
Caveolin-1 Reduces Osteosarcoma Metastases by Inhibiting c-Src Activity and Met Signaling
12
Caveolin-1 Reduces Osteosarcoma Metastases by Inhibiting c-Src Activity and Met Signaling Lara Cantiani, 1 Maria Cristina Manara, 1 Cinzia Zucchini, 2 Paola De Sanctis, 2 Monia Zuntini, 1 Luisa Valvassori, 2 Massimo Serra, 1 Martina Olivero, 3 Maria Flavia Di Renzo, 3 Mario Paolo Colombo, 4 Piero Picci, 1 and Katia Scotlandi 1 1 Laboratory of Oncology Research, Rizzoli Orthopaedic Institute; 2 Center for Molecular Genetic Research ‘‘Fondazione CARISBO’’ at the Istituto di Istologia ed Embriologia Generale, University of Bologna, Bologna, Italy; 3 Laboratory of Cancer Genetics, Institute for Cancer Research and Treatment, University of Turin, Candiolo (Turin), Italy; and 4 Immunotherapy and Gene Therapy Unit, Istituto Nazionale per lo Studio e la Cura dei Tumori, Milan, Italy Abstract Caveolin-1 (Cav-1) is highly expressed in normal osteoblasts. This article reports that Cav-1 down-regulation is part of osteoblast transformation and osteosarcoma progression and validates its role as oncosuppressor in human osteosarcoma. A survey of 6-year follow-up indicates a better overall survival for osteosarcoma expressing a level of Cav-1 similar to osteoblasts. However, the majority of primary osteosarcoma shows significantly lower levels of Cav-1 than normal osteoblasts. Accordingly, Met-induced osteoblast transforma- tion is associated with Cav-1 down-regulation. In vitro , osteo- sarcoma cell lines forced to overexpress Cav-1 show reduced malignancy with inhibited anchorage-independent growth, migration, and invasion. In vivo , Cav-1 overexpression abrogates the metastatic ability of osteosarcoma cells. c-Src and c-Met tyrosine kinases, which are activated in osteosar- coma, colocalize with Cav-1 and are inhibited on Cav-1 over- expression. Thus, Cav-1 behaves as an oncosuppressor in osteosarcoma. Altogether, data suggest that Cav-1 down- modulation might function as a permissive mechanism, which, by unleashing c-Src and Met signaling, enables osteosarcoma cells to invade neighboring tissues. These data strengthen the rationale to target c-Src family kinases and/or Met receptor to improve the extremely poor prognosis of metastatic osteosarcoma. [Cancer Res 2007;67(16):7675–85] Introduction The frequency of primary malignant bone tumors ranks osteosarcoma second after multiple myeloma. Osteosarcoma is a high-grade malignant tumor composed of mesenchymal cells producing osteoid and immature bone with a peak incidence in the second decade of life (1). Although current treatment modalities, which include surgery and neoadjuvant multidrug chemotherapy, significantly improved the 5-year disease-free survival from 10% to 60%–70% (2, 3), no significant new drugs or treatment approaches have been developed during the last 10 years, and the percentages of cure remain unacceptably low for high-risk patients (4). Drug resistance and metastasis are the major causes of treatment failure. Differently from carcinomas, molecular genetics of osteosarcoma progression is not documented and the most accepted prognostic factors are based on response to treatment (percentage of necrosis or P-glycoprotein expression; refs. 5–8) rather than on genetic signature characterizing the development and progression, with few exceptions (9–11). To search for such signature, we did a microarray analysis of genes involved in modulation of osteosarcoma malignancy by comparing osteosarcoma cell lines with different metastasis behavior and identified caveolin-1 ( Cav-1 ) gene whose expression correlates with a lower metastasis potential (12, 13). Cav-1 is the major protein component of caveolae (14), which are abundant in terminally differentiated mesenchymal cells, including adipocytes, endothelial cells, fibroblasts (15), and osteoblasts (16, 17). Caveolae provide physical interaction and compartmentalization of several mem- brane receptors. As part of receptor signaling pathways, Cav-1 plays functions that are important in tumorigenesis and metastasis (see refs. 18–20 for reviews). However, despite significant studies have been done over the last 10 years, its functions in malignancy are very complex and still unclear, as Cav-1 has been described as either a tumor suppressor or a tumor promoter, depending on tumor type and/or stage. Although overexpression of Cav-1 is associated with, and in fact causal in metastasis and progression in some types of carcinomas, particularly prostate cancer (21, 22), in breast cancer as well as fibrosarcoma, Cav-1 has been character- ized as oncosuppressor (23–25). In this article, we validate the role of Cav-1 as oncosuppressor in human osteosarcoma, as we show its down-modulation in more aggressive tumors and show Cav-1 ability to suppress the invasive-metastatic ability of osteosarcoma cells in functional assays in vitro and in vivo . Materials and Methods Cell lines and transfection. The osteosarcoma cell lines U-2 OS, Saos-2, and MG63 were obtained from the American Type Culture Collection. IOR/OS-7 and IOR/OS-9 were obtained at the Laboratorio di Ricerca Oncologica, Istituti Ortopedici Rizzoli (Bologna, Italy) and previously characterized (26). Human osteoblasts (HOS) and their Met-transformed variants were obtained at the Laboratory of Cancer Genetics, Institute for Cancer Research and Treatment, University of Turin [Candiolo (Turin), Italy] and previously characterized (27). pcDNA3 vector was used to construct plasmids expressing mouse Cav-1 (28), which differs from human Cav-1 at only two amino acids and is functionally indistinguishable from the human protein (15), and human antisense Cav-1 (21). Stable transfectants expressing Cav-1 were obtained from U-2 OS, whereas Cav-1–deficient clones were derived from U-2 OS and Saos-2 by using calcium-phosphate transfection method. Cells transfected with the empty vector pcDNA3 were used as negative controls. Transfectants were selected in Iscove’s modified Note: Supplementary data for this article are available at Cancer Research Online (http://cancerres.aacrjournals.org/). L. Cantiani and M.C. Manara contributed equally to this work. Requests for reprints: Katia Scotlandi, Laboratory of Oncology Research, Rizzoli Orthopaedic Institute, Via Di Barbiano 1/10, 40136 Bologna, Italy. Phone: 39-051-63-66- 760; Fax: 39-051-6366-761; E-mail: [email protected]. I2007 American Association for Cancer Research. doi:10.1158/0008-5472.CAN-06-4697 www.aacrjournals.org 7675 Cancer Res 2007; 67: (16). August 15, 2007 Research Article Research. on March 24, 2016. © 2007 American Association for Cancer cancerres.aacrjournals.org Downloaded from
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Caveolin-1 Reduces Osteosarcoma Metastases by Inhibiting c-Src Activity and Met Signaling

Caveolin-1 Reduces Osteosarcoma Metastases by
Inhibiting c-Src Activity and Met Signaling
Lara Cantiani,1Maria Cristina Manara,
1Cinzia Zucchini,
2Paola De Sanctis,
2Monia Zuntini,
1
Luisa Valvassori,2Massimo Serra,
1Martina Olivero,
3Maria Flavia Di Renzo,
3
Mario Paolo Colombo,4Piero Picci,
1and Katia Scotlandi
1
1Laboratory of Oncology Research, Rizzoli Orthopaedic Institute; 2Center for Molecular Genetic Research ‘‘Fondazione CARISBO’’at the Istituto di Istologia ed Embriologia Generale, University of Bologna, Bologna, Italy; 3Laboratory of Cancer Genetics,Institute for Cancer Research and Treatment, University of Turin, Candiolo (Turin), Italy; and 4Immunotherapyand Gene Therapy Unit, Istituto Nazionale per lo Studio e la Cura dei Tumori, Milan, Italy
Abstract
Caveolin-1 (Cav-1) is highly expressed in normal osteoblasts.This article reports that Cav-1 down-regulation is part ofosteoblast transformation and osteosarcoma progression andvalidates its role as oncosuppressor in human osteosarcoma.A survey of 6-year follow-up indicates a better overall survivalfor osteosarcoma expressing a level of Cav-1 similar toosteoblasts. However, the majority of primary osteosarcomashows significantly lower levels of Cav-1 than normalosteoblasts. Accordingly, Met-induced osteoblast transforma-tion is associated with Cav-1 down-regulation. In vitro , osteo-sarcoma cell lines forced to overexpress Cav-1 show reducedmalignancy with inhibited anchorage-independent growth,migration, and invasion. In vivo , Cav-1 overexpressionabrogates the metastatic ability of osteosarcoma cells. c-Srcand c-Met tyrosine kinases, which are activated in osteosar-coma, colocalize with Cav-1 and are inhibited on Cav-1 over-expression. Thus, Cav-1 behaves as an oncosuppressor inosteosarcoma. Altogether, data suggest that Cav-1 down-modulation might function as a permissive mechanism,which, by unleashing c-Src and Met signaling, enablesosteosarcoma cells to invade neighboring tissues. These datastrengthen the rationale to target c-Src family kinases and/orMet receptor to improve the extremely poor prognosis ofmetastatic osteosarcoma. [Cancer Res 2007;67(16):7675–85]
Introduction
The frequency of primary malignant bone tumors ranksosteosarcoma second after multiple myeloma. Osteosarcoma is ahigh-grade malignant tumor composed of mesenchymal cellsproducing osteoid and immature bone with a peak incidence inthe second decade of life (1). Although current treatmentmodalities, which include surgery and neoadjuvant multidrugchemotherapy, significantly improved the 5-year disease-freesurvival from 10% to 60%–70% (2, 3), no significant new drugs ortreatment approaches have been developed during the last10 years, and the percentages of cure remain unacceptably low
for high-risk patients (4). Drug resistance and metastasis are themajor causes of treatment failure. Differently from carcinomas,molecular genetics of osteosarcoma progression is not documentedand the most accepted prognostic factors are based on response totreatment (percentage of necrosis or P-glycoprotein expression;refs. 5–8) rather than on genetic signature characterizing thedevelopment and progression, with few exceptions (9–11). Tosearch for such signature, we did a microarray analysis of genesinvolved in modulation of osteosarcoma malignancy by comparingosteosarcoma cell lines with different metastasis behavior andidentified caveolin-1 (Cav-1) gene whose expression correlates witha lower metastasis potential (12, 13). Cav-1 is the major proteincomponent of caveolae (14), which are abundant in terminallydifferentiated mesenchymal cells, including adipocytes, endothelialcells, fibroblasts (15), and osteoblasts (16, 17). Caveolae providephysical interaction and compartmentalization of several mem-brane receptors. As part of receptor signaling pathways, Cav-1plays functions that are important in tumorigenesis and metastasis(see refs. 18–20 for reviews). However, despite significant studieshave been done over the last 10 years, its functions in malignancyare very complex and still unclear, as Cav-1 has been described aseither a tumor suppressor or a tumor promoter, depending ontumor type and/or stage. Although overexpression of Cav-1 isassociated with, and in fact causal in metastasis and progression insome types of carcinomas, particularly prostate cancer (21, 22), inbreast cancer as well as fibrosarcoma, Cav-1 has been character-ized as oncosuppressor (23–25). In this article, we validate the roleof Cav-1 as oncosuppressor in human osteosarcoma, as we show itsdown-modulation in more aggressive tumors and show Cav-1ability to suppress the invasive-metastatic ability of osteosarcomacells in functional assays in vitro and in vivo .
Materials and Methods
Cell lines and transfection. The osteosarcoma cell lines U-2 OS, Saos-2,and MG63 were obtained from the American Type Culture Collection.
IOR/OS-7 and IOR/OS-9 were obtained at the Laboratorio di Ricerca
Oncologica, Istituti Ortopedici Rizzoli (Bologna, Italy) and previously
characterized (26). Human osteoblasts (HOS) and their Met-transformedvariants were obtained at the Laboratory of Cancer Genetics, Institute for
Cancer Research and Treatment, University of Turin [Candiolo (Turin),
Italy] and previously characterized (27). pcDNA3 vector was used to
construct plasmids expressing mouse Cav-1 (28), which differs from humanCav-1 at only two amino acids and is functionally indistinguishable from the
human protein (15), and human antisense Cav-1 (21). Stable transfectants
expressing Cav-1 were obtained from U-2 OS, whereas Cav-1–deficientclones were derived from U-2 OS and Saos-2 by using calcium-phosphate
transfection method. Cells transfected with the empty vector pcDNA3 were
used as negative controls. Transfectants were selected in Iscove’s modified
Note: Supplementary data for this article are available at Cancer Research Online(http://cancerres.aacrjournals.org/).
L. Cantiani and M.C. Manara contributed equally to this work.Requests for reprints: Katia Scotlandi, Laboratory of Oncology Research, Rizzoli
Orthopaedic Institute, Via Di Barbiano 1/10, 40136 Bologna, Italy. Phone: 39-051-63-66-760; Fax: 39-051-6366-761; E-mail: [email protected].
I2007 American Association for Cancer Research.doi:10.1158/0008-5472.CAN-06-4697
www.aacrjournals.org 7675 Cancer Res 2007; 67: (16). August 15, 2007
Research Article
Research. on March 24, 2016. © 2007 American Association for Cancercancerres.aacrjournals.org Downloaded from

Dulbecco’s medium (IMDM) containing 10% fetal bovine serum (FBS) and500 Ag/mL neomycin (Sigma) and maintained in selective medium.
Cytofluorimetric analysis of Met. Met expression was analyzed by
indirect immunofluorescence using DO-24 antibody against the extracellu-
lar domain of human Met protein (1:50 dilution; ref. 29).Analysis of growth features in monolayer conditions. Doubling time
was determined by daily harvesting of cells after seeding of 20,000 cells/cm2
in IMDM 10% FBS. Cell viability was determined by trypan blue dye
exclusion. Cytotoxicity of DXR on U-2 OS cells and derived clones evaluatedby 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
assays (Invitrogen) was done according to the manufacturer’s instructions.
Soft agar assay. Anchorage-independent growth was determined in
0.33% agarose (SeaPlaque, FMC BioProducts) with a 0.5% agarose underlay.
Cell suspensions (10,000–33,000 cells per 60-mm dish) were plated in
semisolid medium (IMDM 10% FBS plus agar 0.33%) and incubated at 37jCin a humidified 5% CO2 atmosphere. Colonies were counted after 10 days.
Poly-HEMA assay. Six-well plates were treated with poly(2-hydroxye-
thylmethacrylate) (poly-HEMA; Sigma) following the Folkman and Mosconamethod (30). Cells (250,000 per well) were seeded in IMDM 10% FBS and
incubated at 37jC in a humidified 5% CO2 atmosphere. Viable and dead
cells were counted after 24, 48, and 72 h. Detection and quantification of
apoptotic cells was obtained by flow cytometric analysis (FACSCalibur,Becton Dickinson) of Annexin V-FITC–labeled cells according to the
manufacturer’s instructions (Medical & Biological Laboratories).
Motility assay. Motility assay was done using Transwell chambers
(Costar). Cells (105) in IMDM plus 10% FBS with or without different c-Srcinhibitors (PP2, 5 Amol/L; PP3, 5 Amol/L; all provided by Calbiochem) or Src
activator pYEEI (100 Amol/L; Biomol Research Laboratories) were seeded in
the upper compartment, whereas IMDM plus 10% FBS was placed in thelower compartment of the chamber. Experiments were done in triplicate.
Wound-healing assay.Monolayer wounds were made using a pipette tip
on confluent cells. Cell migration was visualized, at regular intervals of time,
at �100 magnification using an inverted microscope (Nikon Diaphot) andphotographed with a Nikon D70s digital camera.
Invasion assay. Cells (5 � 105) were seeded in the upper compartment
of Biocoat Matrigel invasion chambers (BD Biosciences), and assay was
done according to the manufacturer’s instructions. All experiments weremade in triplicate.
Extracellular matrix adhesion assay. Adhesive ability of U-2 OS
transfectants was analyzed by using CytoMatrix cell adhesion strips coatedwith human collagen type I and IV, fibronectin, or laminin (Chemicon
International, Inc.) as described previously (13).
Fluorescence on adherent fixed cells. Immunofluorescence was done
on adherent cells grown on coverslips for 48 h and fixed in 4% para-formaldehyde, permeabilized with 0.15% Triton X-100 in PBS, and incubated
with the following antibodies: a-Cav-1 (1:100 dilution; BD Transduction
Laboratories), a-c-Src (1:10 dilution; Cell Signaling Technology), and
a-h-Met (C-28; 1:10 dilution; Santa Cruz Biotechnology).Western blotting.Western blotting experiments were done as described
previously (13). Membranes were incubated with the following primary
antibodies: anti-Cav-1 polyclonal antibody (1:10,000 dilution; BD Transduc-
tion Laboratories) and anti–phosphorylated Src Tyr416 (1:1,000 dilution; Cell
Signaling Technology). Analysis of c-Src and glyceraldehyde-3-phosphate
dehydrogenase (GAPDH) was also done to verify the total protein as control
(primary antibodies: a-c-Src, diluted 1:1,000; a-GAPDH, diluted 1:2,000,
Santa Cruz Biotechnology). Anti-rabbit secondary antibody conjugated to
horseradish peroxidase was from Amersham.
Immunoprecipitation and c-Src tyrosine kinase activity assay.Equivalent amounts of total cell lysates (800 Ag) were immunoprecipitated
by adding either 1.5 Ag sheep anti-human polyclonal anti-p60 Src antibody
(Chemicon International) and incubated overnight at 4jC. Protein G Plus/
protein A-agarose beads (Calbiochem) were then added to each lysate and
incubation continued for 1 h. Beads were then collected and washed twice
with lysis buffer and once in Src kinase reaction buffer (Upstate). Washed
beads were used directly to evaluate c-Src tyrosine kinase activity using a
nonradioactive ELISA-based assay according to the manufacturer’s
instructions (Chemicon International).
Determination of c-Src and Akt phosphorylation by cell-basedELISA. Cells were seeded into 96-well microplates at the density of
5 � 103 per well in IMDM 10% FBS. Forty-eight hours after seeding, cells
were fixed in 4% formaldehyde at room temperature and hereafter used to
measure Akt and Src phosphorylation using a cell-based CASE ELISA kit(SuperArray) according to the manufacturer’s instructions. To analyze the
status of intracellular mediators after hepatocyte growth factor (HGF)
stimulation, cells were starved for 24 h and then exposed to 40 ng/mL
HGF for 30 min. Absorbance readings were normalized to relative cellnumber as determined by a cell staining solution and to the amount of
total protein.
Tumorigenic and metastatic ability in athymic mice. Female athymic
5-week-old Crl:nu/nu (CD-1) BR mice (Charles River Italia) were used.Tumorigenicity was determined after s.c. injection of 30 � 106 cells, whereas
metastatization was evaluated after i.v. injection of 2 � 106 cells. Tumor
growth was assessed once weekly by measuring tumor volume. Negativemice were checked for 6 months after cell injection. The number of
pulmonary metastases was determined 2 months after cell inoculation.
Experimental procedures were approved by the local ethical committee.
Patients and quantitative PCR analysis of Cav-1. The study includes37 RNA samples extracted from osteosarcoma tumor specimens from
primary lesions of previously untreated patients. Osteosarcoma patients
were seen at the Istituti Ortopedici Rizzoli and treated with consecutive
chemotherapy protocols (IOR/3a and IOR/3b) previously reported in detail(31, 32). All patients provided written informed consent according to local
investigational review board requirements. Investigation was conducted
according to Declaration of Helsinki principles. Adverse events were definedas tumor recurrence at any site or death during remission. Event-free
survival was calculated from the first day of chemotherapy until the date of
first relapse or tumor-related death ( for patients classified as relapsed) or to
the date of the last follow-up examination. Overall survival was calculatedfrom the first day of chemotherapy to death or to the last follow-up
examination.
Total RNA was extracted by Trizol extraction kit (Invitrogen), and the
quality of the RNA samples was determined by electrophoresis and byBioanalyzer (Agilent). Universal Master Mix (Applied Biosystems) was used
with 10 ng cDNA and with 200 nmol/L of primers for evaluation of GAPDH
and Cav-1 expression, respectively. Negative controls without cDNAtemplate were run with each assay. For GAPDH and Cav-1, specific Taqman
assays were purchased from Applied Biosystems (Taqman Gene Expression
Assay, Ref. Seq. Hs99999901_s1 and Hs00184697_m1).
Statistical analysis. Differences among means were analyzed usingStudent’s t test. Fisher’s exact test was used for frequency data. Survival
curves were drawn and evaluated by Kaplan-Meier and log-rank methods.
Results
Met-transformed osteoblasts and osteosarcoma samplesshow lower Cav-1 expression than normal osteoblasts.Osteosarcomas originate from mesenchymal cells having osteo-blastic features. Osteoblasts have caveolae and express both Cav-1and Cav-2 isoforms (16). It has been noted that faster-growingosteoblasts have less caveolae (16) and that Cav-1 expressionincreases along osteoblastic differentiation (12). So we wonderedif osteoblast transformation was associated to Cav-1 change ofexpression. We took advantage of the recently established model ofHOS transformation by the overexpressed MET oncogene (27). It isnoteworthy that the MET oncogene is aberrantly overexpressed inf100% of human osteosarcomas and overexpression is associatedto an aggressive phenotype (33, 34). We found that Cav-1 issignificantly down-regulated in HOSs transformed by stablyoverexpressing either wild-type MET or the MET oncogeneactivated by Y1253D mutation (Fig. 1A and B ; ref. 35). Interestingly,down-modulation was not reverted by the subsequent expressionof a dominant-negative Met kinase, which only partially impairs the
Cancer Research
Cancer Res 2007; 67: (16). August 15, 2007 7676 www.aacrjournals.org
Research. on March 24, 2016. © 2007 American Association for Cancercancerres.aacrjournals.org Downloaded from
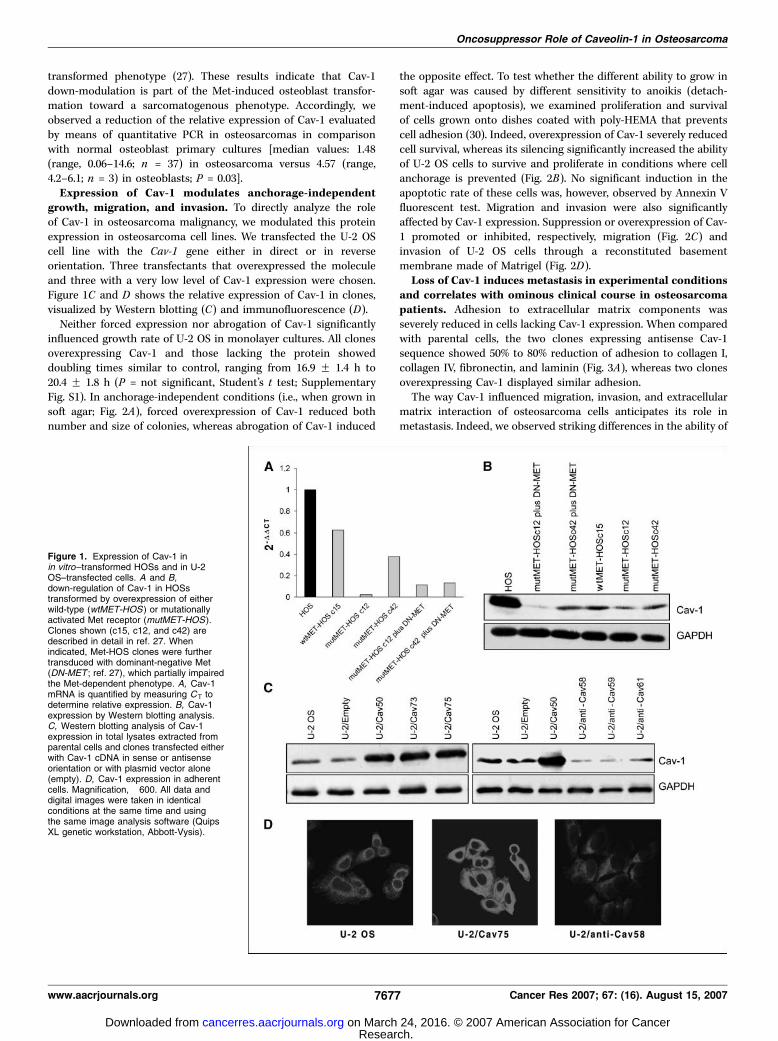
transformed phenotype (27). These results indicate that Cav-1down-modulation is part of the Met-induced osteoblast transfor-mation toward a sarcomatogenous phenotype. Accordingly, weobserved a reduction of the relative expression of Cav-1 evaluatedby means of quantitative PCR in osteosarcomas in comparisonwith normal osteoblast primary cultures [median values: 1.48(range, 0.06–14.6; n = 37) in osteosarcoma versus 4.57 (range,4.2–6.1; n = 3) in osteoblasts; P = 0.03].Expression of Cav-1 modulates anchorage-independent
growth, migration, and invasion. To directly analyze the roleof Cav-1 in osteosarcoma malignancy, we modulated this proteinexpression in osteosarcoma cell lines. We transfected the U-2 OScell line with the Cav-1 gene either in direct or in reverseorientation. Three transfectants that overexpressed the moleculeand three with a very low level of Cav-1 expression were chosen.Figure 1C and D shows the relative expression of Cav-1 in clones,visualized by Western blotting (C) and immunofluorescence (D).Neither forced expression nor abrogation of Cav-1 significantly
influenced growth rate of U-2 OS in monolayer cultures. All clonesoverexpressing Cav-1 and those lacking the protein showeddoubling times similar to control, ranging from 16.9 F 1.4 h to20.4 F 1.8 h (P = not significant, Student’s t test; SupplementaryFig. S1). In anchorage-independent conditions (i.e., when grown insoft agar; Fig. 2A), forced overexpression of Cav-1 reduced bothnumber and size of colonies, whereas abrogation of Cav-1 induced
the opposite effect. To test whether the different ability to grow insoft agar was caused by different sensitivity to anoikis (detach-ment-induced apoptosis), we examined proliferation and survivalof cells grown onto dishes coated with poly-HEMA that preventscell adhesion (30). Indeed, overexpression of Cav-1 severely reducedcell survival, whereas its silencing significantly increased the abilityof U-2 OS cells to survive and proliferate in conditions where cellanchorage is prevented (Fig. 2B). No significant induction in theapoptotic rate of these cells was, however, observed by Annexin Vfluorescent test. Migration and invasion were also significantlyaffected by Cav-1 expression. Suppression or overexpression of Cav-1 promoted or inhibited, respectively, migration (Fig. 2C) andinvasion of U-2 OS cells through a reconstituted basementmembrane made of Matrigel (Fig. 2D).Loss of Cav-1 induces metastasis in experimental conditions
and correlates with ominous clinical course in osteosarcomapatients. Adhesion to extracellular matrix components wasseverely reduced in cells lacking Cav-1 expression. When comparedwith parental cells, the two clones expressing antisense Cav-1sequence showed 50% to 80% reduction of adhesion to collagen I,collagen IV, fibronectin, and laminin (Fig. 3A), whereas two clonesoverexpressing Cav-1 displayed similar adhesion.The way Cav-1 influenced migration, invasion, and extracellular
matrix interaction of osteosarcoma cells anticipates its role inmetastasis. Indeed, we observed striking differences in the ability of
Figure 1. Expression of Cav-1 inin vitro–transformed HOSs and in U-2OS–transfected cells. A and B,down-regulation of Cav-1 in HOSstransformed by overexpression of eitherwild-type (wtMET-HOS ) or mutationallyactivated Met receptor (mutMET-HOS).Clones shown (c15, c12, and c42) aredescribed in detail in ref. 27. Whenindicated, Met-HOS clones were furthertransduced with dominant-negative Met(DN-MET ; ref. 27), which partially impairedthe Met-dependent phenotype. A, Cav-1mRNA is quantified by measuring CT todetermine relative expression. B, Cav-1expression by Western blotting analysis.C, Western blotting analysis of Cav-1expression in total lysates extracted fromparental cells and clones transfected eitherwith Cav-1 cDNA in sense or antisenseorientation or with plasmid vector alone(empty). D, Cav-1 expression in adherentcells. Magnification, �600. All data anddigital images were taken in identicalconditions at the same time and usingthe same image analysis software (QuipsXL genetic workstation, Abbott-Vysis).
Oncosuppressor Role of Caveolin-1 in Osteosarcoma
www.aacrjournals.org 7677 Cancer Res 2007; 67: (16). August 15, 2007
Research. on March 24, 2016. © 2007 American Association for Cancercancerres.aacrjournals.org Downloaded from
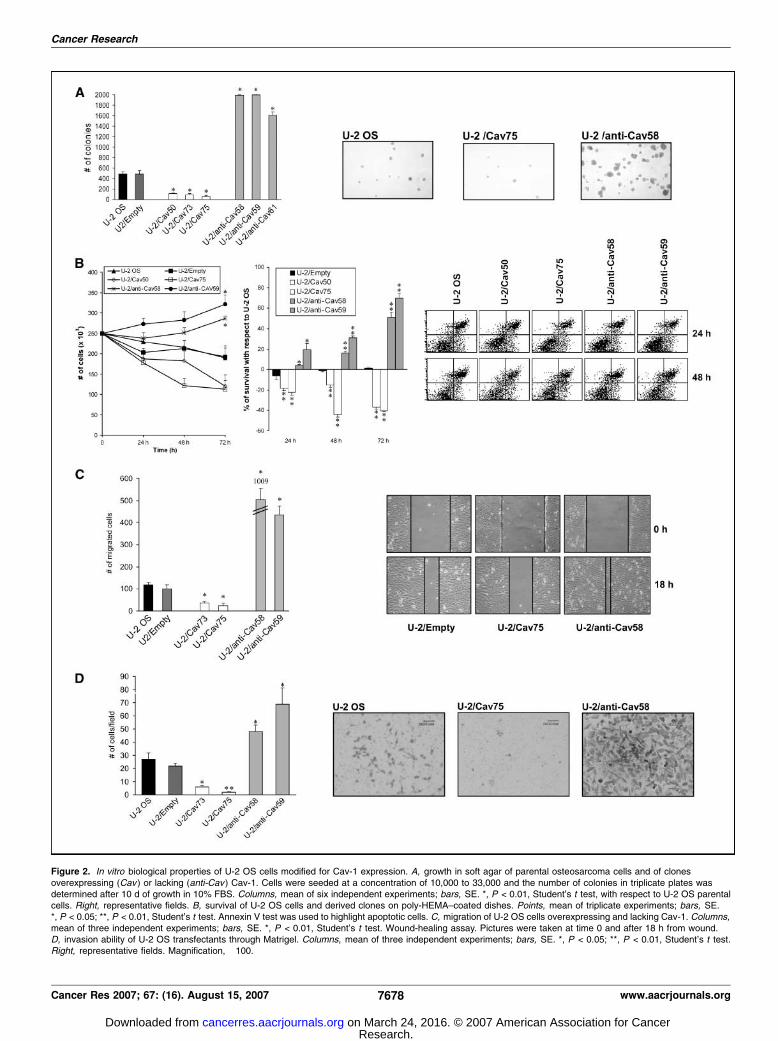
Figure 2. In vitro biological properties of U-2 OS cells modified for Cav-1 expression. A, growth in soft agar of parental osteosarcoma cells and of clonesoverexpressing (Cav ) or lacking (anti-Cav ) Cav-1. Cells were seeded at a concentration of 10,000 to 33,000 and the number of colonies in triplicate plates wasdetermined after 10 d of growth in 10% FBS. Columns, mean of six independent experiments; bars, SE. *, P < 0.01, Student’s t test, with respect to U-2 OS parentalcells. Right, representative fields. B, survival of U-2 OS cells and derived clones on poly-HEMA–coated dishes. Points, mean of triplicate experiments; bars, SE.*, P < 0.05; **, P < 0.01, Student’s t test. Annexin V test was used to highlight apoptotic cells. C, migration of U-2 OS cells overexpressing and lacking Cav-1. Columns,mean of three independent experiments; bars, SE. *, P < 0.01, Student’s t test. Wound-healing assay. Pictures were taken at time 0 and after 18 h from wound.D, invasion ability of U-2 OS transfectants through Matrigel. Columns, mean of three independent experiments; bars, SE. *, P < 0.05; **, P < 0.01, Student’s t test.Right, representative fields. Magnification, �100.
Cancer Research
Cancer Res 2007; 67: (16). August 15, 2007 7678 www.aacrjournals.org
Research. on March 24, 2016. © 2007 American Association for Cancercancerres.aacrjournals.org Downloaded from
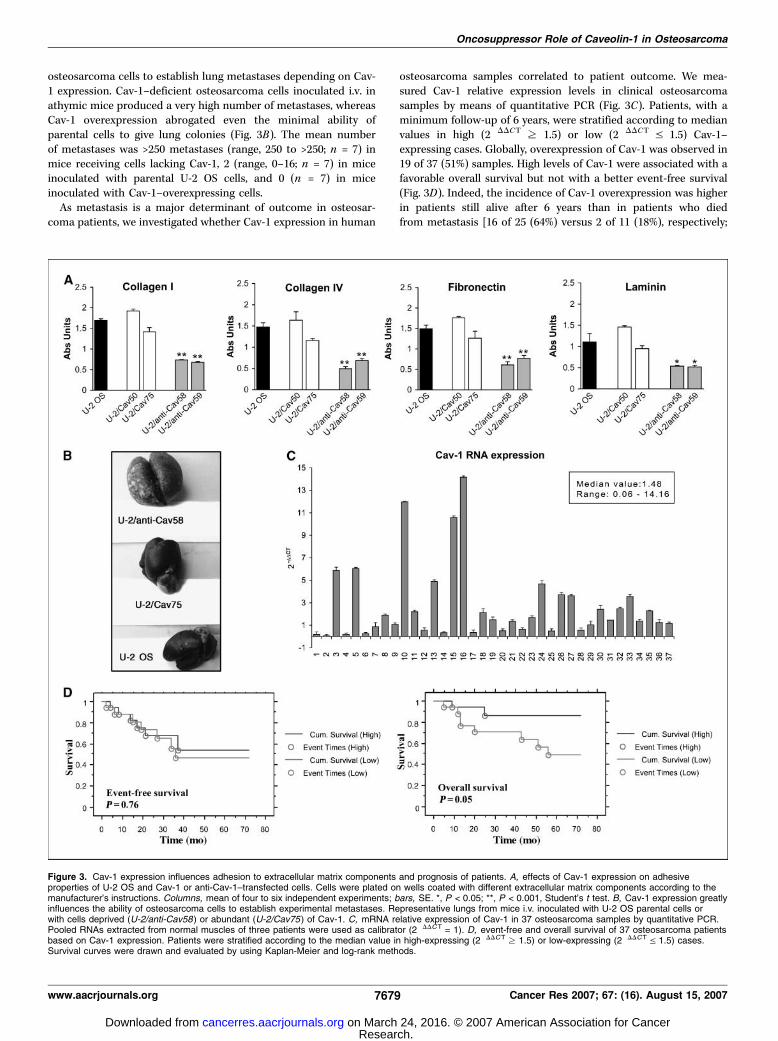
osteosarcoma cells to establish lung metastases depending on Cav-1 expression. Cav-1–deficient osteosarcoma cells inoculated i.v. inathymic mice produced a very high number of metastases, whereasCav-1 overexpression abrogated even the minimal ability ofparental cells to give lung colonies (Fig. 3B). The mean numberof metastases was >250 metastases (range, 250 to >250; n = 7) inmice receiving cells lacking Cav-1, 2 (range, 0–16; n = 7) in miceinoculated with parental U-2 OS cells, and 0 (n = 7) in miceinoculated with Cav-1–overexpressing cells.As metastasis is a major determinant of outcome in osteosar-
coma patients, we investigated whether Cav-1 expression in human
osteosarcoma samples correlated to patient outcome. We mea-sured Cav-1 relative expression levels in clinical osteosarcomasamples by means of quantitative PCR (Fig. 3C). Patients, with aminimum follow-up of 6 years, were stratified according to medianvalues in high (2�DDCT z 1.5) or low (2�DDCT V 1.5) Cav-1–expressing cases. Globally, overexpression of Cav-1 was observed in19 of 37 (51%) samples. High levels of Cav-1 were associated with afavorable overall survival but not with a better event-free survival(Fig. 3D). Indeed, the incidence of Cav-1 overexpression was higherin patients still alive after 6 years than in patients who diedfrom metastasis [16 of 25 (64%) versus 2 of 11 (18%), respectively;
Figure 3. Cav-1 expression influences adhesion to extracellular matrix components and prognosis of patients. A, effects of Cav-1 expression on adhesiveproperties of U-2 OS and Cav-1 or anti-Cav-1–transfected cells. Cells were plated on wells coated with different extracellular matrix components according to themanufacturer’s instructions. Columns, mean of four to six independent experiments; bars, SE. *, P < 0.05; **, P < 0.001, Student’s t test. B, Cav-1 expression greatlyinfluences the ability of osteosarcoma cells to establish experimental metastases. Representative lungs from mice i.v. inoculated with U-2 OS parental cells orwith cells deprived (U-2/anti-Cav58) or abundant (U-2/Cav75) of Cav-1. C, mRNA relative expression of Cav-1 in 37 osteosarcoma samples by quantitative PCR.Pooled RNAs extracted from normal muscles of three patients were used as calibrator (2�DDCT = 1). D, event-free and overall survival of 37 osteosarcoma patientsbased on Cav-1 expression. Patients were stratified according to the median value in high-expressing (2�DDCT z 1.5) or low-expressing (2�DDCT V 1.5) cases.Survival curves were drawn and evaluated by using Kaplan-Meier and log-rank methods.
Oncosuppressor Role of Caveolin-1 in Osteosarcoma
www.aacrjournals.org 7679 Cancer Res 2007; 67: (16). August 15, 2007
Research. on March 24, 2016. © 2007 American Association for Cancercancerres.aacrjournals.org Downloaded from
P = 0.03, Fisher’s exact test], although Cav-1 overexpression wasevenly distributed when the first relapse occurred (52% of positivecases in patients with no evidence of disease versus 48% in relapsedpatients).We considered that patient outcome depends on both tumor
intrinsic malignancy and response to treatments, which very likelydetermine overall and event-free survival, respectively. Thus, westudied the possible association between Cav-1 expression andresponse to chemotherapy. In vitro MTT assay indicated that Cav-1did not significantly modulate osteosarcoma response to doxoru-bicin, one of the leader drugs in the treatment of sarcomas(Supplementary Fig. S2), further confirming that Cav-1 expressioncorrelates with osteosarcoma aggressiveness rather than chemo-sensitivity.Moreover, we studied three cases of patients with high-grade
periosteal osteosarcoma who had wide surgery but no chemother-apy. The two patients whose tumors showed high Cav-1 mRNAexpression (2�DDCT z 1.5) were disease-free after 8 years, whereasthe one with a low Cav-1–expressing tumor had metastasis within
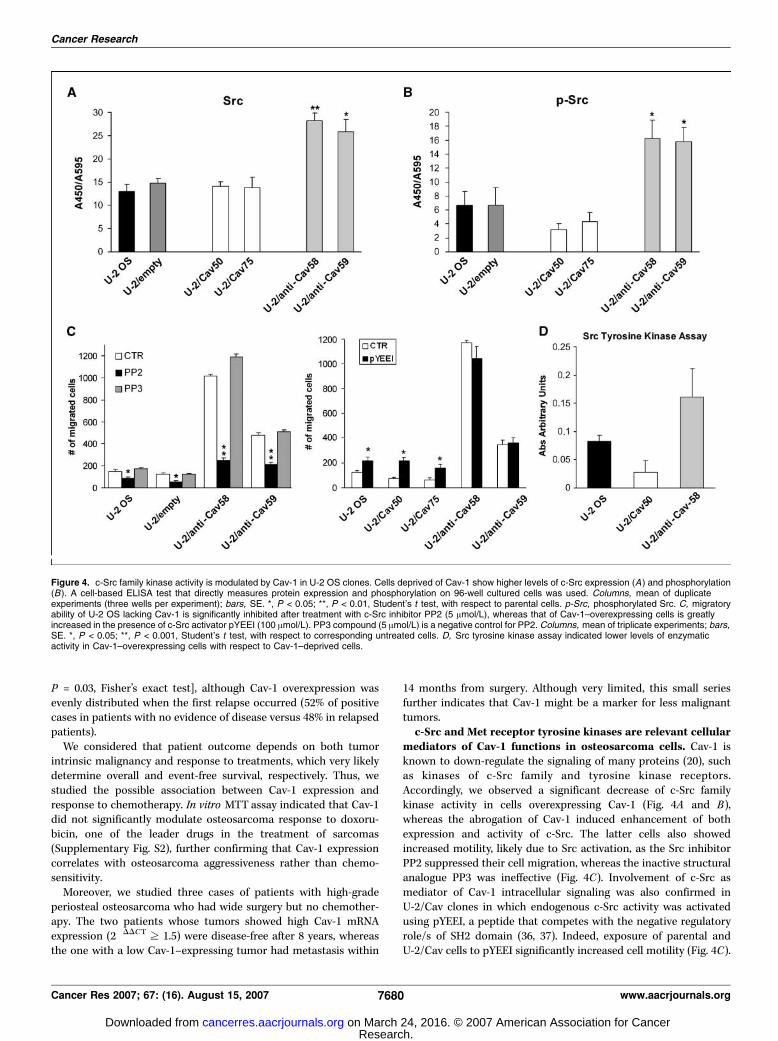
14 months from surgery. Although very limited, this small seriesfurther indicates that Cav-1 might be a marker for less malignanttumors.c-Src and Met receptor tyrosine kinases are relevant cellular
mediators of Cav-1 functions in osteosarcoma cells. Cav-1 isknown to down-regulate the signaling of many proteins (20), suchas kinases of c-Src family and tyrosine kinase receptors.Accordingly, we observed a significant decrease of c-Src familykinase activity in cells overexpressing Cav-1 (Fig. 4A and B),whereas the abrogation of Cav-1 induced enhancement of bothexpression and activity of c-Src. The latter cells also showedincreased motility, likely due to Src activation, as the Src inhibitorPP2 suppressed their cell migration, whereas the inactive structuralanalogue PP3 was ineffective (Fig. 4C). Involvement of c-Src asmediator of Cav-1 intracellular signaling was also confirmed inU-2/Cav clones in which endogenous c-Src activity was activatedusing pYEEI, a peptide that competes with the negative regulatoryrole/s of SH2 domain (36, 37). Indeed, exposure of parental andU-2/Cav cells to pYEEI significantly increased cell motility (Fig. 4C).
Figure 4. c-Src family kinase activity is modulated by Cav-1 in U-2 OS clones. Cells deprived of Cav-1 show higher levels of c-Src expression (A) and phosphorylation(B). A cell-based ELISA test that directly measures protein expression and phosphorylation on 96-well cultured cells was used. Columns, mean of duplicateexperiments (three wells per experiment); bars, SE. *, P < 0.05; **, P < 0.01, Student’s t test, with respect to parental cells. p-Src, phosphorylated Src. C, migratoryability of U-2 OS lacking Cav-1 is significantly inhibited after treatment with c-Src inhibitor PP2 (5 Amol/L), whereas that of Cav-1–overexpressing cells is greatlyincreased in the presence of c-Src activator pYEEI (100 Amol/L). PP3 compound (5 Amol/L) is a negative control for PP2. Columns, mean of triplicate experiments; bars,SE. *, P < 0.05; **, P < 0.001, Student’s t test, with respect to corresponding untreated cells. D, Src tyrosine kinase assay indicated lower levels of enzymaticactivity in Cav-1–overexpressing cells with respect to Cav-1–deprived cells.
Cancer Research
Cancer Res 2007; 67: (16). August 15, 2007 7680 www.aacrjournals.org
Research. on March 24, 2016. © 2007 American Association for Cancercancerres.aacrjournals.org Downloaded from

In addition, kinase assay confirmed a higher c-Src enzymaticactivity in cells lacking Cav-1 (Fig. 4D), whereas those over-expressing Cav-1 showed lower levels of activity according totheir low migration ability. In keeping with these data, a different
osteosarcoma cell line, Saos-2, showed a higher basal level ofCav-1 (Fig. 5A and B), lower amount of c-Src Y416 phosphorylation(Fig. 5B), and minor ability to grow in soft agar and invade Matrigel(Fig. 5C) than U-2 OS. Accordingly, Saos-2 cells lacking Cav-1
Figure 5. Cav-1 expression and c-Src family kinase activity are inversely related and oppositely modulate migration and anchorage-independent growth of Saos-2osteosarcoma cells. A, immunostaining of Cav-1 on adherent U-2 OS and Saos-2 cells. Caveolin is labeled by anti-caveolin antibody and revealed by FITC-conjugatedanti-rabbit secondary antibody. B, evaluation of c-Src kinase activity in U-2 OS and Saos-2 cells by Western blotting and Src tyrosine kinase assay. C, higher expressionof Cav-1 corresponds to lower Y416 c-Src phosphorylation and lower ability of Saos-2 cells to grow in soft agar and to invade Matrigel. *, P < 0.05,Student’s t test. D, loss of Cav-1 in Saos-2 cells, by antisense Cav-1 transduction, confirmed the inverse correlation between Cav-1 expression and c-Src activityas well as osteosarcoma malignancy as indicated by anchorage-independent growth, migration, and invasion. *, P < 0.05; **, P < 0.001, Student’s t test, with respect toparental cell line.
Oncosuppressor Role of Caveolin-1 in Osteosarcoma
www.aacrjournals.org 7681 Cancer Res 2007; 67: (16). August 15, 2007
Research. on March 24, 2016. © 2007 American Association for Cancercancerres.aacrjournals.org Downloaded from

enhanced motility, growth, and survival in anchorage-independentconditions and invasion through Matrigel and displayed a higherlevel of c-Src Y416 phosphorylation than parental Saos-2 cells(Fig. 5D). In addition, Saos-2 cells expressed Met at a lower levelthan U-2 OS osteosarcoma cells (Fig. 6A). The inverse relationshipbetween Cav-1 and Met expression and correlation to malignancywas confirmed in other osteosarcoma cell lines (Table 1) as alsoshown in Met-transformed osteoblasts (see above). In addition, U-2OS clones overexpressing Cav-1 showed lower levels of Metexpression (Fig. 6B), indicating that oncosuppressor activity ofCav-1 might account for Met functions. Moreover, costaining withanti-Cav-1 and anti-Met antibodies showed their colocalization atplasma membrane (Fig. 6C). Therefore, we tested whether Cav-1can inhibit Met functions by down-regulating Met signalingpathway. Both extracellular signal-regulated kinase (ERK)/mito-gen-activated protein kinase (MAPK) and phosphatidylinositol3-kinase (PI3K) are important mediators of proliferation, motility,and invasion of osteosarcoma cells in response to Met ligand (HGF)stimulation (38). In addition, c-Src signaling is activated by HGFtaking part to migration, invasion, and anchorage-independentgrowth of other tumor cell types (39, 40). A colocalization betweenCav-1 and c-Src is here shown (Fig. 6C) in agreement with previousreports (41, 42). Although we did not observe any change in ERK/MAPK activation status of Cav-1–overexpressing U-2 OS (data notshown), the latter cells showed reduced PI3K activation with
respect to parental cells both in basal conditions and, moreimportantly, after exposure to HGF (Fig. 6D). Striking attenuationof c-Src kinase activity was also observed in the U-2 OS model,although the use of anti-Cav Saos-2 cells provided mirroredinformation (Fig. 6D), further confirming that Cav-1 modulatesHGF signaling by negatively regulating both Src and Akt activities.
Discussion
This article investigated the role of Cav-1 in osteosarcomamalignancy. Cav-1 is highly expressed in normal osteoblasts (16). Inthese cells, important signaling molecules (e.g., growth receptors,heterotrimeric G proteins, Src family tyrosine kinase, and MAPKpathway) form cluster within caveolin-rich membrane complexes(16, 17). We show that Cav-1 is down-regulated in HOSstransformed by overexpressing either the wild-type Met, thetyrosine kinase receptor of HGF, or its mutationally activatedversion (27). This correlates with the observation that primaryosteosarcomas have significantly lower levels of Cav-1 than normalosteoblasts.When in osteosarcoma, Cav-1 expression is up-regulated, and
similar to that of normal osteoblasts, patients show a significantlybetter outcome. An association with favorable prognosis wasobserved with overall, but not event-free, survival of osteosarcomapatients. This different clinical effect is, in our opinion, due to the
Figure 6. Met expression and signaling are modulated by Cav-1. A, Met expression in U-2 OS and Saos-2 osteosarcoma cell lines by cytofluorimetry. Openprofile, cells stained with secondary antibody alone; solid profile, cells stained with anti-Met antibody. B, cytofluorimetric profile of Met expression in U-2 OS Cav-1transfectants. A and B, data from an experiment representative of at least two similar experiments. C, immunofluorescent staining of Cav-1 together with Met or Src inSaos-2 osteosarcoma cells. Merged images indicate the extensive formation of Cav-1-Met or Cav-1-Src complexes and colocalization of molecules at cell surface.Digital images were taken in identical conditions using the image analysis software Quips XL genetic workstation. D, Akt and Src phosphorylation status was checkedboth on U-2 OS and Saos-2 models either in basal condition or after HGF stimulation. pAkt, phosphorylated Akt.
Cancer Research
Cancer Res 2007; 67: (16). August 15, 2007 7682 www.aacrjournals.org
Research. on March 24, 2016. © 2007 American Association for Cancercancerres.aacrjournals.org Downloaded from

fact that Cav-1 is a key determinant of biological malignancy ofosteosarcoma cells but does not influence their sensitivity tochemotherapeutic agents. Consistently, we did not observe anyvariation in chemosensitivity to doxorubicin, a major drug in thetreatment of osteosarcoma, in cells deprived or abundant of Cav-1.On the contrary, modulation of Cav-1 expression in osteosarcomacell lines greatly modifies the pattern of anchorage-independentgrowth, migration, invasion, adhesion to extracellular matrixcomponents, and, more importantly, their metastatic ability.Abrogation of Cav-1 dramatically enhanced lung metastasis inathymic mice, in agreement with greater resistance to anoikis andindependence from anchorage for growing, lower adhesion tofibronectin, laminin, and collagens, and a striking motile phenotypein vitro . These results show that Cav-1 suppresses osteosarcomamalignancy and are in line with previous observations infibrosarcoma and other mesenchymal tumors (23) but not withthe recent report on Ewing’s sarcoma that indicates an oncogenicrole for Cav-1 (43). These contradictory findings may reflect thedifferent origin and genetic features of osteosarcoma and othermesenchymal tumors compared with Ewing’s sarcoma, the twoneoplasms being the prototypes of two distinct classes of sarcoma(44), and support the need to study each gene in its appropriatecellular context. This seems to be particularly true for Cav-1 thathas been described as an oncogene or a tumor suppressor genedepending on the cell types. Particularly in prostate carcinoma (22)but also esophageal squamous cell, non–small cell lung carcinoma,and Ewing’s sarcoma (43, 45, 46), up-regulation of the proteincontributes to metastatic phenotype. The opposite seems to occurnot only in osteosarcoma and fibrosarcoma (23) but also inmammary (47) and colon (48) carcinogenesis. All these apparentlycontradictory results might be related to the role of Cav-1 in thedifferentiation of each cell type. More likely, Cav-1 might mainlyplay a permissive role by unleashing the full oncogenic orantioncogenic properties of the proteins, which are sequesteredin caveolae in each tumor type.In osteosarcomas, low Cav-1 expression is unlikely due to genetic
or epigenetic alterations. In fact, although Cav-1 gene maps to aknown fragile site (7q31.1) that shows deletions in a variety ofmalignancies, no mutation of Cav-1 gene nor evidence of itsmethylation within the promoter or in the first two exons has beenreported in osteosarcoma cells (49). More likely, Cav-1 expression is
regulated at the level of transcription, and in osteosarcoma cells, itmay be also further decreased by a feedback circuit. We show thatMet signaling is enhanced in osteosarcoma cells where Cav-1 issuppressed; it is noteworthy that the consistent and highexpression of Met in osteosarcoma (33, 34) could consecutivelycontribute to Cav-1 down-modulation. In addition, Met ligand HGFis abundant in bone tumor microenvironment and might beresponsible of Cav-1 down-regulation through inhibition of itspromoter activity as shown in endothelia and muscle satellite cells(50–52). Conversely, Cav-1 expression might be a marker ofosteoblast differentiation and low Cav-1 an intrinsic property ofthe osteoblast stem cell or progenitor targeted by transformation.Indeed, we observed a direct relationship between Cav-1 expres-sion and level of osteoblastic differentiation in osteosarcoma celllines (12) and, more recently, found evidence of physicalconnection between Cav-1 and CD99 (13), a molecule involved incell-cell adhesion, migration, and apoptosis that was alsoimplicated in osteoblastic differentiation (53, 54). A correlationbetween Cav-1 expression and osteoblastic differentiation was alsoconfirmed when undifferentiated U-2 OS cells were compared withthe osteoblast-like Saos-2 cells. Of note, expression of Met isinversely correlated with osteoblastic differentiation (33), reinforc-ing the view of Met contributing to osteoblast transformation andosteosarcoma progression in association to the down-modulationof Cav-1 and loss of differentiation. The latter hypothesis issupported by data showing that impairment of Met-dependentphenotype in transformed osteoblasts, by means of a dominant-negative Met receptor, does not directly influence Cav-1 expression.Regardless of whether Cav-1 low expression is an intrinsic
property of cells targeted by transformation or a consequence oftransformation, the oncosuppressor role of Cav-1 may pass throughthe modulation of activity of c-Src family tyrosine kinases as well asMet signaling. Cav-1 is known to bind and sequester Src familykinases in an inactive configuration (41, 42). Furthermore, Cav-1enhances inactivation of c-Src by CSK, therefore directly andindirectly shifting the equilibrium in favor of inactive c-Srcconformation (55). Accordingly, Cav-1–deprived osteosarcoma cellsexhibited marked increase in c-Src family tyrosine kinase activity,which significantly contributes to their migration and, very likely,to anchorage-independent growth. In fact, motility of cells lackingCav-1 was strikingly suppressed when c-Src kinase activity was
Table 1. Expression of Cav-1 and Met receptor in human osteosarcoma cell lines in relation to their tumorigenicity in nudemice
Cell line Cav-1 expression* (�DCT) Met expressionc
Tumorigenicityb
% Positive cell Log mean Incidence Latency (days)
U-2 OS �0.44 F 0.1 88.6 19.6 9/10 97 F 8
SaoS-2 0.24 F 0.09 56.5 8.7 0/5 —
MG63 0.88 F 0.05 55.6 9.3 1/5 188OS-7 �1.72 F 0.06 97.2 44.9 4/5 62 F 10
OS-9 �1.24 F 0.07 94.4 33.6 5/5 18 F 7
*Relative mRNA Cav-1 expression evaluated by real-time PCR: �DCT, �(CT Cav-1 � CT GAPDH). Higher �DCT therefore indicates a higher level ofmRNA target expression.cCytofluorimetry evaluation of cells immunostained with DO-24 anti-Met monoclonal antibody. Data from one representative experiment.bCells (30 � 106) were s.c. injected in nude mice. Incidence of positive mice/total.
Oncosuppressor Role of Caveolin-1 in Osteosarcoma
www.aacrjournals.org 7683 Cancer Res 2007; 67: (16). August 15, 2007
Research. on March 24, 2016. © 2007 American Association for Cancercancerres.aacrjournals.org Downloaded from

inhibited. On the other hand, cells overexpressing Cav-1, whichexhibited very low migration capacities, showed low levels of c-Srcactivity. When restored by using a Src-specific activator, cellsconsequently increased their migration. We also found Cav-1overexpression associated to decrease of Met expression andsignaling in U-2 OS and Saos-2 osteosarcoma cells, indicating thatthe oncosuppressor activity of Cav-1 also accounts for Metfunctions. Our hypothesis is that Met is endocytosed from the cellsurface by a caveolae-dependent mechanism in analogy with theCav-1–mediated internalization process that has been described tocounteract epidermal growth factor receptor signaling (56). In thepresence of abundant Cav-1, cellular response to HGF is repressed.Particularly, following HGF stimulation, a reduction in activationstatus of PI3K and c-Src, two pathways involved in HGF-inducedmitogenic and motogenic machinery, was seen (38). This indicatesthat Cav-1, which colocalizes with Met, might act as an inhibitorof HGF/Met signaling. To establish whether inhibitory effects ofCav-1 on c-Src and Met signaling are a reciprocal consequence orrepresent two separate mechanisms accounting for the oncosup-pressive role of this adaptor protein is beyond the purpose ofthis study. However, it is tempting to speculate that Cav-1 mightinduce a coordinated and global inhibition of HGF-activated loopby acting both on c-Src and HGF/Met signaling. Activated c-Srcincreased expression of HGF mRNA and protein and cell scattering
(57). On the other hand, Src is an already known downstreammediator of HGF signaling pathway (58), thus establishing apositive regulatory feedback loop. Cav-1 by inhibiting c-Src activityvia its scaffolding domain interrupts this positive regulation and bydecreasing Met expression further disrupts HGF autocrine loop,therefore depriving osteosarcoma cells of a fundamental pathwayfor their migration and metastasis.Globally, our results show Cav-1 as a crucial regulator of
osteosarcoma malignancy and indicate that Cav-1 down-modula-tion might potentiate both Src family kinase and Met signaling,which provide osteosarcoma cells with the capacity of invadingneighboring tissues. As a consequence, targeted therapies againstc-Src family kinases and/or Met receptor may have a rationale toimprove the extremely poor prognosis of metastatic osteosarcoma.
Acknowledgments
Received 12/27/2006; revised 5/30/2007; accepted 6/7/2007.Grant support: EU grant EuroBoNet 018814 and Associazione Italiana per la
Ricerca sul Cancro (K. Scotlandi and M.F. Di Renzo).The costs of publication of this article were defrayed in part by the payment of page
charges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.
We thank Mordechai Liscovitch (Department of Biological Regulation, WeizmannInstitute of Science, Rehovot, Israel) for providing Cav-1–expressing plasmid andTimothy C. Thompson (Baylor College of Medicine, Texas Medical Center, Houston,TX) for providing antisense Cav-1–expressing plasmid.
Cancer Research
Cancer Res 2007; 67: (16). August 15, 2007 7684 www.aacrjournals.org
References1. Campanacci M. Bone and soft tissue tumors. 2nd ed.Wien (NY): Springer-Verlag; 1999.2. Bramwell VH. The role of chemotherapy in themanagement of non-metastatic operable extremityosteosarcoma. Semin Oncol 1997;24:561–71.3. Bacci G, Longhi A, Fagioli F, Briccoli A, Versari M, PicciP. Adjuvant and neoadjuvant chemotherapy for osteo-sarcoma of the extremities: 27 year experience at RizzoliInstitute, Italy. Eur J Cancer 2005;41:2836–45.4. Weber KL. What’s new in musculoskeletal oncology.J Bone Joint Surg Am 2005;87:1400–10.5. Chan HS, Grogan TM, Haddad G, DeBoer G, Ling V.P-glycoprotein expression: critical determinant in theresponse to osteosarcoma chemotherapy. J Natl CancerInst 1997;89:1706–15.6. Pakos EE, Ioannidis JP. The association of P-glycopro-tein with response to chemotherapy and clinical out-come in patients with osteosarcoma. A meta-analysis.Cancer 2003;98:581–9.7. Serra M, Scotlandi K, Reverter-Branchat G, et al. Valueof P-glycoprotein and clinicopathologic factors as thebasis for new treatment strategies in high-gradeosteosarcoma of the extremities. J Clin Oncol 2003;21:536–42.8. Bacci G, Longhi A, Versari M, Mercuri M, Briccoli A,Picci P. Prognostic factors for osteosarcoma of theextremity treated with neoadjuvant chemotherapy:15-year experience in 789 patients treated at a singleinstitution. Cancer 2006;106:1154–61.9. Khanna C, Khan J, Nguyen P, et al. Metastasis-associated differences in gene expression in a murinemodel of osteosarcoma. Cancer Res 2001;61:3750–9.10. Mintz MB, Sowers R, Brown KM, et al. An expressionsignature classifies chemotherapy-resistant pediatricosteosarcoma. Cancer Res 2005;65:1748–54.11. Laverdiere C, Hoang BH, Yang R, et al. MessengerRNA expression levels of CXCR4 correlate with meta-static behavior and outcome in patients with osteosar-coma. Clin Cancer Res 2005;11:2561–7.12. Zucchini C, Bianchini M, Valvassori L, et al.Identification of candidate genes involved in thereversal of malignant phenotype of osteosarcoma cellstransfected with the liver/bone/kidney alkaline phos-phatase gene. Bone 2004;34:672–9.
13. Manara MC, Bernard G, Lollini P-L, et al. CD99 actsas an oncosuppressor in osteosarcoma. Mol Biol Cell2006;17:1910–21.14. Stan RV. Structure of caveolae. Biochim Biophys Acta2005;1746:334–48.15. Smart EJ, Graf GA, McNiven MA, et al. Caveolins,liquid-ordered domains, and signal transduction. MolCell Biol 1999;19:7289–304.16. Solomon KR, Danciu TE, Adolphson LD, Hecht LE,Hauschka PV. Caveolin-enriched membrane signalingcomplexes in human and murine osteoblasts. J BoneMiner Res 2000;15:2380–90.17. Solomon KR, Adolphson LD, Wank DA, McHugh KP,Hauschka PV. Caveolae in human and murine osteo-blasts. J Bone Miner Res 2000;15:2391–401.18. Carver LA, Schnitzer JE. Caveolae: mining little cavesfor new cancer targets. Nat Rev Cancer 2003;3:571–81.19. Krajewska WM, Maslowska I. Caveolins: structureand function in signal transduction. Cell Mol Biol Lett2004;9:195–220.20. Williams TM, Lisanti MP. Caveolin-1 in oncogenictransformation, cancer, and metastasis. Am J PhysiolCell Physiol 2005;288:494–506.21. Nasu Y, Timme TL, Yang G, et al. Suppression ofcaveolin expression induces androgen sensitivity inmetastatic androgen-insensitive mouse prostate cancercells. Nat Med 1998;4:1062–4.22. Bangma CH, Nasu Y, Ren C, Thompson TC.Metastasis-related genes in prostate cancer. SeminOncol 1999;26:422–7.23. Wiechen K, Sers C, Agoulnik A, et al. Down-regulation of caveolin-1, a candidate tumor suppressorgene, in sarcomas. Am J Pathol 2001;158:833–9.24. Williams TM, Medina F, Badano I, et al. Caveolin-1gene disruption promotes mammary tumorigenesisand dramatically enhances lung metastasis in vivo .Role of Cav-1 in cell invasiveness and matrix metal-loproteinase (MMP-2/9) secretion. J Biol Chem 2004;279:51630–46.25. Sloan EK, Stanley KL, Anderson RL. Caveolin-1inhibits breast cancer growth and metastasis. Oncogene2004;23:7893–7.26. Benini S, Baldini N, Manara MC, et al. Redundancy ofautocrine loops in human osteosarcoma cells. Int JCancer 1999;80:581–8.27. Patane S, Avnet S, Coltella N, et al. MET over-
expression turns human primary osteoblasts intoosteosarcomas. Cancer Res 2006;66:4750–7.28. Fiucci G, Ravid D, Reich R, Liscovitch M. Caveolin-1inhibits anchorage-independent growth, anoikis andinvasiveness in MCF-7 human breast cancer cells.Oncogene 2002;21:2365–75.29. Prat MP, Narsimhn RP, Crepaldi T, Nicotra MR, NataliPG, Comoglio PM. The receptor encoded by the humanc-MET oncogene is expressed in hepatocytes, epithelialcells and solid tumors. Int J Cancer 1991;49:323–8.30. Folkman J, Moscona A. Role of cell shape in growthcontrol. Nature 1978;273:345–9.31. Ferrari S, Mercuri M, Picci P, et al. Nonmetastaticosteosarcoma of the extremity: results of a neoadjuvantchemotherapy protocol (IOR/OS3) with high-dosemethotrexate, intraarterial or intravenous cisplatin,doxorubicin, and salvage chemotherapy based onhistologic tumor response. Tumori 1999;85:458–64.32. Bacci G, Ferrari S, Longhi A, et al. Neoadjuvantchemotherapy for high-grade osteosarcoma of theextremities: long-term results for patients treatedaccording to the Rizzoli IOR/OS-3b protocol. J Chemo-ther 2001;13:93–9.33. Ferracini R, Di Renzo MF, Scotlandi K, et al. The Met/HGF receptor is over-expressed in human osteosarco-mas and is activated by either a paracrine or anautocrine circuit. Oncogene 1995;10:739–49.34. Scotlandi K, Baldini N, Olivero M, et al. Expression ofMet/hepatocyte growth factor receptor gene andmalignant behavior of musculoskeletal tumors. Am JPathol 1996;149:1209–19.35. Di Renzo MF, Olivero M, Martone T, et al. Somaticmutations of the MET oncogene are selected duringmetastatic spread of human HNSC carcinomas. Onco-gene 2000;19:1547–55.36. Wijentunge S, Hughes AD. Activation of endogenousc-Src or a related tyrosine kinase by intracellular(pY)EEI peptide increases voltage-operated calciumchannel currents in rabbit ear artery cells. FEBS Lett1996;399:63–6.37. Porter M, Schindler T, Kuriyan J, Miller WT.Reciprocal regulation of Hck activity by phosphorylationof Tyr(527) and Tyr(416). Effect of introducing a highaffinity intramolecular SH2 ligand. J Biol Chem 2000;275:2721–6.38. Coltella N, Manara MC, Cerisano V, et al. Role of the
Research. on March 24, 2016. © 2007 American Association for Cancercancerres.aacrjournals.org Downloaded from

MET/HGF receptor in proliferation and invasive behav-ior of osteosarcoma. FASEB J 2003;17:1162–4.39. Cutrupi S, Baldanzi G, Gramaglia D, et al. Src-mediated activation of a-diacylglycerol kinase is re-quired for hepatocyte growth factor-induced cellmotility. EMBO J 2000;19:4614–22.40. Atabey N, Gao Y, Yao ZJ, et al. Potent blockade ofhepatocyte growth factor-stimulated cell motility, ma-trix invasion and branching morphogenesis by antago-nists of Grb2 Src homology 2 domain interactions. J BiolChem 2001;276:14308–14.41. Li S, Couet J, Lisanti MP. Src tyrosine kinases, Gasubunits, and H-Ras share a common membrane-anchored scaffolding protein, caveolin. Caveolin bindingnegatively regulates the auto-activation of Src tyrosinekinases. J Biol Chem 1996;271:29182–90.42. Wei Y, Yang X, Liu Q, Wilkins JA, Chapman HA. A rolefor caveolin and the urokinase receptor in integrin-mediated adhesion and signaling. J Cell Biol 1999;144:1285–94.43. Tirado OM, Mateo-Lozano S, Villar J, et al. Caveolin-1(CAV1) is a target of EWS/FLI-1 and a key determinantof the oncogenic phenotype and tumorigenicity ofEwing’s sarcoma cells. Cancer Res 2006;66:9937–47.44. Helman LJ, Meltzer P. Mechanisms of sarcomadevelopment. Nat Rev Cancer 2003;3:685–94.45. Kato K, Hida Y, Miyamoto M, et al. Overexpression of
caveolin-1 in esophageal squamous cell carcinomacorrelates with lymph node metastasis and pathologicstage. Cancer 2002;94:929–33.46. Ho CC, Huang PH, Huang HY, Chen YH, Yang PC, HsuSM. Up-regulated caveolin-1 accentuates the metastasiscapability of lung adenocarcinoma by inducing filopodiaformation. Am J Pathol 2002;161:1647–56.47. Sotgia F, Rui H, Bonuccelli G, Mercier I, Pestell RG,Lisanti MP. Caveolin-1, mammary stem cells, andestrogen-dependent breast cancers. Cancer Res 2006;66:10647–51.48. Mori Y, Cai K, Cheng Y, et al. A Genome-wide searchidentifies epigenetic silencing of somatostatin, tachyki-nin-1, and 5 other genes in colon cancer. Gastroenter-ology 2006;131:797–808.49. Hurlstone AF, Reid G, Reeves JR, et al. Analysis of theCAVEOLIN-1 gene at human chromosome 7q31.1 inprimary tumours and tumour-derived cell lines. Onco-gene 1999;18:1881–90.50. Galbiati F, Volonte D, Engelman JA, et al. Targeteddownregulation of caveolin-1 is sufficient to drive celltransformation and hyperactivate the p42/44 MAPkinase cascade. EMBO J 1998;17:6633–48.51. Liu J, Wang XB, Park DS, Lisanti MP. Caveolin-1expression enhances endothelial capillary tubule forma-tion. J Biol Chem 2002;277:10661–8.52. Volonte D, Liu Y, Galbiati F. The modulation of
caveolin-1 expression controls satellite cell activationduring muscle repair. FASEB J 2005;19:237–9.53. Hamilton D, Mallinger R, Millesi H, Engel A,Baumgartner G, Raderer M. Modulation of CD99/MIC2expression of human AHTO-7 osteoblasts by carcinomacell line-conditioned media. Anticancer Res 2001;21:3909–13.54. Bertaux K, Broux O, Chauveau C, Jeanfils J, DevedjianJC. Identification of CBFA1-regulated genes on SaOs-2cells. J Bone Miner Metab 2005;23:114–22.55. Chong YP, Ia KK, Mulhern TD, Cheng HC. Endogenousand synthetic inhibitors of the Src-family protein tyrosinekinases. Biochim Biophys Acta 2005;1754:210–20.56. Lu Z, Ghosh S, Wang Z, Hunter T. Downregulation ofcaveolin-1 function by EGF leads to the loss of E-cadherin, increased transcriptional activity of h-catenin,and enhanced tumor cell invasion. Cancer Cell 2003;4:499–515.57. Wojcik EJ, Sharifpoor S, Miller NA, et al. A novelactivating function of c-Src and Stat3 on HGFtranscription in mammary carcinoma cells. Oncogene2006;25:2773–84.58. Rahimi N, Hung W, Tremblay E, Saulnier R, Elliott B.c-Src kinase activity is required for hepatocyte growthfactor-induced motility and anchorage-independentgrowth of mammary carcinoma cells. J Biol Chem1998;273:33714–21.
Oncosuppressor Role of Caveolin-1 in Osteosarcoma
www.aacrjournals.org 7685 Cancer Res 2007; 67: (16). August 15, 2007
Research. on March 24, 2016. © 2007 American Association for Cancercancerres.aacrjournals.org Downloaded from

2007;67:7675-7685. Cancer Res Lara Cantiani, Maria Cristina Manara, Cinzia Zucchini, et al. c-Src Activity and Met SignalingCaveolin-1 Reduces Osteosarcoma Metastases by Inhibiting
Updated version
http://cancerres.aacrjournals.org/content/67/16/7675
Access the most recent version of this article at:
Material
Supplementary
http://cancerres.aacrjournals.org/content/suppl/2007/08/16/67.16.7675.DC1.html
Access the most recent supplemental material at:
Cited articles
http://cancerres.aacrjournals.org/content/67/16/7675.full.html#ref-list-1
This article cites 57 articles, 21 of which you can access for free at:
Citing articles
http://cancerres.aacrjournals.org/content/67/16/7675.full.html#related-urls
This article has been cited by 7 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
To request permission to re-use all or part of this article, contact the AACR Publications
Research. on March 24, 2016. © 2007 American Association for Cancercancerres.aacrjournals.org Downloaded from





![[Magnetic resonance and extramural vascular invasion in patients with rectal cancer and liver metastases]](https://static.fdokumen.com/doc/165x107/63379f4e65077fe2dd042c38/magnetic-resonance-and-extramural-vascular-invasion-in-patients-with-rectal-cancer.jpg)














