
Arachidonic acid is a preferred acetyl donor among fatty acids in the acetylation of p-aminobenzoic...
10
BiochimicaL et Biophysics &ta ELSEVIER Biochimica et Biophysics Acta 1257 (1995) 157-166 Arachidonic acid is a preferred acetyl donor among fatty acids in the acetylation of p-aminobenzoic acid by human lymphoid cells Bruno Barbieri a,*, Nikos Papadogiannakis a, Peter Eneroth b, Lars B. Olding a a Department of Immunology, Microbiology, Pathology and Infectious Diseases, Division of Pathology F42, Karolinska Institutet, Huddinge (/nicer@ Hospital, S-141 86 Huddinge, Sweden b Clinical Research Center, Unit for Applied Biochemistry, Nocum, Karolinska Instituter, S-141 86 Huddinge, Sweden Received 27 December 1994; revised 16 March 1995; accepted 20 March 1995 Abstract We have previously reported that human lymphoid cells, such as peripheral blood mononuclear leukocytes (PBML) and the T-cell leukemia line Jurcat, synthesize p-acetamidobenzoic acid from p-aminobenzoic acid (PABA) and a two carbon fragment from arachidonic acid (AA), conceivably derived from P-oxidation. Here we demonstrate that AA is a preferred substrate in this acetylation reaction over other common fatty acids such as palmitic (PA), oleic, linoleic or linolenic. This was unexpected because AA is not considered as a fuel fatty acid. In Jurcat cells, AA is also preferred as a substrate for P-oxidation over PA. In contrast, in PBML, PA was clearly preferred as substrate for P-oxidation over AA, in accordance with previous observations. The difference between Jurcat cells and PBML was not dependent on culture conditions, because phytohemagglutinin and interleukin-2 activated PBML, kept in culture, showed the same PA preference as freshly prepared non-activated PBML. Furthermore, we observed differences between Jurcat cells and PBML in their relative content of fatty acids and in the incorporation of PA and AA into triacylglycerols and phospholipids. Taken together, our results show differences in P-oxidation between Jurcat cells and PBML, and suggest the involvement of peroxisomal, besides mitochondrial, P-oxidation, in the acetylation of PABA with fatty acids as acetyl donors. Keywords: p-Aminobenzoic acid; Arachidonic acid: Fatty acid; P-Oxidation; Lymphoid cell; Jurcat cell 1. Introduction Except for the role of p-aminobenzoic acid (PABA) in folate formation, no specific function has been ascribed to PABA, although it is a constituent of most culture media and is known to improve maintenance of cells in culture [l]. Some decades ago PABA was extensively studied for its biological significance. In some reports PABA showed to have chromotricial effect on gray hair [2]. Other reports Abbreviations: PABA, p-aminobenzoic acid: PACBA, p-acetamido- benzoic acid; AA, arachidonic acid; PBML, peripheral blood mononu- clear leukocytes; FA, fatty acids; PG. prostaglandin; PHA, phytohemag- glutinin; MS, mass spectrometry; PA, palmitic acid; FCS, fetal calf serum: AC-PBML, activated and cultured PBML; OA, oleic acid; LA, linoleic acid; LEA, linolenic acid; TLC, thin-layer chromatography; HPLC, high-pressure liquid chromatography; MeOH, methanol; TG, tri- acylglycerols; PL, phospholipids; PBS, phosphate-buffered saline; PUFA, polyunsaturated fatty acids. * Corresponding author. Fax: +46 8 7795520. showed that PABA could be useful in the treatment of vitiligo, leukemia, rheumatic fever and in ricketsial dis- eases [2]. The mechanisms behind the effects ascribed to PABA were not always understood and statements were also sometimes conflicting. When the role of PABA in folate formation was established, the interest for PABA itself diminished. However, today, esters of PABA are still used as local anesthetics and the acetylated form of PABA, p-acetamidobenzoic acid (PACBA), is one of the compo- nents in the antiviral and immunomodulatory drug Inosine Pranobex [3]. When investigating the metabolism of [l- 14C]arachidonic acid (AA) in neonatal and maternal pe- ripheral blood mononuclear leukocytes (PBML) we found that the cells converted AA to hydroxy fatty acids (FA), prostaglandin (PG) E, and to small amounts of PGF,, and thromboxane B, [4]. However, the major radioactive prod- uct was a substance that was not a known eicosanoid, because cyclooxygenase and lipoxygenase inhibitors did 0005-2760/95/$09.50 0 1995 Elsevier Science B.V. All rights reserved SSDI 0005-2760(95)00070-4
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Arachidonic acid is a preferred acetyl donor among fatty acids in the acetylation of p-aminobenzoic...

BiochimicaL et Biophysics &ta
ELSEVIER Biochimica et Biophysics Acta 1257 (1995) 157-166
Arachidonic acid is a preferred acetyl donor among fatty acids in the acetylation of p-aminobenzoic acid by human lymphoid cells
Bruno Barbieri a, * , Nikos Papadogiannakis a, Peter Eneroth b, Lars B. Olding a a Department of Immunology, Microbiology, Pathology and Infectious Diseases, Division of Pathology F42, Karolinska Institutet, Huddinge (/nicer@
Hospital, S-141 86 Huddinge, Sweden b Clinical Research Center, Unit for Applied Biochemistry, Nocum, Karolinska Instituter, S-141 86 Huddinge, Sweden
Received 27 December 1994; revised 16 March 1995; accepted 20 March 1995
Abstract
We have previously reported that human lymphoid cells, such as peripheral blood mononuclear leukocytes (PBML) and the T-cell leukemia line Jurcat, synthesize p-acetamidobenzoic acid from p-aminobenzoic acid (PABA) and a two carbon fragment from arachidonic acid (AA), conceivably derived from P-oxidation. Here we demonstrate that AA is a preferred substrate in this acetylation reaction over other common fatty acids such as palmitic (PA), oleic, linoleic or linolenic. This was unexpected because AA is not considered as a fuel fatty acid. In Jurcat cells, AA is also preferred as a substrate for P-oxidation over PA. In contrast, in PBML, PA was clearly preferred as substrate for P-oxidation over AA, in accordance with previous observations. The difference between Jurcat cells and PBML was not dependent on culture conditions, because phytohemagglutinin and interleukin-2 activated PBML, kept in culture, showed the same PA preference as freshly prepared non-activated PBML. Furthermore, we observed differences between Jurcat cells and PBML in their relative content of fatty acids and in the incorporation of PA and AA into triacylglycerols and phospholipids. Taken together, our results show differences in P-oxidation between Jurcat cells and PBML, and suggest the involvement of peroxisomal, besides mitochondrial, P-oxidation, in the acetylation of PABA with fatty acids as acetyl donors.
Keywords: p-Aminobenzoic acid; Arachidonic acid: Fatty acid; P-Oxidation; Lymphoid cell; Jurcat cell
1. Introduction
Except for the role of p-aminobenzoic acid (PABA) in folate formation, no specific function has been ascribed to PABA, although it is a constituent of most culture media and is known to improve maintenance of cells in culture [l]. Some decades ago PABA was extensively studied for its biological significance. In some reports PABA showed to have chromotricial effect on gray hair [2]. Other reports
Abbreviations: PABA, p-aminobenzoic acid: PACBA, p-acetamido- benzoic acid; AA, arachidonic acid; PBML, peripheral blood mononu- clear leukocytes; FA, fatty acids; PG. prostaglandin; PHA, phytohemag- glutinin; MS, mass spectrometry; PA, palmitic acid; FCS, fetal calf serum: AC-PBML, activated and cultured PBML; OA, oleic acid; LA, linoleic acid; LEA, linolenic acid; TLC, thin-layer chromatography; HPLC, high-pressure liquid chromatography; MeOH, methanol; TG, tri- acylglycerols; PL, phospholipids; PBS, phosphate-buffered saline; PUFA, polyunsaturated fatty acids.
* Corresponding author. Fax: +46 8 7795520.
showed that PABA could be useful in the treatment of vitiligo, leukemia, rheumatic fever and in ricketsial dis- eases [2].
The mechanisms behind the effects ascribed to PABA were not always understood and statements were also sometimes conflicting. When the role of PABA in folate formation was established, the interest for PABA itself diminished. However, today, esters of PABA are still used as local anesthetics and the acetylated form of PABA, p-acetamidobenzoic acid (PACBA), is one of the compo- nents in the antiviral and immunomodulatory drug Inosine Pranobex [3].
When investigating the metabolism of [l- 14C]arachidonic acid (AA) in neonatal and maternal pe- ripheral blood mononuclear leukocytes (PBML) we found that the cells converted AA to hydroxy fatty acids (FA), prostaglandin (PG) E, and to small amounts of PGF,, and thromboxane B, [4]. However, the major radioactive prod- uct was a substance that was not a known eicosanoid, because cyclooxygenase and lipoxygenase inhibitors did
0005-2760/95/$09.50 0 1995 Elsevier Science B.V. All rights reserved SSDI 0005-2760(95)00070-4

158 Bruno Barbieri et ul. /Biochimicu rt Biophvsicrr Acttr 1257 (19951 157-166
not affect its production. Later we found that a variety of 2.2. Biosynthesis of PACBA with various FA as acetyl cells and cell lines produced the substance [5]. donors
The unidentified metabolite induced an increase in the incorporation of [ 3H]thymidine in phytohemagglutinin (PHA) activated PBML from several recently delivered mothers with a low response to PHA and in PBML from immunodeficient infants [6]. In contrast, no effect was seen on the PHA induced proliferation of PBML from healthy adult blood donors.
To avoid the interference of FA present in the FCS, we substituted FCS with a synthetic serum consisting of 0.75 g bovine albumin, 5 mg insulin and 10 mg transferrin per liter RPM1 1640.
Mass spectrometry (MS) and nuclear magnetic reso- nance spectroscopy revealed that the substance was PACBA, formed through acetylation of PABA by a two- carbon fragment from [ I- 14C]AA [5]. Because acetyl-CoA is known to be the substrate for N-acetylation AA must have underwent P-oxidation before acetylation of PABA. Accordingly every FA that undergoes P-oxidation should be a feasible substrate in the acetylation reaction. How- ever, we will here present data indicating that PBML and the T-cell leukemia line Jurcat prefer AA as a substrate for the acetylation over other common FA, such as palmitic acid (PA). The AA preference follows the respective rate of P-oxidation only for the Jurcat cells. In contrast PBML oxidate PA to a higher degree than AA, but still they prefer AA as an acetyl donor in the acetylation of PABA.
50. IO6 PBML were suspended in 25 ml of this medium with 0.05 mg/ml antibiotic (Gentamycin, Gibco) and each [ 1- ‘jC]FA was added to a concentration of 2.16 PM. The FA investigated were PA (57 Ci/mol), oleic acid (OA) (56 Ci/mol), linoleic acid (LA) (53 Ci/mol), linolenic acid (LEA) (52 Ci/mol), and AA (53 Ci/mol), (All FA from NEN Products, Boston, MA). After 18 h incubation, the cell viability was checked according to the Trypan blue exclusion test, and the cell suspension was centrifuged (430 X g for 15 min). The supematant was collected for determination of PACBA and the radioactivity in the cell pellets counted in a LKB 1214 Rackbeta liquid scintilla- tion counter (Wallac Oy, Turku, Finland).
2.3. AA compared to PA as substrate for acetylation qj PABA
2. Materials and methods
2.1. Sources of cells and culture conditions
In order to examine the different specificities of FA as acetyl donors, we tested AA and PA further. These experi- ments were performed with Jurcat cells and with PBML from six donors. The concentration of PABA in the growth medium was increased to 22.0 PM. The regular, commer- cial RPM1 1640 contains PABA at a concentration of 7.3
PM. Leukocyte enriched buffy coats originated from the
blood of healthy donors at the Huddinge University Hospi- tal. PBML were isolated by means of Ficoll-Isopaque (Pharmacia Biotechnology, Uppsala, Sweden) gradient centrifugation [4]. In this context only unsorted PBML were used.
The Jurcat cells and PBML were incubated in 25 ml RPMI, supplemented with PABA and with the synthetic serum, at a cell concentration of 0.8 and 1.5 . 106/ml, respectively.
In some experiments PBML were brought into a state of continuous proliferation by using PHA-P (Sigma, St Louis, MO) and human interleukin-2 (Lymphocult; Biotest Diag- nostics, Dreieich, Germany) according to the manufac- turer’s protocol (Biotest). Briefly, PBML, 1 . 106/ml, were activated with 1 pg/ml PHA-P in RPM1 1640 growth medium (Gibco, Life Technologies, Paisley, Scotland) with 7.5% fetal calf serum (FCS; Gibco). After 48 h the medium was changed to a medium containing interleukin-2, 20 U/ml, instead of PHA-P. The cells were then cultured in this medium. After 1 week the doubling time for the activated and cultured PBML (AC-PBML) was approx. 26 h. The cells were used in continued experiments at the eighth day of culture if not otherwise indicated.
Jurcat cells and PBML were incubated for 18 h at the [ 1- ‘“C]FA concentrations 2.16 PM (0.063 &i/ml), 6.50 PM (0.063 pCi/ml) and 21.5 PM (0.063 pCi/ml). Unlabeled FA used to dilute the labeled FA was purchased from Sigma. The total amount of PACBA produced in the cultures was isolated and the radioactive PACBA quanti- fied. The cell viability was checked according to the Trypan blue exclusion test.
2.4. Purification of PACBA
The Jurcat cells were obtained from the American Type Culture Collection (ATCC, Rockville, MD), and were cultured in RPM1 1640 medium with 7.5% FCS.
Cell culturing and incubations were performed at 37” C in humidified air with 5% CO,.
Extraction and purification of PACBA was performed as previously described [5], with slight modifications; Su- pernatants from the cell cultures were acidified to pH 3 with formic acid and subsequently extracted three times with an equal amount of ethyl acetate. The organic phase was washed neutral, evaporated and the residue was ap- plied to an open column, 10 X 200 mm, packed with Silicar cc-4 (Mallinkrodt, Paris, KY). The column was eluted with 35 ml hexane/ethyl acetate (9: 1, v/v), 50 ml hexane/ethyl acetate (4:1, v/v) and 40 ml ethyl

Bruno Barbieri et al. / Biochimica et Biophysics Acta 1257 (1995) 157-166 159
acetate/formic acid (20: 1, v/v>. The latter fraction con- taining PACBA was purified by thin layer chromatography (TLC) using the organic phase from ethyl acetate/water/isooctane/acetic acid (11: 10:6:2, v/v). PACBA labeled with [‘“Cl in the carbonyl carbon of the amide was synthetically prepared as previously described [5]. The labeled PACBA and the [ 1- 14ClFA were used as standards. The TLC plate was then scanned with a Berthold LB 2723 thin-layer scanner (Berthold, Berlin, Germany) for the radioactive spot corresponding to PACBA. This material was removed, extracted with ethyl acetate, and quantified by high pressure liquid chromatography (HPLC). The column used was a Supelco C-18, 5 pm (4.6 X 250 mm), and the mobile phase consisted of water/methanol (MeOH)/acetic acid (60:40:0.2, v/v>. The flow rate was constantly 1 ml/min. UV-light absorption was monitored with a LDC/Milton Roy N Series UV detector (LDC Analytical, Riviera Beach, FL) at the absorption maximum 278 nm, and the radioactivity was measured synchronously with a Flow-One A-200 detector (Packard Instrument Company, Meriden, CT). UV-light absorption and radioac- tivity were confirmed as arising from a single substance with the retention volume of 8.05 ml. The identity and purity of the substance in this peak was checked by means of electron impact MS on a Nermag RlO- 10H quadrupole mass analyzer (Delsi Nermag Instruments, Ruil-Malmai- son, France) consisting of a desorption probe, ionization source, quadrupole filter and ion detector. The sample from HPLC was evaporated and dissolved in MeOH and applied on the tungsten filament of the desorption probe with a syringe. Spectra were obtained at 70 eV, and at an ion source temperature of 183” C.
2.5. Distribution of AA and PA between triacylglycerols (TG) and phospholipids fPL)
The cells were incubated at a FA concentration of 2.16 PM (0.063 pCi/ml) for 2 or 18 h. The experiments were performed in 10 ml RPM1 1640 (with the synthetic serum), and the cell number was 15. lo6 and 8. lo6 for PBML and Jurcat cells respectively. After centrifugation, and three washes in fresh medium, the cells from each experi- ment were sonicated in 0.6 ml water for 4 X 10 s with a W-375 microtip sonicator (Heat Systems Ultrasonic, New York, NY). 1.4 ml methanol (MeOH) was added and the homogenate mixture thereafter extracted with 3 X 2 ml petroleum ether (boiling range 40-60” C). After centrifuga- tion at 3000 X g for 30 min the radioactivity in the two organic phases and the protein pellet, from the MeOH fraction, was counted in the liquid scintillation counter.
2.6. Incubation procedure for measurement of FA oxida- tion
CO, produced from the oxidation of l-‘4C-labeled FA was quantified by a modification of two methods previ- ously developed for measurement of the FA P-oxidation
[7,8]. Cells were incubated in 20 ml pots (bottom area, 3.2 cm’) with a Teflon cap and a center well. Liberated 14C02 was trapped in a piece of filter paper soaked with 5 ~1 1 M NaOH. In order to facilitate the removal from the center well, the filter paper was placed in a small container prepared by cutting off the top of a plastic centrifuge tube.
2 ml phosphate buffered saline (PBS) containing 2 . IO6 PBML and [1-14C]FA at a concentration of 6.5 FM (0.36 @Zi/ml) was added to the outer compartment of the pot and the pot was then closed with the Teflon cap.
After incubation for 4 h, 50 ~1 4 M NaOH was added to the filter, and 250 ~1 0.5 M of H,SO, was injected into the cell suspension through the Teflon seal. The pots were left on ice for 3 h, and the contents of the center well were then transferred to a scintillation vial containing 1.5 ml water and left on a shaker at room temperature for 2 h. 15 ml scintillation cocktail was added and the radioactivity measured in the liquid scintillation counter.
Experiments were also carried out to investigate the oxidation of PA and AA when the incubation time was extended to 18 h. In each of these experiments we used; 8 . lo6 PBML, 3 . lo6 AC-PBML and 4. lo6 Jurcat cells. The cells were incubated in 4 ml RPM1 1640 supple- mented with the synthetic serum (see above).
Before the incubation the pots with the cell suspension were left open in the incubator for 2 h to condition the medium, and the [1- 14C]FA was then added. After the 18 h incubation the cell viability was checked, according to the Trypan blue exclusion test.
Blank incubations without cells were included in all experiments to measure the spontaneous decarboxylation of FA. The spontaneous decarboxylation was then sub- tracted from the radioactivity measured in each sample of cells.
2.7. Extraction and methylation of lipids in cells and FCS
10-20. lo6 PBML, AC-PBML, and Jurcat cells were washed three times in PBS, resuspended in 1 ml of dis- tilled water, and disrupted by five freeze-thaw cycles. In parallel, 100 ~1 of FCS was suspended in 0.9 ml distilled water. The total content of lipids in cells and FCS was extracted according to a modified method of Folch [9]. 2.0 ml MeOH, 4.0 ml chloroform and 0.2 ml 0.60 M KC1 in water were added to the cell suspension and the mixture was mixed for 2 min. After centrifugation for 5 min, at 300 X g, the chloroform was collected and both the upper phase (MeOH/water) and the interphase (containing pro- tein like material) was re-extracted with 2.0 ml chloroform containing 2 pg/ml 2,6-bis( 1,l -dimethylethyl)-4-methyl- phenol (Sigma). The total chloroform phase was dried with argon gas in a screw cup equipped, gas-tight tube. To the remaining lipids were added 0.8 ml 0.5 M NaOH in MeOH and the mixture was heated for 5 min at 100” C. After this saponification the product was allowed to cool prior to addition of 1 ml 14% BF,-MeOH reagent (Sigma).
160 Bruno Barbieri et al. /Biochimica et Biophwica Acra 1257 (1995) 157-166
The tubes were then closed firmly and heated for 3 min at 100” C. After cooling, 0.5 ml water and 2.0 ml petroleum ether was added, and the mixture mixed for 1 min. After centrifugation (5 min, at 300 X g) the petroleum ether was collected, and the extraction repeated with further 2.0 ml petroleum ether. The solvent was evaporated by a stream of argon gas and the remaining methyl esters re-dissolved in 100 ~1 hexane for quantitation using gas chromatog- raphy-MS.
2.8. Gas chromatography-MS
For gas chromatographic separation of FA methyl es- ters, a Di 200 gas chromatograph was used (Delsi Instru- ments, Suresnes, France). The column was a Sp 2330, 25 m X 0.25 mm id, (Supelco, Bellefonte, PA) and the split ratio 1: 13. The injector temperature was 250” C, and the oven programmed for an initial temperature of 120” C, for 1 min, and a progression rate of 3 C”/min up to 220” C. The gas chromatograph was connected to the Nermag RIO- 10H for detection and recognition of the FA methyl esters. Spectra were obtained at 70 eV and at the ion source temperature 260” C. To compensate for different ionization efficiency for the FA methyl esters a standard containing 25 or 50 ng/ml of each methylated FA, from PA (16:O) to 22:6(n - 31, was used to calculate the correc- tion factors for the different FA. (The denotation intro-
18
-m-a. -.- b.
-A-c. -V-d.
I * /
/& . :A’
i,::ji-i / ./
tC:_-,// . I 1
PA OA LA LEA AA
[1-14C] FA used as substrate
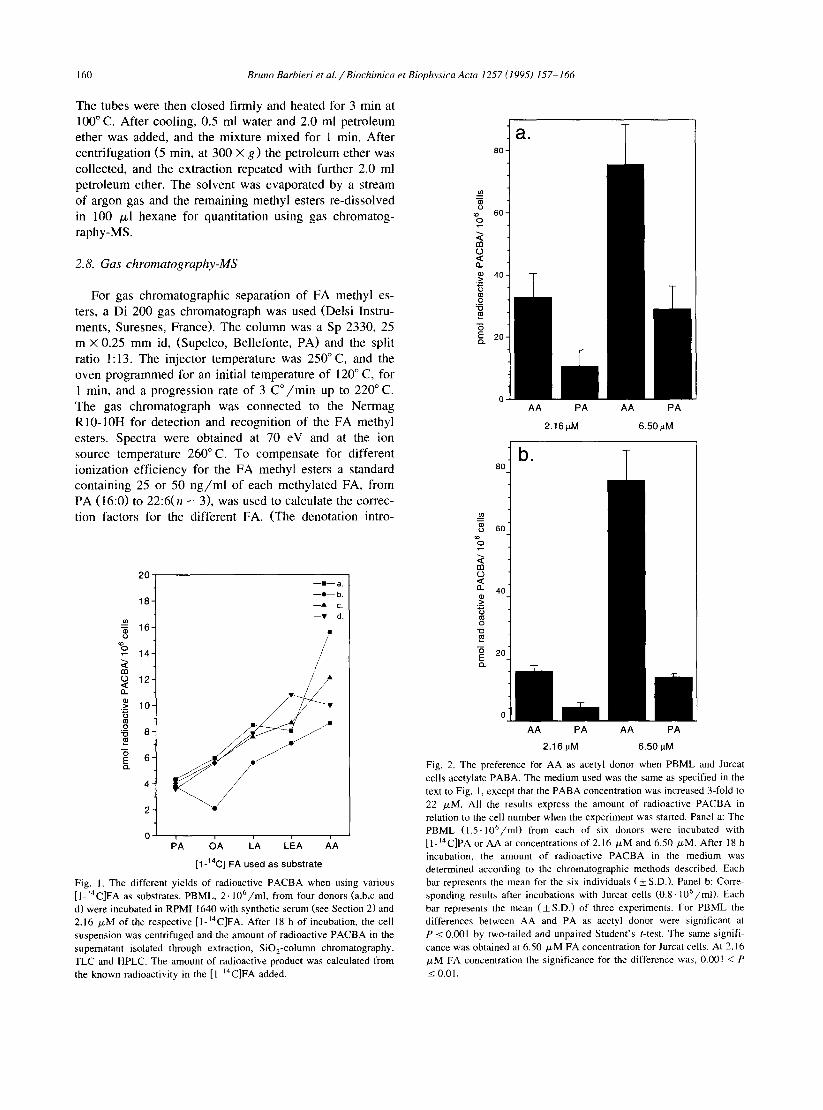
Fig. I. The different yields of radioactive PACBA when using various [ I-‘4C]FA as substrates. PBML, 2. 106/ml, from four donors (a,b,c and d) were incubated in RPM1 1640 with synthetic serum (see Section 2) and 2.16 FM of the respective [ l-‘4C]FA. After 18 h of incubation, the cell suspension was centrifuged and the amount of radioactive PACBA in the supernatant isolated through extraction, SiO,-column chromatography, TLC and HPLC. The amount of radioactive product was calculated from the known radioactivity in the [ l-‘4C]FA added.
a.
AA PA AA PA
2.16pM 6.50 FM
0 il AA PA AA PA
2.16 pM 6.50 pM
J
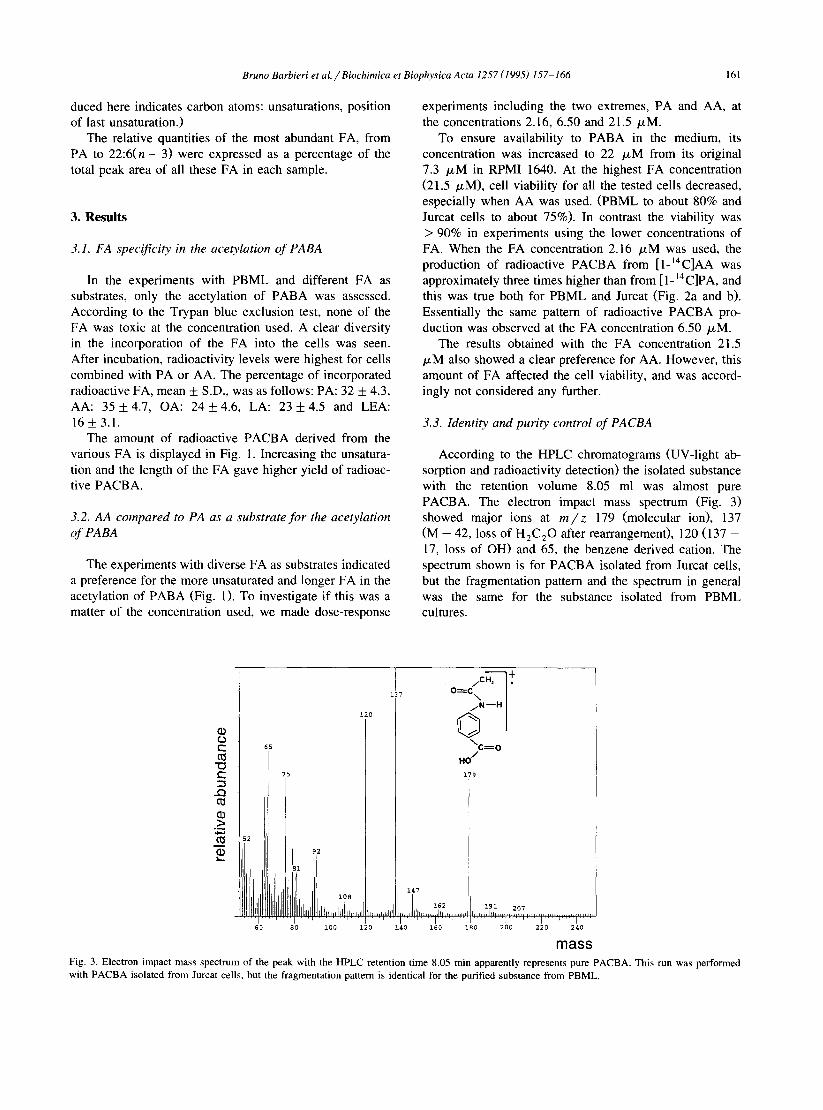
Fig. 2. The preference for AA as acetyl donor when PBML and Jurcat cells acetylate PABA. The medium used was the same as specified in the text to Fig. 1, except that the PABA concentration was increased 3-fold to 22 PM. All the results express the amount of radioactive PACBA in relation to the cell number when the experiment was started. Panel a: The PBML (1.5. lO’/ml) from each of six donors were incubated with [l-‘4C]PA or AA at concentrations of 2.16 FM and 6.50 FM. After 18 h incubation, the amount of radioactive PACBA in the medium was determined according to the chromatographic methods described. Each bar represents the mean for the six individuals (iS.D.). Panel b: Corre- sponding results after incubations with Jurcat cells (0.8. IO’/ml). Each bar represents the mean (f S.D.) of three experiments. For PBML the differences between AA and PA as acetyl donor were significant at P I 0.001 by two-tailed and unpaired Student’s t-test. The same signifi- cance was obtained at 6.50 /.LM FA concentration for Jurcat cells. At 2.16 PM FA concentration the significance for the difference was, 0.001 < P IO.01.
Bruno Barbieri et aL/Biochimica et Biophysics Acta 1257 (1995) 157-166 161
duced here indicates carbon atoms: unsaturations, position experiments including the two extremes, PA and AA, at of last unsaturation.) the concentrations 2.16, 6.50 and 21.5 PM.
The relative quantities of the most abundant FA, from PA to 22:6(n - 3) were expressed as a percentage of the total peak area of all these FA in each sample.
To ensure availability to PABA in the medium, its concentration was increased to 22 PM from its original 7.3 PM in RPM1 1640. At the highest FA concentration (21.5 PM), cell viability for all the tested cells decreased, especially when AA was used. (PBML to about 80% and Jurcat cells to about 75%). In contrast the viability was > 90% in experiments using the lower concentrations of FA. When the FA concentration 2.16 PM was used, the production of radioactive PACBA from [ l-14C]AA was approximately three times higher than from [ 1- I4 C]PA, and this was true both for PBML and Jurcat (Fig. 2a and b). Essentially the same pattern of radioactive PACBA pro- duction was observed at the FA concentration 6.50 PM.
3. Results
3.1. FA specificity in the acetylation of PABA
In the experiments with PBML and different FA as substrates, only the acetylation of PABA was assessed. According to the Trypan blue exclusion test, none of the FA was toxic at the concentration used. A clear diversity in the incorporation of the FA into the cells was seen. After incubation, radioactivity levels were highest for cells combined with PA or AA. The percentage of incorporated radioactive FA, mean f S.D., was as follows: PA: 32 f 4.3, AA: 35 + 4.7, OA: 24 + 4.6, LA: 23 * 4.5 and LEA: 16 f 3.1.
The amount of radioactive PACBA derived from the various FA is displayed in Fig. 1. Increasing the unsatura- tion and the length of the FA gave higher yield of radioac- tive PACBA.
3.2. AA compared to PA as a substrate for the acetylation of PABA
The experiments with diverse FA as substrates indicated a preference for the more unsaturated and longer FA in the acetylation of PABA (Fig. 1). To investigate if this was a matter of the concentration used, we made dose-response
T
L31
The results obtained with the FA concentration 21.5 PM also showed a clear preference for AA. However, this amount of FA affected the cell viability, and was accord- ingly not considered any further.
3.3. Identity and purity control of PACBA
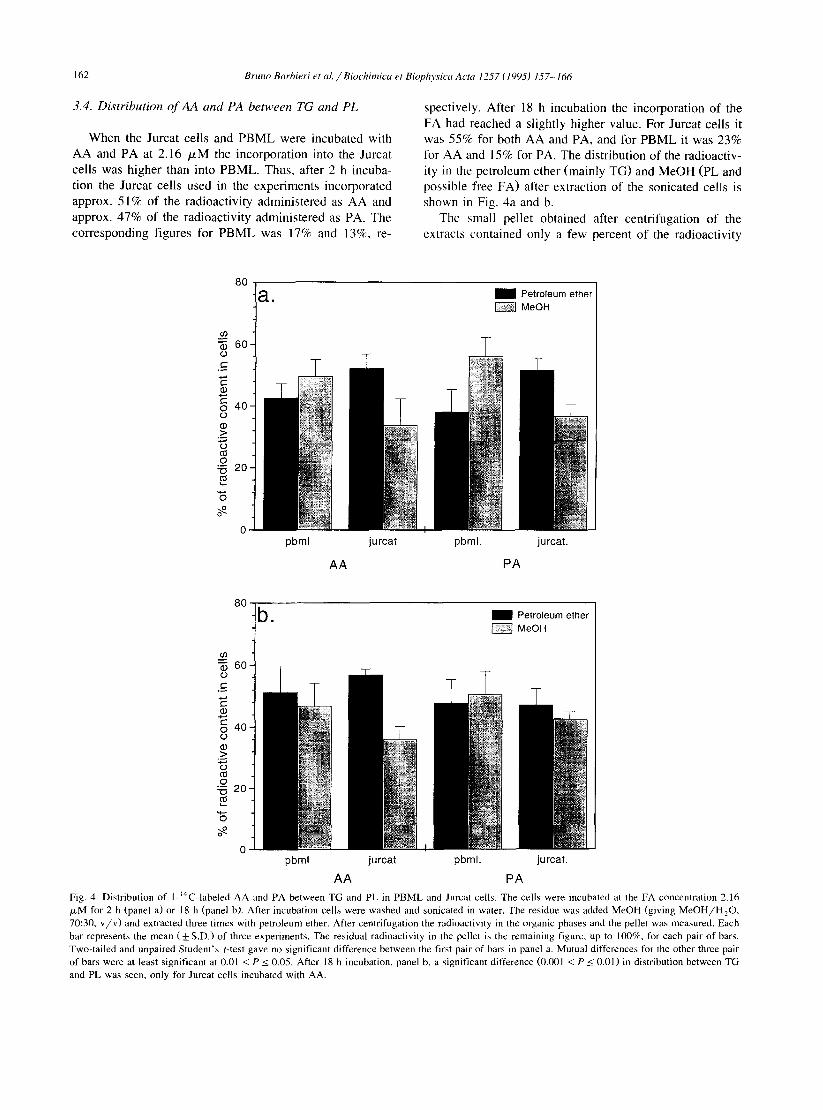
According to the HPLC chromatograms WV-light ab- sorption and radioactivity detection) the isolated substance with the retention volume 8.05 ml was almost pure PACBA. The electron impact mass spectrum (Fig. 3) showed major ions at m/z 179 (molecular ion), 137 (M - 42, loss of H,C,O after rearrangement), 120 (137 - 17, loss of OH) and 65, the benzene derived cation. The spectrum shown is for PACBA isolated from Jurcat cells, but the fragmentation pattern and the spectrum in general was the same for the substance isolated from PBML cultures.
,w . 1 t
o=c \
,N-H
\ c=o HO
/
mass Fig. 3. Electron impact mass spectrum of the peak with the HPLC retention time 8.05 min apparently represents pure PACBA. This run was performed with PACBA isolated from Jurcat cells, but the fragmentation pattern is identical for the purified substance from PBML.
162 Bruno Bnrbirri et al./Biochimica ef Biophysics Actn 1257 (19951 157-166
3.4. Distribution of AA and PA between TG and PL
When the Jurcat cells and PBML were incubated with AA and PA at 2.16 PM the incorporation into the Jurcat cells was higher than into PBML. Thus, after 2 h incuba- tion the Jurcat cells used in the experiments incorporated approx. 5 1% of the radioactivity administered as AA and approx. 47% of the radioactivity administered as PA. The corresponding figures for PBML was 17% and 13%, re-
spectively. After 18 h incubation the incorporation of the FA had reached a slightly higher value. For Jurcat cells it was 55% for both AA and PA, and for PBML it was 23% for AA and 15% for PA. The distribution of the radioactiv- ity in the petroleum ether (mainly TG) and MeOH (PL and possible free FA) after extraction of the sonicated cells is shown in Fig. 4a and b.
The small pellet obtained after centrifugation of the extracts contained only a few percent of the radioactivity
-_
1 a. m Petroleum ether m MeOH
c/l
5 60
c .-
z 75 8 40
a, .g
2
g 20 CJ
5 9 0
0 pbml jurcat pbml. jurcat.
AA PA
80 -b. m Petroleum ether
m MeOH
In
5 60 0 t .-
2 2 3 40
P .s ii g 20 C!
5 9 0
0 pbml jurcat pbml. jurcat.
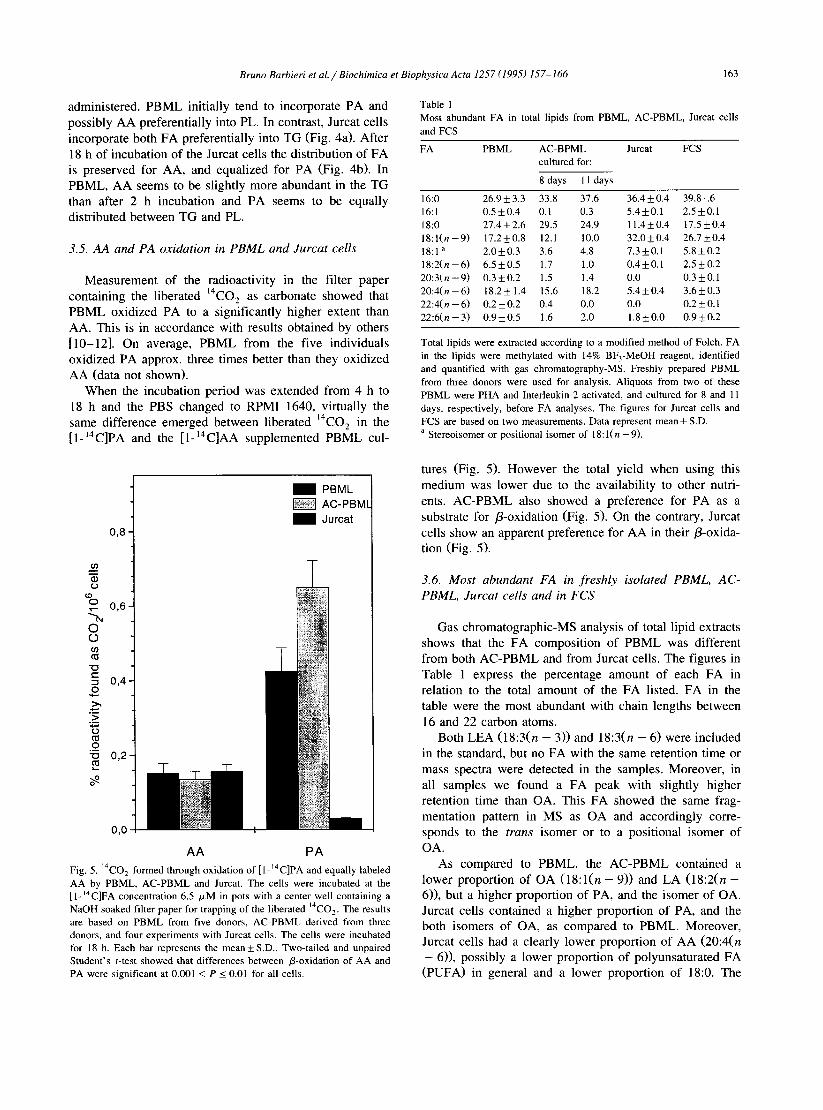
AA PA Fig. 4. Distribution of I- “C-labeled AA and PA between TG and PL in PBML and Jurcat cells. The cells were incubated at the FA concentration 2.16 /.LM for 2 h (panel a) or 18 h (panel b). After incubation cells were washed and sonicated in water. The residue was added MeOH (giving MeOH/H,O, 70:30, v/v) and extracted three times with petroleum ether. After centrifugation the radioactivity in the organic phases and the pellet was measured. Each bar represents the mean ( f S.D.) of three experiments. The residual radioactivity in the pellet is the remaining figure, up to lOO%, for each pair of bars. Two-tailed and unpaired Student’s t-test gave no significant difference between the first pair of bars in panel a. Mutual differences for the other three pair of bars were at least significant at 0.01 < P I 0.05. After 18 h incubation, panel b, a significant difference (0.001 < PI 0.01) in distribution between TG and PL was seen, only for Jurcat cells incubated with AA.
Bruno Barbieri et ai./Biochimica et Biophysics Acta 1257 (1995) 157-166 163
administered. PBML initially tend to incorporate PA and possibly AA preferentially into PL. In contrast, Jurcat cells incorporate both FA preferentially into TG (Fig. 4a). After 18 h of incubation of the Jurcat cells the distribution of FA is preserved for AA, and equalized for PA (Fig. 4b). In PBML, AA seems to be slightly more abundant in the TG than after 2 h incubation and PA seems to be equally distributed between TG and PL.
3.5. AA and PA oxidation in PBML and Jurcat cells
Measurement of the radioactivity in the filter paper containing the liberated 14C02 as carbonate showed that PBML oxidized PA to a significantly higher extent than AA. This is in accordance with results obtained by others [lo-121. On average, PBML from the five individuals oxidized PA approx. three times better than they oxidized AA (data not shown).
When the incubation period was extended from 4 h to 18 h and the PBS changed to RPM1 1640, virtually the same difference emerged between liberated 14C02 in the [I- 14C]PA and the [ 1- 14C]AA supplemented PBML cul-
m PBML
f$f@j AC-PBMI
m Jurcat
T
8
Q,Q
AA PA Fig. 5. 14C0, formed through oxidation of [1-‘4C]PA and equally labeled AA by PBML, AC-PBML and Jurcat. The cells were incubated at the [ I-‘4C]FA concentration 6.5 /.LM in pots with a center well containing a NaOH soaked filter paper for trapping of the liberated “C0,. The results are based on PBML from five donors, AC-PBML derived from three donors, and four experiments with Jurcat cells. The cells were incubated for 18 h. Each bar represents the mean + S.D.. Two-tailed and unpaired Student’s t-test showed that differences between P-oxidation of AA and PA were significant at 0.001 < P I 0.01 for all cells.
Table 1 Most abundant FA in total lipids from PBML, AC-PBML, Jurcat cells and FCS
FA PBML AC-BPML cultured for:
Jurcat FCS
8 days 11 days
16:0 26.9 f 3.3 33.8 37.6 36.4 + 0.4 39.8. .6 16:l 0.5 + 0.4 0.1 0.3 5.4+0.1 2.5*0.1 18:O 27.4+2.6 29.5 24.9 11.4+0.4 17.5kO.4 lS:l(n-9) 17.2kO.8 12.1 10.0 32.0 + 0.4 26.7 to.4 18:l a 2.0 f 0.3 3.6 4.8 7.3*0.1 5.8kO.2 18:2(n-6) 6.5+0.5 1.7 1.0 0.4*0.1 2.5 k 0.2 20:3(n-9) 0.3kO.2 1.5 1.4 0.0 0.3fO.l 20:4(n-6) 18.2+1.4 15.6 18.2 5.4 + 0.4 3.6 f 0.3 22:4(n-6) 0.2iO.2 0.4 0.0 0.0 0.2*0.1 22:6(n-3) 0.9kO.5 1.6 2.0 1.8*0.0 0.9 + 0.2
Total lipids were extracted according to a modified method of Folch. FA in the lipids were methylated with 14% BF,-MeOH reagent, identified and quantified with gas chromatography-MS. Freshly prepared PBML from three donors were used for analysis. Aliquots from two of these PBML were PHA and Interleukin-2 activated, and cultured for 8 and 11 days, respectively, before FA analyses. The figures for Jurcat cells and FCS are based on two measurements. Data represent mean + SD. a Stereoisomer or positional isomer of 18: l(n -9).
tures (Fig. 5). However the total yield when using this medium was lower due to the availability to other nutri- ents. AC-PBML also showed a preference for PA as a substrate for P-oxidation (Fig. 5). On the contrary, Jurcat cells show an apparent preference for AA in their P-oxida- tion (Fig. 5).
3.6. Most abundant FA in freshly isolated PBML, AC- PBML, Jurcat cells and in FCS
Gas chromatographic-MS analysis of total lipid extracts shows that the FA composition of PBML was different from both AC-PBML and from Jurcat cells. The figures in Table 1 express the percentage amount of each FA in relation to the total amount of the FA listed. FA in the table were the most abundant with chain lengths between 16 and 22 carbon atoms.
Both LEA (18:3( n - 3)) and 18:3( n - 6) were included in the standard, but no FA with the same retention time or mass spectra were detected in the samples. Moreover, in all samples we found a FA peak with slightly higher retention time than OA. This FA showed the same frag- mentation pattern in MS as OA and accordingly corre- sponds to the tram isomer or to a positional isomer of OA.
As compared to PBML, the AC-PBML contained a lower proportion of OA (18:l(n - 9)) and LA (18:2(n - 6)), but a higher proportion of PA, and the isomer of OA. Jurcat cells contained a higher proportion of PA, and the both isomers of OA, as compared to PBML. Moreover, Jurcat cells had a clearly lower proportion of AA (20:4(n - 611, possibly a lower proportion of polyunsaturated FA (PUFA) in general and a lower proportion of 18:O. The

164 Bruno Barbieri et al. /Biochimica et Biophwicrr Actrr 1257 f lYY5) 157-166
high proportion of the isomer of OA in both Jurcat cells and AC-PBML seemed to reflect the relatively high amount of this FA in the FCS in which they have been cultured.
4. Discussion
The acetylation of PABA is a common reaction accord- ing to our previous results [5] and takes place in almost every type of tissue [ 131. The reaction mechanism proceeds in a ‘ping pong’ way in which the enzyme, N-acetyltrans- ferase, acetylated by acetyl CoA acetylates the actual amine [14]. Thus, the FA must undergo P-oxidation to form acetyl CoA before acetylation of PABA. Accord- ingly, simple FA like PA and OA would be favored as substrates for the @oxidation and the subsequent N-acety- lation. However, our experiments with PBML incubated with [l- 14C]FA showed that the two essential FA, LA and LEA, as well as AA yielded more radioactive PACBA than the other FA examined. There was no simple correla- tion between the incorporation of the FA into PBML and the formation of radioactive PACBA. For example, incuba- tions with [I-i”C]PA gave high radioactive incorporation but the lowest amount of radioactive PACBA. The per- centage incorporation of the [l- 14C]FA into PBML was not proportional to the respective FA cell content, accord- ing to both our measurements, and others’ [15,16].
The preference for AA as an acetyl donor was further investigated with PBML and with Jurcat cells. The cells were incubated with the most and the least favored FA, i.e., AA and PA, respectively and at higher concentrations than before. Again, AA was clearly the preferred substrate for acetylation of PABA, regardless of the FA concentra- tion or the cell type used.
One would expect that the FA that gave most radioac- tive PACBA also would be preferred as a substrate for the P-oxidation. However, AA is not considered simply as a fuel, especially not when other common fuels are available in excess. Furthermore, when the /l-oxidation of PA and AA in PBML was studied we found that PA was favored as substrate as compared to AA. In contrast, the Jurcat cells showed a significant preference for AA as compared to PA as a substrate in their P-oxidation. Even when PBML were activated to proliferate by PHA-P and inter- leukin-2 and cultured for 8 days, in the same growth medium as used for Jurcat cells, they still showed the same preference for PA in the P-oxidation as freshly prepared PBML.
It has been claimed that the use of labelled FA in metabolic studies could result in labeling of specific pools or lead to FA specific dilutions due to varying content of different endogenous FA. However, the long incubation time used in our experiments should minimize the risk for labeling specific pools [ 171, and the relatively high FA concentrations used should minimize the effect of dilution by endogenous FA.
One possible explanation for the inverse relation be- tween AA as a substrate for P-oxidation and as a substrate for formation of PACBA in PBML might be that AA acetylates PABA by some metabolic route other than the one preceded by conventional P-oxidation. However, such a route has not been described in the literature, as far as we know. Alternatively, a large difference in the abundance of peroxisomes in PBML as compared to Jurcat cells might offer an explanation, providing the peroxisomes have a preference for AA; The main function of peroxisomes is the partial P-oxidation of very long chain FA [ 12,181, dicarboxylic acids [ 191, prostaglandins [20] and hydroxyl- ated SD-cholestanoic acids [21]. The products are chain- shortened acyl-CoA and acetyl-CoA which may leave peroxisomes via pores [22]. Acetyl-CoA, derived from [I-“C]AA, entering the cytoplasm this way could be used by the cytoplasmic N-acetyltransferase to acetylate PABA. Peroxisomal acetyl-CoA formation is probably not propor- tional to AA oxidation measured as ‘“CO? formation in the citric acid cycle because mitochondria are not readily permeable to acetyl-CoA. Thus, a high number of peroxi- somes in PBML could give [ 1- ‘“C]AA derived acetyl-CoA for acetylation of PABA without giving rise to a ‘3C02 production larger than that obtained with [l- ‘“C]PA. Inves- tigations of FA preferences in peroxisomal P-oxidation performed with liver homogenates [23] does not show any AA preference over PA. However the results in this kind of experiments do not necessarily represent the conditions in intact cells [24].
The fate of the acetyl units produced by peroxisomes is not known [2.5]. They may be further oxidized, to some extent. in the mitochondria or used for biosynthetic pur- poses. Our results may indicate one way in which peroxi- somal acetyl-CoA can be used.
Some specific features for the catabolism of AA might be expected since AA is a substrate for production of eicosanoids or is used as a specific building block in the cell membranes as a constituent of phospholipids, pre- dominantly phosphatidylcholine and phosphatidylethanol- amine [ 171. It was early suggested that its slower turnover than other FA reflects the essential character of AA [ 101, and that the marked tendency for AA to enter the phospho- lipid pool [26] would offer an explanation for the usually low catabolic rate of this FA.
In general, TG are less rich in arachidonyl moieties than are PL. The bulk of arachidonate in mammalian cells is esterified in fatty acyl chains of phospholipids [27], and only if the mass of neutral lipids in a tissue exceeds that of phospholipids, e.g. in adipose tissue, might they make a significant contribution to the total AA pool. PLs are important to the structural integrity of the cells and are known to turn over more slowly than TG do [lo]. Our extraction of sonicated Jurcat cells showed that the ra- dioactivity derived from [ 1 - 14C]PA and [ 1- “C]AA initially appeared to a higher extent in the petroleum ether fraction (containing TG) than in the MeOH fraction (containing

Bruno Barbieri et aL/Biochimica et Biophysics Acta 1257 (1995) 157-166 165
PL). For PBML the distribution was the opposite, and tallies with previous reports on human T-lymphocytes [28]. After 18 h incubation with [ l-14C]AA the Jurcat cells still showed the highest radioactivity in the petroleum ether fraction whereas the TG/PL ratio for PBML approached 1 both for AA and PA.
Our measurements of FA in total lipids show that Jurcat cells, cultured with FCS as the only FA source, contain several times more PA than AA. These results are in agreement with other reports [29]. The amount of PA in PBML is slightly higher than the amount of AA. In AC-PBML this relation was somewhat changed. The per- centage PA increased to approximately the same value as in Jurcat cells, but the percentage AA was still approxi- mately the same as in PBML. However, the FA content in AC-PBML still resembles the FA content in PBML more than in Jurcat cells. This indicates that the diversities between PBML and Jurcat cells is not simply due to that Jurcat cells proliferate or to the cells being derived from different FA environment. On the contrary, the differences in P-oxidation, FA distribution after incorporation and FA content seem to reflect genuine differences between benign lymphoid cells and the malignant lymphoid cell line Jurcat.
The preference for AA over the ‘fuel’ FA PA in the P-oxidation might have implications for the homeostasis of the Jurcat cells; It has been shown that PUFA, but not saturated or monounsaturated FA, possess tumoricidal ef- fects [30-321. In addition experiments performed with Jurcat cells clearly indicate that AA, but not OA, suppress [ 3 Hlthymidine incorporation and cell growth [29].
One proposed mechanism for this effect is the forma- tion of free radicals through lipid peroxidation and that malignant cells consequently are thought to be affected as a result of an elevation of toxic peroxidation products [31,32]. Further, it has been claimed that this toxicity would preferentially hit tumour cells because they tend to have less activity of their PUFA desaturase enzymes [30]. Providing tumour cells with PUFA would thereby create a situation where the block in desaturation capacity would be by-passed, leading to selective killing or inhibition of these cells [30,32]. Moreover, it seems to be an abnormal oxygen metabolism in tumor cells where oxygen toxicity is attained, mainly owing to a deficiency of antioxidant de- fences [331.
The high oxidation of AA relative to PA in Jurcat cells could indicate a possible way for the cells to minimize the exposition for radicals formed through lipid peroxidation.
In summary, the present study shows that the N-acetyla- tion of PABA with FA as acetyl donors may be connected to features in FA oxidation that seem to be different between normal lymphoid cells and the T-cell leukemia line Jurcat. Moreover our results demonstrate that the difference in FA content and in P-oxidation between PBML and Jurcat cells is not related to differences in state of proliferation but, rather reflect genuine diversities be- tween these cells.
Acknowledgements
We are indebted to Agnetha Beinhoff and Agneta Sode- rstedt for their invaluable laboratory assistance, to Gunvor Alvelius for skillful assistance with MS and to Aldo Barbieri for constructing the pots for measurement of “c02.
This work was supported by grants from the Karolinska Institutet, the Swedish Cancer Society and Magnus Berg- wall’s Foundation, Stockholm.
References
111
121
[31
[41
151
[61
[71
P31
191
1101 [III
1121
1131
[141 [ISI
1161 1171
[I81 1191
DOI Dll
WI b31
[241
1251
[261
b71
Paul, .I. (1975) in Cell and Tissue Culture, pp. 58, Churchill Living- stone, Edinburgh. Harris, R.S., Wright, L.D., Tavormina, P.A. and Scott C.C. (1954) in The Vitamins (Sebrell, W.H. and Harris, R.S., eds.), Vol. 3. pp. l-85, Academic Press. Kovacs, J.A., Powell, F., Voeller, D. and Allegra, C.J. (1993) Antimicrob. Agents. Chemother. 37, 1227-1231. Johnsen, S.A., Olding, L.B. and Green, K. (1983) Immunol. Lett. 6, 213-218. Barbieri, B., Papadogiannakis, N., Eneroth, P., Hansson, C., Roep- storff, P. and Olding, L.B. (1994) Biochim. Biophys. Acta 1214, 309-3 16. Johnsen, S.A., Green, K., Papadogiannakis N. and Olding, L.B. (1987) Stand. J. Immunol. 25, 425-428. Auestad, N., Korsak, R.A., Morrow, J.V. and Edmond, J. (199 1) J. Neurochem. 4, 1376-1386. Venizelos, N. and Hagenfeldt, L. (1985) Stand. J. Clin. Lab. Invest. 45, 335-340. McDonald-Gibson, R.G. (1987) in Prostaglandins and Related Sub- stances (Benedetto, C., McDonald-Gibson, R.G., Nigam, S. and Slater, T.F., eds.), pp. 259-265, IRL Press, Oxford. Coots, R.H. (1965) J. Lipid Res. 6, 494-497. Coniglio, J.G., Davis, J.T. and Aylward, S. (1964) J. Nutr. 84, 265-27 1. Street, J.M., Johnson, D.W., Singh, H. and Poulos, A. (1989) Biochem. J. 260, 647-655. Gunawardhana, L., Barr, J., Weir, A.J., Brendel, K and Sipes, I.G. (1991) Drug Metabol. Dispos. 3, 648-654. Weber, W.W. and Hein, D.W. (1985) Pharmacol. Rev. 37, 25-79. Marinetti, G.V. and Cattieu, K. (1982) Chem. Phys. Lipids 31, 169-177. Delplanque, B. and Jacotot, B. (1987) Lipids 22, 241-249. Hong, S.L. (1988) in Arachidonate Metabolism in Immunologic Systems (Levine, L., ed.), pp. 99-139, Karger, Basel. Osmundsen, H. (1982) Ann. NY Acad. Sci. 386, 13-29. Kiiivraa, S. and Gregersen, N. (1986) Biochim. Biophys. Acta 876, 515-525. Diczfalusy, U. (1994) Prog. Lipid Res. 33, 403-428. Kase. B.F., Prydz, K., Bjijrkhem, I. and Pedersen, J.I. (1986) Biochim. Biophys. Acta 877, 37-42. Schulz, H. (1991) Biochim. Biophys. Acta 1081, 109-120. Reubsaet, F.A.G., Veerkamp, J.M.F., Trijbels, J.M.F. and Monnens, L.A.H. (1989) Lipids 24, 945-950. Mannaerts, G.P. and Van Veldhoven, P.P. (1993) Verh. K. Acad. Geneeskd. Belg. 55, 45-78 (in Dutch). Reddy, J.K. and Mannaerts, G.P. (1994) Annu. Rev. Nutr. 14, 343-370. Stoffel, W. and Scheid, A. (1967) Hoppe-Seyler’s Z. Physiol. Chem. 348, 205-226 (in German). Irvine, R.F. (1982) Biochem. J. 204, 3-16.

166 Brunn Barbieri et al./ Biochimica et Biophysics Acta 1257 (19951 157-166
1281 Goppelt, M. KGhler, L. and Resch, K. (1985) Biochim. Biophys. Acta 833, 463-472.
[29] Chow, SC., Sisfontes, L., BjGrkhem, I. and Jondal, M. (1989) Lipids 24, 700-704.
1301 Be’gin, M.E., Das, U.N., Ells, G. and Horrobin, D.F. (1985) Prosta- glandins Leukotrienes Med. 19, 177- 186.
[3 I] Smith, D.E. and Salerno, J.W. (1992) Prostaglandins Leukotrienes Essent. Fatty Acids 46, 145-150.
[32] Be’gin, M.E., Ells, G. and Horrobin, D.F. (1988) J. Natl. Cancer. Inst. 80, l88- 194.
[33] Galeotti, T., Borello, S., Minotti, G. and Masotti, L. (1986) Ann. N.Y. Acad. Sci. 488, 468-480.




















