
The Outcomes and Quality of Pancreatic Islet Cells Isolated ...

A Low Protein Diet Alters the Balance of Islet CellReplication and Apoptosis in the Fetal and Neonatal Ratand Is Associated with a Reduced Pancreatic Expressionof Insulin-Like Growth Factor-II*
J. PETRIK, B. REUSENS, E. ARANY, C. REMACLE, C. COELHO, J. J. HOET, AND
D. J. HILL
Lawson Research Institute (J.P., E.A., C.C., D.J.H.), St. Joseph’s Health Centre, London, Ontario N6A4V2, Canada; Departments of Physiology (J.P., C.C., D.J.H.), Medicine (E.A., D.J.H.), and Paediatrics(J.P., D.J.H.), University of Western Ontario, London, Ontario N6A 5A5, Canada; and Laboratoire deBiologie Cellulaire (B.R., C.R., J.J.H.), World Health Organization Collaborating Centre for theDevelopment of the Biology of the Endocrine Pancreas, Universite Catholique de Louvain, B-1348Louvain-La-Neuve, Belgium
ABSTRACTA programmed turnover of pancreatic b cells occurs in the neonatal
rat involving a loss of b cells by apoptosis, and their replacement byislet cell replication and neogenesis. The timing of apoptosis is asso-ciated with a loss of expression of a survival factor, insulin-like growthfactor-II (IGF-II), in the pancreatic islets. Offspring from rats chron-ically fed a low protein isocalorific diet (LP) exhibit a reduced pan-creatic b cell mass at birth and a reduced insulin secretion in later life.This study therefore investigated the impact of LP on islet cell on-togeny in the late fetal and neonatal rat, and any associated changesin the presence of IGFs and their binding proteins (IGFBPs). Preg-nant Wistar rats were fed either LP (8% protein) or normal (C) (20%protein) chow from shortly after conception until the offspring were21 days postnatal (PN). Bromo-deoxyuridine (BrdU) was adminis-tered 1 h before rats were killed and pancreata removed from animalsbetween 19.5 days fetal life and postnatal day 21. Offspring of ratsgiven LP diet had reduced birthweight, pancreatic b cell mass, andpancreas insulin content, with smaller islets compared with controlfed animals, which persisted to weaning. Histological analysis showedthat islets from pups given LP diet had a lower nuclear labeling indexwith BrdU in the b cells, although, paradoxically, more b cells showedimmunoreactivity for proliferating cell nuclear antigen (PCNA). Be-
cause PCNA is present in G1 as well as S phase of the cell cycle, wequantified the number of b cells immunopositive for cyclin D1, amarker of G1, and NEK2, an indicator of cells in G2 and mitosis. Moreb cells in islets from LP-fed animals contained cyclin D1, but lesscontained NEK2 than did those in controls. This suggests that the bcell cycle may have a prolonged G1 phase in LP-fed animals in vivo.Offspring of rats given C diet had a low rate of islet cell apoptosisdetected by the TUNEL method in fetal and neonatal life (1–2%), witha transient increase to 8% at PN day 14. Offspring of rats receivingLP diet demonstrated a significantly greater level of islet cell apo-ptosis at every age, rising to 15% at PN 14. IGF-II mRNA was quan-tified in whole pancreas and was significantly reduced in LP-fedanimals at ages up to PN day 10. IGF-II immunoreactivity within theislets of LP-fed rats was also less apparent, but no changes were seenin immunoreactive IGF-I or IGFBPs-2 to -5. These findings show thatLP diet changes the balance of b cell replication and apoptosis in fetaland neonatal neonatal life, which may involve an altered length of bcell cycle, and contribute to the smaller islet size and impaired insulinrelease seen in later life. A reduced pancreatic expression of IGF-IImay contribute to the lower b cell proliferation rate and increasedapoptosis seen in the fetus and neonate after feeding LP diet. (En-docrinology 140: 4861–4873, 1999)
FETAL UNDERNUTRITION can result in intrauterinegrowth restriction (IUGR), reduced birthweight, and an
increased incidence of adult onset diseases such as type 2diabetes and coronary heart disease (1–3). We have used amodel in which pregnant rats receive throughout gestationa low protein (LP). Birthweight is significantly reduced in theoffspring of LP rats that have altered blood amino acid pro-files, although blood glucose and plasma insulin levels donot differ greatly from controls (4). The pancreatic weight is
significantly lower in LP pups at birth as is mean islet area(5), due in part to a reduced proliferative capacity and also,perhaps, to a reduced vascularity in the endocrine pan-creas (5). These animals have also been studied at adult agefollowing a lactation by mothers receiving normal proteinintake and fed a normal diet after weaning. As adults, theyfeature a slight reduction in the islet cell mass and a lowpancreatic insulin content. An abnormally low release ofinsulin in response to amino acids was found when iso-lated islets were examined in vitro, and lower insulin re-sponse to a oral glucose challenge in vivo was observed infemales (6 – 8). Clearly, dietary experiences in utero canlead to long-lasting functional deficiencies in the endo-crine pancreas and can program a susceptibility to abnor-mal glucose tolerance.
Potentially important events in pancreatic islet ontogenyhave been recognized in the neonatal rat, which may explainwhy this is a period of sensitivity to nutritional insult. In the
Received March 12, 1999.Address all correspondence and requests for reprints to: Dr. D. J. Hill,
Lawson Research Institute, St. Joseph’s Health Centre, 268 GrosvenorStreet, London, Ontario, Canada, N6A 4V2. E-mail: [email protected].
* We are grateful to the Juvenile Diabetes Foundation, the CanadianDiabetes Association, the Medical Research Council of Canada, and theFond National de la Recherche Scientifique of Belgium for financialsupport.
0013-7227/99/$03.00/0 Vol. 140, No. 10Endocrinology Printed in U.S.A.Copyright © 1999 by The Endocrine Society
4861

rat fetus islet cell, mass increases rapidly due to both b cellreplication and recruitment, and maturation of undifferen-tiated b cell precursors within the pancreatic ducts (9, 10).Following birth, the growth rate of all islet cells, including bcells, declines within 3–4 days and continues to decline there-after (10), so that the rate of mitosis in adult pancreatic b cellsis low (11). A wave of apoptosis occurs in neonatal rat isletsbetween 1–2 weeks of age (12), although because the totalpancreatic b cell mass is not substantially changed this sug-gests that a new population of b cells derived from replica-tion or neogenesis compensates for the loss. A similar b cellapoptosis has recently been described in the human fetalpancreas in third trimester (13). This ontogeny of b cells maysignal a change from a population suited to fetal life in whichacute insulin release is not necessary within a stable nutri-tional environment, to b cells with altered glucose thresholdsand the ability to rapidly release insulin under nutritional,endocrine and neural control characteristic of adult meta-bolic control.
We recently showed that the timing of neonatal b cellapoptosis in the rat coincided with a loss of expression ofinsulin-like growth factor-II (IGF-II) within the pancreaticislets, and that IGFs can functionally act as a survival factorsto prevent apoptosis in b cells (14), a role consistent with theactions of IGFs in other cell types (15, 16). There is alsoconsiderable evidence that the IGFs contribute to b cellgrowth, maturation, and function throughout life, and thattheir actions can be modulated by locally produced IGFbinding proteins (IGFBPs). Messenger RNA for IGF-II isabundant in the pancreas of the fetal rat, and declines rapidlywithin 2 weeks of birth, whereas IGF-I mRNA expression islow in the fetus, but increases to adult levels by weaning (17).Isolated islets from the human fetus (18), or rat fetus orneonate (19, 20), release IGF-I and/or -II, which are capableof increasing islet cell DNA synthesis (19–21). The mitogenicaction of IGF-II in isolated fetal rat islets was potentiated byIGFBPs-1 and-2, which together with other IGFBPs are ex-pressed there (19). Both IGF-I and -II can also rapidly mod-ulate insulin release from adult rat islets in a biphasic manner(22, 23).
The IGF axis is highly responsive to nutritional status (24,25). Intrauterine growth restriction in man (26, 27), and ex-perimentally induced growth retardation in the rat fetus(28–30), including protein restriction (31), is associated witha reduction in circulating IGFs and an altered presence ofIGFBPs. Using isolated islets from the fetal rat pancreas oradult hamster, the release of IGFs and IGFBP-1 and -2 wereshown to be enhanced by amino acids and/or glucose(19, 32).
Because of the reorganization of the endocrine pancreasduring the suckling period, and the role of amino acids inb cell development, the purpose of this study was to,firstly, examine how LP diet might alter the balance of isletcell replication and survival in the late fetal and neonatalrat endocrine pancreas; and, secondly, to establish if al-terations in the pancreatic expression of the IGF axis mightbe temporally associated with induced developmentalchanges.
Materials and MethodsAnimal model
The model of pre- and postnatal exposure to a low protein diet hasbeen described by us previously (5). Adult female Wistar rats bred at theCatholic University of Louvain were given food and water ad libitum andwere housed at 24 C with a 60% humidity and a 14-h light, 10-h darkcycle. Nulliparous rats of 200–220 g maintained on standard laboratorydiet were time mated and randomly allocated to one of the two groupson day 1 of gestation until weaning. A control group (C) was providedwith a 20% protein diet and a second group (LP) was given an 8% proteindiet (Hope Farms, Woerden, The Netherlands). The two diets had asimilar fat content and were made isocalorific by the addition of car-bohydrates to the LP diet. We have previously shown that food intakeis not appreciably different between the control and LP groups (5).
At the time of birth, litters from both diet groups were reduced toeight pups, and these remained with the lactating females until theywere killed at up to 21 days. Pregnant rats were anesthetized withpentobarbital (55 mg/kg body weight) on days 19.5 or 21.5 gestation.The abdomen and uterus was opened, and fetal blood samples collectedthrough the axillary vessels; the fetus remaining attached to the mothervia the placenta throughout the procedure. Postnatally, pups were killedby decapitation. Because of the higher incidence of glucose intolerancein the adult female offspring of rats subjected to the LP diet duringpregnancy (8), only female offspring was used in this experiment. Preg-nant mothers or pups were injected sc with 5-bromo-2-deoxyuridine(BrdU, 50 mg/kg body weight in sterile saline, Roche Molecular Bio-chemicals Biochemica, Brussels, Belgium) 1 h before sacrifice to labelnewly formed DNA. At the time of death the animals were weighed andpancreas removed from each animal and either snap-frozen in liquidnitrogen and stored at 280 C, for RNA isolation, or fixed for immuno-histochemistry or in situ hybridization. If pancreata were to be used forhistology they were placed in ice-cold fixative (4% paraformaldehyde in70 mm phosphate buffer, pH 7.4, containing 0.2% glutaraldehyde) for16 h at 4 C, followed by four washes at 4 C in PBS over a 48-h period.Fixed tissues were dehydrated through a graded ethanol series, im-pregnated with butanol and embedded in paraffin. All procedures wereperformed with approval of the animal ethics committees of the CatholicUniversity of Louvain and the University of Western Ontario, and inaccordance with the guidelines of the Canadian Council on Animal Care.
Fifty microliters of blood was collected for glucose and precipitatedin 500 ml HClO4 (0.33N). Glucose concentrations were measured usinga glucose oxidase test kit (Sigma, St. Louis, MO). Plasma was alsoprepared, and the insulin content measured by RIA (INSIK-5 P2796,Sorin Biomedica, Saluggio, Italy), using rat insulin for the standard curve(Novo Nordisk, Copenhagen, Denmark). Pancreas (15–25 mg) was re-moved and homogenized in 5 ml acid-ethanol (0.15 m HCl in 75%[vol/vol] ethanol) and extracted overnight at 4 C. The insulin contentwas determined by RIA and correlated with body weight.
Northern blot analysis
Total RNA was extracted from pancreata as previously described (17).Before hybridization, the integrity and relative amounts of RNA fromeach pancreas were assessed by size separation on 10% agarose TAE(Tris HCl-EDTA) gels with ethydium bromide. Pancreata in which ri-bosomal RNA showed degradation were not used for experiments.
Northern blot hybridization was performed as described previously(17) using between 15 and 20 mg of total RNA following separation bysize on agarose gels. Hybridization was performed with 2 3 106 cpm/mlradiolabeled complementary DNA (cDNA) probe for IGF-I or -II, or 1 3106 cpm/ml for a cDNA encoding 18S ribosomal RNA. Blots wereexposed to x-ray film (Kodak XAR-5, Eastman Kodak Co. Inc., Rochester,NY) at 270 C with intensifying screens for up to 10 days before devel-oping, and were hybridized consecutively with cDNAs for IGF-II fol-lowed by 18S ribosomal RNA. Between consecutive hybridizations, theblots were stripped with 0.01 3 SSC with 0.5% (wt/vol) SDS at 90 C for1 h to remove the previous labeled cDNA probe.
Radiolabeling of cDNA probes with a(32P)-dCTP (ICN Biomedicals,Inc., Irvine, CA) was carried out by random priming using a Pharmaciaoligolabelling kit (Pharmacia LKB Biotechnology, Uppsala, Sweden), tospecific activities of 1–2 3 109 dpm/mg. Separation of radiolabelledcDNA from unincorporated (32P)-dCTP was carried out using
4862 PANCREATIC IGF AXIS AND LOW PROTEIN DIET Endo • 1999Vol 140 • No 10

ProbeQuant G-50 microcolumns (Pharmacia). Complimentary DNAprobes used for hybridization of Northern blots and for in situ hybrid-ization were kindly provided by the following investigators: a 500 bp ratIGF-I cDNA in pGEM Blue (Promega Corp., Madison, WI) encodingexon 3 and part of exon 4 of the rIGF-I gene was provided by Dr. L.Murphy, University of Manitoba, Winnipeg, MT (33); a 807bp mouseIGF-II in pGEM 4z by Dr. G. Bell, University of Chicago, Chicago, IL; anda rat 18S ribosomal RNA cDNA by Dr. D. Denhardt, Rutgers University,Piscataway, NJ The latter was used to determine equality of RNA load-ing and transfer.
In situ hybridization
We performed in situ hybridization of IGF-I and -II mRNAs usinghistological paraffin sections of pancreas as described in detail by uspreviously (14). Kodak NTB-3 photoemulsion diluted 1:1 with water wasapplied subsequently to all sections and exposed for up to 14 days at 4C, then slides were developed in Kodak D19, rinsed in water and fixedin Kodafix. Sections were counter-stained with hematoxylin and eosin.Slides were viewed under dark and lightfield microscopy. As controlsfor nonspecific hybridization, hybridization was also carried out usingsense strand cRNA probes. Where comparisons were made betweenpancreata from animals of different ages all sections were included inthe same hybridization reactions to remove between procedure error.
Antisense riboprobes were prepared from cDNAs for rat (r) IGF-I (agift of Dr. L. Murphy, University of Manitoba, Canada), and mouse (m)IGF-II (a gift of Dr. G. Bell, Univ. of Chicago). The restriction enzymes(BRL, Burlington, ON) and RNA polymerases (Promega Corp.) used tolinearize the plasmids containing these cDNAs and to generate [35S]-radiolabelled riboprobes were: antisense rat IGF I, HindIII/T7; sense ratIGF I, PvuII/SP6; antisense mouse IGF-II, HindIII/TSP6; sense mouseIGF-II EcoRI/T7. Radiolabelled cRNA probes were synthesised usinglinearized riboprobe DNA, a-thio [35S]-UTP, and SP6, T3 or T7 RNApolymerase as described previously (14). To improve cRNA probe pen-etration of tissue sections, limited alkaline hydrolysis was used to reducetranscript size to about 150 bases.
Immunohistochemistry
Histological sections of pancreas (5 mm) were cut from paraffin blocksand mounted on glass microscope slides (Superfrost Plus, Fischer Sci-entific, Nepeon, ON, Canada). Immunohistochemistry was performedto localize IGF-I or -II, IGFBPs, inducible nitric oxide synthase (iNOS),BrdU, proliferating cell nuclear antigen (PCNA), insulin, glucagon, so-matostatin, cyclin D1 and NEK2 within islets by a modified avidin-biotinperoxidase method (34) as described by us previously for pancreas (14).Slides were incubated for 48 h at 4 4]C with either rabbit antihuman IGF-Ior IGF-II (1:2000 dilution) (GroPep Pty. Ltd. Ltd., Adelaide, Australia);rabbit antihuman IGFBP-1, -2, -3, -4 and -5 (all at 1:100 dilution), ormonoclonal antibody against human IGFBP-6 (1:500 dilution) (AustralBiologicals, San Remon, CA); guinea pig antiinsulin antibody (1:500dilution) (provided by Dr. T. J. McDonald, University of Western On-tario, London, Ontario, Canada); rabbit antiporcine glucagon (1:100 di-lution) (C-terminal specific 04A antiserum kindly provided by Dr. R.Ungar, Dallas, TX); rabbit antirat somatostatin (1:100) (DAKO Corp.Laboratories, Mississauga, Ontario, Canada); mouse anti-iNOS anti-serum (1:50 dilution) (Transduction Laboratories, Inc., Lexington, KY);mouse antiproliferating cell nuclear antigen (PCNA) (1:750 dilution)(Sigma, St. Louis, MO); mouse anti-BrdU (labeling kit supplied byZymed Laboratories, Inc., South San Francisco, CA); rabbit anti-NEK2(1 mg/ml dilution) (Zymed Laboratories, Inc.); and mouse anticyclin D1(1:500 dilution) (Zymed Laboratories, Inc.). All antisera were diluted in0.01 m PBS (pH 7.5) containing 2% (wt/vol) BSA and 0.01% (wt/vol)sodium azide (100 ml per slide). Biotinylated goat antirabbit IgG (1:100),goat antimouse IgG (1:100), or mouse antiguinea pig IgG (1:500) (VectorLaboratories, Inc. Burlingame, CA), were used as secondary antibodies.Peptide immunoreactivity was localised by incubation in fresh diami-nobenzidine tetrahydrochloride (DAB tablets, 10 mg, Sigma) with 0.03%(vol/vol) hydrogen peroxide for 2 min and the reaction quenched inexcess 50 mm Tris.HCl pH 7.5. Tissue sections were counter-stained withCarazzi’s hematoxylin.
To establish specificity of staining, the primary antisera for IGF-I or-II were preadsorbed overnight at 4 C with 100 nm homologous antigen
before application to the sections. In each case staining was abolished.Antisera against IGFBPs were preabsorbed with 100 nm of homologousor heterologous IGFBP proteins (Austral). All antisera were found to bespecific as stated by the supplier with the exception of antiserum againstIGFBP-4, which showed cross-reactivity with IGFBP-2. Further controlsincluded substitution of primary antisera with nonimmune serum andomission of the secondary antiserum.
Dual staining for BrdU, PCNA, cyclin D1, or NEK2, and insulin wasperformed by first performing immunohistochemistry for insulin asdescribed above using diaminobenzidine as the chromagen. Beforecounterstaining and dehydration the sections were then subjected toimmunohistochemistry for the second antigen as described with theexception that alkaline phosphatase (blue) was used as the chromogen.Alkaline phosphatase substrate kit III was obtained from Vector Lab-oratories, Inc. Antimouse alkaline phosphatase conjugate (Sigma) wasapplied to each section for 1 h at room temperature, sections washed, andalkaline phosphatase substrate applied for 20 min. Sections were washedand counterstained with Mayer’s haemalum.
Visualization of apoptosis
Immunohistochemistry was performed to localize apoptotic nucleiwithin histological sections of pancreas (35) using the Apoptag in situapoptosis detection kit (Oncor Inc., Gaithersburg, MD), as described indetail by us previously (14). Staining was performed according to themanufacturer’s protocol, following incubation with proteinase K (20mg/ml; Roche Molecular Biochemicals, Dorval, Quebec, Canada) for 15min, washing in distilled water, and the quenching of endogenousperoxidase by incubation in 2% (vol/vol) hydrogen peroxide in PBS for5 min. Color was generated with diaminobenzidine as described forimmunohistochemistry, and the tissue counterstained with Carazzi’shematoxylin for 1 min. Sections were dehydrated in butanol, cleared inxylene and mounted with Permount under glass coverslips. Duel stain-ing for apoptosis and insulin was performed as described above.
Morphometric and statistical analysis
Morphometric analysis was performed using a Carl Zeiss transmittedlight microscope at a magnification of 3250 or 3400. Analyses wereperformed with Northern Eclipse version 2.0 morphometric analysissoftware (Empix Imaging Co., Mississauga, Ontario, Canada). The per-centage of islet cells, or the islet cell area, immunopositive for IGF-I orIGF-II, IGFBPs, insulin, glucagon, somatostatin, BrdU, PCNA, cyclin D1,NEK2 or iNOS; or demonstrating apoptotic nuclei, was calculated ateach age from up to five sections of each pancreas representing pre-dominantly the head regions. This was because we have previouslyshown that the impact of LP diet on islet size is greater in the head thanin the tail of the pancreas (5). Sections chosen contained at least five islets,and pancreata from up to 10 animals were examined for each age.Individual cell area, and total areas of immunoreactive cells within isletswere circled for image analysis and selected by gray-level threshhold.Pancreatic b cell mass was calculated following immunohistochemistryfor insulin on sections obtained throughout pancreata of known weight.The mean b cell volume was found to be 650 mm3. b cell mass wasestimated by calculating the mean percentage area of tissue containingcells immunoreactive for insulin per sectional area of pancreas (five tosix sections per organ). This was then expressed as mg b cell mass basedon the total pancreatic wet weight for individual animals. For Northernblot analysis the ratio of hybridization signal for IGF-II compared with18S rRNA was calculated at each age for each of three separate pancreatafollowing scanning densitometry. Differences between mean values forvariables within individual experiments were compared statistically bytwo way ANOVA, followed by a Scheffe’s test.
ResultsMetabolism
Feeding of LP diet significantly reduced mean fetal bodyweight at 19.5 or 21.5 days gestation, compared with an-imals fed control diet (day 19.5, control 2.03 6 0.03 g, LP1.89 6 0.05 g, P , 0.02; day 21.5, control 4.48 6 0.06 g, LP4.14 6 0.14 g, P , 0.05; mean 6 sem, n 5 15–20). Pups born
PANCREATIC IGF AXIS AND LOW PROTEIN DIET 4863

to mothers receiving LP diet continued to have a lowerbody weight postnatally, and grew at a reduced velocityuntil postnatal day 21 when they had a mean body weight33% less than controls (control 42.02 6 2.31 g, LP 27.80 61.71 g, P , 0.001, n 5 5 per group). The mean pancreaticweight did not significantly differ between control and LPgroups in the fetus and newborn but was significantly lessin the LP group by postnatal day 2. At postnatal day 14 theweight of the pancreas was 46% less in offspring of LP-fedrats compared with controls (Fig. 1A). Blood glucose levelsin fetuses of both LP and control fed rats did not differsignificantly at any age. At gestational age 19.5 days themean (6sem) blood glucose value was 46.5 6 3.5 mg/dl,on the day of birth 96.8 6 4.4 mg/dl, and on postnatal day
10, 123.6 6 2.9 mg/dl (n 5 10 per group). Thereafter, nosubstantial change was seen up to 21 days of age. Meanplasma insulin levels did not significantly differ betweenoffspring of LP and control rats. The combined mean in-sulin values decreased sharply after birth in both dietgroups, being 8.4 6 0.7 ng/ml on day 21.5 gestation, 4.6 60.6 ng/ml on postnatal day 2, and 1.8 6 0.7 ng/ml bypostnatal day 21 (n 5 10 per group). At postnatal day 21the pancreatic insulin content of LP rats was significantlyless than in control animals (control 18.3 6 1.1 mg/pan-creas, LP 13.0 6 1.2, P , 0.05, n 5 8) but this difference wasnot seen if insulin content was related to body weight(control 48.6 6 3.6 mg/100 g body weight, LP 50.8 6 4.2).
FIG. 1. A, Mean (6SEM) pancreaticweight in rats from 21.5 days gestationto 14 days postnatal age following feed-ing of control (open bars) or low protein(LP, shaded bars) diets, and (B) meanislet cell area in pancreata from ani-mals of 19.5 days fetal (F) life, or post-natal (P) days 14 or 21. Figures for pan-creatic weight are derived from fiveanimals at each age, and data for isletarea from 20–25 observations for eachage using five animals. l*, P , 0.001vs. control diet.
4864 PANCREATIC IGF AXIS AND LOW PROTEIN DIET Endo • 1999Vol 140 • No 10

Morphology
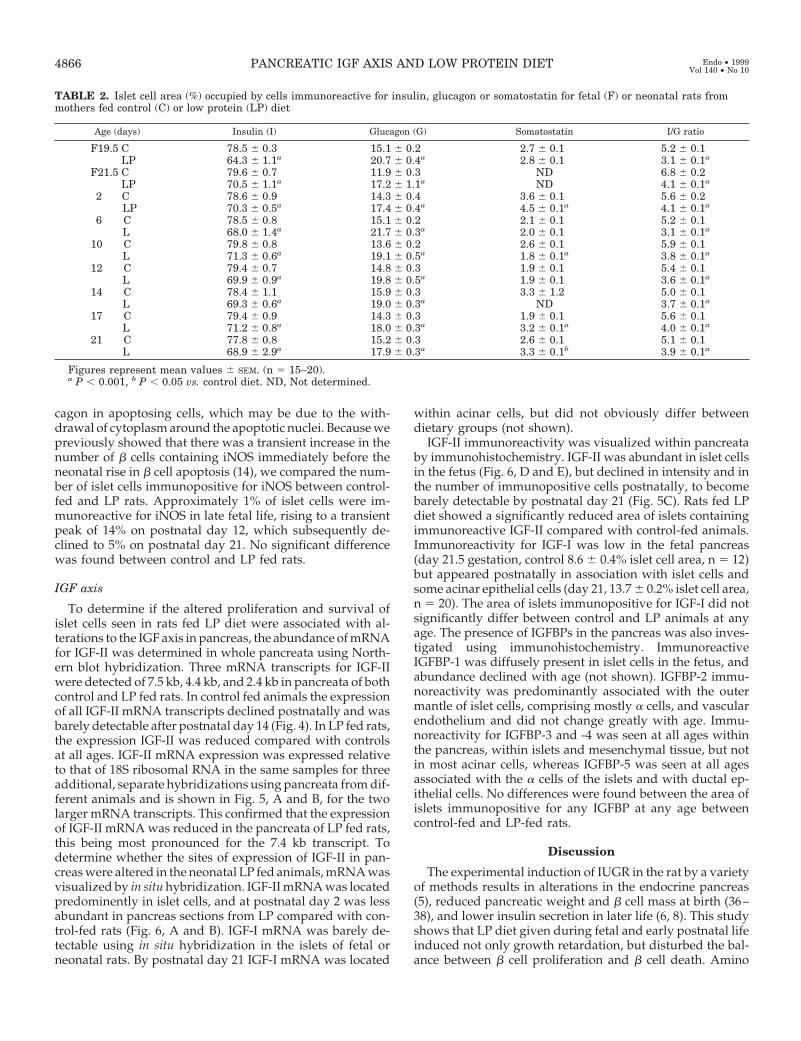
The mean area of islets was compared between LP andcontrol fed rats at 19.5 days gestation, and postnatal days 14and 21. Mean islet area increased from approximately 10,000mm2 in late fetal life to 25,000 mm2 by postnatal day 21 in thecontrol group (Fig. 1B). The mean islet area was significantlyreduced by more than 50% at each age in the LP group. Themean b cell mass per pancreas was calculated and found tobe reduced by greater than 50% in LP compared with controlrats at fetal and neonatal ages, and by 70% at postnatal day14 (Table 1). Analysis of the abundance of endocrine celltypes within the islets showed that the relative distributionsdid not alter with age, but that b cells containing immuno-reactive insulin made up a significantly smaller area of theislets from LP rats at all ages, whereas a cells containingimmunoreactive glucagon constituted a larger area (P ,0.001) (Table 2). Consequently, the insulin/glucagon ratio forrelative immunoreactive islet area was decreased at all agesin LP animals (P , 0.001). The area occupied by somatostatin-containing D cells was not significantly altered between LPand control rat islets at most ages (Table 2). Individual en-docrine cells did not differ in size between control and LPanimals, suggesting that the above results reflected differ-ences in relative cell number. When apoptosing cells withinislets were also considered (Fig. 3D), which did not showimmunoreactivity for any pancreatic endocrine hormone,over 93% of islet cells could be accounted for, the balanceprobably reflecting pancreatic polypeptide-containing cellsthat were not analyzed. Thus, the islets in LP rats weresmaller and contained a relatively smaller proportion of bcells compared with controls throughout fetal and neonatallife.
Cell cycle analysis
Cells traversing S phase of the replication cycle were la-beled in vivo with BrdU before rats were killed, which wasthen visualized within histological sections of pancreas usingimmunocytochemistry (Fig. 2, A and B). The overall labelingindex of islet cells with BrdU was 4–5% in control fed ani-mals, and did not alter with age. The percent islet cells la-beled with BrdU in LP rats was significantly lower (control4.4 6 0.1%, LP 2.6 6 0.1%, n 5 15 representing five animalseach at gestational day 21.5 and postnatal days 14 and 21, P ,0.001). Dual staining for insulin and BrdU showed that mostcells, which labeled with BrdU were b cells, that this pro-portion did not change with age, but that a significantly
lower percentage of cells that were BrdU-positive in LP an-imals were identified as b cells (control 78.0 6 2.3%, LP 62.1 63.5%, P , 0.001, n 5 50 animals for control and 101 for LP).This suggests that the labeling of b cells with BrdU was inproportion to their relative abundance in the islets of bothcontrol and LP animals.
As an additional parameter of islet cell replication, sectionsof pancreas were examined for the presence of PCNA byimmunohistochemistry. PCNA is present in cell nucleiwithin the later part of G1, S, and G2 phases of the cell cycle.In animals fed control diet, the islet cell labeling index forPCNA was between 1% and 4%, with a transient increase atpostnatal day 12. In contrast to findings with BrdU labeling,the percentage of islet cells immunopositive for PCNA wassignificantly greater in LP compared with control rats atevery age up to postnatal day 17 (Fig. 3A). When only b cellswere considered, the percentage of b cells that demonstratedimmunoreactivity for PCNA and insulin was also signifi-cantly greater in islets of LP animals and did not significantlydiffer with age (control 2.3 6 0.8%, LP 3.6 6 0.9%, P , 0.001,n 5 63 for control and 60 for LP). Approximately 60–70% ofcells containing PCNA immunoreactivity were b cells, andthis did not significantly alter with age or with diet (notshown).
To further delineate cell cycle events in islet cells in situ,immunohistochemistry was performed for cyclin D1 and forNEK2. Cyclin D1 is associated with G1 phase of the cell cycle,whereas NEK2 has a cell cycle-dependent expression that ismaximal in G2 and M. Cyclin D1 was visualized in thecytoplasm of a minority of islet cells, and in some acinarepithelial cells (Fig. 2, C and D). The percentage of b cells thatdemonstrated immunoreactivity for cyclin D1 did notchange with age in control-fed animals, but was significantlygreater in LP-fed animals than in the control animals in latefetal life and at postnatal days 14 and 17 (Fig. 3B). NEK2immunoreactivity was seen in the nuclei of a minority of isletcells and in clusters of acinar cells at all ages (Fig. 2, E andF). In rats given LP diet the percentage of b cells demon-strating NEK2 immunoreactivity was significantly reducedbetween postnatal days 10 and 17 (Fig. 3C). Thus, indicatorsof S phase transition and G2/M of the cell cycle for b cells(BrdU and NEK2) showed a decreased incidence in LP ratspostnatally, while indicators of G1 phase (PCNA and cyclinD1) demonstrated an increased incidence.
Apoptosis
The occurrence of apoptosis within islet cells at each agewas examined by molecular histochemistry using theTUNEL method. Apoptosing cells were restricted to the cen-tral, b-cell rich region of the islets (Fig. 2, G and H). In thepancreas of control animals the proportion of islet cells un-dergoing apoptosis was less than 2%, before postnatal day 10,but a transient rise in islet cell apoptosis occurred betweendays 12 and 17, which was maximal at 8% on postnatal day14 (Fig. 3D). In offspring of rats receiving the LP diet, theincidence of islet cell apoptosis was significantly higher atevery age examined, including the transient neonatal rise ondays 12–17. Colocalization immunohistochemistry failed todemonstrate the presence of immunoreactive insulin or glu-
TABLE 1. Beta cell mass (mg) for pancreata from fetal (F) orneonatal rats from mothers fed control (C) or low protein (LP) diet
Age (days) b cell mass (mg)
F21.5 C 1.14 6 0.15LP 0.36 6 0.12a
2 C 2.22 6 0.11LP 0.73 6 0.18a
12 C 3.43 6 0.32LP 1.13 6 0.28b
14 C 5.26 6 0.25LP 1.44 6 0.22b
Figures represent mean values 6 SEM. (n 5 5).a P , 0.05, b P , 0.001 vs. control diet.
PANCREATIC IGF AXIS AND LOW PROTEIN DIET 4865
cagon in apoptosing cells, which may be due to the with-drawal of cytoplasm around the apoptotic nuclei. Because wepreviously showed that there was a transient increase in thenumber of b cells containing iNOS immediately before theneonatal rise in b cell apoptosis (14), we compared the num-ber of islet cells immunopositive for iNOS between control-fed and LP rats. Approximately 1% of islet cells were im-munoreactive for iNOS in late fetal life, rising to a transientpeak of 14% on postnatal day 12, which subsequently de-clined to 5% on postnatal day 21. No significant differencewas found between control and LP fed rats.
IGF axis
To determine if the altered proliferation and survival ofislet cells seen in rats fed LP diet were associated with al-terations to the IGF axis in pancreas, the abundance of mRNAfor IGF-II was determined in whole pancreata using North-ern blot hybridization. Three mRNA transcripts for IGF-IIwere detected of 7.5 kb, 4.4 kb, and 2.4 kb in pancreata of bothcontrol and LP fed rats. In control fed animals the expressionof all IGF-II mRNA transcripts declined postnatally and wasbarely detectable after postnatal day 14 (Fig. 4). In LP fed rats,the expression IGF-II was reduced compared with controlsat all ages. IGF-II mRNA expression was expressed relativeto that of 18S ribosomal RNA in the same samples for threeadditional, separate hybridizations using pancreata from dif-ferent animals and is shown in Fig. 5, A and B, for the twolarger mRNA transcripts. This confirmed that the expressionof IGF-II mRNA was reduced in the pancreata of LP fed rats,this being most pronounced for the 7.4 kb transcript. Todetermine whether the sites of expression of IGF-II in pan-creas were altered in the neonatal LP fed animals, mRNA wasvisualized by in situ hybridization. IGF-II mRNA was locatedpredominently in islet cells, and at postnatal day 2 was lessabundant in pancreas sections from LP compared with con-trol-fed rats (Fig. 6, A and B). IGF-I mRNA was barely de-tectable using in situ hybridization in the islets of fetal orneonatal rats. By postnatal day 21 IGF-I mRNA was located
within acinar cells, but did not obviously differ betweendietary groups (not shown).
IGF-II immunoreactivity was visualized within pancreataby immunohistochemistry. IGF-II was abundant in islet cellsin the fetus (Fig. 6, D and E), but declined in intensity and inthe number of immunopositive cells postnatally, to becomebarely detectable by postnatal day 21 (Fig. 5C). Rats fed LPdiet showed a significantly reduced area of islets containingimmunoreactive IGF-II compared with control-fed animals.Immunoreactivity for IGF-I was low in the fetal pancreas(day 21.5 gestation, control 8.6 6 0.4% islet cell area, n 5 12)but appeared postnatally in association with islet cells andsome acinar epithelial cells (day 21, 13.7 6 0.2% islet cell area,n 5 20). The area of islets immunopositive for IGF-I did notsignificantly differ between control and LP animals at anyage. The presence of IGFBPs in the pancreas was also inves-tigated using immunohistochemistry. ImmunoreactiveIGFBP-1 was diffusely present in islet cells in the fetus, andabundance declined with age (not shown). IGFBP-2 immu-noreactivity was predominantly associated with the outermantle of islet cells, comprising mostly a cells, and vascularendothelium and did not change greatly with age. Immu-noreactivity for IGFBP-3 and -4 was seen at all ages withinthe pancreas, within islets and mesenchymal tissue, but notin most acinar cells, whereas IGFBP-5 was seen at all agesassociated with the a cells of the islets and with ductal ep-ithelial cells. No differences were found between the area ofislets immunopositive for any IGFBP at any age betweencontrol-fed and LP-fed rats.
Discussion
The experimental induction of IUGR in the rat by a varietyof methods results in alterations in the endocrine pancreas(5), reduced pancreatic weight and b cell mass at birth (36–38), and lower insulin secretion in later life (6, 8). This studyshows that LP diet given during fetal and early postnatal lifeinduced not only growth retardation, but disturbed the bal-ance between b cell proliferation and b cell death. Amino
TABLE 2. Islet cell area (%) occupied by cells immunoreactive for insulin, glucagon or somatostatin for fetal (F) or neonatal rats frommothers fed control (C) or low protein (LP) diet
Age (days) Insulin (I) Glucagon (G) Somatostatin I/G ratio
F19.5 C 78.5 6 0.3 15.1 6 0.2 2.7 6 0.1 5.2 6 0.1LP 64.3 6 1.1a 20.7 6 0.4a 2.8 6 0.1 3.1 6 0.1a
F21.5 C 79.6 6 0.7 11.9 6 0.3 ND 6.8 6 0.2LP 70.5 6 1.1a 17.2 6 1.1a ND 4.1 6 0.1a
2 C 78.6 6 0.9 14.3 6 0.4 3.6 6 0.1 5.6 6 0.2LP 70.3 6 0.5a 17.4 6 0.4a 4.5 6 0.1a 4.1 6 0.1a
6 C 78.5 6 0.8 15.1 6 0.2 2.1 6 0.1 5.2 6 0.1L 68.0 6 1.4a 21.7 6 0.3a 2.0 6 0.1 3.1 6 0.1a
10 C 79.8 6 0.8 13.6 6 0.2 2.6 6 0.1 5.9 6 0.1L 71.3 6 0.6a 19.1 6 0.5a 1.8 6 0.1a 3.8 6 0.1a
12 C 79.4 6 0.7 14.8 6 0.3 1.9 6 0.1 5.4 6 0.1L 69.9 6 0.9a 19.8 6 0.5a 1.9 6 0.1 3.6 6 0.1a
14 C 78.4 6 1.1 15.9 6 0.3 3.3 6 1.2 5.0 6 0.1L 69.3 6 0.6a 19.0 6 0.3a ND 3.7 6 0.1a
17 C 79.4 6 0.9 14.3 6 0.3 1.9 6 0.1 5.6 6 0.1L 71.2 6 0.8a 18.0 6 0.3a 3.2 6 0.1a 4.0 6 0.1a
21 C 77.8 6 0.8 15.2 6 0.3 2.6 6 0.1 5.1 6 0.1L 68.9 6 2.9a 17.9 6 0.3a 3.3 6 0.1b 3.9 6 0.1a
Figures represent mean values 6 SEM. (n 5 15–20).a P , 0.001, b P , 0.05 vs. control diet. ND, Not determined.
4866 PANCREATIC IGF AXIS AND LOW PROTEIN DIET Endo • 1999Vol 140 • No 10

FIG. 2. Immunohistochemistry for BrdU(A and B), cyclin D1 (C and D), NEK2 (Eand F), and apoptosis (G and H) in sec-tions of rat pancreas from animals of 14days postnatal age. Animals were main-tained on control diet (A, C, E, and G) orlow protein diet (B, D, F, and H). In Aand B duel staining was performed forinsulin (blue) and BrdU (brown). I, Is-lets; e, exocrine tissue. Arrows indicateimmunoreactivity associated with isletcells. Magnification bar, 10 mm.
PANCREATIC IGF AXIS AND LOW PROTEIN DIET 4867

acids are more potent in the stimulation of fetal islet cellproliferation than is glucose, are more potent insulin secre-togogues (39), and their profiles are altered in the maternalcirculation following the administration of LP diet (4). Areduction in pancreatic weight was not apparent until afterbirth in LP animals, although the mean islet area was sig-nificantly reduced from at least 19.5 days gestation. Theprimary impact of protein restriction is on the proliferationof existing b cells, which differs from observations in pupsfrom maternal caloric restriction where neogenesis is af-fected, whereas proliferation rate was normal (36). We havenot measured islet cell neogenesis in the LP diet model, butone cannot exclude a contribution to the reduced islet cell
mass since when a normal diet is replaced immediately fol-lowing birth, the islets in adult life are larger than in controls,whereas the total volume density of endocrine tissue is de-creased (7, 8). This suggests that the number of islets may bereduced. In the present study b cell mass per pancreas wassubstantially reduced by LP diet, due to islets of smallermean volume. This was due to a relative deficiency of a, b,and somatostatin-secreting cells, although the b cell popu-lation was most severely effected. If nutritional restriction isreversed at birth or maintained until adulthood the isletmorphology remains unaltered (8). In both situations lowerinsulin secretion is apparent in adult life, although this ismore severe if LP diet is continued to adulthood. This sug-
FIG. 3. Percentage of islet cells(mean 6 SEM) demonstrating immuno-reactivity for proliferating cell nuclearantigen (PCNA) (A), cyclin D1 (B),NEK2 (C) or apoptotic nuclei (D) in pan-creata from fetal (F) or postnatal ani-mals following feeding of control (openbars) or low protein (LP, shaded bars)diets. Figures are derived from 12–20observations for each age using three tofive animals. *, P , 0.01 or better vs.control diet.
4868 PANCREATIC IGF AXIS AND LOW PROTEIN DIET Endo • 1999Vol 140 • No 10

gests that LP diet causes a fundamental reprogramming of bcell phenotype with critical periods both in fetal and neonatallife. Our results suggest some possible mechanisms as to howthis may occur.
We showed previously that feeding an LP diet throughoutgestation resulted in a lower nuclear labeling index with [3H]thymidine in islet cells of the newborn (5), findings repro-duced here with BrdU. Labeling with BrdU in b cells withinislets from control-fed animals was consistent with the frac-tional contribution of b cells to these islets, demonstrating anequivalent labeling index for b and nonb cell types. In an-imals fed LP diet significantly less labeling with BrdU waslocated in b cells, consistent with their reduced fractionalarea, and suggesting that the reduction in islet size is pref-erentially linked to a deficit in b cell proliferation. The frac-
tion of cells containing immunoreactivity for PCNA, eitherin all islet cells or specifically in the b cell population was,paradoxically, increased at every age up to postnatal day 17in LP animals. PCNA is an auxiliary protein of DNA poly-merase that begins to accumulate in the nucleus during G1of the cell cycle, is most abundant in S phase and declinesduring G2/M (40). The observation that more b cells con-tained detectable PCNA following LP diet, despite less cellsbeing observed in S phase by incorporation of BrdU, couldbe explained by an altered cell cycle kinetics in which eitherG1 or G2/M phases were extended in LP animals. Alterna-tively, because PCNA is also expressed during DNA repair(41), it is possible that islets from LP-fed rats were subject tooxidative, or some other physiological stress. To distinguishbetween these alternatives we further examined immunore-
FIG. 3. Continued.
PANCREATIC IGF AXIS AND LOW PROTEIN DIET 4869

activity for cyclin D1 and for NEK2. Cyclin D1 is a nuclearprotein that accumulates in G1 and is necessary for transitionfrom G1 to S phase of the cell cycle (42). It associates withPCNA and cyclin-dependent kinase 2 (43). NEK2 is a serine/threonine kinase that accumulates at the G2 to M phasetransition and is necessary for progression into cell mitosis
(44). The fractional increase of b cells in LP rats containingimmunoreactive cyclin D1, and a decrease of NEK2 stainingwould be consistent with a lengthened mean cell cycle du-ration in b cells within islets from LP rats, in which the G1phase of the cycle was extended. However, direct confirma-tion of this would require precise cell cycle kinetic analysisusing isolated b cells from these animals.
A modulation of mean islet area by diet might involve achange in the rate of cell attrition as well as altered prolif-erative rate. A low rate of islet cell apoptosis exists in the fetalrat islet, but this transiently increases at 14 to 17 days post-natal life, to return to minimal levels by weaning (12, 14). Therate of islet cell apoptosis was increased at every age exam-ined in LP rats between late fetal life and weaning, comparedwith control-fed animals. However, the timing of the neo-natal wave of apoptosis was not altered, suggesting that thisevent is qualitatively independent of diet of the mother. It isnot possible to directly show that the apoptozing islet cellsare b cells because they no longer contain immunoreactiveinsulin. As we have described previously (14), this may bedue to a loss of cytoplasm rendering any immunohistochem-ical signal difficult to visualize. However, their predomi-nantly central location within the islets is consistent with themajority having been b cells. In addition, we have previouslydemonstrated that the majority of apoptotic islet cells expressthe glucose transporter protein GLUT2 on the plasma mem-brane, suggesting that they are derived from b cells (45). Thepercent of islet cells undergoing apoptosis in late fetal andearly neonatal life is lower than we reported in a previousstudy of ontogeny, although the ontological pattern is iden-tical (14). This may be due to the use of a separate, inbred ratcolony in the present study. We have shown previously thatthe neonatal wave of islet cell apoptosis is preceded by atransient increase in the presence of iNOS within b cells (14),which may be functionally linked to the developmental ap-optosis. While a similar rise in islet cell iNOS immunoreac-tivity was seen in the LP animals, the percentage of immu-nopositive islet cells did not differ between control and LPdiets. This suggests that the mechanisms underlying an in-creased rate of islet cell apoptosis seen in LP animals areunlikely to be due to increased detectable iNOS.
The transient increase in islet cell apoptosis in the neonatal
FIG. 4. Northern blot hybridization ofmRNAs for IGF-II for total RNA ex-tracted from whole rat pancreata for an-imals of 19.5 or 21.5 days fetal (F) age,or postnatal days 6 to 21 following feed-ing of control (20) or low protein (8) di-ets. Three major mRNA transcriptswere seen of 7.4, 4.4, and 2.4 kb. Thehybridization signal for IGF-II was re-duced at each age for animals given lowprotein diet relative to normal diet. Thesame filter was rehybridized with acDNA for 18S rRNA to estimate the ef-ficiency of loading and transfer of totalRNA. The filter was subjected to alonger autoradiographical expose forsamples from postnatal days 14–21 toamplify the weaker hybridization sig-nals at these ages. Molecular size mark-ers are shown to the left.
FIG. 5. Quantification of messenger RNA for IGF-II (mean 6 SEM)from Northern blot hybridization, expressed as a ratio of the hybrid-ization signals for the 7.4 kb (A), or 4.4 kb (B) mRNA transcripts,relative to the signal for 18S rRNA in the same sample, in pancreatafrom fetal (F) or postnatal animals following feeding of control (openbars) or low protein (LP, shaded bars) diets. Figures are derived fromthree separate hybridizations for each age, each using tissues fromdifferent animals. *, P , 0.05 or better vs. control diet. C, Percentageof islet cells demonstrating immunoreactivity for IGF-II. Figures arederived from 14–20 observations for each age using five animals. *,P , 0.001 vs. control diet.
4870 PANCREATIC IGF AXIS AND LOW PROTEIN DIET Endo • 1999Vol 140 • No 10

rat is temporally related to a rapid loss of expression ofIGF-II mRNA from the pancreas, where it is predomi-nantly expressed within islet cells (14, 17), and we haveshown that endogenous IGF-II acts as a survival factor thatprevents apoptosis in isolated islets from 5-day old rats(14). While an increasing expression of IGF-I mRNA occursin the pancreas postnatally, this is located mainly in theacinar tissue, not in the islets of Langerhans (14). In thepancreata from LP rats, a reduced expression of IGF-IImRNA was found at fetal and neonatal ages, before thedevelopmentally associated decline in IGF-II expressionthat occurs from day 14. It is not known if these changesreflect an effect of diet on gene transcription or on thestability of the mRNAs. For animals receiving both controland LP diet, the major sites of IGF-II expression in thepancreas, assessed by in situ hybridization, were the islets.The number of islet cells that were immunoreactive forIGF-II was also significantly reduced in LP animals, con-firming that differences are also likely to be present at thelevel of the translated protein. These findings are consis-tent with an action of endogenous IGF-II as a survivalfactor for pancreatic b cells, and suggest that the increasedapoptosis seen in islet cells in LP rats may be functionallylinked to a reduced expression of IGF-II. Because IGF-IIwill also increase islet cell DNA synthesis (19, 46), itsrelative paucity in the islets of LP rats may also contribute
directly to reduced b cell proliferation rate and the re-duction in islet size. It has been recently proposed thatamino acids, and in particular branched-chain amino ac-ids, may promote b cell proliferation by stimulating phos-phorylation of PHAS-1 (a regulator of translation initia-tion during mitogenesis) and by facilitating theproliferative effects mediated by growth factors (47).Branched-chain amino acids are most severely depleted inthe plasma of fetuses from LP-fed mothers (4), which couldalso contribute to the lower rate of b cell proliferation inthe pancreas of the LP pups.
An effect of LP diet on pancreatic IGF-II expression maybe direct. We showed previously that the release of immu-noreactive IGF-II from isolated fetal rat islets was potentiatedby amino acids, but not by glucose, whereas the release ofinsulin was increased by both (19). Circulating levels ofIGF-II in the rat fetus, which derive predominantly from anhepatic expression, are not substantially decreased duringmodels of IUGR, although the much lower levels of circu-lating IGF-I are depressed (29, 30). This would suggest thatthe development of the pancreatic islets is more sensitive tochanges in locally expressed IGF-II than to circulatorychanges, and that a paracrine or autocrine action of IGF-II isdominant in this tissue. No significant changes were seen inthe fractional area of islets demonstrating immunoreactivityfor IGF-I or IGFBPs in LP-fed animals. We have shown that
FIG. 6. In situ hybridization to visualize IGF-II mRNA (A–C), and the immunohistochemical localization of IGF-II (D and E) in tissue sectionsof rat pancreas from animals of 21.5 days gestation. Animals were maintained on control diet (A, C, D) or low protein diet (B and E). C, Controlhybridization with a sense strand cRNA. I, Islets; e, exocrine tissue. Arrows indicate either mRNA hybridization signal or immunoreactivityassociated with islet cells. Magnification bar, 10 mm.
PANCREATIC IGF AXIS AND LOW PROTEIN DIET 4871

increased concentrations of amino acids will increaseIGFBP-1, -2, and -3 release from isolated fetal rat islets (19),while the release of IGFBP-1 and -2 from islets is glucose-responsive in the fetal rat or adult hamster (19, 32). Miceexpressing the human IGFBP-1 transgene have an increasedpancreatic islet size with relatively more b cells (48). It ispossible that any changes in IGFBP abundance within theislets of LP rats were too subtle to be detected by immuno-histochemistry alone, and an influence of diet on IGFBPsynthesis and secretion from islet cells is still possible.
In summary, the experiments suggest that LP diet causesa selective deficiency of b cells in the fetus and neonate dueto the combined effects of a reduced b cell proliferation rate,which may involve an altered cell cycle length, and an in-creased incidence of apoptosis. These changes are associatedwith a decreased expression of IGF-II within the pancreaticislets, a growth factor known to act as a b cell mitogen andto prevent apoptosis. It is therefore possible that perturba-tions of prenatal or neonatal nutrition and growth will leadto inappropriate b cell ontogeny in the preweaning period,and result in a population of b cells that may subsequentlybe functionally compromised in later life, contributing toglucose intolerance.
Acknowledgments
We are grateful to Ms. Brenda Strutt for assistance with in situ hy-bridization and Ms. Marie Therese Ahn for skillful technical assistance.
References
1. Hoet JJ, Dahri S, Reusens B, Remacle C 1995 Do NIDDM and cardiovasculardisease originate in utero? In: Baba S, Kenako T (eds) Diabetes 1994. ElsevierScience, Amsterdam, pp 62–71
2. Barker DJP 1997 Fetal undernutrition and adult disease. Endocrinol Metab 4[Suppl B]:39–46
3. Hales CN, Barker DJP 1992 Type 2 (non-insulin-dependent) diabetes mellitus:the thrifty phenotype hypothesis. Diabetologia 35:595–601
4. Reusens B, Dahri S, Snoeck A, Bennis-Taleb N, Remacle C, Hoet JJ 1995Long-term consequences of diabetes and its complications may have a fetalorigin: experimental and epidemiologial evidence. In: Cowett RM (ed) Dia-betes. Nestle Nutrition Workshop Series, Raven Press, New York, vol35:187–198
5. Snoeck A, Remacle C, Reusens B, Hoet JJ 1990 Effect of a low protein dietduring pregnancy on the fetal rat endocrine pancreas. Biol Neonate 57:107–118
6. Dahri S, Cherif H, Reusens B, Remacle C, Hoet JJ 1994 Effect of an isocaloriclow protein diet during gestation in rat on the in vitro insulin secretion by isletsof the offspring. Diabetologia [Suppl] 37:80
7. Dahri S, Snoeck A, Reusens-Billen B, Remacle C, Hoet JJ 1991 Islet functionin offspring of mothers on low protein diet during gestation. Diabetes [Suppl2] 40:115–120
8. Dahri S, Reusens B, Remacle C, Hoet JJ 1995 Nutritional influences on pan-creatic development and potential links with non-insulin-dependent diabetes.Proc Nutr Soc 54:345–356
9. Hill DJ, Hogg J 1991 Growth factor control of pancreatic b cell hyperplasia.In: Herington A (ed) Clinical Endocrinology and Metabolism. Bailliere Tindall,London, pp 689–698
10. Kaung HL 1994 Growth dynamics of pancreatic islet cell populations duringfetal and neonatal development of the rat. Dev Dyn 200:163–175
11. Finegood DT, Scaglia L, Bonner-Weir S 1995 Dynamics of b cell mass in thegrowing rat pancreas. Diabetes 44:249–256
12. Scaglia L, Cahill CJ, Finegood DT, Bonner-Weir S 1997 Apoptosis participatesin the remodeling of the endocrine pancreas in the neonatal rat. Endocrinology138:1736–1741
13. Tornehave D, Larsson L-I 1997 Presence of Bcl-Xl during development of thehuman fetal and rat neonatal endocrine pancreas: correlation to programmedcell death. Exp Clin Endocrinol Diabetes 105:A27
14. Petrik J, Arany E, McDonald TJ, Hill DJ 1998 Apoptosis in the pancreaticislet cells of the neonatal rat is associated with a reduced expression ofinsulin-like growth factor II that may act as a survival factor. Endocrinology139:2994 –3004
15. Jung Y, Miura M, Yuan J 1996 Suppression of IL-1 b converting enzyme-mediated cell death by insulin-like growth factor. J Biol Chem 271:5112–5117
16. Geier A, Haimshon M, Beery R, Lunenfeld B 1992 Insulin-like growth factor-Iinhibits cell death induced by cycloheximide in MCF-7 cells—a model systemfor analyzing control of cell death. In Vitro Cell Dev Biol-Animal 28A:725–729
17. Hogg J, Hill DJ, Han VKM 1994 The ontogeny of insulin-like growth factor(IGF) and IGF binding protein gene expression in the rat pancreas. J MolEndocrinol 13:49–58
18. Hill DJ, Frazer A, Swenne I, Wirdnam PK, Milner RDG 1987 Somatomedin-Cin the human fetal pancreas: cellular localization and release during organculture. Diabetes 36:465–471
19. Hogg J, Han VKM, Clemmons DR, Hill DJ 1993 Interactions of glucose,insulin-like growth factors (IGFs) and IGF binding proteins in the regulationof DNA synthesis by isolated fetal rat islets of Langerhans. J Endocrinol138:401–412
20. Scharfmann R, Corvol M, Czernichow P 1989 Characterization of IGF I pro-duced by fetal rat pancreatic cells. Diabetes 38:686–690
21. Swenne I, Hill DJ, Strain AJ, Milner RDG 1987 Growth hormone regulationof somatomedin-C/insulin-like growth factor I production and DNA repli-cation in fetal rat islets in tissue culture. Diabetes 36:288–294
22. Van Schravendijk CFH, Heylen L, Van Den Brande JL, Pipeleers DG 1990Direct effect of insulin and insulin-like growth factor-I on the secretory activityof rat pancreatic b cells. Diabetologia 33:649–653
23. Hill DJ, Sedran RJ, Brenner SL, McDonald TJ 1997 Insulin-like growthfactor-I (IGF-I) has a dual effect on insulin release from isolated, perifusedadult rat islets of Langerhans. J Endocrinol 153:15–25
24. Thissen JP, Ketelslegers JM, Underwood LE 1994 Nutritional regulation of theinsulin-like growth factors. Endocr Rev 15:80–101
25. Estivariz CF, Zeigler TR 1997 Nutrition and the insulin-like growth factorsystem. Endocrine 7:65–71
26. Lassarre C, Hardouin S, Daffos F, Forestier F, Frankenne F, Binoux M 1991Serum insulin-like growth factor binding proteins in the human fetus. Rela-tionships with growth in normal subjects and in subjects with intrauterinegrowth retardation. Pediatr Res 29:219–226
27. Leger J, Oury JF, Noel M, Baron S, Benali K, Blot P, Czernichow P 1996Growth factors and intrauterine growth retardation. I. Serum growth hor-mone, insulin-like growth factor (IGF)-I, IGF-II, and IGF binding protein 3levels in normally grown and growth-retarded human fetuses during thesecond half of gestation. Pediatr Res 40:94–100
28. De Prins FA, Hill DJ, Fekete M, Robsen DJ, Fieller NRJ, Van Assche FA,Milner RDG 1984 Reduced plasma somatomedin activity and costal cartilagesulfate incorporation activity during experimental growth retardation in thefetal rat. Pediatr Res 18:1100–1104
29. Straus DS, Ooi GT, Orlowski CC, Rechler MM 1991 Expression of the genesfor insulin-like growth factor-I (IGF-I), IGF-II, and IGF-binding proteins-1 and-2 in fetal rat under conditions of intrauterine growth retardation caused bymaternal fasting. Endocrinology 128:518–525
30. Woodall SM, Breier BH, Johnston BM, Gluckman PD 1996 A model ofintrauterine growth retardation caused by chronic maternal undernutrition inthe rat: effects on the somatotrophic axis and postnatal growth. J Endocrinol150:231–242
31. Muaku SM, Underwood LE, Selvais PL, Ketelslegers JM, Maiter D 1995Maternal protein restriction early or late in rat pregnancy has differentialeffects on fetal growth, plasma insulin-like growth factor-I (IGF-I) and liverIGF-I expression. Growth Regul 5:125–132
32. Massa L, Cortizo AM, Gagliardino JJ 1997 Insulin-like growth factor bindingproteins from adult-hamster pancreatic islets: influence of glucose concentra-tion. Diabetes Metab 23:417–423
33. Murphy LJ, Tachibana K, Friesen HG 1988 Stimulation of hepatic insulin-likegrowth factor-I gene expression by ovine prolactin: evidence for intrinsicsomatogenic activity in the rat. Endocrinology 122:2027–2033
34. Hsu SM, Raine L, Fanger H 1981 Use of avidin-biotin peroxidase complex(ABC) in immunoperoxidase techniques: a comparison between ABC andunlabelled antibody (PAP) procedures. J Histochem Cytochem 29:577–580
35. Wijsman JH, Jonker RR, Keijzer R, Van de Velde CJ, Cornelisse CJ, VanDierendonck JH 1993 A new method to detect apoptosis in paraffin sections:in situ end labelling of fragmented DNA. J Histochem Cytochem 41:7–12
36. Garofano A, Czernichow P, Breant B 1997 In utero undernutrition impairs ratbeta-cell development. Diabetologia 40:1231–1234
37. Portha B, Kergoat M, Blondel O, Bailbe D 1995 Underfeeding of rat mothersduring the first two trimesters of gestation does not alter insulin action andinsulin secretion in the progeny. Eur J Endocrinol 133:475–482
38. Alvarez C, Martin MA, Goya L, Bertin E, Portha B, Pascual-Leone AM 1997Contrasted impact of maternal rat food restriction on the fetal endocrinepancreas. Endocrinology 138:2267–2273
39. Swenne I 1985 Glucose-stimulated DNA replication of the pancreatic isletsduring the development of the rat fetus. Effects of nutrients, growth hormoneand triiodothyronine. Diabetes 34:803–807
40. Prosperi E, Scovassi AI, Stivala LA, Bianhi L 1994 Proliferating cell nuclearantigen bound to DNA synthesis sites: phosphorylation ans association withcyclin D1 and cyclin A. Exp Cell Res 215:257–262
41. Prosperi E 1997 Multiple roles of the proliferating cell nulear antigen: DNAreplication, repair and cell cycle control. Prog Cell Cycle Res 3:193–210
4872 PANCREATIC IGF AXIS AND LOW PROTEIN DIET Endo • 1999Vol 140 • No 10

42. Baldin V, Lucas J, Marcote MJ, Pagano M, Draetta G 1993 Cyclin D1 is anuclear protein required for cell cycle progression in G1. Genes Dev 7:812–821
43. Bartkova J, Lucas J, Strauss M, Bartek J 1994 Cell cycle-related variation andrestricted-restricted expression of human cyclin D1 protein. J Pathol172:237–245
44. Fry AM, Nigg EA 1995 Cell cycle. The NIMA kinase joins forces with cdc2. CurrBiol 5:1122–1125
45. Hill DJ, Petrik J, Arany E, McDonald TJ, Delovitch TL 1999 Insulin-likegrowth factors prevent cytokine-mediated cell death in isolated islets of Lang-erhans from pre-diabetic NOD mice. J Endocrinol 161:153–165
46. Rabinovitch A, Quigley C, Russel T, Patel Y, Mintz DH 1982 Insulin andmultiplication stimulating activity (an insulin-like growth factor) stimulateislet b-cell replication in neonatal rat pancreatic monolayer culture. Diabetes31:160–164
47. Xu G, Kwon G, Marshall CA, Lin TA, Lawrence JC 1998 Branched-chainamino acids are essential in the regulation of PHAS-1 and p70 S6 kinase bypancreatic b cells. J Biol Chem 273:28178–28184
48. Rajkumar K, Dheen ST, Murphy LJ 1996 Hyperglycaemia and impairedglucose tolerance in insulin-like growth factor binding protein-1 transgenicmice. Am J Physiol 270:E565–E571
PANCREATIC IGF AXIS AND LOW PROTEIN DIET 4873
Copyright © 2022 FDOKUMEN




















