
A Humanized Mouse Model to Study Hepatitis C Virus Infection, Immune Response, and Liver Disease
11
A Humanized Mouse Model to Study Hepatitis C Virus Infection, Immune Response, and Liver Disease MICHAEL L. WASHBURN,* ,‡ MOSES T. BILITY,* LIGUO ZHANG,* ,§ GRIGORIY I. KOVALEV,* ADAM BUNTZMAN, JEFFERY A. FRELINGER, WALTER BARRY, ¶ ALEXANDER PLOSS, ¶ CHARLES M. RICE, ¶ and LISHAN SU* ,‡,§, *Lineberger Comprehensive Cancer Center, ‡ Curriculum in Genetics and Molecular Biology, University of North Carolina at Chapel Hill, Chapel Hill, North Carolina; § Key Laboratory of Immunity and Infection, Institute of Biophysics, Chinese Academy of Sciences, Beijing, China; Departments of Microbiology and Immunology, University of North Carolina at Chapel Hill, Chapel Hill, North Carolina; and ¶ Center for the Study of Hepatitis C, The Rockefeller University, New York, New York BACKGROUND & AIMS: Studies of hepatitis C virus (HCV) infection, immunopathogenesis, and resulting liver diseases have been hampered by the lack of a small animal model. We developed humanized mice with hu- man immune system and liver tissues to improve the studies of hepatitis C virus pathogenesis and treatment. METHODS: To promote engraftment of human hepa- tocytes, we expressed a fusion protein of the FK506 binding protein (FKBP) and caspase 8 under control of the albumin promoter (AFC8), which induces liver cell death, in Balb/C Rag2 / C-null mice. Cotransplanta- tion of human CD34 human hematopoietic stem cells (HSC) and hepatocyte progenitors into the transgenic mice led to efficient engraftment of human leukocytes and hepatocytes. We then infected these humanized mice (AFC8-hu HSC/Hep) with primary HCV isolates and studied HCV-induced immune responses and liver dis- eases. RESULTS: AFC8-hu HSC/Hep mice supported HCV infection in the liver and generated a human im- mune T-cell response against HCV. HCV infection in- duced liver inflammation, hepatitis, and fibrosis, which correlated with activation of stellate cells and expression of human fibrogenic genes. CONCLUSIONS: AFC8-hu HSC/Hep mice are a useful model of HCV infection, the immune response, and liver disease because they contain human immune system and liver cells. These mice become infected with HCV, generate a specific immune response against the virus, and develop liver diseases that include hepatitis and fibrosis. This model might also be used to develop therapeutics for HCV infection. Keywords: Animal Model of Hepatitis; Human Immunol- ogy; Fibrosis; Virology. O ver 175 million people are chronically infected by hepatitis C virus (HCV), often resulting in hepati- tis, liver fibrosis, cirrhosis, and development of hepato- cellular carcinoma (HCC). 1 The liver consists of unique subsets of antigen presenting cells and lymphocytes. 2,3 It is postulated that HCV infection in the liver leads to impaired immune response that contributes to its chronic infection in 80% of infected patients. Chronic inflammation and impaired T-cell responses in the liver are proposed to lead to liver disease. 4–7 Because of the difficulty of studying HCV infection in human patients, very little is known about how HCV infects patients and evades host immunity to establish chronic infection in the liver. HCV infection triggers robust CD4 and CD8 cytotoxic T-cell (CTL) responses in infected patients. However, the majority of HCV patients fail to resolve HCV infection and become chronically infected, associated with im- paired CD4 and CD8 T-cell functions. 4,7 Sustained CD4 T-helper activity in the blood is a hallmark of infections that are cleared spontaneously in 10%–20% of HCV- infected patients. 4,6,8 Whereas chimpanzees that resolve infection have strong CTL response, those with chronic infection show reduced CTL response. 8,9 The CTL re- sponse in chronically infected HCV patients is also re- duced. 8,10,11 During chronic HCV infection in humans and chimps, regulatory T cells are also induced to subdue the antiviral immune responses. 12,13 Thus, both CD4 and CD8 T cells are involved in HCV immunopathogenesis. However, we understand very little of human immune response to HCV infection because of lack of a robust model to study HCV infection and immunopathogen- esis. 14 Although chimpanzees have played a critical role in studying HCV infection, 15 there are several drawbacks, including low chronic infection rate and lack of liver fibrosis, as well as costs and ethical concerns, that limit Abbreviations used in this paper: AFC8, FKBP-Caspase 8 gene driven by the albumin promoter; AFC8-hu HSC/Hep, AFC8 transgenic mouse transplanted with human CD34 hematopoietic stem cells and hepa- tocyte progenitor cells; Alb, albumin; ALT, alanine aminotransferase; SMA, -smooth muscle actin; CTL, cytotoxic T cell; Fah, fumarylace- toacetate hydrolase; FKBP, FK506 binding protein; HCV, hepatitis C virus; Hep, hepatocyte progenitor cells; HIV, human immunodeficiency virus; HSC, hematopoietic stem cells; IFN, interferon; IL, interleukin; LN, lymph nodes; mAb, monoclonal antibodies; PHA, phytohemagglu- tinin; SCID, severe combined immunodeficiency; TIMP1, tissue inhibi- tor of matrix metalloproteinases 1; TNF, tumor necrosis factor; uPA, urokinase-type plasminogen activator. © 2011 by the AGA Institute 0016-5085/$36.00 doi:10.1053/j.gastro.2011.01.001 BASIC–LIVER, PANCREAS, AND BILIARY TRACT GASTROENTEROLOGY 2011;140:1334 –1344
-
Upload
independentresearcher -
Category
Documents
-
view
1 -
download
0
Transcript of A Humanized Mouse Model to Study Hepatitis C Virus Infection, Immune Response, and Liver Disease

(ma(seHmdcoHtcmidmH
Ko
tc
BA
SIC–LIV
ER,
PA
NCREA
S,A
ND
BILIA
RY
TRA
CT
GASTROENTEROLOGY 2011;140:1334–1344
A Humanized Mouse Model to Study Hepatitis C Virus Infection, ImmuneResponse, and Liver Disease
MICHAEL L. WASHBURN,*,‡ MOSES T. BILITY,* LIGUO ZHANG,*,§ GRIGORIY I. KOVALEV,* ADAM BUNTZMAN,�
JEFFERY A. FRELINGER,� WALTER BARRY,¶ ALEXANDER PLOSS,¶ CHARLES M. RICE,¶ and LISHAN SU*,‡,§,�
*Lineberger Comprehensive Cancer Center, ‡Curriculum in Genetics and Molecular Biology, University of North Carolina at Chapel Hill, Chapel Hill, North Carolina;§ �
Key Laboratory of Immunity and Infection, Institute of Biophysics, Chinese Academy of Sciences, Beijing, China; Departments of Microbiology and Immunology,University of North Carolina at Chapel Hill, Chapel Hill, North Carolina; and ¶Center for the Study of Hepatitis C, The Rockefeller University, New York, New Yorkii
ifi
t
tvvLttu
BACKGROUND & AIMS: Studies of hepatitis C virus(HCV) infection, immunopathogenesis, and resultingliver diseases have been hampered by the lack of a smallanimal model. We developed humanized mice with hu-man immune system and liver tissues to improve thestudies of hepatitis C virus pathogenesis and treatment.METHODS: To promote engraftment of human hepa-tocytes, we expressed a fusion protein of the FK506binding protein (FKBP) and caspase 8 under control ofthe albumin promoter (AFC8), which induces liver celldeath, in Balb/C Rag2�/� �C-null mice. Cotransplanta-tion of human CD34� human hematopoietic stem cellsHSC) and hepatocyte progenitors into the transgenic
ice led to efficient engraftment of human leukocytesnd hepatocytes. We then infected these humanized miceAFC8-hu HSC/Hep) with primary HCV isolates andtudied HCV-induced immune responses and liver dis-ases. RESULTS: AFC8-hu HSC/Hep mice supportedCV infection in the liver and generated a human im-une T-cell response against HCV. HCV infection in-
uced liver inflammation, hepatitis, and fibrosis, whichorrelated with activation of stellate cells and expressionf human fibrogenic genes. CONCLUSIONS: AFC8-huSC/Hep mice are a useful model of HCV infection,
he immune response, and liver disease because theyontain human immune system and liver cells. Theseice become infected with HCV, generate a specific
mmune response against the virus, and develop liveriseases that include hepatitis and fibrosis. Thisodel might also be used to develop therapeutics forCV infection.
eywords: Animal Model of Hepatitis; Human Immunol-gy; Fibrosis; Virology.
Over 175 million people are chronically infected byhepatitis C virus (HCV), often resulting in hepati-
is, liver fibrosis, cirrhosis, and development of hepato-ellular carcinoma (HCC).1 The liver consists of unique
subsets of antigen presenting cells and lymphocytes.2,3 Itis postulated that HCV infection in the liver leads toimpaired immune response that contributes to its
chronic infection in 80% of infected patients. Chronicinflammation and impaired T-cell responses in the liverare proposed to lead to liver disease.4 –7 Because of thedifficulty of studying HCV infection in human patients,very little is known about how HCV infects patients andevades host immunity to establish chronic infection inthe liver.
HCV infection triggers robust CD4 and CD8 cytotoxicT-cell (CTL) responses in infected patients. However, themajority of HCV patients fail to resolve HCV infectionand become chronically infected, associated with im-paired CD4 and CD8 T-cell functions.4,7 Sustained CD4�
T-helper activity in the blood is a hallmark of infectionsthat are cleared spontaneously in 10%–20% of HCV-infected patients.4,6,8 Whereas chimpanzees that resolvenfection have strong CTL response, those with chronicnfection show reduced CTL response.8,9 The CTL re-
sponse in chronically infected HCV patients is also re-duced.8,10,11 During chronic HCV infection in humansand chimps, regulatory T cells are also induced to subduethe antiviral immune responses.12,13 Thus, both CD4 andCD8 T cells are involved in HCV immunopathogenesis.However, we understand very little of human immuneresponse to HCV infection because of lack of a robustmodel to study HCV infection and immunopathogen-esis.14
Although chimpanzees have played a critical role instudying HCV infection,15 there are several drawbacks,ncluding low chronic infection rate and lack of liverbrosis, as well as costs and ethical concerns, that limit
Abbreviations used in this paper: AFC8, FKBP-Caspase 8 gene drivenby the albumin promoter; AFC8-hu HSC/Hep, AFC8 transgenic mousetransplanted with human CD34� hematopoietic stem cells and hepa-ocyte progenitor cells; Alb, albumin; ALT, alanine aminotransferase;
�SMA, �-smooth muscle actin; CTL, cytotoxic T cell; Fah, fumarylace-oacetate hydrolase; FKBP, FK506 binding protein; HCV, hepatitis Cirus; Hep, hepatocyte progenitor cells; HIV, human immunodeficiencyirus; HSC, hematopoietic stem cells; IFN, interferon; IL, interleukin;N, lymph nodes; mAb, monoclonal antibodies; PHA, phytohemagglu-inin; SCID, severe combined immunodeficiency; TIMP1, tissue inhibi-or of matrix metalloproteinases 1; TNF, tumor necrosis factor; uPA,rokinase-type plasminogen activator.
© 2011 by the AGA Institute0016-5085/$36.00
doi:10.1053/j.gastro.2011.01.001

pdtmpcm
t
test
wttdhU
i
weEtCi
c
BA
SIC–L
IVER
,PA
NCREA
S,A
ND
BIL
IARY
TRA
CT
April 2011 HCV PATHOGENESIS IN A HUMANIZED MOUSE MODEL 1335
their utility. A mouse engrafted with human liver cellsand a functional human immune system will be an ex-cellent model to study the virus.14 A number of human-mouse chimeric liver models have been developed butallow analyses of only limited aspects of HCV infectionand pathogenesis.16,17 The albumin (Alb)-urokinase-type
lasminogen activator (uPA)/severe combined immuno-eficiency (SCID) transgenic mouse contains the uPAransgene under control of an Alb promoter, and ho-
ozygous animals are unhealthy and die because ofrofound hypofibrinogenemia and accelerated hepato-yte death, which can be rescued by transplantation ofouse or human hepatocytes.17,18 The Alb-uPA/SCID
mouse with efficient human hepatocyte engraftmentcan be infected with HCV.17 It was reported recentlyhat fumarylacetoacetate hydrolase (Fah)-Rag2-�C-
null mice can also be highly engrafted with humanhepatocytes to support HCV infection.19 –21 However,because of lack of a functional immune system, it isnot possible to study HCV immunopathogenesis, andno liver diseases are observed in the Alb-uPA/SCID orFah-Rag2-�C-null model.17,21
The Balb/C Rag2-�C-null mouse supports develop-ment of a functional human immune system after inject-ing CD34� human hematopoietic stem cells (HSC).22 Tofacilitate engraftment of human liver cells in Balb/CRag2-�C-null mice, we expressed the active Caspase 8fused with FK506 binding domain (FKBP) with induciblesuicidal activity in mouse liver cells under control of thealbumin promoter (AFC8 mice).23 We cotransplanted hu-man hepatocyte progenitor cells (Hep) and CD34� HSCinto the transgenic mice and treated them with the FKBPdimerizer. Human immune cells and human hepatocyteswere both efficiently developed, thereby generating amouse model containing both a human immune systemand human liver cells (AFC8-hu HSC/Hep). AFC8-huHSC/Hep mice supported HCV infection in the liver andgenerated human T-cell response to HCV. In addition,HCV infection induced liver inflammation and fibrosisand correlated with activation of stellate cells and expres-sion of human fibrogenic genes.
Materials and MethodsPlasmidsThe FKBP-Caspase 8 gene was generated by clon-
ing active human Caspase 8 (fragment Ser217-Asp479,kindly provided by Dr Terry Combs, University of NorthCarolina [UNC]-Chapel Hill, NC) into the pC4M-Fv2Evector (Ariad Pharmaceuticals, Cambridge, MA) to ex-press the FKBP-Caspase 8 (FC8) fusion protein.23 Theransgenic construct was generated with the Alb enhanc-r/promoter24 (kindly provided by Dr Snorri Thorges-ons; National Institutes of Health, Bethesda, MD) con-
rolling expression of the FKBP-Caspase 8 gene.Generation of Transgenic Mice With FKBP-Caspase 8 Gene Driven by the Alb PromoterAnimal procedures were approved by the Univer-
sity of North Carolina Institutional Animal Care and UseCommittee. Balb/C Rag2�/��C-null mutant female mice
ere super-ovulated, and the fertilized eggs were isolatedo generate transgenic mice by standard procedures inhe UNC transgenic core facility. FKBP-Caspase 8 generiven by the Alb promoter (AFC8) mice were bred andandled under specific pathogen free conditions in theNC DLAM facility.
Induction of Murine Hepatocyte Death inAFC8 MiceWe injected mice intraperitoneally or intrave-
nously with AP20187 (Ariad Pharmecuticals) or vehicleonly (4% ethanol, 10% polyethylene glycol– 400, 2%Tween-20 in water) at a dose of 5 �g/g body weight atndicated days post-transplant of human cells.23 Alanine
aminotransferase (ALT) levels were measured in theblood of treated mice.
Human Hepatocyte Progenitor Cells andCD34� HSC From Human Fetal LiverHuman hepatocyte progenitor and CD34� cells
ere isolated from 15- to 18-week-old fetal liver tissuessentially as described.25–27 Parenchyma cells (30%pCAM� hepatoblasts/progenitors) were separated from
he nonparenchyma cells (including CD34 HSC cells).D34� cells were isolated by magnetic-activated cell sort-
ng, and the purity was greater than 95%.
Transplantation of AFC8 Mice With HumanCD34 HSC and Liver Progenitor CellsCD34� HSCs (0.5–1 � 106) and Hep progenitor
ells (0.5–1 � 106) from the same donor were coinjectedinto the liver of 1- to 5-day-old newborn AFC8 andlittermate control mice, previously irradiated at 400rad.25,27 We then injected AFC8-hu mice with AP20187(Ariad Pharmecuticals) at a dose of 5 �g/g body weight asdescribed above.
Blood and Tissue AnalysisAFC8-hu HSC/Hep mice were bled to measure
human leukocyte reconstitution at 12–16 weeks post-transplantation. At termination, liver tissue was snapfrozen in RNAlater (Qiagen, Valencia, CA) or fixed in 10%formalin. Liver sections were stained with H&E, SiriusRed, and Fast Green or with antibodies: mouse mono-clonal anti-human � smooth muscle actin (1A4, Dako,Carpinteria, CA), mouse monoclonal anti-human CD45(2B11 � PD7/26; Dako), and anti-human Alb (Dako).Immunoreactivity was determined by incubation with3,3’-diaminobenzidine tetrahydrochloride substrate(Pierce, Rockford, IL) or Vulcan red (Dako) and counter-
stained with hematoxilin.25,27
uifw
shdl
BA
SIC–LIV
ER,
PA
NCREA
S,A
ND
BILIA
RY
TRA
CT
1336 WASHBURN ET AL GASTROENTEROLOGY Vol. 140, No. 4
Human-Specific Gene ExpressionHuman- or mouse-specific quantitative real-time
polymerase chain reaction (PCR) primers were designedusing National Center for Biotechnology Informationprimer design program and Blast database (see Supple-mentary Figures 3 and 6). Gene expression analysis wasexamined using Thermo Scientific SYBR Green real-timePCR reagents. Values shown are relative gene expressionnormalized to specific glyceraldehyde-3-phosphate dehy-drogenase.
HCV Infection of AFC8-hu HSC/Hep MiceAFC8-hu HSC/Hep mice or control mice were
inoculated intravenously with 100 �L of clinical isolates
Figure 1. AFC8 mice support engraftment of human hepatocytes iFKBP-Caspase 8. The chemical dimerizer AP20187 leads to activation oin AFC8 transgenic mice. AFC8 or littermate control mice (4–6 weeksserum at �1, 0, 1, 3, 6, and 7 days postdrug treatment. Data are repretandard deviation. *P � .05. (C) Liver sections from control mice withouuman HSC/Hep (AFC8-hu HSC/Hep) were stained with anti-humaispersed throughout all lobes of the liver. (D) Human Alb positive cells
ittermate control mice were counted, and the percentage of Alb� was calcu
of HCV genotype 1a (1–5 � 107 genome copies/mL) orcontrol human sera. Mouse blood and liver tissues wereharvested to prepare RNA for measuring HCV genomicRNA as reported.28
T-Cell Response AnalysisLeukocytes from various tissues of AFC8-hu mice
were analyzed by fluorescence-activated cell sorter forhuman surface markers. Total spleens and lymph nodecells containing 1 � 105 human CD45� cells were stim-
lated for 20 hours with 10 �g/mL phytohemagglutininn Iscove’s Modified Dulbecco’s Medium (IMDM) � 10%etal bovine serum media (Gibco, Carlsbad, CA). The cellsere then stained for human T cells (CD45, CD3, CD4,
liver. (A) Inducible activation of Caspase 8 through dimerization ofpase 8 and apoptosis. (B) Mouse liver injury can be repeatedly inducedere injected with AP20187 at day 0 and day 6. ALT was measured in
ative of 3 experiments with n � 4 per group. Data represent means �splant (left) and both control (middle) and AFC8 (right) transplanted withantibody. Human hepatocytes were present around the veins and
otal nucleated cells in the liver of humanized AFC8 and nontransgenic
n thef Cas
old) wsentt trann Alband t
lated (n � 16 mice per group, **P � .01).
mst
aiWt1
BA
SIC–L
IVER
,PA
NCREA
S,A
ND
BIL
IARY
TRA
CT
April 2011 HCV PATHOGENESIS IN A HUMANIZED MOUSE MODEL 1337
CD8) and for intracellular human interleukin (IL)-2, tu-mor necrosis factor (TNF)-�, and interferon (IFN)-�. Todetect HCV-specific human T-cell response, human Tcells were expanded by stimulating spleen/lymph node(LN) cells with a pool of 19 HCV core peptides (20meroverlapping by 10) at 10 �g/mL each � 1 �g/mL anti-CD28 monoclonal antibodies (mAb). The cells were thencultured for 8 –10 days with expansion medium (IMDM;10% fetal bovine serum, 10 U/mL human IL-2, and 125ng/mL IL-7). The cells were restimulated with the sameHCV core peptide pool for 18 hours and stained as above.
Statistical AnalysisWe used unpaired 2-tailed Student t tests for all
comparisons. P � .05 is considered significant. All dataare reported as means � standard deviation or standarderror of mean as indicated.
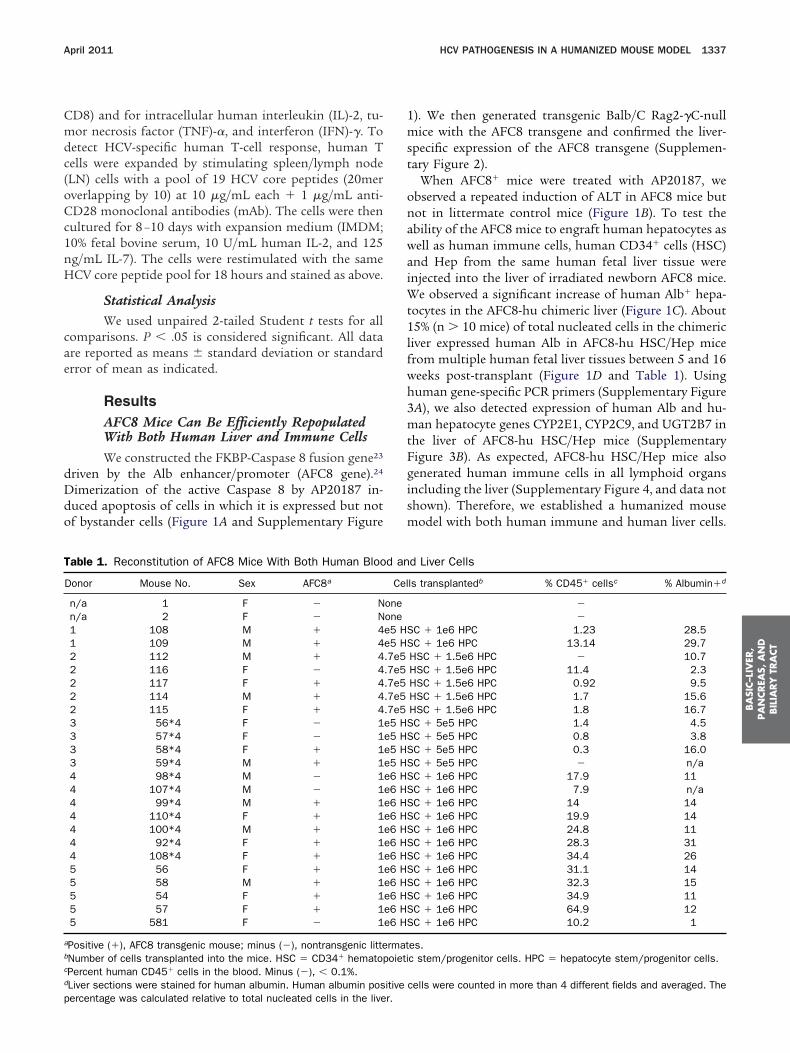
ResultsAFC8 Mice Can Be Efficiently RepopulatedWith Both Human Liver and Immune CellsWe constructed the FKBP-Caspase 8 fusion gene23
driven by the Alb enhancer/promoter (AFC8 gene).24
Dimerization of the active Caspase 8 by AP20187 in-duced apoptosis of cells in which it is expressed but notof bystander cells (Figure 1A and Supplementary Figure
Table 1. Reconstitution of AFC8 Mice With Both Human Bloo
Donor Mouse No. Sex AFC8a
n/a 1 F � Nn/a 2 F � N1 108 M � 41 109 M � 42 112 M � 42 116 F � 42 117 F � 42 114 M � 42 115 F � 43 56*4 F � 13 57*4 F � 13 58*4 F � 13 59*4 M � 14 98*4 M � 14 107*4 M � 14 99*4 M � 14 110*4 F � 14 100*4 M � 14 92*4 F � 14 108*4 F � 15 56 F � 15 58 M � 15 54 F � 15 57 F � 15 581 F � 1
aPositive (�), AFC8 transgenic mouse; minus (�), nontransgenic littebNumber of cells transplanted into the mice. HSC � CD34� hematopcPercent human CD45� cells in the blood. Minus (�), � 0.1%.dLiver sections were stained for human albumin. Human albumin pos
percentage was calculated relative to total nucleated cells in the liver.1). We then generated transgenic Balb/C Rag2-�C-nullice with the AFC8 transgene and confirmed the liver-
pecific expression of the AFC8 transgene (Supplemen-ary Figure 2).
When AFC8� mice were treated with AP20187, weobserved a repeated induction of ALT in AFC8 mice butnot in littermate control mice (Figure 1B). To test theability of the AFC8 mice to engraft human hepatocytes aswell as human immune cells, human CD34� cells (HSC)
nd Hep from the same human fetal liver tissue werenjected into the liver of irradiated newborn AFC8 mice.
e observed a significant increase of human Alb� hepa-ocytes in the AFC8-hu chimeric liver (Figure 1C). About5% (n � 10 mice) of total nucleated cells in the chimeric
liver expressed human Alb in AFC8-hu HSC/Hep micefrom multiple human fetal liver tissues between 5 and 16weeks post-transplant (Figure 1D and Table 1). Usinghuman gene-specific PCR primers (Supplementary Figure3A), we also detected expression of human Alb and hu-man hepatocyte genes CYP2E1, CYP2C9, and UGT2B7 inthe liver of AFC8-hu HSC/Hep mice (SupplementaryFigure 3B). As expected, AFC8-hu HSC/Hep mice alsogenerated human immune cells in all lymphoid organsincluding the liver (Supplementary Figure 4, and data notshown). Therefore, we established a humanized mousemodel with both human immune and human liver cells.
d Liver Cells
ls transplantedb % CD45� cellsc % Albumin�d
��
C � 1e6 HPC 1.23 28.5C � 1e6 HPC 13.14 29.7
HSC � 1.5e6 HPC � 10.7HSC � 1.5e6 HPC 11.4 2.3HSC � 1.5e6 HPC 0.92 9.5HSC � 1.5e6 HPC 1.7 15.6HSC � 1.5e6 HPC 1.8 16.7C � 5e5 HPC 1.4 4.5C � 5e5 HPC 0.8 3.8C � 5e5 HPC 0.3 16.0C � 5e5 HPC � n/aC � 1e6 HPC 17.9 11C � 1e6 HPC 7.9 n/aC � 1e6 HPC 14 14C � 1e6 HPC 19.9 14C � 1e6 HPC 24.8 11C � 1e6 HPC 28.3 31C � 1e6 HPC 34.4 26C � 1e6 HPC 31.1 14C � 1e6 HPC 32.3 15C � 1e6 HPC 34.9 11C � 1e6 HPC 64.9 12C � 1e6 HPC 10.2 1
es.c stem/progenitor cells. HPC � hepatocyte stem/progenitor cells.
cells were counted in more than 4 different fields and averaged. The
d an
Cel
oneonee5 HSe5 HS.7e5.7e5.7e5.7e5.7e5e5 HSe5 HSe5 HSe5 HSe6 HSe6 HSe6 HSe6 HSe6 HSe6 HSe6 HSe6 HSe6 HSe6 HSe6 HSe6 HS
rmatoieti
itive

(5
ws
d
Cca
p
BA
SIC–LIV
ER,
PA
NCREA
S,A
ND
BILIA
RY
TRA
CT
1338 WASHBURN ET AL GASTROENTEROLOGY Vol. 140, No. 4
HCV Infection Leads to Liver Infiltration andHepatitis in AFC8-hu HSC/Hep MiceWe inoculated AFC8-hu mice with patient HCV
isolates (genotype 1a) or control human serum (Table 2).We detected HCV genomic RNA in the liver of about 50%HCV-infected AFC8-hu mice (Figure 2A and Table 2) attermination (1– 4 months postinfection) but were unableto detect HCV viremia in the blood of infected mice (datanot shown). As controls, we did not detect HCV RNA inmock-infected AFC8-hu HSC/Hep mice or in HCV-infected AFC8 mice with no human cell transplant (Fig-ure 2A).
When ALT levels were measured at various times afterHCV infection, significant induction of ALT was detectedin HCV-infected AFC8-hu mice after 2– 4 weeks postin-fection but not in control mice (Figure 2B). Consistentwith elevated ALT levels, we observed significant leuko-cyte infiltration into the liver in HCV infected AFC8-huHSC/Hep mice (Figure 2C). We observed an increase ofhuman CD45� leukocyte, including CD68� macrophagesand CD3� T cells, in the liver of HCV-infected miceFigure 2C, lower panel, and Supplementary Figure
Table 2. HCV Infection and Liver Pathology in AFC8-hu HSC/
Mouse no. HCVHCV particles
inoculated
1 PBS None2 PBS None87 PBS None3 HCV patient 1 �9e54 HCV patient 1 �9e5110d HCV patient 1 �9e5136d,e HCV patient 1 �9e5137d,e HCV patient 1 �9e5138d HCV patient 1 �9e5139d HCV patient 1 �9e5108 HCV patient 1 �9e5109 HCV patient 1 �9e5116f Human sera None117 Human sera None88*4 Human sera None89*4 Human sera None99*4 Human sera None110*4 Human sera None90*4 HCV patient 2 �5e591*4 HCV patient 2 �5e592*4 HCV patient 2 �5e598*4 HCV patient 2 �5e5100*4 HCV patient 2 �5e5108*4 HCV patient 2 �5e5
.i., postinfection.aMice killed because of experimental end points or poor health on dbDetermined by analysis of Sirius Red stained liver sections. 0 � nonumerous septa without cirrhosis; 4 � cirrhosis.cHCV viral load in the liver tissue. �(1–8) mouse number from FiguredAFC8 mice reconstituted with human adult hepatocytes.eMice found dead (analyzed between 12 and 24 hours after death) ofMouse number 116 is a nontransgenic mouse.
A). When total intrahepatic leukocytes were analyzed,
e detected an increase of multiple human leukocyteubsets including CD3�CD4� and CD3�CD8� T cells,
CD3�CD56� natural killer cells, and plasmacytoidendritic cells (CD123�CD4�CD3�) in the liver of
HCV-infected mice (Supplementary Figure 5B). Similarinfiltrations have been reported in HCV-infected hu-mans.29 –31 Thus, AFC8-hu HSC/Hep mice were able tosupport HCV infection in the liver, leading to liverinfiltration and injury.
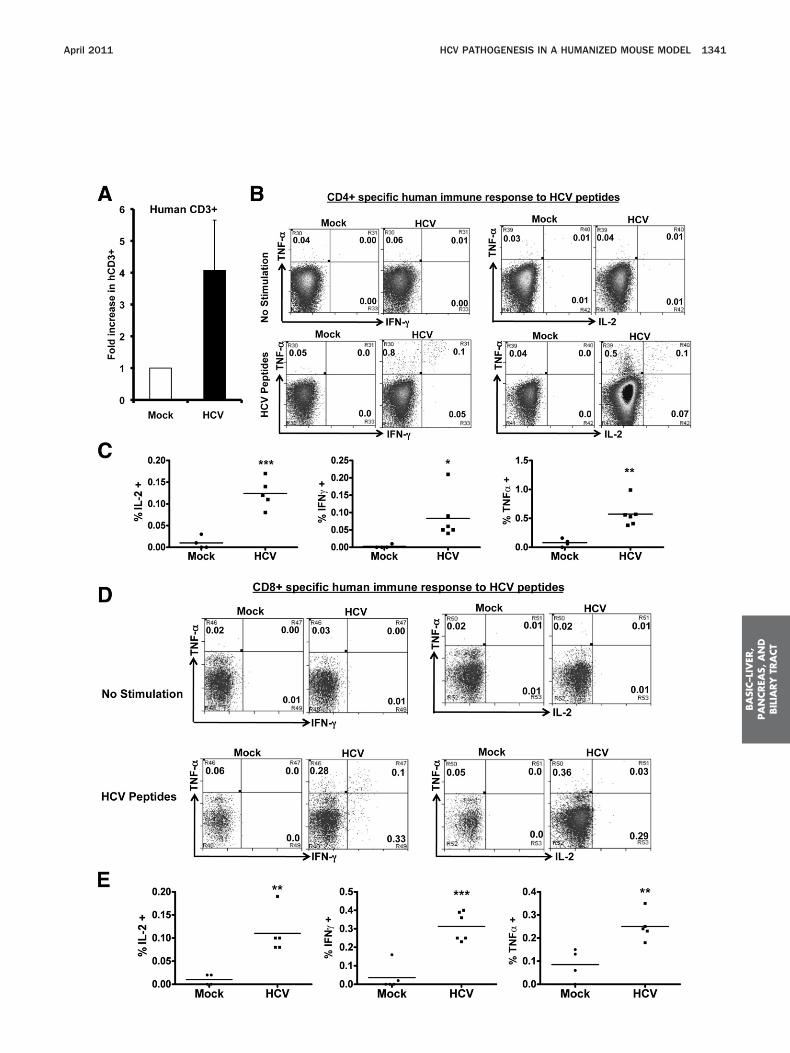
HCV Infection Induces HCV-Specific HumanT-Cell Immune Response
To characterize the human T-cell response inHCV-infected AFC8-hu HSC/Hep mice, we stimulated(phytohemagglutinin for 12 hours) spleen and LN cellsfrom mock- or HCV-infected mice and measured humaneffector cytokines (IL-2, IFN-�, and TNF-�) in human
D4� and CD8� T cells. We observed a significant in-rease of all 3 cytokines in both CD4� (Figure 3A and B)nd CD8� (Figure 3C) T cells from HCV-infected
AFC8-hu HSC/Hep mice. Therefore, HCV infection
Mice
Days p.i.aMetavirscoreb
Viral load in liver tissue(�103 copies/�g RNA)c
80 0 n/d110 0 n/d110 1 n/d*1
110 1 n/d110 1 n/d110 1 18*5
93 2 87*6
89 2 180*8
110 1 n/d*2
80 1 n/d*3
32 3 164*7
80 3 14*4
62 1 n/d48 1 n/d97 1 n/d97 1 n/d69 1 n/d69 1 n/d34 3 32357 2 16897 4 n/d69 1 n/d69 2 34797 2 n/d
ost-HCV injection.sis; 1 � only portal fibrosis; 2 � portal fibrosis with few septa; 3 �
n/d � not detectable.
indicated days postinfection.
Hep
ays pfibro
2A.
n the
primed human T cells in AFC8-hu HSC/Hep mice.

mr
fIHt
wHdi2S
i
BA
SIC–L
IVER
,PA
NCREA
S,A
ND
BIL
IARY
TRA
CT
April 2011 HCV PATHOGENESIS IN A HUMANIZED MOUSE MODEL 1339
To detect HCV-specific T-cell response, we stimulatedthe spleen/LN cells from mock and HCV-infected micewith an overlapping peptide pool from the HCV coreregion in the presence of anti-CD28 mAb. After 8 –10days in culture, we observed a 4- to 5-fold preferentialexpansion of human CD3� T cells from HCV-infected
ice relative to mock control mice (Figure 4A). Whenestimulated with HCV core peptides, both human CD4�
(Figure 4B and C) and CD8� (Figure 4D and E) T cellsrom HCV-infected mice also responded to produceFN-�, IL-2, and TNF-�. Thus, HCV infection in AFC8-huSC/Hep mice induced specific human T-cell response
o HCV antigens.
AFC8-hu HSC/Hep Mice Develop LiverFibrosis After HCV InfectionWe then assessed liver pathology in HCV-infected
Figure 2. HCV infection in AFC8-hu HSC/Hep mice leads to human immnfected with HCV patient isolates (genotype 1a, 1 � 10e5 IU/mouse) orLiver samples were collected at termination time points. (A) HCV genomwere harvested from HCV-infected AFC8-hu or control mice at 32–11liver-derived RNA in triplicates. *P � .05. (B) Increased ALT in sera fromerror of mean (n � 6 mice per group). *P � .05. (C) Representative liveand 2 AFC8-hu/HCV mice were stained to detect leukocyte infiltrationCD68 (bottom panels).
AFC8-hu HSC/Hep mice. About 50% of HCV-infected
AFC8-hu HSC/Hep mice had severe fibrosis throughoutthe liver parenchyma (Figure 5A, top panels, and Table 2),
hereas mock-inoculated AFC8-hu HSC/Hep mice andCV-inoculated AFC8 mice without human transplantsid not develop fibrosis. Infiltrated human CD45� cells
n the liver were detected in the fibrotic regions (Figure). In the absence of a functional immune system, uPA/CID or Fah-Rag-�C-null mice transplanted with only
human hepatocytes supported HCV infection but nosignificant liver fibrosis.17,21 We analyzed HCV infectionand liver pathology in 5 AFC8-hu mice transplanted withonly human adult hepatocytes and did not detect signif-icant liver fibrosis (Table 2). The 2 AFC8-hu Hep micewith low-grade fibrosis were analyzed 12–24 hours afterunexpected death.
In human patients, HCV infection leads to activation
infiltration and liver injury. AFC8-hu HSC/Hep mice or control mice wereol serum. Blood samples were collected at various times after infection.A was detected in the liver of HCV-infected AFC8-hu mice. Liver tissuess postinfection. Values represent HCV RNA copy number/microgram
V-infected AFC8-hu or control mice. Data represent mean � standardtions from control/mock, AFC8-hu/mock, control (no transplant)/HCV,H&E (top panels) or with anti-human CD45 (middle panels) or human
unecontric RN0 day
HCr secwith
of hepatic stellate cells, which contributes to liver fibro-

hai(aa(t
a
Hp
op
BA
SIC–LIV
ER,
PA
NCREA
S,A
ND
BILIA
RY
TRA
CT
1340 WASHBURN ET AL GASTROENTEROLOGY Vol. 140, No. 4
sis.32 We stained liver sections with the mouse anti-uman �-smooth muscle actin (�SMA) mAb to detectctivated stellate cells. We observed activated stellate cellsn the liver of HCV-infected AFC8-hu HSC/Hep miceFigure 5A, bottom panels). Activated hepatic stellate cellslso express extracellular matrix protein Collagen 1A1nd tissue inhibitor of matrix metalloproteinases 1TIMP1), which inhibits degradation of extracellular ma-rix and fibrosis resolution.33 We thus analyzed the ex-
pression of these genes in the liver with human-specificor mouse-specific PCR primers (Supplementary Figure 6).Interestingly, we detected specific induction of humanTIMP1 (Figure 5B) and human Collagen 1A1 (Figure 5C),
Figure 3. HCV infection induces human T-cell response in AFC8-hu mmice at 6–8 weeks postinfection were stimulated for 12–16 hours wit(CD3, CD4, and CD8) and for intracellular human cytokines (IL-2, IFN-�of effector cytokines. Representative fluorescence-activated cell sortecytokine� of CD4 or CD8 T cells. (B) Increase of human CD4 effector Tre shown. (C) Increase of human CD8 effector T cells in HCV infect
expression of effector cytokines. Data represent mean � standard erro***P � .001.
™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™Figure 4. HCV infection induces HCV-specific human T-cell respoHCV-infected AFC8-hu mice after stimulation with HCV Core peptidestimulated with a pool of HCV Core peptides plus anti-CD28 mAb an
CV-infected samples (n � 4) was calculated relative to mock sampleeptides and stained for intracellular human cytokines (IL-2, IFN-�, and T
No stimulation controls show background. (C) Summarized results of indf the cytokines. Representative fluorescence-activated cell sorter plotercent cytokine� of CD4 or CD8 T cells. (E) Shown are summarized da
the mean values. *P � .05; **P � .01; ***P � .001.
but not mouse fibrosis genes, in the liver of some HCV-infected AFC8-hu HSC/Hep mice. These results suggestthat HCV infection activated human stellate cells andinduced expression of human fibrogenic genes in thechimeric liver of AFC8-hu HSC/Hep mice.
DiscussionWe report here a novel mouse model that sup-
ports efficient engraftment of both human immune cellsand human liver cells. The Balb/C Rag2-�C-null mousewith the AFC8 transgene enabled us to inducibly depletemurine hepatocytes. In addition to human immune cells
Spleen and lymph node cells from mock- and HCV-infected AFC8-hutohemagglutinin. Cells were stained with antibodies for human T cellsTNF-�). (A) Human CD3� CD4� T cells were analyzed for expression
s are shown for each cell population. Numbers represent the percentin HCV-infected AFC8-hu mice. Summarized data (n � 4 mice/group)C8-hu mice. Human CD3� CD8� T cells were similarly analyzed forean from 2 cohorts of mice with n � 4 per group. *P � .05; **P � .01;
™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™3n AFC8-hu mice. (A) Preferential expansion of human T cells fromleen and LN cells from mock and HCV-infected AFC8-hu mice weretured for 8–10 days. Fold increase of human CD45�CD3� cells from
3). P � .05. (B–E) The expanded T cells were restimulated with HCV). (B) Human CD4� T cells are analyzed for expression of the cytokines.
al mice are shown. (D) Human CD8� T cells are analyzed for expressionshown for each cell population. Numbers in FACS plots represent theindividual mice from 2 cohorts of mice (n � 4 per group). Lines indicate
ice.h phy, and
r plotcells
ed AFr of m
™™™nse is. Spd culs (n �NF-�ividu
s areta of
BA
SIC–L
IVER
,PA
NCREA
S,A
ND
BIL
IARY
TRA
CT
April 2011 HCV PATHOGENESIS IN A HUMANIZED MOUSE MODEL 1341

cwl
S
pFchtiftica
pTml
gg(HHoHum
h
BA
SIC–LIV
ER,
PA
NCREA
S,A
ND
BILIA
RY
TRA
CT
1342 WASHBURN ET AL GASTROENTEROLOGY Vol. 140, No. 4
in lymphoid and liver organs, AFC8-hu HSC/Hep micewere also efficiently repopulated with human Alb� liverells. AFC8-hu HSC/Hep mice supported HCV infection,hich induced HCV-specific human immune response,
iver infiltration, hepatitis, and fibrosis.In the absence of a functional immune system, uPA/
CID or Fah-Rag-�C-null mice transplanted with onlyhuman hepatocytes supported HCV infection but nosignificant liver fibrosis.17,21 Immunodeficient mice ex-
ressing the uPA transgene in the liver or carrying theah mutation allow human adult hepatocytes to effi-iently repopulate the liver.17–20 These mice have poorealth including neonatal death and, most importantly,hey lack a human immune system. The severe livernjury in these mice may impair the development andunction of human liver and immune cells. To overcomehese deficiencies, the AFC8 mouse enabled us to induc-bly deplete murine hepatocytes through a programmedell death mechanism without bystander cell killing. In
Figure 5. AFC8-hu HSC/Hep mice develop liver fibrosis after HCV infAFC8 (no transplant)/HCV, and 3 AFC8-hu/HCV mice were stained witwere also stained with a mouse monoclonal antibody against human �cells. Human- or murine-specific gene expression of TIMP1 (B) and Collaprimers. Values shown are relative gene expression normalized to humarepresent mean � standard error of mean.
ddition, the Rag2-�C-null mouse in Balb/C background A
ermits efficient engraftment of human immune cells.herefore, AFC8-hu HSC/Hep mice provide the first hu-anized mouse model with both human immune and
iver cells.We inoculated AFC8-hu HSC/Hep mice with HCV
enotype 1a clinical isolates. We were able to detect HCVenomic RNA in the liver of AFC8-hu HSC/Hep miceFigure 2A and Table 2), but we did not detect significant
CV viremia in the blood of HCV-inoculated AFC8-huSC/Hep mice. This may be due to the relative low level
f human hepatocyte engraftment (�15%). To detectCV viremia in the blood, the adult hepatocyte-engraftedPA or Fah models support �50% engraftment of hu-an hepatocytes.17,19 –21 In the future, we will improve
the AFC8-hu HSC/Hep model by optimizing the deathinduction conditions with different doses and times ofdimer injections. It is also reported that the NOD-Scid-�C�/� (NSG) mouse is more permissive in accepting
uman cells.34 It is likely that the introduction of the
n. (A) Representative liver sections from AFC8/mock, AFC8-hu/mock,us Red/Fast Green (top panels) to visualize liver fibrosis. Liver sectionsoth muscle actin (�SMA, bottom panels) to visualize activated stellateA1 (COL1A1) (C) in the liver was measured using species-specific PCRouse glyceraldehyde-3-phosphate dehydrogenase, respectively. Data
ectioh Siri-smogen 1n or m
FC8 transgene into NSG mice will create a better host

akoHh
1
1
1
1
1
1
1
1
1
1
BA
SIC–L
IVER
,PA
NCREA
S,A
ND
BIL
IARY
TRA
CT
April 2011 HCV PATHOGENESIS IN A HUMANIZED MOUSE MODEL 1343
for engrafting human immune and liver cells. To getdifferent sources of human HSC and liver progenitorcells, the AFC8-hu mouse model will be useful to studyfunctional differentiation of human embryonic stem orinduced plural-potent cells to the human liver lineages.The construction of AFC8-hu HSC/Hep mouse withHSC and liver progenitor cells derived from human em-bryonic stem or induced pluripotent cells will be anexciting future direction.
HCV infection appeared to induce human immuneresponses in AFC8-hu HSC/Hep mice. We observed sig-nificant infiltration into the liver of HCV-infectedAFC8-hu HSC/Hep mice, including human T cells, mac-rophages, dendritic cells, and natural killer cells. Not onlydid we observe a preferential expansion of human T cellsfrom HCV-infected AFC8-hu HSC/Hep mice with HCV-derived peptides, but the expanded T cells also expressedIL2, TNF-�, and IFN-� in response to HCV peptide re-stimulation (Figure 4). Therefore, HCV infection inducedHCV-specific human T-cell responses in AFC8-hu HSC/Hep mice. It will be interesting to test whether depletionof human T cells (CD4 and/or CD8) will affect HCVinfection and pathogenesis in AFC-hu HSC/Hep mice. Itwill also be of interest to characterize and study humanT-cell subsets such as regulatory T cells and natural killerT cells in future experiments. Consistent with poor B-cellresponse reported in all humanized mouse models, wefailed to detect HCV-specific human antibodies in HCV-infected AFC8-hu HSC/Hep mice (data not shown).
In human patients, immune responses against HCVare implicated as mediators of liver diseases.35,36 Remark-
bly, HCV-infected AFC8-hu mice developed human leu-ocyte infiltration, hepatitis, and liver fibrosis through-ut the liver parenchyma with bridging septa (Figure 5).uman CD3� T cells and CD68� macrophages, as well asuman Alb� hepatocytes and �SMA� activated stellate
cells, were detected in and near the fibrotic region (Sup-plementary Figure 7), suggesting contribution of humanleukocytes to liver fibrosis. We also showed that HCVinfection led to increased level of activated human stel-late cells in fibrotic livers. Human extracellular matrixprotein Collagen 1A1 and inhibitor of matrix degrada-tion TIMP1 were induced in HCV-infected AFC8-huHSC/Hep mouse livers. It is of interest that the corre-sponding mouse fibrosis-associated genes were not in-duced by HCV infection. The lack of liver fibrosis inchronically infected chimpanzees also suggests a species-specific nature of HCV-induced liver fibrosis.37 TheAFC8-hu HSC/Hep mouse will provide an excellentmodel to elucidate the mechanisms of HCV-induced hu-man liver fibrosis.
Because human immunodeficiency virus (HIV) coinfec-tion occurs in nearly 25% of HCV patients and often leadsto accelerated end-stage liver disease,36,38,39 there is asignificant need for a model system to study HCV/HIV
coinfection. It will be of interest to test how HIV-1 infec-tion will affect HCV infection and pathogenesis in theAFC8-hu HSC/Hep mouse. In addition to HCV infectionand immunopathogenesis, AFC8-hu HSC/Hep mice willalso be useful to study other liver-tropic pathogens suchas hepatitis B virus and malaria.
Supplementary Material
Note: To access the supplementary materialaccompanying this article, visit the online version ofGastroenterology at www.gastrojournal.org, and at doi:
0.1053/j.gastro.2011.01.001.
References
1. Bruno S, Facciotto C. The natural course of HCV infection and theneed for treatment. Ann Hepatol 2008;7:114–119.
2. Crispe IN. Hepatic T cells and liver tolerance. Nat Rev Immunol2003;3:51–62.
3. Racanelli V, Rehermann B. The liver as an immunological organ.Hepatology 2006;43:S54–S62.
4. Bowen DG, Walker CM. Adaptive immune responses in acute andchronic hepatitis C virus infection. Nature 2005;436:946–952.
5. Dustin LB, Rice CM. Flying under the radar: the immunobiology ofhepatitis C. Annu Rev Immunol 2007;25:71–99.
6. Grakoui A, Shoukry NH, Woollard DJ, et al. HCV persistence andimmune evasion in the absence of memory T cell help. Science2003;302:659–662.
7. Rehermann B, Nascimbeni M. Immunology of hepatitis B virusand hepatitis C virus infection. Nat Rev Immunol 2005;5:215–229.
8. Cooper S, Erickson AL, Adams EJ, et al. Analysis of a successfulimmune response against hepatitis C virus. Immunity 1999;10:439–449.
9. Meyer-Olson D, Shoukry NH, Brady KW, et al. Limited T-cell re-ceptor diversity of HCV-specific T-cell responses is associatedwith CTL escape. J Exp Med 2004;200:307–319.
0. Farci P, Shimoda A, Coiana A, et al. The outcome of acutehepatitis C predicted by the evolution of the viral quasispecies.Science 2000;288:339–344.
1. Shoukry NH, Grakoui A, Houghton M, et al. Memory CD8� T cellsare required for protection from persistent hepatitis C virus in-fection. J Exp Med 2003;197:1645–1655.
2. Heeg MH, Ulsenheimer A, Gruner NH, et al. FOXP3(�) expressionin hepatitis C virus-specific CD4(�) T cells during acute hepatitisC. Gastroenterology 2009;137:1280–1288.
3. Sugimoto K, Ikeda F, Stadanlick J, et al. Suppression of HCV-specific T cells without differential hierarchy demonstrated exvivo in persistent HCV infection. Hepatology 2003;38:1437–1448.
4. Legrand N, Ploss A, Balling R, et al. Humanized mice for modelinghuman infectious disease: challenges, progress, and outlook.Cell Host Microbe 2009;6:5–9.
5. Bukh J. A critical role for the chimpanzee model in the study ofhepatitis C. Hepatology 2004;39:1469–1475.
6. Maeda N, Watanabe M, Okamoto S, et al. Hepatitis C virusinfection in human liver tissue engrafted in mice with an infec-tious molecular clone. Liver Int 2004;24:259–267.
7. Mercer DF, Schiller DE, Elliott JF, et al. Hepatitis C virus replica-tion in mice with chimeric human livers. Nat Med 2001;7:927–933.
8. Meuleman P, Libbrecht L, De Vos R, et al. Morphological andbiochemical characterization of a human liver in a uPA-SCID
mouse chimera. Hepatology 2005;41:847–856.
2
2
2
2
2
2
3BA
SIC–LIV
ER,
PA
NCREA
S,A
ND
BILIA
RY
TRA
CT
1344 WASHBURN ET AL GASTROENTEROLOGY Vol. 140, No. 4
19. Azuma H, Paulk N, Ranade A, et al. Robust expansion of humanhepatocytes in Fah-/-/Rag2-/-/Il2rg-/- mice. Nat Biotechnol2007;25:903–910.
0. Bissig KD, Le TT, Woods NB, et al. Repopulation of adult andneonatal mice with human hepatocytes: a chimeric animalmodel. Proc Natl Acad Sci U S A 2007;104:20507–20511.
1. Bissig KD, Wieland SF, Tran P, et al. Human liver chimeric miceprovide a model for hepatitis B and C virus infection and treat-ment. J Clin Invest 2010;120:924–930.
2. Traggiai E, Chicha L, Mazzucchelli L, et al. Development of ahuman adaptive immune system in cord blood cell-transplantedmice. Science 2004;304:104–107.
3. Pajvani UB, Trujillo ME, Combs TP, et al. Fat apoptosis throughtargeted activation of caspase 8: a new mouse model of induc-ible and reversible lipoatrophy. Nat Med 2005;11:797–803.
4. Conner EA, Lemmer ER, Omori M, et al. Dual functions of E2F-1in a transgenic mouse model of liver carcinogenesis. Oncogene2000;19:5054–5062.
5. Jiang Q, Zhang L, Wang R, et al. FoxP3�CD4� regulatory T cellsplay an important role in acute HIV-1 infection in humanizedRag2-/-�C-/- mice in vivo. Blood 2008;112:2858–2868.
26. Schmelzer E, Wauthier E, Reid LM. The phenotypes of pluripotenthuman hepatic progenitors. Stem Cells 2006;24:1852–1858.
27. Zhang L, Kovalev GI, Su L. HIV-1 infection and pathogenesis in anovel humanized mouse model. Blood 2007;109:2978–2981.
28. Lindenbach BD, Meuleman P, Ploss A, et al. Cell culture-grownhepatitis C virus is infectious in vivo and can be recultured invitro. Proc Natl Acad Sci U S A 2006;103:3805–3809.
29. Ahlenstiel G, Titerence RH, Koh C, et al. Natural killer cells arepolarized toward cytotoxicity in chronic hepatitis C in an interferon-alfa-dependent manner. Gastroenterology 2009;138:325–335.
30. Ulsenheimer A, Gerlach JT, Jung MC, et al. Plasmacytoid den-dritic cells in acute and chronic hepatitis C virus infection. Hepa-tology 2005;41:643–651.
31. Yoshizawa K, Abe H, Kubo Y, et al. Expansion ofCD4(�)CD25(�)FoxP3(�) regulatory T cells in hepatitis C virus-related chronic hepatitis, cirrhosis and hepatocellular carcinoma.Hepatol Res 2010;40:179–187.
32. Bataller R, Paik YH, Lindquist JN, et al. Hepatitis C virus core andnonstructural proteins induce fibrogenic effects in hepatic stel-late cells. Gastroenterology 2004;126:529–540.
33. Yoshiji H, Kuriyama S, Yoshii J, et al. Tissue inhibitor of metal-loproteinases-1 attenuates spontaneous liver fibrosis resolutionin the transgenic mouse. Hepatology 2002;36:850–860.
34. Brehm MA, Cuthbert A, Yang C, et al. Parameters for establishinghumanized mouse models to study human immunity: analysis ofhuman hematopoietic stem cell engraftment in 3 immunodefi-cient strains of mice bearing the IL2r�(null) mutation. Clin Immu-nol 2010;135:84–98.
5. Jin Y, Fuller L, Carreno M, et al. The immune reactivity role ofHCV-induced liver infiltrating lymphocytes in hepatocellular dam-
age. J Clin Immunol 1997;17:140–153.36. Kim AY, Chung RT. Coinfection with HIV-1 and HCV—a one-twopunch. Gastroenterology 2009;137:795–814.
37. Bigger CB, Guerra B, Brasky KM, et al. Intrahepatic gene expres-sion during chronic hepatitis C virus infection in chimpanzees.J Virol 2004;78:13779–13792.
38. Martinez-Sierra C, Arizcorreta A, Diaz F, et al. Progression ofchronic hepatitis C to liver fibrosis and cirrhosis in patientscoinfected with hepatitis C virus and human immunodeficiencyvirus. Clin Infect Dis 2003;36:491–498.
39. Sherman KE, Rouster SD, Chung RT, et al. Hepatitis C Virusprevalence among patients infected with human immunodefi-ciency virus: a cross-sectional analysis of the US adult AIDSClinical Trials Group. Clin Infect Dis 2002;34:831–837.
Received September 12, 2010. Accepted January 10, 2011.
Reprint requestsAddress requests for reprints to: Lishan Su, PhD, Lineberger
Comprehensive Cancer Center, University of North Carolina atChapel Hill, Chapel Hill, North Carolina 27599. e-mail:[email protected]; fax: (919) 966-8212.
AcknowledgmentsThe authors thank Drs F. Bai and A. Rogers for help with liver
pathology analysis; L. Chi, T.A. Curtis, and L. Chen for technicalsupport; Drs M. Fried and J. Darling for providing the patient HCVsera (UNC Division of Gastroenterology and Hepatology); Dr L Arnoldfor FACS support; and UNC CFAR for virology support.
Conflicts of interestThe authors disclose no conflicts.
Funding
Supported in part by a UNC UCRF innovation grant; grants fromNIH (AI076142, AA018009, AI077454 and AA018372 to L.S.), animmunology training grant (T32 AI007273 to M.L.W.) and UNCLineberger Comprehensive Cancer Center Postdoctoral TrainingGrant (M.T.B.); and a grant from LCRF (to J.A.F.); from the UNCUniversity Cancer Research Fund innovation grants (to L.S. andJ.A.F.); from Ministry of Science and Technology (2009CB522507,2006CB910901 & KSCX20YW-R-150 to L.Z.); from Ministry ofHealth (2009ZX10604 to L.Z. and 2008ZX10002-011 to L.Z., L.S.);from the Greenberg Medical Research Institute, the Ellison MedicalFoundation, the Starr Foundation, the Ronald A. Shellow MemorialFund, the Richard Salomon Family Foundation, and by a grant fromthe Foundation NIH through the Grand Challenges in Global Healthinitiative (to C.M.R. and A.P.); and a grant from the Center for
Clinical and Translational Research (RR024143 to A.P.).



















