
Beneficial effects of prolactin and laminin on human pancreatic islet-cell cultures

β-catenin signalling in mesenchymal islet-derived precursor cells
L. Ikonomou, E. Geras-Raaka, B. M. Raaka, and M. C. GershengornClinical Endocrinology Branch, National Institute of Diabetes and Digestive and Kidney Diseases,National Institutes of Health
AbstractObjectives—Previously, we characterized human islet-derived precursor cells (hIPCs) asmesenchymal stem cells that migrate out from islets in vitro and can differentiate into functionalislet-like structures following proliferative expansion. Here, we investigate the role of β-cateninsignalling in derivation and proliferation of hIPCs.
Materials and methods—Localization of β-catenin was performed using confocal microscopy.Expression levels of β-catenin target genes were measured by quantitative real-time polymerase chainreaction. Loss-of-function studies were performed using specific short interfering RNAs.
Results—Immunostaining of islet outgrowths revealed translocation of β-catenin from plasmamembranes in intact islets to the nucleus in cells migrating out. There were no nuclear β-catenin-positive cells in intact islets whereas between 35% and 70% of cells in established hIPC culturesexhibited nuclear β-catenin. Transcripts for β-catenin target genes were increased in hIPCs comparedto those in islets. β-Catenin translocated to the cell membrane when hIPCs formed epithelial cellclusters. In proliferating hIPCs, there was a strong correlation between markers of proliferation andnuclear β-catenin. Treatment of hIPCs with the glycogen synthase kinase-3β inhibitor (2′Z,3′E)-6-Bromoindirubin-3′-oxime increased intracellular β-catenin but reduced nuclear β-catenin, and wasassociated with reduced cell proliferation. Finally, knockdown of β-catenin decreased β-catenintarget gene expression and hIPC proliferation.
Conclusions—These results support a functional role for β-catenin during proliferation of hIPCsand suggest that activated β-catenin signalling may also be important during hIPC derivation fromislets.
IntroductionType 1 diabetes is a debilitating autoimmune disease that results in dysfunction of glucosehomeostasis (Bach 1994). A shortage of donors for islet transplantation has brought aboutefforts to isolate and expand progenitor cells from the pancreas (Nir & Dor 2005). Our grouphas characterized a possible mesenchymal-like progenitor cell derived, in vitro, from humanislets (Gershengorn et al. 2004), and a number of laboratories have reported derivation ofsimilar mesenchymal progenitor cell populations from islets (Gao et al. 2005; Lechner et al.2005; Ouziel-Yahalom et al. 2006). Although we previously suggested that these progenitorcells may arise by dedifferentiation of islet endocrine cells by epithelial-to-mesenchymaltransition (Gershengorn et al. 2004), recent work from several laboratories including ours(Atouf et al. 2007; Chase et al. 2007; Morton et al. 2007; Weinberg et al. 2007) suggests thatthese mesenchymal precursors may not be derived from β-cells. This conclusion, however, isbased on studies with mouse cultures and may not apply to human cells. Nevertheless, it is
Correspondence: Marvin C. Gershengorn, Clinical Endocrinology Branch, National Institute of Diabetes and Digestive and KidneyDiseases, National Institutes of Health, 50 South Dr., Rm. 4134, Bethesda, MD 20892-8029, USA. Tel.: 301-451-6305; Fax:301-480-4214; E-mail: E-mail: [email protected].
NIH Public AccessAuthor ManuscriptCell Prolif. Author manuscript; available in PMC 2009 June 1.
Published in final edited form as:Cell Prolif. 2008 June ; 41(3): 474–491. doi:10.1111/j.1365-2184.2008.00527.x.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

perhaps more likely that these precursor cells may represent a pancreas-specific mesenchymalstem cell (MSC) population (Chase et al. 2007; Davani et al. 2007). In any case, the pathwaysthat control the transitions of these mesenchymal-like progenitors to cells with an epithelialphenotype remain largely unknown.
One pathway essential for epithelial-to-mesenchymal transition (Huber et al. 2005; Thiery &Sleeman 2006) and for MSC proliferation (Boland et al. 2004; Etheridge et al. 2004; Cho etal. 2006) in other systems is the canonical Wnt/β-catenin pathway. When Wnt signalling isabsent, β-catenin is part of the ‘destruction complex’ that is composed of scaffolding proteinssuch as adenomatous polyposis coli (APC) and axin, and serine/threonine kinases such asglycogen synthase kinase (GSK)-3β and CK1. This complex hyperphosphorylates β-cateninand marks it for proteasomal degradation (Aberle et al. 1997). Wnt signalling, which can beactivated by Wnt ligand binding to frizzled receptors and LRP5/6 co-receptors (Logan & Nusse2004), inhibits phosphorylation of β-catenin causing it to accumulate in the cytoplasm andsubsequently to translocate to the nucleus. There, β-catenin can associate with members of theT-cell factor/lymphocyte enhancer factor (TCF/LEF) family of DNA binding proteins andinitiate transcription of Wnt-responsive genes. β-catenin signalling can also be activated in aWnt-independent way by growth factor signalling via receptor tyrosine kinases (Monga etal. 2002; Muller et al. 2002) or by cell adhesion using the integrin-linked kinase (Novak etal. 1998; Oloumi et al. 2004). Because β-catenin may also participate in the formation of cell–cell (adherens) junctions by its association with type 1 cadherins, it is clear that its subcellularlocation can dramatically affect processes such as cell differentiation, proliferation andadhesion (Jamora et al. 2003; Nelson & Nusse 2004). In in vitro cell systems, several reportsshow that incubation with growth factors such as insulin-like growth factor-II, epidermalgrowth factor and hepatocyte growth factor induce β-catenin nuclear translocation (Morali etal. 2001; Muller et al. 2002; Lu et al. 2003). Furthermore, recent work shows that vascularsmooth muscle cells exhibit nuclear activation of β-catenin in both in vivo and in vitro modelsof arterial injury (Wang et al. 2002; George & Beeching 2006; Quasnichka et al. 2006).
Because human islet-derived precursor cells (hIPCs) originate during in vitro culture of isletsin a growth factor-rich environment, we hypothesized that β-catenin may play a role in thisprocess. We observed clear transition of β-catenin from surface membrane localization in intactislets to nuclear localization in proliferating mesenchymal-like hIPCs. To investigate whetherthe observed β-catenin translocation activated Wnt responses, we measured mRNA and proteinlevels for known Wnt target genes. Up-regulation of genes such as DKK1, BIRC5, CCND1and FZD7 occurred during transition from islets to hIPCs. Short interfering RNA (siRNA)-mediated knockdown of β-catenin in hIPCs resulted in reduced 5-bromo-2′-deoxyuridine(BrdU) incorporation, indicating that β-catenin signalling controls, at least partially, hIPCproliferation. When hIPCs were induced to cluster and differentiate to epithelial-like cells thatdo not proliferate, β-catenin translocated to the cell membrane. This study, along with ourrecent work (Davani et al. 2007), is consistent with the idea that proliferative hIPCs aremesenchymal-like progenitor cells that exhibit activated β-catenin signalling.
Materials and MethodsReagents
All chemicals were obtained from Sigma-Aldrich (St. Louis, MO, USA) unless otherwiseindicated.
Cell cultureHuman islet-derived precursor cells were maintained in 150-mm tissue culture dishes (Falcon,Becton Dickinson Labware, Franklin Lakes, NJ, USA) in growth medium CMRL-1066 (Gibco,
Ikonomou et al. Page 2
Cell Prolif. Author manuscript; available in PMC 2009 June 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Grand Island, NY, USA) supplemented with 10% foetal bovine serum, Prime (Biosource,Rockville, MD, USA) and 2 mM L-glutamine, 100× (Gibco). Cells were passaged every 3–4days at 80–90% confluence. Briefly, medium was removed and 10 mL trypsinethylenediaminetetraacetic acid, 1× (Cellgro, Mediatech Inc., Herndon, VA, USA) was addedper dish. Trypsin was inactivated by addition of growth medium and cells were collected bycentrifugation (160 g, 5 min). They were then re-suspended in fresh growth medium, countedby means of a Vi-Cell XR analyser (Beckman Coulter Inc., Fullerton, CA, USA) and seededin new dishes at a density of 1.6 × 104/cm2. Passage 2 bone marrow-derived MSCs werepurchased from Cambrex Bio Science (Walkersville, MD, USA) and maintained in proprietarymesenchymal stem cell growth medium. Cells were passaged every 6 days and seeded at adensity of 6000 cells/cm2. Clusters of either hIPCs or MSCs were formed following seedingof 330 000 cells per well of a 6-well plate. Induction media were the respective basal media(CMRL-1066 for hIPCs and mesenchymal stem cell basal medium for mesenchymal cells)supplemented with 2 mM L-glutamine, ITS-A supplement (Invitrogen, Carlsbad, CA, USA) and1% (w/v) fatty acid-free bovine serum albumin (MP Biomedicals, Solon, OH, USA). Cellclusters were harvested by gentle pipetting 4 or 7 days after serum withdrawal, for furtherprocessing.
Quantitative real-time polymerase chain reactionThe RNeasy mini kit (Qiagen Sciences, Germantown, MD, USA) was used for total RNAextraction. hIPC pellets were re-suspended in lysis buffer and processed according to themanufacturer's protocol. Concentration of total RNA was measured using the ND-1000spectrophotometer (NanoDrop Technologies, Wilmington, DE, USA). cDNA was preparedusing the high-capacity cDNA archive kit (Applied Biosystems, Foster City, CA, USA) andquantitative real-time polymerase chain reaction (qRT-PCR) was performed in an Mx3000Pinstrument (Stratagene, La Jolla, CA, USA) using 96-well optical reaction plates and universalPCR master mix (both from Applied Biosystems). Pre-designed TaqMan gene expressionassays from Applied Biosystems were: DKK1, Hs00183740_m1; FZD7, Hs00275833_s1;BIRC5, Hs00153353_ml; MYC, Hs00153408_m1; CCND1, Hs00277039_m1; LEF1,Hs00212390_m1; WNT5A, Hs00180103_m1; FZD2, Hs00361432_s1; DKK3,Hs00247426_m1; TCF4, Hs00162613_m1; GSK3β, Hs00275656_m1; CTNNB1,Hs00170025_m1; JUP, Hs00158408_m1; CLDN3, Hs00265816_s1; CLDN4,Hs00533616_s1; CDH2, Hs00169953_m1; CDH1, Hs00170423_m1. Each PCR reactioncontained cDNA prepared from 100 ng total RNA. Cycle threshold values were normalized toeither 18S rRNA or glyceraldehyde-3-phosphate dehydrogenase (GAPDH).
DKK1 ELISAThe DKK1 ELISA was performed as previously described (Tian et al. 2003). Recombinanthuman DKK1, anti-DKK1 antibody and biotinylated anti-DKK1 antibody were obtained fromR&D Systems (Minneapolis, MN, USA).
ImmunocytochemistryRabbit antihuman C-peptide, antihuman somatostatin and antihuman glucagon (Linco, St.Charles, MO, USA), mouse antihuman β-catenin, mouse antihuman E-cadherin, and rabbitantihistone 3 (phospho S10) (Abcam, Cambridge, MA, USA) and mouse antihuman N-cadherin (BD Biosciences, San Jose, CA, USA) antibodies were used at 1 : 100 dilution inblocking buffer [4% donkey serum in Dulbecco's phosphate-buffered saline (DPBS)]. Rabbitantihuman Ki-67 (Abcam) was used at 1 : 500 dilution; Alexa-Fluor 488 and 546 F(ab')2secondary antibodies (Molecular Probes, Eugene, OR, USA) were added at 1 : 100 dilution.For staining of hIPC monolayers, cells were cultured on two-chamber Lab-Tek Permanoxslides (Nalge Nunc International, Rochester, NY, USA) and were fixed with 4%
Ikonomou et al. Page 3
Cell Prolif. Author manuscript; available in PMC 2009 June 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

paraformaldehyde (EMS, Hatfield, PA, USA) in 1× DPBS and permeabilized with chilled 50%methanol in PBS for 15 min. Whole human pancreas was fixed in 4% paraformaldehyde,embedded in paraffin wax and 10 μm sections were prepared. Embedded sections weredeparaffinized following standard procedures. For antigen retrieval, sections were incubatedin citrate buffer (10 mM sodium citrate, pH 6.0) for 20 min at 95 °C and blocking was performedwith 4% normal donkey serum in 1× DPBS for 30 min at room temperature. Slides were thenincubated with primary antibodies for 1.5–2 h at 37 °C in a humidified chamber, washedextensively with DPBS and incubated with secondary antibodies at 37 °C for 1.5–2 h. Slideswere mounted in Prolong Gold antifade reagent (Invitrogen) containing Hoechst 33342 (10μg/mL) or 4′,6-diamidino-2-phenylindole (10 μg/mL) to visualize nuclei. Confocal imageswere captured with a Zeiss LSM 510 Meta NLO laser scanning inverted microscope. Themicroscope was equipped with Argon/2 (488 nm line), HeNe (546 nm line) (Carl ZeissMicroImaging, Thornwood, NY, USA) and Titanium/Sapphire Chameleon (Coherent, SantaClara, CA, USA) lasers. Slides were viewed with either a 25×, 0.80 ImmCorr DIC or a 40×,1.3 Oil DIC or a 63×, 1.3 Oil DIC or a 100×, 1.4 Oil DIC objectives at room temperature andimages were captured using the LSM510 META software. Confocal micrographs wereconverted to TIFF format and arranged using Photoshop 6.0 (Adobe Systems Incorporated,San Jose, CA, USA).
BIO experimentsA 10-mM solution of (2′Z,3′E)-6-Bromoindirubin-3′-oxime (BIO) in dimethyl sulfoxide waspurchased from Calbiochem (San Diego, CA, USA). hIPCs were seeded in 6-well plates and2-well Permanox chambers in growth media, at a density of 1 × 104 cells/cm2 and let to attachovernight. BIO was diluted in growth media and added to the cells. For experiments measuringBrdU incorporation, BrdU was added to the culture 24 h after BIO addition for a 24-h period.Cells were either trypsin-harvested for cell count by Vi-Cell or ethanol-fixed for cell cycleanalysis (see below) or paraformaldehyde-fixed for immunocytochemistry.
Cell cycle analysisFor cell cycle analysis, hIPCs were harvested with trypsin, collected by centrifugation at 200g for 5 min and re-suspended in PBS. Cells were again collected by centrifugation,monodispersed in 0.5 mL PBS and transferred to tubes containing 4.5 mL of cold 70% ethanol;tubes were stored at −20 °C until analysis. Before analysis, ethanol-suspended cells werecollected by centrifugation at 200 g for 5 min, rinsed with PBS and re-suspended in PBScontaining propidium iodide (50 μg/mL) and RNase A (0.2 mg/mL). A FACS Caliburcytometer operated with CellQuest software (BD Biosciences) was used for data collection.Histogram analysis and calculation of G1, S and G2/M percentages was performed with ModFitLT (Verity Software House, Topsham, ME, USA).
ImmunoblottingCell pellets were dissolved in RIPA buffer (Pierce, Rockford, IL, USA) containing proteaseinhibitor cocktail (Roche Applied Science, Indianapolis, IN, USA). After addition of β-mercaptoethanol, samples were heated at 100 °C for 5 min. Proteins were resolved by 7.5%SDS-PAGE and transferred to nitrocellulose membranes and membranes were blocked with5% non-fat dry milk. After incubation with primary mouse anti-β-catenin (cat. no. 610153, BDBiosciences) and secondary antibodies, bands were visualized with SuperSignalchemiluminescent substrate (Pierce). GAPDH served as loading control.
β-Catenin siRNAValidated Stealth RNAi DuoPak β-catenin siRNAs were purchased from Invitrogen. ForsiRNA delivery by nucleofection, the Human Dermal Fibroblast-Adult Nucleofector kit
Ikonomou et al. Page 4
Cell Prolif. Author manuscript; available in PMC 2009 June 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

(Amaxa Inc., Gaithersburg, MD, USA) was used. A total of 1.0–1.5 × 106 cells per siRNAwere transfected (1.5 μg of plasmid/0.5 × 106 cells). Controls included mock-transfected cellsand cells transfected with a Stealth Negative Control Medium GC siRNA. hIPCs weretransferred to 10-cm culture dishes in growth medium. For siRNA experiments relating to β-catenin target gene expression and hIPC proliferation, cells were co-transfected with one oftwo different siRNAs directed at β-catenin mRNA or both siRNAs, or siRNA buffer and agreen fluorescent protein plasmid (1–1.5 μg of plasmid) and were seeded in 10-cm or 15-cmculture dishes. After 24 h in culture, cells were harvested, sorted by FACS and seeded in 1- or2-chamber Lab-Tek slides in growth medium. After 24 h, BrdU (Amersham Biosciences,Piscataway, NJ, USA) was added to the medium for an additional 24h incubation prior tofixation and staining as described in the ‘Immunocytochemistry’ section above. A monoclonalanti-BrdU antibody was used (Amersham Biosciences). Additional cells were harvested withtrypsin and cell pellets were stored at − 80 °C for RNA isolation, RT-PCR analysis and Westernblotting. Changes in gene expression were quantified by qRT-PCR and cycle threshold valueswere normalized to Ct = 15 of GAPDH. Fold change relative to control siRNA was calculatedbased on the 2(−ΔΔCt) method (Livak & Schmittgen 2001).
Short interfering RNA experiments were also performed using the INTERFERin deliveryreagent (Bridge Bioscience Corp., Portsmouth, NH, USA). Briefly, hIPCs were seeded in 2-chamber Lab-Tek slides or 12-well plates. Following cell attachment overnight, siRNAs werecomplexed with INTERFERin reagent according to the manufacturer's instructions and wereadded to the hIPC monolayer (40 nM final concentration). After 48 h, BrdU was added to themedium and 24 h later samples were fixed and processed as described for the nucleofectionprocedure. Because the effects of one siRNA or the other and combination of both siRNAsyielded the same effects, we combined these data.
ResultsWhen human islets are cultured in growth media, they flatten and mesenchymal-like cellsmigrate out (Gershengorn et al. 2004). The expanded population is highly proliferative andvisually homogeneous after several doublings in culture. hIPCs exhibited nuclear staining forβ-catenin along with some cytoplasmic staining (Fig. 1a). There was a dramatic shift in β-catenin localization in this population relative to intact islets (compare Fig. 1a,c). Nuclear β-catenin staining varied from 30% to about 70% in hIPCs from different donors duringlogarithmic growth (Table 1). In contrast, β-catenin exhibited surface membrane localizationin cells expressing insulin (measured as C-peptide) (β-cells), glucagon (α-cells) andsomatostatin (δ-cells) within islets (Fig. 1c). However, cells that had migrated out of the isletand assumed a mesenchymal-like phenotype were characterized by diminished membrane β-catenin and intense nuclear staining (Fig. 1b). Some cells that had migrated out of islets co-expressed C-peptide (Fig. 1b, arrows), while other cells expressing nuclear β-catenin had nodetectable C-peptide (Fig. 1b, arrowheads). We suggest that the cells that co-express C-peptideand nuclear β-catenin may represent a transient population of mesenchymal cells derived fromhormone-expressing cells similar to those that we (Morton et al. 2007) and others (Weinberget al. 2007) have observed in cultures of mouse islets.
Nuclear localization of β-catenin is a sine qua non for canonical Wnt signalling activation(Cong et al. 2003). Because a large percentage of the expanded hIPC population expressednuclear β-catenin during proliferation (Table 1), we next examined whether β-cateninsignalling was active in established hIPC cultures. A first step was to assess transcriptionalchanges of Wnt target genes. Table 2 shows that mRNA levels of Wnt target genes such asDKK1 (Niida et al. 2004; Gonzalez-Sancho et al. 2005), BIRC5 (survivin) (Zhang et al.2001), CCND1 (Shtutman et al. 1999; Tetsu & McCormick 1999), MYC (He et al. 1998),LEF1 (Filali et al. 2002) and FZD7 (Willert et al. 2002) were higher in hIPCs than in freshly
Ikonomou et al. Page 5
Cell Prolif. Author manuscript; available in PMC 2009 June 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

isolated human islets. Up-regulation of these genes was observed with varying magnitudes insamples from all three donors, suggesting these changes were characteristic of the expansionof mesenchymal pancreatic precursors. Of note, the transcript levels of TCF4, the prototypicalmember of the TCF/LEF family of DNA binding proteins, as well the transcript levels ofWNT5A and FZD2, genes involved in activation of the non-canonical Wnt/Ca2+ pathway(Slusarski et al. 1997), increased in hIPCs compared to islets also (Table 2). mRNA levels ofβ-catenin and GSK-3β were modestly increased during hIPC derivation. Therefore, theobserved β-catenin translocation probably reflects release of membrane-associated β-cateninrather than increased expression of β-catenin.
Because hIPCs may arise from expansion of a mesenchymal stem cell-like population withinhuman islets (Davani et al. 2007), expression levels of Wnt-related genes in hIPCs and bonemarrow-derived MSCs were compared. The molecular signatures of Wnt-related genesexpressed in hIPCs and bone marrow-derived MSCs were similar (Table 2). This result isconsistent with our previous observations that hIPCs share many similarities with bonemarrow-derived MSCs, including the ability to differentiate into adipocytes, chondrocytes andosteocytes (Davani et al. 2007).
One of the most up-regulated Wnt target genes in hIPCs relative to islets was DKK1 (Table 2).DKK1 protein is a potent-secreted inhibitor of the canonical Wnt pathway and it has beensuggested to be part of a negative feedback loop in normal tissues (Gonzalez-Sancho et al.2005). hIPCs produced and secreted DKK1 during proliferation (Fig. 2). Interestingly, theproduction rate of DKK1 correlated with exponential growth and was greatly reduced whencells entered the plateau phase, as indicated by the plateau in its cumulative concentration.Additional experiments revealed that production of DKK1 by hIPCs was switched off whencells were transferred to differentiation media and were induced to cluster (data not shown).Similar observations on changes of DKK1 expression have recently been reported by others(Kayali et al. 2007) when they expanded pancreatic cells using our protocol. Furthermore,DKK1, contrary to DKK3, is not expressed in the adult human pancreas (Hermann et al.2007) and, based on these results, its expression appears to be closely related to themesenchymal state of the cells. Interestingly, DKK1 has been implicated in cell cycle controlof mesenchymal stem cells (Gregory et al. 2003). These results suggest that β-catenin signallingis activated during hIPC derivation and expansion, due to nuclear translocation of β-catenin inmesenchymal hIPCs versus epithelial endocrine cells.
To confirm the importance of nuclear β-catenin for hIPC population growth, we assessedexpression of proliferation markers along with β-catenin in hIPCs during exponential growth(Fig. 3). Phosphorylation of the serine 10 residue of the N-terminal tail of histone H3 occursduring cell cycle progression through mitosis (Nowak & Corces 2004). We used an anti-H3phosphoS10 antibody to identify mitotic cells and these cells showed nuclear β-cateninstaining also. In addition, cells that stained for Ki-67, a marker expressed only in cells in theactive stages of the cell cycle but not in G0 (Scholzen & Gerdes 2000), co-stained with nuclearβ-catenin.
Work from our laboratory has suggested that hIPCs exhibit remarkable plasticity and that theirdifferentiation programme may be initiated by mesenchymal-to-epithelial transition(Gershengorn et al. 2004; Gershengorn et al. 2005). As described previously, hIPCs migratetogether to form epithelial cell clusters (ECC) when monodispersed and transferred to mediumwithout serum. In contrast to its nuclear localization in proliferative hIPC monolayers, β-catenin was localized at the cell periphery in ECCs (Fig. 4a). This translocation wasaccompanied by an increase in mRNA transcript levels of β-catenin and plakoglobin, whichis the main catenin in formation of desmosomes (Fig. 4b). Transcripts for epithelial markersclaudin-3 and claudin-4 also increased. Surprisingly, transcripts for the epithelial gene E-
Ikonomou et al. Page 6
Cell Prolif. Author manuscript; available in PMC 2009 June 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

cadherin decreased (Fig. 4b). Immunostaining of days 4 and 7 cell clusters confirmed that E-cadherin protein was absent (data not shown). Nevertheless, membrane localization of β-catenin (Fig. 4a,c, lower left panel) and N-cadherin (Fig. 4c, upper left panel) in ECCs implythat the latter and another adhesion molecule, or both, may sequester β-catenin at the cellsurface during the initial in vitro epithelialization of hIPCs. Taken together, these resultssuggest that β-catenin participates in the organization of ECCs and are consistent withmesenchymal-to-epithelial transition when hIPCs differentiate into epithelial-like structures.When compared to hIPC ECCs, MSC clusters displayed similar distribution of N-cadherin andβ-catenin (Fig. 4c, compare left and right panels).
We then attempted to modulate β-catenin signalling in hIPCs by changing the distribution ofβ-catenin. We treated hIPCs with BIO, a specific GSK-3β inhibitor that can mimic Wntsignalling by increasing the cellular pool of β-catenin in some systems (Meijer et al. 2003).Paradoxically, the number of hIPCs with distinct nuclear β-catenin was reduced after treatmentwith BIO (Fig. 5a,b), contrary to previous reports on BIO use (Sato et al. 2004). β-Cateninappeared to be mostly perinuclear, with staining intensity proportional to BIO concentration(Fig. 5a, compare middle and lower panels). Treatment of bone marrow-derived MSCs by thesame concentrations of BIO led to similar distribution of β-catenin (data not shown). Nuclearexclusion of β-catenin correlated with decreased proliferation (Fig. 5d, left panel). Cell cycleanalysis showed that BIO addition led to a decrease in the percentage of cells in S phase (Fig.5c). Consistent with this, BIO reduced BrdU incorporation in hIPCs (Fig. 5d, right panel).These results taken together indicate that treatment of proliferating hIPCs with BIO leads tonuclear exclusion of β-catenin, depletion of S-phase cells and over all decreased proliferation.
To confirm the functional roles of β-catenin, we studied the effect of its reduced expressionon β-catenin target genes and hIPC proliferation. We used RNA interference to specificallydown-regulate β-catenin in hIPC cultures. hIPCs were transfected with β-catenin siRNAs orwith control siRNA using either nucleofection or a transfection reagent designed to deliversiRNA oligonucleotides. Two commercial β-catenin siRNAs each led to about 10-foldreduction in β-catenin mRNA levels (Fig. 6b) after 72 h. Specific reduction of β-catenin proteinexpression by these siRNAs was confirmed by Western blot analysis (Fig. 6a). The siRNAsappeared to have similar effects decreasing both protein bands appearing in the region of the95 kDa standard, suggesting that these may represent β-catenin and a degradation or alternativesplice product; there were decreases in DKK1, BIRC5 and FZD7 mRNA levels (Fig. 6b). Theeffect of β-catenin knockdown on hIPC proliferation was assessed by BrdU incorporation (Fig.6c,d). There was a highly significant reduction (∼50%) in the fraction of BrdU-positive cellswhen β-catenin expression was reduced (Fig. 6d). These results clearly show that nuclear β-catenin plays a critical regulatory role in the expression of these genes and the control of hIPCproliferation.
DiscussionShortage of islet donors for the treatment of Type 1 diabetes has led to attempts by manylaboratories to identify and expand progenitor cell populations that have the potential todifferentiate into functional, islet-like structures. Our laboratory has previously described sucha population, mesenchymal hIPCs, which may arise by expansion of cells from the endocrinepancreas (Gershengorn et al. 2004). The present study provides a more detailedcharacterization of hIPCs in terms of β-catenin signalling. In this work, these cells were shownfor the first time to exhibit intense β-catenin nuclear staining. Perhaps more interestingly, cellswith nuclear β-catenin also appeared in islet outgrowths, that is, cells migrating out from islets.Established hIPC cultures contained high proportions of cells with nuclear β-catenin,independent of donor age or sex. The up-regulation of several β-catenin target genes, relativeto their expression levels in islets (Table 2), supports the idea of activation of β-catenin
Ikonomou et al. Page 7
Cell Prolif. Author manuscript; available in PMC 2009 June 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

signalling during hIPC derivation. One of the most up-regulated genes was DKK1; DKK1protein is a potent soluble inhibitor of Wnt signalling (Glinka et al. 1998). That hIPCs exhibitnuclear β-catenin throughout exponential population growth, while continuously secretingDKK1 is consistent with DKK1 being part of a negative feedback loop (Niida et al. 2004;Gonzalez-Sancho et al. 2005). Because DKK1 is not expressed in human pancreas (Hermannet al. 2007) but is highly up-regulated during hIPC expansion (Kayali et al. 2007 and this work),it may have a role as a marker of the mesenchymal state of the cells and its high expression inhIPCs may be a regulatory part of the hIPC proliferation programme.
The concept that β-catenin signalling relates to and is indispensable for hIPC proliferation issupported by several lines of evidence. First, hIPCs co-expressed markers of cell proliferationsuch as Ki-67 and H3phosphoS10 with nuclear β-catenin. When we knocked β-catenin downby means of RNAi, we observed reduction of hIPC proliferation in growth media, as measuredby BrdU incorporation. Hence, it appears that there is a tight link between nuclear localizationof β-catenin in hIPCs and their proliferation. Less direct results suggest that activated β-cateninsignalling may also be important during hIPC derivation from islets.
Some interesting parallels exist between this work and the role of Wnt signalling in vertebratepancreas development. Recent publications suggest that β-catenin signalling has complex rolesin the different stages of pancreatic development (Dessimoz et al. 2005; Murtaugh et al.2005; Papadopoulou & Edlund 2005; Heiser et al. 2006). Wnt signalling appears to be activein pancreatic epithelial cell progenitors and to decline thereafter (Dessimoz et al. 2005;Murtaugh et al. 2005). To determine its role in endocrine pancreas development, β-catenin-mediated signalling was modulated. Canonical Wnt signalling was reduced or abolished indeveloping mice either by overexpression of the cysteine-rich domain of Frz8, which acts asa decoy Wnt receptor (Papadopoulou & Edlund 2005), or by pancreas-specific deletion of β-catenin (Dessimoz et al. 2005; Murtaugh et al. 2005; Wells et al. 2007). Although one studyhas reported a decrease in endocrine cell numbers (Dessimoz et al. 2005), others have not(Murtaugh et al. 2005; Wells et al. 2007) and architecture and function of mature islets has notappeared to be perturbed. In contrast, overexpression of stabilized (unphosphorylated) β-catenin under the control of the Pdx1 promoter (Pdx1Crelate) at E13.5 resulted in increasedsize of the exocrine pancreas without affecting islet size, the latter being attributed to resistanceof pancreatic endocrine cells to nuclear accumulation of β-catenin (Heiser et al. 2006). Asimilar phenomenon was observed when we tried to increase cellular β-catenin by inhibitingGSK-3β activity. hIPCs as well as bone marrow-derived MSCs appear to exclude increasedβ-catenin from the nucleus (Fig. 5 and data not shown). Although this contrasts with the nuclearaccumulation of β-catenin when MSCs were treated with lithium chloride, another GSK-3βinhibitor (Etheridge et al. 2004), it may reflect differences in the mode of action of lithiumchloride and BIO on MSCs. Addition of BIO led to depletion of S-phase cells and decreasedproliferation of hIPCs. On the contrary, a recent study on the effect of GSK inhibitors on ratpancreatic islets, reported an increase in the β-cell replication rate (Mussmann et al. 2007).Thus, GSK inhibition may have different effects on β-cell and mesenchymal islet precursorreplication. Another striking feature of the mature (1-year-old) animals following β-cateninoverexpression in the mouse pancreas was the presence of cells with nuclear localization ofβ-catenin within a subset of islets (Heiser et al. 2006). These cells did not express insulin orany other marker of mature β-cells. Thus, it is possible that nuclear presence of β-catenininduces islet precursor cells to revert to a mesenchymal, proliferative phenotype in vivo also.In a recent publication, addition of Wnt3a, a canonical Wnt ligand, stimulated ex vivoproliferation of human β-cells and increased expression of cyclin D2 (Rulifson et al. 2007).We have observed nuclear β-catenin in C-peptide expressing cells migrating, from cadaverichuman islets. Because these cells may not persist in culture (Morton et al. 2007; Weinberg etal. 2007), Wnt signalling may not be sufficient for β-cell expansion during in vitro culture
Ikonomou et al. Page 8
Cell Prolif. Author manuscript; available in PMC 2009 June 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

(Rulifson et al. 2007), while it appears to have a significant role in the proliferation of MSC-like cells residing in the islets.
The latter idea seems particularly interesting given the similarity of hIPCs and MSCs. hIPCsadhere to plastic and proliferate in serum-containing media. Furthermore, they appear to meetmost of the criteria set to identify MSCs, because they express MSC surface markers (CD105/CD73/CD90), are CD14-, CD19- and CD34-negative and some hIPC preparations have beenshown to be able to differentiate to adipocytes, osteocytes and chondrocytes (Davani et al.2007). Bone-marrow MSCs are established in culture as the fraction of cells from bone aspiratethat adhere to plastic. Wnt signalling has been shown to be active in MSCs (Etheridge et al.2004) and it is known to block adipogenic differentiation of pre-adipocytes (Ross et al.2000) and osteogenic differentiation of adult human MSCs (Boland et al. 2004). In the latterwork, infection of MSCs with an adenovirus encoding Wnt3a, a ligand of the canonical Wntpathway, resulted in increased proliferation and substantially higher cell counts relative tomock-infected cells. Recent work from the same group has indicated that two Wnt pathway-related genes, namely FZD7 and DKK3, are part of the ‘stemness’ gene group for MSCs (Songet al. 2006). These two genes are down-regulated during adipogenesis and osteogenesis ofhMSCs, but their expression is increased when differentiated cells of these lineages revert toan undifferentiated state. Interestingly, it has been recently shown that DKK3 is expressed inhuman β-cells (Hermann et al. 2007) and by in situ hybridization that FZD7 (among genes forother frizzled receptors) is mainly localized in pancreatic islets (Heller et al. 2003). Both genesare highly up-regulated in hIPCs relative to human islets (Table 2). Thus, if hIPCs are an islet-derived stem cell population, insights from MSC biology may be useful for their furthercharacterization.
Certain analogies that exist between hIPCs and other non-transformed cells in culture may alsobe of interest. For example, it has been shown that increased expression of β-catenin and itsnuclear translocation increased proliferation and inhibited apoptosis of vascular smooth musclecells (VSMC) following carotid injury in Sprague-Dawley rats (Wang et al. 2002). Recently,Fat1 cadherin, whose expression is increased in hIPCs relative to islets (unpublishedobservation), has been shown to interact with β-catenin and modulate its proliferative effectwhile increasing migration of VSMCs (Hou et al. 2006). N-cadherin was also found to complexwith β-catenin in the membrane of quiescent VSMCs (Uglow et al. 2003). Serum stimulationof these cells led to adherens junction dismantling and nuclear translocation of β-catenin. Thus,information from VSMC biology, especially with relevance to cadherin–catenin interactions,may help us to gain insights in the regulation of proliferation of islet progenitor cells.
β-Catenin localization may be one of the cues regulating hIPC proliferation and differentiation.Indeed, the reversible translocation of β-catenin from the cell membrane to the nucleus suggeststhat β-catenin may mediate, in part, the remarkable plasticity exhibited by hIPCs. When theyare induced to differentiate by removal of growth factors from the culture media, β-catenintranslocates to the membrane. Because we hypothesize that mesenchymal-to-epithelialtransition is an essential step in the hIPC differentiation programme, enhancement of β-catenintranslocation could produce ECCs that are more stable and poised to differentiate followingtransplantation. In conclusion, in the present work, we show that human mesenchymal islet-derived precursor cells exhibit dynamic β-catenin signalling. Although derivation and invitro expansion of similar populations of proliferative precursor cells have been reported, thisis the first time, to our knowledge, that these cells have been demonstrated to exhibit activationof this major developmental pathway. Because it is known that β-catenin signalling is not activein normal human pancreas, our results may have implications for both pancreas neogenesisand development of better in vitro protocols for progenitor cell expansion and differentiation.
Ikonomou et al. Page 9
Cell Prolif. Author manuscript; available in PMC 2009 June 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

AcknowledgementsThis work was supported by the Intramural Research Program of the National Institute of Diabetes and Digestive andKidney Diseases, National Institutes of Health. The authors thank Behrous Davani for helpful suggestions and criticalreading of the manuscript.
ReferencesAberle H, Bauer A, Stappert J, Kispert A, Kemler R. beta-catenin is a target for the ubiquitin-proteasome
pathway. EMBO J 1997;16:3797–3804. [PubMed: 9233789]Atouf F, Park CH, Pechhold K, Ta M, Choi Y, Lumelsky NL. No evidence for mouse pancreatic β-Cell
epithelial-mesenchymal transition in vitro. Diabetes 2007;56:699–702. [PubMed: 17327438]Bach JF. Insulin-dependent diabetes-mellitus as an autoimmune-disease. Endocr Rev 1994;15:516–542.
[PubMed: 7988484]Boland GM, Perkins G, Hall DJ, Tuan RS. Wnt 3a promotes proliferation and suppresses osteogenic
differentiation of adult human mesenchymal stem cells. J Cell Biochem 2004;93:1210–1230.[PubMed: 15486964]
Chase LG, Ulloa-Montoya F, Kidder BL, Verfaillie CM. Islet-derived fibroblast-like cells are not derivedvia epithelial-mesenchymal transition from Pdx-1 or insulin-positive cells. Diabetes 2007;56:3–7.[PubMed: 17110468]
Cho HH, Kim YJ, Kim SJ, Kim JH, Bae YC, Ba B, Jung JS. Endogenous Wnt signaling promotesproliferation and suppresses osteogenic differentiation in human adipose derived stromal cells. TissueEng 2006;12:111–121. [PubMed: 16499448]
Cong F, Schweizer L, Chamorro M, Varmus H. Requirement for a nuclear function of beta-catenin inWnt signaling. Mol Cell Biol 2003;23:8462–8470. [PubMed: 14612392]
Davani B, Ikonomou L, Raaka BM, Geras-Raaka E, Morton RA, Marcus-Samuels B, Gershengorn MC.Human islet-derived precursor cells are mesenchymal stromal cells that differentiate and mature tohormone-expressing cells in vivo. Stem Cells 2007;25:3215–3222. [PubMed: 17901402]
Dessimoz J, Bonnard C, Huelsken J, Grapin-Botton A. Pancreas-specific deletion of beta-catenin revealsWnt-dependent and Wnt-independent functions during development. Curr Biol 2005;15:1677–1683.[PubMed: 16169491]
Etheridge SL, Spencer GJ, Heath DJ, Genever PG. Expression profiling and functional analysis of Wntsignaling mechanisms in mesenchymal stem cells. Stem Cells 2004;22:849–860. [PubMed:15342948]
Filali M, Cheng NL, Abbott D, Leontiev V, Engelhardt JF. Wnt-3A/beta-catenin signaling inducestranscription from the LEF-1 promoter. J Biol Chem 2002;277:33398–33410. [PubMed: 12052822]
Gao R, Ustinov J, Korsgren O, Otonkoski T. In vitro neogenesis of human islets reflects the plasticity ofdifferentiated human pancreatic cells. Diabetologia 2005;48:2296–2304. [PubMed: 16193291]
George SJ, Beeching CA. Cadherin: catenin complex: a novel regulator of vascular smooth muscle cellbehaviour. Atherosclerosis 2006;188:1–11. [PubMed: 16438974]
Gershengorn MC, Geras-Raaka E, Hardikar AA, Raaka BM. Are better islet cell precursors generatedby epithelial-to-mesenchymal transition? Cell Cycle 2005;4:380–382. [PubMed: 15711124]
Gershengorn MC, Hardikar AA, Wei CJ, Geras-Raaka E, Marcus-Samuels B, Raaka BM. Epithelial-to-mesenchymal transition generates proliferative human islet precursor cells. Science 2004;306:2261–2264. [PubMed: 15564314]
Glinka A, Wu W, Delius H, Monaghan AP, Blumenstock C, Niehrs C. Dickkopf-1 is a member of a newfamily of secreted proteins and functions in head induction. Nature 1998;391:357–362. [PubMed:9450748]
Gonzalez-Sancho JM, Aguilera O, Garcia JM, Pendas-Franco N, Pena C, Cal S, de Herreros AG, BonillaF, Munoz A. The Wnt antagonist DICKKOPF-1 gene is a downstream target of beta-catenin/TCFand is downregulated in human colon cancer. Oncogene 2005;24:1098–1103. [PubMed: 15592505]
Gregory CA, Singh H, Perry AS, Prockop DJ. The Wnt signaling inhibitor dickkopf-1 is required forreentry into the cell cycle of human adult stem cells from bone marrow. J Biol Chem 2003;278:28067–28078. [PubMed: 12740383]
Ikonomou et al. Page 10
Cell Prolif. Author manuscript; available in PMC 2009 June 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

He TC, Sparks AB, Rago C, Hermeking H, Zawel L, da Costa LT, Morin PJ, Vogelstein B, Kinzler KW.Identification of c-MYC as a target of the APC pathway. Science 1998;281:1509–1512. [PubMed:9727977]
Heiser PW, Lau J, Taketo MM, Herrera PL, Hebrok M. Stabilization of beta-catenin impacts pancreasgrowth. Development 2006;133:2023–2032. [PubMed: 16611688]
Heller RS, Klein T, Ling ZD, Heimberg H, Katoh M, Madsen OD, Serup P. Expression of Wnt, frizzled,sFRP, and DKK genes in adult human pancreas. Gene Expr 2003;11:141–147. [PubMed: 14686787]
Hermann M, Pirkebner D, Draxl A, Berger P, Untergasser G, Margreiter R, Hengster P. Dickkopf-3 isexpressed in a subset of adult human pancreatic beta cells. Histochem Cell Biol 2007;127:513–521.[PubMed: 17347849]
Hou R, Liu L, Anees S, Hiroyasu S, Sibinga NES. The Fat1 cadherin integrates vascular smooth musclecell growth and migration signals. J Cell Biol 2006;173:417–429. [PubMed: 16682528]
Huber MA, Kraut N, Beug H. Molecular requirements for epithelial-mesenchymal transition during tumorprogression. Curr Opin Cell Biol 2005;17:548–558. [PubMed: 16098727]
Jamora C, Dasgupta R, Kocieniewski P, Fuchs E. Links between signal transduction, transcription andadhesion in epithelial bud development. Nature 2003;422:317–322. [PubMed: 12646922]
Kayali AG, Flores LE, Lopez AD, Kutlu B, Baetge E, Kitamura R, Hao E, Beattie GM, Hayek A. Limitedcapacity of human adult islets expanded in vitro to redifferentiate into insulin-producing β-cells.Diabetes 2007;56:703–708. [PubMed: 17327439]
Lechner A, Nolan AL, Blacken RA, Habener JF. Redifferentiation of insulin-secreting cells after invitro expansion of adult human pancreatic islet tissue. Biochem Biophys Res Commun2005;327:581–588. [PubMed: 15629153]
Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCRand the 2 (T) (-Delta Delta C) method. Methods 2001;25:402–408. [PubMed: 11846609]
Logan CY, Nusse R. The Wnt signaling pathway in development and disease. Annu Rev Cell Dev Biol2004;20:781–810. [PubMed: 15473860]
Lu ZM, Ghosh S, Wang ZY, Hunter T. Downregulation of caveolin-1 function by EGF leads to the lossof E-cadherin, increased transcriptional activity of beta-catenin, and enhanced tumor cell invasion.Cancer Cell 2003;4:499–515. [PubMed: 14706341]
Meijer L, Skaltsounis AL, Magiatis P, Polychronopoulos P, Knockaert M, Leost M, Ryan XZP, VonicaCA, Brivanlou A, Dajani R, Crovace C, Tarricone C, Musacchio A, Roe SM, Pearl L, Greengard P.GSK-3-selective inhibitors derived from Tyrian purple indirubins. Chem Biol 2003;10:1255–1266.[PubMed: 14700633]
Monga SPS, Mars WM, Pediaditakis P, Bell A, Mule K, Bowen WC, Wang X, Zarnegar R, MichalopoulosGK. Hepatocyte growth factor induces Wnt-independent nuclear translocation of beta-catenin aftermet-ss-catenin dissociation in hepatocytes. Cancer Res 2002;62:2064–2071. [PubMed: 11929826]
Morali OG, Delmas V, Moore R, Jeanney C, Thiery JP, Larue L. IGF-II induces rapid beta-cateninrelocation to the nucleus during epithelium to mesenchyme transition. Oncogene 2001;20:4942–4950. [PubMed: 11526479]
Morton RA, Geras-Raaka E, Wilson LM, Raaka BM, Gershengorn MC. Endocrine precursor cells frommouse islets are not generated by epithelial-to-mesenchymal transition of mature beta cells. Mol CellEndocrinol 2007;270:87–93. [PubMed: 17363142]
Muller T, Bain G, Wang X, Papkoff J. Regulation of epithelial cell migration and tumor formation bybeta-catenin signaling. Exp Cell Res 2002;280:119–133. [PubMed: 12372345]
Murtaugh LC, Law AC, Dor Y, Melton DA. beta-Catenin is essential for pancreatic acinar but not isletdevelopment. Development 2005;132:4663–4674. [PubMed: 16192304]
Mussmann R, Geese M, Harder F, Kegel S, Andag U, Lomow A, Burk U, Onichtchouk D, DohrmannC, Austen M. Inhibition of GSK3 promotes replication and survival of pancreatic beta cells. J BiolChem 2007;282:12030–12037. [PubMed: 17242403]
Nelson WJ, Nusse R. Convergence of Wnt, beta-catenin, and cadherin pathways. Science2004;303:1483–1487. [PubMed: 15001769]
Niida A, Hiroko T, Kasai M, Furukawa Y, Nakamura Y, Suzuki Y, Sugano S, Akiyama T. DKK1, anegative regulator of Wnt signaling, is a target of the beta-catenin/TCF pathway. Oncogene2004;23:8520–8526. [PubMed: 15378020]
Ikonomou et al. Page 11
Cell Prolif. Author manuscript; available in PMC 2009 June 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Nir T, Dor Y. How to make pancreatic beta cells – prospects for cell therapy in diabetes. Curr OpinBiotechnol 2005;16:524–529. [PubMed: 16084716]
Novak A, Hsu SC, Leung-Hagesteijn C, Radeva G, Papkoff J, Montesano R, Roskelley C, GrosschedlR, Dedhar S. Cell adhesion and the integrin-linked kinase regulate the LEF-1 and beta-cateninsignaling pathways. Proc Natl Acad Sci USA 1998;95:4374–4379. [PubMed: 9539744]
Nowak SJ, Corces VG. Phosphorylation of histone H3: a balancing act between chromosomecondensation and transcriptional activation. Trends Genet 2004;20:214–220. [PubMed: 15041176]
Oloumi A, McPhee T, Dedhar S. Regulation of E-cadherin expression and beta-catenin/Tcftranscriptional activity by the integrin-linked kinase. Biochim Biophys Acta 2004;1691:1–15.[PubMed: 15053919]
Ouziel-Yahalom L, Zalzman M, Anker-Kitai L, Knoller S, Bar Y, Glandt M, Herold K, Efrat S. Expansionand redifferentiation of adult human pancreatic islet cells. Biochem Biophys Res Commun2006;341:291–298. [PubMed: 16446152]
Papadopoulou S, Edlund H. Attenuated Wnt signaling perturbs pancreatic growth but not pancreaticfunction. Diabetes 2005;54:2844–2851. [PubMed: 16186384]
Quasnichka H, Slater SC, Beeching CA, Boehm M, Sala-Newby GB, George SJ. Regulation of smoothmuscle cell proliferation by beta-catenin/T-cell factor signaling involves modulation of cyclin D1and p21 expression. Circ Res 2006;99:1329–1337. [PubMed: 17122440]
Ross SE, Hemati N, Longo KA, Bennett CN, Lucas PC, Erickson RL, MacDougald OA. Inhibition ofadipogenesis by Wnt signaling. Science 2000;289:950–953. [PubMed: 10937998]
Rulifson IC, Karnik SK, Heiser PW, Ten Berge D, Chen HN, Gu XY, Taketo MM, Nusse R, Hebrok M,Kim SK. Wnt signaling regulates pancreatic beta cell proliferation. Proc Natl Acad Sci USA2007;104:6247–6252. [PubMed: 17404238]
Sato N, Meijer L, Skaltsounis L, Greengard P, Brivanlou AH. Maintenance of pluripotency in humanand mouse embryonic stem cells through activation of Wnt signaling by a pharmacological GSK-3-specific inhibitor. Nat Med 2004;10:55–63. [PubMed: 14702635]
Scholzen T, Gerdes J. The Ki-67 protein: from the known and the unknown. J Cell Physiol 2000;182:311–322. [PubMed: 10653597]
Shtutman M, Zhurinsky J, Simcha I, Albanese C, D'Amico M, Pestell R, Ben-Ze'ev A. The cyclin D1gene is a target of the beta-catenin/LEF-1 pathway. Proc Natl Acad Sci USA 1999;96:5522–5527.[PubMed: 10318916]
Slusarski DC, Corces VG, Moon RT. Interaction of Wnt and a frizzled homologue triggers G-protein-linked phosphatidylinositol signalling. Nature 1997;390:410–413. [PubMed: 9389482]
Song L, Webb NE, Song YJ, Tuan RS. Identification and functional analysis of candidate genes regulatingmesenchymal stem cell self-renewal and multipotency. Stem Cells 2006;24:1707–1718. [PubMed:16574750]
Tetsu O, McCormick F. Beta-catenin regulates expression of cyclin D1 in colon carcinoma cells. Nature1999;398:422–426. [PubMed: 10201372]
Thiery JP, Sleeman JP. Complex networks orchestrate epithelial-mesenchymal transitions. Nat Rev MolCell Biol 2006;7:131–142. [PubMed: 16493418]
Tian E, Zhan FH, Walker R, Rasmussen E, Ma YP, Barlogie B, Shaughnessy JD. The role of the Wnt-signaling antagonist DKK1 in the development of osteolytic lesions in multiple myeloma. N Engl JMed 2003;349:2483–2494. [PubMed: 14695408]
Uglow EB, Slater S, Sala-Newby GB, Aguilera-Garcia CM, Angelini GD, Newby AC, George SJ.Dismantling of cadherin-mediated cell-cell contacts modulates smooth muscle cell proliferation. CircRes 2003;92:1314–1321. [PubMed: 12775583]
Wang XH, Xiao Y, Mou YS, Zhao Y, Blankesteijn WM, Hall JL. A role for the beta-catenin/T-cell factorsignaling cascade in vascular remodeling. Circ Res 2002;90:340–347. [PubMed: 11861424]
Weinberg N, Ouziel-Yahalom L, Knoller S, Efrat S, Dor Y. Lineage tracing evidence for in vitrodedifferentiation but rare proliferation of mouse pancreatic β-cells. Diabetes 2007;56:1299–1304.[PubMed: 17303800]
Wells JM, Esni F, Boivin GP, Aronow BJ, Stuart W, Combs C, Sklenka A, Leach SD, Lowy AM. Wnt/beta-catenin signaling is required for development of the exocrine pancreas. BMC Dev Biol 2007;7:4.[PubMed: 17222338]
Ikonomou et al. Page 12
Cell Prolif. Author manuscript; available in PMC 2009 June 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Willert J, Epping M, Pollack J, Brown P, Nusse R. A transcriptional response to Wnt protein in humanembryonic carcinoma cells. BMC Dev Biol 2002;2:8. [PubMed: 12095419]
Zhang T, Otevrel T, Gao ZQ, Gao ZP, Ehrlich SM, Fields JZ, Boman BM. Evidence that APC regulatessurvivin expression: a possible mechanism contributing to the stem cell origin of colon cancer. CancerRes 2001;61:8664–8667. [PubMed: 11751382]
Ikonomou et al. Page 13
Cell Prolif. Author manuscript; available in PMC 2009 June 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 1. Localization of β-catenin in expanded human islet-derived precursor cells (hIPCs), isletoutgrowths and mature human pancreas(a) Proliferating hIPCs were fixed 72 h after re-seeding in culture and were stained with amonoclonal antibody against β-catenin. Nuclei were stained with Hoechst 33342. Bar, 20 μm.(b) High purity islet fractions were cultured in 2-chamber Permanox slides in growth media.Islets were fixed after 48 h in culture and stained for β-catenin and C-peptide. A representativefield showing islet outgrowth is presented. Cells co-expressing C-peptide and nuclear β-catenin(arrows) or cells expressing nuclear β-catenin but no detectable C-peptide (arrowheads) canbe seen. Bar, 10 μm. (c) High magnification confocal micrographs of islet endocrine cells.Paraffin wax-embedded sections of human pancreas were deparaffinized and stained for β-catenin (green). Sections were co-stained with C-peptide, glucagon or somatostatin (red) toidentify β-, α- and δ-cells, respectively. Bar, 10 μm.
Ikonomou et al. Page 14
Cell Prolif. Author manuscript; available in PMC 2009 June 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
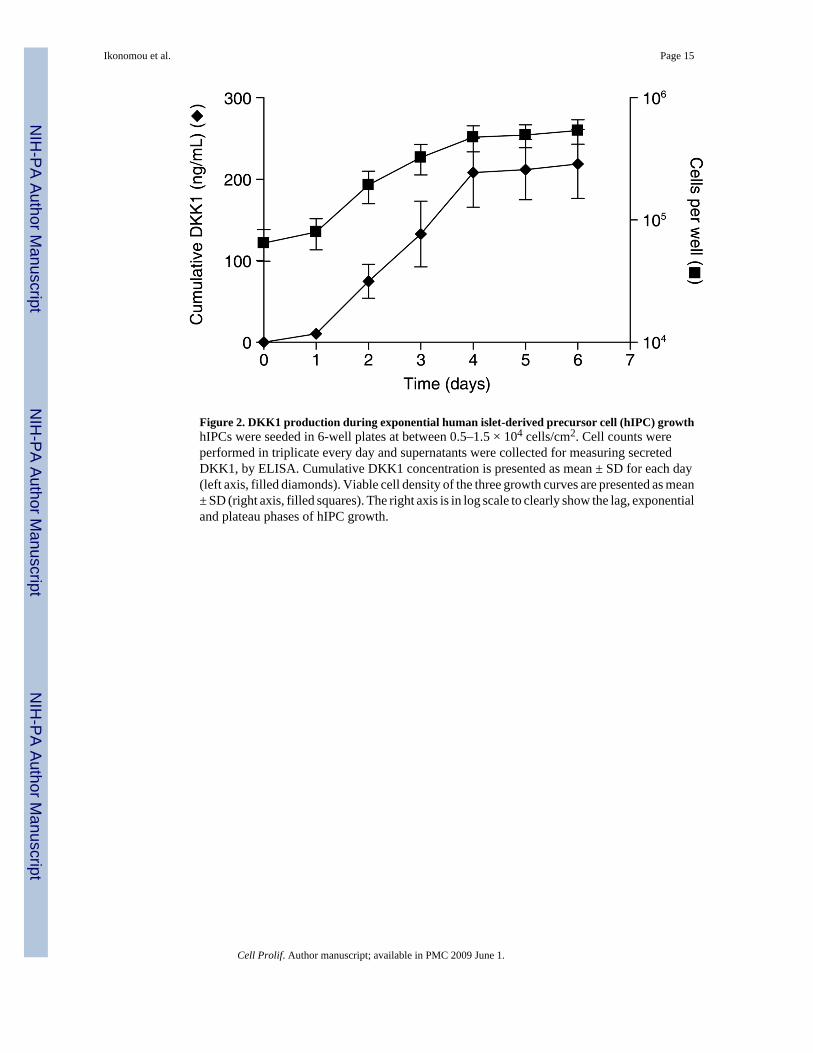
Figure 2. DKK1 production during exponential human islet-derived precursor cell (hIPC) growthhIPCs were seeded in 6-well plates at between 0.5–1.5 × 104 cells/cm2. Cell counts wereperformed in triplicate every day and supernatants were collected for measuring secretedDKK1, by ELISA. Cumulative DKK1 concentration is presented as mean ± SD for each day(left axis, filled diamonds). Viable cell density of the three growth curves are presented as mean± SD (right axis, filled squares). The right axis is in log scale to clearly show the lag, exponentialand plateau phases of hIPC growth.
Ikonomou et al. Page 15
Cell Prolif. Author manuscript; available in PMC 2009 June 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 3. Nuclear β-catenin is co-expressed with proliferation markersHuman islet-derived precursor cells (hIPC) were seeded in 2-well Permanox slides in growthmedium and were stained during exponential growth for H3phosphoS10 (red) (upper panel;bar, 20 μm) or Ki-67 (red) (lower panel; bar, 20 μm) and β-catenin (green). All of the Ki-67and H3phosphoS10-positive cells were found to express nuclear β-catenin.
Ikonomou et al. Page 16
Cell Prolif. Author manuscript; available in PMC 2009 June 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 4. Cell surface translocation of β-catenin during epithelialization of human islet-derivedprecursor cells (hIPC)(a) hIPCs exhibit nuclear β-catenin staining in monolayer culture in growth medium andmembrane staining in epithelial cell clusters (ECC) at day 7. ECCs were harvested either onday 4 or day 7 after switch to differentiation medium and were paraffin wax-embedded. hIPCsfrom three different donors were used to create ECCs. In all experiments, β-catenin had asimilar expression pattern in ECCs. A representative 10-μm section is shown. Bar, 20 μm. (b)Total RNA was prepared from monolayer hIPCs in growth medium or from ECCs after 4 daysin differentiation medium (day 4 ECCs). After cDNA preparation, transcript levels for theindicated genes were determined by qRT-PCR. The fold change relative to day 0 hIPCs wascalculated using the 2(−ΔΔCt) method. (c) N-cadherin and β-catenin staining in day 4 ECCs.Paraffin wax-embedded ECCs of hIPCs or MSCs were deparaffinized and stained for N-cadherin or β-catenin (green). Nuclear counterstain is DAPI. Bar, 20 μm.
Ikonomou et al. Page 17
Cell Prolif. Author manuscript; available in PMC 2009 June 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Ikonomou et al. Page 18
Cell Prolif. Author manuscript; available in PMC 2009 June 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 5. Treatment of human islet-derived precursor cells (hIPCs) with BIO leads to reduced cellgrowth(a) Confocal micrographs of hIPCs treated with BIO. Cells were stained for β-catenin (green)72 h after addition of dimethyl sulfoxide (DMSO) (control, upper panel), 1 μM BIO (middlepanel) or 3 μM BIO (lower panel). Nuclear counterstain was Hoechst 33342. Bar, 20 μm. (b)Quantification of nuclear staining in a typical experiment. (c) Cell cycle analysis of cells treatedwith BIO or DMSO (control). Cells were stained with propidium iodide and analysed by meansof a BD FACS Calibur cytometer. A representative set of data from one of three independentexperiments is shown. (d) For cell number (left panel), hIPCs were seeded in 6-well plates at1 × 104 cells/cm2 (∼0.1 × 106 cells/well). BIO was added at the indicated concentrations afterovernight cell attachment. Cells were harvested 48 h after BIO addition. Six wells were usedfor cell counting and the experiment was performed twice. Results are shown as mean ± SD.Significant statistical differences between control and treated conditions were calculated byone-way ANOVA followed by Dunnett's multiple comparison test (*P < 0.01). For BrdU staining(right panel), hIPCs were seeded in 2-well Permanox chambers, BrdU was added 24 h afterBIO addition and slides were fixed after 24-h incubation. Areas with varied degrees of cellconfluence were included in the analysis. To avoid biased selection of fields, slides were viewedusing the DAPI filter (nuclear staining).
Ikonomou et al. Page 19
Cell Prolif. Author manuscript; available in PMC 2009 June 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 6. Reduced expression of β-catenin inhibits human islet-derived precursor cell (hIPC)proliferation(a) Western blot of total cell extracts for β-catenin 72 h after transfection with a control siRNA,β-catenin siRNA1, β-catenin siRNA2 or pooled β-catenin siRNAs (50 : 50 ratio). The twoprotein bands near the 95 kDa standard are both reduced similarly following siRNA treatmentsand likely reflect β-catenin and a degradation or alternative splice product. The band just belowthe 37 kDa standard is GAPDH, documenting similar protein loading in the four lanes. (b)Levels of DKK1, BIRC5 and FZD7 mRNAs were measured by qRT-PCR. Results are shownas mean ± SEM. (c) Confocal micrographs of BrdU-incorporating hIPCs at 72 h aftertransfection with siRNA1 + siRNA2 (left panel) or Control siRNA (right panel). Bar, 50 μm.(d) BrdU incorporation in hIPCs is decreased following transfection with β-catenin siRNAs.A total number of 500–700 nuclei were counted for each condition in each experiment tocalculate the fraction of BrdU+ cells. Areas with varied degrees of cell confluence wereincluded in the analysis. To avoid biased selection of fields, slides were viewed using the DAPIfilter (nuclear staining). The results of four independent experiments are compiled. Results areshown as mean ± SEM. Significant statistical differences between control siRNA and β-cateninsiRNA were calculated by paired two-tailed t-test (*P < 0.0001).
Ikonomou et al. Page 20
Cell Prolif. Author manuscript; available in PMC 2009 June 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Ikonomou et al. Page 21
Table 1Percentage of nuclear β-catenin-positive cells in exponentially growing human islet-derived precursor cell cultures
Preparation Passage number Time after re-seeding (h) Nuclear β-catenin positivity(%)
A 15 24 68 (189/278)
A 15 72 72 (231/321)
D 3 48 35 (97/274)
E 5 24 32 (88/273)
F 13 48 51 (389/757)
F 13 72 52 (159/306)
Cell Prolif. Author manuscript; available in PMC 2009 June 1.

NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Ikonomou et al. Page 22Ta
ble
2Tr
ansc
riptio
nal c
hang
es fo
r Wnt
sign
allin
g-re
late
d ge
nes f
ollo
win
g hI
PC e
xpan
sion
qRT
-PC
R c
ycle
thre
shol
d
MSC
s
Isle
tsPr
olife
rativ
e hI
PCs
AB
CA
, p7
B, p
5C
, p8
β-C
aten
in ta
rget
gen
esD
KK
129
.729
.527
.520
.720
.222
.520
.4
FZD
723
.725
.323
.518
.019
.119
.519
.7
BIRC
526
.632
.125
.620
.621
.521
.321
.1
MYC
19.9
21.6
22.2
18.9
20.2
19.0
n.m
.
CC
ND
118
.622
.620
.616
.516
.817
.718
.0
LEF1
26.2
28.9
25.7
26.5
25.8
24.2
27.2
Wnt
/Ca2+
pat
hway
WN
T5A
23.8
26.7
21.1
18.7
19.1
16.9
19.4
FZD
226
.830
.125
.721
.522
.120
.822
.3
DK
K3
22.2
25.2
20.2
15.8
16.7
16.9
20.0
TCF4
22.4
24.6
22.1
19.5
19.7
20.0
21.2
GSK
-3β
21.1
20.1
19.4
19.0
19.1
19.3
19.0
CTN
NB1
21.5
22.6
19.7
19.6
20.2
19.9
19.6
hIPC
, hum
an is
let-d
eriv
ed p
recu
rsor
cel
l; M
SC, m
esen
chym
al st
em c
ell;
qRT-
PCR
, qua
ntita
tive
real
-tim
e po
lym
eras
e ch
ain
reac
tion.
Cell Prolif. Author manuscript; available in PMC 2009 June 1.
Copyright © 2022 FDOKUMEN




















