Anatomy and age of the Lothagam mandible
13
Andrew Hill Anatomy and age of the Lothagam Department of Anthropology, Yale mandible New anatomical data strengthen the allocation ofthe Lothagam mandible to the family Hominidae, most similar among known hominids to Australopithecus Steven Ward ufarmis. New biostratigraphic analyses of Lothagam Suidae, coupled with ~epar~rne~~ of A~at~my~ .Mo~th~astern data from the Tugen Hills succession, suggest that the Lothagam hominid is Ohio ~T~~ver~jt~es C&ge of Redstone, older than 5-6 Ma, and is therefore the earliest known member of this taxon. ~oo~s~o~~~, OH 44272, U.S.A. Barbara Brown Dtipartment pfCel1 Biology and Anatomy, Johns Hopkins Medical School, Baltimore, MD 21205, U.S.A. Received lOJuly 1990 Revision received 27 October 1991 and accepted 10 November 1991 iCeywords:Hominidae, Lothagam, Kenya, mandible, dating. Joumal of Human Eaolution (1992) 22,439%45 1 Introduction One major set of questions in paleoanthropolo~ involves the timing of the origin of clades. For example, if speciation is to some extent controlled by environmental factors, then it is essential to be able to establish the times ofcladistic branching so as to make correlations with climatic or other environmental changes documented by the geological or paleontological record. To understand these possible interactions we have to establish the age of branching points with as high a degree of accuracy as possible. In doing this, sometimes specimens are useful that are disappointing from the point ofview of other aims, such as investigations into functional anatomy. With the gready increased sample of Pliocene hominids for comparison, it is now possible to say more about fragmentary and previously neglected material. The Lothagam mandible (KNM-LT 329) is one example of this. Recently it has been discussed in several papers (Kramer, 1986; White, 1986; Hill & Ward, 1988), which have indicated its similarity to Australofiithecus afarensis. As it is probably older than other examples of this taxon, a good deal ofimportance attaches to its exact age, as it may represent the oldest member of the Hominidae so far known, using Hominidae in the sense of the hominoid lineage postdating the ancestor common to them and the African great apes. This paper provides new anatomical data which reinforce the allocation of the mandible to the Hominidae, allied particularly among already known hominids to A~~ralopi~hecus afarensis. In addition, we provide information from the biostratigraphy ofLothagam Suidae that has a bearing on assessments of its age. This is not conclusive, but we also indicate further more detailed work that might strengthen the suggestion. The Lothagam hominid: KNM-LT329 The Lothagam mandible (KNM-LT 329) was found in 1967 by Patterson’s Harvard Expedition. Patterson et al. (1970) originally attributed the specimen to Austra~opithec~ cf. 0047-2484/92/060439+ 13 $03.00/O 0 1992 Academic Press Limited
Transcript of Anatomy and age of the Lothagam mandible

Andrew Hill Anatomy and age of the Lothagam Department of Anthropology, Yale mandible
New anatomical data strengthen the allocation ofthe Lothagam mandible to the family Hominidae, most similar among known hominids to Australopithecus
Steven Ward ufarmis. New biostratigraphic analyses of Lothagam Suidae, coupled with
~epar~rne~~ of A~at~my~ .Mo~th~astern data from the Tugen Hills succession, suggest that the Lothagam hominid is
Ohio ~T~~ver~jt~es C&ge of Redstone, older than 5-6 Ma, and is therefore the earliest known member of this taxon.
~oo~s~o~~~, OH 44272, U.S.A.
Barbara Brown Dtipartment pfCel1 Biology and Anatomy, Johns Hopkins Medical School, Baltimore, MD 21205, U.S.A.
Received lOJuly 1990 Revision received 27 October 1991 and accepted 10 November 1991
iCeywords:Hominidae, Lothagam, Kenya, mandible, dating. Joumal of Human Eaolution (1992) 22,439%45 1
Introduction
One major set of questions in paleoanthropolo~ involves the timing of the origin of clades.
For example, if speciation is to some extent controlled by environmental factors, then it is
essential to be able to establish the times ofcladistic branching so as to make correlations with
climatic or other environmental changes documented by the geological or paleontological
record. To understand these possible interactions we have to establish the age of branching
points with as high a degree of accuracy as possible. In doing this, sometimes specimens are
useful that are disappointing from the point ofview of other aims, such as investigations into
functional anatomy. With the gready increased sample of Pliocene hominids for comparison,
it is now possible to say more about fragmentary and previously neglected material.
The Lothagam mandible (KNM-LT 329) is one example of this. Recently it has been
discussed in several papers (Kramer, 1986; White, 1986; Hill & Ward, 1988), which have
indicated its similarity to Australofiithecus afarensis. As it is probably older than other examples
of this taxon, a good deal ofimportance attaches to its exact age, as it may represent the oldest
member of the Hominidae so far known, using Hominidae in the sense of the hominoid
lineage postdating the ancestor common to them and the African great apes. This paper
provides new anatomical data which reinforce the allocation of the mandible to the
Hominidae, allied particularly among already known hominids to A~~ralopi~hecus afarensis.
In addition, we provide information from the biostratigraphy ofLothagam Suidae that has a
bearing on assessments of its age. This is not conclusive, but we also indicate further more
detailed work that might strengthen the suggestion.
The Lothagam hominid: KNM-LT329
The Lothagam mandible (KNM-LT 329) was found in 1967 by Patterson’s Harvard Expedition. Patterson et al. (1970) originally attributed the specimen to Austra~opithec~ cf.
0047-2484/92/060439+ 13 $03.00/O 0 1992 Academic Press Limited

440 A. HILL ET AL.
Figure 1. Views of KNM-LI’ 329. (a) Occlusal; (b) lateral.
africanus on the basis of its gracile corpus and relatively small M, crown, and despite a
suggestion ofaffinity with Proconsul major (Eckhardt, 1977), there has been a consensus that it
was a hominid, albeit too fragmentary to make a more detailed assignment (Howell, 1978).
The greatly augmented sample of Pliocene mandibular material (Johanson & White, 1980;
White, 1977, 1980; White et al., 1981; White & Johanson, 1982; Hill, 1985a; Ward & Hill,
1987) has made a more definitive assessment possible.
Since 1986, three detailed discussions concerning the morphology, metrics, and taxonomic
status of the specimen have appeared (Kramer, 1986; White, 1986; Hill & Ward, 1988).
While it is not our intention to repeat such descriptive information here, we have continued
our investigations on early hominid mandibular form reported elsewhere (Ward & Hill,
1987; Brown, 1989) and wish to report the results of these observations as they may be
relevant to more confident taxonomic assignment of this specimen. All data and observations
reported here were obtained from the original specimens at the National Museums ofKenya,
Nairobi, and for the Hadar hominids, while they were housed in the Cleveland Museum of
Natural History.
The Lothagam specimen, KNM-LT 329, is a fragmentary right mandibular corpus,
broken anteriorly through the distal root socket of P,. It is broken posteriorly through the
distal socket of M, (Figure 1 a, b) . The crown and root system of M,, as well as the roots of M,,
the mesial root of M, and parts of the distal root of M, are also preserved. The buccal cusps of
M, have been obliterated by wear and the lingual cusps have been flattened. However, from

LOTHAGAM MANDIBLE 441
the occl~sal anatomy that is preserved, it can be determined that the hypoconulid is mesially
appressed to the crown, a feature that White (White, 1986; White et al., 1981) regards as one
characteristic ofslustralokithecus ufarensis. A detailed description of the sub-occlusal anatomy
of KNM-LT 329 is reported in Hill & Ward (1988), but we note here that the preserved
molar roots demonstrate the serrate pattern that seems to be characteristic of A. ufarensis
mandibles (Ward & Molnar, 1978; Ward et al., 1982). However, our recent examination ofa
cast ofOmo 18 (0.18-1967-18) attributed to Paraustralopithecus aethiopicus by Arambourg &
Coppens (1967), seems to show a similar post-canine root pattern. In essence, Omo 18
possesses large A. b&i-like molar roots arranged in an A. ~~rens~s implantation pattern, but
in a mandibular corpus with A. boisei-like contours. This is of some interest, as A. a~~~~nus
does not exhibit a serrate lower molar root pattern. It is therefore possible that the serrate
implantation pattern is primitive for all early hominids. The functional/mechanical basis of
this implantation arrangement is currently unknown.
Overall, lateral corpus surface contours are muted, so that features such as the arrange-
ment of the oblique line, regions of corpus hollowing and other buttressing systems, are
difficult to discern. There is evidence of hollowing under the M, (White, 1986) in a pattern
that is concordant with that described for A. ufurensis, and unlike that of A. africanus (White,
1977). Other features reported to be shared with the A. ufarensis mandibular sample are a
high and narrow extramolar sulcus, hollowing of the lingual corpus beneath the mylohyoid
line and a low, distal extension of the inferior transverse torus (White, 1986; Hill & Ward,
1988).
Metric features of the first molar that have been reported are length and breadth
dimensions (White, 1986; Kramer, 1986; Corruccini & &Henry, 1980), crown area, crown
module (White, 1986) and crown shape index (White, 1986; Kramer, 1986). Comparisons of
these data with early hominids, selected later Miocene hominoids and chimpanzees, ally
KNM-LT 329 with hominids. From Kramer’s data (1986) it also appears that in its crown
shape index, the Lothagam M, overlaps the range for modern chimpanzees. It is difficult to
interpret these observations since we lack any evidence of early post-divergence African ape
teeth. However, it appears that given the fossils we do have, the Lothagam M, is metrically
consistent with attribution to Hominidae.
Metric assessment of corpus dimensions presents a less clear-cut picture. Minimum corpus
breadth reported by Kramer (1986) falls within the ranges for A. a~~ensis and A. africanus
with KNM-LT 329 falling closer to the A. unrigs mean. However, minimum corpus height,
and a shape index derived by dividing breadth by height at the M,-M, contact, places the
Lothagam mandible within the ranges reported for A. afarensis, A. ufr~cun~, the Miocene
hominoid mandible samples from Ravin du Pluie and Rudabanya and 63 chimpanzee
mandibles. In all instances, however, it falls closest to the A. ufurensis mean.
Based on their analyses, White (1986) felt compelled to limit attribution to Hominoidea
indet., while Kramer (1986) suggested attribution to A. ufurensis. There is justification for
both positions. However, while White’s and Kramer’s papers were in press, a new hominoid
mandible fragment, dated at about 5 Ma, from Tabarin in the Lake Baring0 Basin,
was announced (Hill, 1985a), and subsequently described in more detail (Ward & Hill,
1987). As reported by Ward & Hi11 (1987), the Tabarin mandible exhibits dental and
corpus features that suggest affiliation with the A. ufarensis mandible sample. Of particular
interest was the overall pattern of surface contours. We have extended these observations,
by applying ster~ophoto~rammetric procedures to the study of early hominid corpus morphology.
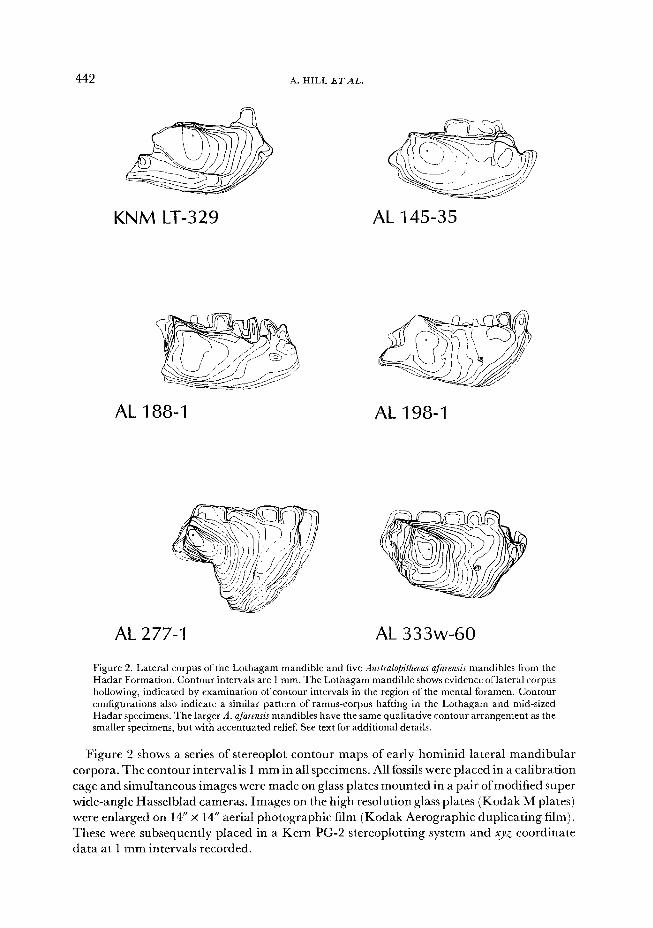
442 A. HILL ET AL.
KNM LT-329
AL 188-I
AL 145-35
AL 198-l
AL 277-l AL 333w-60
Figure 2. Lateral corpus of the Lothagam mandible and five $ustra@ithecus afarensis mandibles from the Hadar Formation. Contour intervals are 1 mm. The Lothagam mandible shows evidence oflateral corpus hollowing, indicated by examination of contour intervals in the region of the mental foramen. Contour configurations also indicate a similar pattern of ramus-corpus hafting in the Lothagam and mid-sized Hadar specimens. The larger A. afarensis mandibles have the same qualitative contour arrangement as the smaller specimens, but with accentuated relief. See text for additional details.
Figure 2 shows a series of stereoplot contour maps of early hominid lateral mandibular
corpora. The contour interval is 1 mm in all specimens. All fossils were placed in a calibration
cage and simultaneous images were made on glass plates mounted in a pair of modified super
wide-angle Hasselblad cameras. Images on the high resolution glass plates (Kodak M plates)
were enlarged on 14” x 14” aerial photographic film (Kodak Aerographic duplicating film).
These were subsequently placed in a Kern PG-2 stereoplotting system and XJZ coordinate
data at 1 mm intervals recorded.

LOTHAGAM MANDIBLE 443
Inspect-ion of the resulting contour maps shows that KNM-LT 329 shares a lateral corpus
surface topographic pattern with the smaller mandibles in the Hadar hominid sample,
represented by AL 145-35, AL 188-l and AL 198-l. Although other mandibles qualify for
inclusion in this group, such as AL 288-la, they tend to be severely damaged in the region
of corpus-ramus hafting, and are therefore unsuitably for contour digitization. The three
A. ufurensis specimens mentioned above are similar in contour distribution to the Lothagam
mandible in the region of the oblique line and the form of the concavity beneath the molars.
Although KNM-LT 329 is broken anterior to M,, we can make some inferences about the
lateral corpus surface beneath the premolars. The preserved contour pattern anterior to the
lateral eminence is consistent with the assumption that the corpus was concave beneath the
premolars. This assessment is supported by the pattern of hollowing around the mental
foramen. The Lothagam mandible therefore appears to manifest an important diagnostic
feature proposed by White et al. (1981) and White (1986) to distinguish A. uf~~~~s among
early hominids. Our observations on molar root implantation, as well as surface contour
patterns, therefore extend the earlier work of White (1986) and Kramer (1986) and support
assignment of the specimen to A. cf. afarensis.
Age
At the time ofits discovery, the Tabarin mandible, dated at just less than 5 Ma, was quite
plausibly the oldest hominid (Hill, 1985a; Ward & Hill, 1987; Hill & Ward, 1988). Although
the Lothagam specimen had long been popularly considered as 5.5 Ma in age, it was not
necessarily a hominid, and various factors suggested it need not be as old as implied. The date
involved considerable interpolation between older and younger radiometric determinations,
coupled with uncertain biostratigraphic information. When the date was first proposed and
became entrenched, this biostratigraphic information was only very loosely diagnostic.
The sedimentary succession forming the Lothagam Group totals 650-720 m (Behrensmeyer,
1976) and is divided into three main units, labeled Lothagam 1,2 and 3. It is underlain by
Pliocene volcanics and a basaltic unit separates Lothagam 2 from Lothagam 3. Lothagam 1 is
the particularly relevant unit here, up to 480 m thick, and itself divided into three informal
lithological members, from bottom to top, A, B and C. Lothagam 1 varies sedimentologically
but predominantly represents fluvial and fluvio-deltaic environments. Lothagam IA is
unfossiliferous. Vertebrate fossils are rare in Lothagam 2.
Whole rock K/Ar dates are quoted by Patterson et al. (1970). A volcanic flow underlying
Lothagam 1 Agave a result of8.3 l&O*25 Ma (Berkeley KA-2294: 8.5 Ma, correcting for new
constants). The basaltic unit or sill, separating units Lothagam 2 and 3, and which is believed
to postdate Lothagam 3, was dated at 3.7 1 + 0.23 Ma (Berkeley KA-2262: 3.8 Ma, corrected).
Behrensmeyer (1976) quoted 3.73 kO.41 Ma for this date. She also refers to other dates on
crystal concentrates from an agglomeratic tuff at the base of Lothagam lC which ranged
from 24 Ma to 12 Ma (FM Consultants), and which clearly bear no relation to the age of the
fauna. The reversed magnetic polarity of the sill, coupled with the K/Ar date, suggests a
correlation with the Gilbert Reversed Epoch of GRTS.
The fauna from Lothagam has been used to suggest time placement relative to other sites.
It was recognized as being older than Kanapoi, slightly older than Kolinga and lower Kaiso,
partly equivalent to Kaperyon, and a little younger than Sahabi. Kaperyon, in the Tugen
Hills succession, was held then to be related to trachytes dated around 5 Ma, but radiometric
dates on these other sites at that time were not available. The commonly quoted date of

444 A. HILL ET AL.
5.5 Ma for Lothagam 1 was therefore no more than an informal guess, and Patterson et al. (1970:920) do not pretend otherwise: “5.0 to 5.5 m.y. would seem to be a plausible estimate
for the age of Lothagam-1”. Despite this, 5.5 Ma became widely accepted and sometimes
used as a standard by which to judge the age of other faunas.
Although the evidence did not justify the precision implied by its later usage, subsequent
paleontological investigations did come to support a date of around 5 Ma to 5.5 Ma in
general. This is the conclusion of Maglio (1970a,b, 1973), from his work on Elephantidae.
This again is primarily based upon the presence of Stegotetrabelodon orbus in both Lothagam 1
and the Tugen Hills Kaperyon beds. Stegotetrabelodon orbus is also reported from the Mpesida
beds, also in the Tugen Hills succession. The other Lothagam 1 elephant, Primelephas
gomphotheroides, was identified by Maglio from a site in the Chemeron Formation. On this
basis, Maglio (1970a,b) suggested an age for Lothagam 1 of between 5 Ma and 5.5 Ma. By
1973 he suggested greater than 5 Ma, probably closer to 5.5 Ma or 6.0 Ma. This slight shift
comes from his beliefs regarding the extent of fauna1 difference between Lothagam 3 and
Lothagam 1, coupled with the difference between the faunas ofLothagam and Kanapoi; the
latter he thought was about 4.0 Ma in age. Cooke & Maglio (1972), summarizing infor-
mation about Suidae and Proboscidea, indicate ages for Lothagam 1 of about 6 Ma to
4.8 Ma, and Hooijer & Maglio (1974) m a work dealing with the Lothagam fossil equids,
put forward an estimate for Lothagam 1 of 6.0 Ma. They suggested it was unlikely to be
younger than this.
Smart (1976) and Behrensmeyer (1976) p oint out that most ofthe fauna on which all these
various estimates are based comes from Lothagam lB, and as fossils derive from more than
300 m of section, material from Lothagam 1C could be somewhat younger.
Ultimately all the estimates also rely on data from the Tugen Hills succession. The date of
5 Ma quoted in this early work for the Kaperyon cannot be taken at face value, because there
is doubt and argument even as to the validity of the Formation (Chapman, 1971; Pickford,
1978; Chapman & Brook, 1978; Hill et al., 1986). Whether it exists in some form or other or
not, Pickford’s arguments and our own subsequent work suggest that at least some of the
rocks formerly attributed to this unit should be allocated to the Lukeino Formation, others to
the Chemeron. As for the Mpesida Beds, we now know that they are bracketed by dates of
636 Ma and 6.2 Ma; the Chemeron sites mentioned are probably between 5.6 Ma and
4.1 ?vIa (Hill, 1985a; Hill et al., 1985, 1986). Consequently, the Elephantidae, formerly
crucial in leading to the acceptance of the 5.5 Ma date for Lothagam 1, in fact imply a range
in age from perhaps > 6 Ma to 4.1 Ma.
Pigs
A number of comprehensive investigations have demonstrated the biostratigraphic value of
Plio-Pleistocene African Suidae (Cooke & Wilkinson, 1978; Harris & White, 1979). In
estimating the age of the Lothagam hominid, the genus Nyanzachoerus is particularly import-
ant. Cooke & Ewer (1972) originally described the Lothagam fossil pigs, including the new
species Nyan<achoerus tulotos and N. plicatus. This publication remains in most essential ways
definitive, although various nomenclatorial changes have occurred. Nyanzachoerus plicatus is
now recognized as a junior synonym of Jv. jaegeri, a species from Hamada Damous, Tunisia
(Coppens, 1971; Cooke, 1976; Cooke & Wilkinson, 1978; Harris & White, 1979; Cooke,
1985)) and N. tulotos is synonymized with N. syrticus, a species from Sahabi, Libya, the type of
which had been for a long time lost (Leonardi, 1952, 1954; Kotsakis & Ingino, 1980; Cooke,

LOTHAGAM MANDIBLE 445
1987). In the following account, these more recent names replace the ones the authors
concerned originally used. Cooke & Ewer (1972) show that _X syrticus and j\r.juegeri are both present in Lothagam 1 C.
This is reiterated in Cooke (1976), Cooke & Wilkinson (I 978) and Cooke ( 1985). Harris &
White (1979), in their study of the African Plio~pleistocene Suidae, suggest that JV. ~~t~c~s
and jv.jasgeri are on an ancestor-descendant lineage. Their work shows that, with only two
exceptions in east Africa, the two taxa are absolutely separated; they are never found at the
same site or level. The two apparent east African exceptions are Ekora and Lothagam 1 C.
The occurrence of a single specimen of N. juegeri along with many of jv. syrticus is recorded
from Lothagam 1 C (Harris & White, 1979; in their text but not in the summary table). Our
recent work shows that there is in fact more than one specimen involved, but all investigations
agree that Lothagam 1 contains two species of the genus ~~ff~~uc~oe~~ that are aImost
universally separated elsewhere.
Harris & White (1979) explain this situation at Ekora by suggesting that a period of time is
represented at the site. They explain their single specimen of N. juegeri at Lothagam 1 C as
being derived from Lothagam 3, where it is also known. Behrensmeyer (1976) points out that
Lothagam IC probably represents a relatively substantial period of time. Consequently an
alternative explanation is that the unit records the putative transition from JV. syrticus to
.N..jaegeri, as Harris & White impiy for the site of Ekora.
Our work in the Tugen Hills, west of Lake Baringo, Kenya, has established appearance
and disappearance times for taxa in the succession. The succession spans the period from
15.5 Ma to l-6 Ma fairly continuously, with only a few gaps (Hill et al., 1985, 1986; Hill,
1985b, 1987). Nyantachoerus .yyrticus is known only in the Lukeino Formation and ~V.jaegeri
only in the lower part of the overlying Chemeron Formation. In many places in the area, the
transition from Lukeino to lower Chemeron is apparently continuous. The units are mostly
separated by lavas ofKaparaina Basalt, which probably represent only a short period oftime.
There are a number of dates on these basaIts, but all center on about 5.6 Ma. Radiometric
ages in the underlying Lukeino and overlying Ghemeron are concordant. This date, 5-6 Ma,
plus or minus a few hundred thousand, would seem to be a good estimate ofthe time ofchange
from JV. syrticus to Jf.jaegeri in the east African succession.
Ifthis transition is monitored by Lothagam 1 C, and ifthe hominid derives from near to this
transition, then there is a possibility of more accurately estimating the age of the hominid
specimen. Accordingly, we have examined the stratigraphic distribution of suid specimens
through the Lothagam succession. This was possible through access to copies of Harvard
MCZ field records in the archives of the Kenya National Museum and from additional notes
and charts kindly provided by Kay Behrensmeyer, who was the geologist on some of those
expeditions. We have also confirmed earlier identifications of relevant specimens.
Relevant data are available for some specimens in the 1967 collection, In this collection
there are 29 identifiable suids for which there is information relating to stratigraphic place-
ment (Table 1). Thirteen specimens come from Lothagam 1B. None is fl..jaegeri, and all
but one or two (which will be discussed below) are attributed to Jv. syrticus. There are 14
specimens from Lothagam 1 C. Two are fl. jaegeri, and these are the specimens retrieved
from highest in the section, stratigraph~caIly just beneath the basaltic unit that separates
Lothagam 1 from the relatively unfossiliferous Lothagam 2. Of the rest, all but one (also to be
discussed below) are attributable to X. syr,rticus. Of the two specimens from Lothagam 3, one
has been attributed to .N.jaecgeri, the other to .Notochoerus euilus. Cooke & Ewer’s (1972) Figure
1 implies a slightly different stratigraphic placement of some particular specimens of
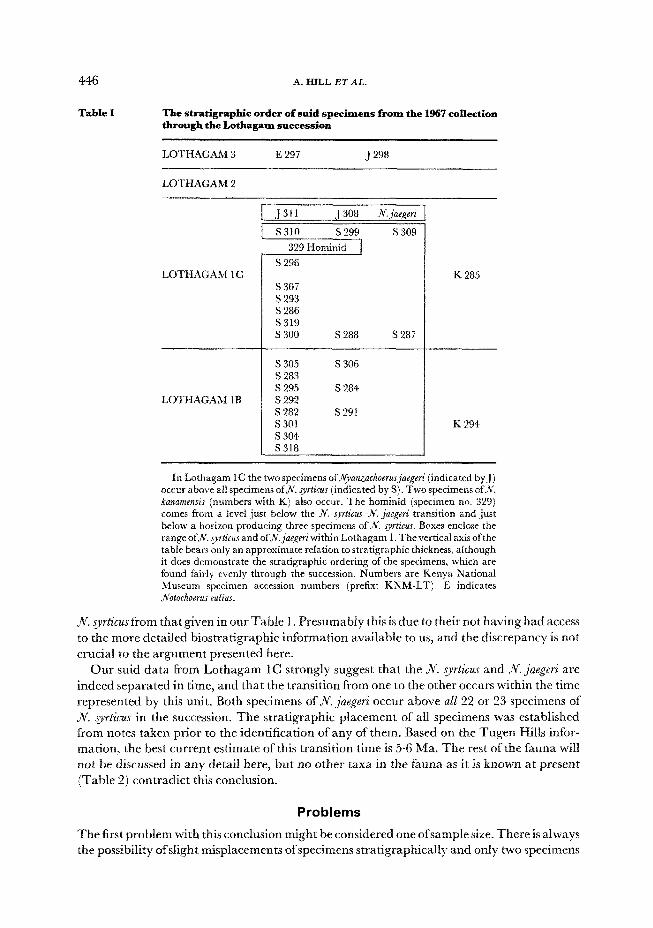
446
Table 1
A. EIILL ET AL.
The stratigrapbic order of suid specimens from the 1967 collection through the Lotbagam succession
LOTHAGAM 3 E 297 J 298
LOTHAGAM 2
LOTHAGAM 1 C
LOTHAGAM 1 B
S 296
s 307 S 293 S 286 s319 s 300 S 288 S 287
s 305 S 306 S 283 s 295 s 284 S 292 s 282 s291 s 301 s 304 5318
K 285
K 294
In Lothagam IC the two specimens OfJVyanZachoerusjaeReri (indicated by J) occur above ail specimens of P. s_vrt;cuS (indicated by S) Two specimens of& ~anu~ef~~s (numbers with K) also occur. The hominid (specimen no. 329) comes from a level just below the ,ni. q&us-R. &e@ transition and just below a horizon producing three specimens of JV. syrticus. Boxes enclose the range of&. syrtkus and ofN.nijaegeri within Lothagam 1, The vertical axis of the table bears only an approximate relation to stratigraphic thickness, although it does demonstrate the stratigraphic ordering of the specimens, which are found fairly evenly through the succession. Numbers are Kenya National Museum specimen accession numbers (prefix: KNM-LT). E indicates Notochoems eulius.
JV. syr~~&~sfrorn that given in our Table 1. presumably this is due to their not having had access
to the more detailed biostratigraphic information available to us, and the discrepancy is not
crucial to the argument presented here.
Our suid data from Lothagam 1C strongly suggest that the N. syrlicus and N. jaegeri are
indeed separated in time, and that the transition from one to the other occurs within the time
represented by this unit. Both specimens of N. jaegeri occur above all 22 or 23 specimens of
Jv. q~ticus in the succession. The stratigraphic placement of all specimens was established
from notes taken prior to the identification of any of them. Based on the Tugen Hills infor-
mation, the best current estimate of this transition time is 5-6 Ma. The rest of the fauna will
not be discussed in any detail here, but no other taxa in the fauna as it is known at present
(Table 2) contradict this conclusion.
Problems
The first problem with this conclusion might be considered one ofsample size. There is always
the possibility ofsfight misplacements ofspecimens stratigraphically and only two specimens

LOTHAGAM MANDIBLE
Table 2 Combined faunaI list for Lotbagam 1B and 1C
Class Order Family/Subfamily Species
447
Pisces
Aves Reptilia
Anseriformes Chelonia
Squamata Crocodilia
Testudinidae Tryonichidae Pelomedusidae Boidae Crocodilidae
Mammalia Primates Cercopithecidae
Carnivora
Proboscidea
Perissodactyla
Artiodactyla
Tubulidentata Rode&a
Hominidae Viverridae Hyaenidae Felidae Machairodontinae Deinotheriidae Gomphotheriidae Elephantidae
Equidae
Rhinocerotidae
Suidae
Hippopotamidae Giraffidae Bovidae
Orycteropodidae Sciuridae
Po&pterus other species
Geochelone sp.
Podocnemis 2 spp. Pythoninae Crocodylus lloidi Euthecodon cf. brumpti cf. Parapapio cf. Cercocebus Australopithecus cf. afarensis ? Viverra gen et sp. indet. Felinae (large sp.) gen et sp. indet. Deinotherium bozasi Anancus kenyensis Stegotetrabelodon orbus Primelephas gomphotheroides Hipparion turkanense Hipparion sitifense Hipparion primigenium Brachypotherium lewisi Ceratotherium praecox Nyanzachoerus syrticus ~yanzachoerusjaegeri flyanzachoerus kanamensis ?another species Hexaprotodon harvardi cf. Paleotragusgermaini Tragelaphus sp. A Tragelaphus sp. % Pachytragus sp. Hippotragini sp. TMiotragocerus sp. Kobus sp. Alcelaphini sp. no”. cf. Aepyceros sp. Gazella sp. A (large) Gazella sp. B (small) cf. Rhynchotragus Leptorycteropusguilielmi k”ububwaxeruspattersoni
This list is based on that given by Smart (1976) with some revisions based on more recent observations and publications. We have also received help from John Harris and Meave Leakey. Non-mammal taxa have been incorporated from Patterson, Behrensmeyer & Sill (1970). Carnivore identifications are based on advice from John Barry (pers. comm). Kubwaxeruspattersoni is a large Protoxerine rodent (Cifelli et al., 1986) formerly identified as an Anomalurid.
of N. juegeri, even if their stratigraphic position is consistent with the suggestion expressed
above, may appear inconclusive. However, even with this small number of N.jaegeri speci-
mens, the likelihood of the observed situation occurring randomly is statistically remote
(Wald-Wolfowitz Runs Test: n, =2, n2= 22, r= 2, x= -4.090, P=O.OOO). This test is also

448 A. HILL ET AL.
relevant to the possibility that the two specimens are derived from Lothagam 3, lying on the
surface of Lothagam 1 sediments. If this were the case, they are likely to occur randomly in
the stratigraphic sequence with respect to other pig specimens. That they do not occur
randomly in the sequence suggests that they are probably not derived.
Another possible problem is the apparent co-occurrence of &. syrticus and N. jaegeri at
another site, Ekora in Kenya (Harris & White, 1979), already discussed.
Thirdly, there is a possible problem internal to the Lothagam sequence. Three suid speci-
mens were not included in the discussion above. Two appear to belong to Nyanzachoerus
kanamensis, which is a species that in general also seems to occur distinctly from Jv. syrticus. In
the Tugen Hills succession, N. kanamensis is known only in the lower Chemeron Formation,
separated by the Kaparaina Basalts from occurrences ofJY. syrticus. Also, according to Harris
& White (1979), the two species only co-occur at Kanam West in Kenya and at Sahabi,
Libya. Kanam is a site where detailed stratigraphic relations and age have not yet been
studied closely. Maybe a span of time is represented there, and at Sahabi too. The situation at
Sahabi is not at present clear, especially as the species Nyan<achoerus devauxi is also found there.
Nyanzachoerus devauxi is the taxon that has been suggested as ancestral to later Nyankachoerus
species (Cooke & Wilkinson, 1978; Cooke, 1987). It has been known until recently from only
two sites, Sahabi, and Dublineau near Bou Hanifia, Algeria. The issue may ultimately be
clarified by its possible presence in the Baynunah Formation, Abu Dhabi, United Arab
Emirates (Hill et al., 1990; Whybrow et al., 1990). Regarding Jv. kanamensis at Lothagam,
however, one specimen, a left M, (KNM-LT 285), is recorded from Lothagam IC. It is
stratigraphically below the hominid (Table 1). The second specimen, KNM-LT 294, appears
to be even more anomalous, occurring at the level of KNM-LT 301 in Lothagam 1B. The
remaining specimen from Lothagam 1B alluded to above is KNM-LT 314, which may not
be N. syrticus, but neither is it N. jaegeri nor a. kanamensis (N. devauxi?), so is probably not
relevant here. It is not shown in Table 1, but it would appear at the level ofKNM-LT 294 and
KNM-LT 301.
It cannot be denied that these two Jv. kanamensis specimens detract from the argument.
Given all our other suid information, these two are anomalous and perhaps suggest, firstly,
that they are derived, or secondly, some mistake has been made in their stratigraphic allo-
cation. But if we concede this for these two suid specimens, we have to consider a similar
possibility for the specimens of JV.jaegeri, although given their location this is statistically less
likely, and we would even have to consider a similar possibility for the isolated hominid itself.
However, the principle of the argument here remains valid, and the evidence regarding Jv.
jaegeri is strongly suggestive, indicating that more robust conclusions could result from
detailed and deliberate collection of suids through Lothagam 1C. Lothagam is currently
being reinvestigated by Meave Leakey and her colleagues, and presumably relevant new
information will be forthcoming.
Age of hominid
This information provides us with an age datum within the Lothagam sequence, but where
does the hominid occur in relation to this apparent transition? The specimen is reported to
derive from high in Lothagam 1 C (Behrensmeyer 1976); this agrees with data in the unpub-
lished field notes. Three X syrticus specimens are found stratigraphically above it (Table 1). If this stratigraphic location of the hominid is accurate, then from our knowledge of the distri-
bution of JV. syrticus in the Tugen Hills succession, it would be greater than 5.6 Ma in age.

LOTHAGAM MANDIBLE 449
Very roughly, it occurs only about 10 m below the three N. yticus specimens, which are at
approximately the same horizon. If these parameters are correct, then it is probably only a
little older than 5.6 Ma. Our work on the Tugen Hills sequence (Deino et al., 1990) indicates
that sedimentation rates in the Ngorora Formation range from about 25 cm/1000 years to
5 cm/1000 years. Using these estimates and adopting the admittedly approximate values for
position in the succession, then it is unlikely that the hominid could be older than 6 Ma,
probably not older than 5.8 Ma. This is an estimate for the hominid specimen; it is likely that
the lower parts of the Lothagam 1 section are older than this.
The Tabarin mandible (Hill, 1985a; Ward & Hill, 1987) occurs in the lower Chemeron
Formation in association with N.jaegeri, and is now radiometrically dated at about 5 Ma.
The Chemeron humerus (Pickford et al., 1983; Hill & Ward, 1988) is in a comparable
stratigraphic position, also associated with N.jaegeri, and about the same age. At just greater
than 5.6 Ma, the Lothagam mandible would thus be the oldest member of the Hominidae so
far known.
Acknowledgements
This paper is part of the work of the Baring0 Paleontological Research Project (BPRP). It is
funded by grants to A.H. from the National Science Foundation (BNS 88-02629), from
the Louise Brown Foundation, the Ingalls Foundation, the Yale Social Sciences Faculty
Research Fund and Mr Clayton Stephenson. A.H. and S.W. also have a grant from the
L. S. B. Leakey Foundation, and S.W. receives additional funding from NSF grant BNS
8’7-18856. We thank Richard Leakey, at the time of this work Director of the National
Museums of Kenya, for permission to study the Lothagam mandible. Kay Behrensmeyer
most generously provided crucial unpublished stratigraphic data for some of the Lothagam
suid specimens. Alfreda Ibui and Mary Mongo gave valuable help in the analysis of the suid
specimens, as did Emma Mbua with the hominid. John Harris and Meave Leakey contri-
buted information to Table 2, and we thank Laura Bishop for her help. We thank the
Government of the Republic of Kenya for research permission.
References
Arambourg, C. & Coppens, Y. (1967). Sur la decouverte, dam le Pl&to&ne infkrieur de la vallCe de I’Omo (Ethiopie), d’une mandibule d’Australopith&%m. C. r. S&nc. Acad. Sci. Paris 265,589-590.
Behrensmeyer, A. K. (1976). Lothagam, Kanapoi and Ekora: a general summary ofstratigraphy and fauna. In (Y. Coppens, F. C. Howell, G. L. Isaac & R. E. Leakey, Eds) Earliest Man and Enuironments in the Lake Rudolf Basin, pp. 163-l 72. Chicago: University ofchicago Press.
Brown, B. (1989). The mandibles ofSiuapithecus. Ph.D. Dissertation, Kent State University, pp. 469. Brown, F. H., McDougall, I., Davies, T. & Maier, R. (1985). A n integrated Plio-pleistocene chronology for the
Turkana basin. In (E. Delson, Ed.) A ncestors: The Hard Evidence, pp. 82-90. New York: Alan R. Liss. Chapman, G. (1971). The geological evolution of the northern Kamasia Hills, Baring0 District, Kenya. Ph.D.
Dissertation, University of London. Chapman, G. & Brook, M. (1978). Chronostratigraphy ofthe Baring0 Basin, Kenya Rift Valley. In (W. W. Bishop,
Ed.) Geologicai Background to Fossil Man, pp. 207-223. London: Scottish Academic Press. Cifelli, R., Ibui, A. K., Jacobs, L. L. & Thorington, R. W. (1986). A giant tree squirrel from the Late Miocene of
Kenya. 3. Mammal. 67,274-283. Cooke, H. B. S. (1976). Suidae from Plio-pleistocenestrata ofthe Rudolfbasin. In (Y. Coppens, F. C. Howell, G. L.
Issac & R. E. F. Leakey, Eds) Earliest Man and Environments in the Lake Rudolf Basin, pp. 251-263. Chicago: University of Chicago Press.
Cooke, H. B. S. (1985). Plio-Pleistocene suidae in relation to African hominid deposits. In (Beden, M. et al., Eds) L’Enuironnement des Hominidbs au Plio-Pl&toc&e. Foundation Singer-Polignac, Paris: Masson.

A. HILL ET AL. 450
Cooke, H. B. S. (1987). Fossil suidae from Sahabi, Libya. In (N. Boaz, Ed.) NeNeogene Paleontology and Geology ofsahabi, pp. 255 -266. New York: Alan R. Liss.
Cddke, H. B. S. & Ewer, R. F. (1972). Fossil suidae from Kanapoi and Lothagam, northwestern Kenya, Bull. Mus. con@. <ool. Hare. 143, 144-296.
Cooke, H. B. S. & Maglio, V. (1972). Plio-Pleistocene stratigraphy in East Africa in relation to proboscidean and suid evolution. In (W. W. Bishop &J. A. Miller, Eds) Calibration of Hominoid Evolution, pp. 303-329. Edinburgh: Scottish Academic Press.
Cooke, H. B. S. & Wilkinson, A. (1978). Suidae and Tayassuidae. In (H. B. S. Cooke & V. Maglio, Eds) Evolution of Aftican Mammals, pp. 435-482. Cambridge: Harvard University Press.
Coppens, Y. (1971). Une nouvelle espkce de Suidi: du Villafranchien du Tunisie. &yNyanzachoerusjaegeri nov. sp. C.r. SLanc Acad. Sci. Paris ser. D. 272: 3264-3267.
Corruccini, R. S. & McHenry, H. M. (1980). Cladometric analysis of Pliocene hominids. 3. hum. Evol. 9,209-22 I,
Deino, A., Tauxe, L., Monaghan, M. & Drake, R. (1990). 40Ar/39Ar age calibration of the litho- and paleomagnetic stratigraphies ofthe Ngorora Formation, Kenya. 3. Geol. 98,567-587.
Eckhardt, R. B. (1977). Hominid origins: the Lothagam problem. Curr. Anthrop. 18,356.
Harris, J. M. & White, T. D. (1979). Evolution ofthe Plio-Pleistocene African Suidae. Trans. Am.phil. SK. 69, l-l 28. Hill, A. (1985a). Early hominid from Baringo, Kenya. Nature 315,222-224. Hill, A. (19856). Les variations de la faune au Miockne r¢ et du Pliocine d’Afrique de l’Est. L’Anthropologie 89,
275-279. Hill, A. (1987). Causes ofperceived fauna1 change in the later Neogene of East Africa. J. hum. Evol. 16,583-596. Hill, A., Drake, R., Tauxe, L., Monaghan M., Barry, J., Behrensmeyer, K., Curtis, G., Fine Jacobs, B., Jacobs, L.,
Johnson, N. & Pilbeam, D. (1985). Neogene palaeontology and geochronology of the Baring0 Basin, Kenya. 3. hum. Evol. 14,744-773.
Hill, A., Curtis, G. & Drake, R. (1986). Sedimentary stratigraphy of the Tugen Hills, Baringo, Kenya. In (L. E. Frostick, R. Renaut, I. Reid &J.-J. Tiercelin, Eds) Sedimentation in the African Rift System, pp. 285-295. Oxford: Blackwell, Geological Society of London.
Hill, A. & Ward, S. (1988). Origin ofthe Hominidae: the record ofAfrican large hominoid evolution between 14 my and 4 my. Yearb. phys. Anthrop. 31,49-83.
Hill, A., Whybrow, P. & Yasin al-Tikriti, W. (1990). Late Miocene primate fauna from the Arabian Peninsular: Abu Dhabi, United Arab Emirates. Am. 3. p/y. Anthrop. 81,240-241.
Hooijer, D. A. & Maglio, V. (1974). H’pp z arion from the late Miocene and Pliocene of northwestern Kenya. <ool.
Verhandel. 143, 1-34. Howell, F. C. (1978). Hominidae. In (V. J. Maglio & H. B. S. Cooke, Eds) EuolutionofAfricanMammals, pp. 154-248.
Cambridge: Harvard University Press. Johanson, D. & White, T. (1980). On the status ofAustralopithecuF afarensis. Science 207, 1104-I 105. Kotsakis, T. & Ingino, S. (1980). 0 sservazioni sui Nyanzachoerus (Suidae, Artiodactyla) de1 Terziaro superiori di
Sahabi (Cirenaica-Libyaj. Boll. Seru. Geol. d’ltalia. c(1979), 391408. Kramer, A. (1986) Hominid-pongid distinctiveness in the Miocene-Pliocene fossil record: the Lothagam mandible.
Am. J.phys. Anthrop. 70,457473. Leonardi, P. (1952). Resti fossili di Sivachoerus de1 giaccimento di Sahabi in Cirenaica (Africa Settentrionale). Atti
Acad. .Naz. Lincei Rend. Ser. 8, 13, 166-169. Leonardi, P. (1954). I suidi di Sahabi nella Sirtica (Africa Settentrionale). Rend. Accad. &az. dei XL Ser. 4,4,75-88. Maglio, V. (1970a). Four new species of Elephantidae from the Plio-Pleistocene of northwestern Kenya. Breuiora.
341, l-43. Maglio, V. (1970b). Early Elephantidae of Africa and a tentative correlation of African Plio-Pleistocene deposits.
Nature 225, 328-332. Maglio, V. (1973). Origin and evolution of the Elephantidae. Trans. Am. Phil. Sot. 63, l-149. Patterson, B., Behrensmeyer, A. K. & Sill, W. D. (1970). Geology ofa new Pliocene locality in northwestern Kenya.
Nature 2!?6,279-284. Pickford, M. (1978). Stratigraphy and mammalian paleontology of the late Miocene Lukeino Formation, Kenya.
In (W. W. Bishop, Ed.) Geological Background to Fossil Man, pp. 263-278. London: Scottish Academic Press. Pickford, M., Johanson, D., Lovejoy, O., White, T. & Aronson, J. (1983). A hominoid humeral fragment from the
Pliocene of Kenya. Am. 3. phys. Anthrop. 60,337-346. Smart, C. (1976). The Lothagam I fauna: its phylogenetic, ecological and biogeographic significance. In (Y.
Coppens, F. C. Howell, G. L. Isaac & R. E. Leakey, Eds) Earliest Man and Environments in the Lake RudolfBasin, pp. 361-369. Chicago: University of Chicago Press.
Ward, S. & Hill, A. (1987). Pliocene hominid partial mandible from Tabarin, Baringo, Kenya. Am. J.phys. Anthrop. 72,21-37.
Ward, S., Johanson, D. & Coppens, Y. (1982). Subocclusal morphology and alveolar process relationships of hominid gnathic elements from the Hadar Formation: 1974-1977 collections. Am. 3. phys. Anthrop. 57,605-630.
Ward, S. & Molnar, S. (1980). Experimental stress analysis of topographic diversity in early hominid gnathic morphology. Am. 3. phys. Anthrop. 53,383-395.

LOTHAGAM MANDIBLE 451
White, T. D. (1977). New fossil hominids from Laetolil, Tanzania: 1976-1979 specimens. Am. J. phys. Anthrop. 46, 197-230.
White, T. D. (1980). Additional fossil hominids from Laetoli, Tanzania: 1976-1979 specimens. Am. J.phys. Anthrop. 53,487-504.
White, T. D. (1986). Australopithecus afaremis and the Lothagam mandible. Anthropos (‘Bmo). 23, 79-90. White, T. D. & Johanson, D. C. (1982). Pliocene hominid mandibles from the Hadar Formation, Ethiopia: 1974-
1977 collections. Am. J.phys. Anthrop. 57,501-544. White, T. D., Johanson, D. C. & Kimbel, W. H. (1981). Australopithecusafriccanus: its phyletic position reconsidered.
5’. Afr. 3, Sci. 77,445-470. Whybrow, P., Hill, A., Yasin al-Tikriti, W. & Hailwood, E. (1990). Late Miocene primate fauna, flora and initial
paleomagnetic data from the Emirate ofAbu Dhabi, United Arab Emirates. 3. hum. Ed. 19,583%588.