
Antifibroblast antibodies from systemic sclerosis patients are internalized by fibroblasts via a...
7
ARTHRITIS & RHEUMATISM Vol. 46, No. 6, June 2002, pp 1595–1601 DOI 10.1002/art.10362 © 2002, American College of Rheumatology Antifibroblast Antibodies From Systemic Sclerosis Patients Are Internalized by Fibroblasts via a Caveolin-Linked Pathway Nicoletta Ronda, 1 Rita Gatti, 1 Roberto Giacosa, 1 Elena Raschi, 2 Cinzia Testoni, 2 Pier Luigi Meroni, 2 Carlo Buzio, 1 and Guido Orlandini 1 Objective. Fibroblast activation is a crucial event in the development of systemic sclerosis (SSc). Antifi- broblast autoantibodies (AFAs), detectable in the sera of SSc patients, are able to induce a proinflammatory phenotype on cultured fibroblasts. This study was un- dertaken to investigate the mechanisms of the interac- tion between AFAs and living fibroblasts. Methods. We coupled to fluorescein 1) IgG puri- fied from AFA-positive and AFA-negative SSc sera (as assessed by cellular enzyme-linked immunosorbent as- say) and 2) single healthy donor and pooled normal IgG. The interaction of IgG with living cultured fibroblasts from healthy individuals and from a patient with SSc was visualized by real-time confocal microscopy. Intra- cellular colocalization of caveolin and internalized AFA- positive IgG was assessed by immunofluorescence. Results. AFA-positive IgG bound to living fibro- blasts and was internalized with a cytoplasmic fibrillar pattern, in contrast to AFA-negative IgG. In the IgG tested, no correlation with antinuclear antibody activity was found. Preincubation of fibroblasts with normal IgG did not affect internalization. Internalized AFA- positive IgG colocalized with caveolin, and internaliza- tion was entirely inhibited by disassembling fibroblast caveolae with filipin. Conclusion. The finding that both normal and pathologic fibroblasts specifically internalized AFA- positive, but not AFA-negative, IgG demonstrates that AFAs in SSc patient sera interact with constitutively expressed membrane molecules on fibroblasts, via an Fc-independent mechanism. The results of colocaliza- tion and inhibition experiments suggest that microdo- mains containing caveolin are involved in the interac- tion between AFAs and fibroblasts. These data, together with the reported ability of AFAs to activate fibroblasts, provide evidence for a role of AFAs in the pathogenesis of SSc. Systemic sclerosis (SSc) is characterized by fibro- sis of skin and internal organs, which develops mainly from lesions of the microvascular system, including endothelial injury, increased vascular wall thickness, and infiltration of monocytes and T cells adjacent to acti- vated fibroblasts (1–3). The initial event leading to inflammatory cell recruitment is not known, but genetic factors (4), infectious agents (3), and microchimerism (5) seem to be involved in stimulating the immune system. The production of autoantibodies, such as anti- nuclear antibodies (ANAs) (anti–topoisomerase I, anti– RNA polymerase III, and anticentromere proteins) (4), and antibodies recognizing surface antigens on lympho- cytes, endothelial cells (6–8), and fibroblasts (9,10) is a notable immunologic abnormality in SSc. In particular, antifibroblast antibodies (AFAs), which induce fibro- blast activation in vitro, are present in 46% of patients with SSc and stimulate monocyte adhesion to fibroblasts by up-regulating intercellular adhesion molecule 1 (ICAM-1), fibroblast secretion of interleukin-1 (IL-1) and monocyte chemotactic protein 1, and the steady- state increase of messenger RNA for cytokines and chemokines (11). Indeed, fibroblasts are abnormally activated in SSc, showing increased production of cyto- kines and growth factors involved in fibrogenesis and abnormal collagen turnover (12). Supported by MURST Cofinanziamento 1998 and FIL2000 (to Drs. Meroni and Orlandini) and by Ricerca Corrente 2000 Istituto Auxologico Italiano (to Dr. Meroni). 1 Nicoletta Ronda, MD, PhD, Rita Gatti, PhD, Roberto Giacosa, MD, Carlo Buzio, MD, Guido Orlandini, MD: University of Parma, Parma, Italy; 2 Elena Raschi, PhD, Cinzia Testoni, PhD, Pier Luigi Meroni, MD: University of Milan, IRCCS Istituto Auxologico Italiano, Milan, Italy. Address correspondence and reprint requests to Nicoletta Ronda, MD, PhD, Department of Clinical Medicine, Nephrology and Health Sciences, University of Parma, Via Gramsci 14, 43100 Parma, Italy. E-mail: [email protected]. Submitted for publication October 9, 2001; accepted in re- vised form March 1, 2002. 1595
Transcript of Antifibroblast antibodies from systemic sclerosis patients are internalized by fibroblasts via a...

ARTHRITIS & RHEUMATISMVol. 46, No. 6, June 2002, pp 1595–1601DOI 10.1002/art.10362© 2002, American College of Rheumatology
Antifibroblast Antibodies From Systemic Sclerosis Patients AreInternalized by Fibroblasts via a Caveolin-Linked Pathway
Nicoletta Ronda,1 Rita Gatti,1 Roberto Giacosa,1 Elena Raschi,2 Cinzia Testoni,2
Pier Luigi Meroni,2 Carlo Buzio,1 and Guido Orlandini1
Objective. Fibroblast activation is a crucial eventin the development of systemic sclerosis (SSc). Antifi-broblast autoantibodies (AFAs), detectable in the seraof SSc patients, are able to induce a proinflammatoryphenotype on cultured fibroblasts. This study was un-dertaken to investigate the mechanisms of the interac-tion between AFAs and living fibroblasts.
Methods. We coupled to fluorescein 1) IgG puri-fied from AFA-positive and AFA-negative SSc sera (asassessed by cellular enzyme-linked immunosorbent as-say) and 2) single healthy donor and pooled normal IgG.The interaction of IgG with living cultured fibroblastsfrom healthy individuals and from a patient with SScwas visualized by real-time confocal microscopy. Intra-cellular colocalization of caveolin and internalized AFA-positive IgG was assessed by immunofluorescence.
Results. AFA-positive IgG bound to living fibro-blasts and was internalized with a cytoplasmic fibrillarpattern, in contrast to AFA-negative IgG. In the IgGtested, no correlation with antinuclear antibody activitywas found. Preincubation of fibroblasts with normalIgG did not affect internalization. Internalized AFA-positive IgG colocalized with caveolin, and internaliza-tion was entirely inhibited by disassembling fibroblastcaveolae with filipin.
Conclusion. The finding that both normal andpathologic fibroblasts specifically internalized AFA-
positive, but not AFA-negative, IgG demonstrates thatAFAs in SSc patient sera interact with constitutivelyexpressed membrane molecules on fibroblasts, via anFc-independent mechanism. The results of colocaliza-tion and inhibition experiments suggest that microdo-mains containing caveolin are involved in the interac-tion between AFAs and fibroblasts. These data, togetherwith the reported ability of AFAs to activate fibroblasts,provide evidence for a role of AFAs in the pathogenesisof SSc.
Systemic sclerosis (SSc) is characterized by fibro-sis of skin and internal organs, which develops mainlyfrom lesions of the microvascular system, includingendothelial injury, increased vascular wall thickness, andinfiltration of monocytes and T cells adjacent to acti-vated fibroblasts (1–3). The initial event leading toinflammatory cell recruitment is not known, but geneticfactors (4), infectious agents (3), and microchimerism(5) seem to be involved in stimulating the immunesystem.
The production of autoantibodies, such as anti-nuclear antibodies (ANAs) (anti–topoisomerase I, anti–RNA polymerase III, and anticentromere proteins) (4),and antibodies recognizing surface antigens on lympho-cytes, endothelial cells (6–8), and fibroblasts (9,10) is anotable immunologic abnormality in SSc. In particular,antifibroblast antibodies (AFAs), which induce fibro-blast activation in vitro, are present in �46% of patientswith SSc and stimulate monocyte adhesion to fibroblastsby up-regulating intercellular adhesion molecule 1(ICAM-1), fibroblast secretion of interleukin-1 (IL-1)and monocyte chemotactic protein 1, and the steady-state increase of messenger RNA for cytokines andchemokines (11). Indeed, fibroblasts are abnormallyactivated in SSc, showing increased production of cyto-kines and growth factors involved in fibrogenesis andabnormal collagen turnover (12).
Supported by MURST Cofinanziamento 1998 and FIL2000(to Drs. Meroni and Orlandini) and by Ricerca Corrente 2000 IstitutoAuxologico Italiano (to Dr. Meroni).
1Nicoletta Ronda, MD, PhD, Rita Gatti, PhD, RobertoGiacosa, MD, Carlo Buzio, MD, Guido Orlandini, MD: University ofParma, Parma, Italy; 2Elena Raschi, PhD, Cinzia Testoni, PhD, PierLuigi Meroni, MD: University of Milan, IRCCS Istituto AuxologicoItaliano, Milan, Italy.
Address correspondence and reprint requests to NicolettaRonda, MD, PhD, Department of Clinical Medicine, Nephrology andHealth Sciences, University of Parma, Via Gramsci 14, 43100 Parma,Italy. E-mail: [email protected].
Submitted for publication October 9, 2001; accepted in re-vised form March 1, 2002.
1595

At present, no information is available on themechanisms of the interaction between AFAs and targetcells. We have developed a system that allows visualmonitoring of cultures of living adherent cells by confo-cal laser scanning microscopy (CLSM), and we haveapplied it to the study of the interaction between severalhuman cell types and normal immunoglobulins (13). Theaim of the present work was to characterize, usingCLSM, the interaction between AFA-positive IgG andliving fibroblasts, the possible processing of IgG, and thecellular structures involved.
PATIENTS AND METHODS
Patients. Sixty-nine SSc patients (58 women and 11men; mean � SD age 51.4 � 14.8 years, mean � SD diseaseduration 9.8 � 3.2 years) were studied. All patients fulfilled theAmerican College of Rheumatology (ACR; formerly, theAmerican Rheumatism Association) criteria for SSc (14).Control sera were from 50 healthy individuals, matched for sexand age.
Cell cultures. Human fibroblasts were obtained by thestandard explant technique from 1) skin from the cheek of a5-year-old donor undergoing plastic surgery, 2) the foreskin ofa healthy individual, and 3) involved skin from a patient withSSc. Cells were cultured in Dulbecco’s modified Eagle’s me-dium (DMEM) supplemented with 10% fetal calf serum andwere used between the fourth and the seventh passages.
IgG preparations. Normal human IgG was obtained inpooled form as used for intravenous therapy (Sandoglobulin;Sandoz, Basel, Switzerland) and as single-donor preparationsobtained by purifying IgG from the sera of 3 healthy subjectsby affinity chromatography on protein G–Sepharose columns(Pharmacia Biotech, Uppsala, Sweden), as previously de-scribed (15). IgG was also purified from 8 patients with SSc.Information on clinical characteristics and serum ANA andAFA activity for each of these 8 patients is presented in Table1. ANA was assessed by indirect immunofluorescence onHEp-2 cells (16), and anti–extractable nuclear antigen (anti-ENA) profiles were investigated as previously described (17).AFA positivity was defined as abnormal binding of serum IgG
to fibroblasts in a cell-based enzyme-linked immunosorbentassay (ELISA) (18). Aliquots from all IgG preparations werecoupled to fluorescein isothiocyanate (IgG-FITC) (19), exten-sively dialyzed in DMEM, and sterile filtered. The IgG con-centration in all samples was measured using high-sensitivityradial immunoprecipitation plates (NorPartigen; Behring,Scoppito, L’Aquila, Italy).
ELISA for AFA detection. Cell-based ELISA was per-formed on living, unfixed fibroblasts as described previously(18), with minor modifications. Briefly, sera were incubated ata 1:25 dilution for 2 hours on confluent fibroblast monolayersin 96-well plates. After washing, the plates were incubated for90 minutes with alkaline phosphatase–conjugated goat anti-human IgG. The reaction was developed by addingp-nitrophenylphosphate in 0.05M Mg-carbonate buffer (pH9.8), and the optical density (OD) at 405 nm was recorded(Titertek Multiskan MCC/340; Labsystems, Helsinki, Finland).Samples were tested in duplicate. The OD value of a positivereference serum at a standard 1:25 dilution was arbitrarilydefined as 100% of fibroblast binding activity. The results oftested samples were expressed as a percentage of this positivereference value, to adjust for interassay variability. An ODmore than 3 SD above the mean OD obtained with sera fromthe 50 healthy individuals (i.e., 22.2%) was considered positive.IgG purified from the sera of 5 AFA-positive and 3 AFA-negative SSc patients was also tested by ELISA, at 100, 50, 25,12.5, 6.2, 3.1, and 1.5 �g/ml.
Confocal laser scanning microscopy. The interactionbetween IgG-FITC and living fibroblasts was studied by CLSM(Multiprobe 2001; Molecular Dynamics, Sunnyvale, CA). Fi-broblasts grown on coverslips were kept at 37°C throughoutthe experiments by using a microincubator placed on themicroscope stage to provide stable, standard culture condi-tions; medium substitution was allowed whenever necessary,without changes of the preselected observation field (13).Normal culture medium was substituted by identical mediumcontaining 400 �g/ml purified IgG-FITC. After 20 minutes,cells were washed with 10 ml of medium in order to remove allunbound IgG. Images were then acquired every 5 minutes. Thesame procedure was repeated for each patient and control IgGsample. To exclude IL-1 interference in the interaction be-tween AFA-positive IgG and fibroblasts, the same experimentwas carried out in the presence of IL-1 receptor antagonist(IL-1Ra; Serotec, Oxford, UK) at 2 �g/ml. Incubation withIL-1Ra was started 30 minutes before addition of IgG andmaintained throughout the experiment.
Immunofluorescence of fixed fibroblasts. Detection ofintracellular IgG. Normal human fibroblasts were grown onslides, incubated with AFA-positive IgG at 400 �g/ml for 2hours, washed with phosphate buffered saline (PBS) at 37°C,and fixed with 100% methanol at 4°C for 2 minutes. Afterwashing with PBS, cells were incubated with a polyclonalanti-human IgG coupled to FITC (Dako, Milan, Italy) or aheavy chain–specific monoclonal anti-human IgG (cloneHP6017; Sigma, St. Louis, MO) followed by an anti-mouse IgGantibody coupled to FITC (Santa Cruz Biotechnology, SantaCruz, CA).
Visualization of caveolin- or clathrin-bearing structures.Normal human fibroblasts grown on slides, with or withoutincubation with AFA-positive IgG at 400 �g/ml for 2 hours,were washed with PBS at 37°C and fixed with 100% methanol
Table 1. Clinical presentation and antibody activity of the systemicsclerosis patients from whom IgG was obtained and purified*
PatientClinical
presentation ANA ENA AFA
1 Diffuse � � �2 Diffuse � � �3 Diffuse � Scl-70 �4 Diffuse � Scl-70 �5 Limited � � �6 Diffuse � Scl-70 �7 Diffuse � � �8 Limited ACA CENP A/B �
* ANA � antinuclear antibody; ENA � extractable nuclear antigen;AFA � antifibroblast antibody; ACA � anticentromere antibody.
1596 RONDA ET AL
at 4°C for 2 minutes. After washing with PBS, cells wereincubated with a rabbit anticaveolin or goat anticlathrin anti-body (both from Santa Cruz Biotechnology), followed byanti-rabbit or anti-goat fluoresceinated antibody (both fromSanta Cruz Biotechnology), respectively.
Double staining of fibroblasts with IgG and for caveo-lae. Normal human fibroblasts were grown on coverslips thatwere mounted on the incubation chamber of the confocalmicroscope. Cells were incubated under standard cultureconditions with 400 �g/ml AFA-positive IgG-FITC for 30minutes and, after washing with culture medium, were ob-served every 5 minutes. Once internalization was evident(usually after a further 30 minutes), images were acquired andcells were washed with PBS at 37°C. After 100% methanol wasflushed through the chamber at 4°C for 2 minutes, fibroblastswere washed with PBS at room temperature and incubatedwith a rabbit anticaveolin antibody, followed by an anti-rabbitIgG antibody coupled to Texas Red (Southern Biotechnology,Birmingham, AL). Samples were excited at appropriate wave-lengths (488 nm and 514 nm), and emission was split with abeam splitter (565 nm) and appropriate barrier filters (530 �30 nm dichroic and 600 long-pass). Thus, the signals fromTexas Red and FITC could be collected on 2 separate photo-multipliers, and images could be digitally overlapped in orderto assess colocalization.
Disassemby of fibroblast caveolae. In some experi-ments, fibroblasts were cultured in the presence of filipin (2�g/ml; Sigma), beginning 1 hour before initiation of otherexperimental conditions and continuing throughout the entireexperiment. Filipin is a sterol-binding molecule which disas-sembles caveolae and their microdomains without killing cells(20).
RESULTS
AFA activity in IgG preparations. To study themechanisms of interaction of AFAs with living fibro-blasts, IgG were purified from 5 SSc sera that were AFApositive and 3 that were AFA negative (as assessed byELISA), from the 69 sera tested. We tested AFA activityin these IgG preparations and confirmed the high-level,
dose-dependent binding in the 5 AFA-positive samples(Tables 1 and 2).
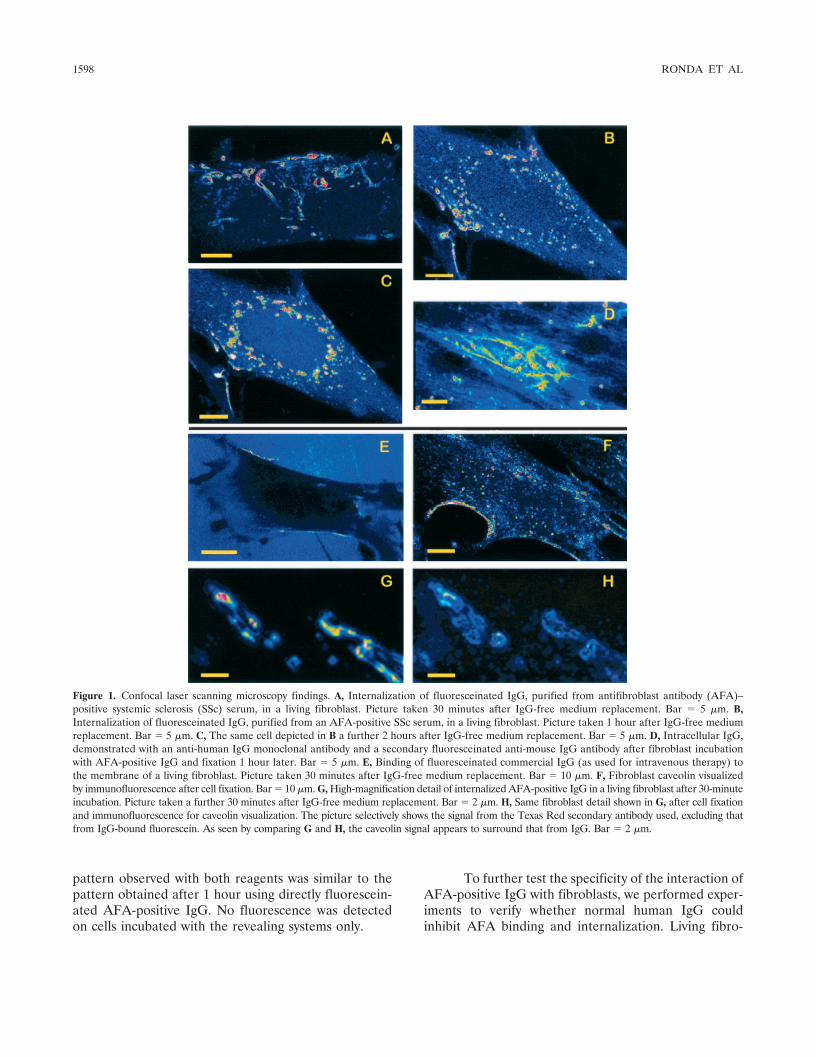
Interaction between AFA-positive IgG and fibro-blasts. We applied CLSM to investigate the interactionbetween AFAs and their target. When incubated withliving fibroblasts, fluoresceinated AFA-positive IgGwere observed to bind to fibroblast cell membranewithin 10 minutes. Interestingly, AFAs were then inter-nalized, with a process that became morphologicallyevident after �30 minutes and evolved during the next 4hours. The first event observed was the appearance offluorescent invaginations in certain regions of the cellmembrane, often presenting as tube-like structures (Fig-ure 1A). Within 1 hour, a network of fluorescent cyto-plasmic filaments became evident; this was followed bythe appearance of a granular pattern �1 hour later(Figure 1B). Finally, more coarse granules appeared,moving to the perinuclear area during the subsequent 2hours (Figure 1C). No additional morphologic modifi-cations were observed upon prolonged observation. Sim-ilar results were obtained in all 5 AFA-positive IgG fromSSc patients. No morphologic differences were observedwhen the interaction of AFA-positive IgG with fibro-blasts was tested using normal skin fibroblasts from 2donors versus fibroblasts from an SSc patient. TheAFA-positive IgG internalization process was not af-fected by pretreating fibroblasts with IL-1Ra.
In order to exclude the possibility that intracell-ular fluorescence was due to molecules other than IgGpossibly copurifying with IgG and thus contaminatingthe preparations, living fibroblasts were incubated withpurified nonfluoresceinated AFA-positive IgG for 1hour, and then were fixed. Intracellular IgG was re-vealed using both a mouse monoclonal anti-human IgGfollowed by an anti-mouse IgG fluoresceinated antibody(Figure 1D) and an anti-human IgG polyclonal fluores-ceinated antibody (results not shown). The fluorescence
Table 2. Binding, to fibroblasts, of IgG purified from selected SSc patients, as assessed by ELISA*
Serumdilution
Patient
1 2 3 4 5 6 7 8
100 �g/ml 100 � 24 100 � 19 92 � 14 1,870 � 184 1,480 � 187 1,690 � 204 1,620 � 221 1,570 � 25450 �g/ml 99 � 18 103 � 16 94 � 10 1,100 � 108 600 � 125 1,080 � 195 1,200 � 178 1,140 � 20825 �g/ml 102 � 25 101 � 20 90 � 10 815 � 74 427 � 84 651 � 89 860 � 74 891 � 6512.5 �g/ml 10 � 15 98 � 15 91 � 11 610 � 75 385 � 68 534 � 72 721 � 69 691 � 616.2 �g/ml 101 � 18 103 � 17 95 � 10 423 � 68 216 � 76 486 � 51 456 � 54 460 � 533.1 �g/ml 97 � 13 97 � 18 93 � 12 210 � 72 162 � 62 215 � 67 205 � 42 264 � 241.5 �g/ml 99 � 13 99 � 17 93 � 9 150 � 65 102 � 54 120 � 58 105 � 31 151 � 32
* Values are the mean � SD optical density from 2 separate experiments. SSc � systemic sclerosis; ELISA � enzyme-linkedimmunosorbent assay.
ANTIFIBROBLAST ANTIBODIES IN SSc 1597
pattern observed with both reagents was similar to thepattern obtained after 1 hour using directly fluorescein-ated AFA-positive IgG. No fluorescence was detectedon cells incubated with the revealing systems only.
To further test the specificity of the interaction ofAFA-positive IgG with fibroblasts, we performed exper-iments to verify whether normal human IgG couldinhibit AFA binding and internalization. Living fibro-
Figure 1. Confocal laser scanning microscopy findings. A, Internalization of fluoresceinated IgG, purified from antifibroblast antibody (AFA)–positive systemic sclerosis (SSc) serum, in a living fibroblast. Picture taken 30 minutes after IgG-free medium replacement. Bar � 5 �m. B,Internalization of fluoresceinated IgG, purified from an AFA-positive SSc serum, in a living fibroblast. Picture taken 1 hour after IgG-free mediumreplacement. Bar � 5 �m. C, The same cell depicted in B a further 2 hours after IgG-free medium replacement. Bar � 5 �m. D, Intracellular IgG,demonstrated with an anti-human IgG monoclonal antibody and a secondary fluoresceinated anti-mouse IgG antibody after fibroblast incubationwith AFA-positive IgG and fixation 1 hour later. Bar � 5 �m. E, Binding of fluoresceinated commercial IgG (as used for intravenous therapy) tothe membrane of a living fibroblast. Picture taken 30 minutes after IgG-free medium replacement. Bar � 10 �m. F, Fibroblast caveolin visualizedby immunofluorescence after cell fixation. Bar � 10 �m. G, High-magnification detail of internalized AFA-positive IgG in a living fibroblast after 30-minuteincubation. Picture taken a further 30 minutes after IgG-free medium replacement. Bar � 2 �m. H, Same fibroblast detail shown in G, after cell fixationand immunofluorescence for caveolin visualization. The picture selectively shows the signal from the Texas Red secondary antibody used, excluding thatfrom IgG-bound fluorescein. As seen by comparing G and H, the caveolin signal appears to surround that from IgG. Bar � 2 �m.
1598 RONDA ET AL

blasts were incubated overnight with a 20-fold excess ofnonfluoresceinated normal pooled IgG (8 mg/ml) beforebeing exposed to fluoresceinated AFA-positive IgG.Alternatively, fluoresceinated AFA-positive IgG wereadded simultaneously with a 20-fold excess of normalpooled IgG to fibroblast cultures and then examined byCLSM. No inhibition of AFA-positive IgG binding andinternalization was observed under either of these con-ditions.
Interaction between fibroblasts and AFA-negative IgG. To verify whether the dynamic interactionswith fibroblasts were specific for AFA-positive IgG, wealso applied CLSM to test binding to fibroblasts ofpurified IgG from normal serum, from pooled commer-cial IgG for intravenous therapeutic use, as well as fromSSc sera that were AFA negative by ELISA. Thesefluoresceinated IgG preparations, incubated with livinghuman fibroblasts at 400 �g/ml under standard cultureconditions, showed membrane binding within 30 min-utes (Figure 1E). However, after prolonged examination(4 hours), neither internalization nor morphologic mod-ification of the fluorescence pattern was observed. Sim-ilar results were obtained using skin fibroblasts from 2healthy donors and from 1 patient with SSc.
Taken together, these results indicate that AFAspresent in the sera of SSc patients, selected on the basisof abnormal binding on cell ELISA, have the capacity torecognize specific targets on the surface of normal andSSc fibroblasts. This specific recognition results in inter-nalization and perinuclear localization, which is appar-ently unrelated to the involvement of Fc� receptors.
Involvement of caveolae in AFA-positive IgGinternalization in fibroblasts. The surprising finding ofAFA-positive IgG internalization in fibroblasts promptedus to test whether internalization was dependent oncaveolin- or clathrin-associated structures. First, we per-formed direct immunofluorescence studies with antica-veolin and anticlathrin antibodies on fixed fibroblasts,with or without previous incubation with AFA-positiveIgG, to show localization of the 2 molecules in restingcells and possible differences in their distribution uponstimulation with AFAs. In both cases clathrin could notbe visualized, while caveolin staining revealed interlac-ing fibrillar structures, mostly close to certain zones ofthe cell membrane, sometimes with a tubular appear-ance (Figure 1F). No differences were observed whencomparing the morphology of caveolin-bearing struc-tures in fibroblasts exposed versus those not exposed toAFAs.
Since the fluorescence pattern of internalizedAFA-positive IgG was similar to that of caveolin-bearing
structures, we performed double staining to verify thepossible colocalization. We incubated living fibroblastswith AFA-positive IgG-FITC for 30 minutes, waited forthe internalization to progress (usually within another 30minutes), and then fixed the cells and performed directimmunofluorescence with a monoclonal anticaveolinantibody and the relevant Texas Red–coupled secondaryantibody. Indeed, a precise correspondence in localiza-tion of IgG and caveolin was found. In particular, inimages obtained at high magnification, IgG signal ap-peared surrounded by that from caveolin (Figures 1Gand H).
In addition, we observed complete inhibition ofAFA-positive IgG membrane binding and internaliza-tion in fibroblasts that were treated with 2 �g/ml filipinstarting 1 hour before the rest of the experiment andcontinuing throughout (results not shown).
These findings strongly indicate that AFA-positive IgG, but not normal IgG or SSc AFA-negativeIgG, specifically bind surface antigens associated withcellular structures that elicit a caveolin-dependent inter-nalization.
DISCUSSION
Whatever the initial event leading to immunesystem stimulation and microvascular inflammation inSSc, fibroblast activation, with abnormal function interms of cytokine/growth factor secretion and collagenturnover, is pivotal in the development of fibrosis, whichis the hallmark of the disease. The possible role ofmembrane-reactive AFAs in stimulating fibroblasts inSSc has been demonstrated (11), and information on themechanism of interaction of these autoantibodies withliving cells may be useful for understanding the fibro-blast activation process and developing new treatmentstrategies.
We have shown previously (21), and confirmed inthis study with confocal laser microscopy on living cells,that normal circulating IgG binds to fibroblast mem-brane but does not enter the cytoplasmic compartment.This might be due to the binding of normal IgG to Fc�receptors expressed on fibroblast membrane, but thepresence of Fc� receptors on resting human fibroblastshas never been reported (22). An alternative explanationcould be the presence, in normal subjects, of a popula-tion of natural autoantibodies reacting with fibroblastantigens that can be revealed morphologically with oursensitive system, allowing real-time confocal image ac-quisition of living cells. Natural autoantibodies consti-tute �60% of circulating immunoglobulins and have
ANTIFIBROBLAST ANTIBODIES IN SSc 1599

been demonstrated to react with several human auto-antigens, both soluble (23) and cellular (15). The mainfunctions of natural autoantibodies appear to be a rolein resistance to infections (24), the regulation of ho-meostasis of immune networks (24), and the control ofphysiologic cell functions (25). Further studies would benecessary to verify whether normal IgG binding tofibroblast membrane, as we have described, has anyfunctional consequence. However, normal IgG does notseem to interfere with fibroblast ICAM-1 expression andcytokine or growth factor secretion (11).
In most cases, quantitative measurement of thebinding of natural autoantibody to autoantigens (e.g., byELISA) shows lower values than those obtained withcorresponding pathologic autoantibodies, and usuallyepitope specificity is also different (15). These differ-ences allow the use of certain pathologic autoantibodiesas markers of autoimmune diseases, even in the pres-ence of a natural autoantibody physiologic counterpart.This is why IgG purified from patients with SSc could beclassified as AFA negative or AFA positive according tonormal or abnormal binding to fibroblasts in a cell-basedELISA. All AFA-negative IgG preparations from SScpatients that we tested reacted as IgG from healthydonors, showing membrane binding to living fibroblastswith no internalization. It is worth noting that thisfinding correlates with the lack of fibroblast activationshown by these IgG (11) and not with ANA or anti-ENAantibodies (which were positive in 2 of the AFA-negativeSSc sera), confirming the lack of correlation betweenAFAs and any of the known ANAs in SSc.
In contrast, all SSc patient IgG preparations thatwere defined as AFA positive based on an abnormalreaction in a cell-based ELISA not only bound to cellmembrane, but were also internalized starting fromdiscrete zones of the cell membrane. We excluded thepossibility that intracellular fluorescence was due tointernalization of material copurifying with IgG. In fact,using both a polyclonal and a monoclonal anti-humanIgG antibody, we demonstrated intracellular IgG byimmunofluorescence on fixed cells that had been previ-ously exposed to nonfluoresceinated AFA-positive IgG.
Fibroblast activation by AFA-positive IgG in-cludes an autocrine loop mediated by IL-1 (11). We thustested the hypothesis that internalization of AFA-positive IgG could be affected by pretreating fibroblastswith IL-1Ra. This was not the case, indicating that IL-1is not involved in this process.
No correlation was found between IgG internal-ization and any of the known ANAs typical of SSc.Internalization of AFA-positive IgG in living fibroblasts
correlates with the ability of the same IgG fractions toactivate fibroblasts (11) and was not related to thepresence of any ANA.
The lack of internalization of AFA-negative IgGexcludes per se the possibility of Fc portion involvementin AFA-positive IgG activity, and this is further excludedby the lack of competitive inhibition of AFA internal-ization by large amounts of normal IgG (by both prein-cubation and coincubation). This latter finding alsoindicates that normal IgG do not interact idiotypicallywith AFAs, as has been reported for other pathologicautoantibodies (26). We found no differences in ourobservations when using normal skin fibroblasts, fibro-blasts from the foreskin of a healthy individual, orfibroblasts from a patient with SSc; thus, we concludethat the molecules recognized are constitutively ex-pressed in all cell types tested.
The time course and morphology of AFA inter-action with fibroblasts are consistent with a receptor–ligand internalization process, in which the receptor–ligand complex induces folding of the cell membraneand formation of endosomes (27). Both caveolae andclathrin-coated pits are specialized cell membrane areasin continuous remodeling involved in receptor–ligandinternalization and signal transduction (20,27), andcould theoretically correspond to the discrete membranezones from which AFA-positive IgG begin to be inter-nalized. Thus, we first analyzed caveolin and clathrindistribution in fibroblasts in our experimental conditionsand visualized a network of filaments and tubules juttingout from the cell membrane into the cytoplasm andcontaining caveolin, while clathrin could not be visual-ized. Caveolin distribution was the same in restingfibroblasts and following incubation with AFA-positiveIgG. The pattern of AFA-positive IgG distribution infibroblasts was very similar to that of caveolin, anddouble staining of fibroblasts to visualize both internal-ized IgG and caveolin demonstrated a colocalization ofthe 2 molecules. In particular, some detailed viewsshowed IgG surrounded by caveolin in a tubule-likeappearance.
These findings strongly suggest that AFA-positive IgG internalization in fibroblasts occurs throughcaveolae-associated receptors, and this idea is confirmedby the complete inhibition of internalization obtainedwhen fibroblasts were treated with filipin. This sterol-binding molecule, widely used to disassemble caveolae,removes plasma membrane cholesterol, inducing flatten-ing of caveolin-rich invaginations, unclustering of recep-tors, and loss of caveolae-mediated endocytosis (20).These effects of filipin may also explain why not only
1600 RONDA ET AL

internalization, but also membrane binding, of AFA-positive IgG was inhibited in filipin-treated fibroblasts.
We thus conclude that abnormal AFAs that aredetectable in the sera of a subpopulation of SSc patientsand are able to activate fibroblasts toward a proinflam-matory state are selectively internalized by fibroblaststhrough an Fc� receptor–independent, caveolae-linkedmechanism. Identification of the putative receptor in-volved could contribute to understanding of the precisemechanisms of fibroblast activation in SSc and develop-ment of new treatment strategies.
REFERENCES
1. Haustein UF, Anderegg U. Pathophysiology of scleroderma: anupdate. J Eur Acad Dermatol Venereol 1998;11:1–8.
2. Sato S. Abnormalities of adhesion molecules and chemokines inscleroderma. Curr Opin Rheumatol 1999;11:503–7.
3. Kahaleh MB, LeRoy EC. Autoimmunity and vascular involvementin systemic sclerosis (SSc). Autoimmunity 1999;31:195–214.
4. Harvey GR, McHugh NJ. Serologic abnormalities in systemicsclerosis. Curr Opin Rheumatol 1999;11:495–502.
5. Nelson JL. Microchimerism and the pathogenesis of systemicsclerosis. Curr Opin Rheumatol 1998;10:564–71.
6. Pignone A, Scaletti C, Matucci-Cerinic M, Vasquez-Abad D,Meroni PL, Del Papa N, et al. Anti-endothelial cell antibodies insystemic sclerosis: significant association with vascular involve-ment and alveolo-capillary impairment. Clin Exp Rheumatol1998;16:527–32.
7. Carvalho D, Savage CO, Black CM, Pearson JD. IgG antiendo-thelial cell autoantibodies from scleroderma patients induce leu-kocyte adhesion to human vascular endothelial cells in vitro:induction of adhesion molecule expression and involvement ofendothelium-derived cytokines. J Clin Invest 1996;97:111–9.
8. Bordon A, Dueyme S, Levy Y, Jamin C, Leroy JP, Piette JC, et al.The binding of some human antiendothelial cell antibodies in-duces endothelial cell apoptosis. J Clin Invest 1998;101:2029–35.
9. Brentnall TJ, Kenneally D, Barnett AJ, de Aizpurua HJ, Lolait SJ,Ashcroft R, et al. Autoantibodies to fibroblasts in scleroderma.J Clin Lab Immunol 1982;8:9–12.
10. Hill MB, Phipps JL, Cartwright RJ, Milford Ward A, Greaves M,Hughes P. Antibodies to membranes of endothelial cells andfibroblasts in scleroderma. Clin Exp Immunol 1996;106:491–7.
11. Del Papa N, Chizzolini C, Raschi E, Testoni C, Facchini A,Gabrielli A, et al. IgG from scleroderma patients react with humanfibroblast membrane antigens and induce a pro-adhesive and apro-inflammatory fibroblast phenotype in vitro [abstract]. ArthritisRheum 1999;42 Suppl 9:S169.
12. Strehlow D, Korn JH. Biology of the scleroderma fibroblast. CurrOpin Rheumatol 1998;10:572–8.
13. Orlandini G, Ronda N, Gatti R, Gazzola GC, Borghetti A.Receptor-ligand internalization. Methods Enzymol 1998;307:340–8.
14. Subcommittee for Scleroderma Criteria of the American Rheu-matism Association Diagnostic and Therapeutic Criteria Commit-tee. Preliminary criteria for the classification of systemic sclerosis(scleroderma). Arthritis Rheum 1980;23:581–90.
15. Ronda N, Haury M, Nobrega A, Kaveri S, Coutinho A, Kazatch-kine MD. Analysis of natural and disease-associated autoantibodyrepertoires: anti-endothelial cell IgG autoantibody reactivity in theserum of healthy individuals and patients with systemic lupuserythematosus. Int Immunol 1994;6:1651–62.
16. Bayer PM, Bauerfeind S, Bienvenue J, Fabien N, Frei PC, GilburdB, et al. Multicenter evaluation study on a new Hep2 ANAscreening enzyme immune assay. J Autoimmun 1999;13:89–93.
17. Brucato A, Frassi M, Franceschini F, Cimaz R, Faden D, PisoniMP, et al. Risk of congenital complete heart block in newborns ofmothers with anti-Ro/SSA antibodies detected by counterimmuno-electrophoresis: a prospective study of 100 women. ArthritisRheum 2001;44:1832–5.
18. Del Papa N, Guidali L, Sironi M, Shoenfeld Y, Mantovani A,Tincani A, et al. Anti–endothelial cell IgG antibodies frompatients with Wegener’s granulomatosis bind to human endothe-lial cells in vitro and induce adhesion molecule expression andcytokine secretion. Arthritis Rheum 1996;39:758–66.
19. Johnstone A, Thorpe R. Immunochemistry in practise. Oxford:Blackwell Scientific Publications; 1982.
20. Schnitzer JE, Oh P, Pinney E, Allard J. Filipin-sensitive caveolae-mediated transport in endothelium: reduced transcytosis, scaven-ger endocytosis, and capillary permeability of selected macro-molecules. J Cell Biol 1994;127:1217–32.
21. Ronda N, Gatti R, Orlandini G, Borghetti A. Binding andinternalization of human IgG by living cultured endothelial cells.Clin Exp Immunol 1997;109:211–20.
22. Gessner JE, Heiken H, Tamm A, Schmidt RE. The IgG Fcreceptor family. Ann Hematol 1998;76:231–48.
23. Hurez V, Kaveri SV, Kazatchkine MD. Expression and control ofthe natural autoreactive repertoire in normal human serum. EurJ Immunol 1993;23:783–9.
24. Coutinho A, Kazatchkine MD, Avrameas S. Natural autoantibod-ies. Curr Opin Immunol 1995;7:812–8.
25. Ronda N, Leonardi S, Orlandini G, Gatti R, Bellosta S, Berini F,et al. Natural anti-endothelial cell antibodies (AECA). J Autoim-mun 1999;13:121–9.
26. Ronda N, Nobrega A, Haury M, Coutinho A, Kazatchkine MD.Selectivity of recognition of variable (V) regions of autoantibodiesby intravenous immunoglobulin (IVIg). Clin Immunol Immuno-pathol 1994;70:124–32.
27. Lodish H, Baltimore D, Berk A, Zipurki SL, Matsudaira P,Darnell J. Molecular cell biology. 3rd ed. New York: ScientificAmerican Books; 1995.
ANTIFIBROBLAST ANTIBODIES IN SSc 1601




















