
Anxiety induced by prenatal stress is associated with suppression of hippocampal genes involved in...
13
Anxiety induced by prenatal stress is associated with suppression of hippocampal genes involved in synaptic function Y. Bogoch,* Y. N. Biala, M. Linial* and M. Weinstock *Department of Biological Chemistry, Life Science Institute, The Hebrew University, Jerusalem, Israel Department of Pharmacology, School of Pharmacy, The Hebrew University, Jerusalem, Israel Abstract Exposure of pregnant women or animals to stress during a critical period of foetal brain development increases the like- lihood of anxiety, depression and learning deficits that are associated with structural alterations in the offspring hippo- campus. In this study, we report the effect of gestational stress in rats on anxiogenic behaviour and hippocampal gene expression of their 23-day-old female offspring. As the rat brain continues to develop after birth, we also used the pro- cedure of handling (H) during the first 10 days of life to reverse the anxiogenic behaviour of prenatally stressed (PS) rats. By means of micro-array analysis on hippocampal extracts, we found that the expression of about 6.1% of 9505 valid genes was significantly altered by prenatal stress (p < 0.05). Of these, 48% were over-expressed and 52% under-expressed. The latter included 300 genes that participate in axonal growth, regulation of ion channels and transporters, trafficking of synaptic vesicles and neurotransmitter release. About 30% of the genes that were down-regulated in PS rats were re- stored to control levels by H. These include genes that play a role in pre-synaptic organization and function. Our results provide a possible relationship between hippocampal gene expression and changes in behaviour resulting from prenatal stress. Keywords: anxiety, exocytosis, gene annotation, handling, micro-array, synapse. J. Neurochem. (2007) 101, 1018–1030. Several reports have linked prenatal stress to a greater incidence of anxiety, affective disorders, attention deficits and schizophrenia (Weinstock 1997; van Os and Selten 1998; Geddes 1999; Linnet et al. 2003; Maccari et al. 2003). In rodents and non-human primates, prenatal stress also increases the incidence of attention deficits (Schneider 1992) and induces anxiogenic and depressive-like behaviour, characterized by increased fear of novelty and impaired coping under adversity (Fride et al. 1986; Fride and Wein- stock 1988) and learned helplessness and anhedonia, respectively (Fride et al. 1986; Alonso et al. 1991; Schneider 1992; Keshet and Weinstock 1995; Milberger et al. 1997). A chronic anxiety state in association with excessive fear of novelty has been linked to alterations in the size of the lateral amygdaloid nucleus in humans (De Bellis et al. 2000) and prenatally stressed (PS) rats (Salm et al. 2004) and also in the size of the synaptic area of the mossy fibres in the dentate gyrus (Belzung 1992). PS non-human primates and rats also show a number of structural changes in the hippocampus, including a decrease in hippocampal synapses (Hayashi et al. 1998), the number of granule cells in the dentate gyrus (Schmitz et al. 2002) and in neurogenesis (Lemaire et al. 2000; Coe et al. 2003). Because a considerable amount of brain development occurs after birth in the rat, it is possible to influence the plasticity of the nervous system and behaviour by an early environmental manipulation known as handling (H). This procedure, first described by Levine et al. (1967), involves removal of the pups from the maternal nest for periods of 3–15 min during the first 10–14 days of life. Its effect on behaviour was detected when compared with that of rats Received May 25, 2006; revised manuscript received August 30, 2006; accepted November 21, 2006. Address correspondence and reprint requests to Marta Weinstock, Department of Pharmacology, School of Pharmacy, Hebrew University, Ein Kerem, Jerusalem 91120, Israel. E-mail: [email protected] Abbreviations used: CH, Control handled; CNH, Control non-han- dled; cRNA, complementary RNA; EPM, elevated plus maze; GO, gene ontology; H, Handling; PS, prenatally stressed; PSH, prenatally stressed handled; PSNH, prenatally stressed non-handled; RT-PCR, reverse transcriptase PCR; SNARE, soluble N-ethylmaleimide-sensitive factor receptor. Journal of Neurochemistry , 2007, 101, 1018–1030 doi:10.1111/j.1471-4159.2006.04402.x 1018 Journal Compilation ȑ 2007 International Society for Neurochemistry, J. Neurochem. (2007) 101, 1018–1030 ȑ 2007 The Authors
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Anxiety induced by prenatal stress is associated with suppression of hippocampal genes involved in...

Anxiety induced by prenatal stress is associated with suppressionof hippocampal genes involved in synaptic function
Y. Bogoch,* Y. N. Biala,� M. Linial* and M. Weinstock�
*Department of Biological Chemistry, Life Science Institute, The Hebrew University, Jerusalem, Israel
�Department of Pharmacology, School of Pharmacy, The Hebrew University, Jerusalem, Israel
Abstract
Exposure of pregnant women or animals to stress during a
critical period of foetal brain development increases the like-
lihood of anxiety, depression and learning deficits that are
associated with structural alterations in the offspring hippo-
campus. In this study, we report the effect of gestational
stress in rats on anxiogenic behaviour and hippocampal gene
expression of their 23-day-old female offspring. As the rat
brain continues to develop after birth, we also used the pro-
cedure of handling (H) during the first 10 days of life to reverse
the anxiogenic behaviour of prenatally stressed (PS) rats. By
means of micro-array analysis on hippocampal extracts, we
found that the expression of about 6.1% of 9505 valid genes
was significantly altered by prenatal stress (p < 0.05). Of
these, 48% were over-expressed and 52% under-expressed.
The latter included �300 genes that participate in axonal
growth, regulation of ion channels and transporters, trafficking
of synaptic vesicles and neurotransmitter release. About 30%
of the genes that were down-regulated in PS rats were re-
stored to control levels by H. These include genes that play a
role in pre-synaptic organization and function. Our results
provide a possible relationship between hippocampal gene
expression and changes in behaviour resulting from prenatal
stress.
Keywords: anxiety, exocytosis, gene annotation, handling,
micro-array, synapse.
J. Neurochem. (2007) 101, 1018–1030.
Several reports have linked prenatal stress to a greaterincidence of anxiety, affective disorders, attention deficitsand schizophrenia (Weinstock 1997; van Os and Selten 1998;Geddes 1999; Linnet et al. 2003; Maccari et al. 2003). Inrodents and non-human primates, prenatal stress alsoincreases the incidence of attention deficits (Schneider1992) and induces anxiogenic and depressive-like behaviour,characterized by increased fear of novelty and impairedcoping under adversity (Fride et al. 1986; Fride and Wein-stock 1988) and learned helplessness and anhedonia,respectively (Fride et al. 1986; Alonso et al. 1991; Schneider1992; Keshet and Weinstock 1995; Milberger et al. 1997). Achronic anxiety state in association with excessive fear ofnovelty has been linked to alterations in the size of the lateralamygdaloid nucleus in humans (De Bellis et al. 2000) andprenatally stressed (PS) rats (Salm et al. 2004) and also in thesize of the synaptic area of the mossy fibres in the dentategyrus (Belzung 1992). PS non-human primates and rats alsoshow a number of structural changes in the hippocampus,including a decrease in hippocampal synapses (Hayashi et al.1998), the number of granule cells in the dentate gyrus
(Schmitz et al. 2002) and in neurogenesis (Lemaire et al.2000; Coe et al. 2003).
Because a considerable amount of brain developmentoccurs after birth in the rat, it is possible to influence theplasticity of the nervous system and behaviour by an earlyenvironmental manipulation known as handling (H). Thisprocedure, first described by Levine et al. (1967), involvesremoval of the pups from the maternal nest for periods of3–15 min during the first 10–14 days of life. Its effect onbehaviour was detected when compared with that of rats
Received May 25, 2006; revised manuscript received August 30, 2006;accepted November 21, 2006.Address correspondence and reprint requests to Marta Weinstock,
Department of Pharmacology, School of Pharmacy, Hebrew University,Ein Kerem, Jerusalem 91120, Israel. E-mail: [email protected] used: CH, Control handled; CNH, Control non-han-
dled; cRNA, complementary RNA; EPM, elevated plus maze; GO, geneontology; H, Handling; PS, prenatally stressed; PSH, prenatally stressedhandled; PSNH, prenatally stressed non-handled; RT-PCR, reversetranscriptase PCR; SNARE, soluble N-ethylmaleimide-sensitive factorreceptor.
Journal of Neurochemistry, 2007, 101, 1018–1030 doi:10.1111/j.1471-4159.2006.04402.x
1018 Journal Compilation � 2007 International Society for Neurochemistry, J. Neurochem. (2007) 101, 1018–1030� 2007 The Authors

from which all contact with humans had been withheldduring the pre-weaning period (Levine et al. 1967; Ader1970). In later studies, H was found to prevent thehyperactivity induced by isolation stress in adult rats(Gentsch et al. 1988) and the increased fear of novelty inPS rats (Wakshlak and Weinstock 1990).
The aim of the present study was to identify the molecularprocesses in the hippocampus underlying anxiety induced byprenatal stress. As the incidence of anxiety and depressionand alterations in hippocampal structure are more prevalentin females (Alonso et al. 1991; Kuehner 1999; Schmitz et al.2002; Gue et al. 2004; Zhu et al. 2004), we looked forchanges induced by prenatal stress in genes and proteins inrats of this gender. To enable us identify the gene changesthat could be linked to anxiogenic behaviour, we determinedwhether any of them were reversed by H.
Materials and methods
Animals and treatments
All experiments were carried out according to the guidelines of the
University Committee for Institutional Animal Care, based on those
of the National Institutes of Health, USA. Female pathogen-free
(SPF) Wistar rats weighing 280–300 g (Harlan Biotech, Jerusalem,
Israel) on day 1 of pregnancy (detected by the presence of a vaginal
plug) were randomly allocated to stress and control (10 rats each)
groups, as previously described (Poltyrev and Weinstock 2004).
Maternal treatmentFrom day 17 to day 22 of gestation, rats were stressed daily in a
random order by three different stressors, restraint, forced swim and
saline injections once on each of 2 of 6 days, to prevent the rats
from adapting to them. Stress was applied to the rats during this
period because it is when key areas of the limbic system develop
(Bayer et al. 1993). Control pregnant females were left undisturbed
in their home cages. The length of gestation was 22.5 days.
Postnatal treatmentAfter birth, all litters were culled to eight pups (no more than three of
one sex and five of the other). From day 1 to day 10 of age, the pups
from half the control and half the stressed litters were handled by
removing them from the maternal cage for only 3 min as described
in Wakshlak and Weinstock (1990). As shown in our earlier study,
this manoeuvre does not significantly affect the behaviour of
controls, if they are handled in the usual manner by experimenters.
The pups were weaned at the age of 21 days and housed in groups of
two to three, according to sex and prenatal treatment. All subsequent
experiments described in the following paragraphs were performed
on the female offspring shortly after weaning in order to avoid the
effect of additional environmental factors on gene expression.
Anxiogenic behaviour
Anxiogenic behaviour was tested in the elevated plus maze (EPM)
at the age of 12 weeks in 11–12 females of each group of control
non-handled (CNH), PS non-handled (PSNH), control handled (CH)
and PS handled (PSH) rats, as previously described (Poltyrev and
Weinstock 2004). The EPM has two open and two closed arms
presenting the rat with a conflict between the desire to explore a
novel situation and its fear of height and open spaces (Handley and
Mithani 1984). Anxiolytic agents like diazepam increase, and
anxiogenic agents decrease the time spent in the open arms of the
maze (Pellow and File 1986). It has been shown that PS rats make
fewer entries into, and spend less time in the open arms of the maze
than controls (Fride and Weinstock 1988; Vallee et al. 1997). Thisbehaviour is consistent with increased anxiety, can be detected more
readily in females at the age of 23–30 days (Fig. 1) and persists into
adulthood (Zagron and Weinstock, unpublished data).
Gene expression in hippocampus
At the age of 23 days, three rats were chosen randomly from each
group of CNH, PSNH, CH and PSH litters. They were decapitated
and the whole hippocampi rapidly dissected out, snap frozen in liquid
nitrogen and then stored at –70 C until RNA isolation. Total RNA of
hippocampus was prepared with Triazol and subsequently cleaned by
the Rneasy Mini Kit (Qiagen, Valencia, CA, USA). RNA quality and
quantity were measured by spectrophotometer with A260/A280 of
1.85–2.1. The RNA was prepared for hybridization using the
protocol recommended by Affymetrix (Santa Clara, CA, USA).
Briefly, total RNA (10 lg) was first reverse transcribed using a T7-
Oligo (O’Donovan et al. 1999). Following Rnase H-mediated
second-strand cDNA synthesis, double-stranded cDNAwas purified
to serve as a template in the subsequent in vitro transcription reaction.The reaction was carried out in the presence of T7 RNA polymerase
and a biotinylated nucleotide analogue and ribonucleotide mix for
complementary RNA (cRNA) amplification and biotin labelling
(BioArray Labeling Kit; Enzoi Diagnostics, Farmingdale, NY, USA).
The biotinylated cRNA targets were cleaned up, fragmented and
hybridized to GeneChip expression arrays (Affymetrix) RAE 230A.
After washing, the chips were stained with a streptavidin–phyco-
erythrin conjugate for fluorescence detection. Hybridization on a test
chip and analysis for the quality of the labelled cRNA were
performed for each of the cRNA samples prior to the hybridization
experiment. Ratios of 3¢/5¢ for glyceraldehyde-3-phosphate dehy-
drogenase and ß-actin, used as a measure of the quality of the RNA
and not as a standard, were confirmed to be within acceptable limits
as recommended by Affymetrix. BioB spike controls were found to
be present on all the arrays. When scaled to a target intensity of 150
Males Females 0
20
40
60
80
100
120
Tim
e in
ope
n ar
ms
(s)
*
Fig. 1 Time spent by young PS and control male and female rats in
the open arms of the EPM. Open bars: control; Grey bars: PS group.
*Significantly different from control, p < 0.01. Each experimental group
comprises eight rats.
Prenatal stress, anxiety and hippocampal genes 1019
� 2007 The AuthorsJournal Compilation � 2007 International Society for Neurochemistry, J. Neurochem. (2007) 101, 1018–1030

(using Affymetrix Microarray Suite 5.0 array analysis software),
scaling factors for all arrays were found to be within acceptable limits
(1.5–2.8), as were the background, Q values and mean intensities.
Reverse transcriptase PCR
Reverse transcriptase (RT)-PCR was performed to validate the
changes in a number of genes from the micro-array using RNA that
was identical to that used for the GeneChip experiment and RNA
prepared from animals taken from the same groups. All reactions were
performed on the same cDNA preparation. For each reaction, the
number of PCR cycles was optimized to match the dynamic range of
the reaction (22–32 cycles). All reaction products were designed to
range between 190 and 270 nucleotides. Pairs of oligonucleotides for
two genes with a difference in amplimer length larger than 50
nucleotides were included in each reaction to reduce experimental
variation. Each set of oligonucleotides was tested at least twice.
Samples were treated for 15 min with 10 U of Rnase-free Dnase
(Qiagen) at 25 �C prior to the RT reaction. The relative amounts of
mRNA were normalized to L19 (housekeeping gene of the large
subunit of the ribosome). Oligonucleotide primers used are for
forward (F) and reverse (R) for the following genes: Stathmin like-2 F:
cggaagctccacgaactcta, R: tagcctcacggttttcctta; SV2b F: aactcccaag-
caaatggatg, R: atcctggaaataccggatca, Rim1 F: cgtcgtcggacacctaagag,
R: tgccctggtaagacttgtg; Synaptotagmin 5 F: gtctgaagaaacggaagacct,
R: cccttggtaaagctgcttag; L19 F: ctgaaggtcaaagggaatgtg, R: ggacagag-
tcttgatgatctc; Rab3a F: cgccagcttgtctcagtttag, R: ggctgtggtgatggttcgg-
ta; Staufen F: tgctgcagaagctatgctgt, R: gcctagagttgtgccagagg.
Western blot
Following separation by sodium dodecyl sulfate (SDS) – polyacryl-
amide gel electrophoresis, extracts of the whole hippocampus from
23-day-old littermates of the rats used for the gene arraywere prepared
for western analyses. Cell lysates were homogenized in solubilization
buffer [Tris 50 mmol/L, NaCl 150 mmol/L, 1% (v/v) NP-40]
supplemented with protease inhibitors (1 mmol/L phenyl methyl
sulphonyl fluoride, 5 lmol/L leupeptin, 10 lg/mL aprotinin and
10 mmol/L EDTA) on ice. The extracts (1 mL/g tissue) were
centrifuged (14 000 g, 20 min, 4�C) and used for separation on
12.5% SDS gels. Protein concentrations were determined by BCA
(Pierce, Rockford, IL, USA) and blotted on nitrocellulosemembranes.
Major synaptic proteins were detected by means of a set of
antibodies to synaptophysin (Sigma, St Louis, MO, USA), syna-
ptojanin (Chemicon, Temecula, CA, USA), synapsin (Chemicon),
synaptotagmin 1 (Alomone Labs, Jerusalem, Israel) and SNAP-25
(kindly contributed by N. Takahashi, Japan). Secondary antibodies
used to detect monoclonal and polyclonal antibodies were Goat amouse and Goat a rabbit coupled to HRP (Jackson, West Grove, PA,
USA). b-tubulin (Sigma) was used for normalization, as it was
unchanged in all three experimental groups. Each experiment was
repeated at least twice. Western blots were quantified and normal-
ized according to controls. Detection method used was enhanced
chemiluminescence based and quantification was performed by
means of ImageGauge.
Data analysis
Differences in behavioural parameters in the EPM between the four
groups of rats were analysed by ANOVA for factorsMATERNAL (stress
or control) and NEONATAL treatment (handled or non-handled).
Duncan’s post hoc test was appliedwhen appropriate, and a differenceof p < 0.05 was considered to be statistically significant.
Changes in gene expression were analysed by means of
Microarray Suite 5.0. We included genes that showed a differential
expression in intensity of >500. We considered genes to be valid for
analysis, if the gene probe was defined as Present (P) according to
Microarray Suite 5.0 for at least two of three samples in each
experimental group. Most (86%) of the genes were assigned P for all
three samples. Genes that did not have a signal intensity of at least
20 in all of the samples were disregarded. Significance of the
difference between CNH and PSNH, PSNH and PSH, and CNH and
CH was tested by Student’s t-test. Only genes that showed a
difference in expression greater than 1.25-fold and statistical
difference of p < 0.05 between groups were considered.
Functional annotation was performed by means of the statistical
model implemented in DAVID (Dennis et al. 2003) and GOTM(Zhang et al. 2004). DAVID is a web-based application that
translates the list of probes to functional annotations that integrate
data from the Gene Ontology (GO) Consortium (Camon et al.2004). These tools apply a statistical model based on annotation
sources (i.e. GO) to rank the coregulated functional categories
among the differentially expressed genes. Ranking was performed
according to a modified Fisher’s exact test referred to as ‘enrichment
score’ (Dennis et al. 2003). The p-value for the ‘enrichment score’
is the probability of finding a certain number of genes that have a
specific annotation by chance (given the number of genes in the set
with this annotation, the total number of genes in the database and
the number of genes in the database with this annotation). Changes
in gene expression between any pair of the experimental groups
were tested for enrichment in functional annotations. No such
functional enrichment was found among genes in the CH group
compared with CNH. Although all gene changes resulting from
prenatal stress were analysed, the major focus was on those that
showed a significantly reduced expression which was restored by H.
Significance of the difference among the various groups in the
levels of the proteins from the western blots of hippocampal extracts
was analysed by ANOVA.
Results
Effect of prenatal stress and handling on behaviour
in the EPM
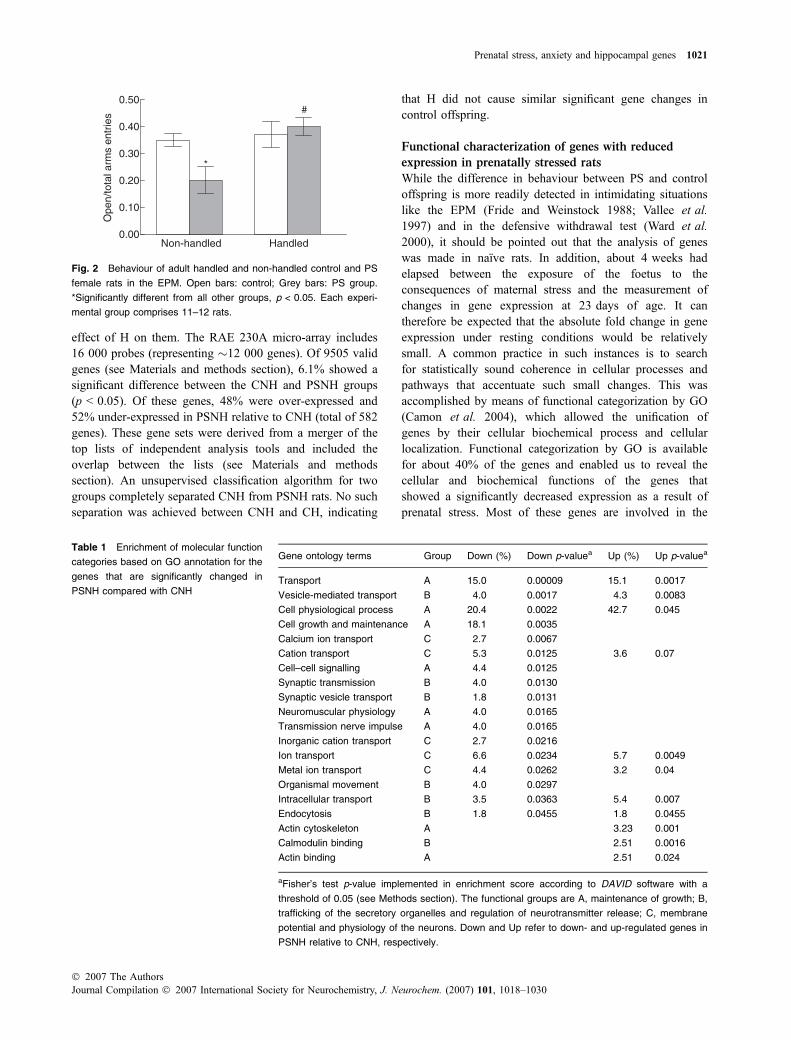
Adult PSNH displayed greater fear than CNH rats in theEPM. This was indicated by the shorter amount of time spentin the open arms and the smaller ratios of the number of open/total arms entries (Fig. 2). There was a significant interactionbetween maternal and neonatal treatment (F(1,43) = 4.61,p < 0.05), as H had a greater effect on PS than on control rats.A post hoc analysis showed that the ratio of open/total armsentries was significantly lower in PSNH rats than in eitherCNH or PSH (p < 0.05), and that there was no significantdifference between these ratios in CNH, PSH or CH rats.
General gene analysis
DNA GeneChip technology (Affymetrix) was used tocompare transcriptional profiles of PSNH and CNH and the
1020 Y. Bogoch et al.
Journal Compilation � 2007 International Society for Neurochemistry, J. Neurochem. (2007) 101, 1018–1030� 2007 The Authors
effect of H on them. The RAE 230A micro-array includes16 000 probes (representing �12 000 genes). Of 9505 validgenes (see Materials and methods section), 6.1% showed asignificant difference between the CNH and PSNH groups(p < 0.05). Of these genes, 48% were over-expressed and52% under-expressed in PSNH relative to CNH (total of 582genes). These gene sets were derived from a merger of thetop lists of independent analysis tools and included theoverlap between the lists (see Materials and methodssection). An unsupervised classification algorithm for twogroups completely separated CNH from PSNH rats. No suchseparation was achieved between CNH and CH, indicating
that H did not cause similar significant gene changes incontrol offspring.
Functional characterization of genes with reduced
expression in prenatally stressed rats
While the difference in behaviour between PS and controloffspring is more readily detected in intimidating situationslike the EPM (Fride and Weinstock 1988; Vallee et al.1997) and in the defensive withdrawal test (Ward et al.2000), it should be pointed out that the analysis of geneswas made in naıve rats. In addition, about 4 weeks hadelapsed between the exposure of the foetus to theconsequences of maternal stress and the measurement ofchanges in gene expression at 23 days of age. It cantherefore be expected that the absolute fold change in geneexpression under resting conditions would be relativelysmall. A common practice in such instances is to searchfor statistically sound coherence in cellular processes andpathways that accentuate such small changes. This wasaccomplished by means of functional categorization by GO(Camon et al. 2004), which allowed the unification ofgenes by their cellular biochemical process and cellularlocalization. Functional categorization by GO is availablefor about 40% of the genes and enabled us to reveal thecellular and biochemical functions of the genes thatshowed a significantly decreased expression as a result ofprenatal stress. Most of these genes are involved in the
Non-handled Handled 0.00
0.10
0.20
0.30
0.40
0.50 O
pen/
tota
l arm
s en
trie
s
*
#
Fig. 2 Behaviour of adult handled and non-handled control and PS
female rats in the EPM. Open bars: control; Grey bars: PS group.
*Significantly different from all other groups, p < 0.05. Each experi-
mental group comprises 11–12 rats.
Table 1 Enrichment of molecular function
categories based on GO annotation for the
genes that are significantly changed in
PSNH compared with CNH
Gene ontology terms Group Down (%) Down p-valuea Up (%) Up p-valuea
Transport A 15.0 0.00009 15.1 0.0017
Vesicle-mediated transport B 4.0 0.0017 4.3 0.0083
Cell physiological process A 20.4 0.0022 42.7 0.045
Cell growth and maintenance A 18.1 0.0035
Calcium ion transport C 2.7 0.0067
Cation transport C 5.3 0.0125 3.6 0.07
Cell–cell signalling A 4.4 0.0125
Synaptic transmission B 4.0 0.0130
Synaptic vesicle transport B 1.8 0.0131
Neuromuscular physiology A 4.0 0.0165
Transmission nerve impulse A 4.0 0.0165
Inorganic cation transport C 2.7 0.0216
Ion transport C 6.6 0.0234 5.7 0.0049
Metal ion transport C 4.4 0.0262 3.2 0.04
Organismal movement B 4.0 0.0297
Intracellular transport B 3.5 0.0363 5.4 0.007
Endocytosis B 1.8 0.0455 1.8 0.0455
Actin cytoskeleton A 3.23 0.001
Calmodulin binding B 2.51 0.0016
Actin binding A 2.51 0.024
aFisher’s test p-value implemented in enrichment score according to DAVID software with a
threshold of 0.05 (see Methods section). The functional groups are A, maintenance of growth; B,
trafficking of the secretory organelles and regulation of neurotransmitter release; C, membrane
potential and physiology of the neurons. Down and Up refer to down- and up-regulated genes in
PSNH relative to CNH, respectively.
Prenatal stress, anxiety and hippocampal genes 1021
� 2007 The AuthorsJournal Compilation � 2007 International Society for Neurochemistry, J. Neurochem. (2007) 101, 1018–1030

following processes: (i) neuronal growth and structuralplasticity; (ii) regulation of membrane potential, mainly ionchannels and transporters; (iii) trafficking of synapticvesicle and the neurotransmitter release machinery. Table 1shows the degree of statistical enrichment according to GO
Table 2 Genes showing reduced expression following PS that was
reversed by H
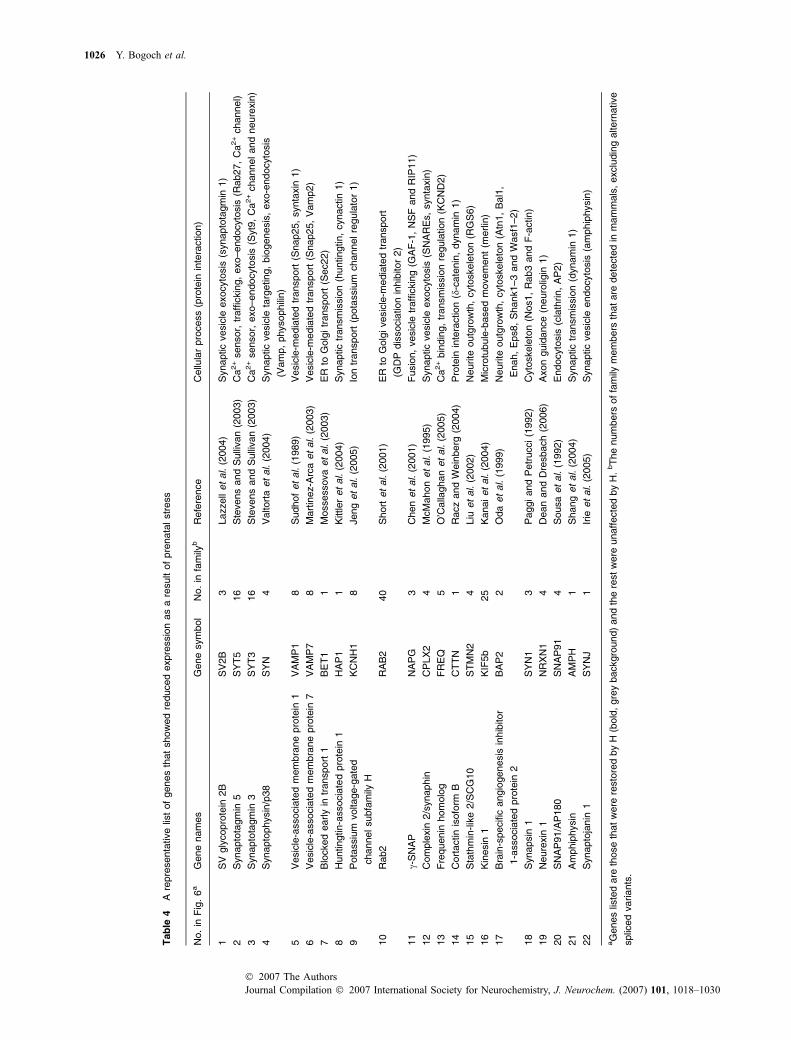
IDa Gene name
1372847_at ACN9 homolog (Saccharomyces cerevisiae)
(predicted)
1368587_at Apolipoprotein C-I
1369122_at Bcl2-associated X protein
1373081_at Brain-specific angiogenesis inhibitor 1-associated
protein 2
1368437_at Carbonic anhydrase 4
1374252_at Centrosomal protein 1 (predicted)
1371395_at Chromobox homolog 3 (HP1 gamma homolog,
Drosophila)
1373256_at Chromodomain helicase DNA-binding protein 3
(predicted)
1372132_at CNDP dipeptidase 2 (metallopeptidase M20 family)
(predicted)
1368584_a_at Complexin 2
1370123_a_at Cortactin isoform B
1372514_s_at Dynein, axonemal, light chain 4 (predicted)
1374016_at Endothelial differentiation, lysophosphatidic acid
GPCR, 2
1370503_s_at Erythrocyte protein band 4.1-like 3
1376578_at Euchromatic histone methyltransferase 1 (predicted)
1367766_at Expressed in non-metastatic cells 2
1372662_at F-box protein 34 (predicted)
1374103_at Frequenin homolog (Drosophila)
1373170_at G protein pathway suppressor 2 (predicted)
1367954_at Glial cell line-derived neurotrophic factor family
receptor alpha 1
1371392_at Glucose phosphate isomerase
1370240_x_at Haemoglobin alpha, adult chain 1
1369868_at Implantation-associated protein
1388240_a_at Integrin alpha 7
1373500_at Leucine-rich PPR-motif containing (predicted)
1375726_at LIM domain only protein 7
1367923_at Lipidosin
1390827_at MAD homolog 3 (Drosophila)
1371607_at Microtubule-associated protein 4
1374468_at Myeloid differentiation primary response gene 88
1369948_at Nerve growth factor receptor-associated
(TNFRSF16) protein 1
1374075_at NSF attachment factor, SNAP, gamma (predicted)
1368261_at Neurexin 3
1371812_at P55
1388076_at PAI-1 mRNA-binding protein
1370214_at Parvalbumin
1371455_at Phosphomannomutase 1 (predicted)
1373243_at Phosphomevalonate kinase (predicted)
1388847_at Positive cofactor 2, multiprotein complex, Q-rich-
associated protein
1370187_at Propionyl coenzyme A carboxylase, beta polypeptide
1370746_at Protein kinase, cAMP dependent, catalytic, beta
(predicted)
1373741_at Pseudouridine synthase 1 (predicted)
1369564_at Rad- and gem-related GTP-binding protein 2
Table 2 Continued
IDa Gene name
1367596_at Ribosomal protein S26
1372240_at Sarcoglycan, alpha (dystrophin-associated
glycoprotein) (predicted)
1368058_at Scaffold attachment factor B
1370116_at Septin 3
1371425_at Serine/arginine repetitive matrix 1 (predicted)
1371487_at SH3 domain binding glutamic acid-rich protein-like 3
(predicted)
1373605_at Similar to 106 kDa O-GlcNAc transferase-interacting
(predicted)
1377656_at Similar to 2600016B03Rik protein (predicted)
1374362_at Similar to 4930566A11Rik protein (predicted)
1373958_at Similar to ALEX3 protein
1373945_at Similar to bruno-like 5, RNA-binding protein
1372157_at Similar to CGI-143 protein (predicted)
1371399_at Similar to over-expressed breast tumour protein
1371863_at Similar to phosphatidylglycerophosphate synthase
(predicted)
1370925_at Similar to potential phospholipid-transporting
ATPase IIB
1372054_at Similar to sex-determination protein homolog Fem1a
1369160_a_at Solute carrier family 4, sodium bicarbonate
cotransporter, 7
1368391_at Solute carrier family 7 (cationic amino acid
transporter), 1
1370042_at Stathmin-like 2
1369251_a_at Synapsin I
1369628_at Synaptic vesicle glycoprotein 2b
1368276_at Synaptophysin
1373865_at Synaptosomal-associated protein, 91 kDa homolog
(mouse)
1369058_at Synaptotagmin 3
1368417_at Synaptotagmin 5
1370691_a_at Thyroid hormone receptor alpha
1371695_at Translocated promoter region (predicted)
1373495_at Ubiquitin-conjugating enzyme E2, J1 (predicted)
1367481_at Vacuolar protein sorting 28 (yeast) (predicted)
1368641_at Wingless-related MMTV integration site 4
1372483_at Zinc finger protein 469 (predicted)
1390649_at Zinc transporter ZnT-3
aID, the Affymetrix GeneChip ID. The list is sorted alphabetically. The
following probes showed a similar trend in their relative expression, but
are associated with un-annotated ESTs supported sequences:
1371321_at, 1371465_at, 1371512_at, 1371954_at, 1372623_at,
1375108_at, 1379073_at, 1372313_at, 1372901_at, 1371472_at,
1372719_at, 1374968_at, 1375192_at, 1376400_at, 1384204_at,
1390132_at, 1391437_at, 1367508_at.
1022 Y. Bogoch et al.
Journal Compilation � 2007 International Society for Neurochemistry, J. Neurochem. (2007) 101, 1018–1030� 2007 The Authors
annotations (p < 0.05) for genes that were decreased orincreased, respectively, by prenatal stress compare withcontrol.
Functional characterization of genes that showed
recovery of their levels of expression following handling
The expression of about 30% of the genes (total of 93 genes),which were down-regulated by prenatal stress (Table 2) andwere significantly up-regulated by H satisfied the statisticalcriteria (p < 0.05). By contrast, these same genes remainedunchanged in the CNH group.
These genes were reanalysed by GOTM (Zhang et al.2004), a tool that presents a subset of the GO hierarchy that isassociated with statistically enriched functional categories.The results of the analysis are shown in Table 3.
The enrichment in GO annotations for the genes that wereanalysed is shown by a directed acyclic graph based on thehierarchy of these annotations. These genes play a role intrafficking in the synapse and in the neurotransmitter releasemachinery (Fig. 3; Table 3). By contrast, other genes thatwere down-regulated and linked to growth and electricalproperties of neurons were not restored by H. (Table 1).
Table 3 Enriched functional categories
according to GOTM of genes, the expres-
sion of which was altered in PSNH and
restored by H
Go term Obsa Expb Ratioc p-valued
Down-regulated (93 genes)
Biological process
Transmission of nerve impulse 7 2.37 2.95 0.008718
Synaptic transmission 7 2.25 3.11 0.006539
Regulation of neurotransmitter levels 4 0.66 6.06 0.004077
Neurotransmitter secretion 3 0.43 6.98 0.008711
Membrane organization and biogenesis 3 0.21 14.29 0.001184
Secretory pathway 6 1.34 4.48 0.002005
Exocytosis 4 0.57 7.02 0.002403
Calcium ion-dependent exocytosis 2 0.14 14.29 0.008633
Vesicle-mediated transport 7 2.41 2.9 0.009429
Synaptic vesicle transport 3 0.3 10 0.003289
Secretion 6 1.75 3.43 0.007488
Molecular function
Death receptor binding 2 0.04 50 0.000460
SNARE binding 2 0.15 13.33 0.009689
Syntaxin binding 2 0.12 16.67 0.006595
Cellular component
Cell projection 5 1.11 4.5 0.004762
Neuron projection 3 0.42 7.14 0.008148
Axon 2 0.15 13.33 0.009680
Cytoplasmic vesicle 5 1.33 3.76 0.009904
Cytoplasmic membrane-bound vesicle 5 1.33 3.76 0.009904
Coated vesicle 5 0.82 6.1 0.001243
Clathrin-coated vesicle 5 0.72 6.94 0.000697
Synaptic vesicle 5 0.44 11.36 0.000070
Synaptic vesicle membrane 2 0.12 16.67 0.006588
Synapse 5 0.99 5.05 0.002856
Up-regulated (125 genes)
Biological process
Transmembrane receptor protein tyrosine
phosphatase signalling pathway
2 0.11 18.18 0.005
Transport 27 16.16 1.62 0.003
Localization 29 18.5 1.57 0.004
Establishment of localization 29 18.41 1.58 0.003
Molecular function
Transmembrane receptor protein
tyrosine phosphatase activity
2 0.1 20 0.004
Cellular component
None
aObserved gene number in the GO category; bExpected gene number in the GO category; cThe
Exp/Obs enrichment. dSignificance of enrichment for the GO category that show a significant
p-value of <0.01.
Prenatal stress, anxiety and hippocampal genes 1023
� 2007 The AuthorsJournal Compilation � 2007 International Society for Neurochemistry, J. Neurochem. (2007) 101, 1018–1030

Table 3 also shows the enrichment in GO annotations forthe 125 genes that were up-regulated by stress and reversedby H. As can be seen, they conform to only a small numberof GO terms that are not associated to any specific process,unlike those genes that were down-regulated by prenatalstress and reversed by H.
Relative expression of genes associated with the
pre-synaptic secretory apparatus
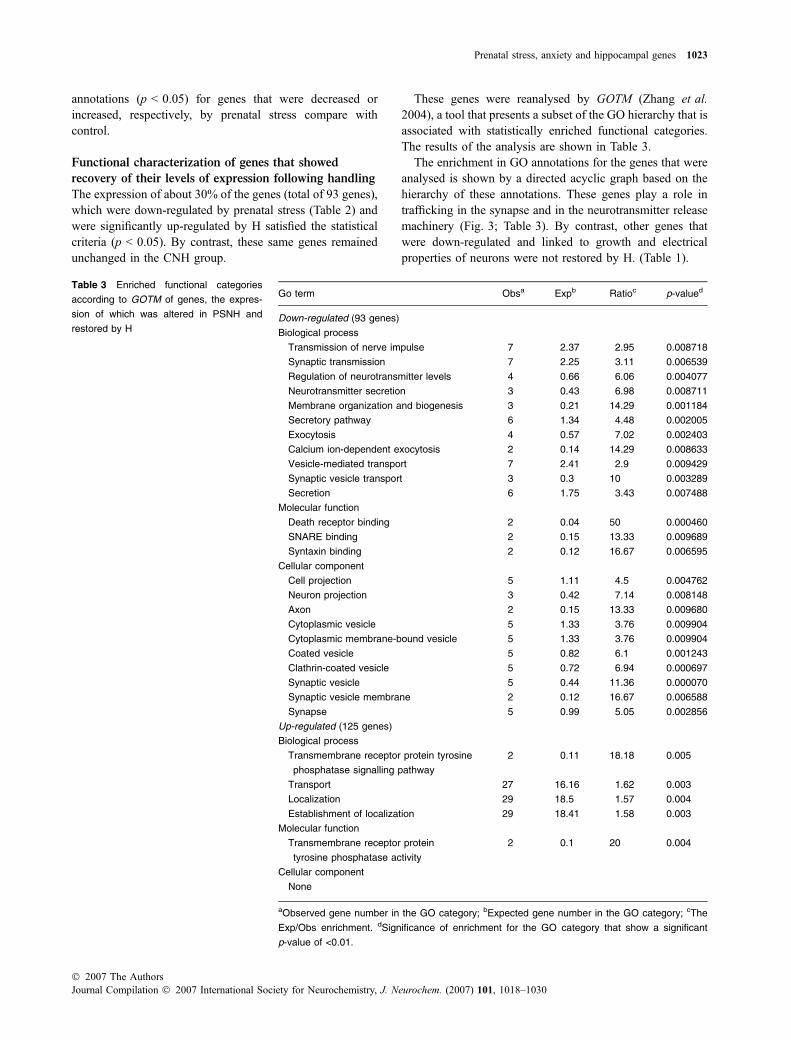
The prevalent functional category that was restored by H isassociated with the structure and function of the synapses.RT-PCR of various genes that have been linked to vesicletrafficking and organization of the pre-synaptic secretoryproteins, e.g. stathmin like-2, rab3A, SV2b and to a lesserextent synaptotagmin 5, validated the results obtained fromthe GeneChip, which were changed by prenatal stress and H,as well as those that were unchanged. The latter groupincluded genes that are involved in dendritic maturation(staufen) and in the regulation of the active zone (Rim1).Results of the RT-PCR reactions and the expression profilefor eight representative genes are shown (Fig. 4).
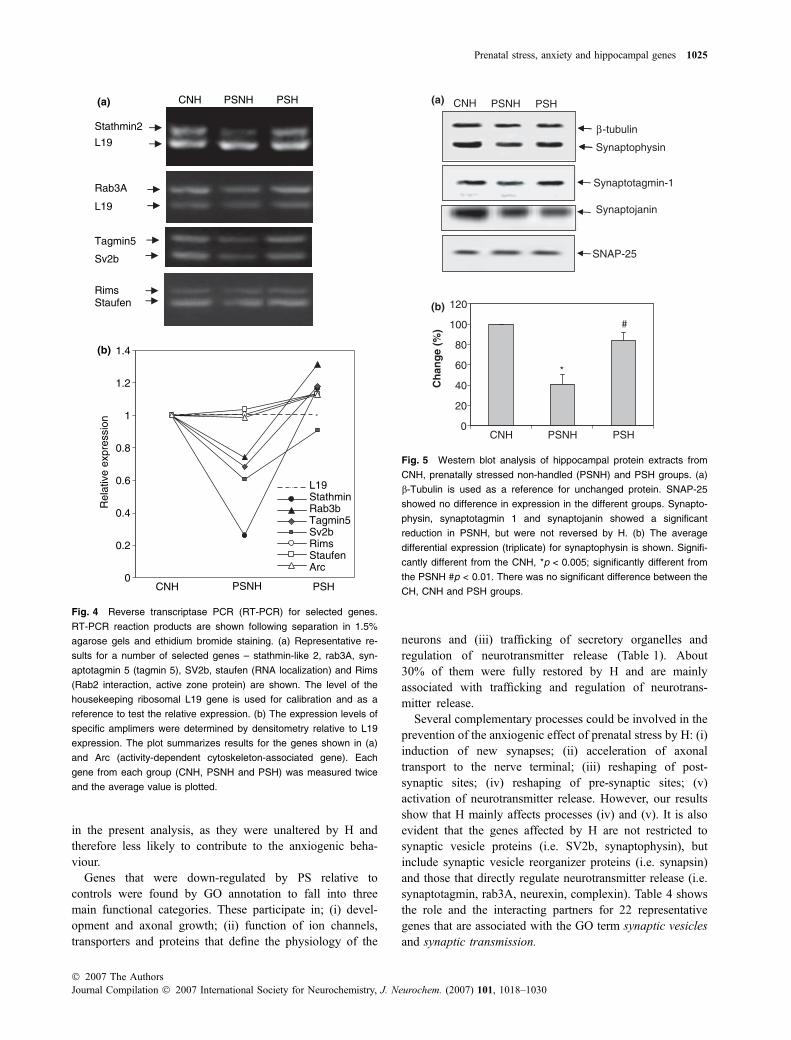
Change in expression of major pre-synaptic proteins
Prenatal stress significantly reduced the levels of synapto-physin, synaptotagmin 1 and synaptojanin as shown bywestern analysis (Fig. 5). H only restored the expression ofthe first two proteins (Figs 5a and b). By contrast, there wereno detectable changes in synapsin 1 and rab3A proteins,despite changes in the levels of gene expression. Synapsin 1is associated with a pre-synaptic cytoskeletal adaptor ofsynaptic vesicles and rab3A, with the regulation of theirpriming (Sudhof 2004). In agreement with the data from theGeneChip analysis, there was no difference in the expressionof soluble N-ethylmaleimide-sensitive factor receptors
(SNAREs), syntaxin 1A, SNAP-25 and VAMP-2 in PSNH,PSH or CNH and CH rats. These results indicate that somebut not all the changes in proteins match the profile of thoseat the mRNA level. They also show that the effects ofprenatal stress and H are very specific and cannot beattributed to a global change in synaptic proteins or in thenumber of synapses.
Discussion
The major new finding in this study is the identification ofspecific genes and proteins in the hippocampus of naıvejuvenile female rats that show altered expression as a resultof prenatal stress. The hippocampus was chosen because ofits important role in the aetiology of anxiety and depressionand because it shows specific structural changes both inhumans (Campbell and MacQueen 2004) and in experimen-tal animal models of these conditions (Lemaire et al. 2000;Coe et al. 2003). By performing the study in weanling rats,we attempted to limit any gene changes to those resultingfrom increased levels of stress hormones in the maternalmilieu and early postnatal rearing influences. We found thatdifferences in the behaviour of PS females from that ofcontrols in the EPM could be detected at this early age with asmaller variance than that in adults, probably because of theabsence of an influence of the oestrus cycle (Marcondeset al. 2001).
Having identified a number of genes that were altered byprenatal stress, we focussed on the genes that no longerdiffered from those of controls in rats that had been subjectedto H, as this manipulation prevented the development ofanxiogenic behaviour. Of 582 genes that were altered byprenatal stress, 52% were under-expressed and 48% over-expressed relative to controls. We did not include these genes
Biological process
Physiological process
Cellular process
Organismal physiological process
Localization
Cell communication Cellular physiological
process
Neurophysiological process
Cell–cell signaling
Cell organization & biogenesis
Transmission of nerve impulse
Secretion Transport Membrane organization
and biogenesis
Synaptic transmission Secretory pathway
Regulation of neurotransmitter
levels
Regulated secretory pathway Synaptic vesicle
transport
Exocytosis
Neurotransmitter secretion Calcium ion-dependant exocytosis
Establishment of localization
Vesicle mediated transport
Fig. 3 A diacyclic graph according to
GOTM. In bold and grey background are
the enriched gene ontology (GO) (p < 0.01)
belonging to ‘biological process’ category.
For details see Table 2. Only 42.5% of the
genes differentially expressed between the
CNH and PSNH rats are annotated by GO.
1024 Y. Bogoch et al.
Journal Compilation � 2007 International Society for Neurochemistry, J. Neurochem. (2007) 101, 1018–1030� 2007 The Authors
in the present analysis, as they were unaltered by H andtherefore less likely to contribute to the anxiogenic beha-viour.
Genes that were down-regulated by PS relative tocontrols were found by GO annotation to fall into threemain functional categories. These participate in; (i) devel-opment and axonal growth; (ii) function of ion channels,transporters and proteins that define the physiology of the
neurons and (iii) trafficking of secretory organelles andregulation of neurotransmitter release (Table 1). About30% of them were fully restored by H and are mainlyassociated with trafficking and regulation of neurotrans-mitter release.
Several complementary processes could be involved in theprevention of the anxiogenic effect of prenatal stress by H: (i)induction of new synapses; (ii) acceleration of axonaltransport to the nerve terminal; (iii) reshaping of post-synaptic sites; (iv) reshaping of pre-synaptic sites; (v)activation of neurotransmitter release. However, our resultsshow that H mainly affects processes (iv) and (v). It is alsoevident that the genes affected by H are not restricted tosynaptic vesicle proteins (i.e. SV2b, synaptophysin), butinclude synaptic vesicle reorganizer proteins (i.e. synapsin)and those that directly regulate neurotransmitter release (i.e.synaptotagmin, rab3A, neurexin, complexin). Table 4 showsthe role and the interacting partners for 22 representativegenes that are associated with the GO term synaptic vesiclesand synaptic transmission.
Stathmin2
L19
L19
Rab3A
RimsStaufen
Tagmin5
Sv2b
0
0.2
0.4
0.6
0.8
1
1.2
1.4
Rel
ativ
e ex
pres
sion
L19StathminRab3bTagmin5Sv2bRimsStaufenArc
PSHPSNHCNH(a)
(b)
PSHPSNHCNH
Fig. 4 Reverse transcriptase PCR (RT-PCR) for selected genes.
RT-PCR reaction products are shown following separation in 1.5%
agarose gels and ethidium bromide staining. (a) Representative re-
sults for a number of selected genes – stathmin-like 2, rab3A, syn-
aptotagmin 5 (tagmin 5), SV2b, staufen (RNA localization) and Rims
(Rab2 interaction, active zone protein) are shown. The level of the
housekeeping ribosomal L19 gene is used for calibration and as a
reference to test the relative expression. (b) The expression levels of
specific amplimers were determined by densitometry relative to L19
expression. The plot summarizes results for the genes shown in (a)
and Arc (activity-dependent cytoskeleton-associated gene). Each
gene from each group (CNH, PSNH and PSH) was measured twice
and the average value is plotted.
β -tubulin
Synaptophysin
SNAP-25
Synaptotagmin-1
Synaptojanin
(a)
0
20
40
60
80
100
120
Ch
ang
e (%
)
(b)
*
#
PSH PSNH CNH
PSH PSNH CNH
Fig. 5 Western blot analysis of hippocampal protein extracts from
CNH, prenatally stressed non-handled (PSNH) and PSH groups. (a)
b-Tubulin is used as a reference for unchanged protein. SNAP-25
showed no difference in expression in the different groups. Synapto-
physin, synaptotagmin 1 and synaptojanin showed a significant
reduction in PSNH, but were not reversed by H. (b) The average
differential expression (triplicate) for synaptophysin is shown. Signifi-
cantly different from the CNH, *p < 0.005; significantly different from
the PSNH #p < 0.01. There was no significant difference between the
CH, CNH and PSH groups.
Prenatal stress, anxiety and hippocampal genes 1025
� 2007 The AuthorsJournal Compilation � 2007 International Society for Neurochemistry, J. Neurochem. (2007) 101, 1018–1030
Tab
le4
Are
pre
senta
tive
list
of
genes
that
show
ed
reduced
expre
ssio
nas
are
sult
of
pre
nata
lstr
ess
No.
inF
ig.
6a
Gene
nam
es
Gene
sym
bol
No.
infa
mily
bR
efe
rence
Cellu
lar
pro
cess
(pro
tein
inte
raction)
1S
Vgly
copro
tein
2B
SV
2B
3Lazzell
et
al.
(2004)
Synaptic
vesic
leexocyto
sis
(synapto
tagm
in1)
2S
ynapto
tagm
in5
SY
T5
16
Ste
vens
and
Sulli
van
(2003)
Ca
2+
sensor,
traffi
ckin
g,
exo–endocyto
sis
(Rab27,
Ca
2+
channel)
3S
ynapto
tagm
in3
SY
T3
16
Ste
vens
and
Sulli
van
(2003)
Ca
2+
sensor,
exo–endocyto
sis
(Syt9
,C
a2+
channeland
neure
xin
)
4S
ynapto
physin
/p38
SY
N4
Valtort
aet
al.
(2004)
Synaptic
vesic
leta
rgeting,
bio
genesis
,exo-e
ndocyto
sis
(Vam
p,
physophili
n)
5V
esic
le-a
ssocia
ted
mem
bra
ne
pro
tein
1V
AM
P1
8S
udhof
et
al.
(1989)
Vesic
le-m
edia
ted
transport
(Snap25,
synta
xin
1)
6V
esic
le-a
ssocia
ted
mem
bra
ne
pro
tein
7V
AM
P7
8M
art
inez-A
rca
et
al.
(2003)
Vesic
le-m
edia
ted
transport
(Snap25,
Vam
p2)
7B
locked
early
intr
ansport
1B
ET
11
Mossessova
et
al.
(2003)
ER
toG
olg
itr
ansport
(Sec22)
8H
untingtin-a
ssocia
ted
pro
tein
1H
AP
11
Kittler
et
al.
(2004)
Synaptic
transm
issio
n(h
untingtin,
cynactin
1)
9P
ota
ssiu
mvoltage-g
ate
d
channelsubfa
mily
H
KC
NH
18
Jeng
et
al.
(2005)
Ion
transport
(pota
ssiu
mchannelre
gula
tor
1)
10
Rab2
RA
B2
40
Short
et
al.
(2001)
ER
toG
olg
ivesic
le-m
edia
ted
transport
(GD
Pdis
socia
tion
inhib
itor
2)
11
c-S
NA
PN
AP
G3
Chen
et
al.
(2001)
Fusio
n,
vesic
letr
affi
ckin
g(G
AF
-1,
NS
Fand
RIP
11)
12
Com
ple
xin
2/s
ynaphin
CP
LX
24
McM
ahon
et
al.
(1995)
Synaptic
vesic
leexocyto
sis
(SN
AR
Es,
synta
xin
)
13
Fre
quenin
hom
olo
gF
RE
Q5
O’C
alla
ghan
et
al.
(2005)
Ca
2+
bin
din
g,
transm
issio
nre
gula
tion
(KC
ND
2)
14
Cort
actin
isofo
rmB
CT
TN
1R
acz
and
Wein
berg
(2004)
Pro
tein
inte
raction
(d-c
ate
nin
,dynam
in1)
15
Sta
thm
in-lik
e2/S
CG
10
ST
MN
24
Liu
et
al.
(2002)
Neurite
outg
row
th,
cyto
skele
ton
(RG
S6)
16
Kin
esin
1K
IF5b
25
Kanaiet
al.
(2004)
Mic
rotu
bule
-based
movem
ent
(merlin
)
17
Bra
in-s
pecifi
cangio
genesis
inhib
itor
1-a
ssocia
ted
pro
tein
2
BA
P2
2O
da
et
al.
(1999)
Neurite
outg
row
th,
cyto
skele
ton
(Atn
1,
Bal1
,
Enah,
Eps8,
Shank1–3
and
Wasf1
–2)
18
Synapsin
1S
YN
13
Paggiand
Petr
ucci(1
992)
Cyto
skele
ton
(Nos1,
Rab3
and
F-a
ctin)
19
Neure
xin
1N
RX
N1
4D
ean
and
Dre
sbach
(2006)
Axon
guid
ance
(neuro
ligin
1)
20
SN
AP
91/A
P180
SN
AP
91
4S
ousa
et
al.
(1992)
Endocyto
sis
(cla
thrin,
AP
2)
21
Am
phip
hysin
AM
PH
1S
hang
et
al.
(2004)
Synaptic
transm
issio
n(d
ynam
in1)
22
Synapto
janin
1S
YN
J1
Irie
et
al.
(2005)
Synaptic
vesic
leendocyto
sis
(am
phip
hysin
)
aG
enes
liste
dare
those
that
were
resto
red
by
H(b
old
,gre
ybackgro
und)
and
the
rest
were
unaff
ecte
dby
H.
bT
he
num
bers
of
fam
ilym
em
bers
that
are
dete
cte
din
mam
mals
,exclu
din
galtern
ative
splic
ed
variants
.
1026 Y. Bogoch et al.
Journal Compilation � 2007 International Society for Neurochemistry, J. Neurochem. (2007) 101, 1018–1030� 2007 The Authors

The assembly of a functional synapse is a fully coordi-nated sequential process that is sensitive to cues from theenvironment (Ziv and Garner 2004). Neurotransmitter releaseis needed for the fine tuning and maintenance of synapticconnections (Verhage et al. 2000) and may be modulated bya few genes such as synaptotagmin 1, synaptophysin andSV2b, all of which were restored by H. These major synapticvesicle proteins participate in multiple protein–protein inter-actions in the synapse (Table 4). Their relative amountsdetermine whether a synaptic vesicle will be engaged into aproductive cycle of docking and fusion. Synaptophysindetermines the accessibility of VAMP-2 to the SNAREcomplex (McMahon et al. 1996) and SV2b determines theaccessibility of synaptotagmin 1 to the SNAREs and the Ca2+
sensitivity of the fusion complexes (Lazzell et al. 2004). Amodel that shows the location and function of representativegenes that were altered by prenatal stress and H is shown(Fig. 6). Although the data do not reveal the identity of the
synapses and neurotransmitters, other studies have shownthat alterations occur in the release of noradrenaline,dopamine, serotonin and glutamate in PS rats (Weinstock2001; Berger et al. 2002).
Several groups of genes down-regulated by prenatal stresswere not restored by H. These include genes that areimportant for axonal transport [i.e. myosin V, kinesin-likeproteins (Okada et al. 1995)], shaping of the post-synapticsites, neurotransmitter receptors, scaffolding proteins con-taining PDZ domains (Kim and Sheng 2004) and neuronalSNAREs. SNARE proteins are transported in vesicles that donot contain other synaptic vesicle proteins (Okada et al.1995) and their accumulation in the growth cone precedessynaptic vesicle protein transport and the maturation of afunctional synapse (Igarashi et al. 1997). The highest changein expression in PSNH relative to CNH was obtained for ionchannels including pre-synaptic voltage-gated Ca2+ type P/Qand several K+ channels. These ion channels regulate theelectrical properties of the neuron, and their down-regulationsuggests a potential decrease in excitability and electricalproperties of the newly formed synapses. However, thesealso were unaffected by H, suggesting that they are lessimportant in determining anxiogenic behaviour in PS rats.Other genes that failed to show a significant level of recoveryfollowing H include synaptojanin, ampiphysin, caveolin thatplay a central role in synaptic vesicle endocytosis (Fig. 6). Ina mature synapse, endocytosis is coupled to the exocytoticmachinery (Linial and Parnas 1996). It is possible that H didnot restore the activity of the endocytotic apparatus, becauseit matures at a later age than that at which our experimentswere performed. Indeed, endocytotic proteins were shown tobe expressed long after the biogenesis of the synapticvesicles and following their accumulation in the synapse(Grabs et al. 2000). We will investigate this possibility bytesting the level of gene expression of this machinery at alater stage of development (12 weeks).
Similar to other studies (Weaver et al. 2006), wholehippocampus was used to monitor changes in gene expres-sion, enabling us to detect genes, proteins and processes thatare differentially expressed as a result of prenatal stress andH. We expect to gain additional information by performingthe analysis in discrete hippocampal regions (Lein et al.2004). To our knowledge, there is only one other report ofthe effect of prenatal stress on gene expression that wasperformed on RNA from the prefrontal cortex of adult males.Its aim was to determine whether there were any similaritiesin the molecular changes that were found in PS rats to thoseseen in schizophrenic and bipolar patients (Kinnunen et al.2003). In spite of the differences in experimental design,gender of the animals, their age and the brain area from thosein the current study, some notable similarities were seenamong the genes that showed a decreased expression as aresult of prenatal stress. These include complexin 2, BDNF,synapsin and ionotropic NMDA 2A receptor. In contrast to
Plasma membrane & endocytosis related proteins
Synaptic vesicle/secretory vesicle membrane proteins
SNARE regulators & ion channels
Cytoskeleton related proteins
Synaptic vesicle
15
17
16
18
Transport organelle
1 2
34567
9
810
11
12
13
14
19
2120
Fig. 6 Schematic representation of a pre-synaptic site indicating
genes that were detected in the current study. A scheme of a synaptic
vesicle and a generic transport organelle is shown. Various proteins
are marked according to their location and function. In bold and grey
are shown those genes that showed a reduced expression as a result
of prenatal stress that was restored by H. The rest were down-regu-
lated by prenatal stress but not restored by H. For additional infor-
mation of these genes see Table 4.
Prenatal stress, anxiety and hippocampal genes 1027
� 2007 The AuthorsJournal Compilation � 2007 International Society for Neurochemistry, J. Neurochem. (2007) 101, 1018–1030

those reported (Kinnunen et al. 2003), we did not detect asignificant change in the expression of genes that make upthe post-synaptic density.
In conclusion, our study shows that prenatal stressoccurring during a critical period of foetal developmentalters the expression of a well-defined set of hippocampalgenes in association with permanent changes in behaviour.Using the procedure of neonatal H, we were able to focus ona defined set of genes, the expression of which was restoredto control levels in association with normalization ofbehaviour. The genes and proteins identified in this wayplay an important role in regulating the rate of pre-synapticmaturation. It remains to be determined whether the changesinduced by prenatal stress in gene expression and behaviourresult from exposure of the foetal brain to excess levels ofmaternal stress hormones and/or changes in maternal atten-tion during the early postnatal period.
Acknowledgements
We thank Yaniv Bledi for helpful suggestions. Analysis of the
Affymetrix data was performed at the Genomic Sequencing Centre
at the Hebrew University. Y.B awarded a fellowship from SCCB, the
Sudarsky Centre for Computational Biology, The Hebrew Univer-
sity of Jerusalem.
References
Ader R. (1970) The effects of early experience on the adrenocorticalresponse to different magnitudes of stimulation. Physiol. Behav. 5,837–839.
Alonso S. J., Arevalo R., Afonso D. and Rodriguez M. (1991) Effects ofmaternal stress during pregnancy on forced swimming test be-havior of the offspring. Physiol. Behav. 50, 511–517.
Bayer S. A., Altman J., Russo R. J. and Zhang X. (1993) Timetables ofneurogenesis in the human brain based on experimentally deter-mined patterns in the rat. Neurotoxicology 14, 83–144.
Belzung C. (1992) Hippocampal mossy fibres: implication in noveltyreactions or in anxiety behaviours Behav. Brain Res. 51, 149–155.
Berger M. A., Barros V. G., Sarchi M. I., Tarazi F. I. and Antonelli M. C.(2002) Long-term effects of prenatal stress on dopamine and glu-tamate receptors in adult rat brain. Neurochem. Res. 27, 1525–1533.
Camon E., Barrell D., Lee V., Dimmer E. and Apweiler R. (2004) TheGene Ontology Annotation (GOA) Database – an integrated re-source of GO annotations to the UniProt Knowledgebase. In SilicoBiol. 4, 5–6.
Campbell S. and MacQueen G. (2004) The role of the hippocampus inthe pathophysiology of major depression. J. Psychiatry Neurosci.29, 417–426.
Chen D., Xu W., He P., Medrano E. E. and Whiteheart S. W. (2001) Gaf-1, a gamma-SNAP-binding protein associated with the mitochon-dria. J. Biol. Chem. 276, 13 127–13 135.
Coe C. L., Kramer M., Czeh B., Gould E., Reeves A. J., Kirschbaum C.and Fuchs E. (2003) Prenatal stress diminishes neurogenesis in thedentate gyrus of juvenile rhesus monkeys. Biol. Psychiatry 54,1025–1034.
De Bellis M. D., Casey B. J., Dahl R. E. et al. (2000) A pilot study ofamygdala volumes in pediatric generalized anxiety disorder. Biol.Psychiatry 48, 51–57.
Dean C. and Dresbach T. (2006) Neuroligins and neurexins: linking celladhesion, synapse formation and cognitive function. Trends Neu-rosci. 29, 21–29.
Dennis G. Jr, Sherman B. T., Hosack D. A., Yang J., Gao W., LaneH. C. and Lempicki R. A. (2003) DAVID: Database forAnnotation, Visualization, and Integrated Discovery. GenomeBiol. 4, P3.
Fride E. and Weinstock M. (1988) Prenatal stress increases anxietyrelated behavior and alters cerebral lateralization of dopamineactivity. Life Sci. 42, 1059–1065.
Fride E., Dan Y., Feldon J., Halevy G. and Weinstock M. (1986) Effectsof prenatal stress on vulnerability to stress in prepubertal and adultrats. Physiol. Behav. 37, 681–687.
Geddes J. (1999) Prenatal and perinatal risk factors for early onsetschizophrenia, affective psychosis, and reactive psychosis. BMJ318, 426.
Gentsch C., Lichtsteiner M., Frischknecht H. R., Feer H. and SiegfriedB. (1988) Isolation-induced locomotor hyperactivity and hypoal-gesia in rats are prevented by handling and reversed by resocial-ization. Physiol. Behav. 43, 13–16.
Grabs D., Bergmann M. and Rager G. (2000) Developmental expressionof amphiphysin in the retinotectal system of the chick: from mRNAto protein. Eur. J. Neurosci. 12, 1545–1553.
Gue M., Bravard A., Meunier J., Veyrier R., Gaillet S., Recasens M.and Maurice T. (2004) Sex differences in learning deficits inducedby prenatal stress in juvenile rats. Behav. Brain Res. 150, 149–157.
Handley S. L. and Mithani S. (1984) Effects of alpha-adrenoceptor ag-onists and antagonists in a maze-exploration model of ‘fear’-motivated behaviour. Naunyn Schmiedebergs Arch. Pharmacol.327, 1–5.
Hayashi A., Nagaoka M., Yamada K., Ichitani Y., Miake Y. and OkadoN. (1998) Maternal stress induces synaptic loss and developmentaldisabilities of offspring. Int. J. Dev. Neurosci. 16, 209–216.
Igarashi M., Tagaya M. and Komiya Y. (1997) The soluble N-ethylma-leimide-sensitive factor attached protein receptor complex ingrowth cones: molecular aspects of the axon terminal development.J. Neurosci. 17, 1460–1470.
Irie F., Okuno M., Pasquale E. B. and Yamaguchi Y. (2005) EphrinB-EphB signalling regulates clathrin-mediated endocytosis throughtyrosine phosphorylation of synaptojanin 1. Nat. Cell Biol. 7, 501–509.
Jeng C. J., Chang C. C. and Tang C. Y. (2005) Differential localization ofrat Eag1 and Eag2 K+ channels in hippocampal neurons. Neuro-report 16, 229–233.
Kanai Y., Dohmae N. and Hirokawa N. (2004) Kinesin transports RNA:isolation and characterization of an RNA-transporting granule.Neuron 43, 513–525.
Keshet G. I. and Weinstock M. (1995) Maternal naltrexone preventsmorphological and behavioral alterations induced in rats by pre-natal stress. Pharmacol. Biochem. Behav. 50, 413–419.
Kim E. and Sheng M. (2004) PDZ domain proteins of synapses. Nat.Rev. Neurosci. 5, 771–781.
Kinnunen A. K., Koenig J. I. and Bilbe G. (2003) Repeated variableprenatal stress alters pre- and postsynaptic gene expression in therat frontal pole. J. Neurochem. 86, 736–748.
Kittler J. T., Thomas P., Tretter V., Bogdanov Y. D., Haucke V., Smart T.G. and Moss S. J. (2004) Huntingtin-associated protein 1 regulatesinhibitory synaptic transmission by modulating gamma-aminobu-tyric acid type A receptor membrane trafficking. Proc. Natl Acad.Sci. USA 101, 12 736–12 741.
Kuehner C. (1999) Gender differences in the short-term course of uni-polar depression in a follow-up sample of depressed inpatients. J.Affect. Disord. 56, 127–139.
1028 Y. Bogoch et al.
Journal Compilation � 2007 International Society for Neurochemistry, J. Neurochem. (2007) 101, 1018–1030� 2007 The Authors

Lazzell D. R., Belizaire R., Thakur P., Sherry D. M. and Janz R. (2004)SV2B regulates synaptotagmin 1 by direct interaction. J. Biol.Chem. 279, 52 124–52 131.
Lein E. S., Zhao X. and Gage F. H. (2004) Defining a molecular atlas ofthe hippocampus using DNA microarrays and high-throughput insitu hybridization. J. Neurosci. 24, 3879–3889.
Lemaire V., Koehl M., Le Moal M. and Abrous D. N. (2000) Prenatalstress produces learning deficits associated with an inhibition ofneurogenesis in the hippocampus. Proc. Natl Acad. Sci. USA 97,11 032–11 037.
Levine S., Haltmeyer G. C., Karas G. G. and Denenberg V. H. (1967)Physiological and behavioral effects of infantile stimulationPhysiol. Behav. 2, 55–59.
Linial M. and Parnas D. (1996) Deciphering neuronal secretion: tools ofthe trade. Biochim. Biophys. Acta 1286, 117–152.
Linnet K. M., Dalsgaard S., Obel C. et al. (2003) Maternal lifestylefactors in pregnancy risk of attention deficit hyperactivity disorderand associated behaviors: review of the current evidence. Am.J. Psychiatry 160, 1028–1040.
Liu Z., Chatterjee T. K. and Fisher R. A. (2002) RGS6 interacts withSCG10 and promotes neuronal differentiation. Role of the Ggamma subunit-like (GGL) domain of RGS6. J. Biol. Chem. 277,37 832–37 839.
Maccari S., Darnaudery M., Morley-Fletcher S., Zuena A. R., Cinque C.and Van Reeth O. (2003) Prenatal stress and long-term conse-quences: implications of glucocorticoid hormones. Neurosci. Bio-behav. Rev. 27, 119–127.
Marcondes F. K., Miguel K. J., Melo L. L. and Spadari-Bratfisch R. C.(2001) Estrous cycle influences the response of female rats in theelevated plus-maze test. Physiol. Behav. 74, 435–440.
Martinez-Arca S., Proux-Gillardeaux V., Alberts P., Louvard D. andGalli T. (2003) Ectopic expression of syntaxin 1 in the ER redirectsTI-VAMP- and cellubrevin-containing vesicles. J. Cell Sci. 116,2805–2816.
McMahon H. T., Missler M., Li C. and Sudhof T. C. (1995) Complexins:cytosolic proteins that regulate SNAP receptor function. Cell 83,111–119.
McMahon H. T., Bolshakov V. Y., Janz R., Hammer R. E., SiegelbaumS. A. and Sudhof T. C. (1996) Synaptophysin, a major synapticvesicle protein, is not essential for neurotransmitter release. Proc.Natl Acad. Sci. USA 93, 4760–4764.
Milberger S., Biederman J., Faraone S. V., Guite J. and Tsuang M. T.(1997) Pregnancy, delivery and infancy complications and atten-tion deficit hyperactivity disorder: issues of gene-environmentinteraction. Biol. Psychiatry 41, 65–75.
Mossessova E., Bickford L. C. and Goldberg J. (2003) SNARE selec-tivity of the COPII coat. Cell 114, 483–495.
O’Callaghan D. W., Haynes L. P. and Burgoyne R. D. (2005) High-affinity interaction of the N-terminal myristoylation motif of theneuronal calcium sensor protein hippocalcin with phosphatidy-linositol 4,5-bisphosphate. Biochem. J. 391, 231–238.
O’Donovan K. J., Tourtellotte W. G., Millbrandt J. and Baraban J. M.(1999) The EGR family of transcription-regulatory factors: pro-gress at the interface of molecular and systems neuroscience.Trends Neurosci. 22, 167–173.
Oda K., Shiratsuchi T., Nishimori H., Inazawa J., Yoshikawa H., Take-tani Y., Nakamura Y. and Tokino T. (1999) Identification of BAI-AP2 (BAI-associated protein 2), a novel human homologue ofhamster IRSp53, whose SH3 domain interacts with the cytoplasmicdomain of BAI1. Cytogenet. Cell Genet. 84, 75–82.
Okada Y., Yamazaki H., Sekine-Aizawa Y. and Hirokawa N. (1995) Theneuron-specific kinesin superfamily protein KIF1A is a uniquemonomeric motor for anterograde axonal transport of synapticvesicle precursors. Cell 81, 769–780.
van Os J. and Selten J. P. (1998) Prenatal exposure to maternal stress andsubsequent schizophrenia: the May 1940 invasion of the Nether-lands. Br. J. Psychiatry 172, 324–326.
Paggi P. and Petrucci T. C. (1992) Neuronal compartments and axonaltransport of synapsin I. Mol. Neurobiol. 6, 239–251.
Pellow S. and File S. E. (1986) Anxiolytic and anxiogenic drugeffects on exploratory activity in an elevated plus-maze: a noveltest of anxiety in the rat. Pharmacol. Biochem. Behav. 24, 525–529.
Poltyrev T. and Weinstock M. (2004) Gender difference in the preven-tion of hyperanxiety in adult prenatally stressed rats by chronictreatment with amitriptyline. Psychopharmacology (Berl.) 171,270–276.
Racz B. and Weinberg R. J. (2004) The subcellular organization ofcortactin in hippocampus. J. Neurosci. 24, 10 310–10 317.
Salm A. K., Pavelko M., Krouse E. M., Webster W., Kraszpulski M. andBirkle D. L. (2004) Lateral amygdaloid nucleus expansion in adultrats is associated with exposure to prenatal stress. Brain Res. Dev.Brain Res. 148, 159–167.
Schmitz C., Rhodes M. E., Bludau M., Kaplan S., Ong P., Ueffing I.,Vehoff J., Korr H. and Frye C. A. (2002) Depression: reducednumber of granule cells in the hippocampus of female, but notmale, rats due to prenatal restraint stress. Mol. Psychiatry 7, 810–813.
Schneider M. L. (1992) Prenatal stress exposure alters postnatal be-havioral expression under conditions of novelty challenge in rhesusmonkey infants. Dev. Psychobiol. 25, 529–540.
Shang W. H., Adachi Y., Nakamura A., Copeland T., Kim S. R. andKamata T. (2004) Regulation of amphiphysin1 by mitogen-acti-vated protein kinase: its significance in nerve growth factorreceptor-mediated endocytosis. J. Biol. Chem. 279, 40 890–40 896.
Short B., Preisinger C., Korner R., Kopajtich R., Byron O. and Barr F. A.(2001) A GRASP55-rab2 effector complex linking Golgi structureto membrane traffic. J. Cell Biol. 155, 877–883.
Sousa R., Tannery N. H., Zhou S. and Lafer E. M. (1992) Characteri-zation of a novel synapse-specific protein. I. Developmentalexpression and cellular localization of the F1-20 protein andmRNA. J. Neurosci. 12, 2130–2143.
Stevens C. F. and Sullivan J. M. (2003) The synaptotagmin C2A domainis part of the calcium sensor controlling fast synaptic transmission.Neuron 39, 299–308.
Sudhof T. C. (2004) The synaptic vesicle cycle. Annu. Rev. Neurosci. 27,509–547.
Sudhof T. C., Baumert M., Perin M. S. and Jahn R. (1989) Asynaptic vesicle membrane protein is conserved from mammals toDrosophila. Neuron 2, 1475–1481.
Vallee M., Mayo W., Dellu F., Le Moal M., Simon H. and Maccari S.(1997) Prenatal stress induces high anxiety and postnatal hand-ling induces low anxiety in adult offspring: correlation withstress-induced corticosterone secretion. J. Neurosci. 17, 2626–2636.
Valtorta F., Pennuto M., Bonanomi D. and Benfenati F. (2004) Synapt-ophysin: leading actor or walk-on role in synaptic vesicle exocy-tosis Bioessays 26, 445–453.
Verhage M., Maia A. S., Plomp J. J. et al. (2000) Synaptic assembly ofthe brain in the absence of neurotransmitter secretion. Science 287,864–869.
Wakshlak A. and Weinstock M. (1990) Neonatal handling reverses be-havioral abnormalities induced in rats by prenatal stress. Physiol.Behav. 48, 289–292.
Ward H. E., Johnson E. A., Salm A. K. and Birkle D. L. (2000) Effects ofprenatal stress on defensive withdrawal behavior and corticotropinreleasing factor systems in rat brain. Physiol. Behav. 70, 359–366.
Prenatal stress, anxiety and hippocampal genes 1029
� 2007 The AuthorsJournal Compilation � 2007 International Society for Neurochemistry, J. Neurochem. (2007) 101, 1018–1030

Weaver I. C., Meaney M. J. and Szyf M. (2006) Maternal care effects onthe hippocampal transcriptome and anxiety-mediated behaviors inthe offspring that are reversible in adulthood. Proc. Natl Acad. Sci.USA 103, 3480–3485.
Weinstock M. (1997) Does prenatal stress impair coping and regulationof hypothalamic-pituitary-adrenal axis Neurosci. Biobehav. Rev.21, 1–10.
Weinstock M. (2001) Alterations induced by gestational stress in brainmorphology and behaviour of the offspring. Prog. Neurobiol. 65,427–451.
Zhang B., Schmoyer D., Kirov S. and Snoddy J. (2004) GOTreeMachine (GOTM): a web-based platform for interpreting setsof interesting genes using Gene Ontology hierarchies. BMC Bio-informatics 5, 16.
Zhu Z., Li X., Chen W., Zhao Y., Li H., Qing C., Jia N., Bai Z. and Liu J.(2004) Prenatal stress causes gender-dependent neuronal loss andoxidative stress in rat hippocampus. J. Neurosci. Res. 78, 837–844.
Ziv N. E. and Garner C. C. (2004) Cellular and molecular mechanisms ofpresynaptic assembly. Nat. Rev. Neurosci. 5, 385–399.
1030 Y. Bogoch et al.
Journal Compilation � 2007 International Society for Neurochemistry, J. Neurochem. (2007) 101, 1018–1030� 2007 The Authors




















