
Multilead ECG Delineation Using Spatially Projected Leads From Wavelet Transform Loops
Upload
independentCategory
view
1download
0
1998 Oxford University Press
Glycobiology vol. 8 no. 7 pp. 695–705, 1998
695
Delineation of the epitope recognized by an antibody specific forN-glycolylneuraminic acid–containing gangliosides
Ernesto Moreno1,2,4, Boel Lanne2, Ana María Vázquez1,Ikuo Kawashima3, Tadashi Tai3, Luis Enrique Fernández1,Karl-Anders Karlsson2, Jonas Ångström2 andRolando Pérez1
1Center of Molecular Immunology, P.O. Box 16040, Havana 11600, Cuba,2Institute of Medical Biochemistry, Göteborg University, Medicinaregatan 9A,S-413 90 Göteborg, Sweden and 3Department of Tumor Immunology, TokyoMetropolitan Institute of Medical Science, Honkomagome, Bunkyo-ku, Tokyo113, Japan
Received on October 31, 1997; revised on February 4, 1998; accepted onFebruary 5, 1998
4To whom correspondence should be addressed
P3 is a mouse monoclonal antibody (mAb) that binds to severalNeuGc-containing gangliosides. It also reacts with antigens ex-pressed in human breast tumors (Vázquez et al. (1995) Hybri-doma, 14, 551–556). In this work, the binding specificity of P3has been characterized in more detail using a panel of glycoli-pids that included several disialylated gangliosides and severalchemical derivatives of NeuGc-GM3. The carboxyl group andthe nitrogen function of sialic acid were found to play import-ant roles in the antibody binding, whereas the glycerol tail ap-pears to be nonrelevant. Molecular modeling was used to ana-lyze the binding data, including the finding that P3 selectivelyrecognizes the internal NeuGc in GD3. For this purpose, con-formational studies of GD3 were performed using moleculardynamics. It was concluded that sialic acid binds the P3 anti-body through its upper face (the one on which the carboxylgroup is exposed) and the C4-C5 side of the sugar ring, where-as none or very little contact between the galactose residue andthe protein is evident. Conformational analysis of GD3 re-vealed that, despite the large flexibility of the NeuGcα8NeuGclinkage, the P3 binding epitope on the external sialic acid is notwell exposed for any of the possible conformations this linkagecan adopt, whereas the internal sialic acid presents the epitopein a proper way for several of these conformations. As a finalresult, a coherent picture of the epitope that fits the wide bind-ing data was obtained.
Key words: epitope/ganglioside/molecular modeling/monoclonal antibody/ NeuGc
Introduction
P3 is a mouse monoclonal antibody (mAb) that binds to severalN-glycolyl-containing gangliosides and also recognizes antigensexpressed in human breast tumors (Vázquez et al., 1995). Only afew monoclonal antibodies (mAbs) specific for NeuGc-containinggangliosides have previously been reported (Nakamura andHanda, 1986; Furukawa et al., 1988; Miyake et al., 1988; Sanai etal., 1988; Watarai et al., 1991; Ozawa et al., 1992; Kawashima etal., 1993). As compared to these antibodies, P3 is unique in
recognizing the N-glycolyl sialic acid on several different types ofsugar chains, as shown in the work presented here, which wasdirected toward characterization of the binding specificity of the P3mAb and delineation of the epitope that it recognizes.
Detailed characterization of the binding specificity of anantibody is important for several applications; for example,antibodies are often used as tools for testing the presence ofmolecules in cells, tissues or chemical preparations. This kind ofinformation becomes especially relevant if the antibody has theability of recognizing tumor cells and, therefore, may be used fordiagnostic and therapeutic purposes.
On the other hand, studying the interactions between anti-bodies or other proteins and the N-acetyl or N-glycolyl types ofsialic acid is also important in order to understand the molecularbasis of the fine distinction that biological systems make betweenthese two species. N-Acetylneuraminic and N-glycolylneurami-nic acids are the most common types of sialic acid found inanimals, occurring in a wide variety of sialoglycoproteins andgangliosides (Corfield and Schauer, 1982). In general, normalhuman tissues yield only NeuAc, while other mammals havesignificant levels of NeuGc. Despite the small structural differ-ence between them (the methyl group in NeuAc being substitutedby a hydroxymethyl group in NeuGc), the two derivatives do notappear to be biologically equivalent.
Knowledge of the way in which a molecule is recognized bya protein at the atomic level is also important for practicalapplications, for example, the design of inhibitors or betterimmunogens, or for theoretical docking studies that may providevaluable information for genetic engineering of a protein bindingsite. X-Ray crystallography of the receptor-ligand complexprovides such information, but it is still far from being a routineprocedure. At present, only one crystal structure of an anti-tumorantibody in complex with its carbohydrate antigen has beenreported: the BR96 antibody in complex with LeY (Jeffrey et al.,1995). A small number of crystal structures of proteins interactingwith N-acetyl sialic acid (alone or as part of a larger molecule) hasbeen reported (Wright, 1990; Sauter et al., 1992; Varghese et al.,1992; Merritt et al., 1994), whereas no complexes containing theN-glycolyl type have been solved to date.
Several reports are found in the literature in which carbohy-drate epitopes have been explored at the atomic level by chemicalmethods. Chemically modified glycolipids have, for example,been used to investigate the epitope recognized by the choleratoxin B-subunit on GM1 (Fishman et al., 1980; Spiegel, 1985;Schengrund and Ringler, 1989; Lanne et al., 1994) and theepitope on NeuGc-GM3 that interacts with the E.coli K99adhesin (Lanne et al., 1995). Molecular modeling applied inconjunction with binding assays may also help to extract moreinformation about binding epitopes (Ångström et al., 1994). Inthis work, a combination of different techniques was used,including binding experiments for a wide panel of NeuGc-con-taining gangliosides and chemical derivatives of sialic acid ofNeuGc-GM3, as well as molecular modeling.
by guest on Decem
ber 17, 2015http://glycob.oxfordjournals.org/
Dow
nloaded from

E.Moreno et al.
696
Table I. Structures and P3 mAb reactivities of glycolipids used in binding experiments
Abbreviation Structurea Binding
TLCb ELISAc
LacCer Galβ4GlcβCer - -
Gangliotri (Gg3) GalNAcβ4Galβ4GlcβCer - -
Gangliotetra (Gg4) Galβ3GalNAcβ4Galβ4GlcβCer - -
Neolactotetra (PG) Galβ4GlcNAcβ3Galβ4GlcβCer - nd
NeuAc-GM4 NeuAcα3GalβCer - -
NeuAc-GM3 NeuAcα3Galβ4GlcβCer - -
NeuAc-GM2 GalNAcβ4(NeuAcα3)Galβ4GlcβCer - -
NeuAc-GM1a Galβ3GalNAcβ4(NeuAcα3)Galβ4GlcβCer - -
NeuAc-GM1b NeuAcα3Galβ3GalNAcβ4Galβ4GlcβCer - -
NeuAc-NeuAc-GD3 NeuAcα8NeuAcα3Galβ4GlcβCer - -
NeuAc-GD2 GalNAcβ4(NeuAcα8NeuAcα3)Galβ4GlcβCer - -
NeuAc-GD1a NeuAcα3Galβ3GalNAcβ4(NeuAcα3)Galβ4GlcβCer - -
NeuAc-GD1b Galβ3GalNAcβ4(NeuAcα8NeuAcα3)Galβ4GlcβCer - -
NeuAc-GT1b NeuAcα3Galβ3GalNAcβ4(NeuAcα8NeuAcα3)Galβ4GlcβCer - -
NeuAc-SPG NeuAcα3Galβ4GlcNAcβ3Galβ4GlcβCer - -
NeuGc-GM3 NeuGcα3Galβ4GlcβCer + ++
NeuGc-GM2 GalNAcβ4(NeuGcα3)Galβ4GlcβCer + ++
NeuGc-GM1a Galβ3GalNAcβ4(NeuGcα3)Galβ4GlcβCer - ±
NeuGc-GM1b NeuGcα3Galβ3GalNAcβ4Galβ4GlcβCer + +
NeuGc-NeuGc-GD3 NeuGcα8NeuGcα3Galβ4GlcβCer + ++
NeuAc-NeuGc-GD3 NeuAcα8NeuGcα3Galβ4GlcβCer + ++
NeuGc-NeuAc-GD3 NeuGcα8NeuAcα3Galβ4GlcβCer - ±
NeuGc-GD1a NeuGcα3Galβ3GalNAcβ4(NeuGcα3)Galβ4GlcβCer + +
NeuGc-GD1c NeuGcα8NeuGcα3Galβ3GalNAcβ4Galβ4GlcβCer - ±
NeuGc-SPGd NeuGcα3Galβ4GlcNAcβ3Galβ4GlcβCer + nd
VI3NeuGcα-nLc6d NeuGcα3Galβ4GlcNAcβ3Galβ4GlcNAcβ3Galβ4GlcβCer + nd
aThe condensed sugar chain representation follows recent recommendations (Sharon, 1987).bThe reactivities for the TLC experiments are given using a qualitative scale (positive or negative).cBinding strengths according to results from ELISA (see Figure 4). (++), (+), and (±) denote strong, moderate, and weak binding, respectively. nd, Notdetermined.dNot shown in the figures.
Results and discussion
Binding of P3 mAb to various glycolipids by HPTLCimmunostaining and ELISA
The binding of P3 mAb to different glycolipids (see Table I) wasdetermined by HPTLC (Figures 1–3) and ELISA (Figure 4).Results are summarized in Table I. The mAb bound strongly toNeuGc-GM3 and NeuGc-GM2, as previously reported (Vázquezet al., 1995). A moderate binding was observed for NeuGc-GM1b, whereas NeuGc-GM1a, which did not bind on the TLCplate, showed a weak reactivity by ELISA (Figures 1 and 4). Thecorresponding NeuAc-containing monosialogangliosides did notreveal any antibody binding.
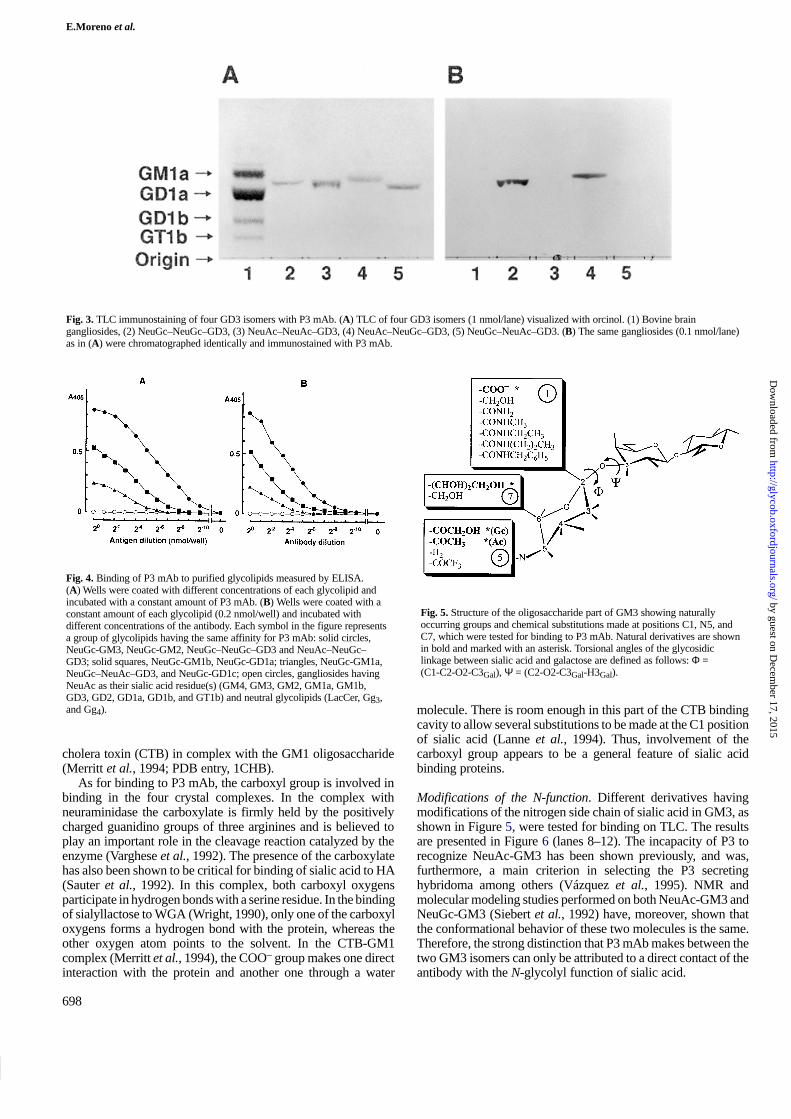
Among the NeuGc-containing disialogangliosides that werestudied, P3 bound moderately to NeuGc-GD1a, whereas a weakbinding was observed to NeuGc-GD1c only by ELISA (Figures2 and 4). Four GD3 isomers: NeuGc–NeuGc–GD3, NeuAc–NeuAc–GD3, NeuAc–NeuGc–GD3, and NeuGc–NeuAc–GD3
were also tested (Figures 3 and 4). NeuGc–NeuGc–GD3 andNeuAc–NeuGc–GD3 were strongly bound by P3 mAb, whereasthe isomer having NeuGc only in the external position showed aweak binding by ELISA and NeuAc–NeuAc–GD3 was notrecognized at all. These results indicate that the inner NeuGcresidue in GD3 is essential for the binding of P3 mAb. Areasonable explanation for this fact was obtained throughmolecular modeling analysis (see below).
P3 also bound strongly to NeuGc-SPG and VI3NeuGcα-nLc6(data not shown). None of the other tested molecules having onlythe N-acetyl type of sialic acid (GM4, GD2, GD1a, GD1b, GT1b,NeuAc-SPG) or neutral glycolipids (LacCer, Gg3, Gg4, and PG)were recognized (data not shown).
Binding of P3 mAb to chemical derivatives of NeuGc-GM3
Figure 5 shows the positions on sialic acid where chemicalsubstitutions were made, as well as the groups that were attached
by guest on Decem
ber 17, 2015http://glycob.oxfordjournals.org/
Dow
nloaded from

Binding specificity of mouse mAb P3
697
Fig. 1. TLC immunostaining of monosialogangliosides with P3 mAb. (A) Ganglioside fraction from bovine brain (10 nmol/lane) and standardmonosialogangliosides (1 nmol/lane) were chromatographed with chloroform/methanol/0.22% CaCl2 in water (55:45:10, v/v/v) and visualized with orcinol. (1)Bovine brain gangliosides, (2) NeuGc-GM3, (3) NeuAc-GM3, (4) NeuGc-GM2, (5) NeuAc-GM2, (6) NeuGc-GM1a, (7) NeuAc-GM1a, (8) NeuGc-GM1b, (9)NeuAc-GM1b. (B) The same gangliosides (0.1 nmol/lane) as in (A) were chromatographed under the same conditions as above and immunostained with P3mAb.
Fig. 2. TLC immunostaining of disialogangliosides with P3 mAb. (A) The ganglioside fraction from bovine brain (10 nmol/lane) and purified disialogangliosideswere chromatographed with chloroform/methanol/0.22% CaCl2 in water (55:45:10, v/v/v) and visualized with orcinol stain. (1) Bovine brain gangliosides, (2)NeuGc–NeuGc–GD3, (3) NeuAc–NeuAc–GD3, (4) NeuGc-GD1a, (5) NeuAc-GD1a. (B) The same gangliosides (0.1 nmol/lane) as in (A) were chromatographedas above and immunostained with P3 mAb.
at each position. The binding responses of P3 mAb to thesederivatives are shown in Figure 6.
Modifications of the carboxyl group. The carboxyl group ofNeuGc-GM3 was modified to alcohol and several amides. Noneof these derivatives was recognized by the antibody, as shown inFigure 6 (lanes 1–6). It should be pointed out, however, that thenegative binding results obtained in these experiments, wherebulky groups had been added to the C1 carbon (such as propyl-and benzylamide), do not necessarily indicate a direct participa-tion of the carboxyl group in the binding interactions, since suchnegative responses might be due to steric interferences caused bythese substituents. The nonbinding of the alcohol and amidederivatives, however, points more strongly to a direct contact ofthe COO– group of sialic acid with the antibody. Nonetheless, thepossibility that binding might be affected as a result ofconformational changes involving the glycosidic linkage close tothe substituted group was also investigated.
Molecular dynamics simulations were thus carried out for theterminal disaccharide of the NeuGc-GM3 alcohol and amidederivatives, as well as for the native molecule, serving as control.
Figure 7 shows the Φ and Ψ trajectories obtained from these runs.For the three molecules, the values of these torsional anglescorresponded to the anticlinal (Φ ≈ -155�, Ψ ≈ -30�) and thesynclinal (Φ ≈ -70�, Ψ ≈ 10�) conformations, as was similarlyobtained by Siebert et al. (1992) for NeuAc- and NeuGc-GM3.The behavior of the glycerol tail of sialic acid was the same forthe three molecules (data not shown), and corresponded also tothat obtained by Siebert et al. (1992). Thus, no conformationalchanges that might account for the nonbinding of the antibody tothese derivatives were found.
It was of interest to compare these results with binding patternsshown by sialic acid with other proteins, especially in those caseswhere crystal data of the complex are available. No complexeswith NeuGc are found in the Protein Data Bank, but four differenttypes of protein have been reported with bound N-Acetyl sialicacid, either alone or as part of a larger ligand: wheat germagglutinin (WGA) in complex with sialyllactose (Wright, 1990;PDB entry, 2WGC), influenza virus hemagglutinin (HA) incomplex with sialyllactose (Sauter et al., 1992; PDB entry,1HGG), influenza virus neuraminidase in complex with NeuAc(Varghese et al., 1992; PDB entry, 2BAT) and the B subunits of
by guest on Decem
ber 17, 2015http://glycob.oxfordjournals.org/
Dow
nloaded from
E.Moreno et al.
698
Fig. 3. TLC immunostaining of four GD3 isomers with P3 mAb. (A) TLC of four GD3 isomers (1 nmol/lane) visualized with orcinol. (1) Bovine braingangliosides, (2) NeuGc–NeuGc–GD3, (3) NeuAc–NeuAc–GD3, (4) NeuAc–NeuGc–GD3, (5) NeuGc–NeuAc–GD3. (B) The same gangliosides (0.1 nmol/lane)as in (A) were chromatographed identically and immunostained with P3 mAb.
Fig. 4. Binding of P3 mAb to purified glycolipids measured by ELISA.(A) Wells were coated with different concentrations of each glycolipid andincubated with a constant amount of P3 mAb. (B) Wells were coated with aconstant amount of each glycolipid (0.2 nmol/well) and incubated withdifferent concentrations of the antibody. Each symbol in the figure representsa group of glycolipids having the same affinity for P3 mAb: solid circles,NeuGc-GM3, NeuGc-GM2, NeuGc–NeuGc–GD3 and NeuAc–NeuGc–GD3; solid squares, NeuGc-GM1b, NeuGc-GD1a; triangles, NeuGc-GM1a,NeuGc–NeuAc–GD3, and NeuGc-GD1c; open circles, gangliosides havingNeuAc as their sialic acid residue(s) (GM4, GM3, GM2, GM1a, GM1b,GD3, GD2, GD1a, GD1b, and GT1b) and neutral glycolipids (LacCer, Gg3,and Gg4).
cholera toxin (CTB) in complex with the GM1 oligosaccharide(Merritt et al., 1994; PDB entry, 1CHB).
As for binding to P3 mAb, the carboxyl group is involved inbinding in the four crystal complexes. In the complex withneuraminidase the carboxylate is firmly held by the positivelycharged guanidino groups of three arginines and is believed toplay an important role in the cleavage reaction catalyzed by theenzyme (Varghese et al., 1992). The presence of the carboxylatehas also been shown to be critical for binding of sialic acid to HA(Sauter et al., 1992). In this complex, both carboxyl oxygensparticipate in hydrogen bonds with a serine residue. In the bindingof sialyllactose to WGA (Wright, 1990), only one of the carboxyloxygens forms a hydrogen bond with the protein, whereas theother oxygen atom points to the solvent. In the CTB-GM1complex (Merritt et al., 1994), the COO– group makes one directinteraction with the protein and another one through a water
Fig. 5. Structure of the oligosaccharide part of GM3 showing naturallyoccurring groups and chemical substitutions made at positions C1, N5, andC7, which were tested for binding to P3 mAb. Natural derivatives are shownin bold and marked with an asterisk. Torsional angles of the glycosidiclinkage between sialic acid and galactose are defined as follows: Φ =(C1-C2-O2-C3Gal), Ψ = (C2-O2-C3Gal-H3Gal).
molecule. There is room enough in this part of the CTB bindingcavity to allow several substitutions to be made at the C1 positionof sialic acid (Lanne et al., 1994). Thus, involvement of thecarboxyl group appears to be a general feature of sialic acidbinding proteins.
Modifications of the N-function. Different derivatives havingmodifications of the nitrogen side chain of sialic acid in GM3, asshown in Figure 5, were tested for binding on TLC. The resultsare presented in Figure 6 (lanes 8–12). The incapacity of P3 torecognize NeuAc-GM3 has been shown previously, and was,furthermore, a main criterion in selecting the P3 secretinghybridoma among others (Vázquez et al., 1995). NMR andmolecular modeling studies performed on both NeuAc-GM3 andNeuGc-GM3 (Siebert et al., 1992) have, moreover, shown thatthe conformational behavior of these two molecules is the same.Therefore, the strong distinction that P3 mAb makes between thetwo GM3 isomers can only be attributed to a direct contact of theantibody with the N-glycolyl function of sialic acid.
by guest on Decem
ber 17, 2015http://glycob.oxfordjournals.org/
Dow
nloaded from

Binding specificity of mouse mAb P3
699
Fig. 6. P3 mAb binding to derivatives and isomers of NeuGc-GM3.(A) Chemical staining of glycolipids (5 µg/lane). (B) Binding of the samecompounds to P3 mAb. (1) NeuGc-GM3 alcohol, (2) NeuGc-GM3 amide,(3) NeuGc-GM3 methylamide, (4) NeuGc-GM3 ethylamide, (5)NeuGc-GM3 propylamide, (6) NeuGc-GM3 benzylamide, (7) NeuGc-GM3,(8) NeuAc-GM3, (9) NeuGc-GM3 (same as (7)), (10) de-N-acetyl-GM3(NeuNH2-GM3), (11) TFA-GM3, (12) TFI-GM3, (13) periodate-oxidizedNeuGc-GM3.
Trifluoroacetylation (Karlsson et al., 1991) of NeuAc-GM3,causing the acetamido moiety of the terminal NeuAc to bereplaced by a trifluoroacetamido group, results in no or very weakbinding to the antibody. Thus, the lack of binding indicates thatthe -CF3 group, despite its hydrophilic properties, cannot beaccommodated within the same binding cavity as the distinctive-CH2OH group of the N-glycolyl form of sialic acid. Further-more, P3 showed a weak binding to de-N-acetyl-GM3(NeuNH2-GM3) on the TLC plate (Figure 6, lane 10), indicatingnot only that a very high specificity for the N-glycolyl groupexists, but also that the interactions with the terminal atoms of thisfunction are important for binding. It should be also taken intoaccount that the amino group in NeuNH2-GM3 might beprotonated, which would change the electrostatic character of thisfunction.
Reported data show that this group also plays an important rolein the binding of sialic acid to other proteins. In the interactionbetween sialic acid and neuraminidase, the CO and NH of theN-acetyl moiety participate in hydrogen bonds with the protein,whereas the CH3 is placed in a hydrophobic pocket. In the wheatgerm agglutinin–sialyllactose complex, the N-acetyl groupmakes the largest number of contacts, being essential for thespecificity of this protein for GlcNAc and NeuAc (Wright, 1990).Both the NH and the CO participate in hydrogen bonds, whereasthe methyl group is in close contact with a tyrosine phenyl ringand the Cβ carbon of a glutamate in the least exposed part of thebinding cavity. In the complex of influenza virus hemagglutininwith sialyllactose, the acetamido nitrogen donates a hydrogenbond to a main chain carbonyl, while the methyl group is inhydrophobic contact with the indole ring of a tryptophan residue.
Fig. 7. Plots of the Φ (+) and Ψ (solid circles) trajectories obtained fromdynamics runs for NeuGc-GM3 alcohol (A), NeuGc-GM3 amide (B), andNeuGc-GM3 (C). See Figure 5 for definitions of angles.
Addition of a hydroxyl group to obtain N-glycolyl neuraminicacid abolishes the binding of the X-31 virus (Higa et al., 1985).However, extension of this side chain by an extra methyl group(N-propionyl analog) does not affect the binding (Sauter et al.,1992). In the CTB–GM1 complex, there is also a hydrogen bonddonated by the NH and hydrophobic contacts of the acetyl groupwith a tyrosine phenyl ring (Merritt et al., 1994). In this case,however, the methyl group is also exposed to the solvent, whichmay explain why NeuGc-GM1 binds to CTB with approximatelythe same strength as the N-acetyl variant (Ångström et al., 1994).
These data indicate that the specificity for the N-acetyl isomerof sialic acid is very dependent on the hydrophobicity around themethyl group. Thus, the marked sensitivity shown by P3 towardschanges of the N-glycolyl function strongly suggests that thisgroup binds into a tight pocket that provides the necessaryelectrostatic interactions in a very specific manner.
Cleavage of the glycerol tail. The reactivity of P3 mAb was notaffected by periodate oxidation of NeuGc-GM3, as shown inFigure 6 (lane 13), suggesting that the C8 and C9 alcohol groupsdo not participate in the binding. This finding agrees with theresults obtained for the GD3 isomers (Figure 4), especially forNeuAc-NeuGc-GD3, in which the internal N-glycolyl sialic acidis recognized by the antibody despite the linkage of the externalNeuAc to the C8 carbon of NeuGc.
by guest on Decem
ber 17, 2015http://glycob.oxfordjournals.org/
Dow
nloaded from

E.Moreno et al.
700
Fig. 8. Models of three NeuGc gangliosides. (A) The anticlinal conformationof GM3 is represented in bold dark lines (with oxygen atoms marked ingray), whereas the synclinal orientation of sialic acid is shown in thin lines.The GM2 structure is obtained by including a GalNAc residue, which isdrawn in bold gray lines. (B) GM1 oligosaccharide, represented in the sameway as GM3 having the anticlinal conformation. A second possibleorientation for the terminal Gal is shown in thin lines. In both panels, theglycolyl oxygen is shown as a small sphere.
The role of this group in binding varies for different proteins.In both the influenza virus neuraminidase and hemagglutinin, the8- and 9-OH of the glycerol side chain participate in hydrogenbonds with protein side chains. It has also been shown thatmodification of this tail reduces the binding capabilities of thevirus (Sauter et al., 1992). In contrast to these two complexes, theglycerol tail in the CTB-GM1 complex is highly exposed to thesolvent and the few interactions it has with the toxin subunit arewater-mediated (Merritt et al., 1994). In the WGA–sialyllactosecomplex, this side chain makes hydrophobic contacts with atyrosine ring, but it is also exposed to the solvent. Binding studiesperformed by Peters et al. (1979) demonstrated that trimmingback the glycerol tail to the C7 carbon improves the binding ofsialic acid to WGA.
Molecular modeling analysis of binding data for naturalglycolipids
Molecular modeling analysis was carried out in order to correlatestructural properties of the glycolipids with binding data, with theaim of extracting additional information about the bindingepitope.
The equally high affinity of P3 mAb for NeuGc-GM3 andNeuGc-GM2 suggests that the GalNAc residue of GM2 does notparticipate in the binding. It should be noted that the conforma-tional behavior of the sialic acid residue differs in these twoglycolipids. The terminal, nonbranched NeuAcα3Galβ disac-charide present in GM3 has been found to be flexible (Poppe etal., 1989; Sabesan et al., 1991; Siebert et al., 1992), existingmainly in two conformations (anticlinal and synclinal), whereasfor the branched NeuAcα3Galβ disaccharide in GM2 (Levery,1991), GM1a (Acquotti et al., 1990), and GD1a (Sabesan et al.,1991) NMR data point to the presence of only the anticlinalorientation. For this conformation, the approach of the antibodyfrom the side of the glycosidic and ring oxygens of sialic acid isnot possible because of the presence of the GalNAc residue (seeFigure 8a).
On the other hand, the terminal galactose of NeuGc-GM1a,which in the preferred conformation (Figure 8b) is highlyexposed at the top of the molecule, denies the antibody access to
the binding epitope from this direction. Yet, a weak binding to thisganglioside was observed in the ELISA experiments, which mustbe due to the existence of a second possible (although much lesspopulated) orientation for the terminal galactose (Imberty et al.,1995), where steric interference is much less pronounced, asshown in Figure 8b.
Therefore, the antibody approaches the gangliosides “from thetop� and recognizes an epitope on sialic acid that involves thecarboxyl group, the N-glycolyl function (especially the terminalatoms, as can be concluded from the weak binding of thede-N-acylated derivative), and most likely also the 4-OH.
Since the internal sialic acid of NeuGc-GM1a produces onlya very weak binding, whereas NeuGc-GM1b shows a moderatereactivity, the likewise moderate binding obtained also forNeuGc-GD1a must be accounted for by the external sialic acid.The weaker binding observed for these two molecules, ascompared with NeuGc-GM3 and NeuGc-GM2, may result froma less favorable orientation of the external NeuGc. In bothNeuGc-GM1b and NeuGc-GD1a this residue is presented mostlyin an orientation (glycosidic angles were taken from Sabesan etal., 1991) where the NeuGc ring is perpendicular to the main bodyof the molecule, with the 4-OH side pointing downwards and thusbeing inaccessible to the antibody. This suggests a possibleengagement of the 4-OH of NeuGc in the interactions with theprotein, as occurs in the complexes between NeuAc or GlcNAcwith the wheat germ agglutinin (Wright, 1990), where the 4-OH,together with the N-acetyl moiety, plays a key role in determiningthe binding specificity. However, this group is not important forthe binding in the other known sialic acid–protein complexes. Itshould be also taken into account that in NeuGc-GM1b andNeuGc-GD1a the external NeuGcα3Galβ disaccharide is3-linked to the third residue (GalNAc), in contrast to NeuGc-GM3 and NeuGc-GM2, where this disaccharide is 4-linked to thecorresponding residue (Glc). The different types of linkage inNeuGc-GM1b and NeuGc-GD1a produce different orientationsof the external sialic acid relative to their respective internalresidues, which might affect the binding of the antibody to thesemolecules.
The weak reactivity of P3 with NeuGc-GD1c can beunderstood on the basis of the considerations made above forNeuGc-GM1b and NeuGc-GD1a (unfavorable orientation of theinternal sialic acid in NeuGc-GD1c), and the results of theconformational analysis carried out for NeuGc–NeuGc–GD3being presented below, which account for the non-binding of theexternal NeuGc being.
The obtained definition of the epitope is illustrated in Figure9 for the GM2 ganglioside.
Conformational analysis of GD3
The selective recognition of the internal N-glycolyl sialic acid inGD3 by P3 mAb was quite an intriguing finding. In order to finda rationale behind this experimental fact and, at the same time, testour conclusions about the binding epitope, a conformationalstudy of the oligosaccharide part of NeuGc–NeuGc–GD3 wascarried out using molecular dynamics, as described in Materialsand methods. Although the minimum energy conformers used asstarting geometries were obtained for the NeuAcα8NeuAcdisaccharide (Brisson et al., 1992), these results, as well as thosefrom our simulations (see below), are not likely to be dependenton the type of N-function since in all the conformers the N-acetylor N-glycolyl groups point away from the critical linkage region.Thus, it is reasonable to assume that the conformational behaviorof the four GD3 isomers should be the same.
by guest on Decem
ber 17, 2015http://glycob.oxfordjournals.org/
Dow
nloaded from

Binding specificity of mouse mAb P3
701
Fig. 10. Structure of NeuGcα8NeuGcα3Galβ (terminal part of NeuGc–NeuGc–GM3) showing the six torsional angles that determine the conformation of thistrisaccharide. In the following definitions, atoms belonging to the external sialic acid are in bold. Φext = (C1-C2-O2-C8), Ψext = (C2-O2-C8-C7), ω8 =(O2-C8-C7-O7), ω7 = (O7-C7-C6-O6), Φint = (C1-C2-O2-C3Gal), Ψint = (C2-O2-C3Gal-H3Gal).
Fig. 9. Model of the terminal trisaccharide of GM2 showing the epitoperecognized by the P3 mAb. The dots represent the solvent surface aroundepitope atoms, as determined from binding data and modeling analysis. Theglycolyl oxygen is shown as a small sphere.
Fourteen 500 ps trajectories were collected (see Materials andmethods). For each run, the values displayed by the torsionalangles of interest (Φint , Ψint , Φext , Ψext , ω7, and ω8; see Figure10), as well as potential energy values, were extracted from thetrajectory files and plotted versus time. The obtained plots wereused to identify the conformational regions that were sampled bythe molecule. Thereafter, average torsional angles and averagepotential energies were calculated for each of these regions (TableII ).
Comparison with results obtained by Brisson et al. (1992)reveals both similarities and differences. Most of the energyminima obtained by Brisson et al. were sampled in our dynamicsruns for both the synclinal and anticlinal orientations of theinternal sialic acid of GD3. The average values obtained by us forthe torsional angles show only small differences relative to thoseobtained by Brisson et al., but the relative potential energies donot correlate well with the values obtained by these authors. Thismay result from choosing average energies to characterize thepotential energy wells instead of using the energies of local
minima, and it may be also a consequence of applying a differentforce field. In general, the energy functions used in calculationsare imprecise (Halgren, 1995), and although the positions of thelocal minima are only slightly altered by modification of theenergy function, the relative depths of the minima may vary.
In Table II , conformers are listed in order of increasing energy,making it evident that the synclinal orientation of the NeuGc-α3Gal linkage was preferred over the anticlinal orientation.However, as pointed out above, these results should be analyzedwith some caution. It should be also taken into account that thebinding experiments were carried out on a solid phase, whereother interactions may favor or disfavor the occurrence of a givengeometry. Therefore, our analysis included all obtained con-formers.
Table II. Conformational regions sampled by GD3 in molecular dynamicssimulations
Conform. PE Total time Φext Ψext w7 ω8 Φint Ψint
ST1 0.0 410 –100 151 62 170 –82 10
ST2 1.1 750 –58 162 65 –177 –73 4
ST3 2.4 140 77 154 62 –178 –69 8
SG2+ 2.5 110 –69 112 63 87 –73 0
SG1+ 2.8 380 –152 94 70 99 –75 1
AT1 4.0 950 –89 150 60 172 –148 –31
SG1– 4.8 270 –158 107 73 –53 –91 –42
AG3– 4.9 650 60 137 40 –74 –160 –25
AG2+ 6.3 240 –70 115 53 69 –155 –27
AG2– 7.4 280 –74 150 49 –65 –159 –18
SG3+ 9.4 110 59 118 57 72 –78 3
AG3+ 10.7 80 64 112 55 76 –154 –26
Conformers are ordered according to the relative values of the averagepotential energies (PE). The standard deviations for the potential energyvalues oscillated between seven and eight kcal/mol for all regions (data notshown). The nomenclature used to name these conformations follows that ofBrisson et al. (1992) for the NeuGcα8NeuGc linkage, with the addition of an“A� or “S� as a first letter to indicate the anticlinal or synclinal conformationof the NeuGcα3Gal linkage. See Figure 10 for definitions of the torsionalangles.
by guest on Decem
ber 17, 2015http://glycob.oxfordjournals.org/
Dow
nloaded from

E.Moreno et al.
702
Fig. 11. Models of two GD3 isomers (gray bold lines) in different conformations superimposed on GM2 (dark thin lines). Glycolyl oxygens of both molecules arerepresented as small spheres. (A) The external NeuGc of NeuGc–NeuAc–GD3 is superimposed on the sialic acid of GM2. Carboxyl oxygens of the two sialicacid residues of GD3 are represented as small dark spheres. In this conformation (ST1), the COO– group of the internal NeuAc stands over the COO– of theexternal NeuGc. (B) The internal NeuGc of NeuAc–NeuGc–GD3 (in the SG1– conformation) is superimposed on the sialic acid of GM2, resulting in the NeuAcresidue lying below the glycerol tail of the GM2 sialic acid, which allows antibody access to the internal NeuGc.
Nonbinding of the external N-glycolyl sialic acid. In order tounderstand why the NeuGc–NeuAc–GD3 isomer does not bind,its external (N-glycolyl) sialic acid was superimposed onto thesialic acid of NeuGc-GM2 and NeuGc-GM1a, for each of theobtained conformers, as illustrated in Figure 11a for the ST1geometry.
In conformers ST1 and AT1, the carboxyl group of the externalsialic acid is masked by the COO- of the internal one, as shownin Figure 11a. A similar situation occurs for conformers SG2+and AG2+, where the N-acetyl group of the internal unit is placedon top of the COO– of NeuGc. Conformations ST2 and SG1– areevidently also nonbinding since in both geometries the carboxylgroup of the external sialic acid points towards the galactose unit,thus being inaccessible to the antibody. The same holds true forthe AG1– conformer, which was unstable during the dynamicsrun.
Conformer AG2– presents the carboxyl group of the externalsialic acid pointing down towards the ceramide. A similarsituation occurs for conformers SG1+ and AG1+ (not stable in thedynamics run), in which the 4-OH side of NeuGc pointsdownwards.
An additional conformational region, ST3, was obtained in thedynamics run that started from the SG3+ geometry. No analogousconformation was reported by Brisson et al. (1992), although avery small energy well at the 10 kcal/mol level containing anapproximate “T3� conformer can be seen in a contour mappresented by these authors. The ST3 conformation is nonbinding,since the carboxyl group of NeuGc points down, towards theceramide moiety, with the 4-OH facing the galactose ring. TheAT2 conformation was found to be unstable during the simula-tion, and only very short excursions from AT1 to this region wereobserved.
In conformers SG3+ and AG3+ the methyl group of theN-acetyl function approaches the 4-OH of the external NeuGc(distance between the methyl carbon of NeuAc and the O4 of
NeuGc = 4.6 Å), which might cause some steric impediments forbinding. Furthermore, the potential energies obtained for thesetwo conformations are high, which correlates well with the NMRdata of Brisson et al. (1992), according to which the populationof the G3+ well must be low in order to reproduce the observedNOEs.
Binding to the internal N-glycolyl sialic acid. The possiblebinding conformations of the two GD3 isomers having NeuGc asthe internal sialic acid were also analyzed by superimposing thissugar onto the sialic acid of GM2 and GM1.
Two of the conformers, ST3 and SG1+, cannot bind becausepart of the external residue covers the ring on the carboxyl sideof the internal NeuGc. In a few other conformations, the externalsialic acid is situated at the top of the molecule, but in a positionsdifferent from the one occupied by the terminal galactose inGM1a, making it difficult to determine whether they can bind theantibody or not. However, in at least six of the obtainedgeometries (SG2+, AG2+, SG1–, AG1–, AG2–, and AG3–) theepitope is presented in a favorable orientation and without sterichindrance from the external sialic acid, as illustrated in Figure 11bfor the SG1– conformer.
Concluding remarks
The combination of binding experiments for a wide panel ofdifferent structures with molecular modeling allowed us todelineate the epitope that is recognized by the P3 antibody. Acoherent picture, fitting the wide range of binding data available,was obtained. For the sialic acid group, the carboxyl and theN-glycolyl functions were shown to be essential for binding, mostlikely by interacting directly with the protein, as in the reportedcrystal complexes containing bound sialic acid. Also, the bindingepitope seen by the P3 antibody is consistent with an approachtowards the antigen “from the top.� This is a prerequisite since inthe immunization procedure followed in the antibody generation
by guest on Decem
ber 17, 2015http://glycob.oxfordjournals.org/
Dow
nloaded from

Binding specificity of mouse mAb P3
703
the NeuGc-GM3 molecules were inserted into liposomes,limiting the number of allowed conformations due to restrictionsimposed by the membrane plane.
It is difficult to draw any conclusions about a possibleparticipation of the Gal residue in the binding to P3 mAb, andwhether the anticlinal orientation of sialic acid is needed forbinding or not. Only two of the available crystal complexes, theWGA– and HA–sialyllactose complexes, provide such compara-tive information. In both complexes, the linkage between sialicacid and galactose exhibits a synclinal geometry. In the case ofWGA, modeling analysis shows that this is the only possibleconformation, since for the anticlinal geometry both the galactoseand glucose residue would bump into the protein. For theinfluenza virus hemagglutinin, however, modeling shows that theanticlinal conformation also is possible and, moreover, allows arelative orientation between the sugar moiety and the protein thatseems more appropriate for a multivalent binding of thehemagglutinin to a cell surface. In this conformation, there can beonly little contact between HA and the galactose, as in theWGA–sialyllactose crystal complex.
The sequence of the variable region of P3 mAb has beenrecently obtained and several models of this region have beenconstructed. Docking studies in conjunction with site directedmutagenesis experiments are currently in progress, aimed atconstructing a model of the P3–NeuGc complex. The delineationof the epitope recognized by P3 on sialic acid is of greatimportance for this purpose since the number of variables in thedocking procedure may be considerably reduced. Furthermore,the extensive and varied binding data that has been obtained mayprovide a valuable test of the consistency of the model.
Materials and methods
Monoclonal antibody
P3 mAb (an IgM antibody) was produced by immunization ofBalb/c mice with liposomes containing NeuGc-GM3 and tetanustoxoid, as previously described (Vázquez et al., 1995).
Preparation of standard glycolipids
The structures of the glycolipids used in this study are shown inTable I. Gangliosides are named according to the nomenclatureof Svennerholm (1964); otherwise, the nomenclature usedfollows IUPAC-IUB recommendations (IUPAC-IUB, 1977;Sharon, 1987).
Neutral glycolipids. LacCer and Gg4 were obtained from GM3and NeuAc-GM1, respectively, by treatment with sialidase (Taiet al., 1983).
N-Acetylated glycolipids. GM1a, GD1a, GD1b, and GT1b wereprepared from bovine brain (Hakomori, 1983). GM2 and GM4were obtained from human brain (Svennerholm, 1976). GD2 wasobtained by transformation of GD1b with bovine testis β-galac-tosidase (Boehringer-Mannheim) (Cahan et al., 1982). GM3 wasisolated from sheep spleen (Svennerholm, 1976). GM1b waskindly donated by Y. Sanai (Tokyo Metropolitan Institute ofMedical Science). NeuAc-SPG was obtained from humanerythrocytes (Hakomori, 1983).
N-Glycolylated glycolipids and GD3 isomers. NeuGc-GM3 wasisolated from sheep spleen (Svennerholm, 1976) while NeuGc-
GM2 was prepared from mouse (Balb/c) liver (Hashimoto et al.,1983). NeuGc-GM1a and NeuGc-GD1a were from mouse (ICR)liver. NeuGc-GM1b and NeuGc-GD1c, from mouse spleen andthymoma, respectively, were donated by K. Nakamura (TokyoMetropolitan Institute of Medical Science). NeuGc-SPG andVI3NeuGcα-nLc6 were obtained from bovine erythrocytes(Chien et al., 1978). The four GD3 isomers, NeuAc–NeuAc–GD3, NeuGc–NeuGc–GD3, NeuAc–NeuGc–GD3 and NeuGc–NeuAc–GD3, isolated from bear erythrocytes, were kindlysupplied by Y. Hashimoto and S. Suziki (Tokyo MetropolitanInstitute of Medical Science).
Ganglioside derivatives
Derivatization of NeuGc-GM3 to amides and the primary alcoholwas achieved as described previously (Nakamura and Handa,1986; Lanne et al., 1995). Methylamine, ethylamine, propyla-mine, benzylamine, and methyl iodide were purchased fromAldrich, Steinheim, Germany. Trifluoro derivatives of the sialicacid and the ceramide of NeuGc-GM3 were produced asdescribed before (Karlsson et al., 1991) yielding one derivativehaving the ceramide modified by a trifluoroacetimide (TFI)moiety and a second derivative in which the fatty acid is replacedby a trifluoroacetamido (TFA) moiety. In both cases theacetamido function of the sialic acid is exchanged for atrifluoroacetamido group. Periodate oxidation of the glycerol tailof NeuGc-GM3 was performed by treating the ganglioside withsodium metaperiodate, followed by borohydride reduction (Vehet al., 1976). The derivatives were purified by ion-exchangechromatography (DEAE-Sepharose CL-6B, Pharmacia,Sweden) or HPLC (silica column), and the products wereidentified by negative ion fast atom bombardment mass spectro-metry (JEOL SX102A, JEOL, Japan) with the following settings:accelerating voltage, 10 kV; resolution, 1000; m/∆m, 10% valley.Xe atom bombardment at 6 keV was employed using triethanola-mine as matrix.
Thin-layer chromatography (TLC)
Pre-coated HPTLC aluminum sheets and glass plates (Merck,Darmstadt, Germany) were used for the separation of glycolipids.The solvent systems used for developing chromatography werechloroform/methanol/0.22% CaCl2 in water (55:45:10, v/v/v) orchloroform/methanol/water (68:35:8, v/v/v). Glycolipids werevisualized with orcinol and resorcinol reagents.
Immunostaining on HPTLC plates
Enzyme immunostaining on HPTLC plates was performed aspreviously reported (Kawashima et al., 1993). Briefly, afterchromatography of the glycolipids, the plates were soaked with0.1% polyisobutylmethacrylate in cyclohexane for 75 s. Afterdrying, the plates were incubated with phosphate-buffered saline(PBS) containing 1% bovine serum albumin for 30 min. Then, theplates were incubated with P3 mAb for 1 h at room temperature.After washing with PBS, the plates were incubated withhorseradish peroxidase–conjugated goat anti-mouse IgM (Cappel,Malvern, PA) for 1 h at room temperature. The plates werewashed again and incubated with the substrate solution consistingof 40 µg/ml O-phenylendiamine (Sigma, St. Louis, MO) in80 mM citrate-phosphate buffer, pH 5.0, containing 0.12% H2O2.The reaction was stopped by dipping the plates in PBS.
To visualize the binding of the P3 mAb to the differentNeuGc-GM3 derivatives, the HPTLC plates were incubated with125I-labeled anti-mouse immunoglobulins (Dakopatts, Denmark)
by guest on Decem
ber 17, 2015http://glycob.oxfordjournals.org/
Dow
nloaded from

E.Moreno et al.
704
as second antibody for 2 h at room temperature, followed bywashing with PBS. Radioactivity on HPTLC plates was detectedby autoradiography (Kodak XAR-5 film, Eastman Kodak,Rochester, NY) and with an excitable phosphorescent screen,which was analyzed using a Bio-Rad GS-250 Molecular Imager(Bio-Rad Laboratories, Inc., USA). For data analysis, thePhosphor Analyst program (Bio-Rad) was used.
Enzyme-linked immunosorbent assay (ELISA)
Solid-phase ELISA was performed as previously described (Taiet al., 1984). In brief, glycolipids were serially diluted in ethanoland applied in a 96-well polystyrene microtiter plate (Dynatech,Alexandria, VA) in volumes of 50 µl/well. P3 mAb was dilutedwith PBS containing 1% human serum albumin. The secondantibody, a peroxidase-conjugated goat anti-mouse IgM, waspurchased from Cappel (Malvern, PA). Three samples of eachexperiment were tested. The standard deviations were less than10% for all values. Background values of absorbance at 405 nmwere less than 0.1.
Molecular modeling
Models of gangliosides carrying NeuGc (GM1a, GM1b, GM2,GM3, GD1a, and GD3) were constructed within Biograf(Molecular Simulations Inc., 1994). Starting values for thedihedral angles over the glycosidic bonds were taken from theliterature (Verulaja and Rao, 1983; Acquotti et al., 1990; Sabesanet al., 1991; Siebert et al., 1992). Molecular dynamics (MD)simulations were carried out using the Dreiding-II force field(Mayo et al., 1990). MD runs for NeuGc-GM3 derivatives werestarted from the anticlinal conformation of the glycosidic linkagebetween sialic acid and galactose (Φ ≈ -155�, Ψ ≈ -30�) and thetorsional angle around the C1-C2 bond of the sialic acid wassearched every 15� in order to choose the minimum energyconformation for the carboxyl group. Complete energy minimiz-ation was carried out before starting the dynamics runs. Theceramide part of GM3 was fixed in these simulations, which wereperformed at 300 K during 1000 ps, using a time step of 0.001 ps.Coordinates were saved every 500 steps. Gasteiger charges anda distance-dependent dielectric constant (ε = 4r) were used. Nohydrogen bond term was included in energy calculations.
MD studies of NeuGc–NeuGc–GD3 were performed usingthe same parameters and conditions given above, except that thetotal run time was 500 ps. Sixteen possible conformations wereconsidered as starting geometries. They were given by thecombination of eight sets of values for the main torsional angles(Φext , Ψext , ω7, and ω8; see Figure 9) involved in theNeuAcα8NeuAc linkage, as found by Brisson et al. (1992), andtwo sets of values for the two torsional angles defining the linkagebetween the internal sialic acid and galactose (Φint and Ψint).Briefly, in the work by Brisson et al. (1992) each of the threestaggered rotamers of ω8 (i.e., g+, g– and anti) yielded several (3,3, and 2, respectively) minimum energy combinations for the Φextand Ψext angles, which were denoted as G1+, G2+, G3+, G1–,G2–, G3–, T1, and T2, respectively. Each of them was combinedin this study with the two energy minima (anticlinal and synclinal)found for the NeuAcα3Gal linkage (Siebert et al., 1992), yieldingconformers AG1+, AG2+, ... ,SG1+, SG2+, ... , ST2. Two of thesestarting geometries (SG2– and SG3–) were not realistic due tosevere steric hindrance, and were therefore discarded. Theremaining 14 conformers were energy-refined prior to thedynamics simulations.
Analysis of known complexes of proteins with sialic acid–containing molecules was carried out using the program Quanta(Molecular Simulations Inc., 1996).
ReferencesAcquotti,D., Poppe,L., Dabrowski,J., von der Lieth,C.-W., Sonnino,S. and
Tettamanti,G. (1990) Three-dimensional structure of the oligosaccharidechain of GM1 ganglioside revealed by distance-mapping procedure: a rotatingand laboratory frame nuclear overhauser enhancement investigation of nativeglycolipid in dimethyl sulfoxide and in water-dodecylphosphocholine sol-utions. J. Am. Chem. Soc., 112, 7772–7778.
Ångström,J., Teneberg,S. and Karlsson,K.-A. (1994) Delineation and comparisonof ganglioside binding epitopes for the toxins of Vibrio cholerae, Escherichiacoli and Clostridium tetani: Evidence for overlapping epitopes. Proc. Natl.Acad. Sci. USA, 91, 11859–11863.
Brisson,J.-R., Baumann,H., Imberty,A., Pérez,S. and Jennings,H.J. (1992) Helicalepitope of the group B meningococcal α(2–8)-linked sialic acid polysacchar-ide. Biochemistry, 31, 4996–5004.
Cahan,L.D., Irie,R.F., Sinyh,R., Cassidenti,A. and Paulson,J.C. (1982) Identifica-tion of human neuroectodermal tumor antigen (OFA-I-2) as ganglioside GD2.Proc. Natl. Acad. Sci. USA, 79, 7929–7933.
Chien,J.-L., Li,S.-C., Laine,R.A. and Li, Y.-T. (1978) Characterization ofgangliosides from bovine erythrocyte membranes. J. Biol. Chem., 253,4031–4035.
Corfield,A.P. and Schauer,R. (1982) Occurrence of sialic acids. Cell Biol.Monogr., 10, 5–50.
Fishman,P.H., Pacuszka,T., Hom,B. and Moss,J. (1980) Modification of ganglio-side GM1. Effect of lipid moiety on choleragen action. J. Biol. Chem., 255,7657–7664.
Furukawa,K., Yamaguchi,H., Oettgen,H.F., Old,L.J. and Lloyd,K.O. (1988)Analysis of the expression of N-glycolyl neuraminic acid-containing ganglio-sides in cells and tissues using two human monoclonal antibodies. J. Biol.Chem., 263, 18507–18512.
Hakomori,S.-i. (1983) In Kanfer,J.N. and Hakomori,S.-i. (eds.), Handbook ofLipid Research. Plenum Press, New York, pp. 1–437.
Halgren,T.A. (1995) Potential energy functions. Curr. Opin. Struct. Biol., 5,205–210.
Hashimoto,Y., Otsuka,H., Sudo,K., Suzuki,A. and Yamakawa,T. (1983) Geneticregulation of GM2 expression in liver of mouse. J. Biochem. (Tokyo), 93,895–901.
Higa,H.H., Rogers,G.N. and Paulson,J.C. (1985) Influenza virus hemagglutininsdifferentiate between receptor determinants bearing N-acetyl-, N-glycolyl-,and N,O-diacetylneuraminic acids. Virology, 144, 279–282.
Imberty,A., Mikros,E., Koca,J., Mollicone,R., Oriol,R. and Pérez,S. (1995)Computer simulation of histo-blood group oligosaccharides: energy maps ofall constituting disaccharides and potential energy surfaces of 14 ABH andLewis carbohydrate antigens. Glycoconjugate J., 12, 331–349.
IUPAC-IUB Commission on Biochemical Nomenclature. (1977) Eur. J. Bio-chem., 79, 11–21.
Jeffrey,P.D., Bajorath,J., Chang,C.Y., Yelton,D., Hellstrom,I., Hellstrom,K.E. andSheriff,S. (1995) The x-ray structure of an anti-tumour antibody in complexwith antigen. Nature Struct. Biol., 2, 466–471.
Karlsson,K.-A., Lanne,B., Pimlott,W. and Teneberg,S. (1991) The resolution intomolecular species on desorption of glycolipids from thin-layer chromato-grams, using combined thin-layer chromatography and fast-atom-bombard-ment mass spectrometry. Carbohydr. Res., 221, 49–61.
Kawashima,I., Ozawa,H., Kotani M., Susuki,M., Kawano,T., Gomobuchi,M. andTai,T. (1993) Characterization of ganglioside expression in human melanomacells: immunological and biochemical analysis. J. Biochem. (Tokyo), 114,186–193.
Lanne,B., Schierbeck,B. and Karlsson,K.-A. (1994) On the role of the carboxylgroup of sialic acid in binding of cholera toxin to the receptor glycosphingoli-pid, GM1. J. Biochem. (Tokyo), 116, 1269–1274.
Lanne,B., Uggla,L., Stenhagen,G. and Karlsson,K.-A. (1995) Enhanced bindingof enterotoxigenic Escherichia coli K99 to amide derivatives of the receptorganglioside NeuGc-GM3. Biochemistry, 34, 1845–1850.
Levery,S.B. (1991) 1H-NMR study of GM2 ganglioside: evidence that aninterresidue amide-carboxyl hydrogen bond contributes to stabilization of apreferred conformation. Glycoconjugate J., 8, 484–492.
Miyake,M., Ito,M., Hitomi,S., Ikeda,S., Taki,T., Kurata,M., Hino,A., Miyake,N.and Kannagi,R. (1988) Generation of two murine monoclonal antibodies thatdiscriminate N-glycolyl and N-acetyl neuraminic acid residues of GM2ganglioside. Cancer Res., 48, 6154–6160.
Mayo,S.M., Olafson,B.D. and Goddard III,W.A. (1990) Dreiding: a generic forcefield for molecular simulations. J. Phys. Chem., 94, 8897–8909.
by guest on Decem
ber 17, 2015http://glycob.oxfordjournals.org/
Dow
nloaded from

Binding specificity of mouse mAb P3
705
Merritt,E.A., Sarfaty,S., van den Akker,F., L’hoir,C., Martial,J.A. and Hol,W.G.J.(1994) Crystal structure of cholera toxin B-pentamer bound to receptor GM1pentasaccharide. Prot. Sci., 3, 166–175.
Nakamura,K. and Handa,S. (1986) Biochemical properties of N-methylamides ofsialic acid in gangliosides. J. Biochem. (Tokyo), 99, 219–226.
Ozawa,H., Kawashima,I. and Tai,T. (1992) Generation of murine monoclonalantibodies specific for N-glycolylneuraminic acid-containing gangliosides.Arch. Biochem. Biophys., 294, 427–433.
Peters,B.P., Ebisu,S., Goldstein,I.J. and Flashner,M. (1979) Interaction of wheatgerm agglutinin with sialic acid. Biochemistry, 18, 5505–5511.
Poppe,L., Dabrowski,J., von der Lieth,C.-W., Numata,M. and Ogawa,T. (1989)Solution conformation of sialosylcerebroside (GM4) and itsNeuAc(α2→3)Galβ sugar component. Eur. J. Biochem., 180, 337–342.
Sabesan,S., Duuss,J.O., Fukunaga,T., Bock,K. and Schauer,R. (1991) NMR andconformational analysis of ganglioside GD1a. J. Am. Chem. Soc., 113,3236–3246.
Sanai,Y., Yamasaki,M. and Nagai,Y. (1988) Monoclonal antibody directed to aHanganutziu-Deicher active ganglioside, GM2(NeuGc). Biochim. Biophys.Acta, 294, 427–433.
Sauter,N.K., Hanson,J.E., Glick,G.D., Brown,J.H., Crowther,R.L., Park,S.-J.,Skehel,J.J. and Wiley,D.C. (1992) Binding of influenza virus hemagglutinin toanalogs of its cell-surface receptor, sialic acid: analysis by proton nuclearmagnetic resonance and x-ray crystallography. Biochemistry, 31, 9609–9621.
Schengrund,C.-L. and Ringler,N.J. (1989) Binding of Vibrio cholerae toxin andthe heat-labile enterotoxin of Escherichia coli to GM1, derivatives of GM1,and nonlipid oligosaccharide polyvaent ligands. J. Biol. Chem., 264,13233–13237.
Sharon,N. (1987) Nomenclature of glycoproteins, glycopeptides and peptidogly-cans. J. Biol. Chem., 262, 13–18.
Siebert,H.-C., Reuter,G., Schauer,R., von der Lieth,C.-W. and Dabrowski,J.(1992) Solution conformations of GM3 gangliosides containing differentsialic acid residues as revealed by NOE-based distance mapping, molecularmechanics, and molecular dynamics simulations. Biochemistry, 31,6962–6971.
Spiegel,S. (1985) Fluorescent derivatives of ganglioside GM1 function asreceptors for cholera toxin. Biochemistry, 24, 5947–5952.
Svennerholm,L. (1964) The gangliosides. J. Lipid Res., 5, 145–155.Svennerholm,L. (1976) Ganglioside isolation. Methods Carbohydr. Chem., 6,
464–474.Tai,T., Paulson,J.C., Cahan,L.D. and Irie,R.F. (1983) Ganglioside GM2 as a
human tumor antigen (OFA-I-1). Proc. Natl. Acad. Sci. USA, 80, 5392–5396.Tai,T., Cahan,L.D., Paulson,J.C., Saxton,R.E. and Irie,R.F. (1984) Human
monoclonal antibody against ganglioside GD2: use in development ofenzyme-linked immunosorbent assay for the monitoring of anti-GD2 incancer patients. J. Natl. Cancer Inst., 73, 627–633.
Varghese,J.N., McKimm-Breschkin,J.L., Caldwell,J.B., Kortt,A.A. and Colman,P.M. (1992) The structure of the complex between influenza virus neuramini-dase and sialic acid, the viral receptor. Proteins, 14, 327–332.
Vázquez,A.M., Alfonso,M., Lanne,B., Karlsson,K.-A., Carr,A., Barroso,O.,Fernández,L.E., Rengifo,E., Lanio,M.E., Alvarez,C., Zeuthen,J. and Pérez,R.(1995) Generation of a murine monoclonal antibody specific againstN-glycolylneuraminic acid-containing gangliosides that also recognizessulfated glycolypids. Hybridoma, 14, 551–556.
Veh,R.W., Corfield,A.P., Sander,M. and Schauer,R. (1976) Neuraminic acid-spe-cific modification and tritium labelling of gangliosides. Biochim. Biophys.Acta, 486, 145–160.
Veluraja,K., and Rao,V.S.R. (1983) Theoretical studies on the conformation ofmonosialogangliosides and disialogangliosides. Carbohydr. Polym., 3,175–192.
Watarai,S., Onuma,M. and Yasuda,T. (1991) Production of monoclonal antibodiesspecific for ganglioside GD3. J. Biochem. (Tokyo), 110, 889–895.
Wright,C.S. (1990) 2.2 Å resolution structure analysis of two refined N-acetylneu-raminyl-lactose-wheat germ agglutinin isolectin complexes. J. Mol. Biol.,215, 635–651.
by guest on Decem
ber 17, 2015http://glycob.oxfordjournals.org/
Dow
nloaded from
Copyright © 2022 FDOKUMEN




















