
Analysis of trans-silencing interactions using transcriptional silencers of varying strength and...
14
Transgenic Research 12: 305–318, 2003. © 2003 Kluwer Academic Publishers. Printed in the Netherlands. 305 Analysis of trans-silencing interactions using transcriptional silencers of varying strength and targets with and without flanking nuclear matrix attachment regions Robert Ascenzi 1,2,3,4,∗ , Bekir Ülker 2,5 , Joselyn J. Todd 1,6 , Dolores A. Sowinski 1 , Carolyn R. Schimeneck 1,7 , George C. Allen 1 , Arthur K. Weissinger 2 & William F. Thompson 1,2,3 1 Department of Botany, North Carolina State University, Raleigh, NC 27695-7612, USA 2 Department of Crop Science, North Carolina State University, Raleigh, NC 27695-7620, USA 3 Department of Genetics, North Carolina State University, Raleigh, NC 27695-7614, USA Received 21 June 2002; accepted 18 September 2002 Key words: matrix attachment region, tobacco, transcriptional gene silencing, transgene expression, trans- silencing Abstract We investigated the effect of the Rb7 matrix attachment region (MAR) on trans-silencing in tobacco plants, com- paring the effects of three transgene silencer loci on ten target loci. Two of the silencer loci, C40 and C190, contain complex and rearranged transgene arrays consisting of 35S:GUS or NOS:NPTII containing plasmids. The third silencer locus, V271, was previously characterized as a complex locus containing rearranged 35S:RiN sequences. Each of these silencers can reduce 35S promoter-driven expression at other loci, albeit with varying efficiencies. The presence of MARs at a target locus does not prevent trans-silencing by the V271 silencer. However, four of seven MAR-containing loci were at least partially resistant to silencing by the C40 and C190 loci. One MAR locus was unaffected by C40, our weakest silencer, and three were silenced only when the silencer locus was maternally inherited. Silencing is progressive in the F1 and F2 generations; two days after germination there is little or no difference between seedlings derived from crosses to silencing or control lines, but seedlings containing silencer loci slowly lose expression during subsequent development. These observations are compatible with the hypothesis that a product of the silencer locus must accumulate before unlinked loci can be affected. However, our silencer loci are themselves silenced for GUS transcription, and coding region homology is not required for their effects on target loci. Our results are consistent with a model in which transcriptional silencing is triggered by transcription of sequences during the early stages of embryo or seedling development. Abbreviations: BC1 – First back-cross generation; F1 – First filial generation; F2 – Second filial generation; GUS – β-Glucuronidase; HC-Pro – Helper component proteinase; R0 – Primary regenerant; SR-1 – Streptomycin resistant 1. ∗ Author for correspondence: E-mail: [email protected] 4 Present address: BASF Plant Science L.L.C., 26 Davis Drive, P.O. Box 13528, Research Triangle Park, NC 27709-3528, USA 5 Present address: Max-Planck Institut für Züchtungsforschung, Department of Biochemistry, Carl-von-Linn´ e Weg 10, D-50829 Cologne, Germany 6 Present address: Cary Academy, 1500 N. Harrison Avenue, Cary, NC 27513, USA 7 Present address: Cato Research, 200 Westpark Corporate Center, 4364 South Alston Avenue, Durham, NC 27713-2280, USA Introduction Homology-dependent silencing phenomena are of two general types. Transcriptional gene silencing (TGS) occurs when promoters share sequence homology, while post-transcriptional gene silencing (PTGS) is dependent on homology within transcribed sequences. While both types of silencing may be associated with methylation of homologous DNA sequences (Matzke
Transcript of Analysis of trans-silencing interactions using transcriptional silencers of varying strength and...

Transgenic Research 12: 305–318, 2003.© 2003 Kluwer Academic Publishers. Printed in the Netherlands.
305
Analysis of trans-silencing interactions using transcriptional silencers ofvarying strength and targets with and without flanking nuclear matrixattachment regions
Robert Ascenzi 1,2,3,4,∗, Bekir Ülker 2,5, Joselyn J. Todd 1,6, Dolores A. Sowinski 1, CarolynR. Schimeneck 1,7, George C. Allen 1, Arthur K. Weissinger 2 & William F. Thompson 1,2,3
1Department of Botany, North Carolina State University, Raleigh, NC 27695-7612, USA2Department of Crop Science, North Carolina State University, Raleigh, NC 27695-7620, USA3Department of Genetics, North Carolina State University, Raleigh, NC 27695-7614, USA
Received 21 June 2002; accepted 18 September 2002
Key words: matrix attachment region, tobacco, transcriptional gene silencing, transgene expression, trans-silencing
Abstract
We investigated the effect of the Rb7 matrix attachment region (MAR) on trans-silencing in tobacco plants, com-paring the effects of three transgene silencer loci on ten target loci. Two of the silencer loci, C40 and C190, containcomplex and rearranged transgene arrays consisting of 35S:GUS or NOS:NPTII containing plasmids. The thirdsilencer locus, V271, was previously characterized as a complex locus containing rearranged 35S:RiN sequences.Each of these silencers can reduce 35S promoter-driven expression at other loci, albeit with varying efficiencies.The presence of MARs at a target locus does not prevent trans-silencing by the V271 silencer. However, four ofseven MAR-containing loci were at least partially resistant to silencing by the C40 and C190 loci. One MAR locuswas unaffected by C40, our weakest silencer, and three were silenced only when the silencer locus was maternallyinherited. Silencing is progressive in the F1 and F2 generations; two days after germination there is little or nodifference between seedlings derived from crosses to silencing or control lines, but seedlings containing silencerloci slowly lose expression during subsequent development. These observations are compatible with the hypothesisthat a product of the silencer locus must accumulate before unlinked loci can be affected. However, our silencerloci are themselves silenced for GUS transcription, and coding region homology is not required for their effects ontarget loci. Our results are consistent with a model in which transcriptional silencing is triggered by transcriptionof sequences during the early stages of embryo or seedling development.
Abbreviations: BC1 – First back-cross generation; F1 – First filial generation; F2 – Second filial generation;GUS – β-Glucuronidase; HC-Pro – Helper component proteinase; R0 – Primary regenerant; SR-1 – Streptomycinresistant 1.
∗Author for correspondence: E-mail: [email protected] address: BASF Plant Science L.L.C., 26 Davis Drive,
P.O. Box 13528, Research Triangle Park, NC 27709-3528, USA5Present address: Max-Planck Institut für Züchtungsforschung,
Department of Biochemistry, Carl-von-Linne Weg 10, D-50829Cologne, Germany
6Present address: Cary Academy, 1500 N. Harrison Avenue,Cary, NC 27513, USA
7Present address: Cato Research, 200 Westpark CorporateCenter, 4364 South Alston Avenue, Durham, NC 27713-2280, USA
Introduction
Homology-dependent silencing phenomena are of twogeneral types. Transcriptional gene silencing (TGS)occurs when promoters share sequence homology,while post-transcriptional gene silencing (PTGS) isdependent on homology within transcribed sequences.While both types of silencing may be associated withmethylation of homologous DNA sequences (Matzke

306
et al., 1989; Ingelbrecht et al., 1994), only TGS isknown to be associated with localized alterations inchromatin structure (e.g., Ye & Signer, 1996). Typ-ically, once TGS is established it persists throughmeiosis and embryogenesis, while PTGS must bere-established in each generation following an initialperiod of gene expression (reviewed in Matzke &Matzke, 1998).
Trans-silencing is a form of TGS that involvesinteraction between unlinked transgenes (Matzkeet al., 1989). Cytosine methylation of the pro-moter sequences often accompanies the silencing.These trans-interactions may be studied by sequentialtransformation (Matzke et al., 1989), or by crossinglines containing silencer loci with lines containing oth-erwise stably expressed target loci (Vaucheret, 1993;Park et al., 1996).
Other types of trans-interactions involving endo-genous genes are paramutation and transvection (re-viewed by Wu & Morris, 1999). Paramutation involvesthe silencing of an active allele by an inactive alleleon a homologous chromosome (reviewed by Hollicket al., 1997; Chandler et al., 2000). In transvection,much like paramutation, allelic copies of genes mayinteract in such a way that α-silenced allele has a neg-ative effect on a well-expressed allele (trans-silencing)or a well-expressed allele has a positive effect on aninactive allele (trans-activation). It is important thatboth alleles are in the same chromosomal location;translocation of one allele eliminates transvection.While there is ample evidence for transvection in fungiand animals, there is only one reported case for trans-vection in plants (Matzke et al., 2001). Importantly,it was found that the trans-activation occurred only athomologous chromosomal sites; transvection was notobserved in lines that had homologous transgenes atectopic sites.
The mechanisms underlying these trans-interactions are unclear, although it seems likely thatrecognition of homologous sequences must involvesome form of nucleic acid pairing. In the case oftransvection, DNA strands from sister chromatids arethought to interact with each other (reviewed in Wu &Morris, 1999). It was originally assumed that DNA–DNA pairing also occurs with trans-silencing (Matzke& Matzke, 1998). However, several lines of evid-ence now suggest that RNA may pair with DNAto cause silencing. For example, Wassenegger et al.(1994) demonstrated that viroid genome replicationcould cause methylation of an integrated viroid cDNA,and Mette et al. (1999) showed that transcription of
the NOS promoter was capable of silencing unlinkedhomologous promoters (Mette et al., 1999, 2000).
Nuclear matrix attachment regions (MARs) aredefined as DNA sequences that exhibit specific bind-ing to nuclear matrix preparations. MARs have beenshown to be important in the proper regulation ofendogenous genes, and to increase expression of trans-genes. For example, in splenic B cells of transgenicmice, an Ig µ gene lacking its endogenous MARis 30–1000-fold less active than a wild-type µ gene(Forrester et al., 1994). In transgenic plant cells, thetobacco Rb7 MAR (Hall et al., 1991) increased the av-erage expression level of a GUS transgene by 60-fold(Allen et al., 1996). Several other animal and plantMARs have been shown to have positive effects on theexpression of transgenes (e.g. Stief et al., 1989; Phi-Van et al., 1990; Schöffl et al., 1993; van der Geestet al., 1994; Peterson et al., 2002). It is not knownhow MARs exert their effects, but several models ex-ist. Most of these have been recently reviewed (Allenet al., 2000).
Because transcriptional silencing has been thoughtto be a chromatin-mediated effect, and MARs arebelieved to resist repressive chromatin effects, itis reasonable to suppose that MARs might preventtrans-silencing. However, Nap et al. (1997) andVaucheret et al. (1998) have presented evidence thatMARs are ineffective against trans-silencing fromstrong silencing loci. In the former study, it wasshown that a chicken MAR (lysozyme A element)was not effective in preventing epistatic dominanceof complex loci with low GUS activity. In the lat-ter study, it was shown that one particular ‘supersilencer’ locus (V271; Vaucheret, 1993) greatly re-duced the expression of transgenes containing 35Spromoters, irrespective of the presence of MARs inthe target transgene constructs. However, it is notclear that silencing by the V271 locus is represent-ative of trans-silencing generally, and much remainsto be learned before mechanistic explanations can beproposed.
In this paper, we describe experiments with theV271 locus and two other trans-silencer loci generatedin the course of our own work. In agreement withVaucheret et al. (1998), we show that the Rb7 MARdoes not prevent trans-silencing by the strong V271locus. However, we also show that this MAR canprovide partial protection from weaker silencers, aneffect that may be important for our understandingof molecular mechanisms of MAR activity and alsowould have implications for achieving coordinated

307
Figure 1. Schematic diagram of the transgenes containing 35S sequences. The orientation of coding sequences are indicated with arrow-heads and the constructs are aligned with respect to the 35S promoter. The control construct pGHNC12 was used to transform tobacco bymicroprojectile bombardment as described in Methods. Transformed plant line names contain the prefix ‘C’ (e.g., C40). Lines transformed bybombardment with the MAR construct pGHNC11 are given names with the prefix ‘M’. Lines designated ‘N-C’ (e.g., N-C9) were transformedwith the control construct, pJSL1, containing the N gene without MARs. These transformations were carried out using Agrobacterium. Plantstransformed with the corresponding MAR construct pJSL3 were also transformed using Agrobacterium, and are designated by the prefix‘N-M’ (e.g., N-M4). Note: The plasmid pGHNC10 (not shown, described in Allen et al., 1996) was co-transformed with either pGHNC11 orpGHNC12 to confer selection; it does not contain promoter, terminator or coding sequences homologous to the plasmids depicted.
transgene expression. Furthermore, we present datathat are consistent that a product(s) from these weak si-lencer loci is required to re-establish silencing in eachgeneration. This result differs from earlier concepts oftranscriptional gene silencing, in which silencing wasthought to involve stable epigenetic changes persis-ting through meiosis (reviewed in Matzke & Matzke,1998).
Materials and methods
Plant material
Nicotiana tabacum cv. Petite Havana SR-1 wastransformed by microprojectile bombardment aspreviously described (Ülker et al., 1999). Briefly,11-day-old-seedlings were bombarded with a mixtureof two plasmids, pGHNC10 and either pGHNC11or pGHNC12. The pGHNC10 carries the selectablemarker nptII under control of the nos promoter, whilepGHNC11 and 12 carry the gusA reporter gene underthe control of the 35S promoter. In pGHNC11, the Rb7MAR flank the 35S:GUS cassette (Hall et al., 1991)(Figure 1). Explants resistant to 100 µg/ml kanamycinwere used to regenerate plantlets. Lines homozygousfor the transgene loci were obtained by selfing R0plants or their BC1 progeny, or by producing doubledhaploid lines (Ülker, 1998). Homozygous plants werereciprocally crossed with each other or with a non-
transformed doubled-haploid plant, or with lines ex-pressing P1/HC-Pro. Specifically, a stamen from oneflower was used to donate pollen to multiple pistilsin each direction. Therefore, any possible non-geneticvariation in expression levels would be due to differ-ences in the position of the flowers pollinated. Cap-sules from each of the flowers from a single cross werepooled. We used the P1/HC-Pro lines U6B (Carringtonet al., 1990) and TEV-B (Verchot et al., 1991), both ofwhich were kindly provided by Dr V. Vance. We ana-lyzed F1 progeny of these crosses, and F2 progeny ofselfed F1 plants. Primary transgenic lines were back-crossed to untransformed SR-1 to produce BC1 linesas described (Ülker et al., 1999). GUS-positive BC1plants were crossed with homozygous V271 plants(Vaucheret, 1993) or untransformed Xanthi controls,and kanamycin resistant seedlings analyzed to producethe data shown in Figure 2(a) and (b).
The lines N-M4 and N-C9 (35S:TSWV-N) wereproduced by Agrobacterium-mediated transformationof haploid plants, followed by chromosome doubling(Ülker, 1998; Collins & Sadasivaiah, 1972).
Plants were typically germinated on GM medium(Valvekens et al., 1988) and transferred to growingmedium (Metro-Mix 200, Scotts, Marysville, OH) fivedays after germination. Plants were then maintained ina growth chamber (16:8 L:D photoperiod, 25◦C) forfour weeks, with assays taking place during this time.Plants were then transplanted and allowed to mature ina greenhouse.

308
Reporter gene assays
β-Glucuronidase (GUS) was assayed in situ by incub-ating plant tissue with X-Gluc overnight as describedby Sieburth and Meyerowitz (1997). GUS activity in
soluble protein extracts was assayed luminometricallywith the substrate Glucuron (Tropix, Bedford, MA),as described previously (Ülker et al., 1999). Samplescontaining 10 µg total soluble protein were assayedin microtiter plates. Standards were constructed withauthentic enzyme standards (Sigma, St. Louis, MO)containing 10 µg protein from a parallel extract of anon-transformed control plant of the same develop-mental age.
Nucleic acids methods
DNA was extracted as described (Ülker et al.,1999), or by a method employing sarkosyl and urea(Cocciolone and Cone, 1993). Southern hybridizationswere performed as described in Ascenzi and Gantt(1999) using a probe derived from the EcoR1/HindIIIfragment containing 35S:GUS:NosT sequences. Asimple PCR-based assay for inverted repeats (IR-PCR)was used. This technique relies on the ability ofa single primer to produce a specific amplificationproduct if the primer sequence is present in an invertedrepeat. In order to increase our chances of obtaininga product, we used nested sets of primers. We per-formed each reaction as follows: approximately 50 ngof genomic DNA was amplified by adding five primersin the sense direction or four primers in the anti-sense direction. The ‘Expand’ Long PCR kit (RocheMolecular Biochemicals, Indianapolis, IN) was usedaccording to the manufacturer’s guidelines with an an-nealing temperature of 50◦C. The sense primers are
←−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−Figure 2. Transcriptional silencing gusA genes in BC1 seedlingscarrying the V271 locus. (a) GUS histochemical staining ofBC1×V271 hybrid seedlings. Kanamycin-resistant seedlings wereobtained from crosses between experimental MAR or control trans-genic lines and a plant containing the V271 silencer locus. Theseseedlings were analyzed by histochemical staining at several timepoints during the first month after germination. Note that the numberof seedlings differs between wells. (b) Effect of the V271 locuson gusA mRNA levels. Seed derived from crosses between lineM56 and either V271 or WT tobacco were germinated and har-vested at various times following germination. RNA was extractedand analyzed using a microscale procedure (Gallo-Meagher et al.,1992). Hybridization signals were quantified using a PhosphorIm-ager. Differences in RNA loading were normalized to 5 µg per laneby probing with an oligonucleotide to the small subunit of ribosomalRNA. (c) GUS is synthesized during germination. Wild-type andtransgenic tobacco seed (Line 1a) carrying TT:GUS, in whichgusA transcription is driven by the tetracycline-suppressible Top10 promoter, were stained for GUS activity 5 days post imbibi-tion. Germination media contained 0 or 0.1 µg/ml tetracycline asindicated.

309
as follows: Y (5′-TAATACGACTCACTATAGGGC-3′); K (5′-AGATAGTGGAAAAGGAAGGTG-3′);P1 (5′-ATGACGCACAATCCCACTAT-3′); X (5′-ACAATGAATCAACAACTCTC-3′); X′ (5′-AGATGGGTTTTTATGATTAGA-3′); and the antisense prim-ers are the following: A (5′-GGAAACAGCTATGACCATGA-3′); L (5′-TGCGGGACTCTAATCATAAAA-3′); P3(5′-TGGTGTAGAGCATTACGCTG-3′); B (5′-AGCCTCTCTAACCATCTGTG-3′). Prim-ers K and L were used as a positive control. Note thatamplification products resulting from sense primersindicate a ‘head:head’ arrangement, while those re-sulting from antisense primers indicate a ‘tail:tail’arrangement. As multiple primers were used the num-ber of products obtained does not necessarily reflectthe number of inverted repeats and are therefore notpresented.
For measuring transcription rates, nuclei were iso-lated from leaves of greenhouse-grown tobacco asdescribed by Cox and Goldberg (1988). Run-on tran-scription assays were performed as detailed in Dehioand Schell (1994) except that 1× 106–2.5× 106 nu-clei were used instead of 1× 107 and the reactionvolume was reduced by a factor of two. Each reac-tion contained 200 µCi of [32P]UTP, 3000 Ci/mmol.In some run-on assays, heparin was added to a finalconcentration of 1.5 mg/ml prior to adding [32P]UTPin order to increase the efficiency of transcription(Mösinger et al., 1987). Labeled RNA with anequivalent number of counts (8.5× 106 cpm) washybridized to replicate Southern blot filters con-taining approximately 5 µg of each of the fol-lowing target sequences: pea 18S DNA, soybeanrbcS (SRS4), E. coli GUS (gusA). The amountof target sequence did not include vector or othernon-transcribed sequences. Hybridization and wash-ing conditions were as described above for South-ern blots. Hybridization signals were quantified byPhosphorImager analysis, and relative gusA tran-scription was standardized against rbcS hybridi-zation signals. Label incorporation into RNA wasreduced by 50% in the presence of α-amanitin,indicating a high level of RNA polymerase IIactivity.
RNA was extracted and analyzed as described byGallo-Meagher et al. (1992), or extracted with TRIzolaccording to the manufacturer’s guidelines (Life Tech-nologies, Rockville, MD) and analyzed as describedby Ascenzi and Gantt (1999). Hybridization was visu-alized and quantitated with a Molecular DynamicsPhosphorImager.
Results
The V271 silencer is not effective immediately
To test whether MARs would prevent trans-silencing,we examined progeny from crosses between tobaccoline V271 (Vaucheret’s line 271) and several of ourpreviously described GUS-expressing lines (Ülkeret al., 1999). The transgene locus in line V271 is com-plex, containing multiple rearranged copies of both35S:RiN (an antisense nitrate reductase sequence) and19S:NPTII transgenes (Vaucheret, 1993). It has beenshown to act as a ‘super-silencer’ for all transgenesdriven by 35S promoters, including those flanked byMARs (Vaucheret et al., 1998).
We crossed plants containing the V271 silencerlocus to plants containing expressed 35S:GUS trans-genes with or without flanking copies of the Rb7MAR element, using the BC1 lines described by Ülkeret al. (1999). Seedlings derived from these crosseswere analyzed by histochemical staining at severaltime points during the first month after germination,and results are shown in Figure 2(a). In MAR lines(MAR lines are indicated with an “M” preceding theline number, while control (non-MAR) lines are indi-cated with a “C”), strong staining was observed 5 daysafter germination, and in some cases visible stainingpersisted for up to 31 days. Most control lines showedmuch weaker initial expression, even in the absence ofthe V271 locus, so it is difficult to evaluate the kine-tics of silencing. However, one control line, C83 (seeTable 1), did show strong staining in the absence of theV271 locus at the earliest time point tested. This lineshowed extensive silencing at 5 days post germination,although low levels of staining could be observed atlater time points (Figure 2(a)).
RNA gel blot analysis of seedlings similar to thoseshown in Figure 2(a) indicated that the V271 locussharply reduced gusA transcript levels at the earli-est time tested, five days after germination (e.g., seeFigure 2(b)). Similar results were obtained for linescontaining constructs with and without MARs. Thusthe persistence of histochemical staining in MARlines is most likely attributable to persistence of GUSprotein synthesized early in germination. If this hy-pothesis is correct, the 35S:GUS transgene must beexpressed in the presence of the V271 locus, albeitonly for a short time.
It remains formally possible that gusA RNA orprotein is carried over from the BC1 parent lines(used as the female parent in these crosses). We

310
Table 1. Summary of transgenic tobacco lines used to provide silencer and target loci. Each line contains the indicated numberof copies of a 35S:GUS reporter construct as well as a NOS:NPTII selectable marker introduced on a separate plasmid byco-bombardment
Line Silencer/ gusA Inverted Repeat Expressione Result of
numbera target copy no.b repeatc orientationd HC-Pro crossf
C40 S 6 Yes H, T No −M80 S 4 Yes H, T Low +C190 S 4 Yes T No −N-M4∗ T 1 ND NA High ND
N-C9∗ T 2 ND NA High +M36 T 2 Yes H Moderate +M56 T 1 No NA High ND
M76 T 6 No NA High ND
C83 T 1 No NA High ND
C91 T 2 No NA High ND
M117 T 3 No NA High +M137 T 2 ND NA Low +∗ 35S:TSWV-N (C= control, M=MAR).a C= pGHNC12 (control); M= pGHNC11 (MAR).b Determined by a combination of quantitative PCR and genomic Southern blot analyses, as described in Methods.c ND= not determined.d H= head:head; T= tail:tail; NA= not applicable.e Expression levels were measured by a combination of histochemical and extractive assays, as described in Methods.f (+) Increased expression when crossed with U6B or TEV-B lines relative to control cross, (−) no effect on expression.
therefore carried out a control experiment with plantsexpressing GUS from the tetracycline-sensitive Top10promoter. This promoter relies on an artificial tran-scription factor, TetVP16, that cannot activate tran-scription in the presence of extremely low levels oftetracycline (Weinmann et al., 1994; Love et al.,2000). Seed from a plant showing strong GUS ex-pression in the absence of tetracycline, Line 1a, wasgerminated on media with or without tetracycline, andthen stained for GUS activity at various times aftergermination. As shown in Figure 2(c), GUS stainingis observed only in seedlings germinated in the ab-sence of tetracycline. The absence of staining whenTOP10:GUS transcription is suppressed during ger-mination indicates that detectable amounts of enzymeare not carried over from the previous generation. TheGUS staining observed in Figure 2(a) must thereforereflect expression of 35S:GUS genes in the presenceof V271 silencer locus during the initial phase of ger-mination. We conclude that trans-silencing by V271does not occur immediately.
Two silenced loci are silenced at thetranscriptional level
As a follow-up of our initial work with cell cul-tures, we have previously shown that 35S:GUS loci
produced by bombardment with MAR-containingconstructs are much less likely to be silenced in thebackcross (BC1) generation than similar loci pro-duced by bombardment with control constructs lack-ing MARs (Ülker et al., 1999). For the experimentsdescribed here, some of the loci used in our previouswork were made homozygous by producing doubledhaploids (see Methods). Two of the resulting homo-zygous control (non-MAR) lines, C40 and C190, werefully silenced and exhibited trans-silencing activity onother loci. Both lines had shown expression in the R0and BC1 generations (Ülker, 1998). As doubled hap-loids, however, neither C40 nor C190 produced anydetectable gusA transcripts or enzyme activity at anydevelopmental stage.
We performed two sets of experiments to deter-mine the nature of the gene silencing in these cases.First, we examined whether the viral suppressor ofPTGS HC-Pro could restore GUS expression in oursilenced GUS lines. We found that the C40 andC190 loci could not be reactivated, whereas silencedlines M137 and M80 became active in the presenceof HC-Pro (Anandalakshmi et al., 1998; Kasschau& Carrington, 1998) (Table 1 and Figure 3). Theseresults are consistent with a recent result that demon-strated that HC-Pro cannot relieve double-stranded

311
Figure 3. Reversal of post-transcriptional silencing of GUS inplants containing HC-Pro. Leaves from equivalent positions fromF1 plants (30 dpg) were stained for GUS as described in Methods.Plants were either crossed with line TEV-B (HC-Pro) or the Xanthibackground (control).
Figure 4. The C40 and C190 silencing loci are transcriptionallysilenced. Nuclear run-on assays were performed as described inMethods. SR-1 is a non-transformed control. Filters contained theentire gusA coding sequence or pea 18S rDNA, as indicated.
RNA-mediate transcriptional silencing (Mette et al.,2001). In addition, nuclear run-on transcription assaysshowed extremely low activity in lines C40 and C190.Figure 4 shows that nuclei from homozygous C40 andC190 plants produce a barely detectable hybridizationsignal, while nuclei from plants containing expressedloci such as C91 and M38 show strong hybridiza-tion signals. We attribute the very weak signals tonon-specific hybridization because we have observedcomparable weak hybridization signals between vec-tor backbone sequences and run-on labeled RNA fromthe non-transformed line, SR-1. Taken together, thesedata indicate that the 35S:GUS transgene loci in thedoubled haploid lines C40 and C190 are silenced atthe transcriptional level.
Characteristics of the target and silencer loci used
Several 35S:GUS lines that showed stable expres-sion were used as target loci to assess trans-silencing.These lines (both control and MAR-containing) wereselected because they showed stable expression in ho-mozygous doubled haploid plants as well as in thehemizygous plants of the R0 and BC1 generations.The characteristics of the loci in this study are shownin Table 1. gusA copy number was determined byquantitative PCR (Ülker et al., 1999) and by Southernblotting. High-expressing control lines C83 and C91contain one or two copies of the transgene, respec-tively. High-expressing MAR lines M56, M117 andM76 contain one, three or six transgene copies. Wealso used two additional MAR lines that showed some-what lower expression levels. These were line M36(moderate expression) and line M137 (low expres-sion), each of which contained two transgene copies.(For comparison, note that the C190 silencer locuscontains four copies of the gusA gene, while the C40silencer locus contains six copies.)
Two 35S:GUS loci function as trans-silencers withvarying strength on MAR and control lines
F1 progeny derived from crosses between target linesand trans-silencer lines were assayed for GUS activityat early and late time points during seedling growth,and compared to progeny of control crosses (targetline× non-transformed control). Root and shoot GUSactivity (staining and the luminometric assay) were as-sayed at two days post germination (2 dpg) and leafGUS activities were assayed at 26 and 50 dpg. At2 dpg, histochemical assays showed that all F1 pro-geny from crosses with the C40 silencer line had GUSstaining intensities comparable to those in progenyof control crosses (Figure 7(a) and data not shown).Because we know there is no carryover of active GUSprotein from the previous generation (Figure 2(c)),these results indicate that expression can occur forsome period of time in the presence of the silencerlocus. Progeny from crosses between target loci andthe C190 silencer line also exhibited GUS stainingat 2 dpg. However, several of the C190 crosses gaverise to progeny that stained slightly less intensely thanthe controls, even at 2 dpg (data not shown). This re-sult suggests that silencing is beginning to be effectiveprior to 2 dpg, a result consistent with other indicationsthat C190 is a more effective silencer than C40 (seebelow).

312
Figure 5. Effect of strong and weak silencer loci, C190 and C40, on well-expressed target loci. All lines are described in Table 1. Crossesare indicated by listing the female parent first. The bars indicate mean and standard deviations of GUS specific activities of extracts from leafpunches (the 4th leaf) of plants harvested 26 days after germination. Each bar represents the average expression for at least three replicateplants.
By 26 dpg, loss of GUS activity was consistentlyobserved in progeny of both sets of crosses. Figure 5shows data from quantitative GUS assays. Leaves con-taining both the C190 silencer and the C91 targetlocus have GUS activity levels less than 27% of thoseseen in progeny of a wild type×C91 cross at 26 dpg.Likewise, F1 seedlings containing both the C190 andM36 loci have less than 25% of the activity seen inM36 control seedlings. The quantitative GUS assaysat 26 dpg clearly show that C190 is a stronger silencerthan C40 (Figure 5). In the case of the M36 targetlocus, silencing by C190 decreases GUS activity toabout 20% of control levels, whereas silencing by C40reduces GUS levels by less than 50% if at all. Incontrast, the progeny of the M36×C40 (silencer de-rived from male) had a higher mean GUS activity levelthan even the control cross. This indicates that C40is ineffective at silencing when derived from the male(differences in silencer activity due to parent of originare discussed further below) and that the M36 locus ispartially resistant to trans-silencing by C40, but notby the stronger C190 locus. Whether the resistanceof M36 is due to the presence of the flanking MARscannot be known. However, another MAR-containingline, M117, is effectively silenced by C40 (Table 2). Asummary of all crosses with C40 and C190 is shownin Table 2.
It is possible that the loss of expression observedin the progeny of the crosses described above re-sults from an additive effect of transgenes at differentloci, rather than from special properties of the silencer
locus. To address this possibility, we analyzed crossesin which M117, a stably expressing line with threecopies of the gusA gene, was substituted for the silen-cing locus C190 (with four copies). When M117 wascrossed with C91 the progeny showed normal GUSactivity, in sharp contrast to the reduced expression inthe progeny of C190×C91 crosses (data not shown).This result indicates that the trans-silencing effect ofC190 is most likely to derive from a special propertyof the locus, rather than simply from increasing thenumber of transgene copies in the genome.
trans-silencing does not require coding regionhomology
The delayed onset of silencing we observe with thesecomplex silencing loci are reminiscent of PTGS,which must be re-established each generation. PTGSis normally triggered by coding region homology (re-viewed in Wolffe & Matzke, 1999), so we askedwhether coding region homology was required forsilencing by our 35S:GUS silencer loci. To test forsilencing mediated by coding region homology, weanalyzed the F1 progeny of crosses between our si-lencers and plants expressing the tomato spotted wiltvirus (TSWV) nucleocapsid protein (N) gene undercontrol of the 35S promoter (Levin, 2001). Homo-logy between this transgene and our silencer locus isrestricted to non-coding regions. If coding region ho-mology is required for silencing, N gene expressionshould be unaffected in progeny of these crosses.
313
Table 2. Summary of GUS histochemical staining observed in F1progeny (26 dpg) from crosses between target lines and the silencerlines C40 and C190
Target locus × C40a × C190
C91 ±/± −/−C83 ±/nd −/−M36 +/± −/−M117 ±/± −/−M56 +/nd −/−M76 +/± −/−M137 nd/nd +/−
a silencer as male/silencer as female. (+) Not different from controlcross. Based on visual inspection. (±) Low, patchy staining. (−)Staining not detectable. nd: not determined.
Table 3. Results of crosses between 35S:TSWV-N line C9 and thesilencer loci C40 and C190
Crossa Viral symptomsb
N-C9×WT Type III (fully resistant)
N-C9×C40 Type 0 (fully susceptible)
N-C9×C190 Type 0 (fully susceptible)
WT×N-C9 Type III (fully resistant)
C40×N-C9 Type 0 (fully susceptible)
C190×N-C9 Type 0 (fully susceptible)
a Crosses are indicated by listing the female parent first.b Consistent observations from four challenged plants.
We assayed TSWV lines N-C9 and N-M4 (Table 1)by crossing each line to lines homozygous for C40or C190 silencer loci. Progeny of crosses betweeneach silencer and line N-C9 showed strongly reducedN gene expression, as evidenced by loss of virusresistance (Table 3). This result indicates that cod-ing region homology is not required for silencing tooccur.
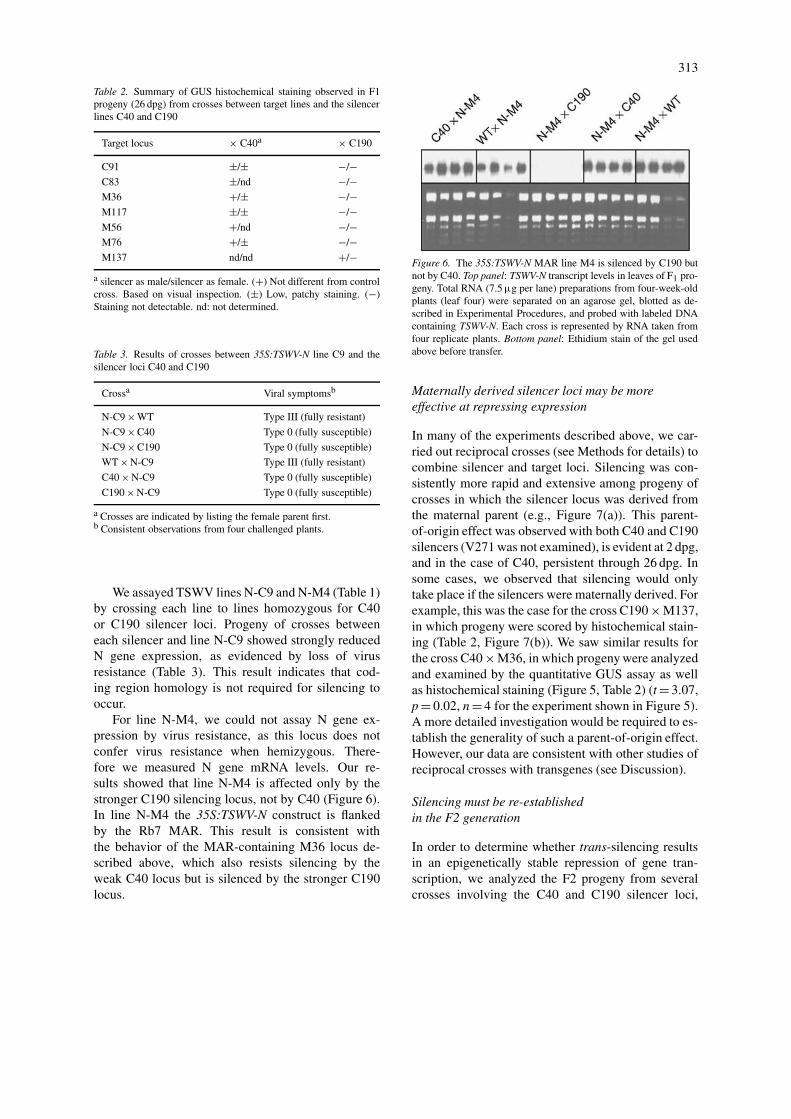
For line N-M4, we could not assay N gene ex-pression by virus resistance, as this locus does notconfer virus resistance when hemizygous. There-fore we measured N gene mRNA levels. Our re-sults showed that line N-M4 is affected only by thestronger C190 silencing locus, not by C40 (Figure 6).In line N-M4 the 35S:TSWV-N construct is flankedby the Rb7 MAR. This result is consistent withthe behavior of the MAR-containing M36 locus de-scribed above, which also resists silencing by theweak C40 locus but is silenced by the stronger C190locus.
Figure 6. The 35S:TSWV-N MAR line M4 is silenced by C190 butnot by C40. Top panel: TSWV-N transcript levels in leaves of F1 pro-geny. Total RNA (7.5 µg per lane) preparations from four-week-oldplants (leaf four) were separated on an agarose gel, blotted as de-scribed in Experimental Procedures, and probed with labeled DNAcontaining TSWV-N. Each cross is represented by RNA taken fromfour replicate plants. Bottom panel: Ethidium stain of the gel usedabove before transfer.
Maternally derived silencer loci may be moreeffective at repressing expression
In many of the experiments described above, we car-ried out reciprocal crosses (see Methods for details) tocombine silencer and target loci. Silencing was con-sistently more rapid and extensive among progeny ofcrosses in which the silencer locus was derived fromthe maternal parent (e.g., Figure 7(a)). This parent-of-origin effect was observed with both C40 and C190silencers (V271 was not examined), is evident at 2 dpg,and in the case of C40, persistent through 26 dpg. Insome cases, we observed that silencing would onlytake place if the silencers were maternally derived. Forexample, this was the case for the cross C190×M137,in which progeny were scored by histochemical stain-ing (Table 2, Figure 7(b)). We saw similar results forthe cross C40×M36, in which progeny were analyzedand examined by the quantitative GUS assay as wellas histochemical staining (Figure 5, Table 2) (t= 3.07,p= 0.02, n= 4 for the experiment shown in Figure 5).A more detailed investigation would be required to es-tablish the generality of such a parent-of-origin effect.However, our data are consistent with other studies ofreciprocal crosses with transgenes (see Discussion).
Silencing must be re-establishedin the F2 generation
In order to determine whether trans-silencing resultsin an epigenetically stable repression of gene tran-scription, we analyzed the F2 progeny from severalcrosses involving the C40 and C190 silencer loci,

314
Figure 7. Parent-of-origin effect on transgene silencing in F1 pro-geny carrying the C190 silencer locus. (a) Representative seedlings(2 dpg) resulting from reciprocal crosses between plants carryingthe C91 locus and plants carrying the C190 silencer locus. Bothparental lines were homozygous for the indicated transgene locus.Progeny seedlings were stained with X-Gluc as described in Meth-ods. Note the absence of staining in the root meristem and reducedintensity of staining in cotyledons of progeny from the C190×C91cross, as compared to seedlings derived from the reciprocal crossC91×C190 or from the WT×C91 cross. (b) Seedlings and leafdiscs from progeny of the C190×M137 cross stained as above.Leaf discs were obtained using a cork borer from the same po-sition on leaf number 3. While very weak residual staining isobserved in cotyledons, there is no staining in leaf primordia of theseven-day-old seedling from the C190×M137 cross.
as shown in Table 4. In all cases, the F1 progenyshowed little or no GUS activity, indicating that si-lencing had been established in the F1 generation(see Figure 5 and Table 2). Previous reports (e.g.,Vaucheret, 1994; Park et al., 1996) have shown thattranscriptional silencing is sometimes epigeneticallystable, persisting for one or more generations afterthe silencer locus is removed by genetic segregation.In contrast, post-transcriptional silencing is typicallyunstable and needs to be re-established in the nextgeneration (e.g., Hobbs et al., 1993; reviewed alsoby Matzke & Matzke, 1998; Vaucheret et al., 1998).If our silencer loci cause epigenetically stable silen-cing, none of the F2 seedlings should express GUS
even though Mendelian segregation should produce asignificant fraction of seedlings containing the targetgusA gene but lacking the silencer locus.
In our crosses, 3/16 of the F2 progeny shouldinherit the target locus but not the silencer. In theabsence of epigenetically stable silencing, these indi-viduals should express GUS throughout their develop-ment. Another 9/16 of the progeny individuals shouldinherit the silencer locus along with the target; theseindividuals might express GUS early in embryo orseedling development but should lose activity whensilencing is subsequently re-established. In our exper-iments, we find that silencing does not persist in theabsence of the silencer (Table 4). Table 4 also showsthat we observed F2 segregation ratios close to 3:1when we tested seedlings at 2 dpg, indicating that alltarget loci are expressed at this time. By 22 dpg, how-ever, the ratio drops dramatically to a value close tothe 3:13 ratio predicted if all plants inheriting the si-lencer locus are silenced. These results are consistentwith the hypothesis that these silencing loci lose theireffectiveness at some point during the sexual cycle,and require a finite time to re-establish a silencingcondition in the next generation.
Discussion
We describe the results of systematic crosses involvingtransgenic silencing loci of varying strengths andtarget loci containing various numbers of transgenecopies with or without MARs. We tested two new 35S-containing silencing loci, C40 and C190 in addition tothe V271 ‘supersilencer’ locus described by Vaucheret(1993). A comparison of all three silencers with thesame target loci (i.e., M36 and C91) shows that si-lencer strength differs, so that V271 > C190 > C40.We do not know the molecular basis for these dif-ferences. However, all three loci are complex andknown to include rearrangements (Table 1; Vaucheretet al., 1993), and it is possible that the number orcomposition of these rearrangements may correlatewith silencing strength. Inverted repeats, in particular,have been associated with other examples of transcrip-tional silencing (e.g., Jakowitsch et al., 1999; Luffet al., 1999).
MARs do not completely eliminate trans-silencing.This result extends the earlier observation that sev-eral different MARs failed to prevent trans effectsfrom the V271 ‘supersilencer’ locus (Vaucheret et al.,1998). Importantly, however, four out of six MAR-

315
Table 4. Chi-squared analysis of expression frequencies in the F2 generation of silencing crosses with the silencerlines C40 and C190
Crossa Observedb Expectedc p valued Expectede p valuef
C40×C83 (d2) 26/9 6.6/28.4 5.2× 10−17 26.3/8.75 9.2× 10−1∗C40×C83(d26) 10/38 9/39 7.1× 10−1∗ 36/12 4.5× 10−18
C40×C91 (d2) 23/10 6.2/26.8 7.1× 10−14 24.8/8.3 4.8× 10−1∗C36×C40 (d2) 25/8 6.2/26.8 5.4× 10−17 24.8/8.3 9.2× 10−1∗M36×C190 (d2) 21/7 5.3/22.8 2.4× 10−14 21/7 1∗M36×C190 (d26) 6/40 8.6/37.4 3.3× 10−1∗ 34.5/11.5 3.0 × 10−22
a Crosses are indicated by listing the female parent first. The time of analysis (in parentheses) is given as dayspost germination.b GUS+/GUS− .c Calculated from the expected frequency of progeny not receiving a silencing locus, silencing effective (3:13ratio).d (∗) Observed value not significantly different from expected value (3:13 ratio).e Calculated from the expected frequency of progeny not receiving a silencing locus, silencing ineffective (3:1ratio).f (∗) Observed value not significantly different from expected value (3:1 ratio).
containing transgene loci we studied show somewhatreduced sensitivity to some silencer loci. Specifically,the MAR-flanked 35S:TSWV line, N-M4, and not N-C9, appears protected from silencing by our weakestsilencer, C40, and the 35S:GUS loci from lines M36and M56 are not silenced when the C40 silencer istransmitted through pollen. In addition, the 35S:GUSlocus in M137 is not silenced even by C190, if thesilencer is paternally derived. These parent-of-origineffects are discussed below. In contrast, all three ofthe target loci without MARs are susceptible to si-lencing from both C40 and C190 derived from eitherparent.
Even though the V271 silencer causes transcrip-tional silencing of all transgenes with a 35S promoter(Vaucheret, 1993), our results indicate that GUS ex-pression persists for a finite time after the silencer andtarget loci have been combined in the same nucleus.For example, Figure 2 shows strong GUS staining inseedlings derived from crosses with V271. Similar re-sults have been reported by Vaucheret (1994). Controlexperiments with the Top10 promoter (Figure 2(c))show that GUS protein or mRNA are not carried overfrom the mother plant. Similarly, we show that thesilencing effect of our C40 and C190 loci must be re-established in each generation. Such requirement is aknown characteristic of post-transcriptional silencingsystems, which depend on continued transcription ofthe affected gene. However, HC-Pro and nuclear run-on data (Figures 2 and 3) are most consistent witha transcriptional silencing mechanism. To reconcile
these two sets of observations, we propose that thesetrans-silencing loci produce one or more aberrant tran-scripts containing 35S promoter sequences, and thatthese ‘promoter transcripts’ cause trans-silencing of35S promoters as suggested by Park et al. (1996),Wassenegger and Pélissier (1998), and Mette et al.(1999, 2001). According to this hypothesis, therewould be a point during the sexual reproduction pro-cess (e.g., gametogenesis or meiosis) at which theTGS-inducing aberrant RNA would no longer be pro-duced. Therefore, this type of TGS, like PTGS, wouldneed to be re-established by renewed production ofthe aberrant RNA at some point subsequent to mei-osis and fertilization. We have not demonstrated thepresence of promoter transcripts in our system. How-ever, promoter-specific RNA transcripts have so farnot been detected in any systems other than those spe-cifically engineered to produce them, and it remainspossible that transcript levels below the threshold ofdetection might mediate silencing in our lines. If si-lencing involves transcripts from the silencer locus,known responses of the 35S promoter may accountfor important features of our system. For example, the35S promoter is relatively inactive in embryos (Frischet al., 1995), which suggests that a decline in silen-cer transcripts might explain the delay we observe inthe onset of silencing after germination. Similar con-siderations could explain the parent-of-origin effectwe observed in reciprocal crosses because the 35Spromoter is known to be poorly expressed in devel-oping pollen (Guerrero et al., 1990). In addition,

316
Vielle-Calzada et al. (2000) have presented enhancertrapping and other evidence indicating that the en-tire paternal genome is poorly expressed during theearliest phases of embryo development; such an ef-fect might help to weaken the silencing effect of apaternally inherited silencer locus. If promoter tran-scripts promote silencing, inhibiting their productionshould reduce silencing. This logic suggests that oneeffect of MARs may be to reduce transcriptional read-through between gene copies, by binding to matrix orother proteins in the nucleus, or simply by acting astranscriptional terminators. According to this model,MARs should decrease the frequency with which com-plex loci acquire silencer activity. The model alsopredicts that MAR loci should remain vulnerable totranscriptional silencing in trans by promoter tran-scripts from another locus. Our data and those ofVaucheret et al. (1998) are consistent with this predic-tion, but the fact that we see partial resistance to trans-silencing in some MAR lines suggests that MARs mayalso influence processes such as DNA methylation andhistone deacetylation which may modify the rate andextent of the response to trans-silencing stimuli. Evid-ence that MARs may counteract silencing by such amechanism was recently provided by Forrester et al.(1999), who observed that both histone deacetylationand DNA methylation of MAR-containing immun-oglobulin transgenes were reduced relative to trans-genes lacking MARs. Future experiments should bedesigned to investigate this hypothesis, as well as totest the possibility that MARs reduce the productionof aberrant transcripts from complex loci.
From the standpoint of engineering plants with co-ordinately regulated genes, MARs may provide someutility via multiple mechanisms. Recently, Mlynárováet al. (2002) have shown that flanking MARs maycreate an artificial chromatin domain allowing co-ordinated expression of two genes. Our findingsshow that even with unlinked loci MARs may pro-tect well-expressed loci from weakly silencing loci,thereby reducing the likelihood of transcriptionalsilencing upon the combination of genes throughcrossing.
Acknowledgements
This work was supported by the NCSU MAR Con-sortium, a research and training grant from the Na-tional Science Foundation/Department of Energy/U.S.Department of Agriculture Collaborative Research in
Plant Biology Program (NSF BIR9420689), and aU.S. Department of Agriculture Competitive ResearchGrant (92-37301-7710) to W.F.T. and G.C.A. B.Ü.received stipend support from the Turkish Repub-lic Ministry of National Education. We thank DrVicki Vance (University of South Carolina) for gener-ously providing us with plants expressing P1/HC-Pro.Thanks also go to Jennifer Levin for the 35S:TSWV-Nlines, and for her comments on the manuscript. KristenHancock and Jennifer Jones provided invaluable as-sistance in maintaining the plants. Brenda Warrick andAngel Harper aided with extractions used for the datain Figure 5.
References
Allen GC, Hall Jr G, Michalowski S, Newman W, Spiker S,Weissinger AK et al. (1996) High-level transgene expression inplant cells: effects of a strong scaffold attachment region fromtobacco. Plant Cell 8: 899–913.
Allen GC, Spiker S and Thompson WF (2000) Use of matrix at-tachment regions (MARs) to minimize transgene silencing. PlantMol Biol 43: 361–376.
Anandalakshmi R, Pruss GJ, Ge X, Marathe R, Mallory AC, SmithTH et al. (1998) A viral suppressor of gene silencing in plants.Proc Natl Acad Sci USA 95: 13079–13084.
Ascenzi R and Gantt JS (1999) Molecular genetic analysis of thedrought-stress inducible linker histone variant in Arabidopsisthaliana. Plant Mol Biol 41: 159–169.
Carrington JC, Freed DD and Oh C-S (1990) Expression of poly-viral polyproteins in transgenic plants reveals three proteolyticactivities required for complete processing. EMBO J 9: 1347–1353.
Chandler VL, Eggleston WB and Dorweiler JE (2000) Paramutationin maize. Plant Mol Biol 43: 121–145.
Cocciolone SM and Cone KC (1993) Pl-Bh, an anthocyaninregulatory gene of maize that leads to variegated pigmentation.Genetics 135: 575–588.
Collins GB and Sadasivaiah RS (1972) Meiotic analysis of hap-loid and doubled haploid forms of Nicotiana otophora and N.tabacum. Chromosoma 38: 387–404.
Cox KH and Goldberg RB (1988) Analysis of plant gene expres-sion. In: Shaw CH (ed.), Plant Molecular Biology: A PracticalApproach. (pp. 1–35) IRL Press, Oxford.
Dehio C and Schell J (1994) Identification of plant genetic lociinvolved in a posttranscriptional mechanism for meioticallyreversible transgene silencing. Proc Natl Acad Sci USA 91:5538–5542.
Forrester WC, Genderen CV, Jenuwein T and Grossschedl R (1994)Dependence of enhancer-mediated transcription of the immun-oglobin µ gene on nuclear matrix attachment regions. Science265: 1221–1225.
Forrester WC, Fernández LA and Grossschedl R (1999) Nuclearmatrix attachment regions antagonize methylation-dependent re-pression of long-range enhancer–promoter interactions. GenesDev 13: 3003–3014.
Frisch DA, van der Geest AHM, Dias K and Hall TC (1995)Chromosomal integration is required for spatial regulation ofexpression from the β-phaseolin promoter. Plant J 7: 503–512.

317
Gallo-Meagher M, Sowinski DA and Thompson WF (1992) Thepea ferredoxin gene exhibits different light responses in pea andtobacco. Plant Cell 4: 383–388.
Guerrero FD, Crossland L, Smutzer GS, Hamilton DA andMascarenhas JP (1990) Promoter sequences from a maize pollen-specific gene direct tissue-specific transcription in tobacco. MolGen Genet 224: 161–168.
Hall Jr G, Allen GC, Loer DS, Thompson WF and Spiker S (1991)Nuclear scaffolds and scaffold-attachment regions in higherplants. Proc Natl Acad Sci USA 88: 9320–9324.
Hobbs SLA, Warkentin TD and DeLong CMO (1993) Transgenecopy number can be positively or negatively associated withtransgene expression. Plant Mol Biol 21: 17–26.
Hollick JB, Dorweiler JE and Chandler VL (1997) Paramutation andrelated allelic interactions. Trends Genet 13: 302–308.
Ingelbrecht I, Van Houdt H, Van Montagu M and Depicker A (1994)Posttranscriptional silencing of reporter transgenes in tobaccocorrelates with DNA methylation. Proc Natl Acad Sci USA 91:10502–10506.
Jakowitsch J, Papp I, Moscone EA, van der Winden J, Matzke M andMatzke AJM (1999) Molecular and cytogenetic characterizationof a transgene locus that induces silencing and methylation ofhomologous promoters in trans. Plant J 17: 131–140.
Kasschau KD and Carrington JC (1998) A counterdefensive strategyof plant viruses: suppression of posttranscriptional gene silen-cing. Cell 95: 461–470.
Levin JS (2001) The effects of matrix attachment regions on RNA-mediated virus resistance. PhD Dissertation, North CarolinaState University, Raleigh, NC, USA.
Love J, Scott AC and Thompson WF (2000) Stringent control oftransgene expression in Arabidopsis thaliana using the Top10promoter system. Plant J 21: 579–588.
Luff B, Pawlowski L and Bender J (1999) An inverted repeat trig-gers cytosine methylation of identical sequences in Arabidopsis.Mol Cell 3: 505–511.
Matzke MA and Matzke AJM (1998) Epigenetic silencing ofplant transgenes as a consequence of diverse cellular defenceresponses. Cell Mol Life Sci 54: 94–103.
Matzke MA, Primig M, Trnovsky J and Matzke AJM (1989) Revers-ible methylation and inactivation of marker genes in sequentiallytransformed tobacco plants. EMBO J 8: 643–649.
Matzke MA, Neuhuber F and Matzke AJM (1993) A variety ofepistatic interactions can occur between partially homologoustransgene loci brought together by sexual crossing. Mol GenGenet 236: 379–386.
Matzke MA, Mette FM, Jakowitsch J, Kanno T, Moscone EA,van der Winden J et al. (2001) A test for transactivationin plants: DNA pairing may lead to trans-activation or si-lencing of complex heteroalleles in tobacco. Genetics 158:451–461.
Mette MF, van der Winden J, Matzke MA and Matzke AJM(1999) Production of aberrant promoter transcripts contributesto methylation and silencing of unlinked homologous promotersin trans. EMBO J 18: 241–248.
Mette MF, Aufsatz W, van der Winden J, Matzke MA andMatzke AJM (2000) Transcriptional silencing and promotermethylation triggered by double-stranded RNA. EMBO J 19:5194–5201.
Mette FM, Matzke AJM and Matzke MA (2001) Resistance ofRNA-mediated TGS to HC-Pro, a viral suppressor of PTGS,suggests alternative pathways for dsRNA processing. Curr Biol11: 1119–1123.
Mlynárová L, Loonen A, Mietkiewska E, Stiekema WJ, Jansen RCand Nap J-P (2002) Assembly of two transgenes in an artifi-
cial chromatin domain gives highly coordinated expression intobacco. Genetics 160: 727–740.
Mösinger E, Batscauer A, Vierstra R, Apel K and Schäfer E (1987)Comparison of the effects of exogenous native phytochrome andin vivo irradiation on in vitro transcription in isolated nuclei frombarley (Hordeum vulgare). Planta 170: 505–514.
Nap J-P, Conner AJ, Mlynárová L, Stiekema WJ and Jansen RC(1997) Dissection of a synthesized quantitative trait to character-ize transgene interactions. Genetics 147: 315–320.
Park Y-D, Papp I, Moscone EA, Iglesias VA, Vaucheret H, MatzkeAJM et al. (1996) Gene silencing mediated by promoter homo-logy occurs at the level of transcription and results in meioticallyheritable alterations in methylation and gene activity. Plant J 9:183–194.
Petersen K, Leah, R, Knudsen S and Cameron-Mills V (2002)Matrix attachment regions (MARs) enhance transformation fre-quencies and reduce variance of transgene expression in barley.Plant Mol Biol 49: 45–58.
Phi-Van L, von Kries JP, Ostertag W and Strätling WH (1990) Thechicken lysozyme 5′ matrix attachment region increases tran-scription from a heterologous promoter in heterologous cells anddampens position effects on the expression of transfected genes.Mol Cell Biol 10: 2302–2307.
Schöffl F, Schroder G, Kliem M and Rieping M (1993) A SAR-sequence containing 395 bp DNA fragment mediates enhanced,gene-dosage-correlated expression of a chimeric heat shock genein transgenic tobacco plants. Transgenic Res 2: 93–100.
Sieburth LE and Meyerowitz EM (1997) Molecular dissectionof the agamous control region shows that cis-elements forspatial regulation are located intragenically. Plant Cell 9:355–365.
Stief A, Winter DM, Strätling WH and Sippel AE (1989) A nuclearDNA attachment region mediates elevated position-independentgene activity. Nature 341: 343–345.
Ülker B (1998) The effects of matrix attachment regions (MARs) ontransgene expression in tobacco plants. PhD Dissertation, NorthCarolina State University, Raleigh, NC, USA.
Ülker B, Allen GC, Thompson WF, Spiker S and Weissinger AK(1999) A tobacco matrix attachment region reduces the loss oftransgene expression in the progeny of transgenic tobacco plants.Plant J 18: 253–263.
Valvekens D, Van Montagu M and van Lijsebettens M (1988)Agrobacterium tumefaciens-mediated transformation of Ara-bidopsis thaliana root explants using kanamycin selection. ProcNatl Acad Sci USA 85: 5536–5540.
van der Geest AHM, Hall GE, Spiker S and Hall TC (1994) Theβ-phaseolin gene is flanked by matrix attachment regions. PlantJ 6: 413–423.
Vaucheret H (1993) Identification of a general silencer for 19S and35S promotors in a transgenic tobacco plant: 90 bp of homologyin the promoter sequence are sufficient for trans-inactiviation. CR Acad Sci Paris 316: 1471–1483.
Vaucheret H (1994) Promoter-dependent trans-inactivation in trans-genic tobacco plants: kinetic aspects of gene silencing and genereactivation. C R Acad Sci Paris 317: 310–312.
Vaucheret H, Elmayan T, Thierry D, van der Geest A, Hall T,Conner AJ et al. (1998) Flank matrix attachment regions (MARs)from chicken, bean, yeast or tobacco do not prevent homology-dependent trans-silencing in transgenic tobacco plants. Mol GenGenet 259: 388–392.
Verchot J, Koonin EV and Carrington JC (1991) The 35-kDaprotein from the N-terminus of the polyviral polyproteinfunctions as a third virus-encoded proteinase. Virology 185:527–535.

318
Vielle-Calzada J-P, Baskar R and Grossniklaus U (2000) Delayedactivation of the paternal genome during seed development.Nature 404: 91–94.
Wassenegger M and Pélessier T (1998) A model for RNA-mediatedgene silencing in higher plants. Plant Mol Biol 37: 349–362.
Wassenegger M, Heimes S, Riedel L and Sanger HL (1994) RNA-directed de novo methylation of genomic sequences in plants.Cell 76: 567–576.
Weinmann P, Gossen M, Hillen W, Bujard H and GatzC (1994) A chimeric transactivator allows tetracycline-
responsive gene expression in whole plants. Plant J 5:559–569.
Wolffe AP and Matzke MA (1999) Epigenetics: regulation throughrepression. Science 286: 481–486.
Wu C-T and Morris JR (1999) Transvection and other homologyeffects. Curr Opin Genet Dev 9: 237–246.
Ye F and Signer ER (1996) RIGS (repeat-induced genesilencing) in Arabidopsis is transcriptional and alterschromatin configuration. Proc Natl Acad Sci USA 93:10881–10886.




















