
Regulation of the CO2-concentrating mechanism in Chlorella vulgaris UAM 101 by glucose
10
PHYSIOLOGIA PLANTARUM 99: 293-301. 1997 Copyrigt, © PftvMotog,, pi,«ar,m mi Primed in Denmark - all rights reserved ISSN 003J-931? Regulation of the C02-concentrating mechanism in Chlorella vulgaris UAM 101 by glucose Arsenio Viliarejo, Maria Isabel Orus and Flor Martinez Vtllarejo, A., OrtJs, M. 1. and Martinez, F 1997. Regulation of the CO:<oncentrating mechanism in Chtorelta vutgaris UAM 101 by glucose. - Fhysiol. Plant. 99: 293-301. ln the green alga Chlorella vulgaris UAM 101, a CO2-eoncentrating mechanism is in- duced when the cells are growing under low CO; conditions. We have investigated the effect of glucose on the induction of this mechanism. Cells adapted to low CO2 in the presence of glucose showed a reduced ability to transpon and fbf extemal inorganic carbon. This reduction wa.s correlated with a decrease in internal carbonic anhydrase activity. 3-0-methyl-glucose, a nonmetabolizable analog of glucose, caused a more dramatic repression of these phenomena. Immunoblot analyses of total cell protein of Chtorelta vulgaris UAM 101 against large subunil of ribulose-1.5-bisphosphate car- boxylase/oxygenase and ribulose 1.5-bisphosphaie-carboxylase/oxygenase activase polyclonal antibodies showed that the expression of these two poly^reptides was af- fected by neither CO^ level, nor glucose or 3-0-raethyl-glucose. Ultrastructure snidies showed that the low COj-induced development of the pyrenoid was aiso affected by glucose. Immunocylochemical data demonstrated that ribulose-1.5-bisphosphale car- boxylase/oxygenase was exclusively located in the pyrenoid matrix. This localization and the density of labeling of the pwenoid region were affected by neither CO2 level nor the presence of glucose. Key words - Carbonic anhydrase. Chlorella. CO;-concentrating mechanism, glucose, pyrenoid, Rubisco. A. Viliarejo (corresponding author [email protected]) et at.. Dept de Biologia. Facuttad de Ciencias. Univ. Autonoma de Madrid. E-28049 Madrid. Spain. lntrod ti bonic anhydrase (CA) is the only low CO2-inducible protein that has been extensively characterized. In addi- Green algae can grow photoautotrophically at very low tion, several lines of evidence suggest the occurrence of CO2 concentrations due to the induction of the CO2-con- intracellular CA(s), which might be components of the centrating mechanism (CCM) (see Coleman 1991, Ka- CCM (Pronina and Semenenko 1984, Husic et al. 1989, plan et al. 1991, Badger and Price 1992). When cells are Sultemeyer et ai. 1990). transferred fixim elevated COj (> 1% [v/v] in air; high in addition, it has been suggested that pyrenoid, a COi-grown cells) to low CO2 concentrations (0.03% large protein complex located in the chloroplast, is an- CO2 in air; low CO2-grown cells) various cellular func- other component of the CCM in unicellular green algae tions change. Low CO2-grown cells exhibit a very high (Kuchitsu et al. 1991, Ramazanov et al. 1994). Pyrenoid photosynthetic affinity for external inorganic carbon morphology undergoes rapid and dramatic changes in [Ko5(Ci)], extremely low CO2 compensation point and response to variations in the CO2 concentration in the high ability to concentrate Ci internally (Badger et al. environment (Miyachi et al. 1986, Kuchitsu et al. 1988. 1980). It has been shown that several polypeptides are Ramazanov et al. 1994). The sheath of starch around the induced while green algae cells increase their affinity for pyrenoid is formed rapidly in response to a decrease in extemal C, (Spalding and Jeffrey 1989, Spalding et aJ. the CO2 concentration in the environment (Miyachi et al. 1991, Ramazanov et al. 1995a,b). The periplasmic car- 1986, Kuchitsu et al. 1988, 1991). The change in Received 7 August, 1996 Physiol. Planl. 99, 1997 293
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Regulation of the CO2-concentrating mechanism in Chlorella vulgaris UAM 101 by glucose

PHYSIOLOGIA PLANTARUM 99: 293-301. 1997 Copyrigt, © PftvMotog,, pi,«ar,m miPrimed in Denmark - all rights reserved ISSN 003J-931?
Regulation of the C02-concentrating mechanism inChlorella vulgaris UAM 101 by glucose
Arsenio Viliarejo, Maria Isabel Orus and Flor Martinez
Vtllarejo, A., OrtJs, M. 1. and Martinez, F 1997. Regulation of the CO:<oncentratingmechanism in Chtorelta vutgaris UAM 101 by glucose. - Fhysiol. Plant. 99: 293-301.
ln the green alga Chlorella vulgaris UAM 101, a CO2-eoncentrating mechanism is in-duced when the cells are growing under low CO; conditions. We have investigated theeffect of glucose on the induction of this mechanism. Cells adapted to low CO2 in thepresence of glucose showed a reduced ability to transpon and fbf extemal inorganiccarbon. This reduction wa.s correlated with a decrease in internal carbonic anhydraseactivity. 3-0-methyl-glucose, a nonmetabolizable analog of glucose, caused a moredramatic repression of these phenomena. Immunoblot analyses of total cell protein ofChtorelta vulgaris UAM 101 against large subunil of ribulose-1.5-bisphosphate car-boxylase/oxygenase and ribulose 1.5-bisphosphaie-carboxylase/oxygenase activasepolyclonal antibodies showed that the expression of these two poly^reptides was af-fected by neither CO^ level, nor glucose or 3-0-raethyl-glucose. Ultrastructure snidiesshowed that the low COj-induced development of the pyrenoid was aiso affected byglucose. Immunocylochemical data demonstrated that ribulose-1.5-bisphosphale car-boxylase/oxygenase was exclusively located in the pyrenoid matrix. This localizationand the density of labeling of the pwenoid region were affected by neither CO2 levelnor the presence of glucose.
Key words - Carbonic anhydrase. Chlorella. CO;-concentrating mechanism, glucose,pyrenoid, Rubisco.
A. Viliarejo (corresponding author [email protected]) et at.. Dept de Biologia.Facuttad de Ciencias. Univ. Autonoma de Madrid. E-28049 Madrid. Spain.
lntrod ti bonic anhydrase (CA) is the only low CO2-inducibleprotein that has been extensively characterized. In addi-
Green algae can grow photoautotrophically at very low tion, several lines of evidence suggest the occurrence ofCO2 concentrations due to the induction of the CO2-con- intracellular CA(s), which might be components of thecentrating mechanism (CCM) (see Coleman 1991, Ka- CCM (Pronina and Semenenko 1984, Husic et al. 1989,plan et al. 1991, Badger and Price 1992). When cells are Sultemeyer et ai. 1990).transferred fixim elevated COj (> 1% [v/v] in air; high in addition, it has been suggested that pyrenoid, aCOi-grown cells) to low CO2 concentrations (0.03% large protein complex located in the chloroplast, is an-CO2 in air; low CO2-grown cells) various cellular func- other component of the CCM in unicellular green algaetions change. Low CO2-grown cells exhibit a very high (Kuchitsu et al. 1991, Ramazanov et al. 1994). Pyrenoidphotosynthetic affinity for external inorganic carbon morphology undergoes rapid and dramatic changes in[Ko5(Ci)], extremely low CO2 compensation point and response to variations in the CO2 concentration in thehigh ability to concentrate Ci internally (Badger et al. environment (Miyachi et al. 1986, Kuchitsu et al. 1988.1980). It has been shown that several polypeptides are Ramazanov et al. 1994). The sheath of starch around theinduced while green algae cells increase their affinity for pyrenoid is formed rapidly in response to a decrease inextemal C, (Spalding and Jeffrey 1989, Spalding et aJ. the CO2 concentration in the environment (Miyachi et al.1991, Ramazanov et al. 1995a,b). The periplasmic car- 1986, Kuchitsu et al. 1988, 1991). The change in
Received 7 August, 1996
Physiol. Planl. 99, 1997 293

pyrenoid morphology is correlated with the redistribu-tion of ribulose-1,5-bisphosphate carboxylase/oxygen-ase (Rubisco), the main component of the pyrenoid(Mckay and Gibbs 1991). Rubisco is more concentratedin the pyrenoid under low CO2 conditions (Kuchitsu etal. 1991). Although these low CO2-inducible phenomenahave been investigated extensively, their functions re-main unclear.
The signal triggering the process of adaptation to lowCO2 conditions has not been characterized. It has beensuggested that the signal could be a detected change inphotosynthetic capacity or metabolite levels (Coleman1991). In earlier studies, algal cultures adapted to lowCO2 conditions in the presence of glucose and/or otherorganic compounds showed a decline in photosyntheticcapacity (Moroney et al. 1987a, Martinez and Onis1991, Shiraiwa and Umino 1991, Umino and Shiraiwa1991). This decline was correlated with a decrease inCA activity and an increase in respiration rates (Uttiinoand Shiraiwa 1991). Recently, Fen and Coleman (1994)reported the suppression by acetate of peripiasmic CAexpression in Chiamydomonas reinhardtii. However, thefactors involved in the suppression by organic com-pounds of the adaptation to low CO2 conditions have notbeen established yet.
In the present stttdy, we investigated the effect of glu-cose on the induction of CCM in Chloretla vutgaris. Wedetermined the rates of C, transport and fixation, CA ac-tivity, protein abundance, and ultrastructure changes incells adapted to low CO2 in the presence of glucose. Weprovide evidence that glucose suppresses the adaptationto low CO2 conditiotts by inhibiting intracellular CA ac-tivity and the development of pyrenoid without affectingRubisco and Rubisco activase levels. Ultrastructure anal-yses provide evidence that Rubisco is exclusively locatedin the pyrenoid of this alga and this localization is af-fected neither by CO2 levels nor the presence of glucose.
Abbreviations — CA, carbonic anhydrase; Ci, inorganic carbon(CO2 + HCOJ); CCM, C02-concentrating mechanism; K,,,(C,),the Ci concentration at which (he rate of COi-dependenl photo-synthetic O2 evoluticm is half-maximal; 3-OMG, 3-0-methyl-glucose; Rubisco, ribulose-l,5-bisphosphate carboxylase/oxy-genase.
Materials and methodsOrganism and culture conditions
The strain UAM 101 oi Chlorella vuigaris (strain SAG9.88 of the Gottingen University Culture Collection),isolated from the effluent of the sugar refinery in Aran-juez (Madrid), was grown in Rodriguez-Lopez A me-dium (Rodriguez-Lopez 1964), in 0.5-1 glass bottles illu-minated with 200 |imol m" s"' continuous light obtainedfrom fluorescent tubes (TDL 150 W; Philips, The Neth-erlands). Fresh cultures were inoculated from algaeaxenically grown on agar slants. Media were bubbledwith air supplemented with 5% (v/v) CO2 at 28°C. Whenlow CO2-grown, glucose/Sow CO2-grown, and 3-OMG/
low CO2-grown cells were to be used, the culture wastransferred from high CO2 conditions to low CO2, lowCO2 plus 28 mM glucose, or low CO2 plus 150 mM 3-OMG for 12 h, respectively.
Measurement of photosynthetic O; evolution
Cells were harvested by centrifugation at 7 500 g for 10min. washed twice in 25 mM CO2-free MES-NaOHbuffer (pH 5.7) and resuspended in the same buffer to achlorophyll density of 2—5 (tg mr'. The cell suspensionswere placed in an illuminated 02-electrode chamber(Hansatech Ltd, Norfolk, UK) and allowed to consumethe dissolved C, of the medium and the intracellular C,pool unti! no net photosynthesis was observed. Bicar-bonate at the indicated concentrations was added and therate of O evolution measured.
Inorganic carhon uptakeC| uptake was determined using the silicone oil centrifu-gation method (Badger et al. 1980). Short- and long-term uptake experiments were carried out.
Short-term assays were performed in the light at 25°Cin microfuge tubes in an Eppendorf centrifuge 5415 C(Eppendorf. Germany). The tubes contained (from bottomto top); 50 Ml 3 M NaOH. 125 fil silicone oil (2;1 [v/vl ofDibutil phthalate and i'i.s[2-ethylhexyl]phthalate. respec-tively; Ruka AG. Buchs. Switzerland), and 200 jt! of algalsuspensions, prepared as mentioned for O2 evolution as-says. Incubations were started by the addition ofNaH ''CO, (2.1 TBq mol') and allowed to proceed for theindicated times. The reaction was stopped by centrifuga-tion for 20 s. Internal C, was calculated from the differ-ence between the total and the acid-stable '''C in the pellet(Viliarejo et al. 1995). The intracellular volume was cal-culated using ["Osorbitol and 'H2O (Werdan et al. J972).
For long-term assays, two different cell suspensionswere used. One was incubated in an illuminated 02-elec-trode chamber (Hansatech Ltd, Norfolk, UK) and al-lowed to consume the dissolved C, of the medium andthe intracellular Q pool until no net photosynthesis wasobserved. Another one was directly used for determiningCi uptake. Both aliquots were incubated in glass vesselswith rubber stoppers in the presence of 10 mAf NaHCO;!(0.2 mM NaH"'CO3 [2.1 TBq mol"'] + 9.8 mMNaHCOi). Where indicated, 2 mA/ glucose was added tothe assays. Cell suspensions were incubated in the light(300 pmol m" s"') at 30°C with shaking for 2.5 h. Ali-quots (200 fil) were taken every 30 min from 0 to 2.5 h.'•"C-acid stable and '''C-acid-labile fractions were deter-mined by using the silicone oil centrifugation techniqueas mentioned above.
Determination ofCA activity
Algae were harvested by centrifugation at 7 500 g for 10min and the pellet was resuspended in ice-cold CA
294 Physiol. PUnt. 99,1997

buffer (75 mM phosphate buffer, pH 8.1, containing 1mM EDTA and 0.1 mM DTT), to a chlorophyll concen-tration of 100 ng ml ' . The measurements were per-formed in intact cells (periplasmic CA) and cell-free ex-tracts (total CA activity). Algal cells were disrupted withan ice-cold French pressure cell (two times, 200 MPa).The cell-free extracts were obtained by centrifugation ofthe homogenate at 16 000 g for 10 min.
CA (EC 4.2.1.1) activity was determined potentiomet-rically in CA buffer by determining the time taken forthe pH to drop from 8.2 to 7.4 at 2°C in a 2-ml sample.The reaction was started by rapid injection of 2 ml ofice-cold CO2-saturated distilled H2O (Ramazanov andCardenas 1992). One unit of CA activity (Wilbur Ander-son, WA, unit) is defined as (tJt^-l)xlO, where l^ and twere the times for the pH change to occur in the nonen-zymatic (buffer) and the enzymatic reactions, respec-tively.
caso et al. (1995). In control experiments, anti-Rubiscoantiserum was replaced with rabbit nonitnmune igG(Sigma), or the antibody was omitted altogether. Thegrids were examined and photographed in a JEOL 1200EX transmission electron microscope. The density of la-beling of each cell section was obtained by determiningthe number of gold particles per pm^ of pyrenoid sec-tioned. Area determinations and gold particle countswere made using a semiautomatic image analyzer (MOPVideoplan, Kontron-Bildanalyse, Munich, Germany).Only micrographs in which pyrenoid was cut in media!section were used for this study.
Other methods
Chlorophyll concentrations were determined spectro-photometricaliy (Holden 1965). Proteins were estimatedby the method of Bradford (1976).
SDS-PAGE and western blot analysisSDS-PAGE was performed with H^/c (w/v) acrylamideconcentration (Laemmli 1970). The immunoblot assaywas performed according to the protocol from Bio-RadLaboratories except that 5% non-fat dry milk was usedto block nitrocellulose. Goat anti-rabbit IgG(H-i-L)horseradish peroxidase conjugate and HRP colour devel-opment reagent was purchased from Bio-Rad Laborato-ries.
Polyclonal antibodies raised against a 37-kDa peri-plasmic CA of Chlamydomonas reinhardlii were kindlyprovided by Dr J. Moroney (Louisiana State University,LA, USA). Polyclonal antibodies raised against the largesubunit of Rubisco of Eugtena gracilis were kindly pro-vided by Prof. C. Ascaso (Consejo Superior de Investi-gaciones Cientificas, Spain). Polyclonal antibodiesraised against Rubisco activase were kindly provided byProf. Z. Ramazanov (Inst. of Applied Algology, Spain).
Electron microscopyFor transmission electron microscopy, cells were fixedin 2.5% (w/v) glutaraldehyde added to 0.05 M cacody-late buffer (pH 7.4) for 2.5 h at 4°C. Samples wererinsed in the same buffer. After rinsing, the sampleswere dehydrated in ethanol, then infiltrated and embed-ded in LR White medium resin (London Resin Co.,Woking, Surrey, UK) (Ramazanov et al. 1994). Thin sec-tions were poststained with uranyl acetate and lead cit-rate (Reynolds 1963), ohserved and photographed in aJEOL 1200 EX (Tokyo, Japan) transmission electron mi-croscope.
Immunogold laheling
Immunolabeling was carried out with anti-Rubisco anti-serum of Euglena gracilis. Immunogold labeling with10-nm gold-protein A was performed according to As-
ResultsEfTect of glucose on photosynthetic O evolution
When high CO2-grown cells of Chloretla vutgaris UAM101 were transferred to low CO2 conditions, the cells'apparent affinity for Ci increased (Tab. 1). This increasewas partially suppressed when the adaptation to low CO2occurred in the presence of glucose or 3-OMG, a non-metabolizable analog of glucose (Tab. 1). The presenceof glucose or 3-OMG during the adaptation to low CO2also reduced the maximal rates of CO2-dependent O2evolution when compared to autotrophic control cells(Fig. 1). 3-OMG caused a more pronounced inhibition ofmaximal O2 evolution rates than glucose did (Fig. 1).
EfTect of glucose on C uptake
When high CO2-grown cells of Chlorelta vulgaris UAM101 were transferred to air, the efficiency in fixing '''CO2improved as a consequence of the high ability to accumu-late and transport '''C, (Fig. 2). After 2 min of incubation,the rate of photosynthetic "CO2 fixation (acid-stable '''Cfraction) in high CO2-grown cells (140 nmoi mg~' Chi)was 50% lower than in low CO2-grown cells (292 nmolmg ' Chi) (Fig. 2B).
The presence of glucose during the adaptation to lowCO2 partially suppressed the increase in the ability totransport and fix "C, (Fig. 2A,B). However, the internalCi pool in these cells reached values i- to 2-fold higher
Tab. 1. Ko5(CJ in Chloretla vulgaris UAM 101 cells growing underdifferent conditions. The procedures were as described in the leg-end to Fig. 1. Data are means ± SD of 5 experiments.
Growth conditions
High COi-grownLow COi-grownGlucose^ow CO2-g3-OMG/low CO2-grown
40 ±2.110 ± 0.530 ±1.430 ±2.4
Physiol. Planl. 99. 1997 295

700
600 'r
rt
300 H
NaHCOg (mM|
Fig. i. The rate of photosynthesis versus inorganic carbon inChlorella vulgaris U.AM 101 cells grown under high COi con-centrations (A) or adapted lo low CO. for 12 h in the presence ofeither no organic carbon sources (•) . 28 mM glucose (#) or 150mM 3-OMG (H). The chlorophyll content was 2—5 |ig ml ' andthe pH was 5.7. Data are means of 5 experiments.
than in IOW-CO2 cells grown withoul organic carbonsource (Fig. 2C).
When 3-OMG was present duting the adaptation tolow CO2. the suppression in the ability to transport andfix C, was more dramatic than in glucose/iow CO2-grown cells. The rates of '•'CO2 fixation in these cells
were much lower than those of glucose/iow CO^-growncells whereas the C, accumulation was only 2.5-foSdlower (Fig. 2B,C). These results agree with those ob-tained in O2 evolution assays.
The glucose-induced decrease in the ability to fix C,was also observed in long-term experiments. Figure 3shows the effect of glucose on the lime course of '*CO;fixation rates in low CO2- and glucose/low CO2-growncells. Incubation with 2 mM glucose caused a decrease in'''CO2 fixation rates of both low CO2- and glucose/lowC02-adapted cells when compai'ed to tlie rates in its ab-sence. This glucose-induced negative effect on '"'CO2 fix-ation appeared immediately after the addition of glucosein glucose/low CO^-adapted ceils, while it appeared about1 h after the addition in low CO2-adapted cells, in whichthe gltjcose uptake system was not previously induced(Fig. 3). On the other hand, glucose/low CO^-adaptedceils incubated without glucose showed a recovery of"CO; fixalion that was faster when tliese cells were nolallowed to consume the internal C, pool (Fig. 38).
EfTect of glucose on C.4 activity
C.A. activity was measured in C. vulgaris UAM 101cells, before and after cellular breakage lo determine Iheactivity of peripiasmic and intraceilular C.^, respec-tively. No CA activity was detected in the suspension ofintact cells (data not shown). Immunoblot analyses usingantibodies raised againsi 37 kDa periplasmic CA ofChlamydomonas reinhardtii show that C. vutgaris UAM
Time (s) Time (s)
Time (s)
Fig. 2. '^d transport i.\).•'C:O, fixalion (B) and C,
accumulation (C) in Cvulgaris VAM 101 cellsgrown tinder high CO;conditions (A), adapted 10low CO; for 12 h in thepresence of either no organiccarbon sotirces (T). 28 mMglucose (•) or 150 mM 3-OMG (B). C, Iraiisport and'"CO2 fixation rales, andiniracellular C, weredetermined foilowint: theaddition of 100 uM NaH"'COjto cells. In gitjcose/low CO^-and 3-OIVIG/iow CO^-growncells, organic compoundswere removed before Iheuptake experiments. Thechlorophyll content was 25 pgml^ and ihepH wa.s 5.7. Dataare means of 5 experiments.
296 Physiol. Ptam. 99, t997
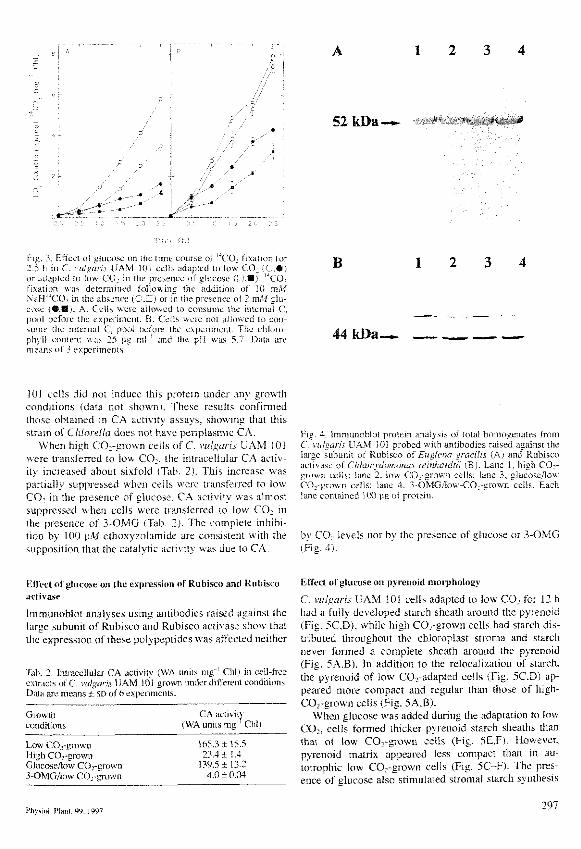
52 kDa.
Fig. 3,1'ttect t»t iiluci'sc on the titne coui>e ot '"CO^ hxjtton lor2..S h in f" xub^aii'. L AM Itil cclK .idaptcd tii l.n\' CO. i : • )or adapted tn k™ CO in thi. pit-scnce ot gluccise C H i "CO^fixalion wa.. dctennitied tollovstng the addilson t5! lu mUNaH^'^CO. in thj absence f' .^ * o\ \\\ the presence ot 2 tn V glu-cose i®.B( \ . CelK "."vCie allowed to consume the mlcmal Cpraii before the experiment B. CclK «ere not alio«i-d to LOti-sume tliL- intertia! C pt)t)l bci<tEC the cxpcnnicnt The chloro-phyll ctsntent v\as 25 pg ml and the pH %\a ^~ Data arcmrans ot 3
B
44 kDa—
101 cells did not induce this protein under any growthconditions (data not shown). These results confirmedthose obtained in C\. activity assays, showing that thisstrain of Chiorelia does not have periplasraic C.4.
When high COrgrown cells ofC. vulgaris UAM 101were transferred to low CO . the intraceliular C.4 activ-ity increased about sixfold (Tab. 2). This increase waspartially suppressed when cells were iransfetred to lowCOi in the presence of glucose. CA activity was almostsuppressed when cells were transferred to lovv CO; inthe presence of 3-OMG (Tab. 2). The cotnplete inliibi-lion by 100 uM elhoxyzolamide are consistenl with thesupposition that the catalytic activity was due to CA.
Fig. 4. lmmunoblo! protein analysis of total homogenates fromC. vulgaris UAM 101 probed with antibodies raised against theliirge subunil of Rubisco of Euglena gracilis {.\) and Rubiscoaclivase of Chlamvdomonas reinhardtii (B). Lane 1. high CO;-grown cells; lane 2. low COj-grown cells; lane 3, glucose/lowCO;-grown cells; lane 4. 3-OMG/low-CO;-grovvn cells. Eachlane eomaiiied UXI pg of protein.
by CO2 levels nor by the presence of glucose or 3-OMG(Fig. 4).
Effect of glucose on the expression of Ru bisco and Rubiscoattivase
inimunoblol analyses using antibodies raised againsl thelarge subunit of Rubisco and Rubisco activase show thatthe expression of these palypeptides was affected neither
Tab. 2. Intracellular CA activity (Vi'A utiits mg ' Ctil) in cell-freeeslracls of C. vulgaris U.Wi lS)'l grov^ii under different conditions.Data ai'e means ± .S0 of 6 experiments.
Growthcotiditions
C.A aclivitv(WA units m g ' Chi)
Low CO;-grownHigh COrgrownGlucose/low CO-grown3-OMG/low COrgrown
165.3 ±15.523.4 ± 1.4
139.5 ±13.24.0±0.fM
Effect of glucose on pyrenoid rjiorphology
C. vulgarts UAM 101 cells adapted to low CO, for 12 hhad a fully developed starch sheath around the pyrenoid(Fig. 5C.D), while high CO;-grovvn cells had starch dis-tributed throughout the chloroplast stroma and starciinever fonned a complete sheath around the pyrenoid(Fig. 5A.B). In addition to the relocalization of starch,the pyrenoid of lovv CO;-adapted cells (Fig. 5C.D) ap-peared more compact and regular than those of high-CO,-grown cells (Fig. 5A,B).
When glucose was added during tlie adaptation to lowCO.. cells forined thicker pyrenoid starch sheaths thanthat of low CO:-gTOwn cells (Fig. 5E,F). However,pyrenoid matrix appeared less compact than in ao-totrophic !ow CO -grown cells (Fig. 5C-F). The pres-ence of glucose also stimulated stromal starch synthesis
Plivsbl. Plant. 99, 199?297
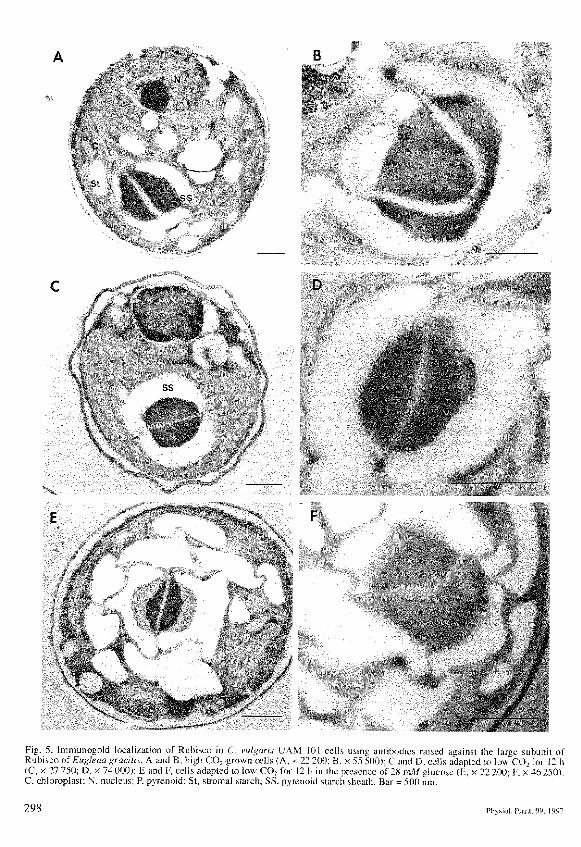
B• * • . • • * . . . 1
\ P
v:... ' • ' ' . ;
Fig. 5. Immunogold localization of Rubisco in C. vuigaris I'AM 101 cells usmp antibodies rjised agatnst the larsze subunit otRubisco of Euglena gracilis. A and B, high COrgrown cells (A. x 22 2fKl; B. y 55 500); C and D. cells adapted to low'cO.. fot 12 h(C, X 27 750; D, x 74 000); E and F, ceils adapted to iow CO; lot 12 h in the presctice of 2H ml/glucose (f . x 22 200. I-, ;< 4h25QlC, chloroplast; N, nucleus; P, p>Tenoid; St. stromal starch; SS. pyrenoid starch sheath. Bat = 500 nm.
298 Phvsiol. I'lant. 99. 1997

Tab. 3. Density of immunogold labeling over pyrenoid in C. vulgar-is LLAM HI! cells after 12 h of growtli under different conditions.Data are means ± Sl). n. Number of cell sections analvzed.
Growlh conditions Gold particles pm "
Low CO-grownHigh CO;-grownGlucose./low CO
37.5 ± 8.437.5 ± 7.438.0 ± B.5
131019
(Fig. 5E.F). 3-OMG caused a similar effeci on pyrenoidmorphology than that caused by glucose (daia tiolshown).
Effect of glucose «n Rubisco localization
Figure 5 also shows the labeling by antibodies againstthe large subutiit of Rubisco in C. vulgaris cells. Thepyrenoid region of the chloropiasl is heavily labeled inihe three culture conditions, whereas ihe rest of the cellwas completely iinlabeied (Fig. 5). Table 3 compares thedensity of labeling of the pyrenoid region. There was nota statistically significanl difference (P< O.l.H) in the la-beling of pyt'etioid region between high and low COi-grovvn cells. The presence of glucose during the adapta-lion 10 low CO; did not affed pyrenoid labeling either(P < 0.01) (Tab. 3). ln conlrol experiments, with notiim-mune antiserum. no labeling was observed (data notshown).
Discussion
The ynicclltjlar green alga Chlorella vulgaris UAM 101.like many olher unicellular algae, can grow photoau-totrophicaliy on very low levels of CO; due lo the induc-tion of the CCM. When high COrgrown cells of C. r«/-gari.s UAM 101 were switched lo low CO.. the cells' ap-parent affinity for C, increased from 40 lo lOpA/iFig. 1.Tab. 1). .An increase in the ability lo traospon. accumu-late and fix C, was also observed (Fig. 2). This increaseill pholos)THhetic capacity was correlaled with the in-duclion of CA activity (Tab. 2l. Our data indicate thaithis increased activity was only due to ititetnal CA formsbecause this strain of C. vulgaris does not have periplas-mic CA.
The induction of alt these phenotnena was correlatedwith changes in pyrenoid tnorphoiogy and the formationof pyrenoid starch shealh as il has been already reportedm Chloretla and other green algae (Miyachi et al. 1986.Kuchitsu et al. 1991. Ramazanov el a!. 1994). In contrastto immunocylochemical data .showing redistribulion ofRubisco during the adaptation lo low CO; (McKay atidGibbs 1991), our data clearly show that in C. vulgarisUAM 101 Rubisco was exclusively located in thepyrenoid in either high or low C02-grown cells (Fig..5A-D). Although pyrenoid appeared lo be more com-pact in low CO2-grown cells, ihe density of labeling ofgold particles was similar under both conditions ("Rib.
3). These results agree with those of the immunoblotanalyses (Fig. 4A) showing that the expression ofRubisco was not affected by CO; levels. In addition, theexpression of Rubisco aclivase, which has been reportedto be located to pyrenoid (McKay et a l 1991), was notaffected by CO; levels either (Fig. 4B).
h has been previously reported that the presence oforganic C sources pattially suppresses the induction ofCCM (Moroney et ai. 1987a. Martinez and OrtJS 1991.Shiraiwa and Umitio 1991, Umino and Shiraiwa 1991.Fell and Coieman 1994). C. vulgaris UAM 101 cellsadapted to low-CO; conditions in the presence of glu-cose showed a reduced COi-dependent photosyntheticcapacity when compared with lovv COi-grown cells (Fig.1, Tab. I). This decline in photosynthetic capacity re-flected the lower ability to transport and fix C. of thesecells (Figs 2 and 3). The presence of 3-OMG, a nonme-tabolizable analog of glucose, during the adaptation tolow CO; resulted in a more dramatic repression than thatcaused by glucose. 3-0MG/1DVV CO;-adapted cellsshowed a very low photosynthetic capacity (Fig. 1) anda tnuch lower ability to tran.spon and fix C when com-pared io glucose/low CO2-adapted cells (Fig. 2). Theability to transport and fix C, strongly reflected the levelof induction of internal CA activit)' (Tab. 2). 3-OMG/low CO;-grovvo cells showing the lowest ability to trans-poti and fix C,. also showed the lowest internal CA ac-ijvity. it has been suggested thai the negative effect oforganic C sources on the ioduction of CCM could be dueto the increased intracellular amoitnts of respiratory CO;ihal would mimic high CO; conditions (Felt and Cole-tiian 1994. Viliarejo et al. 1995). The results obtainedwith the nonmetabolizable analog suggest a direct roleof the organic cotnpound itself on the suppression of theaclivilies involved in CO; supply and ftxation.
The effect of glucose oti the time-course of '"CO; fix-ation depended on whether cells were allowed to con-sume the internal C, pool before the experiment (Fig.3.4) or not (Fig. 3B). The negative effect of glucose onC, fixation appeared as soon as the glucose transpon sys-tem was induced in autolrophic low CO;-grown cellsand was more pronounced in cells previously deprivedof endogenous metabolites. The removal of glucose ing!ucose/low-CO;-grown cells caused a faster and moreefftcietit recovery of Ci fixation values in those cells thatwere not deprived of endogenous metabolites. This factis consistent with the hypothesis thai glucose and notCO; or other end products of its metabolism is responsi-ble for the suppression of the mechanism responsible forthe acquisition of CO;. These data indicate that glucosepartially repressed this mechanism but does not impairthe photosynthefic capacity of these cells.
Since glucose and 3-OMG had no effect on the ex-pression of either Rubisco or Rubisco activase (Fig. 4).we reasoned that the effect of these compounds as inhib-ilors of the ability to transport and fix external C, couldbe a result of glucose or 3-OMG inhibiting the C, trans-pot1 systetn and/or the supply of CO; to the active site of
L Ptiim.299

Rtibisco. It is well established that CA plays an impor-tant role on the efficient fixation of COi during photo-synthesis (Moroney et al. 1985, 1987b, Badger and Price1994). Otir data demonstrate that the presence of glucoseor 3-OMG during adaptation to low CO7 resulted in a re-pression of the itiduction of internal CA activity. This re-pression of itttemal CA activity induced by organic Csources would result in a lower net uptake of C,. In addi-tion, a negative effect of glucose or 3-OMG on the C,transport system can not be ruled out.
The negative effect of organic C sources on the netuptake of C, could also be due to an alteration of eitherpyrenoid morphology or Rubisco distribution. Our datademonstrated that glucose had no effect on Rubisco dis-tribution. This protein was determined to be exclusivelylocated in the pyrenoid matrix (Fig, 5E,F) and with thesame density of labeling as autotrophic cells (Tab. 3). Incontrast to previous reports showing dispersion ofRubisco throughout the stroma in Chlamydomonas cellsadapted to low COi in the presence of organic C sources(Kiss et al. 1986), we have demonstrated that Rubisco islocated in the pyrenoid and this location was affectedneither by CO2 levels nor the presence of glucose. Ultra-structure atialyses showed that the only glucose-inducedalteration was observed at the level of both pyrenoidmorphology and starch distribution. Both stromal andpyrenoid starch showed a great increase in glucose/lowCOi-grown cells, probably due to glucose being accu-mulated as starch granules (Fig. 5E,F). However,pyrenoid starch sheath did not show the organized ap-pearance typical of autotrophic low COi-grown cells(Fig. 5E,F).
It has been previously reported that pyrenoid mayserve as the active site of CO2 ftxation (Ramazanov et al.1994). The alteration of the pyrenoid matrix caused hythe presence of glucose during the adaptation to low CO,conditions could account for the lower net uptake of C;showed by these cells.
In conclusion, our data indicate that the presence oforganic C sources repressed the induction of CCM inChioretla vulgaris UAM 101. A more dramatic repres-sion was shown in the presence of the nonmetabolizableanalog of glucose, suggesting a direct role of the organiccompound itself on the repression of CCM. Organic Csources suppress the induction of CCM by both inhibit-ing intracellular CA and altering pyrenoid morphologywithout affecting the expression of Rubisco and Rubiscoactivase. In addition, our data demonstrated that in thisalga Rubisco was exclusively located to pyrenoid andthis localization was affected neither by CO2 levels northe presence of glucose.
Acknowledgmenis - Authors thank Dr E. Femindez-Valiente forcritical but friendly revision of the manuscript and Prof. Z. Ra-mazanov for his suggestirais. This work was suj^orted by agrant of DGICYT (Grant no. PB93-0274), Spain, and a scholar-ship of Consejeria de Educacion y Ciencia, Comunidad Au-tonoma de Madrid, Spain, to A. V.
References
Ascaso, C, Valladares, F & De los Rios, A. 1995. New ultra-structure aspects of pyrenoids of the lichen photobiont Tre-botixia (Microthamniales, Chlorophyta). - J. Phycol 31;114-119.
Badger, M. R. & Price, G. D. 1992. The CO, concentratingmechanism in cyanobacteria and microaigae. - Physiol.Plant. 84; 606-615.
— & Price, G. D. 1994. The role of carbonic anhydrase in pho-tosynthesis. - Annu. Rev. Plant Physiol. Planl Mol. Biol. 45:369-392.
- . Kaplan, A. & Beny\ J. A. 1980. internal inorganic carbonpool of Chlamvdomonas reinhardlii. - Plant Physiol 66;407-413.
Bradford. M. M. 1976. A rapid and sensitive method for thequantitation of microgram quantities of protein utilizing theprinciple of protein-dye binding. - Anal. Biochem. 72; 24S-254.
Coleman, J. R. 1991. The molecular and biochemica! analysesof COrConcentrating mechanisms in cyanobacteria and mi-croaigae. - Planl Cell Environ. 14; 861-867.
Fett. J. P. & Coleman, J. R. 1994. Regulation of periplasmic car-bonic anhydrase expression in Chtamvdomonas reinhardtiiby acetate and pH. - Plant Physiol 106: 103-108.
Holden, M. 1965. Chlorophylls. -In Chemistry and Biochemis-try of Plant Pigments (f. W. Goodwin, ed.), pp. 3-28. .Aca-demic Press, New York. NY.
Husic, H. D.. Kitayama. M., Togasaki, R. K., Moroney. J. V.Morris. K. L. & Tolbert. N. E. 1989. Identification of intra-cellular carbonic anhydrase in Chtamydomonas reinhardtiiwhich is distinct from ihe periplasmic form of the enz> me. -Plant Physioi 89: 904-909.
Kaplan. A., Schwaiz. R., LiemanHurwitz, J. & Reinhold. L.1991. Physiological and molecular aspects of the inorganiccatbon-concenlrating mechanism in cyanobacleria. - PlanlPhysiol. 97; 851-855.
Kiss. J. Z.. Vasconcelos. A. C. & Triemer. R. E. 1986. Paramy-lon synthesis and chloroplast stnicture associated with nutri-ent levels in Euglena (Euglenophyceae). - J. Phycol. 22;327-333.
Kuchitsu. K.. Tsuzuki, M. & Miyachi. S. 1988. Changes ofstarch localization within the chloropiasl induced bychanges in carbon dioxide concentration during growth ofChtamvdomonas reinhardtii. - Plant Cell Physiol. 29; 1269-1278. '
- , Tsuzuki, M. & Miyachi, S. 1991. Polypeptide compositionand enzyme activities of the pyrenoid and its regulation byCO2 concentration in unicellular green alga. - Can. J. Bot.69; 1062.
Laemmli, U. K. 1970. Cleavage of structural proteins during as-sembly of the head of bacteriophage T4. - Nature 227; 680-685.
Martinez, F. & Orus, M. I. 1991. Interactions between glucoseand inorganic carbon metabolism in Chtoretta vulgarisUAM 101.-Plant Physiol. 95; 1150-1155.
McKay, R. M.L.& Gibbs. S. P. 1991. Composition and functionof pyrenoids; Cytochemica! and immunocytochemical ap-proaches. - Can. J. Bol. 69; 1040-1052.
- , Gibbs, S. P. & Vaughn, K. C. 1991. Rubisco activase ispresent in the pyrenoid of green algae. - Protoplasma 162;38-45.
Miyachi, S., Tsuzuki, M., Maniyama, I., Gantar, M., Miyachi, S.& Matsushima, H. 1986. Effects of CO2 concentration dur-ing growth on the intracellular structure of Chtoretla andScCTe<fe5miu (Chlorophyta). - J. Phycol. 22; 313-319.
Moroney, J. V, Husic, H. D. & Tolbert, N. E. 1985. Effect ofcarbonic anhydrase inhibitors on inorganic carbon accumti-lation by Chtamydomonas reinhardtii - Plant Physiol. 79;177-183.
- , Kitayama, M., Togasaki, R. K. & Tolbert, N. E. 1987a. Ev-idence for inorganic carbon transport by intact chloroplast ofChlamydomona.i reinhardtii. - Plant Physiol. 83; 460-463.
300 Physiol. Plani. 99. 1997

- , Togasaki, R. K., Husic, H. D. & Tolbert, N. E. 1987b. Evi-dence that an internal carbonic anhydrase is present in 5%CO.-grown and air-grown Chtamydomonas - Plant Physiol84; 757-761.
Pronina, N. A. & Semenenko, V. E. 1984. Localization of mem-brane-bound and soluble forms of carbonic anhydrase in theChtoretta cell. - Fiziol. Rast. 31; 241-251,
Ramazanov, Z. & Cardenas, J. T. 1992. Inorganic carbon acrosscell compartments of tbe halotolerant alga Dunalietla sallna.- Physiol Plant. 85; 121-128.
- , Rawat, M., Henk, C, Mason C. Mathews, S. & Moroney,J. 1994. The induction of the COj-concentrating n:iechanismis correlated with the formation of the starch shealh aroundthe pyrenoid of Chlamydomonas reinhardtii. - Planta 195;210-216.
- , Sosa, P A., Henk, M. C, Jimenez del Rio, M.. Gomez-Pinchetti, J. L. & Garcia Reina. G. 1995a. Low-C02-induc-ible protein synthesis in the green alga Dunatietla terti-olecta. - Planta 195; 519-524.
- . Shiraiwa. Y, Jimenez del Rio, M. & Rubio, J. 1995b. Ef-fect of extemal CO, concentrations on prolein synthesis inthe green algae Scenedesmus obliquus (Turp.) Kutz andChlorella vulgaris (Kosikov). - Planla 197; 272-277.
Reynolds, S. 1963. The use of lead citrate at high pH as an efec-tron-opaque stain in electron microscopy. - J. Cell Biol. 17;200-211.
Rodriguez-Lopez, M. 1964. Influence of the inoculum and themedium on the growth of Chlorella pyrenoidosa. - Nature203; 666-667.
Shiraiwa, Y. & Umino, Y. 1991. Effect of glucose on the induc-tion of the carbonic anhydrase and the change in k,n(CO2) ofphotosynthesis in Chlorella vutgaris llh. - Plant C ell Phys-iol. 32; 311-314.
Spalding, M. H. &. Jeffrey, M. 1989. Membrme-associatedpolypeptides indueed in Chtamydomonas by limiting CO2concentrations. - Plant Physiol. 89; 133-137.
- , Winder, T. L., Anderson, J. C , Geraghty, A. M. & Marek,L. F. 1991. Changes in protein and gene expression duringinduction of the C02<oncentrating mechanism in wild-typeand mutant Chlamydomonas. - Can. J. Bot. 69; 1008-1016.
Sultemeyer. D. F., Fock, H. P & Canvin. D. T. 1990. Mass spec-trcmetric measurement of inlracellular carbonic anhydraseactivity in high and low Q cells of Chtamvdomonas. - PlantPhysiol. 94; 1250-1257.
Umino, Y. & Shiraiwa, Y. 1991. Effect of metabolites on car-bonic anhydrase induction in Chtoretta regularis. - J. PlantPhysiol 139; 41^14.
Viliarejo. A., Orus, M. I. & Martinez, F 1995. Coordination ofphotosynthetic and respiratory metabolism in Chlorella vul-garis UAM 101 in the light. - Physiol. Plant. 94; 680-^86.
Werfan, K.. Heldt, H. W. & Geller, G. 1972. Accumulation ofbicarbonate in intact chloroplasts following a pH gradient. -Biochim. Biophys. Acta 283; 430-441.
Edited by H. Senger
Physiol. Plant. 99. 1997301





















