
Regulation of β-catenin trafficking to the membrane in living cells
Upload
independentCategory
view
0download
0
ORIGINAL ARTICLE
Recruitment of adenomatous polyposis coli and b-catenin to axin-puncta
MC Faux, JL Coates, B Catimel, S Cody, AHA Clayton, MJ Layton and AW Burgess
Ludwig Institute for Cancer Research, Melbourne Tumour Biology Branch, Parkville, Victoria, Australia
The adenomatous polyposis coli (APC) tumour suppressoris a multifunctional protein involved in the regulation ofWnt signalling and cytoskeletal dynamics. Little is knownabout how APC controls these disparate functions. In thisstudy, we have used APC- and axin-fluorescent fusionproteins to examine the interactions between theseproteins and show that the functionally distinct popula-tions of APC are also spatially separate. Axin-RFP formscytoplasmic punctate structures, similar to endogenousaxin puncta. Axin-RFP recruits b-catenin destructioncomplex proteins, including APC, b-catenin, glycogensynthase kinase-3-b (GSK3-b) and casein kinase-1-a(CK1-a). Recruitment into axin-RFP puncta sequestersAPC from clusters at cell extensions and this prevents itsmicrotubule-associated functions. The interaction betweenAPC-GFP and axin-RFP within the cytoplasmic puncta isdirect and dramatically alters the dynamic properties ofAPC-GFP. However, recruitment of APC to axin punctais not absolutely required for b-catenin degradation.Instead, formation of axin puncta, mediated by the DIXdomain, is required for b-catenin degradation. AnaxinDDIX mutant did not form puncta, but still mediatedrecruitment of destruction complex proteins and phos-phorylation of b-catenin. We conclude that there aredistinct pools of APC and that the formation of axinpuncta, rather than the axin/APC complex, is essentialfor b-catenin destruction.Oncogene (2008) 27, 5808–5820; doi:10.1038/onc.2008.205;published online 30 June 2008
Keywords: APC; axin; b-catenin
Introduction
Wnt signalling controls crucial stages in cell fatedecisions, tissue development and maintenance andhas been identified as a key signalling pathway in cancer(Logan and Nusse, 2004; Schneikert and Behrens, 2007).When Wnt signalling is suppressed, the scaffold proteinaxin and the tumour suppressor adenomatous polyposis
coli (APC) form a complex that promotes the phos-phorylation of b-catenin by glycogen synthase kinase-3-b(GSK3-b), which consequently targets b-catenin forubiquitination by bTrCP and destruction by theproteasome (Aberle et al., 1997; Orford et al., 1997;Kitagawa et al., 1999). Wnt stimulation and APCmutations disrupt the APC/axin complex and result inincreased levels of b-catenin in the nucleus andcytoplasm (Munemitsu et al., 1995). b-Catenin is knownto interact with members of the Tcf family of transcrip-tion factors to activate transcription of genes involved inproliferation and cellular transformation (Korineket al., 1997).
The function of APC in the regulation of the Wnt/b-catenin pathway has been studied in depth and theaberrant activation of b-catenin as a result of truncatingmutations in APC is recognized as a central event incolon cancer (Polakis, 1997, 2000; Nathke, 2004).However, the spatial and temporal regulation of APC’sinteractions with Wnt signalling components and thecytoskeleton is less well understood. APC is localized atthe basal plasma membrane in association with micro-tubules, to the peripheral ends of cell extensions (Nathkeet al., 1996; Barth et al., 2002). This distribution of APCpoints to a major role for APC in cytoskeletal functionsassociated with cell migration. APC binds to micro-tubules (Munemitsu et al., 1994; Su et al., 1995) andregulates their growth and stability (Kita et al., 2006),and has been linked to the actin cytoskeleton viainteractions with the Rac1/Cdc42 effector proteinIQGAP1 (Watanabe et al., 2004), the Rac-specificexchange factor Asef (Kawasaki et al., 2000) and theRho effector mDia (Wen et al., 2004). b-catenin has alsorecently been implicated in the clustering of APC atcellular protrusions (Sharma et al., 2006). However, it isnot clear whether the ends of APC-loaded microtubulesare the sites of the axin/b-catenin/GSK3-b/APC ‘de-struction’ complex or whether this function takes placeelsewhere in the cell.
The subcellular location of axin is less well definedthan APC. In Xenopus the physiological concentrationof axin is proposed to be exceedingly low (Lee et al.,2003), making detection at the subcellular level difficult.Endogenous axin has been detected in small, punctatecytoplasmic structures that redistribute to large aggre-gates close to the plasma membrane following treatmentwith LiCl (which inhibits GSK3-b, mimicking Wntactivation) (Levina et al., 2004; Wiechens et al., 2004).Ectopically expressed recombinant axin is localized in
Received 12 November 2007; revised 30 April 2008; accepted 30 May2008; published online 30 June 2008
Correspondence: Dr MC Faux, Epithelial Biology Laboratory,Ludwig Institute for Cancer Research, PO Box 2008, RoyalMelbourne Hospital, Parkville, Victoria 3050, Australia.E-mail: [email protected]
Oncogene (2008) 27, 5808–5820& 2008 Macmillan Publishers Limited All rights reserved 0950-9232/08 $32.00
www.nature.com/onc

cytoplasmic vesicles or puncta (Fagotto et al., 1999;Smalley et al., 1999; Schwarz-Romond et al., 2005). TheC-terminal DIX domain of axin is known to mediateself-association, which is thought to be important forthe regulation of b-catenin (Hsu et al., 1999; Kishidaet al., 1999; Sakanaka and Williams, 1999) and hasrecently been shown to associate into polymers that arerequired for puncta formation (Schwarz-Romond et al.,2007a, b). It has also been proposed that axin shuttles inand out of the nucleus (Wiechens et al., 2004) and that itassociates with microtubules and regulates their stabilitythrough dishevelled (Dvl) (Ciani et al., 2004).
Although APC and axin have been shown to interactusing immunoprecipitation (Hart et al., 1998), thesubcellular location of this interaction has not been welldefined. It is possible that only a small proportion ofAPC associates with the axin complex. Recent biochem-ical data point to the existence of discrete populations ofAPC where the APC/axin/b-catenin complex is separatefrom APC interactions with microtubules (Penmanet al., 2005). These studies suggest that the spatialregulation of both APC and axin is a dynamic processbut complex formation at the subcellular level has notbeen explored. In the present study we have used APC-and axin- fluorescent fusion proteins to study axinregulation of APC subcellular localization and toexamine the dynamic interactions of these proteins inliving mammalian cells. We examined whether otherdestruction complex proteins, b-catenin, GSK3-b andcasein kinase-1-a (CK1a) were recruited to axin punctaand whether axin puncta formation, mediated by theDIX domain of axin, is required for complex formationand b-catenin degradation. Our data show that there aredistinct subcellular pools of APC, that axin redistributesAPC from microtubule cell extensions to cytoplasmicaxin puncta, and that this prevents microtubule-associated functions of APC. We show that axin recruits
destruction complex proteins and that formation of theaxin puncta, rather than the axin/APC complex, iscritical for b-catenin degradation.
Results
To study the subcellular location and dynamic proper-ties of axin, we assessed the localization of bothendogenous and recombinant axin fused to monomericred fluorescent protein (RFP) (Figure 1a). Using acommercially available polyclonal axin antibody, weobserved endogenous axin in distinct puncta in thecytoplasm of Madin–Darby canine kidney (MDCK)cells (Figures 1b and c). Both endogenous andrecombinant axin puncta were evenly distributedthroughout the cytoplasm (Figures 1b–e), consistentwith previous reports for endogenous axin in HEK293cells (Levina et al., 2004; Wiechens et al., 2004) andrecombinant axin (Fagotto et al., 1999; Smalley et al.,1999; Schwarz-Romond et al., 2005, 2007b). Axin-RFPwas localized to cytoplasmic puncta when expressedin both MDCK (Figure 1e) and HEK293 T cells(Supplementary Figures S1, S2). Similarly, epitope–tagged axin-HA (Figure 1d) formed cytoplasmic puncta,suggesting that the monomeric RFP tag did not affect itsdistribution.
Axin recruits APC to cytoplasmic punctaEndogenous APC concentrated in clusters at the ends ofmicrotubules in subconfluent epithelial cells (Nathkeet al., 1996; Barth et al., 2002; Figure 2a). Transientlyexpressed APC-GFP was also concentrated in distinctclusters at the peripheral ends of microtubules inMDCK cells (Figure 2b), mimicking the staining ofendogenous APC in these cells (Figure 2a). In some
Figure 1 Localization of axin to cytoplasmic puncta. (a) Schematic representation of domain structures of axin-, axinDRGS- andaxinDDIX-RFP and APC-GFP fusion proteins. (b, c) Endogenous axin is localized to cytoplasmic puncta in Madin–Darby caninekidney (MDCK) cells. Cells were fixed and immunostained for endogenous axin with antibodies to axin-1. (c) High magnification ofcytoplasmic puncta of endogenous axin. (d) MDCK cells transiently expressing axin-HA were fixed and immunostained with anti-HAantibodies. (e) MDCK cells expressing axin-RFP. Shown are single confocal sections.
Recruitment of APC and b-catenin to axin punctaMC Faux et al
5809
Oncogene

cells, APC-GFP also decorated the microtubule network(see Figure 2f). In contrast, in cells expressing axin-RFP,endogenous APC was redistributed to cytoplasmicpuncta, coincident with axin (Figure 2c, arrowheads).In cells not expressing axin-RFP, APC was localized inclusters at cell extensions (Figure 2c, arrows). Expres-sion of the mutant axinDRGS-RFP protein (Figure 1a)that does not bind APC (Behrens et al., 1998; Hart et al.,1998; Kishida et al., 1998) but forms similar puncta towild-type axin, had no effect on the localization ofendogenous APC (Figure 2d, arrows), confirming thatrelocalization of APC depends upon binding to axin.Similarly, when APC-GFP was coexpressed with axin-RFP, APC-GFP redistributed from the cell tips to thepunctate structures containing axin-RFP (Figure 2e).Again, coexpression of APC-GFP with the mutantaxinDRGS-RFP did not lead to redistribution of APC(Figure 2f). Similar results were obtained in HEK293T cells (Supplementary Figures S1 and S2). Thus, axinhas the capacity to recruit APC to cytoplasmic puncta.
Although endogenous APC was detected in axin-RFPpuncta, APC could not be detected in the endogenousaxin puncta observed in MDCK cells (Figures 1b and c):either endogenous axin is not at a sufficient concentra-tion (Lee et al., 2003) to allow recruitment of APC orAPC is present at a level below the threshold ofdetection. Therefore, we examined the levels of axinrequired to sequester APC away from cell extensions.The relative levels of expression of axin-RFP:APC-GFPwere measured based on total fluorescence intensity inindividual cells and were correlated with APC localiza-tion in either the puncta or microtubules (Supplemen-tary Figures S2C and D). Scatter plots revealed twodistinct data clusters corresponding to APC localized tothe microtubule network only when axin expression waslow, and APC localized to axin puncta as the level ofaxin increases (Supplementary Figure S2D). These dataindicate a threshold level of axin expression is requiredfor APC to redistribute to puncta.
We next investigated whether recruitment of APCfrom clusters at the ends of microtubule tips to the axin
puncta affects microtubule-associated functions of APC.The presence of APC on microtubule plus ends in cellextensions has been reported to stabilize and promotethe growth of microtubules (Kita et al., 2006). The lossof APC results in reduced cell protrusions and decreasedmicrotubule stability (Kroboth et al., 2007). Weexamined microtubule assembly activity in untrans-fected, axin-RFP and axinDRGS-RFP transfectedMDCK cells. Cells were incubated on ice for 1 h todepolymerize microtubules and microtubule polymer-ization was then induced by incubation at 30 1Cfollowed by rapid fixation (Figures 2g–j). After 1minincubation at 30 1C, microtubule asters formed inuntransfected cells (Figure 2g) and at 5min, micro-tubules had repolymerized (not shown). In contrast, inaxin-RFP expressing cells, microtubule assembly wasreduced significantly and at 1min, microtubule astershad not formed (Figure 2h). Microtubule assembly inaxinDRGS-RFP expressing cells was similar to controlcells (Figure 2i). We quantitated these differences bycounting the number of cells with microtubule asters(spindles; Figure 2j). Less than 25% of axin-RFP-expressing cells formed asters at 1min compared tomore than 95% in untransfected cells and 75% in axin-DRGS expressing cells (Figure 2j). Expression of axin-RFP therefore inhibited the microtubule assemblyactivity of APC. This was not a direct effect of axinoverexpression, as APC binding and recruitment to axinpuncta was required. Sequestering APC into axin punctatherefore shifts the cellular population of APC awayfrom its microtubule-associated location and function.
Direct interaction between APC-GFP and axin-RFPwithin punctaTo determine whether colocalization of APC and axinwithin the axin puncta involves a direct interaction, wemeasured fluorescence resonance energy transfer(FRET) between APC-GFP and axin-RFP usingfluorescence lifetime imaging microscopy (FLIM). Adecrease in APC-GFP lifetime in the presence of axin-
Figure 2 Recruitment of adenomatous polyposis coli (APC) to cytoplasmic axin puncta. (a) Endogenous APC and (b) APC-GFPlocalize to the ends of microtubules in clusters in Madin–Darby canine kidney (MDCK) cells. APC (anti-APC)/APC-GFP -green, b-tubulin -red, nuclei (4,6-diamidino-2-phenyl indole; DAPI) blue. Shown are single section confocal images. APC (left); compositeimage (middle), scale bar 20 mm; inset (right) at higher magnification of APC clusters, scale bar, 5 mm. (c) In the presence of axin-RFPendogenous APC redistributes to the axin puncta. (d) MDCK cells expressing axinDRGS-RFP. Cells were fixed and immunostainedwith antibodies to APC (green). Shown are single section confocal composite images, scale bar 20 mm. APC is detected at cell tips inuntransfected and axinDRGS-RFP cells (c, d arrows). Distinct APC puncta are present in cells expressing axin-RFP (c, arrowheads),but no APC was associated with axinDRGS-RFP puncta (d). (e) APC-GFP is recruited to axin-RFP puncta in MDCK cells. (f) APC-GFP is not recruited to axinDRGS-RFP puncta. Shown are composite images of Z series projections of cells imaged by laser scanningconfocal microscopy. Scale bar 20 mm. (g–j) Recruitment of APC to axin-RFP puncta prevents microtubule assembly followingmicrotubule depolymerization. MDCK cells were incubated on ice for 1 h to depolymerize microtubules and then at 30 1C for 1min,fixed and immunostained. (g) Microtubule spindle formation occurs in non-axin expressing cells. (h) In axin-RFP expressing cells,microtubule spindle assembly is decreased. (i) Microtubule spindle formation in axinDRGS-RFP expressing cells. (j) Untransfected,axin- and axinDRGS- RFP cells were scored for the presence or absence of microtubule spindles. Microtubule spindle assembly wasreduced in axin-RFP but not in the axinDRGS-RFP cells. (k–n) Measurement of nano-scale interaction of APC-GFP and axin-RFP byfrequency-domain FRET-FLIM. Shown are APC-GFP fluorescence images and fluorescence lifetime images for: (k) APC-GFPexpressed alone; (l, m) APC-GFP coexpressed with (l) axin-RFP and (m) axinDRGS-RFP. APC-GFP fluorescence concentrated at celltips (arrowheads); APC-GFP fluorescence in intracellular puncta with corresponding depressions in fluorescence lifetimes (arrows).Scale bar 20 mm. (n) Mean and s.e. of fluorescence lifetimes of APC-GFP in clusters at cellular extensions and in intracellular puncta inAPC-GFP and axin-RFP co-expressing cells. Statistical analysis revealed that fluorescence lifetime in puncta was reduced compared tofluorescence lifetime in clusters at cellular extensions (***Po0.0001).
Recruitment of APC and b-catenin to axin punctaMC Faux et al
5810
Oncogene

RFP indicates FRET, which implies that the twofluorophores are within 10 nm of each other (Figures2k–n). The fluorescence lifetime of the GFP fluorophorein cells expressing APC-GFP alone was relativelyuniform, with lifetime values in the expected range of2–2.25 ns for GFP (Figure 2k; average lifetime2.03±0.03 ns (n¼ 11), Figure 2n). Fluorescence lifetimewas independent of the fluorescence intensity andtherefore the amount of APC-GFP in the clusters.
When axin-RFP was coexpressed with APC-GFP, theredistribution of APC-GFP to intracellular axin punctawas accompanied by a reduction in APC-GFP fluores-cence lifetime (Figure 2l, arrows, average lifetime1.61±0.03 ns, n¼ 82; Figure 2n) indicating a directinteraction. Coexpression of APC-GFP with the mutantaxinDRGS-RFP protein had no effect on the lifetime ofthe APC-GFP (Figures 2m and n). The interactionbetween axin-RFP and APC-GFP is therefore specific,
Recruitment of APC and b-catenin to axin punctaMC Faux et al
5811
Oncogene

as shown by coprecipitation and deletion mappingstudies (Behrens et al., 1998; Hart et al., 1998), andoccurs within cytoplasmic puncta rather than at the endsof microtubules.
Dynamic behaviour of APC-GFP is influenced byaxin-RFPGFP-tagged APC has previously been shown to movealong the microtubule network towards their plus endsin the periphery of cell extensions (Mimori-Kiyosueet al., 2000). We therefore examined whether binding toaxin altered the dynamics of APC movement within thecell (Figure 3; Supplementary material (movies 1A–C)).APC-GFP was primarily localized at the microtubuletips with some decoration of the microtubule networkwhen present at higher levels (Figure 3a, dashed arrow;Supplementary material (movie 1A); see also Supple-mentary Figure S2B). The movement of humanAPC-GFP was consistent with that of Xenopus APC
(Mimori-Kiyosue et al., 2000) and it also accumulated atspecific structures, possibly associated with microtubules(Figure 3a; arrowheads; Supplementary material(movie 1A)).
In contrast, when APC-GFP was coexpressed withaxin-RFP and relocalized to puncta, there was no longerany detectable tracking, or coordinated or directedmovement of APC-GFP (Figures 3b–d; Supplementarymaterial (movies 1B and C)). Instead, both APC-GFPand axin-RFP were restricted to the puncta and appearto oscillate on the spot. Single particle tracking (SPT)analysis from time-lapse images revealed rapid diffusionof free APC-GFP, whereas APC-GFP particles in theaxin-RFP puncta moved threefold more slowly(Figure 3f). Our analysis includes both combineddiffusion and directed motion. Particle trajectories weredetermined from time-lapse images, represented asmean-squared displacement (MSD) as a function oftime (Figure 3e). The shape of the MSD versus time plotprovides information on the speed and type of motion.
AP
C+
axinA
PC
+axin
AP
C+
axinA
PC
5
<
<
* *
0:00
<
<*
**
6:25
<
<
12:25
MS
D µ
m2 MS
D µ
m2
2
0
4
6
8
10
12
14
Time (seconds)
Time (seconds)
Single particle tracking analysis
APC-GFP
Diffusion µm2/s
6.1 +/- 0.3 x 10-3 1.6 +/- 0.1 x 10-3
8.2 +/- 2.6 x 10-3
17 21
2.8 +/- 1.1 x 10-3Velocity µm2/s
Number
APC-GFP+axinRFP
0 20 40 60 80 1000.0
0.1
0.2
0.3
<<
***
12:256:25
***
0:00
10
10
**
****
20
*
<<
<
<
<*
****
20 40 60 80 100
Figure 3 Axin restricts the dynamic behaviour of adenomatous polyposis coli (APC). Time-lapse analysis of APC-GFP and axin-RFP. Still frames from (Supplementary material (movies 1A–C)) of APC-GFP and axin-RFP in living HEK293 T cells. Merged imagesof GFP and red fluorescent protein (RFP) signal are shown. Elapsed time is indicated in the top left of panel (a) (min:sec). (a) APC-GFP (Movie 1A). Dashed arrows indicate decoration of microtubule network; arrowheads indicate movement of GFP alongmicrotubules. The APC-GFP-structures cluster in several specific regions (single asterisks); some clusters appear to grow and movealong the microtubules before dispersing (double asterisks) or even disappearing (arrows). (b–d) APC-GFP and axin-RFPcoexpression. (b) Movie 1B. GFP signal is now localized in cytoplasmic puncta coincident with axin, visualized as yellow in the mergedimage. Large (arrowheads) and smaller (arrows) axin puncta demonstrate oscillatory movement (Movie 1B). The asterisks denotepuncta that appear to separate into red and green particles and then merge to yellow. (c) The cell on the left from Movie 1C (and athigher magnification in d), demonstrates two GFP puncta that overlap with RFP (visualized as yellow), denoted by open arrowheadsymbol. This cell also displays APC-GFP decoration of the microtubule network (dashed arrows) and accumulation of GFP-particles(*) that appear to move along the microtubules before dispersing (**). The lower level of axin expression in this cell appears only toinfluence a relatively small proportion of APC-GFP; the APC-GFP not bound to axin moves along microtubules in a similar fashion toAPC-GFP expressed alone. This tracking of APC-GFP is visualized in Movie 1C. (e) Single Particle Tracking (SPT) analysis of APC-GFP and axin-RFP. Plot of the mean-squared displacement (MSD) versus time for a highlighted particle (*) undergoing directedmotion from (a) (green triangles) and a highlighted particle from (d) (open arrowhead) undergoing constrained motion (red diamondsand inset). (f) Diffusion and velocity coefficients from SPT analysis.
Recruitment of APC and b-catenin to axin punctaMC Faux et al
5812
Oncogene

A linear plot indicates random Brownian diffusion anupward curvature indicates directed motion and adownward curvature indicates constrained motion.SPT analysis demonstrated that there was a directionalcomponent to the movement in 11 out of 17 APC-GFP-containing spots analysed, as evidenced by the upwardcurvature in the MSD versus time plot (Figure 3e, greentriangles). In contrast, when APC-GFP was colocalizedwith axin-RFP, the levelling of the MSD versus timeplot indicates constrained motion (Figure 3e, reddiamonds, and inset). Both larger (Figure 3b, arrow-heads) and smaller puncta (Figure 3b, arrow) displayedsimilar oscillatory behaviour. Thus, the location andmovement of APC-GFP bound to axin-RFP, wasrestricted compared to the rapid, directed movementof free APC-GFP.
The dynamic behaviour of APC-GFP coexpressedwith axin-RFP was analysed in cells that expresseddifferent levels of axin-RFP (Figures 3c and d;Supplementary material (movie 1C)). Regardless ofaxin-RFP expression levels, APC-GFP dynamics werecontrolled by whether or not APC-GFP was bound toaxin-RFP. In cells expressing high levels of axin-RFP(Figure 3c, arrowhead), the majority of APC-GFP wascolocalized with axin-RFP and its movement was
correspondingly restricted. In cells expressing low levelsof axin-RFP, only a small proportion of total APC-GFPwas colocalized with axin-RFP, however, all of thisproportion had restricted movement (Figures 3c and d,open arrowheads). In contrast, the majority of APC-GFP in these cells was not bound to axin and thedynamics of this proportion was equivalent to that ofAPC-GFP in cells not transfected with axin-RFP(Supplementary material (movie 1C); Figures 3c and d,dashed arrow). Thus, axin influences the dynamics ofaxin-bound APC but has no effect on the proportionthat is not bound. This suggests that two populations ofAPC, one microtubule associated and the other axin-puncta associated, can coexist in the cell.
Recruitment of destruction complex proteins into axinpunctaTo determine whether axin puncta represent theb-catenin destruction complex (Schneikert and Behrens,2007), we examined the ability of axin-RFP to recruitendogenous b-catenin, GSK3-b and CK1-a to axinpuncta (Figure 4). Initially, we confirmed that transientexpression of recombinant axin resulted in downregula-tion of b-catenin in SW480 cells, a colon cancer cell line
50000
40000
30000
20000
10000
0GS
K3β
fluo
resc
ence
in
tens
ity
MD
CK
cells
MG132
β-catenin Axin-RFP
20µ
P-β-cat Axin-RFPP-β-cat Axin-RFP
SW
480 cells
β-catenin Axin-RFP β-catenin Axin-RFP
20µ
Control
20µ
β-catenin Axin-RFP
20µ
P-β-cat Axin-RFP P-β-cat Axin-RFP
MD
CK
cellsM
DC
K cells
GSK3-β Axin-RFPAxin-RFPGSK3-β
20µ
20µ
CK1-α Axin-RFPAxin-RFPCK1-α+ Axin-RFP-
Figure 4 Recruitment of b-catenin, glycogen synthase kinase-3-b (GSK3-b) and casein kinase-1-a (CK1-a) in cells expressing axin-RFP. (a) SW480 and (b) Madin–Darby canine kidney (MDCK) cells expressing axin-RFP were treated with the proteasome inhibitorMG132, and immunostained with antibodies to b-catenin for SW480 cells (a) or phospho-b-catenin (P-b-cat) for MDCK cells (b).Shown are confocal images of b-catenin or P-b-cat, axin-RFP and composite images (b-catenin or P-b-cat—green, axin-RFP—red,nuclei—blue in (a)) for control and treated cells. Arrows indicate axin-RFP expressing cells; arrowheads indicate b-catenin or P-b-catcoincident with axin-RFP. (c) GSK3-b and (d) CK1-a are recruited to axin-RFP puncta in MDCK cells. Shown are confocal images ofMDCK cells expressing axin-RFP and immunostained with antibodies to GSK3-b (c) or CK1-a (d). Composite images show GSK3-bor CK1-a—green, axin-RFP—red. Scale bars, 20 mm. (e) Fluorescence intensity of GSK3b from individual MDCK cells with andwithout axin-RFP expression was measured. Values shown are mean and s.e.m. of GSK3b fluorescence intensity from at least 50 cells.
Recruitment of APC and b-catenin to axin punctaMC Faux et al
5813
Oncogene

containing only truncated APC (Figure 4a), as has beenreported previously (Behrens et al., 1998; Hart et al.,1998). This downregulation was due to proteasome-mediated degradation, as inhibition of the proteasomeby MG132 resulted in accumulation of b-catenin inaxin-containing puncta (Figure 4a, arrowheads). InMDCK cells, the proportion of b-catenin targeted fordegradation was masked by the large population ofmembrane-associated b-catenin, therefore we usedphospho-specific b-catenin antibodies that detect pS45/pT41/pS37/pS33-b-catenin (phospho-b-catenin) to vi-sualize the b-catenin pool that is targeted for degrada-tion (Figure 4b). Again, inhibition of the proteasomeallowed phospho-b-catenin to accumulate in axin-containing puncta (Figure 4b, arrowheads), implyingthat axin puncta represent the destruction complex.Likewise, inhibition of GSK3-b in SW480 cells resultedin increased b-catenin coincident with axin puncta(Supplementary Figure S3A), further implicating theaxin puncta as the site of the destruction complex.Endogenous GSK3-b and CK1-a were also recruited toaxin-RFP-containing puncta in MDCK cells (Figures 4cand d). Similarly, GSK3-b was recruited to axin-RFPpuncta in SW480 cells (Supplementary Figure S3B).Unlike b-catenin, levels of GSK3-b were not regulatedby the destruction complex as quantitation of totalGSK3-b fluorescence intensity shows no differencebetween untransfected and axin-RFP-transfected cells(Figure 4e). Collectively, these results show that theendogenous destruction complex proteins, GSK3-b,CK1-a and APC, are all recruited to axin puncta.Recruitment of b-catenin was only observed if itsdegradation was inhibited by switching off the protea-some, implying that puncta participate in a functionaldestruction complex.
Recruitment of b-catenin into axin punctadoes not require full-length APCThe degradation of b-catenin following transient ex-pression of axin in SW480 cells (Behrens et al., 1998;Hart et al., 1998; Figure 4a) is somewhat surprisinggiven that SW480 cells contain truncated APC proteinthat is not able to bind axin. This implies that expressionof axin allows a functional destruction complex to beformed in the absence of intact APC, but the relativeimportance of axin and APC in forming the destructioncomplex and the mechanism by which expression of axinleads to b-catenin degradation has not been elucidated.We explored the requirement for APC further in cellsother than SW480. The axinDRGS mutant, whichcannot bind APC, was still able to downregulateb-catenin in SW480 cells (Supplementary Figure S4)and to recruit phospho-b-catenin and GSK3-b inMDCK cells (Figure 5). As with wild-type axin-RFP,phospho-b-catenin was detected in axinDRGS puncta(Figure 5a) and levels of phospho-b-catenin wereincreased in these axinDRGS-containing puncta in thepresence of proteasome inhibitors (Figure 5b). Similarly,GSK3-b was also recruited to axinDRGS-containingpuncta (Figure 5c). We conclude that, despite being
recruited to the destruction complex, APC is notabsolutely required for the formation of a functionaldestruction complex.
Formation of axin puncta is necessary for b-catenindegradation but not phosphorylationThe DIX domains of both Dvl2 and axin have recentlybeen shown to mediate both self-polymerization andpuncta formation (Schwarz-Romond et al., 2007a, b).Therefore, we investigated whether axin polymerizationand puncta formation were required for assembly of thedestruction complex, and whether they influence recruit-ment and b-catenin degradation. Previous studies haveshown that the DIX domain is important for b-cateninregulation (Kishida et al., 1999; Sakanaka and Williams,1999), however, it has also been reported that deletion ofthe DIX domain does not affect the ability of axin toalter b-catenin localization (Nakamura et al., 1998). Wetested the ability of axinDDIX-RFP to recruit b-cateninin cells treated with proteasome inhibitors (Figure 6). Aswe had observed previously, expression of axin-RFPresulted in destruction of b-catenin (Figure 6a, left) thatwas prevented by inhibition of the proteasome, resultingin detection of b-catenin in axin puncta (Figure 6a,right). We found that axinDDIX did not form punctabut instead was distributed diffusely in the cytoplasm(Figure 6b), consistent with previous reports that theDIX domain is required for efficient puncta formation(Schwarz-Romond et al., 2007b). Importantly, expres-sion of axinDDIX-RFP did not result in reduced b-catenin, but instead b-catenin was predominantlycytosolic with a distribution that mirrored that ofaxinDDIX (Figure 6b). Thus, axinDDIX appears tosequester b-catenin but this does not lead to itsdegradation. Inhibition of the proteasome did not havea marked effect on b-catenin distribution or levelsimplying that the axin puncta formation is required forefficient degradation of b-catenin. To determine whetheraxin puncta formation is required for b-catenin phos-phorylation, we examined whether phospho-b-cateninwas sequestered in the cytoplasm following expressionof axinDDIX. Indeed, we found increased cytoplasmicphospho-b-catenin in axinDDIX cells compared tountransfected surrounding cells, in both SW480 andMDCK cells (Figures 6c and d). Quantitation offluorescence intensity confirms that the levels ofcytoplasmic phospho-b-catenin are increased in axinDDIX-expressing cells (Figure 6e). AxinDDIX thereforerecruited b-catenin, GSK3-b and CK1 (as implied byphosphorylation of axinDDIX-associated b-catenin).AxinDDIX is also capable of recruiting APC. APCtranslocates from clusters at the ends of microtubules ina proportion of axinDDIX-expressing MDCK cells.This recruitment is not as efficient as recruitment byaxin puncta (Supplementary Figure S5). Theseresults imply that axinDDIX, which still containsbinding sites for b-catenin, APC, GSK3-b and CK1-a,still facilitates b-catenin phosphorylation by GSK3-b,but proteasomal degradation is blocked in the absenceof puncta.
Recruitment of APC and b-catenin to axin punctaMC Faux et al
5814
Oncogene

Discussion
In this paper, we suggest a new paradigm for theassembly of the b-catenin destruction complex. Byexpression of axin-RFP, we were able to detect therecruitment of proteins away from other subcellularlocations and formation of the b-catenin destructioncomplex in association with axin puncta. Our resultsdemonstrate that axin recruits APC into cytoplasmicpuncta, where its movement becomes restricted as aresult of axin binding, and its ability to regulatemicrotubule assembly is decreased significantly. How-ever, in cells overexpressing axin, recruitment anddegradation of b-catenin in axin puncta does notabsolutely require APC. The recruitment of b-catenin,GSK3-b and CK1-a to axin puncta implies that axinpuncta form the site of the classical b-catenin destruc-tion complex. Axin puncta, mediated by the DIXdomain, are not required for recruitment of destructioncomplex proteins or b-catenin phosphorylation, how-ever, b-catenin degradation does not occur unlesspuncta are formed. We believe that axin is important
in the recruitment of APC, b-catenin and GSK3-b andthat axin puncta are crucial for b-catenin degradation.
The ability of axin to form puncta has been reportedpreviously (Fagotto et al., 1999; Smalley et al., 1999;Schwarz-Romond et al., 2005), and recent studies showthat puncta formation is a dynamic process mediated byprotein oligomerization or polymerization (Schwarz-Romond et al., 2007a, b). Our data provide evidence forthe existence of endogenous axin puncta, consistent withthe detection of small punctate structures or spots withaxin antibodies in 293 cells (Levina et al., 2004;Wiechens et al., 2004). The DIX domain of axin hasbeen shown to mediate self-assembly in vitro andcontribute to the signalling function of axin (Kishidaet al., 1999; Sakanaka and Williams, 1999), but is notrequired for recruitment into Dvl2 puncta (Schwarz-Romond et al., 2007b). Instead, our data now show thatthe DIX domain of axin is essential for axin-punctaformation and b-catenin degradation. Polymerization ofDvl2 has been suggested to provide a high localconcentration of Dvl2, thus promoting recruitment ofits binding partners (Schwarz-Romond et al., 2007b). In
MDCK cells
20 µm
20 µm
20 µm
MG
132C
ontrol
P-β-cat
P-β-cat
Axin∆RGS-RFP
Axin∆RGS-RFP
GSK3-β Axin∆RGS Axin∆RGS-RFP GSK3-β
P-β-cat Axin∆RGS
P-β-cat Axin∆RGS
Figure 5 Axin recruitment of b-catenin and glycogen synthase kinase-3-b (GSK3-b) does not require adenomatous polyposis coli(APC). (a) Control or (b) MG132 treated Madin–Darby canine kidney (MDCK) cells expressing axinDRGS-RFP were immunostainedwith antibodies to phospho-b-catenin (P-b-cat). (c) Cells expressing axinDRGS-RFP were immunostained with antibodies to GSK3-b.Shown are confocal images of P-b-cat or GSK3-b, axinDRGS-RFP and composite images (P-b-cat or GSK3-b—green, axin-RFP—red). Arrowheads indicate phospho-b-catenin or GSK3-b coincident with axinDRGS-RFP. Scale bars: 20mm.
Recruitment of APC and b-catenin to axin punctaMC Faux et al
5815
Oncogene
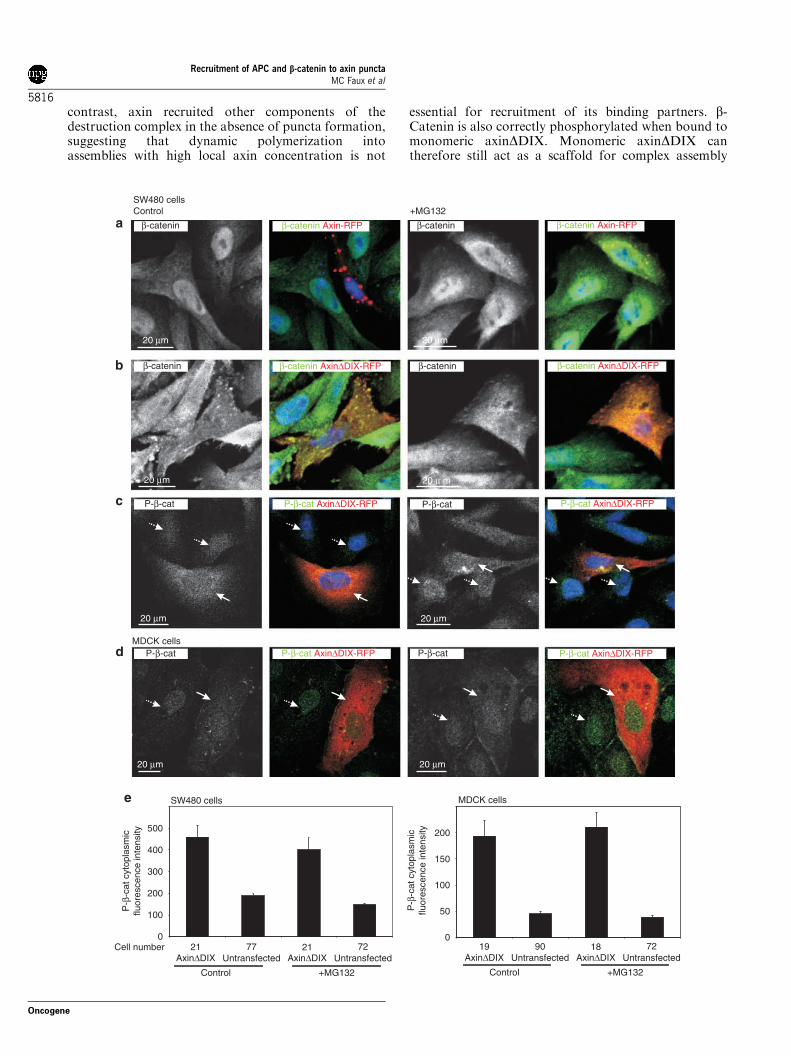
contrast, axin recruited other components of thedestruction complex in the absence of puncta formation,suggesting that dynamic polymerization intoassemblies with high local axin concentration is not
essential for recruitment of its binding partners. b-Catenin is also correctly phosphorylated when bound tomonomeric axinDDIX. Monomeric axinDDIX cantherefore still act as a scaffold for complex assembly
SW480 cells Control
SW480 cells
MDCK cells
MDCK cells
+MG132
20 µm 20 µ m
20 µm 20 µm
20 µm 20 µm
20 µm 20 µm
P-β
-cat
cyt
opla
smic
flu
ores
cenc
e in
tens
ity
P-β
-cat
cyt
opla
smic
flu
ores
cenc
e in
tens
ity
50
100
150
200
0
Control +MG132
Axin∆DIX21
Untransfected77
Axin∆DIX21
Untransfected72
Control +MG132
Axin∆DIX19
Untransfected90
Axin∆DIX18
Untransfected72
100
200
300
400
500
0
β-catenin β-cateninβ-catenin Axin-RFP β-catenin Axin-RFP
β-catenin β-cateninβ-catenin Axin∆DIX-RFP
P-β-cat P-β-catP-β-cat Axin∆DIX-RFP P-β-cat Axin∆DIX-RFP
P-β-cat P-β-catP-β-cat Axin∆DIX-RFP P-β-cat Axin∆DIX-RFP
Cell number
β-catenin Axin∆DIX-RFP
Recruitment of APC and b-catenin to axin punctaMC Faux et al
5816
Oncogene

and b-catenin phosphorylation, but not b-catenindegradation. Puncta formation may be required forthe recruitment, positioning and/or activation of theSkp1/Cul 1/F box ubiquitination machinery (Hart et al.,1999; Latres et al., 1999; Winston et al., 1999).Experiments are currently underway to determinewhether puncta formation is required for b-TrCPrecruitment, b-catenin ubiquitination or proteasomaldegradation.
We propose that axin promotes the formation of anAPC/axin complex at a precise subcellular location thatis distinct from the concentration and function of APCat microtubule tips. This concept is supported by theidentification of at least two functionally different APC-containing protein complexes (Penman et al., 2005).Here, we provide evidence for two distinct subcellularpopulations of APC with different functions. Atmicrotubule extensions, APC regulates microtubuleassembly dynamics whereas in axin puncta, the APC/axin complex potentiates the destruction of b-catenin.Our demonstration that axin redirects APC away frommicrotubule ends, preventing microtubule assembly,suggests that the functions of APC in the regulation ofmicrotubule dynamics and b-catenin stability arespatially separate.
Axin can direct the location and movement of APC,but APC does not appear to influence axin distribution.In Drosophila, APC is also recruited into cytoplasmicpuncta following overexpression of axin (Cliffe et al.,2003). It appears that the amount of axin in the cellalters the balance between APC bound to axin and APCassociated with microtubules. Although the reportedphysiological concentration of axin is low (Lee et al.,2003), even low concentrations of axin (includingendogenous axin) form puncta, implying a high localconcentration. It is likely that the populations ofAPC are governed by whether or not axin is bound,rather than by axin concentration. Importantly,our data show that binding to axin restricted thediffusion and velocity of APC-GFP and preventeddirected motion (Mimori-Kiyosue et al., 2000;Dayanandan et al., 2003; Langford et al., 2006). Thealtered dynamics of APC bound to axin supportsour findings that the axin/APC complex is distinct fromthe rapidly moving APC population at microtubules.Indeed, we found that axin expression alteredmicrotubule function and this was dependent on APCbinding. Recently, Dvl2 has been reported to
alter the dynamic behaviour of axin, possiblychanging the affinity for axin and some of its ligands(Schwarz-Romond et al., 2007b). The constrainedmotion of APC within axin puncta possibly reflects thesize of the polymerized axin complex and/or an alteredconformational state as a result of interactions thatdiffer from the population of APC interacting withmicrotubules.
Intriguingly, recruitment of APC to axin appearsdispensable for b-catenin degradation (this study; Hartet al., 1998; Kishida et al., 1999). However, APC hasbeen shown to influence levels of b-catenin (Munemitsuet al., 1995). It may be that endogenous levels of axin arenot sufficient to regulate the levels of b-catenin in theabsence of wild-type APC but increased axin canindependently promote b-catenin degradation. Indeed,overexpression of Dvl proteins results in Wnt-indepen-dent signalling (Kishida et al., 1999; Smalley et al., 1999;Park et al., 2005) that is dependent on the Dvl DIXdomain (Schwarz-Romond et al., 2007b). In Xenopus,the local concentration of Dvl is important forcanonical signalling (Park et al., 2005). Similarly, forcedoligomerization of LRP6 induces Wnt signalling andbypasses the requirement for Dvl (Cong et al., 2004;Bilic et al., 2007).
The role of APC in the regulation of b-catenin is welldocumented: truncating mutations in APC result inelevated b-catenin levels (Schneikert and Behrens, 2007).However, we have shown previously that stable expres-sion of APC in SW480 cells results in reduced b-catenin/Tcf transcriptional signalling as a result of b-catenintranslocation to the cell periphery and not significantchanges in the total amount of b-catenin (Faux et al.,2004). We propose that axin, rather than APC, is thecrucial player in b-catenin degradation and APC maycontribute by regulating the cytoplasmic b-catenin poolavailable to the destruction complex or by influencingthe rate of b-catenin degradation. APC is mutated in themajority of colon cancers and is the foremost tumoursuppressor in the colon. Given the functions for APC incell migration, cell adhesion and chromosomal stability(Fodde, 2003), the tumour suppressor activity of APCand its role in Wnt signalling is likely to be morecomplex than simply regulating the nuclear levels of b-catenin (Polakis, 2000; Clevers, 2006). The challengenow is to determine how the activities of APC at thecytoskeleton are associated with Wnt signalling andtumour suppression.
Figure 6 Axin puncta formation is required for b-catenin degradation but not b-catenin phosphorylation. (a) Axin-RFP expression inSW480 cells resulted in b-catenin degradation in control cells (left panels) whereas b-catenin was recruited to axin puncta in MG132treated cells (right panels). (b) AxinDDIX-RFP did not form puncta and resulted in sequestration of b-catenin in the cytoplasm, butnot degradation, in both control and MG132-treated SW480 cells. The DIX domain is required for both puncta formation andb-catenin degradation. AxinDDIX expression in (c) SW480 and (d) Madin–Darby canine kidney (MDCK) cells resulted insequestration of P-b-cat in both control (left panels) and MG132-treated cells (right). Shown are confocal images of b-catenin (a, b) orP-b-cat (c, d) and composite images (b-catenin, P-b-cat—green, axin-RFP, axinDDIX-RFP—red, nuclei (46-diamidino-2-phenylindole; DAPI)—blue). Arrows indicate axinDDIX expressing cells where cytoplasmic P-b-cat is increased, dashed arrows denoteuntransfected cells. We found little evidence for axinDDIX-RFP puncta formation in either SW480 or MDCK cells. Scale bar, 20mm.(e) Fluorescence intensity of cytoplasmic P-b-cat from individual SW480 (left graph) and MDCK (right) cells with and withoutaxinDDIX was measured. Values shown are mean±s.e.m. of cytoplasmic P-b-cat fluorescence intensity. Note that proteasomeinhibition had little effect on the cytoplasmic sequestration of b-catenin.
Recruitment of APC and b-catenin to axin punctaMC Faux et al
5817
Oncogene

Materials and methods
Antibodies and plasmidsThe following antibodies were used: anti-APC (APC2,produced in our laboratory (Catimel et al., 2006)), anti-APC(H290; Santa Cruz, Santa Cruz, CA, USA), anti-CK1a(Santa Cruz), anti-axin-1 (Zymed, San Francisco, CA, USA),anti-phospho-b-catenin (Cell Signaling, Danvers, MA, USA,9561, pS33, pS37, pT41 and 9565, pT41, pS45), anti-b-catenin(19920/610153; BD Transduction Laboratories, San Jose, CA,USA), anti-GSK3b (G22320; BD Transduction Laboratories),anti-actin (clone AC-40; Sigma, St Louis, MS, USA), anti-b-tubulin (clone 2.1; Sigma) and anti-HA (262K#2362; CellSignaling). To generate C-terminally tagged APC-GFP,enhanced GFP was subcloned as a BamH1/Not1 fragmentinto pEF-mycAPC (Faux et al., 2004) after removing theSTOP. Expression of GFP at the C terminus did notcompromise the localization of APC nor, as shown previously(Sharma et al., 2006), its colocalization with binding proteinsb-catenin, EB1, KAP3a and DLG-1, suggesting that theC-terminal GFP does not interfere with APC interactions at itsC terminus. For expression of axin-mRFP, the cDNA encodingmonomeric RFP (mRFP1) (Campbell et al., 2002), gift from DrMark Prescot (Monash University, Australia), was subclonedinto pDsRed1-N1 (Clontech, Mountain View, CA, USA),replacing DsRed1 (referred to as pDsRed-mRFP1). The cDNAsencoding full-length mouse axin1 isoform1, 125–956 aa (Zenget al., 1997), was amplified from pCS2-6myc mouse axin (gift ofDr Frank Costantini, Columbia University Medical Centre, NY,USA) and an N-terminally extended mouse axin1 isoform1(12–956), was amplified from pCS2_MT-mouse axin (gift of DrDaniel Capelluto, Denver, USA) and both were subcloned intopDsRed-mRFP1. Both forms of axin-RFP formed punctaand were capable of recruiting APC-GFP. AxinDRGS-RFP(aa 190–353 deleted) and axinDDIX-RFP (aa 875–956 deleted)deletion constructs were created in two parts and subcloned intopDsRed-mRFP1 (Figure 1).
Cell culture, transfections and treatmentsMDCK and HEK293 T cells were grown in Dulbecco’smodified Eagle’s medium supplemented with 10% fetal calfserum (FCS) and 1% penicillin/streptomycin. SW480 cells(Leibovitz et al., 1976) were grown in RPMI supplementedwith 0.00108% 10–2 thioglycerol, 0.1U/ml insulin, 50 mg/mlhydrocortisone, 10% FCS and 1% penicillin/streptomycin.Cells were plated on 1.5mm optical glass coverslips in 35mmdishes. HEK293 T cells were plated on coverslips coated withfibronectin (1 mg/ml). MDCK cells were transfected usingLipofectamine 2000 (Invitrogen, Carlsbad, CA, USA); SW480and HEK293 T cells were transfected using FuGENE 6(Roche Molecular Biochemicals, Mannheim, Germany) with0.1–2 mg plasmid DNA per dish. Microtubule assemblyexperiments were performed as described (Nakamura et al.,2001). Transfected MDCK cells were incubated on ice for60min, and then at 30 1C for 0.5, 1, 2 and 5min and cells werefixed rapidly and immunostained as described (Faux et al.,2004). For inhibition of the proteasome, cells were treated with15 mM MG132 (Sigma) or dimethyl sulphoxide control, 6 h at37 1C. Antibody dilutions were as follows: 1:200 for axin,APC, GSK3-b, CK1-a; 1:400 for b-catenin; anti-pS33,pS37,pT41- and pT41,pS35-phospho-b-catenin antibo-dies were diluted 1:200 and combined to assess phosphorylatedb-catenin (P-b-cat), 1:500 for fluorescently-labelled Alexa-Fluor-488, -405 or -532 secondary antibodies (MolecularProbes Invitrogen, Carlsbad, CA, USA). In the case ofphospho-b-catenin antibodies, all experiments were performedwith anti-pS33,pS37,pT41- and pT41,pS35-phospho-b-catenin
antibodies separately and together with identical results, hencestaining with combined antibodies are shown. Where indi-cated, nuclei were visualized using 46-diamidino-2-phenylindole (Molecular Probes).
Confocal fluorescence microscopy and image analysisDirect fluorescence (GFP and RFP) and immunofluorescencestaining were detected in successive focal planes using Bio-RadMRC-1024 dual system (Hemel Hempstead, UK) and NikonC1 confocal microscopes. Double- and triple-label images weredetected using standard filter sets and laser lines. Cells wereimaged with Nikon Plan Apo � 60 (NA1.4) or Nikon TIRF� 100 (NA1.45) oil immersion lenses. Fluorescence intensitymeasurements were performed using ImageJ (NIH, Bethesda,USA). Following background subtraction, individual cellswere selected and mean fluorescence intensity measured andmultiplied by selected area to obtain total fluorescence. Forquantitative cytoplasmic fluorescence, the fluorescence inten-sity of four randomly selected areas of identical size takenfrom the cytoplasm of individual cells were measured andmean fluorescence intensity determined using ImageJ software.
Fluorescence lifetime imaging micrsoscopy analysisFLIM experiments were carried out as described (Claytonet al., 2005; Hanley and Clayton, 2005). The phase andmodulation were determined from a series of images taken at12 phase settings using software provided by the manufacturerand converted to phase lifetimes with correction for photo-bleaching as described (van Munster and Gadella, 2004).Fluorescein dianion (lifetime 4.1 ns) was used as a reference(Magde et al., 1999). Two approaches were used to analyse theFLIM data. In the first approach the decrease in APC-GFPlifetime in a FLIM image was used to infer the existence ofFRET between APC-GFP and axin-mRFP in axin puncta butnot elsewhere in the cell or not in control cells. In the secondapproach APC-GFP at cellular protrusions were distinguishedfrom cytoplasmic axin-puncta-associated APC-GFP and themean phase lifetime in those two regions were determined for anumber of cells. The cellular population average and standarddeviations were used in an unpaired t-test to determinewhether the lifetimes in the different regions were statisticallydifferent with a 95% confidence interval.
Time-lapse imaging and analysisAPC-GFP and N-terminally extended axin(12–956)-RFPtransfected HEK293 T cells were imaged 24 h post transfectionin indicator-free DME containing 10% FCS in an humidifiedatmosphere containing CO2 saturation at 37 1C using aBioRad MRC-1024 dual system confocal microscope (seeabove) equipped with a Nikon Plan Apo � 60 waterimmersion with coverslip correction collar (NA1.2). Time-lapse images were collected sequentially over 60–90 cyclesevery 12–15 s and saved as uncompressed AVI files. Singleparticles from time-lapse images were filtered and trackedusing the ImageJ plugin ‘Spot Tracker’ (Sage et al., 2005). Theanalysis of the trajectories follows essentially the sameapproach as the Cherry laboratory (Wilson et al., 1996). Foreach track through i images, the MSD is calculated fordifferent time intervals ndt, n¼ 1,2,y, n being the number ofimages between which the displacement is computed, and dt isthe time interval between images. These calculations weremade up to noi/2 to prevent insufficient averaging fromoverlapping segments of the track. The MSD values wereplotted as a function of time (ndt) and the particle mobilityassigned to simple diffusion (diffusion coefficient D, Equationb), diffusionþ directed motion (velocity v, Equation b) (Saxton
Recruitment of APC and b-catenin to axin punctaMC Faux et al
5818
Oncogene

and Jacobson, 1997) motion or an approximation to con-strained diffusion (v is an imaginary number in Equation b).
MSD ¼ 4D ðndtÞ ðaÞ
MSD ¼ 4D ðndtÞ þ v2 ðndtÞ2 ðbÞ
Acknowledgements
We thank Dr M Prescott, Dr F Costantini and Dr D Capellutofor cDNA constructs. MCF, MJL and AWB are supported byNH&MRC program grant 234703; AHAC is an NH&MRCRD Wright Research Fellow.
References
Aberle H, Bauer A, Stappert J, Kispert A, Kemler R. (1997). Beta-catenin is a target for the ubiquitin-proteasome pathway. EMBO J
16: 3797–3804.Barth AI, Siemers KA, Nelson WJ. (2002). Dissecting interactionsbetween EB1, microtubules and APC in cortical clusters at theplasma membrane. J Cell Sci 115: 1583–1590.
Behrens J, Jerchow BA, Wurtele M, Grimm J, Asbrand C, Wirtz Ret al. (1998). Functional interaction of an axin homolog, conductin,with beta-catenin, APC, and GSK3beta. Science 280: 596–599.
Bilic J, Huang YL, Davidson G, Zimmermann T, Cruciat CM,Bienz M et al. (2007). Wnt induces LRP6 signalosomes andpromotes dishevelled-dependent LRP6 phosphorylation. Science
316: 1619–1622.Campbell RE, Tour O, Palmer AE, Steinbach PA, Baird GS,Zacharias DA et al. (2002). A monomeric red fluorescent protein.Proc Natl Acad Sci USA 99: 7877–7882.
Catimel B, Nice EC, Karrlander M, Ross J, Catimel J, Burgess AWet al. (2006). Purification and characterization of a high specificitypolyclonal antibody to the adenomatous polyposis coli tumoursuppressor protein. Biomed Chromatogr 20: 569–575.
Ciani L, Krylova O, Smalley MJ, Dale TC, Salinas PC. (2004).A divergent canonical WNT-signaling pathway regulatesmicrotubule dynamics: dishevelled signals locally to stabilizemicrotubules. J Cell Biol 164: 243–253.
Clayton AH, Walker F, Orchard SG, Henderson C, Fuchs D,Rothacker J et al. (2005). Ligand-induced dimer-tetramer transitionduring the activation of the cell surface epidermal growth factorreceptor-A multidimensional microscopy analysis. J Biol Chem 280:30392–30399.
Clevers H. (2006). Wnt/beta-catenin signaling in development anddisease. Cell 127: 469–480.
Cliffe A, Hamada F, Bienz M. (2003). A role of Dishevelled inrelocating Axin to the plasma membrane during wingless signaling.Curr Biol 13: 960–966.
Cong F, Schweizer L, Varmus H. (2004). Wnt signals across theplasma membrane to activate the beta-catenin pathway by formingoligomers containing its receptors, Frizzled and LRP. Development
131: 5103–5115.Dayanandan R, Butler R, Gordon-Weeks PR, Matus A, Kaech S,Lovestone S et al. (2003). Dynamic properties of APC-decoratedmicrotubules in living cells. Cell Motil Cytoskeleton 54: 237–247..
Fagotto F, Jho E, Zeng L, Kurth T, Joos T, Kaufmann C et al. (1999).Domains of axin involved in protein-protein interactions, Wntpathway inhibition, and intracellular localization. J Cell Biol 145:741–756.
Faux MC, Ross JL, Meeker C, Johns T, Ji H, Simpson RJ et al. (2004).Restoration of full-length adenomatous polyposis coli (APC)protein in a colon cancer cell line enhances cell adhesion. J CellSci 117: 427–439.
Fodde R. (2003). The multiple functions of tumour suppressors: it’s allin APC. Nat Cell Biol 5: 190–192.
Hanley QS, Clayton AH. (2005). AB-plot assisted determination offluorophore mixtures in a fluorescence lifetime microscope usingspectra or quenchers. J Microsc 218: 62–67.
Hart M, Concordet JP, Lassot I, Albert I, del los Santos R, Durand Het al. (1999). The F-box protein beta-TrCP associates withphosphorylated beta-catenin and regulates its activity in the cell.Curr Biol 9: 207–210.
Hart MJ, de los Santos R, Albert IN, Rubinfeld B, Polakis P. (1998).Downregulation of beta-catenin by human Axin and its associationwith the APC tumor suppressor, beta-catenin and GSK3 beta. Curr
Biol 8: 573–581.Hsu W, Zeng L, Costantini F. (1999). Identification of a domain of
Axin that binds to the serine/threonine protein phosphatase 2A anda self-binding domain. J Biol Chem 274: 3439–3445.
Kawasaki Y, Senda T, Ishidate T, Koyama R, Morishita T, IwayamaY et al. (2000). Asef, a link between the tumor suppressor APC andG-protein signaling. Science 289: 1194–1197.
Kishida S, Yamamoto H, Hino S, Ikeda S, Kishida M, Kikuchi A.(1999). DIX domains of Dvl and axin are necessary for proteininteractions and their ability to regulate beta-catenin stability. Mol
Cell Biol 19: 4414–4422.Kishida S, Yamamoto H, Ikeda S, Kishida M, Sakamoto I, Koyama S
et al. (1998). Axin, a negative regulator of the wnt signalingpathway, directly interacts with adenomatous polyposis coliand regulates the stabilization of beta-catenin. J Biol Chem 273:10823–10826.
Kita K, Wittmann T, Nathke IS, Waterman-Storer CM. (2006).Adenomatous polyposis coli on microtubule plus ends in cellextensions can promote microtubule net growth with or withoutEB1. Mol Biol Cell 17: 2331–2345.
Kitagawa M, Hatakeyama S, Shirane M, Matsumoto M, Ishida N,Hattori K et al. (1999). An F-box protein, FWD1, mediatesubiquitin-dependent proteolysis of beta-catenin. EMBO J 18:2401–2410.
Korinek V, Barker N, Morin PJ, van Wichen D, de Weger R, Kinzler KWet al. (1997). Constitutive transcriptional activation by a beta-catenin-Tcf complex in APC�/� colon carcinoma. Science 275:1784–1787.
Kroboth K, Newton IP, Kita K, Dikovskaya D, Zumbrunn J,Waterman-Storer CM et al. (2007). Lack of adenomatouspolyposis coli protein correlates with a decrease in cell migrationand overall changes in microtubule stability. Mol Biol Cell 18:910–918.
Langford KJ, Askham JM, Lee T, Adams M, Morrison EE. (2006).Examination of actin and microtubule dependent APC localisationsin living mammalian cells. BMC Cell Biol 7: 3.
Latres E, Chiaur DS, Pagano M. (1999). The human F box proteinbeta-Trcp associates with the Cul1/Skp1 complex and regulates thestability of beta-catenin. Oncogene 18: 849–854.
Lee E, Salic A, Kruger R, Heinrich R, Kirschner MW. (2003). Theroles of APC and Axin derived from experimental and theoreticalanalysis of the Wnt pathway. PLoS Biol 1: E10.
Leibovitz A, Stinson JC, McCombs III WB, McCoy CE, Mazur KC,Mabry ND. (1976). Classification of human colorectal adenocarci-noma cell lines. Cancer Res 36: 4562–4569.
Levina E, Oren M, Ben-Ze’ev A. (2004). Downregulation of beta-catenin by p53 involves changes in the rate of beta-cateninphosphorylation and Axin dynamics. Oncogene 23: 4444–4453.
Logan CY, Nusse R. (2004). The Wnt signaling pathway indevelopment and disease. Annu Rev Cell Dev Biol 20: 781–810.
Magde D, Rojas GE, Seybold PG. (1999). Solvent dependence of thefluorescence lifetimes of xanthene dyes. Photochem Photobiol 70:737–744.
Mimori-Kiyosue Y, Shiina N, Tsukita S. (2000). Adenomatouspolyposis coli (APC) protein moves along microtubules and
Recruitment of APC and b-catenin to axin punctaMC Faux et al
5819
Oncogene

concentrates at their growing ends in epithelial cells. J Cell Biol 148:505–518.
Munemitsu S, Albert I, Souza B, Rubinfeld B, Polakis P. (1995).Regulation of intracellular beta-catenin levels by the adenomatouspolyposis coli (APC) tumor-suppressor protein. Proc Natl Acad Sci
USA 92: 3046–3050.Munemitsu S, Souza B, Muller O, Albert I, Rubinfeld B, Polakis P.
(1994). The APC gene product associates with microtubules in vivoand promotes their assembly in vitro. Cancer Res 54: 3676–3681.
Nakamura M, Zhou XZ, Lu KP. (2001). Critical role for the EB1 andAPC interaction in the regulation of microtubule polymerization.Curr Biol 11: 1062–1067.
Nakamura T, Hamada F, Ishidate T, Anai K, Kawahara K,Toyoshima K et al. (1998). Axin, an inhibitor of the Wnt signallingpathway, interacts with beta-catenin, GSK-3beta and APC andreduces the beta-catenin level. Genes Cells 3: 395–403.
Nathke IS. (2004). The adenomatous polyposis coli protein: the Achillesheel of the gut epithelium. Annu Rev Cell Dev Biol 20: 337–366.
Nathke IS, Adams CL, Polakis P, Sellin JH, Nelson WJ. (1996). Theadenomatous polyposis coli tumor suppressor protein localizes toplasma membrane sites involved in active cell migration. J Cell Biol
134: 165–179.Orford K, Crockett C, Jensen JP, Weissman AM, Byers SW. (1997).
Serine phosphorylation-regulated ubiquitination and degradation ofbeta-catenin. J Biol Chem 272: 24735–24738.
Park TJ, Gray RS, Sato A, Habas R, Wallingford JB. (2005).Subcellular localization and signaling properties of dishevelled indeveloping vertebrate embryos. Curr Biol 15: 1039–1044.
Penman GA, Leung L, Nathke IS. (2005). The adenomatous polyposiscoli protein (APC) exists in two distinct soluble complexes withdifferent functions. J Cell Sci 118: 4741–4750.
Polakis P. (1997). The adenomatous polyposis coli (APC) tumorsuppressor. Biochim Biophys Acta 1332: F127–F147.
Polakis P. (2000). Wnt signaling and cancer. Genes Dev 14: 1837–1851.Sage D, Neumann FR, Hediger F, Gasser SM, Unser M. (2005).
Automatic tracking of individual fluorescence particles: applicationto the study of chromosome dynamics. IEEE Trans Image Process
14: 1372–1383.Sakanaka C, Williams LT. (1999). Functional domains of axin.
Importance of the C terminus as an oligomerization domain. J Biol
Chem 274: 14090–14093.Saxton MJ, Jacobson K. (1997). Single-particle tracking: applications
to membrane dynamics. Annu Rev Biophys Biomol Struct 26:373–399.
Schneikert J, Behrens J. (2007). The canonical Wnt signalling pathwayand its APC partner in colon cancer development. Gut 56: 417–425.
Schwarz-Romond T, Fiedler M, Shibata N, Butler PJ, Kikuchi A,Higuchi Y et al. (2007a). The DIX domain of Dishevelled confers
Wnt signaling by dynamic polymerization. Nat Struct Mol Biol 14:484–492.
Schwarz-Romond T, Merrifield C, Nichols BJ, Bienz M. (2005). TheWnt signalling effector Dishevelled forms dynamic protein assem-blies rather than stable associations with cytoplasmic vesicles. J Cell
Sci 118: 5269–5277.Schwarz-Romond T, Metcalfe C, Bienz M. (2007b). Dynamic
recruitment of axin by Dishevelled protein assemblies. J Cell Sci
120: 2402–2412.Sharma M, Leung L, Brocardo M, Henderson J, Flegg C, Henderson
BR. (2006). Membrane localization of adenomatous polyposis coliprotein at cellular protrusions: targeting sequences and regulationby beta-catenin. J Biol Chem 281: 17140–17149.
Smalley MJ, Sara E, Paterson H, Naylor S, Cook D, Jayatilake H et al.(1999). Interaction of axin and Dvl-2 proteins regulatesDvl-2-stimulated TCF-dependent transcription. EMBO J 18:2823–2835.
Su LK, Burrell M, Hill DE, Gyuris J, Brent R, Wiltshire R et al.(1995). APC binds to the novel protein EB1. Cancer Res 55:2972–2977.
van Munster EB, Gadella Jr TW. (2004). Suppression of photobleach-ing-induced artifacts in frequency-domain FLIM by permutation ofthe recording order. Cytometry A 58: 185–194.
Watanabe T, Wang S, Noritake J, Sato K, Fukata M, Takefuji M et al.(2004). Interaction with IQGAP1 links APC to Rac1, Cdc42, andactin filaments during cell polarization and migration. Dev Cell 7:871–883.
Wen Y, Eng CH, Schmoranzer J, Cabrera-Poch N, Morris EJ, ChenM et al. (2004). EB1 and APC bind to mDia to stabilizemicrotubules downstream of Rho and promote cell migration. Nat
Cell Biol 6: 820–830.Wiechens N, Heinle K, Englmeier L, Schohl A, Fagotto F. (2004).
Nucleo-cytoplasmic shuttling of Axin, a negative regulator of theWnt-beta-catenin Pathway. J Biol Chem 279: 5263–5267.
Wilson KM, Morrison IE, Smith PR, Fernandez N, Cherry RJ. (1996).Single particle tracking of cell-surface HLA-DR moleculesusing R-phycoerythrin labeled monoclonal antibodies andfluorescence digital imaging. J Cell Sci 109(Part 8):2101–2109.
Winston JT, Strack P, Beer-Romero P, Chu CY, Elledge SJ, HarperJW. (1999). The SCFbeta-TRCP-ubiquitin ligase complex associatesspecifically with phosphorylated destruction motifs in IkappaBalphaand beta-catenin and stimulates IkappaBalpha ubiquitination invitro. Genes Dev 13: 270–283.
Zeng L, Fagotto F, Zhang T, Hsu W, Vasicek TJ, Perry III WL et al.(1997). The mouse Fused locus encodes Axin, an inhibitor of theWnt signaling pathway that regulates embryonic axis formation.Cell 90: 181–192.
Supplementary Information accompanies the paper on the Oncogene website (http://www.nature.com/onc)
Recruitment of APC and b-catenin to axin punctaMC Faux et al
5820
Oncogene
Copyright © 2022 FDOKUMEN




















