
Angiopoietin 1 Promotes Tumor Angiogenesis and Tumor Vessel Plasticity of Human Cervical Cancer in...
11
Angiopoietin 1 Promotes Tumor Angiogenesis and Tumor Vessel Plasticity of Human Cervical Cancer in Mice 1 Winston S. N. Shim,* Ming Teh,† Akanksha Bapna,* Injune Kim,‡ Gou-Young Koh,‡ Peter O. P. Mack,§ and Ruowen Ge* ,2 *Molecular Angiogenesis Laboratory, Department of Biological Sciences, National University of Singapore, Singapore 119260; †Department of Pathology, National University of Singapore, Singapore 119074; §Department of Surgery, Singapore General Hospital, Singapore 169608; and ‡National Creative Research Initiatives Center for Endothelial Cells, Department of Life Science, Pohang University of Science and Technology, San 31, Hyoja-Dong, Nam-Gu, Pohang, 790-784, Korea Angiopoietins have been increasingly implicated to play important roles in blood vessel formation, remod- eling, maturation, and maintenance. However, their roles in tumor angiogenesis and hence tumor growth and metastasis still remain uncertain. In this work, angiopoietin 1 expression was amplified in human cer- vical cancer HeLa cells by stable transfection or re- combinant human adenovirus-mediated gene trans- fer. We show that increased angiopoietin 1 expression promoted in vivo growth of human cervical cancers in mice by promoting tumor angiogenesis and inhibiting tumor cell apoptosis. Furthermore, we also show for the first time that overexpression of angiopoietin 1 also leads to increased tumor vessel plasticity with a large number of vessels lacking periendothelial sup- porting cells. These results indicate that angiopoietin 1 promotes tumor angiogenesis and tumor vessel plas- ticity of human cervical cancer in mice. © 2002 Elsevier Science (USA) Key Words: tumor angiogenesis; angiopoietin 1; ves- sel plasticity. INTRODUCTION Tumor nodules cannot derive nutrients through dif- fusion when greater than 1–2 mm 3 and additional growth is dependent on neovascularization or angio- genesis [1]. Angiogenesis involves sprouting and re- modeling of preexisting blood vessels into a more com- plex network of vasculature through continuous recruiting and pruning of blood vessels. It is an intri- cate process that involves periendothelial cell rear- rangement and matrix degradation with accompanying proliferation, migration, differentiation, and anastomosis of the endothelium [2]. The process is regulated by pro- and antiangiogenic factors which include secreted growth factors from the developing malignant as well as stromal cells. Vascular endothelial growth factor (VEGF) is one such factor, and its involvement in tu- mor angiogenesis has been well demonstrated [3– 6]. Recently, endothelium-specific Tie-2 receptor, through preferential activation, by Ang-1 has been found to play important roles in vascular sprouting and remod- eling as well as endothelial survival, maintenance, and maturation [7–10]. Ang-1 / or Tie-2 / mice died in utero with severe vascular defects that manifested as lack of critical remodeling and absence of close inter- actions between endothelial and periendothelial cells [8]. Recruitment of periendothelial cells such as peri- cytes and smooth muscle cells to the maturing nas- cent vessels is believed to be mediated by endothelial cells (ECs) secreting platelet-derived growth factor B (PDGF-B) and heparin-binding epidermal growth fac- tor (HB-EGF), and leads to the cessation of endothelial cell proliferation and migration as well as the initiation of matrix deposition for the completion of angiogenesis [11, 12]. Recent evidence suggests that the interactions be- tween angiopoietins and VEGF are crucial for physio- logical and pathological angiogenesis [13–17]. Coordi- nated upregulation of Ang-2, a natural antagonist of Tie-2, in cooperation with VEGF has been linked to robust angiogenesis, while in the absence of VEGF and/or Ang-1, Ang-2 has been suggested to induce en- dothelial cell apoptosis and vessel regression [18 –20]. We have previously shown that inhibition of Ang-1 expression in human cervical carcinoma HeLa cells greatly reduces xenografted tumor growth in mice as a result of decreased tumor angiogenesis and increased 1 This work is supported by a Singapore National Medical Re- search Council grant to P.O.P.M. and R.G. (NMRC/004/1994) and a National University of Singapore research grant to R.G. and M.T. (R-154-000-099-112). W.S.N.S. was supported by a National Univer- sity of Singapore graduate scholarship. 2 To whom reprint requests should be addressed at Department of Biological Sciences, National University of Singapore, Block S1A, 05-05, 10 Science Drive 4, Singapore 117543. Fax: 65-6779-2486. E-mail: [email protected]. Experimental Cell Research 279, 299 –309 (2002) doi:10.1006/excr.2002.5597 0014-4827/02 $35.00 299 © 2002 Elsevier Science (USA) All rights reserved.
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Angiopoietin 1 Promotes Tumor Angiogenesis and Tumor Vessel Plasticity of Human Cervical Cancer in...

Experimental Cell Research 279, 299–309 (2002)doi:10.1006/excr.2002.5597
Angiopoietin 1 Promotes Tumor Angiogenesis and Tumor VesselPlasticity of Human Cervical Cancer in Mice1
Winston S. N. Shim,* Ming Teh,† Akanksha Bapna,* Injune Kim,‡ Gou-Young Koh,‡Peter O. P. Mack,§ and Ruowen Ge*,2
*Molecular Angiogenesis Laboratory, Department of Biological Sciences, National University of Singapore, Singapore 119260;†Department of Pathology, National University of Singapore, Singapore 119074; §Department of Surgery, Singapore General
Hospital, Singapore 169608; and ‡National Creative Research Initiatives Center for Endothelial Cells, Department of Life
Angiopoietins have been increasingly implicated toplay important roles in blood vessel formation, remod-eling, maturation, and maintenance. However, theirroles in tumor angiogenesis and hence tumor growthand metastasis still remain uncertain. In this work,angiopoietin 1 expression was amplified in human cer-vical cancer HeLa cells by stable transfection or re-combinant human adenovirus-mediated gene trans-fer. We show that increased angiopoietin 1 expressionpromoted in vivo growth of human cervical cancers inmice by promoting tumor angiogenesis and inhibitingtumor cell apoptosis. Furthermore, we also show forthe first time that overexpression of angiopoietin 1also leads to increased tumor vessel plasticity with alarge number of vessels lacking periendothelial sup-porting cells. These results indicate that angiopoietin1 promotes tumor angiogenesis and tumor vessel plas-ticity of human cervical cancer in mice. © 2002 Elsevier
Science (USA)
Key Words: tumor angiogenesis; angiopoietin 1; ves-sel plasticity.
INTRODUCTION
Tumor nodules cannot derive nutrients through dif-fusion when greater than 1–2 mm3 and additionalgrowth is dependent on neovascularization or angio-genesis [1]. Angiogenesis involves sprouting and re-modeling of preexisting blood vessels into a more com-plex network of vasculature through continuous
1 This work is supported by a Singapore National Medical Re-search Council grant to P.O.P.M. and R.G. (NMRC/004/1994) and aNational University of Singapore research grant to R.G. and M.T.(R-154-000-099-112). W.S.N.S. was supported by a National Univer-sity of Singapore graduate scholarship.
2 To whom reprint requests should be addressed at Department ofBiological Sciences, National University of Singapore, Block S1A,05-05, 10 Science Drive 4, Singapore 117543. Fax: 65-6779-2486.
E-mail: [email protected].299
recruiting and pruning of blood vessels. It is an intri-cate process that involves periendothelial cell rear-rangement and matrix degradation with accompanyingproliferation, migration, differentiation, and anastomosisof the endothelium [2]. The process is regulated bypro- and antiangiogenic factors which include secretedgrowth factors from the developing malignant as wellas stromal cells. Vascular endothelial growth factor(VEGF) is one such factor, and its involvement in tu-mor angiogenesis has been well demonstrated [3–6].
Recently, endothelium-specific Tie-2 receptor, throughpreferential activation, by Ang-1 has been found toplay important roles in vascular sprouting and remod-eling as well as endothelial survival, maintenance, andmaturation [7–10]. Ang-1�/� or Tie-2�/� mice died inutero with severe vascular defects that manifested aslack of critical remodeling and absence of close inter-actions between endothelial and periendothelial cells[8]. Recruitment of periendothelial cells such as peri-cytes and smooth muscle cells to the maturing nas-cent vessels is believed to be mediated by endothelialcells (ECs) secreting platelet-derived growth factor B(PDGF-B) and heparin-binding epidermal growth fac-tor (HB-EGF), and leads to the cessation of endothelialcell proliferation and migration as well as the initiationof matrix deposition for the completion of angiogenesis[11, 12].
Recent evidence suggests that the interactions be-tween angiopoietins and VEGF are crucial for physio-logical and pathological angiogenesis [13–17]. Coordi-nated upregulation of Ang-2, a natural antagonist ofTie-2, in cooperation with VEGF has been linked torobust angiogenesis, while in the absence of VEGFand/or Ang-1, Ang-2 has been suggested to induce en-dothelial cell apoptosis and vessel regression [18–20].
We have previously shown that inhibition of Ang-1expression in human cervical carcinoma HeLa cellsgreatly reduces xenografted tumor growth in mice as a
Science, Pohang University of Science and Technology
n 31, Hyoja-Dong, Nam-Gu, Pohang, 790-784, Korea , Saresult of decreased tumor angiogenesis and increased
0014-4827/02 $35.00© 2002 Elsevier Science (USA)
All rights reserved.

tumor cell apoptosis [21]. In this study, the role ofAng-1/Tie-2 system in cervical cancer angiogenesis andtumor behavior was examined by overexpression ofAng-1 in stably transfected HeLa cell clones or throughadenovirus-mediated gene transfer. We show that in-creased Ang-1 expression promotes the in vivo growthof human cervical cancers by promoting tumor angio-genesis and increasing tumor vessel plasticity.
MATERIALS AND METHODS
Cell lines. The HeLa human cervical adenocarcinoma and 293human embryonic kidney cell lines were obtained from the AmericanType Culture Collection (Rockville, MD). HeLa and 293 cells werecultured in Dulbecco’s minimal essential medium (DMEM) andMEM, respectively, with 10% fetal bovine serum, 100 �g/ml strep-tomycin sulfate, and 100 IU/ml penicillin sulfate (Life Technologies,Gaithersburg, MD) at 37°C in 5% humidified CO2 and 95% air.
Construction of Ang-1 expression construct and establishment ofstable cell lines by transfection. Full-length Ang-1 cDNAs was iso-lated by polymerase chain reaction (PCR) from human heart cDNAusing the Advantage cDNA cloning kit (Clontech, Palo Alto, CA) andcloned into pGemT-Easy vector (Promega, Madison, WI), verified byrestriction digestion analyses and DNA sequencing. It was thensubcloned into HindIII/ClaI sites of pLNCX vector (Clontech) usingPfu DNA polymerase (Stratagene, La Jolla, CA) with primer pair5� CCCAAGCTTGGGTCAGAAGAAAGGAGCAAG 3�, where theunderlined portion represents HindIII restriction site, and 5�CCATCGATGGCTGTCACCCCAAGTAGAG 3�, where the under-lined portion represents ClaI site.
Vectors containing Ang-1 or vector alone (pLNCX) were trans-fected into HeLa cells using the calcium phosphate method as de-scribed by the manufacturer (Clontech). Selective medium contain-ing 600 �g/ml G418 (Life Technologies) was added 48 h later, andviable colonies were selected and expanded. Cell viability was deter-mined by counting and plating the cells into 24-well plates. Eachclone was plated at a density of about 1 � 104 cells per well intriplicate and the experiments were repeated twice. The media werereplaced every 2 days throughout the period. Growth kineticswas monitored over a week by direct cell counting using a hemo-cytometer.
Recombinant adenovirus construction and production. A C-ter-minal hemagglutinin (HA)-tagged Ang-1 was subcloned into BamHI/HindIII sites of pCA13 adenoviral shuttle vector using Pfu DNApolymerase with primer pair 5� CCATCGATGGCGGGATCCCG-TTATCAGAGGCTAGCGTAATCCGGAACATCGTAGGGTAAAA-ATCTAAAGGTCGAATC 3�, where the underlined portion repre-sents BamHI restriction site, the engineered translational termina-tors are in italic, and the complementary HA tag is in boldface, and5� CCCAAGCTTGGGCTAGTTTTCTATGATTCTTCTTCAAACGC 3�,where the underlined portion represents HindIII. The adenoviruswas packaged by cotransfecting Ang-1 containing pCA13 shuttlevector or empty vector with pJM17 Ad5 genomic plasmid (Microbix,Canada) into 293 cells using Lipofectin reagent as described by themanufacturer (Life Technologies). Individual viral clones wereplaque-purified three times; verified and high-titer viral stocks weregenerated by culturing in 150-cm dishes and purified by doublecesium chloride gradient ultracentrifugation. Viral infectious titerwas determined using the endpoint cytopathic effect (cpe) assay(Quantum Biotechnology, Canada) and viral particle count was de-termined with a spectrophotometer at 260 nm. The particle-to-infec-tious titer ratio was below 100.
Isolation of RNA and RT-PCR. Total RNA was isolated fromHeLa and 293 cells and tumor nodules using Trizol reagent (Life
Technologies) following the manufacturer’s instructions. The RNAwas treated with 1 unit of RNase-free DNase (Promega) by incubat-ing at 37°C for 15 min, followed by 70°C for 5 min, to removecontaminating genomic DNA. cDNA was synthesized from 3 �g oftotal RNA using the Access RT kit (Promega) as described by themanufacturer. Preliminary analyses were performed to determinethe exponential phase of the PCR amplification and suitable cyclenumbers were then selected for semiquantitative reverse transcrip-tion (RT)-PCR analyses (data not shown). PCR was performed for30–35 cycles with 30 s at 94°C, 60 s at 60°C, and 60 s at 68°C andended with 7 min at 68°C using primer pair 5� Ang1HRT (5� CAG-GAGGATGGTGGTTTGATGCTT 3�) and 3� Ang1HRT, (5� CTCA-AACGGCTCCAGATTCACGGTC 3�) for Ang-1. To analyze Ad-Ang-1-driven upregulation of Ang-1 expression, primer pair 5� ForAd1S(5� CGGTGAATATTGGCTGGGGAATGAG 3�) and 3� ForAd1S (5�GTAGTGCCACTTTATCCCATTCAG 3�) was used. Primers and PCRconditions of GAPDH are the same as described in Ref. [21].
Tie-2 binding and phosphorylation assays. The Tie-2 in vitrobinding assays were performed using FLAG-Tie-2 and FLAG-Tie-1as previously described [22]. Anti-Ang-1 antibody (Santa Cruz Bio-tech) was used to detect binding of Ang-1. For the Tie-2 phosphory-lation experiment, human umbilical vein endothelial cells (HUVECs)were serum-starved for 24 h before being incubated with concen-trated medium from Ad-Vector- or Ad-Ang-1-infected HEK293 cells.Anti-Tie-2 antibody (Santa Cruz Biotech) was used to immunopre-cipitate the Tie-2 receptor and anti-phosphotyrosine antibody wasused to assess the activation status of the receptor as previouslydescribed [22]. Recombinant Ang-1 protein was collected and concen-trated from conditioned medium of Ad-Ang-1-infected HEK293 cellsusing Centricon concentrators (Amicon, Beverly, MA).
Mouse tumor assays. Six- to eight-week-old female Balb/c nudemice were fed a diet of animal chow and water ad libitum throughoutthe experiment. After cell viability was verified as being �80% withtrypan blue exclusion assay, HeLa cells (5 � 106) in 0.5 ml PBS wereinjected into the peritoneum cavity of the animals. Parental HeLaand control vector-transfected HeLa cells were injected separately ascontrols. Each experimental group consisted of four to seven animalsand the experiment was repeated twice. Animals in all the groupswere sacrificed after 6–7 weeks and carefully dissected tumor nod-ules were meticulously harvested, weighed, and either placed in 10%neutral buffered formalin for paraffin sectioning or snap-frozen forRNA or protein analysis.
The characterized and purified recombinant adenovirus Ad-Ang-1(5 � 109 PFU) was injected intraperitoneally starting from 1 weekafter tumor inoculation on a weekly basis for 4–5 weeks. Separategroups of mice were injected with control virus Ad-Vector or PBSduring the same period as controls.
Immunoblot analysis and immunohistochemistry. Protein ex-pression was analyzed by Western blot using a polyclonal antibodyagainst Ang-1 (Santa Cruz Biotech). Cells were lysed in M-PERsolution (Pierce Chemical Co., Pierce, IL) supplemented with 10�g/ml aprotinin, 1 �g/ml pepstatin, 1 �g/ml leupeptin, and 2 mMPMSF according to the manufacturer. Alternatively, 24-h serum-freeconditioned media were collected and concentrated using Centricon(Amicon). Equal amounts of proteins were loaded and analyzed by10% SDS–PAGE and transferred to nitrocellulose membrane(Amersham–Pharmacia, UK). The membrane was blotted with di-luted Ang-1 in 5% nonfat milk/0.2% Tween-20 in TBS and detectedby diluted HRP-conjugated secondary antibody (Santa Cruz Biotech)and chemiluminescent substrate (Pierce) according to the manufac-turer’s instructions. For immunohistochemical analysis, sectionswere cut at 4 �m, mounted on poly-L-lysine-treated slides, andstained with anti-vWF, anti-desmin (clone D33), anti-muscle-specificactin (clone HHF35) antibodies (Dako, Denmark), anti-� smoothmuscle actin (clone 1A4) (Sigma, St. Louis, MO), anti-Ang-1 (sc-6319), anti-VEGF (A20), anti-PCNA (sc-56) antibodies (Santa Cruz
300 SHIM ET AL.

Biotech). Signals were detected by the streptavidin–biotin system orEnvision Kit (Dako, Denmark) and color was developed in 3,3�-diaminobenzidine (DAB). Treatment procedures for control slideswere similar except that the primary antibody was omitted or pre-immune serum was used.
Microvessel density count. Sections were examined under a lightmicroscope to determine the quality of the staining. This was fol-lowed by careful examination and selection of suitable microscopic-fields for vessel counting following the criteria described by Weidneret al. [23]. Tumors were frequently heterogenous in their microvesselcontent, but areas of highest neovascularization or “hot spots” werelocated by scanning the tumor sections at 40� and 200� magnifica-tion and viewing fields with the highest number of discrete microves-sels stained by vWF were selected for counting at 400� magnifica-tion. A single countable microvessel was defined as any brownimmunostained endothelial cell or cluster of endothelial cells clearlyseparated from adjacent microvessels, tumor cells, and other connec-tive tissue elements. Presence of a lumen or erythrocytes in thelumen was not necessary to define a structure as a microvessel.Vascularity in areas of necrosis was not included. On average, three“hot spots” were selected from each section and three fields werecounted within each spot. Microvessel density (MVD) was expressedas the average number of countable microvessels identified withinthe three 400� fields (corresponding to 0.19 mm2 per 400� field).
Apoptotic index. The apoptotic index (AI) was calculated as thenumber of apoptotic cells per field of neoplastic tissue in the sectionand the apoptotic cells were identified following accepted criteria [24,25]. Cells were counted as apoptotic when they showed markedchromatin condensation and shrinkage of cytoplasm or strongly eo-sinophilic cytoplasm with variable-sized apoptotic bodies which con-tained one or more specks of pyknotic chromatin and were separatedfrom neighboring cells. Cells in necrosis areas or located aroundgrossly necrotic zones were not included in the counting. Two repre-sentative specimens from each group of tumor lines were included forcounting using an Olympus BX4 light microscope with ocular lensfitted with a 10 � 10 counting grid/field and counted under 400�magnification corresponding to an area of 0.0625 mm2. About six tonine random fields were selected from each section. The AI wasexpressed as the average number of counted apoptotic cells withinthe selected fields.
Vessel maturity index. The smooth muscle actin (SMA)-, muscle-specific actin (MSA)-, and desmin-stained sections were carefullyexamined under a microscope to determine the quality of the stain-ing. Then, the total numbers of periendothelial cell-associated andperiendothelial cell-free microvessels were counted under 400� mag-nification. Only individual vessels that were clearly separated fromclusters of vessels were included. Furthermore, only relatively largevessels with well-defined lumens that contained several erythrocyteswere counted. Small capillaries that are frequently sparsely coated
with periendothelial cells and, therefore, may falsely appear as un-covered in these sections were not included in counting. Randomsections from two or three different tumor nodules from each groupwere selected for counting for each staining marker. An independentobserver who was blinded to the outcomes of the experiment wasinvolved in repeating the assessment based on the set criteria. Thevessel maturity index was expressed as the fraction of MSA-, SMA-,or desmin-positive periendothelial cell-associated vessels per crosssection.
Statistical analysis. The data are presented as means � SEM.The statistical software SigmaStat Version 2.0.3 (SPSS, USA) wasemployed in the statistical analyses. One-way ANOVA followed byStudent’s t test was used for sequential pairwise comparison be-tween two groups of samples. A P value �0.05 was consideredstatistically significant.
RESULTS
Generation of Ang-1-Overexpressed StableHeLa Clones
The human cervical carcinoma cell line HeLa natu-rally expresses a moderate level of Ang-1, but does notexpress Ang-2 [21] (Fig. 1 of this work). To study theroles of Ang-1 protein in tumor angiogenesis and tumorgrowth, we constructed stable HeLa cell clones thatconstitutively express high levels of Ang-1. Pooledempty vector transfectants were used as vector-modi-fied control cells for all experiments. RT-PCR andWestern blot analyses revealed a three- to fivefold in-crease in expression of both RNA and protein in twoAng-1 stably transfected clones compared with paren-tal as well as vector-modified HeLa cells (Figs. 1A and1D). All stably transfected clones showed similar levelsof GAPDH expression (Fig. 1B).
Recombinant Ang-1 Protein Is Bioactive
To test the bioactivity of the recombinant Ang-1 pro-tein, Ang-1 cDNA was cloned into human adenovirusexpression vector and packaged into recombinant Ad-Ang-1 adenovirus. Proteins produced from adenovirus-infected 293 cells were assayed for their binding toTie-2 receptor as well as stimulation of Tie-2 phosphor-
FIG. 1. Overexpression of Ang-1 in HeLa cells. (A) Ang-1-overexpressing HeLa cells showed enhanced expression of Ang-1 RNA. (B)Exogenous expression of Ang-1 did not affect GAPDH expression. (C) Negative controls without RT. (D) Western blot analysis using total celllysate showed increased exogenous Ang-1 protein expression. Lanes 1 and 2: vector-modified and parental HeLa; lanes 3 and 4: Ang-1-overexpressed lines, 1H(�)c1 and 1H(�)c2. M, molecular weight marker. Arrow indicates the Ang-1 message and protein.
301Ang-1 STIMULATES CERVICAL CANCER ANGIOGENESIS

ylation activity in HUVECs. As shown in Fig. 2, recom-binant Ang-1 protein preferentially binds to Tie-2 re-ceptor. In addition, it also induced robust and specificphosphorylation of Tie-2 receptor from HUVECs, dem-onstrating strong agonistic activity toward its cogni-tive receptor, while no such induction was observed bythe controls under similar conditions.
In Vivo Cervical Tumor Growth Is Enhanced by Ang-1
Mice bearing HeLa cells that overexpressed Ang-1showed increased tumor burden compared with thosein the control groups of vector-transfected and parentalHeLa cells (P � 0.05) (Fig. 3A). The increased tumorburden was the result of significantly enhanced tumorangiogenesis as determined by MVD count (P � 0.05)(Fig. 3B), and associated decrease in tumor AI (P �0.001) (Fig. 3C). However, no obvious apoptotic endo-thelial cells were detected among all the groups (datanot shown). The increased tumor growth was associ-ated with a drastic upregulation of Ang-1 mRNA intumors (Fig. 4A) and the exogenous Ang-1 expressionwas confirmed by RT-PCR using a combination of vec-tor-specific and gene-specific primers (Fig. 4B). No in-duction of Ang-2 mRNA expression was detected in theAng-1-overexpressed HeLa cancer cells (data notshown). The growth rates of the Ang-1 and vectortransfectants exhibited no significant differences inculture (data not shown), effectively excluding clonalvariation as a possible cause of the observed differ-ences in vivo.
Adenovirus-Mediated Ang-1 Gene Transfer PromotedCervical Cancer Growth
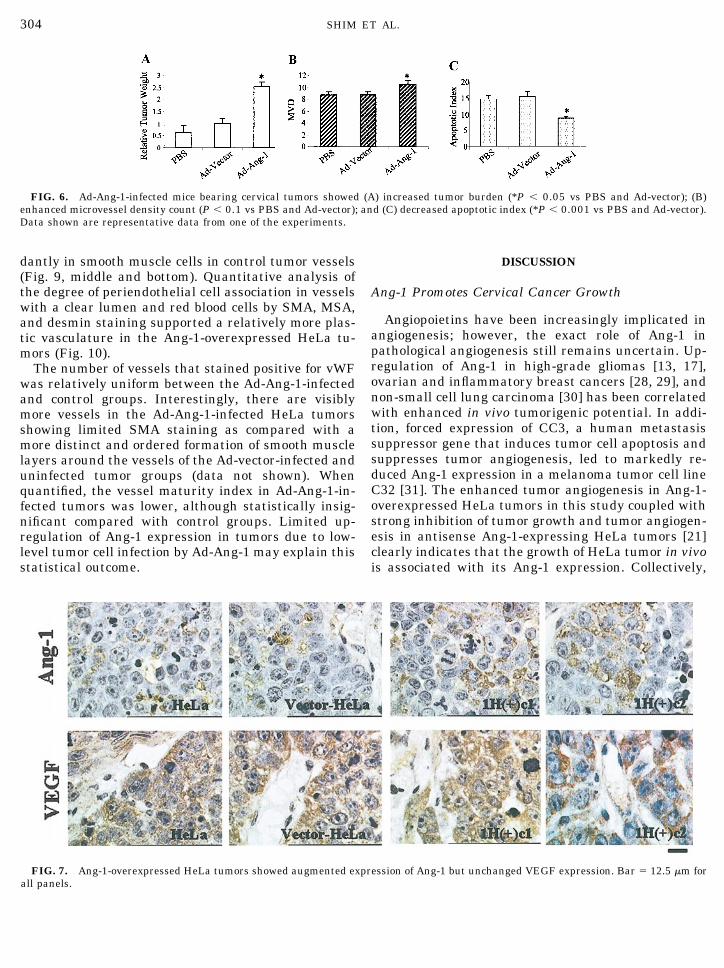
Recombinant human adenovirus carrying Ang-1gene (Ad-Ang-1) expressed large amounts of Ang-1mRNA and protein on infection of HeLa cells comparedwith control virus Ad-Vector-infected and uninfectedparental HeLa cancer cells in vitro (Figs. 5A and 5B).When Ad-Ang-1 was introduced intraperitoneally tocancer-bearing mice, a dramatic increase in tumor bur-den was observed (P � 0.05) (Fig. 6A). This increasedtumor burden was associated with a significantly de-creased AI (P � 0.001) (Fig. 6C). Interestingly, thisdramatic increase in tumor burden was only associatedwith a slight Ang-1 upregulation in Ad-Ang-1-infectedHeLa tumor cells in vivo and a marginally increasedMVD count (P � 0.1) (Figs. 5C, 5D, and 6B).
FIG. 2. Biological activity of recombinant Ang-1 protein. (A) Re-combinant Ang-1 showed strong selective binding to Tie-2 receptor(arrow). Western blot of FLAG-Tie-2 and FLAG-Tie-1 immunopre-cipitated proteins from Ad-vector-infected and Ad-Ang-1 infected 293media, respectively, using anti-Ang-1 antibody [22]. (B) Recombi-nant Ang-1 induced robust autophosphorylation of Tie-2 receptors inHUVECs. Anti-Tie-2 antibody was used to immunoprecipitateHUVEC lysates under various treatments and then probed with antiphosphotyrosine antibody to detect phosphorylated Tie-2 by Westernblot. (C) Western blot using anti-Tie-2 antibody confirming equiva-lent amount of Tie-2 present in the lanes in (B). Ang-1* (100 ng/ml)was included as a positive control [8]. Arrowheads indicate phospho-tyrosine-activated Tie-2 (B) and total Tie-2 (C) protein. CM, condi-tioned medium; IP, immunoprecipitation; WB, Western blot.
FIG. 3. Ang-1-overexpressed cervical tumors showed (A) increased tumor burden (*P � 0.05 vs HeLa and vector-HeLa); (B) enhancedmicrovessel density count (*P � 0.05 vs HeLa and vector-HeLa); and (C) decreased apoptotic index (*P � 0.001 vs HeLa and vector-HeLa).Data shown are representative data from one of the experiments.
302 SHIM ET AL.

Histological and Immunohistochemical Analysesof Ang-1-Overexpressed Cervical Cancers
Immunostaining of formalin-fixed, paraffin-embed-ded sections confirmed the enhanced expression ofAng-1 protein in Ang-1-overexpressed HeLa tumorcells consistent with the RT-PCR results (Fig. 7, top).Ang-1 exhibited localized granular cytoplasmic stain-ing in the tumor cells. No staining was observed in thenecrotic zones and in endothelial cells (data notshown). The staining pattern of Ang-1 is general anddiffused in the tumors in contrast to a more focal stain-ing pattern displayed by VEGF with strong staining intumor cells around well-vascularized regions and ne-crotic zones, reflecting its functional role as a potentangiogenic factor and hypoxia-responsive factor (Fig. 7,middle) [26]. However, no significant difference inVEGF staining between the Ang-1-overexpressed andcontrol tumors was detected. The tumor cell prolifera-tion rate as assessed by proliferating cell nuclear an-tigen (PCNA) staining was also unaffected by the en-hanced tumor growth as a result of increased Ang-1expression (data not shown).
Similar to RT-PCR results, the Ad-Ang-1-infectedtumor cells showed limited increases in Ang-1 expres-sion (Fig. 8, top). However, dramatic upregulation ofAng-1 protein was detected in the liver of Ad-Ang-1-infected mice (Fig. 8, middle). Similar to the stable celllines, no significant differences in tumor VEGF andPCNA staining were detected between the Ad-Ang-1group and control groups (Fig. 8, bottom, and data notshown).
Effect of Ang-1 Overexpression on HeLa TumorAngiogenesis and Tumor Vessel Plasticity
There were considerably more vessels that were vonWillebrand factor (vWF) positive (data not shown) butwith minimum or no SMA staining in Ang-1-overex-pressed tumors compared with control tumors (Fig. 9,top). Consistently, careful examinations also showedminimal staining for MSA and intermediate filamentdesmin in many vessels of Ang-1-overexpressed tu-mors, suggesting scarcity of smooth muscle supportingcells and indicating a state of vessel plasticity [27]. Incontrast, both MSA and desmin were expressed abun-
FIG. 5. Ad-Ang-1 virus-mediated enhanced expression of Ang-1 RNA and protein in infected HeLa cancer cells in vitro and in vivo. (A)RT-PCR. Lanes 1 and 2: Ang-1-expressing adenovirus-infected HeLa, Ad-Ang-1 (25 MOI) and Ad-Ang-1 (100 MOI), respectively; lane 3: mockvirus, Ad-vector-infected HeLa (100 MOI); lane 4: uninfected HeLa. (B) Upregulation of Ang-1 protein expression in Ad-Ang-1-infected HeLatotal cell lysate. Lanes 1–3: Ad-Ang-1-infected, uninfected, and mock virus Ad-vector-infected, respectively. (C) Enhanced Ang-1 expressionin Ad-Ang-1-infected HeLa tumors in vivo. Left: Ang-1; right: GAPDH. Lanes 1–3: Ad-Ang-1-infected, Ad-vector-infected, and uninfected,respectively. (D) RT-PCR analysis confirmed the Ad-Ang-1-driven Ang-1 expression in the infected HeLa tumors. Left: viral expressed Ang-1;right: negative controls without RT. Lanes 1–3: uninfected, Ad-Vector-infected, and Ad-Ang-1-infected tumors, respectively. Arrows indicatethe Ang-1 message and protein.
FIG. 4. Overexpression of Ang-1 resulted in enhanced tumor growth rate in HeLa tumors. (A) Overexpression of Ang-1 as determined byRT-PCR. (B) Enhanced exogenous expression of Ang-1 was mediated by stable expression of Ang-1 from integrated construct. Vector primer:5� CAGGAGGATGGTGGTTTGATGCTT 3�, Ang-1-specific primer: 5� ACCTACAGGTGGGGTCTTTCATTCCC 3�. (C) RNA integrity demon-strated by equivalent GAPDH expression among samples. (D) Negative controls omitting RT as in (C). Lanes 1 and 2: parental andvector-modified HeLa; lanes 3 and 4: Ang-1-overexpressed HeLa, 1H(�)c1, and 1H(�)c2. M, molecular weight marker. Arrow indicatesamplified Ang-1 message in (A) and exogenous sense Ang-1 message in (B).
303Ang-1 STIMULATES CERVICAL CANCER ANGIOGENESIS
dantly in smooth muscle cells in control tumor vessels(Fig. 9, middle and bottom). Quantitative analysis ofthe degree of periendothelial cell association in vesselswith a clear lumen and red blood cells by SMA, MSA,and desmin staining supported a relatively more plas-tic vasculature in the Ang-1-overexpressed HeLa tu-mors (Fig. 10).
The number of vessels that stained positive for vWFwas relatively uniform between the Ad-Ang-1-infectedand control groups. Interestingly, there are visiblymore vessels in the Ad-Ang-1-infected HeLa tumorsshowing limited SMA staining as compared with amore distinct and ordered formation of smooth musclelayers around the vessels of the Ad-vector-infected anduninfected tumor groups (data not shown). Whenquantified, the vessel maturity index in Ad-Ang-1-in-fected tumors was lower, although statistically insig-nificant compared with control groups. Limited up-regulation of Ang-1 expression in tumors due to low-level tumor cell infection by Ad-Ang-1 may explain thisstatistical outcome.
DISCUSSION
Ang-1 Promotes Cervical Cancer Growth
Angiopoietins have been increasingly implicated inangiogenesis; however, the exact role of Ang-1 inpathological angiogenesis still remains uncertain. Up-regulation of Ang-1 in high-grade gliomas [13, 17],ovarian and inflammatory breast cancers [28, 29], andnon-small cell lung carcinoma [30] has been correlatedwith enhanced in vivo tumorigenic potential. In addi-tion, forced expression of CC3, a human metastasissuppressor gene that induces tumor cell apoptosis andsuppresses tumor angiogenesis, led to markedly re-duced Ang-1 expression in a melanoma tumor cell lineC32 [31]. The enhanced tumor angiogenesis in Ang-1-overexpressed HeLa tumors in this study coupled withstrong inhibition of tumor growth and tumor angiogen-esis in antisense Ang-1-expressing HeLa tumors [21]clearly indicates that the growth of HeLa tumor in vivois associated with its Ang-1 expression. Collectively,
FIG. 6. Ad-Ang-1-infected mice bearing cervical tumors showed (A) increased tumor burden (*P � 0.05 vs PBS and Ad-vector); (B)enhanced microvessel density count (P � 0.1 vs PBS and Ad-vector); and (C) decreased apoptotic index (*P � 0.001 vs PBS and Ad-vector).Data shown are representative data from one of the experiments.
FIG. 7. Ang-1-overexpressed HeLa tumors showed augmented expression of Ang-1 but unchanged VEGF expression. Bar � 12.5 �m forall panels.
304 SHIM ET AL.
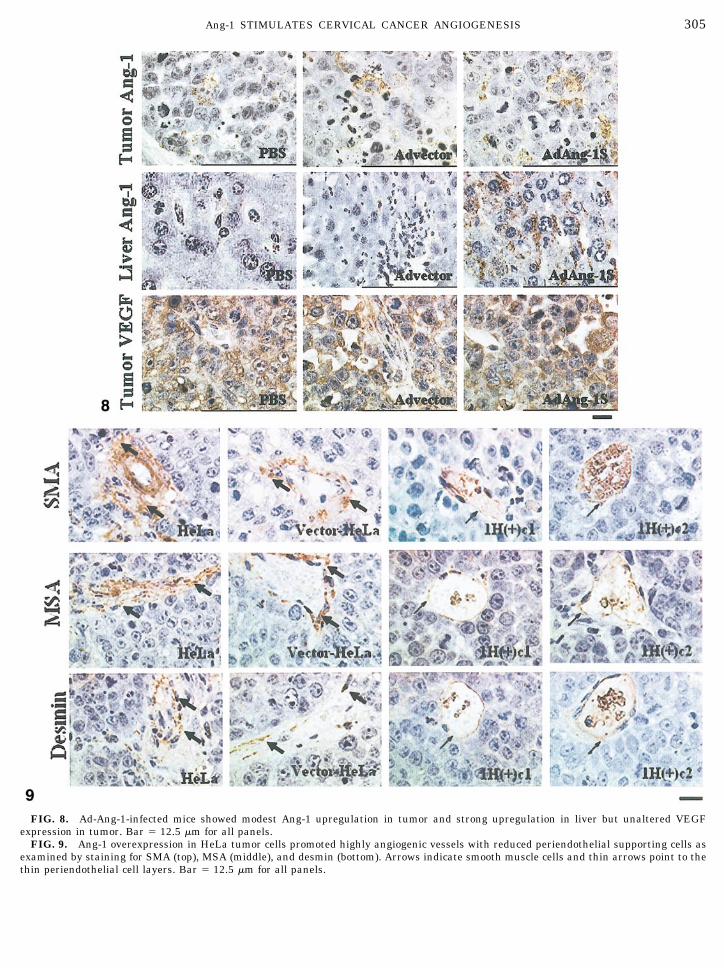
FIG. 8. Ad-Ang-1-infected mice showed modest Ang-1 upregulation in tumor and strong upregulation in liver but unaltered VEGFexpression in tumor. Bar � 12.5 �m for all panels.
FIG. 9. Ang-1 overexpression in HeLa tumor cells promoted highly angiogenic vessels with reduced periendothelial supporting cells asexamined by staining for SMA (top), MSA (middle), and desmin (bottom). Arrows indicate smooth muscle cells and thin arrows point to thethin periendothelial cell layers. Bar � 12.5 �m for all panels.
305Ang-1 STIMULATES CERVICAL CANCER ANGIOGENESIS

the data support Ang-1 as a crucial factor for tumorangiogenesis and tumor progression in this tumor.
A contrasting role for Ang-1 in tumor angiogenesisand tumor growth has, however, also been reported.For example, overexpression of Ang-1 in breast or coloncells was recently reported to be angiostatic and inhib-itory to tumor growth [32, 33]. Furthermore, the an-giostatic effect of Ang-1 in breast cancer xenograft wasshown to be mediated through its vessel-stabilizingfunction by recruiting periendothelial cells [34]. There-fore, the role of Ang-1 in neoplastic progression indifferent tumors and tissues remains elusive with in-creasing evidence of diverging multifunctional and, attimes, contrasting roles played by the Ang-1/Tie-2pathway in various aspects of angiogenesis.
The discrepancy between moderately increasedMVD count and significantly enhanced tumor burdenof Ad-Ang-1-injected mice suggests that the indiscrim-inate Ad-Ang-1 infection of tumor cells, host tissues,and organs as demonstrated in the infected liver (Fig.8) may have led to a general increase in vascularity inthe peritoneal cavity due to widespread Ang-1 expres-sion, and enabled neoplastic growth without robustintratumor neovascularization. In fact, glioma cellsand non-small cell lung carcinomas have been noted togrow without initiating tumor angiogenesis by co-opt-ing the existing rich vascular bed [35, 36]. It is possiblethat some forms of vessel co-option may exist in intra-peritoneally introduced tumor cells since considerableminor tumor nodules were noted along the mesenteryvessels (data not shown).
Ang-1 Promotes Tumor Vessel Plasticity
Immature nascent vessels have been found to dis-play differential rates of periendothelial cell recruit-
ment. Minor arteriole, venule, and capillary brancheshave been observed to lag behind major vessels insmooth muscle cell (SMC) coverage by days to weeks,creating a window of vessel plasticity [27, 37]. Thiswindow period was thought to enable fine-tuning of theneovasculature to adapt to its microenvironmentthrough regulated pruning and remodeling. VEGF hasbeen found to provide critical survival signal to theuncovered vessels during this period [27, 38], henceenabling subsequent establishment of a functional andsustainable vasculature. It is noted that immature ves-sels are functional, at least for the initial period, eventhough they have yet to acquire periendothelial cellcoating [27, 39].
The obvious reduction in the percentage of vesselswith periendothelial cell layers in Ang-1-overexpress-ing tumors was surprising since Ang-1 has been sug-gested to promote the recruitment of periendothelialcells [8]. However, vascular defects relating to lack ofinteractions between the endothelium and surround-ing cells in the Ang-1�/� and Tie-2�/� mutant mice havebeen suggested to be secondary to impaired survivalof the endothelium [40]. Indeed, persistent normalperiendothelial cell recruitment in vessels composedof Tie-2-deficient ECs that subsequently apoptosedstrongly supports this contention [41, 42]. Further-more, transgenic mice that overexpressed Ang-1 in theskin failed to show evidence of enhanced recruitment ofperiendothelial cells to the vessels [43]. Consistently,characteristic tortuous and distinct large-diametervessels promoted by VEGF and Ang-1 through thesonic hedgehog (Shh) signaling pathway in mesenchy-mal cells were found to have minimal numbers ofperiendothelial cells [44]. Collectively, these resultssupport the likelihood of complex control in vessel ma-
FIG. 10. Quantitation of vessel maturation by smooth muscle cells staining using antibody against MSA, SMA, and desmin (*P � 0.05vs HeLa and vector-HeLa).
306 SHIM ET AL.

turity with possible involvement of other crucial mole-cules such as Ephrin/Eph [45–47]. Therefore, Ang-1may function predominantly as a survival factor forangiogenic as well as quiescent endothelia for vesselmaintenance instead of vessel maturation, at least inHeLa tumors.
Indeed, the loss of cell–cell contact between ECneighbors and between ECs and periendothelial cellsobserved in the presence of Ang-1 has been suggestedto be a prerequisite for initiating in vitro and in vivosprouting angiogenesis to form cordlike structure [48].Therefore, the increased vessel plasticity coupled withenhanced MVD count in the Ang-1-overexpressedHeLa tumors suggests a more dynamic angiogenesisprocess with actively migrating, sprouting, and remod-eling endothelium. This is further supported by thefindings that vessel plasticity enables the endotheliumto actively respond to angiogenic factors such as VEGFsince mature vessels are independent of VEGF [37].Indeed, periendothelial cells have been found to inhibitendothelial proliferation and differentiation [49]. Fur-thermore, immature vessels have been associated witha high rate of tumor growth and neovascularization ina growth factor-rich environment in contrast to slow-growing tumors that may contain only a small fractionof immature vessels [38, 50]. Collectively, it may bespeculated that the increased vessel plasticity sanc-tioned by the upregulated Ang-1 protein is instrumen-tal in promoting and sustaining tumor angiogenesis inAng-1-overexpressing HeLa tumors.
The diverse functions of Ang-1/Tie-2 signaling invessel growth and stabilization may be intrinsic todifferent types of tumor and contribute varyingly totumor growth [51]. Furthermore, the existence of var-ious Ang-1 isoforms with related yet contrasting ago-nistic/antagonistic property [52], coupled with poten-tial regulation at the receptor level by Tie-2/Tie-1heterodimerization [53] and by Ang-1-modulated Tie-2downregulation in endothelial as well as tumor cells(Shim et al., unpublished data), adds to the complica-tions of the intricate role of Tie-2 signaling in angio-genesis. Such complex interplay between these diverseroles of Ang-1 may be crucial to the resultant directionof angiogenesis. Careful spatial and temporal exami-nation of the expression of Ang-1 and Tie-2 as well as oftheir antagonist, Ang-2, is needed to illustrate theirimportant roles during distinct phases of tumor devel-opment.
We thank staffs of the Animal Holding Unit at the National Uni-versity of Singapore for maintaining the mice used in this study, andMr. C. K. Ow of the Special Histology Laboratory, Department ofPathology, and Mr. M. S. Loh of the Histology Laboratory, Depart-ment of Biological Sciences, for help in the histology work.
REFERENCES
1. Folkman, J. (1972). Anti-angiogenesis: New concept for therapyof solid tumors. Ann. Surg. 175, 409–416.
2. Bicknell, R. (1994). Vascular targeting and the inhibition ofangiogenesis. Ann. Oncol. 5, S45–S50.
3. Kondos, S., Asano, M., and Suzuki, H. (1993). Significance ofvascular endothelial growth factor/vascular permeability factorfor solid tumor growth, and its inhibition by the antibody.Biochem. Biophys. Res. Commun. 194, 1234–1241.
4. Millauer, B., Shawver, L. K., Plate, K. H., Risau, W., andUllrich, A. (1994). Glioblastoma growth inhibited in vivo by adominant-negative Flk-1 mutant. Nature 367, 576–579.
5. Prewett, M., Huber, J., Li, Y., Santiago, A., O’Connor, W., King,K., Overholser, J., Hooper, A., Pytowski, B., Witte, L., Bohlen,P., and Hicklin, D. J. (1999). Antivascular endothelial growthfactor receptor (fetal liver kinase 1) monoclonal antibody inhib-its tumor angiogenesis and growth of several mouse and humantumors. Cancer Res. 59, 5209–5218.
6. Strawn, L. M., McMahon, G., App, H., Schreck, R., Kuchler,W. R., Longhi, M. P., Hui, T. H., Tang, C., Levitzki, A., Gazit, A.,Chen, I., Keri, G., Orfi, L., Risau, W., Flamme, I., Ullrich, A.,Hirth, K. P., and Shawver, L. K. (1996). Flk-1 as a target fortumor growth inhibition. Cancer Res. 56, 3540–3545.
7. Koblizek, T. I., Weiss, C., Yancopoulos, G. D., Deutsch, U., andRisau, W. (1998). Angiopoietin-1 induces sprouting angiogene-sis in vitro. Curr. Biol. 8, 529–532.
8. Maisonpierre, P. C., Suri, C., Jones, P. F., Bartunkova, S.,Wiegand, S. J., Radziejewski, C., Compton, D., McCain, J.,Aldrich, T. H., Papadopoulos, N., Daly, T. M. J., Davis, S., Sato,T. N., and Yancopoulos, G. D. (1997). Angiopoietin-2, a naturalantagonist for Tie2 that disrupts in vivo angiogenesis. Science277, 55–60.
9. Papapetropoulos, A., Fulton, D., Mahboubi, K., Kalb, R. G.,O’Connor, D. S., Li, F., Altieri, D. C., and Sessa, W. C. (2000).Angiopoietin-1 inhibits endothelial cell apoptosis via the Akt/survivin pathway. J. Biol. Chem. 275, 9102–9105.
10. Wong, A. L., Haroon, Z. A., Werner, S., Dewhirst, M. W., Green-berg, C. S., and Peters, K. G. (1997). Tie2 expression and phos-phorylation in angiogenic and quiescent adult tissues. Circ.Res. 81, 567–574.
11. Beck, L., and D’Amore, P. A. (1997). Vascular development:Cellular and molecular regulation. FASEB J. 11, 365–373.
12. Folkman, J., and D’Amore, P. A. (1996). Blood vessel formation:What is its molecular basis. Cell 87, 1153–1155.
13. Ding, H., Roncari, L., Wu, X., Lau, N., Shannon, P., Nagy, A.,and Guha, A. (2001). Expression and hypoxic regulation ofangiopoietins in human astrocytomas. Neuro-Oncology 3, 1–10.
14. Eggert, A., Ikegaki, N., Kwiatkowski, J., Zhao, H., Brodeur,G. M., and Himelstein, B. P. (2000). High-level expression ofangiogenic factors is associated with advanced tumor stage inhuman neuroblastomas. Clin. Cancer Res. 6, 1900–1908.
15. Goede, V., Schmidt, T., Kimina, S., Kozian, D., and Augustin,H. G. (1998). Analysis of blood vessel maturation processesduring cyclic ovarian angiogenesis. Lab. Invest. 78, 1385–1394.
16. Hazzard, T. M., Christenson, L. K., and Stouffer, R. L. (2000).Changes in expression of vascular endothelial growth factorand angiopoietin-1 and -2 in the macaque corpus luteum duringthe menstrual cycle. Mol. Hum. Reprod. 6, 993–998.
17. Stratmann, A., Risau, W., and Plate, K. H. (1998). Cell type-specific expression of angiopoietin-1 and angiopoietin-2 sug-gests a role in glioblastoma angiogenesis. Am. J. Pathol. 153,1459–1466.
307Ang-1 STIMULATES CERVICAL CANCER ANGIOGENESIS

18. Bunone, G., Vigneri, P., Mariani, L., Buto, S., Collini, P., Pilotti,S., Pierotti, M. A., and Bongarzone, I. (1999). Expression ofangiogenesis stimulators and inhibitors in human thyroid tu-mors and correlation with clinical pathological features. Am. J.Pathol. 155, 1967–1976.
19. Cohen, B., Barkan, D., Levy, Y., Goldberg, I., Fridman, E.,Kopolovic, J., and Rubinstein, M. (2001). Leptin induces angio-poietin-2 expression in adipose tissues. J. Biol. Chem. 276,7697–7700.
20. Yu, Q., and Stamenkovic, I. (2001). Angiopoietin-2 is implicatedin the regulation of tumor angiogenesis. Am. J. Pathol. 158,563–570.
21. Shim, W. S. N., Teh, M., Mack, P. O., and Ge, R. (2001).Inhibition of angiopoietin-1 expression in tumor cells by anantisense RNA approach inhibited xenograft tumor growth inimmunodeficient mice. Int. J. Cancer 94, 6–15.
22. Kim, I., Kim, H. G., Kim, H., Kim, H. H., Park, S. K., Uhm,C. S., Lee, Z. H., and Koh, G. Y. (2000). Hepatic expression,synthesis and secretion of a novel fibrinogen/angiopoietin-re-lated protein that prevents endothelial-cell apoptosis. Biochem.J. 346, 603–610.
23. Weidner, N., Semple, J. P., Welch, W. R., and Folkman, J.(1991). Tumor angiogenesis and metastasis: Correlation in in-vasive breast carcinoma. N. Engl. J. Med. 324, 1–8.
24. Huovinen, R., Warri, A., and Collan, Y. (1993). Mitotic activity,apoptosis and TRPM-2 mRNA expression in DMBA-induced ratmammary carcinoma treated with anti-estrogen toremifene.Int. J. Cancer 55, 685–691.
25. Lipponen, P., and Aaltomaa, S. (1994). Apoptosis in bladdercancer as related to standard prognostic factors and prognosis.J. Pathol. 173, 333–339.
26. Chiarotto, J. A., and Hill, R. P. (1999). A quantitative analysisof the reduction in oxygen levels required to induce up-regula-tion of vascular endothelial growth factor (VEGF) mRNA incervical cancer cell lines. Br. J. Cancer 80, 15–18.
27. Benjamin, L. E., Hemo, I., and Keshet, E. (1998). A plasticitywindow for blood vessel remodeling is defined by pericyte cov-erage of the preformed endothelial network and is regulated byPDGF-B and VEGF. Development 125, 1591–1598.
28. Martoglio, A. M., Tom, B. D. M., Starkey, M., Corps, A. N.,Charnock-Jones, D. S., and Smith, S. K. (2000). Changes intumorigenesis- and angiogenesis-related gene transcript abun-dance profiles in ovarian cancer detected by tailored high den-sity cDNA array. Mol. Med. 6, 750–765.
29. Shirakawa, K., Tsuda, H., Heike, Y., Kato, K., Asada, R., Ino-mata, M., Sasaki, H., Kasumi, F., Yoshimoto, M., Iwanaga, T.,Konishi, F., Terada, M., and Wakasugi, H. (2001). Absence ofendothelial cells, central necrosis, and fibrosis are associatedwith aggressive inflammatory breast cancer. Cancer Res. 61,445–451.
30. Takahama, M., Tsutsumi, M., Tsujiuchi, T., Nezu, K., Kushibe,K., Taniguchi, K., Kotake, Y., and Konishi, Y. (1999). Enhancedexpression of Tie2, its ligand angiopoietin-1, vascular endothe-lial growth factor, and CD31 in human non-small cell lungcarcinomas. Clin. Cancer Res. 5, 2506–2510.
31. NicAmhlaoibh, R., and Shtivelman, E. (2001). Metastasis sup-pressor CC3 inhibits angiogenic properties of tumor cells invitro. Oncogene 20, 270–275.
32. Ahmad, S. A., Liu, W., Jung, Y. D., Fan, F., Wilson, M., Rein-muth, N., Shaheen, R. M., Bucana, C., and Ellis, L. M (2001).The effects of angiopoietin-1 and -2 on tumor growth and an-giogenesis in human colon cancer. Cancer Res. 61, 1255–1259.
33. Hayes, A. J., Huang, W. Q., Yu, J., Maisonpierre, P. C., Liu, A.,Kern, F. G., Lippman, M. E., McLeskey, S. W., and Li, L. Y.(2000). Expression and function of angiopoietin-1 in breast can-cer. Br. J. Cancer 83, 1154–1160.
34. Tian, S., Hayes, A. J., Metheny-Barlow, L. J., and Li, L.-Y.(2002) Stabilization of breast cancer xenograft tumor neovas-cularization by angiopoietin-1. Br. J. Cancer 86, 645–651.
35. Holash, J., Maisonpierre, P. C., Compton, D., Boland, P., Alex-ander, C. R., Zagzag, D., Yancopoulos, G. D., and Wiegand, S. J.(1999). Vessel cooption, regression, and growth in tumors me-diated by angiopoietins and vegf. Science 284, 1994–1998.
36. Pezzella, F., Pastorino, U., Tagliabue, E., Andreola, S., Sozzi,G., Gasparini, G., Menard, S., Gatter, K. C., Harris, A. L., Fox,S., Buyse, M., Pilotti, S., Pierotti, M., and Rilke, F. (1997).Non-small-cell lung carcinoma tumor growth without morpho-logical evidence of neoangiogenesis. Am. J. Pathol. 151, 1417–1423.
37. Korff, T., Kimmina, S., Martiny-Baron, G., and Augustin, H. G.(2001). Blood vessel maturation in a 3-dimensional spheroidalcoculture model: Direct contact with smooth muscle cells regu-lates endothelial cell quiescence and abrogates VEGF respon-siveness. FASEB J. 15, 447–457.
38. Benjamin, L. E., Golijanin, D., Itin, A., Pode, D., and Keshet, E.(1999). Selective ablation of immature blood vessels in estab-lished human tumors follows vascular endothelial growth fac-tor withdrawal. J. Clin. Invest. 103, 159–165.
39. Lindahl, P., Hellstrom, M., Kalen, M., and Betsholtz, C. (1998).Endothelial-perivascular cell signaling in vascular develop-ment: Lessons from knockout mice. Curr. Opin. Lipidol. 9,407–411.
40. Jones, N., Iljin, K., Dumont, D. J., and Alitalo, K. (2001). Tiereceptors: New modulators of angiogenic and lymphangiogenicresponses. Nat. Rev. 2, 257–267.
41. Jones, N., Voskas, D., Master, Z., Sarao, R., Jones, J., andDumont, D. J. (2001). Rescue of the early vascular defects inTek/Tie2 null mice reveals an essential survival function.EMBO Rep. 21, 438–445.
42. Puri, M. C., Partanen, J., Rossant, J., and Bernstein, A. (1999).Interaction of the Tek and Tie receptor tyrosine kinases duringcardiovascular development. Development 126, 4569–4580.
43. Suri, C., McClain, J., Thurston, G., McDonald, D. M., Zhou, H.,Oldmixon, E. H., Sato, T. N., and Yancopoulos, G. D. (1998).Increased vascularization in mice overexpressing angiopoi-etins-1. Science 282, 469–471.
44. Pola, R., Ling, L. E., Silver, M., Corbley, M. J., Kearney, M.,Pepinsky, R. B., Shapiro, R., Taylor, F. R., Baker, D. P., Asa-hara, T., and Isner, J. M. (2001). The morphogen sonic hedge-hog is an indirect angiogenic agent upregulating two families ofangiogenic growth factors. Nat. Med. 7, 706–711.
45. Gale, N. W., Baluk, P., Pan, L., Kwan, M., Holash, J., DeChiara,T. M., McDonald, D. M., and Yancopoulos, G. D. (2001). Eph-rin-B2 selectively marks arterial vessels and neovasculariza-tion sites in the adult, with expression in both endothelial andsmooth-muscle cells. Dev. Biol. 230, 151–160.
46. Wang, H. U., Chen, Z. F., and Anderson, D. J. (1998). Moleculardistinction and angiogenic interaction between embryonic ar-teries and veins revealed by ephrin-B2 and its receptor Eph-B4.Cell 93, 741–753.
47. Shin, D., Garcia-Cardena, G., Hayashi, S. I., Gerety, S., Asa-hara, T., Stavrakis, G., Isner, J., Folkman, J., Gimbrone, M. A.,and Anderson, D. J. (2001). Expression of ephrin-B2 identifies astable genetic difference between arterial and venous vascularsmooth muscle as well as endothelial cells, and marks subsets
308 SHIM ET AL.

of microvessels at sites of adult neovascularization. Dev. Biol.230, 139–150.
48. Audero, E., Cascone, I., Zanon, I., Previtali, S. C., Piva, R.,Schiffer, D., and Bussolino, F. (2001). Expression of angiopoi-etin-1 in human glioblastomas regulates tumor-induced angio-genesis: In vivo and in vitro studies. Arterioscler. Thromb. Vasc.Biol. 21, 536–541.
49. Shepro, D., and Morel, N. M. L. (1993). Pericyte physiology.FASEB J. 7, 1031–1038.
50. Charnock-Jones, D. S., and Burton, G. J. (2000). Placenta vas-cular morphogenesis. Bailliere’s Clin. Obstet. Gynecol. 14, 953–968.
51. Currie, M. J., Gunningham, S. P., Han, C., Scott, P. A. E.,Robinson, B. A., Harris, A. L., and Fox, S. B. (2001). Angiopoi-etin-1 is inversely related to thymidine phosphorylase expres-sion in human breast cancer, indicating a role in vascularremodeling. Clin. Cancer Res. 7, 918–927.
52. Huang, Y. Q., Li, J. J., and Karpatkin, S. (2000). Identificationof a family of alternatively spliced mRNA species of angiopoi-etin-1. Blood 95, 1993–1999.
53. Marron, M. B., Hughes, D. P., Edge, M. D., Forder, C. L., andBrindle, N. P. J. (2000). Evidence of heterotypic interactionbetween the receptor tyrosine kinases Tie-1 and Tie-2. J. Biol.Chem. 275, 39741–39746.
Received January 9, 2002Revised version received June 21, 2002Published online August 19, 2002
309Ang-1 STIMULATES CERVICAL CANCER ANGIOGENESIS




















