
A role for nuclear β-catenin in SNRI antidepressant-induced hippocampal cell proliferation
50
TITLE PAGE Title: A role for nuclear β-catenin in SNRI antidepressant- induced hippocampal cell proliferation. Authors: Ricardo Mostany 1 PhD, Elsa M. Valdizán MD PhD, Angel Pazos MD PhD. Authors’ addresses: Departamento de Fisiología y Farmacología, Instituto de Biomedicina y Biotecnología de Cantabria (IBBTEC), Universidad de Cantabria-CSIC-IDICAN, 39011 Santander, Cantabria, Spain. 1 Present address: Department of Neurology, University of California, Los Angeles. Reed Neurological Research Center, 710 Westwood Plz. Los Angeles, CA 90095 USA. Corresponding author: Dr. Angel Pazos. Department of Physiology and Pharmacology. Universidad de Cantabria. Av. Cardenal Herrera Oria s/n, 39011 Santander (Cantabria), Spain. Tel: +34 942 201 985; Fax: +34 942 201 903; e-mail: [email protected] Angel Pazos 1
Transcript of A role for nuclear β-catenin in SNRI antidepressant-induced hippocampal cell proliferation

TITLE PAGE
Title: A role for nuclear β-catenin in SNRI antidepressant-
induced hippocampal cell proliferation.
Authors: Ricardo Mostany1 PhD, Elsa M. Valdizán MD PhD, Angel
Pazos MD PhD.
Authors’ addresses: Departamento de Fisiología y Farmacología,
Instituto de Biomedicina y Biotecnología de Cantabria
(IBBTEC), Universidad de Cantabria-CSIC-IDICAN, 39011
Santander, Cantabria, Spain.
1 Present address: Department of Neurology, University of
California, Los Angeles. Reed Neurological Research Center,
710 Westwood Plz. Los Angeles, CA 90095 USA.
Corresponding author: Dr. Angel Pazos.
Department of Physiology and Pharmacology. Universidad de
Cantabria. Av. Cardenal Herrera Oria s/n, 39011 Santander
(Cantabria), Spain.
Tel: +34 942 201 985; Fax: +34 942 201 903; e-mail:
Angel Pazos 1

Running title____________________________45 characters
Abstract 250/248 words
Introduction: 750/434
Methods 1500/1459
Text 4197 words
Figures 8
Tables 1
Angel Pazos 2

Abstract sin apartados tienes el anterior después de este
Background: Increasing evidences have been accumulated
suggesting a role for antidepressant drugs (ADs) as
hippocampal neurogenesis enhancers, but the information about
the mechanisms involved in this response is limited. We have
studied in the hippocampus the effects of chronic treatment
with the dual reuptake inhibitor (SNRI) venlafaxine on both
cellular proliferation rate and expression of key effectors of
several signaling pathways.
Methods: Adult Wistar rats were chronically (14 days) treated
either with subcutaneous saline or venlafaxine (10 and 40
mg/kg). Hippocampal cell proliferation was studied by BrdU
immunohistochemistry. -catenin localization was analyzed by
both cellular immunohistochemistry and immunoelectron
microscopy. Protein levels of -catenin, AKT1, ERK1/2,
pERK1/2, CREB and pCREB were determined by Western blot
analysis.
Results: Increased cell proliferation in subgranular zone was
achieved after chronic treatment with venlafaxine (40
mg/kg/day). However, significant increases in the
immunoreactivity of hippocampal β-catenin in SGZ were already
Angel Pazos 3

detected after administration of a lower dose (10 mg/kg/day).
Western blot and immunoelectron microscopy studies
demonstrated an increased presence of β-catenin at the nuclear
level. Cytosolic AKT levels were also increased in
venlafaxine-treated animals. Furthermore, p-ERK2/ERK2 ratio
was increased in the hippocampus of treated animals, while no
differences in the levels of CREB and p-CREB were observed.
Conclusions: Chronic venlafaxine induces a hippocampal
proliferative effect, which involves the activation of
intracellular signaling through Wnt and AKT/PKB pathways,
resulting in an increase of the expression of cell cycle
regulator genes. These results illustrate the complexity of
the intracellular events underlying the neurogenetic responses
of ADs.
Angel Pazos 4

Abstract
Increasing evidences have been accumulated during recent years
suggesting a role for antidepressant drugs (ADs) as
hippocampal neurogenesis enhancers, but the information about
the transductional mechanisms involved in this response is
very limited. We have studied in the adult rat hippocampus the
effects of chronic treatment with the dual reuptake inhibitor
(SNRI) venlafaxine on both cellular proliferation rate and
expression of key effectors of several signaling pathways.
Increased cell proliferation (BrdU incorporation) in
subgranular zone (SGZ) was achieved after chronic treatment
with a high dose (40 mg/kg/day) of venlafaxine. However,
significant increases in the immunoreactivity of hippocampal
β-catenin in SGZ were already detected after administration of
a lower dose of the drug (10 mg/kg/day). Western blot and
immunoelectron microscopy studies demonstrated an increased
presence of β-catenin at the nuclear level. An increase in
cytosolic AKT levels was also observed in venlafaxine-treated
animals. These results suggest that the hippocampal
proliferative effect of chronic venlafaxine, only evident when
both 5-HT and NE reuptake systems are inhibited, requires a
Angel Pazos 5

strong activation of intracellular signaling through Wnt (β-
catenin translocation) and AKT/PKB pathways. This activation
would probably result in an increase of the expression of cell
cycle regulator genes. Furthermore p-ERK2/ERK2 rate was also
increased in the hippocampus of AD-treated animals, while no
differences in the levels of CREB and p-CREB were observed.
These results illustrate the complexity of the intracellular
events underlying the neurogenetic responses of ADs. They also
support the relevance of such effects for the therapeutic
effects of these drugs.
Keywords: venlafaxine, β-catenin, AKT, neurogenesis,
subgranular zone, hippocampus.
Angel Pazos 6

INTRODUCTION
The most common first step in the therapeutic action of
antidepressant drugs (ADs) is the increase in the monoamines
serotonin (5-HT; 5-hydroxytrytamine) and/or norepinephrine
(NE) extracellular levels (Richelson, 1991). This immediate
effect contrasts with the onset of appreciable clinical
improvement, which requires several weeks of AD treatment.
Trying to clarify this apparent mismatch, a large number of
studies about the trophic effects of AD drugs and
electroconvulsive therapy (ECS) have been reported in the
recent years (Altar, et al
2003; Altar, et al 2004; Russo-Neustadt, et al 2004). In this
sense, it is well documented that chronic AD treatment
enhances cell proliferation in adult rodent and nonhuman
primate subgranular zone (SGZ) of hippocampus and that the
time required for the differentiation and maturation of
newborn neurons correlates well with the appearance of
clinical response to the AD treatment (Malberg, et al 2000; D'Sa
and Duman, 2002; Perera, et al 2007). Furthermore, the existence
of a decrease in the hippocampal volume, restored after AD
treatment (Vermetten, et al 2003) has been reported in depressed
Angel Pazos 7

patients by some, but not all authors (Sheline, et al 1996;
Vythilingam, et al 2004). However, the molecular pathways that
control these processes are still unclear. Transduction
pathways promoting cell proliferation are the main targets for
studying the possible mediators for the effect of these drugs
in neurogenesis. Thus, the cAMP cascade has been suggested to
be involved in the neuroproliferative responses induced by ADs
but contradictory results have been reported on this issue
(Nibuya, et al 1996; Manier, et al 2002). In a similar way, the
role of MAPK pathway is under discussion (Tiraboschi, et al 2004;
Fumagalli , et al 2005) and no clear data are available with
respect to AKT/PKB signaling pathway. An emerging candidate to
play a key role in these processes is the Wnt pathway.
Activation of the canonical Wnt pathway leads to the
inhibition of GSK-3, allowing β-catenin to be stabilized in
the cytosol and translocated to the nucleus, where activates
transcription of target genes (Logan and Nusse, 2004). It has
been recently shown that this signaling pathway, a main
regulator of the hippocampal neurogenesis in the adult brain
(Lie, et al 2005), is modulated by chronic ECS treatment (Madsen,
et al 2003).
Angel Pazos 8

The degree of stimulation of cell proliferation varies with
the type of AD drug used (Malberg, et al 2000). The dual reuptake
inhibitor (5-HT and NE, SNRI) venlafaxine blocks both reuptake
processes, with a higher potency for the 5-HT component
(Bëíque, et al 1998) so it is possible to selectively act on the
5-HT transport or to affect both aminergic systems, depending
on the doses assayed for this AD. The goal of the present
study has been to evaluate, in the adult rat hippocampus, the
effects of chronic treatment with venlafaxine at two different
doses on both the hippocampal cellular proliferation and the
expression and cellular distribution of β-catenin, as well as
of other key effectors of several main signaling pathways
(cAMP, MAPK and PKB/AKT).
MATERIALS AND METHODS
Animals
Adult male albino Wistar rats (Harlan, Barcelona, Spain)
weighing 175-200 g were used for the present study. The
animals were housed at 22 ºC with a 12:12 light-dark cycle.
Food and water were provided ad libitum. All experimental
procedures were done according to the Spanish legislation and
Angel Pazos 9

the European Communities Council Directive on “Protection of
Animals Used in Experimental and Other Scientific Purposes”
(86/609/EEC).
Antidepressant treatment and BrdU administration
Rats were implanted subcutaneously with an osmotic minipump
Alzet 2002 (Alza Corp., Palo Alto, CA) which delivered 0.5 µl
per hour during 14 days. Twenty-two animals per group were
treated either with saline, 10 mg/kg/day venlafaxine, or 40
mg/kg/day venlafaxine. Venlafaxine HCl was extracted from
Vandral 75 (Wyeth-Farma, S.A., Madrid, Spain). Minipumps were
removed the fourteenth day of treatment.
For immunohistochemical analysis of cell proliferation,
animals received 5-bromo-2’-deoxyuridine (BrdU; 4 x 75 mg/kg
every 2 hours, i.p.; Sigma, Madrid, Spain) in sterile 0.9%
NaCl solution the last day of antidepressant treatment.
Immunohistochemistry
Twenty-four hours after the last BrdU injection, rats were
deeply anaesthetized with sodium pentobarbital (100 mg/kg,
i.p.) and transcardially perfused with saline followed by 4%
Angel Pazos 10

cold paraformaldehyde in PBS. Brains were removed, postfixed
overnight at 4 ºC and transferred to 30% sucrose in PBS at 4
ºC. Serial coronal sections (45 µm) of the brains were
obtained through the entire hippocampus on a cryostat and
stored at –20 ºC in cryoprotectant solution (25% glycerol, 25%
ethylene glycol in PBS, pH 7.4).
BrdU inmunohistochemistry was carried out following a
previously described protocol (Malberg, et al 2000). Free-
floating sections from animals from the three experimental
groups (n = 9) were incubated for 2 h in 50% formamide/2x SSC
at 65 ºC, followed by incubation in 2 N HCl for 30 min. Then
sections were incubated for 10 min in 0.1 M borate buffer.
After washing in PBS, sections were incubated in 1% H2O2 for 30
min to remove endogenous peroxidases. After several rinses in
PBS, sections were incubated in PBS/0.2% Triton X-100/5% goat
serum (PBS-TS) for 30 min and then incubated with monoclonal
mouse anti-BrdU (1:600; Roche Diagnostics, Barcelona, Spain)
overnight at 4 ºC. After several rinses in PBS-TS, sections
were incubated for 2 h with biotinylated horse anti-mouse IgG
secondary antibody (4 µg/ml; Vector Laboratories, Burlingame,
CA), followed by amplification with avidin-biotin complex
Angel Pazos 11

(Vector Laboratories). BrdU positive (BrdU+) cells were labeled
using DAB as chromogen (Vector Laboratories).
For quantification of BrdU+ cells, every sixth section
throughout the hippocampus was processed and mounted on coded
slides. BrdU+ cells in the SGZ of the dentate gyrus were
counted by an experimenter blinded to the study code under a
light microscope (Carl Zeiss Axioskop 2 Plus) at 400x
magnification. The total number of BrdU+ cells per section was
determined and multiplied by 6 to obtain the total number of
BrdU+ cells per hippocampus. Data were analyzed using ANOVA and
Student-Newman-Keuls post hoc test using GraphPad Prism
(GraphPad Software, Inc. San Diego, CA). Statistical
significance was set at p < 0.05.
β-catenin immunohistochemistry was carried out in adjacent
sections of those used in the BrdU labeling studies (n = 9).
After several rinses in PBS, sections were boiled in
microwaves in 10 mM citric acid, pH 6.0 for 10 min. Following
several rinses in PBS, sections were incubated in methanol and
3% H2O2. Then sections were transferred to blocking buffer
(PBS-TS) for 30 min and incubated for 2 d at 4 ºC with mouse
anti-β-catenin (1:500; BD Biosciences, Madrid, Spain). After
Angel Pazos 12

washes in PBS-TS sections were incubated with biotinylated
horse anti-mouse IgG secondary antibody followed by peroxidase
detection with DAB or FITC conjugated donkey anti-mouse IgG
secondary antibody (1:250; Jackson ImmunoResearch
Laboratories, Inc., West Grove, PA), followed by propidiun
iodide (Sigma) counterstain.
The immunostaining of β-catenin after peroxidase method was
evaluated by an observer blinded to the study code in at least
four sections from each animal corresponding to interaural
stereotaxic coordinates ranging 4.48-5.70 mm (Paxinos and
Watson, 1982). For this purpose, the section with the highest
number of positive clusters in the SGZ was arbitrarily
considered as containing the maximal level of expression. The
remaining sections were ranged according to the following: +,
number of positive clusters < 25% of that observed in the
section with the maximal number; ++, ranging 25-50%; +++,
ranging 50-75%; ++++, > 75% with respect the section with the
highest density.
For BrdU/β-catenin double labeling, frozen sections, 14 µm
thick, were mounted on gelatine-coated slides and fixed in
absolute ethanol for 20 min at –20 ºC (n = 5). Then, sections
Angel Pazos 13

were boiled in microwaves in citric acid followed by several
PBS rinses. Sections were blocked with PBS-TS for 30 min and
then incubated with monoclonal rat anti-BrdU (1:400; Serotec
Ltd., Oxford, UK) overnight at 4 ºC. Sections were incubated
with Texas Red conjugated donkey anti-rat (1:250; Jackson
ImmunoResearch Laboratories, Inc.) secondary antibody for 2 h.
After exhaustive washes in PBS-TS, sections were incubated
overnight with monoclonal mouse anti-β-catenin (1:500) primary
antibody at 4 ºC and then with FITC conjugated donkey anti-
mouse secondary antibody (1:250) for 2 h, followed by DAPI
(Roche Diagnostics) counterstain.
Subcellular fractionation and Western blot
Animals from each experimental group (n= 5) were killed by
decapitation, their brains being removed from skulls and
hippocampi dissected and rapidly frozen at -80 ºC. Every
sample was thawed and homogenized (1:15, 500-600 µl aprox.) by
using a Potter homogenizer provided with a loosely fitting
Teflon pestle in homogenization buffer (10 mM Hepes-HCl, pH
7.9, 1.5 mM MgCl2, 10 mM KCl) containing the following protease
and phosphatase inhibitors: 1 mM PMSF; 10 µl/ml aprotinin; 10
Angel Pazos 14

µg/ml leupetin; 10 µg/ml pepstatin A; 10 µg/ml antipain; 10
µg/ml chymostatin; 5 µg/ml trypsin inhibitor; 1 mM NaV; 1 mM
NaF; 1 mM cantharidin; and 10 µM E-64. After homogenization,
250 µl of homogenate were lysated in lysis buffer
(homogenization buffer containing 1% Igepal; 0.1% sodium
deoxicholate, 0.2% SDS, and 0.1% Triton X-100) 30 min on ice
for the total cell lysate (TCL). The remaining homogenate
(250-300 µl) was processed for subcellular fractionation.
Homogenate was centrifuged at 1000 g for 10 min at 4 ºC, and
the resulting supernatant (S1) and pellet (P1) separated. The
S1 was ultra-centrifuged at 100 000 g for 15 min at 4 ºC,
resulting in a supernatant (S2) cytosolic fraction, that was
lysated and a pellet P2 membrane fraction, resuspended in
buffer containing detergents and protease and phosphatase
inhibitors and centrifuged 10 min at 14 000 g. Nuclear
proteins were isolated by high salt extraction from P1
fraction. P1 fraction was homogenized in 20 mM Hepes pH 7.9,
0.45 M NaCl, 1 mM EDTA containing protease and phosphatase
inhibitors and incubated in ice for 30 min. Solubilized
proteins were recovered in the supernatant after
centrifugation at 14 000 g for 10 min at 4 ºC. Protein
Angel Pazos 15

quantification was performed according to the Lowry method
(Lowry, et al 1951).
Thirty-fifty µg of protein were resolved on 12.5% SDS-PAGE and
transferred to PVDF (non-phosphorylated proteins) or to
nitrocellulose (phosphorylated proteins) membranes. Membranes
were incubated in the following primary antibodies: mouse
anti--catenin (1:1000), mouse anti-GAPDH (1:2000), mouse
anti-histone H1 (1:200), mouse anti-AKT1 (1:1000) and rabbit
anti-ERK1/2 (1:2000), from Santa Cruz Biotechnology, Inc.
Heidelberg, Germany; mouse anti-pERK1/2 (1:10 000) and rabbit
anti-actin (1:2000) from Sigma; rabbit anti-CREB (1:1000) and
rabbit anti-pCREB (1:1000), from Upstate, Charlottesville, VA.
After extensive washings in TBS-T (TBS/0.05% Tween 20)
membranes were incubated with horseradish peroxidase
conjugated secondary antibodies. Secondary antibodies were
detected with ECL Advance kit (GE Healthcare Europe GmbH,
Munich, Germany). Anti-GAPDH (cytosolic marker) and anti-
histone H1 (nuclear marker) antibodies were used to discard
subcellular fraction contaminations. Blot quantitations were
performed by densitometric scanning using Scion Image
Software. The densitometry values were normalized with respect
Angel Pazos 16

to the values obtained with anti-actin antibody. Data for
every sample was the mean of at least two independent
experiments. Statistical analysis was performed using ANOVA
following Student-Newman-Keuls post hoc test.
Immunogold electron microscopy
Vibratome brain sections containing dentate gyrus (400 µm)
were obtained from transcardially perfused (4%
paraformaldehyde) animals from the three experimental groups
(n=3). The tissue samples were then washed with 0.1 M
cacodylate buffer, dehydrated in increasing concentrations of
methanol at –20 ºC, embedded in Lowicryl K4M at -20 ºC and
polymerized under ultraviolet irradiation. Ultrathin sections
were mounted on nickel grids and sequentially incubated with
0.1 M glycine in PBS for 15 min, 5% BSA in PBS for 30 minutes
and mouse anti--catenin (1:50) antibody (diluted in 50 mM
Tris HCl, pH 7.6, containing 1% BSA and 0.1 M glycine) for 1h
at 37 ºC. After washing, the sections were incubated with goat
anti-mouse antibody coupled to 15 nm gold particles (British
BioCell International, Cardiff, UK; 1:50 in PBS containing 1%
BSA). After immunogold labeling, the grids were stained with
Angel Pazos 17

lead citrate and uranyl acetate and examined with a Philips
EM208 electron microscope operated at 60kV.
RESULTS
Effect of venlafaxine on cell proliferation
The analysis of the number of BrdU+ cells showed that chronic
(14 d) venlafaxine treatment produces a significant increase
in cell proliferation in SGZ of dentate gyrus, only when a
high dose (40 mg/kg/day) of this AD was administered (4888 ±
266; 44.8 ± 8.4% increase vs saline group, 3374 ± 355; p <
0.01; Figure 1). No statistically significant changes in cell
proliferation rate were observed in the animals receiving 10
mg/kg/day of venlafaxine (3621 ± 335; 7.3 ± 10.6% increase vs
saline group). Furthermore, significant differences in the
cell proliferation rates were observed when comparing 10 and
40 mg/kg/day venlafaxine dose groups (34.9 ± 7.9%; p < 0.05).
Modifications in the expression patterns of main efector
proteins of signal transduction pathways after venlafaxine
treatment
Angel Pazos 18

Measurement of β-catenin expression in hippocampal total
homogenates revealed significant higher levels of this protein
after chronic treatment with either doses of venlafaxine
compared with saline treated rats (73 ± 13%, p < 0.05 and 175
± 36%, p < 0.001 increases for 10 and 40 mg/kg/day doses
respectively; Figure 2a and 2b). After cellular fractionation,
β-catenin was detected in the membrane-associated and nuclear
fractions. In both of them, significant increases in the
levels of this protein were found after chronic treatment at
either doses of venlafaxine when compared with saline rats,
these increases being more pronounced in the nuclear fraction.
Chronic treatment with 40 mg/kg/day resulted in amounts of
this protein higher than those observed for 10 mg/kg/day in
the two cellular fractions studied (62 ± 25%, p < 0.05 in
membrane-associated fraction, 88 ± 9%, p < 0.001 in nuclear
fraction).
Detectable AKT protein levels were obtained in the cytosolic
and nuclear fractions, as well as in the TCL (Figure 2b and
2c). A dose-dependent increase in the levels of expression of
this protein was observed in the total homogenate (28 ± 11%, p
= n.s.; 84 ± 31%, p < 0.05, for 10 and 40 mg/kg/day doses
Angel Pazos 19

respectively). In the cytosolic subfraction, increased levels
of AKT were found in tissues from animals chronically treated
at either venlafaxine doses (47 ± 13%, p < 0.05 and 69 ± 23%,
p < 0.05). In contrast, a significant dose-dependent decrease
in the amount of AKT was detected in the nuclear fraction (-22
± 6%, p < 0.05 and -38 ± 8%, p < 0.01).
No significant changes in the levels of expression of both
CREB and p-CREB were observed in the nuclear fractions of
animals chronically treated with either dose venlafaxine when
compared with saline group (Figure 3). Phosphorylated
CREB/CREB ratio was not affected either.
Immunoblot analysis of the expression of the two isoforms of
ERK protein (ERK1, ERK2) in total homogenates (Figure 4)
showed no change after chronic treatment with 10 or 40
mg/kg/day venlafaxine with respect to saline administered
rats. On the other hand, increased levels of the
phosphorylated form (activated form) of both isoforms of this
protein were observed in TCL after the AD treatment (96 ± 8%,
p < 0.05 and 141 ± 49%, p < 0.01 for p-ERK1 for 10 and 40
mg/kg/day, respectively. 80 ± 12%, p < 0.05 and 101 ± 38%, p <
0.05 for p-ERK2 for 10 and 40 mg/kg/day, respectively). In
Angel Pazos 20

this sense, p-ERK1/ERK1 and p-ERK2/ERK2 ratios showed at least
a 1.5 fold change after venlafaxine chronic treatment.
Increased immunoreactivity of β-catenin in the SGZ after
chronic treatment with venlafaxine
β-Catenin staining throughout the hippocampus presented a
distribution pattern characterized by a weak immunostaining
over the GCL and a positive immunostained clusters detected in
the SGZ of the dentate gyrus. Saline group presented levels of
immunostaining under 25% with respect to the highest
immunoreactivity found. Animals treated with 10 mg/kg/day
venlafaxine showed a mean number of immunopositive clusters
per section ranging 50-75% with respect the highest
immunoreactivity found. Mean number of immunopositive clusters
for β-catenin in the SGZ of animals treated with 40 mg/kg/day
venlafaxine was over 75% with respect the highest
immunoreactivity found (Table 1 and Figure 5).
The clusters of β-catenin immunolabeling observed in the SGZ
were studied in order to detect nuclear β-catenin presence. No
presence of this protein could be detected in nucleus from SGZ
cells when they were observed under confocal microscopy
Angel Pazos 21

(Figure 6) in any sample from animals from the three
experimental groups. GCL cells nuclei neither were β-catenin
positive when examined.
No nuclear BrdU+/β-catenin+ colocalization could be observed in
any sample analyzed, but all BrdU+ nuclei observed showed high
β-catenin labeling surrounding them and were embedded in β-
catenin clusters (Figure 6).
Detection of nuclear β-catenin in SGZ cells by immunoelectron
microscopy
β-catenin presence in nuclei from SGZ cells could be observed
by immunoelectron microscopy. β-catenin presence and
distribution appeared in DNA euchromatinic regions, with no
presence in nucleoli or heterochromatin, compatible with a
transcription factor distribution pattern. The low density of
gold particles observed in nucleus from SGZ and CGL cells from
animals of saline group strongly contrasts with those observed
after chronic treatment with venlafaxine (Figure 7) showing an
increased number of gold particles together with an increase
in the number of particles per cluster.
Angel Pazos 22

DISCUSSION
The present study demonstrates for the first time that chronic
treatment with the SNRI venlafaxine increases both the
hippocampal cell proliferation and the translocation rate to
the nucleus of β-catenin, a main effector of the Wnt pathway.
The positive effects reported in the last years for AD drugs
and ECS on adult hippocampal cell proliferation (Malberg, et al
2000; Madsen, et al 2000; Czéh, et al 2001; Sairanen, et al 2005;
Dranovsky and Hen, 2006) support a new hypothesis about the
mechanisms through AD exert their therapeutic actions.
Fluoxetine has been the AD drug more widely used in these
studies (Malberg, et al 2000; Sairanen, et al 2005; Malberg and
Duman, 2003; Santarelli, et al 2003). However, it is being
suggested that the use of dual reuptake inhibitors, such as
venlafaxine, could be of special interest in the treatment of
some major depression cases, under certain clinical conditions
(Tzanakaki, et al 2000; Stahl, et al 2002; Poirier and Boyer, 1999;
Mehtonen, et al 2000). Taking into account that venlafaxine shows
a relative preferential pattern of 5-HT transport inhibition
Angel Pazos 23

(Bëíque, et al 1998), we studied the state of cell proliferation
rate after 14 days of treatment, using two doses: 10
mg/kg/day, which mainly acts on 5-HT transporter, and 40
mg/kg/day, a dose that inhibits the reuptake process through
both 5-HT and NE transporters (Bëíque, et al 1998). A
significant increase in cell proliferation in SGZ was observed
in the animals administered with 40 mg/kg/day, while no effect
was found following the administration of 10 mg/kg/day. These
results are in contrast with previous data (Khawaja, et al 2004),
which showed augmentation of cell proliferation in rats
treated for 14 days with 10 mg/kg/day venlafaxine. Two
methodological differences in the experimental design could
explain this discrepancy. First, in Khawaja et al.’s study
(Khawaja, et al 2004) venlafaxine was administered i.p. while we
used s.c. route (osmotic minipumps). Second, in the mentioned
study BrdU was injected along four consecutive days following
the last administration of venlafaxine, while in the present
work the cell proliferation analysis was carried out 24 h
after removing the osmotic minipump. These results suggest
that, depending on several factors (type of drug, route of
administration, duration) the activation of the serotonergic
Angel Pazos 24

system might be not enough to elicit by itself a proliferative
response, thus requiring the concomitant inhibition of NE
transport. Furthermore, they are in good agreement with the
reported ability of venlafaxine to prevent the decrease in
hippocampal cell proliferation induced by chronic stress (Xu,
et al 2006).
These results strongly support that AD drugs are potential
enhancers of hippocampal cell proliferation and neurogenesis.
In order to address the signaling pathways involved, we
studied the effects of venlafaxine on the amount and cellular
distribution of main effector proteins of several signaling
pathways involved in cell proliferation. β-Catenin expression
was increased in both TCL and purified nuclear fraction in
venlafaxine-treated animals in a dose-dependent manner,
suggesting an increase in translocation of β-catenin to cell
nucleus. This finding correlates well with that reported in a
previous study using ECS therapy (Madsen, et al 2003). The
increase in β-catenin immunoreactivity observed in the
proliferating area SGZ after chronic venlafaxine treatment,
together with the fact that BrdU+ cells were embedded in
immunopositive β-catenin clusters further supports the
Angel Pazos 25

relevance of this protein in hippocampal cell proliferation.
Interestingly, GSK-3 inhibitors, which allow stabilization of
-catenin, have been reported to exert antidepressant-like
effects. (Kaidanovich-Beilin, et al 2004; Gould, et al 2006). The
role of β-catenin in affecting gene expression would be
mediated by the interaction with TCF/LEF DNA-binding proteins
(Logan and Nusse, 2004; van de Wetering, et al 1997). Target
genes would include, in addition to cell cycle regulator
genes, as myc and cyclinD1 (He, et al 1998; Shtutman, et al
1999), Wnt signaling genes, thus leading to a fine feedback
regulation of this pathway (Logan and Nusse, 2004). Taking into
account that the adult hippocampal stem/progenitor cells
(AHPs) express receptors for Wnt proteins (Madsen, et al 2003),
it could be suggested that chronic venlafaxine would result in
a double -catenin-dependent activation of AHPs: a direct
activation of neural precursors in SGZ to re-enter in cell
cycle, and an indirect regulation of surrounding
differentiated cells, such as astrocytes, in order to secret
Wnt proteins, that would act on hippocampal progenitor cells,
leading to a further increase in neurogenesis (Lie, et al 2005).
Activated AHP cells are able to divide through mechanisms that
Angel Pazos 26

involve the expression of CyclinD genes, producing daughter
cells that can differentiate to neurons (neurogenesis) or
astrocytes (gliogenesis) (Fig. 8). In this regard, it has been
recently shown that chronic treatment with antidepressants
induces an increase of symmetric divisions of an early
progenitor cell class in the dentate gyrus (Encinas, et al 2006).
However, while the increase in nuclear β-catenin was observed
in animals receiving either doses of venlafaxine (10 and 40
mg/kg/day), we could not detect any clear increase in cell
proliferation after treatment with the lower dose of the drug.
This suggests that a high level of induction of the Wnt
pathway is required to affect cell proliferation.
Alternatively, these results could indicate the involvement of
other signaling pathways in this response. No information
about hippocampal β-catenin expression in tissues from
depressed patients is available, although no changes have been
found in cortical samples (Beasley, et al 2002).
We tried to detect the presence of nuclear β-catenin in SGZ
and GCL cells, as well as in BrdU+ nuclei from SGZ, by
immunofluorescence microscopy techniques. A clear perinuclear
β-catenin staining was detected surrounding BrdU+ nuclei, but
Angel Pazos 27

no nuclear staining was demonstrated as previously reported in
the literature (Madsen, et al 2003; Lucas, et al 1999; Lucas, et al
2001) where the use of immunoelectron microscopy has been
suggested to overcome this limitation (Lucas, et al 2001): our
immunoelectron microscopy studies confirmed the presence of β-
catenin in the nucleus of SGZ cells in all experimental
groups. The pattern of distribution found, in euchromatin and
no nucleolar presence, confirms the role of this protein in
the regulation of target genes expression. In venlafaxine
treated animals, nuclear β-catenin density was higher than in
the saline group, and gold particles usually appeared forming
patches of several units. This is, to our knowledge, the first
report of an increased level of nuclear β-catenin following AD
treatment.
Taking into account that β-catenin is also a component of the
cadherin/catenin complex, stabilizing the actin cytoskeleton
and mediating cell adhesion (Cadigan and Nusse, 1997), the
increased levels of β-catenin detected after venlafaxine
chronic treatment in the membrane-associated fraction could
suggest a positive effect in stabilizing synapses and
increasing dendritic arborization. This agrees with recent
Angel Pazos 28

reports demonstrating, first, that the intracellular level of
cadherin/catenin complex is a limiting factor in dendritic
morphogenesis (Yu and Malenka, 2003) and, second, that AD
treatment increases the formation of dendritic spines in rat
hippocampus (Hajszan, et al 2005 ).
The AKT/PKB signaling is strongly involved in the regulation
of several cell responses, such as cell proliferation, growth
and cell survival (Brazil and Hemmings, 2001). Interestingly,
it has been recently reported the existence of a significantly
lower activity of this protein in occipital cortex of
depressed suicide subjects (Hsiung, et al 2003). Our results
regarding AKT levels after venlafaxine treatment (increase in
cytosolic and decrease in nuclear fractions) suggest that this
protein is being accumulated in the cytosol, in order to be
subsequently activated in the cell membrane. Since GSK-3 is
directly inhibited by AKT (Cross, et al 1995), this recruitment
of cytosolic AKT would lead to a strong inhibition of this
kinase, resulting in further stabilization of β-catenin.
Our results show that CREB levels were not affected by chronic
venlafaxine treatment. A tendency to the decrease in cortical
CREB levels from depressed patients has been reported
Angel Pazos 29

(Dowlatshahi, et al 1998; Yamada, et al 2003), although no study
has been performed in hippocampus. In line with that, several
reports indicate that AD treatment could increase CREB
function, although the level of response appears to depend on
several factors (Tiraboschi, et al 2004; Frechilla, et al 1998;
Thome, et al 2000). Fluoxetine has been reported to be more
effective in increasing the expression and phosphorylation of
hippocampal CREB, when compared to ADs affecting NE uptake
(reboxetine, desipramine) (Tiraboschi, et al 2004; Frechilla, et
al 1998). Furthermore, a reduction in the ability to
phosphorylate CREB in human neuroblasts has been described
following reboxetine and desipramine administration, but not
after venlafaxine treatment (Manier, et al 2002). These results
illustrate the variability of the modifications induced by ADs
on CREB expression, which strongly depend on the particular
drug used.
Few data are available about the effect of ADs on the
components of MAPK pathway (Tiraboschi, et al 2004; Fumagalli ,
et al 2005). Our study agrees with most previously published
results, showing no consistent change in the levels of ERK1/2
in hippocampus after venlafaxine treatment, supporting that
Angel Pazos 30

their expression is not affected by ADs. Regarding pERK1 and
pERK2, data in the literature are contradictory: while no
changes have been reported after treatment with imipramine,
decreases (nuclear pERK1/2), increases (nuclear pERK1) and
lack of changes have been observed as a consequence of
fluoxetine treatment (Tiraboschi, et al 2004; Fumagalli , et al
2005). Although caution is needed when analyzing these results,
our data support the activation of the MAPK pathway following
chronic administration with venlafaxine, and are in agreement
with results from postmortem studies showing a decrease in
brain pERK1/2 levels in depressed patients (Hsiung, et al 2003).
In conclusion, our results indicate that chronic treatment
with venlafaxine induces nuclear translocation of β-catenin,
probably trough activation of Wnt and AKT pathways. This
effect could be mediating the increased hippocampal cell
proliferation observed, thus reinforcing the role of
neurotrophic mechanisms in the therapeutic activity of ADs.
Angel Pazos 31

Disclosure/conflicts of interes
Dr. Pazos reports having received research funding and lecture
fees from Faes Farma S.A.. Drs. Mostany and Valdizán report no
biomedical financial interests or conflicts of interest.
Conferencias??
Angel Pazos 32

Acknowledgements:
We are grateful to FAES FARMA, S.A. (Mr. Antonio Toledo) for
the extraction of venlafaxine. We wish to thank Ms. María
Josefa Castillo, Ms. Paula Díez del Valle, Ms. Rebeca
Madureira, Ms Lourdes Lanza, Mr. Germán Pérez and Ms. Isabel
Ruiz for their technical assistance. We are deeply in debt
with Prof. Miguel Lafarga and Prof. María Teresa Berciano, of
the Departmento de Anatomía y Biología Celular of Universidad
de Cantabria, by their invaluable help in scientific and
technical aspects of confocal and electron microscopy. R.M.
received a postdoctoral fellowship from Fundación Pública
“Marqués de Valdecilla”-IFIMAV. This work has been supported
by CICYT (SAF04-00941), IFIMAV (API 05/23), Alicia Koplowitz
Foundation, the Spanish Ministry of Health, Instituto de Salud
Carlos III, RETICS RD06/0011(REM-TAP Network and FAES FARMA,
S.A.-Universidad de Cantabria contract.
Angel Pazos 33

References
Altar CA, Laeng P, Jurata LW, Brockman JA, Lemire A, Bullard J
et al. (2004). Electroconvulsive seizures regulate gene
expression of distinct neurotrophic signaling pathways. J
Neurosci 24: 2667-2677.
Altar CA, Whitehead RE, Chen R, Wortwein G, Madsen TM, (2003).
Effects of electroconvulsive seizures and antidepressant drugs
on brain-derived neurotrophic factor protein in rat brain. Biol
Psychiatry 54: 703-709.
Beasley C, Cotter D, Everall I, (2002). An investigation of
the Wnt-signalling pathway in the prefrontal cortex in
schizophrenia, bipolar disorder and major depressive disorder.
Schizophr Res 58: 63-67.
Bëíque JC, de Montigny C, Blier P, Debonnel G, (1998).
Blockade of 5-hydroxytryptamine and noradrenaline uptake by
venlafaxine: a comparative study with paroxetine and
desipramine. Br J Pharmacol 125: 526-532.
Brazil DP, Hemmings BA, (2001). Ten years of protein kinase B
signalling: a hard Akt to follow. Trends Biochem Sci 26: 657-664.
Angel Pazos 34

Cadigan KM, Nusse R, (1997). Wnt signaling: a common theme in
animal development. Genes Dev 11: 3286-3305.
Cross DA, Alessi DR, Cohen P, Andjelkovich M, Hemmings BA,
(1995). Inhibition of glycogen synthase kinase-3 by insulin
mediated by protein kinase B. Nature 378: 785-789.
Czéh B, Michaelis T, Watanabe T, Frahm J, de Biurrun G, van
Kampen M et al. (2001) : Stress-induced changes in cerebral
metabolites, hippocampal volume, and cell proliferation are
prevented by antidepressant treatment with tianeptine. Proc Natl
Acad Sci USA 98: 12796-12801.
Dowlatshahi D, MacQueen GM, Wang JF, Young LT, (1998).
Increased temporal cortex CREB concentrations and
antidepressant treatment in major depression. Lancet 352: 1754-
1755.
Dranovsky A, Hen R, (2006). Hippocampal neurogenesis:
regul.ation by stress and antidepressants. Biol.Psychiatry 59:
1136-1143.
D'Sa C, Duman RS, (2002). Antidepressants and
neuroplasticity. Bipolar Disord 4: 183-194.
Angel Pazos 35

Encinas JM, Vaahtokari A, Enikopolov G, (2006). Fluoxetine
targets early progenitor cells in the adult brain. Proc Natl Acad
Sci USA 103: 8233-8238.
Frechilla D, Otano A, Del Rio J (1998). Effect of chronic
antidepressant treatment on transcription factor binding
activity in rat hippocampus and frontal cortex. Prog
Neuropsychopharmacol Biol Psychiatry 22: 787-802.
Fumagalli F, Molteni R, Calabrese F, Frasca A, Racagni G, Riva
MA, (2005). Chronic fluoxetine administration inhibits
extracellular signal-regulated kinase 1/2 phosphorylation in
rat brain. J Neurochem 93: 1551-1560.
Gould TD, Picchini AM, Einat H, Manji HK, (2006). Targeting
glycogen synthase kinase-3 in the CNS: implications for the
development of new treatments for mood disorders. Curr Drug
Targets 7: 1399-1409.
Hajszan T, MacLusky NJ, Leranth C, (2005). Short-term
treatment with the antidepressant fluoxetine triggers
pyramidal dendritic spine synapse formation in rat
hippocampus. Eur J Neurosci 21: 1299-1303.
Angel Pazos 36

He TC, Sparks AB, Rago C, Hermeking H, Zawel L, da Costa LT et
al. (1998). Identification of c-MYC as a target of the APC
pathway. Science 281: 1509-1512.
Hsiung SC, Adlersberg M, Arango V, Mann JJ, Tamir H, Liu KP,
(2003). Attenuated 5-HT1A receptor signaling in brains of
suicide victims: involvement of adenylyl cyclase,
phosphatidylinositol 3-kinase, Akt and mitogen-activated
protein kinase. J Neurochem 87: 182-194.
Kaidanovich-Beilin O, Milman A, Weizman A, Pick CG, Eldar-
Finkelman H, (2004). Rapid antidepressive-like activity of
specific glycogen synthase kinase-3 inhibitor and its effect
on β-catenin in mouse hippocampus. Biol Psychiatry 55: 781-784.
Khawaja X, Xu J, Liang JJ, Barrett JE, (2004). Proteomic
analysis of protein changes developing in rat hippocampus
after chronic antidepressant treatment: Implications for
depressive disorders and future therapies. J Neurosci Res 75: 451-
460.
Lie DC, Colamarino SA, Song HJ, Desire L, Mira H, Consiglio A
et al.(2005). Wnt signalling regulates adult hippocampal
neurogenesis. Nature 437: 1370-1375.
Angel Pazos 37

Logan CY, Nusse R, (2004). The Wnt signaling pathway in
development and disease. Annu Rev Cell Dev Biol 20: 781-810.
Lowry OH, Rosebrough NJ, Farr AL, Randall RJ, (1951). Protein
measurement with the Folin phenol reagent. J Biol Chem 193: 265-
275.
Lucas JJ, Hernandez F, Avila J, (1999). Nuclear localization
of β-catenin in adult mouse thalamus correlates with low
levels of GSK-3β. Neuroreport 10: 2699-2703.
Lucas JJ, Hernandez F, Gomez-Ramos P, Moran MA, Hen R, Avila
J, (2001). Decreased nuclear β-catenin, tau
hyperphosphorylation and neurodegeneration in GSK-3β
conditional transgenic mice. EMBO J 20: 27-39.
Madsen TM, Newton SS, Eaton ME, Russell DS, Duman RS, (2003).
Chronic electroconvulsive seizure up-regulates β-catenin
expression in rat hippocampus: role in adult neurogenesis. Biol
Psychiatry 54: 1006-1014.
Madsen TM, Treschow A, Bengzon J, Bolwig TG, Lindvall O,
Tingstrom A, (2000). Increased neurogenesis in a model of
electroconvulsive therapy. Biol Psychiatry 47: 1043-1049.
Angel Pazos 38

Malberg JE, Duman RS, (2003). Cell proliferation in adult
hippocampus is decreased by inescapable stress: reversal by
fluoxetine treatment. Neuropsychopharmacology 28: 1562-1571.
Malberg JE, Eisch AJ, Nestler EJ, Duman RS, (2000). Chronic
antidepressant treatment increases neurogenesis in adult rat
hippocampus. J Neurosci 20: 9104-9110.
Manier DH, Shelton RC, Sulser F, (2002). Noradrenergic
antidepressants: does chronic treatment increase or decrease
nuclear CREB-P? J Neural Transm 109: 91-99.
Mehtonen OP, Sogaard J, Roponen P, Behnke K, (2000).
Randomized, double-blind comparison of venlafaxine and
sertraline in outpatients with major depressive disorder.
Venlafaxine 631 Study Group. J Clin Psychiatry 61: 95-100.
Nibuya M, Nestler EJ, Duman RS, (1996). Chronic
antidepressant administration increases the expression of cAMP
response element binding protein (CREB) in rat hippocampus. J
Neurosci 16: 2365-2372.
Paxinos G, Watson C (1982): The Rat Brain in Stereotaxic
Coordinates. Academic Press: Sydney.
Angel Pazos 39

Perera TD, Coplan JD, Lisanby SH, Lipira CM, Arif M, Carpio C
et al. (2007). Antidepressant-indiced neurogenesis in the
hippocampus of adult nonhuman primates. J. Neurosci. 27: 4894-4901.
Poirier MF, Boyer P, (1999). Venlafaxine and paroxetine in
treatment-resistant depression. Double-blind, randomised
comparison. Br J Psychiatry 175: 12-16.
Richelson E, (1991). Biological basis of depression and
therapeutic relevance. J Clin Psychiatry 52 Suppl: 4-10.
Russo-Neustadt AA, Alejandre H, Garcia C, Ivy AS, Chen MJ,
(2004). Hippocampal brain-derived neurotrophic factor
expression following treatment with reboxetine, citalopram,
and physical exercise. Neuropsychopharmacology 29: 2189-2199.
Sairanen M, Lucas G, Ernfors P, Castren M, Castren E, (2005).
Brain-derived neurotrophic factor and antidepressant drugs
have different but coordinated effects on neuronal turnover,
proliferation, and survival in the adult dentate gyrus. J
Neurosci 25: 1089-1094.
Santarelli L, Saxe M, Gross C, Surget A, Battaglia F, Dulawa S
et al. (2003). Requirement of hippocampal neurogenesis for the
behavioral effects of antidepressants. Science 301: 805-809.
Angel Pazos 40

Sheline YI, Wang PW, Gado MH, Csernansky JG, Vannier MW,
(1996). Hippocampal atrophy in recurrent major depression.
Proc Natl Acad Sci USA 93: 3908-3913.
Shtutman M, Zhurinsky J, Simcha I, Albanese C, D'Amico M,
Pestell R et al. (1999). The cyclin D1 gene is a target of the
β-catenin/LEF-1 pathway. Proc Natl Acad Sci USA 96: 5522-5527.
Stahl SM, Entsuah R, Rudolph RL, (2002). Comparative efficacy
between venlafaxine and SSRIs: a pooled analysis of patients
with depression. Biol Psychiatry 52: 1166-1174.
Thome J, Sakai N, Shin K, Steffen C, Zhang YJ, Impey S et al.
(2000). cAMP response element-mediated gene transcription is
upregulated by chronic antidepressant treatment. J Neurosci 20:
4030-4036.
Tiraboschi E, Tardito D, Kasahara J, Moraschi S, Pruneri P,
Gennarelli M et al. (2004). Selective phosphorylation of nuclear
CREB by fluoxetine is linked to activation of CaM kinase IV
and MAP kinase cascades. Neuropsychopharmacology 29: 1831-1840.
Tzanakaki M, Guazzelli M, Nimatoudis I, Zissis NP, Smeraldi E,
Rizzo F, (2000). Increased remission rates with venlafaxine
Angel Pazos 41

compared with fluoxetine in hospitalized patients with major
depression and melancholia. Int Clin Psychopharmacol 15: 29-34.
van de Wetering M, Cavallo R, Dooijes D, van Beest M, van Es
J, Loureiro J et al.(1997). Armadillo coactivates transcription
driven by the product of the Drosophila segment polarity gene
dTCF. Cell 88: 789-799.
Vermetten E, Vythilingam M, Southwick SM, Charney DS, Bremner
JD, (2003). Long-term treatment with paroxetine increases
verbal declarative memory and hippocampal volume in
posttraumatic stress disorder. Biol Psychiatry 54: 693-702.
Vythilingam M, Vermetten E, Anderson GM, Luckenbaugh D,
Anderson ER, Snow J et al. (2004). Hippocampal volume, memory,
and cortisol status in major depressive disorder: effects of
treatment. Biol Psychiatry 56: 101-112.
Xu H, Chen Z, He J, Haimanot S, Li X, Dyck L, Li XM, (2006).
Synergetic effects of quetiapine and venlafaxine in preventing
the chronic restraint stress-induced decrease in cell
proliferation and BDNF expression in frat hippocampus.
Hippocampus 16: 551-559.
Angel Pazos 42

Yamada S, Yamamoto M, Ozawa H, Riederer P, Saito T, (2003).
Reduced phosphorylation of cyclic AMP-responsive element
binding protein in the postmortem orbitofrontal cortex of
patients with major depressive disorder. J Neural Transm 110:
671-680.
Yu X, Malenka RC, (2003). β-Catenin is critical for dendritic
morphogenesis. Nat Neurosci 6: 1169-1177.
Angel Pazos 43
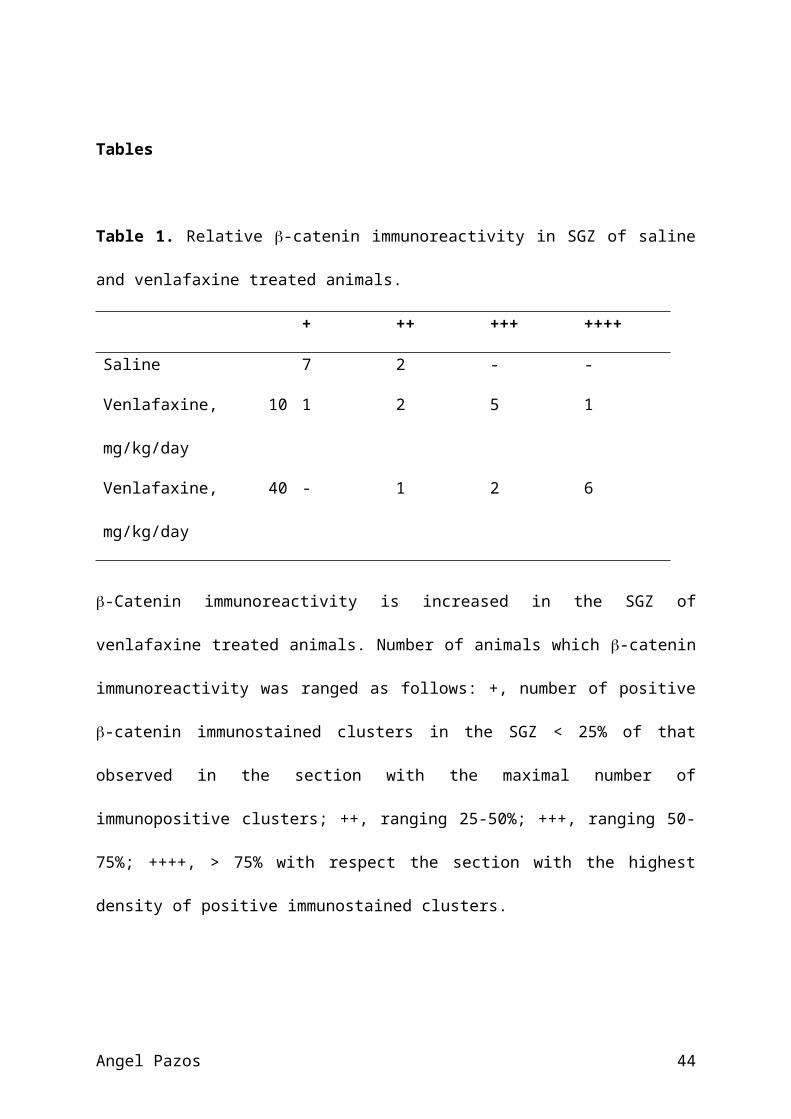
Tables
Table 1. Relative -catenin immunoreactivity in SGZ of saline
and venlafaxine treated animals.
+ ++ +++ ++++
Saline 7 2 - -
Venlafaxine, 10
mg/kg/day
1 2 5 1
Venlafaxine, 40
mg/kg/day
- 1 2 6
-Catenin immunoreactivity is increased in the SGZ of
venlafaxine treated animals. Number of animals which -catenin
immunoreactivity was ranged as follows: +, number of positive
-catenin immunostained clusters in the SGZ < 25% of that
observed in the section with the maximal number of
immunopositive clusters; ++, ranging 25-50%; +++, ranging 50-
75%; ++++, > 75% with respect the section with the highest
density of positive immunostained clusters.
Angel Pazos 44

Titles and legends to figure
Figure 1. Chronic venlafaxine treatment increases the number
of BrdU+ cells in the SGZ of adult rat hippocampus when
administered at a high dose. Representative microphotographs
of BrdU immunohistochemistry from saline (a), 10 mg/kg/day
venlafaxine (b), and 40 mg/kg/day venlafaxine (c) chronically
treated animals. Cresyl violet counter-staining. (d) The
number of BrdU+ cells in the subgranular zone (SGZ) is
significantly increased after chronic treatment with 40
mg/kg/day venlafaxine. The results are the mean ± SEM number
of BrdU+ cells in SGZ. **p < 0.01; *p < 0.05. GCL, granule cell
layer; H, Hilus. Bar = 100 µm
Figure 2. Chronic venlafaxine treatment affects expression of
main effector proteins of Wnt and AKT/PKB signaling pathways.
Graphs represent densitometry levels of β-catenin and AKT
proteins in AD treated animals as a percentage of these
proteins in saline group animals (Mean ± SEM). (a) Venlafaxine
treatment increases β-catenin immunoreactivity in total cell
lysate (TCL), membrane-associated (M) and nuclear (N)
Angel Pazos 45

subcellular fractions of rat hippocampus in Western blot
studies. (b) Representative Western blot analyses of β-catenin
and AKT levels in total cell lysates (TCL) and subcellular
fractions, Citosol (C), membrane-associated (M) and nuclear
(N), from saline, 10 and 40 mg/kg/day venlafaxine chronically
treated animals. Densitometric measurement levels were
normalized to actin protein amounts. Purity of subcellular
fractions used in Western blot assays was assessed by analysis
of GAPDH and histone proteins expression. All samples showed
for comparison were run in the same gel. (c) Venlafaxine
treatment differentially affects AKT protein levels depending
on the dose and the subcellular fraction analyzed. *p < 0.05;
**p < 0.01; ***p < 0.001.
Figure 3. Chronic venlafaxine treatment does not affect
nuclear expression of CREB nor p-CREB. Graphs represent
densitometry levels of CREB as well as p-CREB proteins in AD
treated animals as a percentage of these proteins in saline
group animals, and the p-CREB/CREB levels ratio (all data are
expressed as mean ± SEM). Representative Western blot analyses
of CREB and p-CREB levels in nuclear subcellular fractions
Angel Pazos 46

from saline, 10 and 40 mg/kg/day venlafaxine chronically
treated animals.
Figure 4. Chronic venlafaxine treatment affects the expression
of p-ERK1 and p-ERK2 without modifying the expression of ERK1
or ERK2. Graphs represent densitometry levels of ERK1/2 and p-
ERK1/2 proteins in AD treated animals as a percentage of these
proteins in saline group animals, and the p-ERK1/ERK1 and p-
ERK2/ERK2 levels ratios (all data are expressed as mean ±
SEM). Representative Western blot analyses of ERK1 and ERK2,
and p-ERK1 and p-ERK2 in tissue cell lysates from saline, 10
and 40 mg/kg/day venlafaxine chronically treated animals. *p <
0.05; **p < 0.01; ***p < 0.001.
Figure 5. Representative microphotographs of β-catenin
immunohistochemistry from saline (a), 10 mg/kg/day venlafaxine
(b), and 40 mg/kg/day venlafaxine (c) chronically treated
animals showing increased immunoreactivity after chronic
treatment with 10 and 40 mg/kg/day venlafaxine. Arrows:
positive staining; Bar = 200 µm.
Angel Pazos 47

Figure 6. Left panel shows representative confocal
microphotographs of SGZ from animals from saline group (a) and
40 mg/kg/day venlafaxine group (b). No nuclear presence of β-
catenin could be observed in any animal from these
experimental groups, although an increased β-catenin
immunolabeling was detected in the treated animals. Right
panel shows representative microphotographs of BrdU/β-catenin
double labeling from a 40 mg/kg/day venlafaxine chronically
treated animal. No overlapping could be detected in any sample
using immunohistochemistry techniques (arrow). CGL: granule
cell layer; H: hilus; SGZ: subgranular zone; Bar = 20 µm.
Figure 7. Electron microscopy photomicrographs showing nuclei
from SGZ cells from saline (a), 10 mg/kg/day venlafaxine (b),
and 40 mg/kg/day venlafaxine (c) chronically treated animals.
β-Catenin positive labeling (arrows) is increased in
venlafaxine treated animals. Nu: nucleolus. Bar = 0.2 µm.
Figure 8. Diagram depicting the suggested effects of AD on
Wnt pathway in the SGZ. Adult hippocampal stem/progenitor
cells (AHPs), neurons and glial cells are present in SGZ and
Angel Pazos 48

adjacent GCL. AD treatment increases the extracellular levels
of 5-HT and NE, which, in neurons and glial cells, through
serotonin and adrenergic receptors, elicit a number of
reactions that lead to AKT activation. Inhibition of GSK-3 by
AKT increases β-catenin cytoplasmatic levels and promotes the
traslocation to the nucleus of this protein. In these very
differentiated cells, no expression of cell cycle induction
proteins will take place. However they express Wnt pathway
proteins, among them Wnt receptors agonists, which are
secreted to the extracellular space. There, secreted Wnt
agonists act on Frizzled receptors (FZr) the surrounding
cells, neurons and glia, where they enhance the activation of
the Wnt pathway, and AHPs, where they elicit the expression of
cell cycle related proteins, as CyclinD, promoting cell
division.
Angel Pazos 49

Angel Pazos 50




















