
Differential cellular proliferation underlies heterochronic ...

Fax +41 61 306 12 34E-Mail [email protected]
Original Paper
Cells Tissues Organs DOI: 10.1159/000273266
A Comparative Study of Proliferation and Hepatic Differentiation of Human Adipose-Derived Stem Cells
R. Coradeghini a C. Guida b C. Scanarotti c R. Sanguineti c A.M. Bassi c A. Parodi d P.L. Santi e E. Raposio e
a Nanobiosciences Unit, JRC – European Commission Institute for Health and Consumer Protection, Ispra , b Tissue Engineering Laboratory, Plastic Surgery Division, DICMI, c Analysis and Research of Physiopathology Laboratory, DIMES, General Pathology Section, d Cardiovascular Biology Laboratory, DIMI, and e National Cancer Institute – IST, University of Genoa, Genoa , Italy
tained from liposuction, following two differentiation proto-cols. In the first procedure, medium was supplemented with epidermal growth factor (EGF), basic fibroblast growth fac-tor, hepatocyte growth factor (HGF) and nicotinamide; the second involved the addition of factors such as dexameta-sone, EGF, insulin-transferrin-sodium selenite, HGF, dimethyl sulfoxide and oncostatin. In parallel, we carried out our study in the Hep G2 cell line, as human hepatic differentiated in vitro model. Immunocytochemical analysis and RT-PCR were performed using hepatic markers to evaluate cell differen-
Key Words
Human adipose-derived stem cells � Hepatic differentiation � Proliferation tests � RT-PCR � CFSE staining
Abstract
Human adipose-derived stem cells possess a lot of stem cell characteristics, so they may be considered a source of stem cell population. On the basis of that, we have investigat - ed the hepatic potential of adipose-derived stem cells, ob-
Accepted after revision: September 21, 2009 Published online: December 30, 2009
Dr. Chiara Guida Tissue Engineering Laboratory, Plastic Surgery Division Department of Surgical Sciences (DICMI), University of Genoa Via De Toni 14, IT–16132 Genoa (Italy) Tel. +39 010 353 7311, Fax +39 010 353 7803, E-Mail chiara.guida @ unige.it
© 2009 S. Karger AG, Basel1422–6405/09/0000–0000$26.00/0
Accessible online at:www.karger.com/cto
Abbreviations used in this paper
ADSC adipose-derived stem cells HA human ADSC differentiated with medium AAFP �-fetoprotein HB human ADSC differentiated with medium BbFGF basic fibroblast growth factor HGF hepatocyte growth factorCFSE carboxyl fluorescein succinimidyl ester HNF-4-� hepatocyte nuclear factor-4-�DMEM Dulbecco’s modified Eagle medium MSC mesenchymal stem cellsDMSO dimethyl sulfoxide MTT 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetra-DPBS Dulbecco’s phosphate-buffered saline zolium bromideEDTA ethylenediaminetetracetic acid OSM oncostatin MEGF epidermal growth factor TTR transthyretinGAPDH glyceraldehyde-3-phosphate dehydrogenase

Coradeghini /Guida /Scanarotti /Sanguineti /Bassi /Parodi /Santi /Raposio
Cells Tissues Organs2
tiation. DNA content, MTT test and carboxyl fluorescein suc-cinimidyl ester staining were carried out to evaluate cell pro-liferation. We reported the evidence of basal hepatic marker in undifferentiated adipose-derived stem cells, which con-firmed their multipotency. A strong expression of albumin and � -fetoprotein was observed in hepatic-induced adi-pose-derived stem cells following both differentiation pro-cedures. Morphological aspects of the two types of hepatic adipose-derived stem cells were alike. Proliferation index suggested that the first differentiation procedure promoted better growth than the second. These preliminary findings suggest adipose-derived stem cells may be induced into he-patic lineage, and the most significant difference between the two standard differentiation procedures concerns prolif-eration rate. This aspect is to be considered when adipose-derived stem cells are employed in research and clinical studies. Copyright © 2009 S. Karger AG, Basel
Introduction
The use of stem cells is promising for cell-based ther-apy such as tissue regeneration. Although embryonic stem cells are theoretically highly beneficial, there are some potential limitations of cell regulations and ethical considerations. Therefore, it has been found that adult stem cells like pre-adipocytes or adipose-derived stem cells (ADSC) may be an alternative multipotent stem cell source [Zuk et al., 2002; Strem et al., 2005]. This kind of cells possess many characteristics of mesenchymal stem cells (MSC): low levels of cell senescence even after mul-tiple passages; analogous surface antigen marker profile of bone marrow-derived MSC, consisting in the expres-sion of CD146, CD106, CD105, CD90, CD73, CD54, CD49(d), CD49(e), CD44 [Gronthos et al., 2001; Katz et al., 2005; Wagner et al., 2005; Kern et al., 2006], and vi-mentin, collagen-1, fibronectin [Romanov et al., 2005], while they do not express the hematopoietic and endo-thelial markers, CD45 and CD31, STRO-1 antigen and the human leukocyte antigen, HLA-DR [Gronthos et al., 2001; De Ugarte et al., 2003; Schaffler and Buchler, 2007]; in vitro differentiation potentiality into adipogenic, myo-genic, osteogenic, chondrogenic and pancreatic cells [Zuk et al., 2001; Mizuno and Hyakusoku, 2003; Wagner et al., 2005; Timper et al., 2006; Raposio et al., 2007] as well as into neurogenic cells [Ashjian et al., 2003; Raposio et al., 2007], depending on the microenvironment in which they reside. These lineage-specific differentiations might offer future perspectives in tissue engineering and
reconstruction [Katz et al., 1999; Mizuno and Hyaku-soku, 2003]. ADSC may also be cultivated in an animal product-free environment, maintaining their ability to differentiate in vitro into adipo-, chondro- and osteogen-ic lineages and their typical surface marker expression [Perker et al., 2007]. ADSC are able to secrete angiogen-esis-related cytokines, like vascular endothelial growth factor, granulocyte/macrophage colony-stimulating fac-tor, stromal-derived factor-1 � and hepatocyte growth factor [Meliga et al., 2007]. Moreover, ADSC compared with other MSC have the longest culture survival and the highest proliferation ability [Kern et al., 2006].
It is known that MSC and haematopoietic stem cells have hepatic differentiation capabilities: MSC can differ-entiate into mature hepatocyte-like cells, CK18 seems to have some relationship with Thy-1.1 antigen, and CK18 may be a predictive marker of hepatic differentiation ca-pability of MSC [Shu et al., 2004]. Although the bone marrow is the first source identified to contain MSC, the adipose tissue represents an abundant and accessible source of adult MSC with low cell morbidity and minimal patient discomfort from conventional liposuction proce-dures. Current studies have shown that human and mu-rine stem cells from bone marrow and adipose tissue can effectively be considered as an another supply of multi-potent stem cells, to generate hepatocyte-like cells in vi-tro and in vivo [Seo et al., 2005; Taléns-Visconti et al., 2006; Aurich et al., 2007; Banas et al., 2007; Stock et al., 2008]. Recently, Yamamoto et al. [2008] have reported the gene expression profiles responsible for hepatic differen-tiation of adipose stem cells, using several microarray methods, and the clustering analysis reveals a striking similarity of gene clusters between adipose stem cells dif-ferentiated into hepatic lineage and the whole liver. Many liver diseases and disorders often cause partial/total or-gan failure. Hepatocyte transplantation is an alternative technique to whole liver transplantation for hepatic dis-ease cure, but there is a limited availability of human do-nor organs to isolate enough hepatocytes [Fox and Roy-Chowdhury, 2004]. It is known that the liver can precise-ly regulate its growth and mass, and hepatocytes have a great replication rate and a capability of repopulating the liver. However, a stem-like cell population proliferates when hepatocyte growth is blocked or delayed [Fausto, 2000] and the biliary cells, oval cells, seem to function as facultative stem cells, when hepatocyte regeneration is impaired [Alison and Sarraf, 1998; Zhang et al., 2003]. Moreover, the fetal liver is rich in precursor stem cells. However, little is known about the markers expressed by liver cells during fetal development and it is recognized

Hepatic Human Adipose-Derived Stem Cell Proliferation
Cells Tissues Organs 3
that human hepatic progenitor cells co-express the hema-topoietic stem cell markers CD117 and CD34 [Nava et al., 2005]. In the human fetal liver, high proliferative-poten-tial colony-forming cells are characterized as a compart-ment of primitive progenitors possibly including stem cells [Muench et al., 1994].
In this present study, we analysed the capability of hu-man adult ADSC, obtained from several different volun-tary donors, submitted to suction-assisted lipectomy (that is liposuction), to differentiate in vitro into hepatic lineage, by using two different differentiation proce-dures. For this aim, pre-adypocytes from the same donor were grown with two culture media, each of them was enriched with different growth factors. One medium (medium A) was supplemented with hepatocyte growth factor (HGF), a mesenchymally derived potent mitogen for parenchymal hepatocyte cells which acts via the HGF receptor c-MET, a transmembrane tyrosine kinase pro-tein [Schaffler and Buchler, 2007]. As is well known, stem cells derived from human liver and rat bone mar-row differentiate in mature hepatocytes when cultured in vitro in the presence of HGF and fibroblast growth factor 4, as indicated by the expression of functional cy-tochrome P450, albumin and urea production [Kang et al., 2005; Herrera et al., 2006]. In the second procedure, the me dium (medium B) was supplemented with on-costatin M (OSM), basic fibroblast growth factor (bFGF) and dimethyl sulfoxide (DMSO). In an in vitro study, it has been reported that exposure to OSM, a member of the interleukin-6 cytokine family, increased hepatocyte size, differentiation and formation of bile canaliculi [Lazaro et al., 2003]. bFGF signaling controls a lot of as-pects of embryonic patterning, including liver develop-ment, it is produced by the adjacent cardiac mesoderm and induces a hepatic fate and not the default pancreatic fate [Schwartz et al., 2002]. DMSO is known to maintain the functions of adult hepatocytes in vitro [Kamiya et al., 2002].
Based on these previous findings, the aim of our study was to identify which of the already established differen-tiation procedures could be useful in order to assess a suitable in vitro model for hepatic studies. In order to compare the effects between the two different differen-tiation procedures, we analysed several hepatic markers, such as albumin, � -fetoprotein (AFP), hepatocyte nucle-ar factor-4-alpha (HNF-4- � ) and transthyretin (TTR), and the proliferation index. Furthermore, we examined the same endpoints also in HepG2 cells, as fashionable in vitro model for hepatic differentiation and hepatotoxicity study.
Materials and Methods
Chemicals The chemical reagents, unless otherwise specified, were pur-
chased from Sigma Aldrich (St. Louis, Mo., USA). All cell culture products, unless otherwise specified, were purchased from Euro-clone, Life Sciences Division (Pero, Italy). Cell culture flasks and plates were obtained from Costar (Cambridge, Mass., USA). Rab-bit polyclonal anti-human albumin (A0001) was from Dako (Carpinteria, Calif., USA); rabbit polyclonal anti-human AFP (GTX72748) was from GeneTex Inc. (San Antonio, Tex., USA).
Cell Isolation and Processing Human adipose tissue was obtained from liposuction proce-
dures of 6 healthy female donors (40–50 years) under local anaes-thesia. Our Ethics Committee ensured that the procedure com-plied with national and European bioethical laws regarding the criteria of anonymity, informed consent and safety. As previously described by Zuk et al. [2001], lipoaspirates underwent extracel-lular matrix digestion with type I collagenase solution (2 mg/ml in Dulbecco’s phosphate-buffered saline, DPBS) for 1 h at 37 ° C with gentle shaking. Successively, the cellular suspension was cen-trifuged and the pellet, which contained adipocyte progenitor cells, was incubated with 160 m M ammonium chloride for 10 min to eliminate erythrocytes. The resulting ADSC were resuspended in culture control medium (Dulbecco’s modified Eagle medium, DMEM/Ham’s F12 (1: 1); 10% foetal bovine serum, FBS; gluta-mine 2 m M ; antibiotics) and filtered through a 100- � m nylon mesh to remove cellular debris. Then, the cells were plated in tis-sue sterile flasks in humidified atmosphere at 37 ° C and 5% CO 2 for 24 h (P0 cells). Another part of adipose-derived stem cell cul-tures were maintained in control medium for 3 weeks at subcon-fluent levels. At that moment, cells were harvested with tripsin 0.05% – sodium ethylenediaminetetracetic acid (EDTA) 0.02% solution, counted with a Neubauer chamber and trypan blue solu-tion and re-plated in the same conditions employed before at a density of 2 ! 10 4 cells/cm 2 . As procedure, the culture medium was replaced with fresh medium twice a week. Furthermore, cell cultures were observed daily through microscopy (Eclipse TS100; Nikon, Tokyo, Japan).
Hepatic Differentiation Procedures Hepatic induction was performed over a period of 5 weeks.
After reaching 80% of confluency, ADSC were submitted to the 2 different hepatic differentiation procedures, in humidified at-mosphere at 37 ° C and 5% CO 2 . The first differentiation proce-dure, according to that described by Talens-Visconti et al. [2007], involved the culture of ADSC for 48 h in DMEM serum free with 20 ng/ml epidermal growth factor (EGF) and 10 ng/ml bFGF (Invitrogen Life Technologies, Carlsbad, Calif., USA). A daily check of morphology by microscopy enabled to choose most similar ADSC cultures, derived from 3 different donors, for go-ing on the subsequent end points. For the following 4–5 weeks, the chosen cells were cultured with the differentiation medium A, consisting of DMEM, 10% FBS, 20 ng/ml HGF (PeproTech Inc, Rocky Hill, N.J., USA), 10 ng/ml bFGF and 4.9 m M nicotin-amide.
Following the other differentiation procedure, according to Seo et al. [2005], with minor modification, ADSC were incubated for 2 weeks in DMEM-LG (low glucose) with 5% FBS, 1 n M dexa-
Coradeghini /Guida /Scanarotti /Sanguineti /Bassi /Parodi /Santi /Raposio
Cells Tissues Organs4
methasone, 10 –4 M ascorbic acid 2-phosphate, 10 ng/ml EGF,5 � g/ml insulin, 5 � g/ml transferrin, 5 ng/ml sodium selenite. Hepatogenic differentiation was induced by culturing ADSC for 5 weeks in the same medium without serum, added with 0.1% DMSO, 10 ng/ml HGF and 10 ng/ml OSM (medium B). All culture media were changed twice a week and, during both differentia-tion procedures, all cultures were observed daily through micros-copy to estimate the changes of cell morphology.
HepG2 Cultures As positive control of hepatic differentiation the HepG2 hu-
man hepatoma cell line was employed, purchased from the GMP Cell Bank, Core Facility, National Cancer Institute (IST), Genoa, Italy. HepG2 were maintained in minimum essential Eagle me-dium supplemented with 10% FBS, glutamine 2 m M and antibiot-ics. When HepG2 arrived at confluent levels (80% confluency), cells were detached with tripsin 0.05% – sodium EDTA 0.02% so-lution and seeded at 2 ! 10 4 cells/cm 2 . The culture medium was changed twice a week.
Immunocytochemistry Analysis HepG2, P0 cells, undifferentiated ADSC and ADSC at the end
of differentiation (HA and HB) were seeded in Lab-Tek TM II Chamber Slide TM System (Nunc A/S, Roskilde, Denmark) at a density of 1.5 ! 10 4 cells/well. Cells were fixed at room tempera-ture with 4% paraformaldehyde for 15 min and permeabilized with 0.3% Triton X-100 for 10 min. The immunocytochemical procedure was carried out using EnVision Detection System-HRP/DAB (Dako). Briefly, slides were treated with peroxidase block reagent for 5 min at room temperature and incubated for 30 min with the following antibodies: rabbit polyclonal anti-human albumin (1: 500 dilution) and rabbit polyclonal anti-human AFP (1: 100 dilution). After washing, slides were incubated for 30 min with anti-rabbit-labelled polymers-HRP. Then, 3,3 � -diaminoben-zidine tetrahydrochloride (DAB chromogen) was added for rev-elation system [Van der Loos et al., 1989] and counterstaining
with haematoxylin was performed. Finally, samples were exam-ined under the imaging system microscope (model BX41; Olym-pus Italia Srl, Milan, Italy).
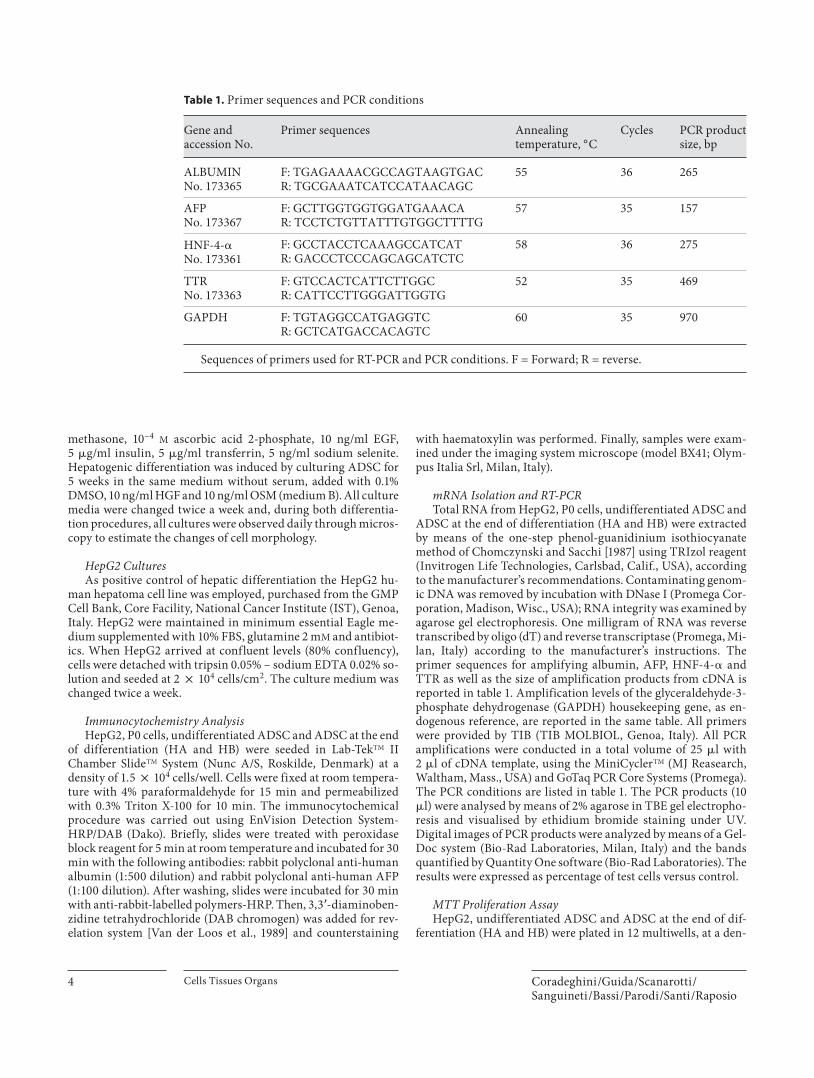
mRNA Isolation and RT-PCR Total RNA from HepG2, P0 cells, undifferentiated ADSC and
ADSC at the end of differentiation (HA and HB) were extracted by means of the one-step phenol-guanidinium isothiocyanate method of Chomczynski and Sacchi [1987] using TRIzol reagent (Invitrogen Life Technologies, Carlsbad, Calif., USA), according to the manufacturer’s recommendations. Contaminating genom-ic DNA was removed by incubation with DNase I (Promega Cor-poration, Madison, Wisc., USA); RNA integrity was examined by agarose gel electrophoresis. One milligram of RNA was reverse transcribed by oligo (dT) and reverse transcriptase (Promega, Mi-lan, Italy) according to the manufacturer’s instructions. The primer sequences for amplifying albumin, AFP, HNF-4- � and TTR as well as the size of amplification products from cDNA is reported in table 1 . Amplification levels of the glyceraldehyde-3-phosphate dehydrogenase (GAPDH) housekeeping gene, as en-dogenous reference, are reported in the same table. All primers were provided by TIB (TIB MOLBIOL, Genoa, Italy). All PCR amplifications were conducted in a total volume of 25 � l with2 � l of cDNA template, using the MiniCycler TM (MJ Reasearch, Waltham, Mass., USA) and GoTaq PCR Core Systems (Promega). The PCR conditions are listed in table 1 . The PCR products (10 � l) were analysed by means of 2% agarose in TBE gel electropho-resis and visualised by ethidium bromide staining under UV. Digital images of PCR products were analyzed by means of a Gel-Doc system (Bio-Rad Laboratories, Milan, Italy) and the bands quantified by Quantity One software (Bio-Rad Laboratories). The results were expressed as percentage of test cells versus control.
MTT Proliferation Assay HepG2, undifferentiated ADSC and ADSC at the end of dif-
ferentiation (HA and HB) were plated in 12 multiwells, at a den-
Table 1. Primer sequences and PCR conditions
Gene andaccession No.
Primer sequences Annealingtemperature, ° C
Cycles PCR productsize, bp
ALBUMINNo. 173365
F: TGAGAAAACGCCAGTAAGTGACR: TGCGAAATCATCCATAACAGC
55 36 265
AFPNo. 173367
F: GCTTGGTGGTGGATGAAACAR: TCCTCTGTTATTTGTGGCTTTTG
57 35 157
HNF-4-�No. 173361
F: GCCTACCTCAAAGCCATCATR: GACCCTCCCAGCAGCATCTC
58 36 275
TTRNo. 173363
F: GTCCACTCATTCTTGGCR: CATTCCTTGGGATTGGTG
52 35 469
GAPDH F: TGTAGGCCATGAGGTCR: GCTCATGACCACAGTC
60 35 970
Sequences of primers used for RT-PCR and PCR conditions. F = Forward; R = reverse.

Hepatic Human Adipose-Derived Stem Cell Proliferation
Cells Tissues Organs 5
sity of 5,000 per well, in the respective culture media. Cell prolif-eration was evaluated in all cell types by 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) test, at 1, 3, 7 and 10 days, as described by Mosmann [1983]. Growth media were changed every 2 days. Briefly, HepG2, control and differentiated ADSC were incubated for 3 h at 37 ° C with 0.5 mg/ml MTT in se-rum-free medium. Mitochondria of viable cells reduce MTT to formazane, which accumulates as blue crystals; these were dis-solved by adding 1 N hydrogen chloride-isopropanol (1: 24 v/v) solution. After 10-min incubation at room temperature, the ab-sorbance of the product was measured by a spectrophotometer at 570 nm.
DNA Assay HepG2, undifferentiated ADSC and ADSC at the end of dif-
ferentiation (HA and HB) were plated in 12 multiwells, at a den-sity of 5,000 per well, in the respective culture media. Cell prolif-eration index was evaluated in all cell types in terms of cellular DNA content, by the method of Rao and Otto [1992]. So, at pre-determined time points (1, 3, 7 and 10 days), growth media were removed and, after washing with warm DPBS, 1 ml of lysis solu-tion (urea 10 M , 0.01% sodium dodecyl sulfate in saline sodium citrate buffer, 0.154 M NaCl, 0.015 M sodium citrate, pH 7) was added to each well. Cell suspensions were incubated at 37 ° C in a shaking bath for 2 h; successively, 1 ml of Hoechst 33,258 dye(1 � g/ � l in saline sodium citrate buffer) was added in the dark. Absorbance was measured by a Perkin Elmer LS5 spectrofluorim-eter (Norwalk, Conn., USA) at excitation and emission wave-lengths of 355 and 460 nm, respectively. Cell proliferation was estimated by referring fluorescence units to a linear standard curve for DNA fluorescence versus cell number.
Carboxyl Fluorescein Succinimidyl Ester Staining Carboxyl fluorescein succinimidyl ester (CFSE) staining was
similarly performed as previously reported by Raposio et al. [2008]. Briefly, 1 day before seeding, HepG2 cells, undifferenti-
ated ADSC, HA and HB at the end of differentiation were washed with DPBS and incubated for 10 min in the dark at 37 ° C and 5% CO 2 with 5 � M CFSE solution in DPBS; successively, the cells were washed with DPBS added with 10% FBS [Urbani et al., 2006]. All types of cells were then seeded in sterile flasks at a density of 4,000 cells/cm 2 in the respective culture media, and at different times after CFSE staining (1, 3, 7 and 10 days), they were harvested, cen-trifuged and washed with DPBS. The samples were acquired us-ing a FACSCalibur flow cytometer and analysis of data was per-formed by Cell Quest (Becton Dickinson, Franklin Lakes, N.J., USA) and ModFit (Mac3.1 SP2; Verity Software House Inc., Top-sham, Me., USA) software.
Results
To determine optimal conditions for hepatic adipose-derived stem cell differentiation, we tested the effectof different cytokines/growth factors on morphologic changes of ADSC growing in the two hepatogenic differ-ent media. Cell morphology did not show significant changes at the initial step of differentiation; during HGF exposure, we observed that both differentiation proce-dures had induced the same changes in cellular mor-phology. Indeed, the cells began to lose the fibroblastic morphology in comparison with undifferentiated ADSC. HA and HB showed very similar remarkable change in morphology, taking up a round and polygonal shape ( fig. 1 a, b). By immunocytochemistry, the expression of albumin and AFP was analysed in P0 cells and HA and HB. In parallel, the same analysis was performed inundifferentiated ADSC and HepG2 cells. HA, HB and
a b
Fig. 1. ADSC after 2 weeks of culture with hepatogenic medium A ( a ) and with hepatogenic medium B ( b ). Scale bars = 50 � m.

Coradeghini /Guida /Scanarotti /Sanguineti /Bassi /Parodi /Santi /Raposio
Cells Tissues Organs6
a
b
c
Fig. 2. Immunostaining with albumin antibody: a in ADSC dif-ferentiated with medium A (scale bar = 10 � m); b in ADSC dif-ferentiated with medium B; c in HepG2 cells. Scale bars = 20 � m.
a
b
c
Fig. 3. Immunostaining with AFP antibody: a in ADSC differen-tiated with medium A; b in ADSC differentiated with medium B; c in HepG2 cells. Scale bars = 20 � m.

Hepatic Human Adipose-Derived Stem Cell Proliferation
Cells Tissues Organs 7
AFP
Albumin
HNF
TTR
GAPDHe
0
*
HepG2
+
C
+
HA
+
HBP0
600
400
200
800
1,000
1,200
1,400
% v
s. P
0
a
Fig. 4. Percentages of levels of albumin ( a ), AFP ( b ), HNF-4- � ( c ) and TTR ( d ) genes in HepG2, undifferentiated ADSC (C), ADSC differentiated with medium A (HA cells) and ADSC differenti-ated with medium B (HB cells) versus those in P0 cells, as control.
e Representative agarose gels. First line: P0 cells; second line: HepG2 cells; third line: undifferentiated ADSC; fourth line: HA cells; fifth line: HB cells. * p ! 0.01 and 0.001 vs. P0; + p ! 0.01 vs. HepG2 (ANOVA followed by Bonferroni’s test).
0
*
HepG2
+
C
+, *
HA
*
HBP0
500
1,000
1,500
2,000
2,500
% v
s. P
0
b
0
*
HepG2
+
C
+
HA
+
HBP0
600
400
200
800
1,000
1,200
% v
s. P
0
c
0
+
HepG2
+
C
+, *
HA
+, *
HBP0
25,0002,000
1,000
55,000
85,000
1,210,000
1,310,000
1,410,000
1,510,000
% v
s. P
0
d

Coradeghini /Guida /Scanarotti /Sanguineti /Bassi /Parodi /Santi /Raposio
Cells Tissues Organs8
HepG2 cells stain positive for both markers ( fig. 2 , 3 ); on the other hand, albumin and AFP were weakly expressed in P0 cells and in undifferentiated ADSC (data not shown).
In all cellular models mRNA levels of 4 hepatocyte-specific genes, such as albumin, AFP, HNF-4- � and TTR, were determined; only for HA and HB cells the analysis was performed after 5 weeks of differentiation ( table 2 ). In P0 cells the albumin gene levels resulted very low ver-sus HepG2, only in HA and HB mRNA levels they reached values 2-fold higher versus control and P0. In HB cells AFP mRNA expression showed a 2.5-fold increase com-pared to HA; undifferentiated ADSC expressed AFP gene at very low level versus HepG2 cells. HNF-4- � gene did not appear to be different in HA and HB. Finally, TTR gene was not detectable in P0 and undifferentiated ADSC, while in HA it reached higher level than in HB (2.5-fold). Figure 4 shows the percentages of levels of each gene in HepG2, undifferentiated ADSC, HA and HB cells versus those in P0 cells, as control.
During 5 weeks of hepatic differentiation, the duplica-tion time of HA cells was 3-fold bigger than that of HB cells, being the density of both cell types at the start dif-ferentiation point of 2 ! 10 4 cells/cm 2 . The proliferation index, extrapolated by MTT test ( fig. 5 a), indicated that, from 3 to 10 days after seeding, the proliferation rate of HA was more strongly enhanced than the other cells. These results were confirmed by DNA content, measured by DNA assay ( fig. 5 b). Moreover, cytofluorimetric pro-liferation analysis was assessed at different time points on undifferentiated ADSC, HepG2, HA and HB cells ( fig. 6 ). At the first day after CFSE staining, all samples displayed
only parental peak (black area), but at the third day un-differentiated ADSC showed virtually 3 generations, HB and HepG2 3 generations and HA 5 generations (greyscale rate). After 7 and 10 days, undifferentiated ADSC prolif-eration rate was less relevant than that of the other cell types; HA cells showed the highest levels, while HepG2 and HB growth index were quite similar.
Discussion
Human ADSC possess characteristics of multipotent adult stem cells since they could differentiate into cells of mesenchymal origin, as adipocytes, osteocytes, chondro-cytes, myocytes and endothelial, neurogenic and pancre-atic cells [Zuk et al., 2001; Ashjian et al., 2003; Mizuno and Hyakusoku, 2003; Wagner et al., 2005; Timper et al., 2006; Raposio et al., 2007]. As shown by literature data, MSC from a predominant source, such as bone marrow, reveal a lower proliferation capacity and frequency than ADSC [Kern et al., 2006]; moreover, from 200 ml of li-poaspirate, it is possible to obtain approximately 40-fold more stem cells than from 40 ml of bone marrow [Strem et al., 2005]. Recently, the hepatogenic potential of rat and human bone marrow stromal cells [Shu et al., 2004; Kang et al., 2005; Taléns-Visconti et al., 2006] as well as ADSC has been investigated [Seo et al., 2005; Taléns-Visconti et al., 2006; Banas et al., 2007, 2009]. Concerning the dona-tion procedure, the isolation ratio and the minimal rejec-tion in donor health, it is conceivable that also ADSC could represent an attractive tool for studies on stem cell therapy for the liver.
In our study, we utilised lipoaspirates from 6 female healthy donors (40–50 years), and after adipose tissue processing an initial observation of cell morphology was performed to assure that samples from different do-nors were quite similar and to exclude those samples were too variable from others. So, all data reported in the present study refer to 3 donors. HA and HB cells have showed very similar morphological changes with both differentiation procedures: in fact, early fibroblast-like cells develop a polygonal morphology. These results are in accordance with those reported by Seo et al. [2005] and Taléns-Visconti et al. [2006]. The analysis of pheno-typic features, gene expression changes and prolifera-tion capacity of HA and HB cells was performed in or-der to verify the better differentiation culture condi-tions. The hepatogenic potentiality of ADSC was confirmed by im munocytochemistry with detection of hepatic-specific markers, such as albumin and AFP; HA
Table 2. Percentages of gene expression
Gene Cell typesHepG2 (control)
C P0 HA HB
Albumin 100 11.1681.22 8.7180.95 20.2982.23 15.8581.74AFP 100 14.7981.60 4.8280.53 32.2883.50 82.4989.00HNF-4-� 100 1.0580.11 1.680.17 5.7680.63 6.5580.72TTR 100 0 0 3.2480.35 1.2580.13
Gene expression of albumin, AFP, HNF-4-� and TTR in un-differentiated ADSC (C), P0 cells, HA and HB at 5 weeks of dif-ferentiation in comparison with positive control cells (HepG2 cells). Values are expressed as percentage of the positive control cells and represent the mean 8 SEM of 3 separate experiments in duplicate.

Hepatic Human Adipose-Derived Stem Cell Proliferation
Cells Tissues Organs 9
Fig. 6. CFSE staining in HA, HB, undiffer-entiated ADSC (C) and HepG2 cells, at dif-ferent times (1, 3, 7 and 10 days). From black to white, colour grading are from old parental to last new cell populations.
0
0.8
0.7
0.6
0.5
0.4
0.3
0.2
0.1
AU (a
bsor
banc
e)
7
**
**
0 1
****3
****
10
**
**
Time (days)a b
0
300
250
200
150
100
50
Cell
num
ber (
×10
3 )
7
*
*
0 1
* *3
* *
10
*
*
Time (days)
Fig. 5. Cell proliferation index, evaluated by MTT test and DNA assay. Undifferentiated ADSC (C), HA and HB cells are cultured for 1, 3, 7 and 10 days. a MTT index is expressed as arbitrary unit (AU), related to the OD at 570 nm. b DNA content index is esti-mated by referring fluorescence units to a linear standard curve
for DNA fluorescence versus cell number. Values represent the mean 8 SEM of 3 different experiments in quadruplicate. * * p ! 0.05; * p ! 0.01 vs. undifferentiated ADSC (ANOVA by Dunnett’s test). i = C; k = HA; g = HB.
Days
0 50 100
150
200
250
1HA
0 50 100
150
200
250
3
0 50 100
150
200
250
5
0 50 100
150
200
250
10
0 50 100
150
200
250
HB
0 50 100
150
200
250
0 50 100
150
200
250
0 50 100
150
200
250
0 50 100
150
200
250
C
0 50 100
150
200
250
0 50 100
150
200
250
0 50 100
150
200
250
0 50 100
150
200
250
HepG2
0 50 100
150
200
250
0 50 100
150
200
250
0 50 100
150
200
250
CFSE

Coradeghini /Guida /Scanarotti /Sanguineti /Bassi /Parodi /Santi /Raposio
Cells Tissues Organs10
References Abe, K, H. Niwa, K. Iwase, M. Takiguchi, M. Mori, S.I. Abè, K. Abe, K.I. Yamamure (1996) Endoderm-specific gene expression in em-bryonic stem cells differentiated to embryoid bodies. Exp Cell Res 229: 27–34.
Alison, M., C. Sarraf (1998) Hepatic stem cells. J Hepatol 29: 676–682.
Ashjian, P.H., A.S. Elbarbary, B. Edmonds, D. DeUgarte, M. Zhu, P.A. Zuk, H.P. Lorenz, P. Benhaim, M.H. Hedrick (2003) In vitro dif-ferentiation of human processed lipoaspirate cells into early neural progenitors. Plast Re-constr Surg 111: 1922–1931.
and HB cells, from all selected samples, showed similar expression pattern surface protein markers. Shiraki et al. [2008] have suggested that stem cell expression of both AFP and albumin indicate a hepatoblast pheno-type, while the lack of only AFP is associated with ma-ture differentiated phenotype (hepatocytes). Our results indicate that HA and HB express hepatoblast pheno-type, while P0 cells represent an early immature differ-entiation step, since these cells are AFP positive and al-bumin negative.
Analysis of gene expression of albumin, AFP, HNF-4- � and TTR, detected by RT-PCR, reveals that undif-ferentiated, P0, HA and HB cells express very low levels of mRNA of those genes, in comparison with well-dif-ferentiated HepG2, even if HA and HB cells show high-er level of expression of the 4 genes than other ADSC-derivative cells. It is relevant that in our study, AFP gene expression appears to be enhanced in HB cells, attesting that these cells are less differentiated than HA, and still maintains multipotential characteristics. As well known, AFP gene expression precedes albumin gene expression in embryonic stem cells during the development of pat-terns of endoderm-specific gene expression [Abe et al., 1996]; moreover, AFP in the liver is a marker of imma-ture (that is, fetal) liver cells or oval cells in adult liver [Brill et al., 1993]. HA and HB showed significantly higher expression of the other two hepatic markers, HNF-4 and TTR, in comparison with undifferentiated P0 cells. HNF-4- � is a very important marker to estab-lish hepatic lineage; it is responsible for the final com-mitment of oval cells to differentiate into hepatocytes, which regenerate the liver parenchyma, it is essential for generation of hepatic epithelium and it is considered the dominant regulator of the epithelial phenotype because its ectopic expression in fibroblasts induces a mesenchy-mal-to-epithelial transition [Nagy et al., 1994]. TTR, a 55-kDa homotetramer, is synthesized in the liver, cho-roid plexus and retinal pigment epithelium and it is a carrier of the thyroid hormone thyroxin. Recently, it has been reported by Banas et al. [2007] that TTR is ex-pressed in ADSC CD105+ differentiated into hepatic
lineage. In our previous study [Raposio et al., 2008], we verified that MTT test, DNA assay and CFSE staining were suitable analyses to evaluate cell proliferation in undifferentiated ADSC, in accordance with other data in the literature [Hemmrich et al., 2006; Liu et al., 2008; Zaminv et al., 2008]. So, we apply the above analyses to evaluate the proliferation rate of HA and HB cells. CFSE possesses the ability to covalently label long-lived intra-cellular molecules with the fluorescent dye, carboxyflu-orescein. Following each cell division, the equal distri-bution of this fluorescent molecule to progeny cells re-sults in halving the fluorescence in each daughter cell [Quah et al., 2007]. Duplication time values extrapolat-ed by cell counting chamber, and trypan blue staining indicated that HA have a 3-fold higher proliferation in-dex than HB cells (data not shown). According to these data, MTT and DNA assay also evidenced a marked proliferation enhancement of HA from 3 to 10 days after seeding. Moreover, 3 days after CFSE staining, HA and HB cells show virtually 5 and 4 generations, respective-ly, and the parental peaks seem to disappear at the same moment in both differentiating cells.
In conclusion, on the basis of the phenotypic charac-teristics and gene patterns of HA and HB, our present results suggest that, under in vitro inducing conditions, ADSC can differentiate toward a hepatic phenotype through two unlike growth media. Our experience high-lights that ADSC differentiated with medium A, with fewer growth factors, show a higher proliferation index, therefore this hepatic differentiation procedure could be considered ideal to achieve a lot of hepatogenic cells for experimental researches and cell hepatic therapy.
Acknowledgments
We would like to thank Prof. R. Fiocca for kind hospitalityin the Laboratory of Pathological Anatomy at the University of Genoa, Italy, as well as Prof. C. Falugi, Dr. M.G. Aluigi, Dr. A. Kunkl, Dr. M.F. Catalano and Dr. M. Curto for their precious help.

Hepatic Human Adipose-Derived Stem Cell Proliferation
Cells Tissues Organs 11
Aurich, I., L.P. Mueller, H. Aurich, J. Luetzken-dorf, K. Tisljar, M.M. Dollinger, W. Schor-mann, J. Walldorf, J.G. Hengstler, W.E. Fleig, B. Christ (2007) Functional integration of he-patocytes derived from human mesenchymal stem cells into mouse livers. Gut 56: 405–415.
Banas, A., T. Teratani, Y. Yamamoto, M. Toku-hara, F. Takeshita, G. Quinn, H. Okochi, T. Ochiya (2007) Adipose tissue-derived mes-enchymal stem cells as a source of human he-patocytes. Hepatology 46: 219–228.
Banas, A., T. Teratani, Y. Yamamoto, M. Toku-hara, F. Takeshita, M. Osaki, T. Kato, H. Okochi, T. Ochiya (2009) Rapid hepatic fate specification of adipose-derived stem cells and their therapeutic potential for liver fail-ure. J Gastroenterol Hepatol 24: 70–77.
Brill, S., P. Holst, S. Sigal, I. Zvibel, A. Fiorino, A. Ochs, U. Somasundaran, L.M. Reid (1993) Hepatic progenitor populations in embryon-ic, neonatal, and adult liver. Proc Soc Exp Biol Med 204: 261–269.
Chomczynski, P., N. Sacchi (1987) Single-step method of RNA isolation by acid guanidini-um thiocyanate-phenol-chloroform extrac-tion. Anal Biochem 162: 156–159.
De Ugarte, D.A., K. Morizono, A. Elbarbary, Z. Alfonso, P.A. Zuk, M. Zhu, J.L. Dragoo, P. Ashjian, B. Thomas, P. Benhaim, I. Chen, J. Fraser, M.H. Hedrick (2003) Comparison of multi-lineage cells from human adipose tis-sue and bone marrow. Cells Tissues Organs 174: 101–109.
Fausto, N. (2000) Liver regeneration. J Hepatol 32: 19–31.
Fox, I.J., J. Roy-Chowdhury (2004) Hepatocyte transplantation. J Hepatol 40: 878–886.
Gronthos, S., D.M. Franklin, H.A. Leddy, P.G. Robey, R.W. Storms, J.M. Gimble (2001) Sur-face protein characterization of human ad-ipose tissue-derived stromal cells. J Cell Physiol 189: 54–63.
Hemmrich, K., M. Meersch, D. von Heimburg, N. Pallua (2006) Applicability of the dyes CFSE, CM-DiI and PKH26 for tracking of human preadipocytes to evaluate adipose tissue engineering. Cells Tissues Organs 184: 117–127.
Herrera, M.B., S. Bruno, S. Buttiglieri, C. Tetta, S. Gatti, M.C. Deregibus, B. Bussolati, G. Ca-mussi (2006) Isolation and characterization of a stem cell population from adult human liver. Stem Cells 24: 2840–2850.
Kamiya, A., N. Kojima, T. Kinoshita, Y. Sakai, A. Miyaijma (2002) Maturation of fetal hepato-cytes in vitro by extracellular matrices and oncostatin M: induction of tryptophan oxy-genase. Hepatology 35: 1351–1359.
Kang, X.Q., W.J. Zang, T.S. Song, X.L. Xu, X.J. Yu, D.L. Li, K.W. Meng, S.L. Wu, Z.Y. Zhao (2005) Rat bone marrow mesenchymal stem cells differentiate into hepatocytes in vitro. World J Gastroenterol 11: 3479–3484.
Katz, A.J., R. Llull, M.H. Hedrick, J.W. Furtrell (1999) Emerging approaches to the tissue en-gineering of fat. Clin Plast Surg 26: 587–603.
Katz, A.J., S.S. Tholpady, H. Shang, R.C. Ogle (2005) Cell surface and transcriptional char-acterization of human adipose-derived ad-herent stromal (hADAS) cells. Stem Cells 23: 412–423.
Kern, S., H. Eichler, J. Stoeve, H. Kluter, K. Bie-back (2006) Comparative analysis of mesen-chymal stem cells from bone marrow, um-bilical cord blood or adipose tissue. Stem Cells 24: 1294–1301.
Lazaro C.A., E.J. Croager, C. Mitchell, J.S. Camp-bell, C. Yu, J. Foraker, J.A. Rhim, G.C. Yeoh, N. Fausto (2003) Establishment, character-ization, and long-term maintenance of cul-tures of human fetal hepatocytes. Hepatolo-gy 38: 1095–1106.
Liu, Q., L. Cen, S. Yin, L. Chen, G. Liu, J. Chang, L. Cui (2008) A comparative study of prolif-eration and osteogenic differentiation of ad-ipose-derived stem cells on akermanite and beta-TCP ceramics. Biomaterials 29: 4792–4799.
Meliga, E., B.M. Strem, H.J. Duckers, P.W. Ser-ruys (2007) Adipose-derived cells. Cell. Transplant 16: 963–970.
Mizuno, H., H. Hyakusoku (2003) Mesengenic potential and future clinical perspective of human processed lipoaspirate cells. J Nip-pon Med Sch 70: 300–306.
Mosmann, T. (1983) Rapid colorimetric assay for cellular growth and survival: application to proliferation and cytotoxicity assays. J Im-munol Methods 65: 55–63.
Muench, M.O., J. Cupp, J. Polakoff, M.G. Roncarolo (1994) Expression of CD33, CD38 and HLD-DR on CD34+ human fetal liver progenitors with a high proliferative poten-tial. Blood 83: 3170–3181.
Nagy, P., H.C. Bisgaard, S.S. Thorgeirsson (1994) Expression of hepatic transcription factors during liver development and oval cell dif-ferentiation. J Cell Biol 126: 223–233.
Nava, S., M. Westgren, M. Jaksch, A. Tibell, U. Broome, B.G. Ericzon, S. Sumitran-Holgers-son (2005) Characterization of cells in the developing human liver. Differentiation 73: 249–260.
Perker, A.M., H. Shang, M. Khurgel, A.J. Katz (2007) Low serum and serum-free culture of multipotential human adipose stem cells. Cytotherapy 9: 637–646.
Quah, B.J., H.S. Warren, C.R. Parish (2007) Monitoring lymphocyte proliferation in vi-tro and in vivo with the intracellular f luores-cent dye carboxyfluorescein diacetate. Nat Protoc 2: 2049–2056.
Rao, J., W.R. Otto (1992) Fluorimetric DNA as-say for cell growth estimation. Anal Biochem 207: 186–192.
Raposio, E., C. Guida, I. Baldelli, F. Benvenuto, M. Curto, L. Paleari, F. Filippi, R. Fiocca, G. Robello, P.L. Santi (2007) Characterization and induction of human pre-adipocytes. Toxicol in vitro 21: 330–334.
Raposio, E., C. Guida, R. Coradeghini, C. Sca-narotti, A. Parodi, I. Baldelli, R. Fiocca, P.L. Santi (2008) In vitro polydeoxyribonucleo-tide affects on human pre-adipocytes. Cell Prolif 41: 739–754.
Romanov, Y.A., A.N. Darevskaya, N.V. Merzli-kina, L.B. Buravkova (2005) Mesenchymal stem cells from human bone marrow and ad-ipose tissue: isolation, characterization and differentiation potentialities. Bull Exp Biol Med 140: 138–143.
Schaffler, A., C. Buchler (2007) Concise review: adipose tissue-derived stromal cells – basic and clinical implications for novel cell-based therapies. Stem Cells 25: 818–827.
Schwartz, R.E., M. Reyes, L. Koodie, Y. Jiang, M. Blackstad, T. Lund, T. Lenvik, S. Johnson, W.S. Hu, C.M. Verfaillie (2002) Multipotent adult progenitor cells from bone marrow dif-ferentiate into functional hepatocyte-like cells. J Clin Invest 109: 1291–1302.
Seo, M.J., S.Y. Suh, Y.C. Bae, J.S. Junga (2005) Differentiation of human adipose stromal cells into hepatic lineage in vitro and in vivo. Biochem Biophys Res Commun 328: 258–264.
Shiraki, N., K. Umeda, N. Sakashita, M. Takeya, K. Kume, S. Kume (2008) Differentiation of mouse and human embryonic stem cells into hepatic lineages. Genes Cells 13: 731–746.
Shu, S.N., L. Wei, J.H. Wang, Y.T. Zhan, H.S. Chen, Y. Wang (2004) Hepatic differentia-tion capability of rat bone marrow-derived mesenchymal stem cells and hematopoietic stem cells. World J Gastroent 10: 2818–2822.
Stock, P., M.S. Staege, L.P. Muller, M. Sgodda, A. Volker, I. Volkmer, J. Lutzkendorf, B. Christ (2008) Hepatocytes derived from adult stem cells. Transplant Proc 40: 620–623.
Strem, B.M., K.C. Hicok, M. Zhu, I. Wulur, Z. Alfonso, R.E. Schreiber, J.K. Fraser, M.H. Hedrick (2005) Multipotential differentia-tion of adipose tissue-derived stem cells. Keio J Med 54: 132–141.
Taléns-Visconti, R., A. Bonora, R. Jover, V. Mi-rabet, F. Carbonell, J.V. Castell, M.J. Gómez-Lechón (2006) Hepatogenic differentiation of human mesenchymal stem cells from adi-pose tissue in comparison with bone marrow mesenchymal stem cells. World J Gastroen-terol 12: 5834–5845.
Taléns-Visconti, R., A. Bonora, R. Jover, V. Mi-rabet, F. Carbonell, J.V. Castell, M.J. Gómez-Lechón (2007) Human mesenchymal stem cells from adipose tissue: differentiation into hepatic lineage. Toxicol in vitro 21: 324–329.
Timper, K., D. Seboek, M. Eberhardt, P. Lin-scheid, M. Christ-Crain, U. Keller, B. Muller, H. Zulewski (2006) Human adipose tissue-derived mesenchymal stem cells differenti-ate into insulin, somatostatin, and glucagon expressing cells. Biochem Biophys Res Com-mun 341: 1135–1140.

Coradeghini /Guida /Scanarotti /Sanguineti /Bassi /Parodi /Santi /Raposio
Cells Tissues Organs12
Urbani, S., R. Caporale, L. Lombardini, A. Bosi, R. Saccardi (2006) Use of CFDA-SE for eval-uating the in vitro proliferation pattern of human mesenchymal stem cells. Cytothera-py 8: 243–253.
Van der Loos, C.M., P.K. Das, J.J. Van den Oord, H.J. Houthoff (1989) Multiple immunoen-zyme staining techniques. Use of f luores-ceinated, biotinylated and unlabelled mono-clonal antibodies. J Immunol Methods 117: 45–52.
Yamamoto, Y., A. Banas, S. Murata, M. Ishikawa, C.R. Lim, T. Teratani, I. Hatada, K. Matsu-bara, T. Kato, T. Ochiya (2008) A compara-tive analysis of the transcriptome and signal pathways in hepatic differentiation of hu-man adipose mesenchymal stem cells. FEBS J 275: 1260–1273.
Wagner, W., F. Wein, A. Seckinger, M. Frank-hauser, U. Wirkner, U. Krause, J. Blake, C. Schwager, V. Eckstein, W. Ansorge, A.D. Ho (2005) Comparative characteristics of mes-enchymal stem cells from human bone mar-row, adipose tissue, and umbilical cord blood. Exp Hematol 33: 1402–1416.
Zaminv, A., I. Ragerdi Kashani, M. Barbarestani, A. Hedayatpuor, R. Mahmoudi, A. Farzaneh Nejad (2008) Osteogenic differentiation of rat mesenchymal stem cells from adipose tis-sue in comparison with bone marrow mes-enchymal stem cells: melatonin as a differen-tiation factor. Iran Biomed J 12: 133–141.
Zhang, Y., X.F. Bai, C.X. Huang (2003) Hepatic stem cells: existence and origin. World J Gas-troenterol 9: 201–204.
Zuk, P.A., M. Zhu, H. Mizuno, J. Huang, J.W. Futrell, A.J. Katz, P. Benhaim, H.P. Lorenz, M.H. Hedrick (2001) Multilineage cells from human adipose tissue: implication for cell-based therapies. Tissue Eng 7: 211–228.
Zuk, P.A., M. Zhu, P. Ashjian, D.A. De Ugarte, J. Huang, H. Mizuno, Z.C. Alfonso, J.K. Fraser, P. Benhaim, M.H. Hedrick (2002) Human adipose tissue is a source of multipotent stem cells. Mol Biol Cell 13: 4279–4295.
Copyright © 2022 FDOKUMEN




















