
Biochemical, physiological and growth changes in response to salinity in callus cultures of Sesuvium...
9
ORIGINAL PAPER Biochemical, physiological and growth changes in response to salinity in callus cultures of Sesuvium portulacastrum L. Vinayak Haribhau Lokhande • Tukaram Dayaram Nikam • Suprasanna Penna Received: 2 June 2009 / Accepted: 20 January 2010 / Published online: 6 February 2010 Ó Springer Science+Business Media B.V. 2010 Abstract In vitro-grown cells of Sesuvium portulacastrum L., an important ‘salt accumulator’ mangrove associate, were incubated on a medium containing different levels of salt, including 0, 100, 200, or 400 mM NaCl, in order to evaluate biochemical, physiological, and growth responses. A significant decrease in callus growth, water status, and cell membrane damage was observed under salt stress. Osmotic adjustment was revealed by the accumulation of inorganic ions, such as sodium (Na ? ), and organic osmolytes (proline, glycine betaine, and total soluble sugars) in NaCl-treated calli compared to control. However, accretion of osmolytes and inorganic ions did not support growth of calli under NaCl stress conditions. The observed reduced growth rate in calli subjected to stress, up to 200 mM NaCl, was coupled with lower catalase and ascorbate peroxidase activities and with a significantly higher superoxide dismutase activity. These findings suggested that S. portulacastrum cell cultures exhibited higher osmotic adjustment to salt stress. Keywords Antioxidative enzymes In vitro Osmolytes Salt tolerance Sesuvium portulacastrum Water status Introduction Sesuvium portulacastrum L., commonly called seapurslane, belongs to the family Aizoaceae, and is a herbaceous facultative halophyte (Lokhande et al. 2009a, b). It grows naturally in the subtropical, Mediterranean, coastal and warmer areas around the world, and in India, it grows along the coastal sides of both eastern and western regions (Lokhande et al. 2009b). The plant plays a major role in the environmental protection such as sand dune fixation, saline soil stabilization, desalination, desert greenification,and landscaping in an ornamental capacity (Menzel and Leith 1999), and acts as an alternative culture to problematic soil in the arid and semiarid regions of the world. Of the var- ious categories of mangroves, Sesuvium comes under ‘salt accumulators’ as the plants accumulate high concentrations of salts in their cells and tissues and overcome salt toxicity by developing succulence (Ashraf 2009). Salinity is one of the most important environmental constraints affecting more than 800 million ha of arable land (Munns and Tester 2008) and, therefore, the agricul- tural productivity (Sairam et al. 2002). Thus, studies aimed at understanding physiological and biochemical mecha- nisms under salt stress has generated considerable interest. Generally, plants under saline conditions are stressed by an increase in the osmotic potential in the rooting medium and by the toxic effect of high concentration of saline ions (Munns 2002). Osmotic adjustment under salt stress is an adaptation mechanism operated by both halophytes and glycophytes in order to maintain their water balance (Flowers and Colmer 2008). The best characterized bio- chemical response of plant cells to osmotic stress is accu- mulation of some inorganic ions such as sodium (Na ? ) and compatible organic solutes like proline, glycine betaine (GB), and soluble sugars (Flowers and Colmer 2008). These compatible solutes can accumulate to high levels without disturbing intracellular biochemistry (Bohnert and Jensen 1996), protecting sub-cellular structures, mitigating oxidative damage caused by free radicals (Attipali et al. V. H. Lokhande T. D. Nikam Department of Botany, University of Pune, Pune 411 007, India V. H. Lokhande S. Penna (&) Functional Plant Biology Section, Nuclear Agriculture & Biotechnology Division, Bhabha Atomic Research Centre, Trombay, Mumbai 400 085, India e-mail: [email protected]; [email protected] 123 Plant Cell Tiss Organ Cult (2010) 102:17–25 DOI 10.1007/s11240-010-9699-3
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Biochemical, physiological and growth changes in response to salinity in callus cultures of Sesuvium...

ORIGINAL PAPER
Biochemical, physiological and growth changes in responseto salinity in callus cultures of Sesuvium portulacastrum L.
Vinayak Haribhau Lokhande •
Tukaram Dayaram Nikam • Suprasanna Penna
Received: 2 June 2009 / Accepted: 20 January 2010 / Published online: 6 February 2010
� Springer Science+Business Media B.V. 2010
Abstract In vitro-grown cells of Sesuvium portulacastrum
L., an important ‘salt accumulator’ mangrove associate,
were incubated on a medium containing different levels of
salt, including 0, 100, 200, or 400 mM NaCl, in order to
evaluate biochemical, physiological, and growth responses.
A significant decrease in callus growth, water status, and cell
membrane damage was observed under salt stress. Osmotic
adjustment was revealed by the accumulation of inorganic
ions, such as sodium (Na?), and organic osmolytes (proline,
glycine betaine, and total soluble sugars) in NaCl-treated
calli compared to control. However, accretion of osmolytes
and inorganic ions did not support growth of calli under NaCl
stress conditions. The observed reduced growth rate in calli
subjected to stress, up to 200 mM NaCl, was coupled with
lower catalase and ascorbate peroxidase activities and with a
significantly higher superoxide dismutase activity. These
findings suggested that S. portulacastrum cell cultures
exhibited higher osmotic adjustment to salt stress.
Keywords Antioxidative enzymes � In vitro � Osmolytes �Salt tolerance � Sesuvium portulacastrum � Water status
Introduction
Sesuvium portulacastrum L., commonly called seapurslane,
belongs to the family Aizoaceae, and is a herbaceous
facultative halophyte (Lokhande et al. 2009a, b). It grows
naturally in the subtropical, Mediterranean, coastal and
warmer areas around the world, and in India, it grows along
the coastal sides of both eastern and western regions
(Lokhande et al. 2009b). The plant plays a major role in the
environmental protection such as sand dune fixation, saline
soil stabilization, desalination, desert greenification,and
landscaping in an ornamental capacity (Menzel and Leith
1999), and acts as an alternative culture to problematic soil
in the arid and semiarid regions of the world. Of the var-
ious categories of mangroves, Sesuvium comes under ‘salt
accumulators’ as the plants accumulate high concentrations
of salts in their cells and tissues and overcome salt toxicity
by developing succulence (Ashraf 2009).
Salinity is one of the most important environmental
constraints affecting more than 800 million ha of arable
land (Munns and Tester 2008) and, therefore, the agricul-
tural productivity (Sairam et al. 2002). Thus, studies aimed
at understanding physiological and biochemical mecha-
nisms under salt stress has generated considerable interest.
Generally, plants under saline conditions are stressed by an
increase in the osmotic potential in the rooting medium and
by the toxic effect of high concentration of saline ions
(Munns 2002). Osmotic adjustment under salt stress is an
adaptation mechanism operated by both halophytes and
glycophytes in order to maintain their water balance
(Flowers and Colmer 2008). The best characterized bio-
chemical response of plant cells to osmotic stress is accu-
mulation of some inorganic ions such as sodium (Na?) and
compatible organic solutes like proline, glycine betaine
(GB), and soluble sugars (Flowers and Colmer 2008).
These compatible solutes can accumulate to high levels
without disturbing intracellular biochemistry (Bohnert and
Jensen 1996), protecting sub-cellular structures, mitigating
oxidative damage caused by free radicals (Attipali et al.
V. H. Lokhande � T. D. Nikam
Department of Botany, University of Pune, Pune 411 007, India
V. H. Lokhande � S. Penna (&)
Functional Plant Biology Section, Nuclear Agriculture &
Biotechnology Division, Bhabha Atomic Research Centre,
Trombay, Mumbai 400 085, India
e-mail: [email protected]; [email protected]
123
Plant Cell Tiss Organ Cult (2010) 102:17–25
DOI 10.1007/s11240-010-9699-3

2004), and maintaining the enzyme activities under salt
stress (Yokoi et al. 2002).
Potassium plays a major role in the biochemical and
physiological processes in plants. Under typical physio-
logical conditions, plant cells require high K? (100–
200 mM) and lower Na? (less than 1 mM) and, accord-
ingly, the high cytosolic K?/Na? ratio to maintain the
osmotic balance (Tester and Davenport 2003) for proper
functioning of the cell. To maintain a high K?/Na? ratio in
the cytosol, plant cells employ primary active transport,
mediated by channels and co-transporters for Na? extru-
sion and/or the intracellular compartmentalization of Na?
into the vacuole (Blumwald 2000). However, under salt
stress, ion ratios are altered by the uncontrolled influx of
Na? through K? pathways. Halophytes being continuously
exposed to high salinity conditions undergo osmotic
adjustment, preferably through higher accumulation of
Na? and its efficient sequestration into the vacuole at low
external water potential as well as by low potassium use
efficiency as compared to glycophytes.
In addition to osmotic stress, salt stress also causes oxi-
dative damage, thereby affecting the cellular membrane
integrity, enzyme activities, and functioning of plant pho-
tosynthetic apparatus (Jitesh et al. 2006). The antioxidant
enzymes like superoxide dismutase (SOD), catalases (CAT),
ascorbate peroxidase (APX) and other enzymes are effi-
ciently involved in scavenging of reactive oxygen species
(ROS) produced by salt stress, and act as one of the main
tolerance mechanisms against oxidative stress in plants.
The response to osmotic and oxidative stresses in plants
has involved a suite of adaptations at cellular, tissue, and
whole plant level (Hasegawa et al. 2000; Borsani et al.
2003). Many studies have revealed that the salt tolerance
mechanism operating at whole plant level also worked at
the cellular level in both halophytes and non-halophytes
(Tonon et al. 2004; Arzani 2008). However, there is mea-
gre information available on the cellular responses to salt
and involvement of antioxidant defence system in man-
groves. Recently, Zhao et al. (2009) reported the effect of
salt stress at cellular level on growth and osmotic regula-
tion in Thellungiella halophila. In Sesuvium, studies at
whole plant level have demonstrated the role of osmolytes
and inorganic ions in osmotic adjustment under salinity
stress (Messedi et al. 2004), but there is no information at
the cellular level on the involvement of these osmolytes
and especially antioxidant enzyme defence capacitance to
salinity tolerance. The assessment of intracellular changes
operating at cellular levels under salt stress can be a useful
means to segregate the specific effect of salt and to identify
specific physiological and biochemical traits associated
with salt tolerance at whole plant level. Thus, the present
investigation was carried out with the objective of using in
vitro cultures to evaluate the responses to salt-induced
stress in Sesuvium. Specifically, we studied the effect of
salt on growth, osmotic adjustment, and antioxidant
defence mechanism to look into the mechanism used by in
vitro-cultured cells of this halophyte, Sesuvium portula-
castrum.
Materials and methods
Plant material
The nodal sectors (*3.0 cm) of S. portulacastrum were
collected from the coastal region of Maharashtra state,
India, and maintained at the Botanic Garden, Department
of Botany, University of Pune, Pune-07 (Lokhande et al.
2009a). The nodal explants (*1.0 cm) from established
plantlets were harvested and surface sterilized with 0.1%
(w/v) mercuric chloride (HgCl2) solution for 5 min, and
rinsed five times with sterile distilled water. The explants
were cultured vertically onto the shoot multiplication
medium (MS; Murashige and Skoog 1962) supplemented
with 20 lM 6-benzyladenine (BA) and 30 g l-1 sucrose.
The pH of the medium was adjusted to 5.8 and solidified
with 0.2% gelrite (Sigma–Aldrich, USA) prior to auto-
claving at 121�C for 15 min. The cultures were incubated
at 25 ± 2�C under a 16-h photoperiod (30 lmol m-2 s-1
PFD) and 70% relative humidity.
Callus induction, proliferation and maintenance
After 4 weeks of culture, callus induction was observed on
the shoot multiplication medium at the lower cut end on
nodal explant and was hard, compact, greenish and non-
proliferated. The callus was incubated on Murashige and
Skoog (MS) (1962) solid medium consisting of 10 lM 2,4-
dichlorophenoxyacetic acid (2,4-D) and 5.0 lM BA and
30 g l-1 sucrose, for its proliferation. After 2 weeks of
culture, the callus underwent proliferation and was soft,
fragile and whitish in nature. Subsequently, the callus was
regularly sub-cultured at intervals of 21 days on the freshly
prepared callus proliferation medium (MS medium con-
taining 10 lM 2,4-D and 5.0 lM BA) and, after mainte-
nance for more than 8 months under control conditions,
was used for salt stress analyses.
Salt stress treatment
For the availability of adequate nutrient and oxygen supply
to inner part of the callus, the calli were cut into small
pieces of approximately 4–5 mm size. Only the fresh,
viable and actively growing and proliferating calli pieces
were weighed (*1.0 g) and cultured on solid callus pro-
liferation medium (MS consisting of 10 lM 2,4-D and
18 Plant Cell Tiss Organ Cult (2010) 102:17–25
123

5.0 lM BA) supplemented in the absence or presence of
various levels of salt (0, 100, 200, or 400 mM NaCl) in
addition to 30 g l-1 sucrose as a source of carbon. The pH
of the medium was adjusted to 5.8 and solidified with 0.2%
gelrite prior to autoclaving at 121�C for 15 min. The
cultures were incubated under controlled conditions as
described earlier. Calli were evaluated after 15 days of salt
treatment for growth rate, accumulation of proline, glycine
betaine, and total soluble sugars. Damage to membrane
was measured in terms of relative electrolytic leakage, and
antioxidant enzyme activities were analyzed for CAT, APX
and SOD.
Relative growth rate measurements
Samples were weighed to calculate relative growth rate
(RGR). Calli were weighed initially at the time of their
transfer (Wi) and finally after 15 days of salt treatment
(Wf), and the mean callus RGR calculated by: [(Wf - Wi)/
Wi] 9 100%.
Measurement of membrane damage rate
Membrane damage was determined in terms of relative
electrolytic leakage (REL) according to the method of
Sullivan (1972). This technique was based on the increase of
cellular membrane permeability and concomitantly greater
electrolyte diffusion out of cells when tissue is injured by a
stress situation. The determination of REL is the measure of
change in the electrical impedance and electrolyte leakage
to detect stress injury of cell membrane. To measure the
REL, calli clumps of approximately equal size were incu-
bated for 24 h in a test tube (25 9 150 mm) containing
10 ml of distilled water at room temperature (25�C) and the
initial electrical conductivity (EC1) was measured after
incubation period. Then, the samples were killed by auto-
claving for 15 min at 121�C temperature to release all the
ions from the tissue and final electrical conductivity (EC2)
was measured when cooled to room temperature. The REL
was calculated as: (EC1/EC2) 9 100.
Determination of tissue water content
The fresh weight (FW) of the calli was determined
immediately after removal from the medium and blotted
with tissue paper to remove the excess water. Dry weight
(DW) was recorded after drying the calli at 60�C in the hot
air oven for 48 h. The percent tissue water content (TWC
%) of the calli was determined using the following
equation:
TWC %ð Þ ¼ FW� DWð Þ=FW½ � � 100
Measurement of osmotic regulation
Free proline content
Free proline content was measured spectrophotometrically
according to the method of Bates et al. (1973). About
500 mg of fresh calli were homogenized in 3% (w/v)
aqueous 50-sulfosalicylic acid, and the residue was
removed by centrifugation at 10,000g for 10 min at 4�C.
The supernatant (2 ml) was reacted with 2 ml each of acid
ninhydrin and glacial acetic acid by incubating at 100�C in
a hot water bath for 1 h. The reaction was terminated in an
ice bath and allowed to cool at room temperature. The
reaction mixture was extracted with 4 ml toluene and
mixed vigorously with a stirrer for 10–15 s. The chromo-
phore containing toluene was aspirated from the aqueous
phase and warmed to room temperature. The optical den-
sity was measured at 520 nm (UV-1700 PharmaSpec, UV–
Visible spectrophotometer; Shimadzu, Japan) using toluene
as a blank. The amount of proline was determined from a
standard curve using L-proline and expressed as lg of
proline g-1 FW.
Glycine betaine (GB) analysis
Accumulation of GB in response to salt stress was deter-
mined according to Grieve and Grattan (1983). The fresh
calli (500 mg) were ground in liquid nitrogen and the finely
ground powder was mechanically shaken with 20 ml of
deionised water for 16 h at 25�C. Samples were filtered,
and filtrates (500 ll) were diluted (1:1) with 2N H2SO4.
The extract was cooled in an ice water for 1 h and then
mixed with 200 ll of I2-KI reagent (mixed with 20%
potassium iodide and 15.7% iodine). The tubes were gently
mixed and stored at 4�C for 16 h followed by centrifuga-
tion at 10,000g for 15 min at 0�C. Periodide crystals were
dissolved in 9.0 ml of 1, 2-dichloroethane, and after 2 h,
absorbance was measured spectrophotometrically at
365 nm (UV-1700 PharmaSpec, UV–Visible spectropho-
tometer; Shimadzu). The GB content (lg g-1 FW) was
determined from a standard curve prepared using glycine
betaine (Sigma–Aldrich) as standard.
Total soluble sugars (TSS) content
TSS was estimated as per the anthrone method (Watanabe
et al. 2000) with some modifications. About 200 mg of
calli were homogenized with ice-chilled 80% ethanol in a
mortar and pestle. The extract was prepared by centrifu-
gation at 5,000g for 10 min at 4�C, and final volume was
adjusted to 10 ml with 80% ethanol. Then 1 ml of super-
Plant Cell Tiss Organ Cult (2010) 102:17–25 19
123

natant was reacted with 3 ml of freshly prepared anthrone
reagent by incubating the reaction mixture for 10 min at
100�C in a hot water bath. The reaction was terminated by
quick cooling in an ice bath and allowed to cool at room
temperature. The optical density was measured spectro-
photometrically at 620 nm (UV-1700 PharmaSpec, UV–
Visible spectrophotometer; Shimadzu). A standard curve
was prepared using D-glucose; the TSS was calculated and
expressed as mg g-1 FW.
Measurement of inorganic ions concentration
For the quantification of major inorganic ions (Na?, K?
and Ca?), calli harvested from four individual petriplates
per treatment were oven-dried at 80�C for 48 h.
Samples (100 mg) from each treatment were acid diges-
ted in 35% (v/v) HNO3 at 100�C for 30 min; and the
ion content was analyzed by flame atomic absorption
spectrophotometry.
Measurement of antioxidant enzyme activities
Extraction
The treated and control calli (500 mg) were homogenized
in 5 ml of ice cold 50 mM sodium phosphate buffer (pH
7.0) containing 0.1 mM EDTA and 1% (w/v) Polyvinyl-
pyrrolidone (PVP) with chilled mortar and pestle. The
homogenate was filtrated with single layered cheese cloth
and centrifuged at 15,000g for 20 min at 4�C. An appro-
priate aliquot/dilution of the supernatant was used as a
crude enzyme(s) for determination of antioxidant enzyme
activities. Soluble protein content in the enzyme extract
was determined according to Bradford (1976) using Bovine
Serum Albumin (BSA) as a standard.
Catalase (CAT) assay
CAT (EC 1.11.1.6) activity was measured by following
the decomposition of hydrogen peroxide (H2O2) as
described by Cakmak and Marschner (1992) with minor
modifications. The activity was measured in a reaction
mixture (1 ml) containing 50 mM phosphate buffer (pH
7.0) and 15 mM H2O2. The reaction was initiated by
adding 50 ll enzyme extract and the activity was deter-
mined as a result of H2O2 decomposition by monitor-
ing the decrease in absorbance at 240 nm (e = 36
mM-1 cm-1) for 2 min at an interval of 15 s. The slope
of readings between the time interval considered as DA
and the enzyme activity was expressed as lKat of CAT
activity mg-1 protein.
Ascorbate peroxidase (APX) assay
APX (EC 1.11.1.11) activity was determined according to
Nakano and Asada (1981). The reaction mixture (1 ml)
contained 50 mM phosphate buffer (pH 7.0), 0.5 mM
ascorbate, 0.1 mM H2O2, and 0.1 mM EDTA. The reaction
was started by adding 100 ll of crude enzyme. Ascorbate
oxidation was monitored for 1 min by measuring the
decrease in absorbance at 290 nm at every 15 s
(e = 2.8 mM-1 cm-1). The enzyme activity was expressed
as lKat of APX activity mg-1 protein.
Superoxide dismutase (SOD) assay
The total SOD (EC 1.15.1.1) was assayed according to
Becana et al. (1986) by inhibition of the photochemical
reduction of nitroblue tetrazolium (NBT). The reaction
mixture (3 ml) containing 50 mM phosphate buffer (pH
7.8) and 0.1 mM EDTA to which an oxygen-generating
system containing 14.3 mM methionine, 82.5 lM NBT,
and 2.2 lM riboflavin, prepared freshly in situ, was added.
The reaction was initiated by adding 100 ll of crude
enzyme. The entire system was kept 30 cm below the light
source (six 15-W fluorescent tubes) for 30 min. The reac-
tion was stopped by switching off the light. For light blank,
all the reactant without enzyme extract was incubated in
light as for the samples, whereas all the reactants along
with 100 ll enzyme extract were incubated in dark for dark
blank. The reduction in NBT was measured by monitoring
the change in absorbance at 560 nm. The readings of light
blank were used in calculation of enzyme units. 1 U SOD
enzyme was defined as amount that produces 50% inhibi-
tion of NBT reduction under the assay condition and
expressed as U SOD activity mg-1 protein.
Statistical analyses
The experiment was laid out in a completely randomized
design (CRD). The analyses were repeated with three inde-
pendent biological samples. The data were analyzed by one-
way analysis of variance (ANOVA) using the statistical
software SPSS 10.0, and the treatment means were compared
by using Duncan’s multiple range test (DMRT) at P B 0.05.
Data were expressed as mean ± standard error (SE).
Results
Effect of salt stress on calli growth, membrane damage
rate (MDR) and water status
Fresh, actively proliferating calli cut into small pieces of
approximately 4–5 mm size were used for the analyses
20 Plant Cell Tiss Organ Cult (2010) 102:17–25
123
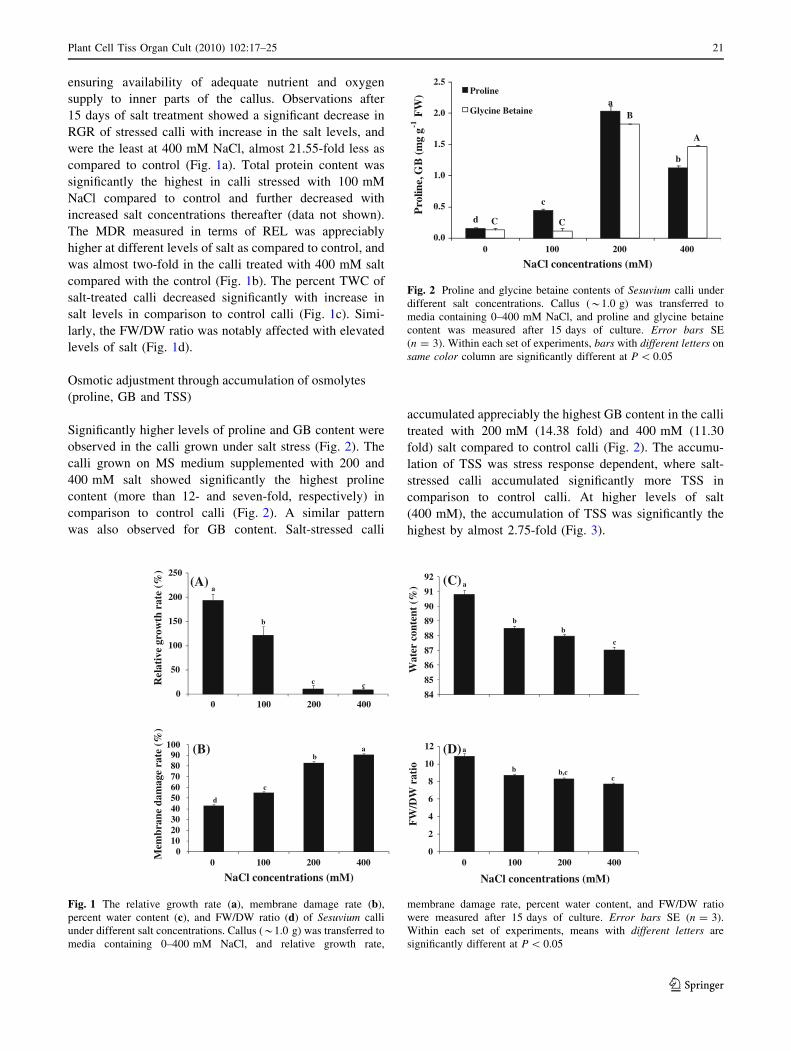
ensuring availability of adequate nutrient and oxygen
supply to inner parts of the callus. Observations after
15 days of salt treatment showed a significant decrease in
RGR of stressed calli with increase in the salt levels, and
were the least at 400 mM NaCl, almost 21.55-fold less as
compared to control (Fig. 1a). Total protein content was
significantly the highest in calli stressed with 100 mM
NaCl compared to control and further decreased with
increased salt concentrations thereafter (data not shown).
The MDR measured in terms of REL was appreciably
higher at different levels of salt as compared to control, and
was almost two-fold in the calli treated with 400 mM salt
compared with the control (Fig. 1b). The percent TWC of
salt-treated calli decreased significantly with increase in
salt levels in comparison to control calli (Fig. 1c). Simi-
larly, the FW/DW ratio was notably affected with elevated
levels of salt (Fig. 1d).
Osmotic adjustment through accumulation of osmolytes
(proline, GB and TSS)
Significantly higher levels of proline and GB content were
observed in the calli grown under salt stress (Fig. 2). The
calli grown on MS medium supplemented with 200 and
400 mM salt showed significantly the highest proline
content (more than 12- and seven-fold, respectively) in
comparison to control calli (Fig. 2). A similar pattern
was also observed for GB content. Salt-stressed calli
accumulated appreciably the highest GB content in the calli
treated with 200 mM (14.38 fold) and 400 mM (11.30
fold) salt compared to control calli (Fig. 2). The accumu-
lation of TSS was stress response dependent, where salt-
stressed calli accumulated significantly more TSS in
comparison to control calli. At higher levels of salt
(400 mM), the accumulation of TSS was significantly the
highest by almost 2.75-fold (Fig. 3).
84
85
86
87
88
89
90
91
92
Wat
er c
onte
nt (
%) (C) a
bb
c
0
2
4
6
8
10
12
FW
/DW
rat
io
NaCl concentrations (mM)
(D)a
b b,cc
0
50
100
150
200
250
Rel
ativ
e gr
owth
rat
e (%
)
(A)a
b
c c
0102030405060708090
100
0 100 200 4000 100 200 400
0 100 200 400
Mem
bran
e da
mag
e ra
te (
%)
NaCl concentrations (mM)
(B) ab
c
d
Fig. 1 The relative growth rate (a), membrane damage rate (b),
percent water content (c), and FW/DW ratio (d) of Sesuvium calli
under different salt concentrations. Callus (*1.0 g) was transferred to
media containing 0–400 mM NaCl, and relative growth rate,
membrane damage rate, percent water content, and FW/DW ratio
were measured after 15 days of culture. Error bars SE (n = 3).
Within each set of experiments, means with different letters are
significantly different at P \ 0.05
d
c
a
b
C C
B
A
0.0
0.5
1.0
1.5
2.0
2.5
0 100 200 400
Pro
line,
GB
(mg
g-1 F
W)
NaCl concentrations (mM)
Proline
Glycine Betaine
Fig. 2 Proline and glycine betaine contents of Sesuvium calli under
different salt concentrations. Callus (*1.0 g) was transferred to
media containing 0–400 mM NaCl, and proline and glycine betaine
content was measured after 15 days of culture. Error bars SE
(n = 3). Within each set of experiments, bars with different letters on
same color column are significantly different at P \ 0.05
Plant Cell Tiss Organ Cult (2010) 102:17–25 21
123
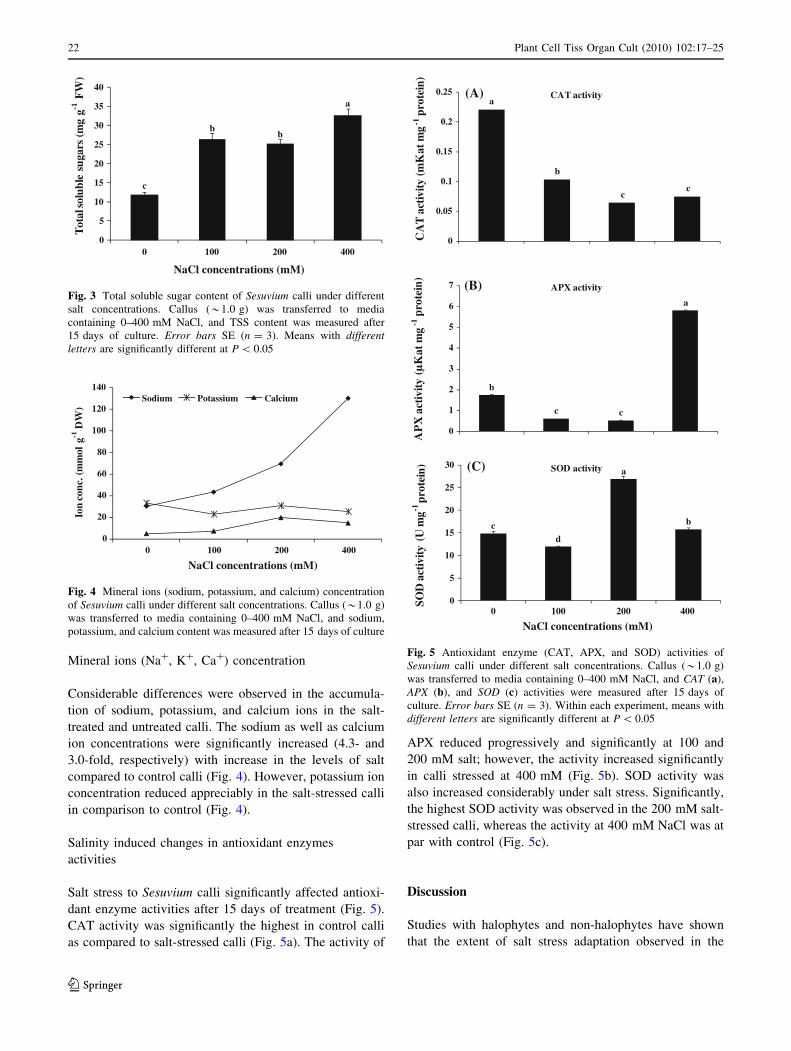
Mineral ions (Na?, K?, Ca?) concentration
Considerable differences were observed in the accumula-
tion of sodium, potassium, and calcium ions in the salt-
treated and untreated calli. The sodium as well as calcium
ion concentrations were significantly increased (4.3- and
3.0-fold, respectively) with increase in the levels of salt
compared to control calli (Fig. 4). However, potassium ion
concentration reduced appreciably in the salt-stressed calli
in comparison to control (Fig. 4).
Salinity induced changes in antioxidant enzymes
activities
Salt stress to Sesuvium calli significantly affected antioxi-
dant enzyme activities after 15 days of treatment (Fig. 5).
CAT activity was significantly the highest in control calli
as compared to salt-stressed calli (Fig. 5a). The activity of
APX reduced progressively and significantly at 100 and
200 mM salt; however, the activity increased significantly
in calli stressed at 400 mM (Fig. 5b). SOD activity was
also increased considerably under salt stress. Significantly,
the highest SOD activity was observed in the 200 mM salt-
stressed calli, whereas the activity at 400 mM NaCl was at
par with control (Fig. 5c).
Discussion
Studies with halophytes and non-halophytes have shown
that the extent of salt stress adaptation observed in the
c
bb
a
0
5
10
15
20
25
30
35
40
0 100 200 400
Tot
al so
lubl
e su
gars
(mg
g-1 F
W)
NaCl concentrations (mM)
Fig. 3 Total soluble sugar content of Sesuvium calli under different
salt concentrations. Callus (*1.0 g) was transferred to media
containing 0–400 mM NaCl, and TSS content was measured after
15 days of culture. Error bars SE (n = 3). Means with differentletters are significantly different at P \ 0.05
0
20
40
60
80
100
120
140
0 100 200 400
Ion
conc
. (m
mol
g-1
DW
)
NaCl concentrations (mM)
Sodium Potassium Calcium
Fig. 4 Mineral ions (sodium, potassium, and calcium) concentration
of Sesuvium calli under different salt concentrations. Callus (*1.0 g)
was transferred to media containing 0–400 mM NaCl, and sodium,
potassium, and calcium content was measured after 15 days of culture
b
c c
a
0
1
2
3
4
5
6
7
AP
X a
ctiv
ity (
µK
at m
g-1
prot
ein) APX activity
a
b
cc
0
0.05
0.1
0.15
0.2
0.25
CA
T a
ctiv
ity (m
Kat
mg
-1pr
otei
n)
CAT activity
cd
a
b
0
5
10
15
20
25
30
0 100 200 400
SOD
act
ivity
(U
mg-1
prot
ein)
NaCl concentrations (mM)
SOD activity
(A)
(B)
(C)
Fig. 5 Antioxidant enzyme (CAT, APX, and SOD) activities of
Sesuvium calli under different salt concentrations. Callus (*1.0 g)
was transferred to media containing 0–400 mM NaCl, and CAT (a),
APX (b), and SOD (c) activities were measured after 15 days of
culture. Error bars SE (n = 3). Within each experiment, means with
different letters are significantly different at P \ 0.05
22 Plant Cell Tiss Organ Cult (2010) 102:17–25
123

whole plant is also exhibited in callus tissue (Tonon et al.
2004; Arzani 2008); however, few in vitro culture systems
have been established from mangrove plants for studying
their salt tolerance at the cellular level (Cherian and Reddy
2003; Amor et al. 2005; Zhao et al. 2009). The study of salt
tolerance mechanism at the field level may not be preferred
as the salinity levels in field vary depending on season and
soil depth. Therefore, salt stress studies are preferably
conducted in soil-less cultures (like callus or axillary
shoots) with nutrient solutions of known salt concentrations
(Vijayan et al. 2003). In vitro culture provides a controlled
and uniform environment for studying physiological and
biological processes in plants particularly at the cellular
level under salt-induced osmotic stress (Ahmad et al.
2007). Therefore, in the present investigation, cellular
growth responses of Sesuvium under salt stress were stud-
ied. The results of the present investigation revealed that
Sesuvium calli respond fairly differently to salt stress in
comparison to responses observed in whole plant under
salinity or drought stress (Messedi et al. 2004; Slama et al.
2008). The higher levels of tissue organization in the whole
plant might efficiently distribute the effect of toxic saline
ions into different parts of the plants through its seques-
tration into the vacuoles and/or extrusion and hence show
better salt tolerance capacity which was not found at the
cellular level. In addition, restriction to entry of excess
toxic ions inside the plant at the root surface reduces the
heritable damage of the whole plant, while in the case of
callus cultures, the unorganised mass of calli is directly
exposed to toxic concentrations of saline ions in the med-
ium and hence loses its compatibility for sequestration of
these ions into the vacuoles. Furthermore, there are possi-
bly several physiological factors operating at the whole
plant level conferring salt tolerance which are lacking in
the dedifferentiated and unorganised callus cultures (Zhang
et al. 2004). This could be the reason that callus cultures
and whole plant respond differentially to salt stress.
One of the strategies developed by plants under salt
stress is to slow down their growth rate, which has been
observed in a number of in vitro systems of halophytes and
non-halophytes (Yasumoto et al. 1999; Khayri 2002; Zhang
et al. 2004; Patade et al. 2008; Zhao et al. 2009). The
reduction in growth not only helps the plant to save the
energy for defence purposes but also limits the risk of
heritable damage (May et al. 1998). The present study has
shown reduction in growth kinetics of Sesuvium calli in
terms of RGR (Fig. 1a), as has been observed in different
plant species: halophytes such as Sonnertia alba (Yasumoto
et al. 1999), Populus euphratica (Zhang et al. 2004),
Thellungiella halophila (Zhao et al. 2009,) and glycophytes
like tobacco (Watad et al. 1991), Saccharum officinarum
(Patade et al. 2008), and Arabidopsis thaliana (Zhao et al.
2009). The hyper-osmotic stress reduces the water
availability to the cell in the medium (Chamandoosti 2007),
as a result of which the FW and water content (Fig. 1c) of
salt-stressed calli decreased in comparison to control calli.
Furthermore, the view about a significant decrease in FW/
DW ratio in the Sesuvium stressed calli (Fig. 1d) was sup-
ported by the reduced FW/DW ratio observed in the callus
cultures of Sonnertia alba (Yasumoto et al. 1999). The
reduction in callus growth (FW and DW) under salt stress
might be a result of nutritional imbalance due to an inter-
ference of excess salt ions, with essential nutrients involved
in both uptake and translocation (Patade et al. 2008). The
reduction in water content of the cell under salt stress was
correlated with the increased cell membrane permeability of
plants growing under salt stress (Tabaei-Aghdaei et al.
2000; Farooq and Azam 2006) and consequently the injury
to the cell membrane which is measured in terms of REL.
Similarly, we observed a progressive increase in REL and
reduction in water content of Sesuvium calli with increasing
salinity levels. This was consistent with the higher values
recorded for REL under salt stress in wheat (Sairam et al.
2002; Farooq and Azam 2006) as well as in callus cultures
of sugarcane (Patade et al. 2008) and tobacco (Watad et al.
1991). The results suggest that, under salt stress conditions,
the membrane permeability of Sesuvium calli increases
producing more leakage of electrolytes outside the cell and
reduction in the growth as well as the water content of the
cell.
The low water content of the cell was apparently
adjusted by accumulating the organic osmolytes such as
proline, GB, and soluble sugars for the survival of the cells
under saline conditions. Accumulation of osmolytes under
salt stress is an indication of oxidative damage which
provides protection to cytosol from dehydration through
the development of compatible cytoplasmic osmoticum. In
the present study, significantly higher accumulation of
proline and GB as observed in the salt-stressed calli
(Fig. 2) might be an adaptive feature for the survival and
maintenance of reduced growth rate and water balance of
Sesuvium calli under salt stress conditions. Proline and GB
act as osmotica in presence of low water content and play a
major role in maintenance of osmotic balance, which was
found to be higher in salt-tolerant species than in salt-
sensitive species (Zhang et al. 2004). Zhao et al. (2009)
also demonstrated an increased accumulation of proline
and GB content in salt-stressed calli as compared to control
calli of mangrove, Thellungiella halophila. Accumulation
of TSS also imparts to the osmotic adjustment of the cell
under saline conditions (Fig. 3). Salinity-induced soluble
sugar accumulation has also been observed in P. euphra-
tica (Watanabe et al. 2000; Zhang et al. 2004). In addition
to accumulation of organic osmolytes, excess Na? accu-
mulation (Fig. 4) in the medium compensates for the loss
of K? ions, which have an added role of an osmoregulatory
Plant Cell Tiss Organ Cult (2010) 102:17–25 23
123

monovalent cation in the salt-stressed calli in comparison
to control calli. The K? outward rectifying channels
(KORCs), a class of low affinity K? channels which opens
during the depolarization of plasma membrane under salt
stress conditions, might mediate the efflux of K? and the
influx of Na? (Maathuis and Sanders 1997; Blumwald
2000) and hence alteration in the normal ratio of K?/Na?.
At the whole plant level in Sesuvium, sodium competes
with the potassium to enter inside the cell by affecting the
potassium absorption efficiency (Slama et al. 2008). The
reduced levels of K? as observed in the Sesuvium calli
under salt stress conditions may reduces osmotic pressure
brought about by K? accumulation within cells thus
affecting the cellular expansion, translocation of fixed
carbon, and turgor pressure of the cell (Szczerba et al.
2009). This might be one of the reasons for reduced growth
of the Sesuvium calli under salt stress condition.
Several studies have demonstrated that salt-tolerant spe-
cies increased their antioxidant enzyme activities and anti-
oxidant content in response to salt stress whereas salt-
sensitive species failed to do so (Yazici et al. 2007). In the
present investigation, CAT activity was significantly the
highest in control as compared to stressed calli (Fig. 5a)
suggesting the declining capacity of calli tissue to scavenge
H2O2 with increasing salt stress. The decrease in CAT
activity under increasing salinity has also been reported in
callus cultures of Suaeda nudiflora (Cherian and Reddy
2003) and Crithmum maritimum (Amor et al. 2005). APX use
ascorbate as a reductant to remove the excess H2O2 gener-
ated under salt stress. Several studies have reported a large
increase in APX activity after salinity stress in glycophytes;
however, studies on the role of APX in tolerance to salt stress
at cellular level in mangroves and other halophytes are
limited. In the present study, APX activity was found to be
low in the control calli as well as low levels of salt (100 and
200 mM NaCl) in the medium, whereas the activity was
significantly the highest in the calli treated with 400 mM salt
(Fig. 5b). The superoxide radicals formed at higher levels of
salt could be scavenged by increased SOD activity, and the
end product of this reaction, H2O2, is partially dismutated by
the increased activities of APX (Parida et al. 2004). Fur-
thermore, the increased SOD activity in stressed calli sug-
gested the view that the superoxide radicals were detoxified
efficiently. An increase in the APX activity and concomitant
increase in SOD activity is an adaptation to remove the
excess H2O2 generated due to increase in SOD activity under
excess salt stress (Jitesh et al. 2006).
Conclusions
The present results indicate that the cellular responses of
Sesuvium to salt tolerance were considerably different from
those reported at whole plant level by Messedi et al.
(2004). The RGR of calli reduced significantly as com-
pared to control calli under salt stress as revealed by
decreased water status, FW/DW ratio, damage to mem-
brane, disturbed osmotic balance, and altered antioxidant
enzyme activities. Accumulation of sodium (Na?) ions and
organic osmolytes (proline, GB, and TSS) in stressed calli
appears to be involved in osmotic adjustment under salt
stress at the cost of reduced growth rate and water content.
Moreover, the growth rate reduction observed in calli
stressed up to 200 mM NaCl was associated with lower
CAT and APX activities and significantly higher SOD
activity. Thus, the cultured cells of Sesuvium appeared to
possess better osmotic adjustment under salt stress. Further,
the results of this study illustrate the potential of using cell
and/or tissue culture for evaluation of salt tolerance at
cellular level, since responses are relatively fast, the gen-
eration times are short, and the environment is controlled.
Acknowledgment The senior author is grateful to the Department
of Atomic Energy (DAE), Board for Research in Nuclear Science
(BRNS) for financial support under the BARC-UOP collaborative
Ph.D. research programme.
References
Ahmad MSA, Javed F, Ashraf M (2007) Iso-osmotic effect of NaCl
and PEG on growth, cations and free proline accumulation in
callus tissue of two indica rice (Oryza sativa L.) genotypes. Plant
Growth Regul 53:53–63
Amor NB, Hamid KB, Debez A, Grignon C, Abdelly C (2005)
Physiological and antioxidant responses of the perennial halo-
phyte Crithmum maritimum to salinity. Plant Sci 168:889–899
Arzani A (2008) Improving salinity tolerance in crop plants: a
biotechnological view. In Vitro Cell Dev Biol Plant 44(5):373–
383
Ashraf MY (2009) Salt tolerance mechanism in halophytes from
Saudi Arabia and Egypt. Res J Agric Biol Sci 5(3):191–206
Attipali RR, Kolluru VC, Munusamy V (2004) Drought induced
responses of photosynthesis and antioxidant metabolism in
higher plants. J Plant Physiol 161:1189–1202
Bates LS, Waldren RP, Teare ID (1973) Rapid determination of free
proline for water stress studies. Plant Soil 39:205–208
Becana M, Moran JF, Iturbe-Ormaetxe I (1986) Iron-dependent
oxygen free radical generation in plants subjected to environ-
mental stress: toxicity and antioxidant protection. Plant Soil
201:137–147
Blumwald E (2000) Sodium transport and salt tolerance in plants.
Curr Opin Cell Biol 12:431–434
Bohnert HJ, Jensen RG (1996) Strategies for engineering water stress
tolerance in plants. Trends Biotechnol 14:89–97
Borsani O, Valpuesta V, Botella MA (2003) Developing salt tolerant
plants in a new century: a molecular biology approach. Plant
Cell Tissue Organ Cult 73:101–115
Bradford MM (1976) A rapid and sensitive method for the
quantitation of microgram quantities of protein utilizing the
principle of protein-dye binding. Analyt Biochem 72:248–254
Cakmak I, Marschner H (1992) Magnesium deficiency and high light
intensity enhance activities of superoxide dismutase, ascorbate
24 Plant Cell Tiss Organ Cult (2010) 102:17–25
123

peroxidase and glutathione reductase in bean leaves. Plant
Physiol 98:1222–1227
Chamandoosti F (2007) Effect of sodium chloride on establishment of
callus and organogenesis in Brassica napus L. Pak J Biol Sci
10(21):3880–3884
Cherian S, Reddy MP (2003) Evaluation of NaCl tolerance in the
callus cultures of Suaeda nudiflora Moq. Biol Plant 46:193–198
Farooq S, Azam F (2006) The use of cell membrane stability (CMS)
technique to screen for salt tolerant wheat varieties. J Plant
Physiol 163:629–637
Flowers TJ, Colmer TD (2008) Salinity tolerance in halophytes. New
Phytol 179:945–963
Grieve CM, Grattan SR (1983) Rapid assay for determination of
water soluble quaternary ammonium compounds. Plant Soil
70:303–307
Hasegawa PM, Bressan RA, Zhu JK, Bohnert HJ (2000) Plant cellular
and molecular responses to high salinity. Ann Rev Plant Physiol
Plant Mol Biol 51:463–499
Jitesh MN, Prashant SR, Sivaprakash KR, Parida AK (2006)
Antioxidative response mechanism in halophytes: their role in
stress defence. J Genet 85(3):237–254
Khayri JMA (2002) Growth, proline accumulation, and ion content in
sodium chloride stressed callus of date palm. In Vitro Cell Dev
Biol Plant 38:79–82
Lokhande VH, Nikam TD, Patade VY, Suprasanna P (2009a)
Morphological and molecular diversity analysis among the
Indian clones of Sesuvium portulacastrum L. Genet Resour Crop
Evol 56:705–717
Lokhande VH, Nikam TD, Suprasanna P (2009b) Sesuvium portul-acastrum (L.) L. a promising halophyte: cultivation, utilization
and distribution in India. Genet Resour Crop Evol 56:741–747
Maathuis FJM, Sanders D (1997) Regulation of K absorption in plant
root cells by external K: interplay of different channel in maize
root stellar cells. J Exp Bot 48:839–846
May MJ, Vernoux T, Leaver C, Van Montagu M, Inze D (1998)
Glutathione homeostasis in plants: implications for environmen-
tal sensing and plant development. J Exp Bot 49:649–667
Menzel U, Leith H (1999) Annex 4: halophyte database vers. 2. In:
Leith H, Moschenko M, Lohmann M, Koyro HW, Hamdy A
(eds) Halophyte uses in different climates, 1. Ecological and
ecophysiological studies. Progress in Biotechnology 13. Back-
huys, Leiden
Messedi D, Labidi N, Grignon C, Abdelly C (2004) Limits imposed
by salt to the growth of the halophyte Sesuvium portulacastrum.
J Plant Nutr Soil Sci 167:720–725
Munns R (2002) Comparative physiology of salt and water stress.
Plant Cell Environ 25:239–250
Munns R, Tester M (2008) Mechanisms of salt tolerance. Annu Rev
Plant Biol 59:651–681
Murashige T, Skoog F (1962) A revised medium for rapid growth and
bioassays with tobacco tissue cultures. Plant Physiol 15:473–497
Nakano Y, Asada K (1981) Hydrogen peroxide is scavenged by
ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell
Physiol 22:867–880
Parida AK, Das AB, Mohanty P (2004) Defence potentials to NaCl in
a mangrove, Bruguiera parviflora: differential changes of
isoforms of some antioxidative enzymes. J Plant Physiol
161:531–542
Patade VY, Suprasanna P, Bapat VA (2008) Effects of salt stress in
relation to osmotic adjustment on sugarcane (Saccharum offici-narum L.) callus cultures. Plant Growth Regul 55(3):169–173
Sairam RK, Rao KV, Srivastava GC (2002) Differential response of
wheat genotypes to longterm salinity stress in relation to
oxidative stress, antioxidant activity and osmolyte concentration.
Plant Sci 163:1037–1046
Slama I, Ghnaya T, Savoure A, Abdelly C (2008) Combined effects
of long-term salinity and soil drying on growth, water relations,
nutrient status and proline accumulation of Sesuvium portulaca-strum. C R Biol 331:442–451
Sullivan CY (1972) Mechanism of heat and drought resistance in
grain sorghum and methods of measurement. In: Rao NGP,
House LR (eds) Sorghum in the seventies. Oxford and IBH, New
Delhi, pp 247–264
Szczerba MW, Britto DT, Kronzucker HJ (2009) K? transport in
plants: physiology and molecular biology. J Plant Physiol
166:447–466
Tabaei-Aghdaei S, Harrison P, Pearee RS (2000) Expression of
dehydration-stress related genes in crown of wheat, grass species
having contrasting acclimation to salt, cold and drought. Plant
Cell Environ 23:561–571
Tester M, Davenport R (2003) Na? tolerance and Na? transport in
higher plants. Ann Bot 91:503–527
Tonon G, Kevers C, Faivre-Rampant O, Grazianil M, Gaspar T(2004)
Effect of NaCl and mannitol iso-osmotic stresses onproline and
free polyamine levels in embryogenic Fraxinusangustifoliacallus. J Plant Physiol 161: 701–708
Vijayan K, Chakraborti SP, Ghosh PD (2003) In vitro screening of
mulberry for salinity tolerance. Plant Cell Rep 22:350–357
Watad AEA, Reuveni M, Bressan RA, Hasegawa PM (1991)
Enhanced net K? uptake capacity of NaCl-adapted cells. Plant
Physiol 95:1265–1269
Watanabe S, Kojima K, Ide Y, Sasaki S (2000) Effects of saline and
osmotic stress on proline and sugar accumulation in Populuseuphratica in vitro. Plant Cell Tissue Organ Cult 63:199–206
Yasumoto E, Adachi K, Kato M, Sano H, Sasamoto H, Baba S,
Ashihara H (1999) Uptake of inorganic ions and compatible
solutes in cultured mangrove cells during salt stress. In Vitro
Cell Dev Biol Plant 35:82–85
Yazici I, Turkan I, Sekmen AH, Demiral T (2007) Salinity tolerance
of purslane (Portulaca oleracea L.) is achieved by enhanced
antioxidant system, lower level of lipid peroxidation and proline
accumulation. Environ Exp Bot 61:49–57
Yokoi S, Bressan RA, Hasegawa PM (2002) The Japan International
Centre for Agricultural Sciences (JIRCAS) Working Report No.
23. In: Iwanaga M (ed) Genetic engineering of crop plants for
abiotic stress. Salt stress tolerance of plants. Japan International
Centre for Agricultural Sciences, Tsukuba, pp 25–33
Zhang F, Yang YL, He WL, Zhao X, Zhang LX (2004) Effects of
salinity on growth and compatible solutes of callus induced from
Populus euphratica. In vitro Cell Dev Biol Plant 40:491–494
Zhao X, Tan HJ, Liu YB, Li XR, Chen GX (2009) Effect of salt stress
on growth and osmotic regulation in Thellungiella and Arabis-opsis callus. Plant Cell Tissue Organ Cult 98(1):97–103
Plant Cell Tiss Organ Cult (2010) 102:17–25 25
123




















