
Thalamic and prefrontal GABA concentrations but not GABAA ...

Ž .Molecular Brain Research 57 1998 310–320
Research report
Antisense inhibition of striatal GABA receptor proteins decreasesA
GABA-stimulated chloride uptake and increases cocaine sensitivity inrats
Joanna Peris ), Bruce J. Jung, Adam Resnick, Paul Walker, Olga Malakhova,Yvonne Bokrand, Donna Wielbo
Departments of Pharmacodynamics and Medicinal Chemistry, Box 100487, UniÕersity of Florida, GainesÕille, FL 32610, USA
Accepted 7 April 1998
Abstract
The functional status of striatal GABA receptors appears to be inversely related to the magnitude of cocaine-induced behaviors.AŽ .Exposure of striatum to antisense oligodeoxynucleotides ASODNs targeted to the mRNAs for the a and the b subunits of the2 3
GABA receptor should decrease expression of receptor proteins and therefore might be expected to increase cocaine sensitivity.A
ASODNs, scrambled ODNs or saline were injected into right lateral ventricle of rats and behavioral responses to cocaine were tested18–20 h after treatment. Animals injected separately with a or b ASODNs exhibited increased behavioral sensitivity to cocaine2 3
compared to rats injected with saline or scrambled ODNs including performing more 3608 turns to the left than to the right. There wassignificantly less GABA-stimulated Cl uptake in right striatum compared to left striatum of ASODN-treated rats with no significantdifference between sides in control animals. Specific binding to benzodiazepine and convulsant sites on the GABA receptor was notA
selectively altered by ASODN treatment. Combined a b ASODN treatment did not affect either cocaine sensitivity or GABA2 3 A
receptor function. There was no difference between the density of Nissl stained cells in the left and right edges of striatum in control orASODN-treated rats indicating the absence of significant neurotoxic effects of the ASODN treatment. Injection of fluorescein-conjugatedASODNs indicated that ASODN is present in striatum at times during which behavioral and neurochemical indices of GABA receptorfunction are decreased. Thus, the functional status of GABA receptors in striatum may be involved in determining cocaine sensitivity.A
q 1998 Elsevier Science B.V. All rights reserved.
Keywords: GABA receptor; Cocaine; Locomotion; Stereotypy; Antisense oligodeoxynucleotide
1. Introduction
One of the important brain regions for mediating thebehavioral effects of initial and repeated cocaine exposure
Ž .is the nigrostriatal dopamine DA projection pathway.This pathway contains a large number of GABAergicneurons in both the cell body and terminal regions. Re-peated injections of cocaine selectively decrease both
w xGABA receptor number 8,30 and GABA-stimulated ClAw xuptake 32 in rat striatum. Furthermore, there is a signifi-
cant negative correlation between the decrease in striatalGABA receptor function and the magnitude of cocaine-A
sensitized behaviors indicating that decreased striatal
) Corresponding author. Fax: q1-352-392-9187; E-mail:[email protected]
GABAergic transmission may be important in the develop-w xment of cocaine sensitization 32 . Additionally, endoge-
nous GABA release is decreased from striatal slices ofw xcocaine-sensitized rats 13 indicating that decreases in
both pre- and postsynaptic GABA transmission may con-tribute to the development of cocaine sensitization. How-ever, in order to indicate a causal relationship betweenGABA receptor function and cocaine sensitivity, furtherA
manipulations of GABA receptor number andror func-A
tion are necessary.The GABA receptorrchloride ionophore is composedA
w xof five polypeptide subunits 27 of up to thirteen differentŽ . w w xxtypes a , b , g and d see Ref. 37 with each1 – 6 1 – 3 1 – 3
of the 13 subtypes displaying a different regional expres-w xsion 7,21,49 . For example, cortical cells express the
mRNA for most of the subtypes of the GABA receptor,A
0169-328Xr98r$19.00 q 1998 Elsevier Science B.V. All rights reserved.Ž .PII S0169-328X 98 00102-8

( )J. Peris et al.rMolecular Brain Research 57 1998 310–320 311
whereas striatal receptors are comprised mostly of a , a ,2 4
b and g subunits. The limited subunit combinations that3
exist in striatum may result in receptors that differ in thepresence of binding sites and the functional interactions ofthese sites in regulating ionophore opening. Since thenumber of 3H-muscimol and 35S-TBPS binding sites are
w xdifferentially determined by subunit composition 53 , it isnot surprising that receptors in striatum have vastly differ-ent ratios of antagonist, agonist and convulsant binding
w xsites compared to cortex 28 . The regional differences inthe composition of striatal GABA receptors may provideA
the mechanism by which repeated cocaine selectively de-creases striatal receptor number and function. Additionally,the limited variety of subunits available in striatum mayaffect the process of receptor homeostasis in this regionsuch that there is a decrease in the ability to alter receptorcomposition in response to various drug treatments orexperimental conditions. Thus, striatum may be more sus-ceptible to the effects of antisense oligodeoxynucleotidesŽ .ASODNs that selectively decrease the translation of par-ticular subunit proteins. The inability of other subunits totake the place of the inhibited proteins may result in amore dramatic loss of receptors in striatum compared toother brain regions treated similarly.
ASODNs have been used to inhibit gene expression in aŽ .number of biological systems see below . These com-
pounds are hypothesized to work by inhibiting transcrip-tional and translational processes of gene expression. An
Ž .ASODN sequence usually 12–18 bases in length targetedto a specific mRNA sequence will bind to the targetmRNA by direct Watson–Crick interactions and inhibit thepost-transcriptional or translational processes thereby in-hibiting protein synthesis. Several practical factors must beconsidered when utilizing ASODNs such as availability forcellular uptake, degree of nuclease resistance and associ-ated neurotoxicity. The mechanism of cellular uptake is
w xthought to be via receptor-mediated endocytosis 23,41,51 .Unmodified or phosphodiester ODNs are degraded readilyby endo- and exonucleases, and therefore have an ex-
w xtremely short half-life in vivo 1,47 . Modification byphosphorothioation substantially enhances nuclease resis-
w xtance while maintaining RNase H activation 20,40,42 . Inaddition, this type of modification imparts a catalytic typebehavior, whereby RNase H degradation of the mRNAportion of the DNA–mRNA duplex frees the ODN strand
w xto bind to other strands of target mRNA 10 . Although themodification of the backbone may increase neurotoxiceffects of ASODN treatment, this is generally not the case
w xunless repeated administration is employed 6 .Ž .The feasibility of intracerebroventricular i.c.v. injec-
tions of ASODN have been recently demonstrated for aw xnumber of neurotransmitter systems 9,43–46,48,55 . In
terms of in vivo ASODN treatments targeted for theGABA receptor, i.c.v. injections of ASODN targeted forA
the mRNA of the g subunit decrease benzodiazepine2w x w x14–16,24 , convulsant and agonist 15,16 binding to the
GABA receptorrionophore and increase convulsantAw xthreshold for benzodiazepine inverse agonists 54 .
Adeno-associated virus vector antisense gene transfer ofcDNA for the a subunit into the inferior colliculus1
decreases zolpidem binding and increases the duration ofw xinduced seizures in rats 50 . As yet, the effects of ASODN
for the GABA receptor a and b subunits present inA
striatal regions have not been studied.In order to further study the relationship between co-
caine sensitization and changes in GABAergic transmis-sion, ASODN treatments that should decrease GABAA
receptors were monitored for their effects on cocainesensitivity. Since the turnover of GABA receptors isA
fairly rapid with 42% of receptors degraded with a half-lifew xof about 4 h 3 , injections of ASODNs for GABAA
receptor protein subunit mRNA should decrease receptornumber in striatum when measured 18–24 h later. Wehypothesize that a decrease in striatal GABA receptorsA
should sensitize animals to cocaine.
2. Materials and methods
2.1. Subjects
Ž .Male Sprague–Dawley rats 200–225 g, Harlan wereused in order to eliminate influences of ovarian steroids oncocaine sensitivity. They were housed in the UF AnimalCare Facility with a 12-h light–dark cycle and free accessto food and water. Rats were anesthetized with ketamine
Ž .HCl 10 mgr kg, i.m. supplemented with Metofane asŽ .needed. Guide cannulae 24 gauge were implanted into
the right lateral ventricle using the following coordinatesw xfrom Bregma 29 : APsy0.2–0.5, laterals3.0–3.5,
dorsalrventrals6.0–6.5. The cannulae were cemented tosurgical screws implanted in the skull using dental cementand then the wound was closed. Cannula were fitted withobturators to maintain patency. After a 3–5 day recoveryperiod from surgery, unilateral injections of ODNs were
Ž .given in 25 mg doses 1 mgrml of isotonic saline . Thisresulted in injection of 5.1 nmol a ODN and 4.8 nmol2
b ODN. In some experiments, behavioral sensitivity to3
cocaine was measured 18 h after ODN injection. Allanimals were killed by decapitation with a guillotine 24 hafter ODN injection and brain tissue was obtained for theexperiments delineated below.
2.2. BehaÕioral assays
Cocaine-induced behaviors were quantified as describedw xpreviously 32,33 . Locomotor and stereotyped behaviors
were rated concurrently using a subjective 5-point scale.Animals were placed in separate chambers which werecylinders made of 0.7 cm wire mesh, measuring 30 cm indiameter and 37.5 cm high. Behavior was rated for 10 speriods every 5 min, beginning 10 min prior to and

( )J. Peris et al.rMolecular Brain Research 57 1998 310–320312
continuing until 80 min after cocaine injection. CocaineŽwas administered to all rats via i.p. injections 15 mgrkg;
.15 mgrml saline . Each animal was rated by an observerwho was blind to the ODN treatment. The following
Ž .stereotypy rating scale was used: 0 no gnawing or loco-motor movements, with or without grooming, normal quiet
Ž .behavior; 1 locomotion, 1–3 s of sniffing over the wireŽ .grid, normal exploratory behavior; 2 4 or more seconds
of licking or sniffing the grid, more rapid locomotorŽ .movement; 3 very rapid locomotion, gnawing on grid,
Ž .repetitive rearing or headbobbing; and 4 restricted loco-motion to a small area with intense gnawing, rearing orhead bobbing. The frequency of each rating during the 80min after injection was recorded. For statistical purposes,the ratings were summed over all trials to give eachsubject one behavioral score for each day of testing.Additionally, the number of right vs. left 3608 turns wasquantified during each 10-s observation period.
2.3. ODN synthesis and sequences
Phosphorothioate-modified ODNs were synthesized bythe Interdisciplinary Center for Biotechnology Research atthe University of Florida using standard phosphoramidite
w xprotocols with oxidation by iodine 2 . The phosphoroth-ioated ODNs were designed to target mRNA section en-coding the divergent intracellular area between putativetransmembrane domains M3 and M4 based on the cDNAsequences of the a and b subunits of the rat GABA2 3 A
w xreceptor 18,52 . The a ASODN was targeted to the2Ž Xsequence from subunits 340–346 5 -TCCATCCCAAGC-
X.CCATCC-3 and the b ASODN was targeted to subunits3Ž X X.380–385 5 -AACGCTGCCTGCAACCTC-3 . As a con-
trol, random sequence phosphorothioate-modified ODNswith the same proportion of bases as the ASODN se-quences were similarly prepared. These scrambled ODNsequences were 5X-CTACCGCACTCTCACCAC-3X for thescrambled a ODN and 5X-CCTACGCGAACGCCATCT-2
3X for the scrambled b ODN. The two ASODN se-3
quences have very low homology with any of the otherknown cDNA sequences found in the Genbank data base.The a ASODN produced high-scoring segment pairs2
with the a subunit of the bovine GABA receptor while2 A
the b ASODN produced high-scoring segment pairs with3
the b subunit of the mouse GABA receptor. The two3 A
scrambled ODN sequences had no homology with anyknown cDNA sequences. Additionally, there was 28%homology between the two ASODN sequences; 17% ho-mology between the a ASODN and scrambled se-2
quences; and, 6% homology between the b ASODN and3
scrambled sequences. There was 28% cross-reactivity be-tween the two ASODN sequences when all 18 base pairswere lined up 5X to 3X ends, and 6% cross-reactivitybetween the two scrambled ODN sequences when lined upin a similar manner. All oligomers were dissolved inisotonic sterile saline, aliquoted and stored at y208C.
2.4. GABA-dependent 36Cl uptake in microsacs
The methods for microsac preparation and Cl uptakew xhave been described previously 31,34 . Cocaine-naive rats
were sacrificed 24 h after ODN treatment and each side ofstriatum was rapidly removed and separately homogenized
Žin 5 ml of ice-cold tissue buffer 145 mM NaCl, 5 mMKCl, 1 mM MgCl , 10 mM dextrose, 1 mM CaCl , 102 2
mM HEPES, 10 mM leupeptin, 10 mM antipain, 10.mgrml aprotinin; pH 7.5 using a glass-teflon homoge-
Ž .nizer Thomas, size C . Homogenates were centrifuged at900=g for 15 min using a Sorvall SA600 rotor, super-natants were discarded, and each pellet was resuspended in5 ml of buffer. The samples were recentrifuged at 900= g
Ž .for 15 min and the final pellet microsacs was suspendedin 2.6 ml tissue buffer to yield 800–1000 mg of proteinper ml.
Ž .Microsacs 200 ml were incubated for 5 min at 348C,36 Ž .then 200 ml of a solution containing Cl 2.0 mCirml
with either 0 or 100 mM GABA were added and vortexedfor 3 s. Uptake was halted by adding 4.8 ml ice-cold bufferthen filtering onto a Whatman GF-C glass microfibre filter.An additional 10 ml of buffer was applied directly onto thefilter, then the amount of 36Cl retained on the filter wasdetermined via liquid scintillation counting. Protein con-
w xcentration of the microsacs was determined 4 .
2.5. QAR of GABA receptor bindingA
Rats were killed 24 h after ODN treatment and 6 h aftermeasurement of cocaine sensitivity. The brains were frozenin powdered dry ice and stored at y808C until sectioning.Thin tissue sections were cut using a cryostat at y208C toobtain coronal sections of 20 mm thickness to includestriatum at three different distances from the site of can-
w xnula placement 29 . Duplicate sections were thaw-mountedonto gelatin-coated slides and stored at y808C.
Specific binding of 35S-TBPS was used to quantify thew xconvulsant binding site on the GABA receptor 35 .A
Slide-mounted sections were thawed, dried at room tem-perature and then transferred to Coplin jars containing
Ž .tissue buffer 258C for 20 min to remove endogenousGABA. Adjacent brain sections were incubated in tissue
35 Žbuffer containing 10 nM S-TBPS 60 Cirmmol, New.England Nuclear either in the absence or presence of 100
mM picrotoxin to define total and non-specific binding.The samples were incubated at 258C for 90 min. Theincubations were terminated by rinsing each section for 5min in ice-cold buffer. Sections were dipped in distilledwater and then were dried on a hot plate at 508C. Autora-diograms of the slide-mounted sections were prepared byexposing the sections along with plastic 14C standards to
Ž .tritium-sensitive film LKB Ultrofilm for 5 days at 258C.These standards have been found to be accurate for quan-
35 w xtification of S radioligand binding 26 .Specific binding of 3H-flunitrazepam was used to quan-
titate the benzodiazepine site on the GABA receptors. InA
( )J. Peris et al.rMolecular Brain Research 57 1998 310–320 313
general, the method described above was used except thattissue was labeled with 2.6 nM 3H-flunitrazepam and 3mM clonazepam was used to define non-specific binding.The samples were incubated at 48C for 30 min followed bya 2 min rinse in ice-cold buffer and then a dip in distilledwater before drying. Tissue samples and plastic tritiumstandards were exposed to film for 7–14 days.
The illuminated image of each autoradiogram was col-lected by a CCD camera with a Nikon lens and digitized
Ž .using the Turnkey Imaging System Imaging Research .The standards included on each piece of film were used tocalibrate radioligand concentration using a Michaelis–Menton function to transform the digitized gray values into
Ž 2 .concentrations fmolrmm of radioligand bound. Thedigitized and standardized images were visualized, areas ofinterest were defined and the total and non-specific bind-ing in these areas were determined. Specific binding wasderived by subtracting non-specific binding from totalbinding. Adjacent sections from each rat brain were stainedfor Nissl bodies using Cresyl violet stain according to thestandard method.
2.6. Fluorescein-conjugated ODNs
In order to study the cellular distribution of ASODNsafter i.c.v. injection, ASODNs for a and b mRNA were2 3
obtained from the ICBR conjugated at the 5X and 3X endswith fluorescein isothiocyanate. Animals were reanes-thetized five days after cannula placement surgery and
Ž .then both ASODNs 25 mg each were administered intothe right lateral ventricle as for unconjugated ODNs. At
Ž .various time points after injection 18 or 24 h , animalswere perfused transcardially with isotonic saline and 4%formaldehyde solution. The brains were removed and storedin 4% formaldehyde solution before freezing. Coronal
Ž .brain sections 30 mm were obtained at 500 mm intervalsstarting at 2.2 to y1.7 mm Bregma which generallyincluded 1.5 mm from either side of the cannula tract.Sections were mounted and examined via laser scanningconfocal microscopy.
2.7. Data analysis
Since we have found previously that summed behav-ioral rankings from successive 10-s observation periods are
w xnormally distributed 32,33 , data from behavioral experi-ments were analyzed using ANOVA followed by Dunnett’sor Newman–Keuls comparisons where appropriate. Con-centration–response curves performed on tissue from thesame animal were compared with ANOVA. Percent con-trol data were analyzed with non-parametric statistics.
3. Results
Guide cannula were surgically implanted into the rightlateral ventricle of each rat 3–5 days before they were
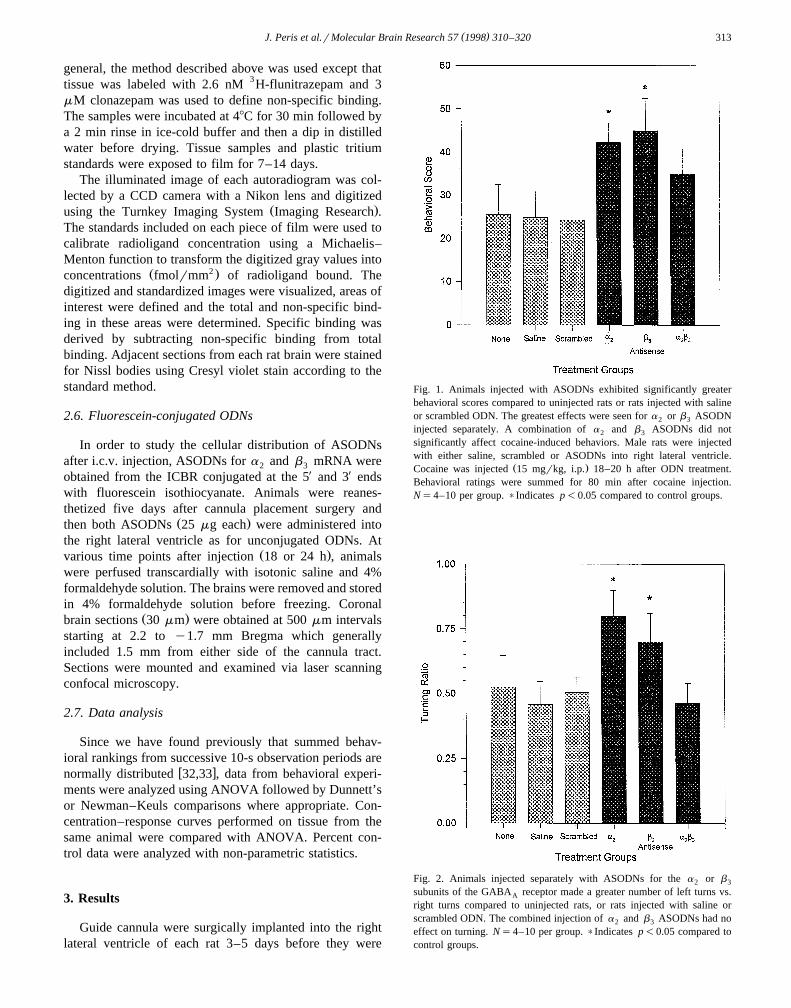
Fig. 1. Animals injected with ASODNs exhibited significantly greaterbehavioral scores compared to uninjected rats or rats injected with salineor scrambled ODN. The greatest effects were seen for a or b ASODN2 3
injected separately. A combination of a and b ASODNs did not2 3
significantly affect cocaine-induced behaviors. Male rats were injectedwith either saline, scrambled or ASODNs into right lateral ventricle.
Ž .Cocaine was injected 15 mgrkg, i.p. 18–20 h after ODN treatment.Behavioral ratings were summed for 80 min after cocaine injection.Ns4–10 per group. )Indicates p-0.05 compared to control groups.
Fig. 2. Animals injected separately with ASODNs for the a or b2 3
subunits of the GABA receptor made a greater number of left turns vs.A
right turns compared to uninjected rats, or rats injected with saline orscrambled ODN. The combined injection of a and b ASODNs had no2 3
effect on turning. Ns4–10 per group. )Indicates p-0.05 compared tocontrol groups.

( )J. Peris et al.rMolecular Brain Research 57 1998 310–320314
Fig. 3. The b ASODN injected separately in right lateral ventricle3
caused a significant 60% decrease in GABA-stimulated Cl uptake in theright striatum compared to the left striatum. A 30% decrease was seen inright striatum of rats treated with both a and b ASODNs but this was2 3
not a statistically significant difference. Male rats were injected witheither saline, scrambled or ASODNs 3–5 days after stereotaxic placementof cannula into right lateral ventricle and then killed 24 after ODNtreatment. Cl uptake was measured in left and right striatum of each rat inthe presence or absence of 100 mM GABA. )Indicates a significantgroup=side interaction. Ns3.
injected i.c.v. with either a combination of the a and b2 3ŽASODNs 25 mg each in 50 ml total isotonic saline;
.approximately 5 nmol each or each ASODN separately.There was no difference in the behavior of animals duringthe 60 min following injection of ASODN, scrambled
Ž .ODN or saline data not shown . Behavioral responses toŽ .cocaine 15 mgrkg, i.p. were measured 18–20 h after
Ž .ODN treatment Fig. 1 . Animals injected with either thea or b ASODNs alone exhibited significantly greater2 3
behavioral scores compared to non-injected rats or rats
Table 2Specific 3H-flunitrazepam binding in brains of male rats treated withASODN targeted for mRNAs of the b subunit of the GABA receptor3 A
Brain region Saline Scrambled b ASODN3
Cortex, left 1.5"0.1 1.2"0.1 1.4"0.2Cortex, right 1.3"0.1 1.1"0.1 1.3"0.1Striatum, left 0.8"0.1 0.6"0.1) 0.6"0.1)
Striatum, right 0.7"0.1 0.6"0.1) 0.6"0.1)
Total binding of 2.6 nM 3H-flunitrazepam was measured using QAR andnon-specific binding was defined by 3 mM clonazepam.Data are means"S.E.M. for Ns6 per group. ) p-0.05 compared tosaline using ANOVA.
injected with saline or scrambled ODN. There was not asignificant increase in the behavioral score of animals
Ž .injected with both a and b ASODNs Fig. 1 . These2 3
data were supported by a significant effect of TreatmentŽ Ž . .F 3,38 s3.31; p-0.05 .
Additionally, animals injected unilaterally with eitherthe a or b ASODNs made a significantly greater num-2 3
Žber of left turns vs. right turns after cocaine injection Fig..2 . Non-injected rats or rats injected unilaterally with
saline or scrambled ODNs exhibited an equal number ofŽ .left vs. right turns Fig. 2 . This difference was supported
by a significant treatment= turn direction interactionŽ Ž . .F 3,38 s3.49; p-0.05 . There was not a significantchange in the turn ratios of animals injected with both a2
Ž .and b ASODNs Fig. 2 . Similar treatment using non-3
phosphorothioated ASODNs of the same sequences had noeffect on cocaine-induced behaviors or the direction of
Ž .turning data not shown .Next, naive male rats were implanted with unilateral
cannula and 3–5 days later, treated with saline, scrambledODN, b ASODN, or a combination of a and b3 2 3
ASODN as described above. Basal and GABA-stimulatedCl uptake were measured in each side of striatum 24 hafter ODN treatment. There was significantly less GABA-stimulated Cl uptake in right striatum compared to leftstriatum of rats treated with b ASODN while there was3
Table 1Specific 35S-TBPS binding in brains of male rats treated with ASODNs targeted for specific mRNAs of GABA receptor subunitsA
Brain region Saline Scrambled b ASODN a b ASODN3 2 3
a aCortex, left 27.3"1.5 22.6"1.6 21.8"2.7 28.7"1.1Cortex, right 24.1"1.0) 18.3"1.7) 18.2"2.1) 21.7"1.6)
a aD. striatum, left 14.4"1.0 11.4"0.8 9.9"1.8 12.6"0.3D. striatum, right 15.8"1.0 9.7"1.3 9.6"1.7 10.7"0.8)
a aM. striatum, left 12.2"0.5 9.9"0.4 8.8"1.6 10.3"0.7M. striatum, right 12.8"1.3 7.6"0.9 7.5"1.5) 8.6"0.6)
VL. striatum, left 15.7"1.5 13.9"1.5 12.7"1.8 13.3"1.3VL. striatum, right 17.5"1.9 11.3"1.0 11.5"1.3 12.0"1.7Globus pallidus, left 59.6"4.4 52.2"8.3 48.6"9.8 70.1"6.4Globus pallidus, right 70.9"4.1 54.3"4.3 47.1"5.6 67.2"6.8
Total binding of 10 nM 35S-TBPS was measured using QAR and non-specific binding was defined by 100 mM picrotoxin.Data are means"S.E.M. for Ns6 per group. ) p-0.05 compared to left side, ap-0.05 for both sides compared to saline group using ANOVA.Msmedial, Dsdorsal, VLsventrolateral.
( )J. Peris et al.rMolecular Brain Research 57 1998 310–320 315
no significant difference between sides in control animalsŽ .or animals treated with both a and b ASODN Fig. 3 .2 3
These findings were supported by a significant interaction
Ž Ž . .of treatment=side F 2,6 s6.88; p-0.05 which wasdue to a significant main effect of side in the b ASODN3
Ž Ž . .group only F 1,2 s34.97; p-0.05 . There were no
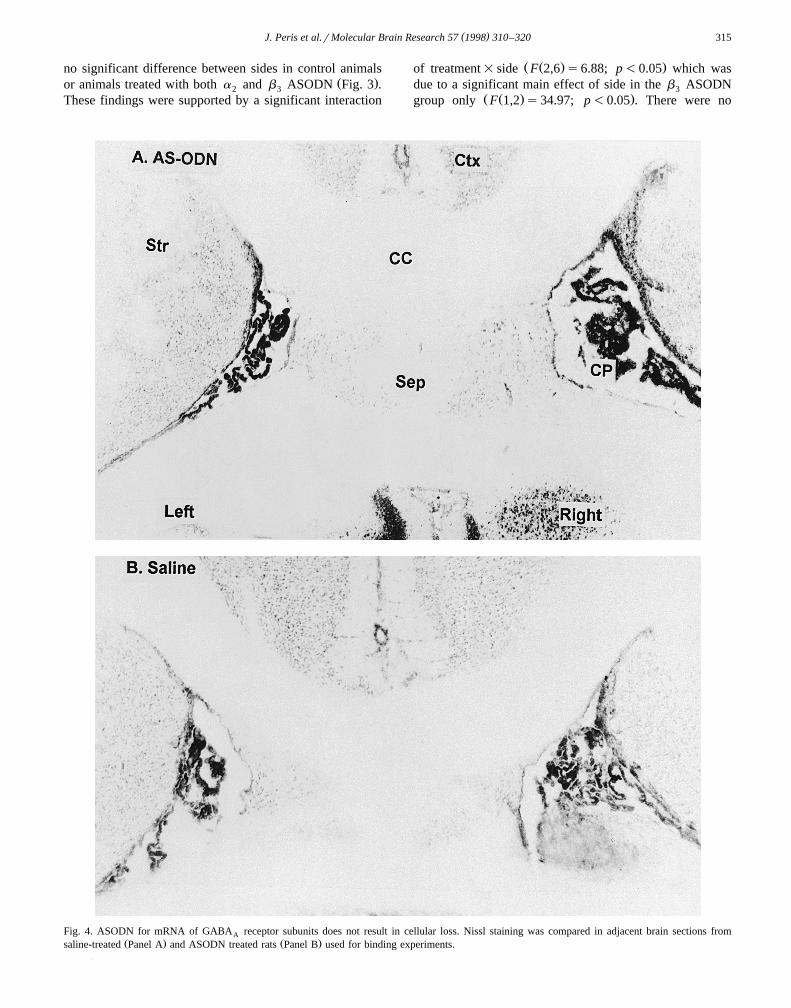
Fig. 4. ASODN for mRNA of GABA receptor subunits does not result in cellular loss. Nissl staining was compared in adjacent brain sections fromAŽ . Ž .saline-treated Panel A and ASODN treated rats Panel B used for binding experiments.
( )J. Peris et al.rMolecular Brain Research 57 1998 310–320316
Table 3Phosphorothioated ASODN for mRNA of the b subunit of the GABA3 A
receptor does not cause neurotoxicity
Group Dorsal Ventral
Left Right Left Right
Saline 67"6 64"6 63"7 65"8ASODN 67"5 69"5 69"4 67"5
Nissl staining was used to visualize the number of cells in striatal areasbordering the lateral ventricle of brains from saline and b ASODN3
treated rats. There was no difference between the number of stained cellsper 0.15 mm2 area selected from left and right edges of striatum fromeither dorsal or ventral sections. Ns4–6.
significant interactions or main effects found when animalstreated with both a and b ASODN were compared to2 3
control rats. There were no significant effects of eitherASODN treatment on basal Cl uptake measured in the
Ž .absence of GABA data not shown .
When specific binding to the convulsant and benzodi-azepine sites on the GABA receptor was measured 24 hA
after ODN injection, there were no decreases in bindingselectively associated with any of the ASODN treatments
Ž .compared to the control treatments Tables 1 and 2 .Specific 35S-TBPS binding was decreased in cortex, dorsalstriatum and medial striatum of rats treated with the b3
ASODN treatment compared to rats injected with salinebut a similar decrease was seen in these regions in rats
Ž .receiving injections of scrambled ODNs Table 1 . Therewas also a significant decrease in the amount of specificbinding of 35S-TBPS on the right side vs. the left side ofcortex, dorsal striatum and medial striatum in rats injected
Ž .with the ASODN treatments Table 1 . However, therewere also significant differences between right and leftsides of cortex in saline and scrambled ODN control ratsŽ .Table 1 . Although there were no significant decreases inright striatum vs. left in control rats, there was a trend for a
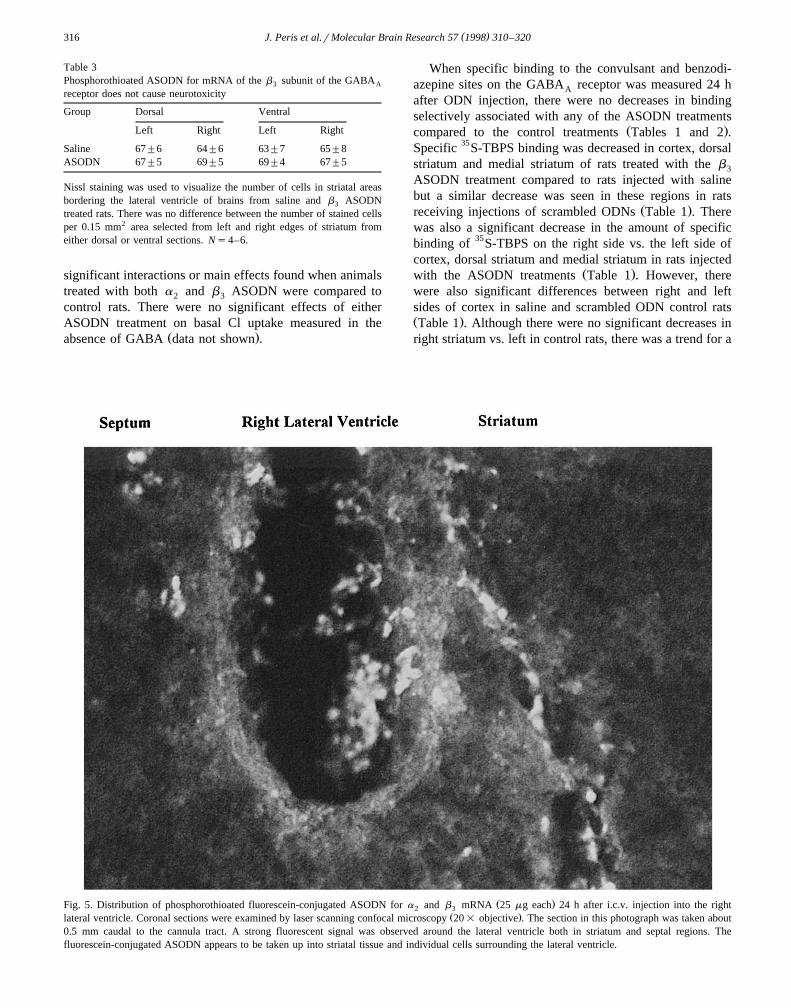
Ž .Fig. 5. Distribution of phosphorothioated fluorescein-conjugated ASODN for a and b mRNA 25 mg each 24 h after i.c.v. injection into the right2 3Ž .lateral ventricle. Coronal sections were examined by laser scanning confocal microscopy 20= objective . The section in this photograph was taken about
0.5 mm caudal to the cannula tract. A strong fluorescent signal was observed around the lateral ventricle both in striatum and septal regions. Thefluorescein-conjugated ASODN appears to be taken up into striatal tissue and individual cells surrounding the lateral ventricle.

( )J. Peris et al.rMolecular Brain Research 57 1998 310–320 317
decrease in these rats which was of the same magnitude asŽ .the decrease in ASODN-treated rats Table 1 . The same
was true when specific 3H-flunitrazepam binding was mea-Ž .sured Table 2 . Although there was a significant decrease
in specific binding in striatum of animals treated withASODN compared to saline treated rats, a similar decrease
Žwas seen in animals treated with scrambled ODN Table.2 .
In order to confirm that ODN treatment did not result incellular damage, Nissl staining was used to visualize thenumber of cells in striatal areas bordering the lateralventricle of brains from saline and ASODN treated ratsŽ .Fig. 4 . There was no difference between the number ofstained cells per 0.15 mm2 area selected from left and rightedges of striatum from either dorsal or ventral horizontal
Ž .sections Table 3 . Similar results were found with alimited number of sections from brains of rats injected
Ž .with the scrambled ODN sequences data not shown .The cellular distribution of i.c.v.-injected ASODNs was
studied using fluorescein-conjugated a and b ASODN2 3Ž .25 mg each administered into the right lateral ventricle.Fluorescent signal was distributed predominantly around
Ž .the right lateral ventricle Fig. 5 although weak fluores-Žcence was present on the left side of some sections data
.not shown . At both 18 and 24 h after injection, fluores-cence was present in all sections extending up to 1.5 mmfrom the cannula tract and was densest in brain regionsnearest the most ventral portion of lateral ventricle both instriatum and lateral septum. Although signal was greatestclosest to the lateral ventricle, there was penetration intoeach structure with some evidence of ASODN uptake into
Ž .individual cells Fig. 5 .
4. Discussion
If the level of GABA -receptor mediated inhibition inAw xstriatum is inversely related to cocaine responsiveness 32 ,
then treatments that decrease GABA receptor number orA
function should ‘sensitize’ animals to cocaine. Animalsinjected in the right lateral ventricle with ASODNs tar-geted for the a or b mRNA of the GABA receptor2 3 A
exhibited significantly greater behavioral scores comparedto control rats. Additionally, these animals made a greaternumber of left turns vs. right turns compared to controlrats. Since separate injection of the a and b ASODNs2 3
both effectively increased cocaine-induced behaviors, weexpected even greater effects when both ASODNs wereinjected together. However, a b ASODN had no effect2 3
on cocaine-induced behaviors and turning.One of the ASODN treatments that affected cocaine
sensitivity also appeared to decrease GABA receptorA
function. There was significantly less GABA-stimulated Cluptake in right striatum compared to left striatum of b3
ASODN treated rats while there was no significant differ-ence between sides in control animals. Injection of a b2 3
ASODN caused a small decrease in GABA receptorA
function in right striatum which was not statistically differ-ent from that in control rats. Data from receptor bindingexperiments did not indicate a selective loss of convulsantor benzodiazepine sites associated with ASODN treatmentin striatum or cortex. Although there were some small butsignificant decreases caused by ASODN treatment, therewere decreases of similar magnitude in animals receivingscrambled ODN sequences. Thus, it appears that the de-crease in receptor function found in animals injected withb ASODN may not have been due to down regulation of3
receptors as a whole but instead to a decrease in the abilityof GABA to functionally stimulate uptake in ASODN-treated tissue perhaps due to a change in receptor composi-tion induced by ASODN treatment. The change in subunitcomposition resulting in decreased receptor function maybe due to decreased translation of the b subunit caused3
directly by the ASODN treatment or it may be due to theeffects of newly synthesized subunits that occur in re-
w w xxsponse to the loss of the b subunits see Ref. 45 . The3
effects of unilateral injection of the a ASODN alone on2
GABA receptor binding and ionophore function were notA
measured in these experiments but are of interest in futureexperiments.
Thus, ASODN treatments that are effective in decreas-ing the function of striatal GABA receptors also increaseA
the behavioral responses elicited by cocaine. Similar re-sults have been obtained using mice that are missing theb subunit of the GABA receptor via genetic knockout3 Aw x11 . These animals exhibit increased cocaine sensitivity aswell as decreased GABA receptor function in striatumAw x36 supporting our hypothesis that a decrease in thenumber of b receptor subunits in striatum can increase3
behavioral responses to cocaine. Additionally, in the pre-sent study, ASODN treatments that successfully caused aunilateral decrease in striatal GABA receptor functionA
resulted in contralateral turning in response to cocaine.These data are consistent with studies finding that unilat-
Žeral picrotoxin or bicuculline injection into striatum which.would inactivate GABA receptors in striatum causesA
w xcontralateral rotation 25 . Thus, our data supports previousevidence in the literature that the functional status ofGABA receptors in striatum may affect cocaine sensitiv-A
ity.It is not clear why the combination of a b ASODN2 3
had only marginal effects on cocaine-induced behaviorsand GABA receptor function. Separate injection of eitherA
a or b ASODN was effective both neurochemically and2 3
behaviorally and it was expected that combined ASODNtreatment should be at least equally effective as the sepa-rate ASODN treatments. At this point, many issues ofantisense technology are not well understood, therefore,combined administration of ODNs may encounter a num-
w w xxber of unknown difficulties see Ref. 5 .The dose of ODNs chosen was selected to maximize
our chances of successfully decreasing mRNA translation.

( )J. Peris et al.rMolecular Brain Research 57 1998 310–320318
Unfortunately, this dose might have increased the risk ofhaving non-selective or neurotoxic effects. Nissl stainingindicated that there was not a significant amount of cellloss in the regions of striatum exposed to ASODN. Addi-tionally, although the injected amount of ASODN washigh, the amount actually reaching and remaining at thesite of action may have been lowered from volume dilutionthereby minimizing neurotoxic effects. These data aresimilar to previous reports in the literature that indicateone-time exposure of these levels of phosphorothioatedODNs does not result in significant amounts of neurotoxic-
w xity 6 .The data from the i.c.v. injection of fluorescein-labeled
ASODN indicate there was significant uptake of label intostriatal cells bordering right lateral ventricle at the timesthat behavioral and neurochemical effects of non-fluo-rescein-labeled ASODN injection were seen. Althoughthere is the possibility that fluorescein labeling may havealtered the distribution of the ASODN, these data indicatethat the dose and route of administration of ASODN wassufficient to continue to have functional effects 18–24 hafter injection without significant neurotoxicity. Similarfindings have been reported using unilateral injection ofsimilar amounts of phosphorothioated ASODN for c-fos
w xmRNA 12 . Unilateral intrastriatal ASODN injections inthat experiment resulted in ipsilateral turning after cocaine
w xinjection with little evidence of neurotoxicity 12 . Thus,the contralateral turning seen in our experiment was proba-bly not due to a non-selective or neurotoxic action ofunilateral injection of phosphorothioated ODNs.
Activation of GABA receptors in striatum decreasesAw xDA release from synaptosomal preparations 38 providing
evidence that presynaptic GABA receptors can inhibit theA
activity of nigrostriatal DA terminals. In support of thishypothesis, striatal DA output and stereotypic movementsare decreased by GABA agonists injected into striatumw w xxsee Ref. 22 . Bilateral striatal injections of GABA antag-
w xonists increase stereotypic jaw movements 19 while uni-lateral striatal injections of antagonists cause contralateral
w xrotation 25 . On the other hand, bicuculline decreases thepercentage of animals exhibiting stereotypic movementsafter an acute injection of 100 mg cocaine or amphetaminew x17 . Thus, the activity of GABA receptors in striatumA
appears to be important for determining the extent ofDA-stimulated stereotypies such as those seen after co-caine sensitization however the exact nature of that in-volvement is still unknown.
GABA receptors may also be located on striatal in-AŽ .terneurons or recurrent collaterals , GABAergic projection
neurons to SNR or GABAergic projection neurons toglobus pallidus thereby complicating how changes in thesereceptors will affect DA transmission and subsequent loco-motor behavior. As mentioned above, a decrease inGABA receptors on DA terminals should enhance DAA
Ž .release and locomotion , whereas a decrease in GABAA
receptors on GABA interneurons would increase inhibition
of DA terminals and decrease locomotion. Similarly, adecrease in GABA receptors on striatonigral efferentsA
should increase locomotion whereas a decrease in GABAA
receptors on striatopallidal neurons should decrease loco-w w xxmotion see Ref. 39 . It is possible that the composition
of GABA receptors varies across these neuronal popula-A
tions in striatum thereby making them selectively suscepti-ble to ASODN inhibition. Thus, it will be very importantto determine the cellular location of GABA receptors andA
their structural composition before concluding that a lossof GABA inhibition should enhance cocaine’s effects.
It is possible that the behavioral effects of the ASODNmight be due to decreases in non-striatal GABA receptorA
function. There were no effects of ASODN treatment todecrease receptor function or receptor binding in cortex,however other regions besides cortex and striatum werenot studied in these assays. The fluorescein-labeledASODN experiments indicated there was significant up-take of fluorescence label into lateral septum as well asinto striatum. Therefore, a decrease in GABA receptorA
function in lateral septum cannot be excluded as an expla-nation for the effect of these ASODNs on cocaine-inducedbehaviors in these experiments.
In summary, unilateral injection of ASODNs selectivefor the particular subunits of the GABA receptor found inA
striatum is effective for decreasing the functional activa-tion of GABA receptors in striatum. Such treatmentsA
result in an increase in the behavioral response to cocaineincluding asymmetric behaviors induced by unilateralchanges in striatal GABA receptor function. These dataA
support the hypothesis that the functional status of theGABA receptor in striatum is important for determiningA
the behavioral response to cocaine. It is possible that thisrelationship may provide important insight into howchanges in the normal functioning of striatum may alterthe response to both initial and repeated exposure tococaine.
Acknowledgements
This research was supported by grants from the Alco-holic Beverage Medical Research Foundation and PHSgrant AA 09115. Bruce Jung was supported by NIAAATraining Grant AA 07561. Technical assistance was pro-vided by Shawn Toffolo.
References
w x1 S. Ahktar, R. Kole, R.L. Juliano, Stability of antisense DNAoligodeoxynucleotide analogs in cellular extracts and sera, Life Sci.
Ž .49 1991 1793–1801.w x2 S.L. Beaucage, M.H. Caruthers, Deoxynucleoside phosphoramidite:
a new class of key intermediates for deoxypolynucleotide synthesis,Ž .Tetrahedron Lett. 22 1981 1859–1864.
w x3 L.A. Borden, D.H. Farb, Mechanism of g-aminobutyric acidrbenzo-

( )J. Peris et al.rMolecular Brain Research 57 1998 310–320 319
diazepine receptor turnover in neuronal cells: evidence for non-Ž .lysosomal degradation, Mol. Pharmacol. 34 1988 354–362.
w x4 M.M. Bradford, A rapid and sensitive method for the quantitation ofmicrogram quantities of protein utilizing the principle of protein-dye
Ž .binding, Anal. Biochem. 72 1976 248–254.w x5 A.D. Branch, A good antisense is hard to find, Trends Biochem. Sci.
Ž .23 1998 45–50.w x6 B.J. Chiasson, J.N. Armstrong, M.L. Hooper, P.R. Murphy, H.A.
Robertson, The application of antisense oligonucleotide technologyŽ .to the brain, Cell. Mol. Neurobiol. 14 1994 507–521.
w x7 J.M. Fritschy, H. Mohler, GABA -receptor heterogeneity in theA
adult rat brain: differential regional and cellular distribution of sevenŽ .major subunits, J. Comp. Neurol. 359 1995 154–194.
w x8 K. Gale, D. Marshall, H. Bernstein, J. Butler, Effects of chroniccocaine administration on nigrostriatal function: neurochemical and
Ž .behavioral changes in rats, Fed. Proc. 40 1981 291.w x9 R. Gyurko, D. Wielbo, M.I. Phillips, Antisense inhibition of AT1
mRNA and angiotensinogen mRNA in the brain of spontaneouslyhypertensive rats reduces hypertension of neurogenic origin, Regul.
Ž .Pept. 49 1993 167–174.w x10 C. Helene, J.J. Toulme, Specific regulation of gene expression by
antisense, sense and antigene nucleic acids, Biochim. Biophys. ActaŽ .1049 1990 99–125.
w x11 G.E. Homanics, T.M. DeLorey, L.L. Firestone, J.J. Quinlan, A.Handforth, N.L. Harrison, M.D. Krasowski, C.E.M. Rick, E.R.Korpi, R. Makela, M.H. Brilliant, N. Hagiwara, C.F. Ferguson, K.Snyder, R.W. Olsen, Mice devoid of g-aminobutyrate type A recep-tor b subunit have epilepsy, cleft palate and hypersensitive behav-3
Ž .ior, PNAS 94 1997 4143–4148.w x12 M.L. Hooper, B.J. Chiasson, H.A. Robertson, Infusion into the brain
of an antisense oligonucleotide to the immediate early gene c-fossuppresses production of Fos and produces a behavioral effect,
Ž .Neuroscience 63 1994 917–924.w x13 B.J. Jung, S.A. Sealey, P.A. Walker, J. Peris, Endogenous GABA
release from striatal slices of cocaine sensitized rats, Soc. Neurosci.Ž .Abstr. 22 1996 929.
w x14 J. Karle, M. Nielsen, Modest reduction of benzodiazepine binding inrat brain in vivo induced by antisense oligonucleotide to GABAA
Ž .receptor g subunit, Eur. J. Pharmacol. 291 1995 439–441.2w x15 J. Karle, M.R. Witt, M. Nielsen, Antisense oligonucleotide to
GABA receptor g subunit induces loss of neurones in rat hip-A 2Ž .pocampus, Neurosci. Lett. 202 1995 97–100.
w x16 J. Karle, M.R. Witt, M. Nielsen, The use of in vivo antisenseoligonucleotide technology for the investigation of brain GABA
Ž .receptors, Neurochem. Int. 31 1997 437–446.w x17 R. Karler, L.D. Calder, L.H. Thai, J.B. Bedingfield, The dopaminer-
gic, glutamatergic, and GABAergic bases for the action of am-Ž .phetamine and cocaine, Brain Res. 671 1995 100–104.
w x18 M. Khrestchatisky, A.J. MacLennan, N.J.K. Tillakaratne, M. Chi-ang, A.J. Tobin, Sequence and regional distribution of the mRNAencoding the a polypeptide of rat GABA receptors, J. Neu-2 A
Ž .rochem. 56 1991 1717–1722.w x19 K. Kikuchi de Beltran, N. Koshikawa, Y. Miwa, M. Kobayashi, J.D.
Stephenson, Further evidence for a functional dorsal–ventral divi-sion of the rat striatum: GABAergic involvement in oral movements,
Ž .Eur. J. Pharmacol. 239 1993 47–54.w x20 R. Kole, R. Shukla, S. Ahktar, Pre-mRNA splicing as a target for
Ž .antisense oligonucleotides, Adv. Drug Deliv. Rev. 6 1991 271–286.w x21 D.J. Laurie, P.H. Seeburg, W. Wisden, The distribution of 13
GABA receptor subunit mRNAs in the rat brain: II. Olfactory bulbAŽ .and cerebellum, J. Neurosci. 12 1992 1063–1076.
w x22 K.G. Lloyd, P. Pichat, B. Scatton, B. Zivkovic, P.L. Morselli, G.Bartholini, The psychopharmacology of GABA synapses: update
Ž .1989, J. Neural. Transm. 29 1990 S13–S28.w x23 S.L. Loke, C.A. Stein, X.H. Zhang, K. Mori, M. Nakanishi, C.
Subasinghe, J.S. Cohen, L. Neckers, Characterization of oligo-Ž .nucleotide transport into living cells, PNAS 86 1989 3474–3478.
w x24 R.D. Mayfield, S. Leonard, T.K. Booker, N.R. Zahniser, Antisenseoligonucleotide to the g subunit of the GABA receptor decreases2 A
mouse brain benzodiazepine binding, Alcohol. Clin. Exp. Res. 18Ž .1994 446.
w x25 J.S. McKenzie, A.D. Shafton, C.A. Stewart, Intrastriatal dopaminer-gic agents, muscarinic stimulation, and GABA antagonism com-
Ž .pared for rotation responses in rats, Behav. Brain Res. 45 1991163–170.
w x 35 32 1426 J.A. Miller, The calibration of S or P with C-labeled brainpaste or 14C-plastic standards for quantitative autoradiography using
Ž .LKB ultrafilm or Amersham hyperfilm, Neurosci. Lett. 121 1991211–214.
w x27 N. Nayeem, T.P. Green, I.L. Martin, E.A. Barnard, Quaternarystructure of the native GABA receptor determined by electronA
Ž .microscopy image analysis, J. Neurochem. 62 1994 815–818.w x28 R.W. Olsen, R.T. McCabe, J.K. Wamsley, GABA receptor sub-A
types: autoradiographic comparison of GABA, benzodiazepine, andŽ .convulsant binding in the rat CNS, J. Chem. Neuroanat. 3 1990
59–76.w x Ž .29 G. Paxinos, C. Watson Eds. , The Rat Brain in Stereotaxic Coordi-
nates, Academic Press, New York, 1986.w x30 M. Pecins-Thompson, J. Peris, Behavioral and neurochemical
changes caused by repeated ethanol and cocaine administration,Ž .Psychopharmacology 110 1993 443–450.
w x31 J. Peris, Antagonist inhibition of GABA receptor-mediated chlo-AŽ .ride uptake is atypical in nigrocollicular areas, Life Sci. 53 1993
707–716.w x32 J. Peris, Repeated cocaine injections decrease the function of striatal
Ž .GABA receptors, J. Pharmacol. Exp. Ther. 276 1996 1002–1008.Aw x33 J. Peris, S.J. Boyson, W.A. Cass, P. Curella, L.P. Dwoskin, G.
Larson, L.-H. Linn, R.P. Yasuda, N.R. Zahniser, Persistence ofneurochemical changes in dopamine systems after repeated cocaine
Ž .administration, J. Pharmacol. Exp. Ther. 253 1990 38–44.w x34 J. Peris, A. Shawley, R. Dawson, K.H. Abendschein, Regulation of
35S-TBPS binding by bicuculline is region specific in rat brain, LifeŽ .Sci. 49 1991 PL49–PL54.
w x35 J. Peris, M.L. Coleman-Hardee, J. Burry, M. Pecins-Thompson,Selective changes in GABAergic transmission in substantia nigraand superior colliculus caused by ethanol and ethanol withdrawal,
Ž .Alcohol. Clin. Exp. Res. 16 1992 311–319.w x36 J. Peris, G.E. Homanics, B.J. Jung, Heterozygous knockout mice for
the GABA receptor b subunit have increased behavioral sensitiv-A 3Ž .ity to cocaine, Soc. Neurosci. Abstr. 23 1997 2411.
w x37 L.E. Rabow, S.J. Russek, D.H. Farb, From ion currents to genomicanalysis: recent advances in GABA receptor research, Synapse 21AŽ .1995 189–274.
w x38 E. Ronken, A.H. Mulder, A.N. Schoffelmeer, Interacting presynaptick-opioid and GABA receptors modulate dopamine release from ratA
Ž .striatal synaptosomes, J. Neurochem. 61 1993 1634–1639.w x39 W. Sommer, R. Rimondini, W. O’Connor, A.C. Hansson, U. Under-
stedt, K. Fuxe, Intrastriatally injected c-fos antisense oligonucleotideinterferes with striatonigral but not striatopallidal GABA transmis-
Ž .sion in the conscious rat, PNAS 93 1996 14134–14139.w x40 C.A. Stein, J.S. Cohen, Oligodeoxynucleotides as inhibitors of gene
Ž .expression: a review, Cancer Res. 48 1988 2659–2668.w x41 C.A. Stein, K. Mori, S.L. Loke, C. Subasinghe, K. Shinozuka, J.S.
Cohen, L. Neckers, Phosphorothioate and normal oligodeoxyribonu-cleotides with SX linked acridine: characterization and preliminary
Ž .kinetics of cellular uptake, Gene 72 1988 333–341.w x42 D.M. Tidd, A potential role for antisense oligonucleotide analogues
in the development of oncogene targeted cancer chemotherapy,Ž .Anticancer Res. 10 1990 1169–1182.
w x43 C. Wahlestedt, E. Golanov, S. Yamamoto, F. Yee, H. Ericson, H.Yoo, C.E. Inturrisi, D.J. Reis, Antisense oligodeoxynucleotides toNMDA-R1 receptor channel protect cortical neurons from excitotox-
Ž .icity and reduce focal ischaemic infarctions, Nature 363 1993260–263.

( )J. Peris et al.rMolecular Brain Research 57 1998 310–320320
w x44 C. Wahlestedt, E.M. Pich, G.F. Koob, F. Yee, M. Heilig, Modula-tion of anxiety and neuropeptide Y–Y1 receptors by antisense
Ž .oligodeoxynucleotides, Science 259 1993 528–531.w x45 B. Weiss, G. Davidkova, S.P. Zhang, Antisense strategies in neuro-
Ž .biology, Neurochem. Int. 31 1997 321–348.w x46 L. Whitesell, D. Geselowitz, C. Chavany, B. Fahmy, S. Walbridge,
J.R. Algers, L.M. Neckers, Stability, clearance, and disposition ofintraventricularly administered oligodeoxynucleotides: implicationsfor therapeutic application within the central nervous system, PNAS
Ž .90 1993 4665–4669.w x47 E.J. Wickstrom, Oligodeoxynucleotide stability in subcellular ex-
Ž .tracts and culture media, Biochem. Biophys. Meth. 13 1986 97–102.
w x48 D. Wielbo, C. Sernia, M.I. Phillips, The distribution and antihyper-tensive effects of phosphorothioated antisense oligodeoxynucleotides
Ž .in SHR, Hypertension 25 1995 314–319.w x49 W. Wisden, D.J. Laurie, H. Monyer, P.H. Seeburg, The distribution
of 13 GABA receptor subunit mRNAs in the rat brain: I. Telen-AŽ .cephalon, diencephalon, mesencephalon, J. Neurosci. 12 1992
1040–1062.w x50 X. Xiao, T.J. McCown, J. Li, G.R. Breese, A.L. Morrow, R.J.
Ž .Samulski, Adeno-associated virus AAV vector antisense gene
transfer in vivo decreases GABA a containing receptors andA 1Ž .increases inferior collicular seizure sensitivity, Brain Res. 756 1997
76–83.w x51 L.A. Yakubov, E.A. Deeva, V.F. Zarytova, E.M. Ivanova, A.S.
Ryte, L.V. Yurchenko, V.V. Vlassov, Mechanism of oligonucleotideŽ .uptake by cells: involvement of specific receptors, PNAS 86 1989
6454–6458.w x52 S. Ymer, P.R. Schofield, A. Draguhn, P. Werner, M. Kohler, P.H.
Seeburg, GABA receptor b subunit heterogeneity: functional ex-AŽ .pression of cloned cDNAs, EMBO J. 8 1989 1665–1670.
w x53 J. Zezula, A. Slany, W. Sieghart, Interaction of allosteric ligandswith GABA receptors containing one, two or three different sub-A
Ž .units, Eur. J. Pharmacol. 301 1996 207–214.w x54 T.J. Zhao, H.C. Rosenberg, T.H. Chiu, Treatment with antisense
oligodeoxynucleotide to the GABA receptor g subunit increasesA 2
convulsive threshold for b-CCM, a benzodiazepine inverse agonist,Ž .in rats, Eur. J. Pharmacol. 306 1996 61–66.
w x55 L.-W. Zhou, S.-P. Zhang, Z.-H. Qin, B. Weiss, In vivo administra-tion of an oligodeoxynucleotide antisense to the D dopamine2
receptor messenger RNA inhibits D dopamine receptor-mediated2
behavior and the expression of D dopamine receptors in mouse2Ž .striatum, J. Pharmacol. Exp. Ther. 268 1994 1015–1023.
Copyright © 2022 FDOKUMEN




















