
The structural molecular biology network of the State of São Paulo, Brazil

1 23
Plant Molecular BiologyAn International Journal on MolecularBiology, Molecular Genetics andBiochemistry ISSN 0167-4412 Plant Mol BiolDOI 10.1007/s11103-011-9814-9
Expression of lorelei-like genes inaposporous and sexual Paspalum notatumplants
Silvina Andrea Felitti, José GuillermoSeijo, Ana María González, MaricelPodio, Natalia Verónica Laspina, LorenaSiena, Juan Pablo Amelio Ortiz, et al.

1 23
Your article is protected by copyright and
all rights are held exclusively by Springer
Science+Business Media B.V.. This e-offprint
is for personal use only and shall not be self-
archived in electronic repositories. If you
wish to self-archive your work, please use the
accepted author’s version for posting to your
own website or your institution’s repository.
You may further deposit the accepted author’s
version on a funder’s repository at a funder’s
request, provided it is not made publicly
available until 12 months after publication.

Expression of lorelei-like genes in aposporous and sexualPaspalum notatum plants
Silvina Andrea Felitti • Jose Guillermo Seijo • Ana Marıa Gonzalez •
Maricel Podio • Natalia Veronica Laspina • Lorena Siena •
Juan Pablo Amelio Ortiz • Silvina Claudia Pessino
Received: 4 December 2010 / Accepted: 23 July 2011
� Springer Science+Business Media B.V. 2011
Abstract Gametophytic apomictic plants form non-
reduced embryo sacs that generate clonal embryos by
parthenogenesis, in the absence of both meiosis and egg-
cell fertilization. Here we report the sequence and
expression analysis of a lorelei-like Paspalum notatum
gene, n20gap-1, which encodes a GPI-anchored protein
previously associated with apomixis in this species. Phy-
logeny trees showed that n20gap-1 was evolutionary rela-
ted to the Arabidopsis thaliana lorelei genes At4g26466
and At5g56170. The lorelei At4g26466 disruption was
shown to be detrimental to sperm cell release in arabid-
opsis. RFLP (Restriction Fragment Length Polymorphism)
analysis revealed the occurrence of several homologous
sequences in the Paspalum notatum genome, exhibiting
polymorphisms genetically linked to apomixis. Real-time
PCR showed that lorelei-family genes present a minor
activity peak at pre-meiosis and a major one at anthesis.
The apomictic genotype analyzed showed a significantly
increased activity at pre-meiosis, post-meiosis and anthesis
with respect to a sexual genotype. In situ hybridization
assays revealed expression in integuments, nucellus and the
egg-cell apparatus. Several n20gap-1 alleles differing
mainly at the 30 UTR sequence were identified. Allele-
specific real-time PCR experiments showed that allele 28
was significantly induced in reproductive tissues of the
apomictic genotype with respect to the sexual genotype at
anthesis. Our results indicate that P. notatum lorelei-like
genes are differentially expressed in representative sexual
(Q4188) and apomictic (Q4117) genotypes, and might play
a role in the final stages of the apomixis developmental
cascade. However, the association of n20gap-1 expression
with the trait should be confirmed in significant number of
sexual and apomictic genotypes.
Keywords Apomixis � Apospory � Lorelei �Paspalum notatum � Plant reproduction
Introduction
Apomixis is an asexual mode of reproduction through
seeds described in more than 400 plant species belonging
to 35 different angiosperm families (Nogler 1984; Ozias-
Akins 2006). The trait is frequently associated with hybrid
origin and/or polyploidy (Richards 2003). Apomictic plants
produce seeds containing clonal embryos with a maternal
genetic constitution. It is highly unlikely that this trait
could have evolved from sexual ancestors through ran-
domly occurring mutations, because it is polyphyletic in
origin and involves the alteration of several independent
developmental steps. Instead, apomixis might have arisen
through deregulation of the sexual developmental pathway
by a mechanism that could comprise both genetic and
epigenetic components (Grossniklaus 2001). This coordi-
nated deregulation could be influenced by global regulatory
changes resulting from hybridization and/or polyploidy
(Grossniklaus 2001).
S. A. Felitti � M. Podio � N. V. Laspina � L. Siena �J. P. A. Ortiz � S. C. Pessino (&)
Laboratorio Central de Investigaciones, Facultad de Ciencias
Agrarias, Universidad Nacional de Rosario, Parque Villarino,
S2125ZAA Zavalla, Provincia de Santa Fe, Argentina
e-mail: [email protected]
J. G. Seijo � A. M. Gonzalez � M. Podio � J. P. A. Ortiz
Facultad de Ciencias Agrarias, Instituto de Botanica del
Nordeste (IBONE), CONICET, Universidad Nacional del
Nordeste, Sargento Cabral 2131, W3402BKG Corrientes,
Argentina
123
Plant Mol Biol
DOI 10.1007/s11103-011-9814-9
Author's personal copy

Due to its potential importance as an enabling technol-
ogy for agriculture, apomixis has been the subject of
exhaustive cytoembryological, cytogenetical and molecular
analyses into its developmental mechanisms (Ozias-Akins
2006). Exploitation of the trait in crop plants would pro-
vide major benefits to agriculture: widespread use and
fixation of hybrid vigor, survival and immediate fixation of
combined genetic resources, including wide-cross progeny
that are unfit when propagated sexually, true seed pro-
duction from crops currently propagated vegetatively and
accelerated breeding programs in response to changing
needs and environments (Spillane et al. 2004).
One of the most frequent apomictic mechanisms is
apospory, a developmental process characterized by the
differentiation of one or several nucellar cells, which ini-
tiate a series of mitotic divisions to generate megaga-
metophytes where all nuclei are non-reduced (2n) (Crane
2001). The resulting 2n egg cells form embryos by par-
thenogenesis. The endosperm can be generated either
autonomously or by pseudogamy (polar nuclei fertiliza-
tion), depending on the species (Crane 2001).
Paspalum notatum Flugge is a rhizomatous subtropical
perennial grass with a highly evolved reproductive strat-
egy, consisting of a combination of vegetative propagation,
sexuality and apomixis (Gates et al. 2004). The species is
organized as an agamic complex including sexual diploid
and aposporous pseudogamous tetraploid cytotypes (Qua-
rin 1992). Sexual tetraploid cytotypes were never collected
from nature, but some plants were artificially produced by
colchicine treatment of diploids and experimental crosses
(Quarin et al. 2003).
The molecular basis of apomixis in P. notatum has been
under examination for more than 10 years. Full genetic
maps were developed at both diploid (sexual) and tetra-
ploid (apomictic) levels (Ortiz et al. 2001; Stein et al.
2007). Fifteen molecular markers completely linked to a
single genomic region controlling apospory were identified
(Stein et al. 2007). The apospory-specific genomic region
(ASGR) showed severe suppression of recombination and
preferential chromosome pairing with one of the three
homologues of the set (Stein et al. 2007). Data derived
from mapping analysis indicated that the P. notatum
chromosomal segment governing apospory could encom-
pass approximately 36 Mbp and might have resulted from
an inversion or a translocation (Stein et al. 2004; Pupilli
et al. 2004; Stein et al. 2007). Similar structural features
(regarding lack of recombination in an extended sequence
and possible occurrence of rearrangements) were reported
for the apomixis-controlling genomic regions of other
aposporous species like Pennisetum ciliare, Pennisetum
squamulatum, Paspalum malacophyllum and Paspalum
simplex (Ozias-Akins et al. 1998; Pupilli et al. 2001; Pupilli
et al. 2004; Akiyama et al. 2005). The non-recombinant
nature of ASGR highly compromised attempts to isolate
genes associated with the trait by direct genetic strategies.
Therefore, expression analyses followed by mapping of
selected candidates were perceived as valid alternative
approaches to facilitate the identification of the apomixis
trigger/s. Several groups focused on the isolation of
sequences differentially expressed in inflorescences of
sexual and apomictic plants (Rodrigues et al. 2003;
Albertini et al. 2004; Laspina et al. 2008; Sharbel et al.
2009; Yamada-Akiyama et al. 2009).
In P. notatum, a comprehensive differential display
analysis revealed 65 transcript sequences differentially
expressed in inflorescences of sexual and apomictic tetra-
ploid plants at late pre-meiotic stages (Laspina et al. 2008).
RFLP (restriction-fragment length polymorphisms) exper-
iments involving one of these sequences (experimental
code N20) showed a marker genetically linked in coupling
to the chromosomal locus governing apospory at a genetic
distance of 22 cM (Laspina et al. 2008). N20 was detected
as a transcript fragment amplified from RNA samples
which originated from sexual individuals (Laspina et al.
2008). Preliminary real-time PCR analysis confirmed a
minor down-regulation in inflorescences of apomictic
plants with respect to sexual ones at late pre-meiotic-
meiotic stages (Laspina et al. 2008). Sequencing revealed
that fragment N20 was homologous to GPI-anchored pro-
teins (Laspina et al. 2008).
The transcript represented by fragment N20 resulted
genetically linked to the apomixis-controlling locus at a
distance of 22 cM (Laspina et al. 2008). Therefore, it does
not qualify as a primary genetic determinant of this
reproductive system. However, its positional association
with the trait could indicate the existence of a cluster of
reproduction-related genes around the ASGR. The influ-
ence of the ASGR proximity on the expression of genes
controlling reproductive processes remains unclear, but
could involve epigenetic phenomenon such as chromatin
structure modification. Our hypothesis is that: (1) full
sequencing of N20 may reveal the existence of different
related alleles/paralogues; (2) Several alleles/paralogues
are represented in the P. notatum genome of sexual and
apomictic plants; (3) N20 is induced at specific stages
during sexual and apomictic development; 4) N20 is dif-
ferentially expressed in a representative sexual genotype
and a representative apomictic genotype at different
developmental stages. The objectives of this work were: (1)
characterize the N20 full cDNA sequence (corresponding
to gene n20gap-1, after N20 GPI-anchored protein-1); (2)
identify different alleles/paralogues; (3) determine gene
copy number in the genome of sexual and apomictic
genotypes; (4) quantify expression in reproductive tissues
of a representative sexual and a representative apomictic
plant, at different developmental stages; (5) analyze
Plant Mol Biol
123
Author's personal copy

reproductive tissue in situ expression in a representative
sexual and a representative apomictic plant, at the devel-
opmental stage of maximal expression and (6) identify
Arabidopsis thaliana putative orthologues and related
mutant germplasm, which could shed light on its potential
role in reproduction
Materials and methods
Plant material
Experiments were conducted on samples originated from
inflorescences of the tetraploid sexual plants Q4188
(2n = 4x = 40) and C4 (2n = 4x = 40), as well as the
tetraploid apomictic plant Q4117 (2n = 4x = 40). Clone
Q4188 is a hybrid derived from a highly sexual genotype
(Q3664) crossed with a natural apomict (Quarin et al.
2003). C4 is recently-formed autotetraploid, obtained after
doubling the chromosome content of a sexual diploid by
the use of colchicine (Quarin et al. 2001). Q4117 is an
obligate apomictic tetraploid clone collected from a natural
population of Southern Brazil (Ortiz et al. 1997). A
P. notatum pseudo-testcross mapping population segre-
gating for apospory (Stein et al. 2007) was used in the
bulked and de-bulked genomic DNA hybridization exper-
iments. This pseudo-testcross population had been obtained
by crossing the fully sexual heterozygous genotype Q4188
(2n = 4x = 40) (female parent) to the fully apomictic
heterozygous genotype Q4117 (2n = 4x = 40) (pollen
donor) (Stein et al. 2007). The total F1 population consisted
of 113 plants, out of which 15 were apomictic and 98
sexual. (Stein et al. 2007), which was in agreement with
previous reports indicating that in P. notatum apospory was
a dominant character with a highly distorted segregation
(Martınez et al. 2003). Ten sexual and ten apomictic F1
hybrids were selected at random to be used in bulked
genomic DNA hybridization experiments. Five sexual and
five apomictic F1 hybrids were selected at random to be
used in de-bulked genomic DNA hybridization experi-
ments (Stein et al. 2007). Plants were maintained in natural
conditions in experimental plots at IBONE (Instituto de
Botanica del Nordeste, Corrientes, Argentina). The com-
plete Paspalum notatum reproductive developmental cal-
endar reported in Laspina et al. (2008) was used for
selecting the appropriate stages for molecular and cyto-
embryological analyses.
Oligonucleotide design and RACE experiments
Two pairs of reverse-oriented gene-specific primers (GSP1
and GSP2) were designed for 30 and 50 RACE reactions,
following the recommendations of the MARATHON
cDNA amplification kit (BD Biosciences Clontech, San
Jose, CA, USA). 30 RACE primers: 50CGAATGCTGTG
CGCCGCTCTCAAGGAA30 and 50CCGTCTTGAGTGTG
GCAGTAGCGTTGTT30; 50 RACE primers: 50CAACAA
CGCTACTGCCACACTCAAGACG30 and 50TACGGCC
GACCCACCATTAGCACCAT30. Oligonucleotides were
23–28 nucleotides long and had from 50 to 70% GC con-
tent with a melting temperature C67�C. PCR reactions
were prepared in a 50 lL final volume mix, containing
2 lL of a MARATHON library product (BD Biosciences
Clontech, San Jose, CA, USA), 19 GoTAQ activity buffer
(Promega, Madison, Wisconsin, USA), 200 lM dNTPs,
0.2 lM gene specific primer (GSP1 and GSP2), 0.2 lM
adaptor-specific primers (AP1 or AP2) and 1.5 U of Go-
TAQ DNA polymerase enzyme (Promega, Madison, Wis-
consin, USA). Initial PCR conditions were the following:
94�C for 1 min followed by 30 cycles of 30 s at 94�C and
4 min at 63�C (both annealing and polymerization tem-
peratures were 63�C). To obtain the final 30 or 50 RACE
product, 2–3 PCR rounds were necessary. Positive and
negative controls were included in each step. Positive
controls consisted of amplifications with two specific oli-
gonucleotides matching in opposite direction that amplified
a small segment within the original sequence fragment.
Negative controls consisted of amplification reactions
using specific and adaptor-complementary primers in the
absence of template DNA. After examination by gel elec-
trophoresis, products were isolated using the SV WIZARD
GEL AND PCR CLEAN UP SYSTEM (Promega, Madi-
son, Wisconsin, USA). Transformation protocols were
taken from the Molecular Cloning Laboratory Manual
(Sambrook and Russell 2001). Plasmids were purified with
the WIZARD PLUS SV MINIPREPS kit (Promega, Mad-
ison, Wisconsin, USA). Insert verification was done by
PCR using the M13 forward and reverse primers and the
following amplification conditions: 94�C for 1 min, 25
cycles of 94�C for 30 s, 63�C for 1 s, 72�C for 1 min.
Sequencing of the 50 and 30 RACE clones was done by
Macrogen Inc (Korea).
Data analysis
Alignments between overlapping 30 and 50 RACE ampli-
fication products and the original sequence were conducted
with ClustalW2 on the EBI-EMBL website (http://www.
ebi.ac.uk/Tools/clustalw2) (Larkin et al. 2007). Analysis of
DNA similarity was carried out using the BLASTn and
BLASTx packages at NCBI (http://www.ncbi.nlm. nih.-
gov/BLAST/) and TAIR (http://www.arabidopsis.org).
Information on characterized germplasm was obtained
from the TAIR website. Conserved domains were detected
with the InterProScan and SMART tools at Expasy (http://
ca.expasy.org/tools/) (Quevillon et al. 2005). The Expasy
Plant Mol Biol
123
Author's personal copy

signal peptide prediction was confirmed by using the Sig-
nalP3 program (Bendtsen et al. 2004). Big-PI Plant Pre-
dictor version 2.1 (Eisenhaber et al. 2003) was used to
detect potential C-terminal GPI modification sites. Thirty-
three (33) sequences encoding plant GPI-anchored proteins
were obtained from NCBI databases (http://www.ncbi.
nlm.nih.gov/). Phylogenetic trees were constructed using
Drawtree application of Phylip version 3.68 (Felsenstein
2005) (with a 2000 bootstrapping). In silico mapping
analysis onto the rice genome was done at the Gramene
webpage (www.gramene.org).
Genomic hybridization analysis
Genomic DNA was extracted from 6 g of young leaves
using the CTAB method (Murray and Thompson 1980).
DNA quality was estimated by measuring the Abs260/280
index. DNA concentration was measured by using a
QUBIT fluorometer (Invitrogen, Carlsbad, USA). Genomic
DNA hybridization and detection was performed as indi-
cated in Stein et al. (2004) using 30 lg of DNA per sample.
To estimate the n20gap copy number, two bulks were
constructed in vitro with DNA from 10 aposporous and 10
non-aposporous (sexual) F1 individuals. Sexual and apo-
mictic bulks (SB and AB) were digested with EcoRI,
HindIII and PstI by using 1.5 U of enzyme per lg of DNA,
overnight at 37�C.
Reproductive tissue in situ hybridization experiments
Inflorescences in developmental stage I (late pre-meiosis)
or VII (anthesis) (Laspina et al. 2008), were fixed in 4%
paraformaldehyde/0.25% glutaraldehyde/0.01 M phos-
phate buffer pH 7.2, dehydrated in an ethanol series and
embedded in paraffin. Specimens were cut into 7 lm thin
sections and placed onto slides treated with poly-L-lysine
100 lg/mL. Paraffin was removed with a xylol series. The
probes used consisted of the sense and antisense version of
clone n20gap-1, including the complete nucleotide
sequence. The plasmid including insert n20gap-1 was lin-
earized using restriction enzymes NcoI or SalI (Promega,
Madison, Wisconsin, USA). Probes were labelled with the
DIG RNA LABELLING KIT (SP6/T7) (Roche Applied
Science, Mannheim, Germany), following the manufac-
turer’s instructions. Template digested with SalI restriction
enzyme was used to produce a probe from the T7 tran-
scription start (SalI probe). Template digested with NcoI
restriction enzyme was used to produce a probe from the
SP6 transcription start (NcoI probe). SalI probes detected
the antisense strand (sense probe), while NcoI probes
detected the sense strand (antisense probe). Probes were
hydrolysed to 150–200 bp fragments. Prehybridization was
carried out in a buffer of 0.05 M Tris–HCl pH 7.5
containing 1 lg/mL proteinase K, in a humid chamber at
37�C for 10 min. Hybridization was carried out in buffer
containing 10 mM Tris–HCl pH 7.5, 300 mM NaCl, 50%
deionized formamide, 1 mM EDTA pH 8.00, 1 9 Den-
hardt, 10% dextransulphate, 600 ng/mL total RNA and
60 ng of the corresponding probe, in a humid chamber at
42�C overnight. Detection was performed following the
instructions of the ROCHE DIG DETECTION KIT (Roche
Applied Science, Mannheim, Germany), using anti DIG
AP and NBT/BCIP as substrates.
Real-time PCR experiments
Real-time PCR reactions were prepared in a final volume
of 25 lL containing 200 nM gene specific primers, 19
REALMIX qPCR (Biodynamics) and 20 ng of reverse-
transcribed RNA (prepared by using SUPERSCRIPT II,
Invitrogen-Life Technologies). Specific PCR primer
pairs were designed by using Primer 3 (http://biotools.
umassmed.edu/bioapps/primer3_www.cgi.). Tubulin-spe-
cific primers were used to amplify the equal-expression
reference (Albertini et al. 2005). All oligonucleotides were
synthesized by IDT (Integrated DNA technologies,
http://www.idtdna.com/Home/Home.aspx). Amplification
efficiency was controlled to be equivalent for samples and
the corresponding internal control. RT (–) and non-tem-
plate controls were incorporated to the assays. Reactions
were performed on two biological replicates (different
RNA extractions), using four to six technical replicates.
Amplifications were performed in an Rotor-Gene Q ther-
mocycler (Quiagen), programmed as follows: 3 min at
95�C, 45 cycles of 15 s at 95�C, 30 s at 63�C, 20 s at 72�C
and 10 s at 78�C, next 5 min at 72�C. A melting curve (86
10 s cycles from 65 to 90�C, the temperature was increased
by 0.3�C after cycle 2) was produced at the end of the
cycling. Differential expression was estimated by using
both the 2-DDCt method (Livak and Schmittgen 2001) and
REST-RG (Relative Expression Software Tool V 2.0.7 for
Rotor Gene, Corbett Life Sciences). The significance of
differential expression was assessed by using non-para-
metric tests (considering Ct values and an amplification
efficiency of 2) and the REST-RG program (considering
take off values and the efficiency for each particular
amplification reaction). Equivalent conclusions were
obtained with both methods. Figures shown here were
based on results obtained using the REST-RG program
(Relative Expression Software Tool V 2.0.7 for Rotor
Gene, Corbett Life Sciences).
GenBank accession numbers
Clone N20: GQ385196; 30 RACE clone 27: GQ385197; 30
RACE clone 28: GQ385198; 30 RACE clone 29:
Plant Mol Biol
123
Author's personal copy

GQ385199; 30 RACE clone 30: GQ385200; 30 RACE clone
32: GQ385201; 30 RACE clone 33: GQ385202; 30 RACE
clone 34: GQ385203; 30 RACE clone 35: GQ385204; 30
RACE clone 36: GQ385205.
Results
N20gap-1 sequence characterization
The P. notatum n20gap-1 sequence originally isolated
from the sexual genotype was introduced in the Gene Bank
with the number GQ385196. It included the complete
coding region and part of the 50 and 30 UTR regions. A
search at the NCBI non redundant databases using Blastn
revealed that the n20gap-1 sequence matched at best the
maize mRNA full-length sequence ZM_BFc0104L23
(Locus BT063764) and the sorghum mRNA sequence
XM_002452592.1, both encoding putative GPI-anchored
proteins (E-value: 0.0). Blastx analysis showed highest
similarity to the Sorghum bicolor hypothetical pro-
tein SORBIDRAFT_04g029590, XP_002452637, and the
maize GPI-anchored protein NP_001148965 (E-value:
6e-63 and 5e-62, respectively). Within the coding region
the sequence was perfectly conserved, except for the initial
ATG codon position, which in the maize clone was pre-
dicted to be two triplets upstream. Additional Blastx sur-
veys in the Arabidopsis sequences databanks at the TAIR
website showed that P. notatum N20GAP-1 was homolo-
gous to the products of the A. thaliana LORELEI-family
genes At5g56170 (E-val: 1e-42), At2g20700 (E-val:
2e-36), At4g26466 (E-val: 6e-35), and At4g28280 (E-val:
1e-31). The representative gene of this family (At4g26466)
had been implicated in the process of egg-cell fertilization
in arabidopsis. LORELEI At4g26466 null mutants display
impaired sperm cell release into the egg cell, a phenotype
reminiscent of feronia/sirene mutants (Capron et al. 2008;
Boisson-Dernier et al. 2008; Tsukamoto et al. 2010). Since
egg-cell fertilization is absent in P. notatum apomictic
plants (embryos are formed through parthenogenesis), but
the central cell is fertilized to produce the endosperm, we
considered relevant to examine expression of n20gap-1
sequence in more detail.
As a preliminar step, the P. notatum N20GAP-1 protein
sequence was characterized by using bioinformatic tools.
The existence of conserved domains was analyzed by using
InterproScan and SMART tools at the Expasy website
(http://ca.expasy.org/tools). A conserved signal peptide
was detected at the amino-terminal region (from amino
acid 1–23). The same region was predicted to be a signal
peptide by using the SignalP3 program (Bendtsen et al.
2004). A transmembrane domain was predicted to be
located between amino acids 7 and 29. A low complexity
domain was predicted between amino acids 151 and 167.
Big-PI Plant Predictor version 2.1 (Eisenhaber et al. 2003)
detected two potential C-terminal GPI modification sites
represented by amino acids 144 and 143.
ClustalW2 alignments of the P. notatum N20GAP-1
protein sequence with homologous sequences from maize,
rice and A. thaliana allowed the detection of highly con-
served regions (Fig. 1). A core segment between amino
acids 44 and 135 was highly similar among all proteins
compared. Flanking N-terminal and C-terminal regions
were more variable.
A survey in the NCBI plant sequence databases facili-
tated the recovery of 33 plant GPI-anchored protein
sequences, representing the best hits in Blastx searches.
Sequences were aligned with ClustalW2 and the program
Phylip-3.68 was used to produce a phylogeny tree with
these 33 sequences (Fig. 2a). As expected, the tree showed
that N20GAP-1 was more similar to GPI-anchored proteins
originated from other Poaceae than those from Arabidopsis
thaliana. It was not possible to judge orthology relation-
ships with A. thaliana from this complete tree, since all
A. thaliana sequences were clustered in two single branches
located at identical distance with respect to N20GAP-1. A
second tree was constructed using only N20GAP-1 and
arabidopsis sequences, in order to reveal putative orthology
relationships (Fig. 2b). From this tree it was possible to
infer that n20gap-1 was a putative orthologue to
At5g56170 or At4g26466, since the program located both
genes at the same distance.
Figure 3 shows an alignment between P. notatum
N20GAP-1 and seven protein sequences originated from the
four A. thaliana LORELEI-like genes. Genes At2g20700 and
At4g28280 give rise to two different proteins each, which
differ in the presence or absence of short protein segments
within the highly variable N-terminal region (marked as a
and b in Fig. 3). The first 70 amino acids show low similarity
levels among all five sequences. The central region, which is
involved in recognition, is very well conserved.
As it was stated, one of the A. thaliana sequences
(At4g26466) corresponds to a gene responsible for an
altered reproductive phenotype in the LORELEI gameto-
phytic mutant (Capron et al. 2008). LORELEI At4g26466
mutants display impaired sperm cell release, a phenotype
reminiscent of feronia/sirene mutants (Capron et al. 2008;
Boisson-Dernier et al. 2008; Tsukamoto et al. 2010). Pollen
tubes reaching LORELEI embryo sacs frequently do not
rupture but continue to grow into the embryo sac, reaching
the central cell and then turning back to the micropyle.
Furthermore, LORELEI embryo sacs continue to attract
additional pollen tubes after arrival of the initial one
(Capron et al. 2008). The remaining three genes (At5g56170,
At2g20700 and At4g28280) were described as Arabidopsis
LORELEI-like proteins, since they are highly similar to
Plant Mol Biol
123
Author's personal copy

At4g26466 (Capron et al. 2008). However, no reproductive
phenotype was associated with null mutations involving
them (Capron et al. 2008; Boisson-Dernier et al. 2008,
Tsukamoto et al. 2010). Mutants with disruptions at
At5g56170 showed no distinctive reproductive features, no
reduced seed set or aborted seed phenotypes (Capron et al.
2008; Tsukamoto et al. 2010). No compensatory increase
in expression was observed in At5g56170 gene in a dis-
rupted At4g26466 background (Tsukamoto et al. 2010). No
increase in the frequency of undeveloped or aborted ovules
was observed in the siliques of double mutants of
At5g56170–At4g26466 compared with single mutants of
At4g26466 (Tsukamoto et al. 2010). These observations
suggest that At5g56170 function is not redundant with the
function of At4g26466 in the female gametophyte (Tsu-
kamoto et al. 2010).
RFLP analysis
RFLP analyses were performed on bulks of 10 F1 sexual
and 10 F1 aposporous plants, which originated from an
apomixis-segregating pseudo-testcross population (Stein
et al. 2007), using three different enzymes (EcoRI, HindIII
and PstI). In silico restriction analysis indicated that the
coding sequence had one restriction site for each one of the
enzymes used. Additional restriction sites occurring within
introns cannot be discarded. Therefore, it is possible that
two or even more bands which originated from the same
allele hybridized with the full-length probe. Hybridization
of DNA digested with EcoRI showed a total of seven
bands, while HindIII and PstI revealed a total of 11 and 5
bands, respectively. Even when the parental genotypes are
highly heterozygous tetraploids, these results seem to
indicate the presence of several members of a gene fam-
ily. In silico mapping analysis on the rice genome
detected three homologues with similar E-values located
on different chromosomes, namely LOC_Os02g48980,
LOC_Os06g19990 and LOC_Os09g12620, which also
evidences a moderate copy number for the gene family
members in the Poaceae. Bands co-segregating with apo-
mixis were readily detected, in agreement with results
reported in Laspina et al. (2008). Our results indicate the
Fig. 1 Multiple ClustalW
alignment of plant
GPI-anchored proteins amino
acid sequences recorded in
public databases. Accessions
and names: Z. mays (Zm1
NP_001148965.1, Zm2
NP_001137106.1 and Zm3
ACG39459.1), O. sativa (Os1
Os02g0721700, Os2
EAY87333.1 and Os3
ABR25668.1) and A. thaliana(At5g56170, encoding At
NP_200428.1). P. notatumN20GAP-1 shares between 80.5
and 50.3% amino acid sequence
identity with proteins from
maize, rice and arabidopsis.
Identical residues were
highlighted in black and similar
residues in grey
Plant Mol Biol
123
Author's personal copy

existence of several paralogues for n20gap in P. notatum,
all of which are detected by hybridizing with an n20gap-1
probe. At least one of the paralogues is located near the
ASGR (apospory specific genomic region). However, the
nature of this linked paralogue remains unknown. The
genomic DNA hybridization analysis is shown in Fig. 4.
Isolation of lorelei-like sequences from inflorescences
of an apomictic genotype
In order to extend the sequence toward the 50 and 30 UTR
regions and/or to isolate alleles/paralogues specifically
expressed in the apomictic genotype, MARATHON-RACE
(Rapid amplification of cDNA ends) (Chenchik et al. 1996)
experiments were carried out from inflorescences of the
apomictic genotype Q4117. Although the 50 RACE experi-
ments did not produce discrete bands, nested gene-specific
primers oriented downstream resulted in the recovery of 30
RACE products, allowing isolation of several unisequences
with highly significant similarity with the original N20
fragment (clones 27, 28, 29, 30, 32, 33, 34, 35 and 36).
Sequence alignments between RACE cDNA fragments and
the original N20 clone are shown in Fig. 5.
Sequences isolated from inflorescences of the apomictic
plant represented different alleles/gene members well-
conserved within the coding region but highly variable at
the 30 UTRs. Sequencing errors were considered not sig-
nificant, since variation involved strings of several nucle-
otides, and/or the same sequences were identified several
times in different sequencing reactions. Three groups of
sequences could be detected based on the 30 UTR similarity
(Family I—represented by clones 27, 28, 32, 33; Family
II—represented by clones 29, 34; Family III—represented
by clone 30). Clones 35 and 36 appeared to be incomplete,
showing exactly the same extension as the original N20
clone, but a slightly variable sequence.
Construction of a phylogenetic tree showed that
sequences representing the different families identified (I,
II and III) showed identical orthology relationship with
arabidopsis LORELEI sequences as the original n20gap-1
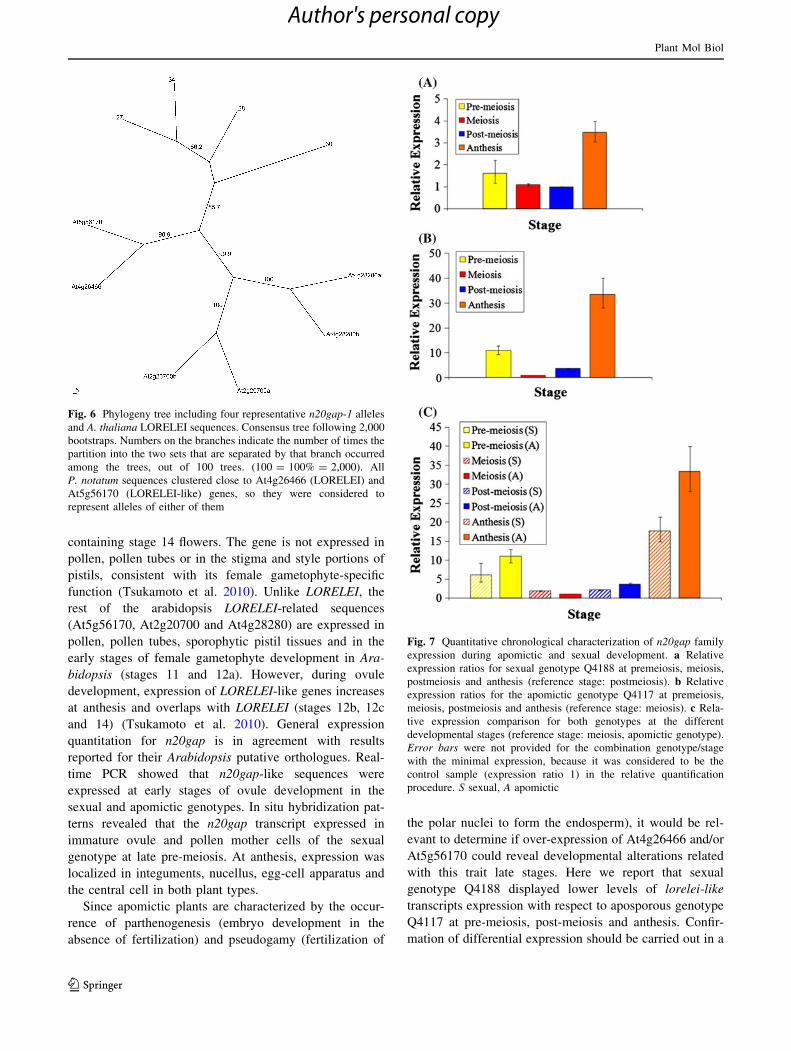
(Fig. 6). Therefore, they were considered to be alleles
originated from either paralogue At4g26466 or At5g56170.
However, the assignment of orthology is only tentative. It
is important to consider that ClustalW2 alignments were
performed only with a partial sequence (30-RACE
sequences encompassed only a part of the coding region
Fig. 2 Phylogeny tree including plant proteins similar to N20GAP-1.
a Tree including 33 plant rice, maize and arabidopsis sequences. As
expected, N20GAP-1 clustered closer to GPI-anchored proteins from
other Poaceae (maize and rice). b Tree including only arabidopsis
sequences. N20GAP-1 grouped preferentially with genes At4g26466
(LORELEI) and At5g56170 (LORELEI-like). Consensus trees fol-
lowing 2,000 bootstraps. The numbers on the branches indicate the
number of times the partition into the two sets that are separated by
that branch occurred among the trees, out of 100 trees
(100 = 100% = 2,000). At arabidopsis thaliana, In ipomoea nil, Osoryza sativa, Pp physcomitrella patens subsp. Patens, Ps picea
sitchensis, Pt populus trichocarpa, Rc ricinus communis, Vr vigna
radiata, Vv vitis vinifera, Zm zea mays
Plant Mol Biol
123
Author's personal copy

and the complete 30 UTR). Full protein sequences should
be used to establish more solid results.
Quantitative chronological characterization
of the n20gap family expression during apomictic
and sexual development
Real-time PCR experiments were conducted in order to
estimate n20gap activity levels at several sexual and
aposporous developmental stages covering the whole
reproductive calendar introduced in Laspina et al. (2008).
Spikelets from genotypes Q4188 (sexual) and Q4117
(apomictic) at pre-meiosis (step 0), late pre-meiosis/mei-
osis (steps I-II), post-meiosis (steps IV-V) and anthesis
(steps VII) were classified based on macromorphology
and microscopic observation of pollen developmental
stages, as indicated in Laspina et al. (2008). Initially, a
primer pair that amplified a short conserved sequence
within the protein coding region was used for amplifica-
tion. This primer pair was designed to target sequences
originated from all lorelei-like transcripts (it does not
discriminate alleles or paralogues). Each amplification
reaction was prepared in triplicate, and at least three
technical replicates were run.
In the sexual genotype (Q4188), the lowest activity was
observed at post-meiosis (Fig. 7a). N20gap was signifi-
cantly up-regulated at anthesis with respect to post-meiosis
(expression ratio: 3.488). No highly significant differential
activity was detected between pre-meiotic, meiotic and
post-meiotic stages. In the apomictic genotype (Q4117),
the lowest activity was detected at meiosis (Fig. 7b).
N20gap was significantly up-regulated at pre-meiosis with
respect to meiosis (expression ratio 10.98), at post-meiosis
with respect to meiosis (expression ratio 3.69) and at
anthesis with respect to meiosis (expression ratio 33.32).
Considering both the apomictic and sexual genotypes, two
activity peaks were detected: a minor one at pre-meiosis
and a major one at anthesis. However, a significantly
higher expression variation was observed for the apomictic
genotype with respect to the sexual one: maximal expres-
sion variation ratios of 3.69 and 33.32 were observed in the
sexual and the apomictic plants, respectively (Fig. 7a, b).
Regulation of expression seems to be more drastic in the
apomictic plant, since the distance between the minimal
and maximal expression level is very much increased. A
chronological comparative expression profile for the apo-
mictic and the sexual genotypes revealed that n20gap was
significantly up-regulated at pre-meiosis, post-meiosis and
Fig. 3 Multiple ClustalW
alignment of amino acid
sequences from arabidopsis
GPI-anchored proteins recorded
in public databases. P. notatumN20GAP-1 shares between 50.9
and 43.2% amino acid sequence
identity with proteins from
arabidopsis. Identical residues
are highlighted in black and
similar residues in grey. Genes
At4g28280 and At2g20700
generate two different proteins
each (a and b)
Plant Mol Biol
123
Author's personal copy

anthesis in the apomictic plant with respect to the sexual
one (Fig. 7c). In agreement with results reported by
Laspina et al. (2008), expression was slightly higher in the
sexual genotype at late pre-meiosis/meiosis. However,
significant differential expression could not be confirmed at
this particular stage. In summary, results showed in
Fig. 7a–c, suggest that n20gap maximal activity takes
place at anthesis, and differential regulation is occurring at
pre-meiosis, post-meiosis and anthesis between the apo-
mictic and the sexual genotypes used in this particular
analysis.
In situ hybridization experiments on reproductive
tissues
For a better characterization of the expression pattern, in situ
hybridization experiments were carried out on reproductive
tissues of the apomictic and sexual genotypes at late
premeiosis and anthesis. DIG-labelled antisense (NcoI-Sp6)
and sense (SalI-T7) probes obtained from n20gap-1 were
used to assess the specific location of expression (Fig. 8).
Probes generated from n20gap-1 have the potential to
hybridize with several P. notatum n20gap paralogues, as it
was shown in RFLP experiments, so the pattern observed
should be considered a combined signal. In situ hybridization
is not a quantitative technique, therefore it is not possible to
infer differential expression unless differences in signal
intensity were drastic.
The developmental phase of the flowers examined cor-
responded to stage I and stage VII in the reproductive
calendar of P. notatum (Laspina et al. 2008). Stage I
immediately precedes the megaspore mother cell differ-
entiation in sexual plants and the emergence of apospory
initials in the nucellus of aposporous plants. Candidate
transcripts displaying differential expression had originally
been isolated from inflorescences at this stage (Laspina
et al. 2008). Stage VII corresponds to anthesis, when a
maximal expression of n20gap was detected.
Hybridization with the N20 NcoI probe (antisense
probe) showed a moderate signal in stage I immature
ovules and pollen mother cells of the sexual genotype,
while the apomictic genotype displayed no significant
signal (Fig. 8). The N20 SalI probe (sense probe) produced
no significant signal in both plant types. A control probe
that originated from a transcript with uniform expression in
differential display experiments (N47) revealed compara-
ble signals in tissues from both plant types (not shown).
At anthesis, hybridization with the N20 NcoI probe
(antisense probe) showed a strong signal in ovules of the
apomictic genotype (Fig. 8). The signal was restricted to
the integuments, the nucellus, and the cytoplasm of the
embryo sac cells (egg cell apparatus and central cell). Lack
of hybridization at inner nucellus and inside the embryo sac
is probably due to the presence of massive vacuoles.
Hybridization in the sexual genotype followed a similar
pattern. The N20 SalI probe (sense probe) produced no
significant signal on both plant types. These results indi-
cated that the sense strand of the n20gap-1 gene is
expressed in integuments, nucellus and embryo sac cells in
both plant types.
Allele-specific expression
The n20gap general primers used in real-time PCR
experiments described in the previous section had been
designed to match conserved sequences included in the
GPI-anchored protein coding region, and did not discrim-
inate different alleles/paralogues within the n20gap family.
However, the prior identification of several alleles of
n20gap-1 allowed specific amplification based on the
Fig. 4 Genomic DNA hybridization analysis of n20gap in sexual and
apomictic P. notatum genotypes. a Sexual (SB) and Apomictic (AB)
bulked genomic DNAs digested with EcoRI, HindIII and PstI and
hybridized with clone n20gap-1 (680 bp) as a probe. Bulks were
made of genomic DNA originating from ten F1 sexual (SB) or ten F1
apomictic (AB) genotypes. Multiple band patterns revealed the
occurrence of several alleles/gene members for n20gap. Several
genetic polymorphisms were observed between bulks. b Hybridization
with n20gap-1 probe on debulked genomic DNA digested with
HindIII. Genomic DNAs originated from 5 sexual and 5 aposporous
F1 progenies from a segregating family. Arrows indicate bands co-
segregating with apospory. The fragment marked with an asterisk had
already been mapped onto the P. notatum genome, and was found to
be linked to the apospory-governing locus at 22 cM (Laspina et al.
2008). The DNA ladder (Lambda-EcoRI-HindIII) migration was
indicated on the left
Plant Mol Biol
123
Author's personal copy

sequence variation observed at the 30 UTR. Additional
primers were designed onto the 30 UTR region in order to
specifically amplify sequences corresponding to four
n20gap-1 alleles (Family I: alleles 27 and 28; Family II:
allele 34; Family III: allele 30). Bands of the expected
molecular weights corresponding to the specific alleles,
which originated from parental plants Q4117 and Q4188
genomic DNAs were observed in polyacrylamide gels (data
not shown). That indicated that all four alleles were present
in both tetraploid parental plants. Specific real-time PCRs
were performed for each allele (27, 28, 30 and 34) on
samples collected from the apomictic (Q4117) and sexual
(Q4188) genotypes at anthesis. Average expression ratios
are shown in Fig. 9.
Allele 27 did not amplify in the first 40 cycles. Allele 28
resulted over-expressed in the apomictic plant with respect
to the sexual one at a highly-significant level (ratio 4.150).
Alleles 30 and 34 were found over-expressed in the apo-
mictic plant with respect to the sexual one at a non-sig-
nificant level (ratios 2.022 and 2.112, respectively)
Fig. 5 Isolation of several
n20gap allelic sequences from
inflorescences of aposporous
P. notatum. The original N20
sequence had been isolated from
sexual plants. 30 RACE
extensions were obtained from a
MARATHON library originated
from an apomictic genotype
(Q4117). Sequences overlapped
with high similarity to the 30 end
of the N20 sequence, and
extended the sequence toward
the 30 UTR region. Three groups
of transcripts could be identified
based on the homology at the 30
UTR (group 1: clones 27, 28,
32, 33; group 2: clones 29, 31,
34 and group 3: clone 30).
Phylogenetic analysis shown in
Fig. 2 panels a and b was done
using the original N20 sequence
(the sequence at the top here).
Phylogenetic analysis shown in
Fig. 6 was done using allelic
sequences 27, 28, 30 and 34,
which represent all allelic
groups detected
Plant Mol Biol
123
Author's personal copy

(Fig. 9). Results indicate that transcripts originated from
n20gap allele 28 are significantly over-expressed in the
apomictic genotype at anthesis, and are probably respon-
sible for the increase of general expression shown in Fig. 7.
Discussion
GPI anchored proteins (GAPs) are broadly distributed
among eukaryotic organisms, including protozoa, fungi,
plants, insects, and mammals (Nosjean et al. 1997). They
vary widely in size, ranging from 12 to 175 kDa (Nosjean
et al. 1997). In mammals, they are generally located on lipid
rafts organized as discrete domains in the outer layer of the
cell membrane, exposed on the cell surface and attached to
the membrane by the glycosylphosphatidylinositol anchor.
GAPs can be released from their anchor into the extracellular
medium in a reaction catalyzed by phospholipase C. These
molecules display diverse biological functions, most of them
characterized in mammals and/or protozoans (Paulick and
Fig. 5 continued
Plant Mol Biol
123
Author's personal copy

Bertozzi 2008), participating in a variety of processes related
with cell to cell adhesion, cell to cell communication and
signal transduction. Some of them behave as enzymes, dis-
playing alkaline phosphatase, 50 nucleotidase, or peptidase
activities, among others (Paulick and Bertozzi 2008).
Limited functional characterization of GPI-anchored
proteins has been performed in plants. Only a few genes
encoding GAPs have been described in terms of function in
A. thaliana. The gene COBRA is required for polarized cell
expansion in the root (Schindelman et al. 2001), and the
gene NDR-1 is involved in signal transduction during dis-
ease resistance (Coppinger et al. 2004). A classical arabi-
nogalactan protein containing a GPI anchor has been
shown to be involved in the initiation of female gameto-
genesis in A. thaliana and has been proposed to be involved
in intercellular signalling between sporophytic and game-
tophytic cells (Acosta-Garcia and Vielle-Calzada 2004).
Characterization of the gametophytic A. thaliana LORELEI
Fig. 5 continued
Plant Mol Biol
123
Author's personal copy

mutants revealed that a GPI-anchored protein encoded by
gene At4g26466 was responsible for the altered phenotype,
consisting of an impaired delivery of the male sperm into
the egg-cell (Capron et al. 2008). Besides pollen tube
reception, lorelei also has a role in double fertilization and
early seed development (Tsukamoto et al. 2010).
Here we present the structural and expression charac-
terization of Paspalum notatum transcript n20gap-1, which
encodes a GPI-anchored protein and had been previously
associated with aposporous apomixis in P. notatum
(Laspina et al. 2008). Full sequencing allowed analysis of
the transcript and its derived protein structure and estima-
tion of their phylogenetic relationships with other plant
GPI-anchored proteins. The n20gap-1 sequence is highly
homologous to all A. thaliana LORELEI family members,
but appears to be more related with genes At4g26466
(LORELEI) and At5g56170 (LORELEI-like). General
primers which amplify all LORELEI-like family tran-
scripts revealed an increased expression in genotype Q4117
(apomictic) with respect to Q4188 (sexual) at premeiosis,
postmeiosis and anthesis. Allele-specific primers revealed
three particular alleles (28, 30 and 34) were overexpressed
in the apomictic genotype at anthesis, but only one of them
(28) at a highly-significant level. This observation suggests
that n20gap-1 expression pattern might be differential
between sexual and apomitic genotypes. However, the
assessment of association of an increased expression of
n20gap-1 with apomixis will require the analysis of a
significant number of sexual and apomictic genotypes. In a
future work we will evaluate expression in a significant
number of inflorescences collected from apomictic and
sexual F1 individuals derived from the cross Q4188 9
Q4117 at anthesis (the stage when maximal activity was
observed)
LORELEI (At4g26466) is expressed in the arabidopsis
synergid cells prior to fertilization, probably taking part in
a signalling mechanism by which the female gametophyte
recognizes the arrival of a compatible pollen tube and
promotes sperm release (Capron et al. 2008). Pollen tubes
reaching lorelei embryo sacs frequently do not rupture but
continue to grow. Furthermore, lorelei embryo sacs con-
tinue to attract additional pollen tubes after arrival of the
initial one (Capron et al. 2008). Transcription was detected
in ovaries from stage 12c and mature female gametophyte
Fig. 5 continued
Plant Mol Biol
123
Author's personal copy
containing stage 14 flowers. The gene is not expressed in
pollen, pollen tubes or in the stigma and style portions of
pistils, consistent with its female gametophyte-specific
function (Tsukamoto et al. 2010). Unlike LORELEI, the
rest of the arabidopsis LORELEI-related sequences
(At5g56170, At2g20700 and At4g28280) are expressed in
pollen, pollen tubes, sporophytic pistil tissues and in the
early stages of female gametophyte development in Ara-
bidopsis (stages 11 and 12a). However, during ovule
development, expression of LORELEI-like genes increases
at anthesis and overlaps with LORELEI (stages 12b, 12c
and 14) (Tsukamoto et al. 2010). General expression
quantitation for n20gap is in agreement with results
reported for their Arabidopsis putative orthologues. Real-
time PCR showed that n20gap-like sequences were
expressed at early stages of ovule development in the
sexual and apomictic genotypes. In situ hybridization pat-
terns revealed that the n20gap transcript expressed in
immature ovule and pollen mother cells of the sexual
genotype at late pre-meiosis. At anthesis, expression was
localized in integuments, nucellus, egg-cell apparatus and
the central cell in both plant types.
Since apomictic plants are characterized by the occur-
rence of parthenogenesis (embryo development in the
absence of fertilization) and pseudogamy (fertilization of
the polar nuclei to form the endosperm), it would be rel-
evant to determine if over-expression of At4g26466 and/or
At5g56170 could reveal developmental alterations related
with this trait late stages. Here we report that sexual
genotype Q4188 displayed lower levels of lorelei-like
transcripts expression with respect to aposporous genotype
Q4117 at pre-meiosis, post-meiosis and anthesis. Confir-
mation of differential expression should be carried out in a
Fig. 6 Phylogeny tree including four representative n20gap-1 alleles
and A. thaliana LORELEI sequences. Consensus tree following 2,000
bootstraps. Numbers on the branches indicate the number of times the
partition into the two sets that are separated by that branch occurred
among the trees, out of 100 trees. (100 = 100% = 2,000). All
P. notatum sequences clustered close to At4g26466 (LORELEI) and
At5g56170 (LORELEI-like) genes, so they were considered to
represent alleles of either of them
Fig. 7 Quantitative chronological characterization of n20gap family
expression during apomictic and sexual development. a Relative
expression ratios for sexual genotype Q4188 at premeiosis, meiosis,
postmeiosis and anthesis (reference stage: postmeiosis). b Relative
expression ratios for the apomictic genotype Q4117 at premeiosis,
meiosis, postmeiosis and anthesis (reference stage: meiosis). c Rela-
tive expression comparison for both genotypes at the different
developmental stages (reference stage: meiosis, apomictic genotype).
Error bars were not provided for the combination genotype/stage
with the minimal expression, because it was considered to be the
control sample (expression ratio 1) in the relative quantification
procedure. S sexual, A apomictic
Plant Mol Biol
123
Author's personal copy

significant number of apomictic and sexual plants, but the
present results suggest that an increased activity of these
candidates could bring out its probable function.
Apospory in P. notatum is governed by one or several
genes located within a single non-recombinant genomic
region of around 36 Mbp, which appears to have suffered
an inversion (Stein et al. 2004). The genetic linkage
between an n20gap gene and the ASGR (apospory specific
genomic region) had been reported previously (Laspina
et al. 2008) and was confirmed here. Since the probes used
to map the n20gap locus detect conserved regions, they did
not discriminate among paralogues, so the nature of the
n20gap-related sequence linked to the ASGR remains
unknown. In future work it should be determined which
one of the paralogues is actually linked to the ASGR. The
linkage between the n20gap gene and the ASGR and its
association with an altered expression needs also to be
investigated further. If only genetic components were
involved in the triggering of apomixis, the lack of full
linkage would have been enough evidence to discard it as
the gene governing the trait. However, if both genetic and
epigenetic factors were acting, n20gap or any other gene
located close to the apo-region should not be excluded as
possible triggers of the trait. The occurrence of particular
phenotypes associated with inversions that cause epigenetic
alterations of large neighbouring regions have been repor-
ted in other organisms (Grewal and Elgin 2002). Moreover,
the proximity of n20gap to the apospory-governing region
Fig. 8 Reproductive tissue in situ hybridization of sexual an
apomictic genotypes. a–d Show hybridization with the N20gap1NcoI probe (antisense probe) on reproductive organs at late-preme-
iosis. a and c Correspond to the apomictic genotype (Q4117), while
b and d to the sexual one (C4). e–l Show hybridization with the
N20gap1 NcoI probe (antisense probe) on reproductive organs at
anthesis. e–j Correspond to the apomictic genotype (Q4117), while
k and l to the sexual one. Expression is localized mainly in
integuments, nucella and embryo sac cells. m and n Show hybrid-
ization with the N20gap1 SalI probe (sense probe) on reproductive
organs of the apomictic and sexual genotype at anthesis, respectively.
ea egg apparatus, es embryo sac, in integuments, nu nucellus, Ovovule, pmc pollen mother cells. Bars: 100 lm
Plant Mol Biol
123
Author's personal copy

could be indicative of the existence of a cluster of repro-
ductive genes around the ASGR. If n20gap genes were
affected by trans-acting factors originated from the neigh-
bouring ASGR, genetic linkage would not be an essential
condition for differential activity. Alternatively, if n20gap
genes were regulated through an epigenetic mechanism
affecting a large region around the non-recombinant ASGR,
the existence of genetic linkage would be necessary for
differential activity. The minor activity peak observed at
early pre-meiosis, just before the differentiation of apospory
initials, together with the partial linkage to the ASGR,
favours the hypothesis that assigns a role of one or several
of the lorelei-like genes at the apospory onset. However, the
drastic activation of n20gap observed at a late develop-
mental stage (anthesis) and the prior evidence involving the
LORELEI family in the fertilization process supports its
function as a downstream participant rather than a trigger of
apomixis.
In the last few years a considerable amount of infor-
mation has been generated from several projects aimed at
exploring contrasting gene expression in inflorescences of
sexual and apomictic plants (Albertini et al. 2004; Laspina
et al. 2008; Cervigni et al. 2008; Sharbel et al. 2009;
Yamada-Akiyama et al. 2009). In parallel, detailed surveys
of transcripts expressed during A. thaliana megagameto-
phyte development also have been carried out, and an
important number of Arabidopsis gametophytic mutants
have been identified and characterized (Pagnussat et al.
2005; Hee-Ju et al. 2005; Steffen et al. 2007). The inte-
gration of information originating from both sources will
contribute to the elucidation of many of the developmental
steps involved in apomixis. Since apomixis frequently
occurs in wild, highly-heterozygous, poorly-characterized
species, the reference to model plants will greatly accel-
erate the discovery of the molecular pathways related to
this trait. However, the triggering of the character, the
regulation of gene activity, the association with polyploidy
and the occurrence of novel genes must of necessity be
examined directly in natural apomicts. It will be difficult or
even impractical to transfer results from model species
when investigating these particular issues.
Acknowledgments Thanks are due to Prof. Camilo Quarin for
kindly providing the plant material used in this work. We thank
Dr. Michael Hayward, Dr. Peggy Ozias-Akins and Dr. Marta Bianchi
for valuable corrections and suggestions that helped to improve the
manuscript. This work was funded by Agencia Nacional de Promo-
cion Cientıfica y Tecnologica, Argentina (ANPCyT PICT 2003
13578, PICT 2007 00476 and PME 2006 03083); Consejo Nacional
de Investigaciones Cientıficas y Tecnicas Argentina (CONICET, PIP
2008 6805); Centro Argentino Brasileno de Biotecnologıa (CABBIO
Proy. 2004 012). Podio M, Laspina N and Siena L received a fel-
lowship from CONICET (Consejo Nacional de Investigaciones
Cientıficas y Tecnicas, Argentina). Felitti SA, Seijo JG., Gonzalez
AM, Ortiz JPA and Pessino SC are career members of CONICET
(Consejo Nacional de Investigaciones Cientıficas y Tecnicas,
Argentina).
References
Acosta-Garcia G, Vielle-Calzada JP (2004) A classical arabinogalac-
tan protein is essential for the initiation of female gametogenesis
in Arabidopsis. Plant Cell 16:2614–2628
Akiyama Y, Hanna WW, Ozias-Akins P (2005) High-resolution
physical mapping reveals that the apospory-specific genomic
region (ASGR) in Cenchrus ciliaris is located on a heterochro-
matic and hemizygous region of a single chromosome. Theor
Appl Genet 111:1042–1051
Fig. 9 Quantitative expression characterization of n20gap-1 allelic
variants during apomictic and sexual development. The relative
expression ratios for different representative alleles (28, 30 and 34) at
anthesis is shown. a Allele 28. b Allele 30. c Allele 34. Expression of
allele 27 was undetectable. Standard deviations were indicated with
lines at the top of the bars. Error bars were not provided for the
sexual genotype, since it presented the minimal expression, and was
therefore considered to be the control sample (expression ratio 1) in
the relative quantification procedure. S sexual, A apomictic
Plant Mol Biol
123
Author's personal copy

Albertini E, Marconi G, Barcaccia G, Raggi L, Falcinelli M (2004)
Isolation of candidate genes for apomixis in Poa pratensis. Plant
Mol Biol 56:879–894
Albertini E, Marconi G, Reale L, Barcaccia G, Proceddu A, Ferranti F,
Falcinelli M (2005) SERK and APOSTART. Candidates genes for
apomixis in Poa pratensis. Plant Physiol 138:2185–2199
Bendtsen JD, Nielsen H, von Heijne G, Brunak S (2004) Improved
prediction of signal peptides-SignalP 3.0. J Mol Biol 340:783–795
Boisson-Dernier A, Frietsch S, Kim TH, Dizon MB, Schroeder JI
(2008) The peroxin loss-of-function mutation abstinence by
mutual consent disrupts male-female gametophyte recognition.
Curr Biol 18:63–68
Capron A, Gourgues M, Neiva LS, Faure J-E, Berger F, Pagnussat G,
Krishnan A, Alvarez-Mejia C, Vielle-Calzada J-P, Lee Y-R, Liu
B, Sundaresan V (2008) Maternal control of male-gamete
delivery in Arabidopsis involves a putative GPI-anchored protein
encoded by the LORELEI Gene. Plant Cell 20:3038–3049
Cervigni GDL, Paniego N, Pessino S, Selva JP, Dıaz M, Spangenberg
G, Echenique V (2008) Gene expression in diplosporous and
sexual Eragrostis curvula genotypes with differing ploidy levels.
Plant Mol Biol 67:11–23
Chenchik A, Mogadam F, Siebert P (1996) A new method for full-
length cDNA cloning by PCR. In: Krieg PA (ed) A laboratory
guide to RNA: isolation, analisis and synthesis. Wiley-Liss, Inc.,
New York, pp 273–321
Coppinger P, Repetti PP, Day B, Dahlbeck D, Mehlert A, Staskawicz
BJ (2004) Overexpression of the plasma membrane localized
NDR1 protein results in enhanced bacterial disease resistance in
Arabidopsis thaliana. Plant J 40:225–237
Crane C (2001) Classification of apomictic mechanisms. In: Savidan
Y, Carman G, Dresselhaus T (eds) Flowering of Apomixis: From
Mechanisms to Genetic Engineering. CIMMYT, IRD, European
Commission DG VI, Mexico, pp 24–43
Eisenhaber B, Wildpaner M, Schultz CJ, Borner GHH, Dupree P,
Eisenhaber F (2003) Glycosylphosphatidylinositol lipid anchor-
ing of plant proteins. Sensitive prediction from sequence- and
genome-wide studies for arabidopsis and rice. Plant Physiol
133:1691–1701
Felsenstein J (2005) PHYLIP (phylogeny inference package) version
3.6. Department of Genome Sciences, University of Washington,
Seattle
Gates RN, Quarin CL, Pedreira CGS (2004) Bahiagrass. In: Moser LE,
Burson BL, Sollenberger LE (eds) Warm-season (C4) grasses,
Agron. Monogr. 45. ASA, CSSA, SSSA, Madison, WI, pp 651–680
Grewal SI, Elgin SC (2002) Heterochromatin: new possibilities for
inheritance of structure. Curr Opin Genet Dev 12:178–187
Grossniklaus U (2001) From sexuality to apomixis: molecular and
genetic approaches. In: Savidan Y, Carman JG, Dresselhaus T
(eds) The flowering of apomixis: from mechanisms to genetic
engineering. Centro Internacional de Mejoramiento de Maız y
Trigo (CIMMYT), Mexico, pp 168–211
Hee-Ju Y, Hogan P, Sundaresan V (2005) Analysis of the female
gametophyte transcriptome of Arabidopsis by comparative
expression profiling. Plant Physiol 139:1853–1869
Larkin MA, Blackshields G, Brown NP, Chenna R, McGettigan PA,
McWilliam H, Valentin F, Wallace IM, Wilm A, Lopez R,
Thompson JD, Gibson TJ, Higgins DG (2007) Clustal W and
Clustal X version 2.0. Bioinformatics 23:2947–2948
Laspina NV, Vega T, Seijo JG, Gonzalez AM, Martelotto LG, Stein J,
Podio M, Ortiz JPA, Echenique VC, Quarin CL, Pessino SC
(2008) Gene expression analysis at the onset of aposporous
apomixis in Paspalum notatum. Plant Mol Biol 67:615–628
Livak KJ, Schmittgen TD (2001) Analysis of relative gene expression
data using real-time quantitative PCR and the 2-DDCt method.
Methods 25:402–408
Martınez EJ, Hopp E, Stein J, Ortiz JPA, Quarin CL (2003) Genetic
characterization of apospory in tetraploid Paspalum notatumbased on the identification of linked molecular markers. Mol
Breed 12(4):319–327
Murray MG, Thompson WF (1980) Rapid isolation of high molecular
weight plant DNA. Nucleic Acids Res 8:4321–4325
Nogler GA (1984) Gametophytic apomixis. In: Johri BM (ed)
Embryology of angiosperms. Springer, Berlin, pp 475–518
Nosjean O, Briolay A, Roux B (1997) Mammalian GPI proteins:
sorting, membrane residence and functions. Biochim Biophys
Acta 1331:153–186
Ortiz JPA, Pessino SC, Leblanc O, Hayward MD, Quarin CL (1997)
Genetic fingerprinting for determining the mode of reproduction
in Paspalum notatum, a subtropical apomictic forage grass.
Theor Appl Genet 95:850–856
Ortiz JPA, Pessino SC, Bhat V, Hayward MD, Quarin CL (2001) A
genetic linkage map of diploid Paspalum notatum. Crop Sci
41:823–830
Ozias-Akins P (2006) Apomixis: developmental characteristics and
genetics. Crit Rev Plant Sci 25:199–214
Ozias-Akins P, Roche D, Hanna WW (1998) Tight clustering and
hemizygosity of apomixis-linked molecular markers in Pennise-tum squamulatum implies genetic control of apospory by
divergent locus that may have no allelic form in sexual gentypes.
Proc Natl Acad Sci USA 95:5127–5132
Pagnussat GC, Yu HJ, Ngo QA, Rajani S, Mayalagu S, Johnson CS,
Capron A, Xie LF, Ye D, Sundaresan V (2005) Genetic and
molecular identification of genes required for female gameto-
phyte development and function in Arabidopsis. Development
132:603–614
Paulick MG, Bertozzi CR (2008) The glycosylphosphatidylinositol
anchor: a complex membrane-anchoring structure for proteins.
Biochemistry 47:6991–7000
Pupilli F, Lambobarda P, Caceres ME, Quarin CL, Arcioni S (2001)
The chromosome segment related to apomixis in Paspalumsimplex is homoeologous to the telomeric region of the long arm
of rice chromosome 12. Mol Breed 8:53–61
Pupilli F, Martınez EJ, Busti A, Calderini O, Quarin CL, Arcioni S
(2004) Comparative mapping reveals partial conservation of
synteny at the apomixis locus in Paspalum spp. Mol Genet
Genomics 270:539–548
Quarin CL (1992) The nature of apomixis and its origin in Panicoid
grasses. Apomixis Newsl 5:8–15
Quarin CL, Espinoza F, Martınez EJ, Pessino SC, Bovo OA (2001) A
rise of ploidy level induces the expression of apomixis in
Paspalum notatum. Sex Plant Reprod 13:243–249
Quarin CL, Urbani MH, Blount AR, Martınez EJ, Hack CM, Burton
GW, Quesenberry KH (2003) Registration of Q4188 and Q4205,
sexual tetraploid germplasm lines of Bahiagrass. Crop Sci
43:745–746
Quevillon E, Silventoinen V, Pillai S, Harte N, Mulder N, Apweiler
R, Lopez R (2005) InterProScan: protein domains identifier.
Nucleic Acids Res 33:W116–W120
Richards AJ (2003) Apomixis in flowering plants: an overview. Philos
Trans R Soc Lond B 358:1085–1093
Rodrigues JC, Cabral GB, Dusi DMA, Mello LV, Rinden D, Carneiro
VT (2003) Identification of differentially expressed cDNA
sequences in ovaries of sexual and apomictic plants of Brachi-aria brizantha. Plant Mol Biol 53:745–757
Sambrook J, Russell DW (2001) Molecular cloning: a laboratory
manual. Cold Spring Harbor Laboratory Press, Cold Spring
Harbor
Schindelman G, Morikami A, Jung J, Baskin TI, Carpita NC,
Derbyshire P, McCann MC, Benfey PN (2001) COBRA encodes
a putative GPI-anchored protein, which is polarly localized and
Plant Mol Biol
123
Author's personal copy

necessary for oriented cell expansion in Arabidopsis. Genes Dev
15:1115–1127
Sharbel T, Voigt M-L, Corral JM, Thiel T, Varshney A, Kumlehn J, Vogel
H, Rotter B (2009) Molecular signatures of apomictic and sexual
ovules in the Boechera holboellii complex. Plant J 58:870–882
Spillane C, Curtis MD, Grossniklaus U (2004) Apomixis technology
development-virgin birth in farmers’ fields. Nat Biotechnol
22(6):687–691
Steffen JG, Kang I, MacFarlane J, Drews GN (2007) Identification of
genes expressed in the Arabidopsis female gametophyte. Plant J
51:281–292
Stein J, Quarin CL, Martınez EJ, Pessino SC, Ortiz JPA (2004)
Tetraploid races of Paspalum notatum show polysomic inheri-
tance with preferential chromosome pairing and suppression of
recombination around the apospory-controlling locus. Theoret
Appl Genet 109:186–191
Stein J, Pessino SC, Martınez EJ, Rodrıguez MP, Siena LA, Quarin
CL, Ortiz JPA (2007) A genetic map of tetraploid Paspalumnotatum Flugge (bahiagrass) based on single-dose molecular
markers. Mol Breed 20:153–166
Tsukamoto T, Qin Y, Huang Y, Dunatunga D, Palanivelu R (2010) A
role for LORELEI, a putative glycosylphosphatidylinositol
anchored protein, in Arabidopsis thaliana double fertilization
and early seed development. Plant J 62:571–588
Yamada-Akiyama H, Akiyama Y, Ebina M, Xu Q, Tsuruta S, Yazaki
J, Kishimoto N, Kikuchi S, Takahara M, Takamizo T, Sugita S,
Nakagawa H (2009) Analysis of expressed sequence tags in
apomictic guineagrass (Panicum maximum). J Plant Physiol
166:750–761
Plant Mol Biol
123
Author's personal copy
Copyright © 2022 FDOKUMEN




















