
Diet and Dietary-Niche Breadth of Diurnal Rain Forest Primates in the Central Western Ghats, India
16
Original Article Folia Primatol 2011;82:283–298 DOI: 10.1159/000336964 Diet and Dietary-Niche Breadth of Diurnal Rain Forest Primates in the Central Western Ghats, India Kuladeep Roy a Mewa Singh a Mridula Singh b a Department of Psychology and b Maharaja’s College, University of Mysore, Mysore, India Key Words Western Ghats Feeding ecology Niche breadth Lion-tailed macaques Bonnet macaques Hanuman langurs Abstract We studied the feeding ecology of 3 sympatric primate species, the lion-tailed ma- caque (Macaca silenus) , bonnet macaque (Macaca radiata) and Hanuman langur (Sem- nopithecus entellus) , in a tropical rain forest of the Central Western Ghats, India. Since the availability of leaves is much higher than that of fruits, we expected that the primarily folivorous langurs would use a larger number of resources than the primarily frugivo- rous macaques. Since fruits are a relatively total resource, unlike leaves, of which only selected parts are consumed, we expected that the primarily frugivorous macaques would use resources more proportionately than the folivorous langurs, resulting in a wider food niche breadth in macaques. We collected data on these primate species over a period of 2 years using scan sampling in a contiguous rain forest. We calculated Levin’s standardised food niche breadth for each species. Langurs used a larger number of tree species than macaques, but their niche breadth was narrower than that of macaques. The majority of their diet (over 50%) came from only a few trees in the case of all three primate species. Long-term data are provided herein for the first time on the feeding ecology of bonnet macaques and Hanuman langurs in a tropical rain forest. Copyright © 2012 S. Karger AG, Basel Introduction The Hanuman langur is one of the most widely distributed primates in the In- dian subcontinent, inhabiting a variety of regions ranging from deserts to rain for- ests and from sea level up to 4,000 m in the Himalayas; the bonnet macaque is en- Received: October 24, 2011 Accepted after revision: January 27, 2012 Published online: March 30, 2012 Mewa Singh Department of Psychology University of Mysore Mysore (India) E-Mail mewasingh @ bsnl.in © 2012 S. Karger AG, Basel 0015–5713/11/0826–0283$38.00/0 Accessible online at: www.karger.com/fpr Fax +41 61 306 12 34 E-Mail [email protected] www.karger.com
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Diet and Dietary-Niche Breadth of Diurnal Rain Forest Primates in the Central Western Ghats, India

Original Article
Folia Primatol 2011;82:283–298 DOI: 10.1159/000336964
Diet and Dietary-Niche Breadth of Diurnal
Rain Forest Primates in the Central Western
Ghats, India
Kuladeep Roy a Mewa Singh a Mridula Singh b
a Department of Psychology and b Maharaja’s College, University of Mysore, Mysore , India
Key Words
Western Ghats � Feeding ecology � Niche breadth � Lion-tailed macaques � Bonnet macaques � Hanuman langurs
Abstract
We studied the feeding ecology of 3 sympatric primate species, the lion-tailed ma-caque (Macaca silenus) , bonnet macaque (Macaca radiata) and Hanuman langur (Sem-nopithecus entellus) , in a tropical rain forest of the Central Western Ghats, India. Since the availability of leaves is much higher than that of fruits, we expected that the primarily folivorous langurs would use a larger number of resources than the primarily frugivo-rous macaques. Since fruits are a relatively total resource, unlike leaves, of which only selected parts are consumed, we expected that the primarily frugivorous macaques would use resources more proportionately than the folivorous langurs, resulting in a wider food niche breadth in macaques. We collected data on these primate species over a period of 2 years using scan sampling in a contiguous rain forest. We calculated Levin’s standardised food niche breadth for each species. Langurs used a larger number of tree species than macaques, but their niche breadth was narrower than that of macaques. The majority of their diet (over 50%) came from only a few trees in the case of all three primate species. Long-term data are provided herein for the first time on the feeding ecology of bonnet macaques and Hanuman langurs in a tropical rain forest.
Copyright © 2012 S. Karger AG, Basel
Introduction
The Hanuman langur is one of the most widely distributed primates in the In-dian subcontinent, inhabiting a variety of regions ranging from deserts to rain for-ests and from sea level up to 4,000 m in the Himalayas; the bonnet macaque is en-
Received: October 24, 2011 Accepted after revision: January 27, 2012 Published online: March 30, 2012
Mewa Singh Department of Psychology University of Mysore Mysore (India) E-Mail mewasingh @ bsnl.in
© 2012 S. Karger AG, Basel0015–5713/11/0826–0283$38.00/0
Accessible online at:www.karger.com/fpr
Fax +41 61 306 12 34E-Mail [email protected]
muellers
Stempel Hoch

Folia Primatol 2011;82:283–298284 Roy /Singh /Singh
demic to southern India, but it is found in towns, temples, roadsides and villages, deciduous forests and rain forests, and the lion-tailed macaque is a typical habitat specialist of the tropical rain forests of the Western Ghats [Kumar, 1987; Ramachan-dran and Joseph, 2000; Singh et al., 2000; Molur et al., 2003]. The feeding ecology of lion-tailed macaques has been described for several regions of the southern Western Ghats. Likewise, the feeding ecology of bonnet macaques and Hanuman langurs has been described for dry-deciduous forests and human-dominated landscapes. In ad-dition to the consumption of a variety of fruits, lion-tailed macaques and bonnet macaques also feed extensively on faunal items, such as arthropods and even some vertebrates (such as bird nestlings, young giant squirrels, Calotes sp., frogs and frog nests) [Kumar, 1987; Krishnamani, 1994; Umapathy, 1997; Kumara et al., 2000; Sushma and Singh, 2006]. Hanuman langurs are mostly folivorous [Hladik, 1977; Sri-vastava, 1992; Koenig, 1996; Koenig et al., 1997; Wasserman and Chapman, 2003].
In none of the studies so far has the feeding ecology of Hanuman langurs and bonnet macaques, inhabiting a rain forest region, been described in detail. It has been shown that the dietary study of one group in a specific habitat and at one point in time only may not give a complete representation of the diet of a species as a whole [Chapman et al., 2004]. In order to get a better understanding of the dietary diver-sity of these primates, we selected a region of the Western Ghats where lion-tailed macaques, bonnet macaques and Hanuman langurs are sympatric in the rain forests. In an earlier study [Singh et al., 2011], we reported that, in this region of the Western Ghats, the food niche overlap between the three primate species over the year was very small, except between bonnet macaques and lion-tailed macaques for a few months when resources were abundant. Since the availability of leaves is much high-er than that of fruits, we expected that the primarily folivorous langurs would use a larger number of resources than the primarily frugivorous macaques. Furthermore, since fruits are a total resource, unlike leaves, of which only selected parts are con-sumed, we expected that the primarily frugivorous macaques would use resources more proportionately than the folivorous langurs, resulting in a wider food niche breadth in macaques than in langurs.
Methods
Study Area and Study Species We conducted this study on the northern limit of the evergreen rain forest of the plains
and low elevations of the Western Ghats of South India [Pascal, 1988] in the Gerusoppa Forest Range (74°42 � 00 � � E to 74°46 � 30 � � E and 14°15 � 15 � � N to 14°19 � 15 � � N), Honnavara Forest Divi-sion, Uttara Kanara district in the state of Karnataka ( fig. 1 ). The annual temperature in this region ranges between a maximum of 33 ° C in April and a minimum of 20 ° C in January. The area receives monsoon rains from the end of May until October, with an annual average of 6,000 mm. July is the rainiest month, with an average rainfall of almost 3,000 mm with 80% average humidity. Champion and Seth [2005] broadly classify this forest as ‘west coast tropical ever-green forest’. In this forest, there is an abundance of heliophilic or light-tolerant species, which may be distinguished as associations of Persea macrantha and species of Diospyros / Holigarna . The forest has legal status as a ‘reserve forest’, with revenue lands interspersed [Kumara and Singh, 2004]. We conducted this study between April 2006 and December 2008.
We selected 1 group each of lion-tailed macaques (n = 21) and Hanuman langurs (n = 7) and 2 groups of bonnet macaques (n = 11; n = 16) ranging in the forests surrounding the village of Matnigadde. All 4 study groups had overlapping home ranges.

Diet of Diurnal Rain Forest Primates in the Central Western Ghats
285Folia Primatol 2011;82:283–298
Data Collection We employed instantaneous scan sampling [Altmann, 1974; Lehner, 1996] to record the
feeding behaviour of individual animals, for a period of 5-min sampling with a 10-min interval during which each individual was observed and the information was collected on a preformat-ted data sheet, recording study species, individual identity, individual activity, substratum used (only when the individual fed on faunal components), plant species (when the individual fed on plant food), plant part eaten and phenophase of the plant part. The faunal components were broadly classified as invertebrates in all figures and tables. We followed each group of each spe-cies of primate for about 5 days every month and collected the data from 6.00 to 18.00 h. We considered November to April as dry season and May to October as wet season.
We collected the plant parts dropped by the animals to ascertain the edible part of the item. We identified the plant species with the help of field guides [Gamble and Fischer, 1916–1935; Pascal and Ramesh, 1997] and, for confirmation, we consulted plant taxonomists of the Department of Forest Biology, College of Forestry, Sirsi, India. We identified most plants to the species level.
Data Analysis We used the � 2 test [Sheskin, 2000] to test for the significance of differences between the
use of resource categories and resource types preferred by the primates, and employed the Z test for proportions to test the seasonal variation in resources and resource consumptions. We con-sidered each plant food item, with its phenophase, as a ‘resource’. For example, if a primate spe-cies fed on both unripe and ripe fruits of a particular tree species, it was considered as two re-
Maharashtra
GoaKarnataka
Forest Division boundary
Aghanashini LTM Conservation Reserve
KeralaTamil Nadu
Andhra Pradesh
74°30'E14
°30'
N14
°20'
N
14°3
0'N
14°2
0'N
74°40'E 74°50'E
74°30'E 74°40'E 74°50'E
Honnavar Forest Division
Sirsi Forest Division
N
50 10km
Fig. 1. Map of the study area.
Co
lor v
ersi
on
avai
lab
le o
nlin
e

Folia Primatol 2011;82:283–298286 Roy /Singh /Singh
sources. For analysis, we combined partially ripe fruit with unripe fruit and leaf flush with young leaves. The major food items from the plant resources were considered as those which were observed to be consumed by a primate species in at least 1% of the feeding scans. We used the data on proportional use of resources to calculate Levin’s standardised measure of niche breadth [Hurlbert, 1978] as follows: B A = (B – 1)/(n – 1) where B A is Levin’s standardised niche breadth, B is Levin’s measure of niche breadth and n is the number of resources. Levin’s measure of niche breadth was calculated as B = 1/ � p j 2 where p j is the proportion of resource j in the diet of the animal species. Niche breadth values are standardised on a scale from 0 to 1 [Hurlbert, 1978].
Results
Feeding Ecology of All Three Primate Species Hanuman langurs used 233 resources from 72 plant species, bonnet macaques
used 75 resources from 37 plant species and lion-tailed macaques used 57 resources from 24 plant species.
Fruits were the major food resource of lion-tailed macaques (78.1% of the total diet), followed by faunal items (13.4%), flowers (6.3%) and ‘other’ items (2.3%; fig. 2 ). The difference in the rate of consumption of these items was significant ( � 2 = 1,077.41, d.f. = 3, p ! 0.01), but there was no seasonal difference in the frequency of use of fruits (z = 0.713, n.s.), flowers (z = 0.445, n.s.), insects (z = 0.135, n.s.) and other food items (z = 1.564, n.s.; fig. 2 ). Overall, lion-tailed macaques preferred ripe fruits (65.9%) to unripe fruits (34.1%; � 2 = 70.74, d.f. = 1, p ! 0.01). However, the consumption of un-ripe fruits was not significantly different between the dry season (34.8%) and the wet season (32.7%; z = 0.53, n.s.). Similarly, the consumption of ripe fruits did not differ
0
Overall Dry season Wet season
Frui
ts
Flow
ers
Inse
cts
Cro
ps
Oth
er
Leav
es
Frui
ts
Flow
ers
Inse
cts
Cro
ps
Oth
er
Leav
es
Frui
ts
Flow
ers
Inse
cts
Oth
er
Cro
ps
Leav
es
Occ
urri
ng
in p
erce
nt
scan
s
10.00
20.00
30.00
40.00
50.00
60.00
70.00
80.00
90.00 Hanuman langurLion-tailed macaqueBonnet macaque
Fig. 2. Different types of resource used by all three study species in different seasons and over the year.

Diet of Diurnal Rain Forest Primates in the Central Western Ghats
287Folia Primatol 2011;82:283–298
between the dry season (65.2%) and the wet season (67.3%) either (z = 0.53, n.s.). Lion-tailed macaques were not observed feeding on leaves or cultivated crops.
The consumption rates of fruits (48.9%), flowers (26.7%), crops (7.7%), inverte-brates (6.8%), leaves (3.6%) and ‘other’ (including pith, dry and green bark) food re-sources (6.2%; fig. 2 ) in bonnet macaques differed significantly ( � 2 = 983.86, d.f. = 5, p ! 0.01). Crops and leaves were consumed by bonnet macaques exclusively in thedry season. The consumption of fruits and invertebrates was significantly higher during the wet season as compared to the dry season (z = 11.94, p ! 0.01, and z = 2.23, p ! 0.05, respectively; fig. 2 ). On the other hand, bonnet macaques consumed flowers and ‘other’ food items with a higher frequency during the dry season than in the wet season (z = 7.65, p ! 0.01, and z = 3.87, p ! 0.01, respectively; fig. 2 ). Although consti-tuting only a small portion of their diet, bonnet macaques consumed more young leaves (86.8%) than mature leaves (13.2%; � 2 = 20.63, d.f. = 1, p ! 0.01). Bonnet ma-caques consumed more ripe fruits (66.7%) than unripe fruits (33.3%; � 2 = 73.67, d.f. = 1, p ! 0.01). Although they consumed more unripe fruits in the dry season (41.1%) than in the wet season (21.8%; z = 5.15, p ! 0.01), the consumption of ripe fruits was higher in the wet season (78.2%) than in the dry season (58.9%; z = 5.15, p ! 0.01).
Hanuman langurs fed on leaves (62.1%), fruits (32.2%), flowers (4.1%) and ‘oth-er’ items (including pith, bark and tendrils, 1.7%; fig. 2 ), and the consumption dif-fered significantly ( � 2 = 2,304.72, d.f. = 3, p ! 0.01). Hanuman langurs did not feed on faunal items and cultivated crops. Leaves and flowers ( fig. 2 ) were consumed more in the wet season than in the dry season (z = 8.30, p ! 0.01, and z = 3.74, p ! 0.01, respectively), but Hanuman langurs consumed more fruits in the wet season than in the dry season (z = 10.11, p ! 0.01; fig. 2 ). They consumed more young leaves (64.0%) than mature leaves (36.0%; � 2 = 117.78, d.f. = 1, p ! 0.01). The consumption of young leaves was not significantly different between the dry season (65.1%) and the wet sea-son (60.6%; z = 1.612, p ! 0.01). There was no significant difference in the consump-tion of mature leaves in the dry season (34.9%) and in the wet season (39.4%; z = 1.612, p ! 0.01). Hanuman langurs consumed more ripe fruits (68.3%) than unripe fruits (31.7%; � 2 = 110.69, d.f. = 1, p ! 0.01). They consumed more unripe fruits in the dry season (35.9%) than in the wet season (26.5%; z = 2.881, p ! 0.01), but they con-sumed more ripe fruits in the wet season (73.5%) than in the dry season (64.1%; z = 2.881, p ! 0.01).
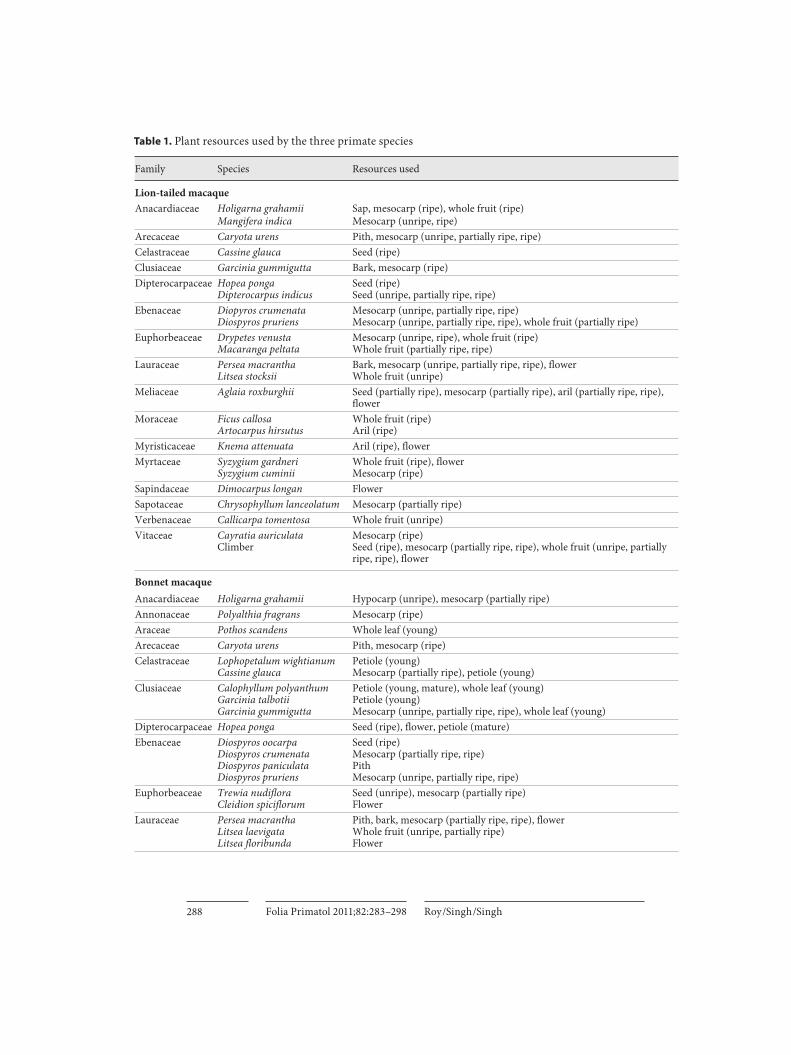
All food plant species and the resources used by lion-tailed macaques, bonnet macaques and Hanuman langurs are shown in table 1 , and the seasonal and yearly selective resources are shown in table 2 . Only 15 plant species, including climbers, provided 50% of the total diet of all three primate species ( table 3 ).
Food Niche Breadth and Comparative Feeding Ecology The overall niche breadths for the major resources of Hanuman langurs, lion-
tailed macaques and bonnet macaques were 0.52, 0.68 and 0.68, respectively. Hanu-man langurs and bonnet macaques had wider niches during the wet season (0.52 for Hanuman langurs and 0.64 for bonnet macaques) than in the dry season (0.43 for Hanuman langurs and 0.57 for bonnet macaques). In the case of lion-tailed ma-caques, the niche narrowed during the wet season (0.41) compared to the dry season (0.58).
In terms of comparative foraging, Hanuman langurs consumed more leaves (62.1%) than bonnet macaques (3.6%; � 2 = 993.87, d.f. = 1, p ! 0.01; fig. 2 ). During the
Folia Primatol 2011;82:283–298288 Roy /Singh /Singh
Family Species Resources used
Lion-tailed macaqueAnacardiaceae Holigarna grahamii Sap, mesocarp (ripe), whole fruit (ripe)
Mangifera indica Mesocarp (unripe, ripe)Arecaceae Caryota urens Pith, mesocarp (unripe, partially ripe, ripe)Celastraceae Cassine glauca Seed (ripe)Clusiaceae Garcinia gummigutta Bark, mesocarp (ripe)Dipterocarpaceae Hopea ponga
Dipterocarpus indicusSeed (ripe)Seed (unripe, partially ripe, ripe)
Ebenaceae Diopyros crumenataDiospyros pruriens
Mesocarp (unripe, partially ripe, ripe)Mesocarp (unripe, partially ripe, ripe), whole fruit (partially ripe)
Euphorbeaceae Drypetes venustaMacaranga peltata
Mesocarp (unripe, ripe), whole fruit (ripe)Whole fruit (partially ripe, ripe)
Lauraceae Persea macranthaLitsea stocksii
Bark, mesocarp (unripe, partially ripe, ripe), flowerWhole fruit (unripe)
Meliaceae Aglaia roxburghii Seed (partially ripe), mesocarp (partially ripe), aril (partially ripe, ripe), flower
Moraceae Ficus callosaArtocarpus hirsutus
Whole fruit (ripe)Aril (ripe)
Myristicaceae Knema attenuata Aril (ripe), flowerMyrtaceae Syzygium gardneri
Syzygium cuminiiWhole fruit (ripe), flowerMesocarp (ripe)
Sapindaceae Dimocarpus longan FlowerSapotaceae Chrysophyllum lanceolatum Mesocarp (partially ripe)Verbenaceae Callicarpa tomentosa Whole fruit (unripe)Vitaceae Cayratia auriculata
Climber Mesocarp (ripe)Seed (ripe), mesocarp (partially ripe, ripe), whole fruit (unripe, partially ripe, ripe), flower
Bonnet macaqueAnacardiaceae Holigarna grahamii Hypocarp (unripe), mesocarp (partially ripe)Annonaceae Polyalthia fragrans Mesocarp (ripe)Araceae Pothos scandens Whole leaf (young)Arecaceae Caryota urens Pith, mesocarp (ripe)Celastraceae Lophopetalum wightianum
Cassine glaucaPetiole (young)Mesocarp (partially ripe), petiole (young)
Clusiaceae Calophyllum polyanthumGarcinia talbotiiGarcinia gummigutta
Petiole (young, mature), whole leaf (young)Petiole (young)Mesocarp (unripe, partially ripe, ripe), whole leaf (young)
Dipterocarpaceae Hopea ponga Seed (ripe), flower, petiole (mature)Ebenaceae Diospyros oocarpa
Diospyros crumenataDiospyros paniculataDiospyros pruriens
Seed (ripe)Mesocarp (partially ripe, ripe)PithMesocarp (unripe, partially ripe, ripe)
Euphorbeaceae Trewia nudifloraCleidion spiciflorum
Seed (unripe), mesocarp (partially ripe)Flower
Lauraceae Persea macranthaLitsea laevigataLitsea floribunda
Pith, bark, mesocarp (partially ripe, ripe), flowerWhole fruit (unripe, partially ripe)Flower
Table 1. P lant resources used by the three primate species
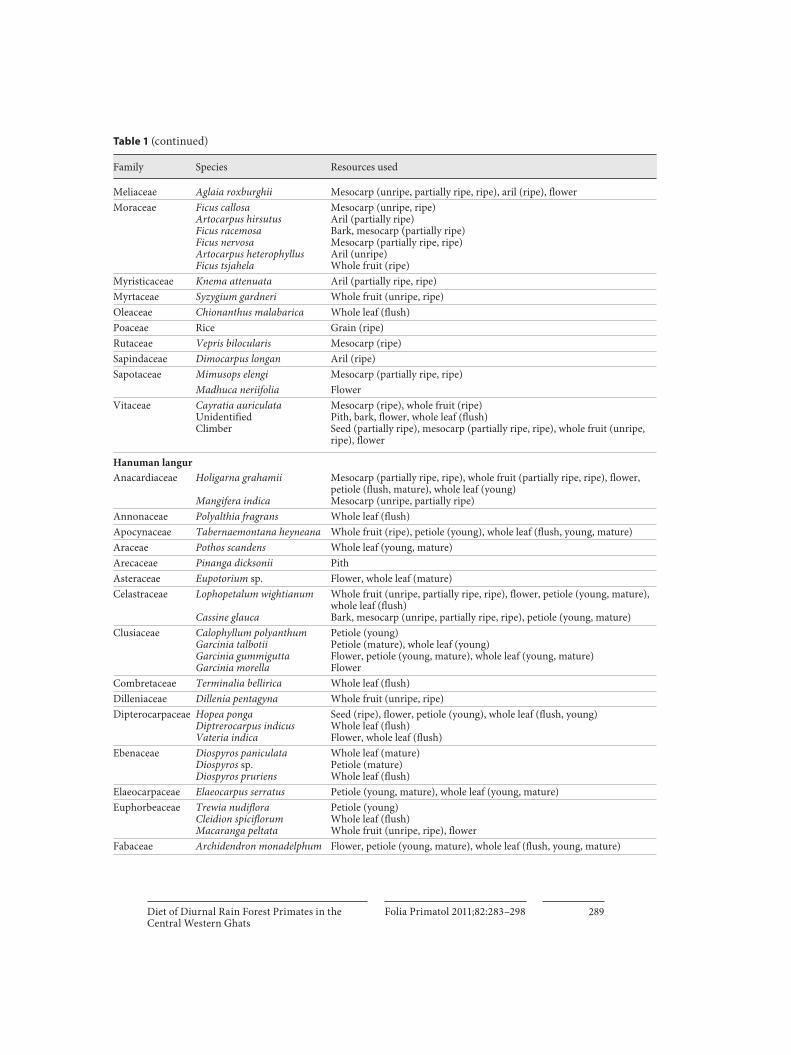
Diet of Diurnal Rain Forest Primates in the Central Western Ghats
289Folia Primatol 2011;82:283–298
Family Species Resources used
Meliaceae Aglaia roxburghii Mesocarp (unripe, partially ripe, ripe), aril (ripe), flowerMoraceae Ficus callosa
Artocarpus hirsutusFicus racemosaFicus nervosaArtocarpus heterophyllusFicus tsjahela
Mesocarp (unripe, ripe)Aril (partially ripe)Bark, mesocarp (partially ripe)Mesocarp (partially ripe, ripe)Aril (unripe)Whole fruit (ripe)
Myristicaceae Knema attenuata Aril (partially ripe, ripe)Myrtaceae Syzygium gardneri Whole fruit (unripe, ripe)Oleaceae Chionanthus malabarica Whole leaf (flush)Poaceae Rice Grain (ripe)Rutaceae Vepris bilocularis Mesocarp (ripe)Sapindaceae Dimocarpus longan Aril (ripe)Sapotaceae Mimusops elengi Mesocarp (partially ripe, ripe)
Madhuca neriifolia FlowerVitaceae Cayratia auriculata
UnidentifiedClimber
Mesocarp (ripe), whole fruit (ripe)Pith, bark, flower, whole leaf (flush)Seed (partially ripe), mesocarp (partially ripe, ripe), whole fruit (unripe, ripe), flower
Hanuman langurAnacardiaceae Holigarna grahamii
Mangifera indica
Mesocarp (partially ripe, ripe), whole fruit (partially ripe, ripe), flower, petiole (flush, mature), whole leaf (young)Mesocarp (unripe, partially ripe)
Annonaceae Polyalthia fragrans Whole leaf (flush)Apocynaceae Tabernaemontana heyneana Whole fruit (ripe), petiole (young), whole leaf (flush, young, mature)Araceae Pothos scandens Whole leaf (young, mature)Arecaceae Pinanga dicksonii PithAsteraceae Eupotorium sp. Flower, whole leaf (mature)Celastraceae Lophopetalum wightianum
Cassine glauca
Whole fruit (unripe, partially ripe, ripe), flower, petiole (young, mature), whole leaf (flush)Bark, mesocarp (unripe, partially ripe, ripe), petiole (young, mature)
Clusiaceae Calophyllum polyanthumGarcinia talbotiiGarcinia gummiguttaGarcinia morella
Petiole (young)Petiole (mature), whole leaf (young)Flower, petiole (young, mature), whole leaf (young, mature)Flower
Combretaceae Terminalia bellirica Whole leaf (flush)Dilleniaceae Dillenia pentagyna Whole fruit (unripe, ripe)Dipterocarpaceae Hopea ponga
Diptrerocarpus indicusVateria indica
Seed (ripe), flower, petiole (young), whole leaf (flush, young)Whole leaf (flush)Flower, whole leaf (flush)
Ebenaceae Diospyros paniculataDiospyros sp.Diospyros pruriens
Whole leaf (mature)Petiole (mature)Whole leaf (flush)
Elaeocarpaceae Elaeocarpus serratus Petiole (young, mature), whole leaf (young, mature)Euphorbeaceae Trewia nudiflora
Cleidion spiciflorumMacaranga peltata
Petiole (young)Whole leaf (flush)Whole fruit (unripe, ripe), flower
Fabaceae Archidendron monadelphum Flower, petiole (young, mature), whole leaf (flush, young, mature)
Table 1 (continued)

Folia Primatol 2011;82:283–298290 Roy /Singh /Singh
Family Species Resources used
Flacourtiaceae Hydnocarpus pentandraFlacourtia montana
Mesocarp (partially ripe, ripe)Whole leaf (flush)
Lauraceae Persea macranthaLitsea sp.Cinnamomum malabathrumLitsea laevigataActinodaphne hookeriLitsea floribundaLitsea stocksii
Mesocarp (unripe, ripe), flowerPetiole (young, mature), whole leaf (flush, young)Whole leaf (flush, young)Whole fruit (unripe, ripe), petiole (mature), whole leaf (flush, young)Whole fruit (ripe), whole leaf (young)Whole fruit (unripe, ripe), flowerWhole fruit (unripe, ripe)
Leeaceae Leea indica Whole leaf (flush, young)Lythraceae Lagerstroemia lanceolata Mesocarp (partially ripe), flower, whole leaf (flush)Meliaceae Dysoxylum malabaricum
Aglaia roxburghiiFlower, whole leaf (flush)Mesocarp (ripe), whole leaf (flush, young)
Moraceae Ficus callosaArtocarpus hirsutus
Ficus racemosaFicus nervosa
Petiole (mature)Mesocarp (unripe), leaf tip (flush), petiole (young, mature), whole leaf (flush)Mesocarp (ripe), whole fruit (unripe)Whole leaf (flush)
Myristicaceae Knema attenuateMyristica dactyloides
Aril (unripe, ripe), petiole (young, mature), whole leaf (flush, young)Mesocarp (ripe)
Myrtaceae Syzygium gardneri
Euginea macrocepalaSyzygium cuminii
Seed (ripe), bark, mesocarp (ripe), whole fruit (unripe, ripe), petiole (mature), whole leaf (flush, young, mature)Whole leaf (flush)Petiole (young)
Olacaceae Strombosia ceylanica Petiole (young)Oleaceae Olea dioica
Chionanthus malabarica
Bark, whole fruit (unripe, partially ripe, ripe), flower, petiole (young, mature), whole leaf (flush, young)Seed (unripe), flower, petiole (mature), whole leaf (flush, young)
Papilionaceae Dalbergia volubilis Whole leaf (flush, young, mature)Piperaceae Piper nigrum Petiole (young, mature), whole leaf (flush, young, mature)Poaceae Ochlandra redii PithRhizophoraceae Carallia brachiata Petiole (young, mature), whole leaf (flush)Rubiaceae Psychotria nigra
Canthium angustifoliumAnthocephalus cadambaRandia rugulosa
Petiole (young), whole leaf (flush, young, mature)Tendrils, whole leaf (flush, young, mature) Whole fruit (ripe)Whole leaf (flush, young, mature)
Rutaceae Vepris bilocularisClausena indica
Whole leaf (mature)Whole leaf (young)
Sapindaceae Dimocarpus longan Pith, aril (unripe, partially ripe, ripe), flower, petiole (young, mature), whole leaf (flush, young)
Sapotaceae Xantolis tomentosaMadhuca neriifolia
Petiole (young), whole leaf (flush, young, mature)Whole leaf (flush, young)
Sterculiaceae Sterculia guttata Seed (unripe), flower, petiole (young), whole leaf (young)Symplocaceae Symplocos racemosa Mesocarp (unripe), petiole (mature), whole leaf (flush)Verbenaceae Callicarpa tomentosa
Vitex altissima
Whole fruit (unripe, ripe), flower, petiole (young, mature), whole leaf (flush, young)Flower, petiole (young), whole leaf (young, mature)
Vitaceae Vitis canarensisUnidentified
Climber
Herb
Mesocarp (ripe), whole leaf (flush)Pith, whole fruit (unripe, ripe), flower, petiole (flush, young, mature), whole leaf (flush, young, mature)Seed (unripe, paripe), tendrils, mesocarp (ripe), whole fruit (unripe, ripe), flower, petiole (flush, young, mature), whole leaf (flush, young, mature)Whole leaf (young)
Table 1 (continued)

Diet of Diurnal Rain Forest Primates in the Central Western Ghats
291Folia Primatol 2011;82:283–298
dry season, the consumption of leaves by Hanuman langurs was also much higher (67.7%) than that of bonnet macaques (5%; � 2 = 809.22; d.f. = 1, p ! 0.01; fig. 2 ). Lion-tailed macaques consumed more fruits (78.1%) than bonnet macaques (48.9%) and Hanuman langurs (32.2%; � 2 = 478.48, d.f. = 2, p ! 0.01; fig. 2 ). In the dry season, the consumption of fruits by lion-tailed macaques was also much higher (77.4%) than by bonnet macaques (37.6%) and Hanuman langurs (25.6%; � 2 = 432.17, d.f. = 2, p ! 0.01; fig. 2 ). During the wet season, however, the consumption of fruits by all three primate species was quite high, with lion-tailed macaques (79.8%) and bonnet macaques (79.5%) consuming more fruits than Hanuman langurs (46.3%; � 2 = 137.30, d.f. = 2, p ! 0.01; fig. 2 ). Flowers were consumed more by bonnet macaques (26.7%) than lion-tailed macaques (6.3%) and Hanuman langurs (4.1%; � 2 = 420.89, d.f. = 2, p ! 0.01; fig. 2 ). A similar pattern of consumption was observed in the dry season, as bonnet macaques consumed a higher proportion of flowers (33.2%) than lion-tailed ma-caques (6.5%) and Hanuman langurs (5.1%; � 2 = 386.48, d.f. = 2, p ! 0.01; fig. 2 ). In the wet season, also, bonnet macaques consumed more flowers (9.4%) than lion-tailed macaques (5.6%) and Hanuman langurs (1.8%; � 2 = 30.22, d.f. = 2, p ! 0.01; fig. 2 ). Lion-tailed macaques fed more on faunal components (13.4%) than bonnet macaques (6.8%; � 2 = 20.75, d.f. = 1, p ! 0.01; fig. 2 ). Similarly, in the dry season, lion-tailed macaques consumed more faunal items (13.2%) than bonnet macaques (5.8%; fig. 2 ; � 2 = 20.90, d.f. = 1, p ! 0.01), but, in the wet season, the difference in the con-sumption of faunal items was not significant (lion-tailed macaques 13.6%; bonnet macaques 9.7%; � 2 = 1.82, d.f. = 1, n.s.; fig. 2 ). The other items, including pith, bark, tendrils and sap, were consumed more by bonnet macaques (6.2%) than lion-tailed macaques (2.3%) and Hanuman langurs (1.7%; � 2 = 52.94, d.f. = 2, p ! 0.01; fig. 2 ). The difference in the consumption of other items was significant in the dry season, as bonnet macaques consumed a higher proportion of other food items (8.0%) than lion-tailed macaques (2.9%) and Hanuman langurs (1.7%; � 2 = 62.13, d.f. = 2, p ! 0.01; fig. 2 ). This difference was not significant among Hanuman langurs (1.8%), bonnet macaques (1.4%) and lion-tailed macaques (0.9%) in the wet season ( � 2 = 0.92, d.f. = 2, n.s.; fig. 2 ).
Discussion
During this study, Hanuman langurs used the largest number of resources from the largest number of plants, followed by bonnet macaques and lion-tailed macaques. This hierarchy in richness of diet of these species fits well with their overall distribu-tion and adaptations to the variety of habitats, with the Hanuman langur being the most habitat generalist and the lion-tailed macaque being the most habitat specialist. The results also supported our prediction that the diet of folivorous langurs would be richer, with a larger number of plant species used, than the diet of frugivorous macaques. Since the variety of edible fruits is always less than the variety of leaves, the macaques used their major resources more equitably than the langurs, support-ing our prediction of a wider food niche breadth in the macaques than in the langur. Our results also supported a general primate pattern of heavier use of only a few re-sources, as over 50% of the total diet of each primate species came from only a few plant species. Among 15 plant species used largely by the primates, most of the plant species are highly abundant and have a high importance value index in the study area

Folia Primatol 2011;82:283–298292 Roy /Singh /Singh
Overall Dry season W et season
resource % scans
resource % scans
resource % scans
Lion-tailed macaqueDifferent insects 22.0 Different insects 22.6 G. gummigutta, mesocarp D. crumenata, mesocarp paripe 12.1 D. crumenata, mesocarp ripe 16.4D. crumenata, mesocarp ripe 9.8 paripe 15.6 A. roxburghii, aril ripe 15.6D. venusta, mesocarp ripe 9.1 D. crumenata, mesocarp ripe 13.0 Different insects 15.6G. gummigutta, mesocarp ripe 7.2 D. venusta, mesocarp ripe 12.2 S. gardneri, whole fruit ripe 10.2C. urens, mesocarp paripe 6.9 D. venusta, whole fruit ripe 7.4 C. urens, mesocarp paripe 8.2S. gardneri, whole fruit ripe 6.0 C. urens, mesocarp paripe 5.7 K. attenuata, flower 3.5D. venusta, whole fruit ripe 5.5 P. macrantha, mesocarp ripe 5.0 A. roxburghii, mesocarpA. roxburghii, aril ripe 5.1 S. gardneri, whole fruit ripe 3.6 paripe 3.5P. macrantha, mesocarp ripe 3.7 H. grahamii, mesocarp ripe 3.6 D. indicus, seed ripe 2.3K. attenuata, flower 2.7 G. gummigutta, mesocarp ripe 2.4 Climber, whole fruit ripe 2.0H. grahamii, mesocarp ripe 2.7 K. attenuata, flower 2.1 F. callosa, whole fruit ripe 2.0D. indicus, seed ripe 1.9 Climber, seed ripe 1.5 Climber, whole fruit paripe 2.0Climber, whole fruit ripe 1.5 D. indicus, seed ripe 1.5 D. crumenata, mesocarpA. roxburghii, mesocarp paripe 1.5 D. venusta, mesocarp unripe 1.4 paripe 1.6P. macrantha, flower 1.2 Climber, whole fruit ripe 1.2 C. glauca, seed ripe 1.6Climber, seed ripe 1.2 D. indicus, seed paripe 1.2 Climber, mesocarp paripe 1.6
M. indica, mesocarp unripe 1.6L. stocksii, whole fruit unripe 1.6S. cumini, mesocarp ripe 1.6D. crumenata, mesocarp unripe 1.2P. macrantha, flower 1.2D. pruriens, mesocarp unripe 1.2D. pruriens, mesocarp paripe 1.2K. attenuata, aril ripe 1.2M. peltata, whole fruit ripe 1.2A. roxburghii, aril paripe 1.2C. auriculata, mesocarp ripe 1.2
Bonnet macaqueDifferent insects 14.9 H. ponga, flower 17.5 A. roxburghii, aril ripe 14.1H. ponga, flower 13.8 Different insects 13.7 Different insects 12.5Rice, grain ripe 8.3 Rice, grain ripe 10.6 Climber, whole fruit ripe 10.0M. neriifolia, flower 8.1 M. neriifolia, flower 10.1 K. attenuata, aril ripe 6.3C. urens, pith 5.2 C. urens, pith 6.6 S. gardneri, whole fruit ripe 6.3Climber, whole fruit ripe 4.8 F. tsjahela, whole fruit ripe 5.3 G. gummigutta, mesocarp ripe 5.9A. roxburghii, aril ripe 4.7 F. callosa, mesocarp ripe 5.0 A. roxburghii, mesocarp ripe 5.6F. callosa, mesocarp ripe 4.1 C. urens, mesocarp ripe 4.9 Climber, mesocarp ripe 5.3F. tsjahela, whole fruit ripe 4.1 H. grahamii, hypocarp unripe 4.6 D. pruriens, mesocarp ripe 5.3C. urens, mesocarp ripe 3.8 F. nervosa, mesocarp paripe 3.6 Climber, flower 4.1Climber, flower 3.7 Climber, flower 3.0 S. gardneri, whole fruit unripe 2.8H. grahamii, hypocarp unripe 3.6 P. macrantha, mesocarp paripe 2.8 T. nudiflora, mesocarp paripe 2.8F. nervosa, mesocarp paripe 2.8 H. grahamii, mesocarp paripe 2.8 P. fragrans, mesocarp ripe 2.8P. macrantha, mesocarp paripe 2.2 Climber, whole fruit ripe 1.9 F. racemosa, mesocarp paripe 2.5
Table 2. S elective resources used by lion-tailed macaques, bonnet macaques and Hanuman langurs (ob-served in 1% or more of the feeding scans)

Diet of Diurnal Rain Forest Primates in the Central Western Ghats
293Folia Primatol 2011;82:283–298
Overall Dry season W et season
resource % scans
resource % scans
resource % scans
H. grahamii, mesocarp paripe 2.2 F. callosa, mesocarp unripe 1.6 L. floribunda, flower 2.5K. attenuata, aril ripe 2.1 D.crumenata, mesocarp paripe 1.4 F. callosa, mesocarp unripe 2.2S. gardneri, whole fruit ripe 2.1 C. malabarica, whole leaf flush 1.4 C. auriculata, mesocarp ripe 2.2F. callosa, mesocarp unripe 2.0 C. spiciflorum, flower 1.2 T. nudiflora, seed paripe 1.9G. gummigutta, mesocarp ripe 2.0 M. elengi, mesocarp paripe 1.1 P. macrantha, flower 1.3Climber, mesocarp ripe 2.0 C. auriculata, whole fruit ripe 1.1 Climber, whole fruit unripe 1.3A. roxburghii, mesocarp ripe 1.9 K. attenuata, aril paripe 1.3D. pruriens, mesocarp ripe 1.8 D. pruriens, mesocarp paripe 1.3
Hanuman langurO. dioica, petiole mature 20.3 O. dioica, petiole mature 20.4 O. dioica, whole fruit ripe 14.9D. longan, whole leaf flush 15.4 D. longan, whole leaf flush 18.8 D. longan, aril ripe 13.2O. dioica, whole fruit ripe 15.2 O. dioica, whole fruit ripe 10.1 O. dioica, petiole mature 7.8D. longan, aril ripe 6.1 Climber, whole leaf young 6.0 V. canarensis, mesocarp ripe 5.0Climber, whole leaf young 6.0 S. gardneri, whole fruit ripe 4.5 A. monadelphum, whole leafS. gardneri, whole fruit ripe 4.2 Climber, whole leaf flush 4.0 flush 4.5Climber, whole leaf flush 4.1 P. macrantha, mesocarp ripe 3.5 P. nigrum, whole leaf mature 4.5P. nigrum, whole leaf mature 3.9 K. attenuata, petiole mature 3.2 L. stocksii, whole fruit ripe 4.3L. laevigata, whole leaf flush 3.7 L. laevigata, whole leaf flush 3.1 D. longan, aril unripe 3.7A. monadelphum, whole leaf C. malabarica, whole leaf flush 2.6 C. glauca, mesocarp ripe 3.2flush 3.2 P. macrantha, mesocarp unripe 2.4 P. scandens, whole leaf mature 2.8L. stocksii, whole fruit ripe 3.2 D. pentagyna, whole fruit D. longan, aril paripe 2.8P. macrantha, mesocarp ripe 2.8 unripe 2.4 V. altissima, petiole young 2.8K. attenuata, petiole mature 2.8 K. attenuata, whole leaf young 2.3 L. laevigata, whole leaf flush 2.6C. malabarica, whole leaf flush 2.3 O. dioica, flower 2.3 A. monadelphum, whole leafV. canarensis, mesocarp ripe 2.3 Unidentified, whole leaf young 2.2 young 2.6D. longan, aril unripe 2.2 Unidentified, petiole mature 2.2 Climber, whole leaf young 2.4Climber, whole leaf mature 2.2 Climber, whole leaf mature 2.2 C. tomentosa, petiole mature 2.1
P. nigrum, whole leaf mature 2.2 C. tomentosa, whole leaf flush 1.7O. dioica, whole leaf young 2.0 C. tomentosa, petiole young 1.7C. tomentosa, whole fruit Climber, whole leaf flush 1.7unripe 1.9 P. nigrum, whole leaf young 1.7O. dioica, whole fruit paripe 1.8 M. indica, mesocarp paripe 1.7
C. tomentosa, whole fruit ripe 1.7H. pentandra, mesocarpparipe 1.7O. redii, pith 1.7T. heyneana, whole leaf young 1.5D. pentagyna, whole fruit ripe 1.5S. gardneri, whole fruit unripe 1.5P. nigrum, petiole mature 1.5L. floribunda, whole fruit ripe 1.5
Paripe = Partially ripe.
Table 2 (continued)

Folia Primatol 2011;82:283–298294 Roy /Singh /Singh
[Roy et al., 2010]. In other words, all the primate species are mostly dependent on those plant species which are abundantly available in that region. The monthly pro-ductivity sampling denoted the presence of different resource parts of those major plant species [Singh et al., 2011]. Although some of the resources are very seasonal (like fruits of Garcinia gummigutta in the wet season or Holigarna grahamii in the dry season), the consumption frequency of those resources was very high.
In some previous studies it has been reported that, although folivorous primates have a higher chance of diet selection because of resource abundance, they are selec-tive by nature and use very few resources for overall consumption [Harris et al., 2009]. Although lion-tailed macaques were not observed to consume leaves, bonnet macaques occasionally consumed leaves, mainly young and whole-leaf flush or their petioles. In previous studies in some other areas, it has also been reported that bon-net macaques fed on leaves and grass [Krishnamani, 1994; Singh, 1999] and a selec-tion of varied food items in different environments [Sinha, 2005]. As defined foli-vores, Hanuman langurs consumed several parts of the foliage, although in the dry season they consumed more leaf flush, probably for its higher water content. Being larger than macaques, Hanuman langurs probably have more digestive efficiency for leafy items [Remis and Dierenfeld, 2004], which allows them to tolerate a higher level of fibre in their food [Remis, 2000, 2003]. Leaves and petioles are a source of protein for them [Milton, 1983; Wasserman and Chapman, 2003].
All three primate species preferred primarily ripe fruits, as is known for other primate species [Dew, 2005], but lion-tailed macaques consumed the most fruit com-pared to the other primate species in this study. Lion-tailed macaques have previ-ously been reported to spend more than half of their total foraging time on fruit
Table 3. C omparative utilization of major plant species by the three primate species
Season Lion-tailed macaque Bonnet macaque Hanuman langur
Overall D. crumenata (21.9) H. ponga (13.8) O. dioica (35.5)D. venusta (14.6) Climber (10.5) D. longan (23.7)G. gummigutta (7.2) C. urens (9.0)C. urens (6.9) Rice (8.3)
M. neriifolia (8.1)A. roxburghii (6.6)
Dry season D. crumenata (28.6) H. ponga (17.5) O. dioica (36.6)D. venusta (21.0) C. urens (11.5) D. longan (18.8)C. urens (5.7) Rice (10.6)
M. neriifolia (10.1)H. grahamii (7.4)
Wet season A. roxburghii (20.3) Climber (20.7) O. dioica (22.7)G. gummigutta (16.4) A. roxburghii (19.7) D. longan (19.7)S. gardneri (10.2) S. gardneri (9.1) P. nigrum (7.7)C. urens (8.2) K. attenuata (7.6)
T otal consumption in percent is given in parentheses.

Diet of Diurnal Rain Forest Primates in the Central Western Ghats
295Folia Primatol 2011;82:283–298
consumption [Kumar, 1987; Singh et al., 1998, 2000]. Bonnet macaques in this study region consumed slightly less fruit compared to in other areas of their occurrence, such as deciduous forests of the Western Ghats [Singh, 1999] and during their occa-sional forays into the rain forests of the Anaimalai Hills [Sushma, 2004]. Among fruits, both macaque species showed a preference for figs, despite the marked sea-sonal variation in their availability, mostly due to their high calcium content [O’Brien et al., 1998; Felton et al., 2008]. Unlike in some other parts of the Western Ghats [Ku-mar, 1987; Sushma, 2004], however, lion-tailed macaques used fruits of only a few fig tree species as their major diet. The three study species consumed fruits mostly in the wet season, although the consumption of some fruit items by lion-tailed ma-caques was highly seasonal, irrespective of their availability.
It has been reported earlier, however, that, unlike Hanuman langurs, some oth-er colobine monkeys, such as capped langurs, consumed flowers as much as fruits [Solanki et al., 2008]. During the post-monsoon season, flowers were a major re-source for lion-tailed macaques and bonnet macaques in other parts of the Western Ghats also [Ganesh and Davidar, 1997; Sushma, 2004]. Although the consumption of similar diets by congeneric species is reported in many other studies [Wegge et al., 2006; Lopez and Vaughan, 2007], in this study area, lion-tailed macaques consumed flowers less frequently than bonnet macaques and the two differed greatly in the choice of tree species.
The faunal items consumed by the macaques included a variety of invertebrates which could not be identified to the species level due to their distance from the ob-server. In this study, Hanuman langurs did not consume faunal resources, but in deciduous forests, they fed on ticks, lice, grasshoppers and other small invertebrates [Mathur et al., 1990; Srivastava, 1991; Koenig and Borries, 2001]. In bonnet ma-caques, in a tropical dry evergreen forest, faunal items constituted about 14.5% of their diet [Krishnamani, 1994], but Singh [1999] reported that there were very few (1.3%) invertebrates in their overall diet in some other forest areas. In this study, bon-net macaques had nearly 7% of faunal food consumption in their annual diet, and that can be considered as a moderate consumption of fauna by bonnet macaques. Maximum consumption of faunal items was observed in lion-tailed macaques, al-though, in this study area, faunal consumption was less than in other parts of the Western Ghats [Sushma, 2004]. Generally, frugivorous primates, especially ma-caques, consume faunal items as their protein supplement. In this study area, there was a high consumption of fruits by the macaques and that probably resulted in less time spent on faunal food intake. Although the plant components, mainly fruits, are rich in carbohydrates, they have very little protein and fat [Herrera, 1982; Cipollini and Levey, 1997; Chapman et al., 2002]. Sushma [2004] suggested that lion-tailed macaques may feed on plant parts of some particular species for the possible presence of insect larvae in them, because these macaques bite the fruits of Hydnocarpus sp. and Ficus beddomei , but do not ingest them completely. According to Redford et al. [1984] also, primates might deliberately select fruits which are infested with insects to feed primarily on insects. So, in this study, it may be that lion-tailed macaques, as well as bonnet macaques, could have increased their intake of faunal items by forag-ing on a variety of fruits and the consumption rates of faunal food increased slightly at the time of scarcity of other plant food items.
Although Hanuman langurs have often been observed to raid cultivated crops in north-western India [Chhangani and Mohnot, 2006], in this study, these langurs

Folia Primatol 2011;82:283–298296 Roy /Singh /Singh
did not feed on crops, despite their availability within the home range. Only bonnet macaques fed on the grains of rice paddies and, in the dry season, it constituted a major component of the total diet. Those crops were sown in the rainy season, and they became ripe in late October and early November. The macaques raided the crops several times a day and, whenever crops were available, they hardly consumed any other food item. This feeding continued until the crops were harvested in Janu-ary or early February. Lion-tailed macaques have never been seen feeding on crops in any of the study areas in the Western Ghats, including in this study.
Whereas the number of food resources consumed by a species indicates richness of its diet, the proportional use of these resources provides a measure of dietary di-versity and food niche breadth. In this study, the Hanuman langur foraged on a much larger number of resources than the macaques, but the food niche breadth was broader in the macaques than in the langur. The abundance and variety of edible fruits are much smaller than leaves. The langur, a primarily folivorous species, fed on leaves of a large number of trees, many of which were visited only occasionally. This resulted in a disproportionate use of these resources. On the other hand, only a few tree species constituted the major fruit resources for the macaques, resulting in a more balanced and proportionate use of these resources. This may explain the broader food niches of the macaques than of the langur.
Acknowledgements
We thank Karnataka Forest Department for the research permit (No. PS CWL.RP.CR-22/2005-06 dated November 23, 2005) and for assistance in the field. Grants for this study were received from the Department of Science and Technology, Government of India (grant No. SR/SO/AS-35/2004), by Mewa Singh and Mridula Singh. We also thank Dr. H.S. Sushma for help during the first year of the study. We thank the editor and three referees for their helpful com-ments.
References
Altmann J (1974). Observational study of behavior: sampling methods. Behaviour 49: 227–267. Champion HG, Seth SK (2005). A Revised Survey of Forest Types of India (reprinted). Dehradun, Natraj. Chapman CA, Chapman LJ, Bjorndal KA, Onderdonk DA (2002). Application of protein to fiber ratios
to predict colobine abundance on different spatial scales. International Journal of Primatology 23: 283–310.
Chapman CA, Chapman LJ, Naughton-Treves L, Lawes MJ, McDowell LR (2004). Predicting folivorous primate abundance: validation of a nutritional model. American Journal of Primatology 62: 55–69.
Chhangani AK, Mohnot SM (2006). Ranging behaviour of Hanuman langurs (Semnopithecus entellus) in three different habitats. Primate Conservation 21: 171–177.
Cipolini ML, Levey DJ (1997). Secondary metabolites of f leshy vertebrate-dispersed fruits: adaptive hy-pothesis and implications for seed dispersal. American Naturalist 150: 346–372.
Dew JL (2005). Foraging, food choice and food processing by sympatric ripe fruit specialists: Lagothrix lagothricha poeppigii and Ateles belzebuth belzebuth . International Journal of Primatology 26: 1107–1135.
Felton AM, Felton A, Wood JT, Lindenmayer DB (2008). Diet and feeding ecology of Ateles chamek in a Bolivian semihumid forest: the importance of Ficus as a staple food resource. International Jour-nal of Primatology 29: 379–403.
Gamble JS, Fischer CEC (1916–1935). Flora of the Presidency of Madras. Dehradun, Shiva Offset Press, 3 volumes.

Diet of Diurnal Rain Forest Primates in the Central Western Ghats
297Folia Primatol 2011;82:283–298
Ganesh T, Davidar P (1997). Flowering phenology and flower predation of Cullenia exarillata (Bomba-caceae) by arboreal vertebrates in Western Ghats, India. Journal of Tropical Ecology 13: 459–468.
Harris TR, Chapman CA, Monfort SL (2009). Small folivorous primate groups exhibit behavioral and physiological effects of food scarcity. Behavioural Ecology 21: 46–56.
Herrera CM (1982). Defense of ripe fruits from pests: its significance in relation to plant-disperser in-teractions. American Naturalist 120: 218–241.
Hladik CM (1977). A comparative study of the feeding strategies of two sympatric species of leaf mon-keys: Presbytis senex and Presbytis entellus . In Primate Ecology: Studies of Feeding and Ranging Behaviour in Lemurs, Monkeys and Apes (Clutton-Brock TH, ed.), pp 323–353. London, Academ-ic Press.
Hurlbert SH (1978). The measurement of niche overlap and some relatives. Ecology 59: 67–77. Koenig A (1996). The fairytale of gentle leaf eaters: food competition and social relationship in female
common langurs (Presbytis entellus) . Primate Report 44: 27. Koenig A, Borries C (2001). Socioecology of Hanuman langurs: the story of their success. Evolutionary
Anthropology 10: 122–137. Koenig A, Borries C, Chalise MK, Winkler P (1997). Ecology, nutrition and timing of reproductive
events in an Asian primate, the Hanuman langur (Presbytis entellus) . Journal of Zoology 243: 215–235.
Krishnamani R (1994). Diet composition of the bonnet macaque (Macaca radiata) in a tropical dry ev-ergreen forest of southern India. Tropical Biodiversity 2: 285–302.
Kumar A (1987). The Ecology and Population Dynamics of the Lion-Tailed Macaque (Macaca silenus) of South India. PhD thesis, Cambridge University.
Kumara HN, Singh M (2004). Distribution and abundance of primates in rainforests of the Western Ghats, Karnataka, India and the conservation of Macaca silenus . International Journal of Prima-tology 26: 1001–1018.
Kumara HN, Singh M, Sharma AK, Singh M, Kumar MA (2000). Faunal component in the diet of lion-tailed macaques. Primate Report 58: 57–66.
Lehner PN (1996). Handbook of Ethological Methods. Cambridge, Cambridge University Press. Lopez JE, Vaughan C (2007). Food niche overlap among neotropical frugivorous bats in Costa Rica.
International Journal of Tropical Biology 55: 301–313. Mathur R, Bhatnagar PS, Manahar BR (1990). Ticks, lice and grasshopper eating by Presbytis entellus .
Human Evolution 5: 531–536. Milton K (1983). Dietary quality and population regulation in a howler monkey population. In The Ecol-
ogy of a Rainforest (Leigh EG, Rand AS, Windsor DM, eds.), pp 273–290. Oxford, Oxford Univer-sity Press.
Molur S, Brandon-Jones D, Dittus W, Eudey A, Kumar A, Singh M, Feeroz MM, Chalise M, Priya P, Walker S (2003). Status of South Asian Primates: Conservation, Assessment and Management Plan (CAMP). Coimbatore, Zoo Outreach Organization/CBSG South Asia.
O’Brien TG, Kinnaird MF, Dierenfeid ES (1998). What’s so special about figs? Nature 392: 668. Pascal JP (1988). Wet Evergreen Forests of the Western Ghats of India: Ecology, Structure, Floristic Com-
position and Succession. Pondicherry, Institut Français de Pondicherry. Pascal JP, Ramesh BR (1997). A Field Key to the Trees and Lianas of the Evergreen Forests of the Western
Ghats. Pondicherry, Institut Français de Pondicherry. Ramachandran KK, Joseph GK (2000). Habitat utilization of lion-tailed macaque (Macaca silenus) in
Silent Valley National Park, Kerala, India. Primate Report 58: 17–25. Redford KH, Da Fonseca GAB, Lacher TE (1984). The relationship between frugivory and insectivory
in primates. Primates 24: 433–440. Remis M, Dierenfeld ES (2004). Digesta passage, digestibility and behavior in captive gorillas under two
dietary regimens. International Journal of Primatology 25: 825–846. Remis MJ (2000). Initial studies on the contributions of body size and gastrointestinal passage times to
dietary flexibility among gorillas. American Journal of Physical Anthropology 112: 171–180. Remis MJ (2003). Are gorillas vacuum cleaners of the forest f loor? The roles of body size, habitat and
food preferences on dietary flexibility and nutrition. In Gorilla Biology: A Multidisciplinary Per-spective (Taylor AB, Goldsmith ML, eds.), pp 385–404. Cambridge, Cambridge University Press.
Roy K, Singh M, Sushma HS, Singh M (2010). Stand structure of a primate rich rainforest region in the central Western Ghats of southern India. Journal of Threatened Taxa 2: 930–939.
Sheskin DJ (2000). Handbook of Parametric and Nonparametric Statistical Procedures, 2nd ed. Boca Raton, Chapman and Hall.
Singh M (1999). Studies on ecology, adaptation and evolution of sociality in non-human primates of South India. Final technical report for research project submitted to the Ministry of Environment and Forests, Government of India.
Singh M, Roy K, Singh M (2011). Resource partitioning in sympatric langurs and macaques in tropical rainforests of the central Western Ghats, South India. American Journal of Primatology 73: 335–346.

Folia Primatol 2011;82:283–298298 Roy /Singh /Singh
Singh M, Singh M, Kumar MA, Kumara HN, De Souza L, Sharma AK (1998). Behaviour of lion-tailed macaque (Macaca silenus) in vulnerable and relatively secure habitats in the rain forests of West-ern Ghats, India. Tigerpaper 25: 19–25.
Singh M, Singh M, Kumar MA, Kumara HN, Sharma AK, Sushma HS (2000). Niche separation in sym-patric lion-tailed macaque (Macaca silenus) and Nilgiri langur (Presbytis johnii) in an Indian tropical rain forest. Primate Report 58: 83–95.
Sinha A (2005). Not in their genes: phenotypic flexibility, behavioural traditions and cultural evolution in wild bonnet macaques. Journal of Biosciences 30: 51–64.
Solanki GS, Kumar A, Sharma BK (2008). Feeding ecology of Trachypithecus pileatus in India. Interna-tional Journal of Primatology 29: 173–182.
Srivastava A (1991). Insectivory and its significance to langur diets. Primates 32: 237–241. Srivastava A (1992). Use of resources by free-ranging female langurs (Presbytis entellus) during different
reproductive phases. Folia Primatologica 59: 157–162. Sushma HS (2004). Resource Utilization and Niche Separation in Sympatric Rainforest Arboreal Mam-
mals. PhD thesis, University of Mysore. Sushma HS, Singh M (2006). Resource partitioning and interspecific interactions among sympatric
rainforest arboreal mammals of the Western Ghats, India. Behavioural Ecology 17: 479–490. Umapathy G (1997). Impact of Habitat Fragmentation on the Arboreal Mammals in the Wet Evergreen
Forests of the Anaimalai Hills, South India. PhD thesis, Bharathiar University. Wasserman MD, Chapman CA (2003). Determinants of colobine monkey abundance: the importance
of food energy, protein and fibre content. Journal of Animal Ecology 72: 650–659. Wegge P, Shrestha AK, Moe SR (2006). Dry season diets of sympatric ungulates in lowland Nepal: com-
petition and facilitation in alluvial tall grasslands. Ecology Research 21: 698–706.
muellers
Stempel Hoch




















