
Alteration of GCN5 levels in maize reveals dynamic responses to manipulating histone acetylation
15
Alteration of GCN5 levels in maize reveals dynamic responses to manipulating histone acetylation Riyaz A. Bhat # , Marcus Riehl #,y , Geraldina Santandrea z , Riccardo Velasco § , Stephen Slocombe , Gu ¨ nter Donn yy , Hans-Henning Steinbiss, Richard D. Thompson zz and Heinz-Albert Becker Max-Planck-Institut fu ¨ r Zu ¨ chtungsforschung, Carl-von-Linne ´ Weg 10, D-50829 Ko ¨ ln, Germany Received 8 July 2002; revised 8 October 2002; accepted 30 October 2002. For correspondence (fax þ49 221 5062 413; e-mail [email protected]). # These authors contributed equally to the manuscript. y Present address: Novoplant, D-06466 Gatersleben, Germany. z Present address: MPI fu ¨ r medizinische Forschung, Jahnstrasse 29, D-69120 Heidelberg, Germany. § Present address: Istituto Agrario di S.Michele all’Adige Via Mach, 1-38010-S. Michele a/Adige Italy. Present address: Centre for Plant Sciences (LIBA), University of Leeds, Leeds LS2 9JT, UK. yy Present address: Aventis Crop Science (Bayer Crop Science), 65929 Frankfurt, Germany. zz Present address: Laboratoire des le ´ gumineuses, URGAP-INRA BP 86510, 21065 Dijon ce ´ dex, France. Summary The role played by histone acetyltransferase (HAT), GCN5, in transcriptional co-activation has been ana- lysed in detail in yeast and mammals. Here, we present the cloning and expression pattern of Zmgcn5, the maize homologue. The enzymatic activity of the recombinant ZmGCN5 was analysed with histone and nucleosome substrates. In situ hybridisation of developing maize kernels using Zmgcn5 as probe shows that the transcript is concentrated in rapidly dividing cells. To investigate the role of ZmGCN5 in the transcription of specific plant genes, direct protein–protein interactions were tested. A cDNA clone encod- ing a putative interacting partner in GCN5-adapter complexes, ZmADA2, was isolated and the interaction between ZmGCN5 and ZmADA2 was confirmed by a GST-spin down experiment. Co-immunoprecipitation of the plant transcriptional activator Opaque-2 and ZmADA2 in nuclear extracts suggests ADA2/GCN5- containing complexes to mediate transcriptional activation by binding of this bZIP factor. For a more general analysis of the effects of histone acetylation on plant gene expression, 2500 ESTs spotted on filters were hybridised with cDNA probes derived either from maize cell lines treated with Trichostatin A (TSA), or from a transgenic line expressing the ZmGCN5 antisense transcript. Several sequences showing marked changes in abundance were confirmed by RNA blot analysis. Inhibition of histone deacetylation with TSA is accompanied by a decrease in the abundance of ZmGCN5 acetylase protein, but by increases in mRNAs for histones H2A, H2B, H3 and H4. The elevated histone mRNA levels were not reflected in increasing histone protein concentrations, suggesting hyperacetylated histones arising from TSA treatment may be preferen- tially degraded and substituted by de novo synthesised histones. The ZmGCN5 antisense material showed suppression of the endogenous ZmGCN5 transcript and the profiling analysis revealed increased mRNA levels for H2A, H2B and H4. Furthermore, in the antisense line, a reduction in the amount of the RPD3-type HD1B-I histone deacetylase protein was observed. A model for linked regulation of histone acetylation and histone mRNA transcription is discussed. Keywords: histone acetyltransferase, co-activator, GCN5, ADA2, maize, Tricnostatin A. Introduction The regulation of eukaryotic gene expression involves interactions of gene-specific transcription factors with chro- matin-associated co-activator proteins. Co-activators mod- ify nucleosomes to facilitate access by the transcription machinery to the DNA. One class changes nucleosome configuration by ATP-dependent re-modelling on the DNA strand, whereas the second class acts via regulating histone acetylation status. Examples of plant genes puta- tively involved in ATP-dependent chromatin re-modelling are the Arabidopsis thaliana DDM1 and MOM loci (Amedeo et al., 2000; Kakutani et al., 1995), which have homology to the SWI2/SNF2 class of proteins and the A. thaliana The Plant Journal (2003) 33, 455–469 ß 2003 Blackwell Publishing Ltd 455
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Alteration of GCN5 levels in maize reveals dynamic responses to manipulating histone acetylation

Alteration of GCN5 levels in maize reveals dynamicresponses to manipulating histone acetylation
Riyaz A. Bhat#, Marcus Riehl#,y, Geraldina Santandreaz, Riccardo Velasco§, Stephen Slocombe��,
Gunter Donnyy, Hans-Henning Steinbiss, Richard D. Thompsonzz and Heinz-Albert Becker�
Max-Planck-Institut fur Zuchtungsforschung, Carl-von-Linne Weg 10, D-50829 Koln, Germany
Received 8 July 2002; revised 8 October 2002; accepted 30 October 2002.�For correspondence (fax þ49 221 5062 413; e-mail [email protected]).#These authors contributed equally to the manuscript.yPresent address: Novoplant, D-06466 Gatersleben, Germany.zPresent address: MPI fur medizinische Forschung, Jahnstrasse 29, D-69120 Heidelberg, Germany.§Present address: Istituto Agrario di S.Michele all’Adige Via Mach, 1-38010-S. Michele a/Adige Italy.��Present address: Centre for Plant Sciences (LIBA), University of Leeds, Leeds LS2 9JT, UK.yyPresent address: Aventis Crop Science (Bayer Crop Science), 65929 Frankfurt, Germany.zzPresent address: Laboratoire des legumineuses, URGAP-INRA BP 86510, 21065 Dijon cedex, France.
Summary
The role played by histone acetyltransferase (HAT), GCN5, in transcriptional co-activation has been ana-
lysed in detail in yeast and mammals. Here, we present the cloning and expression pattern of Zmgcn5, the
maize homologue. The enzymatic activity of the recombinant ZmGCN5 was analysed with histone and
nucleosome substrates. In situ hybridisation of developing maize kernels using Zmgcn5 as probe shows
that the transcript is concentrated in rapidly dividing cells. To investigate the role of ZmGCN5 in the
transcription of specific plant genes, direct protein–protein interactions were tested. A cDNA clone encod-
ing a putative interacting partner in GCN5-adapter complexes, ZmADA2, was isolated and the interaction
between ZmGCN5 and ZmADA2 was confirmed by a GST-spin down experiment. Co-immunoprecipitation
of the plant transcriptional activator Opaque-2 and ZmADA2 in nuclear extracts suggests ADA2/GCN5-
containing complexes to mediate transcriptional activation by binding of this bZIP factor. For a more
general analysis of the effects of histone acetylation on plant gene expression, 2500 ESTs spotted on filters
were hybridised with cDNA probes derived either from maize cell lines treated with Trichostatin A (TSA), or
from a transgenic line expressing the ZmGCN5 antisense transcript. Several sequences showing marked
changes in abundance were confirmed by RNA blot analysis. Inhibition of histone deacetylation with TSA is
accompanied by a decrease in the abundance of ZmGCN5 acetylase protein, but by increases in mRNAs for
histones H2A, H2B, H3 and H4. The elevated histone mRNA levels were not reflected in increasing histone
protein concentrations, suggesting hyperacetylated histones arising from TSA treatment may be preferen-
tially degraded and substituted by de novo synthesised histones. The ZmGCN5 antisense material showed
suppression of the endogenous ZmGCN5 transcript and the profiling analysis revealed increased mRNA
levels for H2A, H2B and H4. Furthermore, in the antisense line, a reduction in the amount of the RPD3-type
HD1B-I histone deacetylase protein was observed. A model for linked regulation of histone acetylation and
histone mRNA transcription is discussed.
Keywords: histone acetyltransferase, co-activator, GCN5, ADA2, maize, Tricnostatin A.
Introduction
The regulation of eukaryotic gene expression involves
interactions of gene-specific transcription factors with chro-
matin-associated co-activator proteins. Co-activators mod-
ify nucleosomes to facilitate access by the transcription
machinery to the DNA. One class changes nucleosome
configuration by ATP-dependent re-modelling on the
DNA strand, whereas the second class acts via regulating
histone acetylation status. Examples of plant genes puta-
tively involved in ATP-dependent chromatin re-modelling
are the Arabidopsis thaliana DDM1 and MOM loci (Amedeo
et al., 2000; Kakutani et al., 1995), which have homology
to the SWI2/SNF2 class of proteins and the A. thaliana
The Plant Journal (2003) 33, 455–469
� 2003 Blackwell Publishing Ltd 455

embryo-defective mutation PICKLE, containing SNF2-like
domains (Ogas et al., 1999).
The histone acetylation status of eukaryotic genomes is
balanced between antagonistic acetylase and deacetylase
activities. The resulting reversible modifications promote,
in the case of N-terminal, histone acetylation access to the
chromatin, whereas deacetylation leads to chromatin con-
densation. Both histone acetylation and deacetylation
occur in plants, and it is likely that these processes regulate
similar functions to those identified in other eukaryotes.
Histone acetyltransferases (HATs) have been classified into
type-A and type-B enzymes. Type-B HATs are at least partly
localised in the cytoplasm, and are thought to be involved
in nucleosome assembly-related histone acetylation (Kolle
et al., 1998; Lusser et al., 1999). Type-A HATs are nuclear-
localised, and are implicated in regulating transcription,
although their involvement in other processes such as
DNA repair and replication is also likely.
In plants, H3 is the most extensively acetylated histone in
contrast to the situation in non-plant eukaryotes where H4
is most highly acetylated (Waterborg, 1990). A further dif-
ference is that in plants, H4 is acetylated at five lysine
residues, including Lys-20, whereas this lysine residue is
methylated rather than acetylated in non-plant H4 histones.
Fractionation of nuclear extracts from germinated maize
embryos yielded three distinct HAT activities (Lechner et al.,
1996), HAT-A1, HAT-A2, and HAT-B. Whereas HAT-B ace-
tylates H4 at Lys-5 and -12, acetylation at Lys-8, -16, and -20
is presumably being carried out by type-A HATs (Kolle et al.,
1998). Several sequence classes of type-A HATs have been
reported, including GCN5, MYST and ESA1 (reviewed in
Lusser et al., 2001).
GCN5 was the first type-A HAT of known function, one of
the several gcn (general control of nitrogen) genes which
are required for the full activity of a bZIP transcription
factor, GCN4, in the yeast cell’s response to amino acid
deficiency. In yeast, GCN5 exists as in multicomponent
complexes up to 2 MD in size. In combination with histone
deacetylases and chromatin re-modelling factors, these
large complexes maintain the dynamic status of the gen-
ome required for co-ordinated gene expression. Direct
evidence for an association of histone acetylation with
promoter regions of actively transcribed plant genes has
been provided by chromatin immunoprecipitation (CHIP)
experiments, using antibodies raised against acetylated H3
and H4 tails (Chua et al., 2001). Genes involved in the
process of plant histone deacetylation have also been
cloned and analysed for their activity (Rossi et al., 1998;
Tian and Chen, 2001; Wu et al., 2000a,b). Targeting a
deacetylase, AtHD2A, to a reporter gene in vivo, caused
its repression (Wu et al., 2000b).
One partner protein for GCN5 in the multicomponent
complexes is the adapter protein ADA2 (Berger et al.,
1992), which in yeast besides binding to GCN5, also directly
interacts with the acidic activation domain of the DNA-
binding transcriptional activator GCN4, and is thought to
furnish a link between the upstream activating sequences
and the transcription machinery (Barlev et al., 1995). The
maize transcriptional activator Opaque-2 (O2), expressed
during late endosperm development, bears a number of
similarities to yeast GCN4 (Hartings et al., 1989). Both
possess bZIP DNA-binding domains and have acidic activa-
tion domains (Schmitz et al., 1997), raising the possibility
that O2 might recruit co-activator complexes via an inter-
action with a maize ADA2 orthologue. Recently, A. thaliana
homologues of GCN5 and ADA2 were isolated and shown
to interact with one another and with a CBF1 (DREB1B), an
AP2/EREBP binding domain-containing transcription factor
involved in cold-regulated gene expression (Stockinger
et al., 2001). We have cloned and analysed the role of
GCN5 and putative partner proteins as central components
of transcriptional co-activation in maize.
Results
Genetic isolation of ZmGCN5 on the basis of an
interaction with the partner protein ADA2
Database searches for plant orthologues of gcn5 and ada2
yielded two A. thaliana sequences, a cDNA for Atgcn5
(AF037442), and an Atada2 genomic sequence (Z97341).
Using primers derived from the Atada2 genomic sequence,
the corresponding A. thaliana Atada2 cDNA was isolated by
RT-PCR. This cDNA was then used as a bait fusion to screen
a maize 7 days after pollination (DAP), endosperm hybriZAP
cDNA library. A total of 800 000 clones were screened, and
16 interacting clones were obtained that were able to grow
on –his plates containing 15 mM 3-amino-1,2,4-triazole (3-
AT). Of the 16 clones obtained, one clone contained a
sequence related to GCN5, and the remaining 15 clone
sequences had no sequence homology to other database
accessions. This clone, which by sequence comparison was
found to be partial, was used to isolate a full-length cDNA
by screening a maize pistil (silk) cDNA library. The cDNA
contained an open reading frame of 1545 bases, predicting
a polypeptide of 515 amino acids (Figure 1a). ZmGCN5
contains a predicted plant nuclear localisation sequence
of four amino acids (situated at amino acids 23–26). In
Figure 1(b), the predicted coding sequence is compared
to that of other members of the GCN5 family. The three
functional domains present in other GCN5 sequences are
shared by ZmGCN5: the catalytic domain at the N-terminus,
a centrally located domain responsible for the interaction
with ADA2, and the C-terminal bromo-domain, which may
interact with the histone N-termini. Although the N-term-
inal region of ZmGCN5 is apparently extended compared to
that of ScGCN5, this extension is unrelated to the PCAF
456 Riyaz A. Bhat et al.
� Blackwell Publishing Ltd, The Plant Journal, (2003), 33, 455–469

domain found at the N-terminus of some GCN5-class HATs,
which has been proposed to be required for acetylation of
histones in nucleosomes (Xu et al., 1998). It is furthermore
unrelated to an extension present in an AtGCN5 N-terminal
sequence (Stockinger et al., 2001) or to the protozoan
parasite homologue TgGCN5 (Sullivan and Smith, 2000).
Organisation and expression of the Zmgcn5 locus
The copy number and organisation of Zmgcn5 sequences
in the maize genome was estimated by Southern hybridisa-
tion (Figure 2a), which indicated that the gene is present as
a single copy in maize. It is however, possible that other
GCN5 like sequences exist in the genome which do not
hybridise or interact with the A. thaliana bait in the yeast
two-hybrid experiment. To examine the expression pattern,
RNA blot hybridisation was carried out with polyAþ RNA
Figure 1. ZmGCN5 open reading frame (ORF) and comparison to othermembers of the GCN5 family.(a) The ZmGCN5 ORF is shown along with the translation product. Intronpositions are indicated by arrows. Italic letters mark the nuclear localisationsequence present in the N-terminus. The HAT domain is indicated by a blackbar, the Ada2 interaction domain by a light bar and the bromo-domain ismarked by a black bar.(b) Protein sequence comparison to other members of the GCN5 family. Allfamily members share the catalytic HAT domain, the Ada2 interactiondomain, and the bromo-domain. Amino acid positions of the domainsare indicated above the diagram. The percentage of identical amino acidresidues is shown on the right. Accession numbers: HsGCN5-L, AF029777;ZmGCN5, AJ428542; AtGCN5, AF031958; ScGCN5, X608628; TgGCN5,AF197953. Hs, Homo sapiens; L, long form; Zm, Zea mays; At, Arabidopsisthaliana; Sc, Saccharomyces cerevisiae; Tg, Toxoplasma gondii.
Figure 2. Zmgcn5, a single copy gene expressed ubiquitously in maize.(a) Southern filters of maize inbreds B73 and A69Y hybridised with Zmgcn5.The inbred lines were digested with different enzymes. Hybridisation wasperformed with the random prime-labelled full-length cDNA.(b) RNA filter hybridisations of Zmgcn5. As a loading control, the filter washybridised with a polyubiquitin probe. The numbers below the lanes givethe division products of the quantified hybridisation signals (Zmgcn5/Ubi-quitin).
� Blackwell Publishing Ltd, The Plant Journal, (2003), 33, 455–469
ZmGCN5, a plant nuclear HAT 457

from various plant tissues and organs (Figure 2b). A single
2.2-kb transcript was detected. Quantification relative to the
ubiquitin transcript used as a control indicates that Zmgcn5
is expressed throughout the plant, with highest abundance
in tissues containing a high proportion of rapidly dividing
cells, such as young leaves, young shoots and the endo-
sperm. The lowest level of mRNA was detected in pollen.
Young maize kernels express Zmgcn5 mainly in the
chalazal zone of the endosperm
To investigate the cellular distribution of Zmgcn5 tran-
scripts, in situ hybridisation was performed with the
Zmgcn5 sense and antisense transcript. Counterstaining
was performed using acetocarmine red. In cross-sections
of 5 DAP developing maize kernels, the ZmGCN5 transcript
is concentrated at the periphery of the embryo sac (Figur-
e 3b,d). No signal can be detected with the sense probe
(Figure 3a,c). A strong signal is already detectable in devel-
oping embryo sacs, probed with antisense RNA 3 DAP
(Figure 3f), with the sense probe showing no signal (Fig-
ure 3e). Furthermore, the ZmGCN5 transcript is more abun-
dant in the basal endosperm (Figure 3b,d) in a cluster of
cells surrounding the embryo which are the first to cellu-
larise and constitute the embryo surrounding region (ESR).
Despite the fact that Northern analysis shows ZmGCN5
expression throughout the plant, in situ hybridisation indi-
cates the transcript is found more abundantly in the mer-
istematic cells.
The N-terminus is responsible for nuclear
targeting of ZmGCN5
The identification of ZmGCN5 as a type-A HAT involved in
transcription implies that the protein is localised in the
nucleus. A potential nuclear localisation sequence (NLS)
was identified in the N-terminus (Figure 1a). To confirm the
existence of a functional NLS, a series of deletions of the
coding sequence were fused to the green fluorescent pro-
tein (GFP) reporter and the resulting plasmids were trans-
fected into tobacco protoplasts. Nuclear localisation of the
GFP signal was only achieved using fusions containing the
N-terminal region of ZmGCN5, such as the full-length
35S::GFP–GCN5 fusion (Figure 3g) and the 35S::N-terminal
domain fusion (1–175) (Figure 3h). Constructs lacking the N-
terminus like the 35S::GCN5 (175–314) (Figure 3i), the
35S::GCN5 (314–515) (Figure 3k), or the 35S:GFP control
(Figure 3l) can be detected in both cytosolic and nuclear
compartments, thus pinpointing a functional nuclear loca-
lisation signal close to the N-terminus of the protein (see
Figure 1a), upstream of the region of homology with
ScGCN5. Due to the presence of a large vacuole, cytoplas-
mically located GFP signal tends to be concentrated around
the nucleus in all protoplasts; a better indication of genuine
nuclear localisation is the absence of signal in peripheral
cytoplasm.
In vitro enzymatic activity of ZmGCN5
To determine whether ZmGCN5 possesses HAT activity,
the full-length protein and a deletion lacking the HAT
domain were expressed in E. coli as C-terminal fusions to
glutathione S-transferase (GST). This enabled the native
fusion polypeptide to be purified by chromatography on
glutathione–agarose columns. The fusion peptide was also
supplied for polyclonal antiserum production in rabbits.
In vitro acetylation assays were carried out using core his-
tones as substrate, and labelling with 3H-acetyl coenzyme
A. A time course of incorporation demonstrated that the
assay was being carried out under substrate excess (Fig-
ure 4a). It could be shown that the HAT activity was con-
ferred by preparations of the full-length GCN5–GST fusion
protein, but not by the fusion without the catalytic HAT
domain (Figure 4b). The absence of significant chemical
acetylation was confirmed by lack of incorporation in con-
trols without enzyme. HAT activity in nuclear extract from
maize endosperm was also able to acetylate the core his-
tone substrate, albeit at a much higher protein concentra-
tion than seen for the recombinant fusion peptide.
ScGCN5, which has been shown to be a component of a
170-kDa complex (Grant et al., 1997), is capable of acetylat-
ing histones in nucleosomes in vivo (Ruiz-Garcia et al.,
1997). Tse et al. (1998) have also reported in vitro acetyla-
tion of nucleosomes by ScGCN5 alone. When the ability of
the recombinant ZmGCN5 to acetylate nucleosomes was
compared to its activity on core histones, only the latter
were acetylated under our experimental conditions (data
not shown).
To investigate the specificity of acetylation of ZmGCN5
on maize core histones after electrophoresis, incorporated3H was examined by fluorography (Figure 4c). The recom-
binant ZmGCN5 appears to acetylate histone H3 preferen-
tially, with less incorporation seen for H4, H2A and H2B. The
nuclear extract showed a similar distribution of label
between the histone species to that seen with ZmGCN5,
but with proportionally stronger signal in histone H4.
Isolation of a maize orthologue ADA2 cDNA,
a partner protein in GCN5 complexes
Database searches identified a Hordeum vulgare EST (clone
BCD450, GenBank accession no. AA231679) homologous to
Atada2. Primers were designed based on this sequence,
and a 226-bp fragment amplified by genomic PCR was used
to screen a barley cDNA library. A cDNA clone approxi-
mately 1 kb in length was used in turn to screen (608C,
4�SSC) 500 000 phages from an amplified maize silk cDNA
library at low stringency. The screen yielded two positive
� Blackwell Publishing Ltd, The Plant Journal, (2003), 33, 455–469
458 Riyaz A. Bhat et al.
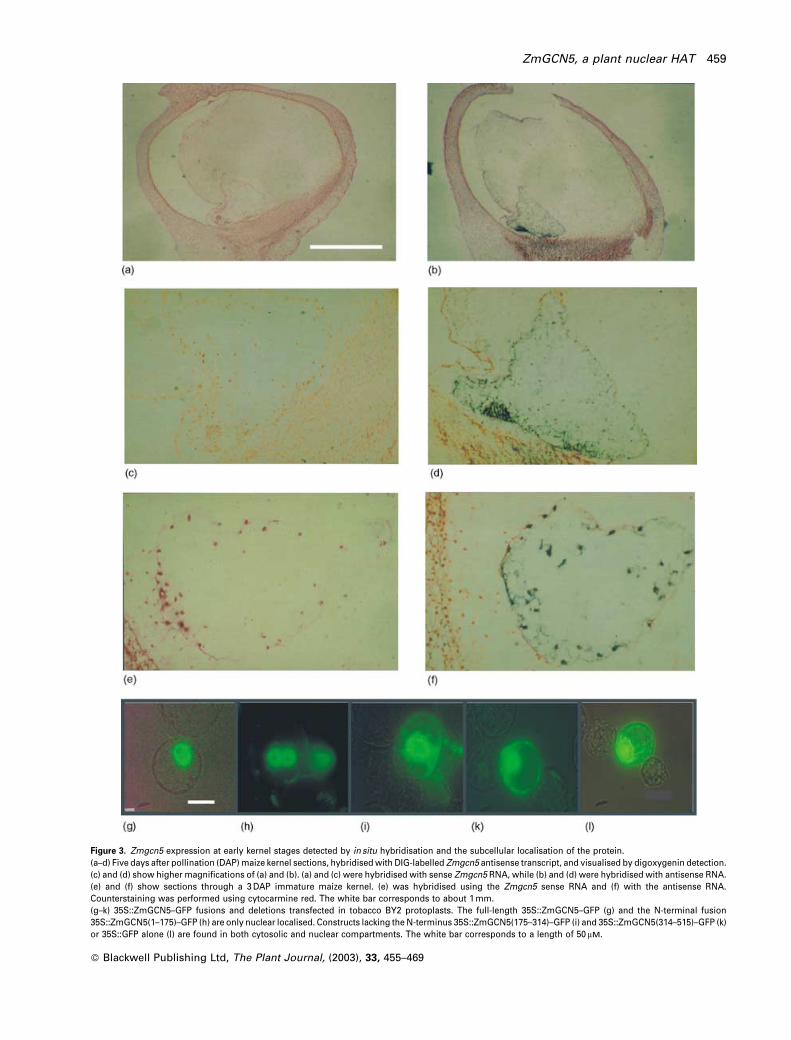
Figure 3. Zmgcn5 expression at early kernel stages detected by in situ hybridisation and the subcellular localisation of the protein.(a–d) Five days after pollination (DAP) maize kernel sections, hybridised with DIG-labelled Zmgcn5 antisense transcript, and visualised by digoxygenin detection.(c) and (d) show higher magnifications of (a) and (b). (a) and (c) were hybridised with sense Zmgcn5 RNA, while (b) and (d) were hybridised with antisense RNA.(e) and (f) show sections through a 3 DAP immature maize kernel. (e) was hybridised using the Zmgcn5 sense RNA and (f) with the antisense RNA.Counterstaining was performed using cytocarmine red. The white bar corresponds to about 1 mm.(g–k) 35S::ZmGCN5–GFP fusions and deletions transfected in tobacco BY2 protoplasts. The full-length 35S::ZmGCN5–GFP (g) and the N-terminal fusion35S::ZmGCN5(1–175)–GFP (h) are only nuclear localised. Constructs lacking the N-terminus 35S::ZmGCN5(175–314)–GFP (i) and 35S::ZmGCN5(314–515)–GFP (k)or 35S::GFP alone (l) are found in both cytosolic and nuclear compartments. The white bar corresponds to a length of 50 mM.
� Blackwell Publishing Ltd, The Plant Journal, (2003), 33, 455–469
ZmGCN5, a plant nuclear HAT 459

clones, of which one possessed an open reading frame.
The Zmada2 cDNA clone was sequenced, and showed
homology to the published ada2 genes from A. thaliana.
Recombinant ZmADA2 was then used to raise anti
bodies.
ZmADA2 interacts with ZmGCN5 in vitro
A GST pull-down assay was set up to investigate the
possibility of a GCN5–ADA2 interaction for the maize pro-
teins (Figure 5a). By using in vitro transcribed and trans-
lated ZmGCN5 protein and recombinant ZmADA2–GST
Figure 4. ZmGCN5 acetylates preferentially histone H3 in core histonepreparations(a) Time course of in vitro acetylation activity of recombinant ZmGCN5–GSTon calf thymus histones.(b) Specificity of in vitro acetylation by recombinant ZmGCN5. Differentincubation sets were used to demonstrate that specific HAT activity on corehistones can only be observed in the presence of the full length ZmGCN5. Asa positive control plant nuclear extract was incubated with calf thymushistones.(c) Acetylation activities of ZmGCN5–GST and maize nuclear extracts onmaize core histones. The 3H-fluorogram shows the acetylation productsafter separation on 12% PAGE.
Figure 5. In vitro interactions of ZmADA2, ZmGCN5, nuclear componentsand O2.(a) ZmADA2–ZmGCN5 interaction. A GST pull-down experiment was per-formed using 35S-labelled ZmGCN5 and ZmADA2–GST. Lane 1 shows thebackground of the transcription-and-translation mix (TþT) when no plas-mid is added to the transcription reaction and the TþT is loaded directly. Anasterisk in (a) and (b) marks an unspecific protein band which appearsindependent of a specific transcript. In lanes 2–5, TþT products were usedwhen a plasmid specific for the transcription of 35S-ZmGCN5 was added tothe transcription reaction. Lane 5 shows that 35S-ZmGCN5 is pulled downonly in the presence of ZmADA2–GST.(b) ZmADA2 antibody recognises 35S-O2 in the presence of nuclear extracts.A co-immunoprecipitation was performed using the 35S-labelled O2, maizenuclear extract and a specific ZmADA2 antibody. Lane1 shows the productof theTþTreactionwherethe35S-O2ispresent (markedbyablackarrow).Theasterisk again marks the background band described in (a). The 35S-O2 speci-fic signal is retained only when the protein-A bound ZmADA2 antibody andthe nuclear extract are both present in the binding reaction (lane 5). In lane 3,the pre-immune serum was used instead of the specific ZmADA2 antibody.
� Blackwell Publishing Ltd, The Plant Journal, (2003), 33, 455–469
460 Riyaz A. Bhat et al.

fusion protein, the interaction between ZmGCN5 and
ZmADA2 could be detected in vitro. The radiolabelled
ZmGCN5 is adsorbed to glutathione–agarose beads only
in the presence of ZmADA2–GST (lane 5). No signal is
retained by the glutathione–agarose when no 35S-GCN5
is used (lane 2), when no ZmADA2–GST is added (lane
3), or when ZmADA2–GST is replaced by GST alone (lane 4).
As an additional control, lane 1 shows the background of
the coupled transcription/translation reaction without the
addition of a plasmid for a specific Zmgcn5 transcript (an
asterisk marks a background band in the translation pro-
ducts which appears even when no plasmid is added to the
transcription mix). Both proteins are therefore functionally
expressed in heterologous systems. In further GST pull-
down experiments, no interaction between 35S-ZmADA2
and the O2 protein, expressed as a GST-fusion, was
observed (data not shown).
Co-immunoprecipitation with AntiADA2 antibody detects
a specific interaction between O2 and components in
nuclear extracts
Since no specific interaction between recombinant O2 and
ZmADA2 proteins was detectable in vitro, and ADA2 is
normally present as part of multicomponent complexes
(Grant et al., 1997), we investigated whether the interaction
may require the presence of additional components. As an
alternative to the incubation of recombinant proteins, pO2,
after in vitro transcription was 35S-labelled by in vitro trans-
lation and then incubated with nuclear extracts. The result-
ing interaction was detected by co-immunoprecipitation
using the ZmADA2 antibody in the presence of protein-A
sepharose (Figure 5b). The interaction was not seen if no
antibody was added or pre-immune serum was used
instead of ZmADA2 antiserum (Figure 5b, lanes 2 and 3),
or if nuclear extract was omitted (lane 4), but required pO2,
antiZmADA2 antibody, and nuclear extract (lane 5). Lane 1
shows the background of the 35S-O2 translation product
loaded directly without co-immunoprecipitation.
ZmGCN5 protein level is regulated by the acetylation
status of the genome
To monitor whether the changes in the chromatin acetyla-
tion status affect the ZmGCN5 amount, maize suspension
cultures were treated with the deacetylase inhibitor Tri-
chostatin A (TSA). After 10 h of TSA treatment, crude
nuclear extracts were subjected to electrophoresis and
analysed by fluorography. During this time the amount
of acetylated histone H4 increases in relation to the cyto-
solic protein immunophilin used as a loading control
(Figure 6a), whereas the total histone fraction is not
affected. On the other hand the level of ZmGCN5 decreases
several fold in response to increasing TSA amounts
(Figure 6b). Therefore, an increase in the amount of acety-
lated H4 protein corresponds with a decreasing ZmGCN5
protein amount, but does not seem to influence the total
histone concentration.
Array analysis reveals histone transcription to be
induced by TSA treatment
To investigate the more global effects of acetylation inhibi-
tion, RNA was extracted at time points between 6 and 12 h
after TSA treatment. The radiolabelled cDNA generated was
then used formedium density array hybridisation using 2500
maize ESTs spotted on nylon filters. The clones showing
major differences in signal intensities are listed in Figure 7(a)
(induced sequences) and Figure 7(b) (repressed sequences).
Interestingly, histones H2A, H2B, H3 and H4 appear several
times in the most-affected clones, indicating the cell is res-
ponding to an increase in the degree of histone acetylation
probably by de novo histone mRNA synthesis. (To increase
clarity tables have been edited. The complete hybridisation
results and the sequences used are accessible at http://
www.mpiz-koeln.mpg.de/�riehl/ArrayDB/MzArrayDB.htm)
The same treated material was then subjected to North-
ern analysis to confirm the major changes in transcript
abundance detected on EST arrays. mRNAs encoding his-
tones H4 and H2, the Cytochrome P450 monooxygenase
and the MFS 18 protein precursor were all strongly upre-
gulated by TSA treatment (Figure 7c). Both non-histone
sequences were randomly chosen. The increased histone
transcription is in contrast to the histone protein levels
which remained relatively constant although the ZmGCN5
protein levels were downregulated several fold (Figure 6).
Reduction in ZmGCN5 acetylase expression results in
gene expression changes partly overlapping with
those seen with TSA treatment
In a second approach specific for the ZmGCN5 contribution
to transcription of specific genes, transgenic antisense lines
of Zmgcn5 were generated. Northern analysis revealed for
two of the lines a reduction in Zmgcn5 transcript abun-
dance compared to the vector-transformed control (Fig-
ure 8a). Microarray analysis of line 4 showed mRNAs for
histones H2A, H2B and H4 to be increased as seen on TSA
treatment, as well as the MFS18 protein precursor (Fig-
ure 8b). Interestingly, histone H1 was found among the
ESTs showing attenuated signals (Figure 8c).
Protein analysis of the antisense cell lines confirmed
the reduction in ZmGCN5 protein concentration (Figure 8d).
To see whether the transcriptome response to TSA treat-
ment includes other proteins of histone metabolism, addi-
tional Western blot analyses were performed. These
revealed the deacetylase HD1B-I (also termed ZmRpd3,
see Lechner et al., 2000) protein level to be reduced whereas
� Blackwell Publishing Ltd, The Plant Journal, (2003), 33, 455–469
ZmGCN5, a plant nuclear HAT 461

deacetylases HD1B-II and HD2 remain relatively unaffected.
The induction of histone transcription and the downregula-
tion of the RPD3-type deacetylase (HD1B-I) protein there-
fore seems to be specific responses to a reduced Zmgcn5
transcript and protein level.
Discussion
Evidence for the role of ZmGCN5 in
co-activator-promoted transcription
There is compelling evidence to support a role for GCN5 in
promoting transcription in higher eukaryotes (Sterner and
Berger, 2000). The genetically isolated ZmGCN5 described
herein, is the first plant HAT which has been analysed for its
nuclear interactions and influence on overall cellular tran-
script profile. ZmGCN5 shows typical features of a member
of the GCN5 family, possessing the catalytic HAT-, ADA2-
interaction and bromo-domain, and being nuclear-loca-
lised. Southern blot hybridisation indicates the presence
of only one copy of Zmgcn5 in the maize genome, although
the possibility of more distantly related homologues which
do not cross-hybridise, cannot be eliminated. Indeed, in
A. thaliana, three gcn5-related sequences have been
detected, and yeast also has more than one type-A HAT,
although these appear to have discrete functions (Clarke
et al., 1999). By in situ hybridisation, Zmgcn5 transcripts
could only be detected in the chalazal region of the endo-
sperm cavity, surrounding the embryo, suggesting expres-
sion is concentrated in transcriptionally active cells. This
observation is supported by the ubiquitous expression
pattern, but with higher transcript abundance in tissues
rich in dividing cells.
The functional properties of ZmGCN5 we have observed
indicate a key role in modulating the acetylation status of
chromatin in plants. As expected for a type-A HAT, the
protein is transported via an NLS sequence to the nucleus.
In contrast to ScGCN5, the NLS employed is not located in
the central portion of the polypeptide, but is close to the
Figure 6. TSA treatment influences histoneacetylation and ZmGCN5 protein level.(a) Histone H4 acetylation increases proportion-ally after TSA treatment. The relative amount ofhistone proteins in TSA-treated and untreatedHE-89 cell lines was visualised after acid extrac-tion of histone fractions, resolved on 18% SDS–PAGE and visualised by Ponceau S staining. H4levels were detected by specific histone H4 anti-body raised against acetylisable positions 5, 8,12 and 16. As a loading control, an antibodyraised against maize immunophilin (66 kDa)was used (Hueros et al., 1998). The numbersbelow give the ratios of ZmGCN5/Immunophilinafter quantification of the ECL reaction.(b) Effect of TSA on ZmGCN5 protein level. TheZmGCN5 antibody was used to detect the pro-tein in the maize cell line HE-89 with and withoutTSA treatment. As a loading control the maizeimmunophilin antibody was used.
� Blackwell Publishing Ltd, The Plant Journal, (2003), 33, 455–469
462 Riyaz A. Bhat et al.

Figure 7. ESTs showing major changes in tran-script levels on Trichostatin A (TSA) treatment.Maize cell line HE-89 was treated with TSA andtotal RNA was extracted from treated and un-treated lines. The radiolabelled cDNA generatedwas used for microarray hybridisations.(a) ESTs with increasing signal intensities.(b) Sequences showing reduced signal intensi-ties.(c) RNA gel blot confirmation for selected arrayclones modulated by TSA treatment. HE-89maize cell line was treated with 5 mM TSA for10–12 h. Fifty micrograms of total RNA fromtreated and untreated material was blotted ona nylon membrane and hybridised with selectedarray clones.
� Blackwell Publishing Ltd, The Plant Journal, (2003), 33, 455–469
ZmGCN5, a plant nuclear HAT 463

Figure 8. Zmgcn5 antisense lines show an increase in histone transcript abundance.(a) Northern analysis of Zmgcn5 antisense lines. Total RNA from vector transformed material and two antisense lines was analysed using the Zmgcn5 cDNA as a probe and ubiquitin as loading control.(b) ESTs showing increased signal intensities in the Zmgcn5 antisense material (line 4).(c) Sequences with reduced signal intensities.(d) HD1B-I, HD1B-II and ZmHD2 deacetylase protein levels in Zmgcn5 antisense lines were detected by immunoblot analysis with specific deacetylase antibodies. The HD1B antibody detects HD1B-I and HD1B-II.
�B
lackw
ell
Pu
blish
ing
Ltd
,T
he
Pla
nt
Jo
urn
al,
(2003),
33,
455–4
69
464
Riy
az
A.
Bh
at
et
al.

N-terminus in a stretch non-homologous with other HAT
sequences. Nuclear localisation signals are often redun-
dant, and it may be that during evolution a site additional to
the yeast sequence emerged, permitting loss of function of
the original NLS via genetic drift.
The enzymatic activity of ZmGCN5 was demonstrated
using the recombinant GST-fusion protein which under
in vitro conditions acetylates free histones but not nucleo-
somes, as reported for other GCN5-type HATs. Acetylation of
histones in nucleosomes requires further component(s) of
the GCN5-coactivator complex. Addition of ZmADA2 was
not sufficient to promote acetylation of nucleosomes by Zm-
GCN5 (unpublished observations), although the interaction
with ZmADA2 could be demonstrated in yeast, and in vitro.
Therefore, other component(s) of the co-activator com-
plexes seem to be required for acetylation of nucleosomes.
The biological role of GCN5-containing complexes may
be difficult to establish from gene knockout phenotypes, for
example, if these are lethal, or if there is redundancy in
functional copies of the coding sequence. In order to estab-
lish a role for GCN5 in maize, we investigated its interaction
with the endosperm-specific transcription factor, O2. We
were unable to show an interaction between O2 and either
ZmGCN5 or ZmADA2 in vitro. Most previous reports indi-
cated ADA2 rather than GCN5 to be the co-activator com-
ponent that interacts directly with the activation domain of
transcription factors. In fact, two Arabidopsis ADA2 pro-
teins have been shown to interact in vitro with the tran-
scription factor CBF1 (Stockinger et al., 2001). However,
ZmADA2 did not interact directly with O2 in our in vitro
assay, although a ZmGCN5–ZmADA2 interaction did occur
under these conditions (Figure 5a).
On the assumption that the interaction might require the
association of more components of the co-activator com-
plex, we probed for interaction of in vitro-labelled O2 protein
with ZmADA2 in nuclear extracts by co-immunoprecipitation
with the ADA2 antiserum. The interaction observed (Fig-
ure 5b) is consistent with the hypothesis that additional com-
ponentsarerequired forZmADA2tostablyassociatedirectly
or indirectly with O2, although it is also possible that apost-
translational modification of ZmADA2 might be necessary.
Contribution of ZmGCN5 to overall nuclear histone
acetylation in maize
To address the biological role of ZmGCN5 in chromatin
modification, we tested the cellular response to changes in
the acetylation status. Interestingly, hyperacetylation
caused by the deacetylase inhibitor TSA and hypoacetyla-
tion generated by the reduction of the endogenous
ZmGCN5 in the antisense lines seem to evoke similar
cellular answers. Firstly, upon changes in the degree of
acetylation, the expression of histone acetylases and/or
deacetylases is affected, presumably as a compensatory
response. Hyperacetylation caused by TSA results in a
reduction in the ZmGCN5 protein. Hypoacetylation in the
ZmGCN5 antisense lines reduces the abundance of
ZmRpd3-type deacetylase protein as well. Cellular mecha-
nisms thus seem to balance acetylation changes by directly
regulating the corresponding enzymes. A second response
to changes in the acetylation status is the increase in core
histone transcription. As depicted in Figure 9, our results
support a dynamic equilibrium between acetylation and
deacetylation processes that keeps an acetylation status
required for co-ordinated gene regulation. Trichostatin A
causing an increase in the degree of histone acetylation by
inhibiting histone deacetylases would lead to increased core
histone transcription. Newly synthesised histones are then
available for de novo acetylation. The overall histone protein
abundance, however, as observed experimentally is not
markedly altered. To redress the balance between acety-
lated and non-acetylated histones, therefore, a fraction of
the acetylated histones has to be preferentially degraded.
The ZmGCN5 knockout in contrast shifts the balance
towards hypoacetylation. Again histone de novo synthesis
and de novo acetylation in this case would enable the cell to
regain an acetylation status required for a working chro-
matin. Furthermore, here a reduction in the linker histone
H1 transcript signal was observed. Histone H1 is thought to
contribute to repression of transcription by increasing the
density of chromatin packaging (Wolffe and Hayes, 1999).
The fact that for H1 changes can only be observed in the
antisense material and not upon TSA treatment might be
due to a more specific cellular response in the antisense
situation. As the total amount of histones remains
unchanged, an increase in histone turnover seems to bal-
ance the equilibrium between acetylated and non- or under-
acetylated histones. The cellular regulation of chromatin
acetylation therefore comprises modifying enzymes as well
as the chromatin components themselves. The mechan-
isms by which the chromatin senses its own state and
regulates the transcription of specific modifying genes
are unknown and would warrant further investigations.
Experimental procedures
Plant material
Maize inbred lines A69Y, B73 and A188 were grown under green-house conditions and tissue samples collected by freezing in liquidnitrogen. The maize cell line HE89 was kindly provided by AventisCrop Sciences (Frankfurt), and cultured as recommended (Moroczet al., 1990).
RNA gel-blot analysis
Poly Aþ RNA was extracted from different maize tissues aspreviously described (Bartels and Thompson, 1983). The RNAwas separated on a 1% agarose/formaldehyde gel along with
� Blackwell Publishing Ltd, The Plant Journal, (2003), 33, 455–469
ZmGCN5, a plant nuclear HAT 465

appropriate molecular size markers (Gibco BRL), and transferredonto Hybond-N (Amersham-Pharmacia). Hybridisation was carriedout at 458C in the presence of 50% formamide with a 32P randomprimer-labelled probe comprising the full-length cDNA sequenceof Zmgcn5 or other sequences investigated.
Southern blot analysis
DNA was isolated from maize inbred lines B73 and A69, digestedwith BamHI, EcoRI and HindIII, and separated by electrophoresison a 0.75% agarose gel. The gel was blotted onto Hybond-N
Figure 9. Cellular response to inhibition of deacetylation by TSA and to reduction of acetylation in Zmgcn5 antisense material to restore an equilibrium extent ofacetylation.Upon TSA treatment, ZmGCN5 is downregulated and increased core histone transcription balances the amount of acetylated histones. In antisense lines withreduced ZmGCN5, the Rpd3-type HD1B-I deacetylase protein amount is downregulated and de novo synthesis of core histones again regulates the degree ofacetylation.
� Blackwell Publishing Ltd, The Plant Journal, (2003), 33, 455–469
466 Riyaz A. Bhat et al.

(Amersham-Pharmacia). Hybridisation was carried out at 658C inthe presence of 10% dextran sulphate and 1% SDS with a 32Prandom primer-labelled probe containing the full-length cDNAsequence of Zmgcn5.
Western analysis
Western analysis was performed as described earlier (Serna et al.,2001). The ZmGCN5 antibody is directed against the C-terminal 206amino acids of the protein. The ZmADA2 antibody is directedagainst the full-length protein (for immunophilin antibody, seeHueros et al., 1998). The antiacetylated Histone 4 antibody recog-nises acetylated lysines 5, 8, 12 and 16 at the histone N-termini(Serotec). Antideacetylase antibodies, HD1B and HD2, were kindlysupplied by P. Loidl (University of Innsbruck).
Histone acetyltransferase assays
For determination of enzyme activity, the enzyme sample (either5 mg of recombinant ZmGCN5 protein, 10 mg maize nuclear extract,or 10 mg chromatin fraction (kindly provided by P. Loidl, UniversityInnsbruck)), was incubated for 30 min at 288C with histones ornucleosomes, in a total volume of 100 ml, containing 50 mM Tris–HCl pH 8.0, 10% glycerol, 1 mM DTT, 10 mM sodium butyrate, and0.2 mCi 3H-Acetyl CoA (5–6 Ci mmol�1). The reaction products wereelectrophoresed on 12% SDS–PAGE and the histones were visua-lised by Coomassie blue staining. 3H-incorporation was detectedby fluorography of the fixed gel. The substrates used for acetyla-tion were: 25 mg calf thymus histones (typ IIA, Sigma), 100 mg maizenucleosomes (Moehs et al., 1988), or 25 mg histones isolated frommaize nucleosomes (Simon and Becker, 1976).
Yeast two-hybrid screen
For two-hybrid analysis the GAL4 system based on the HybriZAP2.1 vector from Stratagene was used. As bait construct, the GAL4DNA-binding domain was fused to an Atada2 sequence lacking thefirst 87 bp. The resulting construct was used to screen a 7 DAPmaize endosperm cDNA library fused to the GAL4 transcriptionalactivation domain (AD). The screen was performed on SD-mediumlacking the amino acids leucine, tryptophan and histidine, in thepresence of 15 mM 3-AT using the yeast reporter strain YRG-2.Colonies were re-screened by re-streaking them on 20 mM 3-ATand by testing for LacZ expression.
Analysis of GFP fusions by transient expression
Full-length (1.54 kb) cDNA of Zmgcn5 was PCR amplified introdu-cing NcoI sites at both ends. The resulting fragment was insertedinto the NcoI site of the vector pGFP-JS (codon optimised versionof GFP, Sheen et al., 1995) to make a C-terminal fusion of Zmgcn5with gfp.
Tobacco cell line BY2, derived from Nicotiana tabacum L. cv.Bright Yellow 2 was propagated and used for protoplast isolationaccording to Nagata et al. (1981). Five millilitres of the 7-day-oldstationary culture was transferred to 95 ml of the fresh culturemedium. After 3 days of culture, cells in the exponential phasewere harvested by passing through a 100-mm sieve and used forprotoplast preparation.
Transfection of BY2 protoplasts was performed according toNegrutiu et al. (1987) with some minor modifications. 10–20 mg ofDNA was mixed with 0.3 ml aliquots of freshly isolated protoplasts
(about 2� 106), followed by mixing with 0.6 ml of PEG solution(25% PEG 1500, 0.1 M MgCl2.6H2O, 0.45 M mannitol, 0.02 M
Hepes, pH¼ 6.0). The PEG–protoplast mixture was incubatedfor 20 min at room temperature and diluted by adding 10 ml ofW5 (154 mM NaCl, 125 mM CaCl2.2H2O, 5 mM KCl, 5 mM sucrose,pH 5.6–6.0). Protoplasts were centrifuged at 100 g for 5 min andthe pellet was re-suspended in 10 ml of K3 (MS base with macroand micro elements, 0.4 M sucrose, 100 mg l�1 inositol, 1 ml l�1
MS vitamins, 250 mg l�1 xylose, pH 5.6). Transfected protoplastswere incubated at 288C in the dark. Sample aliquots for transientGFP fluorescence were taken 18–24 h after the treatment of theprotoplasts.
Protoplasts were examined using either a Leica DMRB or ZeissAxiophot microscope equipped with epifluorescence optics. Greenfluorescent protein specific fluorescence was observed using HQGFP, HQ GFP LP filters (blue light exciter BP 450–490 nm; beamsplitter RKP 510 nm; emitter LP 520 nm).
In situ hybridisation
In situ hybridisation was carried out as previously described (Sernaet al., 2001) using digoxygenin-labelled antisense and senseprobes.
GST spin down experiment
35S-labelled ZmGCN5 protein was prepared by in vitro translationof 50 ng of in vitro transcribed and capped RNA in the presence of35S-methionine using the wheat germ transcription and translationsystem (Promega). For pull-down experiments, 200 ml of recombi-nant ZmAda2–GST protein or GST protein alone was coupled for30 min at 48C to 20 ml of glutathione–agarose beads (Sigma). Thebeads were washed four times, each with 500 ml binding buffer(150 mM NaCl, 20 mM Tris–HCl pH 7.5, 0.1% Igepal Ca 630). Thecoupled proteins were incubated with 5 ml of the in vitro translationreaction mix for 1 h at 48C, and the beads were again washed threetimes, each with 500 ml binding buffer. Bound proteins were elutedin 50 ml SDS sample buffer, separated by electrophoresis on 10%SDS–PAGE and detected by fluorography.
Co-immunoprecipitation
Maize nuclear extract was prepared from 15 g of 15 DAP old maizekernels. After homogenising using a Warring blender, nuclei wereconcentrated by floating on a 2.5-M sucrose cushion and washingwas performed as described (Luthe and Quatrano, 1980). Finally,the nuclei were re-suspended in 1 ml 40 mM HEPES, 350 mM NaCl,10% Glycerol, 1 mM PMSF and 1� plant proteinase inhibitors (PI,Sigma P9599), sonified for 30 sec using a Labson 2000 sonifier athigh intensity (50%) and incubated for 30 min at room tempera-ture with 10 ml DNaseI (from a fresh prepared stock solution of1 mg ml�1, Roche 104159). After centrifugation for 1 min at 3500 g,the supernatant was used for the in vitro binding reaction. The35S-labelled O2 protein was prepared by in vitro transcriptionusing the Pharmacia T-kit and in vitro translation using the wheatgerm system (Promega). The binding reaction was carried out in1 ml of co-immunoprecipitation buffer (50 mM Tris–Cl pH 7.9,150 mM NaCl, 5 mM MgCl2, 1 mM PMSF, 1� PI, 1 mM DTT, 1%NP-40) for 30 min at room temperature (modified from Le Gour-rierec et al., 1999). Afterwards, the interaction was stabilised for30 min at 48C. Each reaction with the presence of different com-ponents contained about 50 ml protein-A sepharose slurry, 50 mlnuclear extract, 20 ml 35S-O2 (transcription and translation product,
� Blackwell Publishing Ltd, The Plant Journal, (2003), 33, 455–469
ZmGCN5, a plant nuclear HAT 467

corresponding to about 150 ng linearised plasmid), 3.5 ml specificZmADA2 antibody or 3.5 ml pre-serum. After binding, the protein-A sepharose beads were spun down at 3500 g for 1 min andwashed three times in 1 ml binding buffer at 48C. Proteins boundto protein-A sepharose were eluted by boiling in 1� Laemmlibuffer and separated by 10% SDS–PAGE. As a control, 3 ml of 35S-O2 were directly loaded. Finally, the gel was subjected to fluor-ography.
Trichostatin A treatment of maize cell line HE-89
Maize suspension cell line HE-89 was cultured weekly as recom-mended (Morocz et al., 1990). For TSA treatment, rapidly dividingcells, 3 days after the subculture, were treated with different con-centrations of TSA (0.5–5.0 mM) and cultured further for 10–12 hunder the same conditions. After the treatment, the cells werepelleted and frozen at �708C. RNA and protein preparations weremade as described (Serna et al., 2001).
Maize transformation, transgenic callus and
suspension cell lines
Transformation of maize suspension cell line HE-89 was kindlycarried out by Anja Horold, Monocot Transformation Group, MPIZ(Koln) and by Maize transformation group, Aventis Crop Sciences(Frankfurt). The transformation procedure was based on the pro-toplast PEG uptake method (Morocz et al., 1990). For selection oftransformants, BASTA resistance was used. After establishingtransgenic callus lines, they were maintained in suspension cul-ture, selecting for resistance to BASTA.
Reverse transcription, labelling and hybridisation
of array filters
Thirty micrograms of total RNA isolated from TSA-treated andnon-treated tissue culture material were labelled in the presence of500 ng oligo-dT Primer, 30 mCi 33P-adCTP and Superscript II reversetranscriptase (Gibco-BRL). After first strand cDNA synthesis, theRNA was hydrolysed at 658C for 30 min in presence of 1% SDS,0.5 M EDTA and 3 M NaOH. Following the neutralisation with 2 NHCl, the cDNA was pelleted down using 3 M Na-acetate, pH 5.3, and10 mg ml�1 t-RNA as a carrier. The incorporation efficiency wasmeasured using a Beckman scintillation counter. Hybridisation ofthe filters was carried out at 658C in Church buffer (0.5 M Na-phosphate pH 7.2, 7% (w/w) SDS, 1 mM EDTA pH 8.0) for at least10 h. Following several washes in 40 mM Na-phosphate pH 7.2 and0.1% SDS, the filters were exposed on a phosphor imager screenover night. (Hoheisel et al., 1991). For each experimental conditionthe hybridisations were repeated three times. Every hybridisationwas performed with three independent replica filters. Images weredeveloped using the Storm 860 scanner (Molecular Dynamics) andexported to Array Vision software (Imaging research Inc.) foranalysis.
Acknowledgements
We thank P. Loidl, A. Lusser and A. Pipal (University Innsbruck) forsupplying maize HAT protein fractions, HD1B and HD2 antibodiesand advice. We are grateful to MPIZ-ADIS staff for assistance inarray preparation and analysis. The work was supported in part bythe Deutsche Forschungsgemeinschaft.
References
Amedeo, P., Habu, Y., Afsar, K., Scheid, O.M. and Paszkowski, J.(2000) Disruption of the plant gene MOM releases transcrip-tional silencing of methylated genes. Nature, 405, 203–206.
Barlev, N.A., Candau, R., Wang, L., Darpino, P., Silverman, N.and Berger, S.L. (1995) Characterization of physical interactionsof the putative transcriptional adaptor, ADA2, with acidic activa-tion domains and TATA-binding protein. J. Biol. Chem. 270,19337–19344.
Bartels, D. and Thompson, R.D. (1983) The characterization ofcDNA clones coding for wheat storage proteins. Nucl. AcidsRes. 11, 2961–2977.
Berger, S.L., Pina, B., Silverman, N., Marcus, G.A., Agapite, J.,Regier, J.L., Triezenberg, S.J. and Guarente, L. (1992) Geneticisolation of ADA2: a potential transcriptional activator requiredfor function of certain acidic activation domains. Cell, 70,251–265.
Chua, Y.L., Brown, A.P. and Gray, J.C. (2001) Targeted histoneacetylation and altered nuclease accessibility over short regionsof the pea plastocyanin gene. Plant Cell, 13, 599–612.
Clarke, A.S., Lowell, J.E., Jacobson, S.J. and Pillus, L. (1999) Esa1pis an essential histone acetyltransferase required for cell cycleprogression. Mol. Cell. Biol. 19, 2515–2526.
Grant, P.A., Duggan, L., Cote, J. et al. (1997) Yeast Gcn5 functionsin two multisubunit complexes to acetylate nucleosomal his-tones: characterization of an Ada complex and the SAGA (Spt/Ada) complex. Genes Dev. 11, 1640–1650.
Hartings, H., Maddaloni, M., Lazzaroni, N., Di Fonzo, N., Motto, M.,Salamini, F. and Thompson, R. (1989) The O2 gene whichregulates zein deposition in maize endosperm encodes a proteinwith structural homologies to transcriptional activators. EMBOJ. 8, 2795–2801.
Hoheisel, J.D., Lennon, G.G., Zehetner, G. and Lerach, H.(1991) Use of high coverage reference libraries of Drosophilamelanogaster for relational data analysis. J. Mol. Biol. 220,903–914.
Hueros, G., Rahfeld, J., Salamini, F. and Thompson, R.D. (1998) Amaize FK506-sensitive immunophilin, mzFKBP-66, is a peptidyl-proline cis-trans isomerase that interacts with calmodulin and a36-kDa cytoplasmic protein. Planta, 205, 121–131.
Kakutani, T., Jeddeloh, J.A. and Richards, E.J. (1995) Character-ization of an Arabidopsis thaliana DNA hypomethylationmutant. Nucl. Acids Res. 23, 130–137.
Kolle, D., Sarg, B., Lindner, H. and Loidl, P. (1998) Substrate andsequential site specificity of cytoplasmic histone acetyltrans-ferases of maize and rat liver. FEBS Lett. 421, 109–114.
Le Gourrierec, J., Li, Y.F. and Zhou, D.X. (1999) Transcriptionalactivation by Arabidopsis GT-1 may be through interaction withTFIIA–TBP–TATA complex. Plant J. 18, 663–668.
Lechner, T., Lusser, A., Brosch, G., Eberharter, A., Goralik-Schra-mel, M. and Loidl, P. (1996) A comparative study of histonedeacetylases of plant, fungal and vertebrate cells. Biochim.Biophys. Acta, 1296, 181–188.
Lechner, T., Lusser, A., Pipal, A., Brosch, G., Loidl, A., Goralik-Schramel, M., Sendra, R., Wegener, S., Walton, J.D. and Loidl, P.(2000) RPD3-type histone deacetylases in maize embryos. Bio-chemistry, 39, 1683–1692.
Lusser, A., Eberharter, A., Loidl, A., Goralik-Schramel, M., Horn-gacher, M., Haas, H. and Loidl, P. (1999) Analysis of the histoneacetyltransferase B complex of maize embryos. Nucl. Acids Res.27, 4427–4435.
Lusser, A., Kolle, D. and Loidl, P. (2001) Histone acetylation: les-sons from the plant kingdom. Trends Plant Sci. 6, 59–65.
� Blackwell Publishing Ltd, The Plant Journal, (2003), 33, 455–469
468 Riyaz A. Bhat et al.

Luthe, D.S. and Quatrano, R.S. (1980) Transcription in isolatedwheat nuclei. Plant Phys. 65, 305–308.
Moehs, C.P., McElwain, E.F. and Spiker, S. (1988) Chromosomalproteins of Arabidopsis thaliana. Plant Mol. Biol. 11, 507–515.
Morocz, S., Donn, G., Nemeth, J. and Dudits, D. (1990) Animproved system to obtain fertile regenerants via maize proto-plasts isolated from a highly embryogenic suspension culture.Theor. Appl. Genet. 80, 721–726.
Nagata, T., Okada, K., Takebe, I. and Matsui, C. (1981) Delivery oftobacco mosaic virus RNA into plant protoplasts mediated byreverse-phase evaporation vesicles (liposomes). Mol. Gen.Genet, 184, 161–165.
Negrutiu, I., Shillito, R., Potrykus, I., Biasini, G. and Sala, F. (1987)Hybrid genes in the analysis of transformation conditions. PlantMol. Biol. 8, 363–373.
Ogas, J., Kaufmann, S., Henderson, J. and Somerville, C. (1999)PICKLE is a CHD3 chromatin-re-modelling factor that regulatesthe transition from embryonic to vegetative development inArabidopsis. Proc. Natl. Acad. Sci. 96, 13839–13844.
Rossi, V., Hartings, H. and Motto, M. (1998) Identification andcharacterization of an RPD3 homologues from maize (Z. maysL.) that is able to complement an rpd3 null mutant of Sacchar-omyces cerevisiae. Mol. Gen. Genet. 258, 288–296.
Ruiz-Garcia, A.B., Sendra, R., Pamblanco, M. and Tordera, V.(1997) Gcn5p is involved in the acetylation of histone H3 innucleosomes. FEBS Lett. 403, 186–190.
Schmitz, D., Lohmer, S., Salamini, F. and Thompson, R.D. (1997)The activation domain of the maize transcription factor Opaque-2 resides in a single acidic region. Nucl. Acids Res. 25, 756–763.
Serna, A., Maitz, M., O’Connell, T. et al. (2001) Maize endospermsecretes a novel antifungal protein into adjacent maternal tissue.Plant J. 25, 687–698.
Sheen, J., Hwang, S., Niwa, Y., Kobayashi, H. and Galbraith, D.W.(1995) Green fluorescent protein as a new vital marker in plantcells. Plant J. 8, 777–784.
Simon, J.H. and Becker, W.M. (1976) A polyethylene glycol/dextranprocedure for the isolation of chromatin proteins (histones and
non-histones) from wheat germ. Biochim. Biophys. Acta, 454,154–171.
Sterner, D.E. and Berger, S.L. (2000) Acetylation of histonesand transcription-related factors. Microbiol. Mol. Biol. Rev. 64,435–459.
Stockinger, E.J., Mao, Y., Regier, M.K., Triezenberg, S.J.and Thomashow, M.F. (2001) Transcriptional adaptor andhistone acetyltransferase proteins in Arabidopsis and theirinteractions with CBF1, a transcriptional activator involvedin cold-regulated gene expression. Nucl. Acids Res. 29,524–1533.
Sullivan, W.J. and Smith, C.K. (2000) Cloning and characterizationof a novel histone acetyltransferase homologue from the pro-tozoan parasite Toxoplasma gondii reveals a distinct GCN5family member. Gene, 242, 193–200.
Tian, L. and Chen, Z.J. (2001) Blocking histone deacetylation inArabidopsis induces pleiotropic effects on plant gene regulationand development. Proc. Natl. Acad. Sci. 98, 200–205.
Tse, C., Georgieva, E.I., Ruiz-Garcia, A.B., Sendra, R. and Hansen,J.C. (1998) Gcn5p, a transcription-related histone acetyltransfer-ase, acetylates nucleosomes and folded nucleosomal arraysin the absence of other protein subunits. J. Biol. Chem. 273,32388–32392.
Waterborg, J.H. (1990) Identification of five sites of acetylation inalfalfa histone H4. Biochemistry, 31, 6211–6219.
Wolffe, A.P. and Hayes, J.J. (1999) Chromatin disruption andmodification. Nucl. Acids Res. 27, 711–720.
Wu, K., Malik, K., Tian, L., Brown, D. and Miki, B. (2000a) Functionalanalysis of a RPD3 histone deacetylase homologue in Arabidop-sis thaliana. Plant Mol. Biol. 44, 167–176.
Wu, K., Tian, L., Malik, K., Brown, D. and Miki, B. (2000b) Functionalanalysis of HD2 histone deacetylase homologues in Arabidopsisthaliana. Plant J. 22, 19–27.
Xu, W., Edmondson, D.G. and Roth, S.Y. (1998) Mammalian GCN5and P/CAF acetyltransferases have homologous amino-terminaldomains important for recognition of nucleosomal substrates.Mol. Cell. Biol. 18, 5659–5669.
Accession numbers: AJ428542 (genomic), AJ428540 (cDNA).
� Blackwell Publishing Ltd, The Plant Journal, (2003), 33, 455–469
ZmGCN5, a plant nuclear HAT 469




















