecological Diversity and Its Measurement, Croom Helm, London, 1988.43. GOTELLI, N. J., GRAVES, G. R., Null Models in Ecology, Smithsonian Institution Press, Washington,
1996.44. ROUSSEAU, R., Evenness as a descriptive parameter for department or faculty evaluation studies, In:
SMET, E. (Ed.), Informatiewetenschap, Belgium, 1998, pp. 135–145.45. LEGENDRE, P., LEGENDRE, L., Numerical Ecology: Developments in Environmental Modeling, 20,
Elsevier Science, Amsterdam, 1998.46. POULIN, R., MARSHALL, L. J., SPENCER, H. G., Metazoan parasite species richness and genetic variation
among freshwater fish species: cause or consequence? International Journal for Parasitology, 30 (2000)697–703.
47. HOEKSTRA, T. W., ALLEN, T. F. H., FLATHER, C. H., Implicit scaling in ecological research, Bioscience,41 (1991) 148–154.




















