
Contrasting ecological breadth of co-occurring annual Polygonum species
Upload
washingtonCategory
view
3download
0
M A J O R A R T I C L E
Zoster Vaccination Increases the Breadth ofCD4+ T Cells Responsive to Varicella ZosterVirus
Kerry J. Laing,1 Ronnie M. Russell,1 Lichun Dong,1 D. Scott Schmid,2 Michael Stern,3 Amalia Magaret,3,4,5 Jürgen G. Haas,6
Christine Johnston,1,5 Anna Wald,1,3,5,7 and David M. Koelle1,3,5,8,9
1Department of Medicine, University of Washington, Seattle; 2Centers for Disease Control and Prevention, Atlanta, Georgia; 3Department of LaboratoryMedicine, and 4Department of Biostatistics, University of Washington, and 5Vaccine and Infectious Diseases Division, Fred Hutchinson Cancer ResearchCenter, Seattle, Washington; 6Division of Infection and Pathway Medicine, University of Edinburgh, United Kingdom; 7Department of Epidemiology, and8Department of Global Health, University of Washington, and 9Benaroya Research Institute, Seattle, Washington
Background. The live, attenuated varicella vaccine strain (vOka) is the only licensed therapeutic vaccine. Boostof varicella zoster virus (VZV)–specific cellular immunity is a likely mechanism of action. We examined memoryCD4+ T-cell responses to each VZV protein at baseline and after zoster vaccination.
Methods. Serial blood samples were collected from 12 subjects vaccinated with Zostavax and immunogenicityconfirmed by ex vivo VZV-specific T-cell and antibody assays. CD4+ T-cell lines enriched for VZV specificity weregenerated and probed for proliferative responses to every VZV protein and selected peptide sets.
Results. Zoster vaccination increased the median magnitude (2.3-fold) and breadth (4.2-fold) of VZV-specificCD4+ T cells one month post-vaccination. Both measures declined by 6 months. The most prevalent responses atbaseline included VZV open reading frames (ORFs) 68, 4, 37, and 63. After vaccination, responses to ORFs 40, 67, 9,59, 12, 62, and 18 were also prevalent. The immunogenicity of ORF9 and ORF18 were confirmed using peptides,defining a large number of discrete CD4 T-cell epitopes.
Conclusions. The breadth and magnitude of the VZV-specific CD4+ T-cell response increase after zostervaccination. In addition to glycoprotein E (ORF68), we identified antigenic ORFs that may be useful componentsof subunit vaccines.
Keywords. epitopes; T lymphocytes; vaccine; varicella zoster virus.
Herpes zoster (shingles) is caused by reactivation of var-icella zoster virus (VZV). This alphaherpesvirus estab-lishes latent infection of neurons innervating skin andother structures during primary infection. Zoster typi-cally presents in a single dermatome as a unilateral,painful, vesicular rash. Chronic, long-lasting pain knownas postherpetic neuralgia (PHN) is common, may re-quire prolonged analgesic use, and can impair quality
of life. Other complications include encephalitis, ocularand cranial nerve dysfunction, and sometimes death.Higher risk for vascular diseases, such as stroke, tran-sient ischemic attack, and myocardial infarction, isalso observed postzoster [1, 2]. Disseminated infectionand complications are more likely in immunocompro-mised and older persons [3, 4].
Recent estimates of zoster rates in high-income coun-tries are 3.4 to 5.0 per 1000 person-years and 8 to 11 per1000 person-years in those ≥65 years old [3]. Zoster in-cidence appears to be rising [5]. Current strategies toprevent zoster rely on therapeutic vaccination of olderadults with a live attenuated (vOka) virus vaccine. Asthe only licensed immunotherapeutic vaccine, this rep-resents an important paradigm for boosting acquiredimmunity without stimulating immunopathology [6].The zoster vaccine is licensed for use in persons ≥50years and recommended for use in those ≥60 years by
Received 9 February 2015; accepted 6 March 2015.Presented in part: American Association of Immunology, Pittsburgh, Pennsylva-
nia, 2–6 May 2014.Correspondence: Kerry J. Laing, PhD, Department of Medicine, University of
Washington, 750 Republican St, Mail Stop 358061, Seattle, WA 98195-8061([email protected]).
The Journal of Infectious Diseases®
© The Author 2015. Published by Oxford University Press on behalf of the InfectiousDiseases Society of America. All rights reserved. For Permissions, please e-mail:[email protected]: 10.1093/infdis/jiv164
CD4+ T-cell Response to Shingles Vaccine • JID • 1
Journal of Infectious Diseases Advance Access published April 17, 2015 by guest on A
pril 19, 2016http://jid.oxfordjournals.org/
Dow
nloaded from

the Advisory Committee on Immunization Practices. Zostervaccination reduces risk for shingles (70% in persons aged50–59 years and 51%–55% in persons ≥60 years) and othercomplications, including PHN (59%–67%) [6–9], thus hasclear benefits yet is only partially protective in aged populations.In addition, as a live attenuated virus, it is not recommended forimmunocompromised persons [4, 9]. There is, therefore, a needfor alternative and improved vaccines.
Alternative vaccines in development include heat-treatedvOka [10] and an adjuvanted VZV glycoprotein E (gE) proteinsubunit [11, 12]. While safe and immunogenic, their protectiveefficacy is largely unknown. gE is a target of neutralizingantibodies, making a gE-subunit vaccine a rational choice forpreventing primary infection. It is also a known CD4+ T-cell an-tigen, but how gE compares to other VZV proteins with regardto cellular immunity is unknown. The current zoster vaccineboosts both arms of the adaptive immune system [13]. Fold in-crease in anti-VZV immunoglobin G (IgG), when accompaniedwith a high absolute titer, correlates with protection against zos-ter [14, 15]. Vaccine-induced T-cell levels may also correlatewith protection [16].
This study was conducted to more fully understand the na-ture of the immune boost from the licensed zoster vaccine. Themechanism(s) of action should be understood to improve effi-cacy, create a vaccine that is efficacious and safe for immune-compromised persons, and extend therapeutic vaccination toother chronic infections. We hypothesized that zoster vaccina-tion would increase the breadth of the T-cell response to VZVin addition to magnitude. By screening blood-derived T cells forreactivity to each VZV protein, an overall increase in CD4+ T-cell breadth was observed. Population-prevalent responses tonovel VZV-derived T-cell antigens were discovered and con-firmed to the peptide level. This information may benefit thedevelopment of future vaccines.
MATERIALS AND METHODS
Subjects and SpecimensTwelve VZV-seropositive adults age 50 or older (seronegativefor human immunodeficiency virus and hepatitis B and C)with no history of zoster or recent VZV exposure were vaccinat-ed. Zostavax was administered (day 0) as a single 0.65-mL dosesubcutaneously in the deltoid region of the upper arm. Bloodwas obtained before (day 0) and after (days 14, 28, 182) vaccina-tion. Additional blood was collected to obtain antigen-presentingcells (APCs). Peripheral blood mononuclear cells (PBMCs) wereisolated from acid citrate dextrose–anticoagulated blood by den-sity centrifugation and cryopreserved within 8 hours [17].Serum samples were collected for anti-VZV IgG titers by end-point glycoprotein enzyme-linked immunosorbent assay [18].Class II human leukocyte antigen (HLA) genotyping was per-formed using next-generation sequencing (Geraghty Lab, Fred
Hutchinson Cancer Research Center, Seattle, Washington) [19].The protocol was approved by the University of Washingtoninstitutional review board, and all participants signed a writteninformed consent.
Viral AntigensExpression constructs containing 71 sequence-verified VZVgenes or fragments were made as detailed elsewhere [20, 21]covering each known VZV open reading frame (ORF) [22].Most genes were from parental Oka and some from a clinicalVZV isolate (Seattle, Washington). Briefly, VZV genes were po-lymerase chain reaction–amplified using primers containingadaptors for cloning into pDONR207 or pDONR221 (Invitro-gen, Grand Island, New York) (Supplementary Table 1). Resul-tant pENTR constructs facilitated subcloning into pDEST203[21].VZV proteins were synthesized as N-terminal 6-His fusionsby cell-free Escherichia coli in vitro transcription translation(IVTT) (Expressway, Invitrogen) and expression confirmed bydot-blot Western via anti–6-His monoclonal antibody (mAb)(Roche, Indianapolis, Indiana) [21]. A negative control antigenset contained IVTT proteins from Francisella tularensis genes,empty pDEST203, or no DNA.
VZV was propagated on human embryonic tonsil (HET)cells. Vaccine Oka strain (vOka, Zostavax, Merck, West Point,Pennsylvania) was reconstituted per the manufacturer to ap-proximately 3 × 104 plaque-forming units (PFUs)/mL assuming19 400 PFU per dose as per the package insert. HET in Eagle’smodified essential medium (EMEM; Corning, Tewksbury,Massachusetts) with 10% fetal calf serum (FCS), 2 mM L-gluta-mine (Hyclone, Logan, Utah), and 1% penicillin/streptomycin(Hyclone) at 80% confluence were infected at a multiplicity ofinfection of 0.01. At 50%–60% cytopathic effect (CPE), trypsin-harvested cells were cocultured with uninfected HET 1:3 (in-fected:uninfected) to generate cell-associated virus, which wascryopreserved. For virus revival, thawed infected cells were over-laid onto uninfected subconfluent HET monolayers at 1:3.
For ultraviolet (UV)–inactivated VZV antigen (UV-VZV),80% confluent HET were overlaid with vOKA-infected HETat 1:6. At 50%–60% CPE, cells removed by scraping were re-suspended in complete EMEM, sonicated, and centrifuged(400g, 10 minutes). Supernatant (100 µL droplets) was exposedto UV light for 30 minutes, pooled, aliquoted, and frozen [23].Mock UV-HET antigen was made in parallel.
Ex Vivo AssaysEx vivo PBMC analyses used duplicate interferon (IFN)–γ–enzyme-linked immunospot (ELISPOT) and intracellular cytokinecytometry (ICC) for interleukin (IL)–2 and IFN-γ [24, 25]. ForELISPOT, overnight-rested PBMCs (5 × 105/well) in R10 (RoswellPark Memorial Institute 1640, 10% FCS, 2 mM L-glutamine, 1%penicillin/streptomycin) were added to anti-IFN-γ (1-D1K-γ;Mabtech, Cincinnati, Ohio)-coated multiscreen-Immobilon-P
2 • JID • Laing et al
by guest on April 19, 2016
http://jid.oxfordjournals.org/D
ownloaded from

ELISPOT plates (Millipore, Billerica, Massachusetts) with UV-VZV (1:100), UV-mock (1:100), medium, or phytohemagglutinin(PHA-P; 1.6 µg/mL; Remel, Kansas City, Kansas). Secreted IFN-γwas detected after 18 hours with 7-B6–1-biotin antibody, strepta-vidin-alkaline phosphatase (AP), and AP–conjugate (Bio-Rad,Hercules, California). Spot-forming units were counted (Immuno-spot, CTL, Shaker Heights, Ohio). For ICC, rested PBMCs (106
PBMCs/well) were incubated (18 hours) with T-cell medium(TCM), costimulatory antibodies (αCD28 and αCD49d; BD, SanJose, California) and antigens: UV-VZV or UV-mock (1:100), me-dium, or staphylococcal enterotoxin B (SEB, 2.5 µg/mL, Sigma)[26]. Brefeldin-A (Sigma) was added after 2 hours. After Live/Dead violet (Invitrogen) stain, cells were treated with fluores-cence-activated cell sorting (FACS) lyse/perm II (BD) and stainedwith: αCD3-PE (S4.1; Invitrogen), αCD4–fluorescein isothiocya-nate (S3.5; Invitrogen), αCD8-PerCP/Cy5.5 (SK1; BD), αIFN-γ-PE/Cy7 (4S.B3;BD),αIL-2-APC(MQ1-17H12;BD).Datacollectedon aBDFACSCanto II cytometerwere compensated/analyzedwithFlowJo (8.6.6; Treestar, Ashland, Oregon). Single liveCD3+CD4+CD8– lymphocytes were assessed for IFN-γ and IL-2.
VZV-reactive T-cell Lines and AssaysUV-VZV (1:200) was added to PBMCs (4 × 106/2 mL TCM).After 4 days (37oC/5% CO2), natural human interleukin-2(nIL-2; Hemagen, Columbia, Maryland) was added (32 U/mL)and replenished twice a week for 2 weeks. Enrichment forVZV reactivity was tested by ICC in 2.5 × 105 T cells combinedwith equal numbers of carboxyfluorescein succinimidylester (CFSE, Invitrogen)–stained autologous APCs [17]. Stainingand analysis was as described above, substituting αCD4-APC/H7(Clone RPA-T4; BD) and dump-gating CFSE-stained cells. VZV-reactivity-enriched T-cell lines were amplified for 2–3 weeks withanti-CD3 mAbs, recombinant IL-2 (Chiron, Emeryville, Califor-nia), and feeders [27], and cryopreserved. To detect responses toeach VZV protein or fragment, a 3H-thymidine incorporationassay was used [28]. Briefly, 105 VZV-enriched T cells were com-bined with 105 irradiated autologous PBMCs. Antigens includedIVTT products (1:2000, duplicates) and whole UV-VZV or UV-mock (1:100, 6 replicates). 3H-thymidine (0.5 µCi, Perkin-Elmer,Boston, Massachusetts) was added at 72 hours, cells harvested 24hours later, and counts per minute determined (TopCount NXT,Perkin-Elmer).
Epitope MappingPeptides (13-mers, overlapping by 9 amino acids) were synthe-sized for VZV ORF9 and ORF18 (Genscript, Piscataway, NewJersey), reconstituted in dimethyl sulfoxide (DMSO), and arrayedin matrices to form pools of ≤12 peptides with each peptide in 2pools. VZV-enriched T cells (105) and autologous irradiatedPBMCs (105) were tested with peptide pools (1 µg/mL eachpeptide), DMSO, medium, or PHA-P (1.6 µg/mL). Positivepools were deconvoluted to identify antigenic peptide(s) [28].
Confirmation included dose-response (serial 10-fold dilutionsof peptide beginning at 1 µg/mL) analysis of single peptides intriplicate using a representative response-confirmed cell lineper subject.
StatisticsPositive response to VZV proteins in lymphoproliferation as-says was determined on a per-cell-line basis surpassing thisthreshold: at least 2.33 × median absolute deviation of all nega-tive-control antigen samples above the median response ofthose same samples [29]. This calculation has a theoreticalfalse-positive rate of 1.0%. VZV antigen wells were scored pos-itive only if both replicates exceeded the threshold. ORFs syn-thesized as fragments (eg, ORF22) were scored positive once ifany fragment was deemed positive.
Antibody titers, T-cell frequencies, and T-cell breadth werecompared between the different time points using the nonpara-metric Friedman’s test for repeated measures. If significant, sub-sequent analysis of measures using Dunn’s test identified whichtime points were different from prevaccine samples. Correlationanalyses were performed using the Spearman’s test. Nonpara-metric tests were used because the data were highly left skewed,thus did not satisfy assumptions of normality. Two-sided P val-ues <.05 were considered statistically significant. Statistical testsand graphs were generated using Prism (v6.04, GraphPad Soft-ware, La Jolla, California).
RESULTS
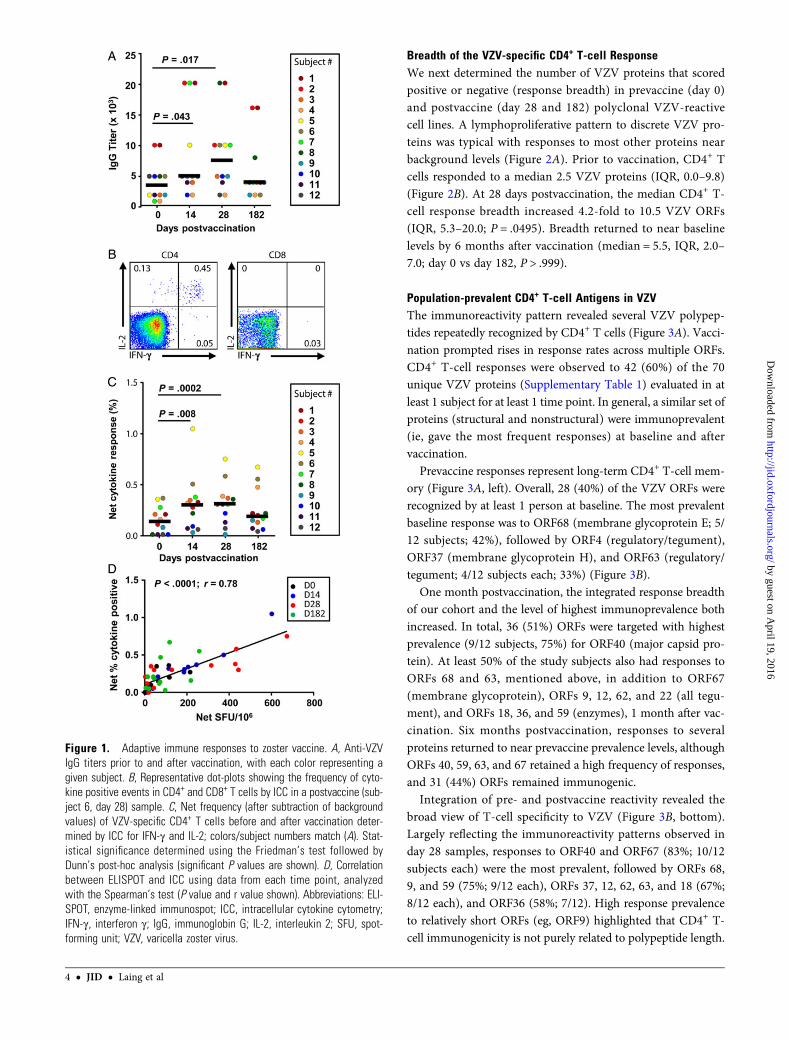
Immunogenicity Ex VivoWe enrolled 12 subjects whomet the Food and Drug Administra-tion indication for zoster vaccination. Among these, 8 (67%) weremen, 10 (83%) werewhite, andmedian agewas 56 (range, 51–67).We first confirmed that each subject was VZV IgG seropositive atbaseline (median titer = 3500; interquartile range [IQR], 2000–5000). Boosting of adaptive immune responses with vaccinationwas readily observed. Postvaccine median anti-VZV IgG titers in-creased 1.4-fold by day 14 (5000; IQR, 4000–17 500; P = .043) and2.1-fold by day 28 (7500; IQR, 4250–10 000; P = .017), returningto 1.1-fold above baseline by 6 months (4000; IQR, 2500–7000;P = .805) (Figure 1A). Ex vivo ICC, verified T-cell reactivity toUV-VZV was boosted and was mediated via CD3+CD4+ cells,but not CD3+CD8+ cells (Figure 1B). Median postvaccine IL-2or IFN-γ responses were >2-fold higher than baseline levels(0.13%; IQR, 0.0–0.25) at day 14 (0.29%; IQR, 0.07–0.36;P = .008) and day 28 (0.30%; IQR, 0.14–0.38; P = .0002), but notat day 182 (0.18%; IQR, 0.08–0.41; P = .173) (Figure 1C). Resultsfor ELISPOT generally mirrored and correlated with theCD3+CD4+ ICC data (net response, r = 0.782; P < .0001) (Fig-ure 1D). Neither ICC (r = 0.206; P = .16) nor ELISPOT(r = 0.120; P = .42) CD4+ T-cell endpoints were correlated withIgG titers (see also Supplementary Table 2).
CD4+ T-cell Response to Shingles Vaccine • JID • 3
by guest on April 19, 2016
http://jid.oxfordjournals.org/D
ownloaded from
Breadth of the VZV-specific CD4+ T-cell ResponseWe next determined the number of VZV proteins that scoredpositive or negative (response breadth) in prevaccine (day 0)and postvaccine (day 28 and 182) polyclonal VZV-reactivecell lines. A lymphoproliferative pattern to discrete VZV pro-teins was typical with responses to most other proteins nearbackground levels (Figure 2A). Prior to vaccination, CD4+ Tcells responded to a median 2.5 VZV proteins (IQR, 0.0–9.8)(Figure 2B). At 28 days postvaccination, the median CD4+ T-cell response breadth increased 4.2-fold to 10.5 VZV ORFs(IQR, 5.3–20.0; P = .0495). Breadth returned to near baselinelevels by 6 months after vaccination (median = 5.5, IQR, 2.0–7.0; day 0 vs day 182, P > .999).
Population-prevalent CD4+ T-cell Antigens in VZVThe immunoreactivity pattern revealed several VZV polypep-tides repeatedly recognized by CD4+ T cells (Figure 3A). Vacci-nation prompted rises in response rates across multiple ORFs.CD4+ T-cell responses were observed to 42 (60%) of the 70unique VZV proteins (Supplementary Table 1) evaluated in atleast 1 subject for at least 1 time point. In general, a similar set ofproteins (structural and nonstructural) were immunoprevalent(ie, gave the most frequent responses) at baseline and aftervaccination.
Prevaccine responses represent long-term CD4+ T-cell mem-ory (Figure 3A, left). Overall, 28 (40%) of the VZV ORFs wererecognized by at least 1 person at baseline. The most prevalentbaseline response was to ORF68 (membrane glycoprotein E; 5/12 subjects; 42%), followed by ORF4 (regulatory/tegument),ORF37 (membrane glycoprotein H), and ORF63 (regulatory/tegument; 4/12 subjects each; 33%) (Figure 3B).
One month postvaccination, the integrated response breadthof our cohort and the level of highest immunoprevalence bothincreased. In total, 36 (51%) ORFs were targeted with highestprevalence (9/12 subjects, 75%) for ORF40 (major capsid pro-tein). At least 50% of the study subjects also had responses toORFs 68 and 63, mentioned above, in addition to ORF67(membrane glycoprotein), ORFs 9, 12, 62, and 22 (all tegu-ment), and ORFs 18, 36, and 59 (enzymes), 1 month after vac-cination. Six months postvaccination, responses to severalproteins returned to near prevaccine prevalence levels, althoughORFs 40, 59, 63, and 67 retained a high frequency of responses,and 31 (44%) ORFs remained immunogenic.
Integration of pre- and postvaccine reactivity revealed thebroad view of T-cell specificity to VZV (Figure 3B, bottom).Largely reflecting the immunoreactivity patterns observed inday 28 samples, responses to ORF40 and ORF67 (83%; 10/12subjects each) were the most prevalent, followed by ORFs 68,9, and 59 (75%; 9/12 each), ORFs 37, 12, 62, 63, and 18 (67%;8/12 each), and ORF36 (58%; 7/12). High response prevalenceto relatively short ORFs (eg, ORF9) highlighted that CD4+ T-cell immunogenicity is not purely related to polypeptide length.
Figure 1. Adaptive immune responses to zoster vaccine. A, Anti-VZVIgG titers prior to and after vaccination, with each color representing agiven subject. B, Representative dot-plots showing the frequency of cyto-kine positive events in CD4+ and CD8+ T cells by ICC in a postvaccine (sub-ject 6, day 28) sample. C, Net frequency (after subtraction of backgroundvalues) of VZV-specific CD4+ T cells before and after vaccination deter-mined by ICC for IFN-γ and IL-2; colors/subject numbers match (A). Stat-istical significance determined using the Friedman’s test followed byDunn’s post-hoc analysis (significant P values are shown). D, Correlationbetween ELISPOT and ICC using data from each time point, analyzedwith the Spearman’s test (P value and r value shown). Abbreviations: ELI-SPOT, enzyme-linked immunospot; ICC, intracellular cytokine cytometry;IFN-γ, interferon γ; IgG, immunoglobin G; IL-2, interleukin 2; SFU, spot-forming unit; VZV, varicella zoster virus.
4 • JID • Laing et al
by guest on April 19, 2016
http://jid.oxfordjournals.org/D
ownloaded from

Confirmation of Responses to VZV ORF9 and ORF18 by PeptideEpitope MappingVZV ORF9, orthologous to tegument protein herpes simplexvirus (HSV) UL49, and ORF18, the homolog of HSV UL40(both known T-cell antigens) [30, 31], were selected for peptideconfirmation of T-cell reactivity. The selection was based onnovelty as VZV T-cell antigens, high CD4+ T-cell immunopreva-lence, and short lengths (302 and 306 amino acids, respectively).Using a matrix peptide pooling approach, a discrete pattern ofpositive and negative responses was observed in the polyclonalVZV-reactive responder cell lines (Figure 4A). Candidatesingle-antigenic peptides identified at the intersections of the pos-itive row and column pools were retested. Reactive and nonreac-tive peptides were clearly differentiated at 1 µg/mL (Figure 4B).Dose-response studies in serial 10-fold dilutions showed thatmost peptides remained stimulatory at 0.1 µg/mL, and lost activ-ity below 0.01 µg/mL (data not shown). Peptide-specific respons-es were detected in all but 1 person among persons with reactivityto ORF9 (n = 9) and ORF18 (n = 8) proteins. Most persons hadmultiple active peptides within ORF9 and ORF18.
We identified 13 antigenic peptides in ORF9 and 14 antigenicpeptides in ORF18 (Figure 4C). Reactivity was regularly observedagainst 2 or more overlapping peptides, such as ORF1853–65 andORF1857–69 for subject 2. Even using a conservative approach inwhich 2 overlapping 13-amino-acid peptides are counted as 1epitope, we noted multiple epitopes within several individuals test-ed for ORF9 (median 2, max 5) and ORF18 (median 1.5, max 7).Several 13-mers were immunogenic in multiple persons (eg,ORF18169–181 in subjects 6, 8, and 10) with shared HLA types—these persons share the HLA-DPB1*04:01 allele but not HLA-DRor -DQ alleles (Supplementary Table 3). Reactivity of subjects 8
and 10, but not subject 6, to the overlapping peptide ORF18165–177suggests a potential heterogeneous response to this region. Somepeptides appear to be relatively promiscuous T-cell activators;for example, ORF9121−133 activated T cells in subjects 3, 7, and8, and ORF9185−197 activated T cells in subjects 2, 6, 7, 8, and 9,among whom there was no consensus HLA allelic variant. No se-quence variations were noted in the epitope-containing regions(Figure 4C) upon examination of VZV sequences in GenBank(2015).
DISCUSSION
In the present report, we evaluated the CD4 T-cell response tozoster vaccination at the level of individual VZV proteins andpeptide epitopes. Antigen-specific responses were readily de-tectable prior to vaccination, suggesting that memory to child-hood varicella is durable, albeit boosting by periodic subclinicalreactivation or exposure, or cross-reactive antigens [32] is alsopossible. Applying a standardized protocol to blood obtainedbefore and after vaccination, we confirmed previous estimatesof the overall antibody and CD4 T-cell boost with vaccination[15, 33] and showed that the ORF level breadth of the CD4T-cell responses approximately quadrupled at 1 month, whichdeclined by 6 months postvaccination theoretically via T-cellrecruitment from blood to the skin or lymphoid tissue, loss dur-ing contraction, or negative regulation (eg, T-regs [34]). In ad-dition to the subunit vaccine candidate antigen ORF68 (gE)currently in clinical development [35], we uncovered severalother population-prevalent CD4 T-cell antigens from VZVthat are, therefore, rational for investigation as vaccine immu-nogens. Little is known about the fine specificity or phenotype
Figure 2. CD4+ T-cell responses to VZV increase in breadth after vaccination. A, Representative plot showing reactivity of a bulk VZV-reactive CD4+ T-cellline to VZV and control proteins (duplicate wells), and to whole UV-treated VZV (UV-VZV) or HET (UV-Mock) antigens. Bars represent mean CPM and errorbars represent standard deviation. The threshold for positivity—calculated as (median response of all negative control antigens) + (2.33 times medianabsolute deviation of all negative control antigens)—is represented by a dashed horizontal line. Both duplicates needed to exceed the threshold forthe ORF to be determined positive. Positive ORFs are named. B, The number of VZV ORFs recognized by CD4+ T-cell lines enriched for reactivity toVZV from samples of 12 study subjects obtained before (D0), 1 (D28), and 6 months (D128) after vaccination. Horizontal bars represent the median ofeach time point. Each subject is shown with 1 color. Statistical significance determined using the Friedman’s test followed by Dunn’s post-hoc analysis(significant P values are shown). Abbreviations: CPM, counts per minute; HET, human embryonic tonsil; ORF, open reading frame; UV, ultraviolet; VZV,varicella zoster virus.
CD4+ T-cell Response to Shingles Vaccine • JID • 5
by guest on April 19, 2016
http://jid.oxfordjournals.org/D
ownloaded from

Figure 3. CD4+ T-cell responses of adult zoster vaccine recipients before (Day 0) and after (Day 28 and Day 182) vaccination. A, VZV ORFs elicitingresponses within polyclonal VZV-specific T-cell lines are indicated by colored boxes. Each column represents a study subject with colors consistent ateach time point; colors/subject numbers match Figure 1A and 1C. Each row represents a VZV ORF. Ranking is from highest to lowest overall responsefrequency. An additional 29 ORFs without responses detected are not shown. B, Frequency of protein-specific responses among zoster vaccine recipientsat before (Day 0) and after (Day 28 and Day 182) vaccination, or at any time point (bottom). Abbreviations: ORFs, open reading frames; VZV, varicella zostervirus.
6 • JID • Laing et al
by guest on April 19, 2016
http://jid.oxfordjournals.org/D
ownloaded from

of VZV-specific T-cell responses [30, 31]. The few CD4 T-cellepitopes, documented in literature or a central database [36]prior to our study, at the well-defined level of short peptides
and HLA class II–restricting alleles were limited to ORFs 4,31, 62, 63, 67, and 68 [30, 37–40]. We greatly expanded thenumber of known VZV-specific CD4 T-cell epitopes with our
Figure 4. Peptides in VZV ORFs 9 and 18 recognized by polyclonal VZV-reactive CD4+ T-cell lines. A, Representative data showing reactivity to ORF9peptide pools for subject 3 at day 28. Each 13-amino-acid peptide was present in 1 row pool and 1 column pool. B, Hits in row and column pools weredeconvoluted by screening each candidate reactive peptide present in both a reactive row and reactive column pool at 1 µg/mL. C, VZV ORFs 9 and 18 CD4+
T-cell epitopes confirmed by single-peptide reactivity. Numbers represent subjects 1–12 as in Figure 3A. Shaded columns represent subjects that had noresponse to the full-length ORF. Black boxes indicate subject reactivity to listed peptides. Abbreviations: CPM, counts per minute; ORFs, open readingframes; VZV, varicella zoster virus.
CD4+ T-cell Response to Shingles Vaccine • JID • 7
by guest on April 19, 2016
http://jid.oxfordjournals.org/D
ownloaded from

studies, targeting VZVORFs 9 and 18, and provide a clear path-way to define epitopes in detail from any ORF reactive in oursystem.
Although we defined T-cell responses in blood, relevantmemory immune responses to VZV could reside in susceptibletissues: zoster can attack any skin site, the liver, retina, middleear, and large arteries, and can cause devastating noncutaneoussyndromes [41]. This clinical variety is explained by VZV latentinfection in and reactivation from extensive sensory or auto-nomic ganglia [42], although VZV reactivates rarely [43]. Therelated alphaherpesviruses, HSV-1 and HSV-2, reactivatemore frequently, provoke antigen-specific T-cell influx to sitesof recurrent infection, and lead to persistent deposition of tissueresident–memory (TRM) T cells [44–46].We have yet to confirmthat zoster vaccine–invoked T cells have chemokine and adhe-sion receptors suitable for migration to relevant tissues or thatVZV-specific TRM persist in ganglia or skin. However, circulat-ing VZV-specific CD4 T cells with a CD45RO+ TCM (centralmemory) pattern—expressing CD27 and CD28 for costimula-tion, CCR7 and variable CD62L for lymph node homing, butlow cutaneous lymphocyte-associated antigen for skin hom-ing—have been described, some of which show signs of recentantigen exposure (CD38+) and possible T-cell exhaustion(PD1+) [47, 48]. Analogous to our proteome-wide HSV studies[27, 45], the VZV tools described herein provide opportunitiesto study acute zoster lesion-infiltrating lymphocytes and TRM.Current efforts to define the HLA restriction of our epitopeswill facilitate tetramer-based study of VZV-specific T-cellphenotype.
Limitations of this study include the relatively small number ofsubjects and time points studied and the lack of vaccine clinicalefficacy data in these subjects. However, repositories from phaseIII trials could provide suitable large cohort specimens for clin-ical end-point correlative studies, for which our methods couldbe suitably simplified and scaled up. We used cell-associatedVZV antigen to purposefully restimulate T cells reactive withboth virion and nonvirion proteins, and the prevalence of re-sponses to enzymes such as ORF18 showed we were successful.However, it is possible that restimulation and expansion of VZV-specific memory CD4 T cells was not unbiased, such that someantigenic specificity was under- or overrepresented, or extremelyrare responses were missed. Nonetheless, we did reliably demon-strate increased response breadth to vaccination.
As a relatively recent vaccination program, insufficient dataexist regarding longevity of protection from the current zostervaccine, although protection is still evident at 7 years with anapparent decline over time [7]. Consistent with our presentedobservations, short-term immunogenicity of zoster vaccination(in clinical trials) approximate a 2-fold boost in both T-cell andB-cell response magnitude, which declines to levels moderatelyabove prevaccine levels within a year [49]; the declines we ob-serve in magnitude and response breadth may explain reduced
protection over time. Although not confirmed mechanistic cor-relates of protection, vaccinees with both a high fold-rise andhigh endpoint titer of anti-VZV IgG after vaccination haveless risk for zoster [15]. A correlation between T-cell boostingand protection has yet to be reported. However, T cells shouldnot be overlooked in the context of vaccination because they areessential in VZV control in the absence of vaccination. Lack ofcorrelation in VZV-specific T-cell magnitude and antibody titerobserved during our investigation (r = 0.206; P = .16) and earlierstudies suggest both arms of adaptive immunity have indepen-dent relationships with zoster, making antibody rises an unlike-ly biomarker for T-cell boosts (or vice versa). We did note arelationship between ex vivo magnitude of VZV-reactive Tcells and response breadth (r = 0.58; P = .0002), suggestingthat dominant responses to single ORFs were not the drivingforce for T-cell elevation following vaccination. Consequently,CD4 T-cell magnitude may serve as a good surrogate for pre-dicting breadth, albeit it is not known if the number, breadth,specificity, or phenotype of CD4 T cells is the most importantfor protection from, or resolution of, zoster or its complications.
In conclusion, the CD4 T-cell response to VZV is polyspe-cific, both before and after therapeutic vaccination of seroposi-tive adults, with myriad VZV proteins documented for the firsttime to elicit specific immunity. Our data support the hypoth-esis that zoster vaccination works at least in part by increasingthe magnitude and breadth of the T-cell response to VZV.
Supplementary Data
Supplementary materials are available at The Journal of Infectious Diseasesonline (http://jid.oxfordjournals.org). Supplementary materials consist ofdata provided by the author that are published to benefit the reader. Theposted materials are not copyedited. The contents of all supplementarydata are the sole responsibility of the authors. Questions or messages regard-ing errors should be addressed to the author.
Notes
Acknowledgments. We thank the research subjects, the staff of the Uni-versity of Washington Virology Research Clinic, and Anne Cent and Rose-mary Obrigewitch for virology assistance.Disclaimer. The views included in this report are those of the authors
and do not necessarily represent the views of the Centers for Disease Controland Prevention.Financial support. This work was supported by the University of Wash-
ington Royalty Research Fund and National Institutes of Health grantsR01AI094019, P01AI30731, and UL1TR000423, and contract HHSN272201400049C.Potential conflicts of interest. All authors: No reported conflicts.All authors have submitted the ICMJE Form for Disclosure of Potential
Conflicts of Interest. Conflicts that the editors consider relevant to the con-tent of the manuscript have been disclosed.
References
1. Breuer J, Pacou M, Gautier A, Brown MM. Herpes zoster as a risk factorfor stroke and TIA: a retrospective cohort study in the UK. Neurology2014; 83:e27–33.
8 • JID • Laing et al
by guest on April 19, 2016
http://jid.oxfordjournals.org/D
ownloaded from

2. Langan SM, Minassian C, Smeeth L, Thomas SL. Risk of stroke follow-ing herpes zoster: a self-controlled case-series study. Clin Infect Dis2014; 58:1497–503.
3. WHO. Varicella and herpes zoster vaccines: WHO position paper, June2014. Wkly Epidemiol Rec 2014; 89:265–87.
4. Weaver BA. The burden of herpes zoster and postherpetic neuralgia inthe United States. J Am Osteopath Assoc 2007; 107:S2–7.
5. Goldman GS, King PG. Review of the United States universal varicellavaccination program: herpes zoster incidence rates, cost-effectiveness,and vaccine efficacy based primarily on the Antelope Valley VaricellaActive Surveillance Project data. Vaccine 2013; 31:1680–94.
6. Oxman MN, Levin MJ, Johnson GR, et al. A vaccine to prevent herpeszoster and postherpetic neuralgia in older adults. N Engl J Med 2005;352:2271–84.
7. Schmader KE, Levin MJ, Gnann JW Jr, et al. Efficacy, safety, and toler-ability of herpes zoster vaccine in persons aged 50–59 years. Clin InfectDis 2012; 54:922–8.
8. Tseng HF, Smith N, Harpaz R, Bialek SR, Sy LS, Jacobsen SJ. Herpeszoster vaccine in older adults and the risk of subsequent herpes zosterdisease. JAMA 2011; 305:160–6.
9. Langan SM, Smeeth L, Margolis DJ, Thomas SL. Herpes zoster vaccineeffectiveness against incident herpes zoster and post-herpetic neuralgiain an older US population: a cohort study. PLOSMed 2013; 10:e1001420.
10. Mullane KM, Winston DJ, Wertheim MS, et al. Safety and immunoge-nicity of heat-treated zoster vaccine (ZVHT) in immunocompromisedadults. J Infect Dis 2013; 208:1375–85.
11. Leroux-Roels I, Leroux-Roels G, Clement F, et al. A phase 1/2 clinicaltrial evaluating safety and immunogenicity of a varicella zoster glyco-protein e subunit vaccine candidate in young and older adults. J InfectDis 2012; 206:1280–90.
12. Chlibek R, Bayas JM, Collins H, et al. Safety and immunogenicity of anAS01-adjuvanted varicella-zoster virus subunit candidate vaccineagainst herpes zoster in adults >=50 years of age. J Infect Dis 2013;208:1953–61.
13. Patterson-Bartlett J, Levin MJ, Lang N, Schodel FP, Vessey R, WeinbergA. Phenotypic and functional characterization of ex vivo T cell responsesto the live attenuated herpes zoster vaccine. Vaccine 2007; 25:7087–93.
14. Levin MJ, Schmader KE, Gnann JW, et al. Varicella-zoster virus-specificantibody responses in 50–59-year-old recipients of zoster vaccine. J In-fect Dis 2013; 208:1386–90.
15. Gilbert PB, Gabriel EE, Miao X, et al. Fold rise in antibody titers bymeasured by glycoprotein-based enzyme-linked immunosorbent assayis an excellent correlate of protection for a herpes zoster vaccine, dem-onstrated via the vaccine efficacy curve. J Infect Dis 2014; 210:1573–81.
16. Levin MJ, Oxman MN, Zhang JH, et al. Varicella-zoster virus-specificimmune responses in elderly recipients of a herpes zoster vaccine. J In-fect Dis 2008; 197:825–35.
17. Laing KJ, Magaret AS, Mueller DE, et al. Diversity in CD8(+) T cellfunction and epitope breadth among persons with genital herpes.J Clin Immunol 2010; 30:703–22.
18. Weinmann S, Chun C, Mullooly JP, et al. Laboratory diagnosis andcharacteristics of breakthrough varicella in children. J Infect Dis2008; 197(Suppl 2):S132–8.
19. Smith AG, Pyo CW, Nelson W, et al. Next generation sequencing to de-termine HLA class II genotypes in a cohort of hematopoietic cell trans-plant patients and donors. Hum Immunol 2014; 75:1040–6.
20. Ceroni A, Sibani S, Baiker A, et al. Systematic analysis of the IgG anti-body immune response against varicella zoster virus (VZV) using a self-assembled protein microarray. Mol Biosyst 2010; 6:1604–10.
21. Jing L, Haas J, Chong TM, et al. Cross-presentation and genome-widescreening reveal candidate T cells antigens for a herpes simplex virustype 1 vaccine. J Clin Invest 2012; 122:654–73.
22. Cohen JI. The varicella-zoster virus genome. Curr Top Microbiol Im-munol 2010; 342:1–14.
23. Koelle DM, Corey L, Burke RL, et al. Antigenic specificities of humanCD4+ T-cell clones recovered from recurrent genital herpes simplexvirus type 2 lesions. J Virol 1994; 68:2803–10.
24. Braun RP, Payne LG, Dong L. Characterization of the IFN-gamma T-cell responses to immediate early antigens in humans with genital her-pes. Virol J 2006; 3:54.
25. Moss NJ, Magaret A, Laing KJ, et al. Peripheral blood CD4 T-cell andplasmacytoid dendritic cell (pDC) reactivity to herpes simplex virus 2and pDC number do not correlate with the clinical or virologic severityof recurrent genital herpes. J Virol 2012; 86:9952–63.
26. Koelle DM. Expression cloning for the discovery of viral antigens andepitopes recognized by T-cells. Methods 2003; 29:213–26.
27. Koelle DM, Chen HB, Gavin MA, Wald A, Kwok WW, Corey L. CD8CTL from genital herpes simplex lesions: recognition of viral tegumentand immediate early proteins and lysis of infected cutaneous cells. J Im-munol 2001; 166:4049–58.
28. Jing L, McCaughey SM, Davies DH, et al. ORFeome approach to theclonal, HLA allele-specific CD4 T-cell response to a complex pathogenin humans. J Immunol Methods 2009; 347:36–45.
29. Chung N, Zhang XD, Kreamer A, et al. Median absolute deviation toimprove hit selection for genome-scale RNAi screens. J Biomol Screen2008; 13:149–58.
30. Ouwendijk WJ, Laing KJ, Verjans GM, Koelle DM. T-cell immunity tohuman alphaherpesviruses. Curr Opin Virol 2013; 3:452–60.
31. Laing KJ, Dong L, Sidney J, Sette A, Koelle DM. Immunology in theClinic Review Series; focus on host responses: T cell responses to herpessimplex viruses. Clin Exp Immunol 2012; 167:47–58.
32. Ouwendijk WJ, Geluk A, Smits SL, Getu S, Osterhaus AD, Verjans GM.Functional characterization of ocular-derived human alphaherpesviruscross-reactive CD4 T cells. J Immunol 2014; 192:3730–9.
33. Weinberg A, Levin MJ. VZV T cell-mediated immunity. In: AbendrothA, Arvin AM, Moffat JF, eds. Varicella-zoster virus. vol. current topicsin microbiology and immunolog. Vol 342. Berlin: Springer, 2010:341–57.
34. Vukmanovic-Stejic M, Sandhu D, Sobande TO, et al. Varicella zoster-specific CD4+Foxp3+ T cells accumulate after cutaneous antigen chal-lenge in humans. J Immunol 2013; 190:977–86.
35. Berkowitz EM, Moyle G, Stellbrink HJ, et al. Safety and immunogenicityof an adjuvanted herpes zoster subunit candidate vaccine in HIV-infect-ed adults: a phase 1/2a randomized, placebo-controlled study. J InfectDis 2014; doi:10.1093/infdis/jiu606.
36. Kleemann P, Distler E, Wagner EM, et al. Varicella-zoster virus glyco-proteins B and E are major targets of CD4+ and CD8+ T cells reconsti-tuting during zoster after allogeneic transplantation. Haematologica2012; 97:874–82.
37. Jones L, Black AP, Malavige GN, Ogg GS. Persistent high frequencies ofvaricella-zoster virus ORF4 protein-specific CD4+ T cells after primaryinfection. J Virol 2006; 80:9772–8.
38. Vita R, Zarebski L, Greenbaum JA, et al. The immune epitope database2.0. Nucleic Acids Res 2010; 38:D854–62.
39. Huang Z, Vafai A, Lee J, Mahalingam R, Hayward AR. Specific lysis oftargets expressing varicella-zoster virus gpI or gpIV by CD4+ humanT-cell clones. J Virol 1992; 66:2664–9.
40. Sadzot-Delvaux C, Arvin AM, Rentier B. Varicella-zoster virus IE63, avirion component expressed during latency and acute infection, elicitshumoral and cellular immunity. J Infect Dis 1998; 178(Suppl 1):S43–7.
41. Arvin AM, Gilden D. Varicella-zoster virus. In: Knipe DM, Howley PM,Cohen JI, Griffin DE, Lamb RA, Martin MA, Racaniello VR, RoizmanB, eds. Fields Virology. Vol. 2. Philadelphia: Wolters Kluwer/LippincottWilliams and Wilkins, 2013:2015–57.
42. Kennedy PG, Cohrs RJ. Varicella-zoster virus human ganglionic laten-cy: a current summary. J Neurovirol 2010; 16:411–8.
43. Levin MJ. Varicella-zoster virus and virus DNA in the blood and oro-pharynx of people with latent or active varicella-zoster virus infections.J Clin Virol 2014; 61:487–95.
44. Zhu J, Peng T, Johnston C, et al. Immune surveillance byCD8alphaalpha+ skin-resident T cells in human herpes virus infection.Nature 2013; 497:494–7.
45. van Velzen M, Jing L, Osterhaus AD, Sette A, Koelle DM, Verjans GM.Local CD4 and CD8 T-cell reactivity to HSV-1 antigens documents
CD4+ T-cell Response to Shingles Vaccine • JID • 9
by guest on April 19, 2016
http://jid.oxfordjournals.org/D
ownloaded from

broad viral protein expression and immune competence in latently in-fected human trigeminal ganglia. PLOS Pathog 2013; 9:e1003547.
46. Koelle DM, Posavad CM, Barnum GR, Johnson ML, Frank JM, Corey L.Clearance of HSV-2 from recurrent genital lesions correlates with infiltrationof HSV-specific cytotoxic T lymphocytes. J Clin Invest 1998; 101:1500–8.
47. Jones L, Black AP, Malavige GN, Ogg GS. Phenotypic analysis of humanCD4+ T cells specific for immediate-early 63 protein of varicella-zostervirus. Eur J Immunol 2007; 37:3393–403.
48. Malavige GN, Jones L, Black AP, Ogg GS. Varicella zoster virus glyco-protein E-specific CD4+ T cells show evidence of recent activation andeffector differentiation, consistent with frequent exposure to replicativecycle antigens in healthy immune donors. Clin Exp Immunol 2008;152:522–31.
49. Vermeulen JN, Lange JM, Tyring SK, et al. Safety, tolerability, and im-munogenicity after 1 and 2 doses of zoster vaccine in healthy adults>/=60 years of age. Vaccine 2012; 30:904–10.
10 • JID • Laing et al
by guest on April 19, 2016
http://jid.oxfordjournals.org/D
ownloaded from
Copyright © 2022 FDOKUMEN

















![[ICPP'14] Yang You et al., Designing a Heuristic Cross-Architecture Combination for Breadth-First Search, 43rd International Conference on Parallel Processing](https://static.fdokumen.com/doc/165x107/631ade94d5372c006e03b0ab/icpp14-yang-you-et-al-designing-a-heuristic-cross-architecture-combination.jpg)


