
Newly recognized Pleistocene human teeth from Tabun Cave, Israel

ORIGINAL INVESTIGATION
Esperanza Rodrıguez Æ Fernanda Romarıs
Sonia Lorenzo Æ Javier Moreno Æ Pedro Bonay
Florencio M. Ubeira Æ Teresa Garate
A recombinant enolase from Anisakis simplex is differentiallyrecognized in natural human and mouse experimental infections
Received: 23 January 2003 / Published online: 28 July 2005� Springer-Verlag 2005
Abstract A 1,963-bp cDNA was isolated from anAnisakis simplex cDNA library by immunoscreeningwith a hyperimmune rabbit serum raised against a crudeextract of A. simplex L3 larvae. The open reading frameencodes a putative protein of 436 amino acid residues,which exhibits high similarity (70–80%) to enolasemolecules from various other organisms, including hel-minth parasites. After subcloning and expression of theA. simplex cDNA in PGEX-4T-3, the resulting gluta-thione S-transferase fusion protein, purified byglutathione-Sepharose-4B chromatography, showedfunctional enolase activity. The immunogenicity of therecombinant A. simplex enolase was analyzed byimmunoblotting using sera obtained from (a) miceimmunized with crude extracts (CE) of A. simplex, orother nematode species, (b) mice immunized withexcretory–secretory (ES) antigens from A. simplex, or(c) mice infected with L3 larvae by the intraperitonealroute. In addition, we used ELISA, to investigate thepresence of IgG1 and IgE antibodies against this mole-cule in sera from patients infected with A. simplex.Mouse sera obtained after infection with L3 or raisedagainst CE antigens, but not sera raised against ESantigens, showed strong reactivity with the recombinant
A. simplex enolase. We also obtained good reactivity inWestern blotting with sera from mice immunized withCE antigens from Ascaris suum and Toxocara canis, butnot with sera from mice immunized with CE antigensfrom Trichuris muris, Trichinella spiralis or Hyster-othylacium aduncum. In contrast to the experimentalinfections/immunizations in mice, we were unable todetect anti-enolase IgE antibodies in sera from humanpatients infected with A.simplex (15 sera), and the levelsof anti-enolase IgG1 antibodies in these sera were lowand apparently nonspecific. These results seem to indi-cate that, during natural infection in humans, A. simplexlarvae do not offer sufficient antigenic stimulus to induceanti-enolase antibodies.
Keywords Anisakis simplex Æ cDNA Æ Enolase ÆAnisakiosis Æ Diagnosis
Introduction
Anisakis simplex is considered to be important for hu-man health, since it is the etiological agent of a painfuldisease known as anisakiosis. Humans may be infectedby ingestion of the third-stage larvae (L3) present in raw,undercooked or marinated fish [30]. Since the first reportof this disease in 1960 [38], thousands of cases have beenreported around the world, mainly in Japan, whereconsumption of raw fish is common [33]. In recent years,hundreds of new cases have been reported in Spain,where anisakiosis can be considered an emergent disease[9]. In Spain, it was recently demonstrated that infec-tions by A. simplex are produced mainly by ingestion ofboquerones (marinated anchovies) [36]. Althoughexperimental infections in nonmarine mammals havedemonstrated that the parasite normally dies 2–4 weeksafter infection with L3 larvae [15], during this period, theparasite normally moults to the L4 stage and inducesclinical manifestations depending on the location of thelarvae in the host (stomach, intestine and ectopic
Note: Nucleotide sequence data reported in this paper were sub-mitted to the GenBank, EMBL and DDBJ databases under theaccession number AJ496792.
E. Rodrıguez Æ J. Moreno Æ T. Garate (&)Present address: Servicio de Parasitologıa, Centro Nacional deMicrobiologıa,Instituto de Salud Carlos III, Ctra. Majadahonda PozueloKm 2,2, 28220 Majadahonda, Madrid, SpainE-mail: [email protected]
F. Romarıs Æ S. Lorenzo Æ F. M. UbeiraLaboratorio de Parasitologıa, Facultad de Farmacia,Universidad de Santiago de Compostela, 15782 Santiago deCompostela, Spain
P. BonayCentro de Biologıa Molecular, Universidad Autonomade Madrid, Cantoblanco, Madrid, Spain
Med Microbiol Immunol (2006) 195: 1–10DOI 10.1007/s00430-005-0236-7

locations). The pathogenetic mechanisms leading tothese clinical manifestations include mechanical traumainduced by L3 larvae at the site of infection [19], andmore importantly hypersensitivity reactions induced byexcretory–secretory (ES) antigens produced both by theL3 and L4 stages, and by somatic antigens producedwhen the larvae die and degrade in host tissues [4]. Themost serious allergic reactions induced by parasiteantigens are type-I IgE-mediated reactions, includingurticaria, angioedema and anaphylaxis (for a review,see [3]), but type-IV hypersensitivity responses are alsocertainly involved [4, 8]. There have been severalstudies on the antigenic composition of the differentlife cycle stages of A. simplex [35], but so far fewantigens have been characterized at the molecular level[1, 2, 20, 25, 29].
Enolase (2-phospho-D-glycerate (2-PGE) hydrolase)is a ubiquitous dimeric glycolytic enzyme that catalyzesthe dehydration of 2-PGE to phosphoenolpyruvate(PEP), an important metabolic intermediate [22]. Eno-lase requires magnesium for both catalysis and stabiliz-ing the dimer, and is probably found in all organismsthat metabolize sugars. Enolases from a variety ofsources, ranging from bacteria to higher vertebrates,show highly conserved amino acid sequence [37]. Inaddition, certain nonglycolytic properties of enolasehave been described, including properties related tosurface expression and plasminogen binding, whichcould indicate that enolases play an important role in theinitiation of disease processes, through modulation ofthe pericellular and intravascular fibrinolytic system [28].In line with this possibility, enolases have been impli-cated in invasive candidiasis, cancer, bacterial diseases,and autoimmune disorders [27]. Furthermore, recentfindings indicate that enolases are important allergeniccomponents of yeasts such as Saccharomyces cerevisiae[6] and Candida albicans [16], and molds such as Cla-dosporium herbarum and Alternaria alternata [32]. In thepresent study, we cloned and sequenced an A. simplexcDNA with high similarity to enolase genes from otherorganisms, including parasites, and analyzed its func-tional activity and antigenic relevance in mice and hu-mans infected with A. simplex. Our results demonstratethat recombinant enolase from A. simplex is functionallyactive and binds human plasminogen but, unlike Can-dida or mold enolases, does not seem to be antigenicallyrelevant for humans. We discuss the possibility that thisreflects structural similarity between the human andA. simplex enolases, and week antigenic stimulation bythis molecule during A. simplex infections.
Material and methods
Construction and immunoscreening of an A. simplexcDNA library
Messenger RNA (mRNA) was obtained from A. simplexL3 using the Fast Track mRNA isolation kit (Invitro-
gen, San Diego, CA, USA). Poly(A)+ RNA from L3was converted into double-stranded cDNA with theZAP-cDNA synthesis kit (Stratagene, La Jolla, CA,USA) and then ligated to the k-ZAP expression vectorusing the Uni-ZAP� XR library kit (Stratagene), fol-lowing the standard Stratagene protocol. The originallibrary was amplified and contained 1.3·1010 pfu/mland 1% nonrecombinant phages. The amplified A.simplex cDNA library was immunoscreened with ahyperimmune rabbit serum raised against crude extractfrom A. simplex L3. Plaques adsorbed onto 10 mMisopropyl thio-b-D-galactoside (IPTG)-impregnatednitrocellulose filters (Schleicher & Schuell, Dassel,Germany) were blocked with 3% BSA in TBS (Tris–HCl 50 mM, NaCl 150 mM, pH 7.5) at 37�C for30 min and then incubated with hyperimmune rabbitserum (1:300 in TBS) at 4�C overnight. The serum hadbeen preadsorbed with an Escherichia coli lysate. Thenitrocellulose filters were washed in TBS containing0.05% Tween-20 (TBS-T), and incubated for 2 h atroom temperature with alkaline–phosphatase-conju-gated goat antirabbit IgG at 1:10,000 (Pierce, Rock-ford, IL, USA). After washing again, the immunecomplexes were visualized using nitroblue tetrazoliumsalt (NBT) and 5 bromo-4-chloro-3-indolyl phosphatep-toluidine salt (BCIP) (Sigma Chemical Corp., StLouis, MO, USA) as substrate.
DNA sequencing
Phagemids were recovered from positive k-ZAP clonesby ‘‘in vivo’’ excision, using helper phage rescue [31].Recombinant pBluescript plasmids were sequencedusing the Big-Dye Terminator Cycle Sequencing ReadyReaction Kit (PE Biosystems, Foster City, CA, USA)and an Applied Biosystems 377 DNA sequencer (PEBiosystems). DNA sequence and predicted amino acidsequence comparisons were carried out with the EMBLand SWISS-PROT databanks using software packagesfrom the Genetic Computer Group [10].
Subcloning, expression and purificationof the recombinant A. simplex enolase
The full cDNA of the A. simplex enolase gene wassubcloned into pGEX-4T-3 expression vector (Amer-sham BioScience, Buckinghamshire, UK), using the SalI and Not I sites. This vector codes for glutathioneS-transferase (GST). Prior to the cloning, the full-lengthcoding sequence of the A. simplex enolase was amplifiedby PCR using the recombinant pBluescript plasmid astemplate and a set of primers containing Sal I and Not Irestriction sites, as well as the ATG initiation codon[Eno-pGEX5¢ (5¢-GAGAGAGAGTCGACATGCCG-ATCACTCGTATTCACGCT-3¢)] and the TAAtermination codon [Eno-pGEX3¢ (5¢-GAGAGAGAG-
2

CGGCCGCGCCACAAATTTAGGCTTGCGG-3¢)] ofthe enolase gene. For expression, the pGEX-4T-3-Enoconstruct was transformed into competent BL21(DE3) E. coli cells (Stratagene). Transformants weregrown at 37�C with vigorous shaking to an OD600 of0.5, then induced with 1 mM IPTG and incubated for4 h. The recombinant protein was purified by gluta-thione-Sepharose-4B chromatography (AmershamBioScience), following the manufacturer’s recommen-dations.
Enzyme activity assay
The enolase activity of the purified recombinant proteinwas measured in a single assay as per Pancholi andFischetti [28]. Different amounts of purified recombinantprotein (5 and 30 lg/ml) were used to measure the in-crease in absorption at 240 nm that occurs as a result ofthe conversion of D(+)2–2-phosphoglycerate (2-PGE)(Sigma) to PEP. This reaction was performed in100 mM HEPES buffer, pH 7.0, containing 10 mMMgSO4 and 7.7 mM KCl, and in the presence of 3 mM2-PGE. Absorbance was monitored spectrophotometri-cally at 5-s intervals for a period of 2 min at 240 nm. Asa positive control, baker’s yeast enolase (Sigma) wasused.
Sera
Sera against crude extract (CE) antigens of the nema-tode species A. simplex (L3 larvae), Ascaris suum (adultmale), Toxocara canis (adults), Hysterothylacium adun-cum (L3 larvae), Trichuris muris (adults), and Trichinellaspiralis (L1 larvae), as well as serum against ES antigensof A. simplex were raised in BCF1 mice using standardimmunization protocols. Briefly, seven groups of fivemice were intraperitoneally immunized with 250 ll/mouse of a 1:1 mixture of complete Freund’s adjuvant(CFA) and a solution containing 25 lg of each antigenpreparation (obtained as previously described [14]). Thesame antigen dose in 125 ll (but without CFA) wasinjected via the retrobulbar venous plexus 21 days post-primary immunization. The mice were bled 14 dayspost-secondary immunization and the sera stored at�30�C until use.
Serum against A. simplex L3–L4 larvae was ob-tained by intraperitoneal infection of BCF1 mice withfive live L3 larvae of this parasite. The mice were bledon day 35 post-infection and the five sera were thenpooled.
Sera from 15 human patients with positive IgG1 andIgE reactions against specific A. simplex antigens (casesnos. 1–15) were selected from our anisakiosis sera col-lection. Serum IgG1 and IgE antibody levels in all pa-tients and 31 human controls (healthy blood donors)were assayed using the standard UA3 ELISA technique[23] (see the section ELISA assays).
SDS-PAGE and immunoblotting
The glutathione-affinity-purified enolase-GST (or GSTalone) was electrophoresed on acrylamide gels(70·80·0.75 mm) in a MiniProtean II cell (Bio-RadLaboratories Hercules, CA, USA) and transferred toPVDF membranes (Immobilon-P, Millipore Iberica SA,Madrid, Spain) in a Trans-Blot SD transfer cell (Bio-Rad Laboratories). After blocking of nonspecific bind-ing sites with 5% nonfat dry milk in TBS-T, individualstrips were incubated with pooled sera (1:50 in TBS-Tcontaining 1% of nonfat dry milk) of BCF1 mice thathad been immunized with CE antigens from differentnematode species, immunized with ES antigens fromA. simplex L3 larvae or infected with A. simplex L3larvae. After washing, strips were incubated with peroxi-dase-conjugated rabbit anti-mouse polyclonal antibody(Dako Diagnosticos SA, Barcelona, Spain; dilution1:800) and developed using 3,3¢-diaminobenzidine tetra-hydrochloride (Sigma) according to the manufacturer’sinstructions.
ELISA assays
(a) UA3 ELISA
IgE or IgG1 antibodies to UA3-recognized specificantigens of A. simplex were determined by antigen-capture ELISA [23]. The wells of polystyrene plates(Corning, New York, USA) were coated overnight at4�C with mAb UA3 (1.4 lg/well). After washing withdH2O, nonspecific binding sites were blocked by incu-bation (1 h at 37�C) with 1% nonfat dry milk in TBS-T.An appropriate dilution of A. simplex CE was then ad-ded, and the plates were incubated for 1 h at 37�C. Afterwashing again, the plates were incubated for 90 min at37�C with human sera (undiluted for IgE determination,or 1:200 in TBS-T containing 1% nonfat dry milk forIgG1 determination). After washing again, bound anti-bodies were detected with the appropriate dilution ofmouse anti-human IgE mAb (Ingenasa, Madrid, Spain)or mouse anti-human IgG1 mAb (Sigma), both labeledwith FITC. Bound Ig-FITC was detected by incubationfirst with peroxidase-conjugated rabbit anti-FITC Ig(dilution 1:1,000) (Dako Diagnosticos, SA) and thenwith o-phenylenediamine (OPD) (Sigma) in citrate-phosphate buffer (100 ll/well). The reaction was stop-ped with 3% H2SO4 (25 ll/well) and the optical densities(ODs) measured at 492 nm in a microtiter plate reader(Titertek Multiskan, Labsystems, Finland).
(b) Capture ELISA for recombinant A. simplex enolase
The levels of IgE or IgG1 antibodies to the recombinantA. simplex enolase were also determined by antigen-capture ELISA. An appropriate dilution of soluble re-combinant enolase-GST or recombinant GST (control)
3

extracted from bacterial lysates was added to the wellsof an ELISA plate coated with goat anti-GST immu-noglobulins (0.5 lg/well; Amersham BioScience). After
a washing step and the addition of human sera (undi-luted for IgE determination, or 1:100 dilution for IgG1determination), the plates were processed as described
4

for the UA3 ELISA. The levels of IgG1 antibodies torecombinant A. simplex enolase in the pooled sera fromBCF1 mice immunized with CE antigens from A. sim-plex L3 larvae were detected in a similar assay. Theserum was diluted 1:100 in TBS-T containing 1% nonfatdry milk and added (100 ll/well) to the wells. Afterwashing, bound serum antibodies were detected with aperoxidase-conjugated rabbit anti-mouse Igs polyclonalserum (Dako Diagnosticos, SA) diluted 1:2,000 in TBS-T (100 ll/well), and the reaction revealed as describedabove.
(c) Plasminogen-binding assay
The plasminogen-binding activity of the purifiedrecombinant protein was evaluated by a solid-phasefluorescent enzyme-linked assay (FELISA). Serial dilu-tions (100 ll) of recombinant A. simplex enolase dilutedin binding buffer (0.1 M NaHCO3 pH 8.5) were addedto the wells of microtiter plates and incubated for 1 h atRT. Remaining unreacted sites were blocked by 1 hincubation in TBS containing 3% gelatin and washedtwice in the same buffer. Thereafter, 100 ll of humanplasminogen (Sigma) at a concentration of 0.15 U/mlwas added to each well and incubated for 90 min at RT,washed three times in TBS-T, and then incubated with100 ll of goat anti-human plasminogen antibodies(dilution 1:2,500; Sigma) for 1 h, and washed three timesin TBS-T. After washing, 100 ll of peroxidase-conju-gated rabbit anti-goat IgG (dilution 1:5,000, Sigma) wasadded; the wells were then incubated for 1 h, and finallywashed three times in TBS-T and once in TBS. After6 min incubation, bound antibodies were detected withQuantablu fluorogenic peroxidase substrate (Pierce) asdescribed by the manufacturer. Fluorescence wasmeasured at 325-nm ex and 420-nm em in a Bertholdmicroplate reader.
Cell proliferation assay
Spleens were obtained aseptically from immunized andcontrol BALB/c mice, passed through a fine sieve, and
washed. Cells were cultured at a density of2·106 cells/ml in 0.2 ml of RPMI 1640 (Gibco, Paisley,UK) supplemented with 100 IU/ml of penicillin, 100 lg/ml of streptomycin, 2 mM L-glutamine, 5·10�5 M2-mercaptoethanol and 10% heat-inactivated fetal calfserum (Biological Industries, Israel). Assays were con-ducted in flat-bottomed 96-well microtiter plates, incu-bated (37�C, 5% CO2) for 3 days with either 3 or 6 lg/ml of purified GST, or 3 or 6 lg/ml of purified enolase-GST, or 6 lg/ml of ConA, or without antigen (blank),and pulsed during the last 20 h with 100 lM of 5-bro-mo-2¢-deoxyuridine (BrdU). BrdU incorporation wasdetermined using the BIOTRAK cell proliferationELISA system (Amersham). The results are expressed asthe mean increase in optical density with respect toblank, in triplicate cultures.
Results
Cloning and expression of the A. simplex enolase
A cDNA expression library in vector kZAPII was con-structed with 5 lg of poly(A)+ RNA from A. simplexL3. Aliquots (2·105 pfu) of the cDNA expression librarywere immunoscreened with hyperimmune rabbit serumraised against crude extract from A. simplex L3, and 25positively immunoreacting clones were isolated andpurified. Recombinant pBluescript plasmids, derivedfrom these clones by helper phage rescue, were then se-quenced. After sequencing, three of these clones showedclose similarity to the members of the enolase family.The clone with the largest cDNA insert, named AniH6,contained a cDNA insert of 1,963 bp with an openreading frame encoding a putative protein of 436 aminoacid residues, with a predicted molecular mass of47.4 kDa and isoelectric point of 6.1. The initiationcodon was assigned by homology to known enolase se-quences (Fig. 1). The cDNA had 62 bp of 5¢ untrans-lated sequence and 590 bp of 3¢ untranslated sequence.Comparison of this sequence with the sequences con-tained in the GenBank-EMBL database revealed highsimilarity with the enolase molecules described in vari-ous organisms (including helminth parasites): Caenor-habditis elegans (82.7%), Onchocerca volvulus (81.4%),T. spiralis (75.2%), Fasciola hepatica (72.6%), Schisto-soma japonicum (70.7%), human (69.8%), mouse(68.9%), A. alternata (66.0%) and C. herbarum (65.1%).The alignment of the deduced amino acid sequence ofthe A. simplex enolase with these sequences is shown inFig. 1.
The deduced aminoacid sequence of the A. simplexenolase included the residues considered to be importantfor Mg2+ binding (Ser40, Asp246, Glu297, Asp325) and thecrucial active-site residues for enzyme activity (Glu211,Lys349, His377). The His377 residue is contained within acharacteristic enolase fingerprint motif 376[SHRSGET-ED]384 located in the C-terminal region, which is highlyconserved in all the enolases described to date [5] (motif
Fig. 1 Alignment of the predicted amino acid sequence of theAnisakis simplex enolase and other enolase sequences, using theClustal method as executed by the Lasergene-MegAlign program(DNASTAR, Inc., Madison, WI, USA). All sequences wereobtained from GenBank protein sequence data (accession numbersin brackets): Caenorhabditis elegans (Q27527); Onchocerca volvulus(AAP81756); Trichinella spiralis (AAK50056); Fasciola hepatica(Q27655); Schistosoma japonicum (AAA29874); Human enolase(P06733); Mus musculus (NP_075608); Alternaria alternata(Q9HDT3) and Cladosporium herbarum (P42040). Residue changeswith respect to the A. simplex enolase are shown in boxes, andconserved residues that participate in catalytic activity and inMg2+-binding are shown in filled boxes. Dashes indicate gapsintroduced to fit the sequences
b
5
and relevant residues are marked in filled boxes in Fig. 1).According to the Prosite database [11], the predictedaminoacid sequence of the A. simplex enolase showedtwo putative N-glycosylation sites located at positions
178[NFT]180 and 307[NWT]309. The sequence was alsoanalyzed with the SignalP V1.1 program [26] and noputative signal peptide was found, although a hydro-phobic domain was observed within the region
33[AAVPSGASTGVH]44, as previously reported byPancholi [27] in human enolase.
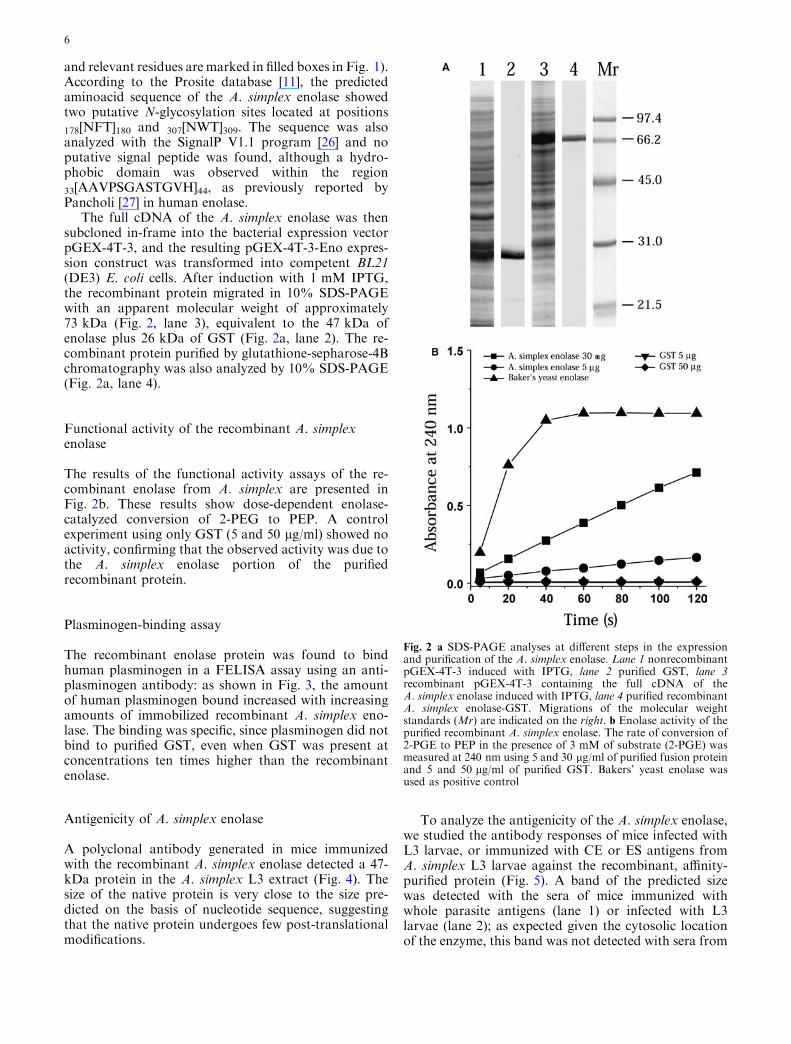
The full cDNA of the A. simplex enolase was thensubcloned in-frame into the bacterial expression vectorpGEX-4T-3, and the resulting pGEX-4T-3-Eno expres-sion construct was transformed into competent BL21(DE3) E. coli cells. After induction with 1 mM IPTG,the recombinant protein migrated in 10% SDS-PAGEwith an apparent molecular weight of approximately73 kDa (Fig. 2, lane 3), equivalent to the 47 kDa ofenolase plus 26 kDa of GST (Fig. 2a, lane 2). The re-combinant protein purified by glutathione-sepharose-4Bchromatography was also analyzed by 10% SDS-PAGE(Fig. 2a, lane 4).
Functional activity of the recombinant A. simplexenolase
The results of the functional activity assays of the re-combinant enolase from A. simplex are presented inFig. 2b. These results show dose-dependent enolase-catalyzed conversion of 2-PEG to PEP. A controlexperiment using only GST (5 and 50 lg/ml) showed noactivity, confirming that the observed activity was due tothe A. simplex enolase portion of the purifiedrecombinant protein.
Plasminogen-binding assay
The recombinant enolase protein was found to bindhuman plasminogen in a FELISA assay using an anti-plasminogen antibody: as shown in Fig. 3, the amountof human plasminogen bound increased with increasingamounts of immobilized recombinant A. simplex eno-lase. The binding was specific, since plasminogen did notbind to purified GST, even when GST was present atconcentrations ten times higher than the recombinantenolase.
Antigenicity of A. simplex enolase
A polyclonal antibody generated in mice immunizedwith the recombinant A. simplex enolase detected a 47-kDa protein in the A. simplex L3 extract (Fig. 4). Thesize of the native protein is very close to the size pre-dicted on the basis of nucleotide sequence, suggestingthat the native protein undergoes few post-translationalmodifications.
To analyze the antigenicity of the A. simplex enolase,we studied the antibody responses of mice infected withL3 larvae, or immunized with CE or ES antigens fromA. simplex L3 larvae against the recombinant, affinity-purified protein (Fig. 5). A band of the predicted sizewas detected with the sera of mice immunized withwhole parasite antigens (lane 1) or infected with L3larvae (lane 2); as expected given the cytosolic locationof the enzyme, this band was not detected with sera from
Fig. 2 a SDS-PAGE analyses at different steps in the expressionand purification of the A. simplex enolase. Lane 1 nonrecombinantpGEX-4T-3 induced with IPTG, lane 2 purified GST, lane 3recombinant pGEX-4T-3 containing the full cDNA of theA. simplex enolase induced with IPTG, lane 4 purified recombinantA. simplex enolase-GST. Migrations of the molecular weightstandards (Mr) are indicated on the right. b Enolase activity of thepurified recombinant A. simplex enolase. The rate of conversion of2-PGE to PEP in the presence of 3 mM of substrate (2-PGE) wasmeasured at 240 nm using 5 and 30 lg/ml of purified fusion proteinand 5 and 50 lg/ml of purified GST. Bakers’ yeast enolase wasused as positive control
6

mice immunized with ES products derived from the L3larvae of A. simplex (lane 3). Two other hyperimmunesera obtained from mice immunized with CE antigensfrom A. suum and T. canis also reacted with the re-combinant protein (lanes 4 and 5, respectively). Incontrast, sera from mice immunized with CE from othernematodes (T. muris, T. spiralis and H. aduncum)showed no reactivity (lanes 6–8). In addition, the abilityof the recombinant A. simplex enolase to induce a T-cell
proliferative response in vitro was also investigated. TheT-cell response induced by the recombinant enolase-GST was no stronger than that induced by the purifiedGST alone (data not shown).
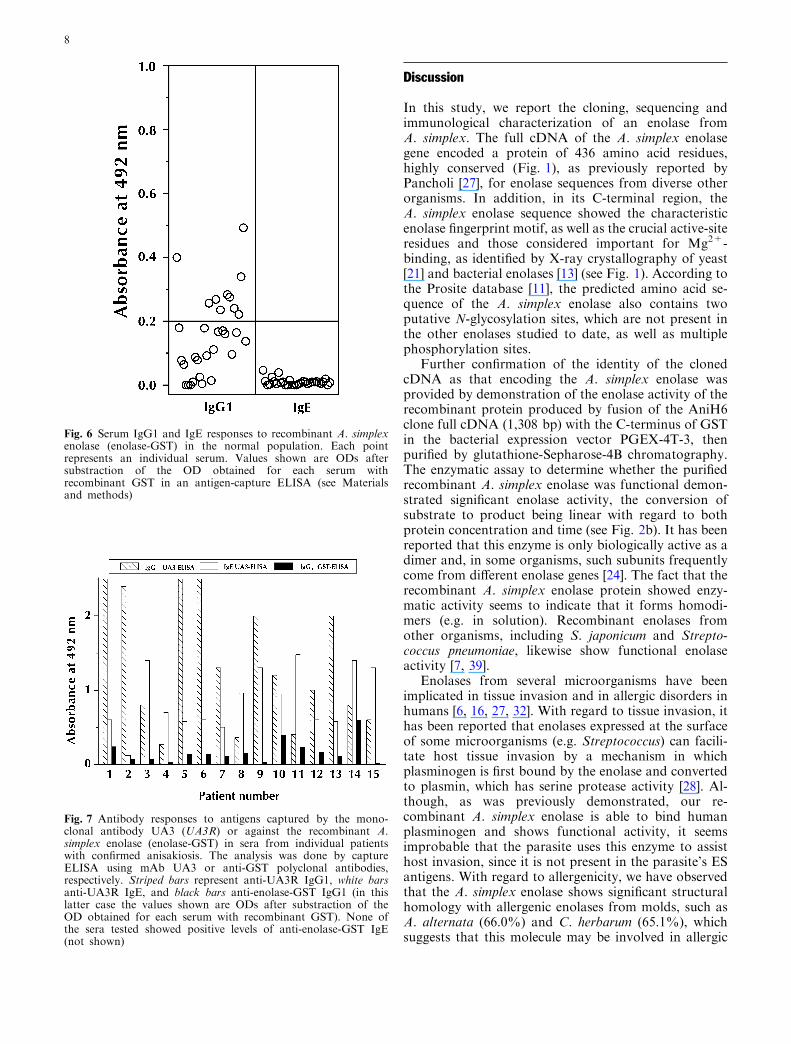
Having shown that the enolase is antigenic inimmunized mice, we looked for IgG1 and IgE antibodiesagainst this molecule in sera from patients infected withA. simplex. As negative controls we used 31 sera fromA. simplex negative subjects (healthy blood donors,Fig. 6). Assuming an initial arbitrary cut-off of 0.2 OD,typical of many ELISA assays [34], we observed thatmost of the sera were negative (mean absorbance 0.155;range: 0.0–0.492). However, 10/31 sera (32%) gave ODvalues higher than cut off. In contrast, all sera werenegative for specific IgE antibodies, with mean ODvalues below 0.05 (range 0.0–0.046). From these data,we calculated the experimental cut-off (mean plus 4 SD)for IgG1 (0.66) and for IgE (0.052). We then analyzed 15individual sera (case nos. 1–15, Fig. 7) from patientswith low-to-high titres of IgG1 and IgE antibodiesagainst specific A. simplex antigens (as demonstrated bythe UA3 ELISA technique [23]). Interestingly, none ofthe sera from anisakiosis patients showed IgE antibodiesagainst the recombinant enolase (mean 0.008; range:0.00–0.069), and these sera were also negative for spe-cific IgG1 antibodies (mean 0.164; range: 0.016–0.599)when compared with the cut off calculated with serafrom uninfected subjects. In addition, the levels of anti-enolase IgG1 antibodies in these sera did not correlate(r=�0.108) with anti-A. simplex antibody levels asdetermined by UA3 ELISA (P>0.7).
Fig. 5 Recognition of the recombinant A. simplex enolase bymouse sera raised against different nematodes. The figure shows animmunoblot analysis of the fusion protein GST A. simplex enolasewith A. simplex CE antigens (lane 1), by infection with L3 larvae(lane 2), or against A. simplex ES antigens (lane 3), or CE antigensfrom Ascaris suum (lane 4), Toxocara canis (lane 5), Trichuris muris(lane 6), Trichinella spiralis (lane 7), or Hysterothylacium aduncum(lane 8). A serum pool from non-immunized mice (lane 9) was usedas negative control. None of the sera used showed reactivity withGST (not shown). Migrations of the molecular weight standards(Mr) are indicated on the left
Fig. 4 Immunoblotting analysis of A. simplex L3 crude extracts(CE) by antibodies raised against the fusion protein GST-A.simplex enolase (lane 1) or GST alone (lane 2)
Fig. 3 Plasminogen-binding activity of the purified recombinantA. simplex enolase, as measured by FELISA. The indicatedamounts of purified GST-enolase (white bars) or GST (black bars)were applied to the wells of the FELISA plate, and plasminogen-binding activity was assayed as described in Material and methods
7
Discussion
In this study, we report the cloning, sequencing andimmunological characterization of an enolase fromA. simplex. The full cDNA of the A. simplex enolasegene encoded a protein of 436 amino acid residues,highly conserved (Fig. 1), as previously reported byPancholi [27], for enolase sequences from diverse otherorganisms. In addition, in its C-terminal region, theA. simplex enolase sequence showed the characteristicenolase fingerprint motif, as well as the crucial active-siteresidues and those considered important for Mg2+-binding, as identified by X-ray crystallography of yeast[21] and bacterial enolases [13] (see Fig. 1). According tothe Prosite database [11], the predicted amino acid se-quence of the A. simplex enolase also contains twoputative N-glycosylation sites, which are not present inthe other enolases studied to date, as well as multiplephosphorylation sites.
Further confirmation of the identity of the clonedcDNA as that encoding the A. simplex enolase wasprovided by demonstration of the enolase activity of therecombinant protein produced by fusion of the AniH6clone full cDNA (1,308 bp) with the C-terminus of GSTin the bacterial expression vector PGEX-4T-3, thenpurified by glutathione-Sepharose-4B chromatography.The enzymatic assay to determine whether the purifiedrecombinant A. simplex enolase was functional demon-strated significant enolase activity, the conversion ofsubstrate to product being linear with regard to bothprotein concentration and time (see Fig. 2b). It has beenreported that this enzyme is only biologically active as adimer and, in some organisms, such subunits frequentlycome from different enolase genes [24]. The fact that therecombinant A. simplex enolase protein showed enzy-matic activity seems to indicate that it forms homodi-mers (e.g. in solution). Recombinant enolases fromother organisms, including S. japonicum and Strepto-coccus pneumoniae, likewise show functional enolaseactivity [7, 39].
Enolases from several microorganisms have beenimplicated in tissue invasion and in allergic disorders inhumans [6, 16, 27, 32]. With regard to tissue invasion, ithas been reported that enolases expressed at the surfaceof some microorganisms (e.g. Streptococcus) can facili-tate host tissue invasion by a mechanism in whichplasminogen is first bound by the enolase and convertedto plasmin, which has serine protease activity [28]. Al-though, as was previously demonstrated, our re-combinant A. simplex enolase is able to bind humanplasminogen and shows functional activity, it seemsimprobable that the parasite uses this enzyme to assisthost invasion, since it is not present in the parasite’s ESantigens. With regard to allergenicity, we have observedthat the A. simplex enolase shows significant structuralhomology with allergenic enolases from molds, such asA. alternata (66.0%) and C. herbarum (65.1%), whichsuggests that this molecule may be involved in allergic
Fig. 7 Antibody responses to antigens captured by the mono-clonal antibody UA3 (UA3R) or against the recombinant A.simplex enolase (enolase-GST) in sera from individual patientswith confirmed anisakiosis. The analysis was done by captureELISA using mAb UA3 or anti-GST polyclonal antibodies,respectively. Striped bars represent anti-UA3R IgG1, white barsanti-UA3R IgE, and black bars anti-enolase-GST IgG1 (in thislatter case the values shown are ODs after substraction of theOD obtained for each serum with recombinant GST). None ofthe sera tested showed positive levels of anti-enolase-GST IgE(not shown)
Fig. 6 Serum IgG1 and IgE responses to recombinant A. simplexenolase (enolase-GST) in the normal population. Each pointrepresents an individual serum. Values shown are ODs aftersubstraction of the OD obtained for each serum withrecombinant GST in an antigen-capture ELISA (see Materialsand methods)
8

reactions induced by A. simplex. However, the results ofour studies of the antigenicity of this molecule in miceand humans were contradictory. Serum from mice in-fected with L3 larvae, or immunized with CE antigensfrom A. simplex L3 larvae, or infected with A. suum orT. canis, reacted with the recombinant A. simplex eno-lase (Fig. 5, lanes 1 and 2). In contrast, sera against ESproducts of A.simplex L3 larvae, or from mice immu-nized with CE from T. muris, T. spiralis and H. adun-cum, showed no reactivity with the enolase. At firstglance, it might appear that the positive reactivity of therecombinant enolase from A. simplex with sera frommice immunized with CE from A. suum and T. canismayreflect the phylogenetic relationship among these threenematodes (all belong to the superfamily Ascaridoidea);however, since sera from mice immunized with the moreclosely related anisakid H. aduncum (likewise of thefamily Anisakidae) tested negative, it seems that phylo-genetic relatedness is not the determinant factor here.
With sera from human anisakiosis patients, we ob-served undetectable or very low levels of IgG1 anti-bodies against recombinant A. simplex enolase, and nodetectable IgE antibodies. Indeed, no significantdifferences were observed in statistical comparisons(Mann–Whitney test) between levels of IgG1 antibodiesto recombinant enolase in healthy individuals andanisakiosis patients (P=0.7165), suggesting that theanti-enolase IgG1 antibodies detected are in fact notinduced by the parasite. Although it is not clear whatsource(s) of immunization might have induced theantibodies reactive with the recombinant enzyme, it canbe speculated that the positive response obtained withnormal sera may be due to sensitization by enolasesfrom microorganisms such as molds and yeast that haveantigenic similarities with A. simplex enolase. Moreintriguing; however, is (a) the absence of anti-enolaseIgG1 antibodies in most of the patients infected withA. simplex, and (b) the complete absence of IgE anti-bodies in sera from these patients. One possible expla-nation is that this molecule shows weak antigenicity inhumans, due to greater structural similarity between thehuman and Anisakis enolases than between human andfungal enolases. However, two facts argue against thispossibility: (a) the A. simplex enolase is differentiallyrecognized in mice and humans, in spite of the fact thatenolases from mice and humans have similar sequencehomologies with the A. simplex enolase (69.8% and68.9%, respectively; see above); and (b) recent data onthe O. volvulus enolase [17], which is more than 80%homologous with the human enolase, indicate that it isrecognized by sera from patients with generalizedonchocercosis, or suffering the hyperreactive form ofthis illness (sowda). Consequently, we hypothesize thatenolase from A. simplex is only recognized by mammals(e.g. mice or humans) after prolonged contact with thehost immune system (i.e. a chronic stimulus). As dem-onstrated in the present study, such contact may occurduring immunization of mice in the presence of Freund’sadjuvant, or when mice are infected with L3 larvae by
the i.p. route. In contrast, natural human infections(acute) are frequently produced by a single larva, andfrequently affect only the mucosa of the digestive tract[18]. The chronic stimulus hypothesis may also explainthe recognition of enolase from O. volvulus by sera ofinfected patients [17], although in this case, the recog-nition of this molecule is probably also favored bypolyclonal activation of the human immune system bythe parasite, as frequently occurs in onchocercosis [12].
In conclusion, the present characterization of the re-combinant A. simplex enolase confirms its enzymaticnature, indicates a high degree of evolutionary conser-vation with respect to other organisms (although epitopedifferences are observed among different nematodegroups), and suggests that it is not relevant in the allergicreactions arising during human Anisakis infections.
Acknowledgments This present work was supported by grantsSAF2002–04057 (Ministerio de Ciencia y Tecnologıa, Spain), 1028/97 (Fondo de Investigacion Sanitaria, Ministerio de Sanidad yConsumo, Spain) and MPY 1337/01 (Instituto de Salud Carlos III,Spain).
References
1. Arrieta I, del Barrio M, Vidarte L, del Pozo V, Pastor C,Gonzalez-Cabrero J, Cardaba B, Rojo M, Mınguez A,Cortegano I, Gallardo S, Aceituno E, Palomino P, Vivanco F.Lahoz C (2000) Molecular cloning and characterization of anIgE-reactive protein from Anisakis simplex: Ani s 1. MolBiochem Parasitol 107:263–268
2. Asturias JA, Eraso E, Martınez A (2000) Cloning and highlevel expression in Escherichia coli of an Anisakis simplextropomyosin isoform. Mol Biochem Parasitol 108:263–267
3. Audıcana MT, Ansotegui IJ, de Corres LF, Kennedy MW(2002) Anisakis simplex: dangerous dead and alive?. TrendsParasitol 18:20–25
4. Audıcana MT, del Pozo MD, Iglesias R, Ubeira FM (2003)Ansiakis simplex and Pseudoterranova decipiens. In: Bier JW,Miliotis MD (eds) International handbook of foodbornepathogens. Marcel Dekker, New York, pp 613–636
5. Babbitt PC, Hasson MS, Wedekin JE, Palmer DRJ, BarrettWC, Reed GH, Rayment I, Ringe D, Kenyon GL, Gerlt JA(1996) The enolase superfamily: a general strategy for enzymecatalyzed abstraction of the a-protons of carboxylic acids.Biochemistry 35:16489–16501
6. Baldo BA, Baker RS (1988) Inhalant allergies to fungi: reac-tions to bakers’ yeast (Saccharomyces cerevisiae) and identifi-cation of bakers’ yeast enolase as an important allergen. IntArch Allergy Appl Immunol 86:201–208
7. Bergmann S, Rohde M, Chhatwal GS, Hammerschmidt S(2001) alpha-Enolase of Streptococcus pneumoniae is a plas-min(ogen)-binding protein displayed on the bacterial cell sur-face. Mol Microbiol 40:1273–1287
8. Conde-Salazar L, Gonzalez MA, Guimaraens D (2002) Type Iand type IV sensitization to Anisakis simplex in 2 patients withhand eczema. Contact Dermatitis 46:361
9. Del Pozo MD, Audicana M, Dıez JM, Munoz D, Ansotegui IJ,Fernandez E, Garcia M, Etxenagusia M, Moneo I, Fernandezde Corres L (1997) Anisakis simplex, a relevant etiologic factorin acute urticaria. Allergy 52:576–579
10. Devereux J, Haeberli P, Smithies O (1984) A comprehensive setof sequence analysis programs for the VAX. Nucleic Acids Res12:387–395
11. Falquet L, Pagni M, Bucher P, Hulo N, Sigrist CJ, HofmannK, Bairoch A (2002) The Prosite database, its status in 2002.Nucleic Acids Res 30:235–238
9

12. Garraud O, Nkenfou C, Bradley JE, Perler FB, Nutman TB(1995) Identification of recombinant filarial proteins capable ofinducing polyclonal and antigen-specific IgE and IgG4 anti-bodies. J Immunol 155:1316–1325
13. Gulick AM, Schmidt DM, Gerlt JA, Rayment I (2001) Evo-lution of enzymatic activities in the enolase superfamily: crystalstructures of the L-Ala-D/L-Glu epimerases from Escherichiacoli and Bacillus subtilis. Biochemistry 40:15716–15724
14. Iglesias R, Leiro J, Ubeira FM, Santamarina MT, SanmartınML (1993) Anisakis simplex: antigen recognition and antibodyproduction in experimentally infected mice. Parasite Immunol15:243–250
15. Iglesias R, Leiro J, Ubeira FM, Santamarina MT, SanmartinML (1995) Anisakis simplex: stage-specific antigens recognizedby mice. J Helminthol 69:319–324
16. Ito K, Ishiguro A, Kanbe T, Tanaka K, Torii S (1995) Char-acterization of IgE-binding epitopes on Candida albicans eno-lase. Clin Exp Allergy 25:529–535
17. Jolodar A, Fischer P, Bergmann S, Buttner DW, Hammers-chmidt S, Brattig NW (2003) Molecular cloning of an alpha-enolase from the human filarial parasite Onchocerca volvulusthat binds human plasminogen. Biochim Biophys Acta1627:111–120
18. Kagei N, Isogaki H (1992) A case of abdominal syndromecaused by the presence of a large number of Anisakis larvae. IntJ Parasitol 22:251–253
19. Kagei N, Orikasa H, Hori E, Sannomiya A, Yasumura Y(1995) A case of hepatic anisakiasis with a literal survey forextra-gastrointestinal anisakiasis. Jpn J Parasitol 44:346–351
20. Kennedy MW, Tierney J, Ye P, McMonagle FA, McIntosh A,McLaughlin D, Smith JW (1988) The secreted and somaticantigens of the third stage larva of Anisakis simplex, andantigenic relationship with Ascaris suum, Ascaris lumbricoides,and Toxocara canis. Mol Biochem Parasitol 31:35–46
21. Lebioda L, Stec B (1991) Mechanism of enolase: the crystalstructure of enolase-Mg2+-2-phosphoglycerate/phosphoenol-pyruvate complex at 2.2 A resolution. Biochemistry 30:2817–2822
22. Lebioda L, Stec B, Brewer JM (1989) The structure of yeastenolase at 2.25-A resolution. An 8-fold beta + alpha-barrelwith a novel beta beta alpha alpha (beta alpha)6 topology.J Biol Chem 264:3685–3693
23. Lorenzo S, Iglesias R, Leiro J, Ubeira FM, Ansotegui I, GarcıaM, Fernandez de Corres L (2000) Usefulness of currentlyavailable methods for the diagnosis of Anisakis simplex allergy.Allergy 55:627–633
24. McAlister L, Holland MJ (1982) Targeted deletion of a yeastenolase structural gene. Identification and isolation of yeastenolase isozymes. J Biol Chem 257:7181–7188
25. Morris SR, Sakanari JA (1994) Characterization of the serineprotease and serine protease inhibitor from the tissue-pene-trating nematode Anisakis simplex. J Biol Chem 269:27650–27656
26. Nielsen H, Brunak S, von Heijne G (1999) Machine learningapproaches to the prediction of signal peptides and otherprotein sorting signals. Protein Eng 12:3–9
27. Pancholi V (2001) Multifunctional alpha-enolase: its role indiseases. Cell Mol Life Sci 58:902–920
28. Pancholi V, Fischetti V (1998) Alpha-enolase, a novel strongplasmin(ogen) binding protein on the surface of pathogenicstreptococci. J Biol Chem 273:14503–14515
29. Perez-Perez J, Fernandez-Caldas E, Maranon F, Sastre J,Bernal ML, Rodrıguez J, Bedate CA (2000) Molecular cloningof paramyosin, a new allergen of Anisakis simplex. Int ArchAllergy Immunol 123:120–129
30. Sakanari JA, Mc Kerrow (1989) Anisakiasis. Clin MicrobiolRev 2:278–284
31. Short JM, Fernandez JM, Sorge JA, Huse WD (1988) LambdaZAP: a bacteriophage lambda expression vector with in vivoexcision properties. Nucleic Acids Res 16:7583–7600
32. Simon-Nobbe B, Probst G, Kajava AV, Oberklofer H, SusaniM, Crameri R, Ferreira F, Ebner C, Breitenbach M (2000) IgE-binding epitopes of enolases, a class of highly conserved fungalallergens. J Allergy Clin Immunol 106:887–895
33. Sugimachi K, Inochuki K, Ooiwa T, Fujino T, Ishii Y (1985)Acute gastric anisakiasis: analysis of 178 cases. J Am MedAssoc 253:1012–1013
34. Tijssen P (1985) Processing of data and reporting of results ofenzyme immunoassays. In: Burdon RH, van Knippenberg PH(eds) Laboratory techniques in biochemistry and molecularbiology: practice and theory of enzyme immunoassays. Else-vier, Amsterdam, 1985:385–421
35. Ubeira FM, Iglesias R (2000) Monoclonal antibodies in thestudy of Anisakis simplex. Allergy Suppl 59:18–27
36. Valinas B, Lorenzo S, Eiras A, Figueiras A, Sanmartin ML,Ubeira FM (2001) Prevalence of and risk factors for IgE sen-sitization to Anisakis simplex in a Spanish population. Allergy56:667–671
37. Van der Straeten D, Rodrigues-Pousada RA, Goodman HM,Van Montagu M (1991) Plant enolase: gene structure, expres-sion, and evolution. Plant Cell 3:719–735
38. Van Thiel PH, Kuipers FC, Roskam TH (1960) A nematodeparasitic to herring, causing acute abdominal syndromes inman. Trop Geogr Med 12:97–113
39. Waine GJ, Becker M, Kalinna B, Yang W, McManus DP(1993) Cloning and functional expression of a Schistosomajaponicum cDNA homologous to the enolase gene family.Biochem Biophys Res Commun 195:1211–1217
10
Copyright © 2022 FDOKUMEN




















