
Trap architecture in carnivorous Utricularia (Lentibulariaceae)
Upload
independentCategory
view
1download
0
Physiological Entomology(1998)23, 227–240
Trap-jaws revisited: the mandible mechanism of theant Acanthognathus
W U L F I L A G R O N E N B E R G , C . R O B E R T O F. B R A N D A˜ O , *B O D O H . D I E T Z * and S T E FA N J U S TTheodor-Boveri-Institut der Universita¨t, Lehrstuhl fur Verhaltensphysiologie und Soziobiologie, Wu¨rzburg, Germany, and
*Museu de Zoologia, Universidade de Sa˜o Paulo, Brazil
Abstract. Ants of the genusAcanthognathusstalk small insects and catch their preyby a strike with their long, thin mandibles. The mandibles close in less than 2.5 msand this movement is controlled by a specialized closer muscle. InAcanthognathus,unlike other insects, the mandible closer muscle is subdivided into two distinct parts:as in a catapult, a large slow closer muscle contracts in advance and provides thepower for the strike while the mandibles are locked open. When the prey touchesspecialized trigger hairs, a small fast closer muscle rapidly unlocks the mandibles andthus releases the strike. The fast movement is steadied by large specialized surfacesin the mandible joint and the sensory-motor reflex is controlled by neurones withparticularly large, and thus fast-conducting, axons.
Key words. Acanthognathus, catch mechanism, fast movement, functionalmorphology, mandible joint, movement control, trap jaws.
Introduction
Speed of action is of obvious advantage for an animal’sevolutionary fitness, whether it is a fast escape manoeuvre, adefensive response or a quick predatory mechanism.Accordingly, there exists a kind of evolutionary arms race inwhich prey and predators try to outrun or overpower eachother by developing ever faster mechanisms. Among the insectsthis has led to astonishing and sometimes bizarre adaptations,such as the leaping jumps of fleas or the inescapable strike ofthe praying mantis (see Alexander, 1988; Gronenberg, 1996a).The ants do not depend on fast escape mechanisms because,as highly social insects, they can rely on the power of theirunited defence and on their stings or noxious secretions.However, some predatory ant genera are equipped with veryfast and highly effective weapons, so-called trap-jaws.
Trap-jaws are mandibles that snap together very rapidly andthat are well suited to catch fast or elusive prey or to attackarmoured or otherwise protected or defensive arthropods. Suchtrap-jaws have repeatedly and independently evolved in threeant subfamilies: the ponerine generaOdontomachusandAnochetus, the formicine genusMyrmoteras, and the myrmicine
Correspondence: Wulfila Gronenberg, Theodor-Boveri-Institut,Lehrstuhl fur Verhaltensphysiologie und Soziobiologie der Universita¨t(Zoologie II), Am Hubland, D-97074 Wu¨rzburg, Germany. E-mail:[email protected]
© 1998 Blackwell Science Ltd 227
tribe Dacetonini, with twenty-four genera (Brown, 1976, 1978;Masuko, 1984; Moffett, 1986). In the ponerines and in twodacetonine genera, the trap-jaw mechanism has been found tobe very similar: as in a catapult, muscular energy is stored inadvance of the mandible strike and is very rapidly releasedwhen particular trigger hairs make contact with the prey. Themechanism is controlled by neurones with very large, and thusfast propagating, axons and specialized muscles (Gronenberget al., 1993; Gronenberg, 1995a,b, 1996b; Gronenberg &Ehmer, 1996).
The Dacetonini form a cosmopolitan tribe of myrmicine antsinhabiting the floor of tropical and subtropical forests. Mostdacetonines ambush and stalk small arthropods, particularlycollembolans, with their mandibles specialized for differentactions. Brown & Wilson (1959) inferred the evolutionarytendencies within the Dacetonini, based on a comparative studyof the morphology, behaviour and habits of the extant species,suggesting that they progressively abandoned their subarborealepigaeic foraging customs in favour of more cryptic and‘terrestrial’ habits. During this evolution the trap-jaws havebeen modified, reduced and even secondarily redeveloped tosuit the particular ecological needs of their bearers (Brown &Wilson, 1959; Ho¨lldobler & Wilson, 1990). Generalized generathen would be oligophagic predators, whereas derived oneswould prey exclusively on collembolans. This tendency wouldbe accompanied by a reduction in overall size, especially themandibles. Accepting this hypothesis,Acanthognathus, withits long and thin mandibles, forms one of the most generalized

228 Wulfila Gronenberget al.
dacetonine genera and should not be specialized on collembolanprey. Dietz & Branda˜o (1993) studied the prey catchingbehaviour of A. rudis and confirmed that they do acceptarthropods other than collembolans, at least under laboratoryconditions. However, in a recent classification proposed forthe Dacetonini, Baroni Urbani & de Andrade (1994), unlikeBrown & Wilson (1959), suggested thatAcanthognathusis infact one of the most specialized genera of the tribe. So theevolution of the dacetonine ants and their mandibles is still amatter of debate.
It was for the dacetonine ants (genusStrumigenys) that acatch mechanism has been proposed for the first time tounderlie the trap-jaw action (Brown & Wilson, 1959). The firstdescription of the muscular action which generates the mandiblestrike is also based on dacetonine ants (genusAcanthognathus,Dietz & Brandao, 1993). Dietz & Brandao (1993) did notpropose a catapult mechanism but suggested that the mandiblestrike is based on rapid contraction and relaxation, respectively,of partly antagonistic mandible closer muscles. Working onthe dacetine generaDaceton and Strumigenys, Gronenberg(1996b), however, found evidence for a catapult mechanismwhich involves energy storage and a catch device. Withouthaving examinedAcanthognathus, Gronenberg (1996b) thoughtit unlikely that two very different trap-jaw mechanisms shouldhave evolved within the same ant tribe. In the present study,the authors of the opposing assumptions together re-examinethe mandible mechanism ofAcanthognathusand try to resolvethe contradictions.
Materials and Methods
Part of a colony (ten workers) ofAcanthognathus rudis(genusrevised by Brown & Kempf, 1969; for taxonomic affinities seeBaroni Urbani & de Andrade, 1994) was collected at theBoraceia Biological Station, Saleso´polis county, Sa˜o Paulostate, Brazil, in primary evergreen cloud forest by Bodo H.Dietz and Christiane I. Yamamoto between 12 and 17 June1997. The ants were compared with specimens of the collectionin the Museo de Zoologia, Sa˜o Paulo, where voucher specimenshave also been deposited. The ants were kept in a moist plasterof Paris nest at 25°C and LD 12:12 h in the laboratory andwere fed on live collembolans.
The behaviour of unrestrained ants was observed in theirforaging arena and was videotaped with standard videoequipment. The mandible movements were further analysedfrom close-up videographs of tethered ants using a high-frequency video camera (NAC HSV400) at 400 frames/s andeither electronic strobe light (strobe flash duration 20µs) or atungsten light source. Ants were waxed to a needle by theirthorax and the defensive mandible snap was released bytouching the antennae and trigger hairs with a pin.
Morphology
The morphology of the head and mandibles was examinedby scanning electron microscopy of air-dried specimens using
© 1998 Blackwell Science Ltd,Physiological Entomology, 23, 227–240
a Zeiss DSM 962 SEM. The internal organization of muscles,apodemes, and nerves within the head was studied fromaldehyde-fixed, plastic-embedded (Durcupan: Fluka, Buchs,Switzerland), and serially sectioned material (3–10µm) stainedwith osmium-ethyl gallate or with methylene blue (Gronenberg,1995a). In addition, sensory afferent axons originating frommandibular sensilla, and motor neurones supplying a dorso-lateral part of the mandible closer muscle, were labelled witha fluorescent dextran tracer (Fluoro Ruby, Molecular Probes,Eugene, OR). The tracer was applied to the stumps of the cutsensilla or muscle fibres and after diffusion periods of 2–6 h,the heads were processed according to standard routines(paraformaldehyde fixation, plastic embedding, epifluorescencemicrography). Graphical reconstruction and microscopicalmeasurements were made using a calibrated camera lucidaattachment to the microscope.
To evaluate the volume of muscles, the structures weretraced on paper using a camera lucida. The respective muscleoutlines were then cut out and the paper pieces weighed.Assuming homogenous weight of the stock paper, the overallvolume of the muscles was thus calculated from the weight ofa reference piece of paper (500 cm2), the section thickness,and the initial magnification.
To assess the number of muscle fibres, each fibre wasindividually traced from section to section and the fibres werethus counted in the smaller muscles. For the large closermuscle, however, all fibres were counted when they insertedat the cuticle. To avoid multiple counting when the region ofcuticle attachment of the same fibre was present in severalsections, this value was corrected by the quotient of mean fibrethickness and section thickness (e.g. fibre thickness 10µm,section thickness 5µm, fibre number corrected by factor 0.5).The sarcomere length of the muscle fibres was measured usingthe camera lucida. As a result of local fibre contraction duringfixation, the sarcomeres were not always of uniform length ina given fibre. To overcome this artefact, the sarcomere lengthwas assessed only from those fibres that could be completelyfollowed within a single section, hence the different contractionstate of the sarcomeres (long and short ones within the samefibre) would cancel out rather than bias the measurement.
Results
Prey-catching behaviour
Acanthognathusgenerally ambush their prey and stalk itvery slowly. In general they keep their mandibles opened mostof the time, even within the nest, in contrast toStrumygenys,which open their mandibles only in the presence of prey (orprobably prey odour) or for defensive purposes. Before the prey-catching behaviour is released byAcanthognathus, antennalcontact with the putative prey is required. Many videosequences were obtained in which, in the absence of previousantennal contact, collembolans virtually walked over orunderneath the ants without being detected. A brief chanceantennal contact with the prey (or with putative chemical cues)will start searching behaviour by the ant. This may take several

Trap-jaw mechanism 229
Fig. 1. Video images (25 frames/s) selectedfrom a single sequence in which the antapproaches, catches and stings a springtail.Numbers give the time elapsed since the startof the sequence (top left: 0 ms) in ms (0–2040) or s (5–14). The strike occurs withinframe 1720 (bottom left), in which theoriginal record showed a faint double-imageof the mandibles. In this frame, the imagecontrast of the mandible region (arrow) hasbeen manipulated in order to reveal thisdouble-image in print.
© 1998 Blackwell Science Ltd,Physiological Entomology, 23, 227–240

230 Wulfila Gronenberget al.
Fig. 2. Schematic representation of the mandible movements as revealed by high-frequency videography. Boxed region of the head on the leftindicates area shown on the right. The opening movement (top right) is slow and takes several video frames; after completion of about 2/3 of theirtrajectory the mandibles are rotated upward around their long axis by about 20° (indicated by a bump in the arrows, symbolising mandiblemovement). Note that mandibles can open even wider when manipulating objects with their accessory processes. The fast closing strike (bottomright) occurs within a single video frame and is preceded by a downward rotation around the mandible’s long axis. Arrows indicate directions ofmovements or rotations, respectively; note that distal mandible teeth point slightly upward before but medially during the strike.
seconds and is shown by the first two frames in Fig. 1 wherethe ant orients towards the collembolan. In the third frame(600 ms) antennal contact with the prey is established and theant then centres her body and particularly her mandibles withrespect to the prey (Fig. 1, frames 600–1080 ms), probablyusing proprioceptive information from the antennal position.Without a further step she then very slowly approaches theprey (frames 1400, 1560 ms) until the prey is touched by thetrigger hairs, long bristles protruding anteriorly from each ofthe opened mandibles (Fig. 2). During this time, the antennaeare withdrawn (between frames 1080 and1400 ms in Fig. 1),which may protect the ant’s most important sense organs frompossible damage by the struggling prey once it is caught. Thefinal approach may take more than 1 s (about 300 ms in Fig. 1,frames 1400–1720 ms), a long time for a swift collembolanto escape.
The strategy of the ant is to disturb the prey as little aspossible. The slight antennal contact arouses the springtail andit may start walking away (compare frame 600 and 760 ms inFig. 1), but it does not release the escape jump of thecollembolan. The very slow final approach of the ant in advanceof the mandible strike seems to serve the same purpose: toavoid any telltale air turbulences which might startle thespringtail immediately before the final assault. The trigger hairs
© 1998 Blackwell Science Ltd,Physiological Entomology, 23, 227–240
serve as a range finder, and once the prey is close enough andwithin reach of the mandibles, it is touched by the hairs andthe strike is very rapidly released.
This speed is demonstrated by the double-image of themandibles in Fig. 1, frame 1720 ms: this frame shows themandibles in the open, as well as in the closed, position withno blur in between, indicating that the time during which themandibles actually move is very short compared to the durationof a video frame (40 ms). Although this finding is notremarkable in itself, the same effect was found in a fewfortuitous frames of the high-frequency video sequences, someof which were taken without a strobe light. In those imagesthe mandibles were ‘frozen’ in the open and closed positionsin the same frame, i.e. within 2.5 ms. As in the normal video(Fig. 1, frame 8), the mandible in the closed position wasbarely visible, indicating that it had been exposed much morebriefly than in the open position. Hence, the mandible movementtakes only a fraction of the entire frame duration, most likelysubstantially less than 1 ms.
Once caught by the fang-like apical teeth of the mandibles,even much larger prey items than the one depicted in Fig. 1did not escape the ant. In the sequence depicted in Fig. 1 theant starts to sting the springtail after about 4 s because theprey then started to wriggle. The entire stinging procedure

Trap-jaw mechanism 231
took more than 6 s, and when stung the prey stopped moving.Larger prey items were stung immediately after the mandiblestrike.
The immobilized collembolans were then carried to adifferent location, often in the vicinity of the nest area. Therethe springtails were kneaded and stroked with the forelegs andthe mandible’s basal teeth (Dietz & Branda˜o, 1993), which arehere referred to as ‘accessory processes’ of the mandibles andwhich are shown in Figs 2, 3a,c,d. These processes are usedto perform ‘normal’ actions for which the long mandibleswould be an awkward hindrance. The prey is kneaded for along time (for several minutes) in a manner appropriate tosqueeze the liquid out of the prey. This liquid is taken up bythe ant with her mouthparts, hence the prey is not eaten entirelyand its husk remains almost intact.
The mandible movement
The movement of the mandibles has been revealed byhigh-frequency videography of tethered ants. At the highmagnification necessary, the optical resolution and the depthof field of the video image were very restricted. Thus, originalhigh-frequency videographs are not shown, but a compositedrawing summarizes these findings (Fig. 2). The openingmovement is relatively slow. It took at least four videoframes (10 ms), but generally considerably longer, to open themandibles to an inter-mandibular angle of 150°, as depictedon the left of Fig. 2. In this position the mandibular triggerhairs point forwards and towards the prey. When closed, themandibular trigger hairs fold into a recession in the mandible(Fig. 3f). The mandibles can be opened even wider, up to aninter-mandibular angle of about 210°, when the ants handleprey, brood or other items with their accessory mandibularprocesses.
High-frequency videographs revealed that during openingand at an inter-mandibular angle ofµ 110°, the mandiblesrotate about 20° around their long axis (direction indicated byarrows in the top right of Fig. 2). Both mandibles can beopened independently and at slightly different velocities, butfor each mandible the position at which the rotation around itslong axis occurs appears to be fixed. Likewise, faster openingmovements seem to be accompanied by faster rotations, andduring the fast openings the rotation occurred within a singleframe (2.5 ms). This suggests (but does not prove) that themandible rotation may result from some mechanical ‘cam’within the mandible joint, rather than from the contraction ofa particular muscle. As will become clear in the section onmuscle morphology, no muscle was found which would havethe appropriate position and temporal properties to generatesuch a fast rotation of the mandible.
This upward rotation of the mandible during the openingmovement is crucial for the mandible strike and its release.Prior to the strike, the mandibles are held at an angle of about150°. In this position the two accessory processes at themandible base, which under normal conditions are used tohandle prey and other items, meet and interlock (Fig. 2) whilethe apical teeth (the fangs) point slightly upwards rather than
© 1998 Blackwell Science Ltd,Physiological Entomology, 23, 227–240
directly towards the prey. The two interlocked accessoryprocesses effectively prevent the mandibles from closing, evenif the powerful mandible closer muscles contract. In order torelease the mandible strike from the locked position, the twoaccessory processes need to be disengaged. This disconnectionof the accessory processes results from their ventrolateralrotation which, in turn, is a consequence of mandible rotationsaround their long axes in a direction opposite to the rotationwhich occurs during opening.
Hence, a mechanism in which mandibles are locked priorto the strike is suggested. The strike is released by a slightdownward rotation of the accessory processes which may thenslide along each other (this situation is depicted in Fig. 3c,d).Although more than fifty snapping movements were analysedat 400Hz, only a few cases were found in which the mandibleswere slightly blurred in the frame preceding the mandibleclosure in a way that could be interpreted as the suggesteddownward rotation. The authors believe that the strike-releasingmandible rotation occurs within much less than 2.5 ms beforethe mandible strike and thus generally within the same videoframe. Subsequently the mandibles are accelerated and snapshut. This entire sequence of events takes less than 2.5 ms,probably less than 1 ms, as explained above. No known muscleis able to contract in such a short time, thus an energy storagemechanism is likely to be involved.
Before the mandibles close entirely, they are decelerated.Although almost the entire strike happens within a single high-frequency video frame, the last 10–15° of the movement mayextend over several frames. Under natural conditions, themandibles do not close so tightly because the prey will becaught between them and prevent further closure. So thedeceleration may usually not occur. If, however, the strike failsand no prey is caught between the striking mandibles, thebraking of the mandibles would protect them from damagethrough collision.
Mandible morphology
Some features of the mandibles ofAcanthognathushavealready been mentioned (Fig. 3): they are very long (longerthan the head width) and slender and bear a three-pronged tipwith long and fang-like apical teeth which penetrate thecollembolan prey. The head, mandibles and antennae arecovered by hairs and bristles of varying length (10–40µm),some of which (most notably those covering the mandibles)are club-like (claret hairs). Comparison with other insects, andants in particular, suggests that these hairs are mechanoreceptors(except for the many chemoreceptor hairs located on the distalantennal segments and on the mouthparts). Some of these hairsensilla are proprioceptors which assess the position of thelimbs (hair-fields on the mandible base or on the antennal base;asterix in Fig. 3h), while one very long hair on each mandibleserves as a trigger hair and releases the strike upon contact(Figs 2, 3g). The trigger hair has a shaft diameter of about3.5µm at its base and is supplied by a single large sensorycell (diameter of cell body: 12µm, of axon: 2.5µm) whichresides close to the hair base at the inner mandible wall.

232 Wulfila Gronenberget al.
Fig. 3. Scanning electron micrographs of the head and mandibles ofAcanthognathusat different magnifications; (a) dorsal view of the head withpartly opened mandibles; (b) lateral view of the frontal half of the head [antennae removed; boxed region enlarged in (k)]; (c) dorso-frontal viewof the mandibles (arrowhead indicates trigger hairs); (d) enlargement of area boxed in (c) showing the accessory mandibular processes and thebasal tooth (arrowhead); (e) pronged tip of the right accessory process boxed in (d); (f) basal tooth with sensory hair (arrowhead) and putativecampaniform sensillum (double arrowhead); (g) trigger hair insertion at the inner face of the mandible; (h) head capsule region where the rightmandible inserts (mandible and other mouthparts removed to reveal joint); asterisk indicates antennal base, arrowhead points at outer joint surfaceand double arrowhead points at inner joint surface); (i) right mandible base separated from head capsule to reveal joint region; arrowhead anddouble arrowhead, respectively, point at regions which slide across the outer or inner joint surface of the head capsule, respectively; to allowcomparison of the different views of the mandible base, asterisks in (i) and (k) mark approximately the same points; (k) lateral view of intact rightmandible joint (mandibles closed); arrows point at cuticular protrusions of the head capsule which supposedly serve as cams and force an upwardrotation onto the trajectory of the mandible opening movement; circles approximately indicate points where the asterisks on the rim of the mandiblebase would contact the head capsule if the mandibles were open; scale bars 250µm (a – c), 50µm (d), 25µm (g – k), 10µm (e, f).

Trap-jaw mechanism 233
The mandibles are asymmetric in that only the right onecarries a peg-like basal tooth close to the mandible base (arrowin Fig. 3d). This peg is supplied by two sensory neurones, thedendrites of which contact two cuticular structures: a hairand a putative campaniform sensillum (single and doublearrowhead, respectively, in Fig. 3f). Hence, it is assumed thatthe basal tooth serves a sensory function. It is touched by theopposite mandible’s accessory process when the mandibles areclosed and may control the closing movement and preventdamage to the closed mandibles by excessive force generatedby the closer muscle. In addition, the basal tooth might aid themanipulation of objects by the accessory processes. When themandibles are closed, the left accessory process rests over theright one, its dorsal surface touching the sensillum of the rightmandible’s basal tooth. In this position, the right accessoryprocess does not touch the left mandible and that is possiblywhy only one of the mandibles is equipped with a basal tooth.
The most important structures in the present context are theaccessory processes (Fig. 3a,c,d,e). When the mandibles close,the process of the right mandible will always slide underneaththat of the left one, which originates slightly more proximalon its mandible than the right one (Fig. 2, inset) which carriesthe basal tooth. Hence, allAcanthognathusare ‘right-handed’.In the closed and half-open position the accessory processespoint slightly downwards, but beyond an inter-mandibularangle of about 110° they are rotated upwards so that their tipsface each other. In this position they are able to handle objects(hence accessory processes). Moreover, at an angle of 150°the tips of the processes meet and can interlock. As shown inFig. 3e, the tips of the processes comprise two slim teethand one opposing wide tooth with a rectangular tip. Uponinterlocking, the rectangular tooth of the right mandible ridesbetween the two slim teeth of the left one in a delicate balance.While in this position the two opposing processes are pressedtogether by the forces acting on the mandibles and form asemi-stable junction, a change in the angle between the twoprocesses lets their tips slip across each other and thus slightdownward rotation unlocks the mandible catch.
To ensure the proper function of this sensitive mechanism,the mandibles require a firm support which prevents anywobbling that might accidentally trip the strike. At the sametime the support has to permit the slight rotation around thelong axis and the wide closing movement. In engineering, suchrequirements would be met by a slide bearing which consistsof large smooth surfaces sliding across each other. InAcanthognathusthe same design is revealed when themandibles are detached from the head capsule. Figures 3h andk show the large cuticular surfaces across which the mandiblebases slide. These parts of the head capsule are devoid of hairsand other textures and appear almost polished, which wouldreduce friction, yet they are not evenly plane surfaces. Rather,they contain shallow grooves and protrusions (Fig. 3h,arrowheads in 3k). We assume that these cuticular protrusionsact like ‘cams’, as the rim of the mandible base (Fig. 3i, k)slides across them. Together, the rim of the mandible base andthe cuticular humps of the sliding surface probably form amechanical guide system that slightly rotates the mandibles.We have tried to indicate this rotation in Fig. 3k. This figure
© 1998 Blackwell Science Ltd,Physiological Entomology, 23, 227–240
shows a row of asterisks on the base of the closed mandible.As the mandible opens, the rim of its base slides across thejoint surface of the head capsule. In the open position the threeasterisks would come to sit adjacent to the points marked bycircles on the joint surface. The difference in the dorso-ventralposition between the row of asterisks on the mandible and therow of circles on the head capsule surface indicates that aslight tilt around the long axis of the mandible has to occurfor the mandible base to occupy this position. This movementis forced upon the mandible by the ‘cams’ indicated byarrowheads in Fig. 3h. An upward rotation is thus superimposedonto the opening trajectory of the mandibles.
The exterior smooth surfaces (arrowheads in Fig. 3h)stabilize the mandibles when the latter are partly or completelyopen. When the mandibles are almost closed, however, theirbases contact these external surfaces only at two points at theirrim (Fig. 3k). In order to steady mandible movements at smallinter-mandibular angles, too, a second set of surfaces isemployed. These comprise a smaller interior smooth surfacewithin the head capsule (partly visible and indicated by doublearrowheads in Fig. 3h) and the roll-like rim (double arrowheadsin Fig. 3i) of an outward-folded part of the mandible base.When the mandibles are closed or only partly opened, thisroll-like rim of the mandible base is pressed against and slidesacross the opposing interior smooth surface of the head capsule(Fig. 3h). Hence, the large amount of opposed sliding surfacesnecessary for stabilization of the mandibles is divided into aninterior and an exterior region. The mandibles ofAcanthognathusare thus stabilized much more elaborately thanthose of other ants, so ensuring rigid control over the widerange of movements.
Internal morphology
As is the case in other ants, and in other trap-jaw ants inparticular, the mandible closer muscle occupies the largest partof the head volume (Fig. 4a-c). This muscle generates andcontrols all the mandibular power, whereas the mandible openermuscle is much smaller (Fig. 4a,c; Table 1) and functionallyless significant. Next to the closer muscle, the brain takes upan important part of the head. This is the case in all small antswhich always have comparatively large brains, whereas inlarge ants the head muscles are more prominent (larger antshave relatively smaller brains).
Unlike any other ant so far histologically examined, and ascorrectly described by Dietz & Branda˜o (1993),Acanthognathushas two different mandible closer muscles oneither side which we will refer to as the slow muscle and thefast muscle (Fig. 4b,c). The slow closer muscle is very large,whereas the fast one is about 10-fold smaller, as is the openermuscle (Table 1). Slow and fast closer muscles connect to thesame apodeme and are thus derived from a single muscle, thecommon mandible closer muscle of other ants. The mandiblecloser apodeme forms a Y-shaped structure (Fig. 4). Each ofthe two muscles connects to a different branch of the ‘Y’(Fig. 4) while the third branch of the apodeme connects themuscle to the mandible base (Fig. 4a). The two branches which

234 Wulfila Gronenberget al.
Fig. 4. Schematic representation of the head ofAcanthognathus; sagittal (c) and frontal (a, b, d, e) views; (a), (d) 300µm and (b), (e) 50µm fromthe dorsal head surface, respectively; (d) and (e) show details of the anterior head region; ap5 mandible closer apodeme, br5 brain, fc5 fast,sc5 slow mandible closer muscle, m5 mandible, o5 mandible opener muscle, sog5 suboesophageal ganglion; scale bar 100µm (a – c), 50µm(d, e).
receive the muscle fibres are rigid and sclerotised (dark-brownin unstained sections), whereas the branch that connects tothe mandible is more flexible and tendon-like (unstained inmicroscopical sections) as indicated by the stippled renderingin Fig. 4a,d. This terminal apodeme branch is not circular but
© 1998 Blackwell Science Ltd,Physiological Entomology, 23, 227–240
elongate in cross-section and thus has the shape of a thick andsturdy ribbon. The lateral sclerotised arm of the apodeme isshort and receives the fast closer muscle fibres, whereas theother sclerotised arm projects through the entire head as itconnects to the bulk of the slow closer muscle fibres.
Trap-jaw mechanism 235
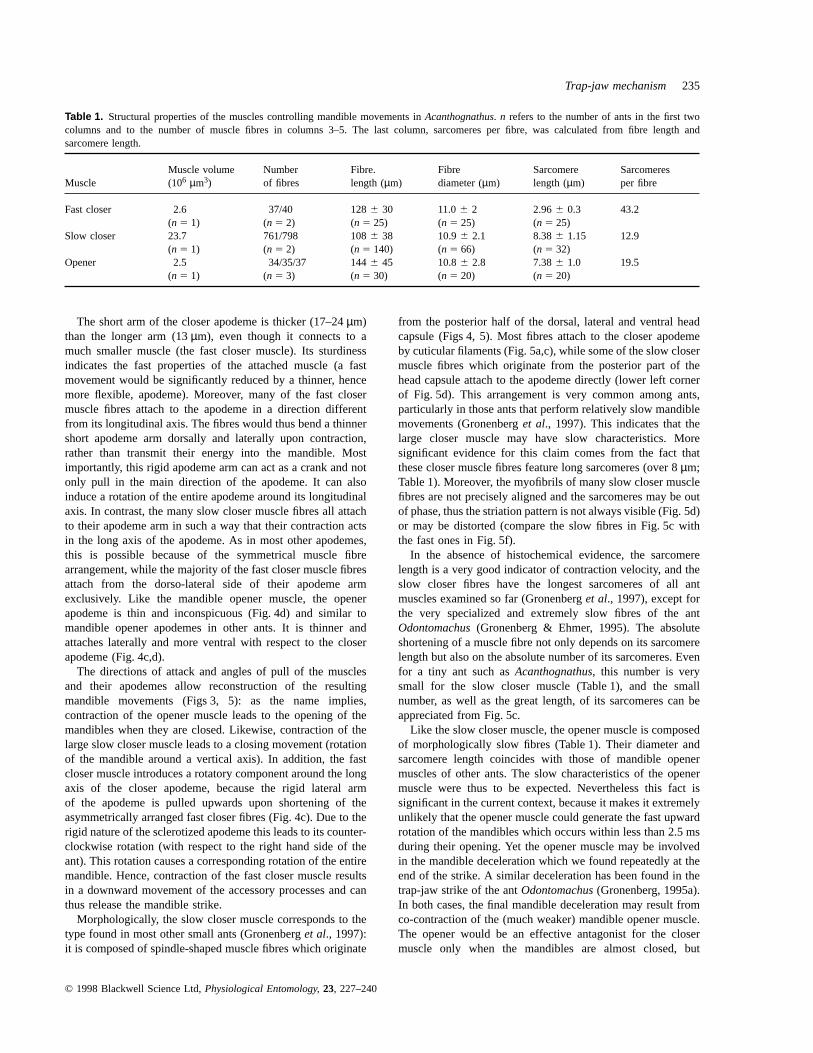
Table 1. Structural properties of the muscles controlling mandible movements inAcanthognathus. n refers to the number of ants in the first twocolumns and to the number of muscle fibres in columns 3–5. The last column, sarcomeres per fibre, was calculated from fibre length andsarcomere length.
Muscle volume Number Fibre. Fibre Sarcomere SarcomeresMuscle (106 µm3) of fibres length (µm) diameter (µm) length (µm) per fibre
Fast closer 2.6 37/40 1286 30 11.06 2 2.966 0.3 43.2(n 5 1) (n 5 2) (n 5 25) (n 5 25) (n 5 25)
Slow closer 23.7 761/798 1086 38 10.96 2.1 8.386 1.15 12.9(n 5 1) (n 5 2) (n 5 140) (n 5 66) (n 5 32)
Opener 2.5 34/35/37 1446 45 10.86 2.8 7.386 1.0 19.5(n 5 1) (n 5 3) (n 5 30) (n 5 20) (n 5 20)
The short arm of the closer apodeme is thicker (17–24µm)than the longer arm (13µm), even though it connects to amuch smaller muscle (the fast closer muscle). Its sturdinessindicates the fast properties of the attached muscle (a fastmovement would be significantly reduced by a thinner, hencemore flexible, apodeme). Moreover, many of the fast closermuscle fibres attach to the apodeme in a direction differentfrom its longitudinal axis. The fibres would thus bend a thinnershort apodeme arm dorsally and laterally upon contraction,rather than transmit their energy into the mandible. Mostimportantly, this rigid apodeme arm can act as a crank and notonly pull in the main direction of the apodeme. It can alsoinduce a rotation of the entire apodeme around its longitudinalaxis. In contrast, the many slow closer muscle fibres all attachto their apodeme arm in such a way that their contraction actsin the long axis of the apodeme. As in most other apodemes,this is possible because of the symmetrical muscle fibrearrangement, while the majority of the fast closer muscle fibresattach from the dorso-lateral side of their apodeme armexclusively. Like the mandible opener muscle, the openerapodeme is thin and inconspicuous (Fig. 4d) and similar tomandible opener apodemes in other ants. It is thinner andattaches laterally and more ventral with respect to the closerapodeme (Fig. 4c,d).
The directions of attack and angles of pull of the musclesand their apodemes allow reconstruction of the resultingmandible movements (Figs 3, 5): as the name implies,contraction of the opener muscle leads to the opening of themandibles when they are closed. Likewise, contraction of thelarge slow closer muscle leads to a closing movement (rotationof the mandible around a vertical axis). In addition, the fastcloser muscle introduces a rotatory component around the longaxis of the closer apodeme, because the rigid lateral armof the apodeme is pulled upwards upon shortening of theasymmetrically arranged fast closer fibres (Fig. 4c). Due to therigid nature of the sclerotized apodeme this leads to its counter-clockwise rotation (with respect to the right hand side of theant). This rotation causes a corresponding rotation of the entiremandible. Hence, contraction of the fast closer muscle resultsin a downward movement of the accessory processes and canthus release the mandible strike.
Morphologically, the slow closer muscle corresponds to thetype found in most other small ants (Gronenberget al., 1997):it is composed of spindle-shaped muscle fibres which originate
© 1998 Blackwell Science Ltd,Physiological Entomology, 23, 227–240
from the posterior half of the dorsal, lateral and ventral headcapsule (Figs 4, 5). Most fibres attach to the closer apodemeby cuticular filaments (Fig. 5a,c), while some of the slow closermuscle fibres which originate from the posterior part of thehead capsule attach to the apodeme directly (lower left cornerof Fig. 5d). This arrangement is very common among ants,particularly in those ants that perform relatively slow mandiblemovements (Gronenberget al., 1997). This indicates that thelarge closer muscle may have slow characteristics. Moresignificant evidence for this claim comes from the fact thatthese closer muscle fibres feature long sarcomeres (over 8µm;Table 1). Moreover, the myofibrils of many slow closer musclefibres are not precisely aligned and the sarcomeres may be outof phase, thus the striation pattern is not always visible (Fig. 5d)or may be distorted (compare the slow fibres in Fig. 5c withthe fast ones in Fig. 5f).
In the absence of histochemical evidence, the sarcomerelength is a very good indicator of contraction velocity, and theslow closer fibres have the longest sarcomeres of all antmuscles examined so far (Gronenberget al., 1997), except forthe very specialized and extremely slow fibres of the antOdontomachus(Gronenberg & Ehmer, 1995). The absoluteshortening of a muscle fibre not only depends on its sarcomerelength but also on the absolute number of its sarcomeres. Evenfor a tiny ant such asAcanthognathus, this number is verysmall for the slow closer muscle (Table 1), and the smallnumber, as well as the great length, of its sarcomeres can beappreciated from Fig. 5c.
Like the slow closer muscle, the opener muscle is composedof morphologically slow fibres (Table 1). Their diameter andsarcomere length coincides with those of mandible openermuscles of other ants. The slow characteristics of the openermuscle were thus to be expected. Nevertheless this fact issignificant in the current context, because it makes it extremelyunlikely that the opener muscle could generate the fast upwardrotation of the mandibles which occurs within less than 2.5 msduring their opening. Yet the opener muscle may be involvedin the mandible deceleration which we found repeatedly at theend of the strike. A similar deceleration has been found in thetrap-jaw strike of the antOdontomachus(Gronenberg, 1995a).In both cases, the final mandible deceleration may result fromco-contraction of the (much weaker) mandible opener muscle.The opener would be an effective antagonist for the closermuscle only when the mandibles are almost closed, but

236 Wulfila Gronenberget al.
Fig. 5. Photomicrographs (osmium stained frontal sections) of the head ofAcanthognathus; planes of sectioning in (a) and (b) correspond to (a)and (b), respectively, of Fig. 4. (c), (d) slow, (e), (f) fast closer muscle fibres (c) and (f) methylene blue-stained); ap5 mandible closer apodeme,br 5 brain, cu5 cuticle of the head capsule, ey5 eye, fc5 fast, sc5 slow mandible closer muscle, sog5 suboesophageal ganglion; scale bars 50µm.
in neither case (Acanthognathusor Odontomachus) did weinvestigate this deceleration any further.
The fast mandible closer muscle is composed of longermuscle fibres with much shorter sarcomeres than found in theslow closer muscle (Fig. 5e,f; Table 1). Its fibre morphologycoincides with that of the fastest mandible closer muscle fibresof other ants (Gronenberget al., 1997) and thus suggests thatthis muscle is much faster than the slow mandible closer. Itsbrief contraction is likely to bring about the slight downwardrotation of the mandibles required to release the strike.
Neuronal control
The suboesophageal ganglion (SOG) controls the mandiblefunction. In ants it is fused ventrally to the brain (Figs 4a,b,5a,b). Besides the two neck connectives which connect theSOG to the ventral nerve cord, the mandibular nerves are thelargest nerves supplying the SOG in any ant. These mandiblenerves bifurcate close to their root at the ganglion (Fig. 4a,e)and give rise to a motor branch which projects laterally toconnect to the slow and the fast closer muscles. The remainderof the nerve proceeds anteriorly towards the mandible. It is
© 1998 Blackwell Science Ltd,Physiological Entomology, 23, 227–240
mainly composed of the afferent sensory axons originatingfrom the many sensilla on the mandible.
Among these sensory axons is one which is thicker than therest (µ 2 µm at the ganglion) and which originates from thelarge cell body associated with the long mandibular triggerhair mentioned above in the section on mandible morphology.The afferent trigger hair neurones project through almost theentire SOG in an antero-posterior direction and give rise tomany axon collaterals decorated with bleb-like swellings,suggesting presynaptic terminals. Because of the small size ofthe mandibles, no success was achieved in staining the onetrigger neurone exclusively, and in the three successfulpreparations we always found the axons of two to four othersensory neurones stained (Fig. 6). Accordingly, the connectionof every single axonal branchlet could not be discriminated.However, the general appearance of the trigger hair afferent’saxonal arborization pattern was not significantly different fromthose of neurones originating from other mandibular hairs,except for the larger axon diameter of the former. This indicatesthat the trigger hair afferents exhibit the same connectivity inthe SOG as do their smaller companions. Their larger axondiameter shows, however, that they have a higher conductionvelocity. Hence the trigger hairs and their neurones are derived
Trap-jaw mechanism 237
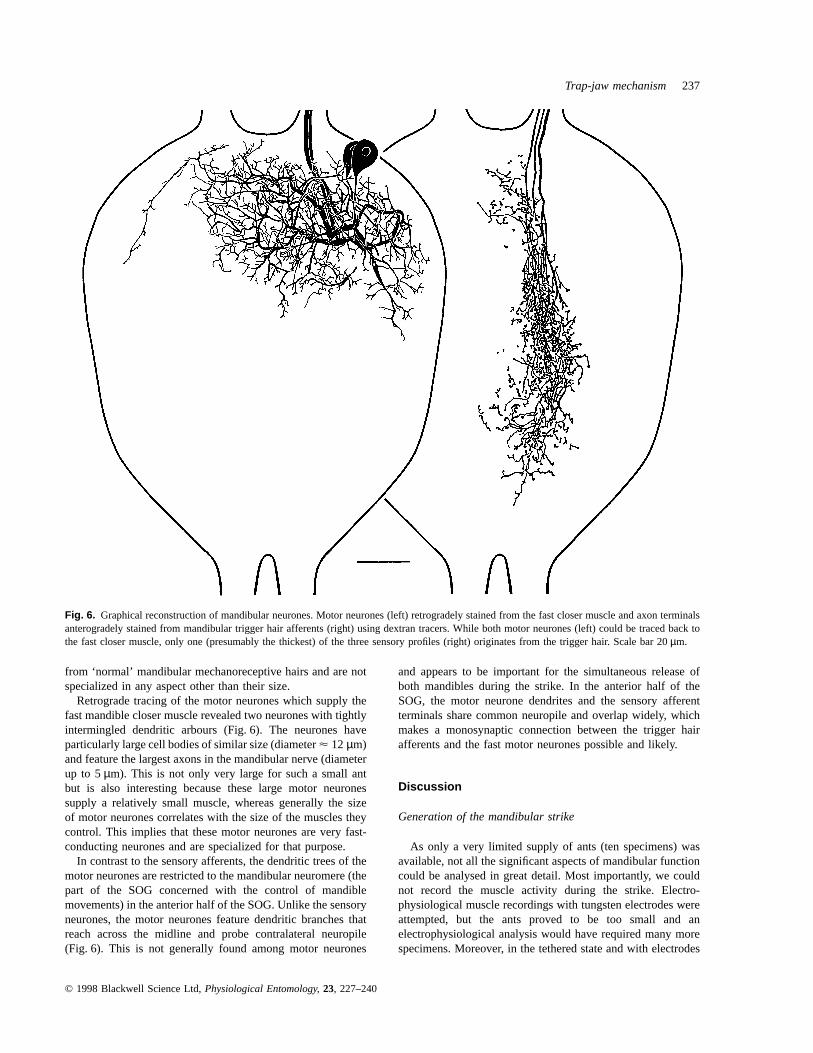
Fig. 6. Graphical reconstruction of mandibular neurones. Motor neurones (left) retrogradely stained from the fast closer muscle and axon terminalsanterogradely stained from mandibular trigger hair afferents (right) using dextran tracers. While both motor neurones (left) could be traced back tothe fast closer muscle, only one (presumably the thickest) of the three sensory profiles (right) originates from the trigger hair. Scale bar 20µm.
from ‘normal’ mandibular mechanoreceptive hairs and are notspecialized in any aspect other than their size.
Retrograde tracing of the motor neurones which supply thefast mandible closer muscle revealed two neurones with tightlyintermingled dendritic arbours (Fig. 6). The neurones haveparticularly large cell bodies of similar size (diameterµ 12 µm)and feature the largest axons in the mandibular nerve (diameterup to 5µm). This is not only very large for such a small antbut is also interesting because these large motor neuronessupply a relatively small muscle, whereas generally the sizeof motor neurones correlates with the size of the muscles theycontrol. This implies that these motor neurones are very fast-conducting neurones and are specialized for that purpose.
In contrast to the sensory afferents, the dendritic trees of themotor neurones are restricted to the mandibular neuromere (thepart of the SOG concerned with the control of mandiblemovements) in the anterior half of the SOG. Unlike the sensoryneurones, the motor neurones feature dendritic branches thatreach across the midline and probe contralateral neuropile(Fig. 6). This is not generally found among motor neurones
© 1998 Blackwell Science Ltd,Physiological Entomology, 23, 227–240
and appears to be important for the simultaneous release ofboth mandibles during the strike. In the anterior half of theSOG, the motor neurone dendrites and the sensory afferentterminals share common neuropile and overlap widely, whichmakes a monosynaptic connection between the trigger hairafferents and the fast motor neurones possible and likely.
Discussion
Generation of the mandibular strike
As only a very limited supply of ants (ten specimens) wasavailable, not all the significant aspects of mandibular functioncould be analysed in great detail. Most importantly, we couldnot record the muscle activity during the strike. Electro-physiological muscle recordings with tungsten electrodes wereattempted, but the ants proved to be too small and anelectrophysiological analysis would have required many morespecimens. Moreover, in the tethered state and with electrodes

238 Wulfila Gronenberget al.
inserted into her muscles,Acanthognathus rudiscould notbe coaxed into releasing the mandible strike at all. Thus,morphological and videographical evidence had to be reliedupon when explaining the way the mandible strike is performed.The following sequence of events is suggested.
As in a catapult, the mandibles are under tension prior tothe strike and are powered by the contraction of the largecloser muscle. This muscle is slow and contracts a long timein advance of the strike to build up tension. Slow contractionproperties of a muscle fibre generally coincide with a largerforce production, so the slow closer muscle appears to bedesigned for maximum force development. AsAcanthognathusslowly stalk their prey, there is ample time to ‘energise’ themandibles by the slow muscle. As in other trap-jaw ants(Gronenberg, 1995a,b, 1996b; Gronenberg & Ehmer, 1996)and fast insect movements in general (Alexander, 1988, 1995;Gronenberg, 1996a), the mechanical energy is probably storedelastically in the closer apodeme, the head capsule, and in themuscle itself until the strike is released.
The mandibles are held in the open position by the opposingaccessory mandibular processes which interlock. Theirconnection is such that a slight downward movement willdisengage the processes and let them slide clear of each other.This downward movement is generated by a contraction of thefast closer muscles which rapidly rotate the closer apodemes andthe mandibles around their long axes. Although physiologicalmeasurements are missing, their structural properties suggestthat the fast closer muscle fibres are well adapted for aparticularly fast contraction. Their morphology coincides withthat of the fastest mandible closer muscle fibres of other ants(Gronenberget al., 1997).
In principle, muscle fibres with short sarcomeres can shortenfaster than ones with long sarcomeres (O’Connoret al., 1982;Costello & Govind, 1983; Stephenset al., 1984; Gu¨nzelet al.,1993). In the fast closer muscles ofAcanthognathus, the greaternumber of sarcomeres per fibre alone would result in a morethan 3-fold increase in shortening velocity, compared to theslow closer fibres. However, it is to be expected that, as inother arthropods (see, e.g. Cochraneet al., 1972; Stokeset al.,1975; Govind & Atwood, 1982; Rathmayer & Maier, 1987;Muller et al., 1992) and in ants in particular, these fast closerfibres are specialized ultrastructurally (Gronenberget al., 1997)and enzymatically (Gronenberg & Ehmer, 1995) for highcontraction speed. Thus, they can probably shorten many timesfaster than the slow closer fibres. This specialization for speedseems particularly efficient because the fast closer muscle isprobably the limiting factor for the latency of the reflex releasedby the prey touching the trigger hairs.
The fast closer muscle contraction is introduced into themandible by a sturdy arm of the apodeme, much thicker andmore sclerotised than would normally be required for such asmall muscle. These lateral arms of the closer apodemes appearto be specialized for their crank action, by which translatorymovement (contraction of the fast closer muscle fibres) isconverted into rotation of the mandible. It is supposed that athinner and less rigid apodeme would be too elastic to transformthe fast muscle twitch properly into a rotation of the apodemeand mandible. When the locking device is thus released, the
© 1998 Blackwell Science Ltd,Physiological Entomology, 23, 227–240
mandibles are rapidly accelerated by the tension previouslybuilt-up.
The fast closer muscles are activated upon stimulation ofthe mandibular trigger hairs through a fast, and probablymonosynaptic pathway which involves particularly largesensory and motor neurones. Such specialized reflex pathwayshave been revealed in all trap-jaw ant genera studied sofar (Odontomachus, Anochetus, Strumigenysand Daceton;Gronenberg, 1995a,b, 1996b; Gronenberg & Ehmer, 1996).The existence of a similar fast neuronal pathway indicates that,in evolutionary terms, it was important forAcanthognathustoestablish a fast reflex in order to snap the mandibles shut assoon as the prey is contacted by the trigger hairs. Otherwise,the collembola might be able to employ their escape jumpwhen touched by the trigger hairs.
The prey-catching mechanism ofAcanthognathusrequires acatch mechanism (the accessory mandible processes) and thefunctional separation of the mandible closer muscle into a fastand a slow component. In addition,Acanthognathushaveevolved a kind of slide-bearing in the mandible joint. Thismechanism is more elaborate than that found in other trap-jawants. It provides an increased stabilization of the mandiblesbecause the catch mechanism is more delicate and allows forthe slight rotation required to release the mandible strike.
Two different trap-jaw mechanisms in dacetonine ants?
Trap-jaws are very effective means for capturing fast andelusive prey such as springtails, as well as protected ordangerous prey [e.g. glue-spraying termite soldiers(Nasutitermes)], and they also form good defensive weapons.For this reason, trap-jaws have evolved independently severaltimes among the ants: in the ponerine generaOdontomachusand Anochetus, in the myrmicine Dacetonini, and in theformicine genusMyrmoteras (Holldobler & Wilson, 1990).They are most abundant among the twenty-four dacetoninegenera, among which trap-jaws have been the subject of manyevolutionary changes (Brown & Wilson, 1959; Masuko, 1984;Holldobler & Wilson, 1990). In some genera, they havebeen greatly reduced (e.g.Smithistruma, Epitritus) or evensecondarily re-elongated from short-mandibulate forms (e.g. insome species ofNeostruma; Brown & Wilson, 1959).
The structure which blocks the trap-jaws before the strikeand thus allows an energy storage mechanism has briefly beendescribed forStrumigenysby Brown & Wilson (1959) and inmore detail for Strumigenysand Daceton by Gronenberg(1996b). Here, a part of the labrum clicks between the openmandibles and thus prevents them from closing. The strike isreleased upon withdrawal of the labrum.
It has already been mentioned that Dietz & Branda˜o (1993)described a different mechanism forAcanthognathus: thelabrum is not involved and the open mandibles are blocked bytheir acessory processes, as outlined in the present account.Dietz & Brandao did not invoke a catapult mechanism in theirinterpretation of the movement. For this reason, Gronenberg(1996b) doubted the existence of such a principally differenttrap-jaw mechanism within the same tribe without further

Trap-jaw mechanism 239
knowledge ofAcanthognathus. In the present account it isdemonstrated that both notions are partly true: the trap-jaws inAcanthognathusrely on a catapult mechanism and yet they aresignificantly different from the design of other dacetoninetrap-jaws.
The classification of the Dacetonini has recently beencritically analysed by Baroni Urbani & de Andrade (1994),based on the description of the first dacetonine fossils fromDominican amber (15–45 million years old). They havedescribed a unique alate queen of anAcanthognathus(A.poinari) that shows basal teeth in both inner margins of themandibles, although the left tooth is decidedly smaller thanthe right one. All six extantAcanthognathusspecies presentonly a single right basal tooth which sometimes is evenvestigial. This suggests that the extant species have lost thetooth in the left mandible, instead of acquiring a new structure.As already mentioned, the possible reason for the loss of theleft basal tooth may be that the right accessory process doesnot touch the left mandible in that particular area. Interestingly,this explanation would suggest that the extinctA. poinari andtheir ancestors may not have been ‘right-handed’, like all theextantAcanthognathusspecies, but ‘ambidextrous’.
The tribe Dacetonini has recently come under revision byBarry Bolton, which is not yet completed (personalcommunication). This revision is partly based on themorphology of the labrum and on the mandibular mechanism,and thus confirms the present finding that the mandibles ofAcanthognathusare indeed powered by a mechanism quitedifferent from that employed byStrumigenysor Daceton. Itwould not be unusual to have two different mandiblemechanisms if the genera are not very closely related. Arelatively close relationship has been implied, however, byBrown & Wilson (1959), who suggest that long mandibles insome dacetonine genera, among themDaceton, Orectognathusand Acanthognathus, are plesiomorphic traits. Hence, thecurrent account may stimulate further taxonomical work onthis issue.
Acknowledgements
We thank Christiane I. Yamamoto for helping to catch the ants,Sonja Schaff for her help with video observation, KarinMuller for histological assistance, Barry Bolton for discussingtaxonomical questions, and two anonymous referees for helpfulsuggestions. This work was supported by DeutscheForschungsgemeinschaft (Gr 933/3 and GK ‘Arthropodenver-halten’). The Brazilian team has been supported by theCoordenadoria para o Aperfeic¸oamento do Ensino Superiorand Fundac¸ao de Amparo a` Pesquisa do Estado de Sa˜o Paulo.
References
Alexander, R. McN. (1988)Elastic Mechanisms in Animal Movement,pp. 81–90. Cambridge University Press, U.K.
Alexander, R. McN. (1995) Leg design and jumping technique for
© 1998 Blackwell Science Ltd,Physiological Entomology, 23, 227–240
humans, other vertebrates and insects.Philosophical Transactionsof the Royal Society London B, 347, 235–248.
Baroni Urbani, C. & de Andrade, M.L. (1994) First description offossil Dacetini ants with a critical analysis of the current classificationof the tribe (Amber Collection Stuttgart: Hymenoptera, Formicidae.VI: Dacetini). Stuttgarter Beitra¨ge zur Naturkunde B, 198, 1–65.
Brown, W.L., Jr (1976) Contributions toward a reclassification of theFormicidae. VI. Ponerinae, tribe Ponerini, subtribe Odontomachiti.Section A. Introduction, subtribal characters, genusOdontomachus.Studia entomologia, 19, 67–171.
Brown, W.L., Jr (1978) Contributions toward a reclassification of theFormicidae. VI. Ponerinae, tribe Ponerini, subtribe Odontomachiti.Section B. GenusAnochetusand bibliography.Studia entomologia,20, 549–652.
Brown, W.L., Jr & Kempf, W.W. (1969) A revision of the NeotropicalDecetini ant genusAcanthognathus(Hymenoptera: Formicidae).Psyche, 76, 87–109.
Brown, W.L., Jr & Wilson, E.O. (1959) The evolution of the dacetineants.Quarterly Review of Biology, 34, 278–294.
Cochrane, D.G., Elder, H.Y. & Usherwood, P.N.R. (1972) Physiologyand ultrastructure of phasic and tonic skeletal muscle fibres in thelocust,Schistocerca gregaria. Journal of Cell Science, 10, 419–441.
Costello, W.J. & Govind, C.K. (1983) Contractile responses of singlefibres in lobster claw closer muscles: correlation with structure,histochemistry and innervation.Journal of Experimental Zoology,227, 381–393.
Dietz, B.H. & Branda˜o, C.R.F. (1993) Comportamento de cac¸a e dietade Acanthognathis rudisBrown & Kempf, com comenta´rios sobrea evolucao da predac¸ao em dacetini (Hymenoptera, Formicidae,Myrmicinae).Revista Brasileira de Entomologia, 37, 683–692.
Govind, C.K. & Atwood, H.L. (1982) Organization of neuromuscularsystems.The Biology of Crustacea, Vol. 3(ed. by D. Bliss), pp. 63–103. Academic Press, New York.
Gronenberg, W. (1995a) The fast mandible strike in the trap-jaw antOdontomachus: temporal properties and morphologicalcharacteristics.Journal of Comparative PhysiologyA, 176, 391–398.
Gronenberg, W. (1995b) The fast mandible strike in the trap-jaw antOdontomachus: motor control.Journal of Comparative PhysiologyA, 176, 399–408.
Gronenberg, W. (1996a) Fast actions in small animals. Springs andclick mechanisms.Journal of Comparative Physiology A, 178,727–734.
Gronenberg, W. (1996b) The trap-jaw mechanism in the dacetine antsDaceton armigerumand Strumigenyssp. Journal of ExperimentalBiology, 199, 2021–2033.
Gronenberg, W. & Ehmer, B. (1995) Tubular muscle fibres in ants andother insects.Zoology, 99, 68–80.
Gronenberg, W. & Ehmer, B. (1996) The mandible mechanism of theant genusAnochetus(Hymenoptera, Formicidae) and the possibleevolution of trap-jaws.Zoology, 99, 183–192.
Gronenberg, W., Paul, J., Just, S. & Ho¨lldobler, B. (1997) Mandiblemuscle fibres in ants: fast or powerful?Cell and Tissue Research,289, 347–361.
Gronenberg, W., Tautz, J. & Ho¨lldobler, B. (1993) Fast trap jaws andgiant neurons in the antOdontomachus. Science, 262, 561–563.
Gunzel, D., Galler, S. & Rathmayer, W. (1993) Fibre heterogeneity inthe closer and opener muscles of crayfish walking legs.JournalExperimental Biology, 175, 267–281.
Holldobler, B. & Wilson, E.O. (1990)The Ants. Belknap Press ofHarvard University Press, Cambridge, Massachussetts.
Masuko, K. (1984) Studies on the predatory biology of oriental dacetineants (Hymenoptera: Formicidae). I. Some Japanese species ofStrumigenys, Pentastruma, and Epitritus, and a Malaysian

240 Wulfila Gronenberget al.
Labidogenys, with special reference to hunting tactics in short-mandibulate forms.Insects socieaux, 31, 429–451.
Moffett, M.W. (1986) Trap-jaw predation and other observations ontwo species ofMyrmoteras (Hymenoptera: Formicidae).Insectssocieaux, 33, 85–99.
Muller, A.R., Wolf, H., Galler, S. & Rathmayer, W. (1992) Correlationof electrophysiological, histochemical, and mechanical properties infibres of the coxa rotator muscle of the locust,Locusta migratoria.Journal of Comparative Physiology, 162, 5–15.
O’Connor, K., Stephens, P.S. & Leferovich, J.M. (1982) Regionaldistribution of muscle fibre types in the asymmetric claws ofCalifornian snapping shrimp.Biological Bulletin, 163, 329–336.
© 1998 Blackwell Science Ltd,Physiological Entomology, 23, 227–240
Rathmayer, W. & Maier, L. (1987) Muscle fibre types in crabs:studies on single identified muscle fibres.American Zoologist, 27,1067–1077.
Stephens, P.J., Lofton, L.M. & Klainer, P. (1984) The dimorphic clawsof the hermit crab,Pagurus pollicaris: properties of the closermuscle.Biological Bulletin, 167, 713–721.
Stokes, D.R., Josephson. R.K. & Price, R.B. (1975) Structural andfunctional heterogeneity in an insect muscle.Journal of ExperimentalZoology, 194, 397–408.
Accepted 20 January 1998
Copyright © 2022 FDOKUMEN




















