
Living outside the box: An updated perspective on diet breadth and sexual division of labor in the...
12
Living outside the box: An updated perspective on diet breadth and sexual division of labor in the Prearchaic Great Basin R.G. Elston a, * , D.W. Zeanah b , B.F. Codding c a Department of Anthropology, University of Nevada, Reno,1664 N. Virginia Street, Reno, NV 89557-0208, USA b Department of Anthropology, California State University Sacramento, 6000 J Street, Sacramento, CA 95819-6106, USA c Department of Anthropology, University of Utah, 270 S.1400 E., Rm. 102, Salt Lake City, UT 84112, USA article info Article history: Available online 7 November 2014 Keywords: Human behavioral ecology Division of labor Patch choice Marginal value theorem PleistoceneeHolocene Transition Western stemmed point tradition abstract A tremendous amount has been learned about the Prearchaic (before 9000 BP) Great Basin since we advocated a perspective of sexual division of labor based on Human Behavioral Ecology a decade ago. Many investigators have taken our advice and a few have challenged our assumptions and inferences. One of the most substantive critiques has been that we misunderstood the paleoenvironmental pa- rameters of ungulate populations during the PleistoceneeHolocene Transition (PHT). Simultaneously, behavioral ecologists have advanced our understanding of sexual division of labor among modern for- agers, but these studies appear to have gone unnoticed by Great Basin prehistorians. We review findings of the last ten years and suggest that the key to understanding patterning in the PHT still relies on understanding (a) variability in men's and women's foraging goals, (b) the abundance and distribution of large prey, (c) how changing environmental parameters effect both the division of labor and the dis- tribution of resources, and (d) the relative influence of search and handling costs on residence time in PHT wetlands. We suggest that consideration of how paleoenvironmental variability structured sexual division of labor remains key to fully understanding Prearchaic lifeways in the Great Basin. © 2014 Elsevier Ltd and INQUA. All rights reserved. 1. Introduction Over ten years ago, Elston and Zeanah (2002) offered an explanation for the seemingly contradictory combination of traits observed in Great Basin archaeological sites dating to the Pleisto- ceneeHolocene Transition (PHT): evidence for high mobility, lithic technology adapted to hunting, and faunal assemblages suggesting broad spectrum foraging. The paper compared applications of various behavioral ecological models (Simms, 1987; Raven and Elston, 1989; Zeanah et al., 1995; Pinson, 1999), using the simu- lated resource structure of Railroad Valley, Nevada (Zeanah et al., 1999) for the early and middle Holocene. Drawing from ethno- graphic studies of diet and patch choice, we proposed that the observed patterns of site location, abundance and content resulted from the different foraging goals and strategies of men and women in the environmental context of the PHT Great Basin. This Pre- archaic pattern differed fundamentally from subsequent Archaic (post 9000 BP) lifeways. Responses were mostly favorable, although there were some mis- interpretations of the piece. Here we clarify the model by updating the original framework with new insights from human behavioral ecology (HBE) and a better understanding of the availability of large prey (ar- tiodactyls). Specifically, we highlight that (1) the key to understanding patterning in the PHT still hinges on understanding variability in men's and women's foraging goals and the resulting division of labor; (2) while some suggest that large prey were too uncommon to have been crucial resources for Prearchaic Great Basin foragers, the concentration of artiodactyls around wetlands actually made hunting large game predictable and reliable as long as foragers were free to move from one wetland basin to another; (3) understanding both the division of labor and the patchiness of large prey provides a clear framework for framing PHT foraging, diminishing returns and how these effected forager mobility and site formation patterns. Before detailing our model of PHT (Prearchaic) lifeways, we first review insights from 2002 and compare how they stand up to additional findings over the last 10 years. 1.1. The PHT record: key observations Elston and Zeanah (2002) made a series of key empirical ob- servations, most of which have held up to subsequent * Corresponding author. E-mail addresses: [email protected], [email protected] (R.G. Elston), [email protected] (D.W. Zeanah), [email protected] (B.F. Codding). Contents lists available at ScienceDirect Quaternary International journal homepage: www.elsevier.com/locate/quaint http://dx.doi.org/10.1016/j.quaint.2014.09.064 1040-6182/© 2014 Elsevier Ltd and INQUA. All rights reserved. Quaternary International 352 (2014) 200e211
-
Upload
nevada-reno -
Category
Documents
-
view
0 -
download
0
Transcript of Living outside the box: An updated perspective on diet breadth and sexual division of labor in the...

lable at ScienceDirect
Quaternary International 352 (2014) 200e211
Contents lists avai
Quaternary International
journal homepage: www.elsevier .com/locate/quaint
Living outside the box: An updated perspective on diet breadth andsexual division of labor in the Prearchaic Great Basin
R.G. Elston a, *, D.W. Zeanah b, B.F. Codding c
a Department of Anthropology, University of Nevada, Reno, 1664 N. Virginia Street, Reno, NV 89557-0208, USAb Department of Anthropology, California State University Sacramento, 6000 J Street, Sacramento, CA 95819-6106, USAc Department of Anthropology, University of Utah, 270 S. 1400 E., Rm. 102, Salt Lake City, UT 84112, USA
a r t i c l e i n f o
Article history:Available online 7 November 2014
Keywords:Human behavioral ecologyDivision of laborPatch choiceMarginal value theoremPleistoceneeHolocene TransitionWestern stemmed point tradition
* Corresponding author.E-mail addresses: [email protected], robertels
[email protected] (D.W. Zeanah), brian.codding@anth
http://dx.doi.org/10.1016/j.quaint.2014.09.0641040-6182/© 2014 Elsevier Ltd and INQUA. All rights
a b s t r a c t
A tremendous amount has been learned about the Prearchaic (before 9000 BP) Great Basin since weadvocated a perspective of sexual division of labor based on Human Behavioral Ecology a decade ago.Many investigators have taken our advice and a few have challenged our assumptions and inferences.One of the most substantive critiques has been that we misunderstood the paleoenvironmental pa-rameters of ungulate populations during the PleistoceneeHolocene Transition (PHT). Simultaneously,behavioral ecologists have advanced our understanding of sexual division of labor among modern for-agers, but these studies appear to have gone unnoticed by Great Basin prehistorians. We review findingsof the last ten years and suggest that the key to understanding patterning in the PHT still relies onunderstanding (a) variability in men's and women's foraging goals, (b) the abundance and distribution oflarge prey, (c) how changing environmental parameters effect both the division of labor and the dis-tribution of resources, and (d) the relative influence of search and handling costs on residence time inPHT wetlands. We suggest that consideration of how paleoenvironmental variability structured sexualdivision of labor remains key to fully understanding Prearchaic lifeways in the Great Basin.
© 2014 Elsevier Ltd and INQUA. All rights reserved.
1. Introduction
Over ten years ago, Elston and Zeanah (2002) offered anexplanation for the seemingly contradictory combination of traitsobserved in Great Basin archaeological sites dating to the Pleisto-ceneeHolocene Transition (PHT): evidence for high mobility, lithictechnology adapted to hunting, and faunal assemblages suggestingbroad spectrum foraging. The paper compared applications ofvarious behavioral ecological models (Simms, 1987; Raven andElston, 1989; Zeanah et al., 1995; Pinson, 1999), using the simu-lated resource structure of Railroad Valley, Nevada (Zeanah et al.,1999) for the early and middle Holocene. Drawing from ethno-graphic studies of diet and patch choice, we proposed that theobserved patterns of site location, abundance and content resultedfrom the different foraging goals and strategies of men and womenin the environmental context of the PHT Great Basin. This Pre-archaic pattern differed fundamentally from subsequent Archaic(post 9000 BP) lifeways.
[email protected] (R.G. Elston),ro.utah.edu (B.F. Codding).
reserved.
Responses were mostly favorable, although there were some mis-interpretations of the piece. Herewe clarify themodel by updating theoriginal frameworkwithnew insights fromhumanbehavioral ecology(HBE) and a better understanding of the availability of large prey (ar-tiodactyls). Specifically,we highlight that (1) the key to understandingpatterning in thePHTstill hingesonunderstandingvariability inmen'sand women's foraging goals and the resulting division of labor; (2)while some suggest that large preywere too uncommon to have beencrucial resources for PrearchaicGreatBasin foragers, the concentrationof artiodactyls around wetlands actually made hunting large gamepredictable and reliable as longas foragerswere free tomove fromonewetland basin to another; (3) understanding both the division of laborand the patchiness of large prey provides a clear framework forframing PHT foraging, diminishing returns and how these effectedforager mobility and site formation patterns.
Before detailing our model of PHT (Prearchaic) lifeways, we firstreview insights from 2002 and compare how they stand up toadditional findings over the last 10 years.
1.1. The PHT record: key observations
Elston and Zeanah (2002) made a series of key empirical ob-servations, most of which have held up to subsequent

R.G. Elston et al. / Quaternary International 352 (2014) 200e211 201
investigations. However, recent work adds some important nu-ances. We highlight the original thinking in 2002, and update itwith new evidence.
1. Most Prearchaic assemblages occur in lowland locations near thewetlands that occupied numerous Great Basin valleys during thePHT (Fig. 1). Despite evidence for limited habitation levelselsewhere (Basgall, 2005; Middleton et al., 2014), this continuesto largely be true.
2. These assemblages typically lack diversity and evidence of pro-longed habitation, storage, or refuse accumulations (Beck andJones, 2009). While we agree that taphonomic factors oper-ating onmany PHT surface sites may have obscured evidence forresidential and storage features, we argue that this is due to theessentially ephemeral nature or absence of such features inshort-term camps used by highly mobile people (Binford, 1980).Substantial features for long-term habitation such as pithouses,prepared hearths, and large, deep storage pits would likely bepreserved to some degree even if subjected to bioturbation andinflation (upward movement of clasts). We observe that PHT-age deposits are often well enough protected in Great Basincaves and rockshelters which buffer taphonomic loss (Surovellet al., 2009) preserving organic artifacts and textiles, but evi-dence from these deposits, invariably thin and containing only
Fig. 1. Pluvial lakes in the Great Basin at their maximum late Pleistocene extent. Frames sproductivity of wetlands would have fluctuated in response to climatic variability, wetlandsthroughout the PHT.
simple hearths, does not support extended occupation (Kirneret al., 1997). While such sites often contain caches of tools, ortextiles that would have been useful to mobile groups cyclingbetween wetlands, caches of food are absent. Variability induration of occupations within particular wetlands is evident inthe use of local toolstones, the extent to which tools of non-localmaterials were curated and recycled, and possibly, the kind oftools manufactured (Duke and Young, 2007; Smith, 2007; Beckand Jones, 2009; Smith, 2011). This may relate to the abundanceof wetlands and distance from one wetland to another.
3. Further investigations have sharpened the distinctions betweenPHT and Archaic use of seeds. Most importantly, Prearchaic use ofground stone milling tools is episodic to altogether absent. Whilethere is strong evidence that PHT foragers used seeds at leastoccasionally, they appear to have usually done so without theaccouterments of milling technology that are “hallmarks”(Jennings,1957) of the later Archaic lifestyle (Basgall, 2008; Beckand Jones, 2009: 143; Goebel et al., 2011; Madsen, 2007; Rhodeet al., 2006; Rhode and Louderback, 2007; Schmitt et al., 2007;Yoder et al., 2010).
4. Tools suggestive of large game hunting and processing dominatePrearchaic chipped stone assemblages. This has largely remainedtrue even though evidence has mounted that stemmed pointswere designed for use in a variety of tasks (Beck and Jones,
how locations of Railroad Valley (a) and Old River Bed (b). Although the size of andpersisted and distance between lake basins would have remained relatively constant

R.G. Elston et al. / Quaternary International 352 (2014) 200e211202
2009). However recent use-wear analyses suggest that functionsincluded use as projectiles for hunting and butchering knives(Duke, 2011; Lafayette and Smith, 2012). Multi-functionalityshould have resulted in increased flexibility and toolstone con-servation, both beneficial for highly mobile adaptations.
5. Diverse toolstone sources in Prearchaic assemblages suggest sub-stantially larger lithic procurement zones for Prearchaic foragersthan typical of later Archaic foragers. Toolstone source profilescontinue to show that catchments exploited by PHT foragers,while likely not so large and probably much more variable thanenvisioned 10 years ago, were substantially larger than theirArchaic successors (Duke and Young, 2007; Smith, 2007, 2010,2011; Duke, 2011; Goebel et al., 2011; Smith and Kielhofer,2011; Jones et al., 2012). It seems most likely that toolstonewas acquired directly by residential mobility, and to a lesserdegree, by trade. Although we do not deny the possibility oflogistic procurement, we see little evidence for it in the formeither of upland base camps or in the use of upland toolstonequarries. Exploitation of such large territories suggests highermobility, lower human population densities, and lower levels ofcompetition for resources than prevailed in the Archaic.
6. Available faunal evidence indicates that Prearchaic foragers hunteda broad array of prey similar to that of later Archaic foragers.Recently analyzed faunal assemblages have continued todemonstrate use of the large artiodactyls present in the GreatBasin today, as well as a variety of smaller game (Hockett, 2007;Pinson, 2007; Goebel et al., 2011; Janetski et al., 2012). However,lack of evidence for bone boiling or marrow processing,commonly evident in Archaic archaeofaunal assemblages, sug-gests that Prearchaic foragers did not usually process fauna asintensively as Archaic foragers.
These six key points paint a concise picture of Prearchaic for-agers as targeting wetlands, utilizing diverse toolstone sources,having low population densities, exploiting large territories, doingmuch the same sorts of things site to site, not using substantialresidential structures, moving residential sites too frequently formidden to accumulate, lacking storage or extensive seed processingequipment, relying on flaked stone tools that whilemultifunctional,suggest a focus on large game hunting and processing, andexploiting diverse flora and fauna, including low-ranked resources.
And yet, the same empirical conundrum exists today as it did in2002: PHT foragers seemingly targeted large game while simulta-neously exploiting diverse, low-ranked resources, all the whileappearing to maintain high residential mobility. As in 2002, wesuggest the first key to understanding this problem is to under-stand the organization of men's and women's subsistence labor.
2. Sexual division of labor
In 2002, Elston and Zeanah benefited from a growing body ofHBE studies on the socioecological context of sexual division oflabor among modern foragers. We drew from the summaries ofHawkes (1990, 1996) and Bleige Bird (1999), and explicitly used thebehavior of contemporary Hadza foragers (O'Connell et al., 1988;Hawkes et al., 1991) e modern big game hunters par excellence e
as an analogy for how Great Basin foragers employed sexual divi-sion of labor to balance a “risk prone” hunting strategy with a “riskaverse” gathering strategy. In our view, the paleoenvironmentalstructure of the PHT did not impose the same spatial conflicts be-tweenmen's and women's foraging opportunities faced by foragerslater in the Archaic (post 9000 BP). Prearchaic men and womenfound their best foraging returns in and near wetland patchesproducing an archaeological record that appeared highly mobileand hunting oriented, yet broad spectrum. We specifically assumed
that Prearchaic sexual division of labor was organized in essentiallythe same manner as it was for ethnographic Great Basin foragers(Elston and Zeanah, 2002: 122e123, cf. Zeanah, 2004).
Since 2002, HBE research on sexual division of labor amonghunteregatherers has advanced significantly allowing a more so-phisticated understanding of its causes and variability. Data from asmall but diverse and extensively studied group of contemporaryforaging societies affirms that the critical differences betweenmen's and women's prey concerns both the variability and mean ofexpected return. Women tend to select prey that can be reliablycaptured and used to provision children. In contrast, men generallytarget high yield resources that are amenable for distributionamong larger social groups, but vulnerable to risks of failed pursuitsand encounters. Debates in HBE over the last several decades haveoften reduced the issue to hunting for food vs. hunting for prestige(Marlowe, 2007; Gurven and Hill, 2009). Are men costly signalingby funding a “public good” with resources risky to acquire? Cansuch “risky pursuits” ever offer a reliable means of provisioningoffspring?
In a recent paper, Codding et al. (2011; Codding, 2012) point outthat hunting for prestige vs. food is a false dichotomy. Both mayoccur, but emphasis on one or the other will vary, even on a day-to-day basis within the same society, depending on the structure ofthe resource base. Specifically, Codding and colleagues demon-strate that the reliability of high-energy resource (e.g., large game)acquisition, determines the extent of conflicts between the goals ofmaximizing energy and minimizing risk, which in turn predicts thedegree to which men's and women's foraging efforts diverge orconverge.
Men's and women's foraging goals differ: men most frequentlymaximize energy without much attention to risk, and womenattempt to minimize risk over any preference for energy. Whenhigh-energy resources can be acquired with low risk of failure,men's and women's foraging strategies tend to converge on similarresources. Convergent foraging strategies are of two kindsdepending on the amount of alloparental support available. Wherelarger groups provide more alloparents, women and men bothtarget high-energy resources (e.g., Agta; Goodman et al., 1985).When groups are small and there are fewer alternative caregivers,men's direct and reliable acquisition of large game is made possibleby women's logistical support d not in the Binfordian sense(Binford, 1980), but in terms of supporting residential moves (e.g.,Ache: Hawkes et al., 1982, 1987) or in processing (e.g., Inuit:Jarvenpa and Brumbach, 2006).
When large, high-energy resources decline in abundance andsuccess becomes uncertain, foragers trade off minimizing risk andmaximizing energy in compensation, and foraging strategiesdiverge by gender. Men are more likely to target high-risk, high-energy resources that can infrequently provision all within thegroup, while women focus on low-energy resources to reliablyprovision offspring and consequently provide the bulk of the re-sources consumed by the foraging group (e.g., Martu, Meriamduring turtle hunting season: Codding et al., 2011; Hadza:Marlowe,2010; Ju/ʼhoansi during dry season: Lee, 1979; and virtually allethnographic Great Basin foragers: Steward, 1938).
The main insight is that whether a group's division of labor ismore likely to converge or diverge depends on the reliability ofhigh-energy resource acquisition. In the Great Basin, that likelydependedmost on the reliability of hunting large prey (Fig. 2). If ourassessment of the resource structure of the PHT Great Basin (dis-cussed below) is correct, we believe that hunting success for higherranked prey, especially ungulates, was actually quite high becausethey were reliably intercepted in and near the wetlands thatoccupied most Great Basin valleys, and there was relatively littlecompetition for these resources because of the low human

Fig. 2. Bivariate plot examining variability in the division of labor as a function ofresource acquisition risk and harvest size proposed for PHT Great Basin foragers (note,though represented as discrete, these should vary continuously; adapted fromCodding, 2012). When available resources provide high-energy returns with low risk(e.g., waterfowl or artiodactyls near the marsh), men's and women's prey sets shouldconverge; when high energy resources are associated with greater acquisition risk, thetargets of men's and women's foraging should diverge with women focusing on lower-energy reliable resources (e.g., eggs, cattail pollen, muskrats, tubers and seeds) whilemen continue to focus on high-energy packages but at higher risk of failure.
R.G. Elston et al. / Quaternary International 352 (2014) 200e211 203
population density. Consequently, sexual division of labor amongPrearchaic hunteregatherers was more convergent than it was forlater Archaic foragers. Ted Goebel et al. (2011:498) were not far offthe mark when they commented that the mortuary associations ofthe Buhl burial suggested a lower degree of sexual division of laborthan we inferred in 2002. Admittedly this is a single example, butprovocative nonetheless.
Given the broad array of small to large sized game representedin PHT faunal assemblages, we can accommodate either the pos-sibility that Prearchaic women regularly contributed to the diet bypursuing a similar array of prey as men (like Agta women;Goodman et al., 1985), or Prearchaic men contributed the bulk ofthe diet through hunting while women either focused on pro-cessing (like many Arctic groups, e.g., Jarvenpa and Brumbach,2006) or played a more logistical role in facilitating hunting (likeAche, e.g., Hawkes et al., 1982). However, we suspect that allopar-enting opportunities were limited in the small groups that seemtypical of the PHT Great Basin and, much like contemporary Ache,Prearchaic women supported the high residential mobility neces-sary to maintain high encounter rates with large game. The Achecase provides a particularly instructive analogy here: while Achewomen do often forage for lower-ranked, reliable resources, theyregularly bypass these opportunities in order to facilitate men'shunting of larger, reliably encountered, and widely shared re-sources (Hawkes et al., 1982). This is precisely the circumstance wepose for the Prearchaic, accounting for the signatures of highmobility, hunting oriented technology, and broad diet. Whether ornot this is true, however, hinges on understanding the abundanceof large prey during the PHT e something that has been the topic ofrecent debate.
2.1. Abundance of large game
Broughton et al. (2008), Hockett (2007), and Pinson (2007) havecritiqued the inference that large game (Artiodactyls) played animportant role in Prearchaic subsistence strategies. All three point
to the low proportion of large ungulates relative to smaller game atPHT sites with preserved faunal assemblages. Leaving aside thequestion of howmany rabbits, squirrels, or grasshoppers it takes toequal the caloric return of one deer, we point out that artiodactylremains are frequently more abundant in PHT components (both inMNI and NISP) than in later Archaic deposits (cf. Pinson, 2007:Table 10.4; Hockett, 2007: Table 11.2). Relative to the entire faunalassemblages frommulticomponent sites with PHT deposits, it is theabundance of small game, not the rarity of large, that accounts forthe relatively low ratios of ungulates in Prearchaic components (aswould be expected with a converging-diverging division of labor,see below). Further, we argue that these relatively low artiodactylindices could be due to field processing thresholds, or samplingbiases reflecting profoundly different PHT and Holocene site for-mation processes. Until these factors are understood and controlledfor both small and large fauna, simple appeals to artiodactyl indicesalone are inconclusive, and we argue that the only conclusiveinference that can be made is that both large and small prey wereimportant prey items of PHT foragers, precisely the phenomenonthat calls for explanation.
More profoundly, Broughton et al. (2008) have challenged ourinterpretation of paleoclimatic evidence that cool, moist climaticconditions of the PHT were capable of supporting sufficient car-rying capacities for large ungulates near wetland patches to makehunting them profitable. Based on climatic reconstructions theyargue that artiodactyl densities were too low during the PHT tohave served as reliable resources, because orbital forcing resultedin greater seasonality during the PHT. Longer, hotter summersand more severe winters limited artiodactyl carrying capacities inthe Great Basin until seasonality ameliorated in the late Holocene.We share Don Grayson's (2011) skepticism that paleoenvir-onmental evidence supports PHT seasonality of the scale thatBroughton and colleagues envision. However, we point out that ifthese reconstructions are accurate, artiodactyls should have,nonetheless, been even more concentrated around marshes andlakes.
Present use of wetlands and riparian zones by large mammals iswell documented. Elk and mule deer use palustrine habitats forshelter and foraging (Lohman, 2004). Several studies describe deerand elk use of riparian habitat (Gammonley, 2004) for water, food,and thermal cover for resting and birthing especially in summerwhen uplands dry out (Leckenby et al., 1982; Marcum and Scott,1985; Raedeke et al., 1988; Walmo, 1981; Wittmer and Calesta,1983). Riparian zones are preferred corridors for deer movement,especially between uplands and valley floors. The marshes and ri-parian zones of the Malheur Lake/Blitzen River/Silvies River systemare home to a large, permanently resident mule deer population,where females oftenwade out to Malheur Lake islands to give birth(Fig. 3).
Anecdotal evidence of use of wetlands from wildlife refugemanagers at Stillwater, Ruby Lake, and Malheur indicate that an-telope and bighorn sheep also make use of wetlands. Antelopewater and forage along the margins of the Malheur and Stillwatermarshes in places where sight lines are undiminished by trees orhigh brush; around A.D.1300, antelopewere apparently driven intothemarsh and processed at the Sheepy East 1 site in Lower KlamathLake (McGuire, 1985). Bighorn sheep come down to wetlandswhere there is nearby escape terrain (e.g., to Stillwater Marsh fromthe Stillwater Mountains via Stillwater point and from the RubyMountains down to Ruby Lake).
While Broughton et al. (2008) argue that greater seasonalitywith harsher winters and hotter summers would have degradedcritical artiodactyl habitat, forage, and water sources, this shouldhave made large game more dependent on wetland habitats, verylikely concentrating animals in and adjacent to the marshes and

Fig. 3. Mule deer in Malheur Lake, c. B. Wheeler, USFWS Volunteer. Elk and mule deeruse palustrine habitats for shelter and foraging and riparian habitat for water, food,thermal cover for resting and birthing especially in summer when uplands dry out.These would have been key locations to intercept large game reliably, even if theiroverall abundance in the environment was relatively low.
R.G. Elston et al. / Quaternary International 352 (2014) 200e211204
riparian zones feeding them, particularly as climate grew dryer andmarshes diminished.
In 2002, Elston and Zeanah were unaware of the extent ofartiodactyl use of wetlands and proposed that large animals wouldhave been hunted from lowland bases in low to mid-elevationbrushy steppe. In many of the narrower Great Basin Valleys, thiswould have entailed foraging trips of less than 10 km, and notrequired (contra Madsen, 2007) extensive logistical trips outsidethe valley. If however, we are correct to assume the presence ofartiodactyls in and immediately adjacent to wetlands, then Pre-archaic hunters could have procured them without incurring ashigh search and transport costs as those paid by Archaic hunters.Human foragers knew where and when animals would most likelybe located, whether they were resident in awetland, moving in andout of it, or visiting its margins. All of these situations could betaken advantage of through encounter or intercept huntingstrategies.
Indeed, if seasonal artiodactyl migration routes circulatedthrough PHT wetlands, Prearchaic hunters could have practicedintercept-hunting strategies to a much higher degree than theirArchaic counterparts. Compared to Middle/Late Holocene Archaicforagers, PHT Prearchaic foragers do not appear to have processedartiodactyl remains as intensivelydthere is no evidence for boneboiling or marrow processing as seen during the Archaic. All thissuggests that success rates with artiodactyls were higher in thePHT, allowing foragers to avoid excessive bone transport to, orprocessing at, the central place.
We find support for the importance of intercept hunting oflarge game in the distribution of sites in Railroad Valley Nevada(Fig. 4). The figure illustrates the distribution of PHT sites andisolates in the region relative to a paleoenvironmental modelshowing the maximum extent of Pleistocene wetlands and sur-rounding landforms. The Paleoenvironmental model was con-structed by combining two independent analyses of the originalRailroad Valley Model (Zeanah et al., 1999). We estimated theminimum and maximum extent of Pleistocene wetlands frommodern soil map units that we reasoned were likely to haveformed at the bottom and margins of lakes (although they includesome eolian soils that clearly postdate Pleistocene wetlands). Thezone of minimum extent is the modern abiotic playa and adjacentsaline soils that support very alkaline vegetation communities. Thehigh salinity of these soils likely results from the desiccation ofshallow lakes and marshes. The riparian soils are simply modern
riparian soil map units. The zone of maximum extent is based onsoil map units currently classified as irregularly inundated, underthe assumption that these were most likely flooded during Pleis-tocene highstands. This overlies a layer of geomorphic landformsdelineated by analysis of air photo stereo pairs. Those landformsrepresenting relict Pleistocene shorelines and lagoons, and fluvialgravel bars are highlighted so they show through the wetlandlayer. The close correspondence of the pluvial/fluvial landformswith the wetland zone derived from soil map units reassures usthat we have derived a reasonable approximation of the extent ofPleistocene wetlands. Young fans are alluvial surfaces that areprobably too young to bear in situ PHT sites and isolates. Old fanand upland landforms are old enough to contain PHT culturalmaterials.
To determine if there are significant differences in PHT versusArchaic site clustering along these different landforms, we ran aseries of point pattern analyses using Ripley's K. Ripley's K calcu-lates the distances between neighboring points at specified spatialextents and determines if the average distance between neighborsin a sample differs significantly from a theoretical randomly ho-mogenous distribution. Here, K[t/o] reports the ratio of the theo-retical or random K value over the observed K value. Potentialvalues range from 0 to 2 with values close to zero suggest signifi-cant clustering, values close to 1 suggest a random distribution andvalues close to 2 suggest significant dispersion. These values reportclustering at a 1-km radii within the valley, asking whether or notsites are more or less likely to have a local neighbor than would beexpected by random chance. Significant departures of an observedvalue from a theoretical value are based on a Monte Carlo simula-tion with 10,000 iterations. Multi-scalar Ripley's K increases theneighborhood size across multiple extents to a maximum radius.
An analysis of local spatial clustering reveals that PHT sites aresignificantly clustered (K[t/o] ¼ 0.05, p < 0.0001) while MiddleHolocene sites exhibit onlymarginally significant local clustering (K[t/o] ¼ 0.33, p ¼ 0.0678). This suggests that neighboring PHT sitesare closer to one another than would be expected by randomchance. Extending this to a multi-scale cluster analysis calculatingRipley's K at increasing distances across the valley shows that PHTsites are significantly clustered throughout almost the entire extentof the valley (Fig. 5; Pelissier and Goreaud, 2010; R, 2014). Incontrast, Middle Holocene sites only reveal significant clustering atscales up to half of the valley, showing essentially a random dis-tribution across half of the extent.
These findings imply that PHT sites were more narrowly focusedon particular habitats within the valley. The strongest clustering ofPHT sites and isolates is along ancient fluvial gravel bars of thecreek flowing into the basin from the north (Zancanella, 1984). Aweaker, but suggestive clustering of sites is apparent near thedrainage flowing from the south. Such locations likely would havebeen suitable for intercept hunting of large game using the riparianzones as migration corridors from surrounding uplands. The largestknown PHT site in the region (Elston et al., 1979), an extensive lithicscatter, is highlighted as a likely location of a central place basecamp by encircling it within a 10 km foraging catchment. Situatedon a relict pluvial gravel spit extending far out into the wetland tointercept the stream feeding the basin, the site provided anelevated dry place ideally situated to access the stream and thesurrounding wetlands and lake to the south. Moreover, it is alsoconveniently located for hunters intercepting game along the ri-parian corridor to the north. Similar PHT site concentrations alongriparian corridors have been observed elsewhere in the Great Basin(Mullins et al., 2013; Duke and King, 2014; Madsen et al., 2015). Wesuggest that this is the sort of site locational strategy that wouldhave occurred under conditions of converging and diverging divi-sion of labor.

Fig. 4. Map of Railroad Valley Archaeological Sites and Paleo-habitat reconstructions during the Prearchaic. Prearchaic sites are significantly more clustered along wetland/riparianhabitats than succeeding Early Archaic sites.
R.G. Elston et al. / Quaternary International 352 (2014) 200e211 205
The crucial point is that the patchy concentration of artiodactylsaround marshes and lakes made their acquisition there reliable;their overall abundance is less important, especially given that withlow human population densities and small group size, PHT foragerswould have been free to travel to new wetland patches as localhunting returns diminished. The large animals in and adjacent towetlands would have constituted low hanging fruit for Prearchaicforagers. However, their local populations should have been rela-tively easily depressed by hunting, particularly given the resourcestructure envisioned by Broughton and colleagues. This would havehad some interesting implications for the patch preferences ofPrearchaic foragers.
2.2. Diminishing returns, marginal value and mobility
As illustrated by the Marginal Value Theorem (MVT) the dura-tion a forager should stay within a patch can be modeled by themarginal value of a diminishing returns curve (Charnov, 1974;Charnov and Orians, 1973; Charnov et al., 1976). The shape andheight of the curve depends on the quantity of resources beingexploited and the rate at which foragers can depress foragingreturns within a patch. The length of time foragers stay within apatch before departing for another depends on the time it takes totravel to an adjacent patch (Fig. 6). The classic formulation of theMVT did not distinguish search and handling time, but Bettinger
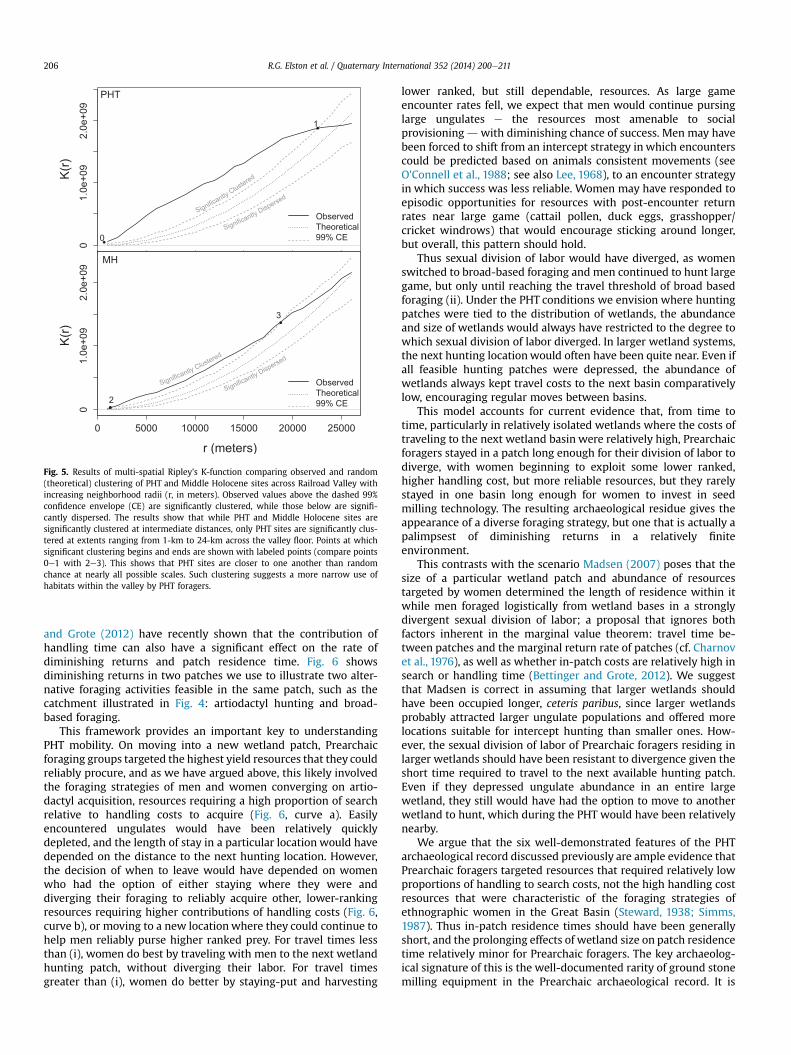
Fig. 5. Results of multi-spatial Ripley's K-function comparing observed and random(theoretical) clustering of PHT and Middle Holocene sites across Railroad Valley withincreasing neighborhood radii (r, in meters). Observed values above the dashed 99%confidence envelope (CE) are significantly clustered, while those below are signifi-cantly dispersed. The results show that while PHT and Middle Holocene sites aresignificantly clustered at intermediate distances, only PHT sites are significantly clus-tered at extents ranging from 1-km to 24-km across the valley floor. Points at whichsignificant clustering begins and ends are shown with labeled points (compare points0e1 with 2e3). This shows that PHT sites are closer to one another than randomchance at nearly all possible scales. Such clustering suggests a more narrow use ofhabitats within the valley by PHT foragers.
R.G. Elston et al. / Quaternary International 352 (2014) 200e211206
and Grote (2012) have recently shown that the contribution ofhandling time can also have a significant effect on the rate ofdiminishing returns and patch residence time. Fig. 6 showsdiminishing returns in two patches we use to illustrate two alter-native foraging activities feasible in the same patch, such as thecatchment illustrated in Fig. 4: artiodactyl hunting and broad-based foraging.
This framework provides an important key to understandingPHT mobility. On moving into a new wetland patch, Prearchaicforaging groups targeted the highest yield resources that they couldreliably procure, and as we have argued above, this likely involvedthe foraging strategies of men and women converging on artio-dactyl acquisition, resources requiring a high proportion of searchrelative to handling costs to acquire (Fig. 6, curve a). Easilyencountered ungulates would have been relatively quicklydepleted, and the length of stay in a particular location would havedepended on the distance to the next hunting location. However,the decision of when to leave would have depended on womenwho had the option of either staying where they were anddiverging their foraging to reliably acquire other, lower-rankingresources requiring higher contributions of handling costs (Fig. 6,curve b), or moving to a new locationwhere they could continue tohelp men reliably purse higher ranked prey. For travel times lessthan (i), women do best by traveling with men to the next wetlandhunting patch, without diverging their labor. For travel timesgreater than (i), women do better by staying-put and harvesting
lower ranked, but still dependable, resources. As large gameencounter rates fell, we expect that men would continue pursinglarge ungulates e the resources most amenable to socialprovisioningdwith diminishing chance of success. Men may havebeen forced to shift from an intercept strategy in which encounterscould be predicted based on animals consistent movements (seeO'Connell et al., 1988; see also Lee, 1968), to an encounter strategyin which success was less reliable. Women may have responded toepisodic opportunities for resources with post-encounter returnrates near large game (cattail pollen, duck eggs, grasshopper/cricket windrows) that would encourage sticking around longer,but overall, this pattern should hold.
Thus sexual division of labor would have diverged, as womenswitched to broad-based foraging and men continued to hunt largegame, but only until reaching the travel threshold of broad basedforaging (ii). Under the PHT conditions we envision where huntingpatches were tied to the distribution of wetlands, the abundanceand size of wetlands would always have restricted to the degree towhich sexual division of labor diverged. In larger wetland systems,the next hunting locationwould often have been quite near. Even ifall feasible hunting patches were depressed, the abundance ofwetlands always kept travel costs to the next basin comparativelylow, encouraging regular moves between basins.
This model accounts for current evidence that, from time totime, particularly in relatively isolated wetlands where the costs oftraveling to the next wetland basin were relatively high, Prearchaicforagers stayed in a patch long enough for their division of labor todiverge, with women beginning to exploit some lower ranked,higher handling cost, but more reliable resources, but they rarelystayed in one basin long enough for women to invest in seedmilling technology. The resulting archaeological residue gives theappearance of a diverse foraging strategy, but one that is actually apalimpsest of diminishing returns in a relatively finiteenvironment.
This contrasts with the scenario Madsen (2007) poses that thesize of a particular wetland patch and abundance of resourcestargeted by women determined the length of residence within itwhile men foraged logistically from wetland bases in a stronglydivergent sexual division of labor; a proposal that ignores bothfactors inherent in the marginal value theorem: travel time be-tween patches and the marginal return rate of patches (cf. Charnovet al., 1976), as well as whether in-patch costs are relatively high insearch or handling time (Bettinger and Grote, 2012). We suggestthat Madsen is correct in assuming that larger wetlands shouldhave been occupied longer, ceteris paribus, since larger wetlandsprobably attracted larger ungulate populations and offered morelocations suitable for intercept hunting than smaller ones. How-ever, the sexual division of labor of Prearchaic foragers residing inlarger wetlands should have been resistant to divergence given theshort time required to travel to the next available hunting patch.Even if they depressed ungulate abundance in an entire largewetland, they still would have had the option to move to anotherwetland to hunt, which during the PHT would have been relativelynearby.
We argue that the six well-demonstrated features of the PHTarchaeological record discussed previously are ample evidence thatPrearchaic foragers targeted resources that required relatively lowproportions of handling to search costs, not the high handling costresources that were characteristic of the foraging strategies ofethnographic women in the Great Basin (Steward, 1938; Simms,1987). Thus in-patch residence times should have been generallyshort, and the prolonging effects of wetland size on patch residencetime relatively minor for Prearchaic foragers. The key archaeolog-ical signature of this is the well-documented rarity of ground stonemilling equipment in the Prearchaic archaeological record. It is
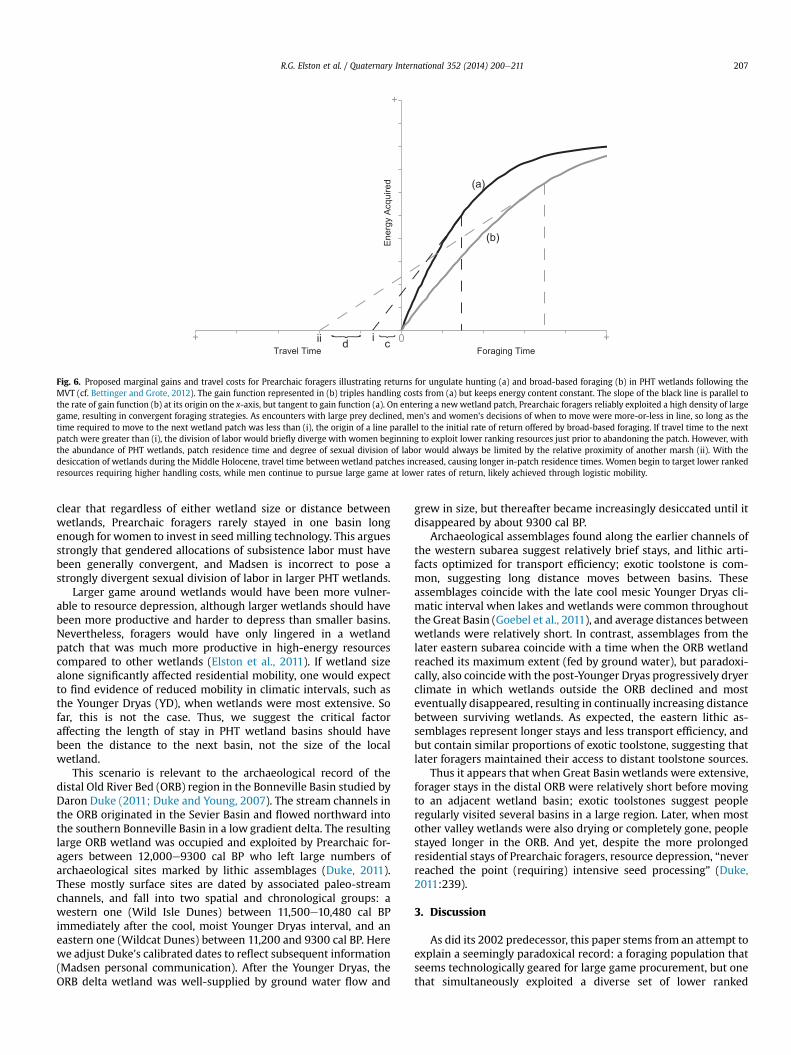
Fig. 6. Proposed marginal gains and travel costs for Prearchaic foragers illustrating returns for ungulate hunting (a) and broad-based foraging (b) in PHT wetlands following theMVT (cf. Bettinger and Grote, 2012). The gain function represented in (b) triples handling costs from (a) but keeps energy content constant. The slope of the black line is parallel tothe rate of gain function (b) at its origin on the x-axis, but tangent to gain function (a). On entering a newwetland patch, Prearchaic foragers reliably exploited a high density of largegame, resulting in convergent foraging strategies. As encounters with large prey declined, men's and women's decisions of when to move were more-or-less in line, so long as thetime required to move to the next wetland patch was less than (i), the origin of a line parallel to the initial rate of return offered by broad-based foraging. If travel time to the nextpatch were greater than (i), the division of labor would briefly diverge with women beginning to exploit lower ranking resources just prior to abandoning the patch. However, withthe abundance of PHT wetlands, patch residence time and degree of sexual division of labor would always be limited by the relative proximity of another marsh (ii). With thedesiccation of wetlands during the Middle Holocene, travel time between wetland patches increased, causing longer in-patch residence times. Women begin to target lower rankedresources requiring higher handling costs, while men continue to pursue large game at lower rates of return, likely achieved through logistic mobility.
R.G. Elston et al. / Quaternary International 352 (2014) 200e211 207
clear that regardless of either wetland size or distance betweenwetlands, Prearchaic foragers rarely stayed in one basin longenough for women to invest in seed milling technology. This arguesstrongly that gendered allocations of subsistence labor must havebeen generally convergent, and Madsen is incorrect to pose astrongly divergent sexual division of labor in larger PHT wetlands.
Larger game around wetlands would have been more vulner-able to resource depression, although larger wetlands should havebeen more productive and harder to depress than smaller basins.Nevertheless, foragers would have only lingered in a wetlandpatch that was much more productive in high-energy resourcescompared to other wetlands (Elston et al., 2011). If wetland sizealone significantly affected residential mobility, one would expectto find evidence of reduced mobility in climatic intervals, such asthe Younger Dryas (YD), when wetlands were most extensive. Sofar, this is not the case. Thus, we suggest the critical factoraffecting the length of stay in PHT wetland basins should havebeen the distance to the next basin, not the size of the localwetland.
This scenario is relevant to the archaeological record of thedistal Old River Bed (ORB) region in the Bonneville Basin studied byDaron Duke (2011; Duke and Young, 2007). The stream channels inthe ORB originated in the Sevier Basin and flowed northward intothe southern Bonneville Basin in a low gradient delta. The resultinglarge ORB wetland was occupied and exploited by Prearchaic for-agers between 12,000e9300 cal BP who left large numbers ofarchaeological sites marked by lithic assemblages (Duke, 2011).These mostly surface sites are dated by associated paleo-streamchannels, and fall into two spatial and chronological groups: awestern one (Wild Isle Dunes) between 11,500e10,480 cal BPimmediately after the cool, moist Younger Dryas interval, and aneastern one (Wildcat Dunes) between 11,200 and 9300 cal BP. Herewe adjust Duke's calibrated dates to reflect subsequent information(Madsen personal communication). After the Younger Dryas, theORB delta wetland was well-supplied by ground water flow and
grew in size, but thereafter became increasingly desiccated until itdisappeared by about 9300 cal BP.
Archaeological assemblages found along the earlier channels ofthe western subarea suggest relatively brief stays, and lithic arti-facts optimized for transport efficiency; exotic toolstone is com-mon, suggesting long distance moves between basins. Theseassemblages coincide with the late cool mesic Younger Dryas cli-matic interval when lakes and wetlands were common throughoutthe Great Basin (Goebel et al., 2011), and average distances betweenwetlands were relatively short. In contrast, assemblages from thelater eastern subarea coincide with a time when the ORB wetlandreached its maximum extent (fed by ground water), but paradoxi-cally, also coincidewith the post-Younger Dryas progressively dryerclimate in which wetlands outside the ORB declined and mosteventually disappeared, resulting in continually increasing distancebetween surviving wetlands. As expected, the eastern lithic as-semblages represent longer stays and less transport efficiency, andbut contain similar proportions of exotic toolstone, suggesting thatlater foragers maintained their access to distant toolstone sources.
Thus it appears that when Great Basin wetlands were extensive,forager stays in the distal ORB were relatively short before movingto an adjacent wetland basin; exotic toolstones suggest peopleregularly visited several basins in a large region. Later, when mostother valley wetlands were also drying or completely gone, peoplestayed longer in the ORB. And yet, despite the more prolongedresidential stays of Prearchaic foragers, resource depression, “neverreached the point (requiring) intensive seed processing” (Duke,2011:239).
3. Discussion
As did its 2002 predecessor, this paper stems from an attempt toexplain a seemingly paradoxical record: a foraging population thatseems technologically geared for large game procurement, but onethat simultaneously exploited a diverse set of lower ranked
R.G. Elston et al. / Quaternary International 352 (2014) 200e211208
resources while appearing to have employed extremely high resi-dential mobility encompassing expansive lithic conveyance zones.Those who argue that large game played only a minor role in Pre-archaic subsistence-settlement strategies address the exploitationof such large territories unconvincingly if at all. What accounts forthe scale of catchments evident in lithic toolstone profiles if nothigh mobility and hunting? If Prearchaic foragers were focused onan array of wetland resources comparable to that exploited byArchaic and ethnographic foragers, there should have been littleneed to leave one wetland for another because wetlands wouldhave been resistant to human-induced resource depression(Zeanah, 2004). If resources became scarce within the dailyforaging range, Prearchaic foragers should simply have residentiallymoved to the next patch in the same wetland as Bob Kelly (1995)modeled for Archaic foragers. Seasonal and inter-annual fluctua-tions in wetland productivity should have been countered in thesame manner as Archaic foragers: settling within the most pro-ductive wetlands, milling and storing seeds to deal with risk, andacquiring toolstone mostly by logistical procurement. If all thiswere true, Prearchaic and Archaic sites and settlement patternswould be essentially identical e but they are not. Toolstoneacquisition embedded in a residentially mobile pattern geared tolarge game acquisition continues to be the only viable explanationposed for the Prearchaic pattern that is supported by ethnograph-ically documented causes of hunteregatherer mobility (cf. Binford,2001; Kelly, 2013).
We argue that understanding the Prearchaic pattern requires adetailed grasp of the sexual division of labor and how it interacteddynamically with variability in the abundance of high ranking re-sources (mainly artiodactyls) and their patchy distribution in thePHT Great Basin. We suggest that Prearchaic foragers targetedwetlands in discrete basins where large game were likely to beabundant and reliably encountered. When encounter rates withlarge game declined, women shifted focus to smaller more reliableresources, but only until a point at which they were better offtraveling with men to the nearest adjacent wetland in order toagain target higher-ranking artiodactyls.
Of course, this systemwas dependent on the lakes and wetlandsin Great Basin valleys fostered by the relatively coolmoist climate ofthe PHT. These formed homogeneous patches containing essen-tially inexhaustible plants, birds, and small animals, along withlarge game, which we arguewould have been tethered towetlands.It is likely that the productivity of wetlands, particularly as largeungulate patches, would have been vulnerable to climate change.Recent dates from Paisley Caves (Jenkins et al., 2012), extend the
8000 10000
-42
-40
-38
-36
-34
Early Holocene
Known Duration of We
6,600
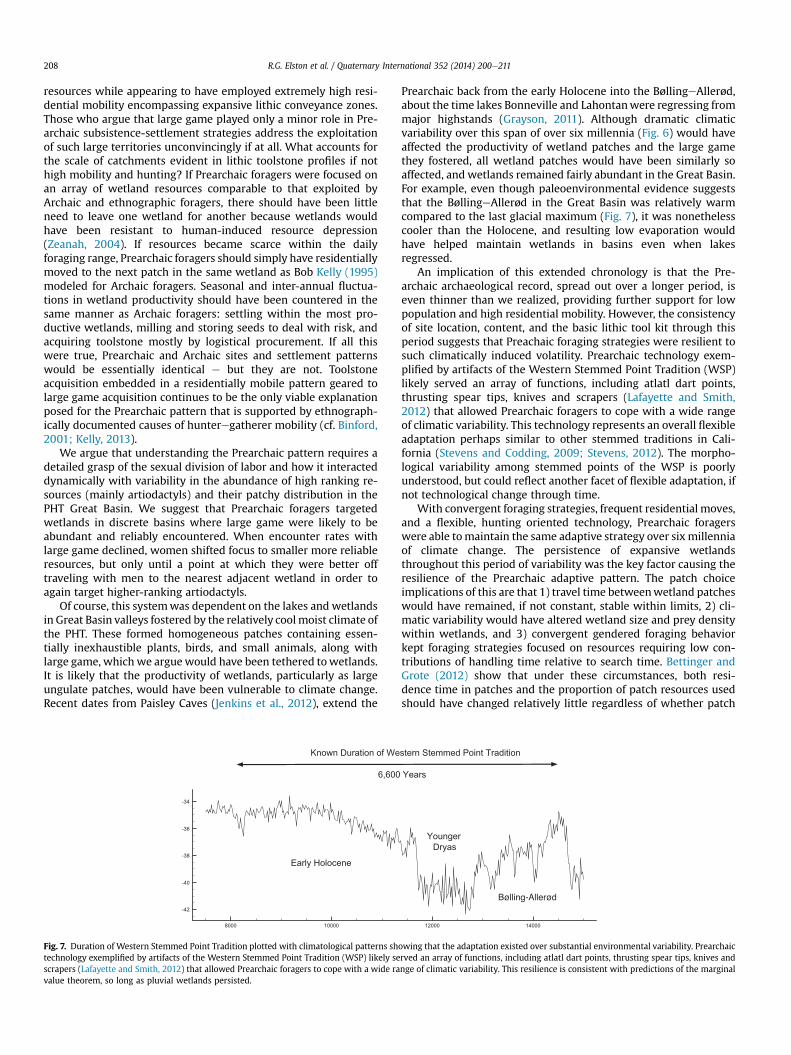
Fig. 7. Duration of Western Stemmed Point Tradition plotted with climatological patterns shtechnology exemplified by artifacts of the Western Stemmed Point Tradition (WSP) likely sescrapers (Lafayette and Smith, 2012) that allowed Prearchaic foragers to cope with a wide ravalue theorem, so long as pluvial wetlands persisted.
Prearchaic back from the early Holocene into the BøllingeAllerød,about the time lakes Bonneville and Lahontanwere regressing frommajor highstands (Grayson, 2011). Although dramatic climaticvariability over this span of over six millennia (Fig. 6) would haveaffected the productivity of wetland patches and the large gamethey fostered, all wetland patches would have been similarly soaffected, and wetlands remained fairly abundant in the Great Basin.For example, even though paleoenvironmental evidence suggeststhat the BøllingeAllerød in the Great Basin was relatively warmcompared to the last glacial maximum (Fig. 7), it was nonethelesscooler than the Holocene, and resulting low evaporation wouldhave helped maintain wetlands in basins even when lakesregressed.
An implication of this extended chronology is that the Pre-archaic archaeological record, spread out over a longer period, iseven thinner than we realized, providing further support for lowpopulation and high residential mobility. However, the consistencyof site location, content, and the basic lithic tool kit through thisperiod suggests that Preachaic foraging strategies were resilient tosuch climatically induced volatility. Prearchaic technology exem-plified by artifacts of the Western Stemmed Point Tradition (WSP)likely served an array of functions, including atlatl dart points,thrusting spear tips, knives and scrapers (Lafayette and Smith,2012) that allowed Prearchaic foragers to cope with a wide rangeof climatic variability. This technology represents an overall flexibleadaptation perhaps similar to other stemmed traditions in Cali-fornia (Stevens and Codding, 2009; Stevens, 2012). The morpho-logical variability among stemmed points of the WSP is poorlyunderstood, but could reflect another facet of flexible adaptation, ifnot technological change through time.
With convergent foraging strategies, frequent residential moves,and a flexible, hunting oriented technology, Prearchaic foragerswere able tomaintain the same adaptive strategy over six millenniaof climate change. The persistence of expansive wetlandsthroughout this period of variability was the key factor causing theresilience of the Prearchaic adaptive pattern. The patch choiceimplications of this are that 1) travel time betweenwetland patcheswould have remained, if not constant, stable within limits, 2) cli-matic variability would have altered wetland size and prey densitywithin wetlands, and 3) convergent gendered foraging behaviorkept foraging strategies focused on resources requiring low con-tributions of handling time relative to search time. Bettinger andGrote (2012) show that under these circumstances, both resi-dence time in patches and the proportion of patch resources usedshould have changed relatively little regardless of whether patch
12000 14000
Younger Dryas
stern Stemmed Point Tradition
Years
owing that the adaptation existed over substantial environmental variability. Prearchaicrved an array of functions, including atlatl dart points, thrusting spear tips, knives andnge of climatic variability. This resilience is consistent with predictions of the marginal

Table 1Aspects of Prearchaic Adaptation.
Temporal Scale Six thousand years, Late Pleistocene to Early HoloceneClimate Cooler than Holocene and relatively moist but variable
at many temporal scalesResource Patches Homogeneous (wetlands), abundantKey Resources Reliable large game, inexhaustible plants and small gameMaximum Range Very largeLithic Technology Hunting-oriented but flexible, easily repurposedPopulation Low, regulated by long birth spacing and
climate-induced infant mortalityResidential Mobility High, short stays, frequent movesGroup Size Small, homogenous, connected by movement through
rangeDivision of Labor Mostly convergent but flexible
R.G. Elston et al. / Quaternary International 352 (2014) 200e211 209
productivity was low or high. Thus, patch choice predicts thatPrearchaic foraging strategies should have been resilient to thepaleoclimatic variability of the PHT irrespective of how that vari-ability affected wetland productivity.
The success of the Prearchaic lifewaywas also predicated on lowhuman population levels and lack of competition for the high-ranking resources in wetland sweet spots. Lack of competition forresources made convergent division of labor and high residentialmobility productive tactics. Low population levels during the PHTwere probably maintained by extreme climatic variability thatcontributed to infant and child mortality operating as a damper onpopulation in a stable limit cycle (Belovsky, 1988; Winterhalderet al., 1988; Elston et al., 2011).
It appears that the long-term success of the Prearchaic lay in thehomogeneity of wetland patches and connectedness of the groupsexploiting them. Everyone did the same kinds of things in the samekinds of places and apparently, groups were not isolated enough todevelop behaviors that caused significant variation in technology.Complex systems with these features can be resilient and highlyresistant to change until some critical threshold (“tipping point”) isreached and the whole system shifts to another mode (Schefferet al., 2012) or fails altogether. Signs that a system is stressedmay appear before the critical threshold in the form of short-termstochastic changes (“flickering,” Scheffer et al., 2012) to or towardthe new mode. Elston et al. (2011) proposed that something likethis could account for the episodic appearance of ground stonetools and pottery in late Pleistocene northern China, and we sug-gest that this could account as well for the infrequent and short-term appearance of ground stone in the Prearchaic Great Basin.
Duke's (2011) analysis of the latest ORB lithic assemblagesdemonstrates a technological response to environmental change,but not one that transformed the entire system. Adjustments inlithic procurement, production and tool maintenance were madepost-YD, but the basic tool kit and its functions remained the sameuntil the entire system failed. Duke was able to observe this in theORB because dating paleochannels in the ORB delta allowed earlyand later surface sites to be sorted out. Although wemight expect asimilar technological response in warmer, dryer pre-YD climateintervals, nothing like it has so far been observed. It is unclearwhether the systemic changes Duke recognizes in the latest ORBlithic assemblages constitute a technological flicker toward a newmode, or a more gradual transition typical of heterogeneous sys-tems with low levels of connectivity (Scheffer et al., 2012).
It was not until wetlands began to dry out in the latter part ofthe Early Holocene that the Prearchaic adaptive strategy reached atipping point. As wetlands disappeared, travel time between thosethat remained would have increased necessitating longer stays inthe wetland patches that remained (Charnov, 1974). While wedisagreewith Broughton and colleagues assessment of the carryingcapacity for ungulates of PHT environments, we accept that, aswetlands disappeared, the Middle Holocene Great Basin wouldhave become a poorer place to reliably find large game. As suc-cessful encounters with large game declined, distance betweenpatches increased, and the more equable climate released thedampening effect on human population, the sexual division of labordiverged permanently, and women began to procure lower yield,but reliably procured resources like seeds. Handling time wouldhave contributed an increasingly high component of women'sforaging cost in patches further prolonging optimal time in patchfor women. We suggest that it is at this point that small seedsregularly entered women's prey breadth making investment ingrinding technology worthwhile (cf. Ugan et al., 2000; Bettingeret al., 2006). In contrast, men would have continued to search forincreasingly unreliably encountered, but high yield artiodactyls inpatches spatially discrete from women's foraging locales. Men
could only accommodate this conflict with greater logistic mobility.Thus the disappearance of wetlands was a critical factor in thetransition from Prearchaic to Archaic subsistence systems.
Intensive seed grinding convincingly appears only at the tran-sition to the Middle Holocene. We will not be surprised ordismayed if future investigations ultimately find PHT millingequipment, but we suggest here that the Middle Holocene prolif-eration of seed grinding implements signals a shift in sexual divi-sion of labor caused both by the desiccation of Great Basinwetlandsand the consequent effects on artiodactyl distributions. We see theproliferation of milling equipment, beginning at the onset of theMiddle Holocene, as a key signature of the reorganization of divi-sion of labor by gender among Great Basin foragers, and the shift toa permanently divergent division of labor as the primary distinc-tion between Prearchaic and Archaic subsistence strategies in theGreat Basin.
4. Conclusion
In this paper we have emphasized the importance of highmobility and convergent division of labor. However, the success ofthis strategy was predicated on several other interrelated factors,summarized in Table 1.
While we hope this contribution aids our understanding of PHTlife-ways, there are several more avenues of investigation that arerequired to fully developing our model. These include (1) greaterprecision in understanding the paleoclimatic record and how itdirectly affected environmental parameters; (2) more precise def-initions of PHT resource structure at different intervals; (3)geographical analyses of site location and structure relative toresource distributions at local and landscape scales; (4) moreresearch on animal migrations and (5) a better understanding oflithic production and use. Following these research avenues shouldprovide crucial answers to still lingering questions regarding PHTadaptations. Nonetheless, we point out that there are a number ofarchaeological testable implications of our framework as itcurrently stands, most of which correlate with both the size of thePHT wetland and the distance to the nearest alternative basin. Weexpect that:
1. Prearchaic sites tend to be oriented to streams and riparianzones feeding wetlands.
2. Accumulations of Prearchaic points bearing signs of impactfractures are most likely to appear in locations that would havebeen most amenable for intercept hunting strategies.
3. While the relative size of toolstone catchments is conditionedby toolstone availability at various scales, in intervals of the PHTwhen wetlands were most numerous and extensive, humanmobility should have been highest, toolstone catchments

R.G. Elston et al. / Quaternary International 352 (2014) 200e211210
largest, and occupational intensity and curation/recyclinglowest.
4. In the waning PHT, as wetlands diminished and distance be-tween surviving ones increased, occupational intensity shouldincrease in sites in isolated wetlands and toolstone assemblagesshould be dominated by local toolstone and have high curationindices.
5. Ground stone milling tools are most likely to appear earliest insmaller, more isolated basins, particularly during dryer climaticintervals of the PHT, but as the Middle Holocene is approached,will more frequently appear in larger wetlands.
We suggest that approaching these questions within theframework we have outlined here provides a clear avenue forfuture work. However, we emphasize the importance of keepinginformed on theoretically driven studies of modern foragers andincorporating their insights into our interpretive frameworks. Weacknowledge that, barring discoveries of larger samples of humanburials such as those at Buhl and Spirit Cave, it is unlikely that wewill be able to directly test our inferences about sexual division oflabor in the PHT Great Basin. Nonetheless, such studies, firmlygrounded within the theoretical framework of HBE and empiricallytested among contemporary foragers, provide a necessary lens forquerying an otherwise opaque and paradoxical archaeologicalrecord.
Acknowledgements
Thanks to Mark Basgall, Robert Bettinger, Darron Duke, DaveMadsen and Geoff Smith for sharing their research. Jim O'Connell,Bruce Winterhalder, Rebecca Bird, Douglas W. Bird and DaveMadsen provided comments on earlier versions of this manuscript,their insights were much appreciated. Michael Drews of GnomonInc. provided invaluable help in tracking down and crosscheckingsite files from Railroad Valley.
References
Basgall, M.E., 2005. Archeological Assessment of Two Early Holocene Sites in theNoble Pass Training Area. Marine Corps Air Ground Combat Center, Sacramento.Twentynine Palms, California Report Submitted to the U.S. Army Corps ofEngineers.
Basgall, M.E., 2008. Variation in the organization of prehistoric milling technologiesof the Northern Mojave Desert, North America. In: Rowan, Y.M., Ebeling, J.R.(Eds.), New Approaches to Old Stones: Recent Studies of Ground Stone Artifacts.Equinox Publishing Ltd, London, pp. 196e212.
Beck, C., Jones, G.T., 2009. The Archaeology of the Eastern Nevada Paleoarchaic, PartI: the Sunshine Locality. In: University of Utah Anthropological Papers No. 126.Salt Lake City.
Belovsky, G.E., 1988. An optimal foraging-based model of hunteregatherer popu-lation dynamics. Journal of Anthropological Archaeology 7, 329e372.
Bettinger, R.L., Grote, M., 2012. Marginal value theorem and hunteregatherer set-tlement response in a changing environment. In: Manuscript in Possession ofthe Authors.
Bettinger, R.L., Winterhalder, B., McElreath, R., 2006. A simple model of techno-logical intensification. Journal of Archaeological Science 33 (4), 538e545.
Binford, L.R., 1980. Willow smoke and dogs' tails: hunteregatherer settlementsystems and archaeological site formation. American Antiquity 45, 4e20.
Binford, L.R., 2001. Constructing Frames of Reference: an Analytical Method forArchaeological Theory Building Using Ethnographic and Environmental DataSets. University of California Press.
Bleige Bird, R., 1999. Cooperation and conflict: the behavioral ecology of the sexualdivision of labor. Evolutionary Anthropology 8 (2), 65e75.
Broughton, Jack M., Byers, David A., Bryson, Reid A., William, Eckerle,Madsen, David B., 2008. Did climatic seasonality control late Quaternaryartiodactyl densities in western North America? Quaternary Science Reviews27, 1916e1937.
Charnov, E.L., 1974. Optimal foraging, the marginal value theorem. TheoreticalPopulation Biology 9, 129e136.
Charnov, E.L., Orians, G., Hyatt, K., 1976. Ecological implications of resourcedepression. American Naturalist 110, 247e259.
Charnov, E.L., Orians, G.H., 1973. Optimal Foraging: Some Theoretical ExplorationsMimio. Department of Biology, University of Utah, Salt Lake City, UT.
Codding, B.F., 2012. Any Kangaroo? On the Ecology, Ethnography and Archae-ology of Foraging in Australia's Arid West Department of Anthropology.Unpublished PhD. dissertation. Department of Anthropology, Stanford Uni-versity, Stanford.
Codding, B.F., Bird, R.B., Bird, D.W., 2011. Provisioning offspring and others: risk-energy trade-offs and gender differences in hunteregatherer foraging strate-gies. Proceedings of the Royal Society B: Biological Sciences 278 (1717),2502e2509.
Duke, D.G., 2011. If the Desert Blooms: a Technological Perspective of PaleoindianEcology in the Great Basin from the Old River Bed, Utah. Department of An-thropology, Unpublished Ph.D. dissertation. University of Nevada, Reno.
Duke, D.G., King, J., 2014. A GIS model for predicting wetland habitat in the GreatBasin at the PleistoceneeHolocene Transition and implications for PaleoindianArchaeology. Journal of Archaeological Science 49, 276e291.
Duke, D.G., Young, D.C., 2007. Episodic permanence in Paleoarchaic Basin selectionand setllement. In: Graf, K.E., Schmitt, D.N. (Eds.), Paleoindian or Paleoarchaic?Great Basin Human Ecology at the PleistoceneeHolocene Transition. Universityof Utah Press, Salt Lake City, pp. 123e155.
Elston, R.G., Davis, J.O., Clerico, R., 1979. Archaeological Site 26NY1908: a LatePleistocene Gravel Bar in Railroad Valley, Nevada. Report Submitted to Bureauof Land Management, Denver Service Center. Archaeological Survey. Universityof Nevada, Reno.
Elston, R.G., Guanghui, D., Dongju, Z., 2011. Late Pleistocene intensification tech-nologies in Northern China. Quaternary International 242 (2), 401e415.
Elston, R.G., Zeanah, D.W., 2002. Thinking outside the box: a new perspective ondiet breadth and sexual division of labor in the Prearchaic Great Basin. WorldArchaeology 34 (1), 103e130.
Goebel, T., Hockett, B., Adams, K.D., Rhode, D., Graf, K., 2011. Climate, environment,and humans in North America's Great Basin during the Younger Dryas,12,900e11,600 calendar years ago. Quaternary International 242 (2), 479e501.
Goebel, Ted, Hockett, Bryan, Adams, Kenneth D., Rhode, David, Graf, Kelly, 2011.Climate, environment, and humans in North America's Great Basin during theYounger Dryas, 12,900-11,600 calendar years ago. Quaternary International 242,479e501.
Gammonley, J.H., 2004. Wildlife of Natural Palustrine Wetlands. In: McKinstry, M.C.,Humert, W.A., Anderson, S.H. (Eds.), Wetland and Riparian Areas of the Inter-mountain West Ecology and Management: Ecology and Management. Univer-sity of Utah Press, Salt Lake City, pp. 123e155.
Goodman, M.J., Griffin, P.B., Estioko-Griffin, A.A., Grove, J.S., 1985. The compatibilityof hunting and mothering among the Agta hunteregatherers of the Philippines.Sex Roles 12 (11e12), 1199e1209.
Grayson, D.K., 2011. The Great Basin: a Natural Prehistory. University of CaliforniaPress, Berkeley.
Gurven, M., Hill, K., 2009. Why do men hunt? A reevaluation of “Man the Hunter”and the sexual division of labor. Current Anthropology 50 (1), 51e62.
Hawkes, K., 1990. Why do men hunt? Some benefits for risky strategies. In:Cashdan, E. (Ed.), Risk and Uncertainty in Tribal and Peasant Economies.Westview Press, Boulder, pp. 145e166.
Hawkes, K., 1996. The evolutionary basis of sex variations in the use of naturalresources: human examples. Population and Environment 18 (2), 161e173.
Hawkes, K., Hill, K., O'Connell, J.F., 1982. Why hunters gather: optimal foraging andthe ache of eastern Paraguay. American Ethnologist 9 (2), 379e398.
Hawkes, K., Kaplan, H., Hill, K., Hurtado, A.M., 1987. Ache at the settlement: con-trasts between farming and foraging. Human Ecology 15 (2), 133e161.
Hawkes, K., O'Connell, J.F., Jones, N.G.B., 1991. Hunting income patterns among theHadza: big game, common goods, foraging goals and the evolution of the hu-man diet philosophical transactions. Biological Sciences 334 (1270), 243e251.
Hockett, B., 2007. Nutritional ecology of Late Pleistocene to Middle Holocene Sub-sistence. In: Graf, K.E., Schmitt, D.N. (Eds.), Paleoindian or Paleoarchaic? GreatBasin Human Ecology at the PleistoceneeHolocene Transition. University ofUtah Press, Salt Lake City, pp. 204e230.
Janetski, J.C., Bodily, M.L., Newbold, B.A., Yoder, D.T., 2012. The Paleoarchaic to EarlyArchaic Transition on the Colorado Plateau: the Archaeology of North creekshelter. American Antiquity 77 (1), 125e159.
Jarvenpa, R., Brumbach, H.J., 2006. Circumpolar Lives and Livelihood: a ComparativeEthnoarchaeology of Gender and Subsistence. University of Nebraska Press,Lincoln.
Jenkins, D.L., Davis, L.G., Stafford, T.W., Campos, P.F., Hockett, B., Jones, G.T.,Cummings, L.S., Yost, C., Connolly, T.J., Yohe, R.M., Gibbons, S.C., Raghavan, M.,Rasmussen, M., Paijmans, J.L.A., Hofreiter, M., Kemp, B.M., Barta, J.L., Monroe, C.,Gilbert, M.T.P., Willerslev, E., 2012. Clovis age western stemmed projectilepoints and human Coprolites at the Paisley caves. Science 337 (6091), 223e228.
Jennings, J.D., 1957. Danger Cave. In: Anthropological Papers Number 27. Universityof Utah Press.
Jones, G., Fontes, L., Horowitz, R., Beck, C., Bailey, D., 2012. Reconsidering Paleo-archaic mobility in the Central Great Basin. American Antiquity 77 (2), 351e367.
Kelly, R.L., 1995. Hunteregatherer lifeways in the Carson desert: a context for bio-archaeology. In: Larsen, C.S., Kelly, R.L. (Eds.), Bioarchaeology of the StillwaterMarsh: Prehistoric Adaptation in the Western Great Basin, vol. 77. AmericanMuseum of Natural History Anthropological Papers, New York, pp. 12e32.
Kelly, R.L., 2013. The Lifeways of HuntereGatherers: the Foraging Spectrum. Cam-bridge University Press.
Kirner, D.L., Burky, R., Selsor, K., George, D., Taylor, R.E., Southon, J.R., 1997. Datingthe Spirit Cave mummy: the value of reexamination. Nevada Historical SocietyQuarterly 40, 54.

R.G. Elston et al. / Quaternary International 352 (2014) 200e211 211
Lafayette, L., Smith, G.M., 2012. Use-wear traces on experimental (replicated) andprehistoric stemmed points in the Great Basin. Journal of California and GreatBasin Anthropology 32 (2), 141e160.
Leckenby, D.A., Sheehy, D.P., Nellis, C.H., Scherzinger, R.J., Luman, I.D., Elmore, W.,Lemos, J.C., Doughty, L., Trainer, C.E., 1982. Wildlife Habitats in Managed Ran-gelands d the Great Basin of Southeastern Oregon: Mule Deer. U.S. ForestService General Technical Report PNW-139.
Lee,R.B.,1968.WhatHuntersdofora living,orhowtomakeoutonscarceresources. In:Lee, R.B., DeVore, I. (Eds.), Man the Hunter. Aldine, Chicago, pp. 30e48.
Lee, R.B., 1979. The !Kung San: Men, Women and Work in a Foraging Society.Cambridge University Press.
Lohman, K., 2004. Wildlife use of riverine wetland habitats. In: McKinstry, M.C.,Humert, W.A., Anderson, S.H. (Eds.), Wetland and Riparian Areas of the Inter-mountain West: Ecology and Management. University of Texas, Austin,pp. 74e86.
Madsen, D.B., 2007. The Paleoachaic to archaic transition in the Great Basin. In:Graf, K.E., Schmitt, D.N. (Eds.), Paleoindian or Paleoarchaic? Great Basin HumanEcology at the PleistoceneeHolocene Transition. University of Utah Press, SaltLake City, pp. 3e20.
Madsen, David B., Schmitt, Dave N., Page, David, 2015. The Paleoarchic Occupationof the Old River Bed Delta. In: Anthropological Papers 128. University of Utah.
Marcum, C.L., Scott, M.D., 1985. Influences of weather on Elk use of springesummerhabitat. The Journal of Wildlife Management 49 (1), 73e76.
Marlowe, F.W., 2007. Hunting and gathering: the human sexual division of foraginglabor. Cross-Cultural Research 41 (2), 170e195.
Marlowe, F.W., 2010. The Hadza. University of California Press, Berkeley.McGuire, K.R., 1985. Test excavation at Sheepy East 1, Lower Klamath Lake, Siskiyou
County, California. Far Western Anthropological Research Group, Inc., Sacra-mento, California. Report submitted to U. S. Bureau of Reclamation.
Middleton, E.S., Smith, G.M., Cannon, W.J., Ricks, M.F., 2014. Paleoindian rock art:establishing the antiquity of Great Basin Carved Abstract petroglyphs in thenorthern Great Basin. Journal of Archaeological Science 43 (0), 21e30.
Mullins, D., Adams, J., Ligman, M., Zingg, C., 2013. A GIS Predictive Model andSample Inventory of 2,150 Acres in Delamar Valley, Lincoln County, Nevada. LSDTechnical Report No.: 105461. Logan Simpson Design Inc., Salt Lake City, UT,p. 148.
O'Connell, J.F., Hawkes, K., Jones, N.B., 1988. Hadza hunting, butchering, and bonetransport and their archaeological implications. Journal of AnthropologicalResearch 44 (2), 113e161.
Pelissier, R., Goreaud, F., 2010. Spatial Point Patterns Analysis v.1.2-10 ads R Library.Pinson, A., 1999. Foraging in Uncertain Times: the Effects of Risk on Subsistence
Behavior during the PleistoceneeHolocene Transition in the Oregon Great Ba-sin. PhD. dissertation. Anthropology Department, University of New Mexico,Albuquerque.
Pinson, A., 2007. Artiodactyl use and adaptive discontinuity across the Paleoarchaic/Archaic Transition in the Northern Great Basin. In: Graf, K.E., Schmitt, D.N.(Eds.), Paleoindian or Paleoarchaic?: Great Basin Human Ecology at the Pleis-toceneeHolocene Transition. University of Utah Press, Salt Lake City,pp. 187e203.
R, 2014. A Language and Environment for Statistical Computing. Vienna.Raedeke, K.J., Taber, R.D., Paige, D.K., 1988. Ecology of large mammals in riparian
systems of Pacific Northwest forests. In: Raedeke, K.J. (Ed.), Streamside Man-agement: Riparian Wildlife and Forestry Interactions, 59. University of Wash-ington Institute of Forest Resources, Seattle.
Raven, C., Elston, R.G., 1989. Prehistoric Human Geography in the Carson Desert.Part I: a Predictive Model of Land Use in the Stillwater Wildlife ManagementArea. U.S. Fish and Wildlife Service, Region 1, Portland, Oregon.
Rhode, D., Louderback, L.A., 2007. Dietary plant use in the Bonneville basin duringthe terminal Pleistocene/early Holocene transition. In: Graf, K.E., Schmitt, D.N.(Eds.), Paleoindian or Paleoarchaic?: Great Basin Human Ecology at the
PleistoceneeHolocene Transition. University of Utah Press, Salt Lake City,pp. 231e247.
Rhode, D., Madsen, D.B., Jones, K.T., 2006. Antiquity of early Holocene small-seedconsumption and processing at Danger Cave. Antiquity 80 (308), 328e339.
Scheffer, M., Carpenter, S.R., Lenton, T.M., Bascompte, J., Brock, W., Dakos, V., van deKoppel, J., van de Leemput, I.A., Levin, S.A., van Nes, E.H., Pascual, M.,Vandermeer, J., 2012. Anticipating critical transitions. Science 338, 344e348.
Schmitt, D.N., Madsen, D.B., Oviatt, C.G., Quist, R., 2007. Late Pleistocene/Early Ho-locene Geomorphology and human occupation of the Old River Bed Delta,Western Utah. In: Graf, K.E., Schmitt, D.N. (Eds.), Paleoindian or Paleoarchaic?:Great Basin Human Ecology at the Pleistocene/Holocene. University of UtahPress, Salt Lake City, pp. 105e122.
Simms, S.R., 1987. Behavioral Ecology and HuntereGatherer Foraging: an Examplefrom the Great Basin. British Archaeological Reports. International Series,Oxford.
Smith, G.M., 2007. Prearchaic mobility and technological activities at the ParmanLocalities, Humboldt County, Nevada. In: Graf, K.E., Schmitt, D.N. (Eds.), Paleo-indian or Paleoarchaic? Great Basin Human Ecology at the Pleistoce-neeHolocene Transition. University of Utah Press, Salt Lake City, pp. 139e155.
Smith, G.M., 2010. Footprints across the black rock: temporal variability in prehis-toric foraging territories and toolstone procurement strategies in the westernGreat Basin. American Antiquity 75 (4), 865e885.
Smith, G.M., 2011. Shifting stones and changing homes: using toolstone ratios toconsider relative occupation span in the northwestern Great Basin. Journal ofArchaeological Science 38 (2), 461e469.
Smith, G.M., Kielhofer, J., 2011. Through the High Rock and beyond: placing the LastSupper Cave and Parman Paleoindian lithic assemblages into a regional context.Journal of Archaeological Science 38 (12), 3568e3576.
Stevens, N.E., 2012. Technological Plascticity and Cultural Evolution along theCentral Coast of California. PhD. dissertation. Department of Anthropology,University of California, Davis.
Stevens, N.E., Codding, B.F., 2009. Inferring the function of stone points from Cal-ifornia's Central Coast. California Archaeology 1, 7e28.
Steward, J., 1938. Basin-Plateau Aboriginal Sociopolitical Groups. In: Bulletin 120.Bureau of American Ethnology, Washington D.C.
Surovell, T.A., Finley, J.B., Smith, G.M., Brantingham, J.P., Kelly, R., 2009. Correctingtemporal frequency distributions for taphonomic bias. Journal of ArchaeologicalScience 36, 1715e1724.
Ugan, A., Bright, J., Rogers, A., 2000. When is technology worth the trouble? Journalof Archaeological Science 30 (10), 1315e1329, 2003.
Walmo, O.C., 1981. Mule and Black-tailed Deer of North America: a Wildlife Man-agement Institute Book. University of Nebraska Press, Lincoln, Nebraska.
Winterhalder, B., Baillargeon, W., Cappelletto, F., Randolph Daniel Jr., I., Prescott, C.,1988. The population ecology of hunteregatherers and their prey. Journal ofAnthropological Archaeology 7 (4), 289e328.
Witmer, G.W., deCalesta, D.S., 1983. Habitat use by female Roosevelt Elk in theOregon Coast Range. The Journal of Wildlife Management 47 (4), 933e939.
Yoder, D.T., Bodily, M.L., Hill, S., Janetski, J.C., Newbold, B.A., 2010. The onset of smallseed processing on the Colorado Plateau. Kiva 75, 425e445.
Zancanella, J.K., 1984. A Cultural Resource Inventory of Land Applied for under theDesert Land Act and Carey Act Applications in Northern Railroad Valley, Nyeand White Pine Counties Nevada. Report on File.
Zeanah, D., Carter, J.A., Elston, R.G., Hammett, J.E., 1995. An Optimal Foraging Modelof HuntereGatherer Land Use in the Carson Desert. Intermountain Research.
Zeanah, D.W., Ingbar, E.E., Zeier, C., 1999. Archaeological Predictive Model, Man-agement Plan and Treatment Plan for Northern Railroad Valley, Nevada. Inter-mountain Research.
Zeanah, D.W., 2004. Sexual division of labor and central place foraging: a model for theCarson Desert ofwestern Nevada. Journal of Anthropological Archaeology 23,1e32.




















