
MUC1 regulates cyclin D1 gene expression through p120 catenin and β-catenin
Upload
independentCategory
view
1download
0
Cell, Vol. 111, 241–250, October 18, 2002, Copyright 2002 by Cell Press
The �-Catenin/TCF-4 Complex Imposes a CryptProgenitor Phenotype on Colorectal Cancer Cells
renewal occurs in these crypts through a coordinatedseries of events involving proliferation, differentiation,and migration toward the intestinal lumen. Pluripotent
Marc van de Wetering,1,7,8 Elena Sancho,1,7,8
Cornelis Verweij,2 Wim de Lau,1,8 Irma Oving,1,8
Adam Hurlstone,1,8 Karin van der Horn,1,8
Eduard Batlle,1,8 Damien Coudreuse,1,8 stem cells at the crypt bottom generate progenitors thatoccupy the lower third of the crypt. Cells in this amplifi-Anna-Pavlina Haramis,1,8 Menno Tjon-Pon-Fong,1,8
Petra Moerer,1,8 Maaike van den Born,1,8 cation compartment divide approximately every 12 hr.Gwen Soete,1,8 Steven Pals,3 Martin Eilers,4 In the midcrypt region, the cells differentiate into one ofRene Medema,5 and Hans Clevers1,6,8 the functional cell types of the colon. At the epithelial1Department of Immunology and Center for surface, cells undergo apoptosis and/or extrusion intoBiomedical Genetics the lumen. The entire process takes approximately 3–5University Medical Center days (Potten and Loeffler, 1990).3584 CX Utrecht The transition of an intestinal epithelial cell into a fullyThe Netherlands transformed, metastatic cancer cell requires mutations2 Department of Molecular Cellular Biology in multiple proto-oncogenes and tumor suppressorVU Medical Center genes (Kinzler and Vogelstein, 1996). The APC gene,1081 BT Amsterdam originally cloned from the rare genetic disorder familialThe Netherlands adenomatous polyposis, is mutated in most sporadic3 Department of Pathology CRCs. The APC protein resides in the destruction com-Academic Medical Center plex, together with GSK3�, axin/conductin, and �-cat-1105 AZ Amsterdam enin. In this complex, phosphorylation by GSK3� targetsThe Netherlands �-catenin for ubiquitination and destruction by the pro-4 Institute for Molecular Biology and Tumor teasome. Wnt signaling inhibits GSK3� activity. Then,
Research �-catenin accumulates in the nucleus, where it binds35033 Marburg members of the TCF family and converts these WNTGermany effectors from transcriptional repressors into activators5 Department of Molecular Biology (reviewed in Bienz and Clevers, 2000). In cancer, truncat-Netherlands Cancer Institute ing mutations in APC and axin/conductin, as well as1066 CX Amsterdam mutations in the GSK3�-target residues in �-catenin, allThe Netherlands lead to the formation of constitutive nuclear �-catenin/
TCF complexes (Korinek et al., 1997; Morin et al., 1997;Rubinfeld et al., 1997; Satoh et al., 2000; Liu et al., 2000).Activating mutations of the WNT pathway are the only
Summary known genetic alterations in early premalignant lesionsin the intestine, such as aberrant crypt foci and small
The transactivation of TCF target genes induced by polyps (Powell et al., 1992). Thus, these mutations mayWnt pathway mutations constitutes the primary trans- initiate the transformation of colorectal epithelial cells.forming event in colorectal cancer (CRC). We show In the intestinal epithelium, Tcf-4 is the most promi-that disruption of �-catenin/TCF-4 activity in CRC cells nently expressed TCF family member (Korinek et al.,induces a rapid G1 arrest and blocks a genetic program 1997). Gene disruption has revealed that Tcf-4 is re-that is physiologically active in the proliferative compart- quired to establish the proliferative progenitors of thement of colon crypts. Coincidently, an intestinal differen- prospective crypts in the embryonic small intestine (Kor-tiation program is induced. The TCF-4 target gene inek et al., 1998). A second TCF family member, TCF-1,c-MYC plays a central role in this switch by direct repres- is expressed in the intestinal epithelium predominantlysion of the p21CIP1/WAF1 promoter. Following disruption of as a dominant-negative isoform, which lacks the N-ter-�-catenin/TCF-4 activity, the decreased expression of minal �-catenin interaction domain. Genetic evidencec-MYC releases p21CIP1/WAF1 transcription, which in turn suggests that Tcf-1 acts as an antagonist of Tcf-4 inmediates G1 arrest and differentiation. Thus, the the formation of polyps in an APC�/� background (Roose�-catenin/TCF-4 complex constitutes the master et al., 1999).switch that controls proliferation versus differentiation To understand the contribution of constitutivein healthy and malignant intestinal epithelial cells.
�-catenin/TCF-4 activity to the colorectal transforma-tion process, we have undertaken a large-scale analysis
Introductionof the downstream genetic program activated by �-cat-enin/TCF in CRC cells. We have subsequently analyzed
The colorectal mucosa contains large numbers of inva-the expression and activities of individual �-catenin/
ginations termed the crypts of Lieberkuhn. EpithelialTCF-4-regulated genes in a physiological context. Ourresults indicate that the �-catenin/TCF-4 complex,6 Correspondence: [email protected] its control over c-MYC and p21CIP1/WAF1 activity,7 These authors contributed equally to this work.inhibits differentiation and imposes a crypt progenitor-8 Present address: Hubrecht Laboratory, Center for Biomedical Ge-
netics, 3584 CT Utrecht, The Netherlands. like phenotype on CRC cells.
Cell242
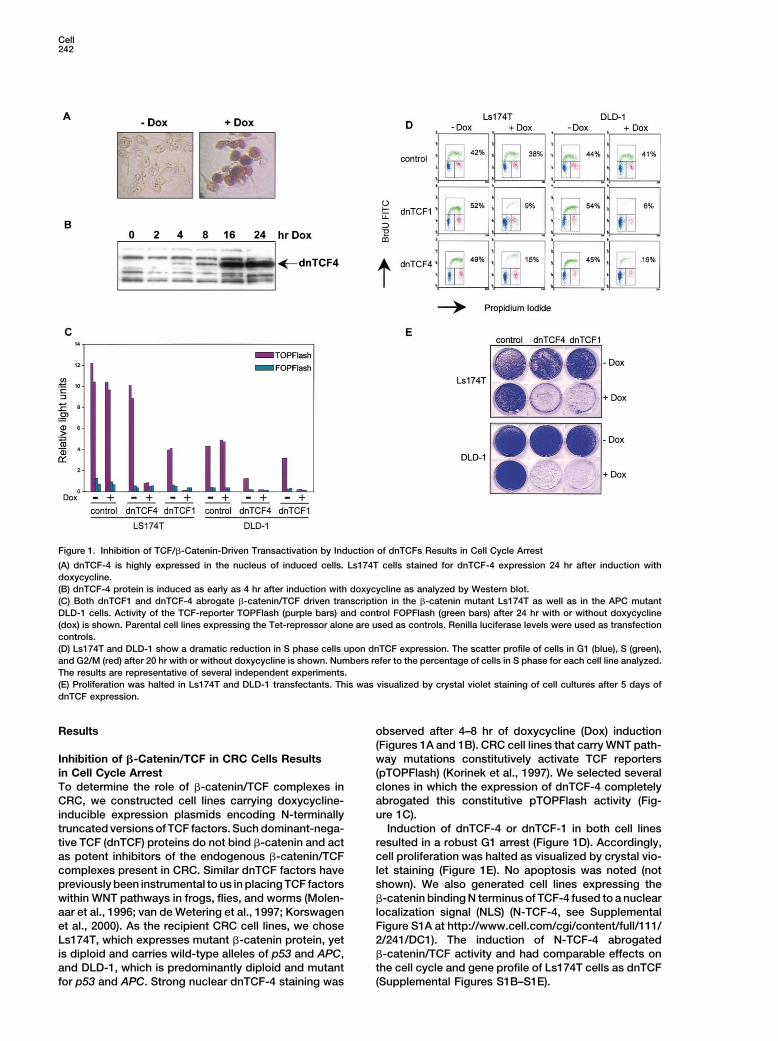
Figure 1. Inhibition of TCF/�-Catenin-Driven Transactivation by Induction of dnTCFs Results in Cell Cycle Arrest
(A) dnTCF-4 is highly expressed in the nucleus of induced cells. Ls174T cells stained for dnTCF-4 expression 24 hr after induction withdoxycycline.(B) dnTCF-4 protein is induced as early as 4 hr after induction with doxycycline as analyzed by Western blot.(C) Both dnTCF1 and dnTCF-4 abrogate �-catenin/TCF driven transcription in the �-catenin mutant Ls174T as well as in the APC mutantDLD-1 cells. Activity of the TCF-reporter TOPFlash (purple bars) and control FOPFlash (green bars) after 24 hr with or without doxycycline(dox) is shown. Parental cell lines expressing the Tet-repressor alone are used as controls. Renilla luciferase levels were used as transfectioncontrols.(D) Ls174T and DLD-1 show a dramatic reduction in S phase cells upon dnTCF expression. The scatter profile of cells in G1 (blue), S (green),and G2/M (red) after 20 hr with or without doxycycline is shown. Numbers refer to the percentage of cells in S phase for each cell line analyzed.The results are representative of several independent experiments.(E) Proliferation was halted in Ls174T and DLD-1 transfectants. This was visualized by crystal violet staining of cell cultures after 5 days ofdnTCF expression.
Results observed after 4–8 hr of doxycycline (Dox) induction(Figures 1A and 1B). CRC cell lines that carry WNT path-way mutations constitutively activate TCF reportersInhibition of �-Catenin/TCF in CRC Cells Results
in Cell Cycle Arrest (pTOPFlash) (Korinek et al., 1997). We selected severalclones in which the expression of dnTCF-4 completelyTo determine the role of �-catenin/TCF complexes in
CRC, we constructed cell lines carrying doxycycline- abrogated this constitutive pTOPFlash activity (Fig-ure 1C).inducible expression plasmids encoding N-terminally
truncated versions of TCF factors. Such dominant-nega- Induction of dnTCF-4 or dnTCF-1 in both cell linesresulted in a robust G1 arrest (Figure 1D). Accordingly,tive TCF (dnTCF) proteins do not bind �-catenin and act
as potent inhibitors of the endogenous �-catenin/TCF cell proliferation was halted as visualized by crystal vio-let staining (Figure 1E). No apoptosis was noted (notcomplexes present in CRC. Similar dnTCF factors have
previously been instrumental to us in placing TCF factors shown). We also generated cell lines expressing the�-catenin binding N terminus of TCF-4 fused to a nuclearwithin WNT pathways in frogs, flies, and worms (Molen-
aar et al., 1996; van de Wetering et al., 1997; Korswagen localization signal (NLS) (N-TCF-4, see SupplementalFigure S1A at http://www.cell.com/cgi/content/full/111/et al., 2000). As the recipient CRC cell lines, we chose
Ls174T, which expresses mutant �-catenin protein, yet 2/241/DC1). The induction of N-TCF-4 abrogated�-catenin/TCF activity and had comparable effects onis diploid and carries wild-type alleles of p53 and APC,
and DLD-1, which is predominantly diploid and mutant the cell cycle and gene profile of Ls174T cells as dnTCF(Supplemental Figures S1B–S1E).for p53 and APC. Strong nuclear dnTCF-4 staining was
�-Catenin/TCF Drives Intestinal Crypt Gene Program243
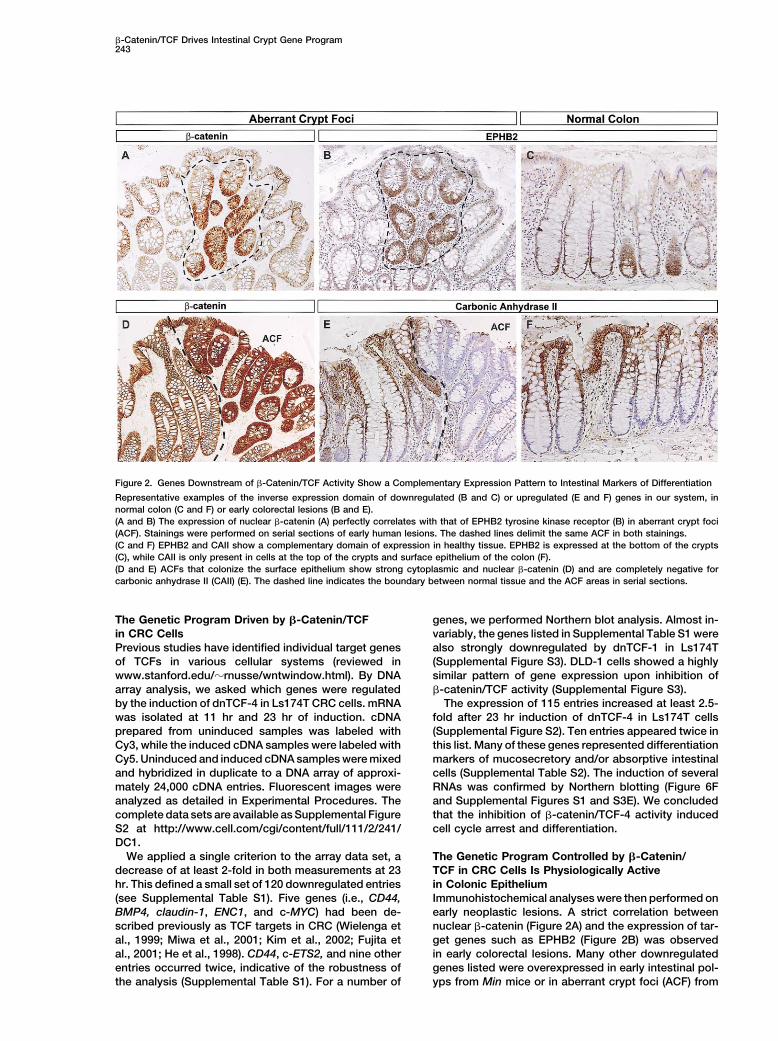
Figure 2. Genes Downstream of �-Catenin/TCF Activity Show a Complementary Expression Pattern to Intestinal Markers of Differentiation
Representative examples of the inverse expression domain of downregulated (B and C) or upregulated (E and F) genes in our system, innormal colon (C and F) or early colorectal lesions (B and E).(A and B) The expression of nuclear �-catenin (A) perfectly correlates with that of EPHB2 tyrosine kinase receptor (B) in aberrant crypt foci(ACF). Stainings were performed on serial sections of early human lesions. The dashed lines delimit the same ACF in both stainings.(C and F) EPHB2 and CAII show a complementary domain of expression in healthy tissue. EPHB2 is expressed at the bottom of the crypts(C), while CAII is only present in cells at the top of the crypts and surface epithelium of the colon (F).(D and E) ACFs that colonize the surface epithelium show strong cytoplasmic and nuclear �-catenin (D) and are completely negative forcarbonic anhydrase II (CAII) (E). The dashed line indicates the boundary between normal tissue and the ACF areas in serial sections.
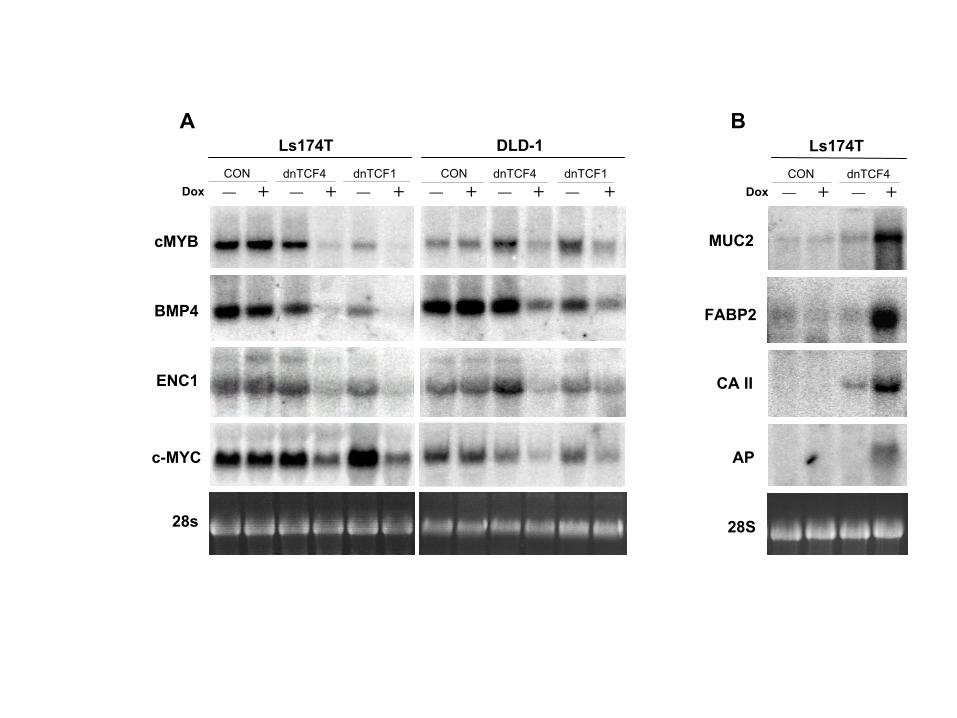
The Genetic Program Driven by �-Catenin/TCF genes, we performed Northern blot analysis. Almost in-variably, the genes listed in Supplemental Table S1 werein CRC Cells
Previous studies have identified individual target genes also strongly downregulated by dnTCF-1 in Ls174T(Supplemental Figure S3). DLD-1 cells showed a highlyof TCFs in various cellular systems (reviewed in
www.stanford.edu/�rnusse/wntwindow.html). By DNA similar pattern of gene expression upon inhibition of�-catenin/TCF activity (Supplemental Figure S3).array analysis, we asked which genes were regulated
by the induction of dnTCF-4 in Ls174T CRC cells. mRNA The expression of 115 entries increased at least 2.5-fold after 23 hr induction of dnTCF-4 in Ls174T cellswas isolated at 11 hr and 23 hr of induction. cDNA
prepared from uninduced samples was labeled with (Supplemental Figure S2). Ten entries appeared twice inthis list. Many of these genes represented differentiationCy3, while the induced cDNA samples were labeled with
Cy5. Uninduced and induced cDNA samples were mixed markers of mucosecretory and/or absorptive intestinalcells (Supplemental Table S2). The induction of severaland hybridized in duplicate to a DNA array of approxi-
mately 24,000 cDNA entries. Fluorescent images were RNAs was confirmed by Northern blotting (Figure 6Fand Supplemental Figures S1 and S3E). We concludedanalyzed as detailed in Experimental Procedures. The
complete data sets are available as Supplemental Figure that the inhibition of �-catenin/TCF-4 activity inducedcell cycle arrest and differentiation.S2 at http://www.cell.com/cgi/content/full/111/2/241/
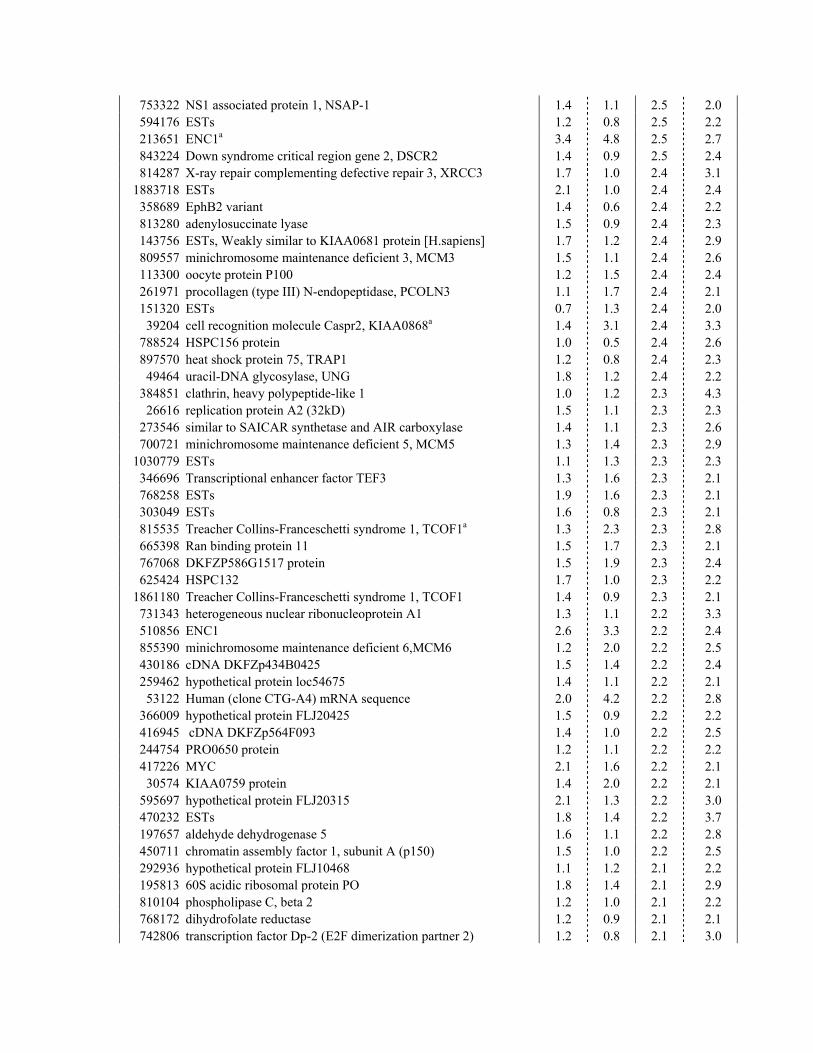
DC1.We applied a single criterion to the array data set, a The Genetic Program Controlled by �-Catenin/
TCF in CRC Cells Is Physiologically Activedecrease of at least 2-fold in both measurements at 23hr. This defined a small set of 120 downregulated entries in Colonic Epithelium
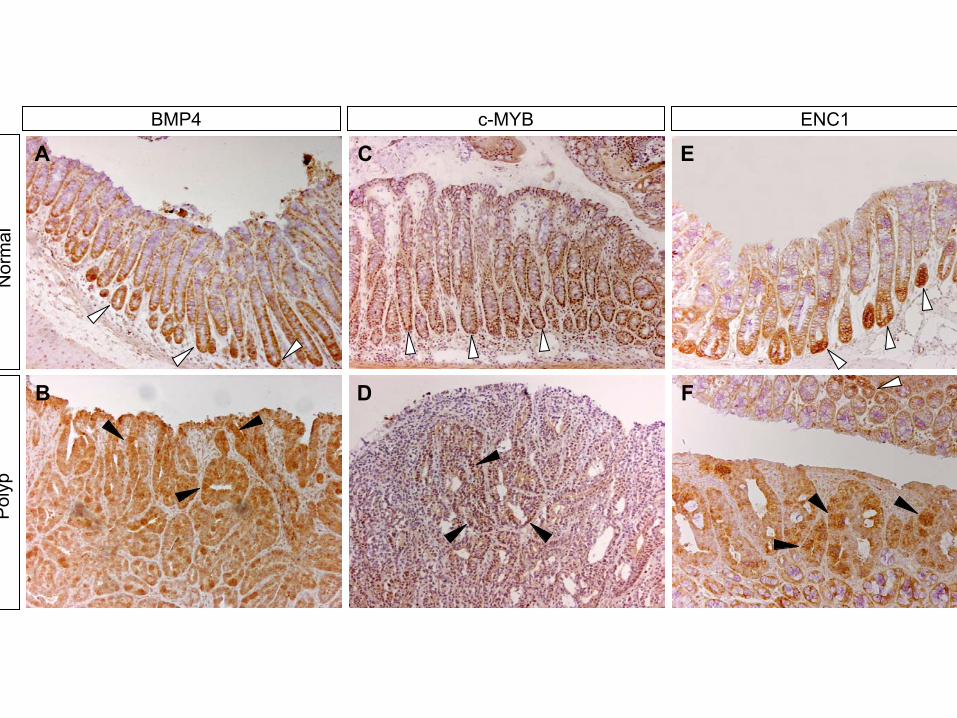
Immunohistochemical analyses were then performed on(see Supplemental Table S1). Five genes (i.e., CD44,BMP4, claudin-1, ENC1, and c-MYC) had been de- early neoplastic lesions. A strict correlation between
nuclear �-catenin (Figure 2A) and the expression of tar-scribed previously as TCF targets in CRC (Wielenga etal., 1999; Miwa et al., 2001; Kim et al., 2002; Fujita et get genes such as EPHB2 (Figure 2B) was observed
in early colorectal lesions. Many other downregulatedal., 2001; He et al., 1998). CD44, c-ETS2, and nine otherentries occurred twice, indicative of the robustness of genes listed were overexpressed in early intestinal pol-
yps from Min mice or in aberrant crypt foci (ACF) fromthe analysis (Supplemental Table S1). For a number of
Cell244
FAP patients (Figure 7, Supplemental Figure S4; Ramsayet al., 1992; Wielenga et al., 1999; Ciclitira et al., 1987;Erisman et al., 1985; Oba et al., 2001; Kitahara et al.,2001; Miwa et al., 2001). More strikingly, all tested targetgenes were expressed also in the proliferative compart-ment of normal colon crypts (EPHB2, Figure 2C; c-MYB,BMP4, ENC1, Figure S4; CD44, Figure 7; EPHB3, notshown). Several other downregulated genes from ourlist (claudin-1, c-MYC, and the Na-K-Cl cotransporterSLC12A2) are reportedly expressed in crypt cells (Miwaet al., 2001; Melhem et al., 1992; Matthews et al., 1998).
A complementary domain of expression was ob-served for genes strongly upregulated upon induction ofdnTCF-4 in Ls174T cells. One such example is carbonicanhydrase II, which was restricted to the top of thecrypts and the surface epithelium (Figure 2F) but wasabsent from polyp cells arising in this area (Figure 2E).
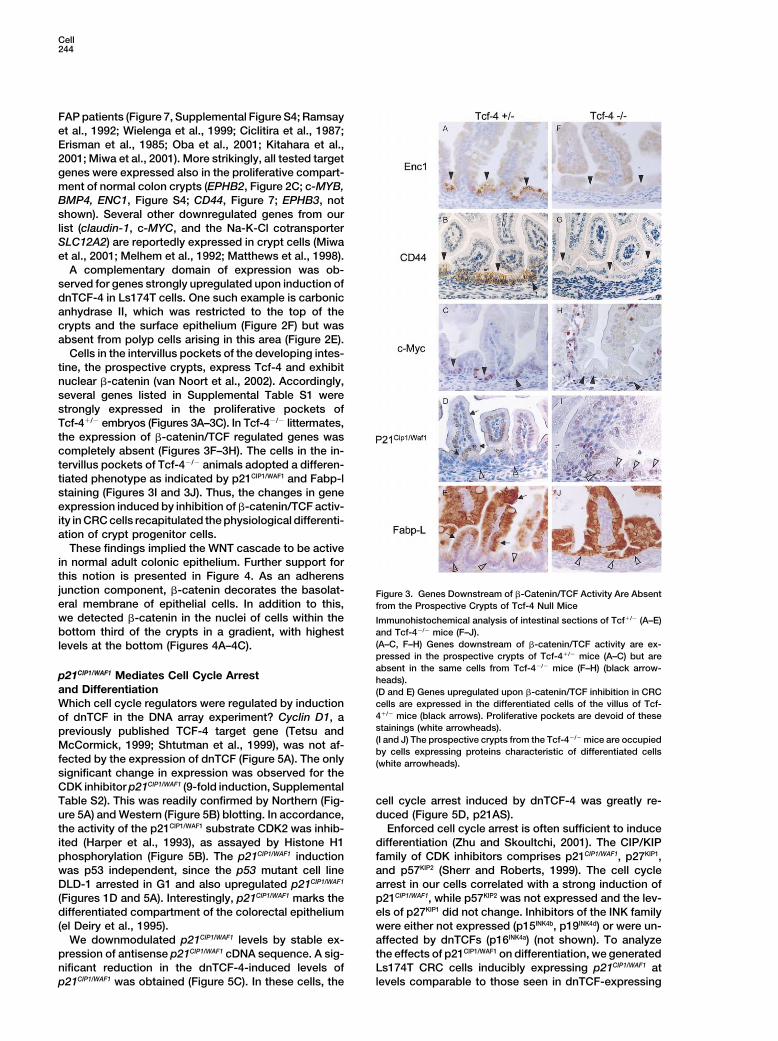
Cells in the intervillus pockets of the developing intes-tine, the prospective crypts, express Tcf-4 and exhibitnuclear �-catenin (van Noort et al., 2002). Accordingly,several genes listed in Supplemental Table S1 werestrongly expressed in the proliferative pockets ofTcf-4�/� embryos (Figures 3A–3C). In Tcf-4�/� littermates,the expression of �-catenin/TCF regulated genes wascompletely absent (Figures 3F–3H). The cells in the in-tervillus pockets of Tcf-4�/� animals adopted a differen-tiated phenotype as indicated by p21CIP1/WAF1 and Fabp-lstaining (Figures 3I and 3J). Thus, the changes in geneexpression induced by inhibition of �-catenin/TCF activ-ity in CRC cells recapitulated the physiological differenti-ation of crypt progenitor cells.
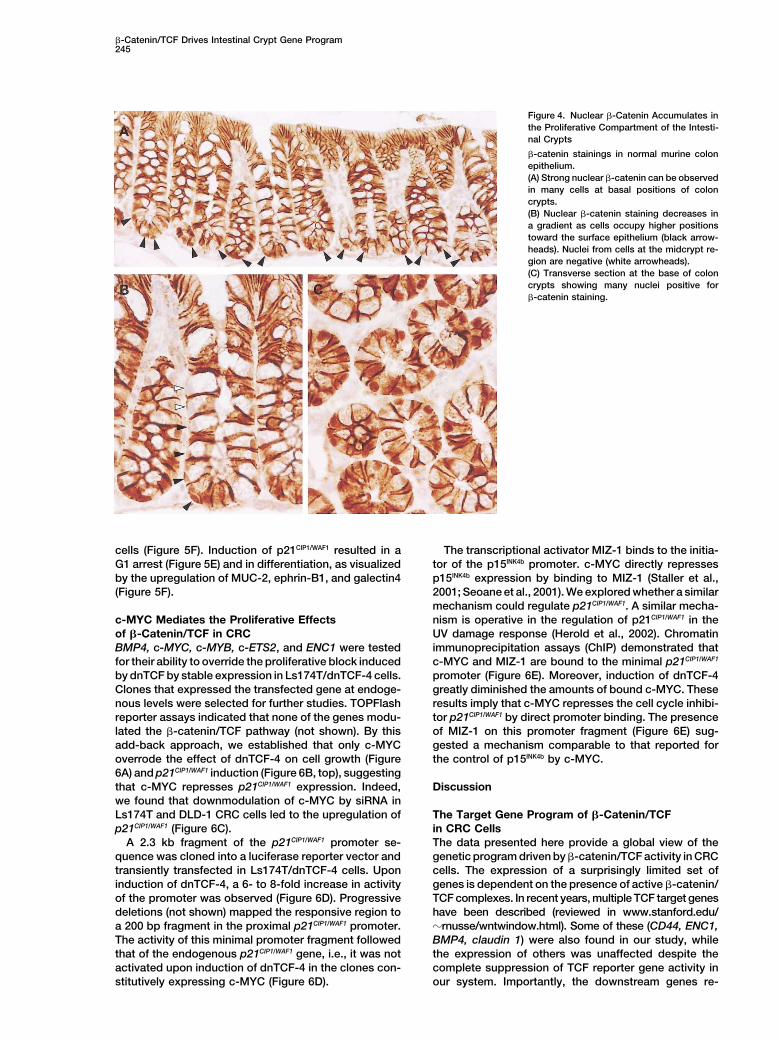
These findings implied the WNT cascade to be activein normal adult colonic epithelium. Further support forthis notion is presented in Figure 4. As an adherensjunction component, �-catenin decorates the basolat- Figure 3. Genes Downstream of �-Catenin/TCF Activity Are Absenteral membrane of epithelial cells. In addition to this, from the Prospective Crypts of Tcf-4 Null Micewe detected �-catenin in the nuclei of cells within the Immunohistochemical analysis of intestinal sections of Tcf�/� (A–E)
and Tcf-4�/� mice (F–J).bottom third of the crypts in a gradient, with highest(A–C, F–H) Genes downstream of �-catenin/TCF activity are ex-levels at the bottom (Figures 4A–4C).pressed in the prospective crypts of Tcf-4�/� mice (A–C) but areabsent in the same cells from Tcf-4�/� mice (F–H) (black arrow-
p21CIP1/WAF1 Mediates Cell Cycle Arrest heads).and Differentiation (D and E) Genes upregulated upon �-catenin/TCF inhibition in CRC
cells are expressed in the differentiated cells of the villus of Tcf-Which cell cycle regulators were regulated by induction4�/� mice (black arrows). Proliferative pockets are devoid of theseof dnTCF in the DNA array experiment? Cyclin D1, astainings (white arrowheads).previously published TCF-4 target gene (Tetsu and(I and J) The prospective crypts from the Tcf-4�/� mice are occupiedMcCormick, 1999; Shtutman et al., 1999), was not af-by cells expressing proteins characteristic of differentiated cells
fected by the expression of dnTCF (Figure 5A). The only (white arrowheads).significant change in expression was observed for theCDK inhibitor p21CIP1/WAF1 (9-fold induction, SupplementalTable S2). This was readily confirmed by Northern (Fig- cell cycle arrest induced by dnTCF-4 was greatly re-
duced (Figure 5D, p21AS).ure 5A) and Western (Figure 5B) blotting. In accordance,the activity of the p21CIP1/WAF1 substrate CDK2 was inhib- Enforced cell cycle arrest is often sufficient to induce
differentiation (Zhu and Skoultchi, 2001). The CIP/KIPited (Harper et al., 1993), as assayed by Histone H1phosphorylation (Figure 5B). The p21CIP1/WAF1 induction family of CDK inhibitors comprises p21CIP1/WAF1, p27KIP1,
and p57KIP2 (Sherr and Roberts, 1999). The cell cyclewas p53 independent, since the p53 mutant cell lineDLD-1 arrested in G1 and also upregulated p21CIP1/WAF1 arrest in our cells correlated with a strong induction of
p21CIP1/WAF1, while p57KIP2 was not expressed and the lev-(Figures 1D and 5A). Interestingly, p21CIP1/WAF1 marks thedifferentiated compartment of the colorectal epithelium els of p27KIP1 did not change. Inhibitors of the INK family
were either not expressed (p15INK4b, p19INK4d) or were un-(el Deiry et al., 1995).We downmodulated p21CIP1/WAF1 levels by stable ex- affected by dnTCFs (p16INK4a) (not shown). To analyze
the effects of p21CIP1/WAF1 on differentiation, we generatedpression of antisense p21CIP1/WAF1 cDNA sequence. A sig-nificant reduction in the dnTCF-4-induced levels of Ls174T CRC cells inducibly expressing p21CIP1/WAF1 at
levels comparable to those seen in dnTCF-expressingp21CIP1/WAF1 was obtained (Figure 5C). In these cells, the
�-Catenin/TCF Drives Intestinal Crypt Gene Program245
Figure 4. Nuclear �-Catenin Accumulates inthe Proliferative Compartment of the Intesti-nal Crypts
�-catenin stainings in normal murine colonepithelium.(A) Strong nuclear �-catenin can be observedin many cells at basal positions of coloncrypts.(B) Nuclear �-catenin staining decreases ina gradient as cells occupy higher positionstoward the surface epithelium (black arrow-heads). Nuclei from cells at the midcrypt re-gion are negative (white arrowheads).(C) Transverse section at the base of coloncrypts showing many nuclei positive for�-catenin staining.
cells (Figure 5F). Induction of p21CIP1/WAF1 resulted in a The transcriptional activator MIZ-1 binds to the initia-tor of the p15INK4b promoter. c-MYC directly repressesG1 arrest (Figure 5E) and in differentiation, as visualized
by the upregulation of MUC-2, ephrin-B1, and galectin4 p15INK4b expression by binding to MIZ-1 (Staller et al.,2001; Seoane et al., 2001). We explored whether a similar(Figure 5F).mechanism could regulate p21CIP1/WAF1. A similar mecha-nism is operative in the regulation of p21CIP1/WAF1 in thec-MYC Mediates the Proliferative Effects
of �-Catenin/TCF in CRC UV damage response (Herold et al., 2002). Chromatinimmunoprecipitation assays (ChIP) demonstrated thatBMP4, c-MYC, c-MYB, c-ETS2, and ENC1 were tested
for their ability to override the proliferative block induced c-MYC and MIZ-1 are bound to the minimal p21CIP1/WAF1
promoter (Figure 6E). Moreover, induction of dnTCF-4by dnTCF by stable expression in Ls174T/dnTCF-4 cells.Clones that expressed the transfected gene at endoge- greatly diminished the amounts of bound c-MYC. These
results imply that c-MYC represses the cell cycle inhibi-nous levels were selected for further studies. TOPFlashreporter assays indicated that none of the genes modu- tor p21CIP1/WAF1 by direct promoter binding. The presence
of MIZ-1 on this promoter fragment (Figure 6E) sug-lated the �-catenin/TCF pathway (not shown). By thisadd-back approach, we established that only c-MYC gested a mechanism comparable to that reported for
the control of p15INK4b by c-MYC.overrode the effect of dnTCF-4 on cell growth (Figure6A) and p21CIP1/WAF1 induction (Figure 6B, top), suggestingthat c-MYC represses p21CIP1/WAF1 expression. Indeed, Discussionwe found that downmodulation of c-MYC by siRNA inLs174T and DLD-1 CRC cells led to the upregulation of The Target Gene Program of �-Catenin/TCF
in CRC Cellsp21CIP1/WAF1 (Figure 6C).A 2.3 kb fragment of the p21CIP1/WAF1 promoter se- The data presented here provide a global view of the
genetic program driven by �-catenin/TCF activity in CRCquence was cloned into a luciferase reporter vector andtransiently transfected in Ls174T/dnTCF-4 cells. Upon cells. The expression of a surprisingly limited set of
genes is dependent on the presence of active �-catenin/induction of dnTCF-4, a 6- to 8-fold increase in activityof the promoter was observed (Figure 6D). Progressive TCF complexes. In recent years, multiple TCF target genes
have been described (reviewed in www.stanford.edu/deletions (not shown) mapped the responsive region toa 200 bp fragment in the proximal p21CIP1/WAF1 promoter. �rnusse/wntwindow.html). Some of these (CD44, ENC1,
BMP4, claudin 1) were also found in our study, whileThe activity of this minimal promoter fragment followedthat of the endogenous p21CIP1/WAF1 gene, i.e., it was not the expression of others was unaffected despite the
complete suppression of TCF reporter gene activity inactivated upon induction of dnTCF-4 in the clones con-stitutively expressing c-MYC (Figure 6D). our system. Importantly, the downstream genes re-
Cell246
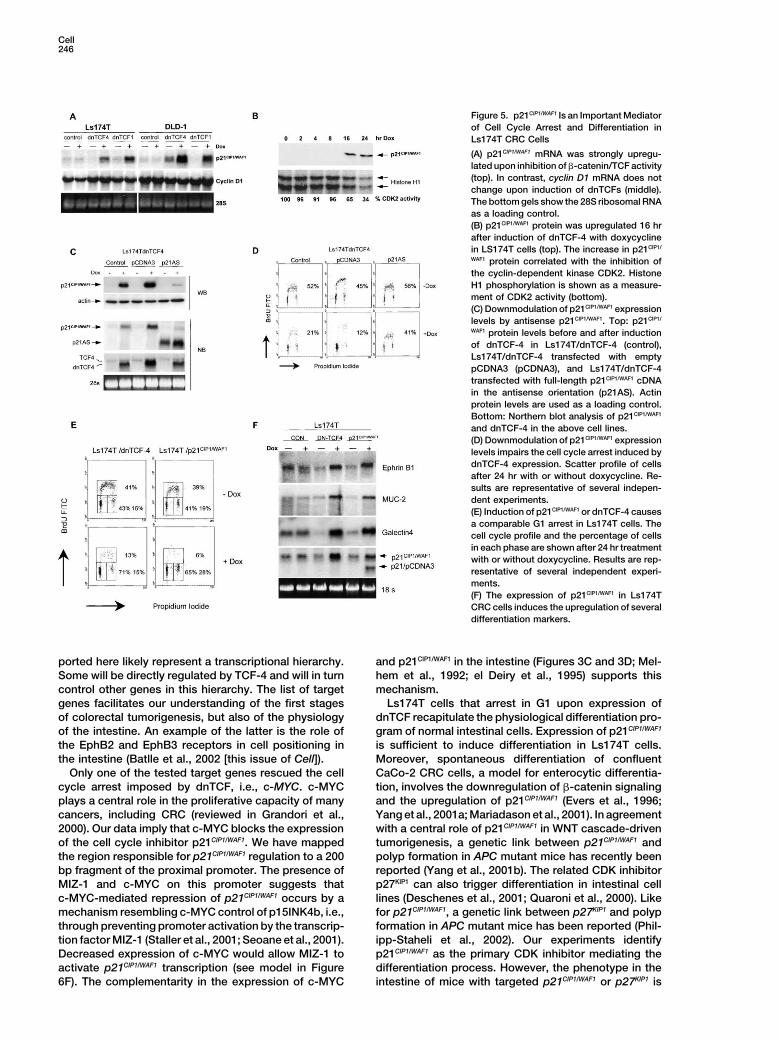
Figure 5. p21CIP1/WAF1 Is an Important Mediatorof Cell Cycle Arrest and Differentiation inLs174T CRC Cells
(A) p21CIP1/WAF1 mRNA was strongly upregu-lated upon inhibition of �-catenin/TCF activity(top). In contrast, cyclin D1 mRNA does notchange upon induction of dnTCFs (middle).The bottom gels show the 28S ribosomal RNAas a loading control.(B) p21CIP1/WAF1 protein was upregulated 16 hrafter induction of dnTCF-4 with doxycyclinein LS174T cells (top). The increase in p21CIP1/
WAF1 protein correlated with the inhibition ofthe cyclin-dependent kinase CDK2. HistoneH1 phosphorylation is shown as a measure-ment of CDK2 activity (bottom).(C) Downmodulation of p21CIP1/WAF1 expressionlevels by antisense p21CIP1/WAF1. Top: p21CIP1/
WAF1 protein levels before and after inductionof dnTCF-4 in Ls174T/dnTCF-4 (control),Ls174T/dnTCF-4 transfected with emptypCDNA3 (pCDNA3), and Ls174T/dnTCF-4transfected with full-length p21CIP1/WAF1 cDNAin the antisense orientation (p21AS). Actinprotein levels are used as a loading control.Bottom: Northern blot analysis of p21CIP1/WAF1
and dnTCF-4 in the above cell lines.(D) Downmodulation of p21CIP1/WAF1 expressionlevels impairs the cell cycle arrest induced bydnTCF-4 expression. Scatter profile of cellsafter 24 hr with or without doxycycline. Re-sults are representative of several indepen-dent experiments.(E) Induction of p21CIP1/WAF1 or dnTCF-4 causesa comparable G1 arrest in Ls174T cells. Thecell cycle profile and the percentage of cellsin each phase are shown after 24 hr treatmentwith or without doxycycline. Results are rep-resentative of several independent experi-ments.(F) The expression of p21CIP1/WAF1 in Ls174TCRC cells induces the upregulation of severaldifferentiation markers.
ported here likely represent a transcriptional hierarchy. and p21CIP1/WAF1 in the intestine (Figures 3C and 3D; Mel-hem et al., 1992; el Deiry et al., 1995) supports thisSome will be directly regulated by TCF-4 and will in turn
control other genes in this hierarchy. The list of target mechanism.Ls174T cells that arrest in G1 upon expression ofgenes facilitates our understanding of the first stages
of colorectal tumorigenesis, but also of the physiology dnTCF recapitulate the physiological differentiation pro-gram of normal intestinal cells. Expression of p21CIP1/WAF1of the intestine. An example of the latter is the role of
the EphB2 and EphB3 receptors in cell positioning in is sufficient to induce differentiation in Ls174T cells.Moreover, spontaneous differentiation of confluentthe intestine (Batlle et al., 2002 [this issue of Cell]).
Only one of the tested target genes rescued the cell CaCo-2 CRC cells, a model for enterocytic differentia-tion, involves the downregulation of �-catenin signalingcycle arrest imposed by dnTCF, i.e., c-MYC. c-MYC
plays a central role in the proliferative capacity of many and the upregulation of p21CIP1/WAF1 (Evers et al., 1996;Yang et al., 2001a; Mariadason et al., 2001). In agreementcancers, including CRC (reviewed in Grandori et al.,
2000). Our data imply that c-MYC blocks the expression with a central role of p21CIP1/WAF1 in WNT cascade-driventumorigenesis, a genetic link between p21CIP1/WAF1 andof the cell cycle inhibitor p21CIP1/WAF1. We have mapped
the region responsible for p21CIP1/WAF1 regulation to a 200 polyp formation in APC mutant mice has recently beenreported (Yang et al., 2001b). The related CDK inhibitorbp fragment of the proximal promoter. The presence of
MIZ-1 and c-MYC on this promoter suggests that p27KIP1 can also trigger differentiation in intestinal celllines (Deschenes et al., 2001; Quaroni et al., 2000). Likec-MYC-mediated repression of p21CIP1/WAF1 occurs by a
mechanism resembling c-MYC control of p15INK4b, i.e., for p21CIP1/WAF1, a genetic link between p27KIP1 and polypformation in APC mutant mice has been reported (Phil-through preventing promoter activation by the transcrip-
tion factor MIZ-1 (Staller et al., 2001; Seoane et al., 2001). ipp-Staheli et al., 2002). Our experiments identifyp21CIP1/WAF1 as the primary CDK inhibitor mediating theDecreased expression of c-MYC would allow MIZ-1 to
activate p21CIP1/WAF1 transcription (see model in Figure differentiation process. However, the phenotype in theintestine of mice with targeted p21CIP1/WAF1 or p27KIP1 is6F). The complementarity in the expression of c-MYC
�-Catenin/TCF Drives Intestinal Crypt Gene Program247
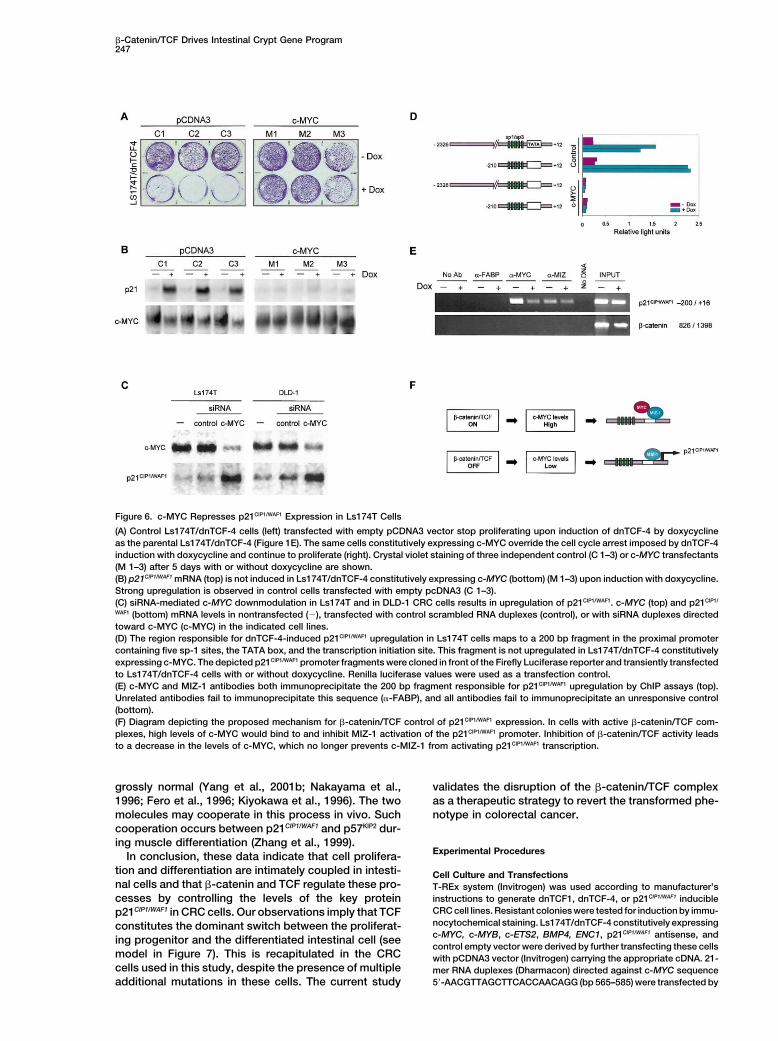
Figure 6. c-MYC Represses p21CIP1/WAF1 Expression in Ls174T Cells
(A) Control Ls174T/dnTCF-4 cells (left) transfected with empty pCDNA3 vector stop proliferating upon induction of dnTCF-4 by doxycyclineas the parental Ls174T/dnTCF-4 (Figure 1E). The same cells constitutively expressing c-MYC override the cell cycle arrest imposed by dnTCF-4induction with doxycycline and continue to proliferate (right). Crystal violet staining of three independent control (C 1–3) or c-MYC transfectants(M 1–3) after 5 days with or without doxycycline are shown.(B) p21CIP1/WAF1 mRNA (top) is not induced in Ls174T/dnTCF-4 constitutively expressing c-MYC (bottom) (M 1–3) upon induction with doxycycline.Strong upregulation is observed in control cells transfected with empty pcDNA3 (C 1–3).(C) siRNA-mediated c-MYC downmodulation in Ls174T and in DLD-1 CRC cells results in upregulation of p21CIP1/WAF1. c-MYC (top) and p21CIP1/
WAF1 (bottom) mRNA levels in nontransfected (�), transfected with control scrambled RNA duplexes (control), or with siRNA duplexes directedtoward c-MYC (c-MYC) in the indicated cell lines.(D) The region responsible for dnTCF-4-induced p21CIP1/WAF1 upregulation in Ls174T cells maps to a 200 bp fragment in the proximal promotercontaining five sp-1 sites, the TATA box, and the transcription initiation site. This fragment is not upregulated in Ls174T/dnTCF-4 constitutivelyexpressing c-MYC. The depicted p21CIP1/WAF1 promoter fragments were cloned in front of the Firefly Luciferase reporter and transiently transfectedto Ls174T/dnTCF-4 cells with or without doxycycline. Renilla luciferase values were used as a transfection control.(E) c-MYC and MIZ-1 antibodies both immunoprecipitate the 200 bp fragment responsible for p21CIP1/WAF1 upregulation by ChIP assays (top).Unrelated antibodies fail to immunoprecipitate this sequence (�-FABP), and all antibodies fail to immunoprecipitate an unresponsive control(bottom).(F) Diagram depicting the proposed mechanism for �-catenin/TCF control of p21CIP1/WAF1 expression. In cells with active �-catenin/TCF com-plexes, high levels of c-MYC would bind to and inhibit MIZ-1 activation of the p21CIP1/WAF1 promoter. Inhibition of �-catenin/TCF activity leadsto a decrease in the levels of c-MYC, which no longer prevents c-MIZ-1 from activating p21CIP1/WAF1 transcription.
grossly normal (Yang et al., 2001b; Nakayama et al., validates the disruption of the �-catenin/TCF complexas a therapeutic strategy to revert the transformed phe-1996; Fero et al., 1996; Kiyokawa et al., 1996). The two
molecules may cooperate in this process in vivo. Such notype in colorectal cancer.cooperation occurs between p21CIP1/WAF1 and p57KIP2 dur-ing muscle differentiation (Zhang et al., 1999).
Experimental ProceduresIn conclusion, these data indicate that cell prolifera-tion and differentiation are intimately coupled in intesti- Cell Culture and Transfectionsnal cells and that �-catenin and TCF regulate these pro- T-REx system (Invitrogen) was used according to manufacturer’s
instructions to generate dnTCF1, dnTCF-4, or p21CIP1/WAF1 induciblecesses by controlling the levels of the key proteinCRC cell lines. Resistant colonies were tested for induction by immu-p21CIP1/WAF1 in CRC cells. Our observations imply that TCFnocytochemical staining. Ls174T/dnTCF-4 constitutively expressingconstitutes the dominant switch between the proliferat-c-MYC, c-MYB, c-ETS2, BMP4, ENC1, p21CIP1/WAF1 antisense, anding progenitor and the differentiated intestinal cell (seecontrol empty vector were derived by further transfecting these cells
model in Figure 7). This is recapitulated in the CRC with pCDNA3 vector (Invitrogen) carrying the appropriate cDNA. 21-cells used in this study, despite the presence of multiple mer RNA duplexes (Dharmacon) directed against c-MYC sequence
5�-AACGTTAGCTTCACCAACAGG (bp 565–585) were transfected byadditional mutations in these cells. The current study
Cell248
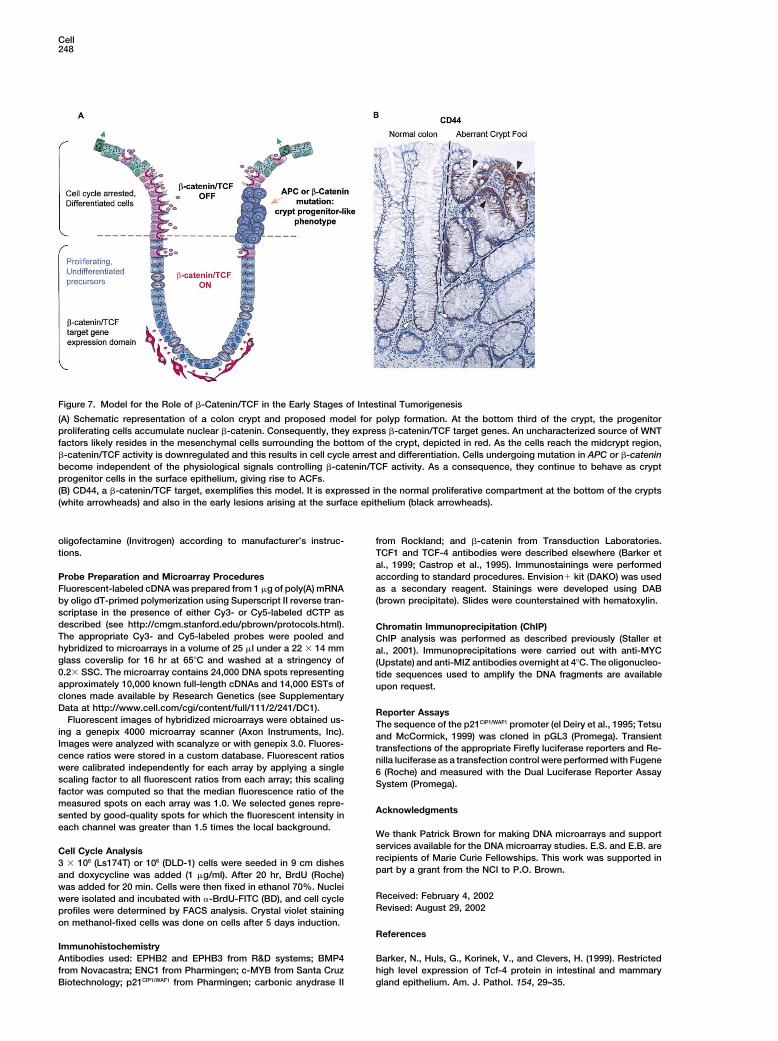
Figure 7. Model for the Role of �-Catenin/TCF in the Early Stages of Intestinal Tumorigenesis
(A) Schematic representation of a colon crypt and proposed model for polyp formation. At the bottom third of the crypt, the progenitorproliferating cells accumulate nuclear �-catenin. Consequently, they express �-catenin/TCF target genes. An uncharacterized source of WNTfactors likely resides in the mesenchymal cells surrounding the bottom of the crypt, depicted in red. As the cells reach the midcrypt region,�-catenin/TCF activity is downregulated and this results in cell cycle arrest and differentiation. Cells undergoing mutation in APC or �-cateninbecome independent of the physiological signals controlling �-catenin/TCF activity. As a consequence, they continue to behave as cryptprogenitor cells in the surface epithelium, giving rise to ACFs.(B) CD44, a �-catenin/TCF target, exemplifies this model. It is expressed in the normal proliferative compartment at the bottom of the crypts(white arrowheads) and also in the early lesions arising at the surface epithelium (black arrowheads).
oligofectamine (Invitrogen) according to manufacturer’s instruc- from Rockland; and �-catenin from Transduction Laboratories.TCF1 and TCF-4 antibodies were described elsewhere (Barker ettions.al., 1999; Castrop et al., 1995). Immunostainings were performedaccording to standard procedures. Envision� kit (DAKO) was usedProbe Preparation and Microarray Procedures
Fluorescent-labeled cDNA was prepared from 1 �g of poly(A) mRNA as a secondary reagent. Stainings were developed using DAB(brown precipitate). Slides were counterstained with hematoxylin.by oligo dT-primed polymerization using Superscript II reverse tran-
scriptase in the presence of either Cy3- or Cy5-labeled dCTP asdescribed (see http://cmgm.stanford.edu/pbrown/protocols.html). Chromatin Immunoprecipitation (ChIP)The appropriate Cy3- and Cy5-labeled probes were pooled and ChIP analysis was performed as described previously (Staller ethybridized to microarrays in a volume of 25 �l under a 22 � 14 mm al., 2001). Immunoprecipitations were carried out with anti-MYCglass coverslip for 16 hr at 65�C and washed at a stringency of (Upstate) and anti-MIZ antibodies overnight at 4�C. The oligonucleo-0.2� SSC. The microarray contains 24,000 DNA spots representing tide sequences used to amplify the DNA fragments are availableapproximately 10,000 known full-length cDNAs and 14,000 ESTs of upon request.clones made available by Research Genetics (see SupplementaryData at http://www.cell.com/cgi/content/full/111/2/241/DC1). Reporter Assays
Fluorescent images of hybridized microarrays were obtained us- The sequence of the p21CIP1/WAF1 promoter (el Deiry et al., 1995; Tetsuing a genepix 4000 microarray scanner (Axon Instruments, Inc). and McCormick, 1999) was cloned in pGL3 (Promega). TransientImages were analyzed with scanalyze or with genepix 3.0. Fluores- transfections of the appropriate Firefly luciferase reporters and Re-cence ratios were stored in a custom database. Fluorescent ratios nilla luciferase as a transfection control were performed with Fugenewere calibrated independently for each array by applying a single 6 (Roche) and measured with the Dual Luciferase Reporter Assayscaling factor to all fluorescent ratios from each array; this scaling System (Promega).factor was computed so that the median fluorescence ratio of themeasured spots on each array was 1.0. We selected genes repre-
Acknowledgmentssented by good-quality spots for which the fluorescent intensity ineach channel was greater than 1.5 times the local background.
We thank Patrick Brown for making DNA microarrays and supportservices available for the DNA microarray studies. E.S. and E.B. areCell Cycle Analysisrecipients of Marie Curie Fellowships. This work was supported in3 � 106 (Ls174T) or 106 (DLD-1) cells were seeded in 9 cm dishespart by a grant from the NCI to P.O. Brown.and doxycycline was added (1 �g/ml). After 20 hr, BrdU (Roche)
was added for 20 min. Cells were then fixed in ethanol 70%. NucleiReceived: February 4, 2002were isolated and incubated with �-BrdU-FITC (BD), and cell cycleRevised: August 29, 2002profiles were determined by FACS analysis. Crystal violet staining
on methanol-fixed cells was done on cells after 5 days induction.References
ImmunohistochemistryAntibodies used: EPHB2 and EPHB3 from R&D systems; BMP4 Barker, N., Huls, G., Korinek, V., and Clevers, H. (1999). Restricted
high level expression of Tcf-4 protein in intestinal and mammaryfrom Novacastra; ENC1 from Pharmingen; c-MYB from Santa CruzBiotechnology; p21CIP1/WAF1 from Pharmingen; carbonic anydrase II gland epithelium. Am. J. Pathol. 154, 29–35.

�-Catenin/TCF Drives Intestinal Crypt Gene Program249
Batlle, E., Henderson, J.T., Beghtel, H., van den Born, M.M.W., San- Kinzler, K.W., Vogelstein, B., and Clevers, H. (1997). Constitutivetranscriptional activation by a beta-catenin-Tcf complex in APC�/�cho, E., Huls, G., Meeldijk, J., Roberston, J., van de Wetering, M.,
Pawson, T., and Clevers, H. (2002). �-catenin and TCF mediate cell colon carcinoma. Science 275, 1784–1787.positioning in the intestinal epithelium by controlling the expression Korinek, V., Barker, N., Moerer, P., van Donselaar, E., Huls, G., Pe-of EphB/ephrinB. Cell 111, this issue, 251–263. ters, P.J., and Clevers, H. (1998). Depletion of epithelial stem-cellBienz, M., and Clevers, H. (2000). Linking colorectal cancer to Wnt compartments in the small intestine of mice lacking Tcf-4. Nat.signaling. Cell 103, 311–320. Genet. 19, 379–383.
Castrop, J., van Wichen, D., Koomans Bitter, M., van de Wetering, Korswagen, H.C., Herman, M.A., and Clevers, H.C. (2000). DistinctM., de Weger, R., van Dongen, J., and Clevers, H. (1995). The human beta-catenins mediate adhesion and signalling functions in C. ele-TCF-1 gene encodes a nuclear DNA-binding protein uniquely ex- gans. Nature 406, 527–532.pressed in normal and neoplastic T-lineage lymphocytes. Blood 86, Liu, W., Dong, X., Mai, M., Seelan, R.S., Taniguchi, K., Krishnadath,3050–3059. K.K., Halling, K.C., Cunningham, J.M., Qian, C., Christensen, E., etCiclitira, P.J., Macartney, J.C., and Evan, G. (1987). Expression of al. (2000). Mutations in AXIN2 cause colorectal cancer with defectivec-myc in non-malignant and pre-malignant gastrointestinal disor- mismatch repair by activating beta-catenin/TCF signalling. Nat.ders. J. Pathol. 151, 293–296. Genet. 26, 146–147.
Deschenes, C., Vezina, A., Beaulieu, J.F., and Rivard, N. (2001). Role Mariadason, J.M., Bordonaro, M., Aslam, F., Shi, L., Kuraguchi, M.,of p27(Kip1) in human intestinal cell differentiation. Gastroenterol- Velcich, A., and Augenlicht, L.H. (2001). Down-regulation of beta-ogy 120, 423–438. catenin TCF signaling is linked to colonic epithelial cell differentia-
tion. Cancer Res. 61, 3465–3471.el Deiry, W.S., Tokino, T., Waldman, T., Oliner, J.D., Velculescu, V.E.,Burrell, M., Hill, D.E., Healy, E., Rees, J.L., Hamilton, S.R., et al. Matthews, J.B., Hassan, I., Meng, S., Archer, S.Y., Hrnjez, B.J., and(1995). Topological control of p21WAF1/CIP1 expression in normal Hodin, R.A. (1998). Na-K-2Cl cotransporter gene expression andand neoplastic tissues. Cancer Res. 55, 2910–2919. function during enterocyte differentiation. Modulation of Cl-secre-
tory capacity by butyrate. J. Clin. Invest. 101, 2072–2079.Erisman, M.D., Rothberg, P.G., Diehl, R.E., Morse, C.C., Spandorfer,J.M., and Astrin, S.M. (1985). Deregulation of c-myc gene expression Melhem, M.F., Meisler, A.I., Finley, G.G., Bryce, W.H., Jones, M.O.,in human colon carcinoma is not accompanied by amplification or Tribby, I.I., Pipas, J.M., and Koski, R.A. (1992). Distribution of cellsrearrangement of the gene. Mol. Cell. Biol. 5, 1969–1976. expressing myc proteins in human colorectal epithelium, polyps,
and malignant tumors. Cancer Res. 52, 5853–5864.Evers, B.M., Ko, T.C., Li, J., and Thompson, E.A. (1996). Cell cycleprotein suppression and p21 induction in differentiating Caco-2 Miwa, N., Furuse, M., Tsukita, S., Niikawa, N., Nakamura, Y., andcells. Am. J. Physiol. 271, G722–G727. Furukawa, Y. (2001). Involvement of claudin-1 in the beta-catenin/
Tcf signaling pathway and its frequent upregulation in human colo-Fero, M.L., Rivkin, M., Tasch, M., Porter, P., Carow, C.E., Firpo, E.,rectal cancers. Oncol. Res. 12, 469–476.Polyak, K., Tsai, L.H., Broudy, V., Perlmutter, R.M., et al. (1996).
A syndrome of multiorgan hyperplasia with features of gigantism, Molenaar, M., van de Wetering, M., Oosterwegel, M., Peterson-tumorigenesis, and female sterility in p27(Kip1)-deficient mice. Cell Maduro, J., Godsave, S., Korinek, V., Roose, J., Destree, O., and85, 733–744. Clevers, H. (1996). XTcf-3 transcription factor mediates beta-
catenin-induced axis formation in Xenopus embryos. Cell 86,Fujita, M., Furukawa, Y., Tsunoda, T., Tanaka, T., Ogawa, M., and391–399.Nakamura, Y. (2001). Up-regulation of the ectodermal-neural cortex
1 (ENC1) gene, a downstream target of the beta-catenin/T-cell factor Morin, P.J., Sparks, A.B., Korinek, V., Barker, N., Clevers, H., Vo-complex, in colorectal carcinomas. Cancer Res. 61, 7722–7726. gelstein, B., and Kinzler, K.W. (1997). Activation of beta-catenin-
Tcf signaling in colon cancer by mutations in beta-catenin or APC.Grandori, C., Cowley, S.M., James, L.P., and Eisenman, R.N. (2000).Science 275, 1787–1790.The Myc/Max/Mad network and the transcriptional control of cell
behaviour. Annu. Rev. Cell Dev. Biol. 16, 653–699. Nakayama, K., Ishida, N., Shirane, M., Inomata, A., Inoue, T., Shi-shido, N., Horii, I., Loh, D.Y., and Nakayama, K. (1996). Mice lackingHarper, J.W., Adami, G.R., Wei, N., Keyomarsi, K., and Elledge, S.J.p27(Kip1) display increased body size, multiple organ hyperplasia,(1993). The p21 Cdk-interacting protein Cip1 is a potent inhibitor ofretinal dysplasia, and pituitary tumors. Cell 85, 707–720.G1 cyclin-dependent kinases. Cell 75, 805–816.
Oba, S.M., Wang, Y.J., Song, J.P., Li, Z.Y., Kobayashi, K., Tsugane,He, T.C., Sparks, A.B., Rago, C., Hermeking, H., Zawel, L., da Costa,S., Hamada, G.S., Tanaka, M., and Sugimura, H. (2001). GenomicL.T., Morin, P.J., Vogelstein, B., and Kinzler, K.W. (1998). Identifica-structure and loss of heterozygosity of EPHB2 in colorectal cancer.tion of c-MYC as a target of the APC pathway. Science 281, 1509–Cancer Lett. 164, 97–104.1512.
Philipp-Staheli, J., Kim, K.H., Payne, S.R., Gurley, K.E., Liggitt, D.,Herold, S., Wanzel, M., Beuger, V., Frohme, C., Beul, D., Hillukkala,Longton, G., and Kemp, C.J. (2002). Pathway-specific tumor sup-T., Syvaoja, J., Saluz, H.-P., Haenel, F., and Eilers, M. (2002). Nega-pression. Reduction of p27 accelerates gastrointestinal tumorigene-tive regulation of the mammalian UV response by Myc through asso-sis in Apc mutant mice, but not in Smad3 mutant mice. Cancer Cellciation with Miz-1. Mol. Cell 10, 509–521.1, 355–368.Kim, J.S., Crooks, H., Dracheva, T., Nishanian, T.G., Singh, B., Jen,Potten, C.S., and Loeffler, M. (1990). Stem cells: attributes, cycles,J., and Waldman, T. (2002). Oncogenic beta-catenin is required forspirals, pitfalls and uncertainties. Lessons for and from the crypt.bone morphogenetic protein 4 expression in human cancer cells.Development 110, 1001–1020.Cancer Res. 62, 2744–2748.
Powell, S.M., Zilz, N., Beazer-Barclay, Y., Bryan, T.M., Hamilton,Kinzler, K.W., and Vogelstein, B. (1996). Lessons from hereditaryS.R., Thibodeau, S.N., Vogelstein, B., and Kinzler, K.W. (1992). APCcolorectal cancer. Cell 87, 159–170.mutations occur early during colorectal tumorigenesis. Nature 359,Kitahara, O., Furukawa, Y., Tanaka, T., Kihara, C., Ono, K., Yana-235–237.gawa, R., Nita, M.E., Takagi, T., Nakamura, Y., and Tsunoda, T.Quaroni, A., Tian, J.Q., Seth, P., and Ap, R.C. (2000). p27(Kip1) is(2001). Alterations of gene expression during colorectal carcinogen-an inducer of intestinal epithelial cell differentiation. Am. J. Physiol.esis revealed by cDNA microarrays after laser-capture microdissec-Cell Physiol. 279, C1045–C1057.tion of tumor tissues and normal epithelia. Cancer Res. 61, 3544–
3549. Ramsay, R.G., Thompson, M.A., Hayman, J.A., Reid, G., Gonda, T.J.,and Whitehead, R.H. (1992). Myb expression is higher in malignantKiyokawa, H., Kineman, R.D., Manova-Todorova, K.O., Soares, V.C.,human colonic carcinoma and premalignant adenomatous polypsHoffman, E.S., Ono, M., Khanam, D., Hayday, A.C., Frohman, L.A.,than in normal mucosa. Cell Growth Differ. 3, 723–730.and Koff, A. (1996). Enhanced growth of mice lacking the cyclin-
dependent kinase inhibitor function of p27(Kip1). Cell 85, 721–732. Roose, J., Huls, G., van Beest, M., Moerer, P., van der Horn, K.,Goldschmeding, R., Logtenberg, T., and Clevers, H. (1999). SynergyKorinek, V., Barker, N., Morin, P.J., van Wichen, D., de Weger, R.,

Cell250
between tumor suppressor APC and the beta-catenin-Tcf4 targetTcf1. Science 285, 1923–1926.
Rubinfeld, B., Robbins, P., El Gamil, M., Albert, I., Porfiri, E., andPolakis, P. (1997). Stabilization of beta-catenin by genetic defectsin melanoma cell lines. Science 275, 1790–1792.
Satoh, S., Daigo, Y., Furukawa, Y., Kato, T., Miwa, N., Nishiwaki, T.,Kawasoe, T., Ishiguro, H., Fujita, M., Tokino, T., et al. (2000). AXIN1mutations in hepatocellular carcinomas, and growth suppression incancer cells by virus-mediated transfer of AXIN1. Nat. Genet. 24,245–250.
Seoane, J., Pouponnot, C., Staller, P., Schader, M., Eilers, M., andMassague, J. (2001). TGFbeta influences Myc, Miz-1 and Smad tocontrol the CDK inhibitor p15INK4b. Nat. Cell Biol. 3, 400–408.
Sherr, C.J., and Roberts, J.M. (1999). CDK inhibitors: positive andnegative regulators of G1-phase progression. Genes Dev. 13, 1501–1512.
Shtutman, M., Zhurinsky, J., Simcha, I., Albanese, C., D’Amico, M.,Pestell, R., and Ben Ze’ev, A. (1999). The cyclin D1 gene is a targetof the beta-catenin/LEF-1 pathway. Proc. Natl. Acad. Sci. USA 96,5522–5527.
Staller, P., Peukert, K., Kiermaier, A., Seoane, J., Lukas, J., Karsunky,H., Moroy, T., Bartek, J., Massague, J., Hanel, F., and Eilers, M.(2001). Repression of p15INK4b expression by Myc through associa-tion with Miz-1. Nat. Cell Biol. 3, 392–399.
Tetsu, O., and McCormick, F. (1999). Beta-catenin regulates expres-sion of cyclin D1 in colon carcinoma cells. Nature 398, 422–426.
van de Wetering, M., Cavallo, R., Dooijes, D., van Beest, M., van Es,J., Loureiro, J., Ypma, A., Hursh, D., Jones, T., Bejsovec, A., et al.(1997). Armadillo coactivates transcription driven by the product ofthe Drosophila segment polarity gene dTCF. Cell 88, 789–799.
van Noort, M., Meeldijk, J., van der Zee, R., Destree, O., and Clevers,H. (2002). Wnt signaling controls the phosphorylation status of beta-catenin. J. Biol. Chem. 277, 17901–17905.
Wielenga, V.J., Smits, R., Korinek, V., Smit, L., Kielman, M., Fodde,R., Clevers, H., and Pals, S.T. (1999). Expression of CD44 in Apcand Tcf mutant mice implies regulation by the WNT pathway. Am.J. Pathol. 154, 515–523.
Yang, W., Velcich, A., Mariadason, J., Nicholas, C., Corner, G., Hous-ton, M., Edelmann, W., Kucherlapati, R., Holt, P.R., and Augenlicht,L.H. (2001a). p21(WAF1/cip1) is an important determinant of intesti-nal cell response to sulindac in vitro and in vivo. Cancer Res. 61,6297–6302.
Yang, W.C., Mathew, J., Velcich, A., Edelmann, W., Kucherlapati,R., Lipkin, M., Yang, K., and Augenlicht, L.H. (2001b). Targeted inacti-vation of the p21(WAF1/cip1) gene enhances Apc-initiated tumorformation and the tumor-promoting activity of a Western-style high-risk diet by altering cell maturation in the intestinal mucosal. CancerRes. 61, 565–569.
Zhang, P., Wong, C., Liu, D., Finegold, M., Harper, J.W., and Elledge,S.J. (1999). P21(CIP1) and p57(KIP2) control muscle differentiationat the myogenion step. Genes Dev. 13, 213–224.
Zhu, L., and Skoultchi, A.I. (2001). Coordinating cell proliferation anddifferentiation. Curr. Opin. Genet. Dev. 11, 91–97.

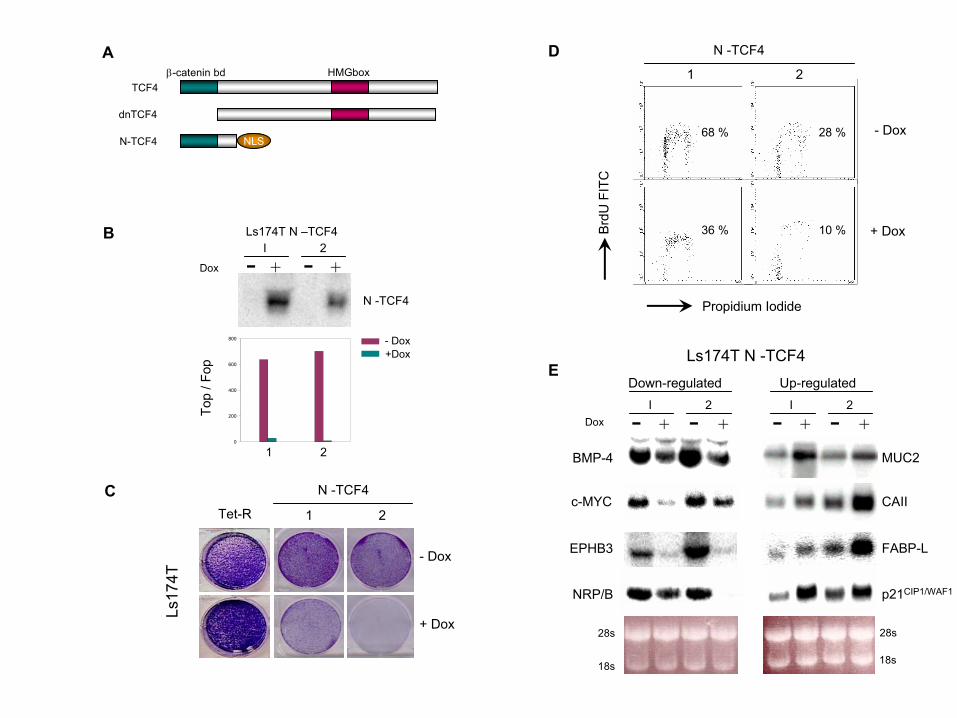
Supplementary Figure and Table Legends for van de Wetering, et al, 2002 Figure S1. Inducible N-TCF Has Similar Effects as dnTCF in Ls174T CRC Cells(A) Diagram depicting the TCF4 constructs used in this study. DnTCF4 lacks the -catenin domain. N-TCF lacks the DNA binding domain but contains the -catenin binding domain fused to a nuclear localization signal (NLS).(B–D) The induction of N-TCF (B, top) abrogates TOPFlash activity (B, bottom, purple bars) and results in cell cycle arrest (C and D). Scatter profile by FACS analysis (C) and crystal violet staining of cell cultures after 5 days of induction (D).(E) The induction of N-TCF causes comparable changes in gene expression to those induced by dnTCF by Northern blot analysis. Results for two independent clones are shown (1, 2). Figure S2. Gene Expression Changes in 24,000 cDNAs upon the Expression of dnTCF4 in Ls174T CRC Cells. Excel File containing the complete data sets of the array hybridizations. Two independent hybridizations of RNAs extracted 11 hrs and 24 hours after induction of dnTCF4 are shown. The hybridizations were performed in duplicate on chips where the same cDNAs were spotted in different order. Figure S3. Northern Blot Analysis of Genes Regulated by -Catenin/TCF Activity(A) Representative examples of several genes downregulated upon 24 hours of induction with doxycyclin in Ls174T and DLD-1 transfectants.The bottom panel shows the 28S ribosomal RNA as a loading control.(B) Abrogation of -catenin/TCF activity induced the mRNA expression of classical differentiation markers such as mucin2 (MUC2), fatty acid binding protein 2 (FABPL), carbonic anhydrase II (CAII), and alkaline phosphatase (AP) mRNAs after 24 hours of dnTCF4 induction with doxycycline. Figure S4. Expression of -Catenin/TCF Target Genes in Normal Colon and Colorectal PolypsImmunohistochemical analysis of the expression of Bmp4 (A and B), c-Myb, (C and D), and Enc1 (E and F) in normal mouse colon (A, C, and E) or colorectal polyps from min mice (B, D, and F). Target genes are highly expressed at the bottom of the normal crypts (white arrowheads) and in colorectal polyps arising at the surface epithelium (black arrowheads). Table S1. Genes Downstream of -Catenin/TCF4 in Ls174T CRC Cells. 120 entries were downregulated at least 2-fold upon the induction of dnTCF4 for 23 hours in two independent experiments. For each experiment, data at 11 hours is also shown. Table S2. Intestinal Markers of Differentiation Are Upregulated upon Abrogation of -Catenin/TCF4 Activity Selected entries of a total of 115 spots upregulated at least 2.5-fold in both hybridizations at the 23 hr time point. These molecules are expressed by differentiated cells in the intestine and most of them are downregulated in CRC (for references, see right column).
HMGboxβ-catenin bdTCF4
dnTCF4
N-TCF4 NLS
A
N -TCF4
Ls174T N –TCF4
Dox - + - +I 2
0
200
400
600
800
Top
/ Fop
- Dox+Dox
1 2
B
Tet-R
N -TCF4
Ls17
4T
- Dox
+ Dox
1 2
C
68 %
36 %
28 %
10 %
Propidium Iodide
Brd
U F
ITC
- Dox
+ Dox
1 2
N -TCF4D
BMP-4
NRP/B
CAII
FABP-L
MUC2
EPHB3
p21CIP1/WAF1
28s
18s
28s
18s
Dox - + - +
Ls174T N -TCF4
I 2
- + - +I 2
Down-regulated Up-regulatedE
c-MYC
++ +— — —dnTCF4 dnTCF1CON
DLD-1
+—++ ——DoxCON dnTCF4 dnTCF1
Ls174T
BMP4
ENC1
cMYB
++ ——DoxCON dnTCF4
Ls174T
CA II
MUC2
AP
FABP2
A B
c-MYC
28S28s
BMP4
A
B
C
D
E
F
Normal
Polyp
c-MYB ENC1
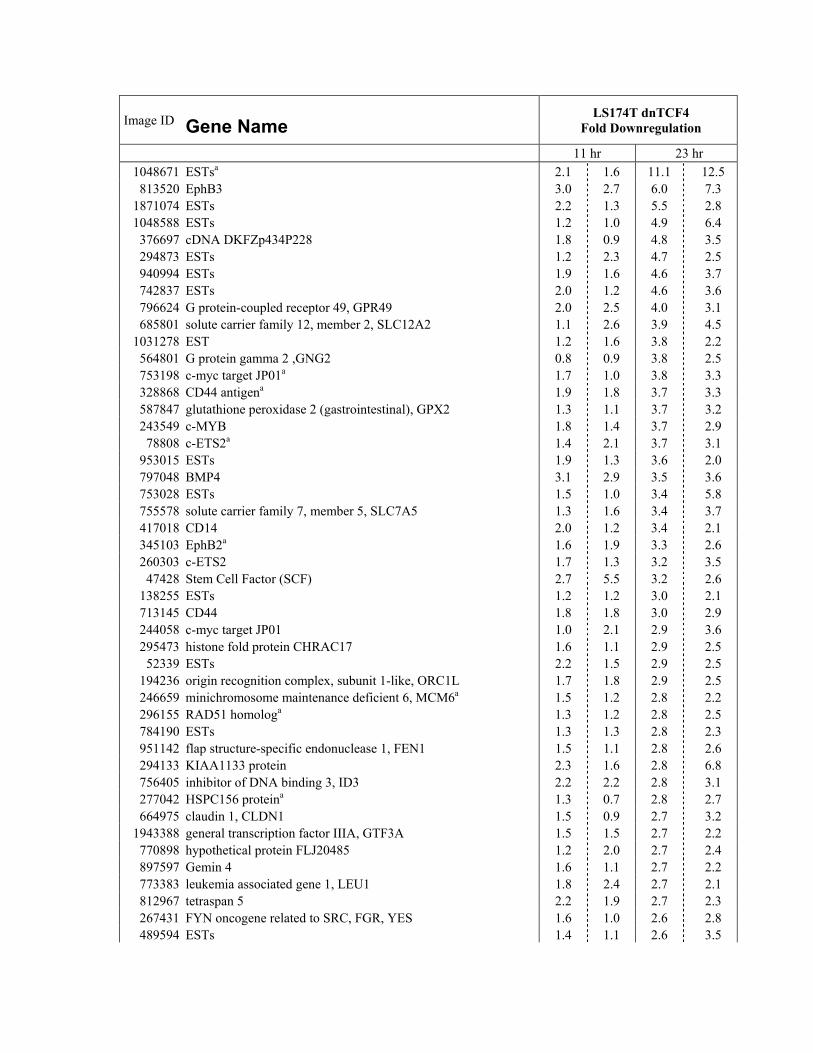
Image ID
Gene Name LS174T dnTCF4
Fold Downregulation
11 hr 23 hr 1048671 ESTsa 2.1 1.6 11.1 12.5 813520 EphB3 3.0 2.7 6.0 7.3
1871074 ESTs 2.2 1.3 5.5 2.8 1048588 ESTs 1.2 1.0 4.9 6.4 376697 cDNA DKFZp434P228 1.8 0.9 4.8 3.5 294873 ESTs 1.2 2.3 4.7 2.5 940994 ESTs 1.9 1.6 4.6 3.7 742837 ESTs 2.0 1.2 4.6 3.6 796624 G protein-coupled receptor 49, GPR49 2.0 2.5 4.0 3.1 685801 solute carrier family 12, member 2, SLC12A2 1.1 2.6 3.9 4.5
1031278 EST 1.2 1.6 3.8 2.2 564801 G protein gamma 2 ,GNG2 0.8 0.9 3.8 2.5 753198 c-myc target JP01a 1.7 1.0 3.8 3.3 328868 CD44 antigena 1.9 1.8 3.7 3.3 587847 glutathione peroxidase 2 (gastrointestinal), GPX2 1.3 1.1 3.7 3.2 243549 c-MYB 1.8 1.4 3.7 2.9 78808 c-ETS2a 1.4 2.1 3.7 3.1
953015 ESTs 1.9 1.3 3.6 2.0 797048 BMP4 3.1 2.9 3.5 3.6 753028 ESTs 1.5 1.0 3.4 5.8 755578 solute carrier family 7, member 5, SLC7A5 1.3 1.6 3.4 3.7 417018 CD14 2.0 1.2 3.4 2.1 345103 EphB2a 1.6 1.9 3.3 2.6 260303 c-ETS2 1.7 1.3 3.2 3.5 47428 Stem Cell Factor (SCF) 2.7 5.5 3.2 2.6
138255 ESTs 1.2 1.2 3.0 2.1 713145 CD44 1.8 1.8 3.0 2.9 244058 c-myc target JP01 1.0 2.1 2.9 3.6 295473 histone fold protein CHRAC17 1.6 1.1 2.9 2.5 52339 ESTs 2.2 1.5 2.9 2.5
194236 origin recognition complex, subunit 1-like, ORC1L 1.7 1.8 2.9 2.5 246659 minichromosome maintenance deficient 6, MCM6a 1.5 1.2 2.8 2.2 296155 RAD51 homologa 1.3 1.2 2.8 2.5 784190 ESTs 1.3 1.3 2.8 2.3 951142 flap structure-specific endonuclease 1, FEN1 1.5 1.1 2.8 2.6 294133 KIAA1133 protein 2.3 1.6 2.8 6.8 756405 inhibitor of DNA binding 3, ID3 2.2 2.2 2.8 3.1 277042 HSPC156 proteina 1.3 0.7 2.8 2.7 664975 claudin 1, CLDN1 1.5 0.9 2.7 3.2
1943388 general transcription factor IIIA, GTF3A 1.5 1.5 2.7 2.2 770898 hypothetical protein FLJ20485 1.2 2.0 2.7 2.4 897597 Gemin 4 1.6 1.1 2.7 2.2 773383 leukemia associated gene 1, LEU1 1.8 2.4 2.7 2.1 812967 tetraspan 5 2.2 1.9 2.7 2.3 267431 FYN oncogene related to SRC, FGR, YES 1.6 1.0 2.6 2.8 489594 ESTs 1.4 1.1 2.6 3.5
753322 NS1 associated protein 1, NSAP-1 1.4 1.1 2.5 2.0 594176 ESTs 1.2 0.8 2.5 2.2 213651 ENC1a 3.4 4.8 2.5 2.7 843224 Down syndrome critical region gene 2, DSCR2 1.4 0.9 2.5 2.4 814287 X-ray repair complementing defective repair 3, XRCC3 1.7 1.0 2.4 3.1
1883718 ESTs 2.1 1.0 2.4 2.4 358689 EphB2 variant 1.4 0.6 2.4 2.2 813280 adenylosuccinate lyase 1.5 0.9 2.4 2.3 143756 ESTs, Weakly similar to KIAA0681 protein [H.sapiens] 1.7 1.2 2.4 2.9 809557 minichromosome maintenance deficient 3, MCM3 1.5 1.1 2.4 2.6 113300 oocyte protein P100 1.2 1.5 2.4 2.4 261971 procollagen (type III) N-endopeptidase, PCOLN3 1.1 1.7 2.4 2.1 151320 ESTs 0.7 1.3 2.4 2.0 39204 cell recognition molecule Caspr2, KIAA0868a 1.4 3.1 2.4 3.3
788524 HSPC156 protein 1.0 0.5 2.4 2.6 897570 heat shock protein 75, TRAP1 1.2 0.8 2.4 2.3 49464 uracil-DNA glycosylase, UNG 1.8 1.2 2.4 2.2
384851 clathrin, heavy polypeptide-like 1 1.0 1.2 2.3 4.3 26616 replication protein A2 (32kD) 1.5 1.1 2.3 2.3
273546 similar to SAICAR synthetase and AIR carboxylase 1.4 1.1 2.3 2.6 700721 minichromosome maintenance deficient 5, MCM5 1.3 1.4 2.3 2.9
1030779 ESTs 1.1 1.3 2.3 2.3 346696 Transcriptional enhancer factor TEF3 1.3 1.6 2.3 2.1 768258 ESTs 1.9 1.6 2.3 2.1 303049 ESTs 1.6 0.8 2.3 2.1 815535 Treacher Collins-Franceschetti syndrome 1, TCOF1a 1.3 2.3 2.3 2.8 665398 Ran binding protein 11 1.5 1.7 2.3 2.1 767068 DKFZP586G1517 protein 1.5 1.9 2.3 2.4 625424 HSPC132 1.7 1.0 2.3 2.2
1861180 Treacher Collins-Franceschetti syndrome 1, TCOF1 1.4 0.9 2.3 2.1 731343 heterogeneous nuclear ribonucleoprotein A1 1.3 1.1 2.2 3.3 510856 ENC1 2.6 3.3 2.2 2.4 855390 minichromosome maintenance deficient 6,MCM6 1.2 2.0 2.2 2.5 430186 cDNA DKFZp434B0425 1.5 1.4 2.2 2.4 259462 hypothetical protein loc54675 1.4 1.1 2.2 2.1 53122 Human (clone CTG-A4) mRNA sequence 2.0 4.2 2.2 2.8
366009 hypothetical protein FLJ20425 1.5 0.9 2.2 2.2 416945 cDNA DKFZp564F093 1.4 1.0 2.2 2.5 244754 PRO0650 protein 1.2 1.1 2.2 2.2 417226 MYC 2.1 1.6 2.2 2.1 30574 KIAA0759 protein 1.4 2.0 2.2 2.1
595697 hypothetical protein FLJ20315 2.1 1.3 2.2 3.0 470232 ESTs 1.8 1.4 2.2 3.7 197657 aldehyde dehydrogenase 5 1.6 1.1 2.2 2.8 450711 chromatin assembly factor 1, subunit A (p150) 1.5 1.0 2.2 2.5 292936 hypothetical protein FLJ10468 1.1 1.2 2.1 2.2 195813 60S acidic ribosomal protein PO 1.8 1.4 2.1 2.9 810104 phospholipase C, beta 2 1.2 1.0 2.1 2.2 768172 dihydrofolate reductase 1.2 0.9 2.1 2.1 742806 transcription factor Dp-2 (E2F dimerization partner 2) 1.2 0.8 2.1 3.0
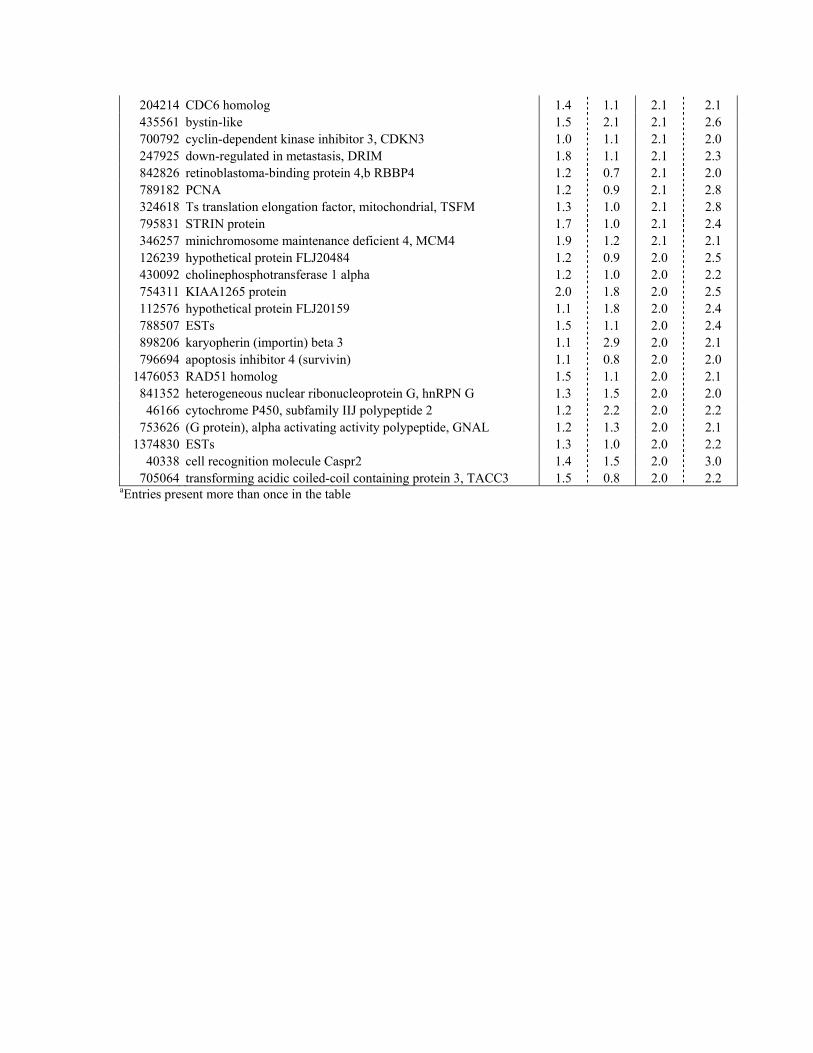
204214 CDC6 homolog 1.4 1.1 2.1 2.1 435561 bystin-like 1.5 2.1 2.1 2.6 700792 cyclin-dependent kinase inhibitor 3, CDKN3 1.0 1.1 2.1 2.0 247925 down-regulated in metastasis, DRIM 1.8 1.1 2.1 2.3 842826 retinoblastoma-binding protein 4,b RBBP4 1.2 0.7 2.1 2.0 789182 PCNA 1.2 0.9 2.1 2.8 324618 Ts translation elongation factor, mitochondrial, TSFM 1.3 1.0 2.1 2.8 795831 STRIN protein 1.7 1.0 2.1 2.4 346257 minichromosome maintenance deficient 4, MCM4 1.9 1.2 2.1 2.1 126239 hypothetical protein FLJ20484 1.2 0.9 2.0 2.5 430092 cholinephosphotransferase 1 alpha 1.2 1.0 2.0 2.2 754311 KIAA1265 protein 2.0 1.8 2.0 2.5 112576 hypothetical protein FLJ20159 1.1 1.8 2.0 2.4 788507 ESTs 1.5 1.1 2.0 2.4 898206 karyopherin (importin) beta 3 1.1 2.9 2.0 2.1 796694 apoptosis inhibitor 4 (survivin) 1.1 0.8 2.0 2.0
1476053 RAD51 homolog 1.5 1.1 2.0 2.1 841352 heterogeneous nuclear ribonucleoprotein G, hnRPN G 1.3 1.5 2.0 2.0 46166 cytochrome P450, subfamily IIJ polypeptide 2 1.2 2.2 2.0 2.2
753626 (G protein), alpha activating activity polypeptide, GNAL 1.2 1.3 2.0 2.1 1374830 ESTs 1.3 1.0 2.0 2.2
40338 cell recognition molecule Caspr2 1.4 1.5 2.0 3.0 705064 transforming acidic coiled-coil containing protein 3, TACC3 1.5 0.8 2.0 2.2
aEntries present more than once in the table
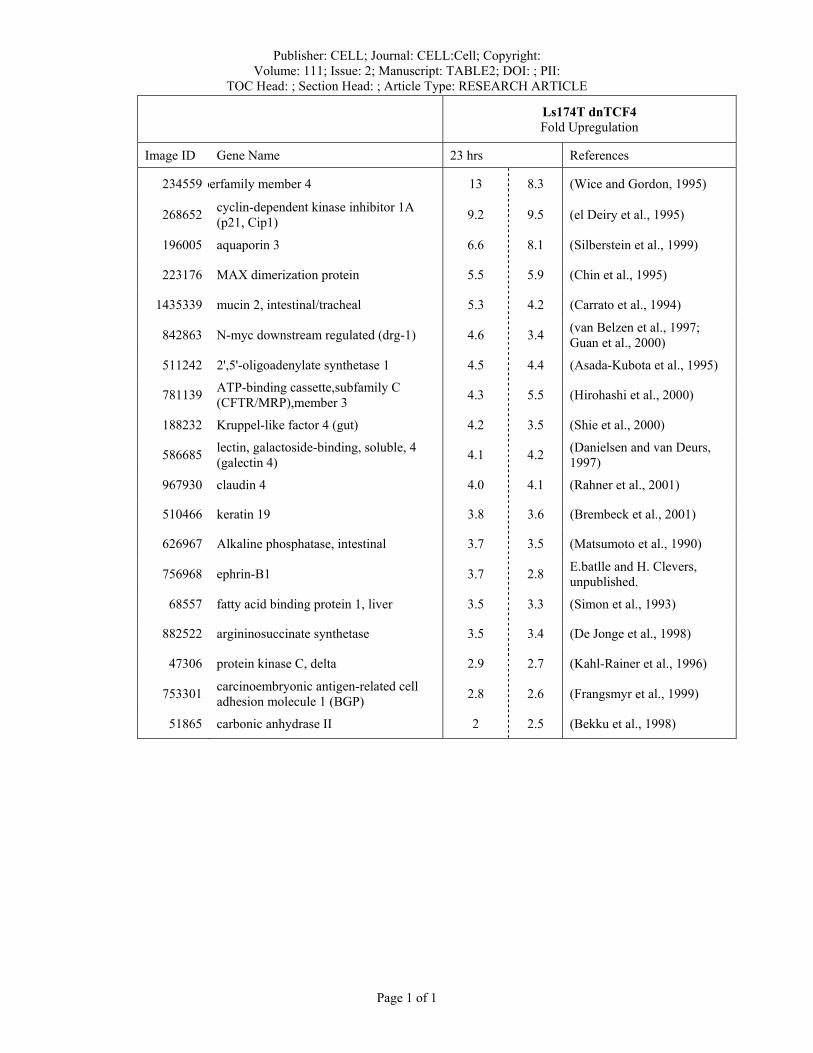
Publisher: CELL; Journal: CELL:Cell; Copyright: Volume: 111; Issue: 2; Manuscript: TABLE2; DOI: ; PII:
TOC Head: ; Section Head: ; Article Type: RESEARCH ARTICLE
Page 1 of 1
Ls174T dnTCF4 Fold Upregulation
Image ID Gene Name 23 hrs References
234559 perfamily member 4 13 8.3 (Wice and Gordon, 1995)
268652 cyclin-dependent kinase inhibitor 1A (p21, Cip1) 9.2 9.5 (el Deiry et al., 1995)
196005 aquaporin 3 6.6 8.1 (Silberstein et al., 1999)
223176 MAX dimerization protein 5.5 5.9 (Chin et al., 1995)
1435339 mucin 2, intestinal/tracheal 5.3 4.2 (Carrato et al., 1994)
842863 N-myc downstream regulated (drg-1) 4.6 3.4 (van Belzen et al., 1997; Guan et al., 2000)
511242 2',5'-oligoadenylate synthetase 1 4.5 4.4 (Asada-Kubota et al., 1995)
781139 ATP-binding cassette,subfamily C (CFTR/MRP),member 3 4.3 5.5 (Hirohashi et al., 2000)
188232 Kruppel-like factor 4 (gut) 4.2 3.5 (Shie et al., 2000)
586685 lectin, galactoside-binding, soluble, 4 (galectin 4) 4.1 4.2 (Danielsen and van Deurs,
1997)
967930 claudin 4 4.0 4.1 (Rahner et al., 2001)
510466 keratin 19 3.8 3.6 (Brembeck et al., 2001)
626967 Alkaline phosphatase, intestinal 3.7 3.5 (Matsumoto et al., 1990)
756968 ephrin-B1 3.7 2.8 E.batlle and H. Clevers, unpublished.
68557 fatty acid binding protein 1, liver 3.5 3.3 (Simon et al., 1993)
882522 argininosuccinate synthetase 3.5 3.4 (De Jonge et al., 1998)
47306 protein kinase C, delta 2.9 2.7 (Kahl-Rainer et al., 1996)
753301 carcinoembryonic antigen-related cell adhesion molecule 1 (BGP) 2.8 2.6 (Frangsmyr et al., 1999)
51865 carbonic anhydrase II 2 2.5 (Bekku et al., 1998)
Copyright © 2022 FDOKUMEN




















