
Biotin deficiency and biotin excess: Effects on the female reproductive system
7
Steroids 74 (2009) 863–869 Contents lists available at ScienceDirect Steroids journal homepage: www.elsevier.com/locate/steroids Biotin deficiency and biotin excess: Effects on the female reproductive system Armida Báez-Salda ˜ na a , Ignacio Camacho-Arroyo b , J. Javier Espinosa-Aguirre c , Teresa Neri-Gómez b , Alberto Rojas-Ochoa f , Christian Guerra-Araiza b , Elena Larrieta f , Paz Vital f,b , Georgina Díaz d , Roberto Chavira e , Cristina Fernandez-Mejia f,∗ a Departamento de Biología Celular, Instituto de Investigaciones Biomédicas, Universidad Nacional Autónoma de México, Mexico b Facultad de Química, Departamento de Biología, Universidad Nacional Autónoma de México, Mexico c Departamento de Medicina y Toxicología, Instituto de Investigaciones Biomédicas, Universidad Nacional Autónoma de México, Mexico d Bioterio, Instituto de Investigaciones Biomédicas, Universidad Nacional Autónoma de México, Mexico e Instituto de Ciencias Médicas y Nutrición Salvador Zubirán, Mexico f Unidad de Génética de la Nutrición, Instituto de Investigaciones Biomédicas, Universidad Nacional Autónoma de México/Instituto Nacional de Pediatría, Mexico article info Article history: Received 28 September 2008 Received in revised form 29 May 2009 Accepted 3 June 2009 Available online 18 June 2009 Keywords: Biotin Estradiol Progesterone Sex steroid receptors CYP Ovary abstract Biotin deficiency and biotin excess have both been found to affect reproduction and cause teratogenic effects. In the reproductive tract, however, the effects of biotin have not been well established yet. We investigated the effects of varying biotin content diets on the oestrus cycle, ovarian morphology, estradiol and progesterone serum levels, and the uterine mRNA abundance of their nuclear receptors, as well as on the activity of the estradiol-degrading group of enzymes cytochrome P450 (CYP) in the liver. Three-week- old female BALB/cAnN Hsd mice were fed a biotin-deficient, a biotin-control, or a biotin-supplemented diet (0, 7.2 or 400 mol of free biotin/kg diet, respectively) over a period of nine weeks. Striking effects were observed in the biotin-deficient group: mice showed arrested estrous cycle on the day of diestrus and changes in ovary morphology. Estradiol serum concentration increased 49.2% in biotin-deficient mice compared to the control group, while the enzymatic activities of CYP1A2 and CYP2B2 increased (P < 0.05). The mRNA abundance of nuclear estrogen and progesterone receptors decreased in the biotin-deficient mice. In the biotin-supplemented group we found that, in spite of a significant (P < 0.05) decrease in the number of primary and Graafian follicles and in CYP1A2 activities, mice exhibited 105.4% higher serum estradiol concentration than the control group. No changes in the expression of the nuclear receptors were observed. No significant differences were observed in serum progesterone among the groups. Our results indicate that both the deficiency and the excess of biotin have significant effects on the female mouse reproductive system. © 2009 Elsevier Inc. All rights reserved. 1. Introduction Biotin is a water-soluble vitamin that acts as a prosthetic group of carboxylases. Unrelated to this role, biotin also regulates gene expression [1,2], and has a wide repertoire of effects on systemic processes such as development [3,4], immunity [5,6], growth [7] and metabolism [8,9]. Several studies have reported that biotin deficiency and biotin excess affect reproductive functions [10–26]. Biotin deficiency during reproduction can result in high rates of abnormal fetal development and growth, and affect rates of absorption and embryonic death in rodents and in fowl [10–15]. In Drosophila melanogaster, biotin deficiency decreased the number of hatched larvae by about 28% [16]. Biotin deficiency decreased serum testosterone and luteinizing hormone concentration, and induced ∗ Corresponding author at: Av. del Iman #1, 4th Floor, Mexico City, CP 04530, Mexico. Tel.: +52 55 5606 35 58; fax: +52 55 56063489. E-mail address: [email protected] (C. Fernandez-Mejia). a significant degree of sloughing of the seminiferous tubule germi- nal epithelium in adult male rats [17]. Biotin supplementation of the diet (1.4 mol/kg diet) of breeding sows increases reproductive performance, induces oestrus and results in earlier conception than sows with control diets (0.13 mol/kg) [18–20], suggesting that, if sows are to reach their full reproductive potential, the dietary formula should include biotin supplementation. On the other hand, excessive amounts of biotin negatively affect reproduction: in insects, reduced fertility is caused by biotin excess in mosquitoes [21], flies [22], and hidebeetles (Der- mestes maculatus) [23]. In rats, acute administration of biotin (204–409 mol/morning and evening) during pregnancy causes resorption of fetuses and placentae [24,25]. Under such conditions, estrogen therapy helped maintain pregnancies to term, suggesting that biotin affects the endogenous production of estrogen [24–26]. Furthermore, atrophy of the corpus luteum, ovary stroma and massive infiltration of leucocytes in the vaginal lumen have also been observed among adult rats injected with 204 mol of biotin [26]. 0039-128X/$ – see front matter © 2009 Elsevier Inc. All rights reserved. doi:10.1016/j.steroids.2009.06.004
Transcript of Biotin deficiency and biotin excess: Effects on the female reproductive system

B
ATPa
b
c
d
e
f
a
ARRAA
KBEPSCO
1
oepad
oaDht
M
0d
Steroids 74 (2009) 863–869
Contents lists available at ScienceDirect
Steroids
journa l homepage: www.e lsev ier .com/ locate /s tero ids
iotin deficiency and biotin excess: Effects on the female reproductive system
rmida Báez-Saldanaa, Ignacio Camacho-Arroyob, J. Javier Espinosa-Aguirrec,eresa Neri-Gómezb, Alberto Rojas-Ochoaf, Christian Guerra-Araizab, Elena Larrieta f,az Vital f,b, Georgina Díazd, Roberto Chavirae, Cristina Fernandez-Mejia f,∗
Departamento de Biología Celular, Instituto de Investigaciones Biomédicas, Universidad Nacional Autónoma de México, MexicoFacultad de Química, Departamento de Biología, Universidad Nacional Autónoma de México, MexicoDepartamento de Medicina y Toxicología, Instituto de Investigaciones Biomédicas, Universidad Nacional Autónoma de México, MexicoBioterio, Instituto de Investigaciones Biomédicas, Universidad Nacional Autónoma de México, MexicoInstituto de Ciencias Médicas y Nutrición Salvador Zubirán, MexicoUnidad de Génética de la Nutrición, Instituto de Investigaciones Biomédicas, Universidad Nacional Autónoma de México/Instituto Nacional de Pediatría, Mexico
r t i c l e i n f o
rticle history:eceived 28 September 2008eceived in revised form 29 May 2009ccepted 3 June 2009vailable online 18 June 2009
eywords:iotinstradiolrogesteroneex steroid receptors
a b s t r a c t
Biotin deficiency and biotin excess have both been found to affect reproduction and cause teratogeniceffects. In the reproductive tract, however, the effects of biotin have not been well established yet. Weinvestigated the effects of varying biotin content diets on the oestrus cycle, ovarian morphology, estradioland progesterone serum levels, and the uterine mRNA abundance of their nuclear receptors, as well as onthe activity of the estradiol-degrading group of enzymes cytochrome P450 (CYP) in the liver. Three-week-old female BALB/cAnN Hsd mice were fed a biotin-deficient, a biotin-control, or a biotin-supplementeddiet (0, 7.2 or 400 �mol of free biotin/kg diet, respectively) over a period of nine weeks. Striking effectswere observed in the biotin-deficient group: mice showed arrested estrous cycle on the day of diestrusand changes in ovary morphology. Estradiol serum concentration increased 49.2% in biotin-deficient micecompared to the control group, while the enzymatic activities of CYP1A2 and CYP2B2 increased (P < 0.05).
YPvary
The mRNA abundance of nuclear estrogen and progesterone receptors decreased in the biotin-deficientmice. In the biotin-supplemented group we found that, in spite of a significant (P < 0.05) decrease in thenumber of primary and Graafian follicles and in CYP1A2 activities, mice exhibited 105.4% higher serumestradiol concentration than the control group. No changes in the expression of the nuclear receptorswere observed. No significant differences were observed in serum progesterone among the groups. Our
them.
results indicate that bothmouse reproductive syste
. Introduction
Biotin is a water-soluble vitamin that acts as a prosthetic groupf carboxylases. Unrelated to this role, biotin also regulates genexpression [1,2], and has a wide repertoire of effects on systemicrocesses such as development [3,4], immunity [5,6], growth [7]nd metabolism [8,9]. Several studies have reported that biotineficiency and biotin excess affect reproductive functions [10–26].
Biotin deficiency during reproduction can result in high ratesf abnormal fetal development and growth, and affect rates of
bsorption and embryonic death in rodents and in fowl [10–15]. Inrosophila melanogaster, biotin deficiency decreased the number ofatched larvae by about 28% [16]. Biotin deficiency decreased serumestosterone and luteinizing hormone concentration, and induced∗ Corresponding author at: Av. del Iman #1, 4th Floor, Mexico City, CP 04530,exico. Tel.: +52 55 5606 35 58; fax: +52 55 56063489.
E-mail address: [email protected] (C. Fernandez-Mejia).
039-128X/$ – see front matter © 2009 Elsevier Inc. All rights reserved.oi:10.1016/j.steroids.2009.06.004
deficiency and the excess of biotin have significant effects on the female
© 2009 Elsevier Inc. All rights reserved.
a significant degree of sloughing of the seminiferous tubule germi-nal epithelium in adult male rats [17]. Biotin supplementation ofthe diet (1.4 �mol/kg diet) of breeding sows increases reproductiveperformance, induces oestrus and results in earlier conception thansows with control diets (0.13 �mol/kg) [18–20], suggesting that,if sows are to reach their full reproductive potential, the dietaryformula should include biotin supplementation.
On the other hand, excessive amounts of biotin negativelyaffect reproduction: in insects, reduced fertility is caused bybiotin excess in mosquitoes [21], flies [22], and hidebeetles (Der-mestes maculatus) [23]. In rats, acute administration of biotin(204–409 �mol/morning and evening) during pregnancy causesresorption of fetuses and placentae [24,25]. Under such conditions,estrogen therapy helped maintain pregnancies to term, suggesting
that biotin affects the endogenous production of estrogen [24–26].Furthermore, atrophy of the corpus luteum, ovary stroma andmassive infiltration of leucocytes in the vaginal lumen have alsobeen observed among adult rats injected with 204 �mol of biotin[26].
8 Steroi
bsoef
2
2
HAiC
mAta28d0oodaptoa0w7rtWcrdsrtitwF
tetieas
tlT
2
d[
64 A. Báez-Saldana et al. /
In this work we compared the effects of diets with varyingiotin content on the oestrus cycle, estradiol and progesteroneerum levels, ovarian morphology, the uterine mRNA abundancef estradiol and progesterone nuclear receptors, as well as onstradiol-degrading enzymes in the liver among BALB/cAnN Hsdemale mice.
. Experimental
.1. Mice
Mice were handled according to the National Institutes ofealth Guide for the Care and Use of Laboratory Animals (Nationalcademy of Sciences, Washington DC, USA, 1996). All methods used
n this study were approved by the Internal Council and the Animalare Committee of the Biomedical Research Institute.
Three-week-old female BALB/cAnN Hsd mice from the ani-al house at the Biomedical Research Institute of the National
utonomous University of Mexico were used. They were main-ained in barrier conditions with 12 h light/dark cycles, andllowed to consume water and food ad libitum. Three lots of1 mice were divided into three experimental groups (n = 7 or), fed 1 of the 3 alternative Harlan Teklad (Madison, WI, USA)iets: biotin-sufficient or -control (TD-01362), biotin-deficient (TD-1363), or biotin-supplemented (TD-02458), containing 16.4, 0r 410 �mol biotin/kg diet, respectively. The complete informationn all diet components has been published elsewhere [6]. Theseiets contained 30% egg-white solids. Egg white contains avidin,glycoprotein that binds biotin, forming a non-covalent com-
lex that is not absorbed into the blood. The 30% egg white inhe diet contributes about 150 mg/kg of avidin. One milligramf avidin is considered to bind 61.4 nmol biotin. Thus, 150 mg ofvidin binds approximately 9.2 �mol biotin. This makes the TD1363 diet, biotin free. The TD 01362 diet was supplementedith 16.4 �mol biotin/kg, therefore it contained approximately
.2 �mol of free biotin/kg, an amount that provides the dailyequirements of the vitamin [27]. Likewise, the TD 02458 diet con-ained 410 �mol biotin/kg, leaving about 400 �mol of free biotin/kg.
e evaluated the amount of free biotin in diets by a commer-ial enzyme immunosorbent assay kit (as described below). Ouresults showed that no free biotin was present in the deficient-iet (−2.1 �mol of free biotin/kg), in the sufficient diet and in theupplemented diets we found 7.8 and 417 �mol of free biotin/kg,espectively. Food consumption was monitored at 8–9 weeks ofreatment: the average food intake was 2.9 ± 0.4 and 3.3 ± 0.7 g/dayn the control and the supplemented group respectively, thus athis age the daily ingestion of biotin was 1.1 and 68 �mol/(kg bodyeight day), in the control and supplemented groups, respectively.
ood consumption in the deficient mice was 2.1 ± 0.1 g/day.Mice were weighed every week. Starting at seven weeks of diet,
he oestrus cycle was evaluated by daily vaginal smears until thend of experimentation. The oestrus cycle was evaluated accordingo the classical criteria of predominant cell type in the smears [28],.e. diestrus day: predominating leukocytes, proestrus: nucleatedpithelial cells, oestrus: cornified epithelial cells, and metaestrus:combination of cells of all stages. Arrested oestrus cycle was con-
idered after four days at the same stage.After nine weeks of the diet, mice were weighed and bled from
he axillary plexus, under ether anesthesia. Following cervical dis-ocation, the wet weight of the liver, and ovaries was determined.he tissues were frozen at −70 ◦C until used for analysis.
.2. Propionyl CoA carboxylase activity
The specific activity of liver propionyl CoA carboxylase wasetermined using a radioenzymatic assay, as previously described5,6].
ds 74 (2009) 863–869
2.3. Biotin concentration analyses
The concentrations of the vitamin were assessed in mice serumwith a commercial enzyme immunosorbent assay kit following themanufacturer’s instructions (ALPCO Diagnostics, Salem, NH, USA).Manufacturer’s human normal status 0.4–0.8 nmol/L. To determinefree biotin in the diets, 0.5 g of diets were suspended in 25 mldistilled/deonized water, insoluble solids were separated by cen-trifugation at 600 × g for 5 min. Proteins in the supernatant wereprecipitated with 30% TCA and pH was neutralized with NaOH.Biotin was determined with a commercial enzyme immunosorbentassay described above.
2.4. Hormone analyses
Estradiol and progesterone concentration were determined inmouse serum with a commercial enzyme-linked immunosorbentassay kit following the manufacturer’s instructions (Diagnostic Sys-tem Labs, Webster, TX, USA). Since only small amounts of blood canbe collected per mouse, sera from mice of each experimental groupwere divided randomly, half of the mice samples were analyzed forestradiol, and the other half for progesterone.
2.5. Histological analysis
Ovaries of five mice from each experimental group weredissected and fixed in 4% paraformaldehyde in 0.1 M sodium phos-phate buffer, pH 7.4. Fixed tissues were dehydrated and embeddedin paraffin. Sections from each tissue were sliced at 5 �m thick-ness and stained with haematoxylin–eosin. The number of folliclesand corpora lutea in the ovaries of each mouse was determined inequal sections from the different tissues at 100× in an OlympusBX41 microscope (Mellville, NY, USA).
2.6. Cytochrome P450 activity
Preparation of S9 and microsomal fraction: Liver S9 fractionswere prepared according to Maron and Ames [29]. Protein con-centration in microsomal fractions was determined as in Bradford[30]. CYP3A2: Erythromycin N-demethylation was assessed spec-trophotometrically as in Alexidis et al. [31], measuring theproduction of formaldehyde. CYP1A2, CYP2B1, CYP2B2: MicrosomalAlkoxyresorufin O-dealkylases activity was measured spectroflu-orometrically by monitoring the formation of resorufin accordingto Refs. [32,33], with modifications in the excitation and emissionwavelengths which were set at 520 and 585 nm, respectively.
2.7. Analysis of mRNA abundance
Total RNA was isolated from the uterus with the single-stepmethod based on guanidine isothiocyanate/phenol/chloroformextraction using TRIzol reagent (Invitrogen, Carlsbad, CA, USA).RNA concentration was determined by absorbance at 260 nm, andits integrity was confirmed by electrophoresis in 1% denaturingagarose gel. Single-stranded cDNA was synthesized from 0.5 �gof total RNA by reverse-transcription reaction with 500 units ofM-MVL-reverse-transcriptase. The relative expression of nuclearreceptors for estradiol-alpha and progesterone in the uterus wasevaluated in real-time PCR with materials and methods sup-plied by Applied Biosystems. Twenty-five nanograms of cDNA
were mixed with TaqMan® universal PCR master mix, unlabeledPCR primers, and TaqMan® MGB probes Esr1 estrogen receptor-alpha: Mm00433149 m1; Pgr: progesterone receptor (TaqMan®Mm00435625 m1), in a total volume of 25 �l. Real-time PCR wasperformed on an ABI Prism 7700 Sequence Detector. The samples

A. Báez-Saldana et al. / Steroids 74 (2009) 863–869 865
Table 1Effects of nine weeks of diet with varying biotin content on food intake, body, ovaries and liver weight.
Body weight (g) Food intake/bodyweight/day (mg/(g day))
Ovaries weight (mg) Ovaries/body weight(mg/g)
Liver weight (mg) Liver weight/bodyweight (mg/g)
Control 20 ± 0.2a 140 + 21a 50 ± 5.6a 2.6 ± 0.3a 985 + 37a 50 + 1.6a
Deficient 16 ± 0.4b 132 + 6a 17 ± 1.5b 1.0 ± 0.1b 804 + 24b 51 + 1.4a
S
n by ond
wufrb[
2
acmAitT
3
3
tagpbbmic
Fimgmda
upplemented 20 ± 0.6a 169 + 38a 44 ± 3.8a
= 12 for each group. Data are expressed as mean + SEM. Significance was assessedifferences (P < 0.05) between groups.
ere analyzed in triplicate and corrected for the 18S ribosomal sub-nit used as internal standard. Standard curves were constructedrom 0.1 to 10 ng of cDNA from untreated mouse uterus. Relativeeceptor expression was calculated from cycle threshold (Ct) valuesy the abundant relative quantification model of the delta–delta Ct34].
.8. Statistics
The PRISM 2.01 program (Graph Pad, La Jolla, CA, USA)nd Statview statistical analysis program V.4.5 (Abacus Con-epts, Berkeley, CA, USA) were used. Data are presented as theean ± SEM Multiple comparisons were evaluated by one-wayNOVA, followed by Student–Newman–Keul’s multiple compar-
son procedure. Body weight comparisons were assessed bywo-way ANOVA (diet, time) analysis of variance, and post hocukey test. The level of significance was P < 0.05.
. Results
.1. Body, ovaries and liver weight
We determined mouse body weight every week (Fig. 1). Mice inhe biotin-control group and biotin-supplemented groups showedsteady weight gain during the nine weeks of the study. Beside
lowing hair, in the biotin-supplemented group we did not noticehysical changes. The growth of biotin-deficient mice showed a
iphasic pattern: at first, weight increased slower than in theiotin-control and biotin-supplemented groups, reaching maxi-um growth at 4 weeks of diet; thereafter, we observed a decreasen body weight. In addition to the smaller size of mice in the defi-ient group, they showed clinical symptoms associated with biotin
ig. 1. Growth curve. Three-week-old female BALB/cAnN Hsd mice were fed vary-ng biotin concentrations. Body weight was measured every week. Values are the
ean ± SEM. n = 14. Circles: control group; squares: biotin-deficient group; trian-les: supplemented group. Significance was assessed by the two-factor repeatedeasurements ANOVA (diet, time) analysis of variance. Letters indicate significant
ifferences, P < 0.05. (a) Different from sufficient, (b) different from supplemented,nd (c) different from sufficient and supplemented.
2.3 ± 0.2a 1010 + 46a 51 + 2.5a
e-way ANOVA analysis of variance. Different letter superscripts indicate significant
deficiency such as alopecia and dermatitis, these signs started tobe observed after 6 weeks on biotin deficiency. The amount offood intake/body weight was not significantly different betweenthe groups (Table 1). Biotin deficiency caused a significant decreasein (P < 0.05) ovarian weight (Table 1). To determine whether theorgan was affected by corporal weight changes induced by the diets,we calculated the ovary weight/body weight ratio of each mouse.We found that this ratio was significantly lower in mice fed thebiotin-deficient diet than in mice fed either the control- or the sup-plemented diet (Table 1), while the ratio of liver weight to bodyweight (Table 1), showed no differences between the groups.
3.2. Biotin status
The enzymatic activity of liver propionyl-CoA carboxylase andbiotin content in serum was used as indicators of biotin sta-tus. Propionyl-CoA carboxylase activity significantly decreased(P < 0.05) in the liver of the biotin-deficient diet animals ascompared with the control group. No significant differencesin PCC activity were observed between the mice receivingeither the biotin-control or the biotin-supplemented diet (biotin-control: 21 ± 1.5; biotin-deficient: 3.9 ± 0.4; biotin-supplemented20 ± 1.1 (nmol CO2 fixed min−1 mg prot−1 g; n = 7 for each group).We also analyzed serum biotin concentration, and observedsignificant differences (P < 0.05) between the groups (biotin-control: 35 ± 4.1; biotin-deficient: 0.7 ± 0.2; biotin-supplemented1272 ± 220 nmol/L; n = 4–5).
3.3. Estrous cycle
All mice fed a biotin-deficient diet exhibited arrested estrouscycle on the day of diestrus. We observed no irregularities in theestrous cycle neither in the control diet group nor in the biotin-supplemented group.
3.4. Serum estradiol and progesterone concentration
Since all biotin-deficient mice manifested arrested estrous cycleat the day of diestrus, we compared the effect of the diets withvarying biotin levels on serum estradiol and progesterone concen-
tration on the day of diestrus. We found that estradiol concentrationwas significantly (P < 0.05) higher in both the deficient and supple-mented groups (Table 2). In the biotin-deficient group, estradiolconcentration was 49% higher than in the control group, and inthe biotin-supplemented group we found a pronounced increase ofTable 2Effect of nine weeks of diet with varying biotin content on serum estradiol andprogesterone concentration on the day of diestrus.
Diet Estradiol (pmol/L) Progesterone (nmol/L)
Control 35 + 2.2a 7.8 + 1.6a
Deficient 53 + 5.2b 9.2 + 1.5a
Supplemented 73 + 6.0c 8.0 + 2.2a
n = 9–10 mice for each group. Data are expressed as mean + SEM. Significance wasassessed by one-way ANOVA analysis of variance. Different letter superscripts indi-cate significant differences (P < 0.05) between groups.
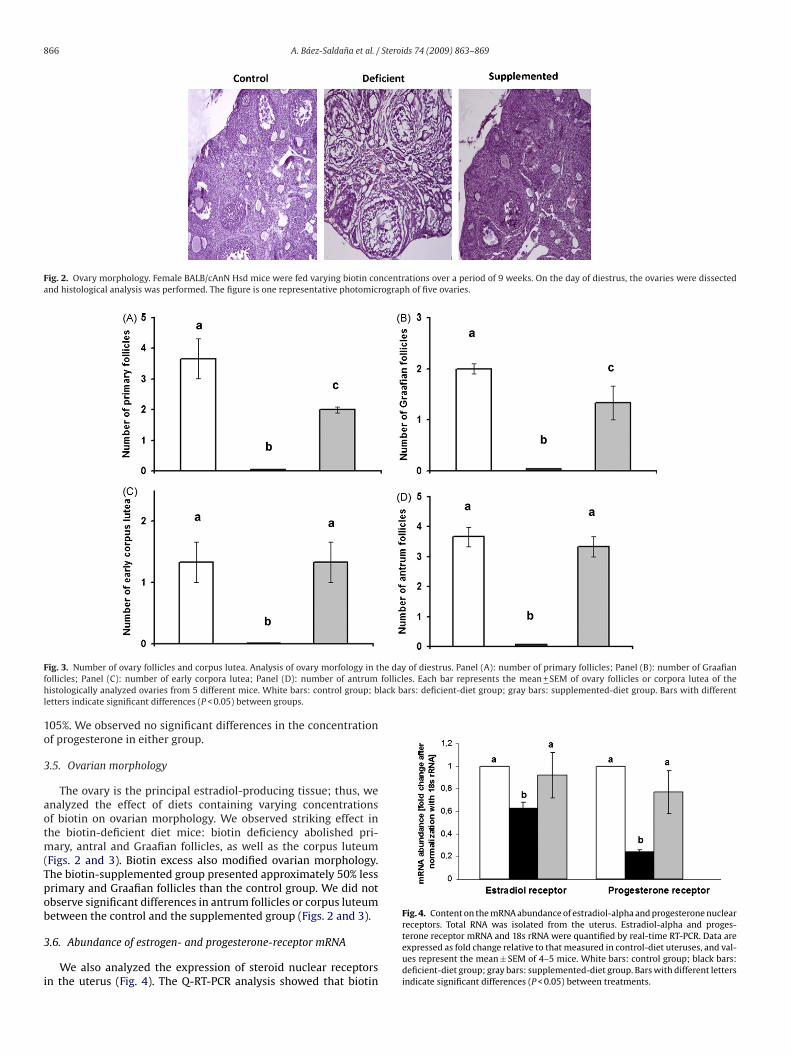
866 A. Báez-Saldana et al. / Steroids 74 (2009) 863–869
Fig. 2. Ovary morphology. Female BALB/cAnN Hsd mice were fed varying biotin concentrations over a period of 9 weeks. On the day of diestrus, the ovaries were dissectedand histological analysis was performed. The figure is one representative photomicrograph of five ovaries.
F he day of diestrus. Panel (A): number of primary follicles; Panel (B): number of Graafianf follicles. Each bar represents the mean + SEM of ovary follicles or corpora lutea of theh lack bars: deficient-diet group; gray bars: supplemented-diet group. Bars with differentl
1o
3
aotm(Tpob
3
i
Fig. 4. Content on the mRNA abundance of estradiol-alpha and progesterone nuclear
ig. 3. Number of ovary follicles and corpus lutea. Analysis of ovary morfology in tollicles; Panel (C): number of early corpora lutea; Panel (D): number of antrumistologically analyzed ovaries from 5 different mice. White bars: control group; b
etters indicate significant differences (P < 0.05) between groups.
05%. We observed no significant differences in the concentrationf progesterone in either group.
.5. Ovarian morphology
The ovary is the principal estradiol-producing tissue; thus, wenalyzed the effect of diets containing varying concentrationsf biotin on ovarian morphology. We observed striking effect inhe biotin-deficient diet mice: biotin deficiency abolished pri-
ary, antral and Graafian follicles, as well as the corpus luteumFigs. 2 and 3). Biotin excess also modified ovarian morphology.he biotin-supplemented group presented approximately 50% lessrimary and Graafian follicles than the control group. We did notbserve significant differences in antrum follicles or corpus luteumetween the control and the supplemented group (Figs. 2 and 3).
.6. Abundance of estrogen- and progesterone-receptor mRNA
We also analyzed the expression of steroid nuclear receptorsn the uterus (Fig. 4). The Q-RT-PCR analysis showed that biotin
receptors. Total RNA was isolated from the uterus. Estradiol-alpha and proges-terone receptor mRNA and 18s rRNA were quantified by real-time RT-PCR. Data areexpressed as fold change relative to that measured in control-diet uteruses, and val-ues represent the mean ± SEM of 4–5 mice. White bars: control group; black bars:deficient-diet group; gray bars: supplemented-diet group. Bars with different lettersindicate significant differences (P < 0.05) between treatments.

A. Báez-Saldana et al. / Steroids 74 (2009) 863–869 867
Table 3Effect of nine weeks of diet with varying biotin content on hepatic cytochrome p450 activities.
Diet CYP1A2 (pmol mg−1 protein−1 min) CYP2B1 (pmol mg−1 protein−1 min) CYP2B2 (pmol mg−1 protein−1 min) CYP3A2 (nmol mg−1 protein−1 min)
Control 32 ± 4.6a 5.3 ± 1.2a 16 ± 1.2a 2.5 ± 0.2a
Deficient 51 ± 3.9b 6.2 ± 0.6a 25 ± 2.8b 2.4 ± 0.9a
S b a a a
n M. Sis
drbn
3
Tddiitid
4
eoamb
eWccdald(vgicwdoscwgs(cetit
tb
upplemented 40 ± 7.1 5.5 ± 0.1
= Pool of 4 different mice from each group. Data are expressed as the mean + SEuperscripts indicate significant differences (P < 0.05) between groups.
eficiency significantly reduced mRNA abundance of progesteroneeceptor and estradiol receptor, while the diet supplemented withiotin did not modify mRNA abundance of neither, progesteroneor estradiol receptor.
.7. Estradiol catabolic enzymes
The catabolism of estradiol largely occurs in the liver [35].o determine whether the increase in estradiol level was due toecreased estradiol degradation, we analyzed the activity of estra-iol CYP catabolic enzymes in the hepatic tissue (Table 3). We found
ncreases in CYP1A2 activity in both the biotin-deficient group andn the biotin-supplemented group (P < 0.05) compared to the con-rol group. Additionally, increased activity of CYP2B2 was also foundn the biotin-deficient mice. No differences in CYP3A activity wereetected among the groups.
. Discussion
The present study allowed a simultaneous comparison of theffects of biotin deficiency and excess on the reproductive systemf female mice. Our results demonstrated that both, the deficiencynd the excess of biotin have significant effects on the femaleouse reproductive system, although some differences were found
etween the effects of these two diets.Mice fed a biotin-supplemented diet presented increased serum
stradiol concentration, and decreased ovarian primary follicles.e observed no significant changes in serum progesterone con-
entration. We did not observe significant changes in the estrousycle, or in the number of corpora lutea in the ovary. Our resultsiffer from a previous report by Paul et al. [26] who found thatdisruption of the estrous cycle and atrophy of the corpora
utea were caused by acute biotin treatment, consisting of twoaily (morning and evening) subcutaneous injections of biotin205 �mol/kg body weight). A number of reasons may explain theariations in response to biotin reported by the Paul et al. [26]roup in contrast with ours. (a) The variation in dose and differencesn administration route: our group of mice received excess biotin,onsuming approximately 68 �mol/(kg body weight day) along 9eeks, while in the Paul et al. studies [26] two subcutaneous biotinoses consisting of 205 �mol/kg body weight were administeredn the day of diestrus. (b) Differences in biotin sensitivity betweenpecies: it has been reported that biotin deficiency occurs moreommonly in mice than in rats [36]. Maternal biotin deficiencyas extremely lethal and teratogenic in mice, but in rats, terato-
enic or fetal losses were not so overt [3,37]. Contrary to this, sometudies [24–26] have reported that excessive amounts of biotin205 �mol/kg body weight) caused resorption of fetuses and pla-enta in rats but did not disturb normal reproductive functions andmbryonic development in ICR mice [4]. Thus, it may be the casehat, contrary to the higher sensitivity of mice with respect to rats
n terms of biotin deficiency; mice are less sensitive to biotin excesshan rats.A relationship between biotin status and estradiol concen-ration has been reported [24,25]. In rats, an acute dose ofiotin (410 �mol/kg body weight), either at the pre-implantation
13 ± 0.9 2.2 ± 0.3
gnificance was assessed by one-way ANOVA analysis of variance. Different letter
or post-implantation stage, caused resorption of fetuses andplacentae. Under such conditions, estradiol therapy maintainedpregnancy with normal fetal and placental growth, suggestingthat biotin-induced loss of pregnancy was produced by estro-gen production blockage. However, Watanabe [4] found thatin ICR pregnant mice, biotin excess (three acute subcutaneousinjections of 205 �mol biotin/kg body weight) or a diet includingsupplementary biotin (4.1 mmol/kg diet) did not change estradiolconcentration. In contrast to Watanabe’s observations, we consis-tently found that BALB/cAnN Hsd mice fed a biotin-supplementeddiet (68 �mol/(body weight day)) showed increased estradiol con-centration.
It has been widely reported that estradiol and progesteroneregulate the expression of their respective receptors [38–41]. Inthe whole uterine homogenates of many mammals’ species it hasbeen reported that estradiol down-regulates its receptor [39–41].In biotin-supplemented mice we found that, albeit the high levelsof serum estradiol, the mRNA abundance of its nuclear receptor inthe uterus was not reduced, suggesting that biotin excess impairsthe normal regulation of estradiol receptor expression by its ligand.Further studies will be designed to generate additional insights intobiotin effects on estrogen receptors.
We observed striking effects in the biotin-deficient group. Alongwith decreased body weight, we observed reductions in ovaryweight and in the relative weight of the ovary (ovary weight/bodyweight). We also found arrested estrous cycle in the day of diestrus,and notable changes in ovarian morphology. Histological analy-sis revealed that biotin deficiency abolished primary, antral andGraafian follicles, as well as the corpus luteum. As previously shown[7] these changes cannot simply ascribed to decreased food intakeor reduced appetite because the relative food ingestion in the con-trol and the biotin-deficient mice is equivalent, however, they mightbe related to decreased levels of IGF-1. Several studies [42–44]have found that maturation of ovarian follicles is associated withhepatic regulation of IGF-I and systemic concentrations of IGF-Ibinding proteins. We have recently reported [7] that biotin defi-ciency decreases serum IGF-1 concentrations; it might be plausiblethen, that the decreased levels of this growth factor accounts for theeffects observed in ovarian follicles in the biotin-deficient mice.
In previous studies [37] biotin deficiency immediately beforemating did not show any effects in rats during gestation. How-ever, rats fed a biotin-free diet for five weeks before being matedwere unable to produce viable products [45]. In our laboratory,we have observed that, conform to the inhibition of ovary matu-ration and arrest on diestrus day when there are not present thephysiological conditions in order to be fertilized and pregnant,mice fed a biotin-deficient diet during eight weeks are incapableto achieve pregnancy. Taken together these observations suggestthat biotin deficiency for only three weeks of the gestational periodof female rats may not be sufficient to produce its overt effecton pregnancy and that longer periods of deficiency are required
to produce the detrimental effects, however, we cannot rule outthe possibility that, as stated previously, differences in the effectsof biotin between mice and rats [3,36,37] may account in the dis-similar effects of the vitamin deficiency in reproduction betweenspecies.
8 Steroi
scdoitd
aTbhrgTt
C[oddibasTtaibet[tt
tcocabi[crbltd[c[rb
A
atCE
[
[[
[
[
[
[
[
[
[
[
[
[
68 A. Báez-Saldana et al. /
Similarly to the observed in the biotin-supplemented group wehow that biotin deficiency increased estradiol concentration. Inontrast to the documented effects of biotin excess on this hormone,escribed above, there are no previous reports describing the effectf biotin deficiency on the serum concentrations of estradiol. Thencrease in serum estradiol is puzzling, because the ovary, which ishe main source of estradiol, was severely damaged in the biotin-eficient group.
In the uterus of biotin-deficient mice we found reduced mRNAbundance of both, progesterone- and estradiol-receptors (Fig. 4).he reduced expression of estradiol-receptor in the uterus coulde related to augmented serum estradiol concentrations [39–41],owever, the reduction in the expression of the progesteroneeceptor cannot be explained by increases in its ligand, since pro-esterone serum levels were not significantly modified (Table 2).aken together, the data suggest that biotin-deficiency may affecthe expression of steroid hormone receptors.
The catabolism of estradiol largely occurs in the liver by theYP1A2, 2B1, 2B2, 3A family and its product is 2-hydroxyestradiol46]. To determine whether the increases in serum estradiolbserved in either the deficient and the supplemented groups wereue to decreased hormone catabolism, we analyzed the effect of theiets with varying biotin levels on several P450 isoenzyme activ-
ties. Contrary to the expected we found that in the liver of theiotin-deficient mice, the activity of CYP1A2 and CYP2B2 increasedbout 60% (P < 0.05) compared with the control group. In the biotin-upplemented group (Table 3), hepatic CYP1A2 activity rose by 25%.hese results rule out the possibility that the increases observed inhe concentrations of serum estradiol in both, the biotin-deficientnd supplemented groups, are related to a decrease in the activ-ty of estradiol catabolic enzymes. It is interesting to note thatiotin supplementation affects one of the enzymes involved instradiol catabolism in extrahepatic tissues [47]. Biotin supplemen-ation increases CYP1B1 expression and activity in lymphocytes47]. Taken together, these findings suggest that biotin is involved inhe modulation of some CYP genes. We are presently investigatinghe implications of these findings.
Biotin deficiency and biotin excess have been useful tools tohrow light on the role of the vitamin in diverse systemic pro-esses [1,2]. It is generally accepted that pharmacological dosesf biotin are harmless [48], despite the fact that pharmacologi-al doses of biotin in commercial formulae have been shown toffect gene expression and physiological functions [1,2,49,50]. Theiotin content in commercially available supplements for humans
s 40–166-fold increase the reference dietary intake of 30 �g/day51]. The present work showed that a 55-fold increase in biotin con-entration in the diet modifies several aspects of the female mouseeproductive system. These observations indicate that the use ofiotin needs to be carefully readdressed in view that pharmaco-
ogical concentrations of biotin affect physiological functions. Onhe other hand, several sources of evidence suggest that a marginalegree of biotin deficiency develops during normal pregnancy52–54]. In mice, degrees of biotin deficiency that are metaboli-ally similar to those seen in pregnant women are very teratogenic55]. The clinical significance of a marginal deficiency in humansemains unclear, but mice studies raise concern about the role ofiotin deficiency on teratogenicity and sterility.
cknowledgements
The authors are grateful to Milena Saqui-Salces for technicalssistance in histological, to Georgina Del Vecchyo for body andissues weight measurements, and Sandra L. Hernández Ojeda forYP analysis. We also thank Isabel Pérez Montfort for correcting thenglish version of the manuscript. Supported by research grants
[
ds 74 (2009) 863–869
from the Dirección General de Asuntos del Personal Académico:IN208605, IN 230905 Universidad Nacional Autónoma de México,and Consejo Nacional de Ciencia y Tecnología CONACyT 52746.
References
[1] Rodriguez-Melendez R, Zempleni J. Regulation of gene expression by biotin. JNutr Biochem 2003;14:680–90.
[2] Zempleni J. Uptake, localization, and noncarboxylase roles of biotin. Annu RevNutr 2005;25:175–96.
[3] Watanabe T, Endo A. Teratogenic effects of maternal biotin deficiency on mouseembryos examined at midgestation. Teratology 1990;42:295–300.
[4] Watanabe T. Morphological and biochemical effects of excessive amountsof biotin on embryonic development in mice. Experientia 1996;52:149–54.
[5] Báez-Saldana A, Díaz G, Espinoza B, Ortega E. Biotin deficiency induces changesin subpopulations of spleen lymphocytes in mice. Am J Clin Nutr 1998;67:431–7.
[6] Báez-Saldana A, Ortega E. Biotin deficiency blocks thymocyte maturation accel-erates thymus involution, and decreases nose-rump length in mice. J Nutr2004;134:1970–7.
[7] Báez-Saldana A, Gutiérrez-Ospina G, Chimal-Monroy J, Fernandez-Mejia C,Saavedra R. Biotin deficiency in mice is associated with decreased serum avail-ability of insulin-like growth factor-I. Eur J Nutr 2009;48:137–44.
[8] Dakshinamurti K. Biotin: a regulator of gene expression. J Nutr Biochem2005;16:419–23.
[9] Fernandez-Mejia C. Pharmacological effects of biotin. J Nutr Biochem 2005;16:424–7.
[10] Cravens WW, McGibbon WH, Sebesta EE. Effect of biotin deficiency on embry-onic development in the domestic fowl. Anat Rec 1994;90:55–64.
[11] Cooper WA, Brown SO. Tissue abnormalities in newborn rats from biotin defi-cient mothers. Tex Sci 1958;10:60–8.
12] Giroud A, Lefebvre J, Depuis R. Carence de biotine et reproduction chez la ratte.Comptes Rendus Soc Biol 1956;150:2066–7.
[13] Watanabe T. Dietary biotin deficiency affects reproductive function and prena-tal development in hamsters. J Nutr 1993;123:2101–8.
[14] Mock DM, Mock NI, Stewart CW, LaBorde JB, Hansen DK. Marginal biotin defi-ciency is teratogenic in ICR mice. J Nutr 2003;133:2519–25.
[15] Mock DM. Marginal biotin deficiency is teratogenic in mice and perhapshumans: a review of biotin deficiency during human pregnancy and effectsof biotin deficiency on gene expression and enzyme activities in mouse damand fetus. J Nutr Biochem 2005;16:435–7.
[16] Landenberger A, Kabil H, Harshman LG, Zempleni J. Biotin deficiency decreaseslife span and fertility but increases stress resistance in Drosophila melanogaster.J Nutr Biochem 2004;15:591–600.
[17] Paulose CS, Thliversi JA, Viswanathan M, Dakshinamurti K. Testicular functionin biotin-deficient adult rats. Horm Metab Res 1989;21:661–5.
[18] Simmins PH, Brooks PH. Supplementary biotin for sows: effect on reproductivecharacteristics. Vet Rec 1983;112:425–9.
[19] Brooks PH, Smith DA, Irwin VC. Biotin-supplementation of diets: the inci-dence of foot lesions, and the reproductive performance in sows. Vet Rec1977;101:46–50.
20] Kornegay ET. Biotin in swine nutrition. Ann N Y Acad Sci 1985;447:112–21.21] Pillai MKK, Madhukar BVR. Effect of biotin on the fertility of the yellow fever
mosquito, Aedes aegypti. Naturwissenschaften 1969;56:218–9.22] Benschoter CA. Effect of dietary biotin on reproduction of the house fly. J Econ
Entomol 1967;60:1326–8.23] Cohen E, Levinson HZ. Disrupted fertility of the hidebeetle Dermestes maculatus
(Deg) due to dietary overdosage of biotin. Experientia 1968;24:367–8.24] Paul PK, Duttagupta PN. The effect of an acute dose of biotin at the pre-
implantation stage and its relation with female sex steroids in the rat. J Nutr SciVitaminol (Tokyo) 1975;21:89–101.
25] Paul PK, Duttagupta PN. The effect of an acute dose of biotin at a post-implantation stage and its relation with female sex steriods in the rat. J Nutr SciVitaminol (Tokyo) 1976;22:181–6.
26] Paul PK, Duttagupta PN, Agarwal HC. Effects of an acute dose of biotin on thereproductive organs of the female rat. Curr Sci 1973;42:206–8.
27] National Research Council. Nutrient Requirements of Laboratory Mice. 4th rev.ed. Washington, DC: National Academic Press; 1995. pp. 47–49.
28] Nelson JN, Felicio LS, Randall PK, Sims C, Finch CE. A longitudinal study of estrouscyclicity in aging C57BL/6J mice: I. Cycle Frequency, length and vaginal cytology.Biol Reprod 1982;27:327–39.
29] Maron DM, Ames BN. Revised methods for the Salmonella mutagenicity test.Mutat Res 1983;113:173–215.
30] Bradford MM. A rapid and sensitive method for the quantification of micro-gram quantities of protein utilizing the principle of protein-dye binding. AnalBiochem 1976;72:248–54.
31] Alexidis AN, Commandeur JN, Rekka EA. Novel piperidine derivatives:inhibitory properties towards cytochrome P450 isoforms, and cytopro-
tective and cytotoxic characteristics. Environ Toxicol Pharmacol 1996;1:81–8.32] Burke MD, Thompson S, Elcombe CR, Halpert J, Haaparanta T, Mayer RT. Ethoxy-,pentoxy-, and benzyloxyphenoxazones and homologues: a series of substratesto distinguish between different induced cytochromes P-450. Biochem Phar-macol 1985;34:3337–45.

Steroi
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
A. Báez-Saldana et al. /
33] Burke MD, Thompson S, Weaver RJ, Wolf CR, Mayer RT. Cytochrome P450specificities of alkoxyresorufin O-dealkylation in human and rat liver. BiochemPharmacol 1994;48:923–36.
34] Livak KJ, Schmittgen TD. Analysis of relative gene expression data usingreal-time quantitative PCR and the 2(−Delta Delta C(T)) method. Methods2001;25:402–8.
35] Tsuchiya Y, Nakajima M, Yokoi T. Cytochrome P450-mediated metabolism ofestrogens and its regulation in human. Cancer Lett 2005;227:115–24.
36] Rabin BS. Inhibition of experimentally induced autoimmunity in rats by biotindeficiency. J Nutr 1983;113:2316–22.
37] Watanabe T, Endo A. Species and strain differences in teratogenic effects ofbiotin deficiency in rodents. J Nutr 1989;119:255–61.
38] Guerra-Araiza C, Camacho-Arroyo I. Progesterone receptor isoforms: functionand regulation. Rev Invest Clin 2000;52:686–91.
39] Cidlowski JA, Muldoon TG. Estrogenic regulation of cytoplasmic recep-tor populations in estrogen-responsive tissues of the rat. Endocrinology1974;95:1621–9.
40] Medlock KL, Lyttle CR, Kelepouris N, Newman ED, Sheehan DM. Estradioldown-regulation of the rat uterine estrogen receptor. Proc Soc Exp Biol Med1991;196:293–300.
41] Morris ID. Changes in brain, pituitary and uterine cytoplasmic oestrogen recep-tors induced by oestradiol-17beta in the ovariectomized rat. J Endocrinol1976;71:343–9.
42] Diskin MG, Mackey DR, Roche JF, Sreenan JM. Effects of nutrition and metabolicstatus on circulating hormones and ovarian follicle development in cattle. Anim
Reprod Sci 2003;78:345–70.43] Spicer LJ, Echternkamp SE. The ovarian insulin and insulin-like growth fac-tor system with an emphasis on domestic animals. Domest Anim Endocrinol1995;12:223–45.
44] Richards JS, Russell DL, Ochsner S, Hsieh M, Doyle KH, Falender AE, etal. Novel signaling pathways that control ovarian follicular develop-
[
[
[
ds 74 (2009) 863–869 869
ment, ovulation, and luteinization. Recent Prog Horm Res 2002;57:195–220.
45] Kennedy C, Palmer LS. Biotin deficiency in relation to reproduction and lacta-tion. Arch Biochem 1945;7:9–13.
46] Zhu BT, Conney AH. Functional role of estrogen metabolism in target cells:review and perspectives. Carcinogenesis 1998;19:1–27. Review.
47] Rodriguez-Melendez R, Griffin JB, Zempleni J. Biotin supplementation increasesexpression of the cytochrome P450 1B1 gene in Jurkat cells, increasing theoccurrence of single-stranded DNA breaks. J Nutr 2004;134:2222–8.
48] Zempleni J. In: Bowman BA, Russell RN, editors. Biotin present knowledge innutrition. Washington, DC: ILSI Press; 2001.
49] Wiedmann S, Eudy JD, Zempleni J. Biotin supplementation increasesexpression of genes encoding interferon-gamma, interleukin-1beta, and3-methylcrotonyl-CoA carboxylase, and decreases expression of the geneencoding interleukin-4 in human peripheral blood mononuclear cells. J Nutr2003;133:716–9.
50] Zempleni J, Helm RM, Mock DM. In vivo biotin supplementation at a phar-macologic dose decreases proliferation rates of human peripheral bloodmononuclear cells and cytokine release. J Nutr 2001;131:1479–84.
51] Food Nutrition Board. Institute of Medicine: Dietary reference intakes forthiamine, riboflavin, niacin, vitamin B6, folate, vitamin B12, panthotenicacid biotin and choline. Washington, DC: National Academy Press; 1998. pp.374–389.
52] Mock DM, Quirk JG, Mock NI. Marginal biotin deficiency during normal preg-nancy. Am J Clin Nutr 2002;75:295–9.
53] Mock DM, Stadler DD. Conflicting indicators of biotin status from a cross-sectional study of normal pregnancy. J Am Coll Nutr 1997;16:252–7.
54] Mock DM, Stadler DD, Stratton SL, Mock NI. Biotin status assessed longitudinallyin pregnant women. J Nutr 1997;127:710–6.
55] Mock DM. Marginal biotin deficiency is common in normal human pregnancyand is highly teratogenic in mice. J Nutr 2009;139:154–7.




















