
Increased susceptibility to amyloid toxicity in familial Alzheimer's fibroblasts

Carcinogenesis vol.17 no.ll pp.2395-2401, 1996
Effect of P-carotene on cell cycle progression of human fibroblasts
Lucia Anna Stivala2, Monica Savio, Ornella Cazzalini,Roberto Pizzala, Laura Rehak, Livia Bianchi,Vanio Vannini and Ennio Prosperi1
Istituto di Patologia Generale 'C. Golgi', Universita di Pavia,Piazza Bona 10, 27100 Pavia and 'Centra di Studio per l'lstochimica,CNR, Piazza Botta 10, 27100 Pavia, Italy2To whom correspondence should be addressed
The uptake of P-carotene (BC) and its effect on the cellcycle progression of normal human fibroblasts in primaryculture were investigated by using two different deliverymethods: exposure to BC solubilized in the organic solventtetrahydrofuran (THF) or to BC incorporated into dipalmi-toylphosphatidylcholine (DPPC) liposomes. Cell cycle pro-gression was evaluated by immunofluorescence detectionand flow cytometric analysis of the proliferating cell nuclearantigen (PCNA). In contrast to THF, which induced amarked reduction in the number of cells in S phase andin the extent of PCNA immunolabeling, DPPC liposomesproved to be an effective delivery system that does notinterfere with cell proliferation. Cellular uptake of 0.23nmol/106 cells was found after 24 h incubation in BC-containing DPPC liposomes. This value increased to 1.2nmol/106 cells after 72 h. After the first day of incubation,the number of cells in S phase was reduced by ~50%, witha consequent accumulation of cells in Gt phase. This effectwas maintained up to 3 days incubation, with no detectableeffects on cell viability. This cell cycle delay was found tobe reversible, returning the percentage of cells in S phaseto the control value 24 h after removal of BC from themedium. In order to determine whether the activity of BCcould be attributed to the molecule itself or to its conversioninto retinoids, the production of BC metabolites wasassessed. Analysis of cellular levels of retinoids failed todemonstrate the presence of retinal, retinol, retinoic acidor retinyl esters during an incubation period of 6 days.These results suggest that in normal human fibroblasts,BC induces a cell cycle delay in the G, phase and that thiseffect is independent of conversion to known retinoids.
Introduction
(J-Carotene (BC*), a carotenoid distributed mostly in greenand yellow vegetables, has been reported to protect cellsagainst cytotoxicity and genotoxicity in both in vitro andin vivo experimental model systems; BC can inhibit celltransformation in vitro and carcinogenesis induced by chemicaland physical agents in animal experimental models (for a
•Abbreviations: BC, P-carotene; THF, tetrahydrofuran; DPPC, dipalmitoyl-phosphatidylcholine; PCNA, proliferating cell nuclear antigen; BrdU, bromo-deoxyuridine; PBS, phosphate buffered saline; LDH, lactate dehydrogenase;DAD, diode array detector, PBT, PBS containing 1% BSA and 0.2%Tween 20; FITC, fluorcsceinisothiocyanate; PI, propidium iodide; MFI, meanfluorescence intensity; DMSO, dimethylsulfoxide; RT, retention time.
© Oxford University Press
review see 1). Moreover, epidemiological studies have sug-gested an association between an increased dietary intake ofcarotenoids and a reduced incidence of cancer (2), even ifrecent results of clinical trials indicated that BC supplementsdo not protect against cancer and might actually increasesmokers' risk of lung tumors (3). Although several anticarcino-genic mechanisms of carotenoids have been proposed, thereare still conflicting opinions and little is known regarding theirmechanisms at the cellular and molecular levels. The protectionafforded by carotenoids can in part be attributed to thequenching of singlet oxygen (4,5). In addition to this mechan-ism, carotenoids has been reported to quench not only singletoxygen, but also to scavenge a variety of free radical species(6). However, BC is considered an 'unusual antioxidant',because its activity depends, in a chemical system, on thepartial pressure of oxygen (7) and, in a cellular system, on itsconcentration (8). Although these observations account forsome of the biological effects of BC, they are not sufficientto explain its antitumor properties. Thus, possible additionalmechanisms have been suggested, such as: conversion tovitamin A, which is a potent modulator of cell differentiationand proliferation (9); an enhancement of immunologicalresponse (10,11); an increase in synthesis of the cytoskeletalproteins (12); an increase in gap junctional communication(13); a decrease in expression of proto-oncogenes (14). Thelast two mechanisms are related to the antiproliferative effectsof BC observed in tumor cell lines (for a review see 12). Onthe other hand, little and contrasting evidence concerning theinhibition of proliferation of normal cells is available (12,15).
BC can be metabolized to retinoids, which are known to bestrong inhibitors of cell proliferation. However, in normaltissues the enzyme 15,15'-dioxygenase, responsible for theconversion of BC to retinoids, has been detected up to nowonly in the small intestine, the liver and corpus luteum (16,17).The ability of the BC molecule itself to inhibit cell growthhas yet to be elucidated.
The aim of this study was to determine the effect of BC oncell cycle progression as such or in relation to the productionof BC metabolites in primary cultures of normal humanfibroblasts.
Because BC is a highly lipophilic compound poorly takenup by most animals (18) and cells in culture (19), twodelivery methods were investigated: (i) exposure of cells toBC solubilized in tetrahydrofuran (THF); (ii) exposure ofcells to BC incorporated into dipalmitoylphosphatydilcholine(DPPC) liposomes. Cellular uptake of BC was determinedby HPLC.
Cell cycle progression was evaluated by immunostaining ofproliferating cell nuclear antigen (PCNA) tightly bound toDNA replication sites (20), which allows the determination ofcells in S phase (21). In addition, the effect of BC on theincorporation of bromodeoxyuridine (BrdU) was assessed byflow cytometry.
2395
by guest on October 14, 2014
http://carcin.oxfordjournals.org/D
ownloaded from

LA^tivala et at
Materials and methods
Chemicals
A stock solution of 5 mM crystalline BC (synthetic aiUtrans, type 1; SigmaCAS no. 7235-40-7) was prepared in THF (BDH). Fresh solutions of BC/THF were prepared on each day of the experiments as needed. The absenceof THF peroxide was monitored by an iodometnc method.
For liposome preparation, 5 mM crystalline BC was dissolved in chloroform(Sigma) and aliquots of 500 nl were used. The liposomes were prepared from20 mg DPPC (Sigma) dissolved in chloroform in the presence or absence ofBC. The lipid mixtures with or without 500 nl BC stock solution were driedwith nitrogen, resuspended in 10 ml phosphate-buffered saline (PBS) andunilamellar vesicles produced by 15 min sonication or, alternatively by filter(1 (im pore size; Nucleopore) extrusion with an extruder device (LipexBiomembranes Inc., Canada). The amount of BC incorporated into liposomeswas determined by extraction of the BC from the liposomes and by subsequentdetection by HPLC. Starting from an initial solution of 250 pM, theconcentration of BC retained by liposomes prepared by sonication was 210|iM (85% of initial concentration), while in the liposomes prepared byextrusion it was 40 uM (16% of initial concentration).
Cell culture and treatments
Human embryonic lung fibroblasts (PEU) were grown in E-MEM medium(Bio-Whittaker) supplemented with 10% fetal calf serum (Gibco, UK), 100IU/ml penicillin, 100 |ig/ml streptomycin and 2.5 ug/ml Fungizone. Cellswere used between the fifth and twentieth passages. The estimated doublingtime for these cells was 30.3 ± 2.5 h. Cell treatment was performed bydiluting BC-containing liposomes 1:10 in culture medium. The mediumcontaining BC was replaced daily in order to avoid formation of oxidationproducts. At 0, 24, 48 and 72 h after initial treatment, the cells were washedtwice with PBS, detached with a standard trypsinization procedure andresuspended in PBS. Cell number was determined by hemocytometer countsand the cell suspension was then divided into aliquots for BC and retinoidderivative determination and for cell cycle analysis. For the determination ofBC metabolites, cell cultures were daily supplemented for a period of 6 days.Cell viability, determined by the release of lactate dehydrogenase (LDH) intothe culture medium (22), was shown to be unaffected by the treatment. Allexperiments were performed at least in triplicate.
Extraction of BC and HPLC determination
After treatment, 1 ml aliquots of cell suspensions (1X 106 cells) were washedthree times with distilled water and subsequently lysed with Triton X-100(10%). BC was extracted twice in ethanol/hexane containing 0.025% butylatedhydroxytoluene as an antioxidant. All solvents (HPLC grade) were providedby BDH (UK). The hexane phase was collected, evaporated under nitrogenand the extract was finally resuspended in HPLC mobile phase (methanol/hexane 85:15) and assayed immediately. Aliquots of 50 ul were injected intothe HPLC system (Hewlett Packard 1090) equipped with a diode arraydetector (DAD).
The analytical procedure to analyze the intracellular concentrations of theBC was an isocratic reverse phase HPLC analysis carried out with a mobilephase flux of 1 ml/min at constant temperature (30°C). The column used forseparation was a Spherisorb ODS 2, 250 mm in length and with a diameterof 4 mm (Hewlett Packard). The DAD was programmed to acquire signals at450 (BC absorption maximum) and 280 nm (tocopherol acetate absorptionmaximum, an internal standard periodically used for extraction and analysisquality control purposes). The entire analytical procedure (treatment, extractionand chromatographic run) was carried out in dim light to avoid carotenoidphoto-autooxidation. For analyte quantitation, a calibration curve was periodic-ally built from pure standard BC and tocopherol acetate (Sigma).
Extraction of BC metabolites and HPLC determination
Pellets containing 1X107 cells were used for quantitation of retinoids. Theextraction method and the analytical procedure to determine the intracellularconcentration of BC metabolites were the same as used for BC, as reportedabove. For better chromatographic resolution of retinoid peaks, a HPLCgradient method modified from the above isocratic one was also used. TheDAD was programmed to acquire signals at 325 (retinol absorption maximum),382 (retinal absorption maximum) and 335 nm (retinoic acid absorptionmaximum). For analyte quantitation, calibration curves were periodically builtfrom pure standard retinol, retinal and retinoic acid (Sigma).
For detection of the presence of retinyl esters, extracts from BC-supple-mented cells, dried with nitrogen, underwent saponification for 6 h in the darkby adding 1 ml of a solution containing 10 mg/ml KOH/CH3OH and 25 ultwice distilled H2O. Cell extracts without BC and extracts with added retinylpalmitate were used as negative and positive controls respectively. Thesaponification procedure proved to be effective since, after 6 h, in the positive
control group no retinyl palmitate was present, while an almost completetrasformation into retinol was detected.
Cell cycle analysisAt the end of incubation, cells were detached using a standard trypsinizationprocedure, counted by hemocytometer, transferred to plastic tubes and washedin PBS. Fixation was performed according to the following protocol (23).Cells were lysed at 4°C for 8 min in a hypotonic solution containing 10 mMTris-HCl, pH 7.4, 2.5 mM MgCl2, 1 mM phenylmethylsulfonylfluoride (BDH)and 0.5% Nonidet P-40 (Sigma). After lysis was complete, cells wereresuspended in saline and cold ethanol was added to a final 70% concentration.Fixed cells were stored at -20°C.
Cells were removed from the fixative, washed in PBS containing 1% BSA(Sigma) and 0.2% Tween 20 (PBT) (Sigma) and incubated for 30 min in100 (il mouse anti-PCNA monoclonal antibody (PC10 clone; Dako, Denmark)diluted 1:100 in PBT. At the end of incubation, cells were washed twice andincubated for 30 min in 100 nl fluoresceinisothiocyanate (hllC)-conjugatedgoat anti-mouse antibody (Sigma) diluted 1:100. After the second antibodyincubation, cells were again washed twice in PBT and resuspended in PBScontaining 5 ug/ml propidium iodide (PI) (Sigma) and 100 |ig/ml RNase A(Sigma). Staining occurred for at least 30 min at room temperature.
Cell cycle distribution was also assessed by determining BrdU incorporationversus DNA content. Cells were incubated with 30 |iM BrdU (Sigma) duringthe last hour of culture, harvested and fixed in cold 70% ethanol. Fixed cellswere washed in PBS, resuspended in 2 N HC1 for 30 min at room temperature,centnfuged at 200 g for 3 min and then resuspended in 0.1 N sodiumtetraborate for 15 min. The samples were then washed in PBS, incubated for15 mm in PBT and for 30 min in 100 |il anti-BrdU monoclonal antibody(Becton Dickinson) diluted 1:10 in PBT After two washes with PBT, cellswere incubated for 30 min in FTTC-conjugated anti-mouse antibody (Sigma)diluted 1:50 in PBT, then washed twice and resuspended in PBS containing5 (ig/ml PI and 100 |ig/ml RNase A.
Flow cytometric measurements
Cells were analyzed with a FACScan (Becton-Dickinson) or with a CoulterXL (Coulter Corp.) flow cytometers. Both instruments were equipped with anargon laser tuned at 488 nm for fluorescence excitation. FTTC fluorescencewas measured with a band-pass filter at 530/30 nm; PI fluorescence wasmeasured with a 585/44 or a 620/30 nm band-pass filter. Ten thousand cellswere measured for each sample. Computer statistical analysis of meanfluorescence intensity (MFT) and graphic representation were performed withLysis II software (Becton-Dickinson) or with XL software (Coulter). Statisticalsignificance analysis was obtained by Student's (-test.
Results
Since BC is poorly water soluble, preliminary studies wereperformed to optimize its solubilization with organic solventswhose concentrations will not interfere with cell cycle progres-sion. Thus, solubilization in dimethylsulfoxide (DMSO),ethanol and THF were assessed. An alternative deliveringsystem with DPPC liposomes was also investigated. To reacha useful concentration of 5 mM in the stock solution, BCcould be solubilized only in THF.
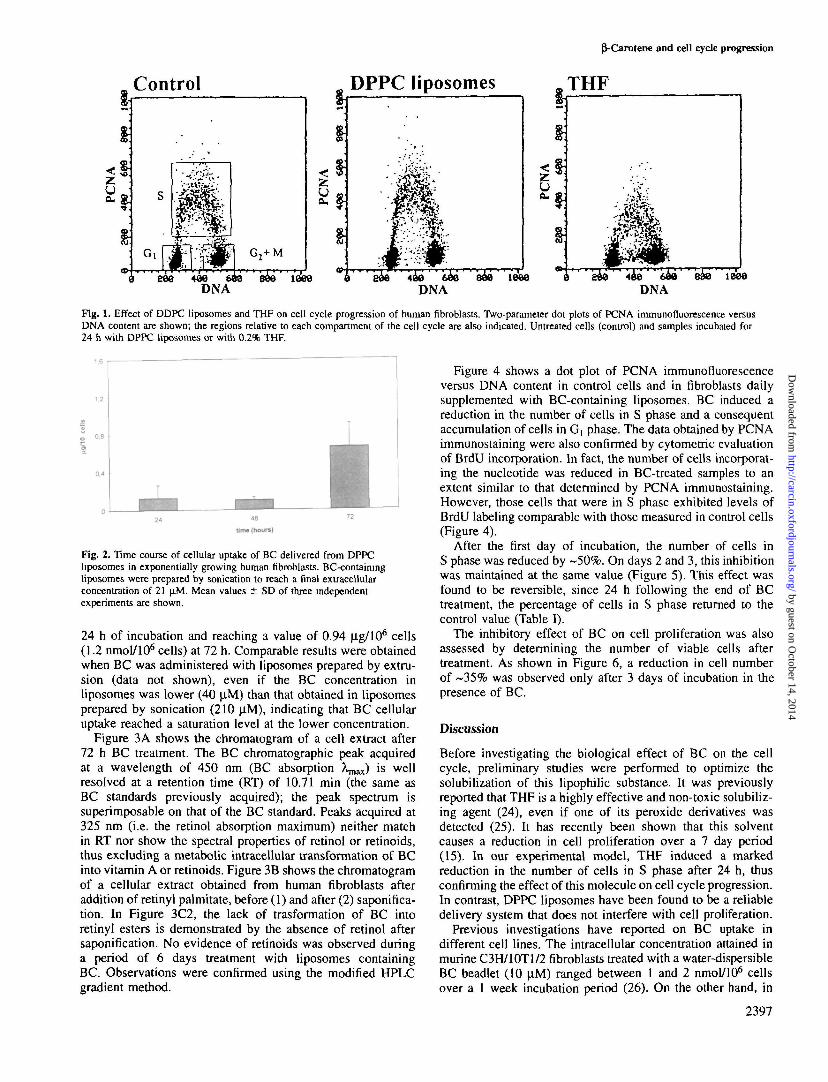
In order to analyze the effect of THF or of DPPC liposomeson the cell cycle, the distribution of PCNA bound to DNAreplication sites was determined. PCNA bound to DNA syn-thesis sites is found only in S phase cells, thus allowingdiscrimination of these cells from those present in the otherphases of the cell cycle, Gj and G2+M (Figure 1, control).
THF was consistently found to remarkably influence cellcycle progression. In fact, this solvent strongly reduced thepercentage of cells in S phase even at the minimal concentration(0.2%) necessary to dissolve BC. In contrast, DPPC liposomesalone were not found to affect cell proliferation (Figure 1).Therefore, subsequent experiments were all performed bydelivering BC dissolved in DPPC liposomes to reach a finalextracellular concentration of 21 (J.M. Figure 2 shows thecellular uptake of BC delivered from liposomes prepared bysonication, after treatment for 24, 48 and 72 h. BC is takenup by exponentially growing human fibroblasts, achievingcellular levels of 0.12 ng/106 cells (0.23 nmol/106 cells) after
2396
by guest on October 14, 2014
http://carcin.oxfordjournals.org/D
ownloaded from
Control DPPC liposomes
pVCarotene and cell cycle progression
THF
436 6*3DNA
1386DNA DNA
Fig. 1. Effect of DDPC liposomes and THF on cell cycle progression of human fibroblasts. Two-parameter dot plots of PCNA immunofluorescence versusDNA content are shown; the regions relative to each compartment of the cell cycle are also indicated. Untreated cells (control) and samples incubated for24 h with DPPC liposomes or with 0.2% THF.
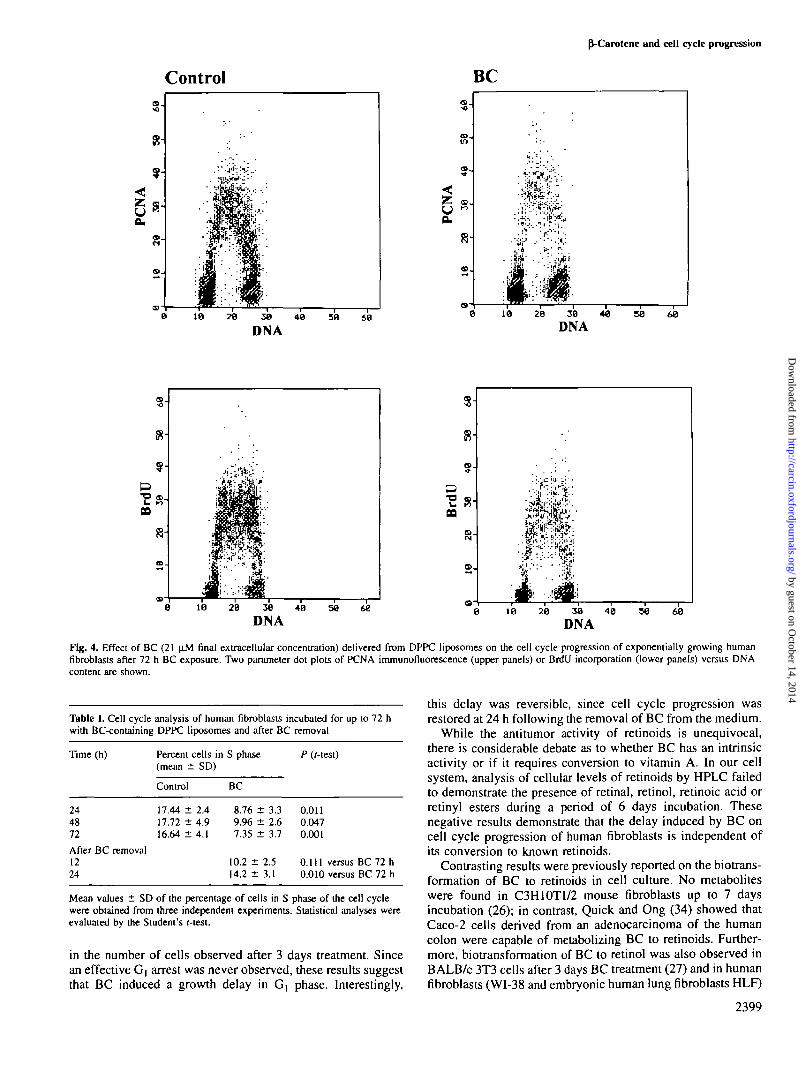
Figure 4 shows a dot plot of PCNA immunofluorescenceversus DNA content in control cells and in fibroblasts dailysupplemented with BC-containing liposomes. BC induced areduction in the number of cells in S phase and a consequentaccumulation of cells in G) phase. The data obtained by PCNAimmunostaining were also confirmed by cytometric evaluationof BrdU incorporation. In fact, the number of cells incorporat-ing the nucleotide was reduced in BC-treated samples to anextent similar to that determined by PCNA immunostaining.However, those cells that were in S phase exhibited levels ofBrdU labeling comparable with those measured in control cells(Figure 4).
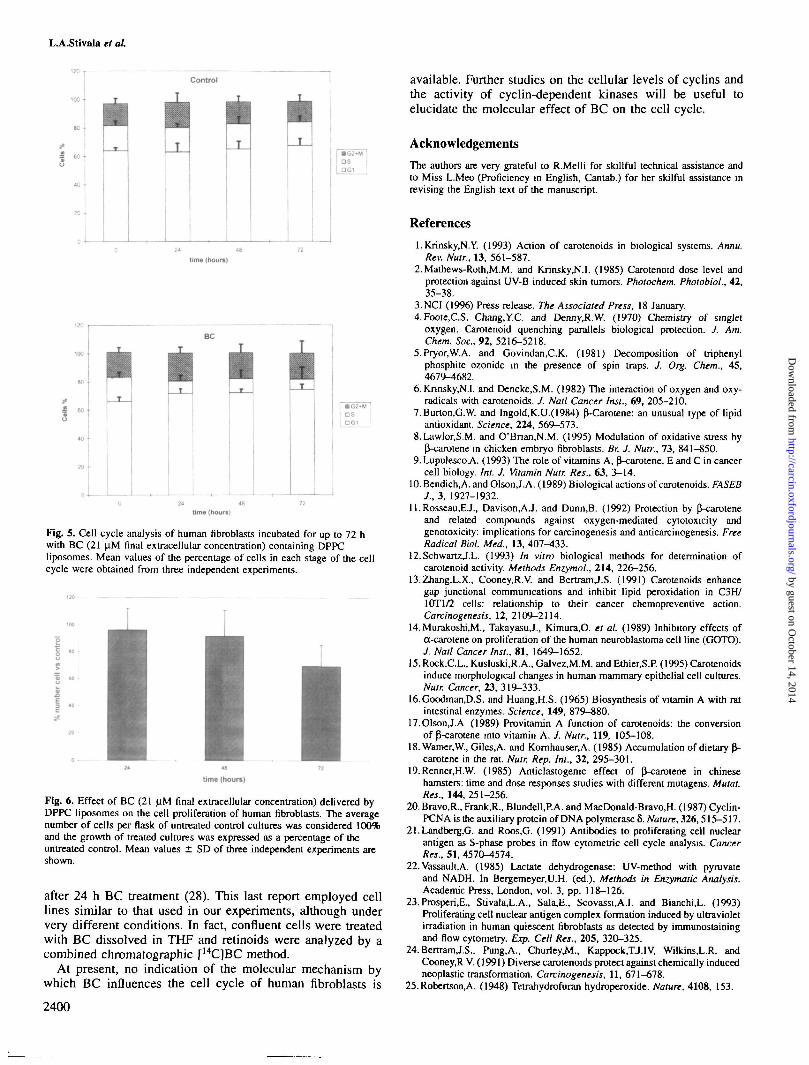
After the first day of incubation, the number of cells inS phase was reduced by ~50%. On days 2 and 3, this inhibitionwas maintained at the same value (Figure 5). This effect wasfound to be reversible, since 24 h following the end of BCtreatment, the percentage of cells in S phase returned to thecontrol value (Table I).
The inhibitory effect of BC on cell proliferation was alsoassessed by determining the number of viable cells aftertreatment. As shown in Figure 6, a reduction in cell numberof ~35% was observed only after 3 days of incubation in thepresence of BC.
Discussion
Before investigating the biological effect of BC on the cellcycle, preliminary studies were performed to optimize thesolubilization of this lipophilic substance. It was previouslyreported that THF is a highly effective and non-toxic solubiliz-ing agent (24), even if one of its peroxide derivatives wasdetected (25). It has recently been shown that this solventcauses a reduction in cell proliferation over a 7 day period(15). In our experimental model, THF induced a markedreduction in the number of cells in S phase after 24 h, thusconfirming the effect of this molecule on cell cycle progression.In contrast, DPPC liposomes have been found to be a reliabledelivery system that does not interfere with cell proliferation.
Previous investigations have reported on BC uptake indifferent cell lines. The intracellular concentration attained inmurine C3H/10T1/2 fibroblasts treated with a water-dispersibleBC beadlet (10 \iM) ranged between 1 and 2 nmol/106 cellsover a 1 week incubation period (26). On the other hand, in
2397
Fig. 2. Time course of cellular uptake of BC delivered from DPPCliposomes in exponentially growing human fibroblasts. BC-containingliposomes were prepared by sonication to reach a final extracellularconcentration of 21 |iM. Mean values ± SD of three independentexperiments are shown.
24 h of incubation and reaching a value of 0.94 (J.g/106 cells(1.2 nmol/106 cells) at 72 h. Comparable results were obtainedwhen BC was administered with liposomes prepared by extru-sion (data not shown), even if the BC concentration inliposomes was lower (40 |iM) than that obtained in liposomesprepared by sonication (210 ^M), indicating that BC cellularuptake reached a saturation level at the lower concentration.
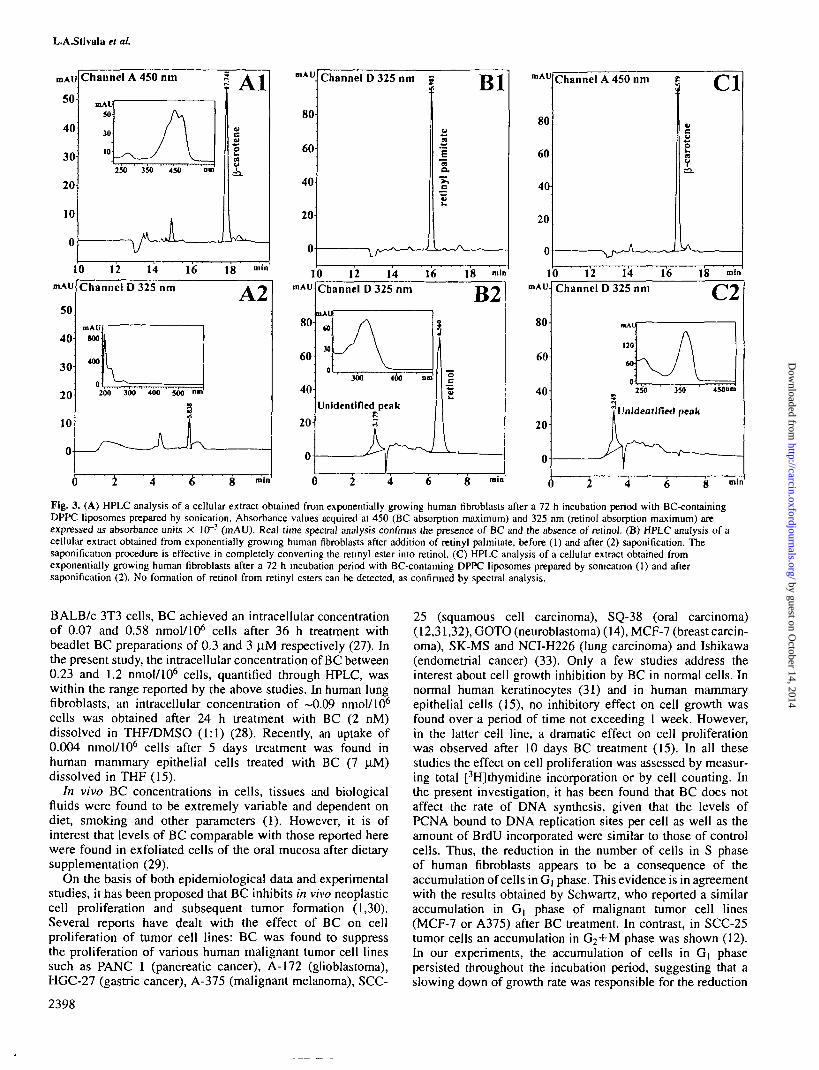
Figure 3A shows the chromatogram of a cell extract after72 h BC treatment. The BC chromatographic peak acquiredat a wavelength of 450 nm (BC absorption A,^) is wellresolved at a retention time (RT) of 10.71 min (the same asBC standards previously acquired); the peak spectrum issuperimposable on that of the BC standard. Peaks acquired at325 nm (i.e. the retinol absorption maximum) neither matchin RT nor show the spectral properties of retinol or retinoids,thus excluding a metabolic intracellular transformation of BCinto vitamin A or retinoids. Figure 3B shows the chromatogramof a cellular extract obtained from human fibroblasts afteraddition of retinyl palmitate, before (1) and after (2) saponifica-tion. In Figure 3C2, the lack of trasformation of BC intoretinyl esters is demonstrated by the absence of retinol aftersaponification. No evidence of retinoids was observed duringa period of 6 days treatment with liposomes containingBC. Observations were confirmed using the modified HPLCgradient method.
by guest on October 14, 2014
http://carcin.oxfordjournals.org/D
ownloaded from
L.A.Stivala el al
1mAU
50
40
30
20
10
0
(
0 12
Channel
mAU800
400
o200
/ " ^
2
14
D325nm
300 400 500
4
16
L6
18
8
m(n
A2
mln
mAU
80
60
40
20
0
Channel D 325 nm j
\_A~^—^-^-
aIaa .
">»Q
ret!
_-> / \
B l
«—̂ -- _ —10
mAU
8fr
60
40
20
u
C
12Channel
nAU
M
JO
/_ /
JOO
14D325nm
AV" 4 0 0 " '
Unidentified peak
kr-7,
i4
16 18 mln
nm
6
B2
etln
o
8 rain
mAU
10 12mAU.
80
60
40
20
0
(
Channel
2
14D325nm
mAL
no
60 A
16
250 3
e?Unidentlfied
J>f~4 6
18
' \
peak
f——
8
rofn
C2
450om
mill
Fig. 3. (A) HPLC analysis of a cellular extract obtained from exponentially growing human fibroblasts after a 72 h incubation period with BC-containingDPPC liposomes prepared by sonication. Absorbance values acquired at 450 (BC absorption maximum) and 325 nm (retinol absorption maximum) areexpressed as absorbance units X 10~3 (mAU). Real time spectral analysis confirms the presence of BC and the absence of retinol. (B) HPLC analysis of acellular extract obtained from exponentially growing human fibroblasts after addition of retinyl palmitate, before (1) and after (2) saponification. Thesaponification procedure is effective in completely converting the reunyl ester into retinol. (C) HPLC analysis of a cellular extract obtained fromexponentially growing human fibroblasts after a 72 h incubation period with BC-containing DPPC liposomes prepared by sonication (1) and aftersaponification (2). No formation of retinol from retinyl esters can be detected, as confirmed by spectral analysis.
BALB/c 3T3 cells, BC achieved an intracellular concentrationof 0.07 and 0.58 nmol/106 cells after 36 h treatment withbeadlet BC preparations of 0.3 and 3 ^M respectively (27). Inthe present study, the intracellular concentration of BC between0.23 and 1.2 nmol/106 cells, quantified through HPLC, waswithin the range reported by the above studies. In human lungfibroblasts, an intracellular concentration of ~0.09 nmol/106
cells was obtained after 24 h treatment with BC (2 nM)dissolved in THF/DMSO (1:1) (28). Recently, an uptake of0.004 nmol/106 cells after 5 days treatment was found inhuman mammary epithelial cells treated with BC (7 (iM)dissolved in THF (15).
In vivo BC concentrations in cells, tissues and biologicalfluids were found to be extremely variable and dependent ondiet, smoking and other parameters (1). However, it is ofinterest that levels of BC comparable with those reported herewere found in exfoliated cells of the oral mucosa after dietarysupplementation (29).
On the basis of both epidemiological data and experimentalstudies, it has been proposed that BC inhibits in vivo neoplasticcell proliferation and subsequent tumor formation (1,30).Several reports have dealt with the effect of BC on cellproliferation of tumor cell lines: BC was found to suppressthe proliferation of various human malignant tumor cell linessuch as PANC 1 (pancreatic cancer), A-172 (glioblastoma),HGC-27 (gastric cancer), A-375 (malignant melanoma), SCC-
2398
25 (squamous cell carcinoma), SQ-38 (oral carcinoma)(12,31,32), GOTO (neuroblastoma) (14), MCF-7 (breast carcin-oma), SK-MS and NCI-H226 (lung carcinoma) and Ishikawa(endometrial cancer) (33). Only a few studies address theinterest about cell growth inhibition by BC in normal cells. Innormal human keratinocytes (31) and in human mammaryepithelial cells (15), no inhibitory effect on cell growth wasfound over a period of time not exceeding 1 week. However,in the latter cell line, a dramatic effect on cell proliferationwas observed after 10 days BC treatment (15). In all thesestudies the effect on cell proliferation was assessed by measur-ing total [3H]thymidine incorporation or by cell counting. Inthe present investigation, it has been found that BC does notaffect the rate of DNA synthesis, given that the levels ofPCNA bound to DNA replication sites per cell as well as theamount of BrdU incorporated were similar to those of controlcells. Thus, the reduction in the number of cells in S phaseof human fibroblasts appears to be a consequence of theaccumulation of cells in G] phase. This evidence is in agreementwith the results obtained by Schwartz, who reported a similaraccumulation in G| phase of malignant tumor cell lines(MCF-7 or A375) after BC treatment. In contrast, in SCC-25tumor cells an accumulation in G2+M phase was shown (12).In our experiments, the accumulation of cells in Gj phasepersisted throughout the incubation period, suggesting that aslowing down of growth rate was responsible for the reduction
by guest on October 14, 2014
http://carcin.oxfordjournals.org/D
ownloaded from
P-Carotene and cell cycle progression
Control BC
0 18 28 38 40 39 60
'•&*:^%
28 38 40
DNA
10 29 38 48 50 68
Fig. 4. Effect of BC (21 pM final extracellular concentration) delivered from DPPC liposomes on the cell cycle progression of exponentially growing humanfibroblasts after 72 h BC exposure. Two parameter dot plots of PCNA immunofluorescence (upper panels) or BrdU incorporation (lower panels) versus DNAcontent are shown.
Table I. Cell cycle analysis of human fibroblasts incubated for up to 72 hwith BC-containing DPPC liposomes and after BC removal
Time (h) Percent cells in S phase P ((-test)(mean ± SD)
244872
After BC removal1224
Control
17.44 ±17.72 ±16.64 ±
2.44.94.1
BC
8.769.967.35
± 3.3± 2.6± 3.7
10.2 ± 2.514.2 ± 3.1
0.0110.0470.001
0.111 versus0.010 versus
BCBC
7272
hh
Mean values ± SD of the percentage of cells in S phase of the cell cyclewere obtained from three independent experiments. Statistical analyses wereevaluated by the Student's r-test.
in the number of cells observed after 3 days treatment. Sincean effective G, arrest was never observed, these results suggestthat BC induced a growth delay in G, phase. Interestingly,
this delay was reversible, since cell cycle progression wasrestored at 24 h following the removal of BC from the medium.
While the antitumor activity of retinoids is unequivocal,there is considerable debate as to whether BC has an intrinsicactivity or if it requires conversion to vitamin A. In our cellsystem, analysis of cellular levels of retinoids by HPLC failedto demonstrate the presence of retinal, retinol, retinoic acid orretinyl esters during a period of 6 days incubation. Thesenegative results demonstrate that the delay induced by BC oncell cycle progression of human fibroblasts is independent ofits conversion to known retinoids.
Contrasting results were previously reported on the biotrans-formation of BC to retinoids in cell culture. No metaboliteswere found in C3H10T1/2 mouse fibroblasts up to 7 daysincubation (26); in contrast, Quick and Ong (34) showed thatCaco-2 cells derived from an adenocarcinoma of the humancolon were capable of metabolizing BC to retinoids. Further-more, biotransformation of BC to retinol was also observed inBALB/c 3T3 cells after 3 days BC treatment (27) and in humanfibroblasts (WI-38 and embryonic human lung fibroblasts HLF)
2399
by guest on October 14, 2014
http://carcin.oxfordjournals.org/D
ownloaded from
L.A.Stivala et al
fl fi Btime {houre)
Fig. 5. Cell cycle analysis of human fibroblasts incubated for up to 72 hwith BC (21 |iM final extracellular concentration) containing DPPCliposomes. Mean values of the percentage of cells in each stage of the cellcycle were obtained from three independent experiments.
1 -•
time (hours)
Fig. 6. Effect of BC (21 |iM final extracellular concentration) delivered byDPPC liposomes on the cell proliferation of human fibroblasts. The averagenumber of cells per flask of untreated control cultures was considered 100%and the growth of treated cultures was expressed as a percentage of theuntreated control. Mean values ± SD of three independent experiments areshown.
after 24 h BC treatment (28). This last report employed celllines similar to that used in our experiments, although undervery different conditions. In fact, confluent cells were treatedwith BC dissolved in THF and retinoids were analyzed by acombined chromatographic [14C]BC method.
At present, no indication of the molecular mechanism bywhich BC influences the cell cycle of human fibroblasts is
2400
available. Further studies on the cellular levels of cyclins andthe activity of cyclin-dependent kinases will be useful toelucidate the molecular effect of BC on the cell cycle.
Acknowledgements
The authors are very grateful to R.Melli for skillful technical assistance andto Miss L.Meo (Proficiency in English, Cantab.) for her skilful assistance inrevising the English text of the manuscript.
References
l.Krinsky.N.Y. (1993) Action of carotenoids in biological systems. Annu.Rev. Nutr., 13, 561-587.
2.Mathews-Roth,M.M. and KrinskyJM.I. (1985) Carotenoid dose level andprotection against UV-B induced skin tumors. Photochem. Photobiol., 42,35-38.
3.NCI (1996) Press release. The Associated Press, 18 January.4.Foote,C.S, Chang.Y.C. and Denny.R.W. (1970) Chemistry of singlet
oxygen. Carotenoid quenching parallels biological protection. J. Am.Chem. Soc., 92, 5216-5218.
5.Pryor,W.A. and Govindan.C.K. (1981) Decomposition of triphenylphosphite ozonide in the presence of spin traps. J. Org. Chem., 45,4679-4682.
6. Knnsky,N.I. and Deneke,S.M. (1982) The interaction of oxygen and oxy-radicals with carotenoids. J. Natl Cancer Inst., 69, 205-210.
7. Burton.G.W. and Ingold,K.U.(1984) f}-Carotene: an unusual type of lipidantioxidant. Science, 224, 569-573.
8. Lawlor.S.M. and O'Bnan,N.M. (1995) Modulation of oxidative stress by^-carotene in chicken embryo fibroblasts. Br. J. Nutr., 73, 841-850.
9. LupulescoA (1993) The role of vitamins A, p^-carotene, E and C in cancercell biology. Int. J. Vitamin Nutr. Res., 63, 3-14.
10. Bendich.A. and Olson J.A. (1989) Biological actions of carotenoids. FASEBJ., 3, 1927-1932.
ll.Rosseau,EJ., Davison.AJ. and Dunn.B. (1992) Protection by ^-caroteneand related compounds against oxygen-mediated cytotoxicity andgenotoxicity: implications for carcinogenesis and anticarcinogenesis. FreeRadical Biol. Med., 13, 407^*33.
12.SchwartzJ.L. (1993) In vitro biological methods for determination ofcarotenoid activity. Methods Enzymol., 214, 226-256.
13.Zhang,L.X., Cooney.R.V. and BertramJ.S. (1991) Carotenoids enhancegap junctional communications and inhibit lipid peroxidation in C3H/10T1/2 cells: relationship to their cancer chemopreventive action.Carcinogenesis, 12, 2109-2114.
14.Murakoshi,M., TakayasuJ., Kimura.O. el al. (1989) Inhibitory effects ofa-carotene on proliferation of the human neuroblastoma cell line (GOTO).J. Natl Cancer Inst., 81, 1649-1652.
15. Rock.C.L., Kusluski,R.A., Galvez.M.M. and Ethier.S.P. (1995) Carotenoidsinduce morphological changes in human mammary epithelial cell cultures.Nutr. Cancer, 23, 319-333.
16.GoodmanJ).S. and Huang.H.S. (1965) Biosynthesis of vitamin A with ratintestinal enzymes. Science, 149, 879-880.
17. OlsonJ.A (1989) Provitamin A function of carotenoids: the conversionof p-carotene into vitamin A. J. Nutr., 119, 105-108.
18. Warner,W., Giles,A. and Komhauser.A. (1985) Accumulation of dietary (1-carotene in the rat. Nutr. Rep. Int., 32, 295-301.
19. Renner.H.W. (1985) Anticlastogenic effect of P-carotene in Chinesehamsters: time and dose responses studies with different mutagens. Mutat.Res., 144,251-256.
20. Bravo.R., Frank,R., Blundell,P.A. and MacDonald-Bravo.H. (1987) Cyclin-PCNA is the auxiliary protein of DNA polymerase 6. Nature, 326,515-517.
21.Landberg,G. and Roos.G. (1991) Antibodies to proliferating cell nuclearantigen as S-phase probes in flow cytometric cell cycle analysis. CancerRes., 51, 4570-4574.
22. Vassault,A. (1985) Lactate dehydrogenase: UV-method with pyruvateand NADH. In Bergemeyer,U.H. (ed.), Methods in Enzymatic Analysis.Academic Press, London, vol. 3, pp. 118-126.
23. Prosperi.E, Stivala,L.A., Sala,E., Scovassi,A.I. and Bianchi,L. (1993)Proliferating cell nuclear antigen complex formation induced by ultravioletirradiation in human quiescent fibroblasts as detected by immunostainingand flow cytometry. Exp. Cell Res., 205, 320-325.
24. BertranUS., Pung.A., Churley,M., Kappock.TJ.IV, Wilkins,L.R. andCooney,R V. (1991) Diverse carotenoids protect against chemically inducedneoplastic transformation. Carcinogenesis, 11, 671-678.
25. Robertson^. (1948) Tetrahydrofuran hydroperoxide. Nature, 4108, 153.
by guest on October 14, 2014
http://carcin.oxfordjournals.org/D
ownloaded from

^-Carotene and cell cycle progression
26. RundhaugJ.E., Pung,A., Read.C.M. and BertramJ.S. (1988) Uptake andmetabolism of p^-carotene and retinal by C3H/10T1/2 cells. Carcinogenesis,9, 1541-1545.
27. Warner,W.G., Wei.R.R., MatusikJ.E., Komhauser.A. and Dunkel.V.C.(1993) fj-Carotene uptake, metabolism, and distribution in BALB/c 3T3cells. Nutr. Cancer, 19, 31^tl.
28.Scita,G., Aponte.G.W. and Wolf.G. (1993) Uptake and cleavage of £-carotene by cultures of rat small intestinal cells and human lung fibroblasts.Methods Enzymol., 214, 21-32.
29.Stich,H.F., Homby.A.P, and Dunn.BP. (1986) Beta-carotene levels inexfoliated human mucosa cells following its oral administration. CancerLett., 30, 133-141.
30. Ziegler.R.G. (1989) A review of epidemiologic evidence that carotenoidsreduce the risk of cancer. J. Nutr., 119, 116-122.
31.SchwartzJ., Singh.R.P., Teicher,B., WrighU-E., Trites.D.H. and Shklar,G.(1990) Induction of a 70 kDa protein associated with the selectivecytotoxicity of fi-carotene in human epidermal carcinoma. Biochem.Biophys. Res. Commun., 169, 941-946.
32. Schwartz J., TanakaJ., Klandekar.V., Herman,T.S. and Teicher.B. (1992)P-Carotene and/or vitamin E as modulator of alkylating agents in SCC-25 human squamous carcinoma cells. Cancer Chemother. Pharmacol., 29,207-213.
33.LevyJ., Bosin,E., Feldman.B., Giat,Y., Miinster^A., Danilenko.M. andSharoni.Y. (1995) Lycopene is a more potent inhibitor of human cancercell proliferation than either oe-carotene or P-carotene. Nutr. Cancer, 24(3), 257-266.
34.Quick,T.C. and Ong,D.E. (1990) Vitamin A metabolism in the humanintestinal Caco-2 cell line. Biochemistry, 29, 11116-11123.
Received on May 15, 1996; revised on July 15, 1996; accepted on July 26, 1996
2401
by guest on October 14, 2014
http://carcin.oxfordjournals.org/D
ownloaded from

by guest on October 14, 2014
http://carcin.oxfordjournals.org/D
ownloaded from
Copyright © 2022 FDOKUMEN




















