
A 38-kilodalton low-CO 2 -inducible polypeptide is associated with the pyrenoid in Chlorella...
10
Abstract. In the green alga Chlorella vulgaris UAM 101, a CO 2 -concentrating mechanism (CCM) is induced when cells are transferred from high (5%) to low (0.03%) CO 2 concentrations. The induction of the CCM is correlated with de-novo synthesis of several polypeptides that remain to be identified. The internal carbonic anhydrase (CA; EC 4.2.1.1) activity increased 6- to 7-fold within 6 h of acclimation to air. When crude homogenates were further separated into soluble and insoluble fractions, nearly all of the CA activity was associated with the membrane fraction. Immunoblot analysis of cell homogenates probed with antibodies raised against the 37-kDa subunit of periplasmic CA of Chlamydomonas reinhardtii showed a cross-reaction with a single 38-kDa polypeptide in both high- and low-CO 2 - grown cells. The up-regulation of the expression of the 38-kDa polypeptide was closely correlated with the increase in internal CA activity. Furthermore, its sub- cellular location was also correlated with the distribu- tion of the activity. Immunoblot analysis of pyrenoid fractions showed that the 38-kDa polypeptide was concentrated in the pyrenoids from low-CO 2 -grown cells but was not present in pyrenoids from high-CO 2 - grown cells. In addition, immunogold labeling experi- ments showed that the protein was mainly associated with membranes crossing the pyrenoid, while it was absent from the pyrenoid matrix. These studies have identified a putative intracellular CA polypeptide asso- ciated with the pyrenoid in Chlorella vulgaris, suggesting that this structure may play an important role in the operation of the CCM and the acclimation to low CO 2 conditions. Key words: Carbonic anhydrase – Chlorella – CO 2 - concentrating mechanism – Photosynthesis – Pyrenoid – Ribulose-1,5-bisphosphate carboxylase/oxygenase Introduction The unicellular green alga Chlorella vulgaris, as well as other green algae, has evolved a CO 2 -concentrating mechanism (CCM) induced by low external levels of CO 2 (see Coleman 1991; Badger and Price 1992). Low- CO 2 -acclimated cells exhibit a high ability to accumulate external inorganic carbon (C i ), elevating the CO 2 concentration at the site of ribulose-1,5-bisphosphate carboxylase/oxygenase (Rubisco). It has been reported that the induction of the CCM is correlated with the de-novo synthesis and/or up-regulation of at least six polypeptides in Chlorella vulgaris (Ramazanov et al. 1995). Although it has been suggested that these polypeptides may be involved in the CCM, there is little information about their location and functional role. Two of these low-CO 2 -inducible polypeptides, 36- and 38-kDa proteins, have been shown to be immuno- logically similar to the periplasmic carbonic anhydrase (pCA) of Chlamydomonas reinhardtii (Coleman et al. 1991; Ramazanov et al. 1995). In earlier studies using Chlorella saccharophila cells, the 38-kDa polypeptide was localized to intact chloroplasts, while the 36-kDa polypeptide has been suggested to represent an extra- cellular carbonic anhydrase (CA; Coleman et al. 1991). All cells of green algae studied so far possess an intracellular CA (Aizawa and Miyachi 1986; Su¨ltemeyer et al. 1993; Badger and Price 1994). Determination of CA activity has provided evidence for the existence of at least two internal CA activities in certain Chlorella species and Chlamydomonas reinhardtii, a soluble CA, located in the cytosol, and an insoluble membrane- bound CA, which is associated with the chloroplast (Miyachi et al. 1983; Pronina and Semenenko 1984; Su¨ltemeyer et al. 1990, 1995). Recently, Karlsson et al. Abbreviations: CA carbonic anhydrase; C i inorganic carbon (CO 2 + HCO 3 ); CCM CO 2 -concentrating mechanism; pCA (periplasmic) carbonic anhydrase; Rubisco ribulose-1,5-bispho- sphate carboxylase/oxygenase; WAU Wilbur-Anderson units Correspondence to: A. Villarejo; E-mail: [email protected]; Fax: 34 (91) 3978344 Planta (1998) 206: 416–425 A 38-kilodalton low-CO 2 -inducible polypeptide is associated with the pyrenoid in Chlorella vulgaris Arsenio Villarejo, M. Isabel Oru´s, Ziyadin Ramazanov, Flor Martı´nez Departamento de Biologı´a, Universidad Auto´noma de Madrid, E-28049 Madrid, Spain Received: 16 July 1997 / Accepted: 26 April 1998
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of A 38-kilodalton low-CO 2 -inducible polypeptide is associated with the pyrenoid in Chlorella...

Abstract. In the green alga Chlorella vulgaris UAM101, a CO2-concentrating mechanism (CCM) is inducedwhen cells are transferred from high (5%) to low(0.03%) CO2 concentrations. The induction of theCCM is correlated with de-novo synthesis of severalpolypeptides that remain to be identi®ed. The internalcarbonic anhydrase (CA; EC 4.2.1.1) activity increased6- to 7-fold within 6 h of acclimation to air. When crudehomogenates were further separated into soluble andinsoluble fractions, nearly all of the CA activity wasassociated with the membrane fraction. Immunoblotanalysis of cell homogenates probed with antibodiesraised against the 37-kDa subunit of periplasmic CA ofChlamydomonas reinhardtii showed a cross-reaction witha single 38-kDa polypeptide in both high- and low-CO2-grown cells. The up-regulation of the expression of the38-kDa polypeptide was closely correlated with theincrease in internal CA activity. Furthermore, its sub-cellular location was also correlated with the distribu-tion of the activity. Immunoblot analysis of pyrenoidfractions showed that the 38-kDa polypeptide wasconcentrated in the pyrenoids from low-CO2-growncells but was not present in pyrenoids from high-CO2-grown cells. In addition, immunogold labeling experi-ments showed that the protein was mainly associatedwith membranes crossing the pyrenoid, while it wasabsent from the pyrenoid matrix. These studies haveidenti®ed a putative intracellular CA polypeptide asso-ciated with the pyrenoid in Chlorella vulgaris, suggestingthat this structure may play an important role in theoperation of the CCM and the acclimation to low CO2
conditions.
Key words: Carbonic anhydrase ± Chlorella ± CO2-concentrating mechanism ± Photosynthesis ± Pyrenoid ±Ribulose-1,5-bisphosphate carboxylase/oxygenase
Introduction
The unicellular green alga Chlorella vulgaris, as well asother green algae, has evolved a CO2-concentratingmechanism (CCM) induced by low external levels ofCO2 (see Coleman 1991; Badger and Price 1992). Low-CO2-acclimated cells exhibit a high ability to accumulateexternal inorganic carbon (Ci), elevating the CO2
concentration at the site of ribulose-1,5-bisphosphatecarboxylase/oxygenase (Rubisco). It has been reportedthat the induction of the CCM is correlated with thede-novo synthesis and/or up-regulation of at least sixpolypeptides in Chlorella vulgaris (Ramazanov et al.1995). Although it has been suggested that thesepolypeptides may be involved in the CCM, there is littleinformation about their location and functional role.
Two of these low-CO2-inducible polypeptides, 36-and 38-kDa proteins, have been shown to be immuno-logically similar to the periplasmic carbonic anhydrase(pCA) of Chlamydomonas reinhardtii (Coleman et al.1991; Ramazanov et al. 1995). In earlier studies usingChlorella saccharophila cells, the 38-kDa polypeptidewas localized to intact chloroplasts, while the 36-kDapolypeptide has been suggested to represent an extra-cellular carbonic anhydrase (CA; Coleman et al. 1991).All cells of green algae studied so far possess anintracellular CA (Aizawa and Miyachi 1986; SuÈ ltemeyeret al. 1993; Badger and Price 1994). Determination ofCA activity has provided evidence for the existence of atleast two internal CA activities in certain Chlorellaspecies and Chlamydomonas reinhardtii, a soluble CA,located in the cytosol, and an insoluble membrane-bound CA, which is associated with the chloroplast(Miyachi et al. 1983; Pronina and Semenenko 1984;SuÈ ltemeyer et al. 1990, 1995). Recently, Karlsson et al.
Abbreviations: CA � carbonic anhydrase; Ci � inorganic carbon(CO2 + HCOÿ3 ); CCM � CO2-concentrating mechanism; pCA �(periplasmic) carbonic anhydrase; Rubisco � ribulose-1,5-bispho-sphate carboxylase/oxygenase; WAU � Wilbur-Anderson units
Correspondence to: A. Villarejo;E-mail: [email protected]; Fax: 34 (91) 3978344
Planta (1998) 206: 416±425
A 38-kilodalton low-CO2-inducible polypeptideis associated with the pyrenoid in Chlorella vulgaris
Arsenio Villarejo, M. Isabel Oru s, Ziyadin Ramazanov, Flor Martõ nez
Departamento de Biologõ a, Universidad Auto noma de Madrid, E-28049 Madrid, Spain
Received: 16 July 1997 /Accepted: 26 April 1998

(1995) identi®ed and isolated an intracellular membrane-bound CA in C. reinhardtii. This polypeptide shows ahigh degree of sequence homology with a-type CAs.These authors reported that more than 95% of theintracellular CA activity was typically found in theinsoluble fraction isolated from low-CO2-acclimatedcells of C. reinhardtii. In addition, Eriksson et al.(1996) identi®ed a 21-kDa low-CO2-inducible polypep-tide as a b-type CA located in the mitochondria isolatedfrom low-CO2-acclimated C. reinhardtii cells.
It has been reported that chloroplastic CA is impor-tant in the photosynthetic CO2 ¯ux of algal cells(SuÈ ltemeyer et al. 1993; Badger and Price 1994; Funkeet al. 1997). A major question concerning the role of thisisoenzyme in the CCM is its precise location within thechloroplast. Several lines of evidence indicate that thechloroplastic CA functions primarily as support of CO2
®xation by Rubisco (Spalding et al. 1983; Moroney et al.1987). Since most, if not all, Rubisco is found in thepyrenoid of green algae (Kuchitsu et al. 1991; Villarejoet al. 1997), the chloroplastic CA may be attachedsomehow to the pyrenoid structure, as in cyanobacteriawhere an internal CA is speci®cally located in thecarboxysome in close proximity to Rubisco (Price et al.1992; Badger and Price 1994). By analogy to thecarboxysomal model, McKay and Gibbs (1991) postu-lated that the pyrenoid may also play a signi®cant role inthe CCM. However, the evidence for association of thechloroplastic CA with pyrenoid is controversial and itslocation is still to be determined.
In this work, we present evidence for the associationof a low-CO2-inducible 38-kDa polypeptide with thepyrenoid in Chlorella vulgaris. Our results indicate thatup-regulation of this protein, which is immunologicallysimilar to the pCA of C. reinhardtii, is correlated withthe induction of internal CA activity. In addition,subcellular-fractionation experiments indicate that the38-kDa protein is mainly associated with the particulatefraction isolated from low-CO2-acclimated cells, whichcontained nearly all of the intracellular CA activity.
Materials and methods
Algal strain and culture conditions. The strain UAM 101 ofChlorella vulgaris (strain SAG 9.88 of the GoÈ ttingen UniversityCulture Collection, Germany) was inoculated in liquid culture at acell density of 4 ´ 104 cells mL)1 and was grown in Rodrõ guez-Lo pez A medium (Rodrõ guez-Lo pez 1964), continuously illumi-nated with an incident photosynthetic photon ¯ux density (PPFD)of 200 lmol m)2 s)1, obtained from ¯uorescent tubes (TDL150 W; Philips, Eindhoven, The Netherlands). The cultures weregrown in 1-L glass bottles at 28 °C and were bubbled with airenriched with 5% CO2 for 2 d. Cultures were then switched tosynchronous growth by using a 12 h light/12 h dark cycle for 3 dand maintained at similar cell density by daily dilution with freshmedium. Cell suspensions were aerated with either a CO2:airmixture [5:95 (v/v), high-CO2] or with air (0.03% CO2, low-CO2).All samples were harvested after 6 h of light.
Cell fractionation. Cells were harvested by centrifugation (5000 g,10 min), washed twice in 25 mM Tris-HCl bu�er (pH 8.0), andresuspended in 10 mL of the same bu�er. The cells were disruptedin a precooled French pressure cell (Aminco, American Instrument
Co., Urbana, Ill, USA) at 200 MPa. The homogenate wascentrifuged (500 g, 5 min) to remove unbroken cells. The superna-tant was then centrifuged at 100 000 g for 2 h (Centrikon T-2070ultracentrifuge, TFT 65.13 rotor; Kontron, Milan, Italy) toseparate soluble and insoluble fractions. The membrane pelletswere resuspended in 25 mM Tris-HCl bu�er (pH 8.0), and thesupernatant was saved as the soluble fraction. The above steps wereperformed at 4 °C.
Determination of CA activity. The activity of CA was potentiomet-rically determined by measuring the time for the pH to decreasefrom 8.2 to 7.4, at 2 °C, in a sample of 2 mL of 75 mM phosphatebu�er (pH 8.1), containing 1 mM EDTA and 0.1 mM dithio-threitol [DTT], upon addition of 2 mL of ice-cold CO2-saturateddistilled H2O (Ramazanov and Ca rdenas 1992). One Wilbur-Anderson unit (WAU) (Wilbur and Anderson 1948) was de®ned as:WAU � (t0/tc)1) 10, where t0 was the time for the pH change withbu�er controls and tc was the time obtained when CA-containingsamples were added.
Photosynthesis assays. The photosynthetic rate of algal cells wasmeasured in 1-mL samples with a Clark-type oxygen electrode(Hansatech, Norfolk, UK). Algae were centrifuged at 5000 g for10 min and the pellet was resuspended in 1 mL of 25 mM CO2-freeMes-NaOH bu�er (pH 5.7) to a chlorophyll density of 10 lg mL)1
and transferred to the electrode chamber, where they were allowedto consume the inorganic carbon of the bu�er and intracellularpool of inorganic carbon (CO2 + HCOÿ3 ) until no net O2 exchangewas observed. Di�erent concentrations of bicarbonate were addedand the rate of O2 evolution measured.
Analysis by SDS-PAGE and western blotting. Total cell homoge-nates and subcellular fractions were subjected to SDS-PAGE [12%(w/v) polyacrylamide; Laemmli 1970]. Gels were stained withCoomasie brilliant blue or transferred electrophoretically to anitrocellulose ®lter (0.2 lm pore size) for western blotting. Theimmunoblot assay was performed according to the protocol fromBio-Rad (Richmond, Calif., USA) except that 5% non-fat dry milkwas used to block nitrocellulose. Antibodies on the ®lters weredetected with a goat anti-rabbit IgG (H+L) horseradish-peroxi-dase-conjugate antibody using 4-chloro-1-naphthol (Bio-Rad) as asubstrate.
Polyclonal antibodies raised against the 37-kDa pCA ofC. reinhardtii were kindly provided by Dr. J. Moroney (LouisianaState University, Baton Rouge, La. USA). Polyclonal antibodiesagainst the large subunit of Rubisco of Euglena gracilis were kindlyprovided by Prof. C. Ascaso (Consejo Superior de InvestigacionesCientõÂ ®cas, Madrid, Spain). Polyclonal antibodies raised againstRubisco activase of C. reinhardtii were kindly provided by Prof. A.Portis (University of Illinois, Urbana, Ill., USA). Polyclonalantibodies raised against D-1 protein of C. reinhardtii wereprovided by Prof. Z. Ramazanov. Polyclonal antibodies raisedagainst ferredoxin-NADP+-reductase of Chlorella fusca was agenerous gift from Dr. M.L. Peleato (Universidad de Zaragoza,Zaragoza, Spain).
Pyrenoid isolation. Pyrenoids were isolated from C. vulgaris cellsaccording to the method of Kuchitsu et al. (1988) with somemodi®cations. High- and low-CO2-grown cells were harvested bycentrifugation, washed twice with 30 mM Hepes-KOH bu�er (pH7.8, bu�er A), and treated with 0.1% (w/v) HgCl2 dissolved inbu�er A for 4 h at 4 °C. After washing and resuspending the Hg2+-treated cells with bu�er A twice, cells were disrupted with an ice-cold French pressure cell (Aminco) at 100 MPa. The homogenatewas incubated with 1% (v/v) Igepal CA-630 (Sigma) at 0 °C for30 min. After incubation, the homogenate was centrifuged at30 000 g for 20 min, and the precipitate was resuspended in bu�erA containing 1% (v/v) Igepal CA-630. After incubation at 0 °C for30 min, the suspension was centrifuged at 30 000 g for 20 min. Theprecipitate (crude pyrenoid fraction) was layered onto a discontin-uous sucrose density gradient (1 M and 2.5 M) and centrifuged at
A. Villarejo et al.: Pyrenoid-associated polypeptides in Chlorella 417
2300 g for 30 min. The layer between 1 M and 2.5 M sucrose wascollected, washed with Igepal CA-630 dissolved in bu�er A andcentrifuged at 30 000 g for 20 min. The precipitate was resuspend-ed in bu�er A and used for SDS-PAGE and transmission electronmicroscopy analysis.
Electron microscopy and immunolocalization. For transmissionelectron microscopy, cells and isolated pyrenoids were ®xed in2.5% (v/v) glutaraldehyde added to 0.05 M cacodylate bu�er (pH7.4) at 4 °C for 2.5 h. Samples were rinsed in the same bu�er. Afterrinsing, the samples were dehydrated in a graded ethanol series,then in®ltrated and embedded in LR White medium resin (LondonResin Co., London, UK) according to Ramazanov et al. (1994).
Thin sections were cut using a diamond knife and mounted onnickel grids. Grids were then ¯oated section side down on drops ofthe following solutions: 2% (w/v) bovine serum albumin (BSA) inTris-bu�ered saline (TBS, 10 mM Tris-HCl and 150 mMNaCl, pH7.4) for 1 h; ®rst antibody diluted 1:500 in TBS/BSA for 90 min;0.2% (w/v) BSA in TBS, ®ve times for 2 min each time; 10 nmgold-protein A diluted 1:25 in TBS containing 0.2% BSA for 1 h;0.2% BSA in TBS, ®ve times for 3 min each time; and TBS, threetimes for 3 min each time (Ascaso et al. 1995).
Grids were poststained with 2% (w/v) aqueous uranyl acetatefor 5 min and lead citrate for 45 s. Sections were viewed andphotographed in a JEOL 1200EX transmission electron microscope(Tokyo, Japan) at 80 kV.
Analytical determinations. Protein concentration was estimatedaccording to Bradford (1976). Chlorophyll was extracted withabsolute methanol and spectrophotometrically determined accord-ing to Holden (1965).
Results
E�ect of external CO2 concentration on CA activity. Theactivity of CA was measured in C. vulgaris cells beforeand after cellular breakage to determine the activity ofextracellular and intracellular CA, respectively. No CAactivity was detected in the suspension of intact cells(data not shown), con®rming our previous resultsshowing that this strain does not have extracellular CA(Villarejo et al. 1997).
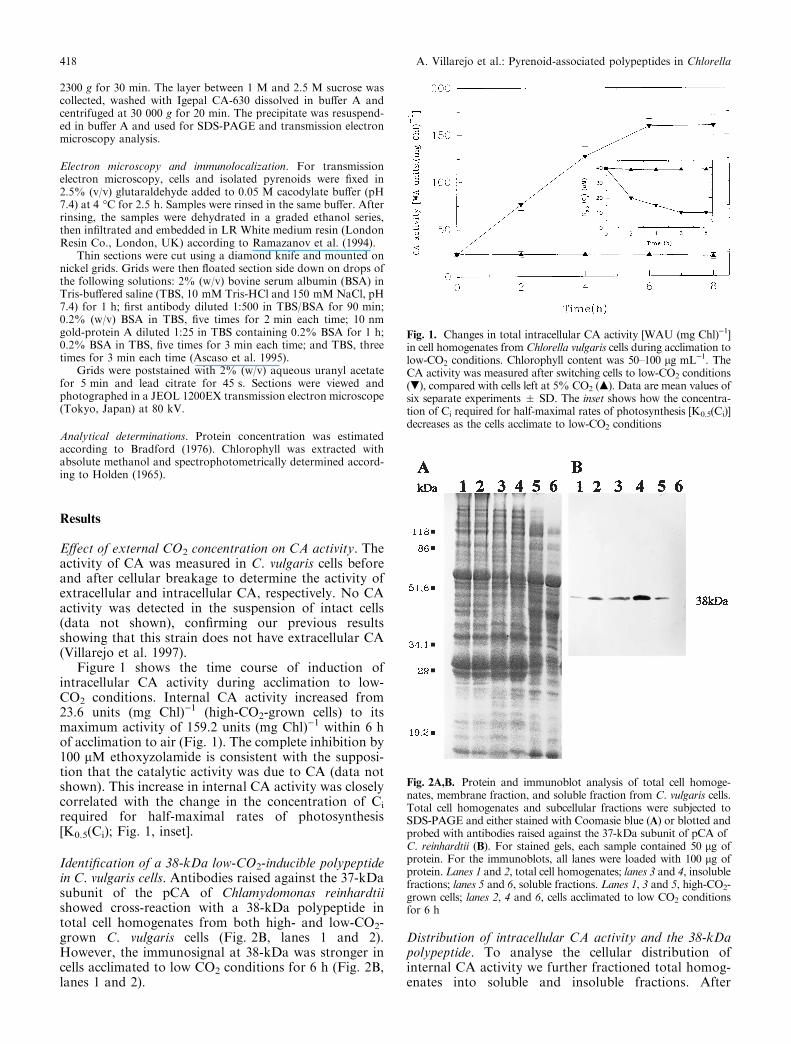
Figure 1 shows the time course of induction ofintracellular CA activity during acclimation to low-CO2 conditions. Internal CA activity increased from23.6 units (mg Chl))1 (high-CO2-grown cells) to itsmaximum activity of 159.2 units (mg Chl))1 within 6 hof acclimation to air (Fig. 1). The complete inhibition by100 lM ethoxyzolamide is consistent with the supposi-tion that the catalytic activity was due to CA (data notshown). This increase in internal CA activity was closelycorrelated with the change in the concentration of Ci
required for half-maximal rates of photosynthesis[K0.5(Ci); Fig. 1, inset].
Identi®cation of a 38-kDa low-CO2-inducible polypeptidein C. vulgaris cells. Antibodies raised against the 37-kDasubunit of the pCA of Chlamydomonas reinhardtiishowed cross-reaction with a 38-kDa polypeptide intotal cell homogenates from both high- and low-CO2-grown C. vulgaris cells (Fig. 2B, lanes 1 and 2).However, the immunosignal at 38-kDa was stronger incells acclimated to low CO2 conditions for 6 h (Fig. 2B,lanes 1 and 2).
Distribution of intracellular CA activity and the 38-kDapolypeptide. To analyse the cellular distribution ofinternal CA activity we further fractioned total homog-enates into soluble and insoluble fractions. After
Fig. 1. Changes in total intracellular CA activity [WAU (mg Chl))1]in cell homogenates from Chlorella vulgaris cells during acclimation tolow-CO2 conditions. Chlorophyll content was 50±100 lg mL)1. TheCA activity was measured after switching cells to low-CO2 conditions(.), compared with cells left at 5% CO2 (m). Data are mean values ofsix separate experiments � SD. The inset shows how the concentra-tion of Ci required for half-maximal rates of photosynthesis [K0.5(Ci)]decreases as the cells acclimate to low-CO2 conditions
Fig. 2A,B. Protein and immunoblot analysis of total cell homoge-nates, membrane fraction, and soluble fraction from C. vulgaris cells.Total cell homogenates and subcellular fractions were subjected toSDS-PAGE and either stained with Coomasie blue (A) or blotted andprobed with antibodies raised against the 37-kDa subunit of pCA ofC. reinhardtii (B). For stained gels, each sample contained 50 lg ofprotein. For the immunoblots, all lanes were loaded with 100 lg ofprotein. Lanes 1 and 2, total cell homogenates; lanes 3 and 4, insolublefractions; lanes 5 and 6, soluble fractions. Lanes 1, 3 and 5, high-CO2-grown cells; lanes 2, 4 and 6, cells acclimated to low CO2 conditionsfor 6 h
418 A. Villarejo et al.: Pyrenoid-associated polypeptides in Chlorella

centrifugation of total homogenates at 100 000 g for 2 hthe majority of the activity was found in the insolublefraction from both high- and low-CO2-grown cells of C.vulgaris, although the CA activity in the pellet was 6- to7-fold higher in cells acclimated to low CO2 for 6 h(Table 1). In low-CO2-grown cells more than 95% of theactivity was found in the insoluble fraction (Table 1),while the activity associated with the soluble fractionwas negligible (Table 1). However, in high-CO2-growncells soluble CA activity contributed about 23% to thetotal internal CA activity (Table 1).
Figure 2 shows protein and immunoblot analysis oftotal cell homogenates and subcellular fractions isolatedfrom high- and low-CO2-grown C. vulgaris cells. Immu-noblot analysis using antibodies raised against pCA ofChlamydomonas reinhardtii indicates that the 38-kDapolypeptide was present in insoluble fractions isolatedfrom both high- and low-CO2-grown cells (Fig. 2B,lanes 3 and 4). The immunosignal at 38-kDa wasstronger in the insoluble fractions with respect to totalcell homogenates, showing that this polypeptide ismainly located in the particulate fraction. However, thisprotein was absent in the soluble fraction isolated fromlow-CO2-grown cells (Fig. 2B, lane 6), while the anti-bodies were able to detect low levels of this polypeptidein the soluble fraction from high-CO2-grown cells(Fig. 2B, lane 5).
Characterization of pyrenoid-enriched fractions. To de-termine the purity and intactness of the isolatedpyrenoids, these fractions were characterized by proteinanalysis and transmission electron microscopy. Figure 3shows the polypeptide pattern of total cell homogenatesand pyrenoid-enriched fractions isolated from high- andlow-CO2-grown C. vulgaris cells. The main componentsof pyrenoid-enriched fractions were 52- and 14-kDapolypeptides (Fig. 3, lanes 3 and 4). These two poly-peptides represented the large and small subunits ofRubisco, as has been previously shown in Chlamydomo-nas reinhardtii (Kuchitsu et al. 1988, 1991).
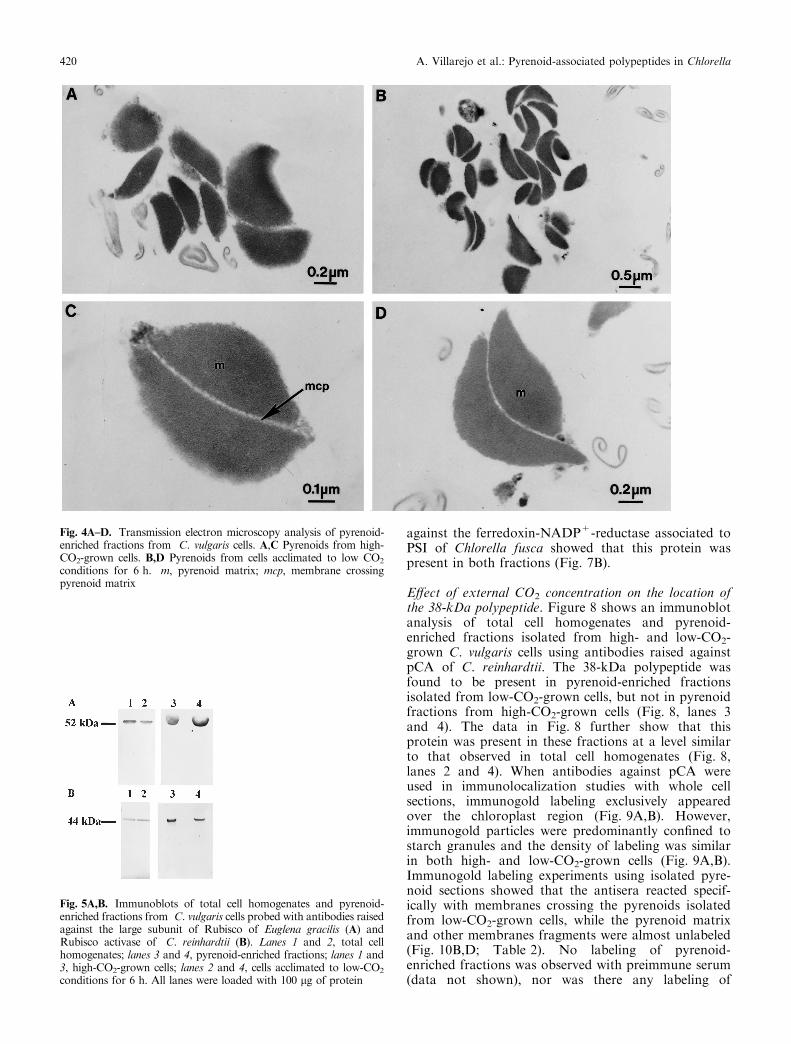
Transmission electron microscopy shows that theisolated fraction was enriched in intact pyrenoids
(Fig. 4A,B). The pyrenoids isolated from high- andlow-CO2-grown cells showed the typical ovoid shapeobserved in cellular sections (see Villarejo et al. 1997).The pyrenoid matrix appeared as an electron-densestructure divided by a channel (Fig. 4C,D). The intact-ness of the pyrenoid structure was con®rmed by thepresence of membranes crossing the pyrenoid matrix inmost of the sections analysed (Fig. 4C), a typicalfeature of the genus Chlorella (Ikeda and Takeda1995; Villarejo et al. 1997). In addition, some mem-brane fragments appeared to be associated with intactpyrenoids (Fig. 4).
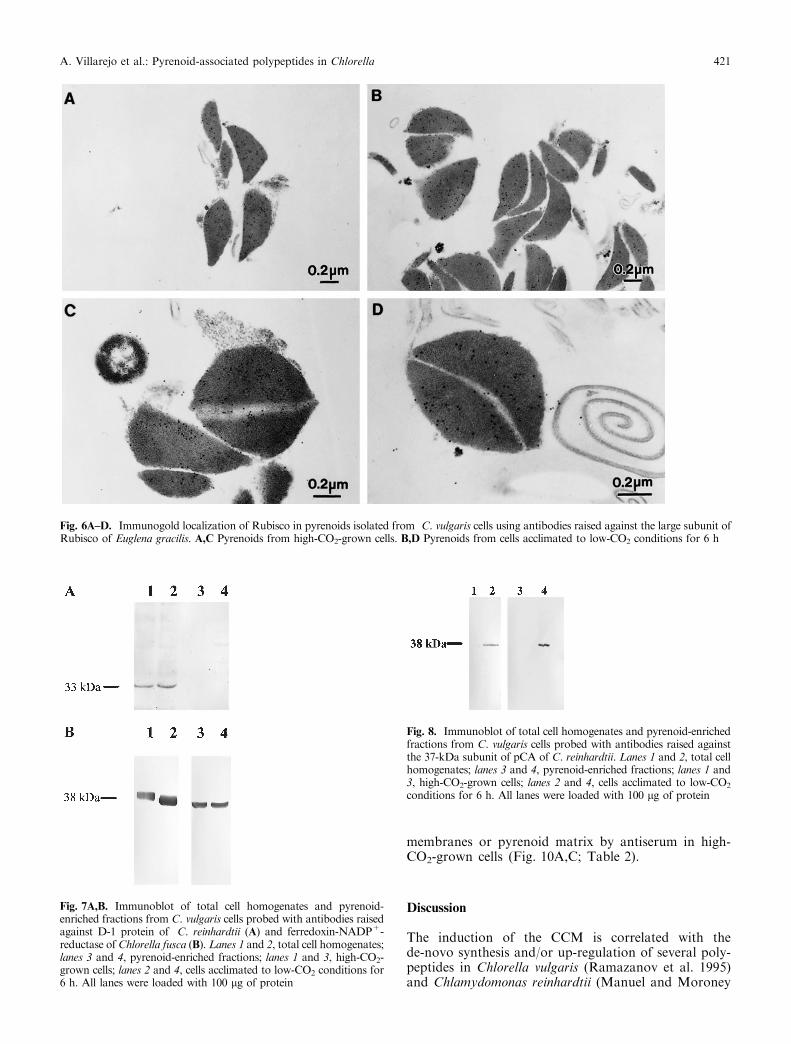
Immunoblot analysis using antibodies raised againstthe large subunit of Rubisco of Euglena gracilis showedthat this polypeptide was enriched in pyrenoid fractionswith respect to total cell homogenates (Fig. 5A). Theseresults were consistent with data from immunogold-labeling experiments, showing that the matrices ofpyrenoids isolated from both high- and low-CO2-growncells were heavily labeled with antiserum (Fig. 6C,D).These results also indicated that protein antigenicity wasnot altered by HgCl2-treatment.We have previouslyshown in cell sections of C. vulgaris that Rubisco isexclusively located in the pyrenoid matrix (Villarejo et al.1997). In addition to the large subunit of Rubisco,immunoblot analysis with antibodies raised againstRubisco activase showed that this polypeptide was alsoenriched in pyrenoid fractions isolated from both high-and low-CO2-grown C. vulgaris cells (Fig. 5B).
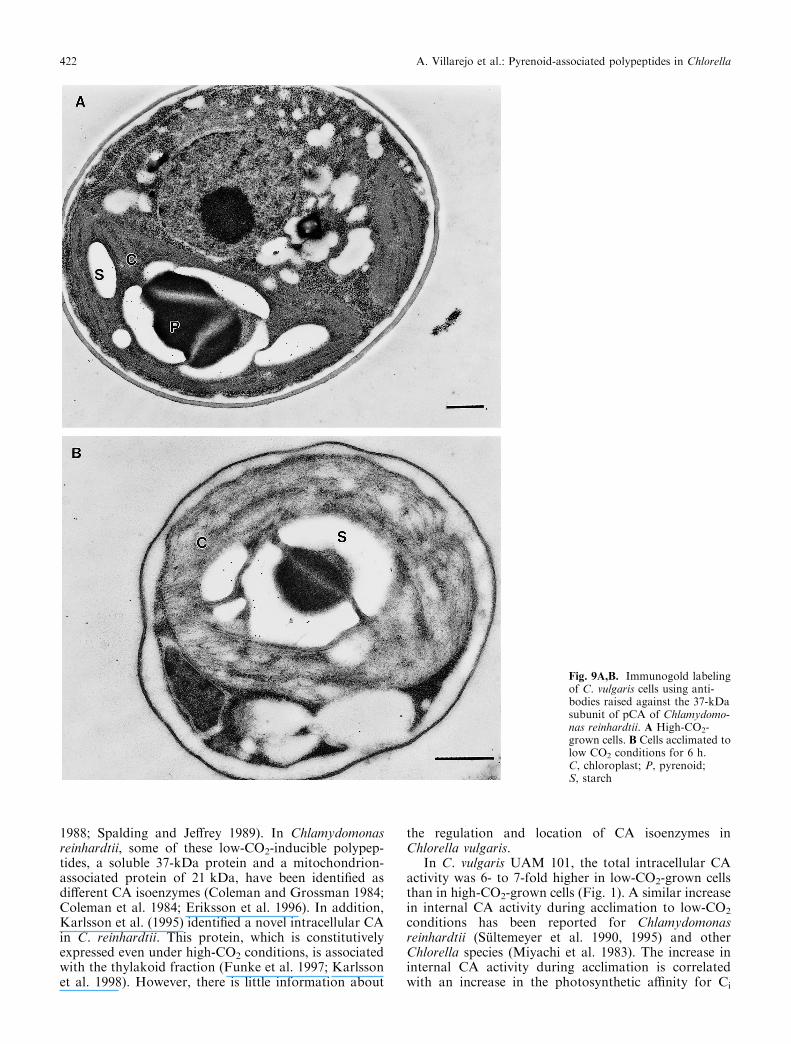
Western blot analysis was used to determine thenature of membranes co-purifying with pyrenoids.Immunoblot analysis of the pyrenoid fractions probedwith antibodies against D-1 protein of PSII showed thatthis polypeptide was absent in pyrenoids isolated fromboth high- and low-CO2-grown C. vulgaris cells(Fig. 7A). Immunoblot analysis with antibodies raised
Fig. 3. Coomasie-blue-stained SDS-PAGE gel of total cell homoge-nates and pyrenoid-enriched fractions from C. vulgaris cells. Lanes 1and 2, total cell homogenates; lanes 3 and 4, pyrenoid-enrichedfractions; lanes 1 and 3, high-CO2-grown cells; lanes 2 and 4, cellsacclimated to low CO2 conditions for 6 h. All lanes were loaded with100 lg of protein
Table 1. Carbonic anhydrase activities [WAU (mg Chl))1] in cellhomogenates from C. vulgaris cells which had been grown eitheron 5% CO2 (high-CO2) or transferred to low CO2 conditions for6 h (low-CO2). After cellular breakage an aliquot was removed tomeasure total intracellular CA; the remaining homogenate wasfurther fractioned into supernatant (soluble CA) and pellet (in-soluble CA; for details, see Materials and methods). Chlorophyllcontent was 100±150 lg mL)1. Numbers in parentheses indicateenzyme activity relative to the enzyme activity in total cell homo-genates. Data are mean values of ®ve separate experiments � SD
Subcellular fraction Activity of CA [WAU (mg Chl))1]
High-CO2 Low-CO2
Total cellhomogenate
23.6 � 1.40 (100%) 159.2 � 7.75 (100%)
Soluble fraction 5.3 � 0.46 (23%) 4.0 � 0.36 (2,5%)Membrane fraction 18.9 � 0.54 (80%) 152.8 � 12.2 (96%)
A. Villarejo et al.: Pyrenoid-associated polypeptides in Chlorella 419
against the ferredoxin-NADP+-reductase associated toPSI of Chlorella fusca showed that this protein waspresent in both fractions (Fig. 7B).
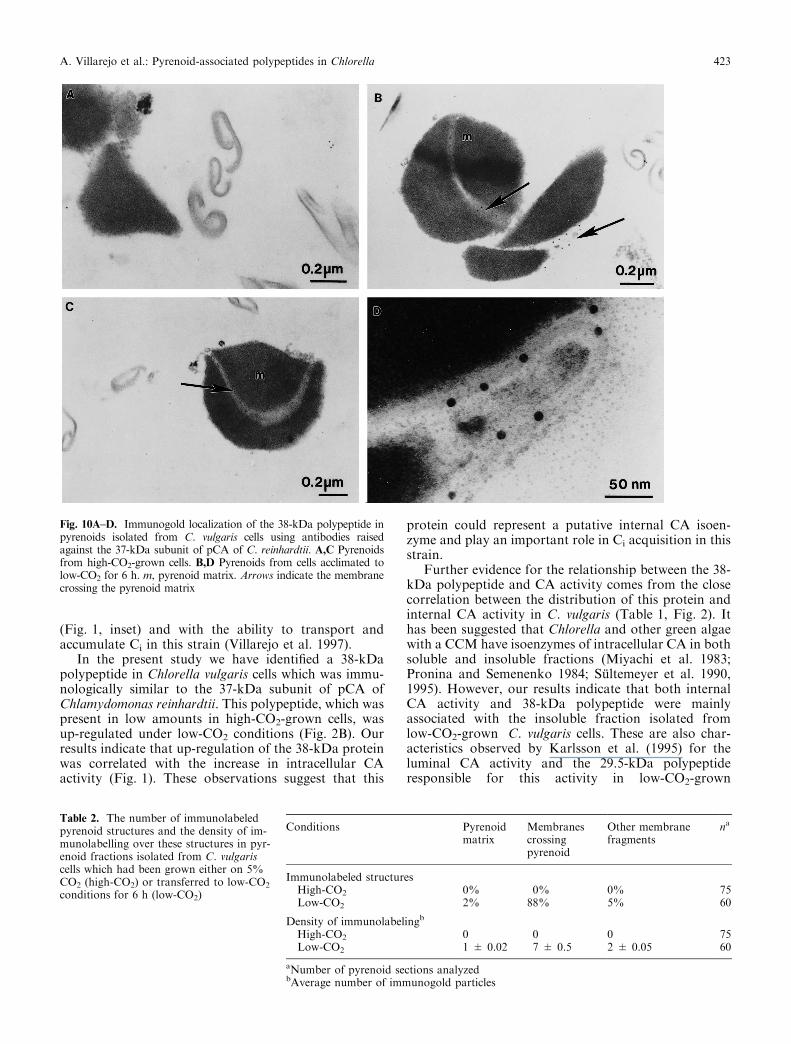
E�ect of external CO2 concentration on the location ofthe 38-kDa polypeptide. Figure 8 shows an immunoblotanalysis of total cell homogenates and pyrenoid-enriched fractions isolated from high- and low-CO2-grown C. vulgaris cells using antibodies raised againstpCA of C. reinhardtii. The 38-kDa polypeptide wasfound to be present in pyrenoid-enriched fractionsisolated from low-CO2-grown cells, but not in pyrenoidfractions from high-CO2-grown cells (Fig. 8, lanes 3and 4). The data in Fig. 8 further show that thisprotein was present in these fractions at a level similarto that observed in total cell homogenates (Fig. 8,lanes 2 and 4). When antibodies against pCA wereused in immunolocalization studies with whole cellsections, immunogold labeling exclusively appearedover the chloroplast region (Fig. 9A,B). However,immunogold particles were predominantly con®ned tostarch granules and the density of labeling was similarin both high- and low-CO2-grown cells (Fig. 9A,B).Immunogold labeling experiments using isolated pyre-noid sections showed that the antisera reacted specif-ically with membranes crossing the pyrenoids isolatedfrom low-CO2-grown cells, while the pyrenoid matrixand other membranes fragments were almost unlabeled(Fig. 10B,D; Table 2). No labeling of pyrenoid-enriched fractions was observed with preimmune serum(data not shown), nor was there any labeling of
Fig. 4A±D. Transmission electron microscopy analysis of pyrenoid-enriched fractions from C. vulgaris cells. A,C Pyrenoids from high-CO2-grown cells. B,D Pyrenoids from cells acclimated to low CO2
conditions for 6 h. m, pyrenoid matrix; mcp, membrane crossingpyrenoid matrix
Fig. 5A,B. Immunoblots of total cell homogenates and pyrenoid-enriched fractions from C. vulgaris cells probed with antibodies raisedagainst the large subunit of Rubisco of Euglena gracilis (A) andRubisco activase of C. reinhardtii (B). Lanes 1 and 2, total cellhomogenates; lanes 3 and 4, pyrenoid-enriched fractions; lanes 1 and3, high-CO2-grown cells; lanes 2 and 4, cells acclimated to low-CO2
conditions for 6 h. All lanes were loaded with 100 lg of protein
420 A. Villarejo et al.: Pyrenoid-associated polypeptides in Chlorella
membranes or pyrenoid matrix by antiserum in high-CO2-grown cells (Fig. 10A,C; Table 2).
Discussion
The induction of the CCM is correlated with thede-novo synthesis and/or up-regulation of several poly-peptides in Chlorella vulgaris (Ramazanov et al. 1995)and Chlamydomonas reinhardtii (Manuel and Moroney
Fig. 6A±D. Immunogold localization of Rubisco in pyrenoids isolated from C. vulgaris cells using antibodies raised against the large subunit ofRubisco of Euglena gracilis. A,C Pyrenoids from high-CO2-grown cells. B,D Pyrenoids from cells acclimated to low-CO2 conditions for 6 h
Fig. 7A,B. Immunoblot of total cell homogenates and pyrenoid-enriched fractions from C. vulgaris cells probed with antibodies raisedagainst D-1 protein of C. reinhardtii (A) and ferredoxin-NADP+-reductase of Chlorella fusca (B). Lanes 1 and 2, total cell homogenates;lanes 3 and 4, pyrenoid-enriched fractions; lanes 1 and 3, high-CO2-grown cells; lanes 2 and 4, cells acclimated to low-CO2 conditions for6 h. All lanes were loaded with 100 lg of protein
Fig. 8. Immunoblot of total cell homogenates and pyrenoid-enrichedfractions from C. vulgaris cells probed with antibodies raised againstthe 37-kDa subunit of pCA of C. reinhardtii. Lanes 1 and 2, total cellhomogenates; lanes 3 and 4, pyrenoid-enriched fractions; lanes 1 and3, high-CO2-grown cells; lanes 2 and 4, cells acclimated to low-CO2
conditions for 6 h. All lanes were loaded with 100 lg of protein
A. Villarejo et al.: Pyrenoid-associated polypeptides in Chlorella 421
1988; Spalding and Je�rey 1989). In Chlamydomonasreinhardtii, some of these low-CO2-inducible polypep-tides, a soluble 37-kDa protein and a mitochondrion-associated protein of 21 kDa, have been identi®ed asdi�erent CA isoenzymes (Coleman and Grossman 1984;Coleman et al. 1984; Eriksson et al. 1996). In addition,Karlsson et al. (1995) identi®ed a novel intracellular CAin C. reinhardtii. This protein, which is constitutivelyexpressed even under high-CO2 conditions, is associatedwith the thylakoid fraction (Funke et al. 1997; Karlssonet al. 1998). However, there is little information about
the regulation and location of CA isoenzymes inChlorella vulgaris.
In C. vulgaris UAM 101, the total intracellular CAactivity was 6- to 7-fold higher in low-CO2-grown cellsthan in high-CO2-grown cells (Fig. 1). A similar increasein internal CA activity during acclimation to low-CO2
conditions has been reported for Chlamydomonasreinhardtii (SuÈ ltemeyer et al. 1990, 1995) and otherChlorella species (Miyachi et al. 1983). The increase ininternal CA activity during acclimation is correlatedwith an increase in the photosynthetic a�nity for Ci
Fig. 9A,B. Immunogold labelingof C. vulgaris cells using anti-bodies raised against the 37-kDasubunit of pCA of Chlamydomo-nas reinhardtii. A High-CO2-grown cells. B Cells acclimated tolow CO2 conditions for 6 h.C, chloroplast; P, pyrenoid;S, starch
422 A. Villarejo et al.: Pyrenoid-associated polypeptides in Chlorella
(Fig. 1, inset) and with the ability to transport andaccumulate Ci in this strain (Villarejo et al. 1997).
In the present study we have identi®ed a 38-kDapolypeptide in Chlorella vulgaris cells which was immu-nologically similar to the 37-kDa subunit of pCA ofChlamydomonas reinhardtii. This polypeptide, which waspresent in low amounts in high-CO2-grown cells, wasup-regulated under low-CO2 conditions (Fig. 2B). Ourresults indicate that up-regulation of the 38-kDa proteinwas correlated with the increase in intracellular CAactivity (Fig. 1). These observations suggest that this
protein could represent a putative internal CA isoen-zyme and play an important role in Ci acquisition in thisstrain.
Further evidence for the relationship between the 38-kDa polypeptide and CA activity comes from the closecorrelation between the distribution of this protein andinternal CA activity in C. vulgaris (Table 1, Fig. 2). Ithas been suggested that Chlorella and other green algaewith a CCM have isoenzymes of intracellular CA in bothsoluble and insoluble fractions (Miyachi et al. 1983;Pronina and Semenenko 1984; SuÈ ltemeyer et al. 1990,1995). However, our results indicate that both internalCA activity and 38-kDa polypeptide were mainlyassociated with the insoluble fraction isolated fromlow-CO2-grown C. vulgaris cells. These are also char-acteristics observed by Karlsson et al. (1995) for theluminal CA activity and the 29.5-kDa polypeptideresponsible for this activity in low-CO2-grown
Fig. 10A±D. Immunogold localization of the 38-kDa polypeptide inpyrenoids isolated from C. vulgaris cells using antibodies raisedagainst the 37-kDa subunit of pCA of C. reinhardtii. A,C Pyrenoidsfrom high-CO2-grown cells. B,D Pyrenoids from cells acclimated tolow-CO2 for 6 h. m, pyrenoid matrix. Arrows indicate the membranecrossing the pyrenoid matrix
Table 2. The number of immunolabeledpyrenoid structures and the density of im-munolabelling over these structures in pyr-enoid fractions isolated from C. vulgariscells which had been grown either on 5%CO2 (high-CO2) or transferred to low-CO2
conditions for 6 h (low-CO2)
Conditions Pyrenoidmatrix
Membranescrossingpyrenoid
Other membranefragments
na
Immunolabeled structuresHigh-CO2 0% 0% 0% 75Low-CO2 2% 88% 5% 60
Density of immunolabelingb
High-CO2 0 0 0 75Low-CO2 1 � 0.02 7 � 0.5 2 � 0.05 60
aNumber of pyrenoid sections analyzedbAverage number of immunogold particles
A. Villarejo et al.: Pyrenoid-associated polypeptides in Chlorella 423

Chlamydomonas cells. The studies providing evidence forseveral intracellular isoenzymes were based on resultsfrom mass-spectrometric techniques, which may havedetected CAs ± immunologically di�erent from pCA ±not readily apparent using the potentiometrical methodthat we used in our experiments.
It has been previously shown that insoluble CAactivity is mainly associated with the chloroplast in bothhigh- and low-CO2-grown cells of Chlamydomonasreinhardtii and that this activity increases during accli-mation to low-CO2 conditions (SuÈ ltemeyer et al. 1995).These results were recently con®rmed by the work ofKarlsson et al. (1998) who showed that the insoluble CApolypeptide is exclusively associated with the thylakoidfraction in both high- and low-CO2-grown cells. How-ever, when total cell homogenate from high-CO2-grownC. vulgaris cells was further fractioned, the CA activitywas found in both soluble and insoluble fractions(Table 1). Furthermore, the 38-kDa polypeptide showedthe same distribution as CA activity in these cells(Fig. 2). Since no cross-reactivity of antisera againstPSII-associated polypeptides was observed in the solublefraction (data not shown), the possibility of somecontamination of this fraction with lighter thylakoidmembranes can be ruled out. There are several possibleexplanations of this di�erential distribution: ®rstly, thelocation of 38-kDa protein changes in response tovariations in the CO2 concentration of the medium;secondly, the 38-kDa polypeptide is associated with aparticulate fraction, possibly in the pyrenoids that maybe disrupted more easily in high-CO2-grown cells. Infact, pyrenoids of high-CO2-grown cells of green algaeappear to be less compact than those of low-CO2-growncells (Ramazanov et al. 1994; Villarejo et al. 1997).
Our results strongly suggest that the 38-kDa low-CO2-inducible polypeptide was mainly concentrated inthe pyrenoid in low-CO2-grown C. vulgaris cells (Fig. 8).In addition, immunogold-labeling experiments indicatethat the 38-kDa polypeptide was mainly associated withmembranes crossing the pyrenoid matrix, while no labelwas observed over the pyrenoid matrix (Fig. 10,Table 2). These membranes only preserved PSI-associ-ated polypeptides, while polypeptides associated withPSII were absent (Fig. 7A,B). These results suggest thatthese membranes were di�erent from stromal thylakoidsand could represent the intrapyrenoid membrane. Infact, the heterogeneity with respect to the distribution ofPSs between stromal and intrapyrenoid thylakoids hasbeen previously reported in the red alga Porphyridiumcruentum (McKay and Gibbs 1991). In earlier studies,Pronina and Semenenko (1984) postulated that much ofthe CA found in Chlorella was tightly bound tothylakoid membranes, although no precise criteria wereobtained indicating the absence of contamination ofthese fractions by other cell membranes. Later, theseauthors showed the correlation between the insolubleCA activity and the presence of PSI-associated poly-peptides (Pronina et al. 1990). Mutants of C. reinhardtiilacking PSI have very low CA activity, while mutantspreserving complexes of PSI have levels of this activitysimilar to those of wild-type cells. Our results con®rm
those of Pronina et al. (1990) and suggest that a putativeCA polypeptide was associated with membranes cross-ing the pyrenoid that only preserved PSI-associatedpolypeptides.
The question arises whether the 38-kDa polypeptideis located in the pyrenoid from high-CO2-grown cellsand is lost during the isolation process or whether thisprotein becomes concentrated in this structure duringacclimation to low CO2 conditions. The intactness ofpyrenoids isolated from high-CO2-grown cells and theabsence of any immunosignal in these fractions argue infavour of the second possibility. In low-CO2-grown cells,the association of the 38-kDa protein with pyrenoidswould completely prevent its solubility. However, inhigh-CO2-grown cells the 38-kDa polypeptide may bemainly associated with stromal thylakoids where thisprotein could be readily dissociated. Unfortunately,immunogold labeling experiments using whole cellsections did not allow us to determine the preciselocalization of this protein in high-CO2-grown cells(Fig. 9). In these experiments, immunogold particleswere predominantly con®ned to starch granules in bothhigh- and low-CO2-grown cells (Fig. 9A,B). The factthat the density of immunolabeling over the chloroplastregion of high-CO2-grown cells was similar to thatobserved in low-CO2-grown cells, which showed ahigher level of the 38-kDa polypeptide, suggested thatthe immunolabeling may be due to an unspeci®c bindingof the antibodies to the starch. Immunocytochemicalanalysis of pyrenoid sections, where no starch granuleswere observed, showed that the immunolabeling patternwas dependent on the growth conditions (Fig. 10).
The role of this pyrenoid-associated polypeptide isspeculative. It has been shown that Chlamydomonasreinhardtii mutants, lacking a chloroplastic CA, overac-cumulate Ci at levels higher than those observed in wild-type cells (Spalding et al. 1983; Moroney et al. 1987). Onthe basis of these results it has been postulated thatchloroplastic CA may be responsible for the dehydrationof the intracellular Ci pool in the pyrenoid, whereRubisco is exclusively located (Moroney and Mason1991; Pronina and Semenenko 1992). Our resultsstrongly suggest that the 38-kDa low-CO2-induciblepolypeptide, which is immunologically similar to thepCA of C. reinhardtii, is associated with the pyrenoid inC. vulgaris. The correlation between the induction of theCCM and the up-regulation of this protein is consistentwith the above-mentioned hypothesis postulating thatthe pyrenoid is not only the site of CO2 ®xation but alsothe site of HCOÿ3 dehydration, playing an essential rolein the operation of CCM. Further studies of the aminoacid sequence and catalytic activity of this protein will becrucial to the elucidation of its function in the CCM inthis strain of Chlorella.
The authors thank Dr. J.M. Gonzalez (Departamento de Cit-ologõ a, Universidad Auto noma de Madrid, Spain) for helpfuldiscussion about electron microscopy techniques. This work wassupported by a grant from Direccio n General de Investigacio n,Ciencia y Tecnologõ a. A.V. was supported by a fellowship ofConsejerõ a de Educacio n y Ciencia, Comunidad Auto noma deMadrid, Spain.
424 A. Villarejo et al.: Pyrenoid-associated polypeptides in Chlorella

References
Aizawa K, Miyachi S (1986) Carbonic anhydrase and CO2
concentrating mechanisms in microalgae and cyanobacteria.FEMS Microbiol Rev 39: 215±233
Ascaso C, Valladares F, De los Rios A (1995) New ultrastructureaspects of pyrenoids of the lichen photobiont Trebouxia(Microthamniales, Chlorophyta). J Phycol 31: 114±119
Badger MR, Price GD (1992) The CO2 concentrating mechanism incyanobacteria and microalgae. Physiol Plant 84: 606±615
Badger MR, Price GD (1994) The role of carbonic anhydrase inphotosynthesis. Annu Rev Plant Physiol Plant Mol Biol 45:369±392
Bradford M (1976) A rapid and sensitive method for the quanti-®cation of microgram quantities of protein utilizing theprinciple of protein-dye binding. Anal Biochem 72: 248±254
Coleman JR (1991) The molecular and biochemical analyses ofCO2-concentrating mechanism in cyanobacteria and microal-gae. Plant Cell Environ 14: 861±867
Coleman JR, Grossman AR (1984) Biosynthesis of carbonicanhydrase in Chlamydomonas reinhardtii during adaptation tolow CO2. Proc Natl Acad Sci USA 81: 6049±6053
Coleman JR, Berry JA, Togasaki RT, Grossman AR (1984)Identi®cation of extracellular carbonic anhydrase of Chlamydo-monas reinhardtii. Plant Physiol 76: 472±477
Coleman JR, Rotatore C, Williams TG, Colman B (1991)Identi®cation and localization of carbonic anhydrase in twoChlorella species. Plant Physiol 95: 331±334
Eriksson M, Karlsson J, Ramazanov Z, GardestroÈ m P, SamuelssonG (1996) Discovery of an algal mitochondrial carbonic anhy-drase: molecular cloning and characterization of a low-CO2-induced polypeptide in Chlamydomonas reinhardtii. Proc NatlAcad Sci USA 93: 12031±12034
Funke RP, Kovar JL, Weeks DP (1997) Intracellular carbonicanhydrase is essential to photosynthesis in Chlamydomonasreinhardtii at atmospheric levels of CO2. Plant Physiol 114: 237±244
Holden M (1965) Chlorophylls. In: Goodwin TW (ed) Chemistryand biochemistry of plant pigments. Academic Press, NewYork, pp 3±28
Ikeda T, Takeda H (1995) Species-speci®c di�erences of pyrenoidsin Chlorella (Chlorophyta). J Phycol 31: 813±818
Karlsson J, Hiltonen T, Husic DH, Ramazanov Z, Samuelsson G(1995) Intracellular carbonic anhydrase of Chlamydomonasreinhardtii. Plant Physiol 109: 533±539
Karlsson J, Clarke AK, Chen Z, Hugghins SY, Park Y, Husic DH,Moroney JV, Samuelsson G (1998) A novel a-type carbonicanhydrase associated with the thylakoid membrane inChlamydomonas reinhardtii is required for growth at ambientCO2. EMBO J 17: 1208±1216
Kuchitsu K, Tsuzuki M, Miyachi S (1988) Characterization of thepyrenoid isolated from unicellular green alga Chlamydomonasreinhardtii: particulate form of Rubisco protein. Protoplasma144: 17±24
Kuchitsu K, Tsuzuki M, Miyachi S (1991) Polypeptide composi-tion and enzyme activities of the pyrenoid and its regulation byCO2 concentration in unicellular green algae. Can J Bot 69:1062±1069
Laemmli UK (1970) Cleavage of structural proteins duringassembly of the head of bacteriophage T4. Nature 227: 680±685
Manuel LJ, Moroney JV (1988) Inorganic carbon accumulation inChlamydomonas reinhardtii: new proteins are made duringadaptation to low CO2. Plant Physiol 88: 491±496
McKay RML, Gibbs SP (1991) Composition and function ofpyrenoids: cytochemical and immunocytochemical approaches.Can J Bot 69: 1040±1052
Miyachi S, Tsuzuki M, Avramova ST (1983) Utilization modes ofinorganic carbon for photosynthesis in various species ofChlorella. Plant Cell Physiol 24: 64±68
Moroney JV, Mason CB (1991) The role of the chloroplast ininorganic carbon acquisition by Chlamydomonas reinhardtii.Can J Bot 69: 1017±1024
Moroney JV, Togasaki RK, Husic DH, Tolbert NE (1987)Evidence that an internal carbonic anhydrase is present in 5%CO2-grown and air-grown Chlamydomonas. Plant Physiol 84:757±761
Price GD, Coleman JR, Badger MR (1992) Association of carbonicanhydrase activity with carboxysomes isolated from the cyano-bacterium Synechococcus PCC7942. Plant Physiol 100: 784±793
Pronina NA, Semenenko VE (1984) Localization of membrane-bound and soluble carbonic anhydrase in the Chlorella cells.Fiziol Rast 31: 241±251
Pronina NA, Semenenko VE (1992) Role of the pyrenoid inconcentration, generation, and ®xation of CO2 in the chloro-plast of microalgae. Soviet Plant Physiol 39: 470±476
Pronina NA, Klyachko-Gurvich GL, Ladygin VG, Semenenko VE(1990) Activity of carbonic anhydrase in Chlamydomonasreinhardtii mutants with di�erent organization of chloroplastphotosynthetic systems. Fiziol Rast 37: 899
Ramazanov Z, Ca rdenas JT (1992) Inorganic carbon across cellcompartments of the halotolerant alga Dunaliella salina. PhysiolPlant 85: 121±128
Ramazanov Z, Rawat M, Henk C, Mason C, Mathews S, MoroneyJV (1994) The induction of the CO2-concentrating mechanismis correlated with the formation of the starch sheath around thepyrenoid of Chlamydomonas reinhardtii. Planta 195: 210±216
Ramazanov Z, Shiraiwa Y, Jime nez M, Rubio J (1995) E�ect ofexternal CO2 concentration on protein synthesis in the greenalgae Scenedesmus obliquus (Turp.) KuÈ tz and Chlorella vulgaris(Kosikov). Planta 197: 272±277
Rodrõ guez-Lo pez M (1964) In¯uence of the inoculum and themedium on the growth of Chlorella pyrenoidosa. Nature 203:666±667
Spalding MH, Je�rey M (1989) Membrane-associated polypeptidesinduced in Chlamydomonas by limiting CO2 concentrations.Plant Physiol 89: 133±137
Spalding MH, Spreitzer RJ, Ogren WL (1983) Carbonic anhy-drase-de®cient mutant of Chlamydomonas reinhardtii requireselevated carbon dioxide concentrations for photoautotrophicgrowth. Plant Physiol 73: 268±272
SuÈ ltemeyer DF, Fock HP, Canvin DT (1990) Mass spectrometricmeasurements of intracellular carbonic anhydrase activity inhigh and low Ci cells of Chlamydomonas. Plant Physiol 94:1250±1257
SuÈ ltemeyer DF, Schmidt C, Fock HP (1993) Carbonic anhydrase inhigher plants and aquatic microorganisms. Physiol Plant 88:179±190
SuÈ ltemeyer DF, Amoroso G, Fock HP (1995) Induction ofintracellular carbonic anhydrase during the adaptation to lowinorganic carbon concentrations in wild-type and ca-1 mutantcells of Chlamydomonas reinhardtii. Planta 196: 217±224
Villarejo A, Oru s MI, Martõ nez F (1997) Regulation of the CO2-concentrating mechanism in Chlorella vulgaris UAM 101 byglucose. Physiol Plant 99: 293±301
Wilbur KM, Anderson NG (1948) Electrometric and colorimetricdetermination of carbonic anhydrase. J Biol Chem 176: 147±154
A. Villarejo et al.: Pyrenoid-associated polypeptides in Chlorella 425




















