
Chronic North American Ginseng Administration Alters ...
141
Western University Western University Scholarship@Western Scholarship@Western Electronic Thesis and Dissertation Repository 7-30-2012 12:00 AM Chronic North American Ginseng Administration Alters Metabolic Chronic North American Ginseng Administration Alters Metabolic and Cardiovascular Variables in the Rat and Cardiovascular Variables in the Rat Megan J. Migchels, The University of Western Ontario A thesis submitted in partial fulfillment of the requirements for the Master of Science degree in Physiology © Megan J. Migchels 2012 Follow this and additional works at: https://ir.lib.uwo.ca/etd Recommended Citation Recommended Citation Migchels, Megan J., "Chronic North American Ginseng Administration Alters Metabolic and Cardiovascular Variables in the Rat" (2012). Electronic Thesis and Dissertation Repository. 792. https://ir.lib.uwo.ca/etd/792 This Dissertation/Thesis is brought to you for free and open access by Scholarship@Western. It has been accepted for inclusion in Electronic Thesis and Dissertation Repository by an authorized administrator of Scholarship@Western. For more information, please contact [email protected].
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of Chronic North American Ginseng Administration Alters ...

Western University Western University
Scholarship@Western Scholarship@Western
Electronic Thesis and Dissertation Repository
7-30-2012 12:00 AM
Chronic North American Ginseng Administration Alters Metabolic Chronic North American Ginseng Administration Alters Metabolic
and Cardiovascular Variables in the Rat and Cardiovascular Variables in the Rat
Megan J. Migchels, The University of Western Ontario
A thesis submitted in partial fulfillment of the requirements for the Master of Science degree in
Physiology
© Megan J. Migchels 2012
Follow this and additional works at: https://ir.lib.uwo.ca/etd
Recommended Citation Recommended Citation Migchels, Megan J., "Chronic North American Ginseng Administration Alters Metabolic and Cardiovascular Variables in the Rat" (2012). Electronic Thesis and Dissertation Repository. 792. https://ir.lib.uwo.ca/etd/792
This Dissertation/Thesis is brought to you for free and open access by Scholarship@Western. It has been accepted for inclusion in Electronic Thesis and Dissertation Repository by an authorized administrator of Scholarship@Western. For more information, please contact [email protected].

i
Chronic North American Ginseng Administration Alters
Metabolic and Cardiovascular Variables in the Rat
(Spine title: Ginseng and Metabolism)
(Thesis format: Monograph)
by
Megan J. Migchels
Graduate Program in Physiology
A thesis submitted in partial fulfillment
of the requirements for the degree of
Master of Science
The School of Graduate and Postdoctoral Studies
The University of Western Ontario
London, Ontario, Canada
© Megan J. Migchels 2012

ii
THE UNIVERSITY OF WESTERN ONTARIO
SCHOOL OF GRADUATE AND POSTDOCTORAL STUDIES
CERTIFICATE OF EXAMINATION
Supervisor Examiners
___________________________________ _______________________________________
Dr. John Ciriello Dr. Doug Jones
Supervisory Committee _______________________________________
Dr. Nica Borradaile
_____________________________________ _______________________________________
Dr. Ed Lui Dr. Paul A. Charpentier
_____________________________________
Dr. Morris Karmazyn
_______________________________________
Dr. Subrata Chakrabarti
_______________________________________
Dr. Stan Leung
The thesis by
Megan J. Migchels
Entitled:
Chronic North American Ginseng Administration Alters
Metabolic and Cardiovascular Variables in the Rat
is accepted in partial fulfillment of the requirements for the degree of
Master of Science
_____________________________________ _______________________________________ Date Chair of the Thesis Examination Board

iii
Abstract and Keywords
Although ginseng has been used for centuries as a medicinal herb, little is known about the
effects of North American ginseng (NAG) on the metabolic syndrome. This study was done to
investigate the effects of NAG on metabolic variables, and whether central neuronal systems
involved in the control of these metabolic variables were altered by NAG treatment. Experiments
were done in adult male Sprague-Dawley rats fed a standard chow or a high fat (45 %kcal) diet.
Animals were given an alcoholic extract of NAG (250 mg/kg body weight) or vehicle (0.9%
saline) daily for 4 weeks by oral gavage. Food and water intake were found not to be altered by
ginseng. However, body weight, cumulative body weight gain, and fat pad mass were
significantly lower in the NAG treated animals compared to the vehicle treated group regardless
of diet. Similarly, NAG treatment reduced plasma leptin, insulin, triglycerides, and noradrenaline
levels in all animal groups. Furthermore, NAG lowered arterial pressure and heart rate in animals
fed a high fat diet. Within the central nervous system, NAG was found to induce the expression
of the immediate early genes Fra-1/2 in brainstem cardiovascular regulatory centers (nucleus of
the solitary tract, and caudal and rostral ventrolateral medulla), and in several hypothalamic
nuclei involved in the control of the circulation and metabolism (paraventricular, arcuate, lateral,
dorsomedial and ventromedial hypothalamic nuclei). Taken together, these data suggest that
central brainstem and hypothalamic circuits involved in autonomic control and ingestive
behaviour are altered by NAG, and suggest that the use of NAG can affect metabolic and
cardiovascular variables.
Keywords: North American ginseng, Panax quinquefolius, metabolic syndrome, blood pressure,
Fos-related antigens

iv
Acknowledgements
“Knowledge is in the end based on acknowledgement.”- Ludwig Wittgenstein
“If we knew what it was we were doing, it would not be called research, would it?”- Albert
Einstein
I would like to thank my supervisor Dr. John Ciriello for the opportunity to work in his
lab. In addition, I would like to thank all of the members of the lab for their support, input,
and friendship throughout my studies. I wish you the best of luck in your future endeavors.
To my family and friends, thank you for believing in me when I didn’t believe in myself.
Words cannot express how grateful I am.
"Sometimes our light goes out but is blown into flame by another human being. Each of us owes
deepest thanks to those who have rekindled this light." - Albert Schweitzer
“I feel a very unusual sensation - if it is not indigestion, I think it must be gratitude.” - Benjamin
Disraeli

v
Table of Contents
Certificate of examination..…………..………………..……………..………………………...…ii
Abstract and keywords…………..……………..……………..……………..…………………...iii
Acknowledgements…………..……………..……………..……………..……………….…...….iv
List of Figures……………………………………………………………………..........……….viii
List of Tables…………………………………...……………………………………………...….x
List of Abbreviations, Symbols, Nomenclature…………...…………..……...……...…..……….xi
Chapter 1: Introduction and Literature Review……………………………………………….1
1.1 Introduction…………..…………..…………..…………..…………..……………..…………1
1.2 Obesity…………..…………..…………..…………..…………..…………..……………..….2
1.2.1 Consequences of Obesity…………..…………..…………..…………..……...….…3
1.2.2 Leptin…………..…...…………..…………..…………..…………..…………...…..4
1.2.2.1 Leptin Resistance…………..…………..…………..…………..……...…..5
1.3 Metabolic syndrome…………..…………..…………..…………..……………………...……5
1.3.1 Insulin…………..…………..…………..…………..…………..…………………...6
1.3.1.1 Insulin resistance…………..…………..……………….………………….8
1.3.2 Dyslipidemia…………..…………..…………..…………..…….………..…………9
1.4 Arterial pressure…………..…………..…………..…………..…………..…..…………..…...9
1.4.1 Baroreceptor Reflex………..…………..…………………..…..………..…………10
1.4.2 Hypertension…………..…………..…………..…………..…………….…………12
1.4.3 Obesity-associated Hypertension…………..…………..…………..………………13
1.5 Ginseng…………..…………..…………..…………..…………..…………..…………...….14
1.5.1 Ginsenosides…………..…………..…………..…………..…………..……...……16
1.5.2 Types of Ginseng…………..…………..…………..…………..…………..………17
1.5.3 Ginseng Metabolism…………..…………..…………..…………..…..…….……..20
1.5.4 The Effect of Ginseng in the Central Nervous System……………………………22
1.6 Metabolic and Cardiovascular effects of Ginseng…………..………………………....……23
1.6.1 Effects of Asian ginseng on Metabolic Variables…………..…………………..…23
1.6.1.1 Asian Ginseng has anti-diabetic effects…………..………...………..….23
1.6.1.2 Asian Ginseng has anti-obesity effects…………..…………..….………25
1.6.1.3 Potential mechanisms responsible for the metabolic effects
of Asian ginseng…………..…………..………..……………..….………….….26

vi
1.6.1.4 The Metabolic effects of Korean Red ginseng………………………….28
1.6.1.5 The potential mechanisms for the effects of Korean Red ginseng……..29
1.6.1.6 The anti-hyperglycemic effects of Ginsenoside Rg1……….…………..30
1.6.2 The Effect of Asian ginseng on Cardiovascular Variables………………..………31
1.6.2.1 The Effect of Asian ginseng on Arterial Pressure……………….………31
1.6.2.2 Potential Anti-hypertensive Mechanisms of Asian Ginseng……….....…32
1.6.2.3 Korean Red ginseng has anti-hypertensive effects……………….…...…33
1.6.2.4 The Potential Anti-hypertensive Mechanisms of Korean Red Ginseng…33
1.6.3 The Effect of NAG on Metabolic Variables…………..………….……..…………33
1.6.3.1 NAG has Anti-Diabetic Properties…………..…………..……..……..…34
1.6.3.2 The Anti-Obesity Effects of NAG…………..…………..……………….35
1.6.3.3 Potential Mechanisms responsible for the metabolic effects of NAG…..35
1.6.3.4 The Metabolic effects of Ginsenoside Rb1…..…………..………………37
1.6.3.5 The Metabolic Effects of Ginsenoside Re…..…………..………….……38
1.6.3.6 The Potential Mechanisms for the Effects of Ginsenoside Re…....……..39
1.6.3.7 The Metabolic Effects of Compound K……..………..……………..…...40
1.6.4 The Effects of NAG on Cardiovascular Variables……..…………..…………….……41
1.6.4.1 The Effects of NAG on Arterial Pressure……..…………..…………..…41
1.6.4.2 Potential Mechanisms Responsible for the Cardiovascular
Effects of NAG….………..………..………..………..…………………..….…42
1.7 Rationale and Hypothesis…………..…………..…………..…………..…………..……..…42
Chapter 2: Materials and Methods…………..…………..…………..……..…………....……45
2.1 Ginseng Production…………..…………..…………..…………..…………..………45
2.2 Animal Care…………..…………..…………..…………..…………..…………..….46
2.3 Treatment…………..…………..…………..…………..…………..………………...46
2.3.1 Cardiovascular measurements…………..…………..………..………..…..46
2.3.2 Metabolism study…………..…………..…………..…………..…………..47
2.4 Tissue Collection…………..…………..…………..…………..…………..………...47
2.5 Biochemical assays…………..…………..…………..…………..…………..………48
2.5.1 Plasma Leptin Quantitation…………..…………..…………..………..…..48
2.5.2 Plasma Insulin Quantitation……………………..…………………………48
2.5.3 Plasma Adiponectin Quantitation…………..…………..……….………...48
2.5.4 Plasma Triglycerides and Total Cholesterol Quantitation…………………49

vii
2.5.5 Plasma Noradrenaline Quantitation……………………………..…………49
2.5.6 Plasma Angiotensin II Quantitation………………………………………..49
2.6 Immunohistochemistry…………..…………..…………..…………..…………....…49
2.7 Data Analysis…………..…………..…………..…………..…………..………….....51
Chapter 3: Results……..…………..…………..…….……..…………..…………..………..…53
3.1 The effects of NAG on markers of the metabolic syndrome…....…..………..….…53
3.1.1 The effect of NAG treatment on Body Weight, Caloric Intake,
and Urine output..............................................................................................…53
3.1.2 The effect of NAG treatment on lipids………...…….….…..………….....56
3.1.3 The effect of NAG on glucose homeostasis…….............……………....…60
3.1.4 The effect of NAG treatment on MAP and HR……..…………….…….…60
3.1.5 The effect of NAG treatment on plasma Noradrenaline and
Angiotensin II levels……………………………………………………………..65
3.2 Central Effects of NAG……..…………..…………..………..…………..…..…...…68
3.2.1 The effect of NAG on brainstem regions……..………………..…….……68
3.2.1.1 Fra-1/2 Expression in brainstem autonomic nuclei…....…...……68
3.2.1.2 Fra-1/2 Immunoreactivity in TH-Labeled neurons……..…….…73
3.2.2 The effect of NAG on neuronal activity in forebrain structures…..…..…..73
3.2.2.1 Fra-1/2 Expression in forebrain circumventricular organs.…..…73
3.2.2.2 Fra-1/2 Expression in hypothalamic nuclei……..…………….…85
Chapter 4: Discussion……..…………..…………..…………..………..…………..………..…92
4.1 The effect of NAG on markers of the metabolic syndrome...………..………………92
4.2 The effect of NAG on AP and HR ……..….…...………..………..…………………94
4.3 Effects of NAG on Brainstem Autonomic Nuclei……...…..…………..…..……..…96
4.3.1 Fra-1/2 Immunoreactivity in TH Labeled Neurons.....………....………….97
4.4 The effect of NAG on neuronal activity in the forebrain………..……..…….....….101
4.4.1 Fra-1/2 Immunoreactivity in the circumventricular organs………..….....101
4.4.2 Fra-1/2 Immunoreactivity in hypothalamic nuclei……….…….…....…...101
4.5 Overall Summary and Conclusion…....……………..…...……………...…….…..102
4.6 Future directions…………..…………………………….…………………..……..103
References…..…………….…………..…..…………..…………..…..…….……..………..…..104
Curriculum Vitae…..…………..…………..…..…………..………..…..……...……..…..……122

viii
List of Figures
Figure Title Page
1.1 Leptin and Insulin signalling in the arcuate nucleus of the hypothalamus…....…7
1.2 The major pathways involved in the baroreceptor reflex…..…………..…..…...11
1.3 Fresh Ginseng shaped like a man…..…………..……..…………..……..………15
1.4 Characteristic ginsenosides present in both Asian and North American
ginseng grouped into protopanaxadiol and protopanaxtriol….………………….18
3.1 The effect of North American ginseng (NAG) on body weight after
28 days of treatment…………………………………………………………..…54
3.2 The effect of North American ginseng (NAG) treatment on cumulative
change in body weight…………………………………………………………...55
3.3 The effects of North American Ginseng (NAG) on fat pad mass……………..…58
3.4 The effect of North American Ginseng (NAG) on plasma leptin and
adiponectin levels….………………………………….………………………….59
3.5 The effect of North American Ginseng (NAG) on plasma lipids ……..………...61
3.6 The effect of North American Ginseng (NAG) on glucose homeostasis…..……62
3.7 The effect of North American Ginseng (NAG) on mean arterial
pressure and heart rate after 28 days of treatment………………………...……..64
3.8 The effect of NAG on systolic and diastolic blood pressure after 28 days of
treatment………………………………………………………………………....66
3.9 The effect of NAG on plasma Noradrenaline and Angiotensin II levels……......67
3.10 Bright-field photomicrographs taken through the region of the caudal
Nucleus of the Solitary Tract (NTS) complex showing the distribution
of Fra-1/2 labeled neurons throughout the complex………………………...…..70
3.11 Bright-field photomicrographs taken through the caudal ventrolateral
medulla (CVLM)………………………………………………………..….……71
3.12 Bright-field photomicrographs taken through the rostral ventrolateral
medulla (RVLM) showing the distribution of Fra-1/2 immunoreactive
neurons…………………………………………………………………………..72
3.13 Bright-field photomicrographs showing examples of neurons in the A2
region of the Nucleus of the Solitary Tract (NTS) complex, expressing
both Fra-1/2 and TH immunoreactivity……………………..…………….……..74
3.14a Camera lucida projection drawings of the Nucleus of the Solitary
Tract (NTS) in animals fed a normal chow diet..…………..……………………75

ix
3.14b Camera lucida projection drawings of the Nucleus of the Solitary
Tract (NTS) in animals fed a high fat diet…………………….…………………76
3.15 Bright-field photomicrographs showing examples of neurons A1 region
of the caudal ventrolateral medulla (CVLM) expressing both
Fra-1/2 and TH immunoreactivity ……………………………………….……...77
3.16 Bright-field photomicrographs showing examples of neurons in the C1
region of the rostral ventrolateral medulla (RVLM) expressing both Fra-1/2
and TH immunoreactivity…………….………………………………………….78
3.17 Camera lucida projection drawings of the Ventrolateral Medulla……………….79
3.18 Bright-field photomicrograph showing examples of neurons in the A5 region
of the expressing Fra-1/2 and TH immunoreactivity………………………...…..80
3.19 Bright-field photomicrographs taken through the subfornical organ (SFO)..…..83
3.20 Bright-field photomicrographs taken through the organum vasculosum of the
lamina terminalis (OVLT) showing Fra-1/2 labeled neurons……………..…….84
3.21 Bright-field photomicrographs taken through the median preoptic nucleus
(MnPO)………………………………………………………………..…………86
3.22 Bright-field photomicrographs expressing Fra-1/2 immunoreactivity taken
through the arcuate nucleus (ARC)……………………………………...……….87
3.23 Bright-field photomicrographs taken through the paraventricular nucleus
of the hypothalamus (PVH) showing the distribution of Fra-1/2 labeled
neurons ………………………………………………….….……………….…...88
3.24 Bright-field photomicrographs taken through the supraoptic nucleus
(SON) showing the distribution of Fra-1/2 labeled neurons………………..….89
3.25 Bright-field photomicrographs taken through the anterior hypothalamic area
(AHA) showing the distribution of Fra-1/2 labeled neurons………………….....90

x
List of Tables
Table Title Page
3.1 The effects of North American ginseng (NAG) treatment on
food and water intake and urine output ……….……………………….……….57
3.2 The effect of North American ginseng (NAG) on organ weights after
28 days of treatment…………………………………………………………..…57
3.3 The effect of North American Ginseng (NAG) on mean arterial
pressure (MAP) throughout the treatment period ……….……………….…..…63
3.4 Table summarizing the number of neurons expressing Fra-1/2 immunoreactivity
in different brainstem regions…………………………………………………...69
3.5 Table summarizing the number of neurons expressing Fra-1/2 immunoreactivity
in addition to labeling with Tyrosine Hydroxylase (TH)……………………..…81
3.6 Table summarizing the number of neurons expressing Fra-1/2 immunoreactivity
in the circumventricular organs………………………………………………….82
3.7 Table summarizing the number of neurons expressing Fra-Li immunoreactivity in
hypothalamic regions…..…………………………………………...……………92

xi
List of Abbreviations, Symbols, Nomenclature
3V Third ventricle
12M hypoglossal nucleus
12N root of hypoglossal nerve
µm micrometre
°C Degrees Celsius
α-MSH alpha-Melanocyte-stimulating hormone
A1 A1 noradrenergic cell group
A2 A2 noradrenergic cell group
a3V anterior third ventricle
A5 A5 noradrenergic cell group
ABC avidin-biotin complex
ac anterior commissure
acetyl-CoA acetyl coenzyme-A
AgRP Agouti-related peptide
AHA Anterior hypothalamic area
AHPA American Herbal Products Association
Akt Protein Kinase B
AMB nucleus ambiguous
AMPK AMP-activated protein kinase
ANOVA analysis of variance
ap area postrema
AP Arterial pressure
ARC arcuate nucleus
ATP adenosine triphosphate
β-cell Beta cell
BMI Body Mass Index
bpm beats per minute
C1 C1 adrenergic cell group
cAMP cyclic adenosine monophosphate
CART Cocaine and amphetamine regulated transcript
cals calories
C/EBPs CCAAT-enhancer-binding proteins
CNS central nervous system
com commissural subnucleus of NTS
CVLM caudal ventrolateral medulla
CVs coefficients of variation
DAB Diaminobenzidine tetrahydrochloride
DBP Diastolic blood pressure
DMD Dorsomedial hypothalamus
DMV dorsal motor nucleus of the vagus
dp dorsal parvocellular nucleus
EIA Enzyme Immunoassay
ELISA enzyme-linked immunosorbent assay
eNOS endothelial nitric oxide synthase

xii
FAS fatty acid synthase
Fra-1/2 Fos-related antigens 1/2
Fx fornix
g gram
g/mm gram per milimeter
G6Pase glucose-6-phosphatase
GLUT1 Glucose Transporter Type 1
GLUT4 Glucose Transporter Type 4
G-Ra ginsenoside Ra
G-Rb1 ginsenoside Rb1
G-Rb2 ginsenoside Rb2
G-Rb3 ginsenoside Rb3
G-Rc ginsenoside Rc
G-Rd ginsenoside Rd
G-Re ginsenoside Re
G-Rf ginsenoside Rf
G-Rg1 ginsenoside Rg1
G-Rg3 ginsenoside Rg3
G-Rh2 ginsenoside Rh2
h hours
H2O2 Hydrogen peroxide
HDL high density lipoprotein
HIV human immunodeficiency virus
HFC high fat diet with vehicle control
HFD high fat diet
HF-NAG high fat diet with treatment of an alcoholic NAG extract
HMG-CoA β-hydroxy-β-methylglutaryl-CoA
HOMA-IR Homeostatic Model of Insulin Resistance
HPLC high performance liquid chromatography
HR heart rate
ICAM-1 intercellular adhesion molecule 1
IGF-1 Insulin-like growth factor 1
IML intermediolateral cell column of the spinal cord
ION inferior olivary nucleus
IR Insulin Receptor
IRS-1 insulin receptor substrate 1
i.p. intraperitoneal
kg kilogram
L litre
LDL Low-density lipoprotein
LPGi nucleus paragiantocellularis
LPL Lipoprotein lipase
LH lateral hypothalamus
LRN Lateral reticular nucleus
LSO lateral superior olive
MAP mean arterial pressure

xiii
MARN magnocellular reticular nucleus
MC4R Melanocortin 4 receptor
MdV Medullary reticular nucleus
ME median eminence
mg milligram
mg/kg milligram per kilogram
mins minutes
ml millilitre
mmHg millimeters of mercury
mmol/L millimol per litre
MnPO median preoptic nucleus
mp medial parvocellular nucleus
mRNA messenger ribonucleic acid
NAG North American Ginseng
Nc Nucleus circularis
ND normal diet
NDC normal diet with vehicle control treatment
ND-NAG normal diet with a treatment of an alcoholic extract of NAG
ng/ml nanogram per mililitre
NO nitric oxide
NPY Neuropeptide Y
NTS nucleus of the solitary tract
NF-κB nuclear factor kappa-light-chain-enhancer of activated B cells
oc optic chiasm
OGIRC Ontario Ginseng Innovation and Research Consortium
OGGA Ontario Ginseng Growers Association
OVLT organum vasculosum of the lamina terminalis
PBS Phosphate buffered saline
pg/ml picograms per millilitre
PI3K Phosphatidylinositol 3-kinase
PKA protein kinase A
pm posterior magnocellular nucleus
POMC Pro-opiomelanocortin
PPARα peroxisome proliferator-activated alpha
PPARɣ peroxisome proliferator-activated receptor gamma
PPD protopanaxadiol ginsenosides
PPT protopanaxatriol ginsenosides
PVH paraventricular nucleus of the hypothalamus
Py pyramidal tract
ROS reactive oxygen species
RPa raphé pallidus
rs rubrospinal tract
RVLM rostral ventrolateral medulla
RXR retinoid X receptor
SBP Systolic blood pressure
Sdl dorsolateral subnucleus of NTS

xiv
SEM Standard error of the mean
SFO subfornical organ
Sg commissural subnucleus of NTS
Slt solitary tract
Sm medial subnucleus of NTS
SNS Sympathetic nervous system
SON Supraoptic nucleus
SREBP-1c Sterol regulatory element-binding protein
TCM Traditional Chinese Medicine
TH Tyrosine Hydroxylase
t-1/2 Half life
UCP2 Uncoupling protein 2
UCP3 uncoupling protein 3
vhc ventral hippocampal commissure
VLDL Very low-density lipoprotein
VMH ventromedial hypothalamus
vp ventral parvocellular nucleus

1
Chapter 1: Introduction and Literature Review
1.1 Introduction
Obesity is a serious medical condition that occurs when excess fat has accumulated to the
point that it exerts an adverse effect on health, ultimately leading to increased morbidity and
mortality (Ogden et al., 2007). In Canada, approximately 53% of women and 68% of men are
overweight or obese (Statistics Canada, 2010). These numbers have increased dramatically over
the past 30 years and mirror the onset and rise of the worldwide obesity epidemic (Finucane et
al., 2011). The pathophysiology associated with obesity is arguably the leading cause of
preventable death in our society (Statistics Canada, 2010).
Numerous strategies have been proposed to treat obesity, including diet therapy, exercise,
and medications that modulate appetite and control lipid digestion. However, conventional drug
therapies have numerous limitations, including contraindications and secondary failures (Sheen,
2003; Nathan et al., 2006). Currently, no medication can provide a cure for obesity: experimental
evidence confirms the simple idea that once the anorexigenic regime is terminated, the previous
weight loss will be followed by a rapid return to initial weight (Bray and Greenway, 1999).
Interestingly, several medicinal herbs are believed to exert positive effects on
components of the metabolic syndrome, such as obesity, blood pressure, hyperlipidemia, and
blood glucose levels with suggested minimal side effects (Cabrera et al., 2006; Dey et al., 2002;
Moreno et al., 2003). In recent years, our health care system has seen a rapid rise in the use of
complementary and alternative medical therapies, that include herbal medicines such as ginseng
(Stavro et al., 2006). The adaptogenic effects of ginseng have been passed down through oral
tradition for centuries. Although the last 50 years has added considerable progress to the modern
study of Asian ginseng, little is known about the cardiovascular and metabolic effects of North

2
American ginseng (NAG), and whether NAG can exert an effect on central neuronal circuitry
involved in the control of these cardiovascular and metabolic variables.
1.2 Obesity
When energy intake exceeds expenditure, the excess energy is stored in adipose tissue.
Consequently, the prolongation of this process results in obesity. It is the increase in adipocyte
cell number (hyperplasia) and/or cell size (hypertrophy) that yields the pathologic lesion seen in
obesity (Bray et al., 1998). An adipocyte is now regarded as an endocrine cell as it has been
shown to produce a large number of peptides, hormones, and small molecules that play a role in
metabolism and cardiovascular regulation (Achike et al., 2011; Zhang et al., 1994).
The pathogenesis of obesity is both complex and multifactorial and is thought to be
related to a gene-environment interaction: that is, susceptible individuals respond to an
environmental scenario wherein there is an increased intake of palatable foods and a decreased
opportunity for energy expenditure (Ogden et al., 2007). The balance between energy intake and
expenditure is influenced by a complex interaction of genetic, environmental, and social factors
(Ogden et al., 2007). However, ultimately, obesity is regulated by the hunger and satiety centers
of the hypothalamus through complex mechanisms that are modulated by both central and
peripheral hormones and cytokines (Achike et al., 2011).
Obesity is typically defined using the Body Mass Index (BMI), an expression of weight
adjusted for height. BMI is calculated by dividing an individual’s weight by the square of their
height [BMI= weight (kg) ÷ height2
(m2)]. Obesity is defined as a BMI ≥ 30, whereas an
overweight classification includes values from 25 to 29.9. Finally, a BMI between 18.5 and 24.9
is indicative of a healthy weight (Ogden et al., 2007). However, it is important to note that the

3
BMI is not an accurate measure of obesity across all populations: BMI tends to overestimate
obesity in muscular people and underestimate in people who have lost body weight (i.e., the
elderly). Therefore, the BMI is often used in conjunction with additional indicators, such as
waist-to-hip ratio. It may be argued that waist-to-hip ratio is a more appropriate measure of
obesity as centrally-distributed adiposity carries a greater health risk, and is suggested to be a
more accurate predictor of metabolic and cardiovascular disease (Janssen et al., 2009; Noble,
2001; Wei et al., 1997). Abdominal obesity is classified as a waist-to-hip ratio greater than 0.90
for men and 0.85 for women (World Health Organization, 2008).
1.2.1 Consequences of Obesity:
Obesity is associated with an increased risk of morbidity and mortality (Ogden et al.,
2007). The morbidities associated with obesity can be broken down into two pathophysiologic
categories. The first category of risks is a result of metabolic changes that are caused by products
secreted from excess fat. These include the development of hypertension, insulin resistance,
Type 2 diabetes mellitus, renal impairment, stroke, cardiovascular disease, hypertriglyceridemia,
atherosclerosis, and low high density lipoprotein (HDL) cholesterol. The second category of
morbidities arises as a result of the increased fat mass itself. These risks include chronic hypoxia
and hypercapnia, obstructive sleep apnea, gout, osteoarthritis, and degenerative joint disease
(Bray, 2004). Approximately 80% of obese adults suffer from one or more of these morbidities
(Must et al., 1999). Correspondingly, a higher body weight is associated with an increased
number of co-morbidities (Ogden et al., 2007), and the BMI and mortality are positively
correlated (Bray et al., 2003). In addition to mortality associated with cardiovascular disease,

4
obese women show an increased mortality from endometrial cancer, while obese men show an
increased mortality from colorectal cancer (Bray, 2004).
It is important to note that central obesity, the accumulation of abdominal fat resulting in
an increase in waist size, poses the greatest health risk. Thus, it is suggested that waist-to-hip
ratio or circumference is a better indicator of cardiovascular and metabolic risk than BMI. The
visceral fat present in central adiposity is quite different from subcutaneous fat. Specifically,
visceral adipose tissue is more metabolically active: this type of adipose tissue contains large
insulin-resistant adipocytes, whereas adipose tissue associated with lower body obesity contains
small, insulin-sensitive adipocytes. Furthermore, visceral adipocytes yield a greater fatty acid
turnover. Large visceral adipocytes have a higher density of adrenergic receptors as compared
with the adipocytes present in lower body obesity. The combination of decrease in insulin-
mediated anti-lipolysis and increase in catecholamine-mediated lipolysis in abdominal adipose
tissue results in increased circulating levels of non-esterified fatty acids in plasma (Sharma,
2002).
1.2.2 Leptin:
Leptin plays an integral role in body weight maintenance through its involvement in the
regulation of food intake and energy expenditure. This peptide hormone is produced primarily in
adipocytes and circulates in the bloodstream in proportion to body fat (Zhang et al., 1994).
Plasma leptin is transported into the central nervous system (CNS) by a unidirectional, saturable
system (Banks et al., 1996). Upon entering the CNS, leptin functions as an adiposity signal to
indicate the level of the body’s energy stores (Friedman and Halass, 1998).

5
Activation of leptin receptors in the arcuate nucleus (ARC) of the hypothalamus lead to
inhibition of the orexigenic peptides Neuropeptide Y (NPY) and Agouti-related peptide (AgRP).
In addition, leptin activates the Pro-opiomelanocortin (POMC) and Cocaine and amphetamine
regulated transcript (CART) pathway to exert an anorexic effect (Friedman and Halass, 1998).
Thus, this adipokine signals satiety. Additionally, leptin functions to increase energy expenditure
and sympathetic tone. Specifically, it has been shown that leptin acts on cardiovascular control
centers in the brain to target sympathetic outflow to the kidney (Mark et al., 2009).
1.2.2.1 Leptin Resistance:
In rodents and humans who lack leptin, severe obesity and hyperphagia occurs, as the
brain no longer receives a signal that the body has adequate fat stores (Friedman and Halaas,
1998). Surprisingly, congenital leptin deficiency in humans is rare. In fact, the majority of obese
individuals exhibit elevated circulating leptin levels. Once obesity is established, exogenous
leptin is relatively ineffective in reducing food intake or adiposity. This indicates the
manifestation of a leptin resistance or insensitivity (Friedman and Halaas, 1998). It is important
to note that this resistance is ‘selective’: in the obese leptin-resistant state, leptin no longer
functions to indicate satiety; however, leptin continues to increase sympathetic excitation. This
selective resistance ultimately contributes to the development of obesity related hypertension
(Rahmouni et al., 2005).
1.3 Metabolic Syndrome
The metabolic syndrome refers to a combination of clinical disorders that increase the risk of
developing diabetes and cardiovascular disease. Like many diseases, the risk of developing the

6
metabolic syndrome depends on the interaction between genetic and environmental variables.
The two most important risk factors for the metabolic syndrome are central obesity and insulin
resistance. Other risk factors include: hypertension, elevated fasting blood glucose, elevated
triglycerides and low HDL cholesterol (A.D.A.M Medical Encyclopedia, 2011).
The underlying cause of the metabolic syndrome remains unidentified. However, this
specific clustering of risk factors is associated with a 2-fold increased risk for cardiovascular
disease, and a 5-fold increased risk for Type 2 diabetes mellitus (Alberti et al., 2009).
Approximately 20-30% of the global adult population is plagued with this condition (Grundy,
2008).
1.3.1 Insulin:
Insulin is a peptide hormone that is produced and secreted by the β-cells of the pancreas.
Insulin is responsible for the maintenance of normal blood glucose levels by promoting glucose
uptake in muscle tissue and adipocytes. In addition, insulin functions to regulate carbohydrate,
protein, and lipid metabolism (Wilcox, 2005).
Like leptin, insulin circulates in the bloodstream in proportion to body fat. Upon entering
the CNS, insulin is involved in appetite regulation, memory, cognition, and olfaction (Wilcox,
2005). Insulin and leptin share a common signalling pathway in the hypothalamus (Fig. 1.1)
(Niswender and Schwartz, 2003). The binding of leptin and insulin to their respective receptors
on the POMC neurons yields the release of alpha-Melanocyte-stimulating hormone (α-MSH). α-
MSH stimulates downstream “catabolic” neurons to reduce food intake and increase energy
expenditure. In addition, insulin and leptin inhibit the potent orexigenic NPY and AgRP
expressing neurons. This inhibition results in a reduced secretion of NPY and AgRP, thus

7
Figure 1.1. Leptin and Insulin signalling in the arcuate nucleus of the hypothalamus. The
binding of leptin and insulin to their respective receptors (insulin, IR, leptin, lepr) on the
proopiomelanocortin (POMC) neurons reduce food intake and increase energy expenditure. In
opposition, insulin and leptin inhibit the neuropeptide Y (NPY) and Agouti related protein
(AgRP) expressing neurons to reduce food intake (modified from Niswender and Schwartz,
2003).

8
decreasing food intake. AgRP is an endogenous antagonist for the Melanocortin 4 receptor
(MC4R) for α-MSH. Thus, increased insulin and leptin signalling increase MC4R agonism
through α-MSH release, and decrease antagonism by suppressing AgRP release. Neural circuits
originating within the ARC control a number of physiological processes including food intake,
energy expenditure, glucose homeostasis, endocrine function, and reproductive function (Fig.
1.1) (Niswender and Schwartz, 2003).
1.3.1.1 Insulin resistance:
Insulin resistance arises as a complication of obesity and is a key component of the
metabolic syndrome. Specifically, it is the accumulation of intra-abdominal fat that correlates
positively with insulin resistance (Cnop, 2002). In the presence of insulin resistance, a normal or
elevated level of circulating insulin produces an attenuated biological response. In individuals
experiencing insulin resistance, the β-cells of the pancreas must secrete more insulin in order to
maintain a state of euglycemia. Eventually, in response to a prolonged hyperfunction, a “β-cell
exhaustion” occurs. When the β-cell reserve capacity of insulin is exceeded by demands from the
metabolism, insulin secretion becomes inadequate. Ultimately, this yields an increase in blood
glucose levels (Miranda et al., 2005).
Insulin resistance is best categorized by the Homeostatic Model Assessment of Insulin
Resistance (HOMA-IR). This states that insulin resistance can be calculated as the product of
fasting insulin level and fasting blood glucose level divided by 22.5 (Matthews et al., 1985).

9
1.3.2 Dyslipidemia:
Dyslipidemia, an abnormal amount of lipid in the blood, is a direct consequence of
insulin resistance. The dyslipidemia present in the metabolic syndrome is characterized by an
elevated triglyceride level, and elevated low density lipoprotein (LDL) cholesterol and very low-
density lipoprotein (VLDL), and a decreased high density lipoprotein (HDL) cholesterol,
(Miranda et al., 2005).
1.4 Arterial pressure
Arterial pressure (AP) is an important indicator of health. Continuous and sufficient AP is
essential for adequate blood flow into all organs: if AP is too low, there will be inadequate
perfusion of organs and when AP is too high, heart disease, stroke, and/or vascular disease will
occur. Thus, AP must be maintained within a narrow range that coincides precisely with the
needs of the tissue (Ackermann, 2004). Regional blood flow can be efficiently regulated by
vascular control mechanisms, such as mechanical forces and chemical stimuli. In addition to this
local control system, AP is also regulated by the CNS (Thomas, 2011). Central neuronal circuitry
regulates cardiovascular function in order to meet the needs of the body as a whole. The neural
control of the cardiovascular system is controlled primarily by the sympathetic nervous system
(SNS), in conjunction with limited, but essential input from the afferent component of the
parasympathetic nervous system. The postganglionic sympathetic neurons that function to
regulate and control cardiovascular targets are primarily noradrenergic (Thomas, 2011).
AP fluctuates within the cardiac cycle. Specifically, AP peaks in systole and is lowest
during diastole. As a result of this constant fluctuation, mean arterial pressure (MAP) is often
used as a representative value of systemic AP. MAP is equal to the sum of the diastolic blood

10
pressure (DBP) plus one-third of the pulse pressure. Pulse pressure is equal to the difference
between systolic blood pressure (SBP) and DBP (Ackermann, 2004).
1.4.1 Baroreceptor Reflex:
Although AP is regulated by multiple organs and systems, the baroreceptor reflex plays a
critical role in both short-term and long-term AP regulation. The function of the baroreceptor
reflex is to stabilize AP despite disturbances to homeostasis. The level of activity of the
baroreceptors at rest is presumed to be the most crucial parameter for long-term AP control.
Baroreceptors are stretch sensitive mechanoreceptors located in the walls of the arteries,
veins, heart, carotid sinus, and aortic arch. Primary afferent fibers from baroreceptors detect
changes in AP (transmural pressure) and relay this information to the brainstem (Stauss, 2002).
The primary afferent fibers of the baroreceptor reflex terminate in the nucleus of the solitary tract
(NTS) (Ciriello et al., 1983). These NTS baroreceptor sensitive neurons project to the caudal
ventrolateral medulla (CVLM). Activation of the CVLM functions to inhibit the actions of the
rostral ventrolateral medulla (RVLM), the primary regulator of sympathetic nervous outflow.
From the RVLM, neurons project directly to sympathetic cardiovascular and vasomotor
preganglionic neurons in the intermediolateral cell column of the spinal cord (IML) (Weston et
al., 2003). In summary, baroreceptors transduce changes in AP and then send this information to
the brainstem where in return it initiates a compensatory mechanism in order to return AP to
within normal ranges (refer to Fig. 1.2; modified from Pilowsky and Goodchild, 2002).
Central catecholaminergic cell groups have been shown to play a crucial role in
cardiovascular control (Guyenet and Cabot, 1981). The C1 adrenergic cell group (C1) is located
within the RVLM and these neurons project to the sympathetic preganglionic neurons of the

11
Figure 1.2. The major pathways involved in the baroreceptor reflex. Open circles indicate
inhibitory neurons while filled circles represent excitatory neurons. Solid lines represent
excitatory pathways while inhibitory pathways represent by dashed lines. IML, intermediolateral
cell column of the spinal cord; RVLM, rostral ventrolateral medulla; CVLM, caudal ventrolateral
medulla; NTS, nucleus of the solitary tract (modified from Pilowsky and Goodchild, 2002).

12
spinal cord and are directly involved in the control of AP (Lipski et al., 1995). However, not all
C1 neurons are under baroreceptor control. In fact, some C1 neurons innervate areas of the
hypothalamus to regulate different aspects of circulation through the release of hormones that
control sodium and water balance and enable the activation of hypothalamic-pituitary axis during
an assortment of physical stresses (Verberne et al., 1999).
The A1 noradrenergic cell group (A1), is located in the CVLM, while the A2
noradrenergic cell group (A2) is located within the NTS. These neurons project to the
vasopressin and oxytocin containing magnocellular neurosecretory cells within the
paraventricular nucleus of the hypothalamus (PVH) and maintain reciprocal connections with
cardiovascular sites within the brainstem. Some of these noradrenergic neurons likely also match
up with the depressor neurons implicated in the baroreflex control of sympathetic outflow
(Pilowsky and Goodchild, 2002). Interestingly, total central catecholamine levels are higher in
hypertension (Goldstein, 1983), but this may reflect an increase in sympathetic outflow,
increased baroreceptor activity attempting to compensate for the increased AP or a combination
of both.
1.4.2 Hypertension:
According to the recommendations from the seventh report of Joint National Committee
of Prevention, Detection, Evaluation, and Treatment of high blood pressure, the classification of
blood pressure for individuals over the age of 18 is as follows (Chobanian et al., 2003):
Normal blood pressure: SBP between 90-119 mmHg, DBP between 60-79mmHg
Pre-hypertensive: SBP between 120-139mmHg, DBP between 80-90mmHg
Stage 1 hypertension: SBP between 140-159mmHg, DBP between 90-99mmHg

13
Stage 2 hypertension: SBP ≥ 160mmHg, DBP ≥100mmHg
Prehypertension indicates that patients are at risk of developing hypertension and lifestyle
modifications are critical to maintain health and prevent the progression of hypertension
(Chobanian et al., 2003).
There are two types of hypertension: essential (or primary) hypertension and secondary
hypertension. Essential hypertension encompasses approximately 95% of hypertensive patients.
Essential hypertension is defined as the diagnosis of hypertension in the absence of any
identifiable secondary cause. It is proposed that factors such as genetic predisposition, obesity,
excess dietary salt intake, and adrenergic tone interact to result in hypertension. Secondary
hypertension arises as a direct result of an identifiable cause, including kidney diseases, tumors,
or endocrine diseases.
Hypertension is one of the most common diseases, and affects approximately 25% of the
Canadian population (Robitaille et al., 2012). In addition, hypertension is one of the most
important modifiable risk factors for various diseases including coronary heart disease, stroke,
congestive heart failure, and end-stage renal disease. As previously indicated, obesity is one of
the gravest medical issues our society is currently faced with and it has a strong correlation with
hypertension (Wilcox, 2005).
1.4.3 Obesity-associated Hypertension:
The associations between obesity and hypertension are well documented (Sironi et al.,
2011; Zalesin et al., 2011). As the prevalence of obesity increased dramatically in the last
decade, a similar trend was seen in hypertension (Public Health Agency of Canada, 2010).
However, the mechanisms involved in the pathogenesis of obesity-associated hypertension

14
remain unclear (Bjorntorp and Rosmond, 2000; Kassab et al., 1995; Landsberg and Krieger,
1989). It has become increasingly clear that central neural mechanisms through the activation of
the SNS play a pivotal role (Lohmeier et al., 2003; Rahmouni et al., 2005), especially as
evidence indicates an impaired baroreceptor reflex function in obesity-associated hypertension
(Grassi et al., 2000; Lohmeier et al., 2003).
1.5 Ginseng
Ginseng refers to the root of several species of plants of the genus Panax. The name
Panax is derived from the Greek words “pan” and “axos” meaning “all-heal” (Yun, 2001).
Ginseng is often referred to as an adaptogen or panacea. This is indicative of the traditional
belief that ginseng has the capacity to heal all aspects of the body, invigorate the body, and
prolong life (Bensky and Gamble, 1993).With origins in Asia, ginseng has been used for at least
2000 years as a medicinal herb. The name ginseng is derived from the Chinese words “Jen
Sheng” meaning “man herb”, describing the root’s humanoid shape (Fig. 1.3; modified from Yun
et al., 2001). Most commonly, it is the root, or an extract prepared from the root, that is
consumed for medicinal purposes; however, beneficial effects have also been reported for the
leaf and berry components of the plant (Attele et al., 2002; Dey et al., 2003; Jung et al., 2005;
Xie et al., 2002A, 2002B, 2004A).
The oldest Traditional Chinese Medicine (TCM) text, “Shen Nong Ben Cao Jing”, states
that ginseng has many pharmacological functions, including restorative, tonic, nootropic, and
anti-aging effects (Xiang et al., 2008). Modern pharmacological research has shown that ginseng
has positive effects in the treatment of cancer, enhances memory, increases immune function,
aids sexual function, and offers some neuroprotective effects (Azike et al., 2011; Dong et al.,

15
Figure 1.3. Fresh ginseng shaped like a man. The name ginseng is derived from the Chinese
words “Jen Sheng” meaning “man herb” describing the root’s humanoid shape (modified from
Yun et al., 2001).

16
2011; Kim et al., 2009A; Persson et al., 2004). Presently, ginseng is one of the most famous, and
well-researched herbs in the world (Blumenthal et al., 2006; Stavro et al., 2006).
1.5.1 Ginsenosides:
Ginseng is made up of many components, including glycoproteins, peptides,
polysaccharides, and ginsenosides. Ginsenosides, also referred to as ginseng saponins, are unique
to the Panax species (Yun, 2001). Over 150 different ginsenosides have been isolated from the
Panax species thus far. Naming ginsenosides follows the template “ginsenoside Rx”, while ‘R’
stands for the root while ‘x’ describes the chromatographic polarity in alphabetical order. For
example, ginsenoside Ra (G-Ra) is the least polar compound (Leung and Wong, 2010).
Evidence suggests that ginsenosides may serve to protect the plant. Ginsenosides have
been shown to be anti-microbial and anti-fungal (Leung and Wong, 2010). In addition, the bitter
taste of ginsenosides may serve as an anti-feedant (Leung and Wong, 2010; Nicol et al., 2002).
The ginsenoside content present in each root varies depending on the Panax species. In
addition, the concentration of ginsenosides depends on the plant age, the preservation method,
the extraction method, the season of harvest, and the quality of the soil (Leung and Wong, 2010).
Ginseng is believed to be most potent after 4-5 years of growth, and is typically harvested at that
age (Leung and Wong, 2010). The majority of harvested ginseng roots are air dried. Most often,
ginseng prepared in this manner is referred to simply as ginseng; however, it can also be referred
to as white ginseng. If the roots are steamed prior to drying the resulting product is referred to as
red ginseng (Kim et al., 2000).
Ginsenosides have the ability to target a myriad of tissues to produce an assortment of
pharmacological responses. This is thought to be a result of their steroid-like structure (Leung

17
and Wong, 2010). All ginsenosides share a common four-ring hydrophobic steroid structure with
attached sugar moieties (i.e., the side chains can be free or coupled with various sugars). It is
these sugar components that are thought to provide the different ginsenosides with their
specificity for various cellular effects (Nah, 1997).
Each ginsenoside may have different pharmacological functions due to their different
chemical structures. It has been shown that single ginsenosides can produce multiple effects in
the same tissue (Odashima et al., 1985). In addition, non-ginsenoside components of ginseng
have been shown to exert pharmacological effects (Sonoda et al., 1998; Wang et al., 2010a;
Wang et al., 2010b). The fact that ginseng is made up of multiple components that may have
differential effects, contributes to the difficulty in elucidating the actions and mechanisms of
ginseng.
There are two categories of ginsenosides: protopanaxadiol ginsenosides (PPD) and
protopanaxatriol ginsenosides (PPT) (Fig. 1.4; modified from Liu et al., 2006). The classification
of each ginsenoside depends on the position of various carbohydrate moieties attached at the
position of carbon 3, 6, and 20 (Liu et al., 2006). PPD ginsenosides include G-Rb1, G-Rb2, and
G-Rb3. PPT ginsenosides include G-Re, G-Rg1, and G-Rf (Liu et al., 2006).
1.5.2 Types of Ginseng:
There are two main types of ginseng recognized worldwide: Asian ginseng, or Panax
ginseng and NAG, or Panax quinquefolius. Asian ginseng is cultivated in China, Korea, and
Japan while NAG grows in the eastern temperate forested areas of North America. Canada is the
largest producer of NAG and the vast majority of ginseng crops are located in Ontario. Ginseng

18
Figure. 1.4. Characteristic ginsenosides present in both Asian and NAG grouped into
protopanaxadiol and protopanaxtriol groups (modified from Liu et al., 2006).

19
production worldwide has been increasing for the last 25 years. Over 1.54 million kg of NAG
were exported from Canada in 1998 alone (Ginseng Ontario, 2012).
According to TCM, Asian ginseng and NAG are thought to embody the complementary
forces of “yin” and “yang”; NAG is calming and nourishing, and provides the “yin”, or cooling
effect that offsets stress, while Asian ginseng is stimulating and invigorating providing the
“yang”, or warmth (Chen et al., 2008). Although both types of ginseng contain ginsenosides,
Asian ginseng and NAG each possess a distinct chemical profile.
One key difference between the two types of ginseng is the ratio of PPD to PPT
ginsenosides: NAG is known to contain higher levels of ginsenoside Rb1 (G-Rb1) and low levels
of ginsenoside Rg1 (G-Rg1), while the opposite is true of Asian ginseng (Fuzzati et al., 2004).
The ratio of G-Rb1:G-Rg1 is used for the identification and standardization of ginseng
products. G-Rb1:G-Rg1 values between 1 and 3 correspond to Asian ginseng, while values
greater than 10 are indicative of NAG (Fuzzati, 2004). The main ginsenosides present in NAG
are G-Rb1, G-Re, and G-Rd, while Asian ginseng contains greater levels of G-Rg1, G-Rb2, and
G-Rc. Furthermore, G-Rf serves as a marker ginsenoside, as it is present in Asian ginseng but
not in NAG (Schlag and McIntosh, 2006). Conversely, ginsenoside F11 is present in NAG and
not in Asian ginseng (Assinewe et al., 2003).
Korean Red ginseng can be prepared by steaming the roots of Panax ginseng at 100°C
for 2-4 hours (h) prior to drying. This is a common method of preparation and yields
ginsenosides (G-Rg3 and G-Rh2) that are not typically present following other extraction or
preparation methods (Popovich and Kitts, 2004). This type of ginseng is suggested to have a
higher medicinal value than Asian ginseng (Kim et al., 2000).

20
NAG contains approximately 3-5 times greater total ginsenoside content than Asian
ginseng (Stavro et al., 2006). The total ginsenoside content of Asian ginseng range from 0.2 to
2% whereas ginsenoside content in NAG roots range between 4 and 10% (Fuzzati, 2004).
Consequently, pharmacological activity has been reported to be stronger for NAG in comparison
to Asian ginseng (Chen et al., 2008).
1.5.3 Ginseng Metabolism:
Ginsenosides possess a bulky molecular structure that results in poor membrane
permeability and renders them prone to degradation. Upon oral consumption, ginsenosides are
exposed to acid hydrolysis, glycosyl elimination, side-reactions, and epimerization at Carbon-20
(Hasegawa, 1996). In addition, ginsenosides are metabolized by intestinal microflora into
metabolites that are capable of being absorbed by the body. For example, PPD ginsenosides (G-
Rb1, G-Rc and G-Rd) are transformed into Compound K, whereas PPT ginsenosides (G-Re, G-
Rg1, and G-Rf) degrade into G-Rh1 and F1 (Tawab et al., 2003). The oral bioavailability of
ginsenosides is thought to be less than 5% (Hasegawa, 2004).
Despite a low oral bioavailability, ginsenosides have been suggested to undergo rapid
absorption by the gastrointestinal tract (Joo et al., 2010; Li et al., 2007; Song et al., 2010; Yan et
al., 2007). Ginsenosides have been shown in the blood stream of canines within 10 minutes
(Song et al., 2010) and in rats within less than one hour after oral administration (Joo et al., 2010;
Li et al., 2007; Xu et al., 2003; Yan et al., 2007). Similar findings have been reported in humans:
ginsenosides are present in plasma one hour after oral administration of ginseng (Tawab et al.,
2003).

21
The precise pharmacokinetics of ginseng has not yet been determined. However,
systemic exposure of G-Rb1 has been shown to be greater than that of other ginsenosides (Song
et al., 2010), According to one study, G-Rb1 plasma concentrations have been shown to peak at
1.5 h following oral administration, and G-Rb1 was shown to remain present in plasma for 72 h
(Xu et al., 2003). However, recent studies have shown G-Rb1, G-Re and G-Rg1 to peak
approximately 45 minutes (mins) after oral administration in rats (Li et al., 2007; Joo et al.,
2010). Yan and colleagues have also shown that G-Rb1 remains present in rat plasma for at least
36 h (Yan et al., 2007). Conversely, PPT ginsenosides are more rapidly degraded: G-Re and G-
Rg1 have been shown to remain within the rat systemic circulation from 1 to 12 hours following
oral administration (Qi et al., 2011; Yan et al., 2007).
It has been suggested that ginsenosides remain in human plasma for approximately 12 h
(Tawab et al., 2003). Compound K, the main metabolite of PPD ginsenosides, was present in
human serum for 7.5 to 11 h after administration (Tawab et al., 2003; Shibata, 2001). However,
it was noted that intestinal degradation of ginsenosides in humans is significantly different
among subjects (Tawab et al., 2003). Another study in humans had similar results and observed
that while in some subjects Compound K was observed as early as 4 h after oral administration
and remained present up to 24 h after, the average absorption occurred between 9-14 h after
administration (Lee et al., 2009A). Furthermore, G-Rb1 was observed in human plasma
approximately 5.5 h after oral administration and remained present for 3 h (Tawab et al., 2003).
As PPT ginsenosides are known to degrade more rapidly, it was not surprising that PPT
metabolites were present 1 h after administration (Tawab et al., 2003). It has been suggested that
with higher dosages, enzymes in the gut may become saturated and thus ginsenosides are able to
enter the blood stream prior to degradation into their metabolized forms. However, with smaller

22
dosages of ginseng, it is more likely that only ginsenoside metabolites will reach the circulation
(Xu et al., 2003). There is evidence to indicate that the metabolites of ginseng are responsible for
many of the pharmacological effects. Comparative experiments using intravenous or abdominal
cavity injections of ginseng metabolites confirm that ginsenoside metabolites provide greater
biological effects than ginsenosides alone (Hasegawa and Uchiyama, 1998; Lee et al., 2005; Yim
et al. 2004).
Pharmacokinetic evaluation of ginseng has proven to be quite difficult, partially due to
the diversity of ginsenosides (Qi et al., 2011). Although trends between studies are common,
differences in sample pre-treatment and analysis may explain some differences in the results
obtained in studies. Furthermore, differences may be observed between administration of a single
ginsenoside versus a prepared extract as competitive absorption and metabolism between the
different components may occur (Qi et al., 2011). Recently, studies have sought to investigate
how to increase and maximize bioavailability of ginsenosides. Possible mechanisms of achieving
this include co-administration with adrenaline (Xiong et al., 2009), intranasal administration (Lu
et al., 2011) or emulsification into lipid-based formula (Xiong et al., 2008).
1.5.4 The Effect of Ginseng in the Central Nervous System:
Despite the fact that brain tissue levels for most ginsenosides have been reported to be
very low, ginseng has been reported to exert an effect within the CNS (Xie et al., 2005B).
Interestingly, it has been suggested that ginseng metabolites, such as Compound K, do not cross
the blood brain barrier (Kim et al., 2003). Nevertheless, the neuroprotective effects of ginseng
are well reported (Li et al., 2010; Mizumaki et al., 2002; Sugaya et al., 1988; Van Kampen et al.,
2003). Ginseng has been shown to possess some protective effects in neurodegenerative disease:

23
such as in Parkinson’s and Alzheimer’s diseases (Lee et al., 2001; Tohda et al., 2002; Van
Kampen et al., 2003). Ginseng has also been shown to prevent learning and memory defects in
aging (Kurimoto et al., 2004; Zhao et al., 2011). Furthermore, ginseng has also been shown to
modulate neurotransmission, and to attenuate neuro-inflammation (Benishin, 1992; Itoh et al.,
1989).
Ginseng has been shown to modulate catcholaminergic activity within the CNS by its
effects on tyrosine hydroxylase expression in the brain (Kim et al., 2010). Additionally, G-Rb1,
the main ginsenoside found within NAG, has been suggested to provide a partial neurotrophic
and neuroprotective effect on catecholaminergic neurons (Hwang and Jeong, 2010; Radad et al.,
2004). Furthermore, G-Rb1 has been reported to increase the release of the neurotransmitter
glutamate within the CNS (Xue et al., 2006), and to interact with excitatory neurotransmitter
receptors (Lee, 2002). In addition, treatment with G-Rb1 has been reported to induce c-fos
expression in brain nuclei such as NTS, ARC, and ventromedial hypothalamus (VMH) (Xiong et
al., 2010).
1.6 The Metabolic and Cardiovascular effects of Ginseng
1.6.1 The Effects of Asian ginseng on Metabolic Variables
1.6.1.1 Asian Ginseng has anti-diabetic effects:
According to TCM, ginseng is effectively used in the treatment of diseases associated
with aging. It is well known that the metabolic syndrome has an increased prevalence in the
older population (Vuksan et al., 2005).
Animal experiments have reported that Asian ginseng possesses anti-hyperglycemic
activity (Dey et al., 2003; Kimura et al., 1999; Lim et al., 2009; Su et al., 2007; Xie et al., 2002;

24
Yun et al., 2004). Treatment with Asian ginseng resulted in decreased plasma glucose (Mollah et
al., 2009; Yun et al., 2004) and increased insulin levels (Dey et al., 2003; Su et al., 2007; Kim et
al.., 1997; Kimura et al., 1999; Su et al., 2007). In addition, Asian ginseng improved glucose
homeostasis and insulin sensitivity (Attele et al., 2002; Kim et al., 2005; Kimura et al., 1999;
Lim et al., 2009; Yun et al., 2004;). Furthermore, Asian ginseng treatment has also been shown
to increase plasma concentrations of C-peptide, a byproduct of insulin production (Su et al.,
2007). Clinical trials strongly support the claim that Asian ginseng possesses anti-diabetic
properties and improves insulin resistance in diabetic individuals (Ma et al., 2008; Sotaniemi et
al., 1995; Vuksan et al., 2008), but has little effect in healthy patients (Reay et al., 2009). Asian
ginseng treatment increased the stimulation of insulin secretion which may serve as a protective
effect by preventing apoptosis of β-cells (Kim et al., 2007). It is suggested that Asian ginseng
enhances insulin secretion by facilitating acetylcholine release from nerve terminals, likely
through stimulation of muscarinic receptors in the pancreas (Su et al., 2007).
It has also been proposed that Asian ginseng reduces the intestinal absorption of glucose
(Chung et al., 2001; Karu et al., 2007). In support of this suggestion, Asian ginseng was found to
inhibit hepatic glucose-6-phosphatase (G6Pase), an enzyme that plays an integral role in
homeostatic regulation of blood glucose levels (Chung et al., 2001). Asian ginseng also increased
messenger ribonucleic acid (mRNA) expression of Glucose Transporter Type 4 (GLUT4) and
Insulin Receptor (IR) suggesting that Asian ginseng increases insulin-regulated glucose transport
(Lim et al., 2009; Mollah et al., 2009). While the anti-diabetic effects of Asian ginseng root have
become clear, it has been suggested that Asian ginseng berry may have a stronger effect in
reducing blood glucose and improving glucose tolerance (Dey et al., 2003).

25
1.6.1.2 Asian Ginseng has anti-obesity effects:
Asian ginseng has been shown to have anti-obesity effects (Dey et al., 2003; Karu et al.,
2007; Kim et al., 2011; Kim and Park, 2003; Lee et al., 2010; Lim et al., 2009; Mollah et al.,
2009; World Health Organization, 1999; Xie et al., 2005A; Xie et al., 2002B; Yang et al., 2010;
Yun et al., 2004; Zierer, 1991). In high fat diet (HFD)-induced models of obesity, chronic Asian
ginseng treatment prevented weight gain (Karu et al., 2007; Kim et al., 2011; Kim and Park,
2003; Lee et al., 2010; Xie et al., 2005A; Yun et al., 2004; Yun et al., 2007; Zierer, 1991).
Similarly, in genetic models of obesity (ob/ob mice and Long Evans Tokushima rats) treatment
with Asian ginseng decreased body weight (Lim et al., 2009; Mollah et al., 2009). Finally, in
human patients with non-insulin dependant diabetes, treatment with Asian ginseng decreased
body weight and improved physical activity (Sotaniemi et al., 1995). While the vast majority of
literature has sought to investigate the effects of Asian ginseng root, it has been reported that
Asian ginseng berry exerts a greater anti-obesity effect (Dey et al., 2003; Xie et al., 2002B). In
fact, one study has shown that Asian ginseng root did not change body weight, while Asian
ginseng berry decreased body weight (Dey et al., 2003).
In addition to effects related to body weight, Asian ginseng has been shown to decrease
body fat mass (Lim et al., 2009; Kim et al., 2011; Yun et al., 2004; Yun et al., 2007).
Specifically, treatment with Asian ginseng decreased the diameter and overall size of adipocytes
(Kim et al., 2011; Yun et al., 2004). In HFD-induced obesity, Asian ginseng treatment has also
been reported to decrease leptin levels (Kim and Park, 2003; Lee et al., 2010).
Although the anti-obesity activity of Asian ginseng has been well-established,
information regarding its effect on food intake is less clear. While one study indicated that
treatment with Asian ginseng decreased food intake, the administration of individual

26
ginsenosides (G-Rg1, G-Re) had no effect (Yang et al., 2010). On the other hand, other studies
have reported no significant changes in food intake following Asian ginseng treatment (Cui et
al., 1998; Kim et al., 2011; Lim et al., 2009).
In HFD-induced obesity, Asian ginseng treatment has also been reported to decrease
plasma triglyceride concentration (Karu et al., 2007; Kim et al., 2011; Kim and Park, 2003; Lee
et al., 2010; Yun et al., 2004). However, following a normal chow diet Asian ginseng did not
alter triglyceride levels (Lim et al., 2009). In a HFD-induced (Kim et al., 2011; Kim and Park,
2003; Xia et al., 2011) or genetic model of obesity (Lim et al., 2009), Asian ginseng decreased
serum cholesterol levels (Kim et al., 2011; Xia et al., 2011). Specifically, LDL cholesterol was
decreased and HDL cholesterol was increased (Lim et al., 2009; Xia et al., 2011). However,
some studies have also reported no change in cholesterol levels following Asian ginseng
treatment (Karu et al., 2007). It may be important to note that in this latter study, the animals did
not experience elevations in cholesterol levels throughout the course of treatment (Karu et al.,
2007). This may suggest that Asian ginseng only modulates plasma cholesterol levels in the
presence of hypercholesterolemia. In humans it was initially reported that Asian ginseng did not
affect triglycerides or total cholesterol levels (Sotaniemi et al., 1995). However, more recent
clinical trials have provided evidence that administration of Asian ginseng extract led to
reduction of total cholesterol, triglycerides, and LDL cholesterol, and increased HDL cholesterol
(Kim and Park, 2003).
1.6.1.3 The potential mechanisms responsible for the metabolic effects of Asian ginseng:
High oxidative stress has been found in patients with a wide range of illnesses, including
Type 2 diabetes mellitus. Asian ginseng has been shown to possess anti-oxidative properties

27
(Kim and Park, 2003). In addition, Asian ginseng decreased production of nitric oxide (NO) and
reactive oxygen species (ROS) (Kim et al., 2007; Kim and Park, 2003). Recent studies have
observed that higher total dietary anti-oxidant intake is correlated with lower levels of glycemic
indices in diabetic individuals (Psaltopoulou et al., 2010). Therefore, it has been suggested that
the anti-oxidant effects of Asian ginseng may lead to the herb’s hypolipidemic actions (Kim and
Park, 2003).
Several mechanisms have been proposed to explain the anti-obesity effects of Asian
ginseng. There are data to suggest that Asian ginseng increases thermogenesis and energy
expenditure (Karu et al., 2007). Furthermore, Asian ginseng has been shown to inhibit the
orexigenic peptide NPY in the hypothalamus (Kim et al., 2005). In addition, Asian ginseng has
been shown to inhibit pancreatic lipase, the primary enzyme to hydrolyze dietary fat (Karu et al.,
2007; Kim et al., 2011). By preventing the absorption of dietary fats, Asian ginseng may reduce
caloric intake and thus exert an anti-obesity effect.
Another potential mechanism for the actions of Asian ginseng involves peroxisome
proliferator-activated receptor gamma (PPARɣ). The nuclear receptor PPARɣ plays a pivotal
role regulating metabolism by affecting gene transcription. Lipoprotein lipase (LPL) is one target
gene of PPARɣ in adipose tissue. Both PPARɣ and LPL are essential in maintaining lipid levels
within a normal range, and expression of these proteins is typically increased in obesity (Mollah
et al., 2009). Interestingly, treatment with Asian ginseng increased mRNA expression of both
PPARɣ (Chung et al., 2001; Lim et al., 2009; Mollah et al., 2009) and LPL (Mollah et al., 2009),
thus potentially facilitating lipid metabolism.

28
1.6.1.4 The metabolic effects of Korean Red ginseng:
Similarly to traditionally prepared Panax ginseng, several studies indicate that Korean
Red ginseng possess anti-hyperglycemic effects, and increases insulin sensitivity (Lee et al.,
2009B; Lee et al., 2012; Park et al., 2005). However, these findings are controversial as in one
study Korean Red ginseng had no effect on blood glucose levels (Lee et al., 2010). Treatment
with Korean Red ginseng was able to delay the deterioration of glucose tolerance and partially
prevent Type 2 diabetes mellitus in a genetically obese and diabetic animal (Otsuka Long-Evans
Tokushima fatty rats) (Lee et al., 2009B). However, Korean Red ginseng has been proposed to
exert an anti-hyperglycemic effect in humans (De Souza et al. 2011, Sievenpiper et al., 2006;
Vuksan et al., 2001). Patients with Type 2 Diabetes were administered Korean Red ginseng in
adjunct to their usual anti-diabetic treatment (Vuksan et al., 2008). It was shown that Korean Red
ginseng improved plasma glucose and insulin levels beyond the effects of conventional oral anti-
hyperglycemic medications (Vuksan et al., 2008). However, a preparation including Korean Red
ginseng in conjunction with other anti-diabetic herbs have been reported to have no effect (Kim
et al., 2012).
In addition to regulating blood glucose, Korean Red ginseng has been shown to regulate
lipid metabolism. Korean Red ginseng consumption decreased body weight gain (De Souza et
al., 2011; Kim et al., 2005; Kim et al., 2009B; Lee et al., 2009B; Lee et al., 2012) and fat pad
mass in an animal models (Kim et al., 2009B). In addition Korean Red ginseng decreased white
adipose tissue mass (Lee et al., 2009B; Lee et al., 2012) and leptin levels (Kim et al., 2009B; Lee
et al., 2012), but did not alter plasma adiponectin concentration (Lee et al., 2012). Korean Red
ginseng treatment in HFD fed mice decreased adipocyte size to a level comparable with low fat
diet fed animals (Lee et al., 2010). Furthermore, Korean Red ginseng increased body temperature

29
suggesting that it affects metabolism and thermogenesis (Lee et al., 2009B). However, in
humans, Korean Red ginseng had no effect on body weight or BMI (Kim et al., 2012).
Although hypothalamic NPY, an orexigenic peptide, has been shown to be down-
regulated following Korean Red ginseng treatment (Kim et al., 2005; Kim et al., 2009), evidence
supporting an effect on food intake is not clear. Although it has been reported that Korean Red
ginseng treatment yields a reduction in food intake (Kim et al.,2005; Kim et al., 2009), other
studies report no change (Lee et al., 2009B; Lee et al., 2012) or an increase in food intake
(Raghavendran et al., 2012).
Following Korean Red ginseng treatment, plasma triglyceride levels were reduced (Kim
et al., 2009B; Lee et al., 2010; Park et al., 2005; Yamamoto et al., 1983) and there was evidence
to suggest Korean Red ginseng decreases total cholesterol levels (Kim et al., 2009B; Park et al.,
2011; Yamamoto et al., 1983). However, another study reported no effect (Lee et al., 2010). In
both rats and human patients fed a high-cholesterol diet, administration of Red ginseng powder
decreased total plasma cholesterol levels and increased HDL cholesterol (Yamamoto et al., 1983;
Yang et al., 2010).
1.6.1.5 The potential mechanisms of the effects of Korean Red ginseng:
Korean Red ginseng has been shown to modulate gene expression of several genes
related to glucose and lipid metabolism. Korean Red ginseng treatment increased the expression
of GLUT4 (Lee et al., 2009B; Lee et al., 2012). Following treatment with HFD, there is
decreased translocation of GLUT4, and decreased phosphorylation of insulin receptor β, insulin
receptor substrate 1(IRS-1), Protein Kinase B (Akt), and Glycogen synthase kinase 3. These
defects in insulin signalling were restored following Korean Red ginseng treatment (Lee et al.,

30
2012). Thus, it was suggested that the anti-diabetic effects of Korean Red ginseng may be partly
the result of enhancing the insulin signalling pathway. Furthermore, Korean Red ginseng
increased the expression of PPARγ coactivator–1α, nuclear respiratory factor–1, cytochrome c,
cytochrome c oxidase–4, and GLUT 4. These findings indicate that Korean Red ginseng
activated AMP-activated protein kinase (AMPK) and glucose utilization in skeletal muscle (Lee
et al., 2009B).
With respect to regulating lipid metabolism, it was observed that Korean Red ginseng
increased mRNA expression of liver peroxisome proliferator-activated alpha (PPARα) and LPL
(Park et al., 2005). Korean Red ginseng also had an effect on the expression of lipogenesis-
related genes: sterol regulatory element-binding protein (SREBP-1c), diglyceride acyltransferase,
and fatty acid synthase (FAS). Following a HFD, these genes show an increased expression;
however, Korean Red ginseng treatment suppressed this increase (Lee et al., 2010). It has also
been suggested that Korean Red ginseng promotes fatty acid oxidation via activation of AMPK
and phosphorylation of acetyl coenzyme-A (acetyl-CoA) muscle carboxylase in skeletal muscle.
AMPK increases glucose uptake and fatty acid oxidation in skeletal muscle cells to increase
adenosine triphosphate (ATP) production while acetyl-CoA carboxylase is the rate-limiting
enzyme of FAS (Lee et al., 2009B). Activation of these pathways may help to explain the anti-
obesity effects of Korean Red ginseng via increasing fatty acid oxidation.
1.6.1.6 The Anti-hyperglycemic Effects of Ginsenoside Rg1:
It is well known that insulin resistance in skeletal muscle is a major contributor to the
development of Type 2 diabetes mellitus (Lee et al., 2011A). Treatment with G-Rg1, the main
ginsenoside in Asian ginseng, significantly increased glucose uptake in insulin-resistant muscle

31
cells. Following treatment with G-Rg1 there was an increased abundance of GLUT4 through the
AMPK pathway (Lee et al., 2011A). Furthermore, the effects of G-Rg1 are abolished following
treatment with an AMPK inhibitor. In vitro studies suggest that G-Rg1 suppresses hepatic
glucose production and the effects may be mediated by the LKB1-AMPK-Fox-01 pathway (Kim
et al., 2010).
1.6.2 The Effect of Asian ginseng on Cardiovascular Variables
1.6.2.1 The Effect of Asian ginseng on Arterial Pressure:
Several decades ago, a study in canines showed that intravenous infusion of Asian
ginseng yields a brief hypotensive response followed by a hypertensive phase. This hypotensive
phase was abolished with atropine, suggesting a potential cholinergic mechanism (Wood et al.,
1964). However, it should be noted that recent studies suggest ginseng metabolites are more
pharmacologically relevant than the ginsenosides themselves (Hasegawa and Uchiyama, 1998;
Lee et al., 2005; Yim et al. 2004).
In addition to variances by method of administration, there is evidence to suggest that the
differential effects of Asian ginseng on arterial pressure (AP) may be attributed to the type of
extract used. An alcoholic extract of Asian ginseng decreased heart rate (HR) and mean arterial
pressure (MAP), while increasing cardiac output and stroke volume. Conversely, aqueous extract
had no effect on HR or MAP, but decreased cardiac output, stroke volume, and central venous
pressure (Lee et al., 1981).
An early observational clinical study suggested a correlation between ginseng and
hypertension (Siegel, 1979). This study followed 133 individuals who self-reported the use of
ginseng, and concluded that 17% of patients became hypertensive within two years. This study

32
(Siegel, 1979) has since been heavily criticized by researchers and herbalists for having a poor
experimental design, as well as uncontrolled, anecdotal, and unverified results. However, the
stigma remains and there is still some concern regarding the effects of ginseng on AP.
According to TCM, it is suggested that Asian ginseng has a balancing effect on AP: no
change in normotensive conditions, and a normalizing effect on abnormal AP (Yue et al., 2007).
Modern pharmacological studies have confirmed this hypothesis in animal models. In
spontaneously hypertensive rats, Asian ginseng inhibited the development of an increased SBP
(Hong et al., 2012; Pan, et al., 2012; Yanai, et al., 2006;), whereas treatment with Asian ginseng
had no effect on the AP of normotensive rats (Yanai, et al., 2006).
Although the majority of literature suggests a beneficial effect of Asian ginseng, there is
suggestion that Asian ginseng is associated with hypertension (Martinez-Mir et al., 2004). A case
study of one elderly man who had been taking Asian ginseng experienced transient ischemic
attacks and hypertensive crisis. Following the cessation of daily Asian ginseng, he did not
experience another attack. However, without further investigation, it is difficult to apply
causality in this event (Martinez-Mir et al., 2004).
1.6.2.2 Potential Anti-hypertensive Mechanisms of Asian Ginseng:
In spontaneously hypertensive rats, the total amount of endothelial nitric oxide synthase
(eNOS) and Akt protein expression was not different between Asian ginseng treated and control;
however, phosphorylation of eNOS and Akt was increased in animals treated with Asian
ginseng. This suggests that the AP lowering effect of ginseng may partially depend on the non-
transcriptional phosphatidylinositol 3-kinase (PI3K)/Akt signalling pathway. In addition, ginseng
treatment reduced aortic wall thickness and intercellular adhesion molecule 1 (ICAM-1) protein

33
in spontaneously hypertensive rats. It should be noted that a higher AP is associated with
increased aortic thickness and ICAM-1 has been shown to increase the progression of
atherosclerosis (Hong et al., 2012).
1.6.2.3 Korean Red ginseng has anti-hypertensive effects:
In contrast to the proposed effects of ginseng according to TCM, there is evidence that
Korean Red ginseng has hypotensive effects in both normotensive and hypertensive animals
(Jeon et al., 2000A, 2000B). This hypotensive effect has been confirmed in clinical trials in
patients suffering from essential hypertension (Han et al., 1998; Sung et al., 2000). However,
Korean Red ginseng had no effect in patients suffering from white coat hypertension (Han et al.,
1998). In addition, Korean Red ginseng in conjunction with patients’ existing anti-hypertensive
medications had no effect on SBP, but did decrease DBP (Rhee et al., 2011).
1.6.2.4 The potential anti-hypertensive mechanisms of Korean Red ginseng:
It has been suggested that Korean Red ginseng exerts a hypotensive effect caused by the
release of endothelial NO, a vasodilator (Jeon et al., 2000A). Furthermore, there is evidence to
suggest that Korean Red ginseng may improve vascular endothelial function in hypertensive
patients (Sung et al., 2000).
1.6.3 The effect of NAG on Metabolic Variables:
The American Botanical Council Clinical Guide to Herbs reports that there are no known
adverse effects for NAG (Blumenthal et al., 2003). It notes that insulin levels may need to be
monitored in patients with diabetes, due to the potential blood sugar modulatory effects of NAG.

34
This guide also indicates that NAG is classified as a “Class 1 herb” according to The American
Herbal Products Association (AHPA) Safety Rating. This classification indicates that NAG “can
be safely consumed when used appropriately” (Blumenthal et al., 2003).
1.6.3.1 NAG has anti-diabetic properties:
NAG root has been reported to have anti-hyperglycemic activity (Quan et al., 2012B; Sen
et al., 2012A, 2012B; Xie et al., 2007, 2009). In addition, treatment with extracts made from
NAG berry, leaf, and associated polysaccharides also decrease blood glucose levels and
improved glucose tolerance (Xie et al., 2002B, 2004A, 2004B). It is interesting to note that
following the administration of polysaccharides isolated from NAG berry, fasting blood glucose
levels remained lower for 30 days following the cessation of treatment (Xie et al., 2004B). This
suggests that the polysaccharides, along with ginsenosides may be exerting these effects.
Furthermore, it has been observed that NAG decreased blood glucose, insulin, and HOMA-IR in
both Type 1 and Type 2 diabetic mice (Sen et al., 2012A).
The efficacy and safety of NAG on glucose metabolism have also been confirmed in
humans in double-blinded placebo-controlled studies (Vuksan et al., 2000). A dosage of NAG
prior to an oral glucose tolerance test lowered postprandial blood glucose in Type 2 diabetic and
non-diabetic patients (Vuksan et al., 2000, 2001). However, NAG did not alter insulin or glucose
in volunteers who had decreased insulin sensitivity as a result of receiving human
immunodeficiency virus (HIV)-protease inhibitors (Andrade et al., 2008).

35
1.6.3.2 The Anti-obesity effects of NAG:
The effects of NAG on body weight have not been well established. Several studies have
observed a decreased body weight following treatment with NAG root (Liu et al., 2010; Sen et
al., 2012A, 2012B; Xie et al., 2009). However, other studies have observed no change (Karaca et
al., 2010) or an increased body weight (Banz et al., 2007). Both NAG berry and leaf have been
shown to decrease body weight (Xie et al., 2002A, 2002B, 2004B, 2007), while a mixture of
stems and leaves have no effect (Liu et al., 2008). Furthermore, polysaccharides isolated from
NAG do not affect body weight (Xie et al., 2004B). In diabetic Zucker rats there was an increase
in body weight following treatment with NAG. Interestingly, this increase in body weight was
not associated with a corresponding increase in body lipids suggesting that it was an increase in
lean body mass (Banz et al., 2007).
NAG has been shown to decrease fat pad mass (Liu et al., 2008, 2010); however, NAG
appears not to alter food intake (Liu et al., 2010). The effect of NAG on lipid metabolism was
first reported over 20 years ago. It was observed that NAG reduced total cholesterol, LDL
cholesterol, and triglyceride levels in liver and serum of avians (Qureshi et al., 1983). However,
the total cholesterol reducing effects of NAG were not reproduced in rodents. Although
treatment with NAG did not affect total cholesterol levels, HFD-fed animals treated with PPD
ginsenosides had LDL cholesterol and plasma triglyceride levels comparable with normal diet
(ND) fed animals (Liu et al., 2010).
1.6.3.3 Potential Mechanisms responsible for the metabolic effects of NAG:
The anti-diabetic effects of NAG have been compared to the commercially available
insulin sensitizer, Rosiglitazone. It was observed that 4 weeks of oral administration of NAG

36
reduced blood glucose to a level comparable with the reduction present in Rosiglitazone-treated
diabetic animals. Furthermore, NAG and Rosiglitazone treated animals showed similar changes
of gene expression of PPAR-α and IRS. However, Rosiglitazone increased the expression of LPL
and uncoupling protein 3 (UCP3). NAG did not alter these genes, though an increased expression
of retinoid X receptor (RXR) was observed. RXR ligands can be effective in lowering blood
glucose. This suggests that the anti-diabetic of NAG and Rosiglitazone may occur via a similar,
yet distinct mechanism involving metabolism (Banz et al., 2007).
Oxidative stress is a critical pathogenic mechanism leading to diabetic complications.
Hyperglycemia in diabetic conditions may cause oxidative stress in pancreatic β-cells which may
lead to their dysfunction, in addition to insulin resistance in peripheral tissues. In β-cells exposed
to oxidative stress, exposure to NAG reduced the resulting oxidant injury. This suggests that
NAG possesses direct scavenging effects and the hypoglycemic properties of NAG may be
related to antioxidant effects in β-cells. (Lin et al., 2008). It has been suggested that NAG
improves β-cell insulin production. NAG increases ATP production, by regulating the level of
uncoupling protein 2 (UCP-2), thus increasing insulin production (Luo and Luo, 2006). In
addition, NAG has been proposed to protect β-cells from apoptosis by reducing caspase-9
activation and raising levels of β-cell lymphoma 2 (Luo and Luo, 2006). It has been reported that
NAG has anti-lipolytic effects on Type 2 diabetic conditions via a signalling pathway distinct
from insulin. This suggests that NAG not only affects pancreatic cells through increasing insulin
production and cell viability, but also through creating an anti-lipolytic effect and targeting
glucose receptors to counter hyperglycemia (Wang et al., 2006).
In Type 1 and Type 2 diabetic animals, there is an increased transcript of heme
oxygenase-1 and nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB),

37
molecular markers of oxidative stress. These increases were prevented in NAG treated animals
(Sen et al., 2012A, 2012B). In diabetes, glucose causes an up-regulation of extracellular matrix
proteins, while this is prevented with NAG treatment (Sen et al., 2012A). This study suggested
that NAG works through a combination of mechanisms involving anti-hyperglycemic and
antioxidant activities. In human studies, two weeks of NAG treatment was shown to reduce
markers of oxidative stress (Lee et al., 2008).
As mentioned, NAG treatment decreased cholesterol levels in avians. This was proposed
to occur though a mechanism related to the suppression of β-hydroxy-β-methylglutaryl-CoA
(HMG-CoA) reductase and cholesterol 7 α-hydroxylase activities (Qureshi et al., 1983). As
pancreatic lipase hydrolyzes 50-70% of total dietary fats, inhibition of this enzyme could
theoretically control obesity. Both in vitro and in vivo studies have shown that ginsenosides
isolated from NAG inhibited pancreatic lipase activity (Liu et al., 2008). Specifically, it was
shown that the pancreatic lipase inhibitory capabilities are due to the PPD ginsenosides
(example: G-Rb1), while the PPT ginsenosides had no effect (Liu et al., 2010). Furthermore,
NAG has been shown to increase body temperature, indicating a potential role in thermogenesis
(Xie et al., 2002, 2004B).
1.6.3.4 The metabolic effects of Ginsenoside Rb1:
There are data to suggest that G-Rb1 has anti-hyperglycemic and insulin-sensitizing
properties (Park et al., 2008; Shang et al., 2007; Xiong et al., 2011). Studies have shown that G-
Rb1 stimulates glucose transport in insulin sensitive cells (Shang, et al., 2007, 2008). This is
hypothesized to occur by promoting translocations of Glucose Transporter Type 1 (GLUT1) and
GLUT4 by partially activating insulin pathways (Shang et al., 2008). In addition, G-Rb1 has

38
been shown to enhance the expression of transcription factors PPARɣ and CCAAT-enhancer-
binding proteins (C/EBPs) thereby altering lipid and glucose metabolism, and promoting
adipogenesis (Shang et al., 2007). It has also been proposed that G-Rb1 promotes glucose-
stimulated insulin secretion via protein kinase A (PKA) that enhances insulin and insulin-like
growth factor 1 (IGF-1) signalling (Park et al., 2008). It has also been observed that G-Rb1
treatment decreased food intake and body weight (Xiong et al., 2010). However, it is postulated
that the anti-hyperglycemic effect of NAG is only partially attributed to these variables, as the
anti-hyperglycemic effects resulting from a G-Rb1 treatment were greater than effects seen in
pair-fed control rats (Xiong et al., 2010).
In HFD-induced obese rats, G-Rb1 initially reduced food intake, however after 24 h G-
Rb1 treated animals had food intake that was no different from control treated animals (Xiong et
al., 2010). Furthermore, G-Rb1 stimulated neuronal activity (as indicated by c-fos expression) in
areas of the brain related to energy homeostasis, particularly, in the ARC and VMH. In addition,
it was shown that G-Rb1 inhibited NPY expression in the hypothalamus (Xiong et al., 2010).
Correspondingly, body weight gain and body fat content were decreased following treatment,
and energy expenditure was increased (Xiong et al., 2010). However, in normal diet fed animals,
G-Rb1 did not alter food intake (Yang et al., 2010).
1.6.3.5 The metabolic effects of Ginsenoside Re:
G-Re is the second most prevalent ginsenoside in NAG. Studies suggest that G-Re is
responsible for the anti-diabetic effects of NAG (Attele et al., 2002; Lee et al., 2011B; Xie et al.,
2005A; Zhang et al., 2008). G-Re enhanced glucose uptake in 3T3-L1 adipocytes (Lee et al.,
2011B; Quan et al., 2012; Zhang et al., 2008). G-Re administration in diabetic (ob/ob) mice was

39
associated with improved blood glucose levels and a decrease in insulin levels. Furthermore,
NAG treatment yielded a greater than 2-fold increase in the rate of insulin-stimulated glucose
disposal. It is interesting to note that these effects remained up to three days following the
cessation of G-Re treatment (Xie et al., 2005).
G-Re has also been proposed to have anti-obesity effects (Attele et al., 2002; Cho et al.,
2006). Treatment with G-Re decreased plasma triglycerides and total cholesterol levels (Cho et
al., 2006; Quan et al., 2012). Another study suggested that treatment with G-Re yielded a
decreased body weight which was attributed to a decline in food intake, increased energy
expenditure, and a rise of body temperature (Attele et al., 2002). However, Xie and colleagues
reported that G-Re had no effect on body weight or body temperature (Xie et al., 2005A).
1.6.3.6 The Potential Mechanisms for the Effects of Ginsenoside Re:
As the anti-diabetic effects of G-Re have been well established, recent studies have
sought to explain the mechanisms of action. G-Re has been shown to activate insulin signalling
at IRS-1 and through PI3K. Furthermore, an increase in GLUT4 mRNA expression was seen
following treatment with G-Re (Lee et al., 2011B; Zhang et al., 2008). It has also been suggested
that G-Re decreases insulin resistance via the inhibition of c-Jun N-terminal kinase and NF-κB
activation (Zhang et al., 2008). In addition, it has been observed that G-Re supresses hepatic
glucose production by induction of an orphan nuclear receptor small heterodimer protein. This
protein is a regulator of cholesterol, lipid, and glucose metabolism (Zhang et al., 2008).
It is well known that diabetes is linked with oxidative stress. It is hypothesized that NAG
may protect against diabetes by adding to the total anti-oxidant defense system. Treatment with
G-Re restored levels of glutathione, an antioxidant (Liu et al., 2012). G-Re acts as an anti-

40
oxidant by scavenging hydrogen peroxide (H2O2) and hydroxyl radicals. Furthermore, NAG is
suggested to have stronger anti-oxidant activity than Asian ginseng, which may be attributed to
the fact that NAG contains more G-Re (Mehendale et al., 2006; Shao et al., 2004).
Although in animal models, G-Re has been shown to have anti-diabetic effects, in obese
and/or diabetic humans, oral administration of G-Re did not improve β-cell function or insulin
sensitivity (Reeds et al., 2012). However, in this clinical study, no ginsenosides were detectable
in the plasma, which perhaps suggests an inappropriate dosage.
Similarly to the suggested mechanisms of G-Rg1 and Korean Red ginseng, it is thought
that G-Re inhibits lipogenesis by suppression of SREBP-1c and the transcription of target genes:
FAS and stearoyl-CoA desaturase. These effects are thought to be mediated by AMPK, as the
effects of G-Re are abolished when cells were treated with an AMPK inhibitor (Quan et al.,
2012).
1.6.3.7 The Metabolic Effects of Compound K:
It is well-known that Compound K is the final metabolite of PPD ginsenosides (Shibata,
2001). It has been observed that insulin secretion was enhanced following Compound K
administration in diabetic (db/db) mice: there was an improvement in glucose tolerance with an
increase in insulin levels present 30 mins after a glucose challenge (Han et al., 2007).
Furthermore, and similarly to PPD ginsenosides, Compound K has been shown to decrease
fasting blood glucose levels in diabetic mice and prevent the destruction of islet cells (Han et al.,
2007). Additionally, Compound K shifted glucose metabolism from glucose production to
glucose utilization in the liver. Finally, an increase in plasma adiponectin levels was observed in

41
these animals which could play a role in the increased insulin sensitivity, as this hormone
controls glucose regulation (Han et al., 2007).
Compound K is suggested to have a synergistic effect with metformin, a common
medication for the treatment of diabetes. The simultaneous treatment with Compound K and
Metformin yielded greater improvements in plasma glucose, insulin, and HOMA-IR than
treatment with either alone (Yoon et al., 2007). In addition, treatment with Compound K led to a
downregulation of the expression of phosphoenolpyruvate carboxykinase and G6Pase. It is
proposed that the hypoglycemic and insulin sensitizing effects of ginseng were caused by the
downregulation of these gluconeogenesis genes in the liver (Li et al., 2012).
1.6.4 The Effects of NAG on Cardiovascular Variables
1.6.4.1 The Effects of NAG on Arterial Pressure:
Several clinical trials in hypertensive humans have sought to clarify the cardiovascular
effects of NAG with respect to AP. It has been suggested that NAG exerts a neutral effect on
blood pressure in both acute (Stavro et al. 2005) and long-term trials in patients with essential
hypertension (Stavro et al., 2006). However, it should be noted that these patients continued
with their anti-hypertensive medications throughout the study, which may be a confounding
variable. An earlier study suggests NAG treatment in Type 2 Diabetic humans decreased both
SBP and DBP (Xu, 2000). In animal studies, diabetic animals exhibited a decreased stroke
volume and cardiac output. In diabetic animals treated with NAG, stroke volume and cardiac
output were no different than non-diabetic animals (Sen et al., 2012B).

42
1.6.4.2 Potential Mechanisms Responsible for the Cardiovascular Effects of NAG:
Essential hypertension is a multifactorial disease that is associated with metabolic
disorders: there is a strong relationship between serum metabolic profiles and AP. For example,
the amino acids serine, threonine, and ornithine are correlated inversely with hypertension.
Following administration of an extract composed primarily of G-Rb1, the levels of these amino
acids increased in spontaneously hypertensive rats. Furthermore, the fatty acids oleic acid and
palmitic acid are directly correlated with hypertension, and were decreased following treatment
with G-Rb1 rich extract. The G-Rb1 based extract treatment corrected metabolic profiles
associated with hypertension, whereas anti-hypertensive medications, though having greater anti-
hypertensive effects, do not correct the serum metabolic profiles (Aa, et al., 2010).
G-Rb1 induces NO generation and has been shown to induce vasodilation in a dose-
dependent manner. It is hypothesized that the vasodilatory effects of NAG occur through
increased phosphorylation of eNOS, as the vasodilatory effects of G-Rb1 were blocked with pre-
incubation with both eNOS and PI3K inhibitors (Pan, et al., 2012). In a similar mechanism to G-
Rg1, G-Rb1 is thought to activate NO by modulating the PI3K/Akt/eNOS pathway and by
increasing l-arginine transport in endothelial cells (Pan, et al., 2012). Additionally, G-Re, the
second most prevalent ginsenoside in NAG, has been shown in clinical studies to induce
vasorelaxation and have anti-oxidant effects (Chen, 2008).
1.7 Rationale and Hypothesis
Asian ginseng has been studied in great detail and the data obtained in these studies
suggest that Asian ginseng possesses anti-diabetic and anti-obesity effects (Jeon, et al., 2000A;
Kim et al., 2010; Lee, et al., 1981; Yanai, et al., 2006; Yang et al., 2010). NAG contains 3-5

43
times greater ginsenoside content than Asian ginseng and there are considerable data suggesting
that G-Rb1 and G-Re, the main ginsenosides of NAG, have beneficial effects in animal models
of obesity and diabetes. However, the effect of NAG on a number of metabolic variables and the
neuronal pathways involved in metabolism and cardiovascular regulation has not been
investigated.
The focus of this thesis was to investigate the effects of NAG on markers of the
metabolic syndrome in vivo. Specifically, physiological parameters such as changes in body
weight, food intake, fat pad mass, specific organ weight, and plasma leptin, adiponectin,
cholesterol, triglycerides, blood glucose, insulin, noradrenaline, and angiotensin II, along with
chronic AP and HR were evaluated. In addition, this study investigated the location, distribution,
and phenotype of central neurons activated in response to NAG treatment. Specifically,
structures that may be involved in mediating the cardiovascular and metabolic effects of the
metabolic syndrome in the brainstem and hypothalamus were investigated. These neuronal
systems were identified using the immunohistochemical detection of the immediate early gene
product Fos-related antigen-1/2 (Fra-1/2)-like proteins (Curran and Franza, 1988; Rauscher et al.,
1988; Shaulian and Karin, 2002). The immediate early gene c-fos has been implicated as one of
the third messengers in the intracellular signal transduction pathway in a variety of cells
including neurons (Morgan et al., 1987, 1990). The basal expression of Fos-related antigen
proteins are relatively low, but can be induced rapidly and transiently by growth factors,
neurotransmitters, second messengers, or membrane depolarization (Lohmeier et al., 2003;
Morgan et al., 1987, 1990; Sagar and Wong, 1988; Solano-Flores et al., 1997).

44
It was hypothesized that chronic NAG treatment reduces the risk of developing the
metabolic syndrome by activating central neuronal systems associated with metabolic and
cardiovascular regulation.

45
Chapter 2: Materials and Methods
2.1 Ginseng Production
Four year old NAG (Panax quinquefolius) roots were provided by the Ontario Ginseng
Growers Association (OGGA). The roots were collected in 2007 from five farms in Ontario,
Canada. Dried root samples were shipped to Naturex (South Hackensack, NJ) to extract the
ginsenosides using a hydroalcoholic process. Ginseng extracts were prepared from each farm and
combined to produce a composite extract (Sen et al., 2012A). In brief, ground ginseng roots were
soaked three times for five hours in an ethanol/water (75:25 v/v) solution at 40°C. The excess
solvent was removed under vacuum at 45°C after the extract was filtered. The extract was
concentrated again until the total solids on a dry basis yielded roughly 60%. These concentrates
were lypophilized at the Ontario Ginseng Innovation and Research Consortium (OGIRC) central
lab at University of Western Ontario in order to make a powder alcoholic ginseng extract to be
used in this study.
The final extract was subject to analysis via high performance liquid chromatography
(HPLC) in order to determine the presence of ginsenosides. HPLC analysis of the extract has
been previously described (Azike et al. 2011). In brief, a Waters 1525 HPLC system with binary
pump and ultraviolet detector and a reversed-phase Inspire C18 column were used (100 mm ×
4.6 mm, i.d. 5 μm) (Dikma Technologies, CA, USA). The gradient program consisted of [A]
water and [B] acetonitrile at a flow of 1.3 ml/min, as follows: 0 min: 80–20%, 0–60 min: 58–
42%, 60–70 min: 10–90%, 70–80 min: 80–20%. The elutes were monitored at 203 nm. Overall,
the extract showed 28.25% ginsensoside content. Major ginsenosides included G-Rb1 and G-Re
while minor ginsenosides included G-Rb2, G-Rg1, G-Rd, and G-Rc. The Rb1/Rg1 ratio was
found to be 56.74%. (Azike, et al. 2011; Guo, et al., 2010; Sen, et al. 2011, 2012,).

46
2.2 Animal Care
Adult, male, Sprague-Dawley rats (8 weeks; 245-327 g; n= 57) were obtained from
Charles River Canada (Saint-Constant, Quebec, Canada). All animals were housed individually,
and allowed a 1-week acclimatization period upon arrival at the facility which was kept at 23 °C
and 65 % humidity with a 12 h light/dark cycle beginning at 0700 h. Animals were fed either a
standard chow (n= 34) or high fat diet (HFD) (45% kcal fat; n= 23) (Research Diets, NJ, U.S.A.)
and water ad libitum. This study was done in accordance with the guidelines set out by the
Canadian Council on Animal Care and The Animal Care Committee at the University of Western
Ontario.
2.3 Treatment
Animals were divided into four treatment groups: normal diet with vehicle control
treatment (NDC; n= 16), normal diet with a treatment of an alcoholic extract of NAG (NDG; n=
18), high fat diet with vehicle control (HFC; n= 11), and high fat diet with treatment of an
alcoholic NAG extract (HFG; n= 12).
A 250 mg/kg of body weight dose of NAG (alcoholic extract dissolved in 0.5ml 0.9%
saline) or a vehicle control (0.5 ml 0.9% saline) was administered daily by oral gavage before the
onset of the active cycle (1900 h). This continued for 28 days and body weight was recorded
daily.
2.3.1 Cardiovascular measurements:
MAP and HR were measured non-invasively by determining the tail blood volume with a
volume pressure recording sensor and an occlusion tail-cuff (CODA System, Kent Scientific,

47
Torrington, CT) (Feng et al., 2008; Sugiyama et al., 2001, 2002). Volume pressure recording is
clinically validated and provides 99% correlation in SBP with telemetry and direct blood
pressure measurements, and a slightly lower correlation in DBP (Feng et al., 2008). Animals
were allowed a training week to acclimatize to the measurement procedure and environment
followed by measurements to collect the blood pressure data for analysis (Sugiyama et al., 2001,
2002).
2.3.2 Metabolism study:
Twenty four hour measurements of food and water intake were taken prior to the first day
of treatment and after 28 days of treatment. In addition, urine was collected over a 24 h period.
This was done before treatment began and after the 28 day treatment period.
2.4 Tissue collection
Animals were fasted for 12 h prior to sacrifice after 28 days of NAG treatment. In all
animals, blood was taken intracardially (5 ml) with 7% EDTA. Fasting blood glucose was
measured immediately using a glucometer (Aviva Accu-Chek; Roche Diagnostics GmbH,
Mannheim, Germany). Plasma was separated by centrifugation at 10,000 rpm. at 4°C for 10 min.
Plasma was frozen at -80°C until the time of analysis. The heart, kidneys, retroperitoneal, and
epididymal fat pads were removed and weighed. In addition, the tibia was removed and organ
weight was standardized to tibia length.

48
2.5 Biochemical assays
2.5.1 Plasma Leptin Quantitation
Plasma leptin levels were measured using an enzyme immunoassay (EIA) kit (leptin EIA kit,
Enzo LifeSciences, Burlington, Ontario), an immunometric EIA that permits leptin
measurements up to 67.2 pg/ml.and with an inter- and intra-assay coefficients of variation (CVs)
of < 11%. Double distilled water was used to prepare the reagents and samples. All samples were
measured in duplicate according to the manufacturer's instructions.
2.5.2 Plasma Insulin Quantitation
Insulin quantitation was performed using a rat specific enzyme-linked immunosorbent assay
(ELISA) (ALPCO Diagnostics, Salem, NH). The sensitivity of this immunoassay was 0.124
ng/ml and with inter- and intra-assay CVs of < 10%. All samples were measured in duplicate
and the assay and analyses were performed according to the instruction of the manufacturer.
Systemic insulin levels were expressed as units per ml of plasma.
2.5.3 Plasma Adiponectin Quantitation
Endogenous plasma adiponectin was measured by ELISA (Phoenix Pharmaceuticals,
Belmont, CA, USA) in accordance with the manufacturer’s instructions. All samples were
measured in duplicate. This assay permits adiponectin measurements up to 8 ng/ml. The
sensitivity of this immunoassay was 0.12 ng/ml with inter- and intra-assay CVs of < 10%.

49
2.5.4 Plasma Triglycerides and Total Cholesterol Quantitation
Plasma triglycerides and total cholesterol levels were measured on a Cobra Mira S
autoanalyzer using reagents and controls from Roche Diagnostics and calibrators from National
Institute of Standards and Technology. Precision testing was done by Digital P and Health Metrx
(Vancouver, Canada).
2.5.5 Plasma Noradrenaline Quantitation
Plasma levels of noradrenaline were determined using a high sensitivity ELISA kit (Rocky
Mountain Diagnostics, Colorado Springs, Colo). The minimum detectable concentration of
noradrenaline was 2.7 pg/ml with inter- and intra-assay CVs of < 15%. All samples were
measured in duplicate according to the manufacturer's instructions.
2.5.6 Plasma Angiotensin II Quantitation
Plasma levels of angiotensin II were determined using a competitive EIA in which a
biotinylated Angiotensin II peptide competes with the peptide in standard solution or samples
(Phoenix Pharmaceuticals Inc., Belmont CA). All samples were measured in duplicate according
to the manufacturer's instructions. Cross-reactivity of this kit antibody to Angiotensin II(1–4),
Angiotensin II(5–8) and Angiotensin I was 0.1, 0.001 and 1.7%, respectively. The lowest
detection limit of this immunoassay was 31 pg/ml with inter- and intra-assay CVs of < 10 %.
2.6 Immunohistochemistry
After 28 days of NAG or control treatment, animals were treated with a final dose of NAG or
vehicle 90 minutes prior to equithesin anesthesia (0.3 ml/100 g body weight, i.p.) followed by

50
perfusion with ice-cold 0.01 M phosphate-buffered saline (PBS, pH 7.2) and 4%
paraformaldehyde fixative. Brains were removed and stored overnight in fixative, followed by 24
h in 5% sucrose in PBS. The following day the brains were transferred to 10% sucrose in PBS
for 24 h, followed by 15% sucrose for 24 hour before processing the tissues for Fra-1/2 and/or
Tyrosine Hydroxylase (TH) immunohistochemistry.
Frozen serial, transverse sections of the brainstem and forebrain were cut at 50 µm on a
cryostat (-21°C). Sections were collected in PBS and after several washes in PBS were placed in
Fra-1/2 (polyclonal rabbit antibody; sc-253; Santa Cruz Biotechnology, Inc., Santa Cruz, CA)
diluted 1:2500 with PBS containing 0.3% Triton X-100 at 4°C for 48-72 h (Messenger et al.,
2012). Sections were placed in goat biotinylated anti-rabbit IgG (Vectastain; Vector
Laboratories, Burlington, Ontario, CAN) diluted in 1:500 PBS/0.3% Triton X-100 for 1 h.
Sections were processed using the avidin-biotin complex (ABC) reagent (Vectastain; Vector
laboratories, Burlington, ON, CAN) in PBS/0.3% Triton X-100 for 75 mins. Following ABC,
sections were set in a solution composed of 0.0006% hydrogen peroxide and 0.02% 3,3’-
diaminobenzidine tetrahydrochloride (DAB; Sigma) in PBS in order to visualize the peroxidase
reaction product (Ciriello et al., 2004; Ciriello, 1999). One of every three sections was mounted
on gelatin-coated slides, air dried, and DPX mountant was applied followed by a cover glass.
Adjacent brainstem sections processed for Fra-1/2 immunoreactivity were washed three
times for ten mins in PBS and placed in primary polyclonal antibody (1:1500) anti-TH (ab112;
AbCam, Cambridge, MA); and processed as described above. However, the TH-
immunoreactivity was visualized with Vector Blue (SK-5300, Vector Laboratories) according to
manufacturer’s instructions.

51
Two types of controls for immunoreactivity were used: omission of the primary antibody
and the pre-absorption of the antisera with an excess of respective antigen. Under these
conditions no immunoreactivity was demonstrated within the tissue sections. The antisera
employed in these studies have already been characterized for specificity (Callsen-Cencic and
Mense, 1999; Guillod-Maximin et al., 2004; Guo et al., 2005).
2.7 Data Analysis
Brainstem and forebrain sections were analyzed and photographed using bright-field
microscopy (Leitz Diaplan microscope). Fra-1/2 labeled neurons at x200 magnification in
different medullary and hypothalamic structures were counted by two independent investigators.
The numbers were expressed as the average number of labeled neurons per section. The
stereotaxic atlas of the rat brain of Paxinos and Watson (1986) was used to identify brain
structures. Specifically, the medullary areas in which neurons were counted included the area
postrema (ap), NTS, CVLM, RVLM, dorsal motor nucleus of the vagus (DMV), raphé pallidus
(RPa), magnocellular reticular nucleus (MARN), and the A5 noradrenergic cell group (A5). In
addition, neurons were counted in the following areas of the forebrain: ARC, PVH, dorsomedial
nucleus of the hypothalamus (DMH), lateral hypothalamus (LH), supraoptic nucleus (SON),
subfornical organ (SFO), organum vasculosum lamina terminalis (OVLT), and median preoptic
nucleus (MnPO). Digital images of neurons within these structures were obtained with a Nikon
DS-Fil camera and NIS Elements Basic Research 3.0 software (Nikon Canada, Mississauga, ON,
Canada).
Statistical analysis was performed using a one-way analysis of variance (ANOVA), or
two-way ANOVA where appropriate. A Bonferroni post-hoc test was done when ANOVA

52
indicated statistical significance. P values < 0.05 were considered to be significant. Results are
presented as mean ± standard error of the mean (SEM). Analyses were performed with Prism
GraphPad statistical software (version 5.0 for Windows).

53
Chapter 3: Results
3.1 The effect of NAG on markers of the metabolic syndrome
3.1.1 The effect of NAG treatment on Body Weight, Caloric Intake, and Urine output:
Body weight at the beginning of the study for all animals was 293 ± 4.5 g. At the end of
the study, NAG animals had a lower body weight (428 ± 10 g) when compared with vehicle
treated animals (440 ± 10 g). HF animals had higher body weight compared with ND animals (p
< 0.05), and NAG treated animals had a lower body weight (489 ± 16 g) as compared with
vehicle (518 ± 13 g) and NAG treated animals (Fig. 3.1).
In addition, cumulative body weight gain in animals on a ND (50 ± 5 g) or HFD (63 ± 3
g) was lower in animals receiving NAG treatment (42 ± 4 g, 50 ± 4 g; NDC and HFC
respectively) after 7 days of treatment (Fig. 3.2a). After the 28 day treatment period, animals fed
a HFD and treated with saline (vehicle) gained significantly (p < 0.05) more weight (207 ± 11 g)
than animals fed HFD and treated with NAG (171 ± 9 g). Similarly, animals fed the ND and
treated with vehicle gained more weight (155 ± 9 g) than NAG treated animals (147 ± 9 g) (Fig.
3.2d). HFD animals treated with NAG gained weight comparable with animals on ND. These
data are summarized in Figure 3.2.
Food caloric intake at the beginning of the study for all animals was 99 ± 2 calories
(cals). At the end of the study, animals on a HFD that received the vehicle (117 ± 9 cals) or NAG
treatment (123 ± 6 cals) had higher (p < 0.05) food intake compared with animals fed a normal
diet. Animals fed a normal diet in conjunction with NAG (98 ± 18 cals) or vehicle treatment (105
± 18 cals) consumed the same amount of calories after 28 days of treatment as initial values.

54
Figure 3.1. The effect of North American ginseng (NAG) on body weight after 28 days of
treatment. Following 28 days of HFD and/or NAG treatment, body weight was higher in all
animals as compared with before treatment. Animals fed a HFD had elevated body weight
compared with ND fed animals. Statistical significance was determined using one-way ANOVA
and two-way ANOVA followed by a Bonferroni post-hoc test. Data are presented as mean ±
SEM. *, p < 0.05 between diet groups. #, p < 0.05 within diet group.
# #

55
Figure 3.2. The effect of North American ginseng (NAG) treatment on cumulative change
in body weight. Measurements were taken at 7 days (a), 14 days (b), 21 days (c), and 28 days
(d). Daily North American Ginseng (NAG) treatment (250mg/kg) decreased weight gain after 7
days (a) of treatment in both normal diet fed and high fat fed animals (# p < 0.05). This
suppression of body weight gain continued throughout the 28 day treatment period (b,c,d).
Animals fed a high fat diet gained more weight than animals fed a normal chow diet (* p < 0.05)
after 14 days of high fat diet. This increased body weight continued throughout the study (c,d).
Statistical significance was determined using a two-way ANOVA followed by a Bonferroni post-
hoc test. Data is presented as mean ± SEM *, p < 0.05 between diet groups. #, p < 0.05 within
diet groups.
a
c
b
d

56
After the 28 day treatment period, animals on a ND that received the vehicle drank
amounts (39 ± 0.5 ml) that were not different from the values following NAG treatment (44 ± 6
ml). Similarly, HFC animals drank (39.6 ± 3 ml) the same amount as NAG treated (36.6 ± 2.2
ml) HFD animals. Furthermore, compared with initial values (15 ± 0.5 ml) there were no
differences in urine output in animals following 28 days of vehicle treatment (13.9 ± 4.5 ml, 23.8
± 5 ml; NDC and HFC respectively) or NAG treatment (14.6 ± 5.5 ml, 17.2 ±3.5 ml; NDG and
HFG respectively). These data are summarized in Table 3.1.
3.1.2 The effect of NAG treatment on lipids:
After the 28 days treatment period, heart and kidney weights of all treatment groups were
not different from one another. This data are summarized in Table 3.2. However, there was a
decreased fat pad mass following NAG treatment. In ND animals, epididymal fat pad mass
(0.122 ± 0.06 g/mm tibia length) was decreased following NAG treatment (0.098 ± 0.34 g/mm
tibia length). Similarly, in HFD animals (0.297 ± 0.114 g/mm tibia length), epididymal fat was
lower in NAG treated animals (0.220 ± 0.089 g/mm tibia length) (Fig. 3.3a). Retroperitoneal fat
pad mass was also decreased following NAG treatment (0.126 ± 0.046 g/mm tibia length, 0.315
± 0.103 g/mm tibia length; ND-NAG and HF-NAG respectively) as compared with vehicle
treatment (0.166 ± 0.080 g/mm tibia length, 0.397 ± 0.144 0.166 ± 0.080 g/mm tibia length;
NDC and HFC respectively) (Fig. 3.3b).
Plasma leptin concentrations were significantly (p < 0.05) higher (10.1 ± 2.0 ng/ml) in
animals fed a HFD as compared with ND fed animals (4.7 ± 0.54 ng/ml). Leptin concentrations
were significantly (p < 0.05) lower after 4 weeks of NAG treatment (2.6 ± 0.4 ng/ml, 6.3 ± 1.1
ng/ml; NDG and HFG respectively) (Fig. 3.4a). Plasma adiponectin levels were not different

57
Table 3.1. The effects of North American ginseng (NAG) treatment on food and water
intake and urine output. Measurements were taken over a 24 hour period. Animals fed a high
fat diet had increased (p < 0.05) caloric intake compared with animals fed a normal chow diet.
North American Ginseng (NAG) treatment had no effects on food and water intake or urine
output. Values are expressed as mean ± SEM. Data were analyzed with a one-way ANOVA. * p
< 0.05 between diets. NDC, normal diet control treatment; NDG, normal diet with North
American Ginseng treatment; HFC, high fat diet control treatment; HFG, high fat diet North
American Ginseng treatment.
Before
After 28 days of treatment
NDC
(n= 8)
NDG
(n= 10)
HFC
(n= 6)
HFG
(n=5)
Caloric intake after
28 days of treatment
(calories)
99 ± 2 105 ± 9 98 ± 18 117 ± 9* 123 ± 6*
Water intake after
28 days treatment
(ml)
36.5 ± 1 39 ± 0.5 44 ± 6 39.6 ± 3 36.6 ± 2.2
Urine output after
28 days of treatment
(ml)
15 ± 0.5 13.9 ± 4.5 14.6 ± 5.5 23.8 ±5 17.2 ± 3.5
Table 3.2. The effect of North American ginseng (NAG) on organ weights after 28 days of
treatment. Values are expressed standardized to length of tibia (mm). Both heart and kidney
weights were the same across all groups. Data is expressed as mean ± SEM. NDC, normal diet
control treatment; NDG, normal diet with North American Ginseng treatment; HFC, high fat diet
control treatment; HFG, high fat diet North American Ginseng treatment
Organ
NDC
(n= 8)
NDG
(n= 10)
HFC
(n= 6)
HFG
(n=5)
Heart (wet weight
in grams/ tibia
length mm)
0.031 ± 0.003 0.031 ± 0.004 0.031 ± 0.013 0.031 ± 0.002
Kidneys (wet
weight in grams/
tibia length mm)
0.088 ± 0.14 0.082 ± 0.14 0.079 ± 0.012 0.065 ± 0.30

58
Figure 3.3. The effects of North American Ginseng (NAG) on fat pad mass. After 4 weeks of
high fat diet feeding, fat pad mass was higher in both retroperitoneal (a) and epididymal (b) fat
pads (* p < 0.05). After 4 weeks of treatment with NAG, fat pad mass was lower in fat pads
(a,b). Fat pad mass is standardized to tibia length (mm). Data are presented as mean ± SEM. *, p
< 0.05 between diet groups. #, p < 0.05 within diet groups.
#
#
#
#
*
*

59
Figure 3.4 The effect of North American Ginseng (NAG) on plasma leptin and adiponectin
levels. Following 28 days of high fat diet plasma leptin concentrations were higher as compared
with normal diet fed animals (a) (* p < 0.05). Leptin concentrations were lower in NAG treated
animals compared with vehicle (# p < 0.05). After 28 days of treatment with NAG, plasma
adiponectin levels were not different in NAG treated animals fed a normal diet or high fat diet
compared with control. Significance was determined using a 2-way ANOVA followed by a
Bonferroni post-hoc test. Data is presented as mean ± SEM. *, p < 0.05 between diets; #, p <0.05
within diets.
#
#
*

60
between control (2.8 ± 0.76 ng/ml) and NAG treated animals (2.9 ± 0.46 ng/ml) fed a ND or
HFD (2.9 ± 0.39 ng/ml, 3.3 ± 0.65 ng/ml; vehicle and NAG treated respectively) (Fig. 3.4b).
NAG lowered plasma triglycerides in animals on a ND (0.30 ± 0.02 mmol/L) and HFD
(0.28 ± 0.05 mmol/L) as compared to vehicle (0.38 ± 0.04 mmol/L, 0.38 ± 0.04 mmol/L; NDC
and HFC respectively (Figure 3.5a). Neither HFD (1.46 ± 0.13 mmol/L) nor NAG treatment
(1.40 ± 0.06 mmol/L, 1.53 ± 0.16 mmol/L; NDG and HFG respectively) altered total cholesterol
levels as compared with control animals (1.44 ± 0.28 mmol/L) (Fig. 3.5b).
3.1.3 The effect of NAG on glucose homeostasis:
Animals treated with NAG (0.67 ± 0.05 ng/ml, 0.51 ± 0.18 ng/ml; NDG and HFG
respectively) had lower fasting insulin levels than HFD (1.25 ± 0.07 ng/ml) or ND (1.13 ± 0.27
ng/ml) vehicle-treated animals (Figure 3.6a). HFD fed animals had higher fasting blood glucose
levels than ND fed animals (Figure 3.6b). Similarly, following NAG treatment, both ND (0.34 ±
0.03) and HFD (0.27 ± 0.08) animals had a decreased insulin resistance compared with vehicle
treated animals (0.67 ± 0.17, 0.76 ± 0.18; ND and HFD respectively) (Fig. 3.6c).
3.1.4 The effect of NAG treatment on MAP and HR:
MAP and HR at the beginning of the study for all animals were 107 ± 2 mmHg and 394 ±
4 bpm, respectively. The MAP for animals throughout the study is presented in Table 3.3. After
the 28 day treatment period, animals on a ND that received the vehicle had MAP (109 ± 2
mmHg) and HR (380 ± 7 bpm) that were not different from the values at the onset of the study
(Fig. 3.7). Similarly, MAP (113 ± 2 mmHg) and HR (386 ± 5 bpm) in animals on a ND and
received NAG treatment were not different from the levels at the onset of the study (Fig. 3.7). On

61
Figure 3.5. The effect of North American Ginseng (NAG) on plasma lipids. After 28 days of
treatment with NAG, plasma triglyceride levels were lower in NAG treated animals fed both a
normal diet and high fat diet compared with vehicle (a). After 28 days of high fat diet and/or
NAG treatment, total cholesterol was not different in any treatment group (b). Data are presented
as mean ± SEM. Statistical significance was determined with a two-way ANOVA followed by a
Bonferroni post-hoc test. *, p < 0.05 between diets.

62
Figure 3.6. The effect of North American Ginseng (NAG) on glucose homeostasis. After 28
days of treatment with NAG, plasma insulin concentration was lower in animals fed normal and
high fat diet compared to vehicle (a) (# p < 0.05). After 28 days, animals fed a high fat diet had
elevated fasting blood glucose levels (b) (* p < 0.05). NAG had no effect on fasting blood
glucose levels (b). The homeostatic model assessment of insulin resistance (HOMA-IR) shows a
decreased insulin resistance following 28 days of NAG treatment on a normal and high fat diet
compared with control (c). Significance was determined using a two-way ANOVA followed by a
Bonferroni post-hoc test (* p < 0.05). Data is presented as mean ± SEM. *, p < 0.05 within diet
groups.
# #
# #
*

63
Table 3.3 The effect of North American Ginseng (NAG) on mean arterial pressure (MAP)
throughout the treatment period. There were no changes in MAP during week 1 or week 2 in
any treatment group. Following 3 weeks of NAG treatment, MAP levels were decreased
compared with vehicle control treated animals. MAP is expressed in millimetres of Mercury
(mmHg). Significance was determined using a Two-way ANOVA (# p < 0.05). Data are
presented as mean ± SEM. #, p < 0.05 within diet groups. NDC, normal diet control treatment;
NDG, normal diet with North American ginseng treatment; HFC, high fat diet control treatment;
HFG, high fat diet with North American ginseng treatment.
Treatment MAP (mmHg)-
Week 1
MAP (mmHg)-
Week 2
MAP (mmHg)-
Week 3
NDC (n=6) 104 ± 1 104 ± 2 107 ± 2
NDG (n=6) 103 ± 2 99 ± 1 100 ± 1#
HFC (n=4) 104 ± 2 102 ± 2 106 ± 1
HFG (n=4) 102 ± 3 101 ± 3 98 ± 3#

64
Figure 3.7. The effect of North American Ginseng (NAG) on mean arterial pressure and
heart rate after 28 days of treatment. After 28 days of high fat diet feeding, diet-induced
overweight rats had higher MAP compared with all other treatments. NAG treated groups and
normal chow fed rats had MAP comparable with MAP before the treatment period. (a).
Similarly, high fat diet-fed overweight rats experienced a higher heart rate, whereas NAG treated
high fat diet fed rats and normal chow-fed animals did not (b). Treatment groups were analyzed
using a one-way ANOVA. Data is presented as mean ± SEM. * p < 0.05.
b

65
the other hand, after the 28 days, HFD animals had MAP (120 ± 2 mmHg) and HR (414 ± 4
bpm) that were significantly (p < 0.05) higher than the initial levels at the onset of the study, and
significantly (p < 0.05) higher than those observed in both NDC and ND-NAG animals.
However, animals fed a HFD and given NAG treatment had MAP (107 ± 2 mmHg) and HR (386
± 4 bpm) that were not different from initial values at the onset of the study, from the values in
the NDC and NDG animals after 28 days of treatment, and were significantly (p < 0.05) lower at
the end of the treatment period than those in the HFD animals that received the vehicle only (Fig.
3.7). In addition, HFD animals had SBP (146 ± 3 mmHg) and DBP that were significantly
different from ND fed animals and NAG treated animals (Fig. 3.8) Furthermore, HFD animals
treated with NAG had significantly lower DBP (80 ± 2 mmHg) as compared with other treatment
groups (Fig. 3.8).
3.1.5 The effect of NAG treatment on plasma Noradrenaline and Angiotensin II levels:
Plasma noradrenaline level in animals on a ND (2.9 ± 0.26 ng/ml) was significantly lower
in animals receiving NAG treatment (2.1 ± 0.25 ng/ml). Noradrenaline levels were significantly
higher (p<0.05) in animals fed a HFD. However, animals fed a HFD and treated with the vehicle
had significantly higher noradrenaline levels (5.6 ± 1.3 ng/ml) than animals fed HFD and treated
with NAG (3.2 ± 0.50 ng/ml) (Fig. 3.9).
Following 28 days of NAG treatment, plasma angiotensin II levels were significantly (p
< 0.05) higher in both NDG (0.70 ± 0.04 ng/ml) and HFG (0.71 ± 0.06 ng/ml) as compared with
vehicle control (0.59 ± 0.02 ng/ml, 0.65 ± 0.04 ng/ml; NDC and HFC respectively) (Fig. 3.9).

66
Figure 3.8. The effect of North American Ginseng (NAG) on systolic and diastolic blood
pressure after 28 days of treatment. After 28 days of high fat diet, diet-induced overweight
rats had higher systolic blood pressure compared with all other treatments. NAG treated groups
and normal chow fed rats had systolic blood pressure comparable with before the treatment
period. (a). Similarly, high fat diet-fed overweight rats experienced an higher diastolic blood
pressure, whereas NAG treated high fat diet fed rats experienced a lower diastolic blood
pressure (b). Treatment groups were analyzed using a one-way ANOVA. Data are presented as
mean ± SEM. * p < 0.05 .
b

67
Figure 3.9. The effect of North American Ginseng (NAG) treatment on plasma
Noradrenaline and Angiotensin II levels. After 28 days of treatment, animals fed a high fat
diet had higher plasma noradrenaline levels (a) (# p < 0.05). Daily NAG treatment (250mg/kg)
resulted in lower noradrenaline levels in both normal diet fed and high fat fed animals (* p <
0.05) (a). Following 28 days of NAG treatment, animals had higher plasma levels of angiotensin
II as compared with control (b). Statistical significance was determined with a two-way ANOVA
followed by a Bonferroni post-hoc test. Data are expressed as mean ± SEM *, p < 0.05 between
diet groups. #, p < 0.05 within diet groups.

68
3.2 Central Effects of NAG
3.2.1 The effect of NAG on brainstem regions
3.2.1.1 Fra-1/2 Expression in brainstem autonomic nuclei:
Induction of Fra-1/2 detected immunohistochemically in brainstem neurons was used as
a marker for neuronal activation. Table 3.4 summarizes the number of Fra-1/2 labeled neurons in
different brainstem autonomic nuclei in animals on ND and HFD after 28 days of either vehicle
or NAG treatment. A small number of neurons expressing Fra-1/2 like immunoreactivity were
found within the ap, NTS, DMV, CVLM, RVLM, RPa, MARN and A5 in animals on a ND
treated with saline (controls) (Table 3.4; Figs. 3.10, 3.11, 3.12). In contrast, in animals on a HFD
and similarly treated with saline, there was a greater than 2-fold higher number of neurons
containing Fra-1/2-like immunoreactivity in the NTS, CVLM, and RVLM, and a lower number
of neurons expressing Fra-1/2 in the MARN (Table 3.4; Figs. 3.10, 3.11, 3.12).
Treatment of animals (ND) with NAG resulted in an almost 2-fold higher number of
neurons expressing Fra-1/2 in the ap, NTS, CVLM, and RVLM (Table 3.4). On the other hand,
NAG treatment of animals on a HFD had a higher number of neurons containing Fra-1/2-like
immunoreactivity almost a 4-fold higher number in ap, NTS, and CVLM (Table 3.4; Figs. 3.10,
3.11, 3.12). In addition, there was a higher number of Fra-1/2 labeled neurons in RVLM in these
animals. In contrast, there was a lower number of labeled neurons in MARN.
Neither diet nor treatment of animals on their respective diets with NAG altered the
number of Fra-1/2 immunoreactive neurons in the DMV, RPa or the A5 region (Table 3.4).

69
Table 3.4. Table summarizing the number of neurons expressing Fra-1/2 immunoreactivity
in different brainstem regions. ND, normal diet; NDG, normal diet ginseng; HF, high fat diet;
HFG, high fat ginseng; Fra-1/2, Fos-related antigens 1/2; ap, area postrema; NTS, nucleus of the
solitary tract; CVLM, caudal ventrolateral medulla; RVLM, rostral ventrolateral medulla; DMV,
dorsal motor nucleus of the vagus; RPa, raphé pallidus; MARN, magnocellular reticular nucleus;
A5, A5 noradrenergic cell group; ND, normal diet control; NDG, normal diet with North
American ginseng treatment; HF, high fat diet control; HFG, high fat diet with North American
ginseng treatment.*, p<0.05 from non-ginseng group for each diet. *, p < 0.05 between diets; #, p
< 0.05 within diet.
Area
of brainstem
Number of Fra-1/2-labeled neurons/section
NDC
(n=5)
NDG
(n=5)
HFC
(n=5)
HFG
(n=6)
ap 13 ± 2 38 ± 4# 13 ± 2 47 ± 7 *
NTS 25 ± 4 35 ± 4 # 48 ±5 * 93 ± 7 * #
CVLM 10 ± 2 19 ± 2 # 16 ± 1 * 39 ± 5 * #
RVLM 16 ± 2 25 ± 3 # 31 ± 2 * 51 ± 4 * #
DMV 1 ± 1 1 ± 1 2 ± 1 3 ± 1
RPa 6 ± 1 6 ± 1 8 ± 1 7 ± 1
MARN 15 ± 2 10 ± 1# 11 ± 2* 11 ± 1*#
A5 23 ± 3 22 ± 3 21 ± 2 23 ± 3

70
Figure 3.10. Bright-field photomicrographs taken through the region of the caudal Nucleus
of the Solitary Tract (NTS) complex showing the distribution of Fra-1/2 labeled neurons
throughout the complex. Note the higher number of Fra-1/2 labeled neurons in the high fat fed
animal after NAG administration. Few neurons express Fra-1/2 in the other three groups of
animals. ap, area postrema; com, commissural subnucleus of NTS; Sdl, dorsolateral subnucleus
of NTS; Sg, commissural subnucleus of NTS; Sm, medial subnucleus of NTS; Slt, solitary tract;
12M, hypoglossal nucleus; ND, normal diet control; NDG, normal diet with North American
ginseng treatment; HF, high fat diet control; HFG, high fat diet with North American ginseng
treatment. Calibration mark of 200 µm in HFG applies to all.

71
Figure 3.11. Bright-field photomicrographs taken through the caudal ventrolateral medulla
(CVLM). Note that ginseng administration induces Fra-1/2 expression in a large number of
CVLM neurons in the animals fed a high fat diet, whereas animals on normal diet with or
without ginseng express few Fra-1/2 labeled neurons. MdV, Medullary reticular nucleus; LRN,
Lateral reticular nucleus; ND, normal diet control; NDG, normal diet with North American
ginseng treatment; HF, high fat diet control; HFG, high fat diet with North American ginseng
treatment. Calibration mark of 400 µm in the HFG photomicrograph applies to all.

72
Figure 3.12. Bright-field photomicrographs taken through the rostral ventrolateral
medulla (RVLM) showing the distribution of Fra-1/2 immunoreactive neurons. Note that
ginseng administration induces Fra-1/2 expression in a large number of RVLM neurons in the
animals fed a high fat diet, whereas animals on normal diet with (b) or without ginseng (a)
express few Fra-1/2 labeled neurons. AMB, nucleus ambiguous; LPGi, nucleus
paragiantocellularis; ND, normal diet control; NDG, normal diet with North American ginseng
treatment; HF, high fat diet control; HFG, high fat diet with North American ginseng treatment;
Fra-1/2, Fos-related antigens 1/2. Calibration mark of 400 µm in the HFG photomicrograph
applies to all.

73
3.2.1.2 Fra-1/2 Immunoreactivity in TH-Labeled neurons:
The number of TH immunoreactive neurons in the NTS/A2 region was higher in animals
on the HFD (47 ± 7 neurons/section) as compared to ND animals (31 ± 4 neurons/section).
Furthermore, the number of TH immunoreactive neurons was lower following NAG treatment in
both ND (15 ± 2 neurons/section) and HFD (30 ± 4 neurons/section). Similarly, in the
CVLM/A1 region, the total number of TH immunoreactive neurons was higher in HFD (25 ± 2
neurons/section) compared with ND (15 ± 1 neurons/section). Within the NTS/A2 (Table 3.5,
Fig. 3.13 and 3.14) and CVLM/A1 (Table 3.5, Figs. 3.15 and 3.16) there were significantly
lower (p < 0.05) number of TH-positive neurons that also expressed Fra-1/2 following NAG
treatment.
In the RVLM/C1 region, TH immunoreactive neurons were higher in the HFD vehicle
(28 ± 4 neurons/section) treated animals in ND and following NAG treatment (37 ± 3
neurons/section) (Table 3.5, Figs. 3.16 and 3.17). Furthermore, there was a significantly higher
number of TH immunoreactive neurons that also expressed Fra-1/2 (5 ± 3 neurons/section) in
HFD-NAG treatment as compared with HFD vehicle (2 ± 2 neurons/section). This data is
summarized in Table 3.5.
3.2.2 The effect of NAG on neuronal activity in forebrain structures
3.2.2.1 Fra-1/2 Expression in forebrain circumventricular organs:
Table 3.6 summarizes the number of Fra-1/2 labeled neurons in different forebrain
circumventricular nuclei in animals on ND and HFD after 28 days of NAG or vehicle treatment.
A small number of neurons expressing Fra-1/2 like immunoreactivity were found within the
MnPO, OVLT, and SFO, in animals treated with vehicle (control). (Table 3.6; Figs. 3.19, 3.20,

74
Figure 3.13. Bright-field photomicrographs showing examples of neurons in the A2 region
of the Nucleus of the Solitary Tract (NTS) complex, expressing both Fra-1/2 and TH
immunoreactivity. Note that many catecholaminergic neurons express both TH and Fra-1/2
immunoreactivity in the vehicle treatment. Following NAG treatment, note the large number of
non-catecholaminergic neurons, and very few neurons that express both TH and Fra-1/2
immunoreactivity. Fra-1/2 only labeled neurons (brown) and TH only labeled neurons (blue) in
all three areas. Inset in HFG corresponds to area (a) outlined in HFG showing examples of
double labelled neurons at greater magnification. Sm, medial subnucleus of NTS; Slt, solitary
tract; DMV, dorsal motor nucleus of the vagus, Fra-1/2, Fos-related antigens 1/2; TH, tyrosine
hydroxylase; ND, normal diet control; NDG, normal diet with North American ginseng
treatment; HF, high fat diet control; HFG, high fat diet with North American ginseng treatment.
Calibration mark of 50 µm in HFG applies to all.

75
Figure 3.14a. Camera lucida projection drawings of the Nucleus of the Solitary Tract
(NTS) in animals fed a normal chow
a

76
Figure 3.14b. Camera lucida projection drawings of the Nucleus of the Solitary Tract
(NTS) in animals fed a high fat diet. Representative projection drawings of the caudal nucleus
of the solitary tract (NTS) show distribution of Fra-1/2 labeled neurons in animals after high fat
diet (HFD) and/or North American ginseng (NAG) treatment. Fra-1/2 immunoreactive neurons
are indicated by the small black dots while those neurons immunoreactive to the
catecholaminergic enzyme TH are indicated by the larger open circles. Neurons immunoreactive
to both Fra-1/2 and TH are indicated by a star. Note the lower number of TH and Fra and TH
immunolabeling following NAG treatment. Furthermore, note the higher number of Fra
immunoreactive neurons following NAG treatment. Note that the majority of the Fra-1/2 labeled
cells were located in the Sm. ap, area postrema; com, commissural subnucleus of NTS; Sdl,
dorsolateral subnucleus of NTS; Sg, commissural subnucleus of NTS; Sm, medial subnucleus of
NTS; Slt, solitary tract; 12M, hypoglossal nucleus; ND, normal diet control; NDG, normal diet
with North American ginseng treatment; HF, high fat diet control; HFG, high fat diet with North
American ginseng treatment; Fra-1/2, Fos-related antigens 1/2; TH, tyrosine hydroxylase.
Calibration marks of 400 µm in the ND and HF drawings apply to all.
b

77
Figure 3.15. Bright-field photomicrographs showing examples of neurons A1 region of the
caudal ventrolateral medulla (CVLM) expressing both Fra-1/2 and TH immunoreactivity
(arrows). Note that in vehicle treatment many catecholaminergic neurons express both TH and
Fra-1/2 immunoreactivity, and there is a large number of non-catecholaminergic neurons
following NAG treatment. Fra-1/2 only labeled neurons (brown) and TH only labeled cells
(blue) in all three areas. Inset in HFG corresponds to area (a) outlined in HFG showing
examples of double labelled neurons at greater magnification. ND, normal diet control; NDG,
normal diet with North American ginseng treatment; HF, high fat diet control; HFG, high fat
diet with North American ginseng treatment; Fra-1/2, Fos-related antigens 1/2; TH, tyrosine
hydroxylase. Calibration mark of 50 µm in the HFG photomicrograph applies to all.
.
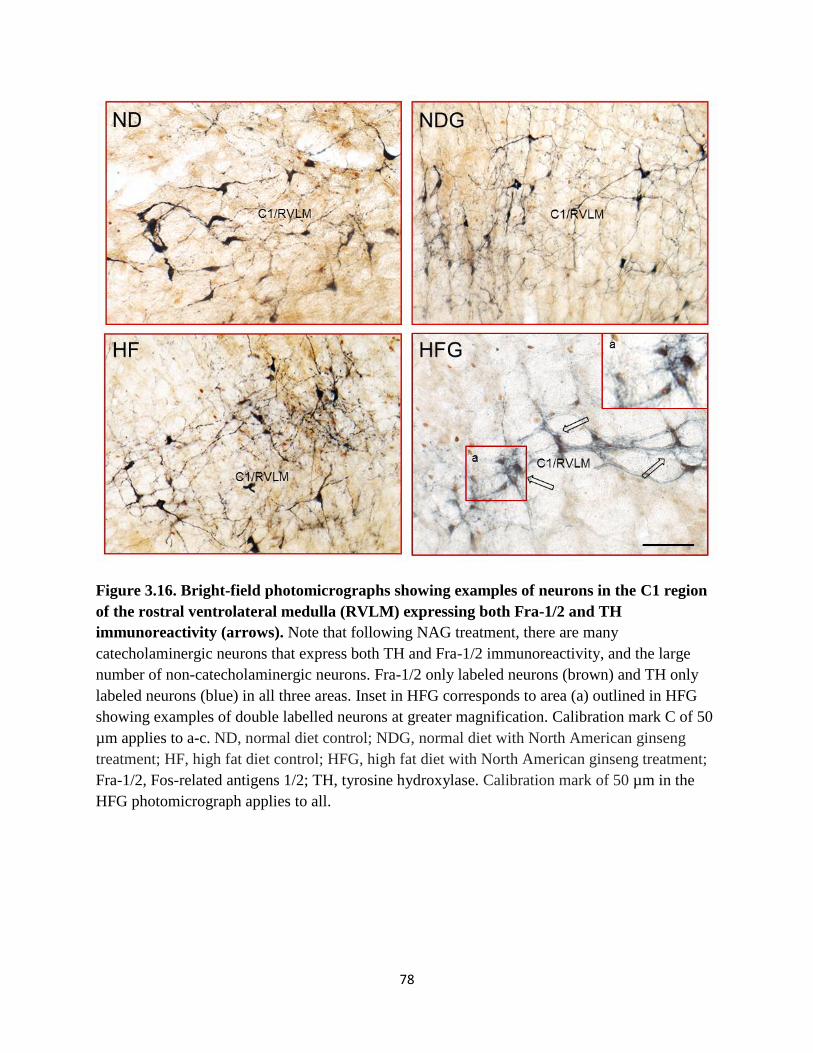
78
Figure 3.16. Bright-field photomicrographs showing examples of neurons in the C1 region
of the rostral ventrolateral medulla (RVLM) expressing both Fra-1/2 and TH
immunoreactivity (arrows). Note that following NAG treatment, there are many
catecholaminergic neurons that express both TH and Fra-1/2 immunoreactivity, and the large
number of non-catecholaminergic neurons. Fra-1/2 only labeled neurons (brown) and TH only
labeled neurons (blue) in all three areas. Inset in HFG corresponds to area (a) outlined in HFG
showing examples of double labelled neurons at greater magnification. Calibration mark C of 50
µm applies to a-c. ND, normal diet control; NDG, normal diet with North American ginseng
treatment; HF, high fat diet control; HFG, high fat diet with North American ginseng treatment;
Fra-1/2, Fos-related antigens 1/2; TH, tyrosine hydroxylase. Calibration mark of 50 µm in the
HFG photomicrograph applies to all.

79
Figure 3.17. Camera lucida projection drawings of the Ventrolateral Medulla.
Representative projection drawings of the caudal ventrolateral medulla (CVLM) show
distribution of Fra-1/2 labeled neurons in animals after high fat diet and/or North American
Ginseng treatment. Fra-1/2 immunoreactive neurons are indicated by the small black dots while
those neurons immunoreactive to the catecholaminergic enzyme TH are indicated by the larger
open circles. Neurons immunoreactive to both Fra-1/2 and TH are indicated by a star.
Note that the majority of the Fra-1/2 labelled neurons were localized in the CVLM and RVLM in
the NAG treated animals. In addition, note the decreased TH and Fra-1/2 TH double labeling in
the CVLM following NAG treatment. LRN, Lateral reticular nucleus; py, pyramidal tract; ION,
inferior olivary nucleus; 12N, root of hypoglossal nerve; AMB, nucleus ambiguous; ND, normal
diet control; NDG, normal diet with North American ginseng treatment; HF, high fat diet
control; HFG, high fat diet with North American ginseng treatment; Fra-1/2, Fos-related antigens
1/2; TH, tyrosine hydroxylase. Calibration mark of 500 µm applies to all.

80
Figure 3.18 Bright-field photomicrograph showing examples of neurons in the A5 region of
the expressing Fra-1/2 and TH immunoreactivity. Note that there are few TH Fra 1/2 double-
labeled neurons. A5, A5 noradrenergic region; LSO, lateral superior olive; rs, rubrospinal tract,
ND, normal diet control; NDG, normal diet with North American ginseng treatment; HF, high fat
diet control; HFG, high fat diet with North American ginseng treatment. Calibration mark of 50
µm in the HFG photomicrograph applies to all.

81
Table 3.5 Table summarizing the number of neurons expressing Fra-1/2 immunoreactivity
in addition to labeling with Tyrosine Hydroxylase (TH). ND, normal diet control; NDG,
normal diet with North American ginseng treatment; HF, high fat diet control; HFG, high fat diet
with North American ginseng treatment; Fra-1/2, Fos-related antigens-1/2; NTS, nucleus of the
solitary tract; CVLM, caudal ventrolateral medulla; RVLM, rostral ventrolateral medulla; A1,
A1 noradrenergic cell group; A2, A2 noradrenergic cell group; C1, C1 adrenergic cell group.
Significance determined using a one-way or two-way ANOVA followed by a Bonferroni post-
hoc test. Data are presented as mean ± SEM. #, p < 0.05 within each diet. *, p < 0.05 between
diets.
Treatment NTS/A2 CVLM/A1 RVLM/C1
TH TH+FRA TH TH+FRA TH TH+FRA
ND 31 ± 4 7 ± 1 15 ± 1 4 ± 1 19 ± 2 1 ± 1
NDG 15 ± 2# 2 ± 1# 17 ± 2 1 ± 1# 23 ± 2 1 ± 1
HFC 47 ± 7* 12 ± 2* 25 ± 2* 7 ± 1* 29 ± 4* 2 ± 2
HFG 30 ± 4#* 6 ± 1#* 28 ± 2* 6 ± 1#* 37 ± 3* 5 ± 3#*

82
Table 3.6. Table summarizing the number of neurons expressing Fra-1/2 immunoreactivity
in the circumventricular organs. Fra-1/2, Fos-related antigens 1/2; ND, normal diet control;
NDG, normal diet with North American ginseng treatment; HF, high fat diet control; HFG, high
fat diet with North American ginseng treatment; MnPO, median preoptic nucleus; OVLT,
organum vasculosum of the lamina terminalis; SFO, subfornical organ; ap, area postrema.
Statistical significance was determined using two-way ANOVA followed by a Bonferroni post-
hoc test. Data are presented as mean ± SEM. # p < 0.05 within diet group.
Area
Number of Fra-1/2 labeled cells/section
NDC
(n=5)
NDG
(n=6)
HFC
(n=5)
HFG
(n=5)
MnPO 13 ± 3 34 ± 7# 5 ± 2 34 ± 7#
OVLT 3 ± 1 14 ± 4# 9 ± 5 28 ± 8#
SFO 19 ± 5 40 ± 6# 19 ± 7 45 ± 8#
ap 13 ± 2 38 ± 4# 13 ± 2 47 ± 7#

83
Figure 3.19. Bright-field photomicrographs taken through the subfornical organ (SFO). Note the higher number of Fra-1/2 labeled neurons in the North American ginseng treated
animals, while there are few neurons that express Fra-1/2 in the control animals. vhc, ventral
hippocampal commissure; 3V, Third ventricle; Fra-1/2, Fos-related antigens 1/2; ND, normal
diet control; NDG, normal diet with North American ginseng treatment; HF, high fat diet
control; HFG, high fat diet with North American ginseng treatment. Calibration mark of 200 µm
in the HFG photomicrograph applies to all.

84
Figure 3.20 Bright-field photomicrographs taken through the organum vasculosum of the
lamina terminalis (OVLT) showing Fra-1/2 labeled neurons. Note the higher number of Fra-
1/2 labeled neurons in animals after ginseng administration. Few neurons express Fra-1/2 in the
vehicle treatment. OVLT, organum vasculosum of the lamina terminalis; a3V, anterior third
ventricle; Fra-1/2, Fos-related antigens 1/2; ND, normal diet control; NDG, normal diet with
North American ginseng treatment; HF, high fat diet control; HFG, high fat diet with North
American ginseng treatment. Calibration mark of 200 µm in the HFG photomicrograph applies
to all.

85
and 3.21). In contrast, in animals treated with NAG there were significantly higher numbers of
neurons containing Fra-1/2-like immunoreactivity in the same regions (Table 3.6; Figs. 3.19,
3.20, and 3.21.).
3.2.2.2 Fra-1/2 Expression in hypothalamic nuclei:
Table 3.7 summarizes the number of Fra-1/2 labeled neurons in different hypothalamic
nuclei in animals on ND and HFD after 28 days of either vehicle or NAG treatment. A small
number of neurons expressing Fra-1/2 like immunoreactivity were found within the ARC, PVH,
SON, DMH, VMH, and LH in animals on a ND or HFD treated with vehicle (controls) (Table
3.7; Figs. 3.22, 3.23, 3.24, and 3.25). In contrast, in animals treated with NAG, there was a
significant increase in the number of neurons containing Fra-1/2-like immunoreactivity within
these nuclei (Table 3.7; Figs. 3.22, and 3.23, 3.24, and 3.25).

86
Figure 3.21 Bright-field photomicrographs taken through the median preoptic nucleus
(MnPO) expressing Fra 1/2 immunoreactivity. Note the higher number of Fra-1/2 labeled
neurons in the NAG treated animals. Few neurons express Fra-1/2 in vehicle treatment.
Calibration. Fx, fornix; ac, anterior commissure; Fra-1/2, Fos-related antigens 1/2; ND, normal
diet control; NDG, normal diet with North American ginseng treatment; HF, high fat diet
control; HFG, high fat diet with North American ginseng treatment. Calibration mark of 200 µm
in the HFG photomicrograph applies to all.

87
Figure 3.22. Bright-field photomicrographs expressing Fra-1/2 immunoreactivity taken
through the arcuate nucleus (ARC). Note the higher number of Fra-1/2 labeled neurons after
NAG administration. VMH, ventral medial hypothalamus; Arc, arcuate nucleus; ME, median
eminence; 3V, third ventricle; Fra-1/2, Fos-related antigens 1/2; ND, normal diet control; NDG,
normal diet with North American ginseng treatment; HF, high fat diet control; HFG, high fat diet
with North American ginseng treatment. Calibration mark of 200 µm in the HFG
photomicrograph applies to all.

88
Figure 3.23. Bright-field photomicrographs taken through the paraventricular nucleus of
the hypothalamus (PVH) showing the distribution of Fra-1/2 labeled neurons. Note the
higher number of Fra-1/2 labeled neurons in the NAG treated animals localized to the posterior
magnocellular and dorsal parvocellular neurons. There are few Fra-1/2 neurons in the vehicle
treated sections. Fx, fornix; LH, lateral hypothalamus; pm, posterior magnocellular nucleus; dp,
dorsal parvocellular nucleus; vp, ventral parvocellular nucleus; mp, medial parvocellular
nucleus; 3V, third ventricle; ND, normal diet control; NDG, normal diet with North American
ginseng treatment; HF, high fat diet control; HFG, high fat diet with North American ginseng
treatment. Calibration mark of 200 µm in the HFG photomicrograph applies to all.

89
Figure 3.24. Bright-field photomicrographs taken through the supraoptic nucleus (SON)
showing the distribution of Fra-1/2 labeled neurons. Note the higher number of Fra-1/2
labeled neurons in the NAG treated animals. There are few Fra-1/2 neurons in the vehicle treated
sections. oc, optic chiasm; ND, normal diet control; NDG, normal diet with North American
ginseng treatment; HF, high fat diet control; HFG, high fat diet with North American ginseng
treatment. Calibration mark of 200 µm in the HFG photomicrograph applies to all.

90
Figure 3.25. Bright-field photomicrographs taken through the anterior hypothalamic area
(AHA) showing the distribution of Fra-1/2 labeled neurons. Note the presence of nucleus
circularis (Nc). ot, optic tract; SON, supraoptic nucleus; LH, lateral hypothalamus; HFG, high fat
diet with North American ginseng treatment. Calibration mark of 200 µm applies to both.

91
Table 3.7. Table summarizing the number of neurons expressing Fra-1/2 immunoreactivity
in hypothalamic regions. Significance was determined using 2-way ANOVA. Data is presented
as mean ± SEM. #, p<0.05 within diet group. NDC, normal diet control; NDG, normal diet with
North American ginseng treatment; HFC, high fat diet control; HFG, high fat with North
American ginseng treatment; PVH, paraventricular nucleus of the hypothalamus; VMH, ventral
medial hypothalamus; ARC, arcuate nucleus; LH, lateral hypothalamus; DMD, dorsomedial
hypothalamus; SON, supraoptic nucleus.
Area
of
hypothalamus
Number of Fra-1/2 labeled neurons/section
NDC
(n=5)
NDG
(n=6)
HFC
(n=5)
HFG
(n=5)
PVN 189 ± 19 233 ± 21# 155 ± 23 266 ± 25#
LH 42 ± 3 64 ± 7# 41 ± 7 50 ± 5#
ARC 83 ± 5 101 ± 7# 84 ± 7 176 ± 16#
VMH 46 ± 4 66 ± 6# 43 ± 7 105 ± 12#
DMD 104 ± 7 160 ± 4# 93 ± 14 121 ± 18#
SON 111 ± 8 127 ± 10# 94 ± 10 151 ± 9#

92
Table 3.7. Table summarizing the number of neurons expressing Fra-1/2 immunoreactivity
in hypothalamic regions. Significance was determined using 2-way ANOVA. Data is presented
as mean ± SEM. #, p<0.05 within diet group. NDC, normal diet control; NDG, normal diet with
North American ginseng treatment; HFC, high fat diet control; HFG, high fat with North
American ginseng treatment; PVH, paraventricular nucleus of the hypothalamus; VMH, ventral
medial hypothalamus; ARC, arcuate nucleus; LH, lateral hypothalamus; DMD, dorsomedial
hypothalamus; SON, supraoptic nucleus.
Area
of
hypothalamus
Number of Fra-1/2 labeled neurons/section
NDC
(n=5)
NDG
(n=6)
HFC
(n=5)
HFG
(n=5)
PVN 189 ± 19 233 ± 21# 155 ± 23 266 ± 25#
LH 42 ± 3 64 ± 7# 41 ± 7 50 ± 5#
ARC 83 ± 5 101 ± 7# 84 ± 7 176 ± 16#
VMH 46 ± 4 66 ± 6# 43 ± 7 105 ± 12#
DMD 104 ± 7 160 ± 4# 93 ± 14 121 ± 18#
SON 111 ± 8 127 ± 10# 94 ± 10 151 ± 9#

93
Chapter 4: Discussion
4.1 The effect of NAG on markers of the metabolic syndrome
The finding in this study that animals on a HFD gained weight (Hall et al., 1993; Mercer
and Trayhurn, 1987; Rahmouni et al., 2005) and had elevated AP and HR compared to those on
ND (Hall et al., 1993; Rahmouni et al., 2005) is in agreement with a number of earlier studies.
However, the finding that NAG treatment was effective in lowering body weight was unexpected
as in the Zucker diabetic fatty rats NAG has been shown to increase body weight, although
cholesterol levels were found to be decreased (Banz et al., 2007). On the other hand,
administration of ginsenosides isolated from NAG has been shown to exert anti-obesity effects
(Liu et al., 2010). Additionally, administration of NAG berry juice has been shown to decrease
body weight in ob/ob mice (Xie et al., 2007).
Interestingly, the lower body weight gain observed in this study occurred independently
of food and water intake. Previous studies using Panax ginseng (Asian ginseng) have reported
similar reductions in body weight gain without a change in food intake (Lee, 2010). It has been
suggested that ginsenosides increase energy consumption and lower body weight by inhibiting
pancreatic lipase thus reducing the absorption of dietary fat (Karu et al., 2007; Liu et al., 2010)
and lowering overall body fat. Thus, it is possible that a similar mechanism is at work in our
studies with NAG. Interestingly, animals treated with NAG in this study also had lower fat pad
mass and leptin levels. Previous studies have suggested that G-Rb1 lowers leptin levels as a
result of the overall reduction in body fat (Liu et al., 2008, 2011; Xiong et al., 2010). As plasma
leptin levels are known to positively correlate with body fat, it is not surprising that NAG treated
animals also had lower plasma leptin levels (Zhang et al., 1994). Furthermore, in corresponding

94
with lower adiposity, NAG lowered plasma triglyceride levels and it has been suggested that this
also may occur as a result of inhibiting pancreatic lipase (Liu et al., 2010).
In this study, NAG had no effect on total cholesterol levels in rats on a ND or HFD. This
is a controversial finding as previous studies have shown lower cholesterol following ginseng
treatment (Kim et al., 2011; Xia et al., 2011). It has been hypothesized that ginseng can decrease
total cholesterol level from an elevated level back to a normal value (Kim et al., 2011; Xia et al.,
2011). However, animals in this study were fed a HFD for only 28 days and this did not yield
any change in total cholesterol levels. As it is known that rodents have the ability to maintain
their cholesterol profile and it is necessary to ‘force’ the atherosclerosis phenotype with a diet
rich in cholesterol, which was not used in this study (Maxwell et al., 2003), it likely that NAG
would not be expected to exert an effect on cholesterol. Interestingly, Liu and colleagues (2010)
reported that although NAG treatment did not alter total cholesterol levels, there was a
significant reduction of LDL cholesterol in NAG treated animals. LDL was not measured in this
study.
NAG treatment was found to be associated with lower plasma insulin levels.
Concomitantly, insulin resistance, as calculated by HOMA-IR, decreased. Other studies have
shown that NAG increases insulin sensitivity. Specifically, it has been proposed that ginseng
reduces blood glucose by an anti-inflammatory mechanism (Zhang et al., 2008). It has been
suggested that G-Re, a major component of NAG, is the component that reduces insulin
resistance via inhibition of c-Jun NH2-Terminal Kinase and NF-kB (Zhang et al., 2008). In
addition, Compound K, a metabolite of NAG, has been shown to have anti-hyperglycemic
properties, though the mechanism of action has not been elucidated (Han et al., 2007; Yoon et
al., 2007).

95
Following NAG treatment, there was no change in plasma adiponectin levels. This was
an unexpected finding as adiponectin is known to regulate energy homeostasis by modulating
glucose and fatty acid metabolism (Dridi and Taouis, 2009) and is normally found in the plasma
in levels opposite to those of leptin (Dridi and Taouis, 2009). Although, it has been reported that
Korean Red ginseng does not alter adiponectin levels in HFD fed animals (Lee et al., 2012), it
has also been reported that Compound K, a metabolite of ginseng, is capable of increasing
circulating adiponectin levels (Han et al., 2007). However, it should be noted that Han and
colleagues (2007) used a db/db diabetic mouse model whereas Lee and colleagues (2012), and
the current study, used HFD-fed rats. However, there are data to indicate that high-molecular
weight adiponectin correlates better with glucose tolerance and insulin resistance (Hara et al.,
2006; Fischer et al., 2005). However, it has been suggested that improvements of insulin
resistance associated with less weight gain are not associated with changes in plasma adiponectin
levels (Abbasi et al., 2009). In the current study treatment with NAG was associated with weight
loss and the lower fat content may account for the lack of difference in plasma adiponectin
levels.
4.2 The effect of NAG on AP and HR
In contrast to earlier observations in clinical studies (Siegel, 1979; Stavro et al., 2005,
2006), the current study has shown that NAG administered by oral gavage in animals on HFD
was effective in preventing an increasing AP and HR to levels similar in animals on a ND only
or on a ND and given NAG. The reason for the difference between the current study and the
clinical studies may be that NAG has a selective effect on the hypertension associated with
weight gain as opposed to essential hypertension as in the clinical studies (Stavro et al., 2005,

96
2006). The finding in the current study that ginseng blocks the increase in AP is consistent with
the previous observation in spontaneously hypertensive rats that intravenous administration of
the G-Rb1, a ginsenoside found in high concentrations in NAG (Wang et al., 2005), lowers AP
(Pan et al., 2012). Consistent with this finding is that administration of capsules containing Asian
ginseng have been shown to improve baroreceptor reflex function (Ai et al., 2010).
Previous studies have shown that weight gain associated with a HFD increases AP,
sympathetic activity, and circulating noradrenaline (Rocchini et al., 2010). The increase
sympathetic nervous activity and the release of noradrenaline contribute to the hypertension
(Rocchini et al., 2010). The current findings are consistent with these findings. Rats on a HFD
gained more weight, had higher MAP, and higher plasma noradrenaline levels as compared to
ND control. Of interest is the finding that in HFD animals treated with NAG, the noradrenaline
levels were lower and MAP was not different than found in controls. This suggests that the lower
plasma noradrenaline, likely due to lower sympathetic activity may have been responsible for the
blockade of the increase in MAP in HFD. In support of this suggestion, ginsenosides from Asian
ginseng have been shown to decrease the noradrenaline-induced blood vessel contractile
responses (Kim et al., 2006). In addition, G-Rb2, G-Rd, and G-Rg1 have been shown to decrease
noradrenaline-induced increases of interleukin-6 (Kim et al, 2003), also suggesting that ginseng
can modulate noradrenaline release.
An unexpected finding in this study was that treatment with NAG resulted in higher
circulating plasma angiotensin II levels. In contrast, Asian ginseng extract has been shown to
inhibit angiotensin-converting enzyme activity (Persson et al., 2006). The function of the higher
angiotensin II following NAG treatment is not clear as NAG treated animals did not have higher
MAP and water intake, or lower urine output.

97
4.3 The effect of NAG on brainstem autonomic nuclei
In animals on HFD, had higher numbers of Fra-1/2 labeled neurons observed in the NTS,
CVLM, and RVLM in comparison to animals on a ND. In addition, there were a lower number
of labeled neurons in the MARN. These findings are supported by an earlier observation of
increased Fra-1/2 labeling in the NTS, CVLM, and RVLM in a model of obesity induced
hypertension in canines (Lohmeier et al., 2003). These earlier findings were interpreted to
indicate that the baroreflex pathway has a continuous suppressive influence on sympathetic
activation in obesity-induced hypertension (Lohmeier et al., 2003). However, it is known that
obesity eventually reduces baroreflex function, which then ultimately leads to increased arterial
pressure and sympathetic nerve activity (Grassi et al., 2000; Lohmeier et al., 2003).
The finding that MARN had a lower number of labeled neurons following NAG
treatment in the ND group is consistent with the notion that MARN is a sympathoexcitory area in
the RVLM, under baroreflex influence (Babic and Ciriello, 2004; Varner et al., 1989, 1992), and
that the integrity of this region is thought to be important in neurogenic hypertension (Varner et
al., 1994). This finding suggests that neurons in this region maintain a tonic excitatory drive on
sympathetic preganglionic neurons controlling vasomotor tone (Babic and Ciriello, 2004; Varner
et al., 1989, 1992, 1994). Interestingly, central hypocretin pathways from the lateral
hypothalamus that are thought to be involved in food intake (Dube et al., 1999; Haynes et al.,
1999) and in sympathetic drive (Samson et al., 1999; Shirasaka et al., 1999) project to this
brainstem region (Ciriello and de Oliveira, 2003). Microinjections of hypocretin into the MARN
have been shown to elicit sympathoexcitatory responses (Ciriello and de Oliveira, 2003).
Treatment of animals on HFD with NAG resulted in a greater than 3-fold higher number
of Fra-1/2 labeled neurons within the ap. In addition, there was almost a 2-fold higher number of

98
Fra-1/2 labeled neurons in the NTS and CVLM, and a smaller number within the RVLM. The
finding of NAG exerting effects on brainstem neurons was not unexpected as it has previously
been shown that ginseng extracts will alter the expression of GluR2 in the nucleus ambiguus (Ai
et al., 2010), increase the central release of the neurotransmitter glutamate (Xue, et al., 2006),
interact with excitatory glutamate receptors (Lee, 2002), and influence the activity of central
catecholaminergic neurons (Hwang and Jeong, 2010; Radad et al., 2004).
The RVLM is known to be the primary regulator of sympathetic nervous activity.
Although we see higher neuronal activation in this area following NAG treatment, it is important
to note that there is not a corresponding higher MAP or HR of these animals. While many
RVLM neurons do project to the spinal cord to increase sympathetic output, other neurons
innervate the hypothalamus. Therefore it is reasonable to suggest that although RVLM is
activated, the neurons that are activated are likely not directly controlling sympathetic outflow.
However, they may represent inhibitory neurons that may be attempting to maintain normal
sympathetic tone, or are involved in relaying integrated information from the NTS to
hypothalamic forebrain structures controlling food intake, water intake, or metabolism.
4.3.1 Fra-1/2 Immunoreactivity in brainstem TH Labeled neurons:
It is well documented that central catecholaminergic neurons play an important role in
cardiovascular control (Esler, et al., 1995, 1997; Guyenet, 1991: Head, 1992; Korner et al., 1987;
Struthers and Dollary, 1985). Electrophysiological and pharmacological studies favour a
sympatho-inhibitory role for noradrenaline containing neurons that project to the spinal cord
(Coote, 1985). On the other hand, adrenergic cells found within the RVLM appear to function as
sympathoexciatrory neurons (Ciriello et al., 1986; Reis et al., 1989). In the present study, it was

99
observed that in animals regardless of diet, there was a difference in expression of TH
immunoreactive neurons in the brainstem following administration of NAG. Within the NTS
NAG the number of TH immunoreactive and TH cells that also expressed Fra-1/2 was lower. As
these cells were observed selectively within the NTS region, it can be assumed that they
represent catecholaminergic cells of the A2 noradrenergic cell group. Thus, noradrenaline
producing cells are affected by NAG, possibly by an effect of NAG on the enzyme TH which
catalyzes the conversion of dopamine to noradrenaline within these neurons. In support of this
finding, Asian ginseng has been shown to modulate catcholaminergic activity within the CNS by
its effects on tyrosine hydroxide expression (Kim et al., 2010). Additionally, the ginsenoside G-
Rb1, found in abundance within NAG, has been suggested to provide a partial neurotrophic and
neuroprotective effect on catecholaminergic neurons (Hwang and Jeong, 2010; Radad et al.,
2004). Of interest is the finding that the number of TH immunoreactive cells doubled-labeled
with Fra-1/2 was significantly smaller than the total number of Fra-1/2 labelled cells observed
with the NTS region. This suggests that neurons containing other transmitters are being affected
by NAG. In support of this suggestion, G-Rb1 has been reported to increase the release of the
neurotransmitter glutamate within the CNS (Xue et al., 2006), and to interact with related
excitatory neurotransmitter receptors (Lee, 2002). In addition, treatment with G-Rb1 has been
reported to induce c-fos expression in brain nuclei such as NTS (Xiong et al., 2010). As
glutamate is known to be the main transmitter within baroreceptor afferents fibers (Bonham and
Chen, 2002), it is likely that an increase in release of glutamate from these afferent pathways
may be responsible for the increased activity of the NTS cells. Interestingly, few noradrenergic
cells in NTS have direct contact with glutamate afferents (Milner and Pickel, 2003).

100
Within the CVLM, NAG had no effect on TH expression in neurons, although with HFD
the number of cells that expressed TH was higher. Cells of the CVLM that express TH are
known to compose the A1 noradrenergic cell group (Reis et al., 1984). These neurons are known
to innervate RVLM neurons (Reis et al., 1984), although most project to hypothalamic sites such
as the PVH, ARC, SON, and LH (Dampney et al., 2003; Day 1989). The finding that with HFD
increased the number of neurons in the CVLM were higher is a novel observation and suggests
that A1 cells may be involved in controlling ingestive behaviours as noradrenaline administered
into PVH is known to alter food intake and metabolism (Welman, 2000). However, it also may
be that these neurons are involved in the control of vasopressin release from the PVH and SON
(Dampney et al., 2003; Day, 1989).
Finally, the RVLM, which is known to only contain adrenergic neurons (Reis et al.,
1984), was found in this study to have a higher number of TH immunoreactive cells and an
higher in the number of TH and Fra-1/2 labelled cells in the animals on HFD, while NAG had
little effect on these neurons. As neurons of the RVLM are known to mainly innervate the
sympathetic preganglionic neurons in the spinal cord, although a smaller number project to the
hypothalamus (Reis et al., 1984), it is likely that the hypertension observed in the HF animals
may be partially due to an increased activity of these TH immunoreactiove neurons in the
RVLM. However, as the hypertension is resolved after NAG treatment, but the activity of these
cells remain above ND control animals, it suggests that either they are not inhibited by NAG and
blood pressure is under the control of a different pathway, or that they subserve a completely
different physiological role.

101
4.4 The effect of NAG on neuronal activity in the forebrain
4.4.1 Fra-1/2 Immunoreactivity in the circumventricular organs:
NAG resulted in higher Fra-1/2 labeling in the circumventricular organs (ap, MnPO,
SFO, and OVLT) suggesting an increase in neuronal activity. The circumventricular organs are
highly vascularized and lack a functional blood–brain barrier (Benarroch, 2011). Thus, these
areas transduce chemical signals from the bloodstream and act as points of communication
between the blood, the cerebrospinal fluid, and neurons (Benarroch, 2011). Although it has been
suggested that Compound K, a metabolite of ginseng, is unable to cross the blood brain barrier,
Compound K and G-Rb1 have been shown to exert effects within the CNS (Kim et al., 2003; Lu
et al., 2011; Murphy and Lee, 2002). This suggests that the ginsenosides must gain access to the
CNS through routes where the blood brain barrier may be “leaky” as in the circumventricular
organs. In addition, several of these structures respond to blood borne chemicals such as
angiotensin II and leptin (Hoyda et al., 2009), which were found to be altered within the plasma
after NAG treatment. Thus it was not unexpected to have higher Fra 1/2 activity within the SFO
as neurons within this structure respond to both angiotensin II and mediate drinking and
cardiovascular responses (Hoyda et al., 2009) and to leptin which may exert an effect on food
intake and cardiovascular responses (Smith and Ferguson, 2012). The ap and OVLT also
function in controlling fluid intake and play a role in food ingestion and cardiovascular function
(Hoyda et al., 2009).
As all these circumventricular structures have connections within the CNS, especially the
hypothalamus (Hoyda et al., 2009; Smith and Ferguson, 2012), it is not surprising that NAG also
had effects on areas associated with cardiovascular regulation and energy metabolism within the
hypothalamus.

102
4.4.2 Fra-1/2 immunoreactivity in hypothalamic nuclei:
Within the hypothalamus several structures associated with cardiovascular regulation,
ingestive behaviour, and metabolism were found to contain a large number of neurons that
expressed Fra-1/2 after NAG administration regardless of diet. This suggests that the diet did not
play a role in altering the activity of these neurons, but rather it was associated with the NAG
administration. Higher neuronal activity was found within the PVH, LH, ARC, SON, VMH, and
DMD. All these structures, except for the PVH and SON which also have roles in cardiovascular
regulation (Coote, 1995), are involved in food intake, metabolism, and energy expenditure
(Blevins and Baskin, 2010; Elmquist, 2001). The ARC nucleus is the main hypothalamic site
known to transduce plasma leptin and insulin signals and activate pathways that control food
intake by the LH, PVH, and DMD (Cammisotto et al., 2010; Konner, 2009; Migrenne et al.,
2011). The PVH on the other hand responds to these ARC signals and activates a series of
neuronal pathways that signal satiety, feeding, or increased metabolism (Ahima and Antwi,
2003). But the PVH exerts more effects than simply on ingestion. In fact, this area is often
considered a major integrating center for a variety of stimuli which then selectively affect a
number of efferent systems that control neuroendocrine, behavioural, and autonomic responses
(Smith and Ferguson, 2008). Within the PVH, there were a number of subnuclei that expressed a
larger number of Fra-1/2 labeled cells after NAG administration. A significant increase in the
number of active neurons was found within the posterior magnocellular, dorsal parvocellular,
and the medial parvocellular subnuclei. The posterior magnocellular subnucleus, like the SON,
contains vasopressin and oxytocin neurosecretory neurons (Coote, 1995). On the other hand, the
parvocellular neurons of the dorsal subnucleus send efferent connections to the spinal cord
sympathetic preganglionic neurons (Swanson and Sawchenko, 1980), whereas those of the

103
medial subnucleus send axons to the median eminence for the release of releasing factors that
control anterior pituitary function (Herman et al., 2008). Interestingly, NAG did not alter activity
of cells of the ventral parvocellular subnucleus of PVH, suggesting that neurons that innervate
the dorsomedial medulla (Swanson and Sawchenko, 1980) are not affected by NAG. These data
could then be interpreted to suggest that NAG has an influence not only on autonomic systems,
but also posterior and anterior pituitary function. Finally, it is of interest that all of these
hypothalamic structures that have higher activity following NAG administration also receive
inputs from the NTS, a brainstem site that also showed increased Fra-1/2 expression after NAG
administration, and is involved in food intake and energy expenditure (Rinaman, 2010).
4.5 Overall Summary and Conclusion
This study has provided evidence indicating that oral administration of NAG may be an
effective approach in preventing the onset of risk factors associated with the development of the
metabolic syndrome in overweight rats. This is based on the observation that NAG treatment
resulted in lower body weight gain and lower fat mass levels, plasma triglycerides, insulin levels,
and insulin resistance. In addition, NAG treatment resulted in lower circulating plasma
noradrenaline and leptin. Furthermore, NAG was able to prevent an increase AP and HR in
animals fed a HFD, and activated brainstem and hypothalamic autonomic nuclei involved in AP
regulation, ingestion of food, metabolism, and energy expenditure. As a result, NAG may
function to alter neuronal activity that contributes to the development of the metabolic syndrome,
especially those that contribute to obesity induced hypertension. Finally, the higher Fra-1/2
activity observed within circumventricular organs suggests a possible route by which
ginsenosides or their metabolites may access the CNS to exert these effects. Overall, this study

104
has laid the groundwork for future investigations of the effects of NAG on central metabolic
pathways. Understanding how NAG can impact the neural and humoral mechanisms that link the
pathophysiologies associated with obesity and the metabolic syndrome may prove to be of
clinical relevance and lead to new therapeutic interventions.
4.6 Future directions
Although the novel findings of this study extend our understanding of the effects of NAG,
they also introduce a number of possibilities for future investigation. The finding that NAG
increases neuronal activation in areas associated with metabolism encourages future
investigation to determine the phenotype of hypothalamic neurons activated by NAG. Although
it is suggested that the circumventricular organs could be a possible route for ginsenosides to
access the CNS, further studies are needed. For example, lesioning the circumventricular organs
will provide more information as to whether this is the access site for ginsenosides. As G-Rb1 is
the main component of NAG and it has been shown to exert effects in the CNS (Lu et al., 2011),
it may be interesting investigate whether the beneficial effects shown in this study are mediated
mainly by this ginsenoside.
In addition, this study suggests that NAG may be beneficial in preventing the onset of
development of risk factors associated with the metabolic syndrome. As approximately 1 in 5
Canadians have already been diagnosed with the metabolic syndrome (Riediger, and Clara,
2011), another logical extension of these findings would be to investigate the effects of NAG as
a treatment option for obesity and the metabolic syndrome once the pathophysiology of disease
has been manifested. The inconvenience of treating each of the components of metabolic

105
syndrome with different therapeutic agents may be avoided with the development of novel agent
that has multiple modes of action such as NAG.

106
References
Aa, J., Wang, G., Hao, H., Huang, Q., Lu, Y., Yan, B., Zha, W., Liu, L., Kang, A., 2010.
Differential regulations of blood pressure and perturbed metabolism by total ginsenosides and
conventional antihypertensive agents in spontaneously hypertensive rats. Acta. Pharmacologica.
Sinica. 31, 930–937.
Ackermann, U., 2004. Regulation of arterial blood pressure. Surgery. 22, 120a-120f.
Achike, F.I., To, N.H., Wang, H., Kwan, C.Y., 2011. Obesity, metabolic syndrome, adipocytes
and vascular function: A holistic viewpoint. Clin. Exp. Pharmaco. Physiol. 38, 1-10.
A.D.A.M. Medical Encyclopedia. Metabolic syndrome. PubMed Health. Updated June 28, 2011.
Accessed August 1, 2012. http://www.ncbi.nlm.nih.gov/pubmedhealth/PMH0004546/
Ahima, R.S., Antwi, D.A., 2008. Brain regulation of appetite and satiety. Endocrinol. Metab.
Clin. North Am. 37, 811-23.
Alberti, K.G., Eckel, R.H., Grundy, S.M., Zimmet, P.Z., Cleeman, J.I., Donato, K.A., Fruchart,
J.C., James, W.P., Loria, C.M., Smith Jr, S.C., 2009. Harmonizing the metabolic syndrome: A
joint interim statement of the International Diabetes Federation Task Force on Epidemiology and
Prevention; National Heart, Lung, and Blood Institute; American Heart Association; World
Heart Federation; International Atherosclerosis Society; and International Association for the
Study of Obesity. Circulation. 120, 1640–1645.
Andrade, A.S., Hendrix, C., Parsons, T.L., Caballero, B., Yuan, C.S., Flexner, C.W., Dobs, A.S.,
Brown, T.T., 2008. Pharmacokinetic and metabolic effects of American ginseng (Panax
quinquefolius) in healthy volunteers receiving the HIV protease inhibitor indinavir. BMC
Complement. Altern. Med. 8, 50.
Assinewe, V., Stavro, M.P., Jenkins, A.L., Leiter, L.A., Francis, T., 2001. American ginseng
(Panax quinquefolius L.) attenuates postprandial glycemia in a time-dependent but not dose-
dependent manner in healthy individuals. Am. J. Clin. Nutr. 73, 753-8.
Attele, A.S., Zhou, Y.P., Xie, J.T., Wu, J.A., Zhang, L., Dey, L., Pugh, W., Rue, P.A.,
Polonsky, K.S., Yuan, C.S., 2002. Antidiabetic effects of Panax ginseng berry extract and the
identification of an effective component. Diabetes. 51,1851-8.
Azike, C.G., Charpentier, P.A., Hou, J., Pei, H., King Lui, E.M., 2011. The Yin and Yang
actions of North American Ginseng root in modulating the immune function of macrophages.
Chin. Med. 27, 21.
Banks, W.A., Kastin, A.J., Huang, W., Jaspan, J.B., Maness, L.M., 1996. Leptin enters the brain
by a saturable system independent of insulin. Peptides. 17, 305 –311.
Banz, W.J., Iqbal, M.J., Bollaert, M., Chickris, N., James, B., Higginbotham, D.A., Peterson, R.,
Murphy, L., 2007. Ginseng modifies the diabetic phenotype and genes associated with diabetes
in the male ZDF rat. Phytomedicine. 14, 681-689.
Benishin, C.G., 1992. Actions of ginsenoside Rb1 on choline uptake in central cholinergic nerve
endings. Neurochem. Int. 21, 1–5.
Bensky, D., Clavey, S., Stoger, E., Gamble, A., 2005. Chinese Herbal Medicine: Materia
Medica, third edition. Eastland Press.

107
Bjorntorp, P., Rosmond R., 2000. Neuroendocrine abnormalities in visceral obesity. Intern. J.
Obesity. Rel. Metab. Dis. 24(suppl 2), S80–S85.
Blevins, J.E., Baskin, D.G., 2010. Hypothalamic-brainstem circuits controlling eating. Forum
Nutr. 63, 133-40.
Blumenthal, M., 2003. The ABC Clinical Guide to Herbs. American Botanical Council, first
edition. American Botanical Council, Austin, TX.
Blumenthal, M., Ferrier, G. K. L., Cavaliere, C., 2006. Total sales of herbal supplements in
United States show steady growth. HerbalGram, 64−66.
Bonham, A.C., Chen, C.Y., Glutamatergic neural transmission in the nucleus tractus solitarius:
N-methyl-D-aspartate receptors. Clin. Exp. Pharmacol. Physiol. 29, 497-502.
Bray, G.A., Fauci, A.S., Braunwald,. E., Isselbacher, K.J., 1998. Obesity. Harrison’s Principles
of Internal Medicine, 14th edition. McGraw- Hill, New Yor,. pp. 454–62.
Bray, G.A., Greenway, F.L., 1999. Current and Potential Drugs for Treatment of Obesity.
Endocr. Rev. 20, 805-875.
Bray, G.A., Ryan, D.H., Harsha, D.W., 2003. Diet, Weight Loss, and Cardiovascular Disease
Prevention. Curr.Treat. Options. Cardiovasc. Med. 5, 259-269.
Bray, G.A., 2004. Medical Consequences of Obesity.J Clin Endocrinol Metab. 89, 2583-2589.
Cammisotto, P.G., Levy, E., Bukowiecki, L.J., Bendayan, M., 2010. Cross-talk between adipose
and gastric leptins for the control of food intake and energy metabolism. Prog. Histochem.
Cytochem. 45, 143-200.
Carmen Cabrera, C., Artacho, R., Giménez, R., 2006. Beneficial Effects of Green Tea—A
Review. J. Am. Coll. Nutr. 25, 79-99.
Chen, X., 1996. Cardiovascular protection by ginsenosides and their nitric oxide releasing action.
Clin. Exp. Pharmacol. Physiol. 23, 728-32.
Chen, C.F., Chiou, W.F., Zhang, J.T., 2008. Comparison of the pharmacological effects of Panax
ginseng and Panax quinquefolium. Acta. Pharmacol. Sin. 29, 1103-8.
Cho, W.C., Chung, W.S., Lee, S.K., Leung, A.W., Cheng, C.H., Yue, K..K., 2006. Ginsenoside
Re of Panax ginseng possesses significant antioxidant and antihyperlipidemic efficacies in
streptozotocin-induced diabetic rats. Eur. J. Pharmacol. 550, 173-9.
Chobanian, A.V., Bakris, G.L., Black, H.R., Cushman, W.C., Green, L.A., Izzo Jr., J.L., Jones,
D.W., Materson, B.J., Oparil, S., Wright Jr., J.T., Roccella, E.J., 2003. The Seventh Report of the
Joint National Committee on Prevention, Detection, Evaluation, and Treatment of High Blood
Pressure: the JNC 7 report. JAMA. 289, 2560-72.
Chung, S.H., Choi, C.G., Park, S.H., 2001. Comparisons between white ginseng radix and rootlet
for antidiabetic activity and mechanism in KKAy mice. Arch. Pharm. Res. 24, 214-8.
Ciriello, J., Rohlicek, C.V., Polosa, C., 1983. Aortic baroreceptor reflex pathway: a functional
mapping using [3H]2-deoxyglucose autoradiography in the rat. J. Auton. Nerv. Syst. 8, 111-128.
Ciriello, J., Caverson, M.M., Polosa, C., 1986. Function of the ventrolateral medulla in the
control of the circulation. Brain Res. 396, 359-91.

108
Cnop, M., Landchild, M.J., Vidal, J., Havel, P.J., Knowles, N.G., Carr, D.R., Wang, F., Hull,
R.L., Boyko, E.J., Retzlaff, B.M., Walden, C.E., Knopp, R.H., Kahn, S.E., 2002. The concurrent
accumulation of intra-abdominal and subcutaneous fat explains the association between insulin
resistance and plasma leptin concentrations : distinct metabolic effects of two fat compartments.
Diabetes. 51, 1005-15.
Coote, J.H., 1985. Noradrenergic projections to the spinal cord and their role in cardiovascular
control. J. Auton. Nerv. Syst. 14, 255-62.
Coote, J.H., 1995. Cardiovascular function of the paraventricular nucleus of the hypothalamus.
Biol Signals. 4, 142-9.
Cui, X., Sakaguchi, T., Ishizuka, D., Tsukada, K., Hatakeyama, K., 1998. Orally administered
ginseng extract reduces serum total cholesterol and triglycerides that induce fatty liver in 66%
hepatectomized rats. J. Int. Med. Res. 26, 181-7.
Curran, T., Franza Jr., B.R., 1988. Fos and Jun: the AP-1 connection. Cell. 55, 395-7.
Dampney, R.A., Polson, J.W., Potts, P.D., Hirooka, Y., Horiuchi, J., 2003. Functional
organization of brain pathways subserving the baroreceptor reflex: studies in conscious animals
using immediate early gene expression. Cell. Mol. Neurobiol. 23, 597-616.
Day, T.A., 1989. Control of neurosecretory vasopressin cells by noradrenergic projections of the
caudal ventrolateral medulla. Prog. Brain Res. 81, 303-17.
De Souza, L.R., Jenkins, A.L., Sievenpiper, J.L., Jovanovski, E., Rahelić, D., Vuksan, V., 2011.
Korean red ginseng (Panax ginseng C.A. Meyer) root fractions: differential effects on
postprandial glycemia in healthy individuals. J. Ethnopharmacol. 137, 245-50.
Dey, L., Attele, A.S., Yuan, C.S., 2002. Alternative therapies for type 2 diabetes. Altern. Med.
Rev. 7, 45-58.
Dey, L., Xie, J.T., Wang, A., Wu, J., Maleckar, S.A., Yuan, C.S., 2003 Anti-hyperglycemic
effects of ginseng: comparison between root and berry. Phytomedicine. 10, 600-5.
Dong, H., Bai, L.P., Wong, V.K., Zhou, H., Wang, J.R., Liu, Y., Jiang, Z.H., Liu, L., 2011. The
in vitro structure-related anti-cancer activity of ginsenosides and their derivatives. Molecules. 16,
10619-30.
Dridi, S., and Taouis, M., 2009. Adiponectin and energy homeostasis: consensus and
controversy. J. Nutr. Biochem. 20, 831-9.
Elmquist, J.K., 2001. Hypothalamic pathways underlying the endocrine, autonomic, and
behavioral effects of leptin. Physiol. Behav. 74, 703-8.
Esler, M.D., Lambert, G.W., Ferrier, C., Kaye, D.M., Wallin, B.G., Kalff, V., Kelly, M.J.,
Jennings, G.L., 1995. Central nervous system noradrenergic control of sympathetic outflow in
normotensive and hypertensive humans. Clin. Exp. Hypertens. 17, 409-23.
Esler, M., Lambert, G., Vaz, M., Thompson, J., Kaye, D., Kalff, V., Kelly, M., Turner, A.,
Jennings, G., 1997. Central nervous system monoamine neurotransmitter turnover in primary and
obesity-related human hypertension. Clin. Exp. Hypertens. 19, 577-90.
Finucane, M.M., Stevens, G.A., Cowan, M.J., Danaei, G., Lin, J.K., Paciorek, C.J., Singh, G.M.,
Gutierrez, H.R., Lu, Y., Bahalim, A.N., Farzadfar, F., Riley, L.M., Ezzati, M., 2011. National,

109
regional, and global trends in body-mass index since 1980: Systematic analysis of health
examination surveys and epidemiological studies with 960 country-years and 9·1 million
participants. Lancet. 377, 557–567.
Fisher, M., Trujillo, M.E., Hanif, W., Barnett, A.H., McTernan, P.G., Scherer, P.E., Kumar, S.,
2005. Serum high molecular weight complex of adiponectin correlates better with glucose
tolerance than total serum adiponectin in Indo-Asian males. Diabetologia. 48, 1084-1087.
Friedman, J.M., Halaas, J.L., 1998. Leptin and the regulation of body weight in mammals.
Nature. 395, 763-70.
Fuzzati, N., 2004. Analysis methods of ginsenosides. J. Chromatogr. Analyt. Technol. Biomed.
Life. Sci. 812, 119-33.
Goldstein, D.S., 1983. Plasma catecholamines and essential hypertension. An analytical review.
Hypertension. 5:86-99
Grassi, G., Seravalle, G., Dell’Oro, R., Turri, C., Bolla, G.B., Mancia, G., 2000. Adrenergic and
reflex abnormalities in obesity-related hypertension. Hypertension. 36, 538–542.
Grundy, S.M., 2008. Metabolic syndrome pandemic. Arterioscler. Thromb. Vasc. Biol. 28, 629–
636.
Guyenet, P.G., 1991. Central noradrenergic neurons: the autonomic connection. Prog. Brain Res.
88, 365-80.
Guyenet, P.G., Cabot, J.B., 1981. Inhibition of sympathetic preganglionic neurons by
catecholamines and clonidine: mediation by an alpha-adrenergic receptor. J. Neurosci. 1, 908–
917.
Guo, J., Gan, X.T., Haist, J.V., Rajapurohitam, V., Zeidan, A., Faruq, N.S., Karmazyn, M., 2010.
Ginseng inhibits cardiomyocyte hypertrophy and heart failure via NHE-1 inhibition and
attenuation of calcineurin activation. Circ. Heart. Fail. 4, 79-88.
Han, K.H., Choe, S.C., Kim, H.S., Sohn, D.W., Nam, K.Y., Oh, B.H., Lee, M.M., Park, Y.B.,
Choi, Y.S., Seo, J.D., Lee, Y.W., 1998. Effect of red ginseng on blood pressure in patients with
essential hypertension and white coat hypertension. Am. J. Chin. Med. 26, 199-209.
Han, G.C., Ko, S.K., Sung, J.H., Chung, S.H., 2007. Compound K enhances insulin secretion
with beneficial metabolic effects in db/db mice. J. Agric. Food. Chem. 55, 10641-8.
Hara, K., Horikoshi, M., Yamauchi, T., Yago, H., Miyazaki, O., Ebinuma, H., Imai, Y., Nagai,
R., Kadowaki, T., 2006. Measurement of the High–Molecular Weight Form of Adiponectin in
Plasma Is Useful for the Prediction of Insulin Resistance and Metabolic Syndrome. Diabetes
Care. 29, 1357-1362.
Hasegawa, H. Uchiyama, M., 1998. Antimetastatic efficacy of orally administered ginsenoside
Rb1 in dependence on intestinal bacterial hydrolyzing potential and significance of treatment
with an active bacterial metabolite. Planta. Med. 64, 696-700.
Hasegawa, H., 2004. Proof of the mysterious efficacy of ginseng: basic and clinical trials:
metabolic activation of ginsenoside: deglycosylation by intestinal bacteria and esterification with
fatty acid. J. Pharmacol. Sci. 95, 153-7.

110
Head, G.A., 1992. Central monoamine neurons and cardiovascular control. Kidney Int. Suppl.
37, S8-13.
Herman, J.P., Flak, J., Jankord, R., 2008. Chronic stress plasticity in the hypothalamic
paraventricular nucleus. Prog. Brain Res. 170, 353-64.
Hong, S.Y., Kim, J.Y., Ahn, H.Y., Shin, J.H., Kwon, O., 2012. Panax ginseng Extract Rich in
Ginsenoside Protopanaxatriol Attenuates Blood Pressure Elevation in Spontaneously
Hypertensive Rats by Affecting the Akt-Dependent Phosphorylation of Endothelial Nitric Oxide
Synthase. J. Agric. Food. Chem. 60, 3086-91.
Hoyda, T.D., Smith, P.M., Ferguson, A.V., 2009. Gastrointestinal hormone actions in the central
regulation of energy metabolism: potential sensory roles for the circumventricular organs. Int. J.
Obes. (Lond). 33 Suppl 1, S16-21.
Hwang, Y.P., Jeong H.G., 2010. Ginsenoside Rb1 protects against 6-hydroxydopamine-induced
oxidative stress by increasing heme oxygenase-1 expression through an estrogen receptor-related
PI3K/Akt/Nrf2-dependent pathway in human dopaminergic cells. Toxicol. Appl. Pharmacol.
242, 18-28.
Itoh, T., Zang, Y.F., Murai, S., Saito, H., 1989. Effects of Panax ginseng root on the vertical and
horizontal motor activities and on brain monoamine-related substances in mice. Planta. Med. 55,
429–433.
Janssen, I., Lam, M., Katzmarzyk, P.T., 2009. Influence of overweight and obesity on physician
costs in adolescents and adults in Ontario, Canada. Obes. Rev. 10, 51-7.
Jeon, B.H., Kim, C.S., Kim, H.S., Park, J.B., Nam, K.Y., Chang, S.J., 2000A. Effect of Korean
red ginseng on blood pressure and nitric oxide production. Acta. Pharmacol. Sin. 21, 1095-100.
Jeon, B.H., Kim, C.S., Park, K.S., Lee, J.W., Park, J.B., Kim, K.J., Kim, S.H., Chang, S.J., Nam,
K.Y., 2000. Effect of Korea red ginseng on the blood pressure in conscious hypertensive rats.
Gen. Pharmacol. 35, 135-41.
Jia, L., Zhao, Y., 2009. Current evaluation of the millennium phytomedicine--ginseng (I):
etymology, pharmacognosy, phytochemistry, market and regulations. Curr. Med. Chem. 16,
2475-84.
Jung, C.H., Seog, H.M., Choi, I.W., Choi, H.D., Cho, H.Y., 2005. Effects of wild ginseng (Panax
ginseng C.A. Meyer) leaves on lipid peroxidation levels and antioxidant enzyme activities in
streptozotocin diabetic rats. J. Ethnopharmacol. 98, 245-50.
Joo, K.M., Lee, J.H., Jeon, H.Y., Park, C.W., Hong, D.K., Jeong, H.J., Lee, S.J., Lee, S.Y., Lim,
K.M., 2010. Pharmacokinetic study of ginsenoside Re with pure ginsenoside Re and ginseng
berry extracts in mouse using ultra performance liquid chromatography/mass spectrometric
method. J. Pharm. Biomed. Anal. 51, 278-83.
Kang, S.Y., Kim, S.H., Schini, V.B., Kim, N.D.,1995. Dietary ginsenosides improve
endothelium-dependent relaxation in the thoracic aorta of hypercholesterolemic rabbit.Gen.
Pharmacol. 26, 483-7.
Karaca, T., Yoruk, M., Yoruk, I.H., Uslu, S., 2010. Effects of Extracts of Green Tea and Ginseng
on Pancreatic Beta Cells and Levels of Serum Glucose, Insulin, Cholesterol and Triglycerides in

111
Rats with Experimentally Streptozotocin-Induced Diabetes: A Histochemical and
Immunohistochemical Study. JAVA. 9, 102-107.
Karu, N., Reifen, R., Kerem, Z., 2007. Weight gain reduction in mice fed Panax ginseng saponin,
a pancreatic lipase inhibitor. J. Agric. Food. Chem. 55, 2824-8.
Kassab, S., Kato, T., Wilkins, F.C., Chen, R., Hall, J.E., Granger, J.P., 1995. Renal denervation
attenuates the sodium retention and hypertension associated with obesity. Hypertension. 25(part
2), 893–897.
Kim, W.Y., Kim, J.M., Han, S.B., Lee, S.K., Kim, N.D., Park, M.K., Kim, C.K., Park, J.H.,
2000. Steaming of ginseng at high temperature enhances biological activity. J. Nat. Prod. 63,
1702-4.
Kim, S.H., and Park, K.S., 2003. Effects of Panax ginseng extract on lipid metabolism in
humans. Pharmacol. Res. 48, 511-3.
Kim, D.H., Jung, J.S., Moon, Y.S., Sung, J.H., Suh, H.W., Kim, Y.H., Song, D.K., 2003.
Inhibition of intracerebroventricular injection stress-induced plasma corticosterone levels by
intracerebroventricularly administered compound K, a ginseng saponin metabolite, in mice. Biol.
Pharm. Bull. 26, 1035-8.
Kim, J.H., Hahm, D.H., Yang, D.C., Kim, J.H., Lee, H.J., Shim, I., 2005. Effect of crude saponin
of Korean red ginseng on high-fat diet-induced obesity in the rat. J. Pharmacol. Sci. 97, 124-31.
Kim, Y., Choi, E.H., Doo, M., Kim, J.Y., Kim, C.J., Kim, C.T., Kim, I.H., 2010. Anti-stress
effects of ginseng via down-regulation of tyrosine hydroxylase (TH) and dopamine β-
hydroxylase (DBH) gene expression in immobilization-stressed rats and PC12 cells. Nutr. Res.
Pract. 4, 270-275.
Kim, H.J., Kang, H.J., Seo, J.Y., Lee, C.H., Kim, Y.S., Kim, J.S., 2011. Antiobesity effect of oil
extract of ginseng. J. Med. Food. 14, 573-583.
Kim, H.Y., Kang, K.S., Yamabe, N., Nagai, R., Yokozawa, T., 2007. Weight gain reduction in
mice fed Panax ginseng saponin, a pancreatic lipase inhibitor. J. Agric. Food. Chem. 55, 8491-7.
Kim, T.H., Jeon, S.H., Hahn, E.J., Paek, K.Y., Park, J.K., Youn, N.Y., Lee, H.L., 2009A. Effects
of tissue-cultured mountain ginseng (Panax ginseng CA Meyer) extract on male patients with
erectile dysfunction. Asian. J. Androl. 211, 356-61.
Kim, J.H., Kang, S.A., Han, S.M., Shim, I., 2009B. Comparison of the antiobesity effects of the
protopanaxadiol- and protopanaxatriol-type saponins of red ginseng. Phytother. Res. 23, 78-85.
Kim, S.J., Yuan, H.D., Chung, S.H., 2010. Ginsenoside Rg1 suppresses hepatic glucose
production via AMP-activated protein kinase in HepG2 cells. Biol. Pharm. Bull. 33, 325-8.
Kim, H.J., Yoon, K.H., Kang, M.J., Yim, H.W., Lee, K.S., Vuksan, V., Sung, M.K., 2012. A six-
month supplementation of mulberry, Korean red ginseng, and banaba decreases biomarkers of
systemic low-grade inflammation in subjects with impaired glucose tolerance and type 2
diabetes. Evid Based Complement Alternat Med. 2012:735191.
Kimura, I., Nakashima, N., Sugihara, Y., Fu-jun, C., Kimura, M., 1999. The antihyperglycaemic
blend effect of traditional chinese medicine byakko-ka-ninjin-to on alloxan and diabetic KK-
CA(y) mice. Phytother. Res. 13, 484-8.

112
Könner, A.C., Klöckener, T., Brüning, J.C., 2009. Control of energy homeostasis by insulin and
leptin: targeting the arcuate nucleus and beyond. Physiol. Behav. 97, 632-8.
Korner, P.I., Head, G.A., Badoer, E., Bobik, A., Angus, J.A., 1987. Role of brain amine
transmitters and some neuromodulators in blood pressure, heart rate, and baroreflex control. J.
Cardiovasc. Pharmacol. 12, S26-32.
Kurimoto, H., Nishijo, H., Uwano, T., Yamaguchi, H., Zhong, Y.M., Kawanishi, K., Ono, T.,
2004. Effects of nonsaponin fraction of red ginseng on learning deficits in aged rats. Physiol.
Behav. 82, 345–355.
Landsberg, L., Krieger, D.R., 1989. Obesity, metabolism and the sympathetic nervous system.
Am. J. Hypertens. 2, 125S–132S.
Lee, D.C., Lee, M.O., Kim, C.Y., Clifford, D.H., 1981. Effect of ether, ethanol and aqueous
extracts of ginseng on cardiovascular function in dogs. Can. J. Comp. Med. 45, 182-7.
Lee, T.F., Shiao, Y.J., Chen, C.F., Wang, L.C., 2001. Effect of ginseng saponins on beta-
amyloid-suppressed acetylcholine release from rat hippocampal slices. Planta. Med. 67, 634–
637.
Lee, J.H., Kim, S.R., Bae, C.S., Kim, D., Hong, H., Nah, S., 2002. Protective effect of
ginsenosides, active ingredients of Panax ginseng, on kainic acid-induced neurotoxicity in rat
hippocampus. Neurosci. Lett. 325, 129-133.
Lee, H.U., Bae, E.A., Han, M.J., Kim, N.J., Kim, D.H., 2005. Hepatoprotective effect of
ginsenoside Rb1 and compound K on tert-butyl hydroperoxide-induced liver injury. Liver Int.
25, 1069-1073.
Lee, L.S., Wise, S.D., Chan, C., Parsons, T.L., Flexner, C., Lietman, P.S., 2008. Possible
differential induction of phase 2 enzyme and antioxidant pathways by american ginseng, Panax
quinquefolius. J. Clin. Pharmacol. 48, 599-609.
Lee, J., Lee, E., Kim, D., Lee, J., Yoo, J., Koh, B., 2009A. Studies on absorption, distribution
and metabolism of ginseng in humans after oral administration. J. Ethnopharmacol. 122, 143-8.
Lee, H.J., Lee, Y.H., Park, S.K., Kang, E.S., Kim, H.J., Lee, Y.C., Choi, C.S., Park, S.E., Ahn,
C.W., Cha, B.S., Lee, K.W., Kim, K.S., Lim, S.K., Lee, H.C., 2009B. Korean red ginseng
(Panax ginseng) improves insulin sensitivity and attenuates the development of diabetes in
Otsuka Long-Evans Tokushima fatty rats. Metabolism. 58, 1170-7.
Lee, Y.S., Cha, B.Y., Yamaguchi, K., Choi, S.S., Yonezawa, T., Teruya, T., Nagai, K., Woo,
J.T., 2010. Effects of Korean white ginseng extracts on obesity in high-fat diet-induced obese
mice. Cytotechnology. 62, 367-76.
Lee, H.M., Lee, O.H., Kim, K.J., Lee, B.Y., 2011A. Ginsenoside Rg1 Promotes Glucose Uptake
Through Activated AMPK Pathway in Insulin-resistant Muscle Cells. Phytother. Res. [Epub
ahead of print]
Lee, O.H., Lee, H.H., Kim, J.H., Lee, B.Y., 2011B. Effect of ginsenosides Rg3 and Re on
glucose transport in mature 3T3-L1 adipocytes. Phytother. Res. 25, 768-73.

113
Lee, S.H., Lee, H.J., Lee, Y.H., Lee, B.W., Cha, B.S., Kang, E.S., Ahn, C.W., Park, J.S., Kim,
H.J., Lee, E.Y., Lee, H.C., 2012. Korean red ginseng (Panax ginseng) improves insulin
sensitivity in high fat fed Sprague-Dawley rats. Phytother. Res. 26, 142-7.
Leung, K.W., and Wong, A.S., 2010. Pharmacology of ginsenosides: a literature review. Chin.
Med. 11, 5-20.
Li, X., Sun, J., Wang, G., Hao, H., Liang, Y., Zheng, Y., Yan, B., Sheng, L., 2007. Simultaneous
determination of panax notoginsenoside R1, ginsenoside Rg1, Rd, Re and Rb1 in rat plasma by
HPLC/ESI/MS: platform for the pharmacokinetic evaluation of total panax notoginsenoside, a
typical kind of multiple constituent traditional Chinese medicine. Biomed. Chromatogr. 21, 735-
46.
Li, Y., Tang, J., Khatibi, N.H., Zhu, M., Chen, D., Zheng, W., Wang, S., 2010. Ginsenoside
Rbeta1 reduces neurologic damage, is anti-apoptotic, and down-regulates p53 and BAX in
subarachnoid hemorrhage. Curr. Neurovasc. Res. 7, 85-94.
Li, W., Zhang, M., Gu, J., Meng, Z.J., Zhao, L.C., Zheng, Y.N., Chen, L., Yang, G.L., 2012.
Hypoglycemic effect of protopanaxadiol-type ginsenosides and compound K on Type 2 Diabetes
mice induced by high-fat diet combining with streptozotocin via suppression of hepatic
gluconeogenesis. Fitoterapia. 83, 192-8.
Lin, E., Wang, Y., Mehendale, S., Sun, S., Wang, C.Z., Xie, J.T., Aung, H.H., Yuan, C.S., 2008.
Antioxidant protection by American ginseng in pancreatic beta-cells. Am. J. Chin. Med. 36, 981-
8.
Lim, S., Yoon, J.W., Choi, S.H., Cho, B.J., Kim, J.T., Chang, H.S., Park, H.S., Park, K.S., Lee,
H.K., Kim, Y.B., Jang, H.C., 2009. Effect of ginsam, a vinegar extract from Panax ginseng, on
body weight and glucose homeostasis in an obese insulin-resistant rat model. Metabolism. 58, 8-
15.
Lipski, J., Kanjhan, R., Kruszewska, B., Smith, M.,1995. Barosensitive neurons in the rostral
ventrolateral medulla of the rat in vivo: morphological properties and relationship to C1
adrenergic neurons. Neuroscience. 69, 601-18.
Liu, Y., Zhang, J.W., Li, W., Ma, H., Sun, J., Deng, M.C., Yang, L., 2006. Ginsenoside
Metabolites, Rather Than Naturally Occurring Ginsenosides, Lead to Inhibition of Human
Cytochrome P450 Enzymes. Toxicol. Sci. 91, 356-364.
Liu, W., Zheng, Y., Han, L., Wang, H., Saito, M., Ling, M., Kimura, Y., Feng, Y., 2008.
Saponins (Ginsenosides) from stems and leaves of Panax quinquefolium prevented high-fat diet-
induced obesity in mice. Phytomedicine. 15, 1140-5.
Liu, R., Zhang, J., Liu, W., Kimura, Y., Zheng, Y., 2010. Anti-Obesity effects of protopanaxdiol
types of Ginsenosides isolated from the leaves of American ginseng (Panax quinquefolius L.) in
mice fed with a high-fat diet. Fitoterapia. 81, 1079-87.
Liu, Y.W., Zhu, X., Li, W., Lu, Q., Wang, J.Y., Wei, Y.Q., Yin, X.X., 2012. Ginsenoside Re
attenuates diabetes-associated cognitive deficits in rats. Pharmacol. Biochem. Behav. 101, 93-8.
Lohmeier, T.E., Warren, S., Cunningham, J.T., 2003. Sustained Activation of the central
baroreceptor pathway in obesity hypertension. Hypertension. 42, 96-102.

114
Lu, T., Jiang, Y., Zhou, Z., Yue, X., Wei, N., Chen, Z., Ma, M., Xu, G., Liu, X., 2011. Intranasal
ginsenoside Rb1 targets the brain and ameliorates cerebral ischemia/reperfusion injury in rats.
Biol. Pharm. Bull. 34, 1319-24.
Luo, J.Z.,and Luo, L., 2006. American ginseng stimulates insulin production and prevents
apoptosis through regulation of uncoupling protein-2 in cultured beta cells. Evid. Based.
Complement. Alternat. Med. 3, 365-72.
Ma, S.W., Benzie, I.F., Chu, T.T., Fok, B.S., Tomlinson, B., Critchley, L.A., 2008. Effect of
Panax ginseng supplementation on biomarkers of glucose tolerance, antioxidant status and
oxidative stress in type 2 diabetic subjects: results of a placebo-controlled human intervention
trial. Diabetes. Obes. Metab. 10, 1125-7.
Matthews, D.R., Hosker, J.P., Rudenski, A.S., Naylor, B.A., Treacher, D.F., Turner, D.C., 1985.
Homeostasis model assessment: insulin resistance and beta-cell function from fasting plasma
glucose and insulin concentrations in man. Diabetologia. 28, 412-9.
Mark, A.L., Agassandian, K., Morgan, D.A., Liu, X., Cassell, M.D., Rahmouni, K., 2009. Leptin
signaling in the nucleus tractus solitarii increases sympathetic nerve activity to the
kidney. Hypertension. 53, 375-380.
Martínez-Mir, I., Rubio, E., Morales-Olivas, F.J., Palop-Larrea, V., 2004. Transient ischemic
attack secondary to hypertensive crisis related to Panax ginseng. Ann. Pharmacother. 38, 1970.
Maxwell, K.N., Soccio, R.E., Duncan, E.M., Sehayek, E., Breslow, J.L., 2003. Novel putative
SREBP and LXR target genes identified by microarray analysis in liver of cholesterol-fed mice.
J. Lipid. Res. 44, 2109-2119.
Messenger, S.A., Moreau, J.M., Ciriello, J., 2012. Intermittent hypoxia and systemic leptin
administration induces pSTAT3 and Fos/Fra-1 in the carotid body. Brain Res. 1446, 56-70.
Mehendale, S.R., Wang, C.Z., Shao, Z.H., Li, C.Q., Xie, J.T., Aung, H.H., Yuan, C.S., 2006.
Chronic pretreatment with American ginseng berry and its polyphenolic constituents attenuate
oxidant stress in cardiomyocytes. Eur. J. Pharmacol. 553, 209-14.
Migrenne, S., Le Foll, C., Levin, B.E., Magnan, C., 2011. Brain lipid sensing and nervous
control of energy balance. Diabetes Metab. 37, 83-8.
Milner, T.A., Pickel, V.M., 2003. Receptor targeting in medullary nuclei mediating baroreceptor
reflexes. Cell. Mol. Neurobiol. 23, 751-60.
Miranda, P.J., DeFronzo, R.A., Califf, R.M., Guyton, J.R., 2005. Metabolic syndrome:
definition, pathophysiology, and mechanisms. Am. Heart. J. 149, 33-45.
Mizumaki, Y., Kurimoto, M., Hirashima, Y., Nishijima, M., Kamiyama, H., Nagai, S., Takaku,
A., Sugihara, K., Shimizu, M., Endo, S., 2002. Lipophilic fraction of Panax ginseng induces
neuronal differentiation of PC12 cells and promotes neuronal survival of rat cortical neurons by
protein kinase C dependent manner. Brain. Res. 20, 254–260.
Mollah, M.L., Kim, G.S., Moon, H.K., Chung, S.K., Cheon, Y.P., Kim, J.K., Kim, K.S., 2009.
Antiobesity effects of wild ginseng (Panax ginseng C.A. Meyer) mediated by PPAR-gamma,
GLUT4 and LPL in ob/ob mice. Phytother. Res. 23, 220-5.

115
Moreno, D.A, Ilic, N, Poulev, A., Brasaemle, D.L., Fried, S.K., Raskin, I., 2003. Inhibitory
effects of grape seed extract on lipases. Nutrition. 19, 876-879.
Morgan, J.I., Cohen, D.R., Hempstead, J.L., Curran, T., 1987. Mapping patterns of c-fos
expression in the central nervous system after seizure. Science. 237, 192-197.
Morgan, J.I., Curran, T., 1990. Inducible proto-oncogenes of the nervous system: their
contribution to transcription factors and neuroplasticity. Prog. Brain. Res. 86, 287-294.
Must, A., Spadano, J., Coakley, E.H., Field, A.E., Colditz, G., Dietz, W.H., 1999. The disease
burden associated with overweight and obesity. JAMA. 282, 1523-9.
Nah, S.Y., 1997. Ginseng: Recent advances and trends. Korean J. Ginseng Sci. 21, 1–12.
Nathan, D.M., Buse, J.B., Davidson, M.B., Heine, R.J., Holman, R.R., Sherwin, R., Zinman, B.,
2006. Management of Hyperglycemia in Type 2 Diabetes: A Consensus Algorithm for the
Initiation and Adjustment of Therapy. Diabetes Care. 29, 1963-1972.
Nicol, R.W., Traquair, J.A., Bernards, M.A., 2002. Ginsenosides as host resistance factors in
American ginseng (Panax quinquefolius). Can. J. Bot. 80, 557-562.
Noble, R.E., 2001. Waist-to-hip ratio versus BMI as predictors of cardiac risk in obese adult
women. West. J. Med. 174, 240–241.
Niswender, K.D., and Schwartz, M.W., 2003. Insulin and leptin revisted: adiposity signals with
overlapping physiological and intracellular signaling capabilities. Front. Neuroendocrinol. 24, 1-
10.
Odashima S., 1986. Reverse transformation of neoplastic cells and phytoglycosides, with special
reference to redifferentiation induction in cancer cells by ginsenosides. Nihon. Rinsho. 44, 245-
53.
Ogden CL, Yanovski SZ, Carroll MD, Flegal KM. (2007). The epidemiology of Obesity.
Gastroeneterology. 132, 2087-2102.
Ginseng Ontario. “Ginseng Production”. http://ginsengontario.com/consumer-
information/ginseng-production/ (3 Jan, 2010).
Pan, C., Huo, Y., An, X., Singh, G., Chen, M., Yang, Z., Pu, J., Li, J., 2012. Panax notoginseng
and its components decreased hypertension via stimulation of endothelial-dependent vessel
dilatation. Vascul. Pharmacol. 56, 150-8.
Park, K.H., Shin, H.J., Song, Y.B., Hyun, H.C., Cho, H.J., Ham, H.S., Yoo, Y.B., Ko, Y.C., Jun,
W.T., Park, H.J., 2002. Possible role of ginsenoside Rb1 on regulation of rat liver triglycerides.
Biol. Pharm. Bull. 25, 457-60.
Park, M.Y., Lee, K.S., Sung, M.K., 2005. Effects of dietary mulberry, Korean red ginseng, and
banaba on glucose homeostasis in relation to PPAR-alpha, PPAR-gamma, and LPL mRNA
expressions. Life. Sci. 77, 3344-54.
Park, S., Ahn, I.S., Kwon, D.Y., Ko, B.S., Jun, W.K.,2008. Ginsenosides Rb1 and Rg1 suppress
triglyceride accumulation in 3T3-L1 adipocytes and enhance beta-cell insulin secretion and
viability in Min6 cells via PKA-dependent pathways. Biosci. Biotechnol. Biochem. 72, 2815-23.
Park, Y., Kwon, H.Y., Shimi, M.K., Rhyu, M.R., Lee, Y., 2011. Improved lipid profile in
ovariectomized rats by red ginseng extract. Pharmazie. 66, 450-3.

116
Persson J, Bringlöv E, Nilsson LG, Nyberg L., 2004. The memory-enhancing effects of Ginseng
and Ginkgo biloba in healthy volunteers.Psychopharmacology (Berl). 172, 430-4.
Persson, I.A., Dong, L., Persson, K., 2006. Effect of Panax ginseng extract (G115) on
angiotensin-converting enzyme (ACE) activity and nitric oxide (NO) production. J.
Ethnopharmacol. 105, 321-5.
Pilowsky, P.M., Goodchild, A.K., 2002. Baroreceptor reflex pathways and neurotransmitters: 10
years on. J. Hypertens. 20, 1675-88.
Pi-Sunyer, F.X., 1991. Health implications of obesity. Am. J. Clin. Nutr. 53, 1595S-1603S.
Popovich, D.G., and Kitts, D.D., 2004. Generation of ginsenosides Rg3 and Rh2 from North
American Ginseng. Phytochemistry. 65, 337-44.
Popovich, D.G., and Kitts, D.D., 2004. Generation of ginsenosides Rg3 and Rh2 from North
American Ginseng. Phytochemistry. 65, 337-44.
Public Health Agency Of Canada. “Report from the Canadian Chronic Disease Surveillance
System: Hypertension in Canada, 2010”. Online www.publichealth.gc.ca. Accessed July 3, 2012.
Qi, L.W., Wang, C.Z., Du, G.J., Zhang, Z.Y., Calway, T., Yuan, C.S., 2011. Metabolism of
ginseng and its interactions with drugs. Curr. Drug. Metab. 12, 818-22.
Quan, L.H., Min, J.W., Sathiyamoorthy, S., Yang, D.U., Kim, Y.J., Yang, D.C., 2012.
Biotransformation of ginsenosides Re and Rg1 into ginsenosides Rg2 and Rh1 by recombinant
β-glucosidase. Biotechnol Lett. [Epub ahead of print].
Qureshi, A.A., Abuirmeileh N., Din, Z.Z., Ahmad, Y., Burger, W.C., Elson, C.E., 1983.
Suppression of cholesterogenesis and reduction of LDL cholesterol by dietary ginseng and its
fractions in chicken liver. Atherosclerosis. 48, 81-94.
Radad, K., Gille, G., Moldzio, R., Saito, H., Rausch, W.D., 2004. Ginsenosides Rb1 and Rg1
effects on mesencephalic dopaminergic cells stressed with glutamate. Brain. Res. 1021, 41-53.
Rahmouni, K., Correia, M.L., Haynes, W.G., Mark, A.L., 2005. Obesity-associated
hypertension: New insights into mechanisms. Hypertension. 45: 9–14.
Raghavendran, H.R.B., Rekha, S., Cho, H.K., Jang, S.S., Son, C.G., 2012. Ginsenoside rich
fraction of Panax ginseng C.A. Meyer improve feeding behavior following radiation-induced
pica in rats. Filoterapia. [Epub ahead of print].
Rauscher 3rd
, F.J., Voulalas, P.J., Franza, B.R. Jr, Curran, T., 1988. Fos and Jun bind
cooperatively to the AP-1 site: reconstitution in vitro. Genes. Dev. 2, 1687-1699.
Reay, J.L., Scholey, A.B., Milne, A., Fenwick, J., Kennedy, D.O., 2008. Panax ginseng has no
effect on indices of glucose regulation following acute or chronic ingestion in healthy volunteers.
Br. J. Nutr. 101, 1673-8.
Reeds, D.N., Patterson, B.W., Okunade, A., Holloszy, J.O., Polonsky, K.S., Klein, S., 2011.
Ginseng and ginsenoside Re do not improve β-cell function or insulin sensitivity in overweight
and obese subjects with impaired glucose tolerance or diabetes. Diabetes. Care. 34, 1071-6.
Reis, D.J., Granata, A.R., Joh, T.H., Ross, C.A., Ruggiero, D.A., Park, D.H., 1984. Brain stem
catecholamine mechanisms in tonic and reflex control of blood pressure. Hypertension. 6, II7-15

117
Reis, D.J., Ruggiero, D.A., Morrison, S.F., 1989. The C1 area of the rostral ventrolateral medulla
oblongata. A critical brainstem region for control of resting and reflex integration of arterial
pressure. Am. J. Hypertens. 2, 363S-374S.
Rhee, M.Y., Kim, Y.S., Bae, J.H., Nah, D.Y., Kim, Y.K., Lee, M.M., Kim, H.Y., 2011. Effect of
Korean red ginseng on arterial stiffness in subjects with hypertension. J. Altern. Complement.
Med. 17, 45-9.
Riediger, N.D., Clara, I. Prevalence of metabolic syndrome in the Canadian adult population.
CMAJ. 183, E1127-E1134.
Rinaman, L., 2010. Ascending projections from the caudal visceral nucleus of the solitary tract to
brain regions involved in food intake and energy expenditure. Brain Res. 2, 1350:18-34.
Robitaille, C., Dai, S., Waters, C., Loukine, L., Bancej, C., Quach, S., Ellison, J., Campbell, N.,
Tu, K., Reimer, K., Walker, R., Smith, M., Blais, C., Quan, H., 2012. Diagnosed hypertension in
Canada: incidence, prevalence and associated mortality. CMAJ. 184, E49-56.
Rocchini, A.P., Yang, J.Q., Smith, M.J., Supiano, M.A., 2010. Serial changes in norepinephrine
kinetics associated with feeding dogs a high-fat diet. J. Clin. Hypertens. (Greenwich). 12, 117-
24.
Sagar, S.M., and Wong, R., 1988. The Scientific Basis of Chinese Medicine and Cancer Care: A
Western Medicine Perspective ALTERNATIVE TREATMENT FOR CANCER © World
Scientific Publishing Co. Pte. Ltd.
Schlag, E.M., McIntosh, M.S, 2006. Ginsenoside content and variation among and within
American ginseng (Panax quinquefolius L.) populations. Phytochemistry. 67, 1510-9.
Sen, S., Chen, S., Feng, B., Wu, Y., Lui, E., Chakrabarti, S., 2011. American ginseng (Panax
quinquefolius) prevents glucose-induced oxidative stress and associated endothelial
abnormalities. Phytomedicine. 18, 1110-7.
Sen, S., Chen, S., Feng, B., Wu, Y., Lui, E., Chakrabarti, S., 2012A. Preventive effects of North
American Ginseng (Panax quinquefolium) on diabetic nephropathy. Phytomedicine.. [Epub
ahead of print].
Sen, S., Chen, S., Wu, Y., Feng, B., Lui, E.K., Chakrabarti, S., 2012B. Preventive effects of
North American Ginseng (Panax quinquefolius) on Diabetic Retinopathy and Cardiomyopathy.
Phytother Res. [Epub ahead of print].
Shang, W., Yang, Y., Jiang, B., Jin, H., Zhou, L., Liu, S., Chen, M., 2007. Ginsenoside Rb1
promotes adipogenesis in 3T3-L1 cells by enhancing PPARgamma2 and C/EBPalpha gene
expression. Life. Sci. 80, 618-25.
Shang, W., Yang, Y., Zhou, L., Jiang, B., Jin, H., Chen, M., 2008. Ginsenoside Rb1 stimulates
glucose uptake through insulin-like signaling pathway in 3T3-L1 adipocytes. J. Endocrinol. 198,
561-9.
Shao, Z.H., Xie, J.T., Vanden Hoek, T.L., Mehendale, S., Aung, H., Li, C.Q., Qin, Y.,
Schumacker, P.T., Becker, L.B., Yuan, C.S., 2004. Antioxidant effects of American ginseng
berry extract in cardiomyocytes exposed to acute oxidant stress. Biochim. Biophys. Acta. 1670,
165-71.

118
Shaulian, E., Karin, M., 2002. AP-1 as a regulator of cell life and death. Nat. Cell. Biol. 4, E131-
E136.
Scheen, A.J., 2003. Current Management Strategies for Coexisting Diabetes Mellitus and
Obesity. Drugs. 63, 1165-1184.
Shibata, S., 2001. Chemistry and cancer preventing activities of ginseng saponins and some
related triterpenoid compounds. J. Korean. Med. Sci. 16, S28-37.
Siegel, R.K., 1979. Ginseng abuse syndrome: Problems with the panacea. JAMA. 241, 1614-
1615.
Sievenpiper, J.L., Sung, M.K., Di Buono, M., Seung-Lee, K., Nam, K.Y., Arnason, J.T., Leiter,
L.A., Vuksan, V., 2006. Korean red ginseng rootlets decrease acute postprandial glycemia:
results from sequential preparation- and dose-finding studies. J. Am. Coll. Nutr. 25, 100-7.
Sironi, A.M., Sicari, R., Folli, F., Gastaldelli, A., 2011. Ectopic fat storage, insulin resistance,
and hypertension. Curr. Pharm. Des. 17, 3074-80.
Smith, P.M., Ferguson, A.V., 2008. Neurophysiology of hunger and satiety. Dev. Disabil. Res.
Rev. 14, 96-104.
Smith, P.M., Ferguson, A.V., 2012. Cardiovascular actions of leptin in the subfornical organ are
abolished by diet-induced obesity. J. Neuroendocrinol. 24, 504-10.
Solano-Flores, L.P., Rosas-Arellano, M.P., Ciriello, J., 1997. Fos induction in central structures
after afferent renal nerve stimulation. Brain. Res. 753, 102-19.
Sonoda, Y., Kasahara, T., Mukaida, N., Shimizu, N., Tomoda, M., Takeda, T., 1998. Stimulation
of interleukin-8 production by acidic polysaccharides from the root of Panax ginseng.
Immunopharmacology. 38, 287-294.
Song, M., Zhang, S., Xu, X., Hang, T., Jia, L., 2010. Simultaneous determination of three Panax
notoginseng saponins at sub-nanograms by LC-MS/MS in dog plasma for pharmacokinetics of
compound Danshen tablets. J. Chromatogr. B. Analyt. Technol. Biomed. Life. Sci. 878, 3331-7.
Sotaniemi, E.A., Haapakoski, E., Rautio, A., 1995. Ginseng therapy in non-insulin-dependent
diabetic patients. Diabetes. Care. 18, 1373-5.
Statistics Canada. (2010) Body Composition of Canadian Adults 2007 to 2009.
http://www.statcan.gc.ca/pub/82-625-x/2010001/article/11091-eng.htm.
Stauss, H.M., 2002. Baroreceptor Reflex Function. Am. J. Physiol. Regul. Integr. Comp. Physiol.
283, R284-6.
Stavro, P.M., Woo, M., Heim, T.F., Leiter, L.A., Vuksan, V., 2005. North American Ginseng
exerts a neutral effect on blood pressure in individuals with hypertension. Hypertension. 46, 406-
411.
Stavro, P.M., Woo, M., Leiter, L.A., Heim, T.F., Sievenpiper, J.L., Vuksan, V., 2006. Long-term
intake of North American Ginseng has no effect on 24-hour blood pressure and renal function.
Hypertension. 47, 791-796.

119
Struthers, A.D., Dollery, C.T., 1985. Central nervous system mechanisms in blood pressure
control. Eur. J. Clin. Pharmacol. 28, 3-11.
Sugiyama, F., Churchill, G.A., Li, R., Libby, L.J., Carver, T., Yagami, K., John, S.W., Paigen,
B.,2002. QTL associated with blood pressure, heart rate, and heart weight in CBA/CaJ and
BALB/cJ mice. Physiol. Genomics.10, 5–12.
Sugiyama, F., Churchill, G.A., Higgins, D.C., Johns, C., Makaritsis, K.P., Gavras, H., Paigen, B.,
2001. Concordance of murine quantitative trait loci for salt-induced hypertension with rat and
human loci. Genomics. 71, 70–77.
Sugaya, A., Yuzurihara, M., Tsuda, T., Yasuda, K., Kajiwara, K., Sugaya, A.E., 1988.
Proliferative effect of ginseng saponin on neurite extension of primary cultured neurons of the rat
cerebral cortex. J. Ethnopharmacol. 22, 173–181.
Swanson, L.W., Sawchenko, P.E., 1980. Paraventricular nucleus: a site for the integration of
neuroendocrine and autonomic mechanisms. Neuroendocrinology. 31, 410-7.
Psaltopoulou, T., D.B. Panagiotakos, D.B., Pitsavos, C., Chrysochoou, C., Detopoulou, P.,
Skoumas, J., Stefanadis, C., 2010. Dietary antioxidant capacity is inversely associated with
diabetes biomarkers: The ATTICA study. Nutr. Metab. Cardiovasc. Dis. 21, 561-567.
Su, C.F., Cheng, J.T., Liu, I.M., 2007. Increase of acetylcholine release by Panax ginseng root
enhances insulin secretion in Wistar rats. Neurosci. Lett. 412, 101-4.
Sung, J., Han, K.H., Zo, J.H., Park, H.J., Kim, C.H., Oh, B.H., 2000. Effects of red ginseng upon
vascular endothelial function in patients with essential hypertension. Am. J. Chin. Med. 28, 205-
16.
Tawab, M.A., Bahr, U., Karas, M., Wurglics, M., Schubert-Zsilavecz, M., 2003. Degradation of
ginsenosides in humans after oral administration. Drug. Metab. Dispos. 31, 1065-71.
Thomas, G.D., 2010. Neural Control of the Circulation. Adv. Physiol. Educ. 35, 28-32.
Tohda, C., Matsumoto, N., Zou, K., Meselhy, M.R., Komatsu, K., 2002. Axonal and dendritic
extension by protopanaxadiol-type saponins from ginseng drugs in SK-N-SH cells. Jpn. J.
Pharmacol. 90, 254–262.
Van Kampen, J., Robertson, H., Hagg, T., Drobitch, R., 2003. Neuroprotective actions of the
ginseng extract G115 in two rodent models of Parkinson’s disease. Exp. Neurol. 184, 21–29.
Verberne, A.J., Stornetta, R.L., Guyenet, P.G., 1999. Properties of C1 and other ventrolateral
medullary neurones with hypothalamic projections in the rat. J. Physiol. 517, 477-94.
Vuksan, V., Sievenpiper, J.L., Koo, V.Y., Francis, T., Beljan-Zdravkovic, U., Xu, Z., Vidgen, E.,
2000. American ginseng (Panax quinquefolius L) reduces postprandial glycemia in nondiabetic
subjects and subjects with type 2 diabetes mellitus. Arch. Intern. Med. 160, 1009-13.
Vuksan, V., Sievenpiper, J.L., Wong, J., Xu, Z., Beljan-Zdravkovic, U., Arnason, J.T.,
Assinewe, V., Stavro, M.P., Jenkins, A.L., Leiter, L.A., Francis, T., 2001. American ginseng
(Panax quinquefolius L.) attenuates postprandial glycemia in a time-dependent but not dose-
dependent manner in healthy individuals. Am. J. Clin. Nutr. 73, 753-8.

120
Vuksan, V., Sievenpiper, J.L., Wong, J., Xu, Z., Beljan-Zdravkovic, U., Arnason, J.T., 2005.
Herbal remedies in the management of diabetes: lessons learned from the study of ginseng. Nutr.
Metab. Cardiovasc. Dis. 15,149-60.
Vuksan, V., Sung, M.K., Sievenpiper, J.L., Stavro, P.M., Jenkins, A.L., Di Buono, M., Lee, K.S.,
Leiter, L.A., Nam, K.Y., Arnason, J.T., Choi, M., Naeem, A., 2008. Korean red ginseng (Panax
ginseng) improves glucose and insulin regulation in well-controlled, type 2 diabetes: results of a
randomized, double-blind, placebo-controlled study of efficacy and safety. A..Nutr. Metab.
Cardiovasc. Dis. 18, 46-56.
Wang, H., Reaves, L.A., Edens, N.K., 2006. Ginseng extract inhibits lipolysis in rat adipocytes
in vitro by activating phosphodiesterase 4. J. Nutr. 136, 337-342.
Wang, J., Flaisher-Grinberg, S., Li, S., Liu, H., Sun, L., Zhou, Y., Einat, H., 2010A.
Antidepressant-like effects of the active acidic polysaccharide portion of ginseng in mice. J.
Ethnopharmacol. 132, 65-9.
Wang, J., Li, S., Fan, Y., Chen, Y., Liu, D., Cheng, H., Gao, X., Zhou, Y., 2010B. Anti-fatigue
activity of the water-soluble polysaccharides isolated from Panax ginseng C. A. Meyer. J.
Ethnopharmacol. 130, 421-3.
Wei, M., Gaskill, S.P., Haffner, S.M., Stern, M.P., 1997. Waist circumference as the best
predictor of noninsulin dependent diabetes mellitus (NIDDM) compared to body mass index,
waist/hip ratio and other anthropometric measurements in Mexican Americans--a 7-year
prospective study. Obesity. Research. 5, 16-23.
Wellman, P.J., 2000. Noradrenaline and the control of food intake. Nutrition. 16, 837-42.
Weston, M., Wang, H., Stornetta, R.L., Sevigny, C.P., Guyenet, P.G., 2003. Fos expression by
glutamatergic neurons of the solitary tract nucleus after phenylephrine-induced hypertension in
rats. J. Comp. Neurol. 460, 525-41.
Wilcox, G., 2005. Insulin and insulin resistance. Clin. Biochem. Rev. 26, 19-39.
Wood, W.B., Roh, B.L., White, R.P., 1964. Cardiovascular Actions of Panax Ginseng in Dogs.
Jpn. J. Pharmacol. 14, 284-94.
World Health Organization (WHO), 1999. Consultation on Obesity. Obesity: Preventing and
Managing the Global Epidemic. WHO Technical Report Series 894. Geneva, World Health
Organization.
World Health Organization, 1999. Definition, Diagnosis, and Classification of Diabetes Mellitus
and its Complications: Report of a WHO Consultation. Geneva, World Health Organization.
World Health Organization, 2011. Waist Circumference and Waist–Hip Ratio. Report of a WHO
Expert Consultation, Geneva, World Health Organization.
Xia, W., Sun, C., Zhao, Y., Wu, L., 2011. Hypolipidemic and antioxidant activities of sanchi
(radix notoginseng) in rats fed with a high fat diet. Phytomedicine. 18, 516-20.

121
Xiang, Y.Z, Shang, H.C., Gao, X.M., Zhang B.,L., 2008. A Comparison of the ancient use of
ginseng in traditional Chinese medicine with modern pharmacological experiments and clinical
trials. Phytother. Res. 22, 851–858.
Xie, J.T., Aung, H.H., Wu, J.A., Attel, A.S., Yuan, C.S., 2002A. Effects of American ginseng
berry extract on blood glucose levels in ob/ob mice. Am. J. Chin. Med. 30, 187-94.
Xie, J.T., Zhou, Y.P., Dey, L., Attele, A.S., Wu, J.A., Gu, M., Polonsky, K.S., Yuan, C.S.,
2002B. Ginseng berry reduces blood glucose and body weight in db/db mice. Phytomedicine. 9,
254-8.
Xie, J.T., Mehendale, S.R., Wang, A., Han, A.H., Wu, J.A., Osinski, J., Yuan, C.S., 2004A.
American ginseng leaf: ginsenoside analysis and hypoglycemic activity. Pharmacol. Res. 49,
113-7.
Xie, J.T., Wu, J.A., Mehendale, S., Aung, H.H., Yuan, C.S., 2004B. Anti-hyperglycemic effect
of the polysaccharides fraction from American ginseng berry extract in ob/ob mice.
Phytomedicine. 11, 182-7.
Xie, J.T., Wang, C.Z., Wang, A.B., Wu, J., Basila, D., Yuan, C.S., 2005A. Antihyperglycemic
effects of total ginsenosides from leaves and stem of Panax ginseng. Acta. Pharmacol. Sin. 26,
1104-10.
Xie, H.T., Wang, G.J., Sun, J.G., Tucker, I., Zhao, X.C., Xie, Y.Y., Li, H., Jiang, X.L., Wang,
R., Xu, M.J., Wang, W., 2005B. High performance liquid chromatographic-mass spectrometric
determination of ginsenoside Rg3 and its metabolites in rat plasma using solid-phase extraction
for pharmacokinetic studies. J. Chromatogr. B. Analyt. Technol. Biomed. Life. Sci. 818, 167-73.
Xie, J.T., Wang, C.Z., Ni, M., Wu, J.A., Mehendale, S.R., Aung, H.H., Foo, A., Yuan, C.S.,
2007. American ginseng berry juice intake reduces blood glucose and body weight in ob/ob
mice. J. Food. Sci. 72, S590-4.
Xie, J.T., Wang, C.Z., Li, X.L., Ni, M., Fishbein, A., Yuan, C.S., 2009. Anti-diabetic effect of
American ginseng may not be linked to antioxidant activity: comparison between American
ginseng and Scutellaria baicalensis using an ob/ob mice model. Fitoterapia. 80, 306-11.
Xiong, J., Guo, J., Huang, L., Meng, B., Ping, Q., 2008. The use of lipid-based formulations to
increase the oral bioavailability of Panax notoginseng saponins following a single oral gavage to
rats. Drug. Dev. Ind. Pharm. 34, 65-72.
Xiong, J., Sun, M., Guo, J., Huang, L., Wang, S., Meng, B., Ping, Q., 2009. Enhancement by
adrenaline of ginsenoside Rg1 transport in Caco-2 cells and oral absorption in rats. J. Pharm.
Pharmacol. 61, 347-52.
Xiong, Y., Shen, L., Liu, K.J., Tso, P., Xiong, Y., Wang, G., Woods, S.C., Liu, M., 2010.
Antiobesity and antihyperglycemic effects of ginsenoside Rb1 in rats. Diabetes. 59, 2505-12.
Xu, Zheng. “ROLE OF NITRIC OXIDE IN THE TREATMENT OF TYPE 2 DIABETES
WITH AMERICAN GINSENG (Panax quinquefolius L.” MSc. Thesis. University of Toronto.
2000.
Xu, Q.F., Fang, X.L., 2003. Pharmacokinetics and bioavailability of ginsenoside Rb1 and Rg1
from Panaxnotoginseng in rats. J Ethnopharmacol. 84, 187-192.

122
Xue, J.F., Hu, J.F., Liu, Z.J., Chen, H., Zhang, J.T., Chen, N.H., 2006. The mechanism of
ginsenosides Rg1 and Rb1 promoting glutamic acid release from PC12 cells.Yao. Xue. Xue.
Bao. 41, 1141-1145.
Yan, B., Wang, G., A, J., Xie, L., Hao, H., Liang, Y., Sun, J., Li, X., Zheng, Y., 2007.
Construction of the fingerprints of ginseng stem and leaf saponin reference substances and
spiked plasma sample by LC-ESI/MS and its application to analyzing the compounds absorbed
into blood after oral administration of ginseng stem and leaf saponin in rat. Biol. Pharm. Bull.
30, 1657-62.
Yanai, K., Sato, K., Masuda, S., Ikeda, M., Kinae, N., 2006. Utilization study of stems and
leaves of Tienchi Ginseng. I. Anti-hypertensive effect of stems and leaves of Tienchi Ginseng on
stroke-prone spontaneously hypertensive rat (SHRSP). Biosci. Biotechnol. Biochem. 70, 2501-
2507.
Yang, C.Y., Wang, J., Zhao, Y., Shen, L., Jiang, X., Xie, Z.G., Liang, N., Zhang, L., Chen, Z.H.,
2010. Anti-diabetic effects of Panax notoginseng saponins and its major anti-hyperglycemic
components. J Ethnopharmacol. 130, 231-6.
Yamamoto, M., Uemura, T., Nakama, S., Uemiya, M., Kumagai, A.,1983. Serum HDL-
cholesterol-increasing and fatty liver-improving actions of Panax ginseng in high cholesterol
diet-fed rats with clinical effect on hyperlipidemia in man. Am. J. Chin. Med. 11, 96-101.
Yim, J.S., Kim, Y.S., Moon, S.K., Cho, K.H., Bae, H.S., Kim, J.J., Park, E.K., Kim, D.H., 2004.
Metabolic activities of ginsenoside Rb1, baicalin, glycyrrhizin and geniposide to their bioactive
compounds by human intestinal microflora. Biol. Pharm. Bull. 27, 1580-3.
Yue, P.Y., Mak, N.K., Cheng, Y.K., Leung, K.W., Ng, T.B., Fan, D.T., Yeung, H.W., Wong,
R.N., 2007. Pharmacogenomics and the Yin/Yang actions of ginseng: anti-tumor,
angiomodulating and steroid-like activities of ginsenosides. Chin. Med. 15, 2-6.
Yoon, S.H., Han, E.J., Sung, J.H., Chung, S.H., 2007. Anti-diabetic effects of compound K
versus metformin versus compound K-metformin combination therapy in diabetic db/db mice.
Biol. Pharm. Bull. 30, 2196-200.
Yun, T.K.J., 2001. Brief introduction of Panax ginseng C.A. Meyer. Korean. Med. Sci. Suppl,
S3-5.
Yun, S.N., Moon, S.J., Ko, S.K., Im, B.O., Chung, S.H., 2004. Wild ginseng prevents the onset
of high-fat diet induced hyperglycemia and obesity in ICR mice. Arch. Pharm. Res. 27, 790-6.
Yun, S.N., Ko, S.K., Lee, K.H., Chung, S.H., 2007. Vinegar-processed ginseng radix improves
metabolic syndrome induced by a high fat diet in ICR mice. Arch. Pharm. Res. 30, 587-95.
Zalesin, K.C., Franklin, B.A., Miller, W.M., Peterson, E.D., McCullough, P.A., 2011. Impact of
obesity on cardiovascular disease. Med. Clin. North. Am. 95, 919-37.
Zhang, Y., Proenca, R., Maffei, M., Barone, M., Leopold, L., Friedman, J.M., 1994. Positional
cloning of the mouse obese gene and its human homologue. Nature 372 : 425 –432.
Zhang, Z., Li, X., Lv, W., Yang, Y., Gao, H., Yang, J., Shen, Y., Ning, G., 2008. Ginsenoside Re
reduces insulin resistance through inhibition of c-Jun NH2-terminal kinase and nuclear factor-
kappaB. Mol. Endocrinol. 22, 186-95.

123
Zhao, H.F., Li, Q., Li, Y., 2011. Long-term ginsenoside administration prevents memory loss in
aged female C57BL/6J mice by modulating the redox status and up-regulating the plasticity-
related proteins in hippocampus. Neuroscience. 183, 189-202.
Zierer, R., 1991. Prolonged infusion of Panax ginseng saponins into the rat does not alter the
chemical and kinetic profile of hormones from the posterior pituitary. J Ethnopharmacol. 34,
269-74.

124
Curriculum Vitae
Megan J Migchels
EDUCATION
Master of Science Physiology 2010 - 2012
University of Western Ontario, London, ON
Department of Physiology and Pharmacology
Supervisor: Dr. John Ciriello
Focus: Cardiovascular Neurobiology
Thesis: Chronic North American Ginseng Administration Alters Metabolic and Cardiovascular
Variables in the Rat
Bachelor of Medical Science (Honors) 2004 - 2009
University of Western Ontario, London, ON
Major: Medical Science
Major: Biology
Areas of concentration: Physiology, Biology, French
AWARDS
Recipient of 1st prize for a trainee oral presentation at the Cultivating Natural Bioactives
International Conference 2012, valued at $175, for the presentation: The effect of NAG
on central circuits involved in the metabolic syndrome
Recipient of Jamieson Graduate Student Travel Award (2011) valued at $1400: provided
to an exceptional graduate student to present their research findings at an international
conference
Recipient of 2nd
prize at the Physiology & Pharmacology Research Day, UWO
(November 8, 2011) valued at $100 for the poster: Chronic North American Ginseng
(NAG) Treatment Following Diet Induced Obesity Alters the Expression of Fos-related
Proteins in Medullary Cardiovascular Areas
Schulich Graduate Scholarship 2010-2012 valued at $14,008: Recognizes outstanding
scholastic achievement among graduate students
Dyche Memorial Scholarship 2004 valued at $500: Awarded to a student who excels in
science and is active in the school community

125
CERTIFICATIONS AND TRAINING
Biosafety Training 2011
Advanced Rodent Surgical Techniques 2010
Comprehensive WHMIS 2009
Animal Care and Use Training 2009
PRESENTATIONS & MEETINGS
Cultivating Natural Bioactives International Conference, London, ON (July 9-12, 2012):
oral presentation
OGIRC Journal Club, UWO, (May 18, 2012): oral presentation
OGIRC Retreat, London, ON (November 27-28, 2011): poster presentation
Society for Neuroscience conference, Washington, DC (November 12-16, 2011): poster
presented
Physiology & Pharmacology Research Day, UWO (November 8, 2011): poster presented
OGIRC Annual Retreat, Windsor, ON (June 2011): oral presentation
OGIRC Journal Club, UWO (January 24, 2011): oral presentation
Society for Neuroscience conference, San Diego, CA (November 13-17, 2010)
Physiology & Pharmacology Research Day, UWO (November 9, 2010): poster presented
OGIRC/CICMR joint conference, Ottawa, ON (October 23-25, 2010): poster presented
OGIRC Journal Club, UWO (February 25, 2010): oral presentation
PROFESSIONAL AFFILIATIONS
Member of Canadian Obesity Network 2011-present
Member of Society for Neuroscience 2010-present
Member of American Physiological Society 2010-present
Member of the Ontario Ginseng Innovation and Research Consortium 2009-present
WORK EXPERIENCE
Teaching Assistantship 2010-2011
University of Western Ontario
Department of Physiology and Pharmacology
Personal Support Worker 2008-2010
London, ON

126
REFERRED PUBLICATIONS
Published Abstracts
MIGCHELS, M.J., CIRIELLO, J. The effect of NAG on central circuits involved in the
metabolic syndrome. (Cultivating Natural Bioactives Conference, London, ON; July 9-
11, 2012).
MIGCHELS, M.J., IQBAL, W.I., MOREAU, J.M. and CIRIELLO, J. (2011) Ingestion
of North American Ginseng Decreases Circulating Levels of Leptin and Insulin. FASEB
J. 25, 853.8
MIGCHELS, M.J., IQBAL, W., MOREAU, J.M., MESSENGER, S.A. and CIRIELLO,
J. Chronic North American Ginseng treatment following diet induced obesity alters the
expression of Fos-related proteins in medullary cardiovascular areas. Program No.
717.10. Neuroscience Meeting Planner. Washington, DC: Society for Neuroscience,
2011. Online.
[J. CIRIELLO, J. M. MOREAU, S. A. MESSENGER, W. IQBAL, M. J. MIGCHELS].
[Circulatory leptin increases the activity of neurons in the nucleus of the solitary tract
(NTS) projecting to rostral ventrolateral medulla (RVLM)]. 502.09. 2011 Neuroscience
Meeting Planner. Washington, DC: Society for Neuroscience, 2011. Online.
[J. M. MOREAU, S. A. MESSENGER, M. J. MIGCHELS, W. IQBAL, J. CIRIELLO].
[Medullary autonomic areas contain leptin receptors and are activated in response to
circulating leptin]. 283.07. 2011 Neuroscience Meeting Planner. Washington, DC:
Society for Neuroscience, 2011. Online.
[S. A. MESSENGER, J. M. MOREAU, W. IQBAL, M. J. MIGCHELS, J. CIRIELLO].
[Intermittent hypoxia and circulating levels of leptin induce fos and pSTAT3
immunoreactivity in carotid body.]. 502.10. 2011 Neuroscience Meeting Planner.
Washington, DC: Society for Neuroscience, 2011. Online.
[W. IQBAL, J. M. MOREAU, M. J. MIGCHELS, S. A. MESSENGER, J. CIRIELLO].
[Gestational chronic intermittent hypoxia exposure alters growth and brain development
in neonatal sprague-dawley rats]. Program No 35.02. 2011 Neuroscience Meeting
Planner. Washington, DC: Society for Neuroscience, 2011. Online.
MIGCHELS, M.J., IQBAL, W.I., MOREAU, J.M. and CIRIELLO, J. Chronic North
American Ginseng Administration Alters Arterial Pressure and Metabolic Variables in
the Rat. (OGIRC CICMR Annual Conference, Ottawa, ON; October 23-25, 2010).
Iqbal W, Moreau JM, Migchels MJ, Ciriello J. Stanniocalcin-1 inhibits the Angiotensin-
II mediated dipsogenic response of the subfornical organ. Neuroscience Abstract Viewer,
No. 598.10, 2010. Online (Society for Neuroscience, San Diego, CA; November 13 – 17,
2010).
Moreau JM, Iqbal W, Migchels MJ, Ciriello, J. Nesfatin-1 in the subfornical organ elicits
a dipsogenic effect. Neuroscience Abstract Viewer, No. 598.16, 2010. Online (Society
for Neuroscience, San Diego, CA; November 13 – 17, 2010).
Iqbal W, Moreau JM, Migchels MJ, Messenger SA, Ciriello J. Peripheral chemoreceptor
excitation activates brainstem nesfatin-1-expressing neurons. Proc. of the Ontario
Hypertension Society Annual Spring Conference, No. P2-17, 2010. (Ontario
Hypertension Society Annual Spring Conference, Alliston, ON; May 28 – 30, 2010).




















