
Biology of the cell
28
75 How Do the Cells of a Growing Plant Know in Which Direction to Elongate? Sometimes questions that seem simple can be devilishly dif- ficult to answer. Imagine, for example, that you are holding a green blade of grass in your hand. The grass blade has been actively growing, its cells dividing and then stretching and elongating as the blade lengthens. Did you ever wonder how the individual cells within the blade of grass know in what direction to grow? To answer this deceptively simple question, we will first need to provide answers to several others. Like Sherlock Holmes following a trail of clues, we must approach the an- swer we seek in stages. Question One. First, we need to ask how a blade of grass is able to grow at all. Plant cells are very different from ani- mal cells in one key respect: every plant cell is encased within a tough cell wall made of cellulose and other tough building materials. This wall provides structural strength and protection to the plant cell, just as armor plate does for a battle tank. But battle tanks can’t stretch into longer shapes! How is a plant cell able to elongate? It works like this. A growing cell first performs a little chemistry to make its wall slightly acidic. The acidity acti- vates enzymes that attack the cell wall from the inside, rear- ranging cellulose cross-links until the wall loses its rigidity. The cell wall is now able to stretch. The cell then sucks in water, creating pressure. Like blowing up a long balloon, the now-stretchable cell elongates. Question Two. In a growing plant organ, like the blade of grass, each growing cell balloons out lengthwise. Stating this more formally, a botanist would say the cell elongates parallel to the axis along which the blade of grass is extend- ing. This observation leads to the second question we must answer: How does an individual plant cell control the di- rection in which it elongates? It works like this. Before the stretchable cell balloons out, tiny microfibrils of cellulose are laid down along its inside sur- face. On a per weight basis, these tiny fibrils have the tensile strength of steel! Arrays of these cellulose microfibrils are organized in bands perpendicular to the axis of elongation, like steel belts. These tough bands reinforce the plant cell wall laterally, so that when the cell sucks in water, there is only one way for the cell to expand—lengthwise, along the axis. Question Three. Now we’re getting somewhere. How are the newly made cellulose microfibrils laid down so that they are oriented correctly, perpendicular to the axis of elongation? It works like this. The complicated enzymic machine that makes the cellulose microfibrils is guided by special guiderails that run like railroad tracks along the interior sur- face. The enzyme complex travels along these guiderails, laying down microfibrils as it goes. The guiderails are con- structed of chainlike protein molecules called microtubules, assembled into overlapping arrays. Botanists call these ar- rays of microtubules associated with the interior of the cell surface “cortical microtubules.” Question Four. But we have only traded one puzzle for another. How are the cortical microtubules positioned cor- rectly, perpendicular to the axis of elongation? It works like this. In newly made cells, the microtubule assemblies are already present, but are not organized. They simply lie about in random disarray. As the cell prepares to elongate by lessening the rigidity of its cell wall, the micro- tubule assemblies become organized into the orderly trans- verse arrays we call cortical microtubules. Question Five. Finally, we arrive at the question we had initially set out to answer. How are microtubule assemblies aligned properly? What sort of signal directs them to ori- ent perpendicular to the axis of elongation? THAT is the question we need to answer. Part II Biology of the Cell Seeing cortical microtubules. Cortical microtubules in epidermal cells of a fava bean are tagged with a flourescent protein so that their ordered array can be seen. Real People Doing Real Science
Transcript of Biology of the cell

75
How Do the Cells of a GrowingPlant Know in Which Direction to Elongate? Sometimes questions that seem simple can be devilishly dif-ficult to answer. Imagine, for example, that you are holdinga green blade of grass in your hand. The grass blade hasbeen actively growing, its cells dividing and then stretchingand elongating as the blade lengthens. Did you ever wonderhow the individual cells within the blade of grass know inwhat direction to grow?
To answer this deceptively simple question, we will firstneed to provide answers to several others. Like SherlockHolmes following a trail of clues, we must approach the an-swer we seek in stages.
Question One. First, we need to ask how a blade of grassis able to grow at all. Plant cells are very different from ani-mal cells in one key respect: every plant cell is encasedwithin a tough cell wall made of cellulose and other toughbuilding materials. This wall provides structural strengthand protection to the plant cell, just as armor plate does fora battle tank. But battle tanks can’t stretch into longershapes! How is a plant cell able to elongate?
It works like this. A growing cell first performs a littlechemistry to make its wall slightly acidic. The acidity acti-vates enzymes that attack the cell wall from the inside, rear-ranging cellulose cross-links until the wall loses its rigidity.The cell wall is now able to stretch. The cell then sucks inwater, creating pressure. Like blowing up a long balloon,the now-stretchable cell elongates.
Question Two. In a growing plant organ, like the bladeof grass, each growing cell balloons out lengthwise. Statingthis more formally, a botanist would say the cell elongatesparallel to the axis along which the blade of grass is extend-ing. This observation leads to the second question we mustanswer: How does an individual plant cell control the di-rection in which it elongates?
It works like this. Before the stretchable cell balloons out,tiny microfibrils of cellulose are laid down along its inside sur-face. On a per weight basis, these tiny fibrils have the tensilestrength of steel! Arrays of these cellulose microfibrils are
organized in bands perpendicular to the axis of elongation,like steel belts. These tough bands reinforce the plant cell walllaterally, so that when the cell sucks in water, there is only oneway for the cell to expand—lengthwise, along the axis.
Question Three. Now we’re getting somewhere. Howare the newly made cellulose microfibrils laid down so thatthey are oriented correctly, perpendicular to the axis ofelongation?
It works like this. The complicated enzymic machine thatmakes the cellulose microfibrils is guided by specialguiderails that run like railroad tracks along the interior sur-face. The enzyme complex travels along these guiderails,laying down microfibrils as it goes. The guiderails are con-structed of chainlike protein molecules called microtubules,assembled into overlapping arrays. Botanists call these ar-rays of microtubules associated with the interior of the cellsurface “cortical microtubules.”
Question Four. But we have only traded one puzzle foranother. How are the cortical microtubules positioned cor-rectly, perpendicular to the axis of elongation?
It works like this. In newly made cells, the microtubuleassemblies are already present, but are not organized. Theysimply lie about in random disarray. As the cell prepares toelongate by lessening the rigidity of its cell wall, the micro-tubule assemblies become organized into the orderly trans-verse arrays we call cortical microtubules.
Question Five. Finally, we arrive at the question we hadinitially set out to answer. How are microtubule assembliesaligned properly? What sort of signal directs them to ori-ent perpendicular to the axis of elongation? THAT is thequestion we need to answer.
Part IIBiology of the Cell
Seeing cortical microtubules. Cortical microtubules inepidermal cells of a fava bean are tagged with a flourescentprotein so that their ordered array can be seen.
Rea
l Peo
ple
Doi
ng R
eal S
cienc
e
The Experiment
This question has been addressed experimentally in a sim-ple and direct way in the laboratory of Richard Cyr atPennsylvania State University. Rigid plant cells conductmechanical force well from one cell to another, and CarolWymer (then a graduate student in the Cyr lab) suspectedsome sort of mechanical force is the signal guiding corticalmicrotubule alignment
Wymer set out to test this hypothesis using centrifuga-tion. If cortical microtubules are obtaining their positionalinformation from an applied force, then their alignmentshould be affected by centrifugal force, and should be im-possible if the integrity of the cell wall (which is supposedlytransmitting the mechanical force) is perturbed with chem-icals that prevent cell wall formation.
Wymer, along with others in the Cyr lab, started out withcells that were not elongated. She isolated protoplasts (cellswithout walls) from the tobacco plant, Nicotiana tabacum, byexposing the plant cells to enzymes that break down the cellwall, creating a spherical plant cell. If allowed to grow in cul-ture, these protoplasts will eventually re-form their cell walls.
In order to examine the effects of directional force on theelongation patterns of plant cells, Wymer and co-workersexposed the tobacco protoplasts to a directional force gener-ated by a centrifuge. Prior experiments had determinedthat centrifugation at the low speeds used in these experi-ments does not disrupt the integrity or shape of the proto-plasts. The protoplasts were immobilized for centrifugationby embedding them in an agar medium supported in amold. The embedded protoplasts were spun in a centrifugeat 450 rpm for 15 minutes. Following centrifugation, theembedded cells were cultured for 72 hours, allowing for cellelongation to occur.
Following centrifugation, fluorescently tagged micro-tubule antibody was applied to the protoplasts, which werethen examined with immunofluorescence microscopy formicrotubule orientation.
To confirm the involvement of microtubules as sensorsof directional force in cell elongation, some protoplastswere incubated prior to centrifugation with a chemical her-bicide, APM, which disrupts microtubules.
The Results
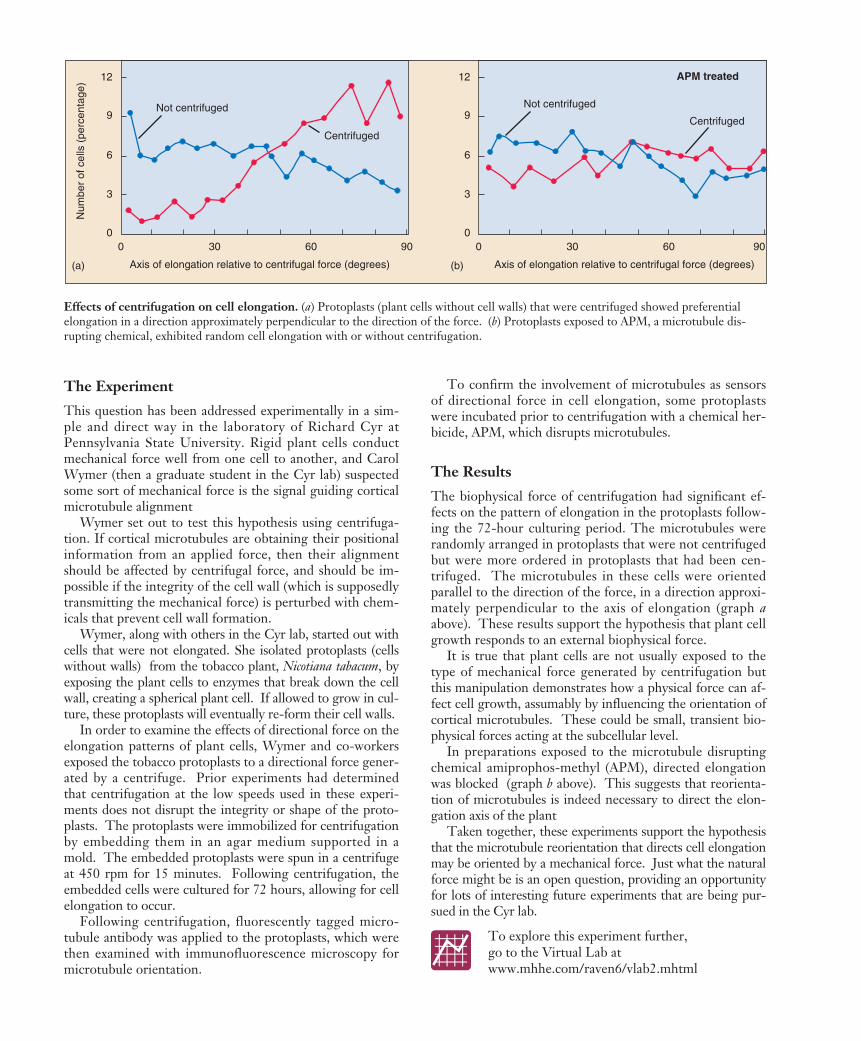
The biophysical force of centrifugation had significant ef-fects on the pattern of elongation in the protoplasts follow-ing the 72-hour culturing period. The microtubules wererandomly arranged in protoplasts that were not centrifugedbut were more ordered in protoplasts that had been cen-trifuged. The microtubules in these cells were orientedparallel to the direction of the force, in a direction approxi-mately perpendicular to the axis of elongation (graph aabove). These results support the hypothesis that plant cellgrowth responds to an external biophysical force.
It is true that plant cells are not usually exposed to thetype of mechanical force generated by centrifugation butthis manipulation demonstrates how a physical force can af-fect cell growth, assumably by influencing the orientation ofcortical microtubules. These could be small, transient bio-physical forces acting at the subcellular level.
In preparations exposed to the microtubule disruptingchemical amiprophos-methyl (APM), directed elongationwas blocked (graph b above). This suggests that reorienta-tion of microtubules is indeed necessary to direct the elon-gation axis of the plant
Taken together, these experiments support the hypothesisthat the microtubule reorientation that directs cell elongationmay be oriented by a mechanical force. Just what the naturalforce might be is an open question, providing an opportunityfor lots of interesting future experiments that are being pur-sued in the Cyr lab.
Axis of elongation relative to centrifugal force (degrees)
6
9
Num
ber
of c
ells
(pe
rcen
tage
) 12
0 30 60 90
3
0
Axis of elongation relative to centrifugal force (degrees)
6
9
12
0 30 60 90
3
0
Not centrifuged
CentrifugedNot centrifuged
Centrifuged
(b)(a)
APM treated
Effects of centrifugation on cell elongation. (a) Protoplasts (plant cells without cell walls) that were centrifuged showed preferentialelongation in a direction approximately perpendicular to the direction of the force. (b) Protoplasts exposed to APM, a microtubule dis-rupting chemical, exhibited random cell elongation with or without centrifugation.
To explore this experiment further, go to the Virtual Lab atwww.mhhe.com/raven6/vlab2.mhtml
77
5Cell Structure
Concept Outline
5.1 All organisms are composed of cells.
Cells. A cell is a membrane-bounded unit that containsDNA and cytoplasm. All organisms are cells or aggregates ofcells, descendants of the first cells.Cells Are Small. The greater relative surface area of smallcells enables more rapid communication between the cellinterior and the environment.
5.2 Eukaryotic cells are far more complex thanbacterial cells.
Bacteria Are Simple Cells. Bacterial cells are small andlack membrane-bounded organelles.Eukaryotic Cells Have Complex Interiors. Eukaryoticcells are compartmentalized by membranes.
5.3 Take a tour of a eukaryotic cell.
The Nucleus: Information Center for the Cell. Thenucleus of a eukaryotic cell isolates the cell’s DNA.The Endoplasmic Reticulum: Compartmentalizing theCell. An extensive system of membranes subdivides the cellinterior.The Golgi Apparatus: Delivery System of the Cell. Asystem of membrane channels collects, modifies, packages,and distributes molecules within the cell.Vesicles: Enzyme Storehouses. Sacs that contain enzymesdigest or modify particles in the cell, while other vesiclestransport substances in and out of cells.Ribosomes: Sites of Protein Synthesis. An RNA-proteincomplex directs the production of proteins.Organelles That Contain DNA. Some organelles withvery different functions contain their own DNA.The Cytoskeleton: Interior Framework of the Cell. Anetwork of protein fibers supports the shape of the cell andanchors organelles.Cell Movement. Eukaryotic cell movement utilizescytoskeletal elements.Special Things about Plant Cells. Plant cells have a largecentral vacuole and strong, multilayered cell walls.
5.4 Symbiosis played a key role in the origin of someeukaryotic organelles.
Endosymbiosis. Mitochondria and chloroplasts may havearisen from prokaryotes engulfed by other prokaryotes.
All organisms are composed of cells. The gossamerwing of a butterfly is a thin sheet of cells, and so is the
glistening outer layer of your eyes. The hamburger ortomato you eat is composed of cells, and its contents soonbecome part of your cells. Some organisms consist of a sin-gle cell too small to see with the unaided eye (figure 5.1),while others, like us, are composed of many cells. Cells areso much a part of life as we know it that we cannot imaginean organism that is not cellular in nature. In this chapterwe will take a close look at the internal structure of cells. Inthe following chapters, we will focus on cells in action—onhow they communicate with their environment, grow, andreproduce.
FIGURE. 5.1The single-celled organism Dileptus. The hairlike projectionsthat cover its surface are cilia, which it undulates to propel itselfthrough the water (1000×).
78 Part II Biology of the Cell
5.1 All organisms are composed of cells.
2 X 10-4 mm2 X 10-2 mm
2 X 101 mm
2 X 100 mm
2 X 10-1 mm
2 X 10-3 mm
CellsWhat is a typical cell like, and what would we find inside it?The general plan of cellular organization varies in the cellsof different organisms, but despite these modifications, allcells resemble each other in certain fundamental ways. Be-fore we begin our detailed examination of cell structure,let’s first summarize three major features all cells have incommon: a plasma membrane, a nucleoid or nucleus, andcytoplasm.
The Plasma Membrane Surrounds the Cell
The plasma membrane encloses a cell and separates itscontents from its surroundings. The plasma membrane is aphospholipid bilayer about 5 to 10 nanometers (5 to 10 bil-lionths of a meter) thick with proteins embedded in it.Viewed in cross-section with the electron microscope, suchmembranes appear as two dark lines separated by a lighterarea. This distinctive appearance arises from the tail-to-tailpacking of the phospholipid molecules that make up themembrane (see figure 3.18). The proteins of a membranemay have large hydrophobic domains, which associate withand become embedded in the phospholipid bilayer.
The proteins of the plasma membrane are in largepart responsible for a cell’s ability to interact with its en-vironment. Transport proteins help molecules and ionsmove across the plasma membrane, either from the envi-ronment to the interior of the cell or vice versa. Receptorproteins induce changes within the cell when they comein contact with specific molecules in the environment,such as hormones. Markers identify the cell as a particu-lar type. This is especially important in multicellular
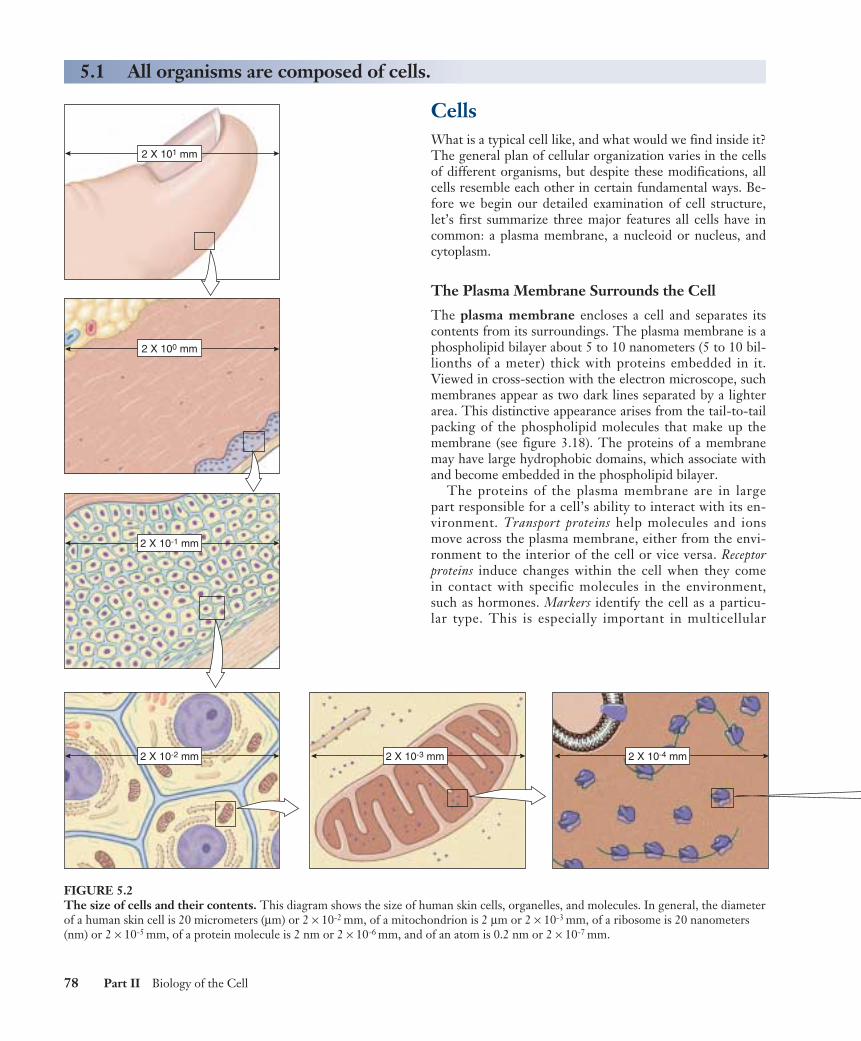
FIGURE 5.2The size of cells and their contents. This diagram shows the size of human skin cells, organelles, and molecules. In general, the diameterof a human skin cell is 20 micrometers (µm) or 2 × 10-2 mm, of a mitochondrion is 2 µm or 2 × 10-3 mm, of a ribosome is 20 nanometers(nm) or 2 × 10-5 mm, of a protein molecule is 2 nm or 2 × 10-6 mm, and of an atom is 0.2 nm or 2 × 10-7 mm.

Chapter 5 Cell Structure 79
2 X 10-7 mm2 X 10-5 mm 2 X 10-6 mm
organisms, whose cells must be able to recognize eachother as they form tissues.
We’ll examine the structure and function of cell mem-branes more thoroughly in chapter 6.
The Central Portion of the Cell Contains the Genetic Material
Every cell contains DNA, the hereditary molecule. Inprokaryotes (bacteria), most of the genetic material lies ina single circular molecule of DNA. It typically resides nearthe center of the cell in an area called the nucleoid, butthis area is not segregated from the rest of the cell’s interiorby membranes. By contrast, the DNA of eukaryotes iscontained in the nucleus, which is surrounded by twomembranes. In both types of organisms, the DNA containsthe genes that code for the proteins synthesized by the cell.
The Cytoplasm Comprises the Rest of the Cell’sInterior
A semifluid matrix called the cytoplasm fills the interior ofthe cell, exclusive of the nucleus (nucleoid in prokaryotes)lying within it. The cytoplasm contains the chemical wealthof the cell: the sugars, amino acids, and proteins the cell usesto carry out its everyday activities. In eukaryotic cells, thecytoplasm also contains specialized membrane-boundedcompartments called organelles.
The Cell Theory
A general characteristic of cells is their microscopic size.While there are a few exceptions—the marine alga Acetabu-laria can be up to 5 centimeters long—a typical eukaryoticcell is 10 to 100 micrometers (10 to 100 millionths of ameter) in diameter (figure 5.2); most bacterial cells are only1 to 10 micrometers in diameter.
Because cells are so small, no one observed them untilmicroscopes were invented in the mid-seventeenth century.Robert Hooke first described cells in 1665, when he used amicroscope he had built to examine a thin slice of cork, anonliving tissue found in the bark of certain trees. Hookeobserved a honeycomb of tiny, empty (because the cellswere dead) compartments. He called the compartments inthe cork cellulae (Latin, “small rooms”), and the term hascome down to us as cells. The first living cells were observeda few years later by the Dutch naturalist Antonie vanLeeuwenhoek, who called the tiny organisms that he ob-served “animalcules,” meaning little animals. For anothercentury and a half, however, biologists failed to recognizethe importance of cells. In 1838, botanist Matthias Schlei-den made a careful study of plant tissues and developed thefirst statement of the cell theory. He stated that all plants“are aggregates of fully individualized, independent, sepa-rate beings, namely the cells themselves.” In 1839,Theodor Schwann reported that all animal tissues also con-sist of individual cells.
The cell theory, in its modern form, includes the fol-lowing three principles:
1. All organisms are composed of one or more cells, andthe life processes of metabolism and heredity occurwithin these cells.
2. Cells are the smallest living things, the basic units oforganization of all organisms.
3. Cells arise only by division of a previously existingcell. Although life likely evolved spontaneously in theenvironment of the early earth, biologists have con-cluded that no additional cells are originating sponta-neously at present. Rather, life on earth represents acontinuous line of descent from those early cells.
A cell is a membrane-bounded unit that contains theDNA hereditary machinery and cytoplasm. Allorganisms are cells or aggregates of cells.
Cells Are SmallHow many cells are big enough to seewith the unaided eye? Other than eggcells, not many. Most are less than 50micrometers in diameter, far smallerthan the period at the end of this sen-tence.
The Resolution Problem
How do we study cells if they are toosmall to see? The key is to understandwhy we can’t see them. The reason wecan’t see such small objects is thelimited resolution of the human eye.Resolution is defined as the minimumdistance two points can be apart andstill be distinguished as two separatedpoints. When two objects are closertogether than about 100 micrometers,the light reflected from each strikes thesame “detector” cell at the rear of theeye. Only when the objects are fartherthan 100 micrometers apart will thelight from each strike different cells,allowing your eye to resolve them astwo objects rather than one.
Microscopes
One way to increase resolution is to increase magnification,so that small objects appear larger. Robert Hooke andAntonie van Leeuwenhoek were able to see small cells bymagnifying their size, so that the cells appeared larger thanthe 100-micrometer limit imposed by the human eye.Hooke and van Leeuwenhoek accomplished this feat withmicroscopes that magnified images of cells by bendinglight through a glass lens. The size of the image that fallson the sheet of detector cells lining the back of your eyedepends on how close the object is to your eye—the closerthe object, the bigger the image. Your eye, however, isincapable of focusing comfortably on an object closer thanabout 25 centimeters, because the eye is limited by the sizeand thickness of its lens. Hooke and van Leeuwenhoekassisted the eye by interposing a glass lens between objectand eye. The glass lens adds additional focusing power.Because the glass lens makes the object appear closer, theimage on the back of the eye is bigger than it would bewithout the lens.
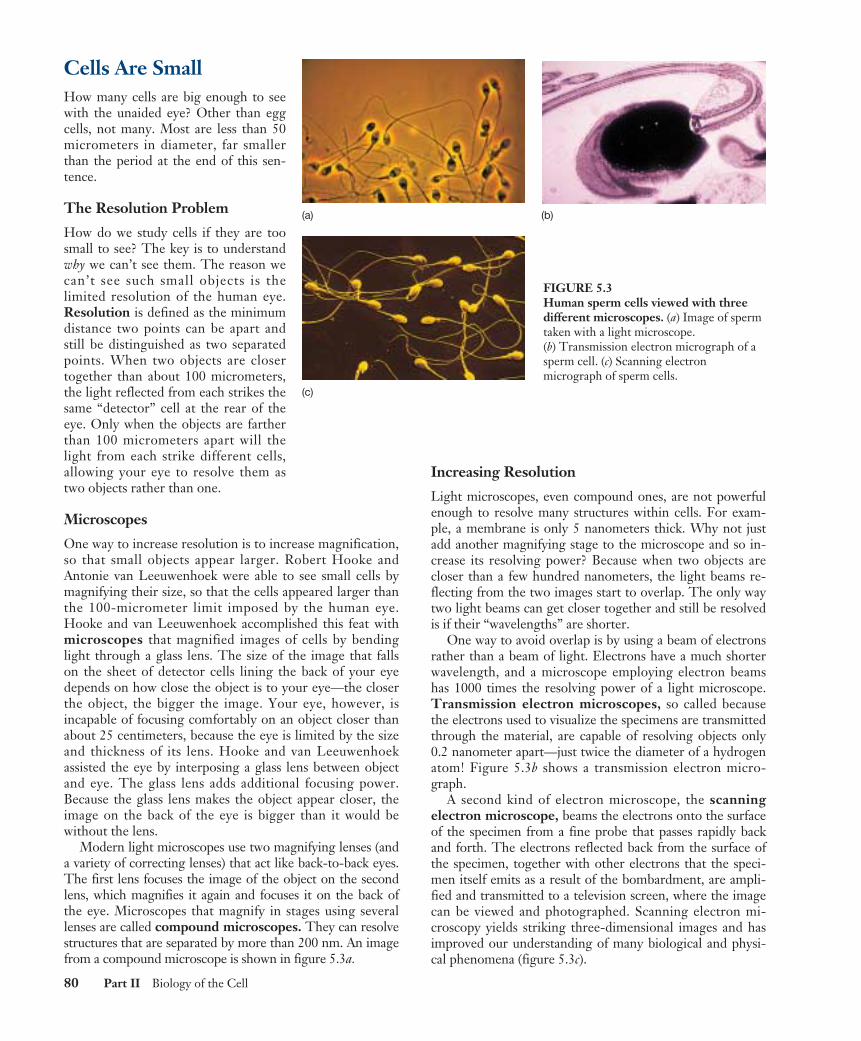
Modern light microscopes use two magnifying lenses (anda variety of correcting lenses) that act like back-to-back eyes.The first lens focuses the image of the object on the secondlens, which magnifies it again and focuses it on the back ofthe eye. Microscopes that magnify in stages using severallenses are called compound microscopes. They can resolvestructures that are separated by more than 200 nm. An imagefrom a compound microscope is shown in figure 5.3a.
Increasing Resolution
Light microscopes, even compound ones, are not powerfulenough to resolve many structures within cells. For exam-ple, a membrane is only 5 nanometers thick. Why not justadd another magnifying stage to the microscope and so in-crease its resolving power? Because when two objects arecloser than a few hundred nanometers, the light beams re-flecting from the two images start to overlap. The only waytwo light beams can get closer together and still be resolvedis if their “wavelengths” are shorter.
One way to avoid overlap is by using a beam of electronsrather than a beam of light. Electrons have a much shorterwavelength, and a microscope employing electron beamshas 1000 times the resolving power of a light microscope.Transmission electron microscopes, so called becausethe electrons used to visualize the specimens are transmittedthrough the material, are capable of resolving objects only0.2 nanometer apart—just twice the diameter of a hydrogenatom! Figure 5.3b shows a transmission electron micro-graph.
A second kind of electron microscope, the scanningelectron microscope, beams the electrons onto the surfaceof the specimen from a fine probe that passes rapidly backand forth. The electrons reflected back from the surface ofthe specimen, together with other electrons that the speci-men itself emits as a result of the bombardment, are ampli-fied and transmitted to a television screen, where the imagecan be viewed and photographed. Scanning electron mi-croscopy yields striking three-dimensional images and hasimproved our understanding of many biological and physi-cal phenomena (figure 5.3c).
80 Part II Biology of the Cell
(a) (b)
(c)
FIGURE 5.3Human sperm cells viewed with threedifferent microscopes. (a) Image of spermtaken with a light microscope.(b) Transmission electron micrograph of asperm cell. (c) Scanning electronmicrograph of sperm cells.

Why Aren’t Cells Larger?
Most cells are not large for practical reasons. The most im-portant of these is communication. The different regions ofa cell need to communicate with one another in order forthe cell as a whole to function effectively. Proteins and or-ganelles are being synthesized, and materials are continuallyentering and leaving the cell. All of these processes involvethe diffusion of substances at some point, and the larger acell is, the longer it takes for substances to diffuse from theplasma membrane to the center of the cell. For this reason,an organism made up of many relatively small cells has anadvantage over one composed of fewer, larger cells.
The advantage of small cell size is readily visualized interms of the surface area-to-volume ratio. As a cell’ssize increases, its volume increases much more rapidlythan its surface area. For a spherical cell, the increase insurface area is equal to the square of the increase in di-ameter, while the increase in volume is equal to the cubeof the increase in diameter. Thus, if two cells differ by afactor of 10 cm in diameter, the larger cell will have 102,or 100 times, the surface area, but 103, or 1000 times,the volume, of the smaller cell (figure 5.4). A cell’s sur-face provides its only opportunity for interaction with
the environment, as all substances enter and exit a cellvia the plasma membrane. This membrane plays a keyrole in controlling cell function, and because small cellshave more surface area per unit of volume than largeones, the control is more effective when cells are rela-tively small.
Although most cells are small, some cells are nonethelessquite large and have apparently overcome the surface area-to-volume problem by one or more adaptive mechanisms.For example, some cells have more than one nucleus, allow-ing genetic information to be spread around a large cell.Also, some large cells actively move material around theircytoplasm so that diffusion is not a limiting factor. Lastly,some large cells, like your own neurons, are long and skinnyso that any given point in the cytoplasm is close to theplasma membrane, and thus diffusion between the insideand outside of the cell can still be rapid.
Multicellular organisms usually consist of many smallcells rather than a few large ones because small cellsfunction more efficiently. They have a greater relativesurface area, enabling more rapid communicationbetween the center of the cell and the environment.
Chapter 5 Cell Structure 81
Cell radius(r)
Surface area(4�r 2)
Volume(4–3�r 3)
1 cm
12.57 cm2
4.189 cm3
10 cm
1257 cm2
4189 cm3
FIGURE 5.4Surface area-to-volume ratio. As a cell gets larger, its volume increases at a faster rate than its surface area. If the cell radius increases by10 times, the surface area increases by 100 times, but the volume increases by 1000 times. A cell’s surface area must be large enough tomeet the needs of its volume.
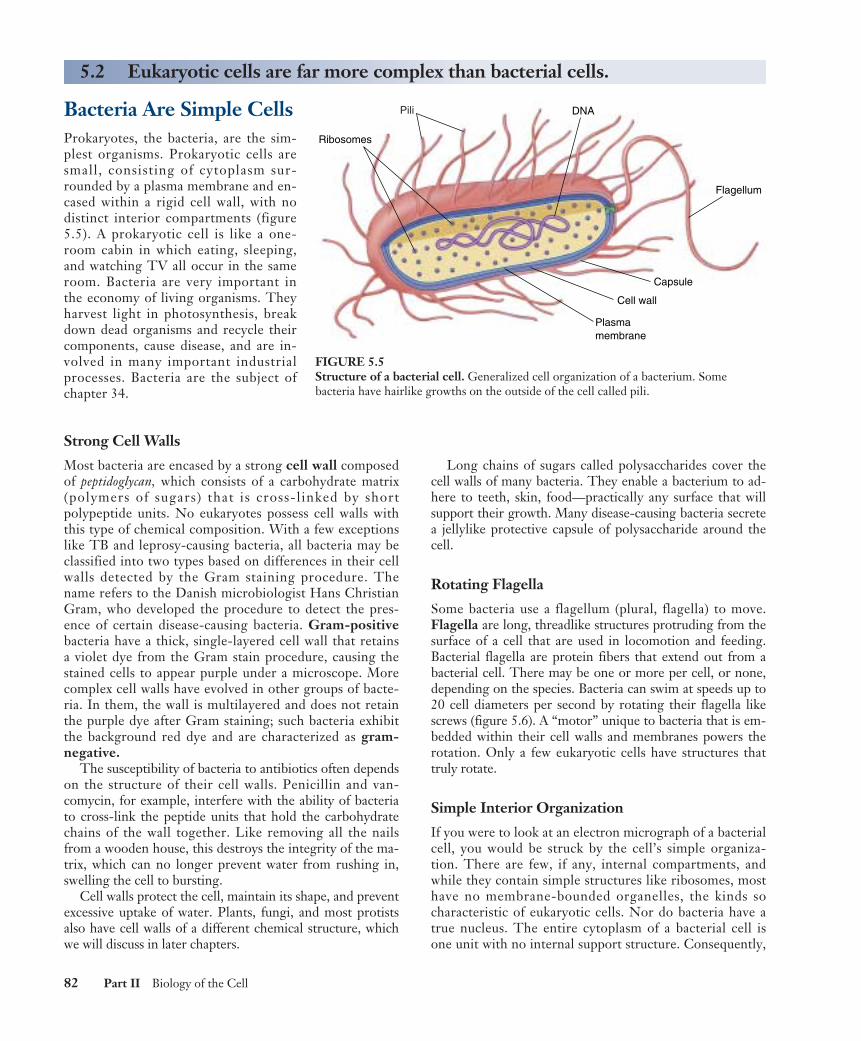
Bacteria Are Simple CellsProkaryotes, the bacteria, are the sim-plest organisms. Prokaryotic cells aresmall, consisting of cytoplasm sur-rounded by a plasma membrane and en-cased within a rigid cell wall, with nodistinct interior compartments (figure5.5). A prokaryotic cell is like a one-room cabin in which eating, sleeping,and watching TV all occur in the sameroom. Bacteria are very important inthe economy of living organisms. Theyharvest light in photosynthesis, breakdown dead organisms and recycle theircomponents, cause disease, and are in-volved in many important industrialprocesses. Bacteria are the subject ofchapter 34.
Strong Cell Walls
Most bacteria are encased by a strong cell wall composedof peptidoglycan, which consists of a carbohydrate matrix(polymers of sugars) that is cross-linked by shortpolypeptide units. No eukaryotes possess cell walls withthis type of chemical composition. With a few exceptionslike TB and leprosy-causing bacteria, all bacteria may beclassified into two types based on differences in their cellwalls detected by the Gram staining procedure. Thename refers to the Danish microbiologist Hans ChristianGram, who developed the procedure to detect the pres-ence of certain disease-causing bacteria. Gram-positivebacteria have a thick, single-layered cell wall that retainsa violet dye from the Gram stain procedure, causing thestained cells to appear purple under a microscope. Morecomplex cell walls have evolved in other groups of bacte-ria. In them, the wall is multilayered and does not retainthe purple dye after Gram staining; such bacteria exhibitthe background red dye and are characterized as gram-negative.
The susceptibility of bacteria to antibiotics often dependson the structure of their cell walls. Penicillin and van-comycin, for example, interfere with the ability of bacteriato cross-link the peptide units that hold the carbohydratechains of the wall together. Like removing all the nailsfrom a wooden house, this destroys the integrity of the ma-trix, which can no longer prevent water from rushing in,swelling the cell to bursting.
Cell walls protect the cell, maintain its shape, and preventexcessive uptake of water. Plants, fungi, and most protistsalso have cell walls of a different chemical structure, whichwe will discuss in later chapters.
Long chains of sugars called polysaccharides cover thecell walls of many bacteria. They enable a bacterium to ad-here to teeth, skin, food—practically any surface that willsupport their growth. Many disease-causing bacteria secretea jellylike protective capsule of polysaccharide around thecell.
Rotating Flagella
Some bacteria use a flagellum (plural, flagella) to move.Flagella are long, threadlike structures protruding from thesurface of a cell that are used in locomotion and feeding.Bacterial flagella are protein fibers that extend out from abacterial cell. There may be one or more per cell, or none,depending on the species. Bacteria can swim at speeds up to20 cell diameters per second by rotating their flagella likescrews (figure 5.6). A “motor” unique to bacteria that is em-bedded within their cell walls and membranes powers therotation. Only a few eukaryotic cells have structures thattruly rotate.
Simple Interior Organization
If you were to look at an electron micrograph of a bacterialcell, you would be struck by the cell’s simple organiza-tion. There are few, if any, internal compartments, andwhile they contain simple structures like ribosomes, mosthave no membrane-bounded organelles, the kinds socharacteristic of eukaryotic cells. Nor do bacteria have atrue nucleus. The entire cytoplasm of a bacterial cell isone unit with no internal support structure. Consequently,
82 Part II Biology of the Cell
5.2 Eukaryotic cells are far more complex than bacterial cells.
Flagellum
Cell wall
Plasmamembrane
Capsule
Ribosomes
DNAPili
FIGURE 5.5Structure of a bacterial cell. Generalized cell organization of a bacterium. Somebacteria have hairlike growths on the outside of the cell called pili.

the strength of the cell comes primarily from its rigid wall(see figure 5.5).
The plasma membrane of a bacterial cell carries out someof the functions organelles perform in eukaryotic cells. Whena bacterial cell divides, for example, the bacterial chromosome,a simple circle of DNA, replicates before the cell divides. Thetwo DNA molecules that result from the replication attach tothe plasma membrane at different points, ensuring that eachdaughter cell will contain one of the identical units of DNA.Moreover, some photosynthetic bacteria, such as cyanobacte-ria and Prochloron (figure 5.7), have an extensively foldedplasma membrane, with the folds extending into the cell’sinterior. These membrane folds contain the bacterialpigments connected with photosynthesis.
Because a bacterial cell contains no membrane-boundedorganelles, the DNA, enzymes, and other cytoplasmic con-stituents have access to all parts of the cell. Reactions are notcompartmentalized as they are in eukaryotic cells, and thewhole bacterium operates as a single unit.
Bacteria are small cells that lack interior organization.They are encased by an exterior wall composed ofcarbohydrates cross-linked by short polypeptides, andsome are propelled by rotating flagella.
Chapter 5 Cell Structure 83
Bacterial cell wall
Flagellin
Rotarymotor
(b)
Sheath
(a)
(c)
FIGURE 5.6Bacteria swim by rotating their flagella. (a) The photograph is of Vibrio cholerae, the microbe that causes the serious disease cholera. Theunsheathed core visible at the top of the photograph is composed of a single crystal of the protein flagellin. (b) In intact flagella, the core issurrounded by a flexible sheath. Imagine that you are standing inside the Vibrio cell, turning the flagellum like a crank. (c) You wouldcreate a spiral wave that travels down the flagellum, just as if you were turning a wire within a flexible tube. The bacterium creates thiskind of rotary motion when it swims.
FIGURE 5.7Electron micrograph of a photosynthetic bacterial cell.Extensive folded photosynthetic membranes are visible in thisProchloron cell (14,500×). The single, circular DNA molecule islocated in the clear area in the central region of the cell.
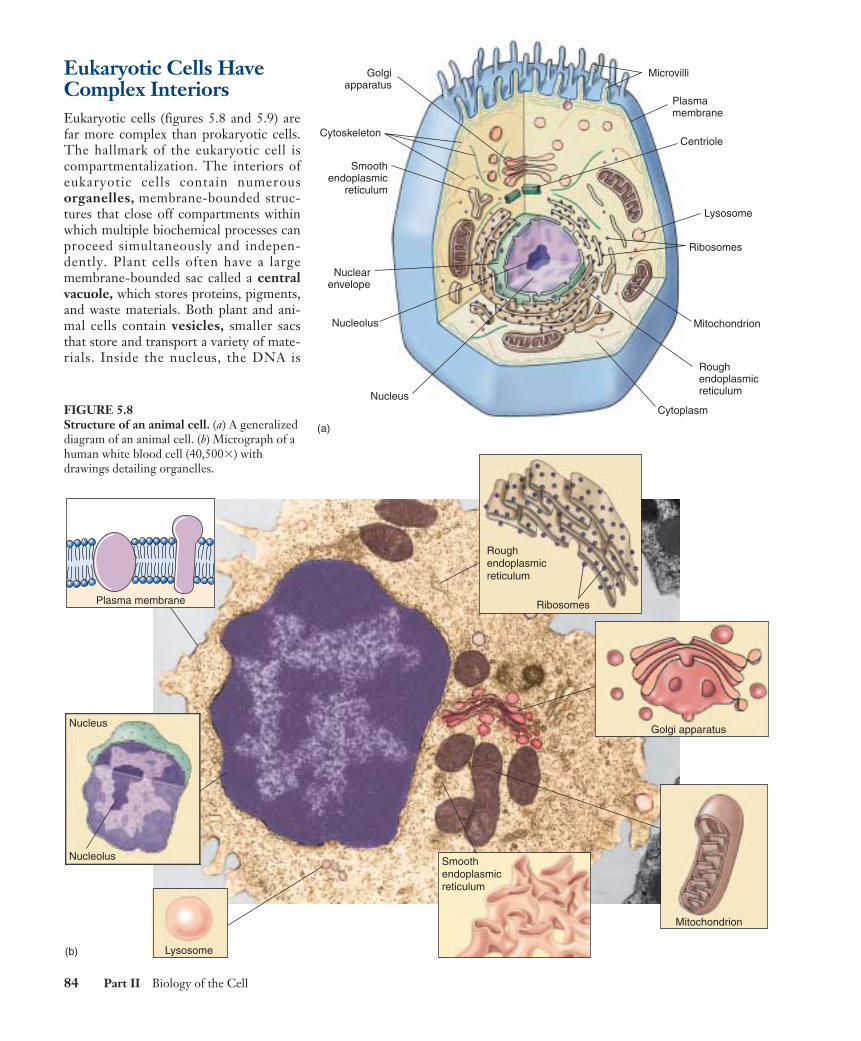
Eukaryotic Cells HaveComplex InteriorsEukaryotic cells (figures 5.8 and 5.9) arefar more complex than prokaryotic cells.The hallmark of the eukaryotic cell iscompartmentalization. The interiors ofeukaryotic cells contain numerousorganelles, membrane-bounded struc-tures that close off compartments withinwhich multiple biochemical processes canproceed simultaneously and indepen-dently. Plant cells often have a largemembrane-bounded sac called a centralvacuole, which stores proteins, pigments,and waste materials. Both plant and ani-mal cells contain vesicles, smaller sacsthat store and transport a variety of mate-rials. Inside the nucleus, the DNA is
84 Part II Biology of the Cell
Centriole
Lysosome
Mitochondrion
Ribosomes
Roughendoplasmicreticulum
CytoplasmNucleus
Nucleolus
Nuclearenvelope
Smoothendoplasmic
reticulum
Cytoskeleton
Golgiapparatus
Plasmamembrane
Microvilli
(a)
Plasma membrane
Nucleolus
Nucleus
Lysosome
Ribosomes
Roughendoplasmicreticulum
Golgi apparatus
Mitochondrion
Smoothendoplasmicreticulum
(b)
FIGURE 5.8Structure of an animal cell. (a) A generalizeddiagram of an animal cell. (b) Micrograph of ahuman white blood cell (40,500�) withdrawings detailing organelles.
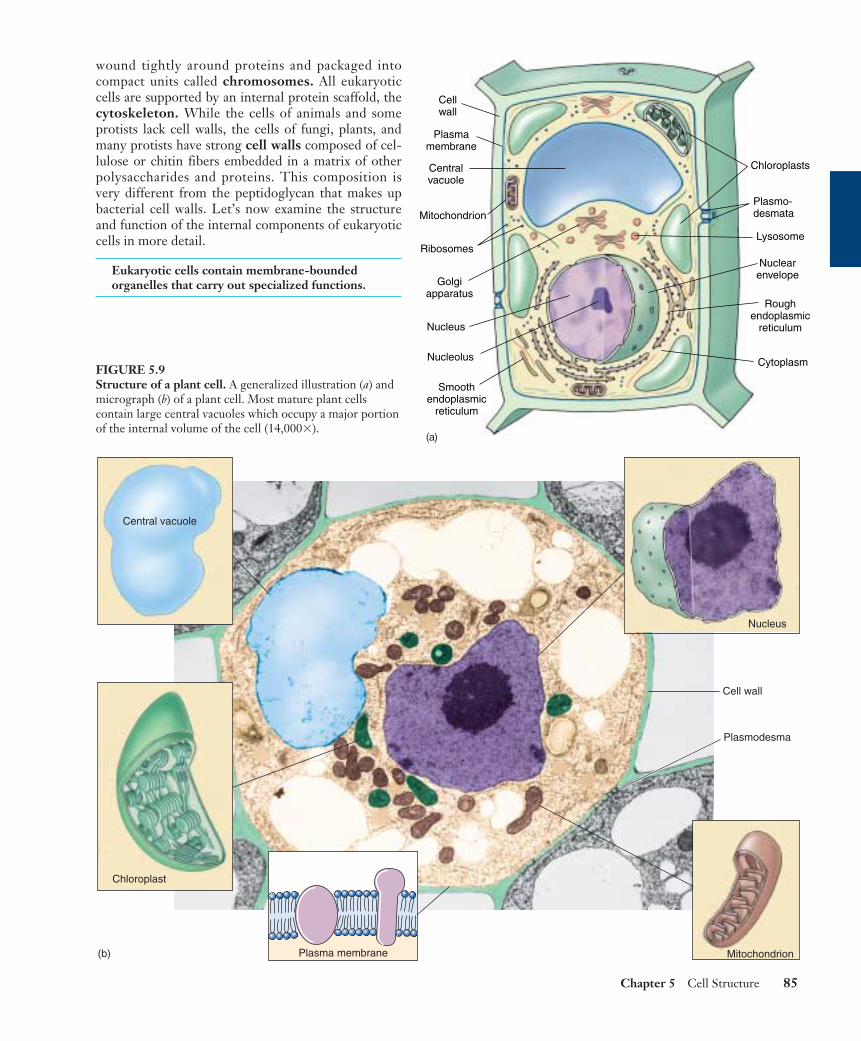
wound tightly around proteins and packaged intocompact units called chromosomes. All eukaryoticcells are supported by an internal protein scaffold, thecytoskeleton. While the cells of animals and someprotists lack cell walls, the cells of fungi, plants, andmany protists have strong cell walls composed of cel-lulose or chitin fibers embedded in a matrix of otherpolysaccharides and proteins. This composition isvery different from the peptidoglycan that makes upbacterial cell walls. Let’s now examine the structureand function of the internal components of eukaryoticcells in more detail.
Eukaryotic cells contain membrane-boundedorganelles that carry out specialized functions.
Chapter 5 Cell Structure 85
Plasma membrane Mitochondrion
Nucleus
Chloroplast
Central vacuole
(b)
Cell wall
Plasmodesma
FIGURE 5.9Structure of a plant cell. A generalized illustration (a) andmicrograph (b) of a plant cell. Most mature plant cellscontain large central vacuoles which occupy a major portionof the internal volume of the cell (14,000�).
Cellwall
Plasmamembrane
Centralvacuole
Mitochondrion
Ribosomes
Golgiapparatus
Nucleus
Nucleolus
Chloroplasts
Nuclearenvelope
Roughendoplasmic
reticulum
Cytoplasm
Smoothendoplasmic
reticulum
Plasmo-desmata
Lysosome
(a)

The Nucleus: Information Centerfor the CellThe largest and most easily seen organelle within a eukary-otic cell is the nucleus (Latin, for kernel or nut), firstdescribed by the English botanist Robert Brown in 1831.Nuclei are roughly spherical in shape and, in animal cells,they are typically located in the central region of the cell(figure 5.10). In some cells, a network of fine cytoplasmic
filaments seems to cradle the nucleus in this position. Thenucleus is the repository of the genetic information thatdirects all of the activities of a living eukaryotic cell. Mosteukaryotic cells possess a single nucleus, although the cellsof fungi and some other groups may have several to manynuclei. Mammalian erythrocytes (red blood cells) lose theirnuclei when they mature. Many nuclei exhibit a dark-staining zone called the nucleolus, which is a region whereintensive synthesis of ribosomal RNA is taking place.
86 Part II Biology of the Cell
5.3 Take a tour of a eukaryotic cell.
(c)
Cytoplasm
Pore
Nucleus
FIGURE 5.10The nucleus. (a) The nucleus is composed of a double membrane, called a nuclear envelope, enclosing a fluid-filled interior containingthe chromosomes. In cross-section, the individual nuclear pores are seen to extend through the two membrane layers of the envelope; thedark material within the pore is protein, which acts to control access through the pore. (b) A freeze-fracture scanning electron micrographof a cell nucleus showing nuclear pores (9500×). (c) A transmission electron micrograph (see figure 6.6) of the nuclear membrane showing anuclear pore.
Nuclearpores
Nuclearpore
Nuclearenvelope
Nucleoplasm Outer membrane
Inner membrane
Nucleolus
(a)
Pore
(b)
The Nuclear Envelope: Getting In and Out
The surface of the nucleus is bounded by two phospho-lipid bilayer membranes, which together make up thenuclear envelope (see figure 5.10). The outer mem-brane of the nuclear envelope is continuous with thecytoplasm’s interior membrane system, called the endo-plasmic reticulum. Scattered over the surface of thenuclear envelope, like craters on the moon, are shallowdepressions called nuclear pores. These pores form 50to 80 nanometers apart at locations where the two mem-brane layers of the nuclear envelope pinch together.Rather than being empty, nuclear pores are filled withproteins that act as molecular channels, permitting certainmolecules to pass into and out of the nucleus. Passage isrestricted primarily to two kinds of molecules: (1) proteinsmoving into the nucleus to be incorporated into nuclearstructures or to catalyze nuclear activities; and (2) RNAand protein-RNA complexes formed in the nucleus andexported to the cytoplasm.
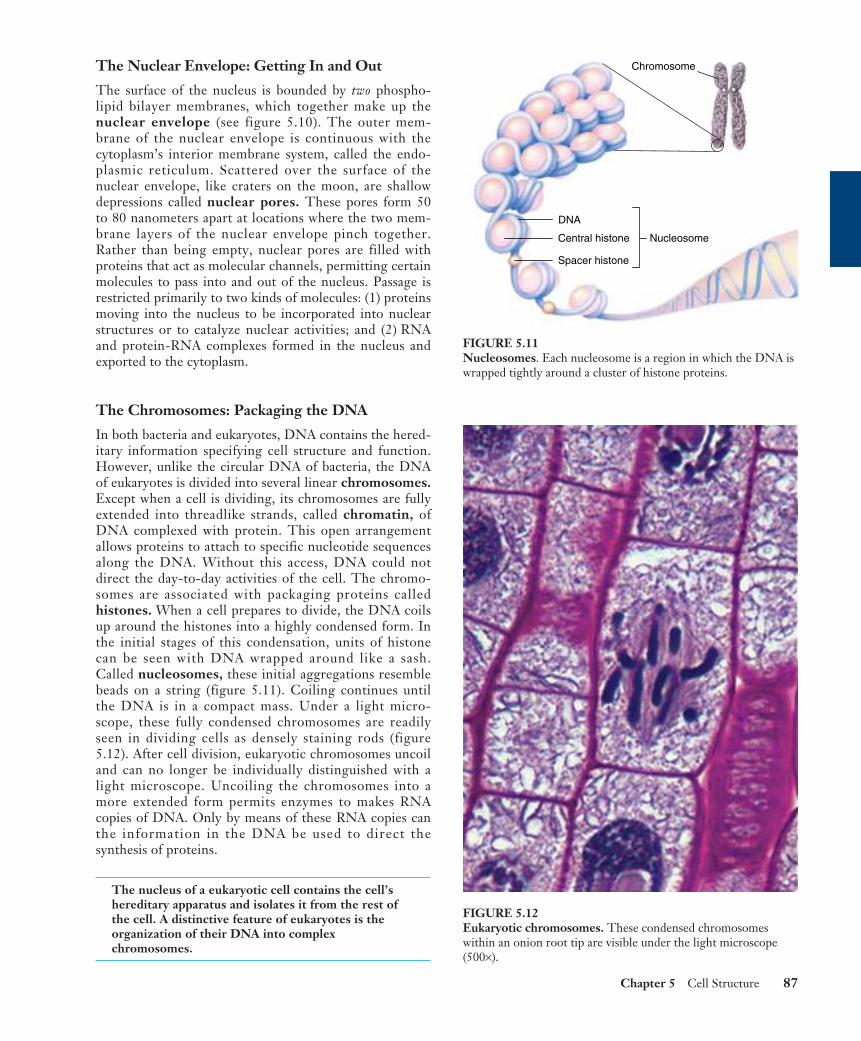
The Chromosomes: Packaging the DNA
In both bacteria and eukaryotes, DNA contains the hered-itary information specifying cell structure and function.However, unlike the circular DNA of bacteria, the DNAof eukaryotes is divided into several linear chromosomes.Except when a cell is dividing, its chromosomes are fullyextended into threadlike strands, called chromatin, ofDNA complexed with protein. This open arrangementallows proteins to attach to specific nucleotide sequencesalong the DNA. Without this access, DNA could notdirect the day-to-day activities of the cell. The chromo-somes are associated with packaging proteins calledhistones. When a cell prepares to divide, the DNA coilsup around the histones into a highly condensed form. Inthe initial stages of this condensation, units of histonecan be seen with DNA wrapped around like a sash.Called nucleosomes, these initial aggregations resemblebeads on a string (figure 5.11). Coiling continues untilthe DNA is in a compact mass. Under a light micro-scope, these fully condensed chromosomes are readilyseen in dividing cells as densely staining rods (figure5.12). After cell division, eukaryotic chromosomes uncoiland can no longer be individually distinguished with alight microscope. Uncoiling the chromosomes into amore extended form permits enzymes to makes RNAcopies of DNA. Only by means of these RNA copies canthe information in the DNA be used to direct thesynthesis of proteins.
The nucleus of a eukaryotic cell contains the cell’shereditary apparatus and isolates it from the rest ofthe cell. A distinctive feature of eukaryotes is theorganization of their DNA into complexchromosomes.
Chapter 5 Cell Structure 87
Central histone
Spacer histone
Nucleosome
Chromosome
DNA
FIGURE 5.11Nucleosomes. Each nucleosome is a region in which the DNA iswrapped tightly around a cluster of histone proteins.
FIGURE 5.12Eukaryotic chromosomes. These condensed chromosomeswithin an onion root tip are visible under the light microscope(500×).

The Endoplasmic Reticulum:Compartmentalizing the CellThe interior of a eukaryotic cell is packed with membranes(table 5.1). So thin that they are invisible under the low re-solving power of light microscopes, this endomembranesystem fills the cell, dividing it into compartments, channel-ing the passage of molecules through the interior of the cell,and providing surfaces for the synthesis of lipids and someproteins. The presence of these membranes in eukaryoticcells constitutes one of the most fundamental distinctionsbetween eukaryotes and prokaryotes.
The largest of the internal membranes is called theendoplasmic reticulum (ER). The term endoplasmic means“within the cytoplasm,” and the term reticulum is Latin for“a little net.” Like the plasma membrane, the ER iscomposed of a lipid bilayer embedded with proteins. Itweaves in sheets through the interior of the cell, creating aseries of channels between its folds (figure 5.13). Of themany compartments in eukaryotic cells, the two largest arethe inner region of the ER, called the cisternal space, andthe region exterior to it, the cytosol.
Rough ER: Manufacturing Proteins for Export
The ER surface regions that are devoted to protein synthe-sis are heavily studded with ribosomes, large molecularaggregates of protein and ribonucleic acid (RNA) that trans-late RNA copies of genes into protein (we will examineribosomes in detail later in this chapter). Through the elec-tron microscope, these ribosome-rich regions of the ERappear pebbly, like the surface of sandpaper, and they aretherefore called rough ER (see figure 5.13).
The proteins synthesized on the surface of the rough ERare destined to be exported from the cell. Proteins to be ex-ported contain special amino acid sequences called signalsequences. As a new protein is made by a free ribosome(one not attached to a membrane), the signal sequence ofthe growing polypeptide attaches to a recognition factorthat carries the ribosome and its partially completed proteinto a “docking site” on the surface of the ER. As the proteinis assembled it passes through the ER membrane into theinterior ER compartment, the cisternal space, from which itis transported by vesicles to the Golgi apparatus (figure5.14). The protein then travels within vesicles to the innersurface of the plasma membrane, where it is released to theoutside of the cell.
88 Part II Biology of the Cell
Table 5.1 Eukaryotic Cell Structures and Their Functions
Structure Description Function
Cell wallCytoskeletonFlagella (cilia)
Plasma membrane
Endoplasmic reticulum
Nucleus
Golgi apparatus
Lysosomes
Microbodies
Mitochondria
Chloroplasts
Chromosomes
NucleolusRibosomes
Outer layer of cellulose or chitin; or absentNetwork of protein filamentsCellular extensions with 9 + 2 arrangement of pairs ofmicrotubulesLipid bilayer with embedded proteins
Network of internal membranes
Structure (usually spherical) surrounded by doublemembrane that contains chromosomesStacks of flattened vesicles
Vesicles derived from Golgi apparatus that containhydrolytic digestive enzymesVesicles formed from incorporation of lipids andproteins containing oxidative and other enzymesBacteria-like elements with double membrane
Bacteria-like elements with membranes containingchlorophyll, a photosynthetic pigmentLong threads of DNA that form a complex withproteinSite of genes for rRNA synthesisSmall, complex assemblies of protein and RNA, oftenbound to endoplasmic reticulum
Protection; supportStructural support; cell movementMotility or moving fluids over surfaces
Regulates what passes into and out of cell;cell-to-cell recognitionForms compartments and vesicles;participates in protein and lipid synthesisControl center of cell; directs proteinsynthesis and cell reproductionPackages proteins for export from cell;forms secretory vesiclesDigest worn-out organelles and cell debris;play role in cell deathIsolate particular chemical activities fromrest of cell“Power plants” of the cell; sites of oxidativemetabolismSites of photosynthesis
Contain hereditary information
Assembles ribosomesSites of protein synthesis
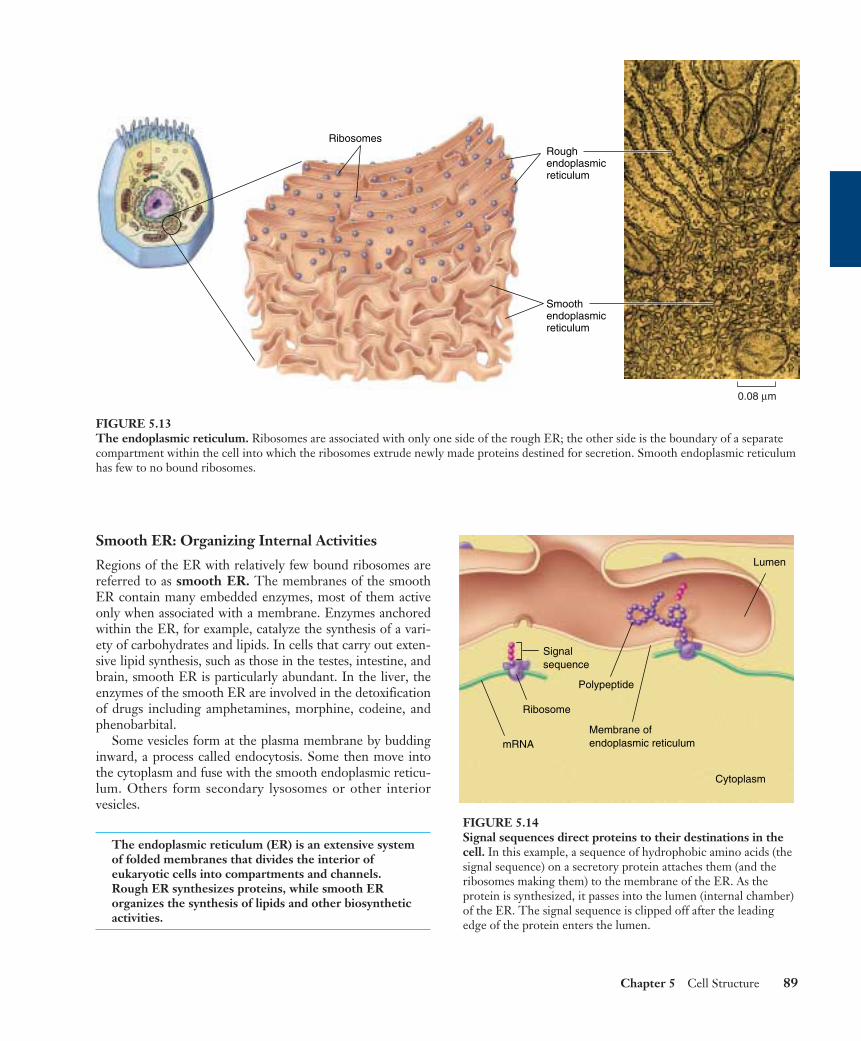
Smooth ER: Organizing Internal Activities
Regions of the ER with relatively few bound ribosomes arereferred to as smooth ER. The membranes of the smoothER contain many embedded enzymes, most of them activeonly when associated with a membrane. Enzymes anchoredwithin the ER, for example, catalyze the synthesis of a vari-ety of carbohydrates and lipids. In cells that carry out exten-sive lipid synthesis, such as those in the testes, intestine, andbrain, smooth ER is particularly abundant. In the liver, theenzymes of the smooth ER are involved in the detoxificationof drugs including amphetamines, morphine, codeine, andphenobarbital.
Some vesicles form at the plasma membrane by buddinginward, a process called endocytosis. Some then move intothe cytoplasm and fuse with the smooth endoplasmic reticu-lum. Others form secondary lysosomes or other interiorvesicles.
The endoplasmic reticulum (ER) is an extensive systemof folded membranes that divides the interior ofeukaryotic cells into compartments and channels.Rough ER synthesizes proteins, while smooth ERorganizes the synthesis of lipids and other biosyntheticactivities.
Chapter 5 Cell Structure 89
0.08 µm
RibosomesRough endoplasmic reticulum
Smooth endoplasmic reticulum
FIGURE 5.13The endoplasmic reticulum. Ribosomes are associated with only one side of the rough ER; the other side is the boundary of a separatecompartment within the cell into which the ribosomes extrude newly made proteins destined for secretion. Smooth endoplasmic reticulumhas few to no bound ribosomes.
Cytoplasm
Lumen
mRNA
Ribosome
Membrane ofendoplasmic reticulum
Polypeptide
Signalsequence
FIGURE 5.14Signal sequences direct proteins to their destinations in thecell. In this example, a sequence of hydrophobic amino acids (thesignal sequence) on a secretory protein attaches them (and theribosomes making them) to the membrane of the ER. As theprotein is synthesized, it passes into the lumen (internal chamber)of the ER. The signal sequence is clipped off after the leadingedge of the protein enters the lumen.

The Golgi Apparatus: DeliverySystem of the CellAt various locations within the endomembrane system,flattened stacks of membranes called Golgi bodies occur,often interconnected with one another. These structuresare named for Camillo Golgi, the nineteenth-centuryItalian physician who first called attention to them. Thenumbers of Golgi bodies a cell contains ranges from 1 ora few in protists, to 20 or more in animal cells and severalhundred in plant cells. They are especially abundant inglandular cells, which manufacture and secrete sub-stances. Collectively the Golgi bodies are referred to asthe Golgi apparatus (figure 5.15).
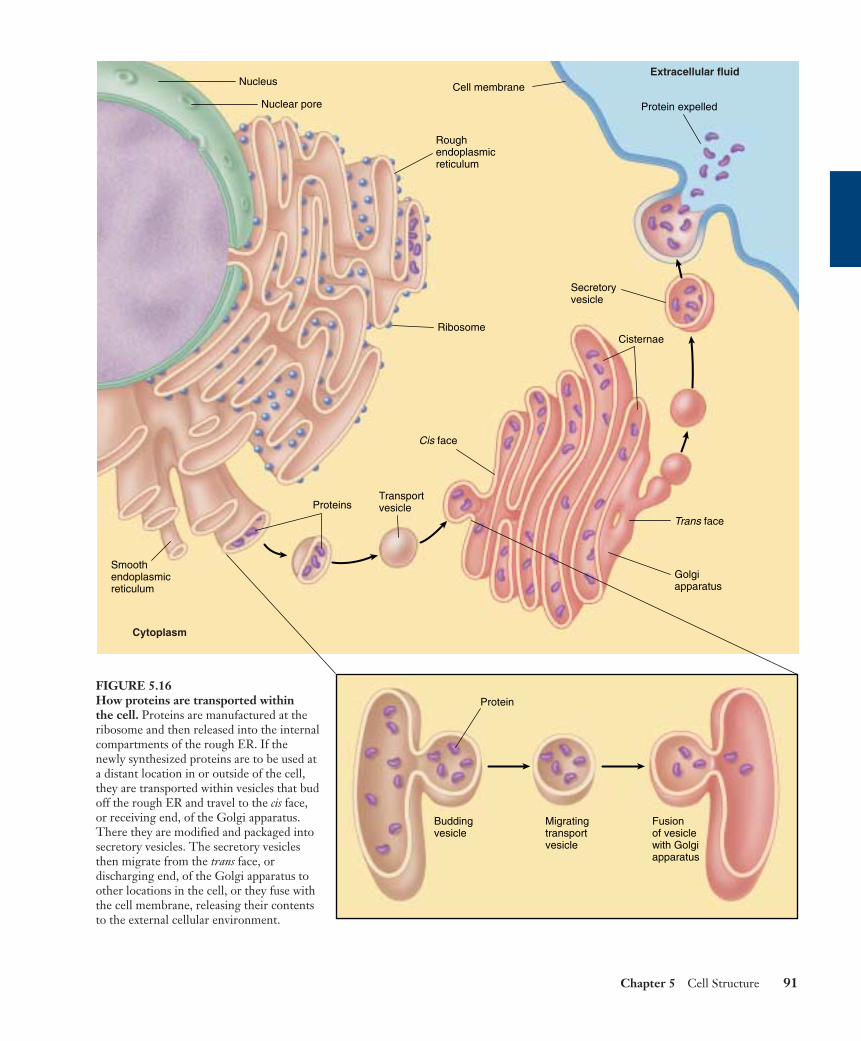
The Golgi apparatus functions in the collection, packag-ing, and distribution of molecules synthesized at one placein the cell and utilized at another location in the cell. AGolgi body has a front and a back, with distinctly differentmembrane compositions at the opposite ends. The front, orreceiving end, is called the cis face, and is usually locatednear ER. Materials move to the cis face in transport vesiclesthat bud off of the ER. These vesicles fuse with the cis face,emptying their contents into the interior, or lumen, of theGolgi apparatus. These ER-synthesized molecules then passthrough the channels of the Golgi apparatus until theyreach the back, or discharging end, called the trans face,where they are discharged in secretory vesicles (figure 5.16).
Proteins and lipids manufactured on the rough andsmooth ER membranes are transported into the Golgi ap-
paratus and modified as they pass through it. The mostcommon alteration is the addition or modification of shortsugar chains, forming a glycoprotein when sugars are com-plexed to a protein and a glycolipid when sugars are boundto a lipid. In many instances, enzymes in the Golgi appara-tus modify existing glycoproteins and glycolipids made inthe ER by cleaving a sugar from their sugar chain or modi-fying one or more of the sugars.
The newly formed or altered glycoproteins and glycol-ipids collect at the ends of the Golgi bodies, in flattenedstacked membrane folds called cisternae (Latin,“collecting vessels”). Periodically, the membranes of thecisternae push together, pinching off small, membrane-bounded secretory vesicles containing the glycoproteinand glycolipid molecules. These vesicles then move toother locations in the cell, distributing the newlysynthesized molecules to their appropriate destinations.Liposomes are synthetically manufactured vesicles thatcontain any variety of desirable substances (such asdrugs), and can be injected into the body. Because themembrane of liposomes is similar to plasma and organellarmembranes, these liposomes serve as an effective andnatural delivery system to cells and may prove to be ofgreat therapeutic value.
The Golgi apparatus is the delivery system of theeukaryotic cell. It collects, packages, modifies, anddistributes molecules that are synthesized at onelocation within the cell and used at another.
90 Part II Biology of the Cell
Secretory vesicles
Vesicle 0.57 µm
FIGURE 5.15The Golgi apparatus. The Golgi apparatus is a smooth, concave membranous structure located near the middle of the cell. It receivesmaterial for processing on one surface and sends the material packaged in vesicles off the other. The substance in a vesicle could be forexport out of the cell or for distribution to another region within the same cell.
Chapter 5 Cell Structure 91
Buddingvesicle
Fusionof vesiclewith Golgiapparatus
Migratingtransportvesicle
Protein
ProteinsTransportvesicle
Golgiapparatus
Secretoryvesicle
Smoothendoplasmicreticulum
Roughendoplasmicreticulum
Nuclear pore
Nucleus
CisternaeRibosome
Trans face
Cis face
Cell membrane
Protein expelled
Cytoplasm
Extracellular fluid
FIGURE 5.16How proteins are transported within the cell. Proteins are manufactured at theribosome and then released into the internalcompartments of the rough ER. If thenewly synthesized proteins are to be used ata distant location in or outside of the cell,they are transported within vesicles that budoff the rough ER and travel to the cis face,or receiving end, of the Golgi apparatus.There they are modified and packaged intosecretory vesicles. The secretory vesiclesthen migrate from the trans face, ordischarging end, of the Golgi apparatus toother locations in the cell, or they fuse withthe cell membrane, releasing their contentsto the external cellular environment.

Vesicles: Enzyme StorehousesLysosomes: Intracellular Digestion Centers
Lysosomes, membrane-bounded diges-tive vesicles, are also components of theendomembrane system that arise fromthe Golgi apparatus. They contain highlevels of degrading enzymes, which cat-alyze the rapid breakdown of proteins,nucleic acids, lipids, and carbohydrates.Throughout the lives of eukaryotic cells,lysosomal enzymes break down old or-ganelles, recycling their component mol-ecules and making room for newlyformed organelles. For example, mito-chondria are replaced in some tissuesevery 10 days.
The digestive enzymes in lysosomesfunction best in an acidic environment.Lysosomes actively engaged in digestionkeep their battery of hydrolytic enzymes(enzymes that catalyze the hydrolysis ofmolecules) fully active by pumping protons into their inte-riors and thereby maintaining a low internal pH. Lyso-somes that are not functioning actively do not maintain anacidic internal pH and are called primary lysosomes. When aprimary lysosome fuses with a food vesicle or other or-ganelle, its pH falls and its arsenal of hydrolytic enzymes isactivated; it is then called a secondary lysosome.
In addition to breaking down organelles and other struc-tures within cells, lysosomes also eliminate other cells thatthe cell has engulfed in a process called phagocytosis, a spe-cific type of endocytosis (see chapter 6). When a whiteblood cell, for example, phagocytizes a passing pathogen,lysosomes fuse with the resulting “food vesicle,” releasingtheir enzymes into the vesicle and degrading the materialwithin (figure 5.17).
Microbodies
Eukaryotic cells contain a variety of enzyme-bearing,membrane-enclosed vesicles called microbodies. Micro-bodies are found in the cells of plants, animals, fungi, andprotists. The distribution of enzymes into microbodies isone of the principal ways in which eukaryotic cells orga-nize their metabolism.
While lysosomes bud from the endomembrane system,microbodies grow by incorporating lipids and protein,then dividing. Plant cells have a special type of microbodycalled a glyoxysome that contains enzymes that convertfats into carbohydrates. Another type of microbody, aperoxisome, contains enzymes that catalyze the removalof electrons and associated hydrogen atoms (figure 5.18).
If these oxidative enzymes were not isolated within micro-bodies, they would tend to short-circuit the metabolism ofthe cytoplasm, which often involves adding hydrogen atomsto oxygen. The name peroxisome refers to the hydrogenperoxide produced as a by-product of the activities of theoxidative enzymes in the microbody. Hydrogen peroxide isdangerous to cells because of its violent chemical reactivity.However, peroxisomes also contain the enzyme catalase,which breaks down hydrogen peroxide into harmlesswater and oxygen.
Lysosomes and peroxisomes are vesicles that containdigestive and detoxifying enzymes. The isolation ofthese enzymes in vesicles protects the rest of the cellfrom inappropriate digestive activity.
92 Part II Biology of the Cell
Cytoplasm
Phagocytosis
Foodvesicle
Golgiapparatus
Lysosomes
Plasmamembrane
Digestion of phagocytizedfood particles
or cells
Endoplasmicreticulum
Transportvesicle
Old or damagedorganelle
Breakdownof old
organelleExtracellular
fluid
FIGURE 5.17Lysosomes. Lysosomes contain hydrolytic enzymes that digest particles or cells takeninto the cell by phagocytosis and break down old organelles.
0.21 µm
FIGURE 5.18A peroxisome.Peroxisomes arespherical organellesthat may contain alarge diamond-shapedcrystal composed ofprotein. Peroxisomescontain digestive anddetoxifying enzymesthat produce hydrogenperoxide as a by-product.
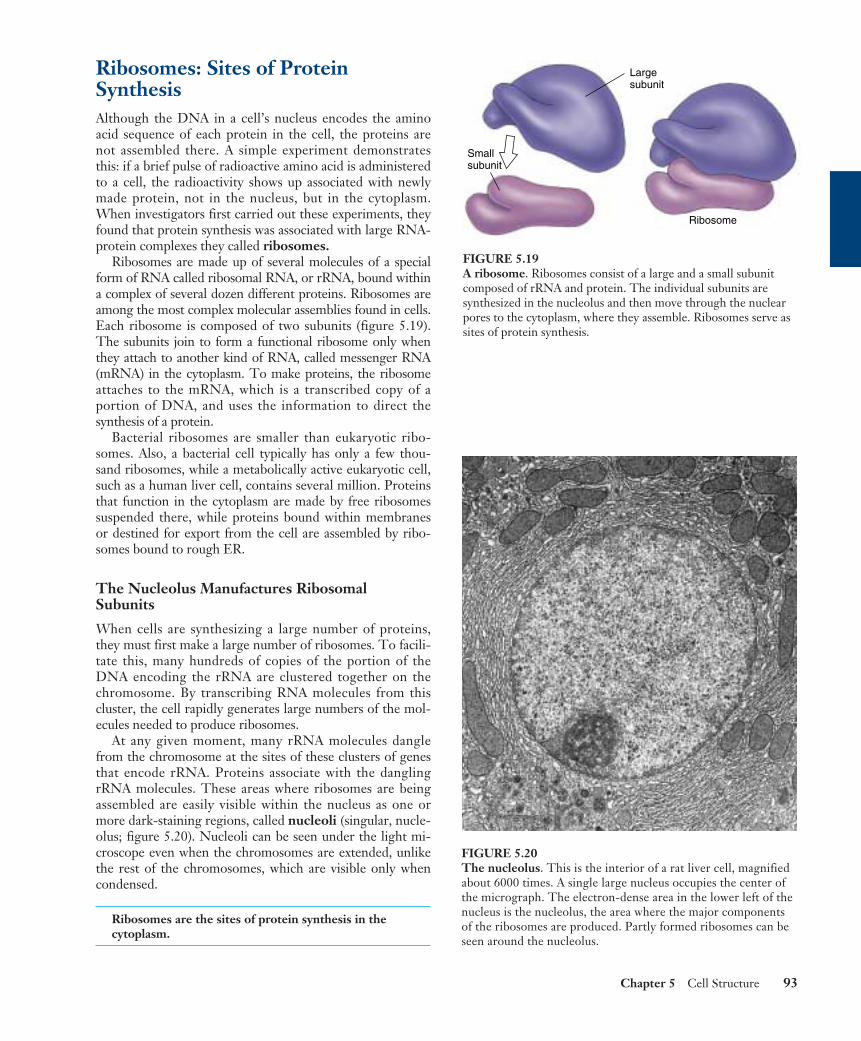
Ribosomes: Sites of ProteinSynthesisAlthough the DNA in a cell’s nucleus encodes the aminoacid sequence of each protein in the cell, the proteins arenot assembled there. A simple experiment demonstratesthis: if a brief pulse of radioactive amino acid is administeredto a cell, the radioactivity shows up associated with newlymade protein, not in the nucleus, but in the cytoplasm.When investigators first carried out these experiments, theyfound that protein synthesis was associated with large RNA-protein complexes they called ribosomes.
Ribosomes are made up of several molecules of a specialform of RNA called ribosomal RNA, or rRNA, bound withina complex of several dozen different proteins. Ribosomes areamong the most complex molecular assemblies found in cells.Each ribosome is composed of two subunits (figure 5.19).The subunits join to form a functional ribosome only whenthey attach to another kind of RNA, called messenger RNA(mRNA) in the cytoplasm. To make proteins, the ribosomeattaches to the mRNA, which is a transcribed copy of aportion of DNA, and uses the information to direct thesynthesis of a protein.
Bacterial ribosomes are smaller than eukaryotic ribo-somes. Also, a bacterial cell typically has only a few thou-sand ribosomes, while a metabolically active eukaryotic cell,such as a human liver cell, contains several million. Proteinsthat function in the cytoplasm are made by free ribosomessuspended there, while proteins bound within membranesor destined for export from the cell are assembled by ribo-somes bound to rough ER.
The Nucleolus Manufactures Ribosomal Subunits
When cells are synthesizing a large number of proteins,they must first make a large number of ribosomes. To facili-tate this, many hundreds of copies of the portion of theDNA encoding the rRNA are clustered together on thechromosome. By transcribing RNA molecules from thiscluster, the cell rapidly generates large numbers of the mol-ecules needed to produce ribosomes.
At any given moment, many rRNA molecules danglefrom the chromosome at the sites of these clusters of genesthat encode rRNA. Proteins associate with the danglingrRNA molecules. These areas where ribosomes are beingassembled are easily visible within the nucleus as one ormore dark-staining regions, called nucleoli (singular, nucle-olus; figure 5.20). Nucleoli can be seen under the light mi-croscope even when the chromosomes are extended, unlikethe rest of the chromosomes, which are visible only whencondensed.
Ribosomes are the sites of protein synthesis in thecytoplasm.
Chapter 5 Cell Structure 93
Smallsubunit
Largesubunit
Ribosome
FIGURE 5.19A ribosome. Ribosomes consist of a large and a small subunitcomposed of rRNA and protein. The individual subunits aresynthesized in the nucleolus and then move through the nuclearpores to the cytoplasm, where they assemble. Ribosomes serve assites of protein synthesis.
FIGURE 5.20The nucleolus. This is the interior of a rat liver cell, magnifiedabout 6000 times. A single large nucleus occupies the center ofthe micrograph. The electron-dense area in the lower left of thenucleus is the nucleolus, the area where the major componentsof the ribosomes are produced. Partly formed ribosomes can beseen around the nucleolus.
Organelles ThatContain DNAAmong the most interesting cellorganelles are those in additionto the nucleus that containDNA.
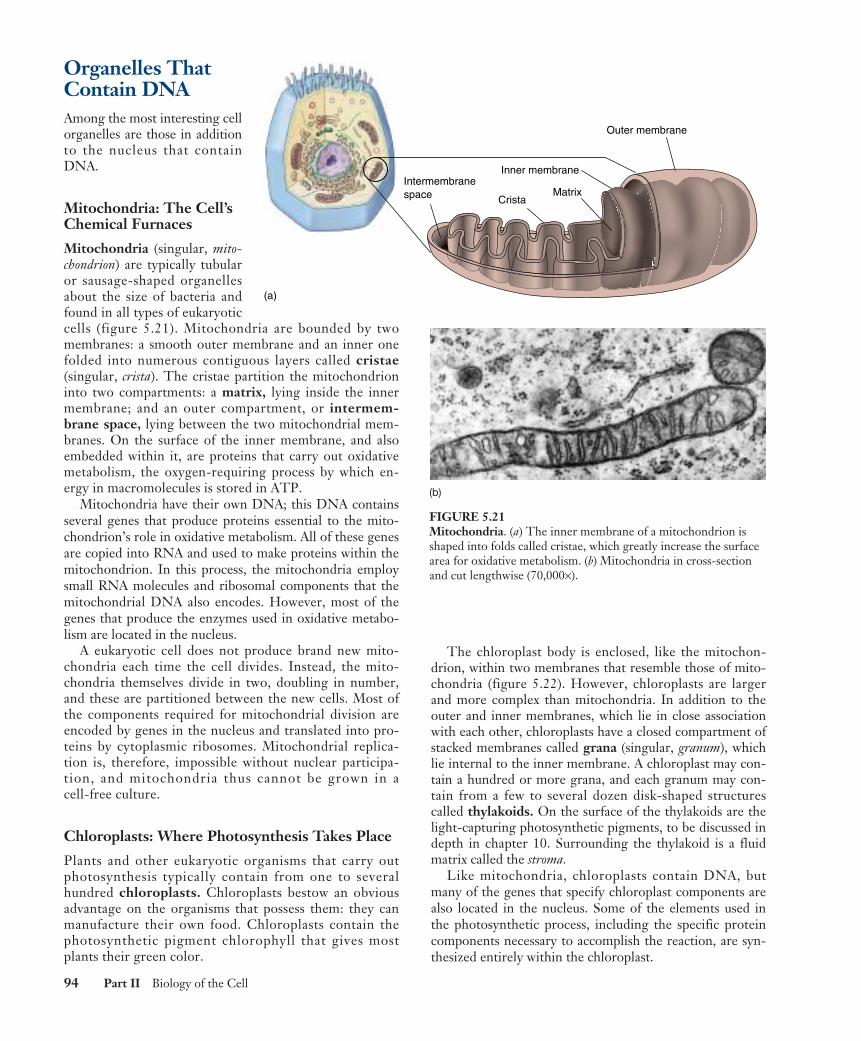
Mitochondria: The Cell’sChemical Furnaces
Mitochondria (singular, mito-chondrion) are typically tubularor sausage-shaped organellesabout the size of bacteria andfound in all types of eukaryoticcells (figure 5.21). Mitochondria are bounded by twomembranes: a smooth outer membrane and an inner onefolded into numerous contiguous layers called cristae(singular, crista). The cristae partition the mitochondrioninto two compartments: a matrix, lying inside the innermembrane; and an outer compartment, or intermem-brane space, lying between the two mitochondrial mem-branes. On the surface of the inner membrane, and alsoembedded within it, are proteins that carry out oxidativemetabolism, the oxygen-requiring process by which en-ergy in macromolecules is stored in ATP.
Mitochondria have their own DNA; this DNA containsseveral genes that produce proteins essential to the mito-chondrion’s role in oxidative metabolism. All of these genesare copied into RNA and used to make proteins within themitochondrion. In this process, the mitochondria employsmall RNA molecules and ribosomal components that themitochondrial DNA also encodes. However, most of thegenes that produce the enzymes used in oxidative metabo-lism are located in the nucleus.
A eukaryotic cell does not produce brand new mito-chondria each time the cell divides. Instead, the mito-chondria themselves divide in two, doubling in number,and these are partitioned between the new cells. Most ofthe components required for mitochondrial division areencoded by genes in the nucleus and translated into pro-teins by cytoplasmic ribosomes. Mitochondrial replica-tion is, therefore, impossible without nuclear participa-tion, and mitochondria thus cannot be grown in acell-free culture.
Chloroplasts: Where Photosynthesis Takes Place
Plants and other eukaryotic organisms that carry outphotosynthesis typically contain from one to severalhundred chloroplasts. Chloroplasts bestow an obviousadvantage on the organisms that possess them: they canmanufacture their own food. Chloroplasts contain thephotosynthetic pigment chlorophyll that gives mostplants their green color.
The chloroplast body is enclosed, like the mitochon-drion, within two membranes that resemble those of mito-chondria (figure 5.22). However, chloroplasts are largerand more complex than mitochondria. In addition to theouter and inner membranes, which lie in close associationwith each other, chloroplasts have a closed compartment ofstacked membranes called grana (singular, granum), whichlie internal to the inner membrane. A chloroplast may con-tain a hundred or more grana, and each granum may con-tain from a few to several dozen disk-shaped structurescalled thylakoids. On the surface of the thylakoids are thelight-capturing photosynthetic pigments, to be discussed indepth in chapter 10. Surrounding the thylakoid is a fluidmatrix called the stroma.
Like mitochondria, chloroplasts contain DNA, butmany of the genes that specify chloroplast components arealso located in the nucleus. Some of the elements used inthe photosynthetic process, including the specific proteincomponents necessary to accomplish the reaction, are syn-thesized entirely within the chloroplast.
94 Part II Biology of the Cell
Intermembranespace
Inner membrane
Outer membrane
MatrixCrista
membranee
Inner membrane
MatrixCrista
(a)
(b)
FIGURE 5.21Mitochondria. (a) The inner membrane of a mitochondrion isshaped into folds called cristae, which greatly increase the surfacearea for oxidative metabolism. (b) Mitochondria in cross-sectionand cut lengthwise (70,000×).
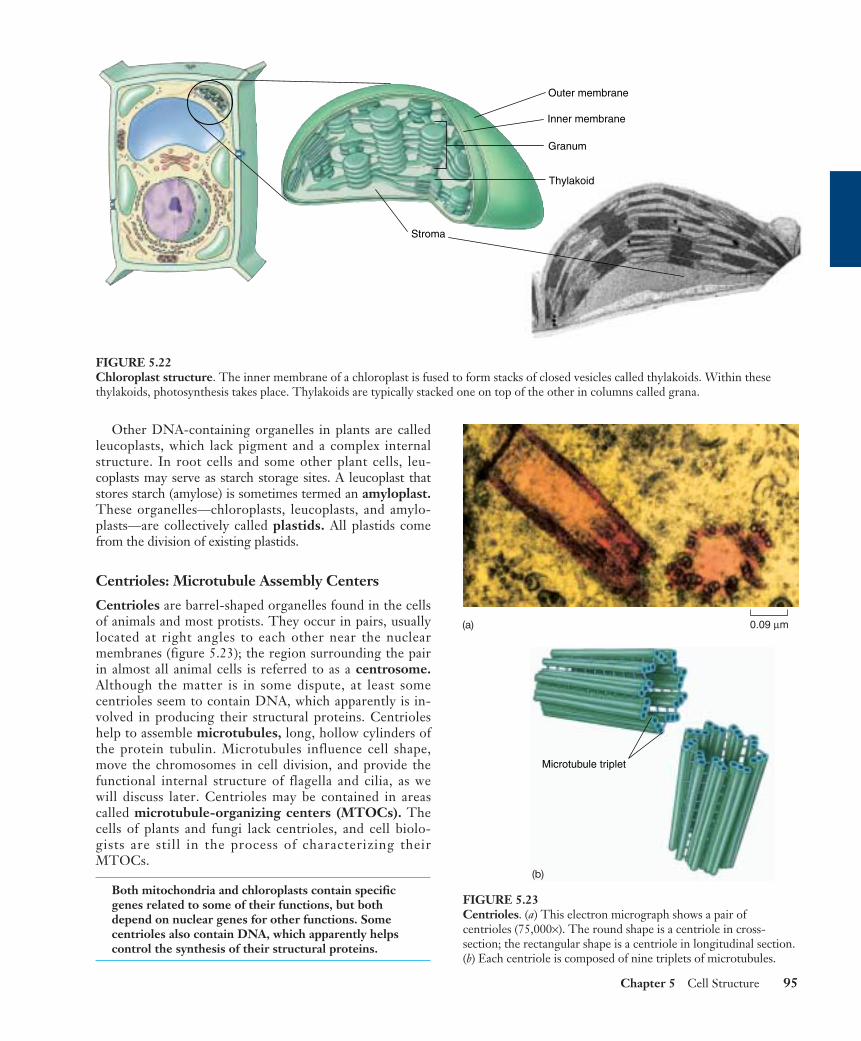
Other DNA-containing organelles in plants are calledleucoplasts, which lack pigment and a complex internalstructure. In root cells and some other plant cells, leu-coplasts may serve as starch storage sites. A leucoplast thatstores starch (amylose) is sometimes termed an amyloplast.These organelles—chloroplasts, leucoplasts, and amylo-plasts—are collectively called plastids. All plastids comefrom the division of existing plastids.
Centrioles: Microtubule Assembly Centers
Centrioles are barrel-shaped organelles found in the cellsof animals and most protists. They occur in pairs, usuallylocated at right angles to each other near the nuclearmembranes (figure 5.23); the region surrounding the pairin almost all animal cells is referred to as a centrosome.Although the matter is in some dispute, at least somecentrioles seem to contain DNA, which apparently is in-volved in producing their structural proteins. Centrioleshelp to assemble microtubules, long, hollow cylinders ofthe protein tubulin. Microtubules influence cell shape,move the chromosomes in cell division, and provide thefunctional internal structure of flagella and cilia, as wewill discuss later. Centrioles may be contained in areascalled microtubule-organizing centers (MTOCs). Thecells of plants and fungi lack centrioles, and cell biolo-gists are still in the process of characterizing theirMTOCs.
Both mitochondria and chloroplasts contain specificgenes related to some of their functions, but bothdepend on nuclear genes for other functions. Somecentrioles also contain DNA, which apparently helpscontrol the synthesis of their structural proteins.
Chapter 5 Cell Structure 95
Outer membrane
Inner membrane
Granum
Thylakoid
Stroma
FIGURE 5.22Chloroplast structure. The inner membrane of a chloroplast is fused to form stacks of closed vesicles called thylakoids. Within thesethylakoids, photosynthesis takes place. Thylakoids are typically stacked one on top of the other in columns called grana.
0.09 µm
Microtubule triplet
FIGURE 5.23Centrioles. (a) This electron micrograph shows a pair ofcentrioles (75,000×). The round shape is a centriole in cross-section; the rectangular shape is a centriole in longitudinal section.(b) Each centriole is composed of nine triplets of microtubules.
(a)
(b)
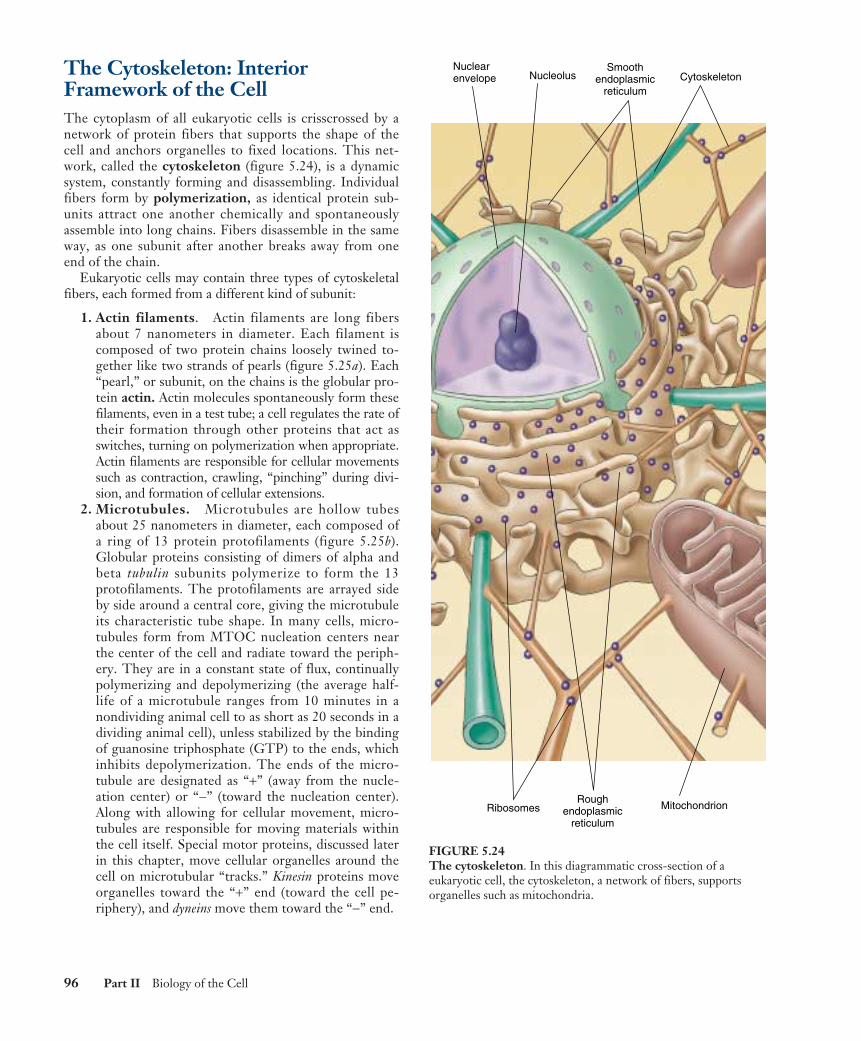
The Cytoskeleton: InteriorFramework of the CellThe cytoplasm of all eukaryotic cells is crisscrossed by anetwork of protein fibers that supports the shape of thecell and anchors organelles to fixed locations. This net-work, called the cytoskeleton (figure 5.24), is a dynamicsystem, constantly forming and disassembling. Individualfibers form by polymerization, as identical protein sub-units attract one another chemically and spontaneouslyassemble into long chains. Fibers disassemble in the sameway, as one subunit after another breaks away from oneend of the chain.
Eukaryotic cells may contain three types of cytoskeletalfibers, each formed from a different kind of subunit:
1. Actin filaments. Actin filaments are long fibersabout 7 nanometers in diameter. Each filament iscomposed of two protein chains loosely twined to-gether like two strands of pearls (figure 5.25a). Each“pearl,” or subunit, on the chains is the globular pro-tein actin. Actin molecules spontaneously form thesefilaments, even in a test tube; a cell regulates the rate oftheir formation through other proteins that act asswitches, turning on polymerization when appropriate.Actin filaments are responsible for cellular movementssuch as contraction, crawling, “pinching” during divi-sion, and formation of cellular extensions.
2. Microtubules. Microtubules are hollow tubesabout 25 nanometers in diameter, each composed ofa ring of 13 protein protofilaments (figure 5.25b).Globular proteins consisting of dimers of alpha andbeta tubulin subunits polymerize to form the 13protofilaments. The protofilaments are arrayed sideby side around a central core, giving the microtubuleits characteristic tube shape. In many cells, micro-tubules form from MTOC nucleation centers nearthe center of the cell and radiate toward the periph-ery. They are in a constant state of flux, continuallypolymerizing and depolymerizing (the average half-life of a microtubule ranges from 10 minutes in anondividing animal cell to as short as 20 seconds in adividing animal cell), unless stabilized by the bindingof guanosine triphosphate (GTP) to the ends, whichinhibits depolymerization. The ends of the micro-tubule are designated as “+” (away from the nucle-ation center) or “−” (toward the nucleation center).Along with allowing for cellular movement, micro-tubules are responsible for moving materials withinthe cell itself. Special motor proteins, discussed laterin this chapter, move cellular organelles around thecell on microtubular “tracks.” Kinesin proteins moveorganelles toward the “+” end (toward the cell pe-riphery), and dyneins move them toward the “−” end.
96 Part II Biology of the Cell
Nuclearenvelope Nucleolus
Ribosomes MitochondrionRough
endoplasmicreticulum
Smoothendoplasmic
reticulumCytoskeleton
FIGURE 5.24The cytoskeleton. In this diagrammatic cross-section of aeukaryotic cell, the cytoskeleton, a network of fibers, supportsorganelles such as mitochondria.

3. Intermediate filaments. The most durable ele-ment of the cytoskeleton in animal cells is a systemof tough, fibrous protein molecules twined togetherin an overlapping arrangement (figure 5.25c). Thesefibers are characteristically 8 to 10 nanometers in di-ameter, intermediate in size between actin filamentsand microtubules (which is why they are called in-termediate filaments). Once formed, intermediatefilaments are stable and usually do not break down.Intermediate filaments constitute a heterogeneousgroup of cytoskeletal fibers. The most commontype, composed of protein subunits called vimentin,provides structural stability for many kinds of cells.Keratin, another class of intermediate filament, isfound in epithelial cells (cells that line organs andbody cavities) and associated structures such as hair
and fingernails. The intermediate filaments of nervecells are called neurofilaments.
As we will discuss in the next section, the cytoskeletonprovides an interior framework that supports the shape ofthe cell, stretching the plasma membrane much as the polesof a circus tent. Changing the relative length of cytoskeletonfilaments allows cells to rapidly alter their shape, extendingprojections out or folding inward. Within the cell, theframework of filaments provides a molecular highway alongwhich molecules can be transported.
Elements of the cytoskeleton crisscross the cytoplasm,supporting the cell shape and anchoring organelles inplace. There are three principal types of fibers: actinfilaments, microtubules, and intermediate filaments.
Chapter 5 Cell Structure 97
(b) Microtubule(a) Actin filament
(c) Intermediate filament
Mitochondrion Microtubule
Intermediate filament
Ribosome
Rough endoplasmicreticulum
Actin filament
Cell membrane
FIGURE 5.25Molecules that make up the cytoskeleton. (a) Actin filaments. Actin filaments are made of two strands of the fibrous protein actintwisted together and usually occur in bundles. Actin filaments are ubiquitous, although they are concentrated below the plasma membranein bundles known as stress fibers, which may have a contractile function. (b) Microtubules. Microtubules are composed of 13 stacks oftubulin protein subunits arranged side by side to form a tube. Microtubules are comparatively stiff cytoskeletal elements that serve toorganize metabolism and intracellular transport in the nondividing cell. (c) Intermediate filaments. Intermediate filaments are composed ofoverlapping staggered tetramers of protein. This molecular arrangement allows for a ropelike structure that imparts tremendousmechanical strength to the cell.

Cell MovementEssentially all cell motion is tied to the movement of actinfilaments, microtubules, or both. Intermediate filaments actas intracellular tendons, preventing excessive stretching ofcells, and actin filaments play a major role in determiningthe shape of cells. Because actin filaments can form and dis-solve so readily, they enable some cells to change shapequickly. If you look at the surfaces of such cells under a mi-croscope, you will find them alive with motion, as projec-tions, called microvilli in animal cells, shoot outward fromthe surface and then retract, only to shoot out elsewheremoments later (figure 5.26).
Some Cells Crawl
It is the arrangement of actin filaments within the cell cy-toplasm that allows cells to “crawl,” literally! Crawling is asignificant cellular phenomenon, essential to inflamma-tion, clotting, wound healing, and the spread of cancer.White blood cells in particular exhibit this ability. Pro-duced in the bone marrow, these cells are released intothe circulatory system and then eventually crawl out ofcapillaries and into the tissues to destroy potentialpathogens.
Cells exist in a gel-sol state; that is, at any given time,some regions of the cell are rigid (gel) and some are morefluid (sol). The cell is typically more sol-like in its interior,and more gel-like at its perimeter. To crawl, the cell cre-ates a weak area in the gel perimeter, and then forces thefluid (sol) interior through the weak area, forming apseudopod (“false foot”). As a result a large section of cy-toplasm oozes off in a different direction, but still remainswithin the plasma membrane. Once extended, the pseudo-pod stabilizes into a gel state, assembling actin filaments.Specific membrane proteins in the pseudopod stick to thesurface the cell is crawling on, and the rest of the cell isdragged in that direction. The pressure required to forceout a developing pseudopod is created when actin filamentsin the trailing end of the cell contract, just as squeezing awater balloon at one end forces the balloon to bulge out atthe other end.
Moving Material within the Cell
Actin filaments and microtubules often orchestrate their ac-tivities to affect cellular processes. For example, during cellreproduction (see chapter 11), newly replicated chromo-somes move to opposite sides of a dividing cell because theyare attached to shortening microtubules. Then, in animalcells, a belt of actin pinches the cell in two by contractinglike a purse string. Muscle cells also use actin filaments tocontract their cytoskeletons. The fluttering of an eyelash, theflight of an eagle, and the awkward crawling of a baby all de-pend on these cytoskeletal movements within muscle cells.
Not only is the cytoskeleton responsible for the cell’sshape and movement, but it also provides a scaffold thatholds certain enzymes and other macromolecules in definedareas of the cytoplasm. Many of the enzymes involved incell metabolism, for example, bind to actin filaments; so doribosomes. By moving and anchoring particular enzymesnear one another, the cytoskeleton, like the endoplasmicreticulum, organizes the cell’s activities.
Intracellular Molecular Motors
Certain eukaryotic cells must move materials from one placeto another in the cytoplasm. Most cells use the endomem-brane system as an intracellular highway; the Golgi appara-tus packages materials into vesicles that move through thechannels of the endoplasmic reticulum to the far reaches ofthe cell. However, this highway is only effective over shortdistances. When a cell has to transport materials throughlong extensions like the axon of a nerve cell, the ER high-ways are too slow. For these situations, eukaryotic cells havedeveloped high-speed locomotives that run along micro-tubular tracks.
Four components are required: (1) a vesicle or or-ganelle that is to be transported, (2) a motor moleculethat provides the energy-driven motion, (3) a connectormolecule that connects the vesicle to the motor mole-cule, and (4) microtubules on which the vesicle will ridelike a train on a rail. For example, embedded within themembranes of endoplasmic reticulum is a protein calledkinectin that bind the ER vesicles to the motor protein ki-nesin. As nature’s tiniest motors, these motor proteins lit-erally pull the transport vesicles along the microtubulartracks. Kinesin uses ATP to power its movement toward
98 Part II Biology of the Cell
FIGURE 5.26The surfaces of some cells are in constant motion. Thisamoeba, a single-celled protist, is advancing toward you, itsadvancing edges extending projections outward. The movingedges have been said to resemble the ruffled edges of a skirt.
the cell periphery, dragging the vesicle with it as it travelsalong the microtubule. Another vesicle protein (or per-haps a slight modification of kinesin—further research isneeded to determine which) binds vesicles to the motorprotein dynein, which directs movement in the oppositedirection, inward toward the cell’s center. (Dynein is alsoinvolved in the movement of eukaryotic flagella, as dis-cussed below.) The destination of a particular transportvesicle and its contents is thus determined by the natureof the linking protein embedded within the vesicle’smembrane.
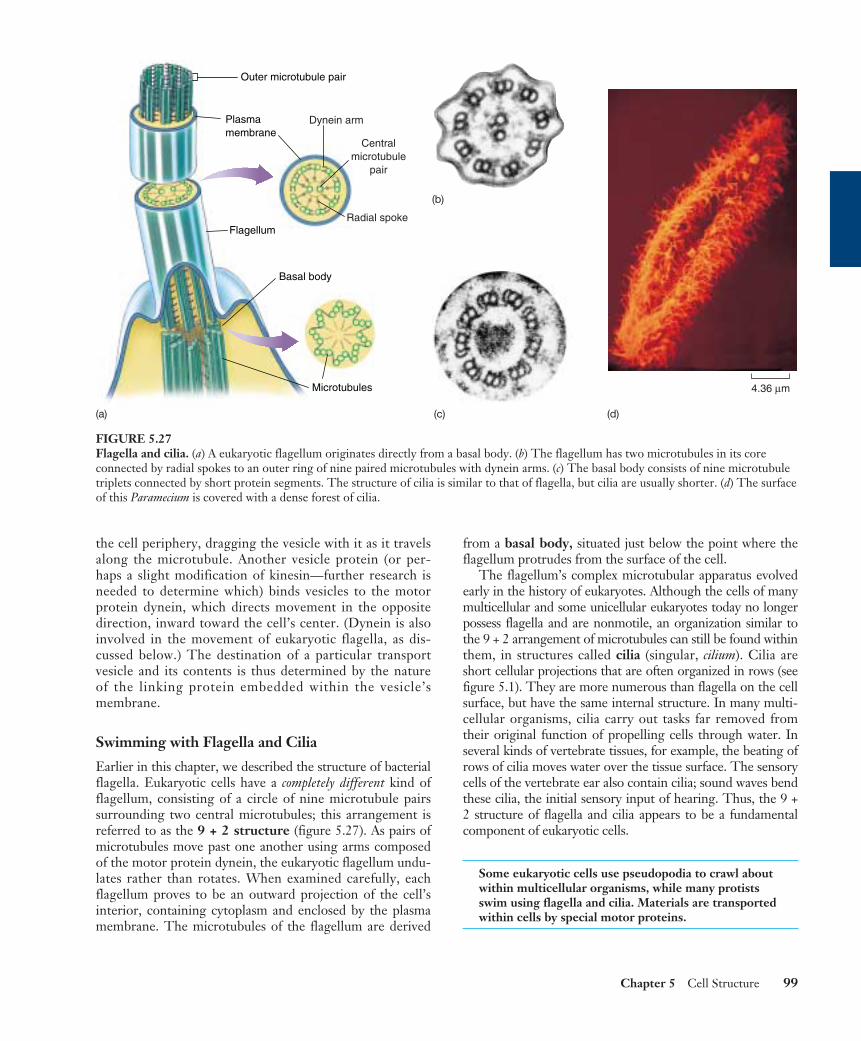
Swimming with Flagella and Cilia
Earlier in this chapter, we described the structure of bacterialflagella. Eukaryotic cells have a completely different kind offlagellum, consisting of a circle of nine microtubule pairssurrounding two central microtubules; this arrangement isreferred to as the 9 + 2 structure (figure 5.27). As pairs ofmicrotubules move past one another using arms composedof the motor protein dynein, the eukaryotic flagellum undu-lates rather than rotates. When examined carefully, eachflagellum proves to be an outward projection of the cell’sinterior, containing cytoplasm and enclosed by the plasmamembrane. The microtubules of the flagellum are derived
from a basal body, situated just below the point where theflagellum protrudes from the surface of the cell.
The flagellum’s complex microtubular apparatus evolvedearly in the history of eukaryotes. Although the cells of manymulticellular and some unicellular eukaryotes today no longerpossess flagella and are nonmotile, an organization similar tothe 9 + 2 arrangement of microtubules can still be found withinthem, in structures called cilia (singular, cilium). Cilia areshort cellular projections that are often organized in rows (seefigure 5.1). They are more numerous than flagella on the cellsurface, but have the same internal structure. In many multi-cellular organisms, cilia carry out tasks far removed fromtheir original function of propelling cells through water. Inseveral kinds of vertebrate tissues, for example, the beating ofrows of cilia moves water over the tissue surface. The sensorycells of the vertebrate ear also contain cilia; sound waves bendthese cilia, the initial sensory input of hearing. Thus, the 9 +2 structure of flagella and cilia appears to be a fundamentalcomponent of eukaryotic cells.
Some eukaryotic cells use pseudopodia to crawl aboutwithin multicellular organisms, while many protistsswim using flagella and cilia. Materials are transportedwithin cells by special motor proteins.
Chapter 5 Cell Structure 99
Outer microtubule pair
Microtubules
Flagellum
Basal body
Plasmamembrane
Dynein arm
Radial spoke
Centralmicrotubule
pair
(a)
(b)
(c) (d)
4.36 µm
FIGURE 5.27Flagella and cilia. (a) A eukaryotic flagellum originates directly from a basal body. (b) The flagellum has two microtubules in its coreconnected by radial spokes to an outer ring of nine paired microtubules with dynein arms. (c) The basal body consists of nine microtubuletriplets connected by short protein segments. The structure of cilia is similar to that of flagella, but cilia are usually shorter. (d) The surfaceof this Paramecium is covered with a dense forest of cilia.
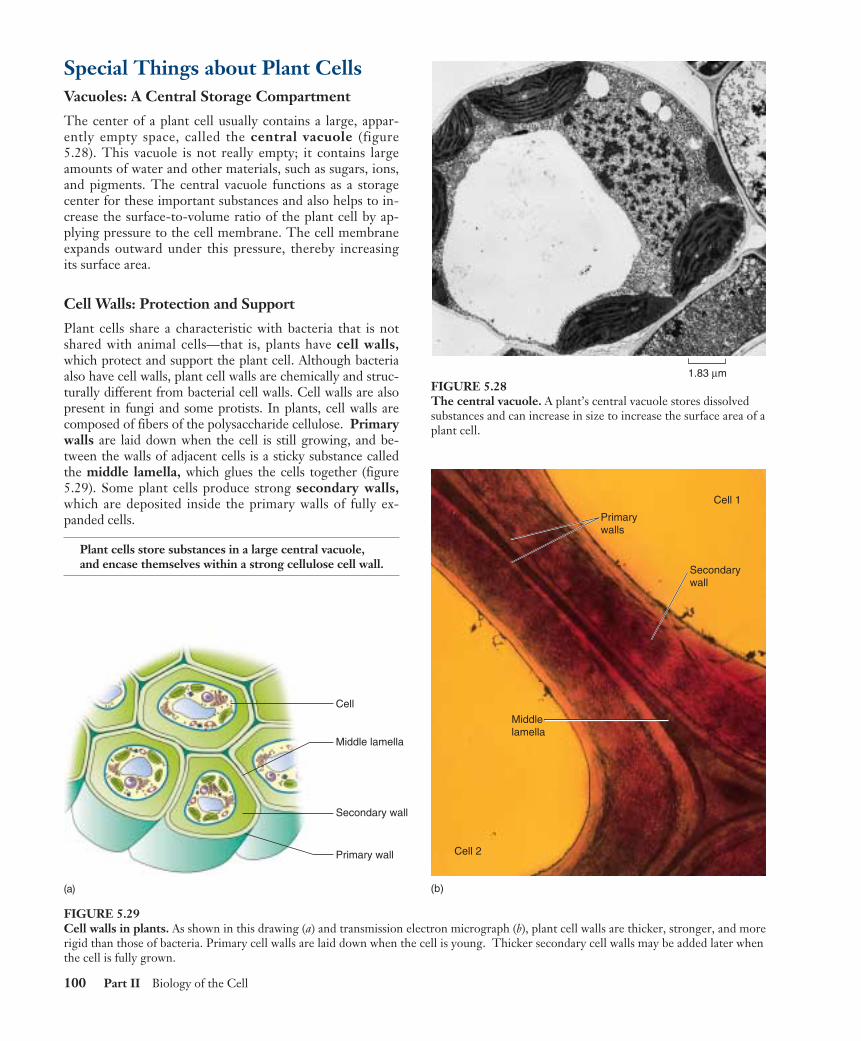
Special Things about Plant CellsVacuoles: A Central Storage Compartment
The center of a plant cell usually contains a large, appar-ently empty space, called the central vacuole (figure5.28). This vacuole is not really empty; it contains largeamounts of water and other materials, such as sugars, ions,and pigments. The central vacuole functions as a storagecenter for these important substances and also helps to in-crease the surface-to-volume ratio of the plant cell by ap-plying pressure to the cell membrane. The cell membraneexpands outward under this pressure, thereby increasingits surface area.
Cell Walls: Protection and Support
Plant cells share a characteristic with bacteria that is notshared with animal cells—that is, plants have cell walls,which protect and support the plant cell. Although bacteriaalso have cell walls, plant cell walls are chemically and struc-turally different from bacterial cell walls. Cell walls are alsopresent in fungi and some protists. In plants, cell walls arecomposed of fibers of the polysaccharide cellulose. Primarywalls are laid down when the cell is still growing, and be-tween the walls of adjacent cells is a sticky substance calledthe middle lamella, which glues the cells together (figure5.29). Some plant cells produce strong secondary walls,which are deposited inside the primary walls of fully ex-panded cells.
Plant cells store substances in a large central vacuole,and encase themselves within a strong cellulose cell wall.
100 Part II Biology of the Cell
Cell
Middle lamella
Primary wall
Secondary wall
FIGURE 5.29Cell walls in plants. As shown in this drawing (a) and transmission electron micrograph (b), plant cell walls are thicker, stronger, and morerigid than those of bacteria. Primary cell walls are laid down when the cell is young. Thicker secondary cell walls may be added later whenthe cell is fully grown.
Primary walls
Cell 1
Secondary wall
Cell 2
(b)
Middle lamella
(a)
1.83 µmFIGURE 5.28The central vacuole. A plant’s central vacuole stores dissolvedsubstances and can increase in size to increase the surface area of aplant cell.

Chapter 5 Cell Structure 101
EndosymbiosisSymbiosis is a close relationship between organisms of differ-ent species that live together. The theory of endosymbiosisproposes that some of today’s eukaryotic organelles evolvedby a symbiosis in which one species of prokaryote was en-gulfed by and lived inside another species of prokaryote thatwas a precursor to eukaryotes (figure 5.30). According to theendosymbiont theory, the engulfed prokaryotes providedtheir hosts with certain advantages associated with their spe-cial metabolic abilities. Two key eukaryotic organelles arebelieved to be the descendants of these endosymbioticprokaryotes: mitochondria, which are thought to have origi-nated as bacteria capable of carrying out oxidative metabo-lism; and chloroplasts, which apparently arose from photo-synthetic bacteria.
The endosymbiont theory is supported by a wealth ofevidence. Both mitochondria and chloroplasts are sur-rounded by two membranes; the inner membrane proba-bly evolved from the plasma membrane of the engulfedbacterium, while the outer membrane is probably derivedfrom the plasma membrane or endoplasmic reticulum ofthe host cell. Mitochondria are about the same size as mostbacteria, and the cristae formed by their inner membranesresemble the folded membranes in various groups of bac-teria. Mitochondrial ribosomes are also similar to bacterialribosomes in size and structure. Both mitochondria and
chloroplasts contain circular molecules of DNA similar tothose in bacteria. Finally, mitochondria divide by simplefission, splitting in two just as bacterial cells do, and theyapparently replicate and partition their DNA in much thesame way as bacteria. Table 5.2 compares and reviews thefeatures of three types of cells.
Some eukaryotic organelles are thought to have arisenby endosymbiosis.
5.4 Symbiosis played a key role in the origin of some eukaryotic organelles.
FIGURE 5.30Endosymbiosis. This figure shows how a double membrane mayhave been created during the symbiotic origin of mitochondria orchloroplasts.
Table 5.2 A Comparison of Bacterial, Animal, and Plant Cells
Bacterium Animal Plant
EXTERIOR STRUCTURESCell wallCell membraneFlagella
INTERIOR STRUCTURESERRibosomesMicrotubulesCentriolesGolgi apparatusNucleusMitochondriaChloroplastsChromosomesLysosomesVacuoles
Present (protein-polysaccharide)PresentMay be present (single strand)
AbsentPresentAbsentAbsentAbsentAbsentAbsentAbsentA single circle of DNAAbsentAbsent
AbsentPresentMay be present
Usually presentPresentPresentPresentPresentPresentPresentAbsentMultiple; DNA-protein complexUsually presentAbsent or small
Present (cellulose)PresentAbsent except in sperm of a few species
Usually presentPresentPresentAbsentPresentPresentPresentPresentMultiple; DNA-protein complexPresentUsually a large single vacuole
Chapter 5 Summary Questions Media Resources
5.1 All organisms are composed of cells.
• The cell is the smallest unit of life. All living thingsare made of cells.
• The cell is composed of a nuclear region, which holdsthe hereditary apparatus, enclosed within thecytoplasm.
• In all cells, the cytoplasm is bounded by a membranecomposed of phospholipid and protein.
102 Part II Biology of the Cell
1. What are the three principlesof the cell theory?2. How does the surface area-
to-volume ratio of cells limit thesize that cells can attain?
5.2 Eukaryotic cells are far more complex than bacterial cells.
• Bacteria, which have prokaryotic cell structure, donot have membrane-bounded organelles within theircells. Their DNA molecule is circular.
• The eukaryotic cell is larger and more complex, withmany internal compartments.
3. How are prokaryotesdifferent from eukaryotes interms of their cell walls, interiororganization, and flagella?
• A eukaryotic cell is organized into three principal zones:the nucleus, the cytoplasm, and the plasma membrane.Located in the cytoplasm are numerous organelles,which perform specific functions for the cell.
• Many of these organelles, such as the endoplasmicreticulum, Golgi apparatus (which gives rise tolysosomes), and nucleus, are part of a complexendomembrane system.
• Mitochondria and chloroplasts are part of the energy-processing system of the cell.
• The cytoskeleton encompasses a variety of fibrousproteins that provide structural support and performother functions for the cell.
• Many eukaryotic cells possess flagella or cilia having a9 + 2 arrangement of microtubules; sliding of themicrotubules past one another bends these cellularappendages.
• Cells transport materials long distances within thecytoplasm by packaging them into vesicles that arepulled by motor proteins along microtubule tracks.
4. What is the endoplasmicreticulum? What is its function?How does rough ER differ fromsmooth ER?5. What is the function of the
Golgi apparatus? How do thesubstances released by the Golgiapparatus make their way toother locations in the cell?6. What types of eukaryotic
cells contain mitochondria?What function do mitochondriaperform?7. What unique metabolic
activity occurs in chloroplasts?8. What cellular functions do
centrioles participate in?9. What kinds of cytoskeleton
fibers are stable and which arechangeable?10. How do cilia compare witheukaryotic flagella?
5.3 Take a tour of a eukaryotic cell.
• Present-day mitochondria and chloroplasts probablyevolved as a consequence of early endosymbiosis: theancestor of the eukaryotic cell engulfed a bacterium,and the bacterium continued to function within thehost cell.
11. What is the endosymbionttheory? What is the evidencesupporting this theory?
5.4 Symbiosis played a key role in the origin of some eukaryotic organelles.
• Exploration: Cell Size
• Surface to Volume
• Art Activities: -Animal Cell Structure-Plant Cell Structure-NonphotosyntheticBacterium
-Cyanobacterium
• Scientists on Science:The Joy of Discovery
• Art Quizzes:-Nucleosomes-Rough ER andProtein Synthesis
-Protein Transport
• Art Activities: -Anatomy of theNucleus
-Golgi ApparatusStructure
-MitochondrionStructure
-Organization ofCristae
-Chloroplast Structure-The Cytoskeleton-Plant Cell
• Endomembrane• Energy Organelles• Cytoskeleton
BIOLOGY
RAVEN
JOHNSON
S I X T H
E D I T I O N
www.mhhe.com/raven6/resources5.mhtml




















