
Mural cell-derived laminin-α5 plays a detrimental role in ...
A
T
i
(
pf©
K
1
fddscmsctddc
PT
0d
Molecular and Cellular Endocrinology 263 (2007) 120–133
Beneficial effects of prolactin and laminin on humanpancreatic islet-cell cultures
Leticia Labriola a, Wagner R. Montor a, Karin Krogh a, Fernando H. Lojudice a,Tercio Genzini b, Anna C. Goldberg a, Freddy G. Eliaschewitz b, Mari C. Sogayar a,∗
a Chemistry Institute, University of Sao Paulo, Brazilb Albert Einstein Hospital, Sao Paulo, Brazil
Received 31 January 2006; received in revised form 15 September 2006; accepted 16 September 2006
bstract
The problem of pancreas donor shortage could be addressed through in vitro islet-cell proliferation prior to transplantation into diabetic patients.herefore, we set out to evaluate the effects of prolactin (rhPRL) and laminin on primary cultures of human pancreatic islets.Our results showed that rhPRL induced an increase in islet-cell number and in cumulative insulin secretion (p < 0.01). However, glucose-induced
nsulin secretion was enhanced only in the presence of both laminin and rhPRL.In addition, we describe, for the first time in human islets, the PRL-induced activation of JAK2, and signal transducer and activator of transcription
STAT) 1, 3 and 5.
Our results demonstrate a significant beneficial effect of rhPRL and laminin on human islets and support widely held notion that the closerhysiological stimuli and environment of beta cells are mimicked, the better are the results in cell proliferation and secretory function, both essentialor successful islet transplantation.
2006 Elsevier Ireland Ltd. All rights reserved.
; Lam
lgeliaaprai
eywords: Diabetes; Human Pancreatic islets; Beta-cell proliferation; Prolactin
. Introduction
Type 1 diabetes is an insulin-deficient condition resultingrom the autoimmune destruction of pancreatic beta cells. Thus,iabetic patients require lifelong insulin therapy, but often thisoes not prevent the commonest complications of diabetes. Inome cases, transplantation of isolated pancreatic islets fromadaveric organ donors is a promising possibility for opti-al treatment of type 1 diabetes. However, this approach is
everely limited by the shortage of organ donors. Ex vivo islet-ell culture prior to transplantation is as an attractive alterna-ive, but maintenance of human islets in culture has been a
ifficult task. Therefore, stimulation of islet proliferation andifferentiation in vitro remains a major scientific and clinicalhallenge.∗ Corresponding author at: Chemistry Institute, University of Sao Paulo, Av.rof. Lineu Prestes 748, Room 964, Sao Paulo 05508-900, SP, Brazil.el.: +55 11 3091 3820; fax: +55 11 3091 3820.
E-mail address: [email protected] (M.C. Sogayar).
iasr
bmpI
303-7207/$ – see front matter © 2006 Elsevier Ireland Ltd. All rights reserved.oi:10.1016/j.mce.2006.09.011
inin; Insulin; STAT
Hormones of the growth hormone (GH) family, such as pro-actin (PRL) and placental lactogen (PL), are the most potentrowth factors for pancreatic beta cells reported to date (Nielsent al., 2001). The lactogenic hormones PL and PRL stimu-ate DNA synthesis, insulin production, and glucose-dependentnsulin secretion in pancreatic islets of fetal and neonatal ratsnd mice and adult humans (Brelje et al., 1993, 1994; Breljend Sorenson, 1991; Swenne et al., 1987). In rat fet uses and inregnant mothers, the long isoform of the pancreatic prolactineceptor (PRLR) is expressed predominantly (Asfari et al., 1995)nd, upon binding of PRL to this receptor, Janus kinase 2 (JAK2)s activated. This in turn will lead to phosphorylation, dimer-zation and nuclear translocation of the signal transducer andctivation of transcription (STAT), which bind to specific con-ensus DNA sequences in the promoters of target genes, therebyegulating their transcription (Bole-Feysot et al., 1998).
Recently, it has been shown that lactogen induction of
eta-cell aggregation, coupled with localized beta-cell growth,ay contribute to the growth of islet-cell mass that occurs inregnancy and in the perinatal period (Fleenor et al., 2000).n addition, Freemark et al. (2002) have recently shown that

llular
palg
micepjte
lswp(if(lcc
wicam
chttpsi
2
2
nIPdbeatdrcLfiM(fltC2
2
o1csa1(tocap(
TR
I
1234567
M
L. Labriola et al. / Molecular and Ce
rolactin receptor (PRLR) deficiency is accompanied by isletnd beta-cell hypoplasia, reduced pancreatic insulin mRNAevels, blunted insulin secretory response to glucose and mildlucose intolerance.
The ability to maintain pancreatic islets in culture provides aethod for in vitro islet proliferation and storage, prior to clin-
cal transplantation. However, one major disadvantage of isletulture is the loss of tissue mass that occurs over time (Fragat al., 1998; Weber et al., 1997). Given the importance of trans-lanting large amounts of islets, any loss of islet tissue mass mayeopardize the chances of a successful graft. Therefore, factorshat could protect the islets from apoptosis, or at least reduce theffects of deleterious agents, would be beneficial.
In fact, in adults, it has been proposed that islet extracellu-ar matrix (ECM) protein interactions are required for beta-cellurvival and function (Wang and Rosenberg, 1999). Severalorks have shown the importance of laminin, the major com-onent of the basal membrane of epithelial and endothelial cellsColognato and Yurchenco, 2000), in enhancing adult rat isletsnsulin secretion in vitro (Bosco et al., 2000) or promoting dif-erentiation of mouse fetal pancreatic cells into beta cells in vitroJiang et al., 1999). Furthermore, it has been demonstrated thataminin is likely to be in close association with islet endocrineells since it was detected within the islets and associated withapillaries (Jiang et al., 2002).
The goal of the work presented here was to determinehether treatment of primary cultures of human pancreatic
slets with rhPRL and/or laminin would result in increased beta-ell mass without loss of their functional properties, with theim of defining new strategies to increase the final islet-cellass.Here, we report, for the first time, that rhPRL induces islet-
ell proliferation and basal insulin release in primary cultures ofuman pancreatic islets and that addition of laminin increaseshe glucose-induced secretory function. We further report, for
he first time in human pancreatic islets, the protein level andhosphorylation state of JAK2, STATs 1, 3 and 5, as an initialtep towards an understanding of the molecular mechanismsnvolved in the action of PRL.iBr1t
able 1esponse of human islet preparations to rhPRL treatment
slet preparation Donor data Pancreas coldischaemia (h)
Islet preparation
Age Sex Purity (%) Glucose stimu(24 h after isol
40 M 4 90 ± 5 1.5 ± 0.546 M 8 90 ± 5 3.7 ± 1.048 F 7 90 ± 7 1.2 ± 0.670 M 8 80 ± 6 2.4 ± 0.849 F 7 80 ± 5 1.2 ± 0.548 F 7 75 ± 5 3.1 ± 0.751 F 6 60 ± 7 2.7 ± 1.1
ean ± S.E. 50 ± 3 7 ± 1 80 ± 5 2.3 ± 0.9
a Fold stimulation relative to the vehicle (control), considered as 1.b Fold increase in glucose stimulation index relative to the vehicle (control), consid
Endocrinology 263 (2007) 120–133 121
. Materials and methods
.1. Cell cultures
Human pancreas from adult brain-dead donors (mean age 50 ± 3 years,= 7) were harvested in conformity with Brazilian regulations and the local
nstitutional Ethical Committee. Donor characteristics are shown in Table 1.ancreatic islets were isolated after ductal distension of the pancreas andigestion of the tissue with Liberase (Roche Diagnostics, Indianapolis, IN)y the automated method of Ricordi et al. (1988) with modifications (Shapirot al., 2000). Islets were purified in a continuous Ficoll density gradient inCOBE 2991 cell processor (Gambro, Lakewood, CO). The islet prepara-
ions used in this study exhibited 80 ± 5% purity, which was determined byithizone staining, and islet-cell viability, evaluated using the live/dead fluo-escent method, based on the incorporation of either acridine orange by liveells (Sigma, ST. Louis, MO) or propidium iodide by dead cells (Sigma, ST.ouis, MO), was usually greater than 80%. Upon isolation, the islets wererst maintained in CMRL 1066 medium (5.6 mM glucose) (Mediatech-Cellgro,iami, FL) supplemented with 100 units/mL penicillin and 5% fetal calf serum
FCS) (Cultilab, Campinas, SP, Brazil) in non-adherent T75 flasks in the free-oating state (Falcon, Becton Dickinson Labware, NJ, USA) for 24–48 h, and
hen transferred to adherent flasks and cultured in CMRL 1066 (Mediatech-ellgro, Miami, FL) supplemented with 100 units/mL penicillin and 5% FCS for4–48 h.
.2. Proliferation assay
For proliferation assays, cultured islet cells were trypsinized in a solutionf 1 g/L trypsin (Life Technologies Inc., Grand Island, NY), 1 mM EDTA,40 mM NaCl and 2.7 mM KCl in phosphate-buffered saline (PBS) withoutalcium or magnesium (pH 7.4) for 1 min at room temperature. The celluspension was then diluted 1:10 in CMRL 1066 supplemented with 10% FCSnd centrifuged for 1 min at 800 × g; the cell pellet was resuspended in CMRL066 supplemented with 10% FCS and cells were counted in a hemocytometerNeubauer chamber). After trypsinization, islet-cell viability was evaluated byhe live/dead fluorescent method (Sigma, St. Louis, MO), showing an averagef 85% live cells. A 104 cells/well were plated in 24-well adherent dishes andultured for 48 h in CMRL 1066 and supplemented with 100 units/mL penicillinnd 5% FCS, in the presence or in the absence of mouse laminin-1 (10 �g/mL)urified from murine Engelbreth–Holm–Swarm (EHS) tumor basal membraneInvitrogen, San Diego, CA), or in dishes coated with Matrigel® solution
n CMRL 1066 medium (50:1)/cm2 (Collaborative Biomedical Products,edford, MA) (Kleinman et al., 1982). Before starting the incubation withecombinant human prolactin (rhPRL), cells were serum-starved in CMRL066 medium supplemented with 0.1% FCS for 24 h. On the following day,he cells were incubated with CMRL supplemented with 0.5% FCS and either
Islet adherent cultures upon 4-dayrhPRL treatment (300 ng/mL)
lation indexation)
Folda proliferation Folda accumulatedinsulin release
Glucosestimulation indexb
1.9 ± 0.7 3.5 ± 0.4 0.3 ± 0.13.4 ± 0.2 4.3 ± 0.4 0.7 ± 0.41.7 ± 0.4 8.0 ± 1.0 3.0 ± 0.81.6 ± 0.4 6.2 ± 0.7 1.3 ± 0.71.2 ± 0.5 5.7 ± 0.6 1.0 ± 0.52.8 ± 0.7 6.4 ± 0.8 1.3 ± 0.51.5 ± 0.9 6.1 ± 0.5 0.9 ± 0.4
2.0 ± 0.9 5.7 ± 1.2 1.2 ± 0.9
ered as 1.

1 llular
rP
c
2
iSen
2
oa
wac(ogaii
2
rCEcmEb0
2
leARudt(u3sT
sde(PCtfA
emaTet
2
l5p0taa
2
j(Tmsw(ombaws
(acTsetbpBp
3smw
2
2aifici(
22 L. Labriola et al. / Molecular and Ce
hPRL prolactin (300 ng/mL) produced in our laboratory (Lawson et al., 1982;ereira et al., 2001; Sapin et al., 2001) or vehicle for 4 days.
Cells were harvested with trypsin–EDTA, fixed with 3.7% formaldehyde andounted in an electronic CC530 cell counter (CELM, Sao Paulo, SP, Brazil).
.3. Accumulated insulin release
Conditioned media was collected from each well culture, after incubationn the presence or absence of rhPRL (300 ng/mL) for 4 days as described underection 2.2 above, and the accumulated insulin release, was quantified by thelectrochemiluminescence assay ELECSYS (Sapin et al., 2001) (Roche Diag-ostics, Indianapolis, IN). The results were normalized by cell number.
.4. Glucose-induced insulin secretion
Islet cells were cultured in adherent wells in the presence or in the absencef rhPRL (300 ng/mL) for 4 days, as previously described in the Section 2.2bove.
Cells were then washed with PBS and incubated for 2 h with RPMI 1640ithout glucose (Cultilab, Campinas, Brazil) supplemented with 0.5% BSA
nd 2.8 or 20 mM glucose. Secreted insulin was quantified in conditioned mediaollected from each well by the electrochemiluminescence assay ELECSYSRoche Diagnostics, Indianapolis, IN). The stimulation index for each groupf cultures was calculated by dividing the value of insulin secretion in 20 mMlucose by that obtained upon 2.8 mM glucose stimulation. Results are presenteds relative increase in the glucose stimulation index, where the index obtainedn the control (vehicle) was set to 1. The data shown represent the mean of sevenndependent experiments (n = 3 for each experiment) ± S.D.
.5. Insulin content
Islet-derived cells were cultured and incubated in the presence or absence ofhPRL (300 ng/mL) for 4 days, as described above for the proliferation assay.ells were then washed with PBS and detached with 0.25% trypsin and 0.2%DTA. The cell pellet was resuspended in 100 �L of PBS. A fraction of thisell suspension was mixed with glacial acetic acid containing 0.2% human albu-in and lysed for insulin determination by the electrochemiluminescence assayLECSYS (Roche Diagnostics, Indianapolis, IN). The results were normalizedy either cell number or DNA content. The limit of detection of this assay was.05 ng/106 cells.
.6. Real-time polymerase chain reaction (real-time PCR)
Cells were harvested and total RNA were prepared as previously pub-ished (Sogayar et al., 2004), with minor modifications. RNA integrity waslectrophoretically verified by ethydium bromide staining and through thebs260 nm/Abs280 nm absorption ratio >1.95. cDNA was generated from theNA samples using SuperScript (Invitrogen, Carlsbad, CA). The primerssed for gene amplification by quantitative real-time PCR experiments wereesigned with the Primer Express 2.0 Software (Applied Biosystems, Fos-er City, California, USA) connected to the ABI ABI-5700 ThermocyclerApplied Biosystems, Foster City, CA, USA). The following primers weresed in this study: human insulin (F: 5′-TGCGGGGAACGAGGCTTCTTCTA-′; R: 5′-AGGGACCCCTCCAGGGCCAAG-3′), hypoxanthine phosphoribo-yl transferase (HPRT) (F: 5′-GAACGTCTTGCTCGAGATGTGA-3′; R: 5′-CCAGCAGGTCAGCAAAGAAT-3′).
In comparative gene expression analysis, the accuracy depends on primerets having equal efficiency, which can be determined by the slope of a stan-ard curve. The corresponding real-time PCR efficiency (E) of one cycle in thexponential phase was calculated according to the equation: E = 10(−1/slope)Rasmussen et al., 2001). To quantify the products formed during the real-time
CR reaction, the SYBR® Green Dye reagent (Applied Biosystems, Fosterity, CA, USA) was used (Lekanne Deprez et al., 2002). Management of thehermocycler and assessment of data generated during amplification were per-ormed using the ABI-Manager SDS program (Roche, Indianapolis, IN, USA).ll real-time PCR reactions were performed in triplicates (n = 4 independent
LdDUM
Endocrinology 263 (2007) 120–133
xperiments). For the mathematical model, it was necessary to manually deter-ine the crossing points (Ct) for each transcript. Ct was defined as the point
t which the fluorescence rises appreciably above the background fluorescence.he relative expression of a target gene was determined in comparison to a ref-rence gene (Pfaffl, 2001), which was chosen to be a stable and unregulatedranscript, e.g. a housekeeping gene transcript, such as HPRT.
.7. Preparation of total cell protein extract
Primary cultures of human islets were washed with ice-cold PBS and thenysed by scraping at 4 ◦C in a lysis buffer (10 mM Tris, pH 7.5, 150 mM NaCl,mM EDTA, 1 mM EGTA, 1 mM DTT, 10 �g/mL aprotinin, 10 �g/mL leu-eptin, 5 �g/mL pepstatin, 1 mM PMSF, 25 mM NaF, 1% NP-40, 0.1% SDS,.5% sodium deoxycholate and 1 mM Na3VO4). The homogenate was cen-rifuged for 30 min at 12,000 × g and the supernatant fraction was then collectednd stored at −70 ◦C. The protein content in these extracts was quantified usingBio-Rad kit (Bio-Rad, Richmond, CA).
.8. Western blot analysis and immunoprecipitation
Total extracts were prepared from primary cultures of pancreatic islets sub-ected to the different treatments described in each experiment. Equal amounts100 �g) of proteins from each extract were solubilized in sample buffer (60 mMris–HCl [pH 6.8], 2% sodium dodecyl sulfate (SDS), 10% glycerol, 0.01% bro-ophenol blue) and subjected to SDS–polyacrylamide gel (7.5%) electrophore-
is (PAGE). Proteins were electrotransferred to nitrocellulose membranes, whichere blocked and then incubated with the following antibodies: anti-JAK2
Santa Cruz Biotechnology Inc., Santa Cruz, CA), anti-STAT3 mouse mon-clonal antibody; anti-STAT1 rabbit polyclonal antibody; anti-STAT5 mouseonoclonal antibody (BD transduction Laboratories, San Diego, CA). The mem-
ranes were then incubated with horseradish peroxidase-conjugated secondaryntibody (Vector Laboratories, Burlingame, CA). Enhanced chemiluminescenceas performed according to the manufacturer’s instructions (Amersham Bio-
ciences, Buckinghamshire, UK).For JAK2, STATs 5, 3 and 1 tyrosine phosphorylation analysis, cell lysates
1 mg of total cell proteins) were incubated with 4 �g of either anti-JAK2,nti-STAT3, anti-STAT1 or anti-STAT5 antibodies. Thereafter, the immuno-omplexes were captured with protein A–G agarose (SIGMA, St. Louis, MO).he beads were washed three times with lysis buffer, boiled for 10 min inample buffer and subjected to SDS–PAGE on a 7.5% gel. Proteins werelectroblotted onto nitrocellulose membranes and tyrosine–phosphorylated pro-eins were detected by Western blotting using an anti-phosphotyrosine anti-ody Clone PY69 (BD transduction Laboratories), followed by horseradisheroxidase-conjugated rabbit anti-mouse immunoglobulin (Vector laboratories,urlingame, CA). As a loading control, the membranes were stripped and re-robed with either anti-JAK2, anti-STAT3, 1 or 5 antibodies.
Quantitative densitometry was carried out with a Hewlett Packard (hpscanjet500) scanner and the Image Quant 5.2 (Molecular Dynamics, Amersham Bio-ciences, Buckinghamshire, UK) software. The volume density of the chemilu-inescent bands (n = 3 experiments performed with different islet preparations)as calculated as O.D. × mm2 after background correction.
.9. Immunofluorescence
Cultured islet cells were trypsinized and plated onto glass coverslips in a4-well plate at a density of 104 cells/well. After the 4-day treatment protocollready described in Section 2.2, cells were fixed and permeabilized for 10 minn PBS containing 4% paraformaldehyde and 0.3% Triton X-100 at 37 ◦C. Afterxation, the cultures were washed with PBS, incubated in blocking solution (PBSontaining 2% BSA) and then incubated with the following primary antibod-es diluted in blocking solution: human anti-insulin rabbit polyclonal antibodyICN, Aurora, OH), human anti-insulin mouse monoclonal antibody (Zymed
aboratories, San Francisco, CA), rabbit polyclonal antibody against (homeoboxomain transcription factor) Pdx1 (Kawaguchi et al., 2002) (kindly donated byr. C.V. Wright, Department of Cell Biology, School of Medicine, Vanderbiltniversity), human anti-cytokeratin 19 mouse monoclonal antibody (ICN, Cosaesa, CA) and human anti-vimentin mouse monoclonal antibody (Amersham
llular
BflaeGFe5citttp
ietm
2
2t
foarBbp
2
tdtt
3
3
Ftcra
L. Labriola et al. / Molecular and Ce
iosciences, Buckinghamshire, UK). The primary antibodies were detected withuorescent labeled secondary antibodies (Vector Laboratories, Burlingame, CA)nd DAPI was used for nuclear staining. The immunostained coverslips werexamined under a confocal laser scanning microscope LSM 510 (Carl ZeissmbH, Jena, Germany) with a 40× oil immersion objective. Images of DAPI,ITC and Texas Red fluorophores were taken with the corresponding sets ofxcitation and emission filters (351 nm/385–470 nm, 488 nm/505–530 nm and43 nm/>560 nM, respectively). For each experimental condition, at least 100ells of each replicate (n = 3) were visually inspected for the intracellular local-zation of the fluorescence signal, by two independent observers, with one ofhem unaware of sample identity. The agreement between findings obtained byhe two observers was always greater than 90%. Images selected were represen-ative of the majority of the cells present on each experiment (n = 6 experimentserformed with different islet preparations).
Control experiments to test for the specificity of the primary antibodiesncluded incubation with preimmune mouse and rabbit sera. In addition, cov-rslips were plated with NIH-3T3, as a negative control, and incubated withhe rabbit polyclonal anti-insulin antibody. All confocal immunofluorescence
icroscopy images presented correspond to single optical sections.
.10. BrdU labelling
Cultured islet cells were trypsinized and plated on to glass coverslips in a4-well plate at a density of 104 cells/well. The cells were submitted to the 4-dayreatment protocol described in the Proliferation assay section above. Twenty-
i
t
ig. 1. Primary cultures of human pancreatic islets characterization. Cells obtained fo attach for 24 h in CMRL 1066 medium supplemented with 5% FCS and then theells were labeled with antibodies against Pdx1 (top and bottom left panel), insulin (tight panel). Slides incubated with rabbit and mouse IgGs (bottom left panel) and NIHs controls for the primary antibodies specificity. In all cases nuclear staining was pe
Endocrinology 263 (2007) 120–133 123
our hours before the end of the experiment, cells were cultured in the presencef 10 �M BrdU (Amersham Biosciences, Buckinghamshire, UK). After fixingnd blocking, BrdU, insulin and Pdx1 labeling were detected by immunofluo-escence, using the following antibodies: anti-BrdU (Amersham Biosciences,uckinghamshire, UK), anti-insulin rabbit polyclonal or rabbit polyclonal anti-ody against Pdx1. Microscopic examination was carried out as described in therevious section.
.11. Statistical analysis
Data are presented as the mean ± S.E. Each experiment was repeated at leasthree times with duplicate or triplicate values within each group. The statisticalifferences between group means were tested, after confirming the normal dis-ribution of data, by ANOVA, followed by Tukey’s test or by unpaired two-tailed-test. p < 0.05 was considered statistically significant.
. Results
.1. Effect of human prolactin on cell proliferation and
nsulin secretionPrevious results from our laboratory characterized the short-erm (up to 28 days) adherent islet-cell cultures derived from
rom human islet primary cultures were plated on glass coverslips and allowedcultures were serum starved for 24 h. After fixation and permeabilization, theop middle and right panel), vimentin (top middle panel) or cytokeratin 19 (top3T3 cells incubated with anti-insulin antibody (bottom right panel) were used
rformed with DAPI. A representative field (40×) is shown.

124 L. Labriola et al. / Molecular and Cellular
Fig. 2. Effect of human prolactin (hPRL) on cell proliferation of human isletprimary cultures. Cells (104 cells/well) obtained from human islet primary cul-tures were incubated in adherent dishes for 4 days in CMRL medium containing5.6 mM glucose and 0.5% FCS supplemented with different concentrations ofrop
ssUwww
odcCccp
hiSrierfirv
atrlfictio
eus
ioac(f
iogw
amnntv
suwpo
3
EeomhB
mstw
sewvdSm
w
hPRL. Cells were then harvested and counted. Data are presented as the meanf three independent experiments (n = 4 for each experiment) ± S.D. (b vs. a:< 0.001).
everal human islet preparations, with respect to their expres-ion of various islet-cell markers (Maria-Engler et al., 2004).sing confocal microscopy and semi-quantitative RT-PCR, itas shown that a high proportion of cells were insulin-positive,hile very few cytokeratin 19 (CK19)- or vimentin-positive cellsere observed (Maria-Engler et al., 2004).In agreement with these earlier results, the adherent cultures
btained from primary cultures of human pancreatic islets, asescribed in Methods, exhibited 88 ± 2% (n = 7) insulin-positiveells, 86 ± 4% (n = 7) Pdx1-positive cells, 1.3 ± 0.5% (n = 7)K19-positive cells and 0.8 ± 0.2% (n = 7) vimentin-positiveells (Fig. 1). It is important to emphasize that the adherent isletultures used in the present study, were never maintained for aeriod of time longer than 21 days.
Since, it has been already described that the significance oformones of the GH family is mainly indicated by the increasedslet-cell mass in both rodents and humans (Nielsen et al., 1992;orenson et al., 1997), we first studied the in vitro effects ofecombinant human prolactin (rhPRL) on human pancreaticslets. To this end, islet cells were incubated for 4 days withither vehicle or increasing concentrations (20–300 ng/mL) ofhPRL, obtained as described in Section 2. After formaldehydexation, the number of cells was determined. As shown in Fig. 2,hPRL-induced cell proliferation in a dose-dependent manner (aersus b: p < 0.001).
We then investigated the long-term effects of rhPRL onccumulated insulin release and insulin intracellular content inhe presence of basal glucose concentration (5.6 mM). Insulinelease and insulin content were determined in cultures stimu-ated in the presence or in the absence of rhPRL (300 ng/mL)or 4 days. Since a high variability between independent exper-ments was detected, we were forced to express the insulin
ontent and release data relative to the plastic and vehicle cul-ure condition, as the internal standard. Thus, for example,nsulin content in rhPRL-treated cells was detected in the rangef 200–2000 ng/106 cells, depending on the islet preparationlott
Endocrinology 263 (2007) 120–133
mployed. Despite the high variability of data, absolute val-es of accumulated insulin release, insulin content and glucosetimulation index were reported later on this section.
The results showed that rhPRL treatment of these islet cellsnduced a significant (5.7 ± 1.2-fold) (p < 0.01, n = 7) increasever the control cultures, incubated with vehicle only (Fig. 3And Table 1). As expected, the insulin content was also signifi-antly increased upon rhPRL treatment both at the protein level2.6 ± 0.1-fold) (p < 0.01, n = 3) and at the mRNA (2.1 ± 0.3-old; p < 0.05, n = 4) (Fig. 3B and C).
In order to determine the effects of rhPRL on glucose-inducednsulin secretion, cells were cultured for 4 days in the absencer in the presence of rhPRL (300 ng/mL). After a washing step,lucose-stimulated insulin release was determined in cells thatere incubated for 2 h with glucose (2.8 or 20 mM).Even though the phenotype of regulated insulin secretion
fter glucose stimulation was maintained in multiple experi-ents, the results presented in Fig. 3D showed that rhPRL was
ot able to increase glucose-stimulated insulin release. In fact,o significant effect on the stimulation index was detected inhe rhPRL-treated cells compared to the control, incubated withehicle only (Fig. 3D and Table 1).
Despite the fact that a correlation has been noted betweenex of fetal pancreas donors and the increase in insulin secretionpon PRL stimulation of free-floating islets (Piper et al., 2004),e were not able to establish any correlation between any of thearameters analyzed and the sex or age of organ donors, underur culture conditions (Table 1).
.2. Effect of laminin on islet-cell proliferation and function
Accumulated evidence has highlighted the importance ofCM signals for pancreatic development and function (Lelievret al., 1996; Wang and Rosenberg, 1999). In particular, the rolef laminin, one of the major (80%) components of the base-ent membrane of epithelial cells or dispersed rat islet cells,
as recently been described as important (Baker et al., 1996;osco et al., 2000).
Therefore, we investigated the influence of laminin on pri-ary cultures of human pancreatic islets. We first performed the
ame proliferation assay as referred to in Fig. 2, but with the addi-ion (or not) of laminin or Matrigel® to the cultures. Matrigel®
as used as an alternative source of ECM components.We found that laminin did not alter the capacity of rhPRL to
timulate cell proliferation, while Matrigel® inhibited cell prolif-ration even in the presence of rhPRL (p < 0.001) (Fig. 4). In fact,hile the number of cells plated on the plastic and incubated withehicle doubled, the number of cells plated in Matrigel®-coatedishes did not alter during the 4-day PRL treatment (Fig. 4).ince the presence of Matrigel® in the culture counteracted ourain goal, it was discarded for further experiments.In order to characterize the proliferating cells in our cultures,
e performed BrdU incorporation assays and insulin or Pdx1
abeling of the cells cultured in the presence or in the absencef laminin and treated (or not) with rhPRL (300 ng/mL) duringhe 4-day period. Insulin and Pdx1 labeling was similar in allhe groups studied. A mean of 87 ± 2% (n = 7) and 86.3 ± 1.3%
L. Labriola et al. / Molecular and Cellular Endocrinology 263 (2007) 120–133 125
Fig. 3. Effect of human prolactin (hPRL) on the secretory function of human islet primary cultures. (A) Accumulated insulin release was measured, in conditionedmedia obtained from cultures treated either with rhPRL (300 ng/mL) or vehicle (control). Results are presented as relative increase in the accumulated insulin release,compared to control, which was set to 1. The data represent the mean of seven independent experiments (n = 3 for each experiment) ± S.D. (a vs. b: p < 0.01). (B)Insulin content in cell lysates from cultures treated either with rhPRL (300 ng/mL) or vehicle was measured by an electrochemiluminescence assay (ELECSYS,Roche). All measurements were standardized against total DNA content. Results are presented as relative increase, with the insulin content in the control (vehicle)treatment was set to 1. The data represent the mean of three independent experiments (n = 3 for each experiment) ± S.D. (a vs. b: p < 0.01). (C) Insulin mRNAcontent in cell lysates from cultures treated with either rhPRL (300 ng/mL) or vehicle was measured by real-time PCR reaction using the SYBR® Green Dye reagent(Applied Biosystems, Foster City, CA, USA). To quantify the products formed during the real-time PCR reaction, management of the thermocycler and assessmentof data generated during amplification were performed using the ABI-Manager SDS program (Roche, Indianapolis, IN, USA). All real-time PCR reactions wereperformed in triplicates (n = 4 independent experiments). The relative expression of a target gene was determined in comparison to a reference gene (HPRT). Resultsare presented as relative increase in the insulin mRNA level, where the value obtained in the control (vehicle) was set to 1. The data shown represent the mean offour independent experiments ± S.D. (a vs. b: p < 0.05). (D) Glucose-induced insulin secretion assay was performed in cells treated with either rhPRL (300 ng/mL)or vehicle. After a washing step, cells were incubated for 2 h with RPMI without glucose supplemented with 0.5% BSA and either 20 or 2.8 mM glucose. Insulin wast ults at dex oo < 0.0
(tstoncIp
o
aotcu(
hen measured by an electrochemiluminescence assay (ELECSYS, Roche). Reshe ratio of insulin secreted in 20 mM glucose vs. 2.8 mM glucose), where the inf seven independent experiments (n = 3 for each experiment) ± S.D. (a vs. b: p
n = 7) of the cells were positive for insulin and Pdx1, respec-ively (Table 2), regardless of the cell treatment. However, aignificant increase in both the BrdU labeling index and inhe proportion of double (insulin and BrdU)-stained cells wasbserved (Table 2 and Fig. 5), upon addition of rhPRL (p < 0.01,= 6), whether in the presence or in the absence of laminin, indi-ating the rhPRL-induced proliferation of insulin-positive cells.
t is important to note that all insulin-positive cells were alsoositive for Pdx1 and vice versa (data not shown).We also analyzed the morphology of islet cells growingn uncoated plastic or on laminin-coated dishes. We were
rfap
re presented as relative increase in the glucose-stimulation index (calculated asbtained in the control (vehicle) was set to 1. The data shown represent the mean1).
ble to show that laminin (10 �g/mL) induced the formationf cell-clusters, which grew to a large size during the 4-dayreatment with rhPRL (Fig. 6B black arrows). Culturing theells in plastic dishes only led to a change in cell densitypon rhPRL treatment, but no cluster formation was observedFig. 6A).
In an attempt to characterize the cells involved in this rear-
angement, staining for insulin and the homeobox transcriptionactor Pdx1, an early marker for pancreatic organogenesis andmarker for beta and for a subset of delta cells in the adultancreas (Oster et al., 1998), was performed with cells cultured
126 L. Labriola et al. / Molecular and Cellular
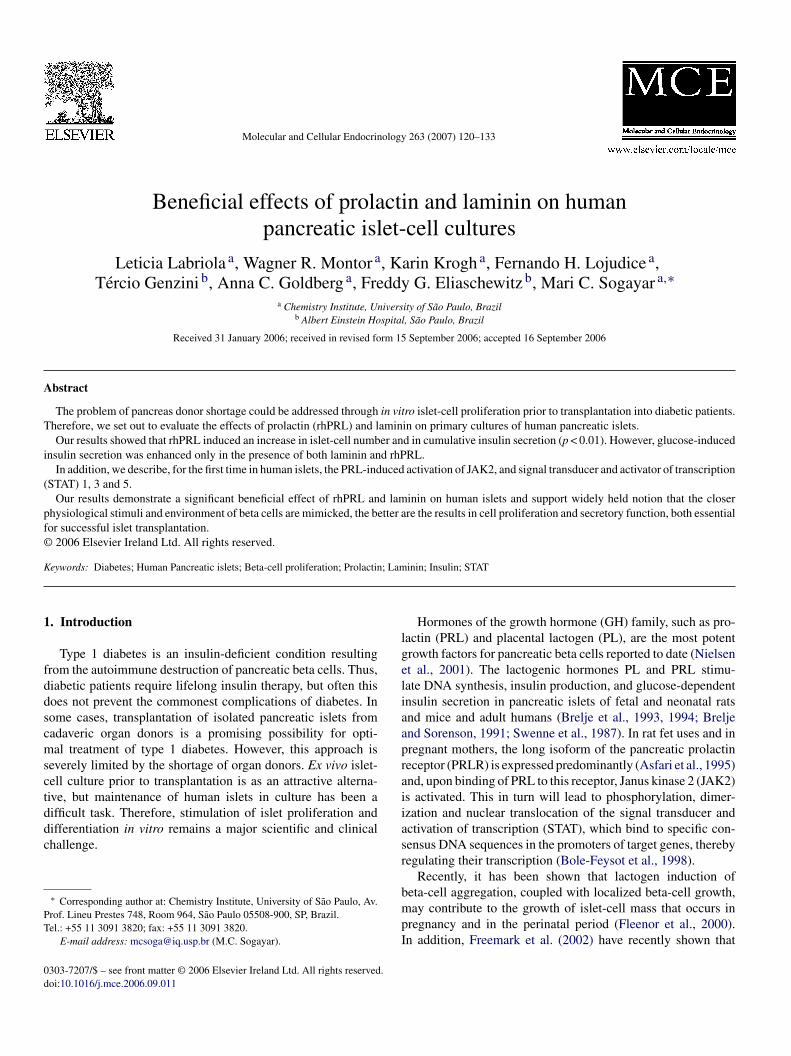
Fig. 4. Effect of human prolactin (rhPRL) and extracellular matrix compo-nents on the proliferation of human islet primary cultures. Cells (104 cells/well),obtained from human islet primary cultures, were plated on plastic, in thepresence or absence of laminin-1 (10 �g/mL) or Matrigel® (50 �L/cm2) andincubated for 4 days in CMRL medium containing 0.5% FCS supplementedwith either rhPRL (300 ng/mL), or vehicle (control). Cells were then harvestedand counted. Results are presented as relative increase, with the results obtainedfov
ir
8imonmi
oag
so(or1ltc4ipl
iwip2uti
35
ogenetcWatt
TM
C
PPLL
CCis
or the control (uncoated plastic plus vehicle) set as 1. Data represent the meanf seven independent experiments (n = 3 for each experiment) ± S.D. (a vs. b; as. c; c vs. b: p < 0.001).
n the presence or absence of laminin and treated (or not) withhPRL (300 ng/mL) during 4 days.
Fig. 6C and D show a high proportion of stained cells. Thus,5.8 ± 2.3% and 86.8 ± 1.4% of the cells were positive fornsulin and Pdx1, respectively (n = 7), independent of cell treat-
ent. Nevertheless, CK19- and vimentin-positive cells werenly rarely observed (1.5 ± 0.7% and 1.0 ± 2.5%, respectively,= 6) and were negative for insulin or Pdx1 (data not shown),aintaining a similar pattern of expression as the one observed
n Fig. 1.
The difference in cell aggregation induced by laminin wasur first finding suggesting that this ECM protein might playrole in in vitro human islet function. In order to investi-
ate the effect of laminin on PRL-induced cumulative insulin
rStp
able 2orphometric analysis of primary cultures of human adult islet cells immunostained
ondition BrdU labeling index(% of total cells)
Insulin positive(% of total cells)
Pdx1 positive (%of total cells)
lastic + vehicle 23.1 ± 3.2 a 82.8 ± 1.0 82.8 ± 1.0lastic + hPRL 51.25 ± 3.2 b 88.1 ± 7.5 85.7 ± 3.5aminin + vehicle 25.6 ± 1.1 a 84.7 ± 4.4 88.3 ± 0.8aminin + hPRL 54.6 ± 3.9 b 88.7 ± 2.6 88.3 ± 0.3 5
ells (104 cells/well), obtained from human islet primary cultures, were plated on plMRL medium containing 0.5% FCS supplemented with either rhPRL (300 ng/mL),
mmunostaining with antibodies to BrdU and insulin or Pdx1 was then performed. Aeven independent experiments ± S.E.M. (a vs. b: p < 0.01).
Endocrinology 263 (2007) 120–133
ecretion, cells were plated in the absence or in the presencef laminin (10 �g/mL) and then treated with either rhPRL300 ng/mL) or vehicle for 4 days. As shown in Fig. 6A, additionf laminin abrogated the accumulated insulin release induced byhPRL (plastic + vehicle: 26 ± 6 ng/106 cells; plastic + rhPRL:23.37 ± 37 ng/106 cells; laminin + vehicle: 21 ± 6 ng/106 cells;aminin + rhPRL: 50 ± 13 ng/106 cells). Interestingly, neitherhe insulin content of cells incubated in the presence of vehi-le (plastic + vehicle: 432 ± 97 ng/106 cells; laminin + vehicle:11 ± 119 ng/106 cells), nor the significant (p < 0.01) rhPRL-nduced two-fold increase in insulin content was altered in theresence of laminin (plastic + rhPRL: 1230 ± 369 ng/106 cells;aminin + rhPRL: 1067 ± 357 ng/106 cells) (Fig. 7A and B).
Conversely, when we studied the role of laminin in glucose-nduced insulin secretion, we found that the stimulation indexas increased about four-fold only in the case of cells cultured
n the presence of both laminin and rhPRL (stimulation indexlastic + vehicle: 2.4 ± 0.3; stimulation index plastic + rhPRL:.8 ± 0.3; stimulation index laminin + vehicle: 2.21 ± 0.4; stim-lation index laminin + rhPRL: 11 ± 2) (Fig. 7C). Altogether,hese results indicate that laminin may be primarily involved innsulin secretion induced by high glucose concentrations.
.3. Effect of human prolactin on JAK2 and STATs 1, 3 andprotein expression and state of phosphorylation
In order to look into the molecular basis for the action of PRLn primary cultures of human pancreatic islets, we first investi-ated the expression levels of JAK2 and STATs 1, 3 and 5. To thisnd, total protein extracts were prepared from cells treated (orot) with rhPRL (300 ng/mL) for several periods of time. West-rn blot analyses performed on whole cell lysates with antibodieso JAK2 and to STATs 1, 3 and 5 and the appropriate positiveontrols confirm the presence of these proteins (data not shown).e then studied the levels of tyrosine phosphorylation of JAK2
nd of STATs 1, 3 and 5 upon rhPRL treatment, as a marker forheir activation (Levy and Darnell, 2002). Cell extracts (1 mg ofotal protein) from islet cultures that were treated (or not) with
hPRL, were immunoprecipitated with anti-JAK2 or with anti-TATs 1, 3 and 5 antibodies and the phosphotyrosine content ofhese proteins was determined by Western blotting with an anti-hosphotyrosine antibody. Fig. 8A–C shows that while tyrosine
for BrdU and insulin or Pdx1
Double positive
Insulin and BrdU(% of total cells)
Pdx1 and BrdU(% of total cells)
Insulin and BrdU(% of insulin-positive cells)
Pdx1 and BrdU(% of Pdx1-positive cells)
21.3 ± 3.3 a 19.6 ± 2.4 a 25.7 ± 4.1 a 23.6 ± 5.5 a46.5 ± 2.8 b 51.9 ± 2.0 b 48.7 ± 7.2 b 60.6 ± 5.5 b26.7 ± 2.0 a 21.6 ± 1.8 a 31.5 ± 6.4 a 24.4 ± 2.6 a2.9 ± 2.6 b 48.0 ± 2.7 b 59.6 ± 5.1 b 54.4 ± 5.7 b
astic, uncoated or coated with laminin (10 �g/mL) and incubated for 4 days inor vehicle (control). BrdU (0.1 mmol/L) was added 24 h before fixation. Doublet least 100 cells were counted in each replicate. The data represent the mean of

L. Labriola et al. / Molecular and Cellular Endocrinology 263 (2007) 120–133 127
Fig. 5. Effect of rhPRL and extracellular matrix components on BrdU incorporation of human islet primary cultures. Cells (104 cells/well), obtained from humani g/mLv fixat(
pcpitofrkrmmdt
4
tlit
iobat2eilp
mdPist
slet primary cultures, were incubated in the presence or absence of laminin (10 �
ehicle (control) or rhPRL (300 ng/mL). BrdU (10 �M) was added 24 h beforegreen) was performed. A representative field (600×) was photographed.
hosphorylation of these proteins was almost undetectable inells maintained in 0.5% FCS, rhPRL strongly induced thehosphorylation levels of JAK2 and of STATs 1, 3 and 5. Thencrease of JAK2 tyrosine phosphorylation induced by rhPRLreatment was observed as early as 10 min after the additionf the hormone and was maintained, although at a lower level,or up to 2 h of treatment (Fig. 8D). Interestingly, the phospho-ylation state of these STATs displayed two different types ofinetics. As shown in Fig. 8A–C, the STAT5 tyrosine phospho-ylation level peaked after 10 min of PRL treatment, and wasaintained for up to 2 h, whereas STATs 1 and 3 reached theiraxims at 30 min, but, after 2 h, their phosphorylation states
ecreased to levels comparable to those obtained after 10 min ofreatment.
. Discussion
In this article, we describe the beneficial effects of a new cul-
ure protocol for human pancreatic islets, combining rhPRL andaminin. The results clearly demonstrate significantly increasedslet growth without loss of functionality, since the cells retainedhe insulin secretory response upon glucose stimulation.cbfll
) for 4 days in CMRL medium containing 0.5% FCS supplemented with eitherion and the double immunostaining with antibodies to BrdU (red) and insulin
Isolation of pancreatic islets and the ability to maintain themn tissue culture have been essential not only in the developmentf islet-cell transplantation as a clinical treatment for type 1 dia-etes mellitus, but also for their storage prior to transplantationnd for studies of islet biochemistry and physiology. A con-roversy surrounds human beta-cell replication (Beattie et al.,002; Hayek et al., 1995; Lefebvre et al., 1998), since the vari-ty of cell populations among islet cultures and differences inslet isolation protocols may interfere with the process of estab-ishing whether a growth factor is directly involved in beta-cellroliferation or in insulin secretion.
In the present study, we used human islet cultures with aajority of insulin and Pdx1-positive cells, and CK19-positive
uct cells and vimentin-positive cells being only rarely observed.revious work from our laboratory using confocal microscopy
n short and long-term human islet primary cultures have alsohown, beta cells-enriched cultures exhibiting a granular dis-ribution of insulin and very few CK19- and vimentin-positive
ells (Maria-Engler et al., 2004). A possible explanation for thiseta-cell enrichment is that the islets were cultured in a free-oating state for 48 h prior to plating and presented an outerayer of dead cells and an inner core of live cells, according to

128 L. Labriola et al. / Molecular and Cellular Endocrinology 263 (2007) 120–133
Fig. 6. Morphology of primary cultures of human pancreatic islets after rhPRL and laminin treatment. (A) Cells (104 cells/well), obtained from human islet primarycultures, were incubated for 4 days in CMRL medium containing 0.5% FCS supplemented with either vehicle (control) or rhPRL (300 ng/mL). A representativefield (×100) was photographed under a phase-contrast Nikon microscope. (B) Cells (104 cells/well), obtained from human islet primary cultures, were incubatedfor 4 days in CMRL medium containing 0.5% FCS and laminin-1 (10 �g/mL) supplemented with either rhPRL (300 ng/mL) or vehicle (control). A representativefield (100×) was photographed (arrows indicate islet-like cell clusters) under a phase-contrast Nikon microscope. (C) Cells (104 cells/well), obtained from humanislet primary cultures, were incubated for 4 days in CMRL medium containing 0.5% FCS supplemented with either vehicle (control) or rhPRL (300 ng/mL). Thecells were labeled with antibodies to insulin or Pdx1. Nuclei were labeled using DAPI dye (blue). A representative field (40×) is shown. (D) Cells (104 cells/well),obtained from human islet primary cultures, were incubated for 4 days in CMRL medium containing 0.5% FCS and laminin-1 (10 �g/mL) supplemented with eitherrhPRL (300 ng/mL) or vehicle (control). The cells were labeled with antibodies to insulin or Pdx1. White arrows indicate cells negative for the staining. Nuclei werelabeled using DAPI dye (blue). A representative field (40×) is shown.
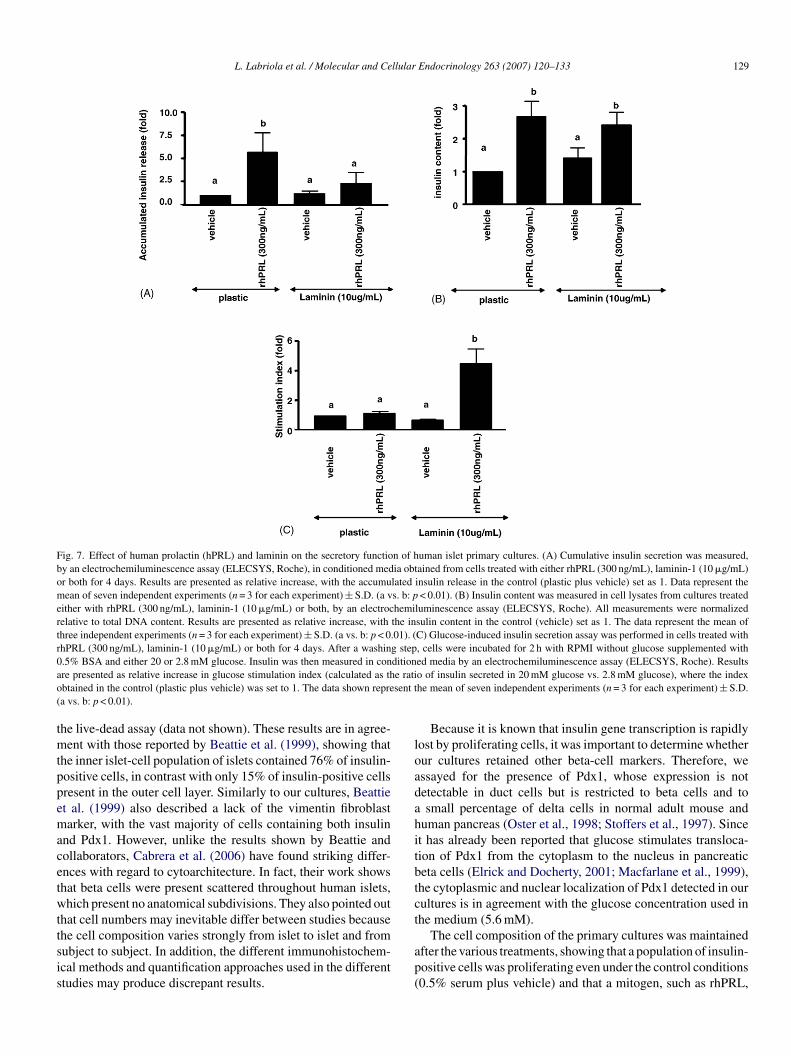
L. Labriola et al. / Molecular and Cellular Endocrinology 263 (2007) 120–133 129
Fig. 7. Effect of human prolactin (hPRL) and laminin on the secretory function of human islet primary cultures. (A) Cumulative insulin secretion was measured,by an electrochemiluminescence assay (ELECSYS, Roche), in conditioned media obtained from cells treated with either rhPRL (300 ng/mL), laminin-1 (10 �g/mL)or both for 4 days. Results are presented as relative increase, with the accumulated insulin release in the control (plastic plus vehicle) set as 1. Data represent themean of seven independent experiments (n = 3 for each experiment) ± S.D. (a vs. b: p < 0.01). (B) Insulin content was measured in cell lysates from cultures treatedeither with rhPRL (300 ng/mL), laminin-1 (10 �g/mL) or both, by an electrochemiluminescence assay (ELECSYS, Roche). All measurements were normalizedrelative to total DNA content. Results are presented as relative increase, with the insulin content in the control (vehicle) set as 1. The data represent the mean ofthree independent experiments (n = 3 for each experiment) ± S.D. (a vs. b: p < 0.01). (C) Glucose-induced insulin secretion assay was performed in cells treated withrhPRL (300 ng/mL), laminin-1 (10 �g/mL) or both for 4 days. After a washing step, cells were incubated for 2 h with RPMI without glucose supplemented with0 ditiona e ratio sent t(
tmtppemacetwttsis
loadahitbtct
.5% BSA and either 20 or 2.8 mM glucose. Insulin was then measured in conre presented as relative increase in glucose stimulation index (calculated as thbtained in the control (plastic plus vehicle) was set to 1. The data shown reprea vs. b: p < 0.01).
he live-dead assay (data not shown). These results are in agree-ent with those reported by Beattie et al. (1999), showing that
he inner islet-cell population of islets contained 76% of insulin-ositive cells, in contrast with only 15% of insulin-positive cellsresent in the outer cell layer. Similarly to our cultures, Beattiet al. (1999) also described a lack of the vimentin fibroblastarker, with the vast majority of cells containing both insulin
nd Pdx1. However, unlike the results shown by Beattie andollaborators, Cabrera et al. (2006) have found striking differ-nces with regard to cytoarchitecture. In fact, their work showshat beta cells were present scattered throughout human islets,hich present no anatomical subdivisions. They also pointed out
hat cell numbers may inevitable differ between studies because
he cell composition varies strongly from islet to islet and fromubject to subject. In addition, the different immunohistochem-cal methods and quantification approaches used in the differenttudies may produce discrepant results.ap(
ed media by an electrochemiluminescence assay (ELECSYS, Roche). Resultso of insulin secreted in 20 mM glucose vs. 2.8 mM glucose), where the indexhe mean of seven independent experiments (n = 3 for each experiment) ± S.D.
Because it is known that insulin gene transcription is rapidlyost by proliferating cells, it was important to determine whetherur cultures retained other beta-cell markers. Therefore, wessayed for the presence of Pdx1, whose expression is notetectable in duct cells but is restricted to beta cells and tosmall percentage of delta cells in normal adult mouse and
uman pancreas (Oster et al., 1998; Stoffers et al., 1997). Sincet has already been reported that glucose stimulates transloca-ion of Pdx1 from the cytoplasm to the nucleus in pancreaticeta cells (Elrick and Docherty, 2001; Macfarlane et al., 1999),he cytoplasmic and nuclear localization of Pdx1 detected in ourultures is in agreement with the glucose concentration used inhe medium (5.6 mM).
The cell composition of the primary cultures was maintainedfter the various treatments, showing that a population of insulin-ositive cells was proliferating even under the control conditions0.5% serum plus vehicle) and that a mitogen, such as rhPRL,

130 L. Labriola et al. / Molecular and Cellular Endocrinology 263 (2007) 120–133
Fig. 8. Protein expression and tyrosine phosphorylation levels of JAK2, STATs 1, 3 and 5. Primary cultures of human pancreatic islets, plated in the absence oflaminin, and cultured in CMRL supplemented with 0.5% FCS were treated with rhPRL for 10 min, 30 min or 2 h. Cells were then lysed and STATs 1 (a), 3 (b), 5(c) and JAK2 (d) were immunoprecipitated from 1 mg of total proteins. Immunocomplexes were subjected to SDS–PAGE and analyzed by Western blotting with ananti-pTYR mAb. The membranes were stripped and re-probed with the same antibodies that were used for the immunoprecipitations to confirm that nearly equala e of aa . b: p
icaotataec
bhetpwt8ase
p
pn
aeimgbotbnp
srlto
mounts of each protein were loaded. The immunoblots shown are representativs the mean ± S.E.M. of three independent experiments (a vs. b: p < 0.001, c vs
ncreased their proliferation. Analysis of insulin content in theseultures by quantitative real-time PCR, immunofluorescencend electrochemiluminescence assays demonstrated the abilityf these cells to store significant amounts of insulin. It is impor-ant to note that the pattern of response was very reproduciblemong the seven different islet preparations tested. However,he high variability (up to 10 times) of the absolute values ofccumulated insulin release and insulin content between differ-nt islet preparations forced us to express the data relative to theontrol cultures (plastic and vehicle).
These results are in agreement with those reported presentedy Hayek et al. (1995) showing that most beta cells in theuman pancreas retain the capacity to replicate in vitro whenxposed to mitogenic stimuli, such as those provided by hepa-ocyte growth factor HGF/SF and 804G matrix, which containsredominantly laminin-5. However, unlike Hayek et al. (1995),e found a higher rate of proliferation upon rhPRL treatment
han these authors obtained upon treatment with HGF/SF and04G matrix, since approximately 20% of cells were BrdU-nd insulin- or Pdx1-positive. The different media composition,
erum concentration and matrix components used here mightxplain the differences observed in cell proliferation rates.Recently, Beattie et al. (2002) also reported an increase inroliferation of human adult islets growing in fibrin gels in the
rred
total of three independent experiments. The densitometric results are presented< 0.001). W: Western blot. IP: immunoprecipitation.
resence of growth factors, such as HGF/SF, hrFGF4 and nicoti-amide, without loss of cell function.
Regarding the effects of PRL on islet-cell biology, our resultsre in agreement with early investigations performed in rat mod-ls, showing that long-term PRL stimulation of islets in vitro andn vivo upregulates glucose metabolism and associated enzy-
atic pathways (Weinhaus et al., 1996), and stimulates beta-cellrowth and insulin secretion (Sorenson et al., 1997). As reportedy Brelje et al. (2002), the PRL receptor (PRLR) was detectednly in the insulin-containing beta cells of rat islets. If this pat-ern were maintained in human adult islets, our cultures shoulde more responsive to rhPRL treatment. Accordingly, the phe-otype of the predominant beta-cell type in the expanded cellopulations is consistent with a beta-cell origin.
In agreement with our results, Brelje et al. (1993, 2004) havehown that continuous treatment with PRL for 1 or 7 days led,espectively, to a two- or five-fold increase in rat islet-cell pro-iferation. Even if we cannot completely rule out the possibilityhat a small percentage of the generated cells could be the resultf a differentiation process from ductal or other precursor cells
esiding within the islet itself, Dor et al. (2004) have recentlyeported that, in the mouse, beta-cell formation from undiffer-ntiated precursors ceases at a specific time during embryonicevelopment or early postnatal life and that new beta cells arise
llular
fs
cstToitpdttMsdwie
ta(sft(wtltwstaFcwl(
ihS
raoici(
toa
ig
rpoUti1
tip(
ptbiatrvpr
nnra
sio
mtn
A
alStTdCwU
L. Labriola et al. / Molecular and Ce
rom pre-existing beta cells. These authors also pointed out theignificant proliferative potential of differentiated beta cells.
Furthermore, it has recently been demonstrated, in primaryultures of rat islets, that PRL has the ability to induce DNAynthesis and to prevent long-term glucose-induced desensitiza-ion of insulin secretory responses (Tian and Laychock, 2003).he present study shows a PRL-induced increase in the numberf insulin-positive and Pdx1-positive cells, as shown by BrdUncorporation and insulin or Pdx1 labeling. However, rhPRLreatment could not prevent the desensitization process in ourrimary cultures of human islets maintained on uncoated plasticishes. Since the loss of islet-stabilizing factors, such as cell-o-cell or cell-to-substrate interactions, seems to induce cellso change their commitment (Bosco et al., 1989; Salomon and
eda, 1986; Schmied et al., 2000) and it has been widely demon-trated that the extracellular matrix plays a crucial role in cellifferentiation through rearrangement of the cytoskeletal net-ork (Levenberg et al., 2002), we decided to culture the cells
n the presence of an ECM protein (laminin-1 purified from thextracellular matrix of a mouse sarcoma cell line).
Laminin-1 is reported to be involved in duct lineage selec-ion in the development of mouse embryonic pancreas (Crisera etl., 2000) and to promote beta-cell differentiation in cell cultureJiang et al., 1999). This is consistent with our findings, whichhow that addition of laminin to islet-cell cultures is essentialor the organization of cell clusters presenting a high propor-ion of insulin- and Pdx1-positive cells. However, Gao et al.2003) reported that formation of three-dimensional structuresas arrested when adult pancreatic duct cells were cultured in
he presence of laminin-rich matrix produced by the 804G celline. The different cell population and source of laminin used inhe present study may be the reason for the cellular response thate observed. We have shown that rhPRL is able to increase the
ize of the cell clusters formed in the presence of laminin andhis might be the cause for recovery of the secretory function,
result, which is in agreement with previously reported data.urthermore, it has been shown that disruption of the normalell-to-cell contact is associated with impaired insulin secretion,hereas preservation or the re-establishment of the intercellu-
ar contacts within clusters restores an almost normal functionHalban et al., 1982; Weir et al., 1984).
Meda and collaborators have shown, in isolated rat pancreaticslets, that cell-to cell adhesion and/or junction coupling regulateormone secretion of individual beta cells (Bosco et al., 1989;alomon and Meda, 1986).
Recently, rat beta cells were allowed to attach to the laminin-ich matrix produced by the 804G rat carcinoma cell line, withn increase in insulin secretory response to secretagogues beingbserved (Bosco et al., 2000). These results demonstrate thatslet-cell–matrix interactions can improve the sensitivity of betaells to glucose, suggesting that outside-in signaling throughntegrins plays a major role in regulation of beta-cell functionBosco et al., 2000).
The role of extracellular matrix was also related to the long-erm survival of pancreatic beta cells. In fact, by using a modelf rat pancreatic islet cultured on 804G matrix, Hammar etl. (2004) have shown that this matrix protects against IL-1�-
R
A
Endocrinology 263 (2007) 120–133 131
nduced cell death, by inducing the expression of anti-apoptoticenes.
As an initial step in the study of the molecular mechanisms ofhPRL action in human islets, we have shown, for the first time inrimary cultures of human islets, the expression and activationf JAK2 and STATs 1, 3 and 5, the main targets of rhPRL action.nderstanding the role of PRL in beta-cell regulation is impor-
ant since PRL is a major pregnancy hormone, and increasednsulin secretion is observed during pregnancy (Sorenson et al.,997).
The relevance of describing the presence and the activa-ion state of JAK2 and STAT family protein-family membersn human islets lies in the fact that in rat and mouse models, thisathway is involved in cell proliferation and insulin secretionCarter-Su et al., 1996).
We were also able to describe three different time-dependenthosphorylation profiles upon rhPRL stimulation. The long-erm activation of STAT5, observed in our experiments, mighte related to the increase in islet function since, at least in ratslets, STAT5B induced an increase in the expression of PRLRfter continuous PRL stimulation, which was essential to main-ain the long-term activation of STAT5B (Brelje et al., 2004). Itemains to be elucidated whether these different kinetics of acti-ation are present in free-floating islets. These different kineticrofiles might be in part responsible for the different cellularesponses induced by rhPRL stimulation.
In summary, the results presented here lead us to propose aew culture protocol for adult human pancreatic islets that allowsot only cell proliferation, achieving a final three-fold expansionate of insulin-positive cells in 4 days, but also insulin storage andppropriate insulin release in response to physiological signals.
We also describe, for the first time, as an initial step in thetudy of molecular mechanisms involved in PRL action in humanslets, the presence and activation of JAK2 and STAT proteins,ne of the principal pathways involved in the action of PRL.
Further studies along these lines may lead to the develop-ent of new strategies for increasing the islet mass available for
ransplantation and even to induce regeneration of the endoge-ous beta-cell mass in diabetic patients.
cknowledgements
We are especially grateful to Prof. Dr. Celia R. da Silva Garciand Marcos L. Gazarini Dutra (Laboratorio de Biologia Celu-ar e Molecular do Plasmodium, University of the Sao Paulo,P, Brazil) for help with the confocal microscopy. We sincerely
hank Prof. Dr. S. Efrat for critical reading of the manuscript.he excellent technical assistance provided by Patricia Barrosos Santos, Irenice Cairo da Silva, Zizi de Mendonca, Deboraosta and Sandra Regina Souza is also acknowledged. This workas supported by grants from FAPESP, CNPq, FINEP and PRP-SP.
eferences
sfari, M., De, W., Postel-Vinay, M.C., Czernichow, P., 1995. Expressionand regulation of growth hormone (GH) and prolactin (PRL) receptors

1 llular
B
B
B
B
B
B
B
B
B
B
B
C
C
C
C
D
E
F
F
F
G
H
H
H
J
J
K
K
L
L
L
L
L
L
M
M
N
N
O
P
32 L. Labriola et al. / Molecular and Ce
in a rat insulin producing cell line (INS-1). Mol. Cell. Endocrinol. 107,209–214.
aker, S.E., DiPasquale, A.P., Stock, E.L., Quaranta, V., Fitchmun, M., Jones,J.C., 1996. Morphogenetic effects of soluble laminin-5 on cultured epithelialcells and tissue explants. Exp. Cell Res. 228, 262–270.
eattie, G.M., Itkin-Ansari, P., Cirulli, V., Leibowitz, G., Lopez, A.D., Bossie, S.,Mally, M.I., Levine, F., Hayek, A., 1999. Sustained proliferation of PDX1+cells derived from human islets. Diabetes 48, 1013–1019.
eattie, G.M., Montgomery, A.M., Lopez, A.D., Hao, E., Perez, B., Just, M.L.,Lakey, J.R., Hart, M.E., Hayek, A., 2002. A novel approach to increasehuman islet cell mass while preserving beta-cell function. Diabetes 51,3435–3439.
ole-Feysot, C., Goffin, V., Edery, M., Binart, N., Kelly, P.A., 1998. Prolactin(PRL) and its receptor: actions, signal transduction pathways and phenotypesobserved in PRL receptor knockout mice. Endocr. Rev. 19, 225–268.
osco, D., Meda, P., Halban, P.A., Rouiller, D.G., 2000. Importance of cell-matrix interactions in rat islet beta-cell secretion in vitro: role of alpha6beta1integrin. Diabetes 49, 233–243.
osco, D., Orci, L., Meda, P., 1989. Homologous but not heterologous contactincreases the insulin secretion of individual pancreatic B-cells. Exp. CellRes., 184.
relje, T.C., Parsons, J.A., Sorenson, R.L., 1994. Regulation of islet beta-cellproliferation by prolactin in rat islets. Diabetes 43, 263–273.
relje, T.C., Scharp, D.W., Lacy, P.E., Ogren, L., Talamantes, F., Robertson,M., Friesen, H.G., Sorenson, R.L., 1993. Effect of homologous placentallactogens, prolactins, and growth hormones on islet B-cell division andinsulin secretion in rat, mouse, and human islets: implication for placentallactogen regulation of islet function during pregnancy. Endocrinology 132,879–887.
relje, T.C., Sorenson, R.L., 1991. Role of prolactin versus growth hormone onislet B-cell proliferation in vitro: implications for pregnancy. Endocrinology128, 45–57.
relje, T.C., Stout, L.E., Bhagroo, N.V., Sorenson, R.L., 2004. Distinctive rolesfor prolactin and growth hormone in the activation of STAT5 in pancreaticislets of Langerhans. Endocrinology 145, 4162–4175.
relje, T.C., Svensson, A.M., Stout, L.E., Bhagroo, N.V., Sorenson, R.L., 2002.An immunohistochemical approach to monitor the prolactin-induced acti-vation of the JAK2/STAT5 pathway in pancreatic islets of Langerhans. J.Histochem. Cytochem. 50, 365–383.
abrera, O., Berman, D.M., Kenyon, N.S., Ricordi, C., Berggren, P.O., Caicedo,A., 2006. The unique cytoarchitecture of human pancreatic islets has impli-cations for islet cell function. Proc. Natl. Acad. Sci. U.S.A. 103, 2334–2339.
arter-Su, C., Schwartz, J., Smit, L.S., 1996. Molecular mechanism of growthhormone action. Annu. Rev. Physiol. 58, 187–207.
olognato, H., Yurchenco, P.D., 2000. Form and function: the laminin familyof heterotrimers. Develop. Dyn. 218, 213–234.
risera, C.A., Kadison, A.S., Breslow, G.D., Maldonado, T.S., Longaker, M.T.,Gittes, G.K., 2000. Expression and role of laminin-1 in mouse pancreaticorganogenesis. Diabetes 49, 936–944.
or, Y., Brown, J., Martinez, O.I., Melton, D.A., 2004. Adult pancreatic betacells are formed by self-duplication rather than stem-cell differentiation.Nature 429, 41–46.
lrick, L.J., Docherty, K., 2001. Phosphorylation-dependent nucleocytoplasmicshuttling of pancreatic duodenal homeobox-1. Diabetes 50, 2244–2252.
leenor, D., Petryk, A., Driscoll, P., Freemark, M., 2000. Constitutive expressionof placental lactogen in pancreatic beta cells: effects on cell morphology,growth, and gene expression. Pediatr. Res. 47, 136–142.
raga, D.W., Sabek, O., Hathaway, D.K., Gaber, A.O., 1998. A comparison ofmedia supplement methods for the extended culture of human islet tissue.Transplantation 65, 1060–1066.
reemark, M., Avril, I., Fleenor, D., Driscoll, P., Petro, A., Opara, E., Kendall,W., Oden, J., Bridges, S., Binart, N., Breant, B., Kelly, P.A., 2002. Targeteddeletion of the PRL receptor: effects on islet development, insulin produc-
tion, and glucose tolerance. Endocrinology 143, 1378–1385.ao, R., Ustinov, J., Pulkkinen, M.A., Lundin, K., Korsgren, O., Otonkoski,T., 2003. Characterization of endocrine progenitor cells and critical factorsfor their differentiation in human adult pancreatic cell culture. Diabetes 52,2007–2014.
P
Endocrinology 263 (2007) 120–133
alban, P.A., Wollheim, C.B., Blondel, B., Meda, P., Niesor, E.N., Mintz, D.H.,1982. The possible importance of contact between pancreatic islet cells forthe control of insulin release. Endocrinology 111, 86–94.
ammar, E., Parnaud, G., Bosco, D., Perriraz, N., Maedler, K., Donath, M.,Rouiller, D.G., Halban, P.A., 2004. Extracellular matrix protects pancreaticbeta-cells against apoptosis: role of short- and long-term signaling pathways.Diabetes 53, 2034–2041.
ayek, A., Beattie, G.M., Cirulli, V., Lopez, A.D., Ricordi, C., Rubin, J.S.,1995. Growth factor/matrix-induced proliferation of human adult beta-cells.Diabetes 44, 1458–1460.
iang, F.X., Cram, D.S., DeAizpurua, H.J., Harrison, L.C., 1999. Laminin-1promotes differentiation of fetal mouse pancreatic beta-cells. Diabetes 48,722–730.
iang, F.X., Naselli, G., Harrison, L.C., 2002. Distinct distribution of lamininand its integrin receptors in the pancreas. J. Histochem. Citochem. 50,1625–1632.
awaguchi, Y., Cooper, B., Gannon, M., Ray, M., MacDonald, R.J., Wright,C.V., 2002. The role of the transcriptional regulator Ptf1a in convertingintestinal to pancreatic progenitors. Nat. Genet. 32, 128–134.
leinman, H.K., McGarvey, M.L., Liotta, L.A., Robey, P.G., Tryggvason, K.,Martin, G.R., 1982. Isolation and characterization of type IV procollagen,laminin, and heparan sulfate proteoglycan from the EHS sarcoma. Biochem-istry 21, 6188–6193.
awson, D.M., Sensui, N., Haisknlender, D.H., Gala, R.R., 1982. Rat lymphomabioassay for prolactin: observations on its use and comparison with radioim-munoassay. Life Sci. 31, 3063–3070.
efebvre, V.H., Otonkoski, T., Ustinov, J., Huotari, M.A., Pipeleers, D.G.,Bouwens, L., 1998. Culture of adult human islet preparations with hepa-tocyte growth factor and 804G matrix is mitogenic for duct cells but not forbeta-cells. Diabetes 47, 134–137.
ekanne Deprez, R.H., Fijnvandraat, A.C., Ruijter, J.M., Moorman, A.F., 2002.Sensitivity and accuracy of quantitative real-time polymerase chain reactionusing SYBR green I depends on cDNA synthesis conditions. Anal. Biochem.307, 63–69.
elievre, S., Weaver, V.M., Bissell, M.J., 1996. Extracellular matrix signalingfrom the cellular membrane skeleton to the nuclear skeleton: a model of generegulation. Recent Prog. Horm. Res. 51, 417–432.
evenberg, S., Golub, J.S., Amit, M., Itskovitz-Eldor, J., Langer, R., 2002.Endothelial cells derived from human embryonic stem cells. Proc. Natl.Acad. Sci. U.S.A. 99, 4391–4396.
evy, D.E., Darnell, J.E.J., 2002. Stats: transcriptional control and biologicalimpact. Nat. Rev. Mol. Cell Biol. 3, 651–662.
acfarlane, W.M., McKinnon, C.M., Felton-Edkins, Z.A., Cragg, H., James,R.F., Docherty, K., 1999. Glucose stimulates translocation of the home-odomain transcription factor PDX1 from the cytoplasm to the nucleus inpancreatic beta-cells. J. Biol. Chem. 274, 1011–1016.
aria-Engler, S.S., Correa-Giannella, M.L.C., Labriola, L., Krogh, K., Colin,C., Lojudice, F.H., Aita, C.A., Oliveira, E.M.C., Correa, T.C.S., Silva,I.C., Genzini, T., Miranda, M.P., Noronha, I.L., Vilela, L., Coimbra, C.N.,Mortara, R.A., Mares Guia, M., Eliaschewitz, F.G., Sogayar, M.C., 2004.Co-localization of nestin and insulin and expression of islet cell markersin long-term human pancreatic nestin positive cells. J. Endocrinol. 183,455–467.
ielsen, J.H., Galsgaard, E.D., Moldrup, A., Friedrichsen, B.N., Billestrup, N.,Hansen, J.A., Lee, Y.C., Carlsson, C., 2001. Regulation of beta-cell mass byhormones and growth factors. Diabetes 50 (Suppl. 1), S25–S29.
ielsen, J.H., Moldrup, A., Billestrup, N., Petersen, E.D., Allevato, G., Stahl,M., 1992. The role of growth hormone and prolactin in beta cell growth andregeneration. Adv. Exp. Med. Biol. 321, 9–20.
ster, A., Jensen, J., Serup, P., Galante, P., Madsen, O.D., Larsson, L.I., 1998.Rat endocrine pancreatic development in relation to two homeobox geneproducts (Pdx-1 and Nkx 6.1). J. Histochem. Cytochem. 46, 707–715.
ereira, C.A., Pouliquen, Y., Rodas, V., Massotte, D., Mortensen, C., Sogayar,
M.C., Menissier-de Murcia, J., 2001. Optimized insect cell culture for theproduction of recombinant heterologous proteins and baculovirus particles.Biotechniques 31, 1262–1268.faffl, M.W., 2001. A new mathematical model for relative quantification inreal-time RT-PCR. Nucl. Acids Res. 29, e45.

llular
P
R
R
S
S
S
S
S
S
S
S
T
W
W
W
L. Labriola et al. / Molecular and Ce
iper, K., Brickwood, S., Turnpenny, L.W., Cameron, I.T., Ball, S.G., Wilson,D.I., Hanley, N.A., 2004. Beta cell differentiation during early human pan-creas development. J. Endocrinol. 181, 11–23.
asmussen, L.D., Ekelund, F., Hansen, L.H., Sorensen, S.J., Johnsen, K., 2001.Group-specific PCR primers to amplify 24S a-subunit rRNA genes fromkinetoplastida (Protozoa) used in denaturing gradient gel electrophoresis.Microb. Ecol. 42, 109–115.
icordi, C., Lacy, P.E., Finke, E.H., Olack, B.J., Scharp, D.W., 1988. Automatedmethod for isolation of human pancreatic islets. Diabetes 37, 413–420.
alomon, D., Meda, P., 1986. Heterogeneity and contact-dependent regulationof hormone secretion by individual B cells. Exp. Cell Res. 162, 507–520.
apin, R., Le Galudec, V., Gasser, F., Pinget, M., Grucker, D., 2001. Elecsysinsulin assay: free insulin determination and the absence of cross-reactivitywith insulin lispro. Clin. Chem. 47, 602–605.
chmied, B.M., Ulrich, A., Matsuzaki, H., Ding, X., Ricordi, C., Moyer, M.P.,Batra, S.K., Adrian, T.E., Pour, P.M., 2000. Maintenance of human islets inlong-term culture. Differentiation 66, 173–180.
hapiro, A.M., Lakey, J.R., Ryan, E.A., Korbutt, G.S., Toth, E., Warnock, G.L.,Kneteman, N.M., Rajotte, R.V., 2000. Islet transplantation in seven patientswith type 1 diabetes mellitus using a glucocorticoid-free immunosuppressiveregimen. N. Engl. J. Med. 343, 230–238.
ogayar, M.C., Camargo, A.A., Bettoni, F., Carraro, D.M., Pires, L.C., Parmi-giani, R.B., Ferreira, E.N., de Sa Moreira, E., do Rosario, D., de O Latorre,M., Simpson, A.J., Cruz, L.O., Degaki, T.L., Festa, F., Massirer, K.B., Filho,F.C., Camargo, L.P., Cunha, M.A., De Souza, S.J., Faria, M.J., Giuliatti,S., Kopp, L., de Oliveira, P.S., Paiva, P.B., Pereira, A.A., Pinheiro, D.G.,Puga, R.D., S de Souza, J.E., Albuquerque, D.M., Andrade, L.E., Baia, G.S.,Briones, M.R., Cavaleiro-Luna, A.M., Cerutti, J.M., Costa, F.F., Costanzi-Strauss, E., Espreafico, E.M., Ferrasi, A.C., Ferro, E.S., Fortes, M.A.,
Furchi, J.R., Giannella-Neto, D., Goldman, G.H., Goldman, M.H., Gruber,A., Guimaraes, G.S., Hackel, C., Henrique-Silva, F., Kimura, E.T., Leoni,S.G., Macedo, C., Malnic, B., Manzini, B.C.V., Marie, S.K., Martinez-Rossi,N.M., Menossi, M., Miracca, E.C., Nagai, M.A., Nobrega, F.G., Nobrega,M.P., Oba-Shinjo, S.M., Oliveira, M.K., Orabona, G.M., Otsuka, A.Y., Paco-W
Endocrinology 263 (2007) 120–133 133
Larson, M.L., Paixao, B.M., Pandolfi, J.R., Pardini, M.I., Passos Bueno,M.R., Passos, G.A., Pesquero, J.B., Pessoa, J.G., Rahal, P., Rainho, C.A.,Reis, C.P., Ricca, T.I., Rodrigues, V., Rogatto, S.R., Romano, C.M., Romeiro,J.G., Rossi, A., Sa, R.G., Sales, M.M., Sant’Anna, S.C., Santarosa, P.L.,Segato, F., Silva, W.A.J., Silva, I.D., Silva, N.P., Soares-Costa, A., Sonati,M.F., Strauss, B.E., Tajara, E.H., Valentini, S.R., Villanova, F.E., Ward, L.S.,Zanette, D.L., Initiative, L.-F.T.F., 2004. A transcript finishing initiative forclosing gaps in the human transcriptome. Genome Res. 14, 1413–1423.
orenson, R.L., Brelje, T.C., Roth, C., 1997. Adaptation of islets of Langerhansto pregnancy: beta-cell growth, enhanced insulin secretion and the role oflactogenic hormones. Horm. Metab. Res. 29, 301–307.
toffers, D.A., Thomas, M.K., Habener, J.F., 1997. Homeodomain protein IDX-1: a master regulator of pancreas development and insulin gene expression.Trends Endocrinol. Metab. 8, 145–151.
wenne, I., Hill, D.J., Strain, A.J., Milner, R.D., 1987. Effects of human placentallactogen and growth hormone on the production of insulin and somatomedinC/insulin-like growth factor I by human fetal pancreas in tissue culture. J.Endocrinol. 113, 297–303.
ian, Y., Laychock, S.G., 2003. Prolactin regulates adenylyl cyclase and insulinsecretion in rat pancreatic islets. Mol. Cell. Endocrinol. 204, 75–84.
ang, R.N., Rosenberg, L., 1999. Maintenance of beta-cell function and sur-vival following islet isolation requires re-establishment of the islet–matrixrelationship. J. Endocrinol. 163, 181–190.
eber, M., Deng, S., Kucher, T., Shaked, A., Ketchum, R.J., Brayman, K.L.,1997. Adenoviral transfection of isolated pancreatic islets: a study ofprogrammed cell death (apoptosis) and islet function. J. Surg. Res. 69,23–32.
einhaus, A.J., Stout, L.E., Sorenson, R.L., 1996. Glucokinase, hexokinase,glucose transporter 2, and glucose metabolism in islets during pregnancy
and prolactin-treated islets in vitro: mechanisms for long term up-regulationof islets. Endocrinology 137, 1640–1649.eir, G.C., Halban, P.A., Meda, P., Wollheim, C.B., Orci, L., Renold, A.E.,1984. Dispersed adult rat pancreatic islet cells in culture: A, B, and D cellfunction. Metabolism 33, 447–453.
Copyright © 2022 FDOKUMEN




















