
Xklp15 a chromosomal xenopus kinesin-like protein essential for spindle organization and chromosome...
11
Cell, Voi. 81, 117-127, April 7, 1995, Copyright © 1995 by Cell Press Xklpl, a Chromosomal Xenopus Kinesin-like Protein Essential for Spindle Organization and Chromosome Positioning Isabelle Vernos, 1,= Joe Raats, 2,3, 4 Tatsuya Hirano, s Janet Heasman, 3, e Eric Karsenti, 1,T and Chris Wylie 3,8 1Cell Biology Programme ~Cell Biophysics Programme European Molecular Biology Laboratory Meyerhofstrasse 1 69117 Heidelberg Federal Republic of Germany 3Institute for Human Genetics 6Departments of Cell Biology and Neuroanatomy 8Department of Pediatrics University of Minnesota School of Medicine Minneapolis, Minnesota 55455 ~Department of Pharmacology University of California, San Francisco San Francisco, California 94143-0450 4Wellcome/Cancer Research Campaign Institute Tennis Court Road Cambridge CB2 1QR England Summary Xklpl is a novel Xenopus kinesin-like protein with a motor domain at the amino terminus, nuclear localiza- tion sequences in the stalk, and a putative zinc finger- like sequence in the tail. It is nuclear during interphase and chromosomal during mitosis. During late ana- phase, a fraction of the protein relocalizes to the spin- dle interzone and accumulates in the midbody during telophase. Depletion of Xklpl protein by antisense oligo knockout in oocytes leads to defective mitosis during the first cell cycles following fertilization. The bipolarity of spindles assembled in vitro in the pres- ence of anti-Xklpl antibodies is unstable, and the chro- mosomes fail to congress on the metaphase plate. Introduction Mitosis proceeds in three major steps: assembly of the mitotic spindle, separation of the sister chromatids, and cytokinesis. A complete understanding of the principles underlying the morphogenesis and function of the mitotic spindle requires the identification of the molecules regulat- ing microtubule dynamics and of the microtubule-depen- dent motors involved in each step of mitosis. Apart from the localization of dynein at kinetochores, the microtubule- dependent motor proteins that have been identified and localized so far in the spindle belong to the kinesin family (kinesin-like proteins [KLPs]; Pereira and Goldstein, 1994; Bloom and Endow, 1995). Although the number of mitotic KLPs identified is already 21sabelle Vernos and Jos Raats have contributedequally to this work and should both be regarded as first authors. quite high, some mitotic events that are likely to require the activity of motors still lack their partners. For example, it has been postulated that the polar ejection forces, which form part of the mechanism leading to the congression of chromosomes on the metaphase plate, are produced by plus end-directed motors associated with the chromo- some arms (Theurkauf and Hawley, 1992; Zhang et al., 1990). Also, microtubule motors are predicted to be in- volved in spindle pole function and antiparallet microtu- bule sliding in anaphase and telophase. Finally, the kineto- chore motors that mediate chromosome distribution have not been unambiguously identified (Gorbsky, 1992). We previously reported partial sequences of four kinesin-like transcripts in Xenopus laevis oocytes (Vernos et al., 1993). H ere, we present the characterization of Xen- opus kinesin-like protein 1 (Xklpl). The protein organiza- tion is conventional, but features in its sequence and its localization pattern in cells indicate that it belongs to a novel family of KLPs. Xklpl is nuclear in interphase and remains on the chromosomes during mitosis. At late ana- phase, part of the protein migrates toward the spindle mid- zone and becomes associated with the midbody. Deple- tion of the Xklpl mRNA from oocytes by use of the antisense technique leads to abnormal mitosis. Meta- phase spindles assembled in frog egg extracts in the pres- ence of anti-Xklpl antibodies are unstable, with chromo- somes scattered throughout the spindle. Results Xklpl Sequence and Structural Organization Using a polymerase chain reaction (PCR)-based ap- proach, we previously identified four novel members of the kinesin family of motor proteins in Xenopus (Vernos et al., 1993). One of them, Xklpl, has been fully se- quenced. The predicted protein has 1226 amino acids, an estimated relative molecular mass of 1.39 x 105 Da and a pl of 8 (Figure la). The overall organization of the predicted protein is similar to that of kinesin and many other mem- bers of the family (Figure lb). A putative motor domain located at the amino-terminal end of the protein is linked to the 210 amino acid tail domain by a 540 amino acid region predicted to be m-helical. The Pepcoil program (Lu- pas et al., 1991) shows that this region may be involved in coiled-coil interactions (Figure lb). Xklpl Contains Nuclear Localization Signals, a Zinc Finger-Like Domain, and a Consensus Cdc2 Phosphorylation Site The analysis of the Xkipl protein sequence showed that it contains fou r sequences that conform to the consensus for nuclear localization (nuclear localization signals [NLSs]) (Dingwall and Laskey, 1991). These sequences (Figures la and 1c) are organized in tandem and are local- ized in the region predicted to be a-helical (Figure lb). We used the region comprising amino acids 1050-1226, corresponding to the predicted tail region, to search the
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Xklp15 a chromosomal xenopus kinesin-like protein essential for spindle organization and chromosome...

Cell, Voi. 81, 117-127, April 7, 1995, Copyright © 1995 by Cell Press
Xklpl, a Chromosomal Xenopus Kinesin-like Protein Essential for Spindle Organization and Chromosome Positioning Isabelle Vernos, 1, = Joe Raats, 2, 3, 4 Tatsuya Hirano, s Janet Heasman, 3, e Eric Karsenti, 1, T and Chris Wylie 3, 8 1Cell Biology Programme ~Cell Biophysics Programme European Molecular Biology Laboratory Meyerhofstrasse 1 69117 Heidelberg Federal Republic of Germany 3Institute for Human Genetics 6Departments of Cell Biology and Neuroanatomy 8Department of Pediatrics University of Minnesota School of Medicine Minneapolis, Minnesota 55455 ~Department of Pharmacology University of California, San Francisco San Francisco, California 94143-0450 4Wellcome/Cancer Research Campaign Institute Tennis Court Road Cambridge CB2 1QR England
Summary
Xklpl is a novel Xenopus kinesin-like protein with a motor domain at the amino terminus, nuclear localiza- tion sequences in the stalk, and a putative zinc finger- like sequence in the tail. It is nuclear during interphase and chromosomal during mitosis. During late ana- phase, a fraction of the protein relocalizes to the spin- dle interzone and accumulates in the midbody during telophase. Depletion of Xklpl protein by antisense oligo knockout in oocytes leads to defective mitosis during the first cell cycles following fertilization. The bipolarity of spindles assembled in vitro in the pres- ence of anti-Xklpl antibodies is unstable, and the chro- mosomes fail to congress on the metaphase plate.
Introduction
Mitosis proceeds in three major steps: assembly of the mitotic spindle, separation of the sister chromatids, and cytokinesis. A complete understanding of the principles underlying the morphogenesis and function of the mitotic spindle requires the identification of the molecules regulat- ing microtubule dynamics and of the microtubule-depen- dent motors involved in each step of mitosis. Apart from the localization of dynein at kinetochores, the microtubule- dependent motor proteins that have been identified and localized so far in the spindle belong to the kinesin family (kinesin-like proteins [KLPs]; Pereira and Goldstein, 1994; Bloom and Endow, 1995).
Although the number of mitotic KLPs identified is already
21sabelle Vernos and Jos Raats have contributed equally to this work and should both be regarded as first authors.
quite high, some mitotic events that are likely to require the activity of motors still lack their partners. For example, it has been postulated that the polar ejection forces, which form part of the mechanism leading to the congression of chromosomes on the metaphase plate, are produced by plus end-directed motors associated with the chromo- some arms (Theurkauf and Hawley, 1992; Zhang et al., 1990). Also, microtubule motors are predicted to be in- volved in spindle pole function and antiparallet microtu- bule sliding in anaphase and telophase. Finally, the kineto- chore motors that mediate chromosome distribution have not been unambiguously identified (Gorbsky, 1992).
We previously reported partial sequences of four kinesin-like transcripts in Xenopus laevis oocytes (Vernos et al., 1993). H ere, we present the characterization of Xen- opus kinesin-like protein 1 (Xklpl). The protein organiza- tion is conventional, but features in its sequence and its localization pattern in cells indicate that it belongs to a novel family of KLPs. Xklpl is nuclear in interphase and remains on the chromosomes during mitosis. At late ana- phase, part of the protein migrates toward the spindle mid- zone and becomes associated with the midbody. Deple- tion of the Xklpl mRNA from oocytes by use of the antisense technique leads to abnormal mitosis. Meta- phase spindles assembled in frog egg extracts in the pres- ence of anti-Xklpl antibodies are unstable, with chromo- somes scattered throughout the spindle.
Results
Xklpl Sequence and Structural Organization Using a polymerase chain reaction (PCR)-based ap- proach, we previously identified four novel members of the kinesin family of motor proteins in Xenopus (Vernos et al., 1993). One of them, Xklpl, has been fully se- quenced. The predicted protein has 1226 amino acids, an estimated relative molecular mass of 1.39 x 105 Da and a pl of 8 (Figure la). The overall organization of the predicted protein is similar to that of kinesin and many other mem- bers of the family (Figure lb). A putative motor domain located at the amino-terminal end of the protein is linked to the 210 amino acid tail domain by a 540 amino acid region predicted to be m-helical. The Pepcoil program (Lu- pas et al., 1991) shows that this region may be involved in coiled-coil interactions (Figure lb).
Xklpl Contains Nuclear Localization Signals, a Zinc Finger-Like Domain, and a Consensus Cdc2 Phosphorylation Site The analysis of the Xkipl protein sequence showed that it contains fou r sequences that conform to the consensus for nuclear localization (nuclear localization signals [NLSs]) (Dingwall and Laskey, 1991). These sequences (Figures l a and 1 c) are organized in tandem and are local- ized in the region predicted to be a-helical (Figure lb).
We used the region comprising amino acids 1050-1226, corresponding to the predicted tail region, to search the

Cell 118
M~EGIPYRYALRCRPLVPK~C~T~YgGEQ~V 40 I"~G~XSTT~DYVFDP SAEQEEYYNSAVAPL Tr GLFKGY~ 80 ATVLAYGQTGSGKTYSMGGA~RTT~A 120 LFRE IHQ RPE%~EFI~ETSYLE ~ ILDLLYAARDKTNT 160 I SIREDPKEGI~ICGLTERDVKTALDTLSCLEOGNSSRTV 280 ASTAt~t ~ S SR S~All~1~ S ~GDKI~SI~ 5TJ~ ~D 240 LAGSERQI3Y/X~G~RLK~GI S II~GLLCLGNV~ SALGDE 280 SKX~SKLTRLLODSLGGNSRTLMIACVSPAD SN 320 ~E~L/~.RY~DRARK]3~KP~R]TDPQAAELQRLKLOVO 360 E I ~ Q A E [ ~ S ~ L Q Sr.,,l~X'm :~k'r~" "zOO
~kA~CE~.0.RL~ETJ2~D0.]~K~R YL v .L~I~.,QQ ~I'~.L Q DE 480 S SGIJ~GS IEAMD~X ~ ~'~ Y~;~KR S ~ ROAOL ~J~L]~L~ XT-Xt~fIQNDR~ZQ ~ S61D
O.EIQGI~MQR~LI~OI~3~DAEK~RTWEQOJ~.'Zl~ ~t J.QLKE 680
L(~QKr.~ SQ SKG~AASRY/~T~L~LVS T 760 ~.AQR~uI~LLEDI%K ILA~D .L~,QLKQZ'IDAG]~ Il~3~ ~.R 8 0 0 RT~q~AELE~ .~x ~TKQ ~ ~.~EI~LR SAQ~ADLQQ 840 ~,,DAD GEEEII'VI~R~"I'I SN"r'.~ xrc.AIA"TL ITELVS SK 81B0 VAGSKLE S S~KQI~U~YADLOT~ J J S 920 Q~T.~t ILYLLS~0ASVP ~fm~r.L/AE~IT 9 6 0
r .M .T ,~VASGKKLI~r/~P Ju~EICSPDS~P~D~I~XPRG~R 4 0 4 0 ~._S.~.AV'n,_E~LL S'Z S'Z S ' ~ s ~ . _ _ ~ z : ~ _ _ _s-x~. _ s - ~ IOSO
KVLRE ~$0 IG~TLS IKL~ST~SASASTMESQENQTS I 1200 ~u~ Y~CNSNTS~FSGCSAITEDE
b 1.o 1 .o
° 1 !i ~ 0 . 4 O.4
O.Z 0.2
o,o , 2~00 , L 1 .o
uO-mlZ
C Xklpl Xklpl Xklpl Xklpl Nucleoplasmin N1/N2 N038
(u 1 to lZSl)
S'r.~CK T,,m,
589 K1~rmqaKLs ~ 603 1~Llqel egqm~ e 2/~i~ 704 L~Kt eeaasan~RiK 715 z~LiKeal qRqKeame/~
155 KRpaat K K a g g ~ 534 ~t eeesplKdKda.~ 142 ~%lapds a SKVlDRKKtR
/ / -
/ : J I I I I ] I * ~ r I
:G.P~A~a 1 to 1211)
. . . .
I I I I I I I I It111 I I I I I I : I: e ~dp,la. . . . . . . . . . Ocsca~Ksm,.mrs~ . . . . . . . . ~ , Q ~
C-X(2,4)-C-X L~H-X( 3,5)-H Zinc-frog= C2H2
Figure I. Primary Sequence and Structural Features of Xklpl
(a) Xklpl protein sequence. The basic residues that are characteristic of the NLS in the stalk are boxed, and the linking amino acids are underlined. The cysteine-rich region in the tail is indicated by a broken box. The putative cdc2 phosphorylation site at position 1157 is in bold and underlined. The amino acids are numbered on the right. (b) Coiled-coil structure prediction (Lupas et al., 1991). A schematic representation of Xklpl protein structure is shown. The region pre- dicted to be coiled coil is shaded. The NLSs and the cysteine-rich regions are indicated as black bars. (c) Comparison of the four NLSs found in the Xklpl protein sequence with NLSs from known Xenopus nuclear proteins. The essential basic amino acids at both ends of the NLS are shown in uppercase and bold. (d) Dot plot comparison of the Xklpl sequence to KIF4 and to KLP3A using COMPARE and DOTPLOT (Devereux et al., 1984). Xklpl and KIF4 are highly conserved throughout their entire length (identity, 60O/o). Three regions are highly conserved between Xklpl and KLP3A: the motor domain, the NLS region, and the cysteine region (indicated with an arrow). The window size used was 30 and the stringency 20. (e) Alignment of the cysteine regions of Xklpl, KLP3A, and KIF4. Identi-
EMBL SwissProt database by using the program FASTA (Pearson and Lipman, 1988). The only significant homol- ogy found was with type I and type II metallothionein se- quences from different organisms (40% identity within an overlap of 32 amino acids and 71.8% homology in the same region). Strikingly, most of the conserved amino acids are cysteines (data not shown) that in the metallothi- oneins bind metal ions, suggesting that this region in Xklpl may also be a metal-binding domain. The cysteine-rich region in Xklpl (amino acids 1084-1127) is much larger than the region of homology with the metallothioneins. We found that part of this region conforms to the consensus sequence for C2H2 zinc fingers (Figure le), but the amino acids in the loops are not optimal for DNA-interacting zinc fingers (Rosenfeld and Margalit, 1993). Finally, close to this motif, there is a consensus cdc2 phosphorylation site (TPTKK) (Figure la) at position 1157 (Nigg, 1993).
Xklpl has no striking homology with most known KLPs except for two recently reported proteins, KIF4 from mouse (Sekine et al., 1994) and KLP3A from Drosophila melano- gaster (M. Goldberg, personal communication). As shown on the dot plots of Figure ld, Xklpl and KIF4 show a strik- ing homology, with 60% identity and 74% similarity throughout their whole length. Xklpl has also a high ho- mology with KLP3A, although it is more restricted to the motor domain (50% identity and 60% similarity), the NLS domain (43% identity and 51% similarity), and a small region in the tail that corresponds to the cysteine-rich do- main (31% similarity). In the three proteins, the positions of the nine cysteines of this domain are entirely conserved, indicating that they may perform related fu nctions in Xeno- pus, mouse, and Drosophila (Figure le).
Xklpl Is Mostly Present in Gametes and Dividing Cells To identify and characterize the Xklpl protein, we raised a polyclonal antibody (Ab 65) and a monoclonal antibody (MAb P10) against a fusion protein containing the carboxy- terminal end of the protein and a monoclonal antibody (MAb K4) against a synthetic peptide from a nonconserved sequence in the motor domain (Figure 2a). All antibodies recognized the same single band on Western blots of high speed supernatants from Xenopus eggs (Figure 2b). This protein was also recognized by a polyclonal antibody raised against a synthetic peptide corresponding to a highly conserved sequence in the motor domain of proteins of the kinesin family (named KMTBX, for kinesin microtubule- binding domain of Xenopus, although this region is rather vague). This antibody also reacted specifically with a num- ber of proteins on Western blots of high speed superna- tants from Xenopus eggs (Figure 2b, lane 1 and 2).
Figure 2c shows that Xklpl was detected by MAb P10 in oocytes, eggs, testis, and very faintly in the brain tissue
cal residues are indicated with a bar, and conservative changes are indicated with two dots. The positions of the nine cysteines are con- served and indicated with stars• Additionally, Xklpl sequence contains a consensus sequence for a C2H2 zinc finger in the region indicated.

Xklpl, a Chromosome-Associated Kinesin-Like Protein 119
mAb K4 Ab65 / mAb P10 ~ y ~ ~ .................
i i ~ i | . ! i | i
MOTOR STALK TAIL
tar ~ " ~ ' ~ C _ ! f l 97 ~.:_.. ,: , E
66~ ~ i
[
Figure 2. Antibodies to Xklpl and Tissue Distribution (a) Anti-Xklpl antibodies. The peptide used to generate MAb K4 and the fusion protein used to generate Ab 65 and MAb P10 are shown on the schematic representation of Xklpl protein. (b) Western blot of Xenopus egg high speed supernatants stained with different antibodies. For KMTBX and MAb K4, plus indicates that the antigen was added to the reaction, and minus indicates that it was not. The open arrow on the left points to the band identified as Xklpl. Molecular weight markers are indicated on the left (x 1000). All the antibodies recognize the same band. Additionally, KMTBX reacts with a reproducible set of proteins that may be related to kinesin. (c) Western blot of adult tissue extracts stained with MAb K4. Equiva- lent amounts of proteins from the tissue extracts were loaded. Xklpl can only be detected in eggs, oocytes, testis, XL177 cells, and faintly in brain.
extract, as well as in the Xenopus cell line XL177, but not in any other tissue. The fact that Xklpl was found prominently in gametes and tissue culture cells suggested that it is mostly involved in cell growth and division.
Microtubule Binding Properties of Xklpl We found that in the presence of AMP-PNP, about 60% of the protein pelleted with endogenous, taxol-stabilized microtubules. About 50% of the protein was released from microtubules by one incubation with 10 mM Mg-ATP and most of it after a second incubation with ATP. The effi- ciency of release was enhanced by adding 0.5 M NaCI to the ATP, and unlike the case for other KLPs, about 50% of Xklpl was eluted from microtubules by 0.5 M salt alone (data not shown). This is similar to the microtubule binding properties of KIF4 (Sekine et al., 1994), as expected from the high level of conservation between the two proteins.
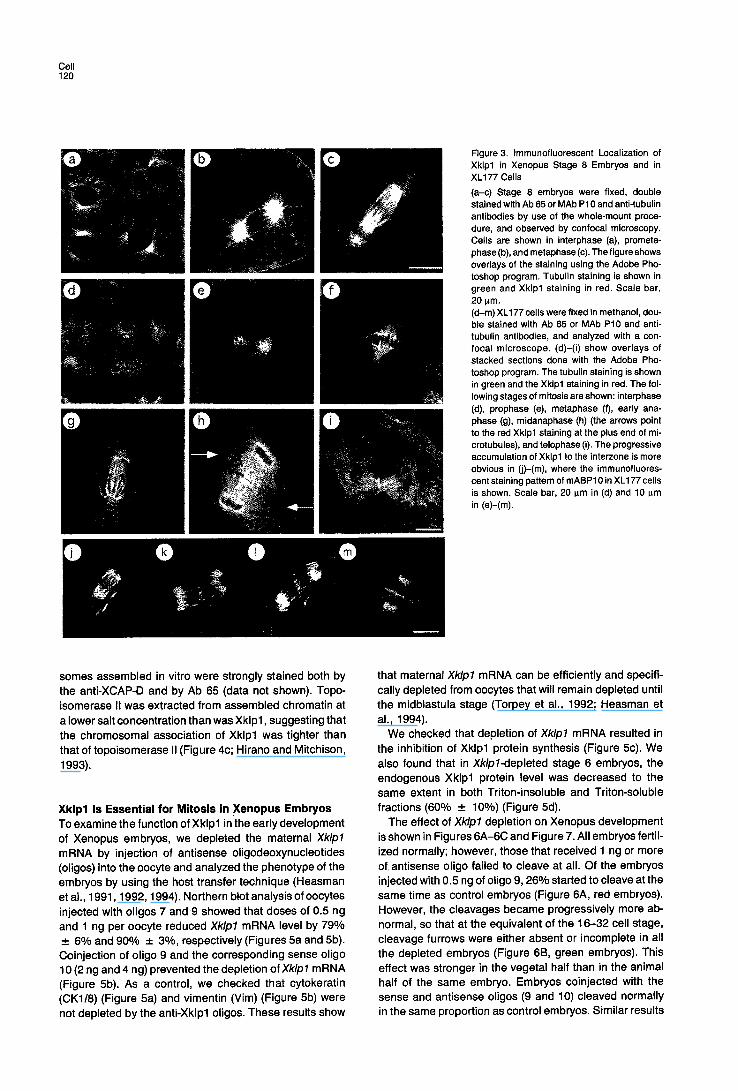
Xklpl Is Localized in the Nucleus, on Mitotic Chromosomes, and at the Spindle Interzone The subcellular distribution of Xklpl was determined by confocal microscopy of whole-mount stained Xenopus e m- bryos (stage 8/9; Figures 3a, 3b, and 3c). Xklpl was de- tected on chromosomes throughout the cell cycle (red color in Figure 3). There was also some staining on the
microtubules (Figure 3; yellow color due to overlap of green anti-tubulin and red anti-Xklpl stainings). The mid- body became strongly stained in telophase (data not shown).
To extend these results, we performed an immunofluo- rescence study on the Xenopus cell line XL177 (Miller and Daniel, 1977). During interphase, Xklpl was detected in the nucleus (Figure 3d). In most cases, the protein seemed to colocalize tightly with DNA foci strongly labeled with Hoechst (data not shown). During prophase, Xklpl stain- ing became very intense on the condensing chromosomes (Figure 3e; Hoechst chromosomal staining not shown). This intense chromosomal staining persisted throughout mitosis (Figures 3f-3i). After the separation of the chroma- tids during anaphase (Figures 3g and 3j), an additional staining appeared as elongated bars in the spindle in- terzone, and as dots at the plus end of nonoverlapping spindle microtubules (Figures 3h and 3k-31). This staining became progressively more intense during anaphase and telophase, when the protein seemed to migrate along in- terzonal microtubules (Figures 3j-3m). Finally, Xklpl be- came concentrated into a disc oriented perpendicular to the spindle axis as determined by the observation of serial confocal sections. This disc was localized in the region of reduced tubulin staining, and its diameter decreased at the same time as the cleavage furrow formed (Figure 3i). The protein localized at the interzone during telophase became incorporated into the midbody at the end of cyto- kinesis.
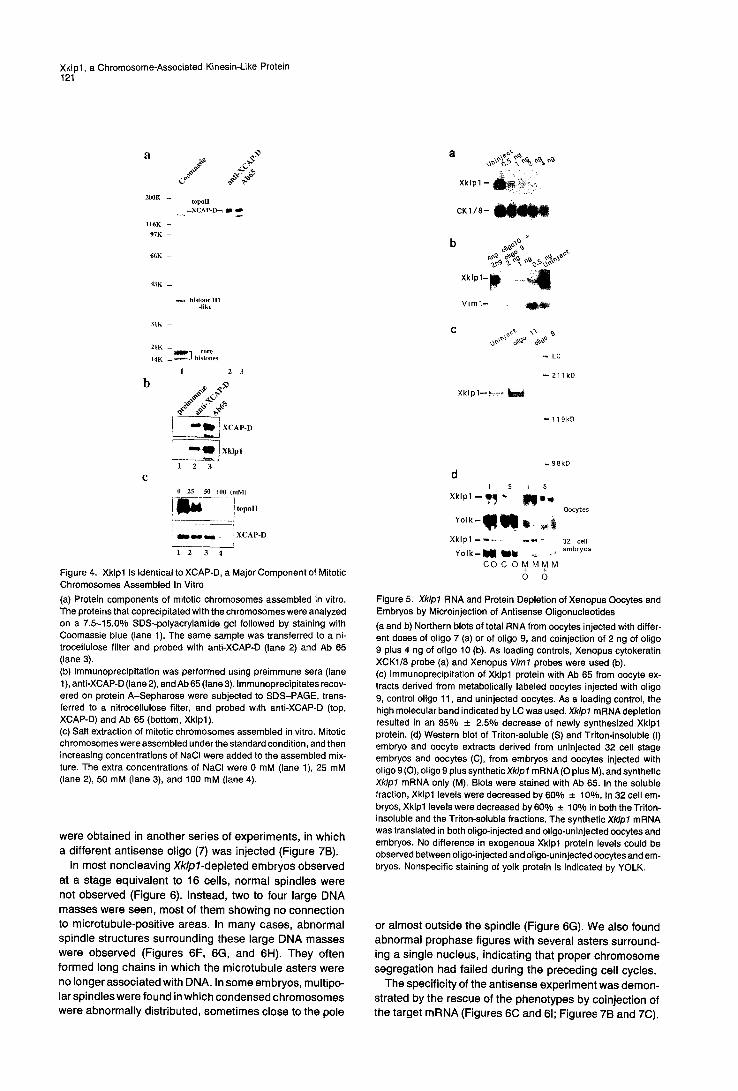
Xklpl Is a Major Component of Mitotic Chromosomes Assembled In Vitro Xklpl was also found associated with mitotic chromo- somes by a completely independent approach aimed at identifying proteins involved in chromatin condensation. Mitotic chromosomes assembled from sperm chromatin in a mitotic extract were isolated by sedimentation through a sucrose cushion (Hirano and Mitchison, 1994). As shown in Figure 4a (lane 1), in addition to histones and topoisom- erase II, a subset of polypeptides was reproducibly recov- ered in the high molecular weight range. One major chro- mosomal polypeptide (T. H., unpublished data) named XCAP-D (for Xenopus chromosome-associated polypep- tide D) was used to raise a mouse polyclonal antibody. The anti-XCAP-D specifically recognized a 150/155 kDa doublet band in the chromosomal fraction (Figure 4a, lane 2) and a 150 kDa single band in a total egg extract (data not shown). A Xenopus oocyte expression library was screened by using this antibody and a positive clone with a 0.9 kb insert isolated. The nucleotide sequence of the cDNA fragment was found identical to that of the Xklpl cDNA between nucleotides 850 and 1766, indicating that the two proteins identified by completely different strate- gies were the same. This was finally confirmed, because Ab 65 reacted with the 150 kDa band immunoprecipitated by the anti-XCAP-D (Figure 4b, lane 2, bottom), while the anti-XCAP-D antibody reacted with immunoprecipitated Xklpl (Figure 4b, lane 3, top). Thus, Xklpl was identical to XCAP-D, and it was a major chromosomal protein. Ac- cordingly, the whole chromosome arms of mitotic chromo-
Cell 120
Figure 3. immunofluorescent Localization of Xklpl in Xenopus Stage 8 Embryos and in XL177 Cells (a-c) Stage 8 embryos were fixed, double stained with Ab 65 or MAb P10 and anti-tubulin antibodies by use of the whole-mount proce- dure, and observed by confocal microscopy. Cells are shown in interphase (a), prometa- phase (b), and metaphase (c). The figure shows overlays of the staining using the Adobe Pho- toshop program. Tubulin staining is shown in green and Xklpl staining in red. Scale bar, 20 p.m. (d-m) XL177 cells were fixed in methanol, dou- ble stained with Ab 65 or MAb P10 and anti- tubulin antibodies, and analyzed with a con- focal microscope. (d)-(i) show overlays of stacked sections done with the Adobe Pho- toshop program. The tubunn staining is shown in green and the Xklpl staining in red. The fol- lowing stages of mitosis are shown: interphase (d), prophase (e), metaphase (t), early ana- phase (g), midanaphase (h) (the arrows point to the red Xklpl staining at the plus end of mi- crotubules), and telophase (i). The progressive accumulation of Xklpl to the interzone is more obvious in (j)-(m), where the immunofluores- cent staining pattern of mABP10 in XL177 cells is shown. Scale bar, 20 p.m in (d) and 10 I~m in (e)-(m).
somes assembled in vitro were strongly stained both by the anti-XCAP-D and by Ab 65 (data not shown). Topo- isomerase II was extracted from assembled chromatin at a lower salt concentration than was Xklpl, suggesting that the chromosomal association of Xklpl was tighter than that of topoisomerase II (Figure 4c; Hirano and Mitchison, 1993).
Xklpl Is Essential for Mitosis in Xenopus Embryos To examine the function of Xklpl in the early development of Xenopus embryos, we depleted the maternal Xklpl mRNA by injection of antisense oligodeoxynucleotides (oligos) into the oocyte and analyzed the phenotype of the embryos by using the host transfer technique (Heasman et al., 1991,1992, 1994). Northern blot analysis of oocytes injected with oligos 7 and 9 showed that doses of 0.5 ng and 1 ng per oocyte reduced Xklpl mRNA level by 79% _+ 6% and 90% -+ 3°/0, respectively (Figures 5a and 5b). Coinjection of oligo 9 and the corresponding sense oligo 10 (2 ng and 4 ng) prevented the depletion ofXklpl mRNA (Figure 5b). As a control, we checked that cytokeratin (CK1/8) (Figure 5a) and vimentin (Vim) (Figure 5b) were not depleted by the anti-Xklpl oligos. These results show
that maternal Xklpl mRNA can be efficiently and specifi- cally depleted from oocytes that will remain depleted until the midblastula stage (Torpey et al., 1992; Heasman et al., 1994).
We checked that depletion of Xklpl mRNA resulted in the inhibition of Xklpl protein synthesis (Figure 5c). We also found that in Xklpl-depleted stage 6 embryos, the endogenous Xklpl protein level was decreased to the same extent in both Triton-insoluble and Triton-soluble fractions (600/0 - 10°/0) (Figure 5d).
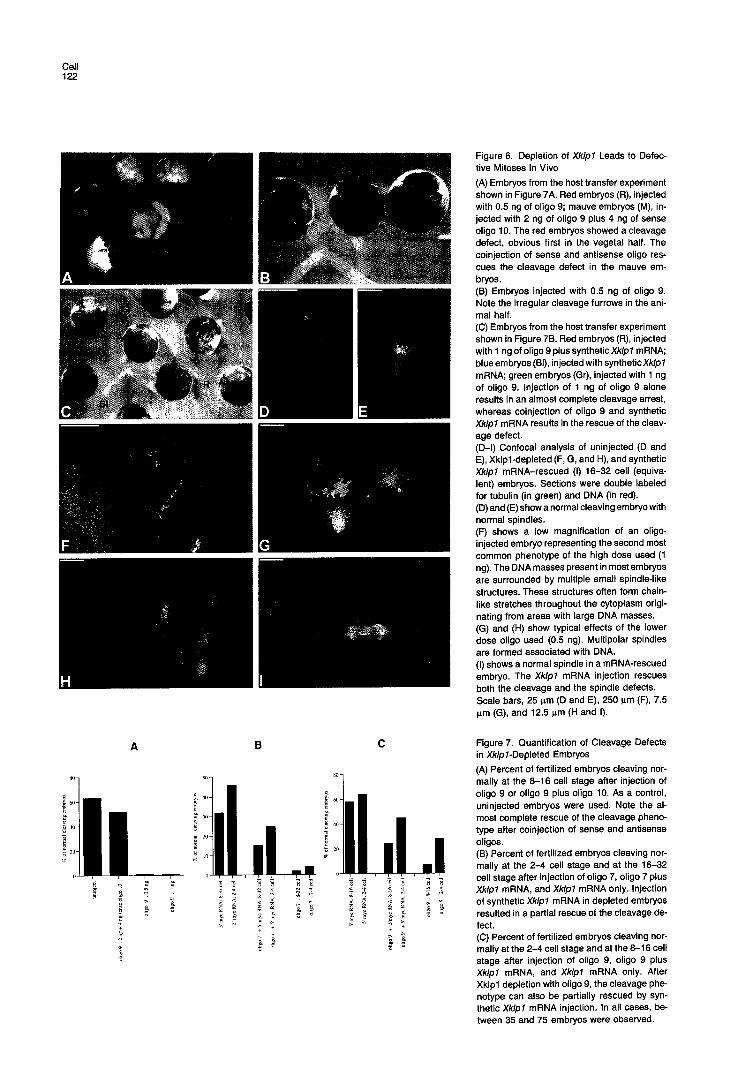
The effect of Xklpl depletion on Xenopus development is shown in Figures 6A-6C and Figure 7. All embryos fertil- ized normally; however, those that received 1 ng or more of antisense oligo failed to cleave at all. Of the embryos injected with 0.5 ng of oligo 9, 26% started to cleave at the same time as control embryos (Figure 6A, red embryos). However, the cleavages became progressively more ab- normal, so that at the equivalent of the 16-32 cell stage, cleavage furrows were either absent or incomplete in all the depleted embryos (Figure 6B, green embryos). This effect was stronger in the vegetal half than in the animal half of the same embryo. Embryos coinjected with the sense and antisense oligos (9 and 10) cleaved normally in the same proportion as control embryos. Similar results
Xklpl, a Chromosome-Associated Kinesin-Like Protein 121
a
200K -- _topoH v--XCAP-D~ ~
l I 6 K -
97K --
66K --
43K --
historic H1 -like
31K -
21K - core
I4K - ~ histories
I 2 3
b ~ ~,¢
I ~ ~1~ I KCAP.D
r e R q ~ 1 X k l p l
1 2 3
c 0 25 50 100 tmM)
!i= --!,°.°,, ! ~ a , , , ~ ] XCAP-D ~ - 2 ~- 4
Figure 4. Xklpl Is Identical to XCAP-D, a Major Component of Mitotic Chromosomes Assembled In Vitro (a) Protein components of mitotic chromosomes assembled in vitro. The proteins that coprecipitated with the chromosomes were analyzed on a 7.5-15.0% SDS-polyacrylamide gel followed by staining with Coomassie blue (lane 1). The same sample was transferred to a ni- trocellulose filter and probed with anti-XCAP-D (lane 2) and Ab 65 (lane 3). (b) Immunoprecipitation was performed using preimmune sera (lane 1), anti-XCAP-D (lane 2), and Ab 65 (lane 3). Immunoprecipitates recov- ered on protein A-Sepharose were subjected to SDS-PAGE, trans- ferred to a nitrocellulose filter, and probed with anti-XCAP-D (top, XCAP-D) and Ab 65 (bottom, Xklpl). (c) Salt extraction of mitotic chromosomes assembled in vitro. Mitotic chromosomes were assembled under the standard condition, and then increasing concentrations of NaCI were added to the assembled mix- ture. The extra concentrations of NaCI were 0 mM (lane 1), 25 mM (lane 2), 50 mM (lane 3), and 100 mM (lane 4).
were obta ined in another series of exper iments, in which a dif ferent ant isense ol igo (7) was injected (Figure 7B).
In most noncleaving Xklpl-depleted embryos observed at a stage equiva lent to 16 cells, normal spindles were not observed (Figure 6). Instead, two to four large DNA masses were seen, most of them showing no connect ion to microtubule-posi t ive areas. In many cases, abnormal spindle structures surrounding these large DNA masses were observed (Figures 6F, 6G, and 6H). They often formed long chains in which the microtubule asters were no longer associated with DNA. In some embryos, mult ipo- lar spindles were found in which condensed chromosomes were abnormal ly distr ibuted, somet imes close to the pole
C K l / 8 - ~
Vim'J- , ~
C ~ ~
- - LC
--211kD
--119kD
.98kD
d I S t S
X k l p l - - . ~ ~, M m q
Y o l k - J l~ ~ ~ - ' c o +M MM+ M
o o
Oocytes
32 cell embryos
Figure 5. Xklpl RNA and Protein Depletion of Xenopus Oocytes and Embryos by Microinjection of Antisense Oligonucleotides (a and b) Northern blots of total RNA from oocytes injected with differ- ent doses of oligo 7 (a) or of oligo 9, and coinjection of 2 ng of oligo 9 plus 4 ng of oligo 10 (b). As loading controls, Xenopus cytokeratin XCKI/8 probe (a) and Xenopus Vim1 probes were used (b). (c) Immunoprecipitation of Xklpl protein with Ab 65 from oocyte ex- tracts derived from metabolically labeled oocytes injected with oligo 9, control oligo 11, and uninjected oocytes. As a loading control, the high molecular band indicated by LC was used. Xklpl mRNA depletion resulted in an 85% ± 2.5% decrease of newly synthesized Xklpl protein. (d) Western blot of Triton-soluble (S) and Triton-insoluble (I) embryo and oocyte extracts derived from uninjected 32 cell stage embryos and oocytes (C), from embryos and oocytes injected with oligo 9 (O), oligo 9 plus synthetic Xk/p I mRNA (O plus M), and synthetic Xk/pl mRNA only (M). Blots were stained with Ab 65. In the soluble fraction, Xklpl levels were decreased by 60% ± 100. In 32 cell em- bryos, Xklpl levels were decreased by 60% ± 10% in both the Triton- insoluble and the Triton-soluble fractions. The synthetic Xklpl mRNA was translated in both oligo-injected and oligo-uninjected oocytes and embryos. No difference in exogenous Xklpl protein levels could be observed between oligo-injected and oligo-uninjected oocytes and em- bryos. Nonspecific staining of yolk protein is indicated by YOLK.
or almost outs ide the spindle (Figure 6G). We also found abnormal prophase figures with several asters surround- ing a single nucleus, indicat ing that proper ch romosome segregat ion had fai led during the preceding cell cycles.
The specif ici ty of the ant isense exper iment was demon- strated by the rescue of the phenotypes by coinject ion of the target mRNA (Figures 6C and 61; Figures 7B and 7C).
Cell 122
A B C
40 ~ 60
40
~o
=
Figure 6. Depletion of Xklpl Leads to Defec- tive Mitoses In Vivo
(A) Embryos from the host transfer experiment shown in Figure 7A. Red embryos (R), injected with 0.5 ng of oligo 9; mauve embryos (M), in- jected with 2 ng of oligo 9 plus 4 ng of sense oligo 10. The red embryos showed a cleavage defect, obvious first in the vegetal half. The coinjection of sense and antisense oligo res- cues the cleavage defect in the mauve em- bryos. (B) Embryos injected with 0.5 ng of oligo 9. Note the irregular cleavage furrows in the ani- mal half. (C) Embryos from the host transfer experiment shown in Figure 7B. Red embryos (R), injected with 1 ng of oligo 9 plus synthetic Xklpl mRNA; blue embryos (BI), injected with synthetic Xklpl mRNA; green embryos (Gr), injected with 1 ng of oligo 9. injection of 1 ng of oligo 9 alone results in an almost complete cleavage arrest, whereas coinjection of oligo 9 and synthetic Xklpl mRNA results in the rescue of the cleav- age defect. (D-I) Confocal analysis of uninjected (D and E), Xklpl-depleted (F, G, and H), and synthetic Xklpl mRNA-rescued (I) 16-32 cell (equiva- lent) embryos. Sections were double labeled for tubulin (in green) and DNA (in red). (D) and (E) show a normal cleaving embryo with normal spindles. (F) shows a low magnification of an oligo- injected embryo representing the second most common phenotype of the high dose used (1 ng). The DNA masses present in most embryos are surrounded by multiple small spindle-like structures. These structures often form chain- like stretches throughout the cytoplasm origi- nating from areas with large DNA masses. (G) and (H) show typical effects of the lower dose oligo used (0.5 ng). Multipolar spindles are formed associated with DNA. (I) shows a normal spindle in a mRNA-rescued embryo. The Xklpl mRNA injection rescues both the cleavage and the spindle defects. Scale bars, 25 p.m (D and E), 250 p.m (F), 7.5 p.m (G), and 12.5 I.tm (H and I).
Figure 7. Quantification of Cleavage Defects in Xklpl-Depleted Embryos
(A) Percent of fertilized embryos cleaving nor- mally at the 8-16 cell stage after injection of oligo 9 or oligo 9 plus oligo 10. As a control, uninjected embryos were used. Note the al- most complete rescue of the cleavage pheno- type after coinjection of sense and antisense oligos. (B) Percent of fertilized embryos cleaving nor- mally at the 2-4 cell stage and at the 16-32 cell stage after injection of oligo 7, oligo 7 plus Xk/pl mRNA, and Xklpl mRNA only. Injection of synthetic Xklpl mRNA in depleted embryos resulted in a partial rescue of the cleavage de- fect. (C) Percent of fertilized embryos cleaving nor- mally at the 2-4 cell stage and at the 8-16 cell stage after injection of oligo 9, oligo 9 plus Xklpl mRNA, and Xklpl mRNA only. After Xklpl depletion with oligo 9, the cleavage phe- notype can also be partially rescued by syn- thetic Xklpl mRNA injection. In all cases, be- tween 35 and 75 embryos were observed.
Xklpl, a Chromosome-Associated Kinesin-Like Protein 123
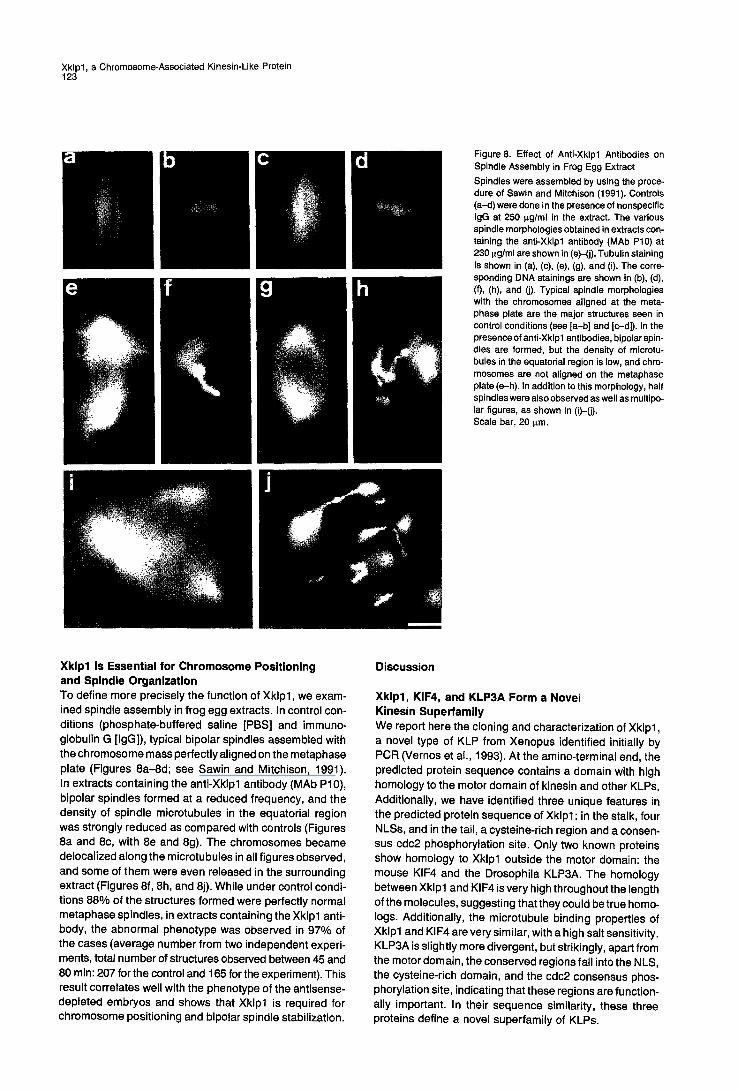
b d Figure 8. Effect of Anti-Xklpl Antibodies on Spindle Assembly in Frog Egg Extract Spindles were assembled by using the proce- dure of Sawin and Mitohison (1991). Controls (a-d) were done in the presence of nonspecific IgG at 250 ~g/ml in the extract. The various spindle morphologies obtained in extracts con- taining the anti-Xklpl antibody (MAb P10) at 230 p.g/ml are shown in (e)-(j). Tubulin staining is shown in (a), (c), (e), (g), and (i). The corre- sponding DNA stainings are shown in (b), (d), (f), (h), and (j). Typical spindle morphologies with the chromosomes aligned at the meta- phase plate are the major structures seen in control conditions (see [a-b] and [c-d]). In the presence of anti-Xklpl antibodies, bipolar spin- dles are formed, but the density of microtu- bules in the equatorial region is low, and chro- mosomes are not aligned on the metaphase plate (e-h). In addition to this morphology, half spindles were also observed as well as multipo- lar figures, as shown in (i)-(j). Scale bar, 20 p.m.
Xklpl Is Essential for Chromosome Positioning and Spindle Organization To define more precisely the function of Xklpl, we exam- ined spindle assembly in frog egg extracts. In control con- ditions (phosphate-buffered saline [PBS] and immuno- globulin G [IgG]), typical bipolar spindles assembled with the chromosome mass perfectly aligned on the metaphase plate (Figures 8a-8d; see Sawin and Mitchison, 1991). In extracts containing the anti-Xklpl antibody (MAb P10), bipolar spindles formed at a reduced frequency, and the density of spindle microtubules in the equatorial region was strongly reduced as compared with controls (Figures 8a and 8c, with 8e and 8g). The chromosomes became delocalized along the microtubules in all figures observed, and some of them were even released in the surrounding extract (Figures 8f, 8h, and 8j). While under control condi- tions 88% of the structures formed were perfectly normal metaphase spindles, in extracts containing the Xklpl anti- body, the abnormal phenotype was observed in 97% of the cases (average number from two independent experi- ments, total number of structures observed between 45 and 80 rain: 207 for the control and 165 for the experiment). This result correlates well with the phenotype of the antisense- depleted embryos and shows that Xklpl is required for chromosome positioning and bipolar spindle stabilization.
Discussion
Xklpl, KIF4, and KLP3A Form a Novel Kinesin Superfamily We report here the cloning and characterization of Xklpl, a novel type of KLP from Xenopus identified initially by PCR (Vemos et al., 1993). At the amino-terminal end, the predicted protein sequence contains a domain with high homology to the motor domain of kinesin and other KLPs. Additionally, we have identified three unique features in the predicted protein sequence of Xklpl: in the stalk, four NLSs, and in the tail, a cysteine-rich region and a consen- sus cdc2 phosphorylation site. Only two known proteins show homology to Xklpl outside the motor domain: the mouse KIF4 and the Drosophila KLP3A. The homology between Xklpl and KIF4 is very high throughout the length of the molecules, suggesting that they could be true homo- logs. Additionally, the microtubule binding properties of Xklpl and KIF4 are very similar, with a high salt sensitivity. KLP3A is slightly more divergent, but strikingly, apart from the motor domain, the conserved regions fall into the NLS, the cysteine-rich domain, and the cdc2 consensus phos- phorylation site, indicating that these regions are function- ally important. In their sequence similarity, these three proteins define a novel superfamily of KLPs.

Cell 124
Tissue Distribution and Subcellular Localization of Xklpl and KIF4 Xklpl and KIF4 have been isolated by PCR from a Xeno- pus oocyte cDNA library and a 5-day-old murine brain cDNA library, respectively. Xklpl is mostly present in oo- cytes and during early development as well as in testis, while KIF4 is present in juvenile mouse tissues but not in adult tissues. It seems that in Xenopus, Xklpl is present in minute amounts in brain extracts (see Figure 2c). We conclude that the main function of these proteins is associ- ated with cell division.
Sekine et al. (1994) report that in juvenile neurones, about 50% of KIF4 is associated with the microsomal frac- tion. We have preliminary results in Xenopus egg extracts indicating that part of Xklpl could associate with vesicles (I. V. and E. K., unpublished data). We were able to distin- guish some vesicular staining in the cytoplasm of XL177 cells and embryos by immunofluorescence, but we could not decide whether this was real. The staining seen on spindle microtubules, in particular at the microtubule plus ends in late anaphase, may well correspond to vesicles involved in cleavage furrow formation. The main discrep- ancy between the localization reported for Xklpl and that for KIF4 concerns the chromosomal association of Xklpl .
Nuclear and Chromosomal Localization of Xklpl The presence of several NLSs in the Xklpl protein se- quence suggests that it is transported very efficiently into the nucleus (Dingwall and Laskey, 1991). In agreement with this, two different anti-Xklpl antibodies stain the nu- clei of Xenopus tissue culture and embryonic cells. The potential homolog of Xklpl in the mouse, KIF4, also con- tains NLSs in its stalk domain, and Sekine et al. (1994) detect it weakly in the nucleus. By contrast, Xklpl is de- tected on chromosomes, whereas KIF4 is not.
There are seve ral reasons to think that the chromosomal localization of Xklpl is real. In prophase, the chromo- somes are already strongly stained, although the nuclear envelope has not yet disassembled, ruling out a potential contamination of chromosomes by cytoplasmic stores of Xklpl during fixation. We usually fix cells and embryos with methanol alone or low glutaraldehyde concentrations in the presence of Triton. Although both methods are known to increase accessibility of difficult antigens to anti- bodies, the chemical reactions involved in protein fixation are entirely different; yet both methods allow visualization of the chromosomal staining. Sekine et al. (1994) fix the cells by using paraformaldehyde and glutaraldehyde fol- lowed by permeabilization. This kind of fixation strongly cross-links both soluble and cytoskeletal proteins in the cells, forming a thick gel that prevents accessibility of many antigens to antibodies. We think that this is the rea- son why they could not see KIF4 on the chromosomes. To test this possibility, we repeated the staining of XL177 cells with anti-Xklpl antibodies (both monoclonal and poly- clonal) after fixation, using the procedure reported by Sek- ine et al. (1994). We found the same kind of staining they report: poor staining on spindle microtubules and no stain- ing on the chromosomes, with some staining of interphasic nuclei and diffuse cytoplasmic fluorescence.
Our immunofluorescence data, together with the direct biochemical analysis carried out entirely independently, demonstrate that Xklpl is associated with chromosomes. There are two features in the sequence of Xklpl that could be involved in the tight binding to chromatin that we ob- serve. The first feature is found in the region between the NLS sequences. It could be required for interaction with chromatin-associated proteins or directly with DNA. An- other feature potentially involved in the association with DNA is the cysteine-rich domain found in the tail of the protein. This domain contains the consensus separation of cysteines and histidines found in C2H2 zinc fingers. These structures are found in transcription factors and are known to mediate the interaction of these proteins with DNA. However, in the Xklpl sequence, the amino acids found in the loops do not fulfill the requirements for true DNA-interacting zinc fingers (Rosenfeld and Margalit, 1993). The alignment of Xklpl , KIF4, and KLP3A shows that the positions of the nine cysteines in this region are entirely conserved. It is almost certain that this domain binds metal molecules and forms a zinc finger-like struc- ture that may participate in the targeting of this class of KLPs to specific subcellular compartments.
Relocalization of Xklpl during Mitosis Xklpl is mostly associated with chromatin until anaphase. The immunofluorescence images seen in XL177 cells sug- gest that the protein begins to move along the spindle microtubules during late anaphase A and then migrates toward the equatorial region while the chromosomes move apart during ana-telophase. Some posttranslational modi- fication may occur on Xklpl to induce this movement. It is interesting to note that there is a cdc2 consensus phos- phorylation site in the tail of the protein (see Figure 1) that may be important in this process (preliminary data indicate that Xklpl is phosphorylated).
Xklpl localization to the interzone occurs very progres- sively and is not correlated with a reduction in chromo- somal staining. This timing of relocalization suggests that Xklpl may not be required for antiparallel microtubule slid- ing in anaphase B.
Xklpl Function The antisense experiments show that Xklpl depletion re- sults in an inhibition of cell division with formation of aber- rant spindle figures. The occurrence of many asters not associated with chromosome masses suggests that cen- trosomes continue to duplicate without engaging in spin- dle assembly (Gard et al., 1990). In addition, the low num- ber of nuclei found at the 16 cell stage indicates a failure in chromosome segregation. These results suggest that Xklpl is required for normal spindle function and that the cleavage inhibition observed is a consequence of this defect.
The in vitro analysis of spindle assembly shows that Xklpl is indeed essential for normal spindle function. The establishment of a bipolar spindle does not seem to require Xklpl function, although it seems essential for the stabili- zation of bipolarity. Sawin et al. (1991) have described in the same system that microtubule density is significantly greater in spindles than in half spindles. The microtubule

Xklpl, a Chromosome-Associated Kinesin-Like Protein 125
densi ty in b ipolar spindles formed in the presence of Xk lp l ant ibody is much lower than that found in control bipolar spindles, in part icular in the equator ia l region (Figure 8). This suggests that Xk lp l could be involved in stabi l izing a populat ion of spindle microtubules required to secure the bipolar state. In fact, Sawin and Mitchison (1991) have reported that some microtubules run a longside of chroma- tin wi thout any obv ious point of insertion. The local izat ion of Xk lp l on the ch romosome arms suggests that it could stabi l ize some spindle microtubules that make contact with the chromosomes. This could be one of the steps in the hierarchy of events leading to microtubule stabi l izat ion by chromat in postulated by Sawin and Mitchison (1991) and required to form the cross-br idges between the anti- paral lel microtubules of a b ipolar spindle.
In addit ion to its function in b ipolar spindle stabi l ization, Xk lp l is required for posi t ioning chromosomes on the metaphase plate. The first phases of mitosis from pro- metaphase to ear ly anaphase are character ized by an os- ci l lat ion of chromosomes, based on rapid switching of the direct ion of movemen t toward and away from the poles (Rieder and Salmon, 1994). These osci l lat ions are proba- bly essential for the congression of ch romosomes on the equator ia l plate. The po leward mot ion of chromosomes is often attr ibuted to the act iv i ty of motor proteins local ized at the k inetochores (Hyman and Mitchison, 1991). The movements away from the poles seem to be produced by a combinat ion of motor activi t ies present at the k inetochores and of eject ion forces produced by the polar arrays of microtubules pushing on the ch romosome arms (Rieder and Salmon, 1994). The nonk inetochore microtubules are dynamic, and it has been suggested that, in the growing state, they genera te enough force to push the chromo- some arms away from the pole. Another possibi l i ty that has been put forward (Theurkauf and Hawley, 1992; Zhang et al., 1990) involves the act iv i ty of plus end-d i rec ted motors associated with ch romosome arms. Since Xk lp l is a novel KLP in showing such a local izat ion and its inact ivat ion leads to delocal ized chromosomes, it is l ikely to be in- vo lved in chromosomal congression onto the metaphase
plate. The egg extract contains soluble and insoluble pools of Xk lp l . We have not demonst ra ted here that it is the chromosomal form of Xk lp l that is involved in chromo- somal congression, but this is the simplest hypothesis.
Al though the c leavage inhibit ion observed in ol igo- t reated embryos may simply be a consequence of abnor- mal spindle assembly, we cannot rule out that Xk lp l has an addit ional funct ion in c leavage furrow format ion as well, since c leavages were b locked in ant isense-treated em- bryos.
It has long been thought that ch romosome arms do not play an act ive role in spindle assembly and organizat ion, the k inetochores being the only region interact ing with spindle microtubules (Mazia, 1987; Kirschner and Mitchi- son, 1986). Over the last ten years, this idea has been chal lenged by several observat ions (Karsenti et al., 1984; Nicklas and Gordon, 1985; Sawin and Mitchison, 1991; Rieder and Salmon, 1994). Xk lp l is a novel example of a protein local ized on ch romosome arms with an essential role in spindle organizat ion.
Experimental Procedures
Isolation and Sequencing of Xklpl cDNA The Xklpl cDNA clones purified from a Xenopus oocyte cDNA library (Vernos et al., 1993) were subcloned into M13mp18 and M13mp19. Single-stranded DNA was sequenced by using Sequenase (United States Biochemical Corporation).
Production of Xklpl Fusion Proteins and Antibodies A fragment of Xklpl cDNA corresponding to the tail and part of the stalk (nucleotides 2790-3739) was ligated into the p-real-c2 vector (New England Biolabs). The fusion protein was purified by affinity chro- matography on amylose resin and used to immunize rabbits. The se- rum was affinity purified on the fusion protein. The same fusion protein was used to immunize mice, and a monoclonal antibody (MAb P10) was produced by standard methods.
The peptide FREIHQRPEWEFN (amino acids 122-134) from the motor domain of Xklpl was cross-linked to KLH and used to produce monoclonal antibodies (MAb K4).
A synthetic peptide (HIPYRESKLTRILQDSLGGRTK) correspond- ing to a highly conserved region in the motor domain of kinesins was used to produce a polyclonal antibody, KMTBX.
Preparation of Motor Proteins from Eggs Egg extracts (10,000 g) (Verde et al., 1991) were diluted three times in cold motor buffer (CMB; 100 mM K-PIPES, 0.5 mM EGTA, 2.5 mM magnesium acetate, 1 mM dithiothreitol [DI-T; pH 7] and protease inhibitors) and centrifuged at 180,000 × g for 90 rain. Taxol (20 ~M) and AMP-PNP (1.5 raM) were added for 40 rain at room temperature. The mixture was then loaded on a 15% sucrose cushion in CMB with 5 ~_M taxol and centrifuged at 22°C, 30,000 x g for 20 rain. The pellet was washed once and resuspended in CMB with 5 ~M taxol containing either 10 mM Mg-ATP, 10 mM Mg-ATP/0.5 M NaCI, or 0.5 M NaCI. After incubation for 30 rain at room temperature, the elutsd proteins were separated from the microtubules by centdfugation at 30,000 x g for 15 rain at room temperature.
Western Blotting and Extracts from Adult Tissues Western blotting were carried out according to standard procedures (Harlow and Lane, 1988). Quantitation of protein levels was performed with a Molecular Dynamics Personal Densitometer SI. Adult tissue extracts were prepared as previously described (Andersen et al., 1994).
Immunofluorescence on Xenopus Embryos Embryos were staged according to the protocols of Nieuwkoop and Faber (1956) and fixed in methanol. The whole-mount staining proce- dure was performed according to the protocols of Gard st al. (1990).
For sections, the embryos were fixed in methanol and embedded in polyethylene glycol distearate wax. Sections (35 ~m) were dewaxed in xylene-ethanol, rehydrated, and stained with the anti-tubulin anti- body (MD1A; ICN Biomedicals, Lisle, It.) and propidium iodide.
Immunofluorescence on XL177 Cells XL177 cells (Miller and Daniel, 1977) were fixed for 10 rain in methanol at-20°C. Primary antibodies in PBS with 1% bovine serum albumin (BSA) were applied for 15 min at room temperature. Ab 65 or MAb P10 was used indifferently and mixed with either the anti-tubulin anti- body from Amersham or a rabbit anti-tubulin antibody (a gift from T. Kreis). The coverslips were washed in PBS three times for 5 rain and the secondary antibodies applied for 15 rain. Secondary antibodies were purchased from Dianova (Hamburg, Federal Republic of Ger- many). The DNA was stained with Hoechst (5 ~g/ml) (Sigma), and the coverslips were mounted in Mowiol (Hoechst).
Confocal Microscopy and Image Acquisition, Processing, and Data Analysis Confocal fluorescence microscopy was performed with the compact confocal scanning laser beam microscope (CCM) developed at the European Molecular Biology Laboratory (Stelzer et al., 1989). The 529 and 476 laser lines of an argon-ion laser (Spectra-physics) were used for excitation of rhodamine-Texas red- and fluorescein-labeled sam- ples, respectively. Confocal series of mitotic cells were collected at

Cell 126
around 0.5 p.m steps, and image analysis was performed by using the Adobe Photoshop program in a Macintosh Quadra. Images for figures were contrast-enhanced in the Adobe Photoshop program and copied to Canvas (Deneba Software, Miami, FL) for final graphic presentation. Figures were digitally printed by using a dye sublimation printing pro- cess (Kodak).
Thick embryo sections were observed by using a confocal micro- scope (Bio-Rad Microsciences) fitted to an Olympus BH2RFCA.
Plasmid Construction for the Antisense Experiment The complete Xenopus Xklpl coding sequence was amplified by PCR and cloned into pSP64XI3M (Krieg and Melton, 1984) introducing the human c-Myc epitope. PCR-mediated site-directed mutagenesis (SDM) was used to introduce four base 3 mutations at the annealing site for oligo 9. Sequence analysis of the resulting vector, pXklpl myc5'sdm ~, revealed one base substitution that introduced the amino acid substitu- tion of G285D. All DNA preparations used for in vitro mRNA synthesis were purified on CsCI gradients.
Oligonucleotides Used for the Antisense Experiment Oligos 7 (5'-ATGCCCTCATCCTTCCCCAT-3') and 9 (5'-ACAGTTGGC- TCATI'CTCCTG-3') were selected as the most efficient in depleting Xklpl mRNA by Northern blot analysis. These oligos (7 and 9), the sense oligo 10 (5'-CAGGAGAATGAGCCAACTGT-3'), and a control oligo, 11 (5'-CGAACACGGAGAGTATAAGG-3) (directed against Xklp4; Vernos et al., 1993), were synthesized in a modified form in which the 5'and 3'terminal four bases were linked by methoxyethylphosphorami- date linkages instead of the conventional phosphodiester bonds (Inte- grated DNA Technologies, Incorporated). The modified oligos were di- luted in sterile distilled water, and 5-15 nl was injected at 0.1-0.5 ng/nl.
Oocytes and Embryos for the Antisense Experiments injected and control oocytes were cultured for 32-48 hr after injection, matured, labeled with vital dyes, and transferred to a host female by use of the host transfer technique as described previously (Heasman et al., 1994, 1991). Embryos from each transfer experiment were ana- lyzed by both Northern and Western blotting to ensure that mRNA knockout, exogenous protein expression, or both occurred properly.
Xenopus Oocyte Labeling, Extraction, and Immunoprecipitstion of Motor Proteins Triton-soluble and -insoluble fractions from oocytes and embryos were prepared according to the protocols of Torpey et al. (1990).
[~S]methionine-labeled total oocyte extract was obtained by cultur- ing manually defolliculated oocytes in 3.5 cm dishes in 1.5 ml of methio- nine-free 50% Leibovitz L15 supplemented with BSA (40 rag/100 ml) and 500 p_Ci of [~S]methionine (44.6 TBq/mMol specific activity; New England Nuclear-Du Pont). After an 18 hr incubation, oocytes were matured by adding progesterone (final concentration, 2 pM) for 8 hr. Mature oocytes were washed 5 x in PBS and transferred to a douncer. An equal volume of acetate buffer (100 mM potassium acetate, 2.5 mM magnesium acetate, 60 mM EGTA, 0.01 mg/ml cytochalasin D, 1 mM DTT [pH 7.2], 4°C) was added. After douncing, 900 p~l of CMB was added. The mixture was centrifuged for 90 rain at 180,000 x g, 4°C, and the middle phase was removed. To this fraction, one volume of solubilization buffer (SOL; 50 mM Tris [pH 7.5], 150 mM NaCI, 1% NP-40, protease inhibitors [PMSF, aprotinin, leupeptin, pepstatin]) was added. For precipitation of Xklpl, protein A-Sepharose beads were incubated with Ab 65 in 1 ml of SOL buffer 2 hr at room temperature. After extensive washing in SOL buffer, beads were incubated with the radiolabeled extract containing equal amounts of radioactivity (cpm) overnight at 4°C. Precipitates were washed extensively in SOL buffer and subjected to 5o/0 SDS-polyacrylamide gel electrophoresis (PAGE). The gels were fluorographed, dried, and exposed to Fuji XR5 film. Quantification of signal after fluorography was performed with a Molecular Dynamics Personal Densitometer SI.
Northern Blot Analysis Oocyte RNA was extracted as described previously (Gurdon et al., 1985). Electrophoresis and Northern blotting were performed as de- scribed by Hopwood et al. (1989), by using 1.5 oocyte equivalents per lane. Control probes were XCK1/8 coding sequence (Franz and Franke, 1986) or the Xenopus Vim1 clone (Torpey et al., 1990). Probes
were radiolabeled by use of the random prime method described by Feinberg and Vogelstein (1983). Quantitation of mRNA signals was performed with a Molecular Dynamics Personal Densitometer SI.
In Vitro mRNA Synthesis All pXklpl myc5'sdm c templates were linearized with BamHI and tran- scribed in vitro by using SP6 polymerase (Krieg and Melton, 1984; Krieg and Melton, 1987). Five to fifteen nl were injected into the equato- rial zone of the oocytes at 100-500 pg/nl.
Spindle Assembly in Extracts Spindles were assembled by following the method of Sawin and Mitchi- son (1991) and Murray (1991). Control nonspecific IgG (at 2.5 mg/ml), MAb P10 (at 2.3 mg/ml), or PBS was added to the mitotic extract at a 1:10 dilution and incubated on ice for 30 min. The mixture was then split in two tubes, and one of them was kept on ice. Sperm nuclei (2500 nuclei/id) and tetramethylrhodamine (TMR)-Iabeled bovine brain tubulin (Hyman et al., 1990) were added to the other tube and the mixture incubated at 20°C for 10 rain. Calcium was then added at a final concentration of 0.3 mM to induce the mitotic extract to enter interphase and the mixture incubated at 20°C. At 80 rain after calcium addition, the mitotic extract kept on ice was added to the interphase one to induce mitosis again. Samples were taken every 15 min and analyzed by fluorescence microscopy. Usually 1.2 ILl was spotted on a microscope slide, and 6 p.I of formaldehyde fix (Murray, 1991) was deposited on top and squashed gently under a coverslip.
Acknowledgments
Correspondence should be addressed to I. V. We thank Sharyn Endow for highlighting to us the possible relationship between Xklpl and KLP3A, Mike Goldberg for communicating results prior to publication, and Tim Mitchison and Michael Glotzer for critical reading of the manu- script. We thank D r. G. Evans for his generous supply of 9E10 antibody, Alan Sawyer for help in the preparation of antibodies, Regis Tourne- bize for help with in vitro spindle assembly, Lucinda Vickers and Kim Goldstone for their technical assistance, and the Stelzer group in Hei- delberg for help with confocal microscopy. T. H. is grateful to Tim Mitchison for his generous support and encouragement and to Arshad Desai for his help with immunofluorescent staining of tissue culture cells. We thank the Wellcome Trust, the Institute for Human Genetics of the University of Minnesota, and the Harrison Fund for financial support. The work done in Heidelberg was supported by a Human Frontier Science Program grant to E. K. (RG 350•94). The work done in San Francisco was supported by the Tobacco-Related Disease Re- search Program in the University of California. I. V. was supported by a Wellcome Trust Travelling Fellowship and a Human Capital Mobility Fellowship (ICT 930645). T. H. is a Leukemia Society of America Spe- cial Fellow.
Received December 2, 1994; revised February 3, 1995.
References
Andersen, S. S. L., Buendia, B., Dominguez, J. E., Sawyer, A., and Karsenti, E. (1994). Effect on microtubule dynamics of XMAP230, a microtubule-associated protein present in Xenopus/aevis eggs and dividing cells. J. Cell Biol. 127, 1289-1299.
Bloom, G. S., and Endow, S. A. (1995). Motor proteins I: kinesins. In Protein Profile, Volume 1, P. Sheterline, ed. (London: Academic Press), in press.
Devereux, J., Haeberli, P., and Smithies, O. (1984). A comprehensive set of sequence analysis programs for the VAX. Nucl. Acids Res. 12, 387-395.
Dingwall, C., and Laskey, R. A. (1991). Nuclear targeting sequences: a consensus? Trends Biochem. Sci. 16, 478-481.
Feinberg, A. P., and Vogelstein, B. (1983). A technique for radiola- belling DNA restriction endonuclease fragments to high specific activ- ity. Addendum: Anal. Biochem. 137, 266-267.
Franz, J. K., and Franke, W. W. (1986). Cloning of a cDNA and amino acid sequences of a cytokeratin expressed in oocytes of Xenopus laevis. Proc. Natl. Acad. Sci. USA 83, 6475-6479.

Xklpl, a Chromosome-Associated Kinesin-Like Protein 127
Gard, D. L., Hafezi, S., Zhang, T., and Doxsey, S. J. (1990). Centrosome duplication continues in cycloheximide-treated Xenopus blastulae in the absence of a detectable cell cycle. J. Cell Biol. 110, 2033-2042.
Gorbsky, G. J. (1992). Chromosome motion in mitosis. Bioassays 14, 73-80.
Gurdon, J. B., Fairman, S., Mohun, T. J=, and Brennan, S. (1985). Activation of muscle specific actin genes in Xenopus development by an induction between animal and vegetal cells of a blastula. Cell 41, 913-922.
Harlow, E., and Lane, D. (1988). Antibodies: A Laboratory Manual (Cold Spring Harbor, New York: Cold Spring Harbor Laboratory Press).
Heasman, J., Torpey, N., and Wylie, C. C. (1991). Fertilization of cul- tured Xenopus oocytes and use in studies of maternally inherited mole- cules. Meth. Cell Biol. 36, 213-235.
Heasman, J., Torpey, N., and Wylie, C. C. (1992). The role of intermedi- ate filaments in early Xenopus development studied by antisense depletion of maternal mRNA. Development (Suppl.) 116, 119-125.
Heasman, J., Ginsberg, D., Geiger, B., Goldstone, K., Partt, T., Yo- shida-Noro, C., and Wylie, C. (1994). A functional test for maternally inherited cadherin in Xenopus shows its importance in cell adhesion at the blastula stage. Development 120, 49-57.
Hirano, T., and Mitchison, T. J. (1993). Topoisomerase II does not play a scaffolding role in the organization of mitotic chromosomes assembled in Xenopue egg extracts. J. Cell Biol. 120, 601-612.
Hirano, T., and Mitchison, T. (1994). A heterodimeric coiled-coil protein required for mitotic chromosome condensation in vitro. Cell 79, 449- 458.
Hopwood, N. D., Pluck, A., and Gurdon, J. B. (1989). MyoD expression in the forming somites is an early response to mesoderm induction in Xenopus embryos. EMBO J. 8, 3409-3417.
Hyman, A. A., and Mitchison, T. (1991). Two different microtubule- based motor activities with opposite polarities in kinetochores. Nature 351,206-211. Hyman, A. A., Drechsel, D., Kellogg, D., Salser, S., Sawin, K., Steffen, P., Wordeman, L., and Mitchison, T. (1990). Preparation of modified tubulines. Meth. Enzymol. 196, 303-319.
Karsenti, E., Newport, J., Hubble, R., and Kirschner, M. W. (1984). Interconversion of metaphase and interphase microtubule arrays, as studied by the injection of centrosomes and nuclei into Xenopus eggs. J. Cell Biol. 98, 1730-1745.
Kirschner, M. W., and Mitchison, T. J. (1986). Beyond self assembly: from microtubules to morphogenesis. Cell 45, 329-342.
Krieg, P. A., and Melton, D. A. (1984). Functional messenger RNAs are produced by Sp6 in vitro transcription of cloned cDNAs. Nucl. Acids Res. 12, 7057-7070.
Krieg, P. A., and Melton, D. A. (1987). In vitro RNA synthesis with SP6 RNA polymerase. Meth. Enzymol. 155, 397-415.
Lupas, A., Van Dyke, M., and Stock, J. (1991). Predicting coiled coils from protein sequences. Science 252, 1162-1164.
Mazia, D. (1987). The chromosome cycle and the centrosome cycle in the mitotic cycle. Int. Rev. Cytol. 100, 49-92.
Miller, L., and Daniel, J. C. (1977). Comparison of in vivo and in vitro ribosomal RNA synthesis in nucleolar mutants of Xenopus/aevis. In Vitro 13, 557-567.
Murray, A. W. (1991). Cell cycle extracts= Meth. Cell Biol. 36, 573- 597.
Nicklas, R. B., and Gordon, G. W. (1985). The total length of spindle microtubule depends on the number of chromosomes present. J. Cell Biol. 100, 1-7.
Nieuwkoop, P. D., and Faber, J. (1956). Normal Table of Xenopus /aevis (Amsterdam: Daudin).
Nigg, E. A. (1993). Targets of cyclin-dependent protein kinases. Curr. Opin. Cell. Biol. 5, 187-193.
Pearson, W. R., and Lipman, D. J. (1988). Improved tools for biological sequence comparison. Proc. Natl. Acad. Sci. USA 85, 2444-2448.
Pereira, A., and Goldstein, L S. B. (1994). The kinesin superfamily.
In Microtubules, J. S. Hyams and C. W. Lloyd, eds. (New York: Wiley- Lies), pp. 269-284.
Rieder, C. L., and Salmon, E. D. (1994). Motile kinetochores and polar ejection forces dictate chromosome position on the vertebrate mitotic spindle. J. Cell Biol. 124, 223-233.
Rosenfeld, R., and Margalit, H. (1993). Zinc fingers: conserved proper- ties that can distinguish between spurious and actual DNA-binding motifs. J. Biomol. Struct. Dyn. 11, 557-569.
Sawin, K. E., and Mitchison, T. J. (1991). Mitotic spindle assembly by two different pathways in vitro. J. Cell Biol. 112, 925-940.
Sekine, Y., Okada, Y., Noda, Y., Kondo, S., Aizawa, H., Takemura, R., and Hirokawa, N. (1994). A novel microtubule-based motor protein (KIF4) for organelle transport, whose expression is regulated develop- mentally. J. Cell Biol. 127, 187-201.
Stelzer, E. H. K., Stricker, R., Pick, R., Storz, C., and H&nninen, P. (1989). Confocal fluorescence microscopes for biological research. Proc. Soc. Photo-opt. Instr. Eng. 1028, 146-151.
Theurkauf, W. E., and Hawley, R. S. (1992). Meiotic spindle assembly in Drosophila females: behavior of non-exchange chromosomes and the effects of mutations in the nod kinesin-like protein. J. Cell Blot. 116, 1167-1180.
Torpey, N. P., Heasman, J., and Wylie, C. C. (1990). Identification of vimentin and novel vimentin-related proteins in Xenopus oocytes and early embryos. Development 110, 1185-1195.
Torpey, N. P., Wylie, C. C., and Heasman, J. (1992). The function of maternal cytokeratin in Xenopus development. Natu re 357, 413-415.
Verde, F., Berrez, J. M., Antony, C., and Karsenti, E. (1991). Taxol induced microtubule asters in mitotic extracts of Xenopus eggs: re- quirement for phosphorylated factors and cytoplasmic dynein. J. Cell Biol. 112, 1177-1187.
Vernos, I., Heasman, J., and Wylie, C. (1993). Multiple kinesin-like transcripts in Xenopus oocytes. Dev. Biol. 157, 232-239.
Zhang, P., Knowles, B. A., Goldstein, L. S. B., and Hawley, R. S. (1990). A kinesin-like protein required for distributive chromosome seg- regation in Drosophila. Cell 62, 1053-1062.
GenBank Accession Number
The GenBank accession number for the sequence reported in this paper is X82012.



















