
Generation of Human Islets Through Expansion and Differentiation of Non-islet Pancreatic Cells...
9
Generation of Human Islets Through Expansion and Differentiation of Non-islet Pancreatic Cells Discarded (Pancreatic Discard) After Islet Isolation Ivan Todorov, PhD, Keiko Omori, MD, Michael Pascual, BS, Jeffery Rawson, BS, Indu Nair, PhD, Luis Valiente, BS, Tommy Vuong, BS, Takeru Matsuda, MD, PhD, Chris Orr, BS, Kevin Ferreri, PhD, Craig V. Smith, MD, Fouad Kandeel, MD, PhD, and Yoko Mullen, MD, PhD Objectives: Islet transplantation is hampered by the shortage of donor tissues. Our objective was to generate islet-like cell clusters (ICCs) from cultures of non-islet pancreatic cells. Methods: The starting cultured cells came from the non-islet fractions of human pancreases after enzymatic digestion and purification for the purpose of islet isolation. Initially, these cells expanded in monolayer cultures and became confluent on collagen-coated flasks. After trypsination and suspension of these cells in a defined islet differentiation medium, the cells aggregated to form ICCs. Results: The initial cell population consisted of less than 1% of insulin-positive cells, 44% amylase-positive cells, and 41% cytoke- ratin (CK) 7Ypositive, or CK19 + cells, but PDX-1 + cells were absent. Cells from later stages of the monolayer cultures showed signs of dedifferentiation/transdifferentiation. At the time of har- vesting, more than 90% of the cells were positive for CK 7/19 and PDX-1, but less than 1% of the cells were insulin-positive. After aggregation, the ICCs appeared redifferentiated, and contained glucose-responsive, insulin-secreting cells with an insulin content measuring 20% of that found in freshly isolated islets isolated from the same pancreas. ICCs transplanted into athymic mice and re- moved after 4 months did acquire the morphology of mature islets, indicating further maturation of the ICCs in vivo after transplan- tation. Human C-peptide was detected in recipient animal sera. Conclusion: Using the specified culture methods, non-islet pancreas cells can generate cell clusters resembling islets. These ICCs, ob- tained from fractions of the pancreas that are otherwise discarded, continue to differentiate after transplantation to become mature islets. Key Words: ex vivo human islet generation, islet precursors, non-islet pancreatic cell differentiation (Pancreas 2006;32:130Y138) R ecent advances in clinical islet transplantation 1 have led to the expectation that $-cell replacement therapy may provide a possible cure for type 1 diabetes. However, a major hurdle is the limited availability of donor pancreata. Various lines of investigation have been conducted to alleviate the islet shortage as discussed in a number of reviews. 2Y6 The use of xenogeneic islets, especially from swine, has been studied for a number of years, but the unresolved problems of the xenogeneic immune barrier and the risks of xenoinfections have remained as major challenges. 7 Insulin-producing cells have been generated from embryonic stem cells, 8,9 but in addition to the controversies 10,11 and ethical issues, a teratoma formation was recently reported after transplanta- tion of embryonal stem cells (ES)-generated ICCs. 12 Adult stem cells, including bone marrow cells, and neural stem cells were also reported as a source to generate islets, 13 but this has yet to be clearly demonstrated. 5 Transdifferentiation of hepatocytes 14 or epithelial cells in the intestine 15 has also resulted in insulin-producing cell generation in vitro and may be potential sources of islets. Others have attempted to bio- engineer $ cells in the generation of artificial islets. Despite many reports, only a few have convincingly shown the reversal of diabetes by islets generated by these approaches. 14,16,17 The human pancreas from healthy individuals contains approximately 1 million islets, which represent approximately 1% to 2% of the total pancreatic mass, and the remaining 98% of the cells consists mainly of acinar and ductal cells. In the preparation of islets for transplantation, islets are purified or enriched by a density gradient centrifugation. The remaining non-islet fractions are usually discarded (the pancreatic discard). Studies from several laboratories 18Y20 have suggested the potential of non-islet cells of the human pancreas to differentiate into insulin-producing cells. Islet-like cell clusters (ICCs) have also been generated in vitro from pancreatic cells using an extracellular matrix (Matrigel). 18,20,21 However, insulin expression levels in the resulting $ cells were low. Furthermore, the use of Matrigel may prevent this approach from clinical application. ICCs have been generated by culturing ductal epithelium of the pancreas from prediabetic non-obese diabetic (NOD) mice. 19 When transplanted into diabetic NOD mice, these ICCs were fully developed and able to reverse hyperglycemia. Others have speculated that non-islet pancreatic cells, such as acinar cells, dedifferentiate or transdifferentiate into ductlike cells or ORIGINAL ARTICLE 130 Pancreas & Volume 32, Number 2, March 2006 Received for publication July 21, 2005; accepted October 21, 2005. From the Southern California Islet Cell Resources Center, Department of Diabetes, Endocrinology and Metabolism, City of Hope National Medi- cal Center/Beckman Research Institute, Duarte, CA. This work was supported by a grant from the Nora Eccles Treadwell Foundation and the ICR. Reprints: Ivan Todorov, PhD, SC-ICR Center/DEM, City of Hope National Medical Center/Beckman Research Institute, 1500 E. Duarte Road, Duarte, CA 91010 (e-mail: [email protected]). Copyright * 2006 by Lippincott Williams & Wilkins Copyr ight © Lippincott Williams & Wilkins. Unauthor iz ed reproduction of this article is prohibited.
Transcript of Generation of Human Islets Through Expansion and Differentiation of Non-islet Pancreatic Cells...

Generation of Human Islets Through Expansion andDifferentiation of Non-islet Pancreatic Cells Discarded
(Pancreatic Discard) After Islet IsolationIvan Todorov, PhD, Keiko Omori, MD, Michael Pascual, BS, Jeffery Rawson, BS, Indu Nair, PhD,
Luis Valiente, BS, Tommy Vuong, BS, Takeru Matsuda, MD, PhD, Chris Orr, BS, Kevin Ferreri, PhD,
Craig V. Smith, MD, Fouad Kandeel, MD, PhD, and Yoko Mullen, MD, PhD
Objectives: Islet transplantation is hampered by the shortage of
donor tissues. Our objective was to generate islet-like cell clusters
(ICCs) from cultures of non-islet pancreatic cells.
Methods: The starting cultured cells came from the non-isletfractions of human pancreases after enzymatic digestion andpurification for the purpose of islet isolation. Initially, thesecells expanded in monolayer cultures and became confluenton collagen-coated flasks. After trypsination and suspensionof these cells in a defined islet differentiation medium, thecells aggregated to form ICCs.Results: The initial cell population consisted of less than 1% of
insulin-positive cells, 44% amylase-positive cells, and 41% cytoke-
ratin (CK) 7Ypositive, or CK19+ cells, but PDX-1+ cells were
absent. Cells from later stages of the monolayer cultures showed
signs of dedifferentiation/transdifferentiation. At the time of har-
vesting, more than 90% of the cells were positive for CK 7/19 and
PDX-1, but less than 1% of the cells were insulin-positive. After
aggregation, the ICCs appeared redifferentiated, and contained
glucose-responsive, insulin-secreting cells with an insulin content
measuring 20% of that found in freshly isolated islets isolated from
the same pancreas. ICCs transplanted into athymic mice and re-
moved after 4 months did acquire the morphology of mature islets,
indicating further maturation of the ICCs in vivo after transplan-
tation. Human C-peptide was detected in recipient animal sera.
Conclusion: Using the specified culture methods, non-islet pancreas
cells can generate cell clusters resembling islets. These ICCs, ob-
tained from fractions of the pancreas that are otherwise discarded,
continue to differentiate after transplantation to become mature
islets.
Key Words: ex vivo human islet generation, islet precursors,
non-islet pancreatic cell differentiation
(Pancreas 2006;32:130Y138)
R ecent advances in clinical islet transplantation1 have ledto the expectation that $-cell replacement therapy may
provide a possible cure for type 1 diabetes. However, a majorhurdle is the limited availability of donor pancreata. Variouslines of investigation have been conducted to alleviate theislet shortage as discussed in a number of reviews.2Y6 The useof xenogeneic islets, especially from swine, has been studiedfor a number of years, but the unresolved problems of thexenogeneic immune barrier and the risks of xenoinfectionshave remained as major challenges.7 Insulin-producing cellshave been generated from embryonic stem cells,8,9 but inaddition to the controversies10,11 and ethical issues, ateratoma formation was recently reported after transplanta-tion of embryonal stem cells (ES)-generated ICCs.12 Adultstem cells, including bone marrow cells, and neural stem cellswere also reported as a source to generate islets,13 but this hasyet to be clearly demonstrated.5 Transdifferentiation ofhepatocytes14 or epithelial cells in the intestine15 has alsoresulted in insulin-producing cell generation in vitro and maybe potential sources of islets. Others have attempted to bio-engineer $ cells in the generation of artificial islets. Despitemany reports, only a few have convincingly shown the reversalof diabetes by islets generated by these approaches.14,16,17
The human pancreas from healthy individuals containsapproximately 1 million islets, which represent approximately1% to 2% of the total pancreatic mass, and the remaining98% of the cells consists mainly of acinar and ductal cells.In the preparation of islets for transplantation, islets arepurified or enriched by a density gradient centrifugation. Theremaining non-islet fractions are usually discarded (thepancreatic discard). Studies from several laboratories18Y20
have suggested the potential of non-islet cells of thehuman pancreas to differentiate into insulin-producingcells. Islet-like cell clusters (ICCs) have also been generatedin vitro from pancreatic cells using an extracellular matrix(Matrigel).18,20,21 However, insulin expression levels inthe resulting $ cells were low. Furthermore, the use ofMatrigel may prevent this approach from clinical application.ICCs have been generated by culturing ductal epithelium of thepancreas from prediabetic non-obese diabetic (NOD) mice.19
When transplanted into diabetic NOD mice, these ICCs werefully developed and able to reverse hyperglycemia. Othershave speculated that non-islet pancreatic cells, such as acinarcells, dedifferentiate or transdifferentiate into ductlike cells or
ORIGINAL ARTICLE
130 Pancreas & Volume 32, Number 2, March 2006
Received for publication July 21, 2005; accepted October 21, 2005.From the Southern California Islet Cell Resources Center, Department of
Diabetes, Endocrinology and Metabolism, City of Hope National Medi-cal Center/Beckman Research Institute, Duarte, CA.
This work was supported by a grant from the Nora Eccles TreadwellFoundation and the ICR.
Reprints: Ivan Todorov, PhD, SC-ICR Center/DEM, City of Hope NationalMedical Center/Beckman Research Institute, 1500 E. Duarte Road,Duarte, CA 91010 (e-mail: [email protected]).
Copyright * 2006 by Lippincott Williams & Wilkins
Copyr ight © Lippincott Williams & Wilkins. Unauthorized reproduction of this article is prohibited.

even hormone-positive cells.22Y24 The presence of stem cellsin the pancreas has also been demonstrated.6 Nestin-positivecells derived from isolated human islets were reported to havethe ability to differentiate ex vivo into pancreatic endocrine,exocrine, and hepatocyte phenotypes.4,25,26 However, nestinas a marker for islet progenitor cells is highly controversial,27
and several lines of compelling evidence have demonstrated therise of islet cells from precursors that do not express nestin,during embryonic development, in the adult pancreas, or duringislet regeneration.28,29 Data demonstrating the plasticity ofadult stem cells and adult differentiated cells and thetransdifferentiation of these cells into various cell types,even crossing the lineage boundary, have been reported.30,31
In this report, we describe an in vitro culture proce-dure for the generation of ICCs from human pancreaticdiscard through processes of selection, expansion, anddifferentiation, without the use of a matrix. ICCs generatedby this method are ready for transplantation within 3 to 4weeks. These ICCs contain approximately 20% as much in-sulin as the purified islets prepared from the same pancreasand are capable of releasing insulin in response to high-glucose stimulation in vitro. After transplantation into non-diabetic immunodeficient mice, the ICCs continuouslymature, acquire adult islet-like morphology, and releaseinsulin as indicated by the presence of human C-peptide in therecipient sera.
MATERIALS AND METHODS
Pancreatic Cell PreparationThe use of human pancreatic tissue in this study was
approved by the institutional review committee at the City ofHope/Beckman Research Institute. Human pancreata wereprocured from heart-beating cadaveric multiorgan donors bythe local organ procurement agency (OneLegacy, LosAngeles, Calif ) with appropriate informed consent, includingthe research use approval of the tissue. Pancreata wereprocessed for islets using a 2-step digestion method32
following the standard operation procedures of the SouthernCalifornia Islet Cell Resources Center of the City of HopeNational Medical Center. Liberase-digested pancreatic tissuewas placed on Euro-Ficoll solutions to separate islet and non-islet fractions by a discontinuous gradient centrifugation on aCOBE2991 cell processor (Gambro, Lakewood, Colo). Isletsmigrated in the interface between layers of the densities 1.056and 1.086(g/mL). Cells in Euro-Ficoll solutions of densityhigher than 1.096 g/mL contained less than 1% islets andwere not used for transplantation and discarded. This fractionwas named as Bpancreatic discard.^ The pancreatic discardrepresented the majority of the digested pancreatic tissue andhad a volume far greater than that of the purified islets (920vs. 1Y3 mL in the packed cell volume). The pancreatic discardcells were collected for this study after completing isletisolation and washed with Hanks balanced salt solution(Mediatech, Herndon, Va) before starting culture.
Cell Culture MethodsCells in the pancreatic discard were cultured through 2
consecutive steps: the first step was for cell selection and
expansion (proliferation culture) and the second step for celldifferentiation (differentiation culture). Cells and cell clumpsobtained after washing the pancreatic discard were placed intissue culture flasks coated with collagen I (BD Biosciences,Bedford, Mass) in Ham’s F12 culture medium (Mediatech) sup-plemented with 5 mmol/L nicotinamide (Sigma, St Louis, Mo),nonessential amino acids (Sigma), antibiotics (Sigma), and10% fetal bovine serum (FBS, Omega Scientific, Tarzana,Calif). The medium was changed daily for 1 to 2 days andthereafter every 2 to 3 days. When monolayer-grown cellsreached confluence (usually 4Y7 days), they were harvestedusing trypsin (Sigma).
Cell differentiation was initiated by promoting singlecells harvested from monolayer cultures to form cellaggregates. The monolayer-grown cells from the proliferationcultures were suspended in the pancreatic cell differentiationmedium (PCDM) and placed in nontissue culture-coated Petridishes (Fisher, Pittsburgh, Pa). The PCDM was a MCDB153medium (Sigma), supplemented with 1 mmol/L calciumchloride (Sigma), 10 mmol/L nicotinamide, 20 ng/mL gastrin(Sigma), 5 ng/mL keratinocyte growth factor (KGF) (R&DSystems, Minneapolis, Minn), 20 ng/mL insulin-like growthfactor 1 (Sigma), 20 ng/mL exendin 4 (Bachem, Torrance,Calif), 0.1 mmol/L ascorbic acid (Sigma), and 2% FBS, butinsulin was not added. Petri dishes containing the suspendedcells were placed on an orbital shaker (Bellcoglass, Vineland,NJ) to promote cell clustering. The medium was changedevery 2 days. The cells formed aggregates within 24 to 48hours, and these aggregates were stable for a period of up to1 month. Cell clusters were harvested at a desired time toassess various parameters.
ImmunocytochemistryImmunocytochemical analyses were performed on cells
grown on coverslips or cell clusters attached to polylysine-coated coverslips and fixed with cold methanol and acetone.Alternatively, cell clusters were fixed with formalin andembedded first in agar and then in paraffin for preparation ofhistology sections. The following primary antibodies wereused for immunocytochemical staining: rabbit anti-humanamylase (Sigma), mouse anti-human cytokeratin (CK) 7(Dako, Carpinteria, Calif), mouse anti-human CK19 (Dako),guinea pig anti-human insulin (Linco Research, St Charles,Mo), rabbit anti-human glucagon (Linco), rabbit anti-humansomatostatin (Sigma), rabbit anti-human PDX-1 (2 differentantibodiesVkind gifts from Dr Chris Wright, Vanderbilt Uni-versity, Nashville, Tenn, and Dr Marc Montmini, Salk In-stitute, La Jolla, Calif ), rabbit anti-human MCM2 (hMCM233),and mouse anti-human nestin (BD Biosciences, San Diego,Calif ). Corresponding secondary antibodies against each firstantibody/immunoglobulin were either conjugated with fluo-rescein (FITC) or Texas red (all from Jackson Immuno-Research Laboratories, West Grove, Pa). All samples werecounterstained for DNA with DAPI (Sigma). Stained sampleswere embedded in Vectashield (Vector Laboratories, Burlin-game, Calif) and examined under an Olympus BX51 fluo-rescent microscope, equipped with a Pixera 600CL cooledCCD camera (Olympus America, Melville, NY). A picture foreach individual color was separately captured using Pixera
Pancreas & Volume 32, Number 2, March 2006 Generation of Human Islets by Expansion and Differentiation
* 2006 Lippincott Williams & Wilkins 131
Copyr ight © Lippincott Williams & Wilkins. Unauthorized reproduction of this article is prohibited.

Viewfinder 3.0 acquisition software (Pixera, Los Gatos, Calif)and processed using Adobe Photoshop 7.0 software (Adobe,San Jose, Calif). Merged images are shown in all figures.
Measurements of ICC Insulin ContentsCell aggregates were harvested from differentiation
cultures and washed with phosphate-buffered saline (Media-tech). After 5 washes, the cell pellet was snap-frozen in liquidnitrogen and stored at j80-C until insulin extraction andmeasurements of insulin and DNA. For insulin and DNAmeasurements, the frozen stored samples were thawed andsonicated in water. The lysate was obtained by centrifugingthe sonicated material at 12,000g for 3 minutes using anEppendorf centrifuge (Eppendorf North America, Westbury,NY). Insulin was measured using an ELISA kit (Alpco,Windham, NH) and DNA using a Pico Green Kit (MolecularProbes, Eugene, Ore). Insulin contents in ICCs were ex-pressed as insulin amount (ng)/DNA (6g).
In Vitro Insulin Release AssaysTo assess the insulin release function of ICCs, a static
incubation assay was performed. The static incubation assayswere performed on 5 batches of ICCs generated from dif-ferent pancreatic discards obtained from 5 consecutive isletisolations. Before the assay, ICCs were washed and cul-tured overnight in RPMI1640 medium (Sigma) containing3.3 mmol/L glucose and 2% FBS (basal medium). Before thestatic incubation assay, the ICCs were washed 5 times withbasal medium and then incubated in basal medium. After30 minutes, the medium was collected for insulin assay (firstlow-glucose sample) and then replaced by high-glucose stim-ulation medium (RPMI1640 containing 19.3 mmol/L glucoseand 2% FBS) to incubate ICCs for another 30 minutes.Collection of the supernatant (high-glucose sample) wasfollowed by 5 washes with basal medium, and then the ICCswere again incubated with basal medium for 30 minutes toobtain a second low-glucose sample. This was followed by30 minute incubation in high-glucose medium containing10 mmol/L theophylline (high-glucose plus theophyllinesample) and then the third incubation in basal medium (thirdlow-glucose sample) after an additional 5 washes with basalmedium. Collected samples were frozen and stored untilmeasurement of human insulin. The stimulation index (SI)was calculated as insulin released into the stimulation mediumdivided by insulin released into the basal medium.
Transplantation of ICCs intoImmunodeficient Mice
In vivo survival, function, and growth/maturation of exvivoYgenerated ICCs were examined by transplanting them in-to immunodeficient mice, either athymic or NOD/severecombined immunodeficiency mice. Approximately 2000 ICCs,containing an estimated number of 4 million cells, weretransplanted under the renal capsule of normal (nondiabetic)mice. At the time of graft removal, human C-peptide levels in therecipient serum were measured 30 minutes after an intraperito-neal injection of 2 g/kg glucose. The kidney carrying the ICCgraft was removed at various intervals after transplantation, fixedin formalin, and processed for immunohistochemical examina-tion. Human C-peptide was measured using an ELISA kit(ALPCO, Windham, NH).
Statistical AnalysisData are expressed as average T SD. Differences
between groups were analyzed by the unpaired Studentt test. A value of P less than 0.05 was considered significant.
RESULTS
Pancreatic Cells Expand and Dedifferentiate/Transdifferentiate in Monolayer Cultures
Days 0 to 1 Monolayer Cultures
The starting material, pancreatic discard, contained70% to 80% (74.89% T 0.77%) of cells stained for the aci-nar cell marker amylase, 20% to 30% (28.59% T 0.72%)for CK7+ or CK19+ ductal cell markers, and less than 1%(0.35% T 0.34%) of cells were positive for insulin and alsofor the other islet hormones (Table 1). After being placedin collagen IYcoated flasks using PCPM medium, cells thatdid not attach to the plastic surface were mostly acinar cellsand were washed away when the medium was changed af-ter overnight culture (day 1, day 0 is the day of starting theculture). The cells that had attached to the flask bottom stainedeither for amylase or CK7, and their percentages were similar,changing significantly from the starting material (P G 0.05;amylase-positive vs. CK7+ cells = 43.59% T 6.32% vs. 41.08% T2.89%, n = 3), and less than 0.15% of the cells were insulin-positive or other pancreatic hormoneYpositive (Fig. 1).Some amylase-positive cells were also costained for CK7(12% T 5.6%) at this stage of culture, indicative of
TABLE 1. Cell Composition of Discard Fraction, Monolayer Cultures, and Aggregates
Insulin Glucagon Amylase CK7 PDX1 MCM2
Non-islet fraction 0.35 T 0.34 nd 74.89 T 0.77 28.59 T 0.72 nd nd
d1 nd nd 43.59 T 6.32 41.08 T 2.89 nd nd
d2 nd nd 29.97 T 19.68 66.0 T 4.27 n/a 4.5 T 2.9
d5 nd nd 6.72 T 1.91 92.51 T 4.96 96.13 T 1.23 31.07 T 11.9
Aggregates 12.71 T 2.31 9.19 T 2.62 nd 63.63 T 14.30 n/a nd
N = 4; for each experiment, 600 to 1200 cells were counted at more than 4 random fields.Data are given as % of total cells T SD.nd Indicates not detected (e0.15); n/a, data not shown; d1, d2, and d5, monolayer cultures on days 1, 2, and 5, respectively.
Todorov et al Pancreas & Volume 32, Number 2, March 2006
132 * 2006 Lippincott Williams & Wilkins
Copyr ight © Lippincott Williams & Wilkins. Unauthorized reproduction of this article is prohibited.

dedifferentiation or transdifferentiation of acinar cells tocells of the ductal phenotype. Staining for hMCM2, a proteininvolved in the regulation of DNA replication and used as amarker for cell proliferation,33 was totally negative on day 1.
Day 2 Monolayer Cultures
Cells rapidly spread in a monolayer in this short pe-riod, with a significant increase in CK7+ cells (66.0 T 4.27,P G 0.05 compared with day 1). The amylase-positive cellnumber decreased markedly, and the amylase-CK7 double-positive cell number increased to 22% T 10.6%. A few cellsexpressed hMCM2 (4.5% T 2.91%), and most of these cellswere also CK7+.
Days 4 to 7 Monolayer Cultures
Cells were nearly confluent with an expansion factorof 2 (as compared with the cell number initially attached tothe flask). Over 90% of the cells expressed the ductal cellmarkers CK7 and CK19 (92.51% T 4.96% for CK7, n = 3),and only a few cells were stained for amylase (6.72% T1.91%, n = 3), all of which also stained for CK7 (Fig. 1B),again indicating acinar cell dedifferentiation or trans-differentiation. The percent of cells that were combinedinsulin-positive and glucagon-positive was less than 0.3%.Most of the monolayer cells (96.13% T 1.23%, n = 3) alsoexpressed the pancreatic lineage marker, PDX 1, both in thenucleus and cytoplasm (Fig. 1D). Less than 1% of the cellsat this stage stained for the nerve progenitor cell markernestin (data not shown).
Monolayer Grown Cells Form Aggregates,Differentiate, and Express Insulin inDifferentiation Culture
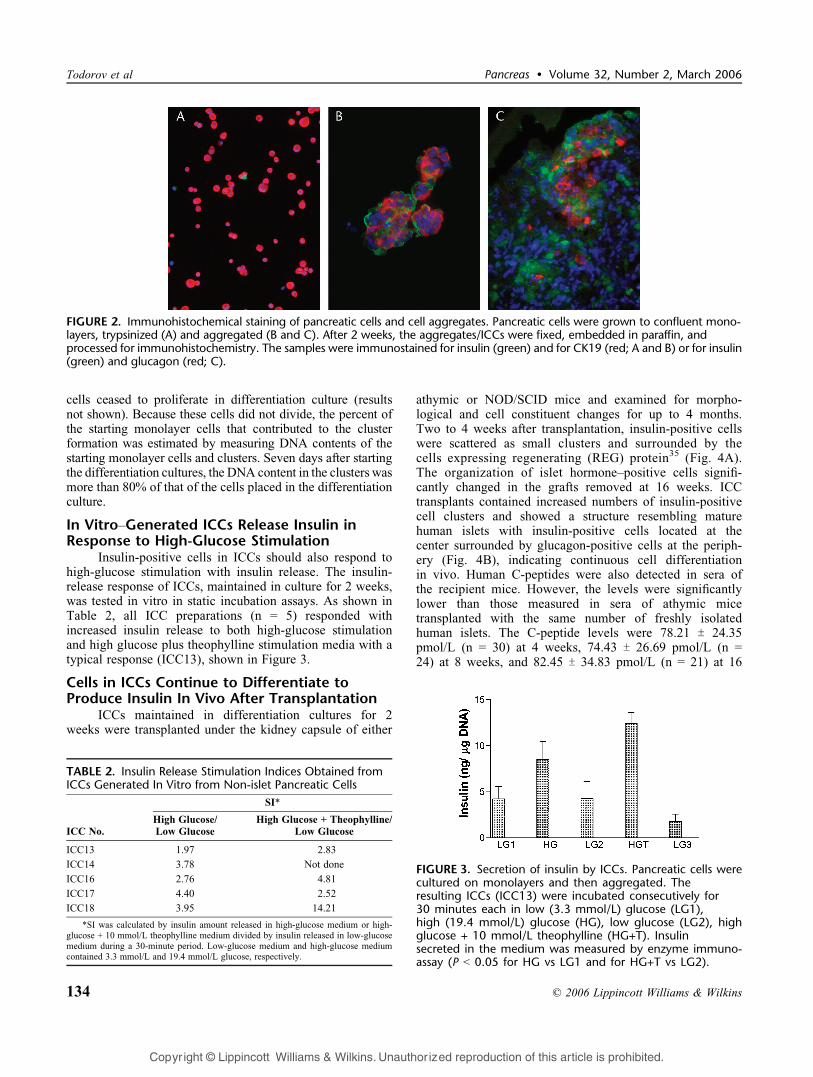
When monolayer cultures reached 80% to 90%confluence, the cells were placed in suspension cultures. Topromote cell-to-cell adhesion/attachment, the calcium con-centration was increased in PCDM. The cell-cell contact wasalso facilitated by orbital shaking of culture flasks. Within 24to 48 hours, the cells started to form aggregates whichincreased in size, measuring between 50 and 250 6m indiameter. As time advanced, cell clusters also became stableand remained stable in culture for over 3 weeks. As shown inFig. 2, monolayers contained only trace number of insulin-positive cells. After 2 weeks, many cell aggregates started tostain positively with dithizone, a zinc-chelating dye, widelyused to detect islets,34 indicating the presence of insulin-producing cells (results not shown). As shown in Figures 2Band C, immunocytochemical staining detected insulin-positiveand glucagon-positive cells in cell aggregates (insulin-positivevs glucagon-positive cells = 12.7% T 2.3% vs. 9.2% T 2.6% ofthe total cell population, n = 3). The cells expressing the ductalphenotype, which was the major phenotype at the initiation ofdifferentiation cultures, decreased significantly from 92.51% T4.96% to 63.63% T 14.3% (P G 0.01) during the 2 weeks ofdifferentiation culture. Cells double-positive for insulin andCK19 were occasionally found. Because the appearance ofthese cell aggregates resembled islets and they containedinsulin-positive cells, we called them ICCs. None of the cells inthe aggregates at any stage stained for hMCM2, indicating that
FIGURE 1. Immunocytochemical staining of cultured pancreatic cells. Pancreatic cells were cultured on collagen IVcoated coverslipsand processed for immunocytochemistry on the first day (A and C) and at the time of reaching confluence (days 5 to 7; B and D).The cells were immunostained for amylase (green; A and B), CK7 (red; A-D), and for PDX1 (green; C and D).
Pancreas & Volume 32, Number 2, March 2006 Generation of Human Islets by Expansion and Differentiation
* 2006 Lippincott Williams & Wilkins 133
Copyr ight © Lippincott Williams & Wilkins. Unauthorized reproduction of this article is prohibited.
cells ceased to proliferate in differentiation culture (resultsnot shown). Because these cells did not divide, the percent ofthe starting monolayer cells that contributed to the clusterformation was estimated by measuring DNA contents of thestarting monolayer cells and clusters. Seven days after startingthe differentiation cultures, the DNA content in the clusters wasmore than 80% of that of the cells placed in the differentiationculture.
In VitroYGenerated ICCs Release Insulin inResponse to High-Glucose Stimulation
Insulin-positive cells in ICCs should also respond tohigh-glucose stimulation with insulin release. The insulin-release response of ICCs, maintained in culture for 2 weeks,was tested in vitro in static incubation assays. As shown inTable 2, all ICC preparations (n = 5) responded withincreased insulin release to both high-glucose stimulationand high glucose plus theophylline stimulation media with atypical response (ICC13), shown in Figure 3.
Cells in ICCs Continue to Differentiate toProduce Insulin In Vivo After Transplantation
ICCs maintained in differentiation cultures for 2weeks were transplanted under the kidney capsule of either
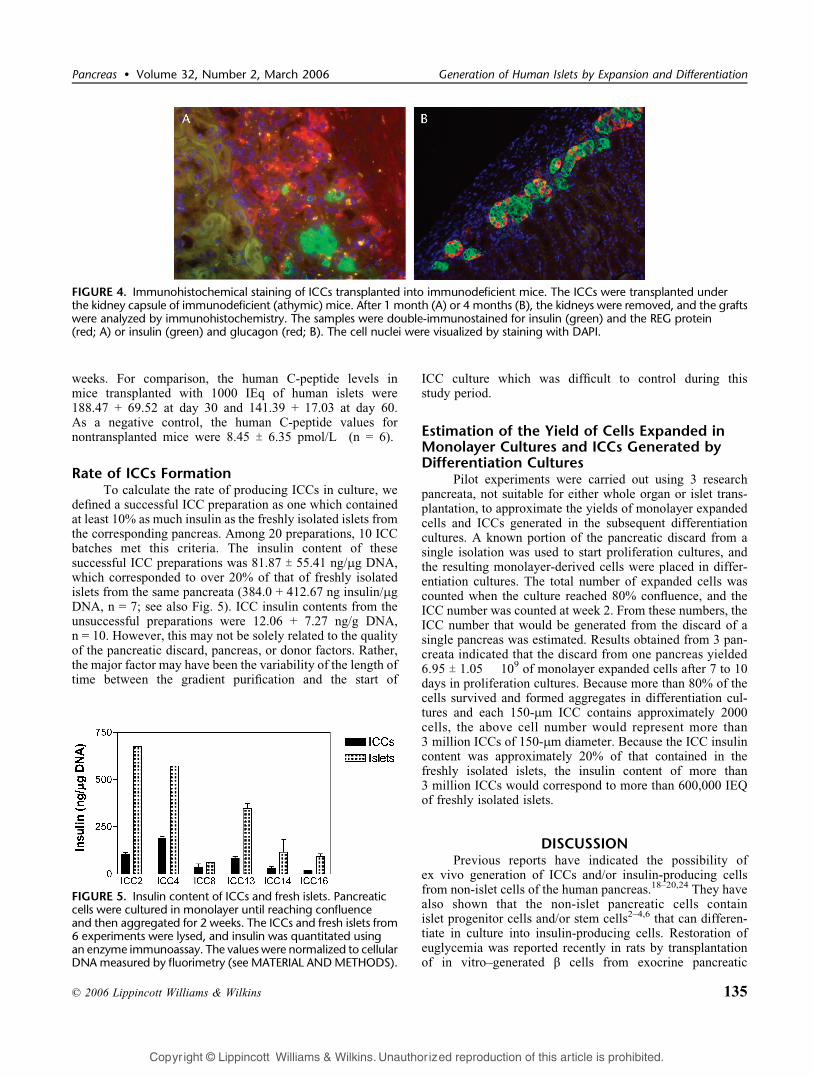
athymic or NOD/SCID mice and examined for morpho-logical and cell constituent changes for up to 4 months.Two to 4 weeks after transplantation, insulin-positive cellswere scattered as small clusters and surrounded by thecells expressing regenerating (REG) protein35 (Fig. 4A).The organization of islet hormoneYpositive cells signifi-cantly changed in the grafts removed at 16 weeks. ICCtransplants contained increased numbers of insulin-positivecell clusters and showed a structure resembling maturehuman islets with insulin-positive cells located at thecenter surrounded by glucagon-positive cells at the periph-ery (Fig. 4B), indicating continuous cell differentiationin vivo. Human C-peptides were also detected in sera ofthe recipient mice. However, the levels were significantlylower than those measured in sera of athymic micetransplanted with the same number of freshly isolatedhuman islets. The C-peptide levels were 78.21 T 24.35pmol/L (n = 30) at 4 weeks, 74.43 T 26.69 pmol/L (n =24) at 8 weeks, and 82.45 T 34.83 pmol/L (n = 21) at 16
TABLE 2. Insulin Release Stimulation Indices Obtained fromICCs Generated In Vitro from Non-islet Pancreatic Cells
ICC No.
SI*
High Glucose/Low Glucose
High Glucose + Theophylline/Low Glucose
ICC13 1.97 2.83
ICC14 3.78 Not done
ICC16 2.76 4.81
ICC17 4.40 2.52
ICC18 3.95 14.21
*SI was calculated by insulin amount released in high-glucose medium or high-glucose + 10 mmol/L theophylline medium divided by insulin released in low-glucosemedium during a 30-minute period. Low-glucose medium and high-glucose mediumcontained 3.3 mmol/L and 19.4 mmol/L glucose, respectively.
FIGURE 2. Immunohistochemical staining of pancreatic cells and cell aggregates. Pancreatic cells were grown to confluent mono-layers, trypsinized (A) and aggregated (B and C). After 2 weeks, the aggregates/ICCs were fixed, embedded in paraffin, andprocessed for immunohistochemistry. The samples were immunostained for insulin (green) and for CK19 (red; A and B) or for insulin(green) and glucagon (red; C).
FIGURE 3. Secretion of insulin by ICCs. Pancreatic cells werecultured on monolayers and then aggregated. Theresulting ICCs (ICC13) were incubated consecutively for30 minutes each in low (3.3 mmol/L) glucose (LG1),high (19.4 mmol/L) glucose (HG), low glucose (LG2), highglucose + 10 mmol/L theophylline (HG+T). Insulinsecreted in the medium was measured by enzyme immuno-assay (P G 0.05 for HG vs LG1 and for HG+T vs LG2).
Todorov et al Pancreas & Volume 32, Number 2, March 2006
134 * 2006 Lippincott Williams & Wilkins
Copyr ight © Lippincott Williams & Wilkins. Unauthorized reproduction of this article is prohibited.
weeks. For comparison, the human C-peptide levels inmice transplanted with 1000 IEq of human islets were188.47 + 69.52 at day 30 and 141.39 + 17.03 at day 60.As a negative control, the human C-peptide values fornontransplanted mice were 8.45 T 6.35 pmol/L (n = 6).
Rate of ICCs FormationTo calculate the rate of producing ICCs in culture, we
defined a successful ICC preparation as one which containedat least 10% as much insulin as the freshly isolated islets fromthe corresponding pancreas. Among 20 preparations, 10 ICCbatches met this criteria. The insulin content of thesesuccessful ICC preparations was 81.87 T 55.41 ng/6g DNA,which corresponded to over 20% of that of freshly isolatedislets from the same pancreata (384.0 + 412.67 ng insulin/6gDNA, n = 7; see also Fig. 5). ICC insulin contents from theunsuccessful preparations were 12.06 + 7.27 ng/g DNA,n = 10. However, this may not be solely related to the qualityof the pancreatic discard, pancreas, or donor factors. Rather,the major factor may have been the variability of the length oftime between the gradient purification and the start of
ICC culture which was difficult to control during thisstudy period.
Estimation of the Yield of Cells Expanded inMonolayer Cultures and ICCs Generated byDifferentiation Cultures
Pilot experiments were carried out using 3 researchpancreata, not suitable for either whole organ or islet trans-plantation, to approximate the yields of monolayer expandedcells and ICCs generated in the subsequent differentiationcultures. A known portion of the pancreatic discard from asingle isolation was used to start proliferation cultures, andthe resulting monolayer-derived cells were placed in differ-entiation cultures. The total number of expanded cells wascounted when the culture reached 80% confluence, and theICC number was counted at week 2. From these numbers, theICC number that would be generated from the discard of asingle pancreas was estimated. Results obtained from 3 pan-creata indicated that the discard from one pancreas yielded6.95 T 1.05 � 109 of monolayer expanded cells after 7 to 10days in proliferation cultures. Because more than 80% of thecells survived and formed aggregates in differentiation cul-tures and each 150-6m ICC contains approximately 2000cells, the above cell number would represent more than3 million ICCs of 150-6m diameter. Because the ICC insulincontent was approximately 20% of that contained in thefreshly isolated islets, the insulin content of more than3 million ICCs would correspond to more than 600,000 IEQof freshly isolated islets.
DISCUSSIONPrevious reports have indicated the possibility of
ex vivo generation of ICCs and/or insulin-producing cellsfrom non-islet cells of the human pancreas.18Y20,24 They havealso shown that the non-islet pancreatic cells containislet progenitor cells and/or stem cells2Y4,6 that can differen-tiate in culture into insulin-producing cells. Restoration ofeuglycemia was reported recently in rats by transplantationof in vitroYgenerated $ cells from exocrine pancreatic
FIGURE 5. Insulin content of ICCs and fresh islets. Pancreaticcells were cultured in monolayer until reaching confluenceand then aggregated for 2 weeks. The ICCs and fresh islets from6 experiments were lysed, and insulin was quantitated usingan enzyme immunoassay. The values were normalized to cellularDNAmeasured by fluorimetry (see MATERIAL ANDMETHODS).
FIGURE 4. Immunohistochemical staining of ICCs transplanted into immunodeficient mice. The ICCs were transplanted underthe kidney capsule of immunodeficient (athymic) mice. After 1 month (A) or 4 months (B), the kidneys were removed, and the graftswere analyzed by immunohistochemistry. The samples were double-immunostained for insulin (green) and the REG protein(red; A) or insulin (green) and glucagon (red; B). The cell nuclei were visualized by staining with DAPI.
Pancreas & Volume 32, Number 2, March 2006 Generation of Human Islets by Expansion and Differentiation
* 2006 Lippincott Williams & Wilkins 135
Copyr ight © Lippincott Williams & Wilkins. Unauthorized reproduction of this article is prohibited.

tissue. However, most of the previous culture methodswere not suitable for a large-scale production of ICCs in acontrollable manner. Our objective was to generate cellclusters containing $ cells from human pancreatic discard ina controllable in vitro procedure, which has the potentialto be expanded into a large-scale production of ICCs forclinical application.
Our pancreatic cell culture methods contained 2 essen-tial steps. The first step used monolayer cultures to select andexpand particular pancreatic cell populations, and the secondstep used suspension cultures to facilitate the formation ofcell aggregates to induce cell differentiation. In proliferationcultures, the cells need to attach rapidly to the bottom offlasks. Collagen I coating of the flasks and the extracellularmatrix molecules provided by serum-supplemented medium,for example, fibronectin and vitronectin, facilitated cell ad-herence, and approximately 20% of the cells and cell clumpsin the pancreatic discard attached to the flasks. The remainingcells not attached to the flask were washed away when themedium was changed. Through this initial stage, cells withthe ductal phenotype were selected, as indicated by the in-creased CK7+/CK19+ cell numbers from 30% in the initiallyplated population to 40% in the attached cells. During thisstage, the dedifferentiation/transdifferentiation of acinar cellsto the ductal phenotype also occurred as indicated by thepresence of cells double-positive for amylase and CK7, withmore than 90% of the cells expressing CK7 when culturesreached confluence. In the earlier stages (1Y2 days), mono-layer cells contained a very low number of replicating cells(Table 1), indicating that the marked increase of the CK7+
cells was mostly caused by acinar cell dedifferentiation,rather than the proliferation of CK7+ ductal cells. Some de-differentiating amylase-positive cells also stained for theproliferation marker and CK7 (results not shown), indicatinglow levels of proliferation. These findings confirmed theresults previously reported by several other groups.22,23,36,37
At the end of the monolayer cultures, more than 90% of thecells were positive for CK7 and CK19 and also expressed thepancreatic transcription factor, PDX1, a decisive moleculeexpressed by the cells that differentiate into the pancreaticendocrine lineage.38,39 These results support that the mono-layer culture method we developed selectively expands cellsof the pancreatic endocrine lineage within a short period,while acinar cells were either eliminated or dedifferentiated/transdifferentiated to a cell type that expresses CK7/CK19and PDX1.
The key components initiating cell differentiation inthe second step of our culture process were the cell-cell contact resulting from cell aggregation. Several factorsin our differentiation medium contributed to supporting cell-cell contact, preserving stable cell aggregates, and facilitat-ing cell differentiation in the aggregates. An increasedcalcium concentration is essential for the formation of cellaggregates, suggesting the involvement of the cadherinfamily members in this process essential for the aggregationof islet cells.40,41 Nicotinamide is known as a potent inducerof endocrine cell differentiation.42 Exendin 4, a stable analogof the glucagon-like peptide 1, has also been shown toinduce pancreatic cell differentiation.43 Our differentiation
medium also contained insulin-like growth factor 1, KGF/FGF7, and gastrin. IFG-I prevents pancreatic cell apopto-sis.44 KGF/FGF7 is an important factor for ductal cellproliferation and differentiation,18,45 and gastrin has beenshown to stimulate $-cell neogenesis from transdifferentiat-ed pancreatic cells.46,47
After 2 weeks in differentiation cultures, the ICCscontained both insulin-positive and glucagon-positive cells.Because our differentiation medium did not contain insulin,the positive insulin staining represented the presence ofintracellular insulin and insulin production. However, theinsulin level measured in the ICCs was far lower than that inislets freshly isolated from the corresponding pancreas. TheseICCs released insulin in response to high-glucose stimulation,demonstrating that, unlike fetal islets,48 these cells werefunctionally mature. After transplantation into immunodefi-cient mice, the differentiation and maturation process in theICCs further progressed as shown by the increased insulin-positive cell numbers and acquisition of the mature adult isletmorphology (Fig. 4B). In the earlier stages of transplantation,insulin-positive cells were surrounded by cells expressing theREG 1 protein. Because the REG 1 protein has been shownto facilitate $/islet cell growth35 or differentiation of pancrea-tic cells into $ cells,49 REG protein might have played a rolein the ICC differentiation/maturation. The morphologicalmaturation of ICCs required a long period, comparable to thatreported for the maturation of neonatal50 and fetal islets.51,52
Although insulin was released from ICC grafts as demon-strated by the presence of human C-peptide in recipients’sera, human ICCs transplanted after 2 weeks in differentiationcultures did not reverse diabetes in athymic mice. This maybe attributable to the small number of insulin-positive cellsand the limited space under the renal capsule for placingICCs. Reversal of diabetes was reported in mice by trans-planting in vitroYgenerated $ cells from rat exocrine pan-creatic cells.24 Studies are in progress to examine the abilityof in vitroYgenerated human ICCs to restore euglycemia indiabetic NOD/SCID mice.
In summary, we have developed a new procedure forgenerating cell clusters ex vivo from human pancreaticdiscard that contains cells producing insulin and glucagon.Our method has advantages over previously publishedmethodologies, especially in the following 2 areas: (1) ourmonolayer culture can select and expand cells of thepancreatic endocrine lineage and are harvested readily tostart differentiation cultures and (2) stable cell clusters/ICCsare formed, and cell differentiation is initiated without theneed for an extracellular matrix, such as Matrigel, and thegenerated ICCs are easily harvested at a desired time. The totalnumber of ICCs that can be generated from the discardof a single pancreas is estimated to be over 3 million, whichcorresponds to over 600,000 IEQ when calculated by theinsulin content of ICCs. After transplantation, cell differen-tiation and maturation continue, ICCs form the morphology ofmature islets, and thus, their ability to produce insulin isexpected to increase. Finally, the scale of both selection/expansion and differentiation cultures can readily be expandedto process a large amount of cells. ICCs could be readyfor transplantation in 3 to 4 weeks. Demonstration of the
Todorov et al Pancreas & Volume 32, Number 2, March 2006
136 * 2006 Lippincott Williams & Wilkins
Copyr ight © Lippincott Williams & Wilkins. Unauthorized reproduction of this article is prohibited.

ability of ICCs to reverse diabetes would be a further steptoward the use of in vitroYgenerated ICCs in clinicaltransplantation.
ACKNOWLEDGEMENTThe authors thank Drs Chris Wright, Vanderbilt
University, and Marc Montmini, Salk Institute, for the kindgifts of anti-PDX1 antibodies; Dr Elisabeth Stein for criticalreading of the manuscript; and Sofia Loera and TinaMontgomery, City of Hope Pathology Laboratory, for theirexcellent assistance in preparation of the histology samples.
REFERENCES1. Shapiro AM, Lakey JR, Ryan EA, et al. Islet transplantation in seven
patients with type 1 diabetes mellitus using a glucocorticoid-freeimmunosuppressive regimen. N Engl J Med. 2000;343:230Y238.
2. Bonner-Weir S, Sharma A. Pancreatic stem cells. J Pathol. 2002;197(4):519Y526.
3. Lechner A, Habener JF. Stem/progenitor cells derived from adult tissues:potential for the treatment of diabetes mellitus. Am J Physiol EndocrinolMetab. 2003;284:E259YE266.
4. Zhang YQ, Sarvetnick N. Development of cell markers for theidentification and expansion of islet progenitor cells. Diabetes Metab ResRev. 2003;19(5):363Y374.
5. Hussain MA, Theise ND. Stem-cell therapy for diabetes mellitus. Lancet.2004;364(9429):203Y205.
6. Trucco M. Regeneration of the pancreatic beta cell. J Clin Invest. 2005;115(1):5Y12.
7. Fishman JA, Patience C. Xenotransplantation: infectious risk revisited.Am J Transplant. 2004;4(9):1383Y1390.
8. Lumelsky N, Blondel O, Laeng P, et al. Differentiation of embryonicstem cells into insulin secreting structures similar to pancreatic islets.Science. 2001;292:1389Y1394.
9. Hori Y, Rulifson IC, Tsai BC, et al. Growth factor inhibitors promotedifferentiation for insulin producing tissue from embryonic stem cells.Proc Natl Acad Sci U S A. 2002;99:16105Y16110.
10. Hansson M, Tonning A, Frandsen U, et al. Artifactual insulin releasefrom differentiated embryonic stem cells. Diabetes. 2004;53(10):2603Y2609.
11. Kania G, Blyszczuk P, Wobus AM. The generation of insulin-producingcells from embryonic stem cellsYa discussion of controversial findings.Int J Dev Biol. 2004;48(10):1061Y1064.
12. Fujikawa T, Oh SH, Pi L, et al. Teratoma formation leads to failure oftreatment for type I diabetes using embryonic stem cellYderivedinsulin-producing cells. Am J Pathol. 2005;166(6):1781Y1791.
13. Hori Y, Gu X, Xie X, et al. Differentiation of insulin-producing cellsfrom human neural progenitor cells. PLoS Med. 2005;2(4):e103.
14. Kojima H, Fujimiya M, Matsumura K, et al. NeuroD-betacellulin genetherapy induces islet neogenesis in the liver and reverses diabetes inmice. Nat Med. 2003;9(5):596Y603.
15. Suzuki A, Nakauchi H, Taniguchi H. Glucagon-like peptide 1 (1Y37)converts intestinal epithelial cells into insulin producing cells. Proc NatlAcad Sci U S A. 2003;100:5034Y5039.
16. Zalzman M, Gupta S, Giri RK, et al. Reversal of hyperglycemia in miceby using human expandable insulin-producing cells differentiated fromfetal liver progenitor cells. Proc Natl Acad Sci U S A. 2003;100(12):7253Y7258.
17. Cao LZ, Tang DQ, Horb ME, et al. High glucose is necessary forcomplete maturation of Pdx1-VP16Yexpressing hepatic cells intofunctional insulin-producing cells. Diabetes. 2004;53(12):3168Y3178.
18. Bonner-Weir S, Taneja M, Weir GC, et al. In vitro cultivation ofhuman islets from expanded ductal tissue. Proc Natl Acad Sci U S A.2000;97:7999Y8004.
19. Ramiya VK, Maraist M, Arfors KE, et al. Reversal of insulin-dependentdiabetes using islets generated in vitro from pancreatic stem cells. NatMed. 2000;6:278Y282.
20. Gao R, Ustinov J, Pulkkinen MA, et al. Characterization of endocrineprogenitor cells and critical factors for their differentiation in humanadult pancreatic cell culture. Diabetes. 2003;52(8):2007Y2015.
21. Lechner A, Nolan AL, Blacken RA, et al. Redifferentiation ofinsulin-secreting cells after in vitro expansion of adult human pancreaticislet tissue. Biochem Biophys Res Commun. 2005;327(2):581Y588.
22. Bouwens L. Transdifferentiation versus stem cell hypothesis for theregeneration of islet beta-cells in the pancreas. Microsc Res Tech. 1998;43:332Y336.
23. Gmyr V, Kerr-Conte J, Belaich S, et al. Adult human cytokeratin19Ypositive cells reexpress insulin promoter factor 1 in vitro: furtherevidence for pluripotent pancreatic stem cells in humans. Diabetes.2000;49(10):1671Y1680.
24. Baeyens L, De Breuck S, Lardon J, et al. In vitro generation ofinsulin-producing beta cells from adult exocrine pancreatic cells.Diabetologia. 2005;48(1):49Y57.
25. Zulewski H, Abraham EJ, Gerlach MJ, et al. Multipotentialnestin-positive stem cells isolated from adult pancreatic isletsdifferentiate ex vivo into pancreatic endocrine, exocrine and hepaticphenotypes. Diabetes. 2001;50:521Y533.
26. Abraham EJ, Kodama S, Lin JC, et al. Human pancreatic isletYderivedprogenitor cell engraftment in immunocompetent mice. Am J Pathol.2004;164(3):817Y830.
27. Street CN, Lakey JR, Seeberger K, et al. Heterogenous expression ofnestin in human pancreatic tissue precludes its use as an islet precursormarker. J Endocrinol. February 2004;180(2):213Y225.
28. Selander L, Edlund H. Nestin is expressed in mesenchymal and notepithelial cells of the developing mouse pancreas. Mech Dev. 2002;113(2):189Y192.
29. Humphrey RK, Bucay N, Beattie GM, et al. Characterization andisolation of promoter-defined nestin-positive cells from the human fetalpancreas. Diabetes. 2003;52(10):2519Y2525.
30. Masson S, Harrison DJ, Plevris JN, et al. Potential of hematopoietic stemcell therapy in hepatology: a critical review. Stem Cells. 2004;22(6):897Y907.
31. Quesenberry PJ, Dooner G, Colvin G, et al. Stem cell biology and theplasticity polemic. Exp Hematol. 2005;33(4):389Y394.
32. Kenmochi T, Miyamoto M, Une S, et al. Improved quality and yield ofislets isolated from human pancreata using a two-step digestion method.Pancreas. 2000;20(2):184Y190.
33. Todorov IT, Werness BA, Wang HQ, et al. HsMCM2/BM28: a novelproliferation marker for human tumors and normal tissues. Lab Invest.1998;78(1):73Y78.
34. Latif ZA, Noel J, Alejandro R. A simple method of staining fresh andcultured islets. Transplantation. 1988;45(4):827Y830.
35. Okamoto H. The REG gene family and REG proteins: with specialattention to the regeneration of pancreatic beta-cells. J HepatobiliaryPancreat Surg. 1999;6(3):254Y262.
36. Hall PA, Lemoine NR. Rapid acinar to ductal transdifferentiation incultured human exocrine pancreas. J Pathol. 1992;166(2):97Y103.
37. Vila MR, Lloreta J, Real FX. Normal human pancreas cultures displayfunctional ductal characteristics. Lab Invest. 1994;71(3):423Y431.
38. Ferber S, Halkin A, Cohen H, et al. Pancreatic and duodenal homeoboxgene 1 induces expression of insulin genes in liver and amelioratesstreptozotocin-induced hyperglycemia. Nat Med. 2000;6(5):568Y572.
39. Edlund H. Pancreatic organogenesisVdevelopmental mechanisms andimplications for therapy. Nat Rev Genet. 2002;3:524Y532.
40. Cirulli V, Baetens D, Rutishauser U, et al. Expression of neural celladhesion molecule (N-CAM) in rat islets and its role in islet cell typesegregation. J Cell Sci. 1994;107(pt 6):1429Y1436.
41. Dahl U, Sjodin A, Semb H. Cadherins regulate aggregation of pancreaticbeta-cells in vivo. Development. 1996;122(9):2895Y2902.
42. Otonkoski T, Beattie GM, Mally MI, et al. Nicotinamide is a potentinducer of endocrine differentiation in cultured human fetal pancreaticcells. J Clin Invest. 1993;92(3):1459Y1466.
43. Egan JM, Bulotta A, Hui H, et al. GLP-1 receptor agonists are growthand differentiation factors for pancreatic beta cells. Diabetes Metab ResRev. 2003;19:115Y123.
44. Liu W, Chin-Chance C, Lee EJ, et al. Activation ofphosphatidylinositol 3-kinase contributes to insulin-like growth factorIYmediated inhibition of pancreatic beta-cell death. Endocrinology.2002;143(10):3802Y3812.
Pancreas & Volume 32, Number 2, March 2006 Generation of Human Islets by Expansion and Differentiation
* 2006 Lippincott Williams & Wilkins 137
Copyr ight © Lippincott Williams & Wilkins. Unauthorized reproduction of this article is prohibited.

45. Movassat J, Beattie GM, Lopez AD, et al. Keratinocyte growth factorand beta-cell differentiation in human fetal pancreatic endocrineprecursor cells. Diabetologia. 2003;46(6):822Y829.
46. Wang TC, Bonner-Weir S, Oates PS, et al. Pancreatic gastrin stimulatesislet differentiation of transforming growth factor alphaYinducedductular precursor cells. J Clin Invest. 1993;92(3):1349Y1356.
47. Rooman I, Lardon J, Bouwens L. Gastrin stimulates beta-cell neogenesisand increases islet mass from transdifferentiated but not from normalexocrine pancreas tissue. Diabetes. 2002;51(3):686Y690.
48. Tsunoda T, Furui H, Klandorf K, et al. Functional maturation ofporcine fetal pancreatic explants. Transplant Proc. 1989;21(1 pt 3):2667Y2668.
49. Zenilman ME, Magnuson TH, Swinson K, et al. Pancreatic thread proteinis mitogenic to pancreatic-derived cells in culture. Gastroenterology.1996;110:1208Y1214.
50. Korbutt CG, Elliott JF, Ao Z, et al. Large scale isolation, growth, andfunction of porcine neonatal islet cells. J Clin Invest. 1996;97:2119Y2129.
51. Shiogama T, Mullen Y, Klandorf H, et al. Growth and functionalmaturation of cryopreserved human fetal pancreata. Diabetes. 1986;38(suppl 1):284.
52. Beattie GM, Lopez AD, Otonkoski T, et al. Transplantation of humanfetal pancreas: fresh vs. cultured fetal islets or ICCS. J Mol Med.1999;77(1):70Y73.
Todorov et al Pancreas & Volume 32, Number 2, March 2006
138 * 2006 Lippincott Williams & Wilkins
Copyr ight © Lippincott Williams & Wilkins. Unauthorized reproduction of this article is prohibited.




















